
Templat tugas akhir S1 Lab Molekuler Ibu Maria Ulfah MSc Agr dan penghuni Zoo Corner yang saya...
20
KERAGAMAN RUAS EKSON 2 DAN 3 GEN PENYANDI ENZIM α-KETO DEHIDROGENASE E1-α RANTAI ASAM AMINO BERCABANG (BCKDHA) PADA SAPI MADURA RIA MARIA DEPARTEMEN BIOLOGI FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR BOGOR 2014
Transcript of Templat tugas akhir S1 Lab Molekuler Ibu Maria Ulfah MSc Agr dan penghuni Zoo Corner yang saya...

KERAGAMAN RUAS EKSON 2 DAN 3 GEN PENYANDI
ENZIM α-KETO DEHIDROGENASE E1-α RANTAI ASAM
AMINO BERCABANG (BCKDHA) PADA SAPI MADURA
RIA MARIA
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2014


PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Keragaman Ruas
Ekson 2 dan 3 Gen Penyandi Enzim α-Keto Dehidrogenase E1-α Rantai Asam
Amino Bercabang (BCKDHA) pada Sapi Madura adalah benar karya saya dengan
arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada
perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya
yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam
teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut
Pertanian Bogor.
Bogor, Juli 2014
Ria Maria
NIM G34090088

ABSTRAK
RIA MARIA. Keragaman Ruas Ekson 2 dan 3 Gen Penyandi Enzim α-Keto
Dehidrogenase E1-α Rantai Asam Amino Bercabang (BCKDHA) pada Sapi
Madura. Dibimbing oleh ACHMAD FARAJALLAH dan DYAH
PERWITASARI.
Sapi madura merupakan sapi lokal Indonesia dari hasil persilangan sapi
india (Bos indicus) dan banteng (Bos javanicus). Kompleks enzim Branched
chain α-keto acid dehydrogenase (BCKDH) terdapat di membran bagian dalam
mitokondria sel eukariot. yang berperan dalam mengubah branched chain amino
acids (BCAAs), yaitu isoleusin, leusin, dan valin ke bentuk α-keto. Kompleks
enzim ini terdiri atas tiga subunit yang salah satunya adalah subunit E1-α yang
disebut branched chain α-keto acid dehydrogenase E1-α (BCKDHA). Maple
syrup urine disease (MSUD) merupakan kelainan genetik yang salah satunya
diakibatkan oleh kejadian mutasi pada subunit E1-. Penelitian ini bertujuan
untuk menganalisis keragaman ruas ekson 2 – 3 gen penyandi enzim BCKDHA
pada sapi madura. Hasil analisis keragaman ruas ekson 2 – 3 gen BCKDHA
ditemukan dalam populasi sapi madura J adalah identik antar individunya. Selain
itu ruas ekson 2 – 3 gen BCKDHA sapi madura juga identik dengan sapi taurin
(Bos taurus).
Kata kunci: BCKDHA, ekson 2 – 3, ruminants, sapi madura.
ABSTRACT
RIA MARIA. Diversity of Segment Exon 2 and 3 encoding genes enzymes
α-Keto dehydrogenase E1-α Branched Chain Amino Acids (BCKDHA) in
Madura Cattle. Supervised by ACHMAD FARAJALLAH and DYAH
PERWITASARI.
Madura cattle is a cattle from the local Indonesian cattle crossbreeding India (Bos
indicus) and banteng (Bos javanicus). Enzyme complex branched chain α-keto
acid dehydrogenase (BCKDH) located in the inner mitochondrial membrane of
eukaryotic cells. BCKDH has a role in changing branched chain amino acids
(BCAAs), which is isoleucine, leucine and valine to α-keto form. This enzyme
complex consists of three enzyme subunits, one which is E1- α subunit called
branched chain α-keto acid dehydrogenase E- α (BCKDHA). Maple syrup urine
disease (MSUD) is a genetic disorder caused by one occurrence of mutation in the
E1- α subunit. This study aimed to determine the diversity of segment 2 – 3 exon
encoding BCKDHA gene in madura cattle. The result revealed that segment of
exon 2 – 3 BCKDHA gene in madura cattle J population were found to be
identical among them. Furthermore the segment of exon 2 – 3 BCKDHA gene
madura cattle J was identical with taurine cattle (Bos taurus).
Keywords: BCKDHA, 2 – 3 exon, ruminants, madura cattle.

Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains
pada
Departemen Biologi
KERAGAMAN RUAS EKSON 2 DAN 3 GEN PENYANDI
ENZIM α-KETO DEHIDROGENASE E1-α RANTAI ASAM
AMINO BERCABANG (BCKDHA) PADA SAPI MADURA
RIA MARIA
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2014


Judul Skripsi: Keragaman Ruas Ekson 2 dan 3 Gen Penyandi Enzim α-Keto
Dehidrogenase E1-α Rantai Asam Amino Bercabang (bckdha)
pada sapi madura Nama : Ria Maria
NIM : G34090088
Disetujui oleh
Dr Ir Achmad Farajallah, MSi
Pembimbing I
Dr Ir R.R. Dyah Perwitasari, MSc
Pembimbing II
Diketahui oleh
Dr Ir Iman Rusmana, MSi
Ketua Departemen
Tanggal Lulus:

PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas
segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang
dipilih dalam penelitian yang dilaksanakan sejak bulan Januari 2013 ini ialah
Genetika Molekuler, dengan judul Keragaman Ruas Ekson 2 dan 3 Gen
BCKDHA (branched chain α-keto acid dehidrogenase E1-α) pada Sapi Madura.
Terima kasih penulis ucapkan kepada Bapak Dr Ir Achmad Farajallah MSi
dan Ibu Dr Ir R.R. Dyah Perwitasari Msc selaku dosen pembimbing yang telah
memberikan arahan dan bimbingannya selama penyelesaian penelitian ini. Terima
kasih juga kepada Prof Dr Ir Alex Hartana selaku dosen penguji atas saran dan
kritiknya dalam penyelesaian karya ilmiah ini. Ungkapan terima kasih tak
terhingga disampaikan kepada ayahanda Achmad Bakrie dan Ibunda Penih beserta
keluarga atas segala doa, dukungan dan kasih sayangnya yang memberi semangat
kepada penulis. Di samping itu, penghargaan penulis sampaikan kepada segenap
penghuni Lab Molekuler Ibu Maria Ulfah MSc Agr dan penghuni Zoo Corner
yang saya hormati. Terima kasih juga saya ucapkan kepada teman-teman tercinta
Kurrataa’yun, Eci, Elok, Dewi, Yovita dan keluarga Biologi 46 yang terus
memberikan saya semangat. Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2014
Ria Maria

DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
METODE 3
Sampel DNA 3
Amplifikasi dan Visualisai DNA 3
Perunutan Produk PCR 3
Analisis Data 3
HASIL DAN PEMBAHASAN 4
Hasil 4
Amplifikasi Ruas Ekson 2 – 3 gen BCKDHA 4
Analisis Varian Nukleotida Ruas Ekson 2 – 3 Gen BCKDHA 4
Pembahasan 5
SIMPULAN 6
DAFTAR PUSTAKA 6
LAMPIRAN 8
RIWAYAT HIDUP 10

DAFTAR TABEL
1 Ukuran ruas ekson 2 – 3 gen BCKDHA pada sapi madura J dan spesies
pembanding. 5
2 Variasi nukleotida ekson 2 – 3 gen BCKDHA dari sampel sapi madura
J terhadap nukleotida gen BCKDHA spesies pembanding. 5
DAFTAR GAMBAR
1 Lokasi beberapa kompleks enzim di lintasan metabolisme intermediet
karbohidrat, lemak dan asam amino sejak di sitoplasma sampai matriks
mitokondria. 1
2 Amplikon ruas ekson 2 – 3 gen BCKDHA diatas gel poliakrilamida 6%
setelah pewarnaan perak. 4 3 Kromatogram menunjukkan puncak-puncak tunggal pada hasil sekuens. 4
DAFTAR LAMPIRAN
1 Gambar sapi madura 8
2 Database DNA polled samples 8
3 Posisi penempelan primer AF318 dan AF319 pada peta organisasi gen
BCKDHA sapi Bos taurus (NW001493616) 9
4 Posisi penempelan primer AF318 dan AF319 9

PENDAHULUAN
Latar Belakang
Indonesia memiliki keanekaragaman sumber daya genetik hewan ternak lokal
yang tinggi, terutama sapi. Sapi pada umumnya dimanfaatkan manusia sebagai
penyedia kebutuhan protein hewani. Jenis sapi lokal yang diternakan di Indonesia
salah satunya adalah sapi madura. Sapi madura merupakan hasil persilangan dari sapi
zebu (Bos indicus) dan banteng (Bos. javanicus) (Nijman et al. 2003). Sapi madura
memiliki ciri kepala bertanduk, gumba berkembang baik pada jantan, warna bulu
merah bata, terdapat lingkaran warna putih pada daerah bawah kaki, bokong dan
moncong (Lampiran 1) (Huitema 1986). Sapi madura tergolong sebagai sapi potong
yang memiliki kualitas daging yang baik, resisten terhadap stres pada lingkungan
panas dan infestasi kutu ternak (Payne dan Hodges 1997). Selain itu, sapi madura juga
sangat efisien terhadap pakan. Pakan yang biasanya digunakan sederhana dari rumput
liar namun kualitas daging dan pertumbuhannya sangat baik. Sapi madura memiliki
kemampuan metabolisme makanan yang sangat baik, oleh karena itu efisiensi
metabolisme dari sapi madura sangat menarik untuk diteliti. Kelainan genetik pada
metabolisme sapi madura juga akan menurunkan kualitas daging dan kesehatan sapi
tersebut.
Metabolisme adalah sekumpulan perubahan kimiawi yang mendukung
kehidupan dalam sel-sel dari organisme hidup termasuk pencernaan dan transportasi
zat ke dalam dan di antara sel yang berbeda. Reaksi enzimatik ini memungkinkan
organisme untuk tumbuh dan berkembang biak, menjaga struktur, dan merespon
lingkungan. Dalam hal ini serangkaian reaksi dalam sel disebut metabolisme perantara
atau metabolisme intermediet. Kompleks enzim branched chain α-keto-acid
dehydrogenase (BCKD) merupakan kompleks enzim yang berada di membran dalam
mitokondria pada eukariota (Gambar 1) (Hutson 1988).
Keterangan : BCKD - branched-chain α-keto acid dehydrogenase. PDC - pyruvate
dehydrogenase complex. _KGDC -ketoglutarate dehydrogenase complex. AA- asam
amino, FA- asam lemak, TCA cycle - siklus asam sitrat, BCAA - branched-chain
amino acids (Sumber : Patel dan Harris 1995).
Gambar 1 Lokasi beberapa kompleks enzim di lintasan metabolisme intermediet
karbohidrat, lemak dan asam amino sejak di sitoplasma sampai matriks
mitokondria.

2
Kompleks enzim BCKDH sangat conserved pada Mamalia (Yeaman 1986).
Kompleks enzim BCKDH berperan dalam mengubah branched chain amino acids
(BCAAs), yaitu isoleusin, leusin, dan valin menjadi bentuk α-keto sehingga bisa
masuk ke dalam Siklus Kreb. Kompleks enzim BCKD tersusun empat subunit yaitu 2-
oxoisovalerate dehydrogenase (E1-α dan E1-β), dihydrolipoamide acyltransferase
(E2), dan dihydrolipoamide dehydrogenase (E3). Subunit E1-α disandikan oleh gen
branched chain α-keto-acid dehydrogenase E1-α (BCKDHA) (Pettit et al. 1978).
Lokasi gen BCKDHA terletak di kromosom nomor 18 pada sapi taurin (B. taurus)
(Elsik et al. 2009). Subunit E1-α dari kompleks enzim metabolisme BCKD disandikan
oleh gen yang ada di dalam genom inti, ditranskripsi menjadi RNA, ditranslasikan
menjadi polipeptida di sitoplasma dan ditranslokasikan ke matriks mitokondria
(Morris 2006). Pada membaran bagian dalam mitokondria, subunit BCKDHA ini
bergabung dengan subunit-subunit lainnya membentuk kompleks enzim.
Defisiensi dari enzim BCKDHA dilaporkan oleh Zhang et al. (1989) sebagai
penyebab penyakit Maple Syrup Urine Disease (MSUD). Akibat dari akumulasi kadar
BCAAs dalam darah akan menjadi racun bagi tubuh. Hal tersebut dapat mengganggu
sistem saraf pusat yang ditunjukkan dengan dismyelinasi, edema serebral, dystonia,
koma, keterbelakangan mental, masalah kejiwaan, dan bahkan kematian dalam
beberapa minggu setelah dilahirkan (Friedrich et al. 2012). MSUD tipe 1A disebabkan
mutasi gen BCKDHA di ruas ekson 2 yang menyandikan 2-oxoisovalerate
dehydrogenase pada anak sapi Polled Hereford. Mutasi tersebut merupakan mutasi titik
akibat terjadinya substitusi nukleotida 248C/T pada kodon 6 dan menyebabkan stop kodon
prematur dari sub unit E1-α kompleks BCKD (Zhang et al. 1989; Zhang et al. 1990;
Zhang et al. 1991). Mutasi yang terjadi pada gen BCKDHA selain di ruas ekson 2 pada
sapi belum pernah dilaporkan (Zhang et al. 1990). Beberapa titik mutasi ruas ekson 2
pada subunit E1-α ini telah dikembangkan menjadi uji klinis penyakit MSUD berbasis
PCR pada sapi perah dan manusia (Zhang et al. 1990).
Keragaman yang dipelajari dalam penelitian ini adalah keragaman ruas ekson 2
– 3 gen BCKDHA terkait pengaruhnya terhadap kendali fungsi pengikatan antar
subunit dan juga terkait efisiensi metabolisme makanan pada sapi madura. Gen
BCKDHA memiliki ruas peptida transit yang disandikan oleh bagian ujung 5’ ekson 1
dan ruas bagian ujung 3’ pada ekson 2 – 9 yang akan menyandikan polipeptida untuk
membentuk struktur kuartener dengan sub unit lainnya (Chuang et al. 1993). Jika
terdapat mutasi pada nukleotida bagian ekson maka akan mempengaruhi efisiensi pada
peptida transit, situs pelekatan dengan subunit lainnya dan situs katalitik. Perubahan-
perubahan ketiganya ditunjukkan dengan kisaran efisiensi BCKDH dalam
memetabolisme BCAAs.
Tujuan Penelitian
Penelitian ini bertujuan untuk menganalisis keragaman ruas ekson 2 – 3 gen
penyandi enzim BCKDHA.

3
METODE
Penelitian ini dilaksanakan pada bulan Juni-Oktober 2013 di bagian Fungsi
Hayati dan Perilaku Hewan, Departemen Biologi, Fakultas Matematika dan Ilmu
Pengetahuan Alam, Institut Pertanian Bogor.
Sampel DNA
Sampel DNA sapi madura yang digunakan adalah koleksi dari Dr. R.R Dyah
Perwitasari, FMIPA IPB, yang diperoleh dari kabupaten Bangkalan, Madura.
Sebanyak 10 sampel DNA yang diekstraksi dari sampel darah dicampur menjadi satu
(pooled DNA samples) yang kemudian diberi label J (Lampiran 2).
Amplifikasi dan Visualisai DNA
Amplifikasi ruas ekson 2 – 3 gen BCKDHA dilakukan secara in vitro
menggunakan mesin PCR ESCO Swift Maxi Thermal Cycler. Pereaksi PCR yang
digunakan untuk volume reaksi 25 μl terdiri atas sampel DNA sekitar 50 ng sebanyak
1 µl, GoTaq® Green Master Mix 2X (Promega) (1 unit Taq Polimerase, 400 μM
dNTP, 3 mM MgCl2), primer forward AF318 (5’-agcacccccacaggtggcag-3’) dan
primer reverse AF319 (5’-cctgtcttgtggtccttagacc-3’) (lampiran 3) masing-masing 0,1
μM. Kondisi PCR yang digunakan adalah: predenaturasi 95 °C dua menit, kemudian
dilanjutkan 30 siklus terdiri dari denaturasi 95 °C 45 detik, suhu penempelan primer
58 °C satu menit, pemanjangan DNA pada suhu 72 °C satu menit, dan diakhiri
pemanjangan DNA pada suhu 72 °C lima menit. Produk PCR diamati menggunakan
metode polyacrilamide gel electrophoresis (PAGE) dengan konsentrasi akrilamid 6%
yang dilanjutkan dengan pewarnaan perak (Byun et al. 2009). Komposisi penyusun
gel adalah larutan akrilamid (akrilamid:bis akrilamid = 29:1) dan larutan buffer 1x
TBE (Tris HCl 10 mM, asam borat 1M, EDTA 0,1 mM, pH 8.0).
Perunutan Produk PCR
Produk PCR yang berupa pita tunggal di atas gel poliakrilamid dan berukuran
516 pasang basa (pb) dijadikan sampel dalam reaksi perunutan nukleotida. Reaksi
PCR perunutan nukleotida menggunakan primer yang sama seperti amplifikasi awal
dengan metode sequencing big dye terminator. Output perunutan nukleotida berupa
kromatogram yang dilakukan oleh lembaga komersial jasa sequencing. Pembacaan
runutan nukleotida menggunakan program ABI Prism 3700-Avant Generic Analyzer.
Analisis Data
Runutan nukleotida yang diperoleh kemudian diedit dan saling disejajarkan
dengan runutan DNA referensi yang ada dalam database GenBank
(http://ncbi.nlm.nih.gov) menggunakan program BioEdit versi 7.1.11. dan program
Clustal W 1.8 yang tertanam dalam program Moleculer Evolutionary Genetics
Analysis versi 5.0.5 (MEGA 5). Runutan nukleotida gen BCKDHA referensi yang
diambil sebagai pembanding adalah dari B. taurus (sapi taurine) no. akses
NW_001493616, Camelus ferus (unta) no. akses NW006211451, Capra hircus
(kambing) no. akses NC_022310, dan Ovis aries (domba) no. akses NC_019471.
Penghitungan jarak genetik dilakukan berdasarkan model subtitusi Kimura-2-
parameter (K2P).
4
HASIL DAN PEMBAHASAN
Hasil
Amplifikasi Ruas Ekson 2 – 3 gen BCKDHA
Ruas ekson 2 – 3 gen BCKDHA pada sapi madura J berhasil diamplifikasi
dengan metode PCR menggunakan pasangan primer forward AF318 dan reverse
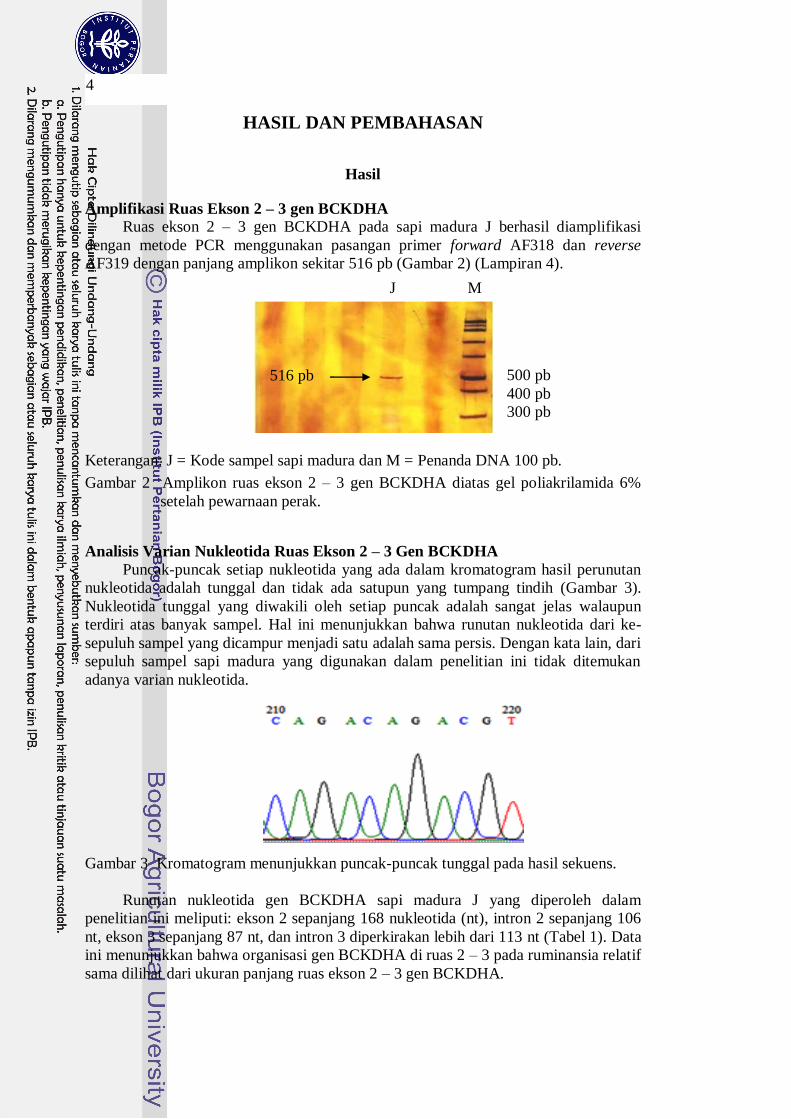
AF319 dengan panjang amplikon sekitar 516 pb (Gambar 2) (Lampiran 4).
Keterangan: J = Kode sampel sapi madura dan M = Penanda DNA 100 pb.
Gambar 2 Amplikon ruas ekson 2 – 3 gen BCKDHA diatas gel poliakrilamida 6%
setelah pewarnaan perak.
Analisis Varian Nukleotida Ruas Ekson 2 – 3 Gen BCKDHA
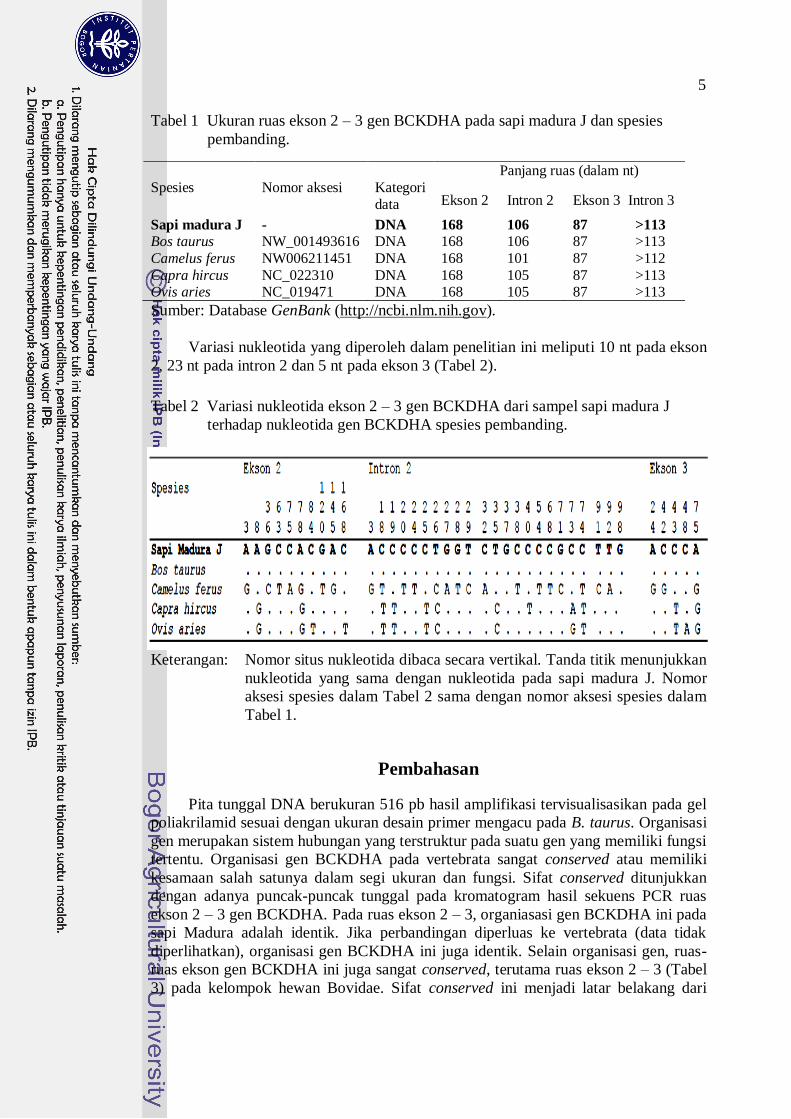
Puncak-puncak setiap nukleotida yang ada dalam kromatogram hasil perunutan
nukleotida adalah tunggal dan tidak ada satupun yang tumpang tindih (Gambar 3).
Nukleotida tunggal yang diwakili oleh setiap puncak adalah sangat jelas walaupun
terdiri atas banyak sampel. Hal ini menunjukkan bahwa runutan nukleotida dari ke-
sepuluh sampel yang dicampur menjadi satu adalah sama persis. Dengan kata lain, dari
sepuluh sampel sapi madura yang digunakan dalam penelitian ini tidak ditemukan
adanya varian nukleotida.
Gambar 3 Kromatogram menunjukkan puncak-puncak tunggal pada hasil sekuens.
Runutan nukleotida gen BCKDHA sapi madura J yang diperoleh dalam
penelitian ini meliputi: ekson 2 sepanjang 168 nukleotida (nt), intron 2 sepanjang 106
nt, ekson 3 sepanjang 87 nt, dan intron 3 diperkirakan lebih dari 113 nt (Tabel 1). Data
ini menunjukkan bahwa organisasi gen BCKDHA di ruas 2 – 3 pada ruminansia relatif
sama dilihat dari ukuran panjang ruas ekson 2 – 3 gen BCKDHA.
J M
516 pb 500 pb
400 pb
300 pb
5
Tabel 1 Ukuran ruas ekson 2 – 3 gen BCKDHA pada sapi madura J dan spesies
pembanding.
Spesies
Nomor aksesi
Kategori
data
Panjang ruas (dalam nt)
Ekson 2 Intron 2 Ekson 3 Intron 3
Sapi madura J - DNA 168 106 87 >113
Bos taurus NW_001493616 DNA 168 106 87 >113
Camelus ferus NW006211451 DNA 168 101 87 >112
Capra hircus NC_022310 DNA 168 105 87 >113 Ovis aries NC_019471 DNA 168 105 87 >113
Sumber: Database GenBank (http://ncbi.nlm.nih.gov).
Variasi nukleotida yang diperoleh dalam penelitian ini meliputi 10 nt pada ekson
2, 23 nt pada intron 2 dan 5 nt pada ekson 3 (Tabel 2).
Tabel 2 Variasi nukleotida ekson 2 – 3 gen BCKDHA dari sampel sapi madura J
terhadap nukleotida gen BCKDHA spesies pembanding.
Keterangan: Nomor situs nukleotida dibaca secara vertikal. Tanda titik menunjukkan
nukleotida yang sama dengan nukleotida pada sapi madura J. Nomor
aksesi spesies dalam Tabel 2 sama dengan nomor aksesi spesies dalam
Tabel 1.
Pembahasan
Pita tunggal DNA berukuran 516 pb hasil amplifikasi tervisualisasikan pada gel
poliakrilamid sesuai dengan ukuran desain primer mengacu pada B. taurus. Organisasi
gen merupakan sistem hubungan yang terstruktur pada suatu gen yang memiliki fungsi
tertentu. Organisasi gen BCKDHA pada vertebrata sangat conserved atau memiliki
kesamaan salah satunya dalam segi ukuran dan fungsi. Sifat conserved ditunjukkan
dengan adanya puncak-puncak tunggal pada kromatogram hasil sekuens PCR ruas
ekson 2 – 3 gen BCKDHA. Pada ruas ekson 2 – 3, organiasasi gen BCKDHA ini pada
sapi Madura adalah identik. Jika perbandingan diperluas ke vertebrata (data tidak
diperlihatkan), organisasi gen BCKDHA ini juga identik. Selain organisasi gen, ruas-
ruas ekson gen BCKDHA ini juga sangat conserved, terutama ruas ekson 2 – 3 (Tabel
3) pada kelompok hewan Bovidae. Sifat conserved ini menjadi latar belakang dari

6
desain penelitian ini, yaitu semua sampel dijadikan satu (DNA pooling). Teknik DNA
pooling juga dilakukan oleh Sham (2002), Mahfud (2009) dan Herodita (2009). Ukuran ruas ekson 2 – intron 3 gen BCKDHA pada hewan ruminansia relatif
sama (Tabel 1). Variasi nukleotida menunjukkan bahwa Sapi madura J identik dengan
Bos taurus (NW001493616). Sebagaimana gen-gen vital bagi metabolisme, nilai
keragaman ruas-ruas ekson gen BCKDHA sangat rendah (Ibeagha-Awemu et al.
2008) karena bersifat sangat conserved. Keragaman yang dipelajari dalam penelitian
ini adalah keragaman ruas ekson 2 – 3 gen BCKDHA terkait pengaruhnya terhadap
kendali fungsi pengikatan antar subunit dan juga terkait efisiensi metabolisme
makanan pada sapi madura.
Sapi madura umumnya merupakan hasil persilangan dari sapi zebu sebagai
induk dan banteng sebagai pejantan, namun ada beberapa jenis sapi madura yang
merupakan hasil persilangan dari sapi zebu sebagai induk betina dan sapi taurine
sebagai pejantan. Kesamaan fenotipe dapat dilihat dari warna bulu antara sapi taurin
dengan sapi madura yang berwarna merah kecoklatan (Maksum 1993). Berdasarkan
kromosom Y ditemukan beberapa sampel sapi madura merupakan keturunan dari sapi
taurin (Kusdiantoro et al. 2009).
SIMPULAN
Pada populasi sapi madura tidak ditemukan keragaman di ruas ekson 2 – 3 gen
BCKDHA. Organisasi gen BCKDHA pada ruas ekson 2 – 3 pada sapi madura sama
dengan anggota ruminansia lainnya dari segi ukuran, komposisi runutan nukleotida
dan fungsinya.
DAFTAR PUSTAKA
Byun SO, Fang Q, Zhou H, Hickford JGH. 2009. An effective method for silver
staining DNA in large numbers of polyacrylamide gels. Anal Biochem. 385:174-
175.
Chuang JL, Cox RP, Chuang DT. 1993. Characterization of the promoter-regulatory
region and structural organization of E1α gene (BCKDHA) of human branched-
chain α-ketoacid dehydrogenase complex. J Biol Chem. 268(11):8309-8316.
Elsik CG, Tellam RL, Worley KC. [Bovine Genome Sequencing and Analysis
Consortium]. 2009. The genome sequence of taurine cattle: a window to ruminant
biology and evolution. Science. 324(5926):522-528.
Friedrich T, Lambert AM, Masino MA, Downes GB. 2012. Mutations of zebrafish
dihydrolipoamide branched-chain transacylase E2 results in motor dysfunction and
models maple syrup urine disease. Disease Model and Mechanism. 5:248-258.
Herodita LU. 2009. Identifikasi defisiensi bovine leucocite adhesion deficiency
(BLAD) pada sapi friesan-holstein (FH) dari peternakan di jawa-bali [tesis]. Bogor
(ID): Institut Pertanian Bogor.

7
Hutson SM, Fenstermacher D, Mahar C. 1988. Role of mitochondrial transamination
in branched chain amino acid metabolism. The Journal of Biological Chemistry.
263(8):3618-3625.
Huitema H. 1986. Peternakan di Daerah Tropis; Arti Ekonomi dan Kemampuannya,
Penelitian di Beberapa Daerah di Indonesia. Terjemahan: Peni Hardjosworo.
Jakarta (ID): Yayasan Obor Indonesia.
Ibeagha-Awemu EM, Kgwatalala P, Ibeagha AE, Zhao X. 2008. A critical analysis of
disease-associated DNA polymorphisms in the genes of cattle, goat, sheep, and pig.
Mamm Genome. 19:226-245. Kusdiantoro M, Olsson M, Tol HTA, Mikko S, Vlamings BH, Andersson G, Martinez HR,
Purwantara B, Paling W, Colender B et al. 2009. The origin of indonesian cattle. PLoS
ONE. 4:1-5.
Mahfud K. 2009. Deteksi Dini Kelainan Genetik Complex Vertebral Malformation
dan Factor XI Deficiency pada Sapi Perah Friesian-Holstein [tesis]. Bogor (ID):
Institut Pertanian Bogor.
Maksum K. 1993. Hasil-hasil penelitian sapi madura di Sub Balai Penelitian Ternak
Grati-Pasuruan. Di dalam: Hasil Penelitian dan Pengembangan Sapi Madura.
Prosiding Pertemuan Ilmiah; Sumenep (ID), 11-12 Oktober 1992.
Morris SM. 2006. Branched-chain amino acids: metabolism, physiological function,
and application. The Journal of Nutrition. 136:254-255.
Nijman IJ, Otsen M, Verkaar ELC, de Ruijter C, Hanekamp E, Ochieng JW,
Shamshad S, Rege JEO, Hanotte O, Barwegen MW et al. 2003. Hybridization of
banteng (Bos javanicus) and zebu (Bos indicus) revealed by mitochondrial DNA,
satellite DNA, AFLP and microsatellites. Heredity. 90:10-16.
Patel MS, Harris RA. 1995. Mammalian α-keto acid dehydrogenase complexes: gene
regulation and genetic defects [review]. FASEB J. 9: 1164-1172.
Payne WJA, Hodges J. 1997. Tropical Cattle: Origins, Breed, and Breeding Policies.
Oxford: Blackwell Science Ltd.
Pettit FH, Yeaman SJ, Reed LJ. 1978. Purification and characterization of branched
chain α- ketoacid dehydrogenase complex of bovine kidney. Proc NatL Acad Sci.
75:4881-4885.
Sham P, Bader JS, Craig I, O’Donovsan M, Owen M. 2002. DNA pooling: a tool for a
larfe-scale association studies. Nar Rev Genet. 3(11):862:871.
Yeaman SJ. 1986. The mammalian 2-oxoacid dehydrogenases: a complex family.
Trends Biochem Sci: 11:293-296.
Zhang B, Edenberg HJ, Crabb DW, Harris RA. 1989. Evidence for both a regulatory
mutation and a structural mutation in a family with maple syrup urine disease. J
Clin Invest. 83(4):1425-9.
Zhang B, Healy PJ. Zhao Y, Ciabb DW, Hams RA. 1990. Premature translation
termination of the pre-Ela subunit of the branched chain a-ketoacid dehydrogenase
as a cause of maple syrup urine desease in Polled Hereford calves. J. Biol. Chem.
265:2425-2427.
Zhang B, Zhao Y, Harris RA, Crab DW. 1991. Molecular defects in the E1 alpha
subunit of the branched-chain alpha-ketoacid dehydrogenase complex that cause
maple syrup urine disease. Mol Biol Med. 8(1): 39-47.
8
LAMPIRAN
Lampiran 1 Gambar sapi madura
(Sumber: http://ditjennak.deptan.go.id)
Lampiran 2 Database DNA polled samples
Kode
sampel
Sumber
ektraksi
Darah yang
diambil
Asal Tipe Sapi
S1 Darah 200 µl Bangkalan Pedaging
S2 Darah 200 µl Bangkalan Pedaging
S3 Darah 200 µl Bangkalan Pedaging
S4 Darah 200 µl Bangkalan Pedaging
S5 Darah 200 µl Bangkalan Pedaging
S6 Darah 200 µl Bangkalan Pedaging
S7 Darah 200 µl Bangkalan Pedaging
S8 Darah 200 µl Bangkalan Pedaging
S9 Darah 200 µl Bangkalan Pedaging
S10 Darah 200 µl Bangkalan Pedaging

9
Lampiran 3 Posisi penempelan primer AF318 dan AF319 pada peta organisasi gen
BCKDHA sapi Bos taurus (NW001493616)
Lampiran 4 Posisi penempelan primer AF318 dan AF319
1 agcaccccca caggtggcag caacagcagc acttctcgtc cctggatgac
AF318-primer forward
51 aagccgcagt tcccaggggc ctcagcggag ttcatagaca agctcgaatt 101 catccagccc aatgtcatct ctgggatccc catctaccgg gtcatggacc
151 ggcagggcca gatcatcaac cccagcgagg atccccacgt aagaggccac
201 ctccccccga ccctgtgcct cccatgccca agccccttgc ccatctcctc
251 tctggcccca gctggcccac gtctgtctgt gcctctgtct gcagctgccc
301 caggagaagg tgctcaaatt ctacaagagc atgaccctgc tcaacaccat
351 ggaccgcatc ctctatgaat cccagaggca ggtgcgtggg gacaggctag
401 ggagggggcc tgggattacc tgaggtcctg cccaccctac ctgtgtctgg
451 gccaaaagac agcgcccaaa gaagagggag tggaatggag atccctgtct
AF319-
501 tgtggtcctt agacc Primer reverse

10
RIWAYAT HIDUP
Penulis dilahirkan di Bekasi pada tanggal 30 September 1991. Penulis
merupakan putri bungsu dari empat bersaudara dari pasangan Bapak Achmad Bakrie
dan Ibu Penih.
Penulis lulus dari SMAN 1 Tambun Utara, Bekasi pada tahun 2009 dan
melanjutkan pendidikan di Departemen Biologi, Fakultas Matematika dan Ilmu
Pengetahuan Alam, Institut Perttanian Bogor melalui jalur Undangan Seleksi Masuk
IPB (USMI).
Selama masa studi di IPB penulis aktif di berbagai organisasi kemahasiswaan.
Pada tahun 2009 sebagai anggota Klub Cinta Lingkungan dan Dewan Mushola
Asrama Putri A3 TPB, tahun 2010-2011 sebagai sekretaris divisi Biosains Himpunan
Mahasiswa Biologi (HIMABIO) dan pengurus rohis kelas biologi angkatan 46, tahun
2011 sebagai anggota UKM bela diri Taekwondo IPB. Penulis juga terlibat dalam
beberapa kepanitiaan kegiatan kampus pada tahun 2010 seperti Lomba Cepat Tepat
Biologi (LCTB) “Pesta Sains Nasional 2010” divisi Publikasi, Dekorasi dan
Dokumentasi, Green Society divisi Dana Usaha dan Konsumsi, Masa Pengenalan
Departemen Biologi angkatan 47 (MORFOLOGI 47) divisi Pendamping Junior Kelas,
tahun 2011 sebagai sekretaris divisi LCTB, sekretaris pada acara Seminar Nasional;
“Kesehatan Reproduksi Manusia”, sekretaris Pesta Karya ilmiah Mahasiswa club
(PKM club), Musyawarah Wilayah Jawa I Ikatan Himpunan Mahasiswa Biologi
Indonesia (IKAHIMBI) divisi penginapan, Panitia Pemilihan Raya Himabio 2011
divisi acara dan tahun 2012 sebagai panitia IPB Green Living Movement (IGLM) 2nd
divisi Photography and Film Contest.
Penulis pernah mengikuti program magang mahasiswa di Pusat Konservasi
Tumbuhan Kebun Raya Bogor pada divisi khusus pembibitan tanaman selama libur
alih semester 2010. Penulis telah melakukan Studi Lapangan pada tahun 2011 di
Hutan Pendidikan Gunung Walat IPB (HPGW), Sukabumi Jawa Barat dengan judul
“Ragam Ektomikoriza pada Pinus dan Meranti di Gunung Walat” dibawah bimbingan
Ir. Agustin Wydia Gunawan, M.Si. Selain itu penulis telah melakukan Praktik
Lapangan di PT. Godang Tua Jaya, Bantar Gebang, Bekasi dengan judul
“Pengomposan Sampah Organik dengan Teknik Open Windrow (Aerob) di Tempat
Pengolahan Sampah Terpadu (TPST) Bantar Gebang” dibawah bimbingan Dr. Ir. Utut
Widyastuti, M.Si. Penulis juga pernah menjadi asisten praktikum Fungsi Hayati dan
Perilaku Hewan tahun 2013. Selama masa studi penulis juga menerima Beasiswa
Bantuan Mahasiswa (BBM).





![PLAGIAT MERUPAKAN TINDAKAN TIDAK TERPUJI1].pdf · ΨTeman-teman Paduan Suara Fakultas Angel’s Voice angkatan tua. Noy, Sella, Anton, Damar, Siska, Krisna, Avi, Kak Chika, Mba’](https://static.fdocument.org/doc/165x107/5d199b0c88c993495f8d0c68/plagiat-merupakan-tindakan-tidak-terpuji-1pdf-teman-teman-paduan-suara-fakultas.jpg)










