
TGF-bi Promotes Islet b-Cell Function and Regeneration
13
of April 12, 2019. This information is current as Regeneration -Cell Function and β i Promotes Islet β TGF- Wu Bing Han, Shijie Qi, Bing Hu, Hongyu Luo and Jiangping http://www.jimmunol.org/content/186/10/5833 doi: 10.4049/jimmunol.1002303 April 2011; 2011; 186:5833-5844; Prepublished online 6 J Immunol References http://www.jimmunol.org/content/186/10/5833.full#ref-list-1 , 19 of which you can access for free at: cites 50 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2011 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 12, 2019 http://www.jimmunol.org/ Downloaded from by guest on April 12, 2019 http://www.jimmunol.org/ Downloaded from
Transcript of TGF-bi Promotes Islet b-Cell Function and Regeneration

of April 12, 2019.This information is current as
Regeneration-Cell Function andβi Promotes Islet βTGF-
WuBing Han, Shijie Qi, Bing Hu, Hongyu Luo and Jiangping
http://www.jimmunol.org/content/186/10/5833doi: 10.4049/jimmunol.1002303April 2011;
2011; 186:5833-5844; Prepublished online 6J Immunol
Referenceshttp://www.jimmunol.org/content/186/10/5833.full#ref-list-1
, 19 of which you can access for free at: cites 50 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
TGF-bi Promotes Islet b-Cell Function and Regeneration
Bing Han,* Shijie Qi,* Bing Hu,† Hongyu Luo,* and Jiangping Wu*,‡
TGF-bi is a secreted protein and is capable of binding to both extracellular matrix (ECM) and cells. It thus acts as a bifunctional
molecule enhancing ECM and cell interactions, a lack of which results in dysfunction of many cell types. In this study, we
investigated the role of TGF-bi in the function and survival of islets. Based on DNA microarray followed by quantitative PCR
confirmation, TGFbi gene showed drastic increase in expression in islets after culture. We demonstrated that recombinant TGF-bi
could preserve the integrity and enhance the function of cultured islets. Such a beneficial effect was mediated via signaling through
FAK. Exogenous TGF-bi was capable of sustaining high-level FAK phosphorylation in isolated islets, and FAK knockdown by
small interfering RNA in islets resulted in compromised islet function. TGF-bi transgenic (Tg) islets showed better integrity and
insulin release after in vitro culture. In vivo, b-cell proliferation was detectable in Tg but not wild-type pancreata. At age above 12
mo, Tg pancreata contained giant islets. Tg mice displayed better glucose tolerance than that of the controls. Tg islets were more
potent in lowering blood glucose when transplanted into syngeneic mice with streptozotocin-induced diabetes, and these trans-
planted islets also underwent regeneration. Our results indicate that TGF-bi is a vital trophic factor promoting islet survival,
function, and regeneration. At least some of its beneficial effect was mediated by signaling through FAK. The Journal of
Immunology, 2011, 186: 5833–5844.
In vivo, islets are situated in a complex environment, withconstant interaction and support from the extracellular matrix(ECM) and neighboring cells. Loss of such interaction and
support is detrimental to islet function and viability. It is wellknown that isolated islets lose their morphologic integrity afterseveral days of culture, followed by b-cell apoptosis. Dispersedb-cells have reduced insulin-secretion capability, and reaggre-gation restores their near-normal insulin secretion rate (1–3). TheECM is certainly critical for the above-described phenomena, asits RGD motif can promote b-cell survival via signaling througha5b1 and aVb3 on the b-cell surface (4–6). In addition, the RGDmotif of the ECM can interact with and signal through a1b1integrin on b-cells and enhance insulin secretion (7). b-cells alsoexpress a3b1, which, like a5b1 and aVb3, can bind to RGDof the ECM (8, 9). Cell–cell interaction is probably equally es-sential for b-cell function, as insulin-secreting MIN6 cells showsignificantly enhanced insulin release when aggregated (10). Both
E-cadherin and N-cadherin are implicated in b-cell/b-cell andb-cell/non–b-cell interactions. The former is expressed on bothb-cells and non–b-cells in islets (10, 11), and the latter is ex-pressed in a subpopulation of b-cells producing high levels ofinsulin (12). Transgenic (Tg) overexpression of dominant negativeE-cadherin displaces both endogenous E-cadherin and N-cadherinon b-cells and results in loss of islet formation in the pancreas(13).These findings suggest that measures preserving and promoting
b-cell–ECM interaction and methods enhancing b-cell inter-actions with surrounding cells will likely ameliorate the survivaland function of native and transplanted islets.TGFbi (also known as big-h3) is a gene cloned from TGF-b–
stimulated A549 lung adenocarcinoma cells (14). TGF-bi proteinhas an N-terminal secretory signal (aa 1–23), four FAS1 homol-ogous internal domains, and a cell attachment site (RGD) at its Cterminus (15). TGF-bi is a secreted protein that is also found inthe cytoplasm and nucleus of cells (16). It is produced by severalcell types, such as smooth muscle cells, keratinocytes, fibroblasts(16), and renal proximal tubular epithelial cells (17). It is upreg-ulated in the kidney in diabetic conditions due to TGF-b or highglucose stimulation (18, 19), but whether and how such upregu-lation affects the progress of diabetic nephropathy is not clear. Insitu hybridization shows that TGF-bi is also expressed in theheart, blood vessels, intestine, eyes, and other tissues of mesen-chymal origin (20), indicating that this gene plays a general role invarious organs during ontogeny.Although the biological significance of cytoplasmic and nuclear
TGF-bi presence is not clear to date, we have a basic under-standing of secreted TGF-bi function. TGF-bi binds to the ECMdue to interaction of the YH motif in its FAS1 domains withECM collagen I, II, IV, and VI (21, 22). Its FAS1 domains caninteract with a3b1, aVb3, and aVb5 integrins on the cell surface(17, 23, 24). Its RGD domain at the C terminus binds to integrins.Although the types of its interacting integrins have not beenpinpointed (25, 26), in theory, the RGD domain should be able tointeract with a5b1, a8b1, aVb3, aVb5, aVb6, aVb8, and aIIb3(27). TGF-bi is also known to bind to a1b1, a6b4, a7b1 (28–30),but domain(s) involved in such associations have not been re-
*Laboratoire Immunologie, Centre de Recherche, Centre Hospitalier de l’Univer-site de Montreal2Hopital Notre-Dame, Montreal, Quebec H2L 4M1, Canada;†Quest Diagnostics, Tucker, GA 30084; and ‡Service Nephrologie, Centre de Re-cherche, Centre Hospitalier de l’Universite de Montreal2Hopital Notre-Dame, Mon-treal, Quebec H2L 4M1, Canada
Received for publication July 8, 2010. Accepted for publication March 8, 2011.
This work was supported by grants from the Canadian Institutes of Health Research(MOP57697 and MOP69089), Genome Canada/Quebec, the Heart and Stroke Foun-dation of Quebec, the Quebec Ministry of Economic Development, Innovation andExportation (PSR-SIIRI-069), and the J.-Louis Levesque Foundation (to J.W.), andby a group grant from Fonds de la Recherche en Sante du Quebec for Transfusionaland Hemovigilance Medical Research.
The array data presented in this article have been deposited into the Gene ExpressionOmnibus database (http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi) under accessionnumber GSE27547.
Address correspondence and reprint requests to Dr. Jiangping Wu, Laboratory ofImmunology, Centre Hospitalier de l’Universite de Montreal2Hopital Notre-Dame, Pavillion DeSeve, Room Y-5616, 1560 Sherbrooke Street East, Montreal,QC H2L 4M1, Canada. E-mail address: [email protected]
Abbreviations used in this article: ECM, extracellular matrix; NIRILOD, non-immune-related islet loss or dysfunction; qPCR, quantitative PCR; PCNA, prolif-erating cell nuclear Ag; RT/qPCR, reverse transcription/real-time PCR; siRNA, smallinterfering RNA; STZ, streptozotocin; Tg, transgenic; WT, wild-type.
Copyright� 2011 by The American Association of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1002303
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

ported. Thus, secreted TGF-bi acts as a bifunctional moleculeenhancing ECM and cell interactions, a lack of which results indysfunction of many cell types.Indeed, when TGF-bi is coated on solid phase, it induces ad-
hesion, reduces proliferation, and inhibits the differentiation ofkeratinocytes (24). Solid-phase TGF-bi also enhances osteoblastadhesion, suppresses their differentiation (23), and blocks cornealepithelial cell adhesion and spreading (31). In this case, it is worthmentioning that TGF-bi point mutation results in many differenttypes of corneal dystrophy in humans (32). These biologicaleffects of TGF-bi could be explained by triggering of the cellsurface integrin signaling pathways. In contrast, a soluble TGF-bifragment containing its C-terminal RGD domain induces Chinesehamster ovary cell apoptosis (25). In this case, it is likely that thesoluble fragment interferes with normal cell–cell interactions be-tween integrins and fibronectin, as the RGD domain could bindto integrins and dampen their association with fibronectins. Thissuggests that TGF-bi is not only necessary for ECM–cell inter-ction but might also regulate cell–cell interaction, both of whichare essential for the well-being of cells.The effect of TGF-bi on islet survival, function, and regen-
eration, relative to its roles in islet b-cell–ECM and b-cell–neighboring cell interactions, has never been investigated and isthe focus of our project.
Materials and MethodsIslet isolation
The islet isolation procedure has been detailed in an earlier publication (33).
DNA microarray analysis
Six independent batches of islets were isolated. For each batch, islets fromfour C57BL/6 mice were pooled and were divided into three groups, whichwere harvested at 0 h, 24 h, and 48 h after culture. Τotal RNAwas extractedfrom them with Qiagen RNeasy Micro Kits. After reverse transcription,cDNA served as a template for PCR-based cRNA amplification. It wasthen reverse-transcribed to produce second-generation cDNA, which wasused to generate biotin-labeled cRNA probes. The probes were hybridizedon GeneChip Mouse Genome 430 2.0 Array (Affymetrix, Santa Clara,CA), which contains 39,000 transcripts. DNA microarray analysis wasperformed at the McGill University Genome Quebec Innovation Centre.For each time point, six chips were hybridized with cRNA from the sixbatches of isolated islets. For samples collected at 24 and 48 h (six batchesfor each time point), if the mean signal strength of a gene had above 4-folddifference over that of the 0-h samples, it was selected for reverse PCRconfirmation. Details of the experimental protocols and data analysisprocedure can be found at http://www.affymetrix.com/support/downloads/manuals/expression_analysis_technical_manual.pdf.
Reverse transcription/real-time PCR
Total RNA from islets was extracted with TRIzol (Invitrogen, Carlsbad, CA).RNAwas reverse-transcribed into cDNAwithM-MLVReverse TranscriptaseKits. Qiagen Quanti-Test SYBR Green PCR Kits were used for real-timePCR amplification of cDNA templates. The PCR amplification programwas as follows: 95˚C, 15 min, 1 cycle; 95˚C, 10 s, 56˚C, 15 s, 72˚C, 20 s, 45cycles. The primers for DNA microarray data verification and TGF-bimRNA quantification are listed in Table I. The quantitative PCR (qPCR)samples were in duplicate. In addition to melting curve analysis, the qPCRproducts were also resolved by agarose gel electrophoresis to ascertain thatthe expected band-size was obtained. The data were expressed as ratiosbetween the signal of the mRNA in question and that of b-actin.
Evaluation of cultured islet integrity
Islets were cultured in F12K complete medium containing 10% FBS inthe absence or presence of recombinant human TGF-bi (5 mg/ml; R&DSystems, Minneapolis, MN). They were observed under a microscope oncea day. An islet was considered to have lost its integrity if .50% of itscapsule had disappeared. The islet integrity index was calculated as fol-lows: integrity index = 100% 3 (number of total islets 2 number of isletswith lost integrity)/number of total islets.
Flow cytometry
Flow cytometry was performed to quantify islet apoptosis, FAK phos-phorylation, and proliferation. The islets were first dispersed by trypsindigestion. For apoptosis analysis, the cells were stained with FITC–annexinV. For FAK phosphorylation, the cells were fixed and permeabilized withBD Cytofix/Cytoperm Fixation/Permeabilization Kit (BD Biosciences, SanDiego, CA) and then stained with rabbit anti-mouse phospho-FAK Ab(Cell Signaling Technology, Beverly, MA), rat anti-insulin Ab (R&DSystems), and proper second Abs. For b-cell proliferation, the mice wereinjected with BrdU i.p. (1 mg/mouse, once every other day for three times)and sacrificed 2 d after the last injection. The dispersed islet cells werestained with FITC BrdU Flow Kit (BD Biosciences) and rat anti-insulin Abfor analysis by two-color flow cytometry.
Immunoblotting
pCEP4–TGF-bi–transfected or vector-transfected NIT-1 cells were treatedwith IFN-g and IL-1b for 24 and 48 h and then lysed. Seventy microgramsof lysate protein was loaded to each lane in 15% SDS-PAGE for cleavedcaspase 3 and 8 or 10% SDS-PAGE for cleaved caspase 9. After trans-ferring the proteins from the gel to polyvinylidene fluoride membranes, themembranes were hybridized with rabbit Abs against cleaved fragments ofmouse caspases 3, 8, and 9. The Abs were all from Cell Signaling Tech-nology (Danvers, MA). All Abs were used at a 1:1000 dilution. The pre-hybridization and hybridization procedures were performed accordingto recommended protocols of Cell Signaling Technology. The signalswere detected with SuperSignal West Pico Chemiluminescent Substrate(Thermo Scientific, Rockford, IL).
FAK siRNA transfection
Mouse FAK small interfering RNA (siRNA) was synthesized by Inte-grated DNA Technologies (Coralville, IA). The sequences of the siRNAwere rArGrArArArUrArGrCrUrGrArUrCrArArGrU and rUrUrGrGrUr-CrUrArCrUrUrGrArUrCrArGrC. The sequences of the control siRNAwere rCrUrUrCrC rUrCrUrCrU rUrUrCrUrC rUrCrCrCrU rUrGrUGA andrUrCrArCrA rArGrGrGrA rGrArGrArA rArGrArGrA rGrGrArAr GrGrA.Paired siRNA was first heated to 94˚C for 2 min and then cooled to roomtemperature for annealing. The annealed FAK siRNA was transfected intoislet cells with X-tremeGENE transfection reagent (Roche DiagnosticsGmbH, Mannheim, Germany) at a final concentration of 80 nM siRNA for100 islets/0.3 ml/well in 48-well plates. FAK mRNA knockdown wasconfirmed by reverse transcription/real-time PCR (RT/qPCR) 48 h aftertransfection. The primers for this purpose were 59-ACCATCCCTAAC-CATTGCCGAGAA-39 and 59-ACTTTGGTATTGATGGCAACGCCC-39.
Insulin release assay
The assay was conducted as reported elsewhere (33). The data are presentedas the mean + SD of ratios of insulin levels upon low (2.8 mM) versus high(16.7 mM) glucose stimulation.
Generation of TGF-bi Tg mice
Human full-length TGF-bi cDNA (MGC-1598) was excised from thevector with BamHI/XbaI and cloned into BamHI/XhoI sites in vector pAC(34) between the human b-actin promoter and b-actin polyA signals. Theresulting construct was named pAC-TGF-bi. The junction and orientationof the insert were verified by sequencing. The 8.0-kb PciI/SspI fragmentcontaining the b-actin promoter, TGF-bi cDNA, and b-actin polyA signalwas excised and injected into fertilized C57BL/6 or C3H3 C57BL/6 eggs.
Tail DNA PCR was undertaken to genotype Tg mice. The 59 and 39primers were 59-TTCCC TATTG TGACA GAGCC ATGGT G-39 and 59-CACTG CATTC TAGTT GTGGT TTGTC C-39, respectively, for de-tection of a 112-bp band. The following PCR conditions were used: 94˚C,30 s; 58˚C, 1 min; 72˚C, 1 min; 30 cycles.
TGFbi ELISA
Human TGFbi ELISA kits were purchased from R&D Systems, and assayswere performed according to the manufacturer’s instructions. The sampleswere in duplicate, and assay sensitivity was 6 pg/ml.
Determination of the relative copy number of the TGF-bitransgene
Tail DNA from two lines of Tg mice and wild-type (WT) mice were assayedfor relative copy numbers of human TGF-bi transgene by qPCR. The sameprimers and condition as described earlier for genotyping the Tg mice wereused in qPCR. Mouse EphB6 gene was used as an internal control in qPCRusing the forward primer 59-AAGCCATAGCAGTGCCTCAGAACA-39
5834 THE ROLE OF TGF-bi IN b-CELL FUNCTION AND GROWTH
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

and reverse primer 59-TCCAGAGCTAGAACTGATGACCCT-39, with thesame qPCR amplification condition as described earlier. The relative copynumbers of the transgene are determined by ratios of the human TGF-bitransgene signal versus the endogenous mouse EphB6 gene signal. As WTmice contained no human TGF-bi transgene, its ratio is near zero.
Immunohistochemistry and immunofluorescence
The same paraffin-fixed sections of pancreata of Tg and WT mice werestained by proliferating cell nuclear Ag (PCNA) immunohistochemistry andinsulin immunofluorescence. The rabbit anti-insulin mAb and mouse anti-PCNA mAb were both from Cell Signaling Technology (Danvers, MA)and were used at a 1:100 dilution. DyLight 488 anti-rabbit IgG Ab (1:100;Jackson ImmunoResearch Laboratories, West Grove, PA) and HRP-linkedanti-mouse IgG Ab (1:200; GE Healthcare, Little Chalfont Bucking-hamshire, U.K.) were used as secondary Abs, respectively. Signals in im-munohistochemistry were revealed using DAB Substrate kit (BD Bio-sciences). For transplanted islets, paraffin-fixed kidneys under the capsuleof which the islets were transplanted were consecutively sectioned; onesection was stained with rabbit anti-insulin mAb and the following sectionwas stained with mouse anti-PCNA mAb. The sections were then reactedwith HRP-conjugated anti-rabbit IgG Ab (1:200; GE Healthcare) or HRP-conjugated anti-mouse IgG Ab, respectively, followed by 3,39-diamin-obenzidine tetrahydrochloride substrate as described above. The standardprotocols from Cell Signaling Technology were used for the stainingprocedures.
Glucose tolerance tests
The Tg and WT mice were then fasted for 16 h and injected i.v. with D-glucose (1.2 mg/g body weight) in PBS. Blood samples from the tail veinwere collected at 15, 30, 60, 90, and 120 min after injection for glucosemeasurements with a glucose meter (Bayer, Toronto, ON, Canada).
Syngeneic islet transplantation
Diabetes was chemically induced by injecting C57BL/6 male mice (6–10wk old) with streptozotocin (STZ; 90 mg/kg) three times every other day.Three weeks after STZ injection, blood glucose was monitored daily withan Ascensia Contour glucose meter until it was above 20 mM for 2 con-secutive days. Three hundred Tg or WT islets of the C57BL/6 backgroundin 20 ml PBS were transplanted under the left renal capsule of these di-abetic mice. The surgical procedure is described elsewhere (35). Recipientblood glucose was monitored daily for 7 d and then once every 3 d untilday 30.
FAK siRNA transfection
Mouse FAK siRNA was synthesized by Integrated DNA Technologies.The sequences of the siRNA were rArGrArArArUrArGrCrUrGrArUrCrAr-ArGrU and rUrUrGrGrUrCrUrArCrUrUrGrArUrCrArGrC. The sequencesof the control siRNA were rCrUrUrCrC rUrCrUrCrU rUrUrCrUrC rUrCr-CrCrU rUrGrUGA and rUrCrArCrA rArGrGrGrA rGrArGrArA rArGr-ArGrA rGrGrArAr GrGrA. Paired siRNA was first heated to 94˚C for2 min and then cooled to room temperature for annealing. The annealedFAK siRNA was transfected into islet cells with X-tremeGENE trans-fection reagent (Roche Diagnostics GmbH) at a final concentration of 80nM siRNA for 100 islets/0.3 ml/well in 48-well plates. FAK mRNAknockdown was confirmed by RT/qPCR 48 h after transfection. The pri-mers for this purpose were 59-ACCATCCCTAACCATTGCCGAGAA-39and 59-ACTTTGGTATTGATGGCAACGCCC-39.
ResultsModulation of TGF-bi expression in islets after isolation
To identify molecules that are detrimental to or protective of isletsurvival after their isolation, microarray analysis was performed
Table I. Primers used in RT/qPCR
Gene Name 59 Primer 39 Primer
Fgg TGATAAGCATGCAGTCCACA GCATAGGTCAGGCGGTATTTRgs4 TCCTGCGAACACAGTTCTTC AACGATTCAGCCCATTTCTTSdpr TGTCCAGGTGAAGAGACTGG ACTGGCAGGGATCTCACTTTLy6a CTTCTTGTGGCCCTACTGTG TCAAATGGGACTCCATAGCAH2-Bf AGACTCTCAGGCCCATCTGT TTTGACATCCTTCACAGGGACmkor1 GAAGCCCTGAGGTCACTTG CCAGGCTCTGCATAGTCAAANrp1 GTGGCACAGGTGATGACTTC GTCGGGAACTCTGATTGGATTfpi2 TATACTGGCTGTGGTGGGAA GGCTTCTTCCAGCCTTTAACCrybb3 TGACCTGGTTCTCCTCCAGT ACACCGTCACCTTGTAGCTGTimp3 ACACGGAAGCCTCTGAAAGT GTCCCACCTCTCCACAAAGTSerpine1 GCCAACAAGAGCCAATCAC AAGCAAGGGCTGAAGACATCRgs5 ATTCTCCTCCAGAAGCCAGA CAGTGAGGGCTTGTGTGCCyp7b1 ACACAATGACCCGGAAATCT CTTCAGCCTCTTCCCTCCTT2210010C04Rik TCACTGCTACAAATCCCGAA GCATCAATGAATTGCTCACCCol5a2 TGGGTTTACAAGGACAGCAA CCAGGTTCACCATCTTCTCCGsdm1 TGCACAAGGAGAGGAAACTG CCACCTCCATCACCACATAGIfld2 AGCAACTGTGAGAGGACGAA GCTTGTCCCACAGAGAGTCALgals3 ATGCTGATCACAATCATGGG GCAACATCATTCCCTCTCCTMki67 TGAGGCTGAGACATGGAGAC TTCCATCCTCATGATTTCCALcn2 AAACAGAAGGCAGCTTTACGA TCTGATCCAGTAGCGACAGCCpb1 TGACCGTCCACTCCTACTCA ATGCAGAGTGGAAAGCTCCTGvin1 AGGCCACTAAGGACTCCAGA GCTGAGGCTCATCAGTGAAAPpap2b GTACAGGAGGCCAGGAAGTC GAGCAAAGTGAACTGCAGGA1810010M01Rik CCCTCTTAGTCCTTCTTTGTGC CTGGTTGCCAGAATGAGAGAD5Ertd593e CAAAGCAGAGCATCTTCGAG CAGGAACAAACTCTCTGGCASycn CTTTGGAGACTGGTCCGACT TGGGTAGACAGAGCCTCAAGHook1 GGGAGGTTGCTTCAGCTAAT CTCATCAGCTCCTGAATTGC1190002H23Rik AGCAACTTCTCCATGAAGCC GTCGAACTCGCACAGCACClps GTAAGAGCAGATGCTGCCAA GGACACCGGTAGTAGATCCCD14Ertd732e GCACTTTCCGGACTAGGTGT CAGATTTGTTTGGCGTCTTGCav CTCCGAGGGACATCTCTACAC CGCGTCATACACTTGCTTCTCd68 GAGGTTGTGACGGTACCCAT CGCCATGTAGTCCAGGTAGAGadd45a AAGACCGAAAGGATGGACAC GTCTACGTTGAGCAGCTTGGIgfbp5 AAGCTTCCCTCCAGGAGTTC GGAGGGCTTACACTGCTTTCZfp36 CCATCTACGAGAGCCTCCAG TATGTTCCAAAGTCCTCCGAHba-a1 AAGCCCTGGAAAGGATGTTT GACCTTCTTGCCGTGACCTgfbi TCATTTCCACCATCACCAAC AGCACGGTATTGAGTCCTGAVim ACCCTGCAGTCATTCAGACA TCCTGGATCTCTTCATCGTGCxcl1 CCGAAGTCATAGCCACACTC GTGCCATCAGAGCAGTCTGTGsdm1 TGCACAAGGAGAGGAAACTG CCACCTCCATCACCACATAG
The Journal of Immunology 5835
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
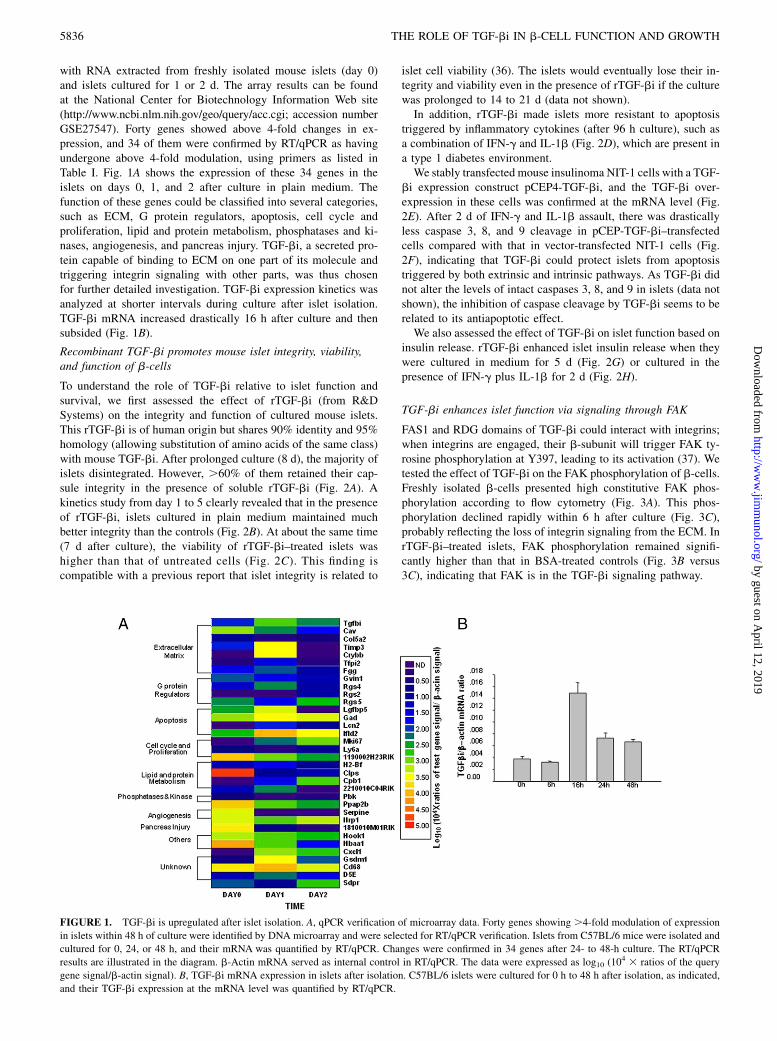
with RNA extracted from freshly isolated mouse islets (day 0)and islets cultured for 1 or 2 d. The array results can be foundat the National Center for Biotechnology Information Web site(http://www.ncbi.nlm.nih.gov/geo/query/acc.cgi; accession numberGSE27547). Forty genes showed above 4-fold changes in ex-pression, and 34 of them were confirmed by RT/qPCR as havingundergone above 4-fold modulation, using primers as listed inTable I. Fig. 1A shows the expression of these 34 genes in theislets on days 0, 1, and 2 after culture in plain medium. Thefunction of these genes could be classified into several categories,such as ECM, G protein regulators, apoptosis, cell cycle andproliferation, lipid and protein metabolism, phosphatases and ki-nases, angiogenesis, and pancreas injury. TGF-bi, a secreted pro-tein capable of binding to ECM on one part of its molecule andtriggering integrin signaling with other parts, was thus chosenfor further detailed investigation. TGF-bi expression kinetics wasanalyzed at shorter intervals during culture after islet isolation.TGF-bi mRNA increased drastically 16 h after culture and thensubsided (Fig. 1B).
Recombinant TGF-bi promotes mouse islet integrity, viability,and function of b-cells
To understand the role of TGF-bi relative to islet function andsurvival, we first assessed the effect of rTGF-bi (from R&DSystems) on the integrity and function of cultured mouse islets.This rTGF-bi is of human origin but shares 90% identity and 95%homology (allowing substitution of amino acids of the same class)with mouse TGF-bi. After prolonged culture (8 d), the majority ofislets disintegrated. However, .60% of them retained their cap-sule integrity in the presence of soluble rTGF-bi (Fig. 2A). Akinetics study from day 1 to 5 clearly revealed that in the presenceof rTGF-bi, islets cultured in plain medium maintained muchbetter integrity than the controls (Fig. 2B). At about the same time(7 d after culture), the viability of rTGF-bi–treated islets washigher than that of untreated cells (Fig. 2C). This finding iscompatible with a previous report that islet integrity is related to
islet cell viability (36). The islets would eventually lose their in-tegrity and viability even in the presence of rTGF-bi if the culturewas prolonged to 14 to 21 d (data not shown).In addition, rTGF-bi made islets more resistant to apoptosis
triggered by inflammatory cytokines (after 96 h culture), such asa combination of IFN-g and IL-1b (Fig. 2D), which are present ina type 1 diabetes environment.We stably transfected mouse insulinoma NIT-1 cells with a TGF-
bi expression construct pCEP4-TGF-bi, and the TGF-bi over-expression in these cells was confirmed at the mRNA level (Fig.2E). After 2 d of IFN-g and IL-1b assault, there was drasticallyless caspase 3, 8, and 9 cleavage in pCEP-TGF-bi–transfectedcells compared with that in vector-transfected NIT-1 cells (Fig.2F), indicating that TGF-bi could protect islets from apoptosistriggered by both extrinsic and intrinsic pathways. As TGF-bi didnot alter the levels of intact caspases 3, 8, and 9 in islets (data notshown), the inhibition of caspase cleavage by TGF-bi seems to berelated to its antiapoptotic effect.We also assessed the effect of TGF-bi on islet function based on
insulin release. rTGF-bi enhanced islet insulin release when theywere cultured in medium for 5 d (Fig. 2G) or cultured in thepresence of IFN-g plus IL-1b for 2 d (Fig. 2H).
TGF-bi enhances islet function via signaling through FAK
FAS1 and RDG domains of TGF-bi could interact with integrins;when integrins are engaged, their b-subunit will trigger FAK ty-rosine phosphorylation at Y397, leading to its activation (37). Wetested the effect of TGF-bi on the FAK phosphorylation of b-cells.Freshly isolated b-cells presented high constitutive FAK phos-phorylation according to flow cytometry (Fig. 3A). This phos-phorylation declined rapidly within 6 h after culture (Fig. 3C),probably reflecting the loss of integrin signaling from the ECM. InrTGF-bi–treated islets, FAK phosphorylation remained signifi-cantly higher than that in BSA-treated controls (Fig. 3B versus3C), indicating that FAK is in the TGF-bi signaling pathway.
FIGURE 1. TGF-bi is upregulated after islet isolation. A, qPCR verification of microarray data. Forty genes showing .4-fold modulation of expression
in islets within 48 h of culture were identified by DNA microarray and were selected for RT/qPCR verification. Islets from C57BL/6 mice were isolated and
cultured for 0, 24, or 48 h, and their mRNA was quantified by RT/qPCR. Changes were confirmed in 34 genes after 24- to 48-h culture. The RT/qPCR
results are illustrated in the diagram. b-Actin mRNA served as internal control in RT/qPCR. The data were expressed as log10 (104 3 ratios of the query
gene signal/b-actin signal). B, TGF-bi mRNA expression in islets after isolation. C57BL/6 islets were cultured for 0 h to 48 h after isolation, as indicated,
and their TGF-bi expression at the mRNA level was quantified by RT/qPCR.
5836 THE ROLE OF TGF-bi IN b-CELL FUNCTION AND GROWTH
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
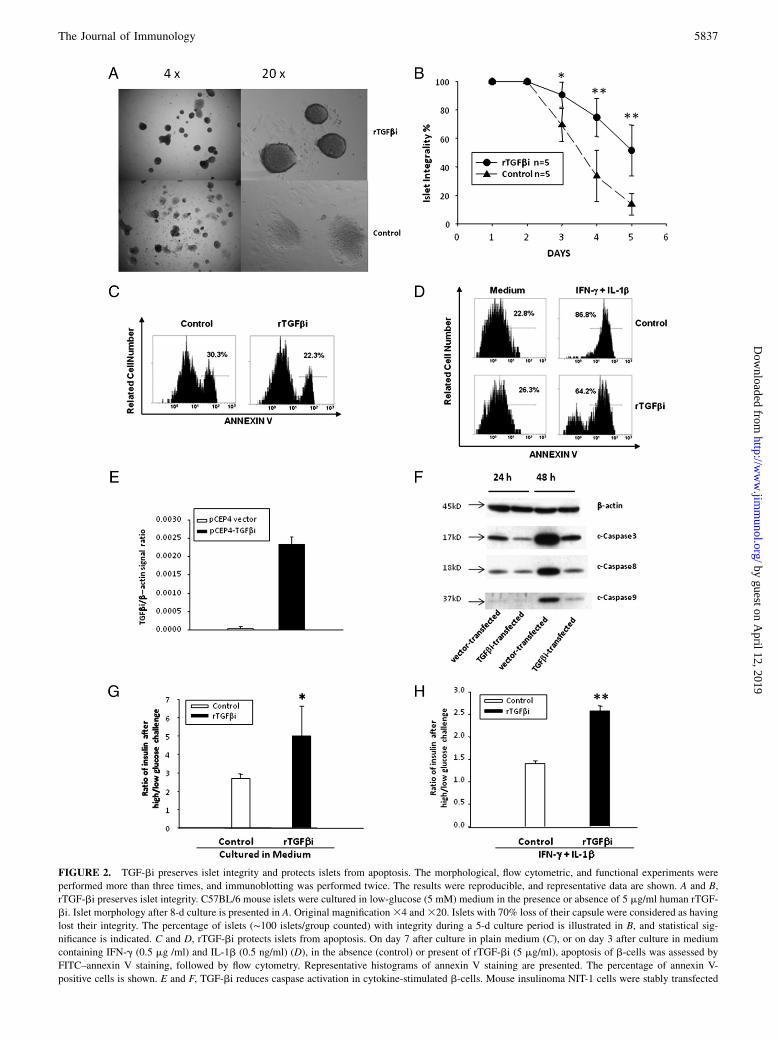
FIGURE 2. TGF-bi preserves islet integrity and protects islets from apoptosis. The morphological, flow cytometric, and functional experiments were
performed more than three times, and immunoblotting was performed twice. The results were reproducible, and representative data are shown. A and B,
rTGF-bi preserves islet integrity. C57BL/6 mouse islets were cultured in low-glucose (5 mM) medium in the presence or absence of 5 mg/ml human rTGF-
bi. Islet morphology after 8-d culture is presented in A. Original magnification34 and320. Islets with 70% loss of their capsule were considered as having
lost their integrity. The percentage of islets (∼100 islets/group counted) with integrity during a 5-d culture period is illustrated in B, and statistical sig-
nificance is indicated. C and D, rTGF-bi protects islets from apoptosis. On day 7 after culture in plain medium (C), or on day 3 after culture in medium
containing IFN-g (0.5 mg /ml) and IL-1b (0.5 ng/ml) (D), in the absence (control) or present of rTGF-bi (5 mg/ml), apoptosis of b-cells was assessed by
FITC–annexin V staining, followed by flow cytometry. Representative histograms of annexin V staining are presented. The percentage of annexin V-
positive cells is shown. E and F, TGF-bi reduces caspase activation in cytokine-stimulated b-cells. Mouse insulinoma NIT-1 cells were stably transfected
The Journal of Immunology 5837
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
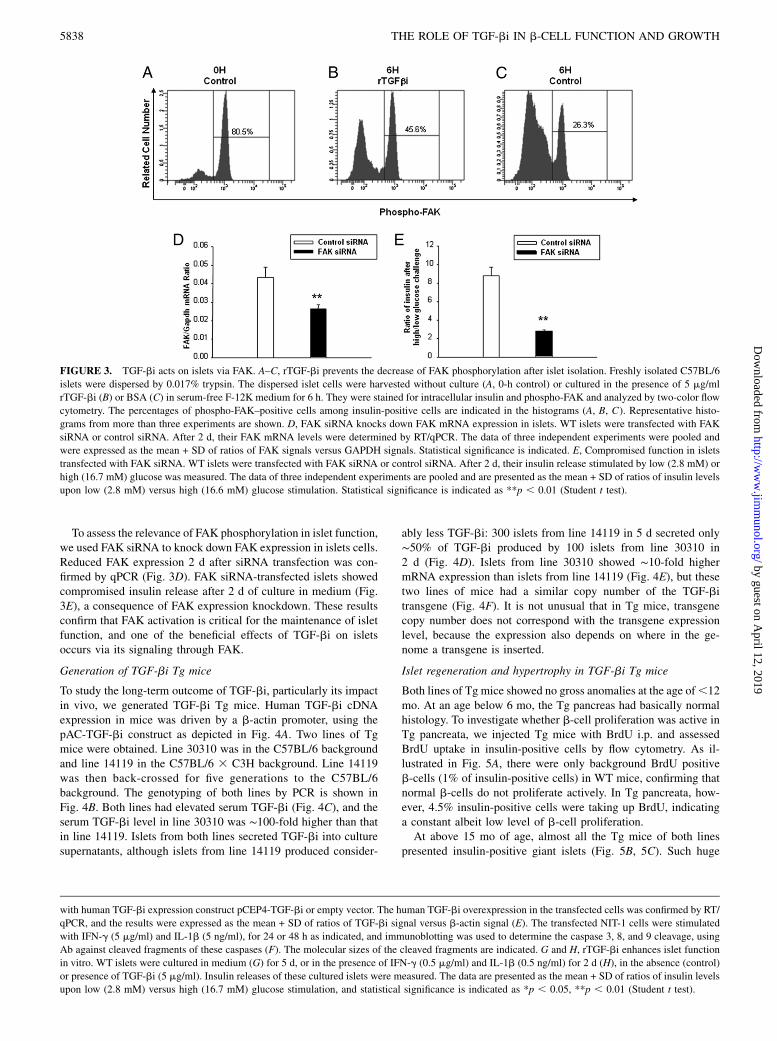
To assess the relevance of FAK phosphorylation in islet function,we used FAK siRNA to knock down FAK expression in islets cells.Reduced FAK expression 2 d after siRNA transfection was con-firmed by qPCR (Fig. 3D). FAK siRNA-transfected islets showedcompromised insulin release after 2 d of culture in medium (Fig.3E), a consequence of FAK expression knockdown. These resultsconfirm that FAK activation is critical for the maintenance of isletfunction, and one of the beneficial effects of TGF-bi on isletsoccurs via its signaling through FAK.
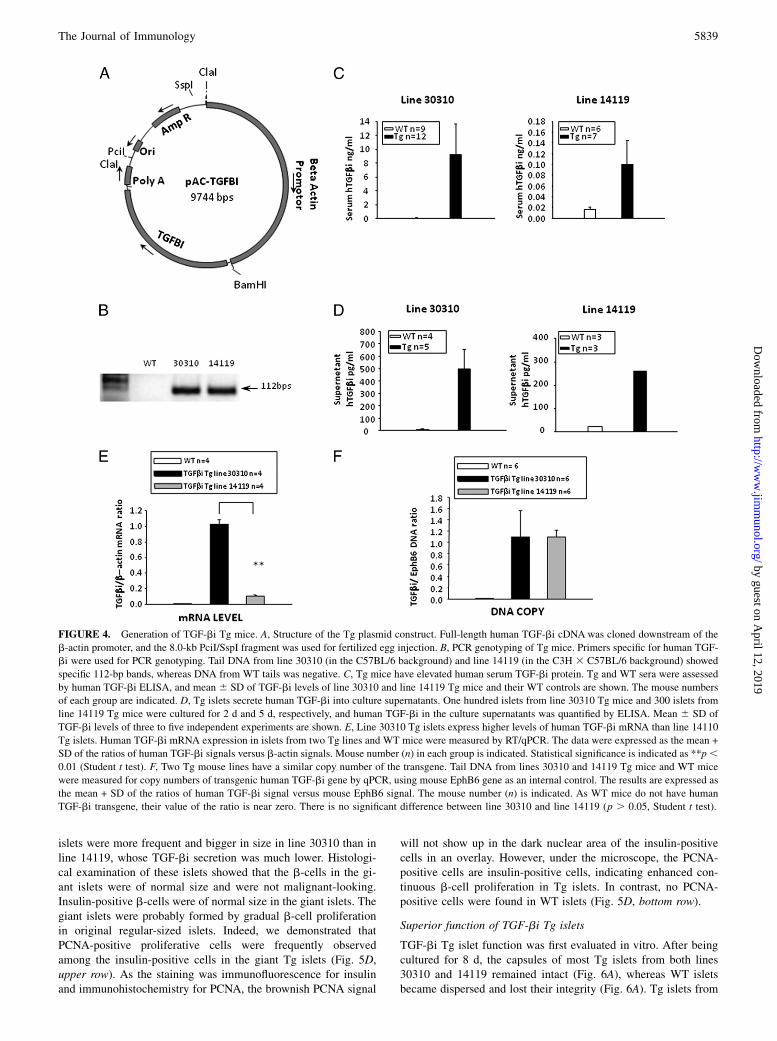
Generation of TGF-bi Tg mice
To study the long-term outcome of TGF-bi, particularly its impactin vivo, we generated TGF-bi Tg mice. Human TGF-bi cDNAexpression in mice was driven by a b-actin promoter, using thepAC-TGF-bi construct as depicted in Fig. 4A. Two lines of Tgmice were obtained. Line 30310 was in the C57BL/6 backgroundand line 14119 in the C57BL/6 3 C3H background. Line 14119was then back-crossed for five generations to the C57BL/6background. The genotyping of both lines by PCR is shown inFig. 4B. Both lines had elevated serum TGF-bi (Fig. 4C), and theserum TGF-bi level in line 30310 was ∼100-fold higher than thatin line 14119. Islets from both lines secreted TGF-bi into culturesupernatants, although islets from line 14119 produced consider-
ably less TGF-bi: 300 islets from line 14119 in 5 d secreted only∼50% of TGF-bi produced by 100 islets from line 30310 in2 d (Fig. 4D). Islets from line 30310 showed ∼10-fold highermRNA expression than islets from line 14119 (Fig. 4E), but thesetwo lines of mice had a similar copy number of the TGF-bitransgene (Fig. 4F). It is not unusual that in Tg mice, transgenecopy number does not correspond with the transgene expressionlevel, because the expression also depends on where in the ge-nome a transgene is inserted.
Islet regeneration and hypertrophy in TGF-bi Tg mice
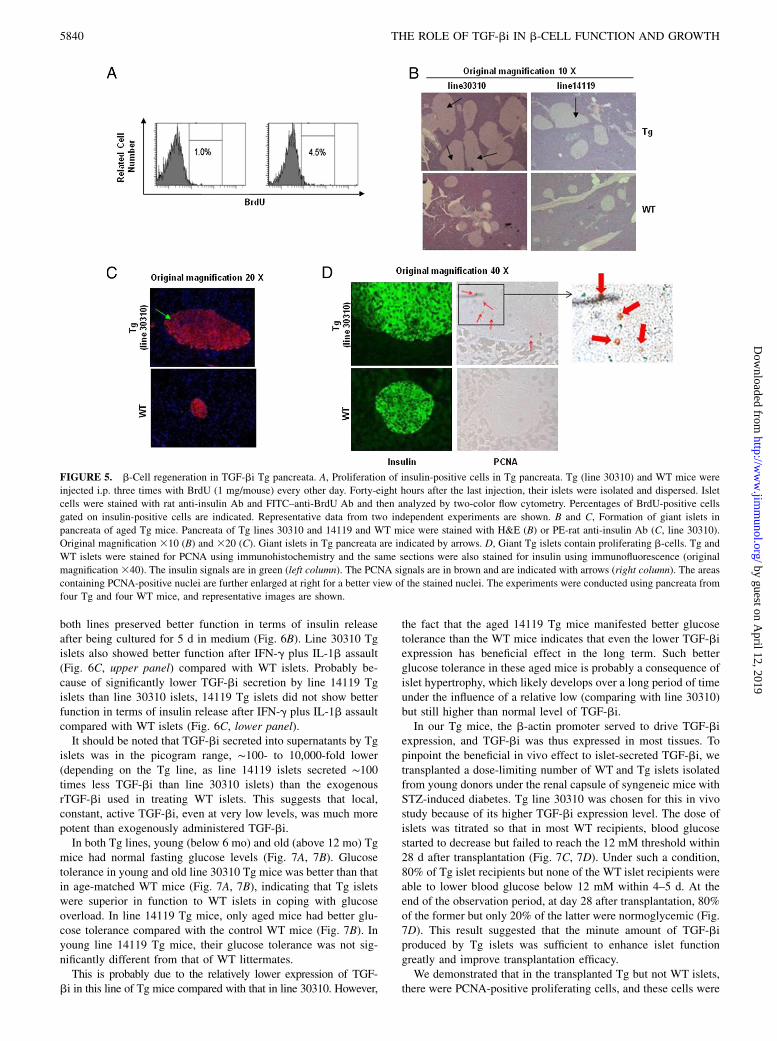
Both lines of Tg mice showed no gross anomalies at the age of,12mo. At an age below 6 mo, the Tg pancreas had basically normalhistology. To investigate whether b-cell proliferation was active inTg pancreata, we injected Tg mice with BrdU i.p. and assessedBrdU uptake in insulin-positive cells by flow cytometry. As il-lustrated in Fig. 5A, there were only background BrdU positiveb-cells (1% of insulin-positive cells) in WT mice, confirming thatnormal b-cells do not proliferate actively. In Tg pancreata, how-ever, 4.5% insulin-positive cells were taking up BrdU, indicatinga constant albeit low level of b-cell proliferation.At above 15 mo of age, almost all the Tg mice of both lines
presented insulin-positive giant islets (Fig. 5B, 5C). Such huge
with human TGF-bi expression construct pCEP4-TGF-bi or empty vector. The human TGF-bi overexpression in the transfected cells was confirmed by RT/
qPCR, and the results were expressed as the mean + SD of ratios of TGF-bi signal versus b-actin signal (E). The transfected NIT-1 cells were stimulated
with IFN-g (5 mg/ml) and IL-1b (5 ng/ml), for 24 or 48 h as indicated, and immunoblotting was used to determine the caspase 3, 8, and 9 cleavage, using
Ab against cleaved fragments of these caspases (F). The molecular sizes of the cleaved fragments are indicated. G and H, rTGF-bi enhances islet function
in vitro. WT islets were cultured in medium (G) for 5 d, or in the presence of IFN-g (0.5 mg/ml) and IL-1b (0.5 ng/ml) for 2 d (H), in the absence (control)
or presence of TGF-bi (5 mg/ml). Insulin releases of these cultured islets were measured. The data are presented as the mean + SD of ratios of insulin levels
upon low (2.8 mM) versus high (16.7 mM) glucose stimulation, and statistical significance is indicated as *p , 0.05, **p , 0.01 (Student t test).
FIGURE 3. TGF-bi acts on islets via FAK. A–C, rTGF-bi prevents the decrease of FAK phosphorylation after islet isolation. Freshly isolated C57BL/6
islets were dispersed by 0.017% trypsin. The dispersed islet cells were harvested without culture (A, 0-h control) or cultured in the presence of 5 mg/ml
rTGF-bi (B) or BSA (C) in serum-free F-12K medium for 6 h. They were stained for intracellular insulin and phospho-FAK and analyzed by two-color flow
cytometry. The percentages of phospho-FAK–positive cells among insulin-positive cells are indicated in the histograms (A, B, C). Representative histo-
grams from more than three experiments are shown. D, FAK siRNA knocks down FAK mRNA expression in islets. WT islets were transfected with FAK
siRNA or control siRNA. After 2 d, their FAK mRNA levels were determined by RT/qPCR. The data of three independent experiments were pooled and
were expressed as the mean + SD of ratios of FAK signals versus GAPDH signals. Statistical significance is indicated. E, Compromised function in islets
transfected with FAK siRNA. WT islets were transfected with FAK siRNA or control siRNA. After 2 d, their insulin release stimulated by low (2.8 mM) or
high (16.7 mM) glucose was measured. The data of three independent experiments are pooled and are presented as the mean + SD of ratios of insulin levels
upon low (2.8 mM) versus high (16.6 mM) glucose stimulation. Statistical significance is indicated as **p , 0.01 (Student t test).
5838 THE ROLE OF TGF-bi IN b-CELL FUNCTION AND GROWTH
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
islets were more frequent and bigger in size in line 30310 than inline 14119, whose TGF-bi secretion was much lower. Histologi-cal examination of these islets showed that the b-cells in the gi-ant islets were of normal size and were not malignant-looking.Insulin-positive b-cells were of normal size in the giant islets. Thegiant islets were probably formed by gradual b-cell proliferationin original regular-sized islets. Indeed, we demonstrated thatPCNA-positive proliferative cells were frequently observedamong the insulin-positive cells in the giant Tg islets (Fig. 5D,upper row). As the staining was immunofluorescence for insulinand immunohistochemistry for PCNA, the brownish PCNA signal
will not show up in the dark nuclear area of the insulin-positivecells in an overlay. However, under the microscope, the PCNA-positive cells are insulin-positive cells, indicating enhanced con-tinuous b-cell proliferation in Tg islets. In contrast, no PCNA-positive cells were found in WT islets (Fig. 5D, bottom row).
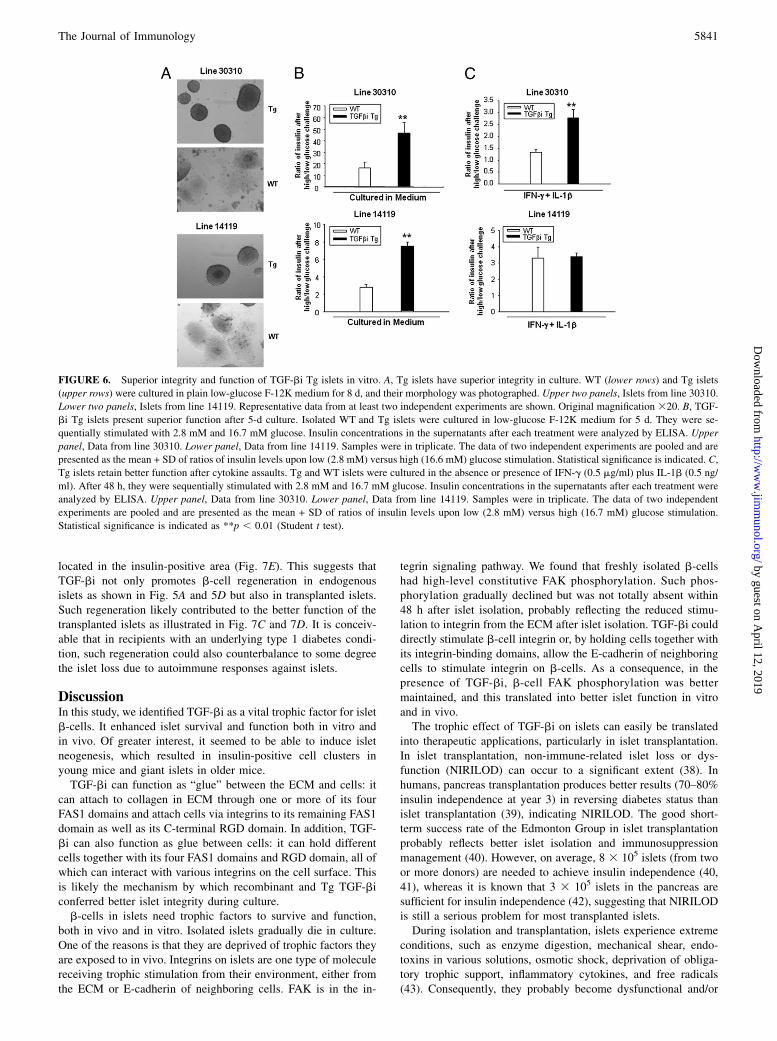
Superior function of TGF-bi Tg islets
TGF-bi Tg islet function was first evaluated in vitro. After beingcultured for 8 d, the capsules of most Tg islets from both lines30310 and 14119 remained intact (Fig. 6A), whereas WT isletsbecame dispersed and lost their integrity (Fig. 6A). Tg islets from
FIGURE 4. Generation of TGF-bi Tg mice. A, Structure of the Tg plasmid construct. Full-length human TGF-bi cDNA was cloned downstream of the
b-actin promoter, and the 8.0-kb PciI/SspI fragment was used for fertilized egg injection. B, PCR genotyping of Tg mice. Primers specific for human TGF-
bi were used for PCR genotyping. Tail DNA from line 30310 (in the C57BL/6 background) and line 14119 (in the C3H 3 C57BL/6 background) showed
specific 112-bp bands, whereas DNA from WT tails was negative. C, Tg mice have elevated human serum TGF-bi protein. Tg and WT sera were assessed
by human TGF-bi ELISA, and mean 6 SD of TGF-bi levels of line 30310 and line 14119 Tg mice and their WT controls are shown. The mouse numbers
of each group are indicated. D, Tg islets secrete human TGF-bi into culture supernatants. One hundred islets from line 30310 Tg mice and 300 islets from
line 14119 Tg mice were cultured for 2 d and 5 d, respectively, and human TGF-bi in the culture supernatants was quantified by ELISA. Mean 6 SD of
TGF-bi levels of three to five independent experiments are shown. E, Line 30310 Tg islets express higher levels of human TGF-bi mRNA than line 14110
Tg islets. Human TGF-bi mRNA expression in islets from two Tg lines and WT mice were measured by RT/qPCR. The data were expressed as the mean +
SD of the ratios of human TGF-bi signals versus b-actin signals. Mouse number (n) in each group is indicated. Statistical significance is indicated as **p,0.01 (Student t test). F, Two Tg mouse lines have a similar copy number of the transgene. Tail DNA from lines 30310 and 14119 Tg mice and WT mice
were measured for copy numbers of transgenic human TGF-bi gene by qPCR, using mouse EphB6 gene as an internal control. The results are expressed as
the mean + SD of the ratios of human TGF-bi signal versus mouse EphB6 signal. The mouse number (n) is indicated. As WT mice do not have human
TGF-bi transgene, their value of the ratio is near zero. There is no significant difference between line 30310 and line 14119 (p . 0.05, Student t test).
The Journal of Immunology 5839
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
both lines preserved better function in terms of insulin releaseafter being cultured for 5 d in medium (Fig. 6B). Line 30310 Tgislets also showed better function after IFN-g plus IL-1b assault(Fig. 6C, upper panel) compared with WT islets. Probably be-cause of significantly lower TGF-bi secretion by line 14119 Tgislets than line 30310 islets, 14119 Tg islets did not show betterfunction in terms of insulin release after IFN-g plus IL-1b assaultcompared with WT islets (Fig. 6C, lower panel).It should be noted that TGF-bi secreted into supernatants by Tg
islets was in the picogram range, ∼100- to 10,000-fold lower(depending on the Tg line, as line 14119 islets secreted ∼100times less TGF-bi than line 30310 islets) than the exogenousrTGF-bi used in treating WT islets. This suggests that local,constant, active TGF-bi, even at very low levels, was much morepotent than exogenously administered TGF-bi.In both Tg lines, young (below 6 mo) and old (above 12 mo) Tg
mice had normal fasting glucose levels (Fig. 7A, 7B). Glucosetolerance in young and old line 30310 Tg mice was better than thatin age-matched WT mice (Fig. 7A, 7B), indicating that Tg isletswere superior in function to WT islets in coping with glucoseoverload. In line 14119 Tg mice, only aged mice had better glu-cose tolerance compared with the control WT mice (Fig. 7B). Inyoung line 14119 Tg mice, their glucose tolerance was not sig-nificantly different from that of WT littermates.This is probably due to the relatively lower expression of TGF-
bi in this line of Tg mice compared with that in line 30310. However,
the fact that the aged 14119 Tg mice manifested better glucosetolerance than the WT mice indicates that even the lower TGF-biexpression has beneficial effect in the long term. Such betterglucose tolerance in these aged mice is probably a consequence ofislet hypertrophy, which likely develops over a long period of timeunder the influence of a relative low (comparing with line 30310)but still higher than normal level of TGF-bi.In our Tg mice, the b-actin promoter served to drive TGF-bi
expression, and TGF-bi was thus expressed in most tissues. Topinpoint the beneficial in vivo effect to islet-secreted TGF-bi, wetransplanted a dose-limiting number of WT and Tg islets isolatedfrom young donors under the renal capsule of syngeneic mice withSTZ-induced diabetes. Tg line 30310 was chosen for this in vivostudy because of its higher TGF-bi expression level. The dose ofislets was titrated so that in most WT recipients, blood glucosestarted to decrease but failed to reach the 12 mM threshold within28 d after transplantation (Fig. 7C, 7D). Under such a condition,80% of Tg islet recipients but none of the WT islet recipients wereable to lower blood glucose below 12 mM within 4–5 d. At theend of the observation period, at day 28 after transplantation, 80%of the former but only 20% of the latter were normoglycemic (Fig.7D). This result suggested that the minute amount of TGF-biproduced by Tg islets was sufficient to enhance islet functiongreatly and improve transplantation efficacy.We demonstrated that in the transplanted Tg but not WT islets,
there were PCNA-positive proliferating cells, and these cells were
FIGURE 5. b-Cell regeneration in TGF-bi Tg pancreata. A, Proliferation of insulin-positive cells in Tg pancreata. Tg (line 30310) and WT mice were
injected i.p. three times with BrdU (1 mg/mouse) every other day. Forty-eight hours after the last injection, their islets were isolated and dispersed. Islet
cells were stained with rat anti-insulin Ab and FITC–anti-BrdU Ab and then analyzed by two-color flow cytometry. Percentages of BrdU-positive cells
gated on insulin-positive cells are indicated. Representative data from two independent experiments are shown. B and C, Formation of giant islets in
pancreata of aged Tg mice. Pancreata of Tg lines 30310 and 14119 and WT mice were stained with H&E (B) or PE-rat anti-insulin Ab (C, line 30310).
Original magnification 310 (B) and 320 (C). Giant islets in Tg pancreata are indicated by arrows. D, Giant Tg islets contain proliferating b-cells. Tg and
WT islets were stained for PCNA using immunohistochemistry and the same sections were also stained for insulin using immunofluorescence (original
magnification 340). The insulin signals are in green (left column). The PCNA signals are in brown and are indicated with arrows (right column). The areas
containing PCNA-positive nuclei are further enlarged at right for a better view of the stained nuclei. The experiments were conducted using pancreata from
four Tg and four WT mice, and representative images are shown.
5840 THE ROLE OF TGF-bi IN b-CELL FUNCTION AND GROWTH
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
located in the insulin-positive area (Fig. 7E). This suggests thatTGF-bi not only promotes b-cell regeneration in endogenousislets as shown in Fig. 5A and 5D but also in transplanted islets.Such regeneration likely contributed to the better function of thetransplanted islets as illustrated in Fig. 7C and 7D. It is conceiv-able that in recipients with an underlying type 1 diabetes condi-tion, such regeneration could also counterbalance to some degreethe islet loss due to autoimmune responses against islets.
DiscussionIn this study, we identified TGF-bi as a vital trophic factor for isletb-cells. It enhanced islet survival and function both in vitro andin vivo. Of greater interest, it seemed to be able to induce isletneogenesis, which resulted in insulin-positive cell clusters inyoung mice and giant islets in older mice.TGF-bi can function as “glue” between the ECM and cells: it
can attach to collagen in ECM through one or more of its fourFAS1 domains and attach cells via integrins to its remaining FAS1domain as well as its C-terminal RGD domain. In addition, TGF-bi can also function as glue between cells: it can hold differentcells together with its four FAS1 domains and RGD domain, all ofwhich can interact with various integrins on the cell surface. Thisis likely the mechanism by which recombinant and Tg TGF-biconferred better islet integrity during culture.b-cells in islets need trophic factors to survive and function,
both in vivo and in vitro. Isolated islets gradually die in culture.One of the reasons is that they are deprived of trophic factors theyare exposed to in vivo. Integrins on islets are one type of moleculereceiving trophic stimulation from their environment, either fromthe ECM or E-cadherin of neighboring cells. FAK is in the in-
tegrin signaling pathway. We found that freshly isolated b-cellshad high-level constitutive FAK phosphorylation. Such phos-phorylation gradually declined but was not totally absent within48 h after islet isolation, probably reflecting the reduced stimu-lation to integrin from the ECM after islet isolation. TGF-bi coulddirectly stimulate b-cell integrin or, by holding cells together withits integrin-binding domains, allow the E-cadherin of neighboringcells to stimulate integrin on b-cells. As a consequence, in thepresence of TGF-bi, b-cell FAK phosphorylation was bettermaintained, and this translated into better islet function in vitroand in vivo.The trophic effect of TGF-bi on islets can easily be translated
into therapeutic applications, particularly in islet transplantation.In islet transplantation, non-immune-related islet loss or dys-function (NIRILOD) can occur to a significant extent (38). Inhumans, pancreas transplantation produces better results (70–80%insulin independence at year 3) in reversing diabetes status thanislet transplantation (39), indicating NIRILOD. The good short-term success rate of the Edmonton Group in islet transplantationprobably reflects better islet isolation and immunosuppressionmanagement (40). However, on average, 8 3 105 islets (from twoor more donors) are needed to achieve insulin independence (40,41), whereas it is known that 3 3 105 islets in the pancreas aresufficient for insulin independence (42), suggesting that NIRILODis still a serious problem for most transplanted islets.During isolation and transplantation, islets experience extreme
conditions, such as enzyme digestion, mechanical shear, endo-toxins in various solutions, osmotic shock, deprivation of obliga-tory trophic support, inflammatory cytokines, and free radicals(43). Consequently, they probably become dysfunctional and/or
FIGURE 6. Superior integrity and function of TGF-bi Tg islets in vitro. A, Tg islets have superior integrity in culture. WT (lower rows) and Tg islets
(upper rows) were cultured in plain low-glucose F-12K medium for 8 d, and their morphology was photographed. Upper two panels, Islets from line 30310.
Lower two panels, Islets from line 14119. Representative data from at least two independent experiments are shown. Original magnification 320. B, TGF-
bi Tg islets present superior function after 5-d culture. Isolated WT and Tg islets were cultured in low-glucose F-12K medium for 5 d. They were se-
quentially stimulated with 2.8 mM and 16.7 mM glucose. Insulin concentrations in the supernatants after each treatment were analyzed by ELISA. Upper
panel, Data from line 30310. Lower panel, Data from line 14119. Samples were in triplicate. The data of two independent experiments are pooled and are
presented as the mean + SD of ratios of insulin levels upon low (2.8 mM) versus high (16.6 mM) glucose stimulation. Statistical significance is indicated. C,
Tg islets retain better function after cytokine assaults. Tg and WT islets were cultured in the absence or presence of IFN-g (0.5 mg/ml) plus IL-1b (0.5 ng/
ml). After 48 h, they were sequentially stimulated with 2.8 mM and 16.7 mM glucose. Insulin concentrations in the supernatants after each treatment were
analyzed by ELISA. Upper panel, Data from line 30310. Lower panel, Data from line 14119. Samples were in triplicate. The data of two independent
experiments are pooled and are presented as the mean + SD of ratios of insulin levels upon low (2.8 mM) versus high (16.7 mM) glucose stimulation.
Statistical significance is indicated as **p , 0.01 (Student t test).
The Journal of Immunology 5841
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
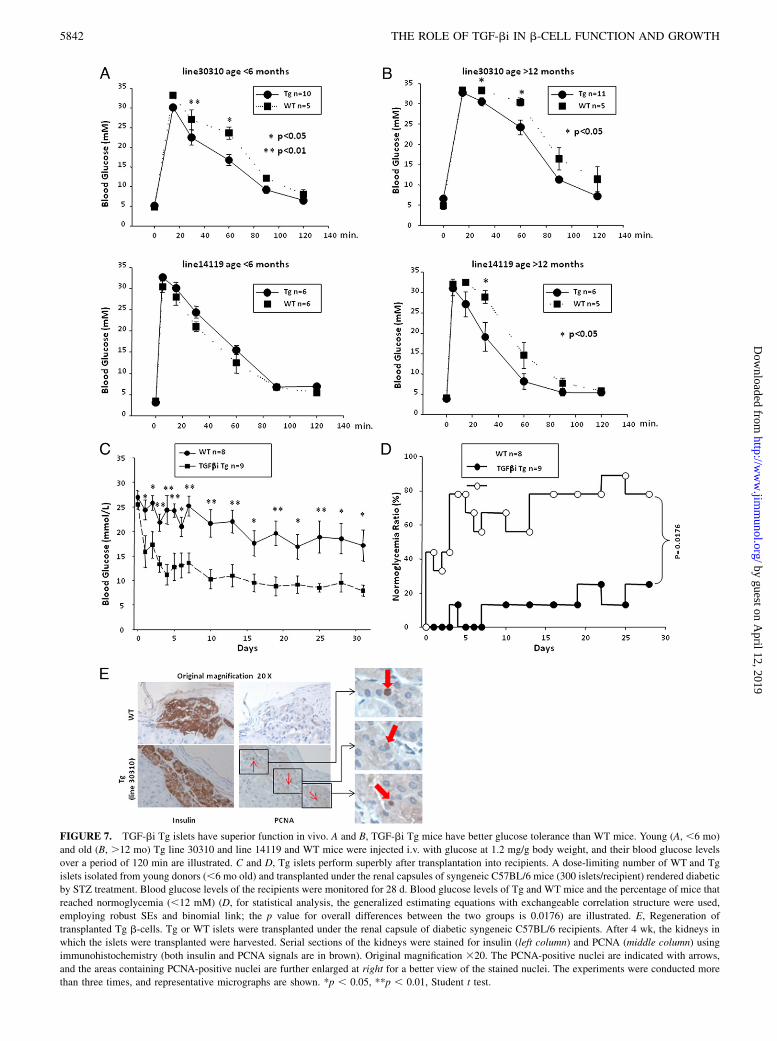
FIGURE 7. TGF-bi Tg islets have superior function in vivo. A and B, TGF-bi Tg mice have better glucose tolerance than WT mice. Young (A, ,6 mo)
and old (B, .12 mo) Tg line 30310 and line 14119 and WT mice were injected i.v. with glucose at 1.2 mg/g body weight, and their blood glucose levels
over a period of 120 min are illustrated. C and D, Tg islets perform superbly after transplantation into recipients. A dose-limiting number of WT and Tg
islets isolated from young donors (,6 mo old) and transplanted under the renal capsules of syngeneic C57BL/6 mice (300 islets/recipient) rendered diabetic
by STZ treatment. Blood glucose levels of the recipients were monitored for 28 d. Blood glucose levels of Tg and WT mice and the percentage of mice that
reached normoglycemia (,12 mM) (D, for statistical analysis, the generalized estimating equations with exchangeable correlation structure were used,
employing robust SEs and binomial link; the p value for overall differences between the two groups is 0.0176) are illustrated. E, Regeneration of
transplanted Tg b-cells. Tg or WT islets were transplanted under the renal capsule of diabetic syngeneic C57BL/6 recipients. After 4 wk, the kidneys in
which the islets were transplanted were harvested. Serial sections of the kidneys were stained for insulin (left column) and PCNA (middle column) using
immunohistochemistry (both insulin and PCNA signals are in brown). Original magnification 320. The PCNA-positive nuclei are indicated with arrows,
and the areas containing PCNA-positive nuclei are further enlarged at right for a better view of the stained nuclei. The experiments were conducted more
than three times, and representative micrographs are shown. *p , 0.05, **p , 0.01, Student t test.
5842 THE ROLE OF TGF-bi IN b-CELL FUNCTION AND GROWTH
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

undergo substantial apoptosis in culture (44, 45). Upon trans-plantation into diabetic patients, they immediately face unfavor-able conditions, such as high serum lipid and glucose levels,which are now believed to be toxic to b-cells, at least according tosome researchers (46, 47), as well as inflammatory cytokines re-leased by local inflammatory responses. These in vivo factorscompound the gravity of islet dysfunction and apoptosis. Con-sistent with this notion, immune-isolated, microencapsulated isletsinjected in the dog peritoneal cavity fail within 6 mo (48), andislets placed into vascularized, biohybrid pancreas fail at 1 y (49).Histological evaluation reveals a major loss of islet mass in thesedevices (48, 49), demonstrating the destruction of islets in theabsence of cellular immune responses. Therefore, NIRILOD islikely one of the major factors contributing to low islet trans-plantation efficiency.Our study of TGF-bi indicates that it could replenish some of
the lost trophic support in isolated islets and render to them sur-vival advantage and resistance to inflammatory cytokine assaults.This finding could be easily translated into clinical applications.Isolated islets could be incubated with TGF-bi for 24 h beforetheir transplantation. Even better, a suitable local TGF-bi ex-pression system, such as lentivirus, could be used to allow isolatedislets to express TGF-bi locally and continuously for a longer termafter transplantation. Both ways should be able to significantlyincrease islet transplantation efficacy, as we have demonstratedwith TGF-bi–expressing transplanted Tg islets. As describedabove, NIRILOD is caused by multiple factors. TGF-bi treatmentwill likely ameliorate but not eliminate NIRILOD. A combinationof several approaches, in addition to providing trophic support,will be needed to reduce NIRILOD to negligible levels.The de novo formation of insulin-positive cell clusters and later
huge islets in Tg mice is extremely interesting. It has raised thepossibility of regenerating islets in type 1 diabetes and in the latestage of type 2 diabetes as a way of delaying the onset or evenreversing the progress of diabetes. Of course, to achieve sucha goal, suitable, long-term TGF-bi overexpression locally in thepancreas will need to be developed, and any potential TGF-bi–caused tumor risk should be excluded. Although TGF-bi can in-duce epithelial cells to mesenchymal cell transition, which is in-volved in fibrosis in some chronic diseases (50), there is no reportto date documenting any role of TGF-bi in fibrosis. Indeed, our Tgmice, even the aged ones, manifested no signs of fibrosis in anyorgans (data not shown). Therefore, the role of TGF-bi in fibrosisdoes not seem to act via TGF-bi, and this relieves another concernfor the clinical application of TGF-bi in the future.
DisclosuresThe authors have no financial conflicts of interest.
References1. Bosco, D., L. Orci, and P. Meda. 1989. Homologous but not heterologous con-
tact increases the insulin secretion of individual pancreatic B-cells. Exp. CellRes. 184: 72–80.
2. Halban, P. A., S. L. Powers, K. L. George, and S. Bonner-Weir. 1987. Spontaneousreassociation of dispersed adult rat pancreatic islet cells into aggregates with three-dimensional architecture typical of native islets. Diabetes 36: 783–790.
3. Hopcroft, D. W., D. R. Mason, and R. S. Scott. 1985. Structure-function rela-tionships in pancreatic islets: support for intraislet modulation of insulin secre-tion. Endocrinology 117: 2073–2080.
4. Pinkse, G. G., W. P. Bouwman, R. Jiawan-Lalai, O. T. Terpstra, J. A. Bruijn, andE. de Heer. 2006. Integrin signaling via RGD peptides and anti-beta1 antibodiesconfers resistance to apoptosis in islets of Langerhans. Diabetes 55: 312–317.
5. Matter, M. L., and E. Ruoslahti. 2001. A signaling pathway from the alpha5beta1and alpha(v)beta3 integrins that elevates bcl-2 transcription. J. Biol. Chem. 276:27757–27763.
6. Zhang, Z., K. Vuori, J. C. Reed, and E. Ruoslahti. 1995. The alpha 5 beta 1integrin supports survival of cells on fibronectin and up-regulates Bcl-2 ex-pression. Proc. Natl. Acad. Sci. USA 92: 6161–6165.
7. Kaido, T., M. Yebra, V. Cirulli, and A. M. Montgomery. 2004. Regulation ofhuman beta-cell adhesion, motility, and insulin secretion by collagen IV and itsreceptor alpha1beta1. J. Biol. Chem. 279: 53762–53769.
8. Wang, R. N., and L. Rosenberg. 1999. Maintenance of beta-cell function andsurvival following islet isolation requires re-establishment of the islet-matrixrelationship. J. Endocrinol. 163: 181–190.
9. Wang, R. N., S. Paraskevas, and L. Rosenberg. 1999. Characterization of integrinexpression in islets isolated from hamster, canine, porcine, and human pancreas.J. Histochem. Cytochem. 47: 499–506.
10. Hauge-Evans, A. C., P. E. Squires, S. J. Persaud, and P. M. Jones. 1999. Pan-creatic beta-cell-to-beta-cell interactions are required for integrated responses tonutrient stimuli: enhanced Ca2+ and insulin secretory responses of MIN6pseudoislets. Diabetes 48: 1402–1408.
11. Cirulli, V., D. Baetens, U. Rutishauser, P. A. Halban, L. Orci, and D. G. Rouiller.1994. Expression of neural cell adhesion molecule (N-CAM) in rat islets and itsrole in islet cell type segregation. J. Cell Sci. 107: 1429–1436.
12. Bernard-Kargar, C., N. Kassis, M. F. Berthault, W. Pralong, and A. Ktorza. 2001.Sialylated form of the neural cell adhesion molecule (NCAM): a new tool for theidentification and sorting of beta-cell subpopulations with different functionalactivity. Diabetes 50(Suppl 1): S125–S130.
13. Dahl, U., A. Sjødin, and H. Semb. 1996. Cadherins regulate aggregation ofpancreatic beta-cells in vivo. Development 122: 2895–2902.
14. Skonier, J., M. Neubauer, L. Madisen, K. Bennett, G. D. Plowman, andA. F. Purchio. 1992. cDNA cloning and sequence analysis of beta ig-h3, a novelgene induced in a human adenocarcinoma cell line after treatment with trans-forming growth factor-beta. DNA Cell Biol. 11: 511–522.
15. Skonier, J., K. Bennett, V. Rothwell, S. Kosowski, G. Plowman, P. Wallace,S. Edelhoff, C. Disteche, M. Neubauer, H. Marquardt, et al. 1994. beta ig-h3:a transforming growth factor-beta-responsive gene encoding a secreted proteinthat inhibits cell attachment in vitro and suppresses the growth of CHO cells innude mice. DNA Cell Biol. 13: 571–584.
16. Billings, P. C., D. J. Herrick, U. Kucich, B. N. Engelsberg, W. R. Abrams,E. J. Macarak, J. Rosenbloom, and P. S. Howard. 2000. Extracellular matrix andnuclear localization of beta ig-h3 in human bladder smooth muscle and fibroblastcells. J. Cell. Biochem. 79: 261–273.
17. Park, S. W., J. S. Bae, K. S. Kim, S. H. Park, B. H. Lee, J. Y. Choi, J. Y. Park,S. W. Ha, Y. L. Kim, T. H. Kwon, et al. 2004. Beta ig-h3 promotes renalproximal tubular epithelial cell adhesion, migration and proliferation through theinteraction with alpha3beta1 integrin. Exp. Mol. Med. 36: 211–219.
18. Gilbert, R. E., J. L. Wilkinson-Berka, D. W. Johnson, A. Cox, T. Soulis,L. L. Wu, D. J. Kelly, G. Jerums, C. A. Pollock, and M. E. Cooper. 1998. Renalexpression of transforming growth factor-beta inducible gene-h3 (beta ig-h3) innormal and diabetic rats. Kidney Int. 54: 1052–1062.
19. Lee, S. H., J. S. Bae, S. H. Park, B. H. Lee, R. W. Park, J. Y. Choi, J. Y. Park,S. W. Ha, Y. L. Kim, T. H. Kwon, and I. S. Kim. 2003. Expression of TGF-beta-induced matrix protein betaig-h3 is up-regulated in the diabetic rat kidney andhuman proximal tubular epithelial cells treated with high glucose. Kidney Int. 64:1012–1021.
20. Schorderet, D. F., M. Menasche, S. Morand, S. Bonnel, V. Buchillier, D. Marchant,K. Auderset, C. Bonny, M. Abitbol, and F. L. Munier. 2000. Genomic charac-terization and embryonic expression of the mouse Bigh3 (Tgfbi) gene. Biochem.Biophys. Res. Commun. 274: 267–274.
21. Hashimoto, K., M. Noshiro, S. Ohno, T. Kawamoto, H. Satakeda, Y. Akagawa,K. Nakashima, A. Okimura, H. Ishida, T. Okamoto, et al. 1997. Characterizationof a cartilage-derived 66-kDa protein (RGD-CAP/beta ig-h3) that binds to col-lagen. Biochim. Biophys. Acta 1355: 303–314.
22. Hanssen, E., B. Reinboth, and M. A. Gibson. 2003. Covalent and non-covalentinteractions of betaig-h3 with collagen VI. Beta ig-h3 is covalently attached tothe amino-terminal region of collagen VI in tissue microfibrils. J. Biol. Chem.278: 24334–24341.
23. Thapa, N., K. B. Kang, and I. S. Kim. 2005. Beta ig-h3 mediates osteoblastadhesion and inhibits differentiation. Bone 36: 232–242.
24. Oh, J. E., J. K. Kook, and B. M. Min. 2005. Beta ig-h3 induces keratinocytedifferentiation via modulation of involucrin and transglutaminase expressionthrough the integrin alpha3beta1 and the phosphatidylinositol 3-kinase/Aktsignaling pathway. J. Biol. Chem. 280: 21629–21637.
25. Kim, J. E., S. J. Kim, H. W. Jeong, B. H. Lee, J. Y. Choi, R. W. Park, J. Y. Park,and I. S. Kim. 2003. RGD peptides released from beta ig-h3, a TGF-beta-induced cell-adhesive molecule, mediate apoptosis. Oncogene 22: 2045–2053.
26. Ruoslahti, E. 1996. RGD and other recognition sequences for integrins. Annu.Rev. Cell Dev. Biol. 12: 697–715.
27. Hynes, R. O. 2002. Integrins: bidirectional, allosteric signaling machines. Cell110: 673–687.
28. Ohno, S., M. Noshiro, S. Makihira, T. Kawamoto, M. Shen, W. Yan,Y. Kawashima-Ohya, K. Fujimoto, K. Tanne, and Y. Kato. 1999. RGD-CAP((beta)ig-h3) enhances the spreading of chondrocytes and fibroblasts via integ-rin alpha(1)beta(1). Biochim. Biophys. Acta 1451: 196–205.
29. Kim, M. O., S. J. Yun, I. S. Kim, S. Sohn, and E. H. Lee. 2003. Transforminggrowth factor-beta-inducible gene-h3 (beta(ig)-h3) promotes cell adhesion ofhuman astrocytoma cells in vitro: implication of alpha6beta4 integrin. Neurosci.Lett. 336: 93–96.
30. Ferguson, J. W., B. S. Thoma, M. F. Mikesh, R. H. Kramer, K. L. Bennett,A. Purchio, B. J. Bellard, and R. G. LeBaron. 2003. The extracellular matrixprotein betaIG-H3 is expressed at myotendinous junctions and supports musclecell adhesion. Cell Tissue Res. 313: 93–105.
The Journal of Immunology 5843
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

31. Kim, J. E., S. J. Kim, B. H. Lee, R. W. Park, K. S. Kim, and I. S. Kim. 2000.
Identification of motifs for cell adhesion within the repeated domains of transforming
growth factor-beta-induced gene, betaig-h3. J. Biol. Chem. 275: 30907–30915.32. Klintworth, G. K. 2009. Corneal dystrophies. Orphanet J. Rare Dis. 4: 7.33. Wu, Y., B. Han, H. Luo, R. Roduit, T. W. Salcedo, P. A. Moore, J. Zhang, and
J. Wu. 2003. DcR3/TR6 effectively prevents islet primary nonfunction after
transplantation. Diabetes 52: 2279–2286.34. Han, B., P. A. Moore, J. Wu, and H. Luo. 2007. Overexpression of human decoy
receptor 3 in mice results in a systemic lupus erythematosus-like syndrome.
Arthritis Rheum. 56: 3748–3758.35. Zhang, N., A. Richter, J. Suriawinata, S. Harbaran, J. Altomonte, L. Cong,
H. Zhang, K. Song, M. Meseck, J. Bromberg, and H. Dong. 2004. Elevated
vascular endothelial growth factor production in islets improves islet graft vas-
cularization. Diabetes 53: 963–970.36. Luther, M. J., E. Davies, D. Muller, M. Harrison, A. J. Bone, S. J. Persaud, and
P. M. Jones. 2005. Cell-to-cell contact influences proliferative marker expression
and apoptosis in MIN6 cells grown in islet-like structures. Am. J. Physiol.
Endocrinol. Metab. 288: E502–E509.37. Parsons, J. T. 2003. Focal adhesion kinase: the first ten years. J. Cell Sci. 116:
1409–1416.38. Kenmochi, T., M. Miyamoto, and Y. Mullen. 1996. Protection of mouse islet
isografts from nonspecific inflammatory damage by recipient treatment with
nicotinamide and 15-deoxyspergualin. Cell Transplant. 5: 41–47.39. Wahoff, D. C., B. E. Papalois, J. S. Najarian, D. M. Kendall, A. C. Farney,
J. P. Leone, J. Jessurun, D. L. Dunn, R. P. Robertson, and D. E. Sutherland. 1995.
Autologous islet transplantation to prevent diabetes after pancreatic resection.
Ann. Surg. 222: 562–575, discussion 575–579.40. Shapiro, A. M., J. R. Lakey, E. A. Ryan, G. S. Korbutt, E. Toth, G. L. Warnock,
N. M. Kneteman, and R. V. Rajotte. 2000. Islet transplantation in seven patients
with type 1 diabetes mellitus using a glucocorticoid-free immunosuppressive
regimen. N. Engl. J. Med. 343: 230–238.
41. Witkowski, P., S. B. Zakai, A. Rana, Z. Sledzinski, and M. A. Hardy. 2006.Pancreatic islet transplantation, what has been achieved since Edmonton break-through. Ann. Transplant. 11: 5–13, discussion 32–43.
42. Gruessner, A. C., and D. E. Sutherland. 2005. Pancreas transplant outcomes forUnited States (US) and non-US cases as reported to the United Network forOrgan Sharing (UNOS) and the International Pancreas Transplant Registry(IPTR) as of June 2004. Clin. Transplant. 19: 433–455.
43. Rosenberg, L. 1998. Clinical islet cell transplantation. Are we there yet? Int. J.Pancreatol. 24: 145–168.
44. Rosenberg, L., R. Wang, S. Paraskevas, and D. Maysinger. 1999. Structural andfunctional changes resulting from islet isolation lead to islet cell death. Surgery126: 393–398.
45. Thomas, F. T., J. L. Contreras, G. Bilbao, C. Ricordi, D. Curiel, andJ. M. Thomas. 1999. Anoikis, extracellular matrix, and apoptosis factors inisolated cell transplantation. Surgery 126: 299–304.
46. Lupi, R., F. Dotta, L. Marselli, S. Del Guerra, M. Masini, C. Santangelo,G. Patane, U. Boggi, S. Piro, M. Anello, et al. 2002. Prolonged exposure tofree fatty acids has cytostatic and pro-apoptotic effects on human pancreaticislets: evidence that beta-cell death is caspase mediated, partially dependenton ceramide pathway, and Bcl-2 regulated. Diabetes 51: 1437–1442.
47. Prentki, M., E. Joly, W. El-Assaad, and R. Roduit. 2002. Malonyl-CoA signaling,lipid partitioning, and glucolipotoxicity: role in beta-cell adaptation and failurein the etiology of diabetes. Diabetes 51(Suppl 3): S405–S413.
48. Soon-Shiong, P., E. Feldman, R. Nelson, J. Komtebedde, O. Smidsrod, G. Skjak-Braek, T. Espevik, R. Heintz, and M. Lee. 1992. Successful reversal of spon-taneous diabetes in dogs by intraperitoneal microencapsulated islets. Trans-plantation 54: 769–774.
49. Lanza, R. P., S. J. Sullivan, and W. L. Chick. 1992. Perspectives in diabetes. Islettransplantation with immunoisolation. Diabetes 41: 1503–1510.
50. Rygiel, K. A., H. Robertson, H. L. Marshall, M. Pekalski, L. Zhao, T. A. Booth,D. E. Jones, A. D. Burt, and J. A. Kirby. 2008. Epithelial-mesenchymal transitioncontributes to portal tract fibrogenesis during human chronic liver disease. Lab.Invest. 88: 112–123.
5844 THE ROLE OF TGF-bi IN b-CELL FUNCTION AND GROWTH
by guest on April 12, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from


















