Tesis de Licenciatura - biblio.uabcs.mxbiblio.uabcs.mx/tesis/te2977.pdf · Lista de Anexos Anexo I...
68
UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR ÁREA DE CONOCIMIENTOS EN CIENCIAS DEL MAR DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA Tesis de Licenciatura “Efecto in vitro del β-caroteno natural obtenido a partir de Dunaliella salina (Chlorophyta) sobre la línea celular de cáncer MDA-MB-231” QUE COMO REQUISITO PARA OBTENER EL Título de Biólogo Marino PRESENTA: Ilhui Rocío Gómez Pacheco DIRECTOR: Jorge Olmos Soto LA PAZ, BAJA CALIFORNIA SUR, AGOSTO DE 2013
Transcript of Tesis de Licenciatura - biblio.uabcs.mxbiblio.uabcs.mx/tesis/te2977.pdf · Lista de Anexos Anexo I...

UNIVERSIDAD AUTÓNOMA DE BAJA CALIFORNIA SUR
ÁREA DE CONOCIMIENTOS EN CIENCIAS DEL MAR
DEPARTAMENTO ACADÉMICO DE BIOLOGÍA MARINA
Tesis de Licenciatura
“Efecto in vitro del β-caroteno natural obtenido a partir de
Dunaliella salina (Chlorophyta) sobre la línea celular de cáncer
MDA-MB-231”
QUE COMO REQUISITO PARA OBTENER EL
Título de Biólogo Marino
PRESENTA:
Ilhui Rocío Gómez Pacheco
DIRECTOR:
Jorge Olmos Soto
LA PAZ, BAJA CALIFORNIA SUR, AGOSTO DE 2013

Dedicatoria
A
Mi mamá
Que nunca deja de sorprenderme con lo maravillosa que es.
Abuelita Lupita
Que siempre cuido y amó a todos. Te llevo siempre conmigo.
Mi abuela Toña,
El amor de mis amores. Gracias por estar siempre a mi lado.
Javier y Andrés,
El regalo más bonito que tuve por mis padres. Los adoro.
A mi tía Laura Lilían,
Porque las batallas más grandes, solo se pueden ganar con amor a la vida.
A mis tías María Elena e Ilhui,
Por inspirarme siempre a seguir estudiando.
Mi papá,
Y mis abuelos Sergio y Carlos
Por todo su cariño.

Agradecimientos académicos Al Dr. Jorge Olmos Soto por la dirección de esta tesis y por el apoyo brindado;
pero sobre todo, por aceptarme y darme la oportunidad de aprender cosas
nuevas.
A la M.C. Rosalía Contreras Flores técnico del laboratorio, durante el cultivo de
Dunaliella salina y en toda mi estancia como estudiante del laboratorio. Gracias
nuevamente.
A la M.C. Silvia Viviana Pitones Rubios, por su enorme ayuda técnica y asesoría
durante la fase de cultivo celular, en la elaboración de los ensayos y por la
bibliografía compartida. Muchas gracias sin tu ayuda no podría haber realizado
esta tesis.
Al M.C. Eduardo Morales Guerrero por toda su asesoría y ayuda brindada al
realizar los análisis por HPLC.
Al M.C. Viktor Iván Rodríguez Abdalá, por su ayuda en el lenguaje de
programación R.
Al Dr. Jose Luis Ortiz Galindo y al Dr. Noé Díaz Viloria, por la extensa revisión de
la tesis. Mil gracias por todo.
Al comité revisor tesis: Dr. Sergio Flores Ramírez, M.C. Marco Antonio Medina
López, al Dr. Noé Díaz Viloria y al Dr. Carlos Sánchez Ortiz.
A Beti y al Dr. Alejandro Gómez por su ayuda en los trámites administrativos. A la
UABCS y al CICESE.

Agradecimientos personales
A mi mamá, por todo su amor y todo el tiempo que me ha dedicado desde el día
en que nací. Muchas gracias por estar siempre a mi lado, en las buenas y en las
malas, por regañarme siempre que lo necesité, Por enseñarme todo cuanto sé.
A Silvia Viviana Pitones, por tu amistad, tu tiempo, tu compañía y gran ayuda no
solo académica sino personal. Siempre estaré en deuda contigo, eres una persona
muy especial y valiosa. Gracias por todo.
A José Luis Ortiz Galindo, muchas gracias tío por todo el cariño, tiempo y apoyo,
que me has dado siempre, por ser parte de nuestra familia.
A mi tía María Elena Gómez Rojo, muchas gracias por todo tu cariño, apoyo y
ánimos. Te quiero mucho madrina.
A toda mi hermosa familia que siempre me ha apoyado desde lejos.
A las familias Norzagaray-Navarro y Silva-Cruz por ser como mi familia.
A Laura Camacho, muchas gracias por animarme a hacer la tesis en ensenada; y
por tu gran amistad. Te quiero mucho.
A Magaly Gómez, Malissa Fragoso, Angelina Lazcano, Betsabé Luna, Diana
Zaleta, Mirelle Dávila, Rocío González, Casandra Gálvez, Marcela Valdovinos,
Christian por su amistad.
A Iván Rodríguez, muchas gracias por estar a mi lado.

Índice
Lista de figuras i
Lista de cuadros iiI
Lista de anexos iii
Glosario iv
Resumen vi
1. Introducción 1
2. Antecedentes 2
2.1. Los carotenoides 2
2.2. Dunaliella salina 5
2.3. Desarrollo del cáncer 8
3. Justificación 12
4. Hipótesis 12
5. Objetivo general 13
5.1. Objetivos particulares 13
6. Materiales y Métodos 14
7. Resultados 20

7.1 Cultivo de Dunaliella salina 20
7.2 β-Caroteno 21
7.3 Curva de crecimiento 26
7.4 Ensayo de viabilidad celular con β-caroteno 26
8. Discusión 30
9. Conclusiones 34
10. Recomendaciones 35
11. Bibliografía 36
12. Anexos 41

i
Lista de figuras Figura 1 Molécula de β-caroteno. 5
Figura 2 Dunaliella salina. Contraste de fases con
fluorescencia, objetivo 100X.
5
Figura 3 Línea celular MDA-MB-231. Microscopio de
Contraste de fases, objetivo 10X.
10
Figura 4. Línea celuar HaCat. Microscopio de Contraste
de fases, objetivo de 10X.
11
Figura 5. Curva de crecimiento poblacional del cultivo de
Dunaliella salina hasta alcanzar la fase
estacionaria.
20
Figura 6 Cromatogramas obtenidos por HPLC a una
longitud de onda de 450 nm.
22
Figura 7 Cromatogramas sobrepuestos del estándar y el
extracto natural de D. salina.
23
Figura 8 Curva de calibración por HPLC para estimar la
concentración de β-caroteno en el extracto
natural.
25
Figura 9 Resultados del ensayo de viabilidad celular
realizado con β-caroteno sobre MDA-MB-231.
29
Figura 10 Resultados del ensayo de viabilidad celular
realizado con β-caroteno sobre HaCat.
29
Figura 11 Variación dentro del tratamiento de 2h con β-
caroteno en la línea celular MDA-MB-231
51
Figura 12 Variación dentro de grupos con el tratamiento
de 2h con β-caroteno.
52

ii
List
a de cuadros
Figura 13 Variación dentro del tratamiento de 24h con β-
caroteno en la línea celular MDA-MB-231.
54
Figura 14 Variación dentro del tratamiento de 24h con β-
caroteno en la línea celular MDA-MB-231:
1)Células con BS, 2) Células con BN.
54
Figura 15 Variación dentro del tratamiento de 24h con β-
caroteno en la línea celular HaCat: 1)Células
HaCat con BS, 2)Células HaCat con BN.
55
Figura 16 Variación entre grupos con el tratamiento de
24h con β-caroteno. Células MDA-MB-231 con
BS, 2) Células MDA-MB-231 con BN, 3)Células
HaCat con BS, 4) Células HaCat con BN.
56

iii
Lista de cuadros
Cuadro I Incrementos en la concentración de NaCl
realizados en el cultivo L.
15
Cuadro II Esquema general del ensayo con β-caroteno. 18
Cuadro III Mediciones de absorbancia obtenidas en el
cromatógrafo HPLC al analizar el estándar
sintético de trans β-caroteno y el extracto
natural de D. salina.
24
Lista de Anexos Anexo I Cultivo de D. salina. 41
Anexo II Soluciones y medios empleados para el cultivo
de las líneas celulares.
46
Anexo III Ensayo con β-caroteno. 48
Anexo IV Ensayo colorimétrico de MTT. 50
Anexo V Resultados del ANOVA realizado con el
programa R.
51

iv
Glosario
Adenocarcinoma: Cáncer que empieza en las células glandulares. Las células
glandulares se encuentran en el tejido que reviste algunos órganos internos y que
producen y liberan sustancias en el cuerpo. La mayoría de los cánceres de mama,
páncreas, pulmón, próstata y colon son adenocarcinomas (INC, 2013).
Antioxidante: Es una molécula capaz de retardar o prevenir la oxidación (pérdida
de electrones) de otra molécula, sin llegar a perder su estabilidad (Holum, 1999).
Pro-oxidante: Es una molécula capaz de reducir a otra molécula (Holum, 1999).
Medio suplementado: Medio que ha sido complementado o enriquecido con suero
fetal bovino; y que por ello cuenta con los nutrientes, cofactores y hormonas de
crecimiento necesarios para el mantenimiento de un cultivo celular (Pitones-Rubio
y Olmos-Soto, 2010).
Confluencia: La cantidad de células que crecen sobre la superficie de una caja o
placa de cultivo; esta generalmente se expresa en porcentajes (Langdon, 2004).
Cultivo primario: El cultivo iniciado a partir de células, tejidos u órganos tomados
directamente de un organismo (Langdon, 2004).
Pase: Consiste en tomar una fracción de células a partir de un cultivo establecido
para colocarlas en una nueva placa o caja que cuenta con el medio y los
nutrientes necesarios para poder iniciar un nuevo sub-cultivo celular (Langdon,
2004).

v
Línea celular continua: Son células de un tipo único (humano, animal o vegetal),
que han sido sub-cultivadas a partir de un cultivo primario del cual derivan. Estas
células tienen alteraciones genéticas que les dan la capacidad de proliferar
indefinidamente si se les brindan las condiciones ambientales y los nutrientes
esenciales para su mantenimiento (Langdon, 2004).

vi
Resumen
Dunaliella salina es una microalga Cholorophyta produce de forma natural grandes
cantidades de β-caroteno. En humanos se ha observado que esta molécula tiene
distintas funciones biológicas como estabilizador de membrana, antioxidante y
precursor de la vitamina A; además puede actuar como quimio-protector y
anticancerígeno. Por ello el objetivo de este trabajo fue demostrar si el β-caroteno
tiene un efecto in vitro sobre la supervivencia de la línea celular para cáncer de
mama MDA-MB-231. Para ello se realizó un ensayo en el que se aplicaron dos
tratamientos de 6 µg/µl de β-caroteno, cada uno con dos condiciones de
incubación con distintas duraciones de tiempo (2 y 24 horas). Los tratamientos
consistieron en un extracto natural obtenido a partir de D. salina y en trans β-
caroteno sintético. Ambos tratamientos disminuyeron significativamente la
supervivencia de las células de MDA-MB-231; sin embargo el efecto se vio
potenciado cuando se aplicó β-caroteno de origen natural. El mayor efecto se
observó con un tiempo de incubación de 2 horas, en el cual la supervivencia de las
células de MDA-MB-231 fue de un 27.9 %.

1
1 Introducción En los océanos del mundo habita una gran diversidad de microorganismos
conformada por bacterias, virus, hongos, protozoarios y algas. Estos
microorganismos producen en conjunto, un reservorio natural de sustancias bio-
activas, que son producto de su metabolismo primario y secundario. Estas
sustancias son muy novedosas, ya que tienen amplias aplicaciones en la industria
y en las ciencias de la salud (Ireland et al., 1993; Paniagua-Michel, 2009).
Entre estos microorganismos se encuentra Dunaliella salina, una microalga
Chlorophyta que es uno de los eukariotas más extremófilos que se conoce y que
bajo condiciones de estrés salino, alta irradianza y falta de nutrientes, produce
grandes cantidades de β-caroteno, para proteger la clorofila a y el DNA nuclear
(Ben-Amotz et al., 1987; Borowitzka, 1990; Olmos-Soto et al., 2002; Ben-Amotz et
al., 2009).
El β-caroteno es una molécula que presenta una fuerte actividad antioxidante,
capaz de captar y destruir los radicales producidos por la quimioterapia durante el
tratamiento del cáncer. Por esta razón, se le atribuye la propiedad de agente
quimio-preventivo y anticancerígeno (Bendich y Olson, 1989; Lamson y Brignall,
1999). A este caroteno, también se le ha atribuido la capacidad de detener el
proceso de metástasis en algunas clases de cáncer, como lo son: el de piel, boca
y pulmón (Venugopal, 2009). Sin embargo, en lo que concierne a la aparición de
este último tipo de cáncer, hay estudios que consideran que el consumo del β-
caroteno junto con el tabaco y el alcohol es un factor de riesgo (Leo y Lieber,
1999).
En México el tipo de cáncer más común en mujeres mayores de 20 años es el
carcinoma de mama (CaMa) (INEGI, 2013). Debido a lo anterior, en este trabajo

2
se busca probar si el β-caroteno producido por D. salina, al que ya se le han
atribuido actividades anticancerígenas (Borowitska, 1990; Ben-Amotz, 2009;
Venugopal, 2009); presenta un efecto in vitro sobre una línea celular de cáncer de
mama.
2 Antecedentes
2. 1 Los carotenoides
Los carotenoides son una familia de pigmentos de color amarillo- naranja o rojo
que proveen de un intenso color a muchas especies de organismos (Asker et al.,
2012). Estos pigmentos son los más ampliamente distribuidos en la naturaleza, ya
que se les encuentra en las plantas, las bacterias, las algas, los hongos y los
animales; sin importar si se trata de especies marinas, terrestres o de agua dulce
(Mínguez-Mosquera et al., 2008; Venugopal, 2009). Hasta la fecha se han
descubierto más de 700 carotenos distintos en la naturaleza (Asker et al., 2012),
entre ellos el β-caroteno.
2. 1. 1 Función biológica
En los organismos fotosintéticos los carotenoides sirven como pigmentos
accesorios, ayudando a absorber la luz y transfiriendo la energía a la clorofila
(Asker et al., 2012). Otra función de los carotenoides en organismos fototróficos y
no-fototróficos, es la de proteger a las células contra daños en el núcleo causados
por la producción de radicales libres, y que puedan originar mutaciones u otras
alteraciones genéticas (Bendich y Olson, 1989).
En los miembros del reino animal incluyendo a los humanos, vacas, aves, peces y
algunos crustáceos, los carotenoides no pueden ser sintetizados y tienen que ser

3
adquiridos en la dieta. Algunas de sus principales funciones son: Actuar como un
precursores de la Vitamina A en todas sus formas activas (retinol, retinal y ácido
retinoico) (Holum, 1999; Venugocopal, 2009; Asker et al., 2012), como
antioxidantes y pro-oxidantes (Krinsky, 1988; Bendich y Olson, 1989).
Además, en los seres humanos los carotenoides son constituyentes normales de
la sangre y algunos tejidos. Así, en el cuerpo de una persona bien nutrida la
cantidad total de carotenoides varía de 100-150 mg. De ese total el 1% está
presente en el suero sanguíneo, el 80-85% en el tejido adiposo, el 8-12% en el
hígado (8-12%) y el 2-3% en los músculos. Los carotenoides que se encuentran
en mayor cantidad en el suero humano son el β-caroteno, el α-caroteno, la
cryptoxantina, el licopeno y la luteína de los cuales el β-caroteno es el más
abundante (15 al 30%) (Bendich y Olson, 1989).
2. 1. 2 Uso de los carotenoides en la industria y ciencias de la salud
Los carotenoides se emplean en la elaboración de alimento para aves de corral,
para organismos cultivados por acuacultura y como colorante en la manufactura
de alimentos para humanos (Venugopal, 2009). También se emplean como
complemento nutricional y fuente de provitamina-A, de las cuales el β-caroteno
tiene la mayor actividad. También se les emplea como aditivo en cosméticos
(Holum, 1999).
En ciencias de la salud hay estudios que muestran que en humanos, los
carotenoides reducen los niveles de colesterol, homocisteína, agregación
plaquetaria y presión sanguínea, teniendo una función cardio-protectora debido a
su alta actividad antioxidante. Hay evidencia que indica, que también ayudan a
prevenir la aparición de úlceras gástricas, combatir enfermedades neuro-
degenetarivas, a la activación de las células T del sistema inmune, y que protegen

4
a las células-β de la presencia de radicales libres en pacientes con diabetes
mellitus (Holum, 1999; Herchberg, 2005; Mínguez-Mosquera et al., 2008;
Venugopal, 2009).
En relación al cáncer existen estudios epidemiológicos y oncológicos, en los que
se muestra que algunos carotenoides como la Aztaxantina, el Lycopeno y el β-
caroteno tienen una actividad anticancerígena, ya sea como agentes
quimiopreventivos contra el desarrollo del cáncer de piel, boca, pulmón y seno,
como inhibidores del proceso de metástasis en casos de cáncer de hígado
(Dimitrov et al., 1988; Mínguez-Mosquera et al., 2008; Emeish, 2012).
Además, se ha observado que la administración de β-caroteno de origen natural
potencia el efecto de la quimioterapia y la radioterapia, e incluso incrementa la
supervivencia en pacientes enfermos de cáncer (Lamson y Brignall, 1999).
2. 1.3 β-caroteno: Molécula y propiedades generales
El β-caroteno (C40H56) tiene un peso molecular de 536.9 umas y es uno de los
carotenoides más ampliamente distribuido en la naturaleza (Figura 1). Es un
hidrocarburo poli-insaturado compuesto por dos moléculas de retinol. Al tratarse
de una molécula lipofílica, es insoluble en soluciones acuosas a menos que
tengan cierto grado de polaridad, como los solventes orgánicos: acetona y
diclorometano (Venugopal, 2009).
La presencia de dobles enlaces conjugados en el β-caroteno hace que presente
isómeros geométricos cis y trans (E/Z), que pueden ínter-convertirse cuando se
encuentran en solución y que presentan sus enlaces dobles en distintas
posiciones. Estos cambios en la geometría afectan el punto de fusión, el color, la
solubilidad, la estabilidad y un poco sus propiedades ópticas de absorción de luz

5
(Venugopal, 2009). Así, aunque presentan un máximo de absorción a una longitud
de onda de 450 nm, esta es interferida por la presencia de sus anillos β y enlaces
dobles (Mínguez-Mosquera et al., 2008). Cuando la molécula del β-caroteno se
encuentra en solución con presencia de luz y oxígeno se oxida, puede polimerizar
y actuar agente quelante, además de formar isómeros (Venugopal, 2009).
Figura 1. Molécula de β-caroteno
2. 2 Dunaliella salina
2. 2. 1 Taxonomía
De acuerdo al ITIS (2003), la clasificación de la especie es la siguiente:
División Chlorophyta
Clase Chlorophyceae
Orden Volvocales
Family Dunaliellaceae
Género Dunaliella
Especie Dunaliella salina (Teodoresco, 1905).

6
Figura 2. Dunaliella salina. Contraste de fases con fluorescencia, objetivo
100X.
2. 2. 2 Características generales de la especie
D. salina es una microoalga verde unicelular que tiene una forma ovoide, dos
flagelos apicales y un color que varía de verde-amarillo a rojo profundo (Parra et
al., 1990). Esta microalga pertenece al género Dunaliella conformado por 27
especies, las cuales comparten las siguientes características: Carecen de una
pared celular rígida de polisacáridos, están rodeadas por una delgada membrana
plasmática cubierta por una capa mucilaginosa, tienen un solo cloroplasto,
pirenoide, un núcleo y nucléolo (Borowitzka, 1990; Wilcox y Graham, 2000; Ben-
Amotz et al., 2009). Sin embargo solo D. salina y Dunaliella bardawil poseen la
capacidad de acumular grandes cantidades de β-caroteno y glicerol en su interior,
cuando se encuentra en ambientes extremos hipersalinos y en los que hay una
intensa radiación solar (Olmos-Soto et al., 2012).
2. 2. 3 Estudios nutricionales con Dunaliella
Entre los antecedentes más importantes se encuentran los de Ben-Amotz y
colaboradores (1987) y Ben-Amotz y Avron (1992). En estos trabajos se encontró
que D. salina produce una combinación de isómeros: 15-cis-β-caroteno (10%), 9-

7
cis-β-caroteno (41%), trans β-caroteno (42%) y 2 isómeros sin identificar (7%).
Ben-Amotz y Levi (1996) realizaron un estudio in vivo con humanos y observaron
que la mezcla isomérica de β-caroteno producida por Dunaliella bardawil fue más
absorbida, con respecto a otras formas de β-caroteno trans de origen sintético que
fueron administradas. También descubrieron, que tuvo una actividad antioxidante
más potente.
Diversos investigadores (Borowitska, 1990; Venugopal, 2009), mencionaron a D.
salina como una fuente importante de β-caroteno y complemento nutricional para
humanos, por su fuerte actividad pro vitamina A, entre otros beneficios.
2. 2. 4 Estudios previos en el noroeste de México
Se han aislado e identificado bioquímicamente varias cepas locales pertenecientes
al género Dunaliella, entre ellas D. salina en lagunas hipersalinas costeras
ubicadas en los estados de Baja California (San Quintin, La Salina) y Baja
California Sur (Salina de Guerrero Negro) (Gutiérrez-Millán, 1996; Sánchez-
Castrejón, 1998).
En el 2004, Capa-Robles (2004) estudió la fisiología de la especie para inducir la
producción de β-caroteno con fines comerciales. Para ello, estudió el efecto de la
salinidad, así como la presencia o ausencia de vitaminas en condiciones no
estresantes de cultivo; además de que identificó la presencia de tres tipos de β-
caroteno: 1)trans-β-caroteno, 2)α-caroteno y 3)de tipo no trans.
Olmos y colaboradores (2002) identificaron molecularmente a la especie por
medio del método de intron-zising finger-printing, demostrando que las especies
de Dunaliella tiene un tamaño específico, de 18S ADNr (Olmos-Soto et al., 2012).

8
Contreras-Flores (2005) estudió la variabilidad intraespecie y en función a ella
evaluó la acumulación de β-caroteno bajo distintas condiciones de cultivo.
2. 3 Desarrollo del cáncer
Cáncer es el nombre general que se da a más de 100 enfermedades distintas pero
que presentan una característica: Todas se originan a partir de células anormales
o neoplásicas que proliferan de forma continua y descontrolada, formando masas
de tejido llamados tumores. Algunas de estas células adquieren la habilidad de
invadir nuevos tejidos y órganos causando la enfermedad del cáncer (INC, 2013).
La proliferación celular que da origen al cáncer es ocasionada por una alteración
de los eventos que comprenden el ciclo celular, y que dependen de los niveles
apropiados de la transcripción y traducción de ciertos genes, cuyas mutaciones o
fallas han sido estrechamente relacionadas con el desarrollo del cáncer. Estos
genes se dividen en dos clases: 1) Los proto-oncogenes, o genes cuyas proteínas
estimulan la división celular e inhiben la muerte celular programada o apoptosis; y
2) Los genes supresores de tumores, cuyas proteínas restringen el crecimiento
celular o conllevan a la apoptosis (Lodish et al., 2002). Las mutaciones en estos
genes son ocasionadas por diversos factores que dañan el ADN nuclear (Baba,
2007).
Los factores carcinógenos pueden ser de tipo físico, como la radiación UV-A y UV-
B, los rayos X, y rayos Gama; químico, debido a la presencia de epóxidos,
nitrosaminas, hidrocarburos aromáticos y minerales (As, Cr, Cd y Ni); biológico, lo
que incluye a sustancias liberadas por parásitos, plantas, o microorganismos, o
que se encuentran en alimentos y medicinas (Flavonoides, taninos, Ochratoxina A,
caspacianina); y genético (Baba, 2007). Los factores genéticos pueden tener

9
distintos orígenes como son: mutaciones heredas en ciertos genes (genes BRCA-
1 y BRCA-2), la acción de algunos tipos de virus (Papilomavirus, herpesvirus y
adenovirus), y la liberación de especies reactivas de Oxígeno (1O2 ) y otros
radicales libres (1OH, -NO) durante el metabolismo celular (Baba, 2007; Musolino
et al., 2007; Fiaschi y Chiaruggi, 2012).
El nombre que se le da a cada tipo de cáncer está determinado por el tipo de
tejido y órgano en el cual se origina. Así, existe cáncer de seno, piel, de hígado,
hueso y pulmón, entre muchos otros (INC, 2013).
2. 3. 1 Cáncer de mama
El cáncer de mama (CaMa) es causado cuando hay una proliferación acelerada,
desordenada y no controlada de las células de los tejidos de la glándula mamaria,
y es causado por mutaciones que ocurren en los genes que actúan normalmente
suprimiendo o estimulando la continuidad del ciclo celular (Rodríguez-Bandala,
2010).
De los tipos de cáncer que existen, el CaMa junto con el cáncer de próstata, son
de los más agresivos, ya que hacen metástasis en los huesos y la mayoría de los
pacientes mueren cuando esto sucede (Kowalski et al., 2003).
2. 3. 2. Incidencia del Cáncer de Mama en México
El cáncer es una enfermedad que en muchos casos va asociada a la edad y a la
senectud, por lo que desde el último siglo ha ido incrementándose junto con la
edad de las personas y está en continuo aumento; siendo el cáncer de mama uno
de los más comunes en el mundo (WCR, 2008). En México las cifras son
alarmantes, ya que desde 2010 el CaMa es el tipo de cáncer más común en
mujeres mayores de 20 años (24%) y actualmente es la segunda causa de muerte

10
natural en mujeres de 35 años y más (INEGI, 2013).
2. 4 Líneas celulares
MDA-MB-231 (ATCC® HTB-26)
Esta línea celular fue aislada a partir de una paciente femenina de 51 años, de
origen caucásico, que presentaba un adenocarcinoma grado III de glándula de
mama; por lo que fueron elegidas como modelo para estudiar el efecto en ellas del
β-caroteno de origen natural. Estas células carecen de la capacidad de
diferenciarse, no forman tejidos, presentan un número cromosómico alterado (64-
69) y su forma es epitelial adherente (Cailleau et al., 1974).
Figura 3. Línea celular MDA-MB-231. Microscopio de Contraste de fases,
objetivo 10X

11
HaCat
Es una línea celular inmortal de queratinocitos humanos (QHA), cuyas células
conservan la capacidad de diferenciarse y crecer de forma semejante al tejido de
la piel humana (NHEQ) (Lehman, 1997). Debido a sus características las células
de esta línea fueron empleadas como modelo, para ver el efecto del β-caroteno en
células normales.
Figura 4. Línea celuar HaCat. Microscopio de Contraste de fases, objetivo de
10X.

12
3 Justificación Muchos de los tratamientos contra el cáncer en general y no solo de mama, son
altamente invasivos y poseen muchos efectos secundarios. Por ello encontrar
biomoléculas de origen natural como el β-caroteno producido por D. salina que
puedan actuar como agentes preventivos, o curativos en el tratamiento, ya sea
que inhiban la proliferación celular descontrolada o puedan inducir la apoptosis de
las células neoplásicas, sin causar efectos tan dañinos como la quimio o
radioterapia, aún es una panacea en la lucha contra todos los tipos de cáncer que
existen.
Por otro lado, hay estudios previos que indican que el β-caroteno puede ser un
factor que predisponga a algunos tipos de cáncer y otros que indican que tiene
una función preventiva. Este estudio puede ayudar a probar in vitro si esta
molécula: 1) Tiene un efecto en la supervivencia de las células de cáncer de
mama y 2) Si su origen sintético o natural causa alguna diferencia.
4 Hipótesis
Debido a que en otras especies de Dunaliella (Dunaliella bardawil) se demostró
una mayor absorción y una actividad antioxidante más potente con respecto al β-
caroteno de origen sintético; el β-catoteno natural obtenido a partir de Dunaliella
salina puede tener un mayor efecto en la supervivencia de las células MDA-MB-
231 que β-caroteno de origen sintético.

13
5 Objetivo General
Demostrar el efecto que tendrá el β-caroteno de origen natural presente en un
extracto obtenido a partir de la microoalga D. salina en la supervivencia de la línea
celular MDA-MB-231.
5. 1 Objetivos particulares
-Estandarizar una fase móvil que permita detectar y cuantificar por medio de
cromatografía líquida de alta resolución (HPLC), el β-caroteno sintético (comercial)
y de origen natural.
-Cuantificar el β-caroteno natural presente en el extracto natural de D. salina.
-Evaluar el efecto del β-caroteno natural y sintético en la supervivencia de la línea
celular MDA-MB-231.
-Evaluar si hay un efecto diferencial entre el β-caroteno natural y sintético en la
línea celular HaCat.
-Observar si el tiempo de incubación con ambos tratamientos con β-caroteno,
también causa un efecto sobre ambas líneas celulares.

14
6 Materiales y Métodos
6. 1 Dunaliella salina
La cepa de D. salina (UTEX LB2538) empleada para este estudio, proviene de un
lugar conocido como La Salina, localizada 35 km al norte de la ciudad de
Ensenada, B.C. (Contreras-Flores, 2005) y forma parte de la colección del
laboratorio de Microbiología Molecular del Centro de Investigación Científica y de
Educación Superior de Ensenada, B. C. lugar en donde se realizó este trabajo.
6. 1. 1 Condiciones iniciales de Cultivo
En dos matraces Erlenmeyer de 250 ml se colocaron por separado dos inóculos
iniciales de aproximadamente 13,800 células de D. salina a los que se les nombró
como cultivos F y L . A cada cultivo se les agregó 50 ml de un medio nutritivo, el
cual fue elaborado con la mezcla de los de medios cultivo para microalgas F2
(Guillard y Ryter, 1962) y Erdschreiber's (1:1) a una concentración 1.0 M de NaCl.
Los cultivos F y L fueron mantenidos con un fotoperiodo de luz-oscuridad de 12
horas, una temperatura de 25°C y un flujo luminoso de 20 quanta m-2 seg-1 hasta
que alcanzaron su fase de crecimiento estacionario. Durante este periodo, cada
cultivo fue agitado manualmente por la mañana y por la tarde, para favorecer el
intercambio gaseoso (Ver ANEXO I).
6. 1. 2. Cálculo de la densidad poblacional
Cada tres días se realizaron conteos directos a los cultivos F y L al microscopio
óptico con ayuda de una cámara de Neubauer. Para ello, se tomó una alícuota de
1 ml a partir de cada cultivo y se le agregaron 70 μl de solución de Lugol ácido
para inmovilizar las células y contarlas.
15
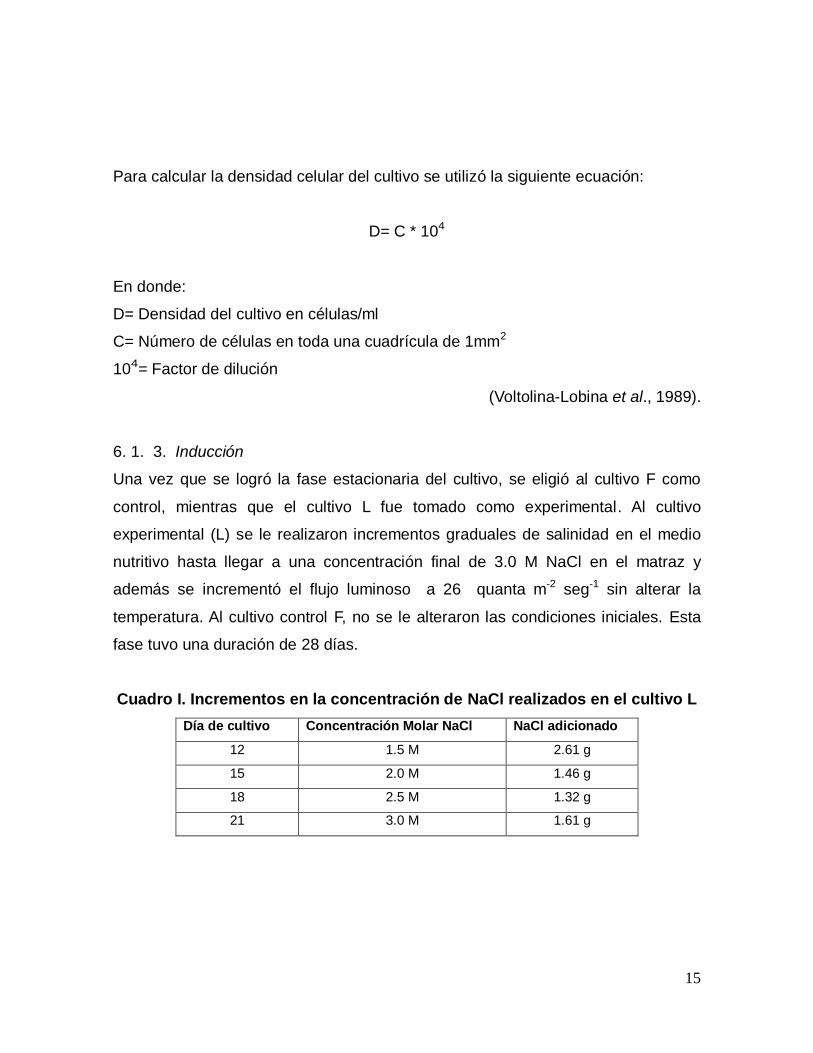
Para calcular la densidad celular del cultivo se utilizó la siguiente ecuación:
D= C * 104
En donde:
D= Densidad del cultivo en células/ml
C= Número de células en toda una cuadrícula de 1mm2
104= Factor de dilución
(Voltolina-Lobina et al., 1989).
6. 1. 3. Inducción
Una vez que se logró la fase estacionaria del cultivo, se eligió al cultivo F como
control, mientras que el cultivo L fue tomado como experimental. Al cultivo
experimental (L) se le realizaron incrementos graduales de salinidad en el medio
nutritivo hasta llegar a una concentración final de 3.0 M NaCl en el matraz y
además se incrementó el flujo luminoso a 26 quanta m-2 seg-1 sin alterar la
temperatura. Al cultivo control F, no se le alteraron las condiciones iniciales. Esta
fase tuvo una duración de 28 días.
Cuadro I. Incrementos en la concentración de NaCl realizados en el cultivo L
Día de cultivo Concentración Molar NaCl NaCl adicionado
12 1.5 M 2.61 g
15 2.0 M 1.46 g
18 2.5 M 1.32 g
21 3.0 M 1.61 g

16
6. 2 Betacaroteno
En este estudio se emplearon dos fuentes de β-caroteno: Una de origen natural
contenido en un extracto de D. salina obtenido con solventes orgánicos; cuya
elaboración no se describe ampliamente por motivos de patente. El segundo tipo
fue β-caroteno trans de origen sintético, fabricado por la empresa Sigma®.
6. 2. 1 Detección y cuantificación por HPLC
Para cuantificar e identificar el β-caroteno natural y sintético se empleó la técnica
de Cromatografía líquida de alta resolución (HPLC). El análisis se realizó en un
cromatógrafo Hewlett Packard serie 1100 con inyector manual, un sistema
cuaternario para solventes y un detector múltiple para longitud de onda. Se
empleó una columna Zorbax C8 de fase inversa con un tamaño de partícula de 5
μm, una elución isocrática con metanol al 100% a una velocidad de flujo de 0.5
ml/min y 25 minutos de corrida.
Para cuantificar el β-caroteno natural presente en el extracto, se realizaron
diluciones con metanol al 100% del estándar sintético (0.5, 1, 1.5 y 3 µg/µl) con
las cuales se obtuvo una curva de calibración con las mediciones obtenidas en el
cromatógrafo.
6.3 Cultivo celular
6. 3. 1 Condiciones de crecimiento
Para su crecimiento, las líneas HaCat y MDA-MB-231 (ATTC® HTB-26®) fueron
sembradas en cajas de cultivo (Corning®) con 7 ml del medio RPMI-1640
(GIBCO®) suplementado con 10% de SFB (Suero fetal bóvino) inactivado

17
(GIBCO®) y 1% del antibiótico comercial anti-ant (Invitrogen®). Las líneas
celulares siempre fueron incubadas en un ambiente estéril, a una temperatura de
37°C y una atmósfera húmeda con 4% de CO2 (Ver Anexo II).
6. 3. 2 Desprendimiento de líneas celulares
Cuando los cultivos presentaron una confluencia aproximada de un 85%, las
células fueron lavadas con 2 ml PBS y se les aplicó un tratamiento con una
solución de tripsina-EDTA y Verseno (1:1) (1 ml) (Ver Anexo II). Enseguida se
tomó una porción de las células desprendidas, y se les sembró en una caja nueva
con medio RPMI-1640 suplementado. Para realizar los ensayos posteriores con el
β-caroteno, solo se emplearon las células obtenidas a partir de un cuarto pase,
siguiendo lo propuesto por Pitones-Rubio y Olmos-Soto, 2010.
6. 3. 3. Curva de crecimiento
Para estimar la cantidad de células adecuada para realizar el ensayo con β-
caroteno fue necesario obtener una curva de crecimiento basada en la
confluencia. Para esto se utilizó una placa de cultivo de 96 pozos (Corning®), en
donde se colocaron en cada pozo, diferentes cantidades de células de MDA-MB-
231 (1000, 2000, 3000, 4000 y 5000) en 100 µl de medio RPMI suplementado, las
cuales fueron incubadas durante 4 horas a una temperatura de 70°C y una
atmósfera húmeda de CO2 al 4%. Transcurrido ese tiempo, a las células se les
realizó el arresto celular, para lo cual se les cambió el medio a RPMI sin
suplementar. Después con ayuda de un microscopio invertido Axiovert Zeiss se
observó de forma visual la confluencia celular. De inmediato, las células
nuevamente fueron incubadas durante 20 horas bajo las mismas condiciones de
crecimiento. Al cabo de ese tiempo la confluencia fue revisada nuevamente.
18
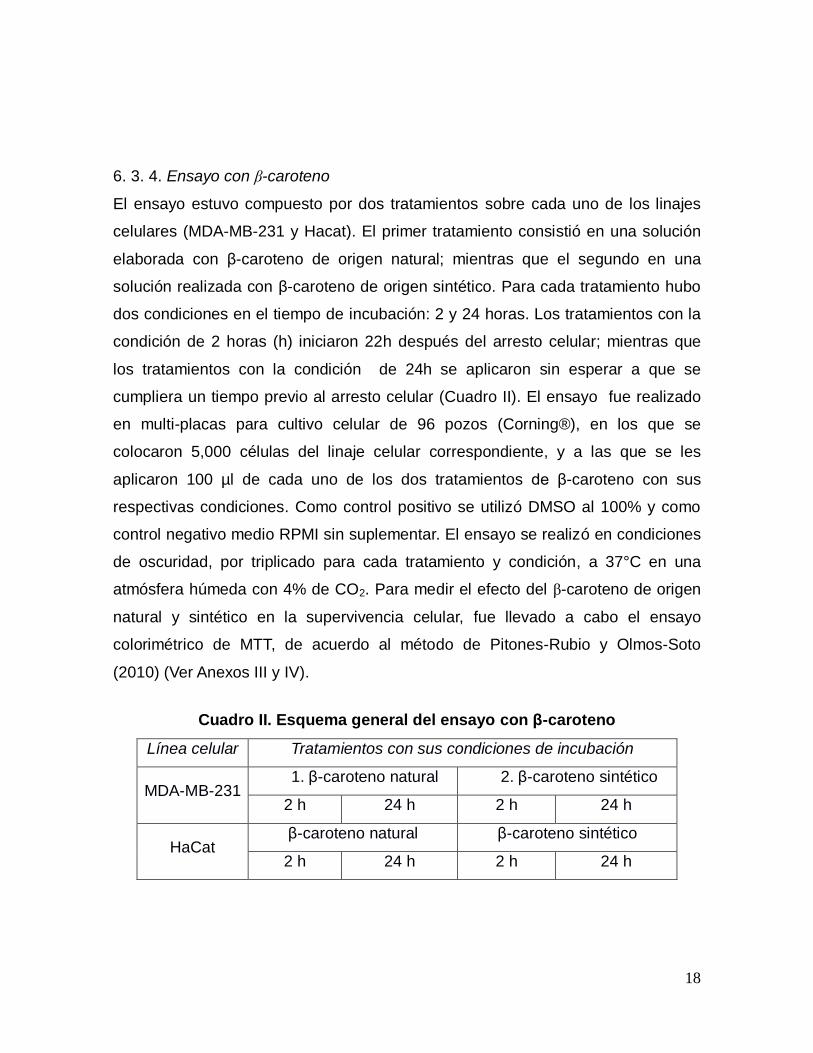
6. 3. 4. Ensayo con β-caroteno
El ensayo estuvo compuesto por dos tratamientos sobre cada uno de los linajes
celulares (MDA-MB-231 y Hacat). El primer tratamiento consistió en una solución
elaborada con β-caroteno de origen natural; mientras que el segundo en una
solución realizada con β-caroteno de origen sintético. Para cada tratamiento hubo
dos condiciones en el tiempo de incubación: 2 y 24 horas. Los tratamientos con la
condición de 2 horas (h) iniciaron 22h después del arresto celular; mientras que
los tratamientos con la condición de 24h se aplicaron sin esperar a que se
cumpliera un tiempo previo al arresto celular (Cuadro II). El ensayo fue realizado
en multi-placas para cultivo celular de 96 pozos (Corning®), en los que se
colocaron 5,000 células del linaje celular correspondiente, y a las que se les
aplicaron 100 µl de cada uno de los dos tratamientos de β-caroteno con sus
respectivas condiciones. Como control positivo se utilizó DMSO al 100% y como
control negativo medio RPMI sin suplementar. El ensayo se realizó en condiciones
de oscuridad, por triplicado para cada tratamiento y condición, a 37°C en una
atmósfera húmeda con 4% de CO2. Para medir el efecto del β-caroteno de origen
natural y sintético en la supervivencia celular, fue llevado a cabo el ensayo
colorimétrico de MTT, de acuerdo al método de Pitones-Rubio y Olmos-Soto
(2010) (Ver Anexos III y IV).
Cuadro II. Esquema general del ensayo con β-caroteno
Línea celular Tratamientos con sus condiciones de incubación
MDA-MB-231 1. β-caroteno natural 2. β-caroteno sintético
2 h 24 h 2 h 24 h
HaCat β-caroteno natural β-caroteno sintético
2 h 24 h 2 h 24 h

19
6.3.5 Análisis estadísticos
Las absorbancias obtenidos en el ensayo final de MTT fueron analizadas con los
programas Calc de Libre Oppen Office y con R (Ihaka y Gentleman, 1996). En el
primero, se realizaron los estadísticos básicos y se probó la normalidad y
homocedasticidad de los datos. Con el programa R se realizaron varios análisis de
variancia (ANDEVA), mediante el uso de matrices, con el fin de establecer si
existió una diferencia entre los tratamientos de β-caroteno con base al estadístico
F.
20
7 Resultados
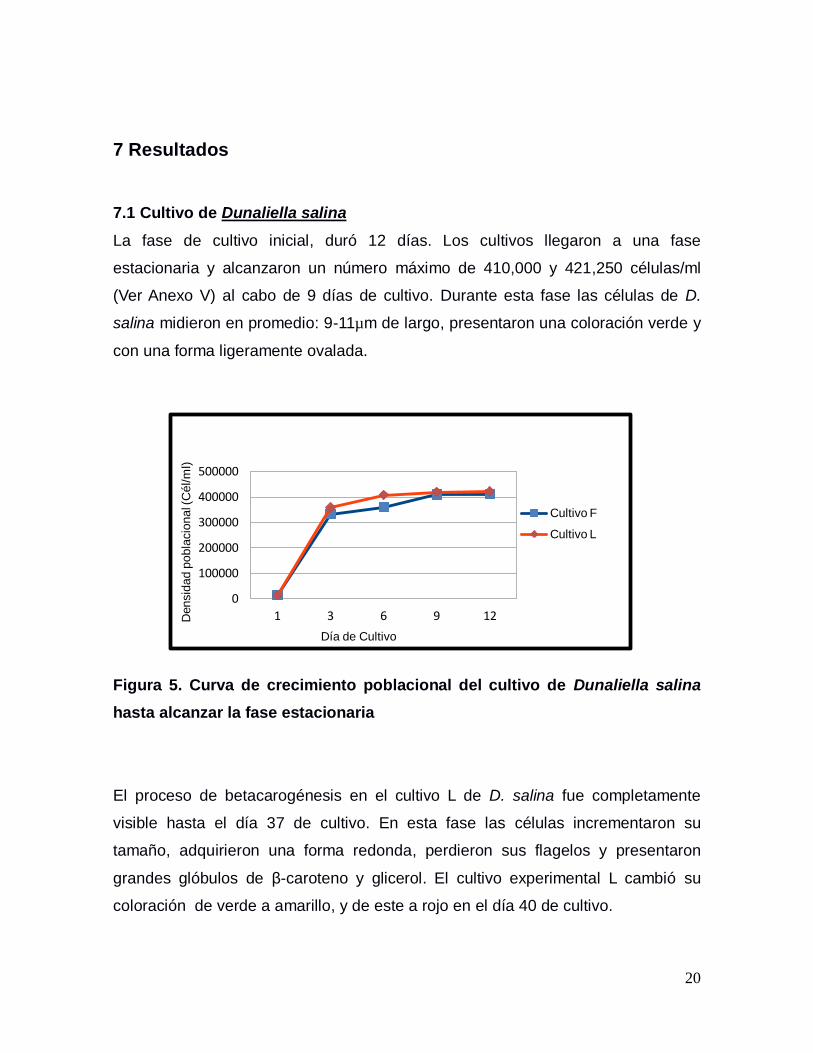
7.1 Cultivo de Dunaliella salina
La fase de cultivo inicial, duró 12 días. Los cultivos llegaron a una fase
estacionaria y alcanzaron un número máximo de 410,000 y 421,250 células/ml
(Ver Anexo V) al cabo de 9 días de cultivo. Durante esta fase las células de D.
salina midieron en promedio: 9-11µm de largo, presentaron una coloración verde y
con una forma ligeramente ovalada.
Figura 5. Curva de crecimiento poblacional del cultivo de Dunaliella salina
hasta alcanzar la fase estacionaria
El proceso de betacarogénesis en el cultivo L de D. salina fue completamente
visible hasta el día 37 de cultivo. En esta fase las células incrementaron su
tamaño, adquirieron una forma redonda, perdieron sus flagelos y presentaron
grandes glóbulos de β-caroteno y glicerol. El cultivo experimental L cambió su
coloración de verde a amarillo, y de este a rojo en el día 40 de cultivo.
1 3 6 9 12
0
100000
200000
300000
400000
500000
Día de Cultivo
Densid
ad p
obla
cio
nal (
Cél/m
l)
Cultivo F
Cultivo L

21
7.2 β-caroteno
En los cromatogramas obtenidos por HPLC el β-caroteno presentó su mayor pico
de absorción a una longitud de onda de 450 nm. Al analizar el estándar sintético
se observó un pico mayor correspondiente al trans β-caroteno, el cual tuvo un
8.105 min. En el extracto natural, se identificó un pico muy semejante, pero con un
tiempo de retención de 8.432 min. (Figura 6). Sin embargo, al sobreponer los
cromatogramas obtenidos con dos diluciones del estándar y una muestra del
extracto natural, se pudo confirmar la presencia trans β-caroteno en D. salina tal y
como se puede ver en la Figura 7.
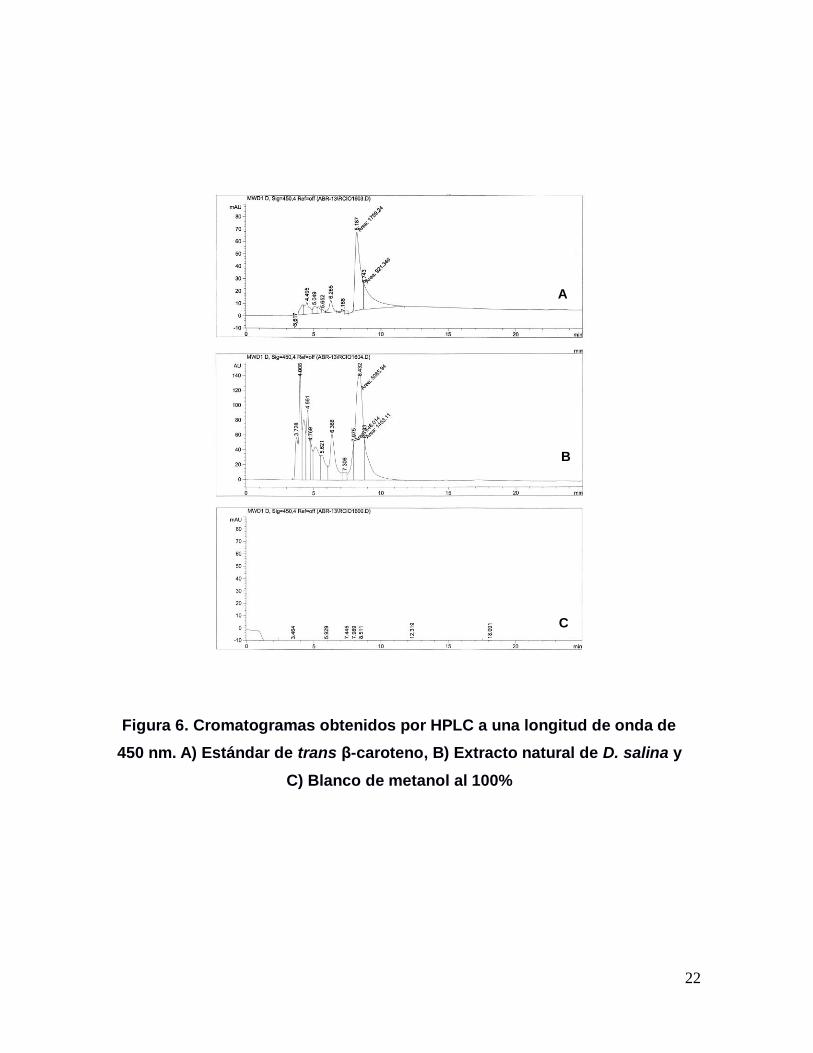
22
Figura 6. Cromatogramas obtenidos por HPLC a una longitud de onda de
450 nm. A) Estándar de trans β-caroteno, B) Extracto natural de D. salina y
C) Blanco de metanol al 100%
A
B
C
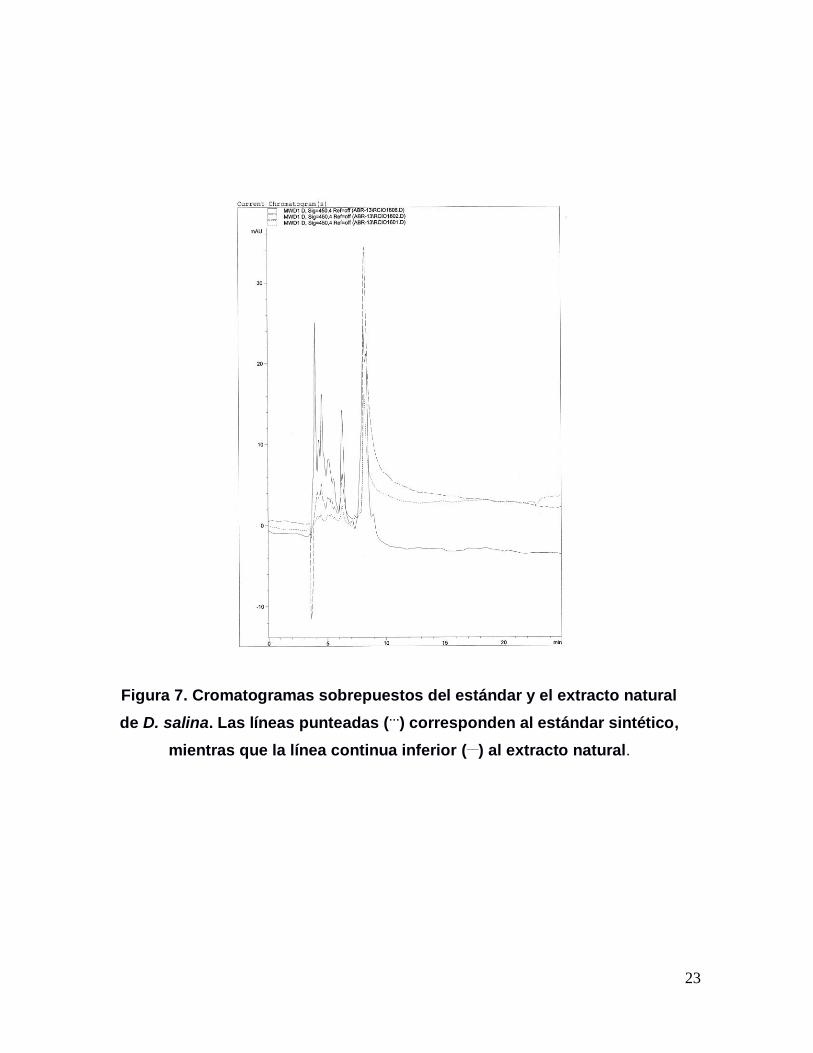
23
Figura 7. Cromatogramas sobrepuestos del estándar y el extracto natural
de D. salina. Las líneas punteadas (…) corresponden al estándar sintético,
mientras que la línea continua inferior (__) al extracto natural.
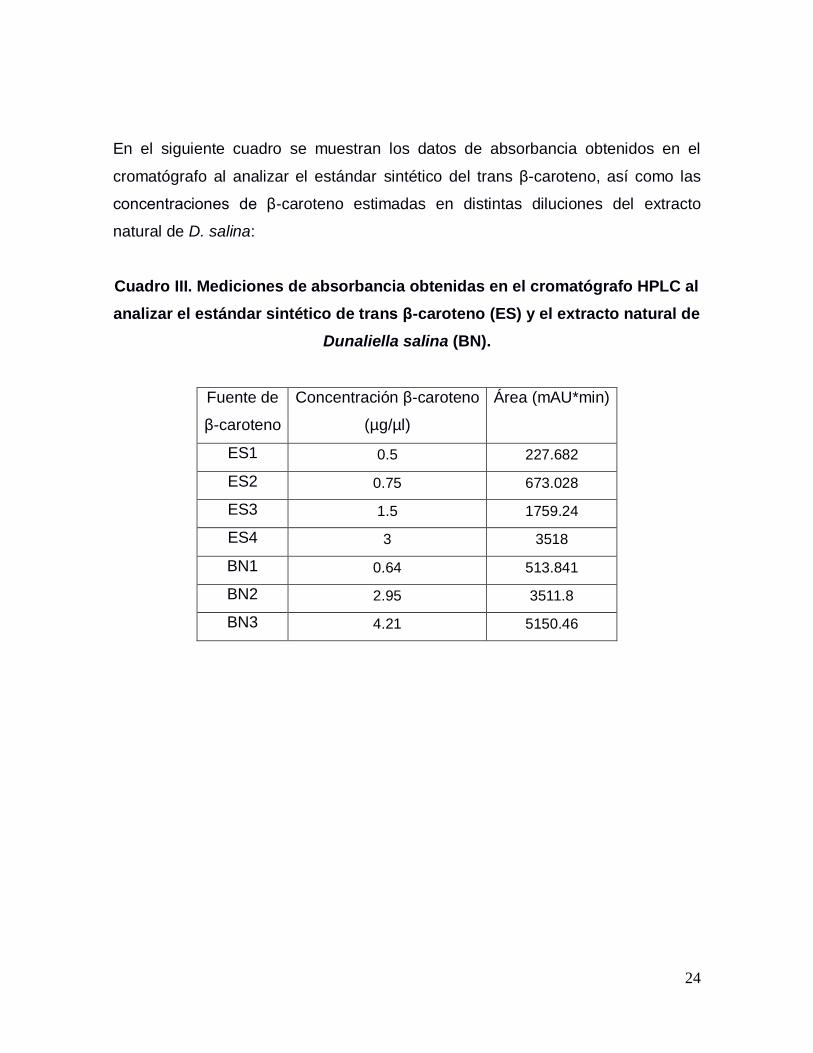
24
En el siguiente cuadro se muestran los datos de absorbancia obtenidos en el
cromatógrafo al analizar el estándar sintético del trans β-caroteno, así como las
concentraciones de β-caroteno estimadas en distintas diluciones del extracto
natural de D. salina:
Cuadro III. Mediciones de absorbancia obtenidas en el cromatógrafo HPLC al
analizar el estándar sintético de trans β-caroteno (ES) y el extracto natural de
Dunaliella salina (BN).
Fuente de
β-caroteno
Concentración β-caroteno
(µg/µl)
Área (mAU*min)
ES1 0.5 227.682
ES2 0.75 673.028
ES3 1.5 1759.24
ES4 3 3518
BN1 0.64 513.841
BN2 2.95 3511.8
BN3 4.21 5150.46
25
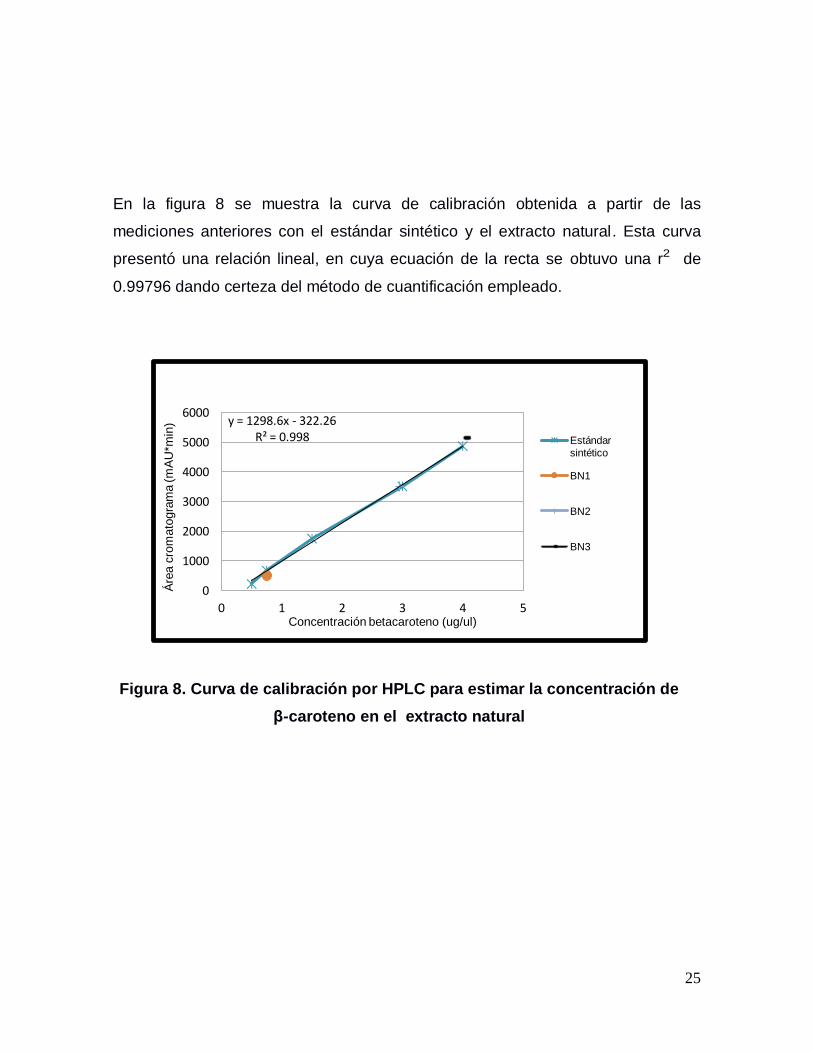
En la figura 8 se muestra la curva de calibración obtenida a partir de las
mediciones anteriores con el estándar sintético y el extracto natural. Esta curva
presentó una relación lineal, en cuya ecuación de la recta se obtuvo una r2 de
0.99796 dando certeza del método de cuantificación empleado.
Figura 8. Curva de calibración por HPLC para estimar la concentración de
β-caroteno en el extracto natural
y = 1298.6x - 322.26 R² = 0.998
0 1 2 3 4 5
0
1000
2000
3000
4000
5000
6000
Concentración betacaroteno (ug/ul)
Áre
a c
rom
ato
gra
ma (m
AU
*min
)
Estándar
sintético
BN1
BN2
BN3

26
7.3 Curva de crecimiento
Al realizar la curva, se determinó que la cantidad óptima de células para realizar
los ensayos fue de 5,000 células debido a las confluencias iniciales y finales
obtenidas, como se puede observar en el siguiente cuadro:
Cuadro IV. Resultados obtenidos para la curva de crecimiento con la línea
celular MDA-MB-231.
Células/ml Confluencia 4h Confluencia 20h
MDA-MB-231
1000 10% 20%
2000 15-20% 35%
3000 20-25% 50%
4000 30-35% 65-70%
5000 40-45% 85-90%
7.4 Ensayo de viabilidad celular con β-caroteno
Al analizar estadísticamente los resultados obtenidos, los datos presentaron en
todos los casos una distribución normal, al realizar intervalos de confianza para un
α de 0.05.
7.4.1 Tratamiento de β-caroteno: 2 horas
En la línea celular MDA-MB-231 el β-caroteno natural tuvo un fuerte efecto, ya que
la supervivencia fue del 27.9%, contra una supervivencia del 64.45% originado por
el trans β-caroteno sintético (Figura 9). Al realizar el ANDEVA se obtuvo una F=

27
1148.3 con un valor de P=4.45e-6 (P <0.05). Esto demostró que la diferencia fue
estadísticamente significativa entre tratamientos (Ver Figura 11 en Anexo 5). Al
comparar contra el control interno de la propia línea celular MDA-MB-231 (control
negativo) nuevamente existió diferencia significativa. Además la diferencia más
grande se presentó en las células tratadas con β-caroteno natural.
En la línea celular HaCat, la supervivencia fue de un 96.04% y de un 89.13% al
ser tratadas con el β-caroteno natural y sintético respectivamente (Figura 10). El
ANDEVA realizado indicó que no existió una diferencia significativa entre la
supervivencia de las células HaCat tratadas con ambas fuentes de β-caroteno
(Figura 12).
Por medio de una cuarta comparación entre ambas líneas celulares (MDA-MB-231
y HaCat) tratadas con β-caroteno (natural y sintético) durante 24h, se tuvo una
F=55.526 con una P= 1.065e-6 (P<0.05); por lo que nuevamente se encontró una
diferencia estadísticamente significativa. Esto se puede observar con mayor
detenimiento en la Figura 13, en donde se ve una marcada diferencia entre grupos
y en el caso de MDA-MB-231 entre tratamientos.
7. 4. 2 Tratamiento de β-caroteno: 24 horas
En las células de MDA-MB-231 que fueron sometidas al de β-caroteno natural
durante 24h, hubo una supervivencia del 70.98%; mientras que al ser tratadas con
el de origen sintético fue de un 96.74% (Figura 9). Al realizar el ANDEVA se obtuvo
una F=474.32 con un valor de P=2.63e-6 (P<0.05), por lo que existió una
diferencia entre ambos tratamientos (Ver figura 14 en Anexo V). Al comparar
nuevamente contra el control interno de la línea celular, nuevamente existió
diferencia significativa, y la diferencia más grande se presentó entre las células
MDA-MB-231 control interno y las tratadas con β-caroteno natural, corroborando el

28
resultado anterior (Ver Figura 17).
En la línea celular HaCat la supervivencia fue de un 79.71% para el β-caroteno
natural y 83.3% para el sintético durante 24 horas (Figura 10). El ANDEVA indicó
que no existió una diferencia significativa entre ambas condiciones (Figura 15).
Sin embrago, al realizar una cuarta prueba considerando a las células de control
interno para HaCat, se encontró diferencia significativa con un valor de F=20.878
con una P=0.00019 (P< 0.05). En este caso la mayor variación se pudo observar
entre las células del control negativo y las células tratadas con β-caroteno
sintético.
Al comparar los resultados de ambas líneas celulares (MDA-MB-231) tratadas con
ambos tipos de β-caroteno por 24 horas, nuevamente se encontró una diferencia
entre los tratamientos. Se obtuvo una F=30.198 con un valor de P=0.001 (P<0.05).
Ver Figura 16, en el Anexo V.
Finalmente, de acuerdo a los resultados obtenidos en ambos tratamientos con β-
caroteno (2 y 24 horas), se acepta la hipótesis planteada en este trabajo.
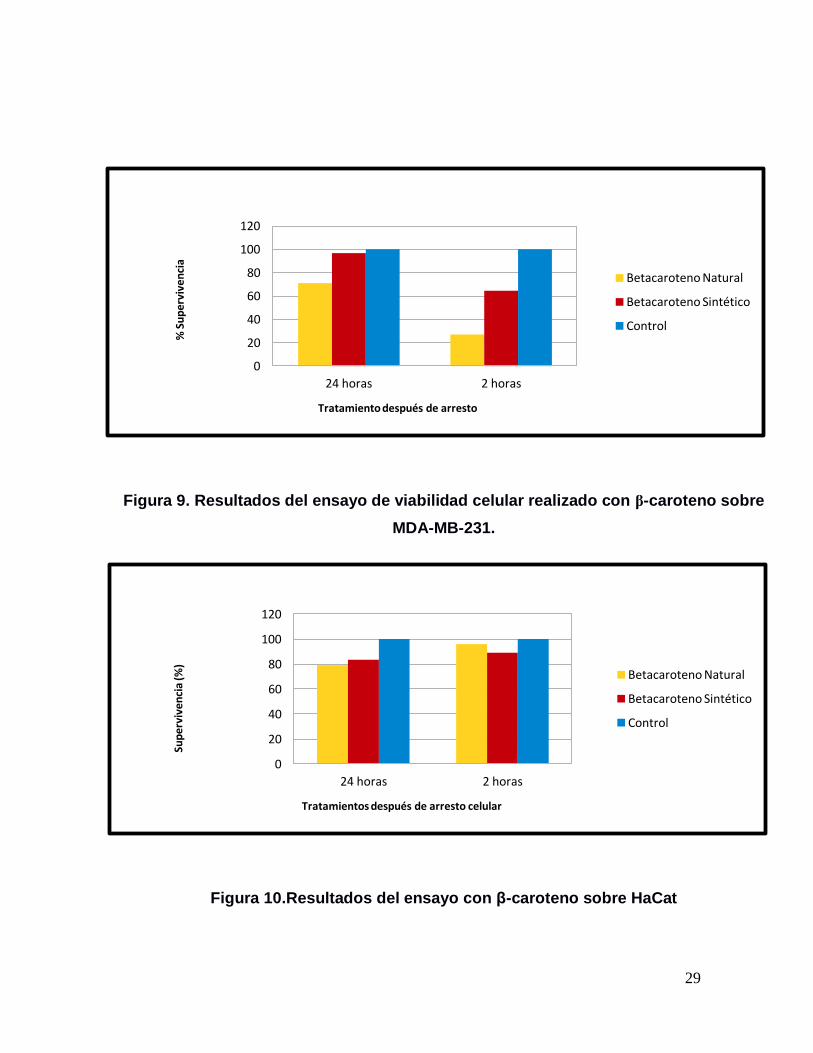
29
Figura 9. Resultados del ensayo de viabilidad celular realizado con β-caroteno sobre
MDA-MB-231.
Figura 10.Resultados del ensayo con β-caroteno sobre HaCat
24 horas 2 horas
0
20
40
60
80
100
120
Tratamiento después de arresto
% S
up
ervi
ven
cia
Βetacaroteno Natural
Betacaroteno Sintético
Control
24 horas 2 horas
0
20
40
60
80
100
120
Tratamientos después de arresto celular
Sup
ervi
ven
cia
(%)
Βetacaroteno Natural
Betacaroteno Sintético
Control

30
8 Discusión
Los resultados obtenidos en el ensayo mostraron un claro efecto del β-caroteno de
origen natural sobre las células de cáncer de mama MDA-MB-231, y que además
fue significativamente mayor con respecto al efecto que causó el trans β-caroteno
de origen sintético. Con base en este resultado podríamos platearnos la siguiente
pregunta: ¿De qué forma puede interactuar la molécula de β-caroteno con las
células cancerígenas para causarles una disminución de su supervivencia? Una
forma podría ser por medio de su actividad antioxidante (Dimitrov et al., 1988;
Bendich y Olson, 1989; Mínguez-Mosquera et al., 2008).
En 1924 se descubrió que las células cancerígenas presentan una alteración en el
metabolismo celular, llamado efecto Warburg y que las distingue de las células
normales. Debido a esta alteración las células de cáncer llevan a cabo la gilcólisis
anaeróbica (convirtiendo la glucosa en lactato) para obtener su energía y realizar
sus actividades vitales (Lehninger, 1995; Dang, 2012). Sin embargo, este proceso
es menos eficiente, de forma que las mitocondrias de las células cancerígenas
deben trabajar más y por ende tienen una mayor actividad. Cada vez que las
mitocondrias llevan a cabo sus actividades nuevamente liberan especies reactivas
de oxígeno -1O2 (ROS); por lo que en donde hay células cancerígenas se forma un
microambiente químico en el que hay condiciones de hipoxia y una mayor cantidad
de ROS (Dang, 2012).
Las ROS tienen un papel importante durante el desarrollo del cáncer, activando o
inhibiendo la actividad de distintos proto-oncogenes y genes supresores de
tumores. Un caso es el del oncogen Myc, que en presencia de ROS es sobre-
expresado en las células de cáncer. Myc es un regulador central del crecimiento

31
celular, de forma que un fallo es un su funcionamiento acelera el metabolismo, y la
proliferación celular. Otro oncogen, que se ve afectado por la presencia de ROS es
el Factor inducible a la hipoxia (HIF-1); este es un factor trans-cripsional que
induce continuamente la activación de la glicólisis anaerobia (efecto Warburg) y
tiene una sobre actividad metabólica, que las prepara para poder proliferar
continuamente (Dang, 2012). También se sabe que las ROS disminuyen la
expresión de distintos genes supresores de tumores, entre los cuales se encuentra
el gen que codifica la Tensin-fosfatasa homóloga (PTEN). La expresión de esta
enzima, detiene la proliferación celular. Un gen supresor de tumores que también
se ve afectado por la presencia de ROS en células cancerígenas es P53 (Lau et
al., 2011; Fiachi y Chiarugui, 2012).
Como ha sido ampliamente descrito, la molécula de β-caroteno posee un largo
esqueleto poli-insaturado, es decir que cuenta con muchos dobles enlaces,
situados en distintos ángulos y posiciones geométricas, lo que le hace capaz (al
oxidarse) de atrapar y destruir, moléculas altamente reactivas, como las especies
reactivas de oxígeno activado (Holum, 1999). Es por eso, que una actividad
antioxidante que atrape ROS por parte del β-caroteno pudo disminuir la
supervivencia de las células MDA-MB-231 en este trabajo, al dañar directamente
su metabolismo, y alterar señalizaciones que les permiten evadir la apoptosis e
inducir la proliferación celular; ya sea que altere las condiciones de hipoxia y
modifique el ambiente químico externo en el que se encuentran las células o bien
que actúe dentro de ellas al ser asimilado.
Por otro lado, Minguez-Mosquera (2008) además de otros autores, mencionan que
los carotenoides pueden, tener una actividad anticancerígena sin actuar como
antioxidantes; interactuando directamente con receptores celulares que se
encuentran embebidos en las membranas celulares (Bendich y Olson, 1989;

32
Lamson y Brignall, 1999; Heber y Lu, 2002. A su vez, esos receptores celulares
podrían intervenir en la señalización para la progresión del ciclo celular, evitando la
proliferación característica del cáncer (Heber y Lu, 2002). Si esto ocurrió en el
ensayo realizado, uno de los receptores involucrados podrían ser Cadherina-11 y
E-cadherina.
E-cadherina es una proteína de membrana de importancia fundamental para el
mantenimiento y la formación de tejidos; ya que junto con otras proteínas
conforman las uniones intercelulares (uniones GAP). Cadherina-11, es un receptor
celular que se expresa en las células de MDA-MB-231 una vez que estas han
sufrido un proceso epigenetico, y jugando un importante papel en el desarrollo de
la metástasis (Onder et. al., 2008; Benton et al., 2009; Lau et al., 2011). Una
diferencia a nivel de receptores como estos también podría explicar, porque el β-
caroteno tuvo un efecto sobre las células cancerígenas de mama y no en las
células HaCat. Además se sabe, que la expresión de Cadherina-11 en células de
cáncer obedece a un aumento en su proliferación (Qureshi et al., 2006), pudiendo
explicar porque el efecto del β-caroteno fue mayor en el ensayo de 2 horas, ya que
en este caso la confluencia celular era cercana al 100%.
La diferencia que existió en el efecto causado con respecto al origen de la
molécula de β-caroteno se debe a que D. salina produce de forma natural, una
mezcla de isómeros: 15-cis-β-caroteno (10%), 9-cis-β-caroteno (41%), trans β-
caroteno (42%) y además de otros isómeros sin identificar (7%). Esta combinación
de isómeros hace que sean más fácilmente asimilados y metabolizado por las
células humanas que las fuentes sintéticas de tipo trans (Ben-Amotz, 1992); Esto
también podría explicar porque tuvo un efecto mayor. Además, aunque
actualmente la producción comercial de carotenoides es en su mayoría llevada a
cabo por síntesis química con alto grado de pureza y bajo costo; en ella a menudo

33
quedan algunos precursores de las reacciones o -productos no biológicos que
pueden tener efectos no deseados (Asker et al., 2012). Por ello, en este trabajo se
consideró importante utilizar una fuente sintética de β-caroteno y comparar su
efecto contra uno de origen natural.
Cualquiera que haya sido el mecanismo de acción del β-caroteno, la actividad
anticancerígena reportada en este trabajo coincide con lo ya expuesto por Bendich
y Olson (1989); Lamson y colaboradores (1989), Venugocopal (2009) y Ben-Amotz
(2009) entre otros en distintos tipos de cáncer.

34
9 Conclusiones
-Fue posible inducir la β-carotenogénesis en D. salina por medio de estrés salino
bajo condiciones de laboratorio.
-El β-caroteno obtenido a partir de D. salina tuvo in vitro una actividad negativa en
la supervivencia de las células cancerígenas MDA-MB-231 en todos los
tratamientos.
-El β-caroteno trans sintético afectó en menor grado la supervivencia de las
células cancerígenas MDA-MB-231 en todos los tratamientos.
-En células normales HaCat el β-caroteno sintético tuvo un mayor efecto negativo
en la supervivencia, con respecto al de origen natural en el tratamiento de 2
horas.
-En células normales HaCat tratadas durante 24 horas con el β-caroteno natural la
diferencia fue mínima.
-El β-caroteno de origen natural obtenido a partir de D. salina puede ser un buen
candidato para la prevención y el tratamiento del cáncer de mama.

35
10 Recomendaciones
-Optimizar las condiciones de cultivo y de estrés para D. salina.
-Obtener cultivos masivos de D. salina para lograr una mayor producción de β-
caroteno en menos tiempo.
-Confirmar el efecto del β-caroteno producido naturalmente por D. salina en otras
líneas celulares de cáncer.
-Estudiar los mecanismos mediante los cuales el β-caroteno de origen natural
puede interactuar con las células de cáncer..
-Estudiar las rutas metabólicas que inducen la β-carotenogénesis en D. salina
desde un punto de vista genético, con el fin de generar cepas super-
productoras del pigmento.

36
Bibliografía
Asker, D., Awad, T., Beppu, T. y K. Ueda. 2012. Isolation, characterization, and
diversity of novel radiotolerant carotenoid-producing bacteria. En: Microbial
Crotenoids from Bacteria and Michroalgae. Methods and Protocols. Barredo, J.
(Ed). Human Press. Estados Unidos. 355 pp.
Ames y Shigenaga. 1992. Molecular Biology of free Radicals scavenging systems.
Cold Spring harbor. Estados Unidos. 128 pp.
Baba, A. I. 2007. Comparative Oncology. The publishing house of the Rumanian
Academy. Rumania. 782 pp.
Ben-Amotz, A. y M. Avron. 1992. Dunaliella: Physiology, Biochemistry and
Biotechnology. CRC Press. Estados Unidos. 240 pp.
Ben-Amotz, A. y Y. Levi. 1996. Bioavailabity of a natural isomer mixture compared
with synthetic all trans β-carotene in human serum. Am. J. Clin. Nutr. 63: 729-
34.
Ben-Amotz, A., L. Amnon, y M. Avron. 1987. Steroisomers of β-carotene and
Phytoene in the Alga Dunaliella bardawill. Plant Physiol. 86: 1286-1291.
Ben-Amotz, A.,, J. Polle, y D.V. Subba-Rao. 2009. The Alga Dunaliella
Biodiversity, Physiology and Biotecnology. CRC Press. Estados Unidos. 550
pp.
Bendich, A. y J. Olson. 1989. Biological actions of carotenoids. FASEB J., 3(8):
1927-1932.
Benton, G.; Crooke, E. y J. George. 2009. Laminin-1 induces E-cadherin
expression in 3-dimensional cultured breast cancer cells by inhibiting DNA
methyltransferase 1 and reversing promoter methylation status. FASEB 23:
3884-3893.
Borowitzka, M. 1990. The mass culture of Dunaliella salina, pp. 63-80. En:

37
Technical resource papers. Regional workshop on the culture and utilization of
seaweeds 2. Regional seafarming development and demostration project,
FAO Network of Aquaculture Centres in Asia: Bangkok, Thailand.
Capa-Robles, W. 2004. Caracterización de vías metabólicas de formación de
isoprenoides y beta-caroteno durante la carotenogénesis de D. salina BC02.
Tesis de maestría. CICESE. 86 pp.
Cailleau, R.; Young, R.; Olivé, M. y J. Reeves. 1974. Breast tumor cell lines form
pleural efussions. J. Natl. Cancer Inst 53: 661-674.
Contreras-Flores, R. 2005. Identificación molecular de cepas de Dunaliella salina,
evaluación de la acumulación de β-caroteno y determinación de la variabilidad
genética intraespecie. Tesis de maestría. CICESE. 71 pp.
Dang, C. 2013. Links between metabolism and cáncer. Genes Dev. 26: 877-890.
Dimitrov, N., C. Meyer,, D. Ullrey,; W. Chenowerth,, W. Malone,, C.Boone y G. Fink.
1988. Bioavility of B-carotene in humans. Am. J. Clin. Nutr. 48: 298-304.
Emeish, S. 2012. Production of natural β-carotene from Dunaliella living in the
Dead Sea. JJEES 4(2): 23-27.
Fiaschi, T. y P. Chiarugi. 2012. Oxidative stress, tumor microenviroment, and
metabolic reprograming: A diabolic liaison. International Journal of Cell
Biology 2012: 1-8.
Guillard, R. L. y J. Rither. 1962. Studies on marine planktonic diatoms. Can. J.
Microbiol. 8: 229-239.
Gutierrez-Millán, L. 1996. Ácido desoxirribonucleico, proteínas y pigmentos
durante la carotenogénesis inducida de una nueva cepa de Dunaliella salina
(Chlorophyta) aislada de una laguna salina hipersalina de Baja California.
Tesis de maestría. CICESE. 82 pp.
Heber, D. y Q.Y. Lu. 2002. Overview of mechanism of action of Lycopene. Exp.
Biol. Med. 227: 920-923.
Herchberg, S. 2005. The history of β-caroteno and cancers: from observational to

38
intervention studies. What lessons can be drawn for future research on
polyphenols?. Am. J. Clin. Nutr. 81: 218-222.
Holum, J. 1999. Fundamentos de química general, orgánica y bioquímica para
ciencias de la salud. LIMUSA-Colección de textos del IPN. México. 865 pp.
Ihaka R. y R. Gentleman. 1996. R: a language for data analysis and graphics. J.
Comput Graph. Stat. 5: 299–314.
INEGI. 2013. Estadística a propósito del día mundial del cáncer. En:
http://www.inegi.con,mx (consultado el 9 de junio de 2013).
INC. 2013. Instituto Nacional del Cáncer de Estados Unidos. En:
http://www.cancer.gov/ (consultado el 14 de mayo de 2013).
Ireland, C.M.; Copp, B.; Foster, M.; Macdonald, L.; Radisky, D. and J.C. Swerser.
1993. En: Marine Biotechnology, Volume 1: Pharmeceutical and bioactive
Natural Products. Haltaway, H. y O. Zabosrky (Eds). P. 1-11.
Kowalski, P.J.; Rubin, M.A. y C. Kleer. 2003. E-cadherin expression in primary
carcinomas of the breast and its distant metastasis. Breast Cancer Reserch.
5: R217-R222.
Krinsky, N. 1988. Antioxidant functions of carotenoids. Free Radical Biology and
Medicine 7: 617-635.
Lamson, D.W. y M. S. Brignall. 1999. Antioxidants in Cancer therapy; their actions
and interactions with oncologic therapies. Alt. Med. Rev., 4(5): 304-329.
Langdon, S. 2004. Cancer cell culture. Humana Press. Estados Unidos. 361 pp.
Lehmann, B. 1997. HaCat cell line as a model system for Vitamin D3 metabolism
in human skin.J. Inv. Derm. 108: 72-82.Lau, M.; Klausen, C. y P. Leung. 2011.
E-cadherin inhibits tumor cell growth by suppressing PI3K/Akt signaling via β-
catenin-Egr-1mediated PTEN expression. Oncogene: 1-14
Leo, A. y C. Lieber. 1999. Alcohol, vitamin A, and β-carotene: adverse interactions
including heptotoxity and carcinogenicity. Am. J. Clin. Nutr. 69: 1071-85.
Lehninger, A. 1995. Bioqímica. Omega. España. 1116 pp.

39
Lodish, H.; Matthew, P.; Scott, M.; Matsudaira, P.; Darnell, J.; Zipursky, L.; Kaiser,
C.; Berk, A. y M. Krieger. 2002. Biología molecular y celular. Panamericana.
México. 863 pp.
Mínguez-Mosquera, M., D. Hornero-Méndez, y A. Pérez-Gálvez. 2008.
Carotenoids and provitamin A in functional foods. En: Methods of analysis for
functional food and nutraceuticals. (Ed.) W.J. Hurst, CRC Press. Estados
Unidos. 532 pp.
Musolino, A.; Bella, A.; Bortesi, B.; Michiaria, M.; Naldi, N.; Zanelli, P.; Neri, T. Y A.
Ardizzoni. 2007. BRCA mutations, molecular markers and clinical variables in
early one set breast cancer. The Breast, 16: 280-292.
Olmos-Soto, J., J. Paniagua-Michel, R. Contreras y L. Trujillo. 2002. Molecular
identification of β-carotene hyper-producing strains of Dunaliella from saline
environments using species-specific oligonucleotides. Biotechnol. Lett. 24:
365-369.
Olmos-Soto, J.; Paniagua-Michel, J.; Contreras, R. y L. Ochoa. 2012. DNA
Fingerpringting Intron Method to acomplish specific, rapid, and sensitive
identification of Carotenogenic Dunaliella especies, pp. . En: Microbial
Carotenoids from Bacteria and Michroalgae. Methods and Protocols. Bárredo
(Eds). Human Press. Estados Unidos.
Onder, T.; Gupta, P. y S. Mani. 2008. Loss of E-cadherin promotes metástasis vía
multiple downstream transcriptional pathways. Can. Res 68: 3648-3654.
Paniagua-Michel, J. 2009. Productos naturales marinos, metabolitos con actividad
biológica, pp. 147-186. En: Biotecnología marina. Capítulo 6. (Eds) J.
Paniagua-Michel. AGT-Editor S.A. P.
Pitones-Rubio, S. y J. Olmos-Soto. 2010. Manual técnico de cultivo celular.
Laboratorio de Microbiología Molecular. Centro de Investigación Científica y de
educación superior de Ensenada, B. C. 23 pp.
Qureshi, H., M. Linden,y U. Raju. 2006. E-cadherin status in breast cáncer

40
correlates with histologic type but does not correlate with established
prognostic parameters. Am. J. Clin. Pathol. 125: 377-385.
Rodríguez -Bandala, C. 2010. Identificación del receptor SV2 isoforma en cáncer
de mama in vitro e in vivo. Tesis de Maestría en Ciencias de la Salud.
Escuela Superior de Medicina del Instituto Politécnico Nacional. México. 64 pp.
Sánchez-Castrejón, E. 1998. Ácido desoxirribunocleico, proteína, y pigmentos
durante la carotenogénesis inducida en una nueva cepa de Dunaliella salina
(CLOROPHYTA), aislada de una laguna hipersalina de Baja California,
México. Tesis de maestría. CICESE. 81 pp.
Venugocopal, V. 2009. Marine Products for Healthcare. Functional and
bioactive nutraceutical compounds form the ocean. CRC. Estados Unidos.
525 pp.
Voltolina-Lobina; D; Buckle-Ramírez y E. Morales-Guerrero. 1989. Manual de
metodologías y alternativas para el cultivo de microalgas. Segunda edición.
CICESE. 60 pp.
Wilcox, L. y L. Graham. 2000. Algae. University of Michigan. Prentice Hall.
Estados Undos.640 pp.
World Cancer Report. 2008. World Health Organization. International Agency
for Research on Cancer. Francia. 508 pp.

41
ANEXO I
Cultivo de D. salina
a) Medio nutritivo
Este medio consistió en una combinación de los medios Erdschreiber´s y F/2 en
una proporción 1:1 y a una concentración de 1.0 M de NaCl. Para preparar 1L total
es necesario:
1. Combinar los componentes.
2. Guardar a temperatura ambiente.
Cuadro V. Componentes del medio de cultivo nutritivo
Componente Cantidad
Medio Ersdschraver´s 1.0 M NaCl 500 ml
Medio F/2 1.0 M NaCl 500 ml
b) Medio Erdschreiber´s modificado 1.0 m NaCl.
Para preparar 1 l de medio se debe:
1. Agregar a 3 l de agua de mar (30-35 ppt) pasteurizada, cada uno de los
componentes en el orden especificado con exceptuando las vitaminas.
2. Agitar vigorosamente y vaciar en un recipiente con tapa.
3. Esterilizar en autoclave durante 30 minutos.
4. Cuando el medio esté temperado agregar la solución de vitamina B12.
5. Guardar a 4°C.
42
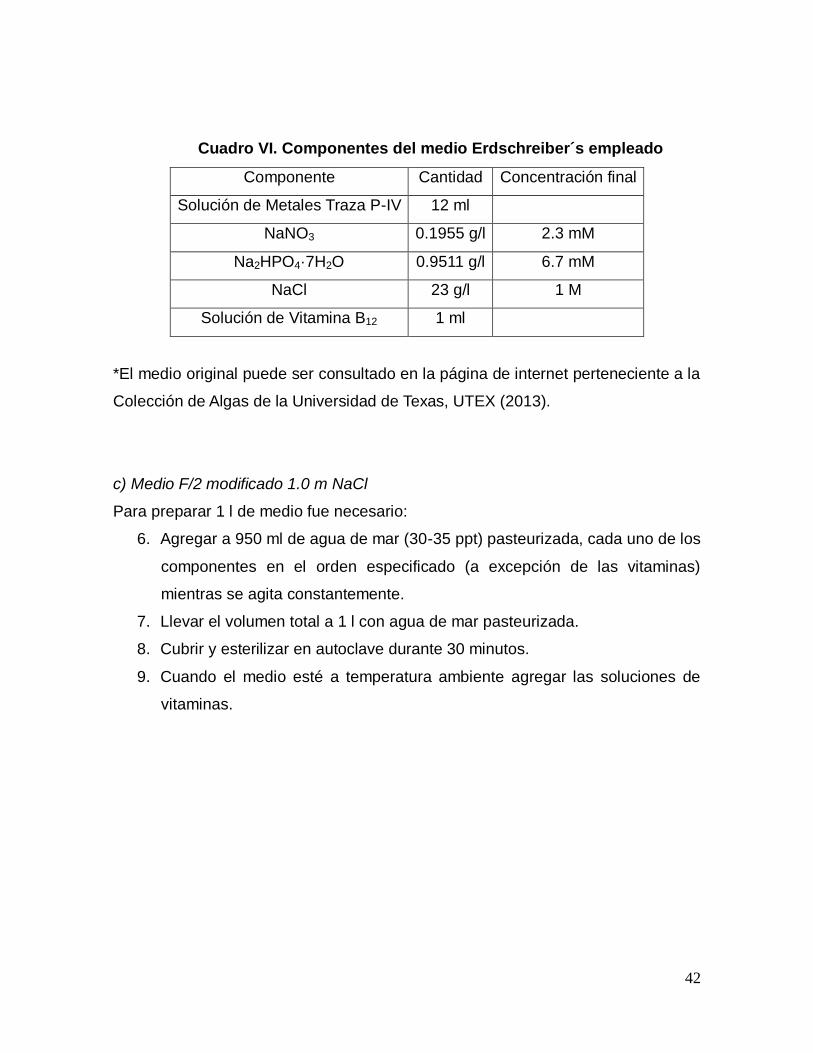
Cuadro VI. Componentes del medio Erdschreiber´s empleado
Componente Cantidad Concentración final
Solución de Metales Traza P-IV 12 ml
NaNO3 0.1955 g/l 2.3 mM
Na2HPO4·7H2O 0.9511 g/l 6.7 mM
NaCl 23 g/l 1 M
Solución de Vitamina B12 1 ml
*El medio original puede ser consultado en la página de internet perteneciente a la
Colección de Algas de la Universidad de Texas, UTEX (2013).
c) Medio F/2 modificado 1.0 m NaCl
Para preparar 1 l de medio fue necesario:
6. Agregar a 950 ml de agua de mar (30-35 ppt) pasteurizada, cada uno de los
componentes en el orden especificado (a excepción de las vitaminas)
mientras se agita constantemente.
7. Llevar el volumen total a 1 l con agua de mar pasteurizada.
8. Cubrir y esterilizar en autoclave durante 30 minutos.
9. Cuando el medio esté a temperatura ambiente agregar las soluciones de
vitaminas.
43
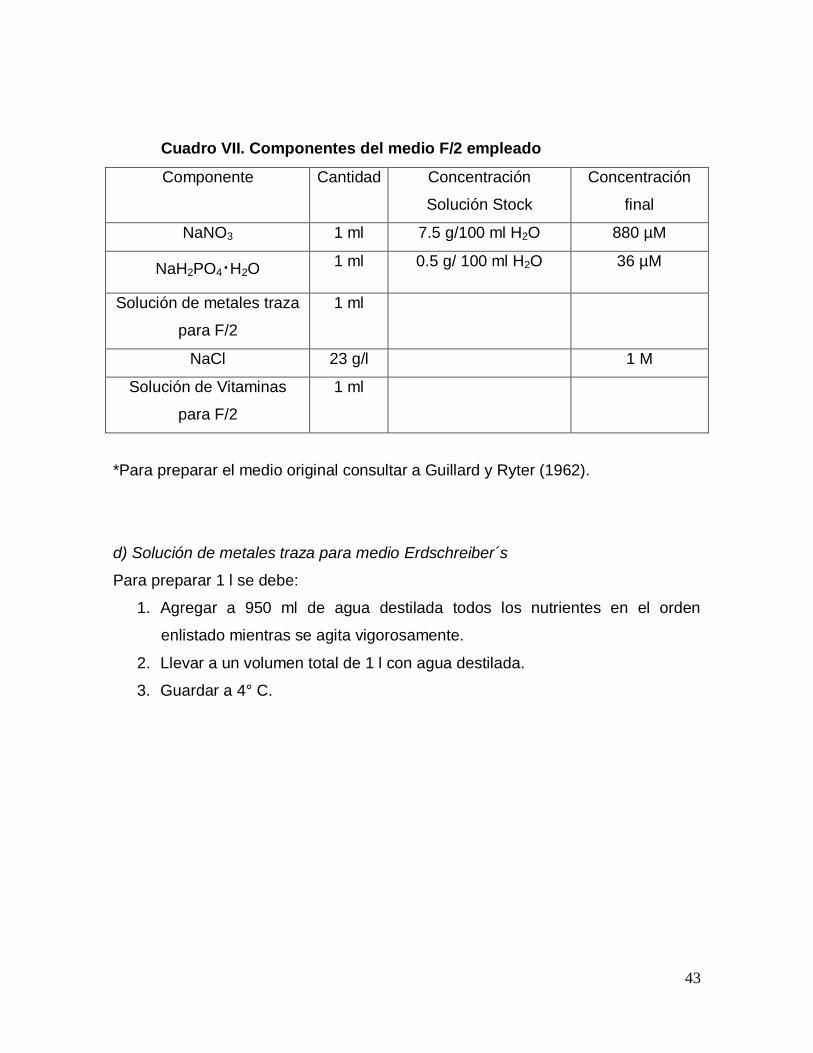
Cuadro VII. Componentes del medio F/2 empleado
Componente Cantidad Concentración
Solución Stock
Concentración
final
NaNO3 1 ml 7.5 g/100 ml H2O 880 µM
NaH2PO4·H2O 1 ml 0.5 g/ 100 ml H2O 36 µM
Solución de metales traza
para F/2
1 ml
NaCl 23 g/l 1 M
Solución de Vitaminas
para F/2
1 ml
*Para preparar el medio original consultar a Guillard y Ryter (1962).
d) Solución de metales traza para medio Erdschreiber´s
Para preparar 1 l se debe:
1. Agregar a 950 ml de agua destilada todos los nutrientes en el orden
enlistado mientras se agita vigorosamente.
2. Llevar a un volumen total de 1 l con agua destilada.
3. Guardar a 4° C.

44
Cuadro VIII. Solución de metales traza para preparar el medio Erdschreiber´s
Componente Cantidad Concentración final
Na2EDTA·2H2O 0.75 g/l 2 mM
FeCl3·6H2O 0.097 g/l 0.36 mM
MnCl2·4H2O 0.041 g/l 0.21 mM
ZnCl2 0.005 g/l 0.037 mM
CoCl2 0.002 g/l 0.0084 mM
Na2MoO4·2 H 2O 0.004 gl 0.018 mM
e) Solución de Vitamina B12 para medio Erdschreiber´s
Para preparar 200 ml es necesario:
1. Prepara 200 ml del amortiguador HEPES (50mM)
2. Ajustar el ph a 7.8.
3. Agregar la Vitamina B12 (0.1 mN) y esperar a que se disuelva por completo.
4. Esterilizarla solución por medio de un filtro Millipore de 0.45 µm.
5. Guardar es oscuridad a 4°C.
Cuadro IX. Vitaminas adicionadas en el medio Erdschreiber´s
Componente Cantidad
Amortiguador HEPES pH 7.8 2.4 g/200 ml H20
Vitamina B12 0.027 g/200 ml H20
f) Solución de metales traza para medio F/2
Para preparar 500 ml se debe:
1. Colocar en un vaso de precipitado 450 ml de agua destilada.
2. Calentar el agua en hasta casi alcanzar el hervor y en agitación constante

45
agregar los componentes.
3. Llevar a un volumen total de 500 ml con agua destilada
Cuadro X. Solución de metales traza para preparar el medio F/2
Componente Cantidad Concentración final
ZnSO4·7H2O 23 mg/500 ml 0.08 µM
MnSO4·H2O 152 mg/500 ml 0.9 µM
Na2MoO4·2H 2O 7.3 mg/500 ml 0.03 µM
CoSO4·7H2O 14 mg/500 ml 0.05 µM
CuCl2·2H 2O 6.8 mg/500 ml 0.04 µM
Fe(NH4)2(SO4)2·6H2O 4.6 mg/500 ml 11.7 µM
Na2EDTA·2H2O 4.4 mg/500 ml 11.6 µM
g) Solución de Vitaminas para medio F/2
Para preparar 200 ml es necesario:
1. Prepara 200 ml del amortiguador HEPES (50mM)
2. Ajustar el ph a 7.8.
3. Agregar los componentes y esperar a que se disuelvan por completo.
4. Esterilizarla por medio de un filtro Millipore de 0.45 µm.
5. Guardar es oscuridad a 4°C.
6.
Cuadro XI. Solución de Vitaminas adicionadas en el medio F/2
Componente Cantidad
Amortiguador HEPES pH 7.8 2.4 g/ 200 ml H20
Vitamina B12 0.027 g/ 200 ml H20
Biotina 0.005 g/ 200 ml H20
Tiamina 0.22 g/ 200 ml H20

46
ANEXO II
Soluciones y medios empleados para el cultivo de las líneas celulares
a) Solución PBS
Para preparar una solución de PBS 10X se deben mezclar en 950 ml de agua
destilada, los componentes mostrados a continuación:
Cuadro XII. Componentes del buffer PBS
Reactivo Cantidad NaCl 8 g
KCl 2 g Na2HPO4 17 mg
KH2PO4 1.63 g
Posteriormente, se debe titular hasta llegar a pH 7.4 con HCl ó NaOH, llevar a 1L
con agua destilada y esterilizar a 15 lb de presión durante 30 minutos en
autoclave.
b) Verseno
Para preparar 60 ml, se debe pesar el EDTA y disolverlo en PBS estéril en
constante agitación.:
Cuadro XIII. Reactivos para preparar EDTA/PBS [1 mM].
Reactivo Cantidad
EDTA 0.0228 g PBS 60 ml
Una vez preparada la solución, esta debe ser esterilizada por medio de un filtro
Millipore de 0.2 µm.

47
c) Suero fetal bovino
Antes de añadir el SFB (GIBCO®) al medio RPMI-1640, este fue inactivado. Para
ello es necesario colocarlo en un baño maría y calentarlo a 55°C durante 20
minutos.
d) Medio RPMI-1640 (Invitrogen ®)
Para preparar 50 ml de medio suplementado es necesario:
1. Tomar 45 ml de medio RPMI y agregar cada uno de los componentes.
2. Filtrar el medio por medio de un filtro Millipore de 0.2 µm con ayuda de una
bomba de vacío.
3. Cerrar y guardarlo a 4°C.
Cuadro XIV. Medio RPMI suplementado.
Componente Cantidad RPMI-1640 45 ml
SFB inactivado 5 ml Ant-Ant 500 µl

48
Anexo III
Ensayo con β-caroteno
a) Solución β-caroteno natural
Para preparar las soluciones de β-caroteno natural:
1. Se mezclaron en un tubo falcon de 15 ml los componentes enumerados en la
siguiente tabla:
Cuadro XV. Solución A elaborada a partir del extracto de D. salina.
Reactivos Cantidades
CH3Cl2 100 μl DMSO 40 μl
Extracto 12 μl EtOH 248 μl
Medio RPMI sin suplementar
2.6 ml
Volumen final 3 ml
2. En otro tubo falcon de 15 ml se colocaron 9 ml de medio RPMI con o sin
suplementar según el caso necesario.
3. Calentó en baño maría a 37° C durante 3 minutos y agregar 1 ml de la mezcla
4. Agregar 1 ml de la solución A preparada.
5. Mantener en baño de hielo y oscuridad, ya que la solución es fotolábil.
b) β-caroteno sintético
Para preparar las soluciones de β-caroteno sintético:
6. Se mezclaron en un tubo falcon de 15 ml los componentes enumerados en la
siguiente tabla.

49
Cuadro XVI. Solución B de β-caroteno trans sintético.
Reactivos Cantidades
CH3Cl2 50 μl DMSO 20 μl
β-caroteno trans 18 mg
EtOH 112 μl Medio RPMI-1640 Sin suplementar
2800
Volumen final 3 ml
7. En otro tubo falcon de 15 ml se colocaron 9 ml de medio RPMI con o sin
suplementar según el caso necesario.
8. Calentó en baño maría a 37° C durante 3 minutos y agregar 1 ml de la mezcla
9. Agregar 1 ml de la solución B preparada.
10. Mantener en baño de hielo y oscuridad, ya que la solución es fotolábil.
* La concentración final de todas las soluciones empleadas en los tratamientos fue
de 6 µg/µl de β-caroteno.

50
Anexo IV
Ensayo colorimétrico de MTT
Para realizar el ensayo se debe:
1. Preparar el reactivo de MTT (Sal de bromuro de 3-(4,5-dimetil-2-tiazolil)-2, 5
difeniltetrazolio) con una concentración de 5mg/ml. Para ello se mezclan en
completa oscuridad los siguientes componentes:
Cuadro XVII. Componentes del Reactivo de MTT.
Componente Cantidad
Sal de MTT 25 mg
PBS estéril 5 ml
2 Con ayuda de una micro-peta agregar 50 ul de de medio RPMI sin suplementar
en cada pozo de la placa en la que se llevó a cabo el ensayo (muestras, blancos y
controles).
3 Enseguida adicionar 50 ul reactivo de la sal de MTT en cada pozo. Este paso
debe hacerse sin presencia de luz blanca.
4 Agitar suavemente la placa para mezclar la sal de MTT con el medio de cultivo.
5 Introducir en la incubadora durante 2 horas y media, a una temperatura de 37°C
y a un 5% CO2.
6 Sacar la placa de la incubadora y con cuidado extraer el sobrenadante de cada
de pozo. Este paso debe de hacerse con sumo cuidado, ya que se debe evitar
quitar los cristales formados en el fondo de cada pozo.
7 Adicionar 100 ul de isopropanol a cada pozo e incubar tres minutos.
8 Agitar suavemente la placa durante 10 minutos, hasta observar que haya una

51
coloración (azul-roja) que sea homogénea.
9 Leer en un micro-lector de placas a una absorbancia de 490 nm.
ANEXO V
“Resultados del ANOVA realizado con el programa R”
1. Comandos empleados en R, y resultados obtenidos en el tratamiento de 2 Horas con β-caroteno para cada línea celular
a) MDA
>y1=c(.249, .258, .264) y>2=c(.107, .108, .109) >y=c(y1,y2) >n=rep(3,2) >group=rep(1:2,n) >data=data.frame(y=y,group=factor(group)) >fit=lm(y ~ group,data) >anova(fit) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 1 0.033302 0.033302 1148.3 4.524e-06 *** Residuals 4 0.000116 0.000029 Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ’ 1
Figura 11. Variación dentro del tratamiento de 2h con β-caroteno en la línea
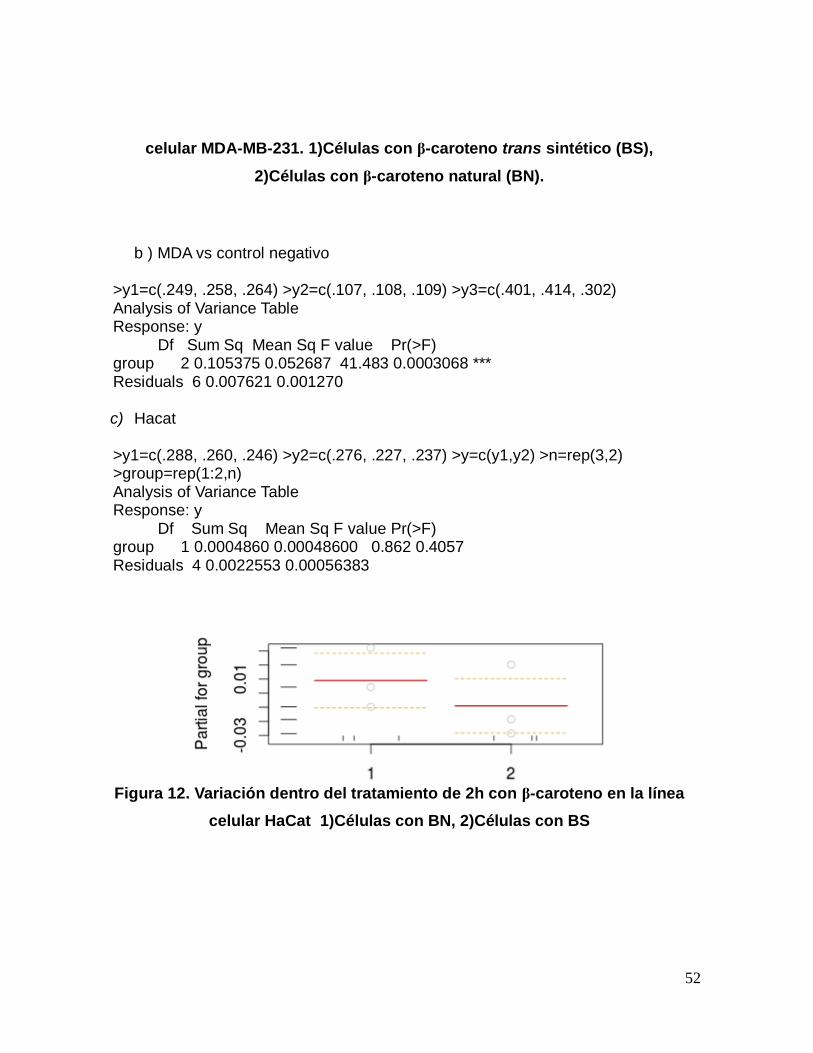
52
celular MDA-MB-231. 1)Células con β-caroteno trans sintético (BS),
2)Células con β-caroteno natural (BN).
b ) MDA vs control negativo
>y1=c(.249, .258, .264) >y2=c(.107, .108, .109) >y3=c(.401, .414, .302) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 2 0.105375 0.052687 41.483 0.0003068 *** Residuals 6 0.007621 0.001270 c) Hacat
>y1=c(.288, .260, .246) >y2=c(.276, .227, .237) >y=c(y1,y2) >n=rep(3,2) >group=rep(1:2,n) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 1 0.0004860 0.00048600 0.862 0.4057 Residuals 4 0.0022553 0.00056383
Figura 12. Variación dentro del tratamiento de 2h con β-caroteno en la línea
celular HaCat 1)Células con BN, 2)Células con BS

53
d) HaCat con control interno
>y1=c(.276, .227, .236) >y2=c(.288, .260, .246) >y3=c(.296, .271, .275) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 2 0.0017709 0.00088544 .0154 0.214 Residuals 6 0.0026360 0.00043933 e) MDA-MB-231 Vs HaCat Beta Natural
>y1=c(.107, .108, .109) >y2=c(.276, .227, .236) >y3=c(y1,y2) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 1 0.0287042 0.0287042 84.259 0.0007822 *** Residuals 4 0.0013627 0.0003407 f) MDA-MB-231 Vs HaCat (Todos los tratamientos)
>y1=c(.249, .258, .264) >y2=c(.107, .108, .109) >y3=c(.276, .227, .236) >y4=c(.288, .260, .246) >y=c(y1,y2,y3,y4) >n=rep(3,4) >group=rep(1:4,n) >data=data.frame(y=y,group=factor(group)) >fit=lm(y ~ group,data) >anova(fit) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 3 0.049793 0.0165976 55.526 1.065e-05 *** Residuals 8 0.002391 0.0002989
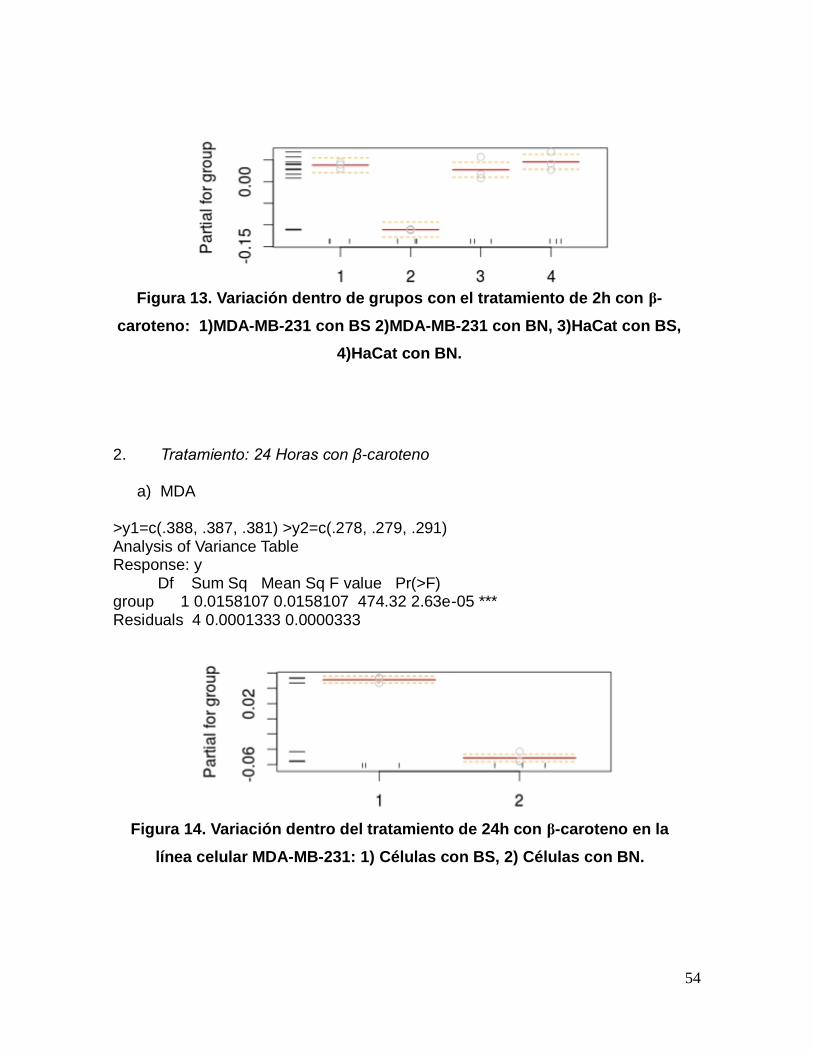
54
Figura 13. Variación dentro de grupos con el tratamiento de 2h con β-
caroteno: 1)MDA-MB-231 con BS 2)MDA-MB-231 con BN, 3)HaCat con BS,
4)HaCat con BN.
2. Tratamiento: 24 Horas con β-caroteno
a) MDA
>y1=c(.388, .387, .381) >y2=c(.278, .279, .291) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 1 0.0158107 0.0158107 474.32 2.63e-05 *** Residuals 4 0.0001333 0.0000333
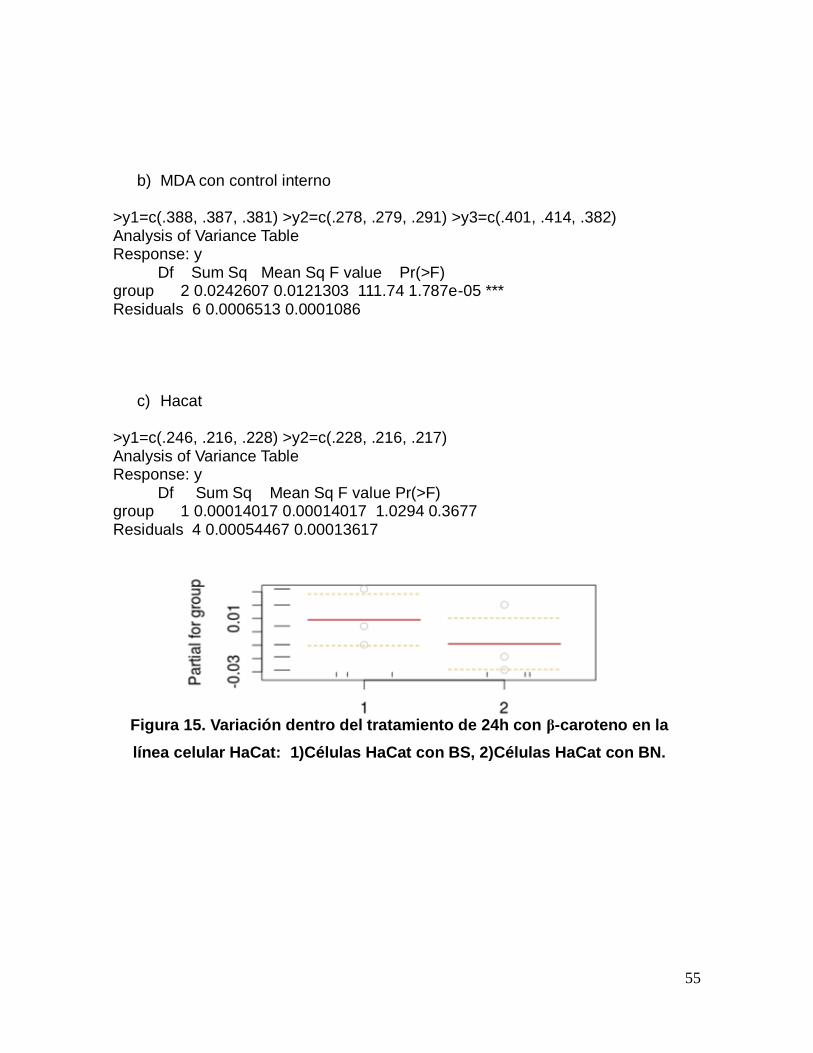
Figura 14. Variación dentro del tratamiento de 24h con β-caroteno en la
línea celular MDA-MB-231: 1) Células con BS, 2) Células con BN.
55
b) MDA con control interno
>y1=c(.388, .387, .381) >y2=c(.278, .279, .291) >y3=c(.401, .414, .382) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 2 0.0242607 0.0121303 111.74 1.787e-05 *** Residuals 6 0.0006513 0.0001086
c) Hacat
>y1=c(.246, .216, .228) >y2=c(.228, .216, .217) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 1 0.00014017 0.00014017 1.0294 0.3677 Residuals 4 0.00054467 0.00013617
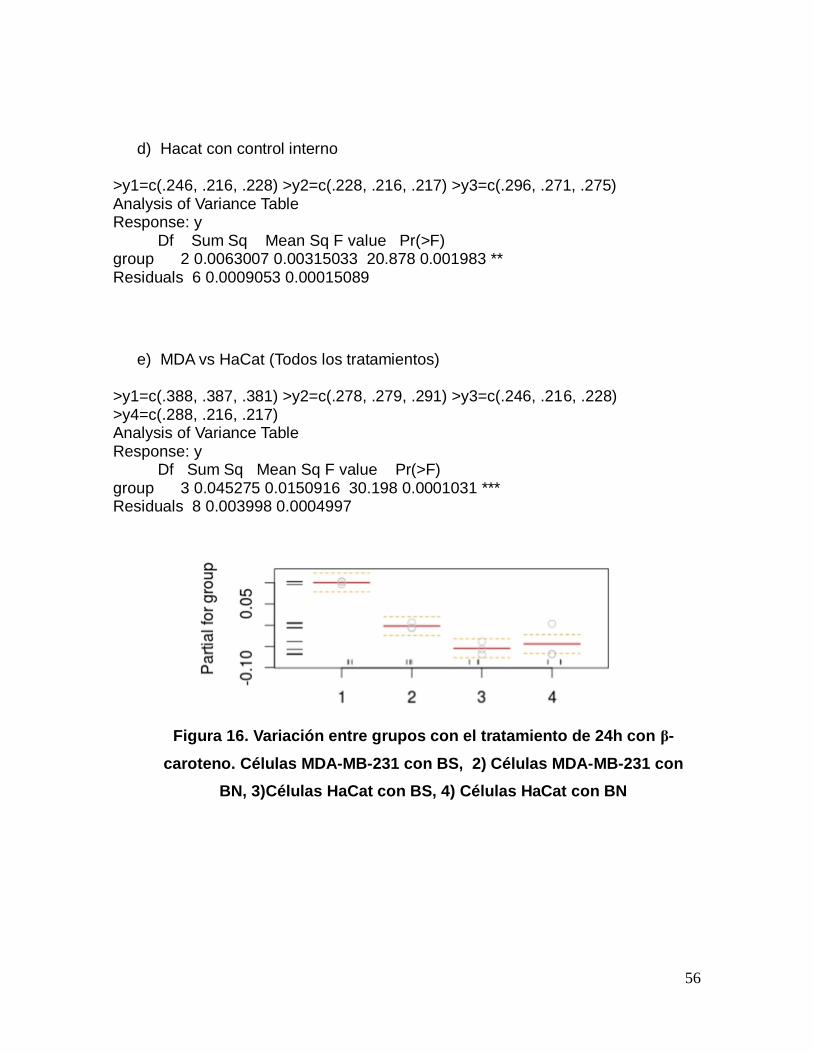
Figura 15. Variación dentro del tratamiento de 24h con β-caroteno en la
línea celular HaCat: 1)Células HaCat con BS, 2)Células HaCat con BN.
56
d) Hacat con control interno
>y1=c(.246, .216, .228) >y2=c(.228, .216, .217) >y3=c(.296, .271, .275) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 2 0.0063007 0.00315033 20.878 0.001983 ** Residuals 6 0.0009053 0.00015089
e) MDA vs HaCat (Todos los tratamientos) >y1=c(.388, .387, .381) >y2=c(.278, .279, .291) >y3=c(.246, .216, .228) >y4=c(.288, .216, .217) Analysis of Variance Table Response: y Df Sum Sq Mean Sq F value Pr(>F) group 3 0.045275 0.0150916 30.198 0.0001031 *** Residuals 8 0.003998 0.0004997
Figura 16. Variación entre grupos con el tratamiento de 24h con β-
caroteno. Células MDA-MB-231 con BS, 2) Células MDA-MB-231 con
BN, 3)Células HaCat con BS, 4) Células HaCat con BN