Technische Universität Darmstadt July 2010 Fachbereich ...
37
ACARBOSE DaMocles – the students project 0 Technische Universität Darmstadt July 2010 Fachbereich Chemie Acarbose Students project of: Valerie Spieler, Mirja Schulze, Tran Nam Trung and Johanna Tüshaus http://www.guardianchemist2u.com/gpic/IMGP6751.JPG
Transcript of Technische Universität Darmstadt July 2010 Fachbereich ...

ACARBOSE
DaMocles – the students project 0
Technische Universität Darmstadt July 2010
Fachbereich Chemie
Acarbose
Students project of: Valerie Spieler, Mirja Schulze, Tran Nam Trung and Johanna Tüshaus
http://www.guardianchemist2u.com/gpic/IMGP6751.JPG

ACARBOSE
DaMocles – the students project 1
"Imagination is more important than knowledge."
Albert Einstein (1879-1955)

ACARBOSE
DaMocles – the students project 2
Index:
1.Introduction……………………………………………………….3
2.Production of acarbose……………………………….........5
3.Biosynthesis of acarbose……………………………….......8
4.Total synthesis of acarbose……………………………….13
5.Introduction of α-Glucosidase
and α-Glucosidase-inhibitors……………………………….20
6.α-Glucosidase inhibitions by acarbose
6.1.CGTase inhibiton .……………………………………25
6.2. Barley α-Amylase inhibition ………………… 29
7.Medical effects of acarbose ……………………………..33

ACARBOSE
DaMocles – the students project 3
1. Introduction.
Metabolites produced by bacteria are always interesting objects of research
for biologists and pharmacologists. Many screenings are designed for the
detection of desired properties among these metabolites, such as antibiotic
effect, inhibition of enzymes etc.
Acarbose is an α-Glucosidase-inhibitor, first found in the screening among the
product of various Actinomycetes. As an important drug for the treatment of
diabetes type II, its structure, properties, mechanism of inhibitions as well as
the biosynthetic pathway of acarbose are well researched. The brand names of
acarbose are Glucobay© (in Germany) and Precose© (in USA).
Acarbose is produced in a multi-step batch fermentation with developed strains derived from Actinoplanes sp. SE50, followed by a complex multi-step purification process (the detailed production process will be described later on). Under optimal conditions, up to 1g/L acarbose can be obtained. Among the side-products, component C is the most significant. Due to the structural similarity, component C can’t be removed easily from acarbose, and the purified product can contain up to 2% component C contamination[1].
Fig.1: Structure of acarbose (A) and related compounds [1]

ACARBOSE
DaMocles – the students project 4
Some information about acarbose [2] [3]:
Name: Acarbose
IUPAC Name: (2R,3R,4R,5S,6R)-5-[(2R,3R,4R,5S,6R)-5-[(2R,3R,4S,5S,6R)-3,4-
dihydroxy-6-methyl-5-[[(1S,4S,5S,6S)-4,5,6-trihydroxy-3-
(hydroxymethyl)-1-cyclohex-2-enyl]amino]oxan-2-yl]oxy-3,4-dihydroxy-
6-(hydroxymethyl)oxan-2-yl]oxy-6-(hydroxymethyl)oxane-2,3,4-triol
CAS-Number: 56180-94-0
Chemical formula: C25H43NO18
Molecular mass: 645.61
Solubility: very soluble in water, soluble in methanol, practically insoluble
in methylene chloride
Predicted Water Solubility: 148 mg/mL
Appearance: white to yellowish, amorphous powder hygroscopic
Specific rotation: [αD]20: +1680 to +1830
Usage: anti-diabetic drug
Brand names: Glucobay© (Germany), Precose© (USA)

ACARBOSE
DaMocles – the students project 5
2. Production of acarbose
2.1. Actinoplanes sp. SE50, the producers of acarbose
Bacteria of the „Actinoplanes sp.“ species belong to the genus “Actinomyces“,
which are also called “ray fungus”. This name derives from their nature, much
like fungi, to produce the so-called mycelium, the branching filaments.
Actinoplanes sp. also produce spores. They are the dormant form of the
bacteria, in which the whole metabolism is switched off. Even after many years
and extreme conditions, the spores can still germinate to totally proficient
bacteria. Actinoplanes sp. are Gram-positive bacteria with a high GC-content
(70-73%). These bacteria are found in many various habitats: different types of
soils, marine habitats, internal waters… They are everywhere. Actinoplanes is
not the only „famous“ species of this genus. Other species are well-known for
their production of antibiotics. [4]
The industrially used bacteria belong to the elite mutants, which were
cultivated in 1960s at the university of Marburg. They are originally descended
from a soil sample, taken from a coffee plantation in Kenya. The wild strains
were treated with various mutation-causing agents. The strains with highest
producing yield were chosen and isolated. The producing yield can be thus
enhanced 500 times[5].
2.2. Industrial production process of acarbose (text extracted from[1])
Acarbose-type compounds were found in a screening at Bayer AG in a program designed for the detection of inhibitors for mammalian intestinal α-amylase, sucrase and maltase activities among the products of various actinomycetes. The active ingredient of Glucobay, a weakly basic acarbose, was produced in a multi-step batch fermentation with developed strains derived from Actinoplanes sp. SE50 and by use of feeding protocols. A multi-stage purification process was carried out at the Bayer plant at Wuppertal–Elberfeld. The sterile fermentation took place in 30–100 m3 fermenters in a high-concentration medium, mainly based on starch hydrolysates (10% or higher) with or without added maltose (low glucose levels) under continuous control and applying a feed of nutrients to keep the main C-source (poly-,oligomaltodextrins) at a high concentration throughout.

ACARBOSE
DaMocles – the students project 6
Fig. 2: Industrial production of acarbose[1]
The basic downstream processing of the industrial production process involved seven steps:
1. The ion content of the culture broth was lowered by mixing it with both a highly acidic cation exchanger and an anion exchanger. Under this condition, most (more than 80%) of the acarbose bound to the cation exchanger.
2. The resins were separated from the broth and mycelia by passing the mixture through a sieve screw centrifuge, in which a washing step (deionized water) also took place.
3. Elution by 0.1 M sodium acetate occurred in kettles containing a suction head with nozzle sieves in the bottom, connected via a pump and rotameter to a column line.
4. Both inhibitor and contaminants were adsorbed and thereby separated via a column line of three ionexchange columns (cation/anion/cation exchangers in acidic/basic/acidic forms, respectively) of which the first was used for

ACARBOSE
DaMocles – the students project 7
desalination, the second for elevating the pH to higher than 3.0 and the third for binding the acarbose and its related components.
5. The last column was eluted by a gradient of 0.01–0.05 M mineral acids and a fractionated collection was made of the different components, the acarbose fraction of which was cut off.
6. The final creation of a slightly colored powder was achieved by volume reduction in a vacuum and the pH was brought to a value of about pH 5–7 by a mixture of cation and anion exchange resins, filtration through a sterile filter and freeze- or spray-drying.
7. Later, several improvements of the fermentation process and downstream processing were published, e.g. replacement of step 6 of the above procedure by an upscale of a quasi-HPLC procedure (e.g. columns of 11 m3 bed volume, 2.5 m diameter, 3 m height) via the development of a sulfonated bead cation exchanger using specially developed macroporous, hard and resistant polymers based on aromatic compounds with at least one vinyl group and one hydrophilic monomer (e.g. 6–20% divinylbenzene, 5-25% methoxymethacrylamide, among others).This allowed a more rapid flow and smaller elution volumes, combined with a stronger separation.
No exact values for the final acarbose concentrations in the harvested broth of the fermentation have been reported for the actual production strains. The deposited derivative Actinoplanes sp.SE50/110 produces up to 1 g acarbose/l under optimal culture conditions (Rauenbusch and Schmidt 1978). As a side-product, with increasing relative concentration during the elongated fermentation, component C seems to be unavoidable. Due to its high structural similarity to acarbose, component C is hard to separate. The biotechnological production process requires 2–3 weeks; and the purified product still contains some contamination by component C (below 2%).

ACARBOSE
DaMocles – the students project 8
3. Biosynthesis of acarbose
Though acarbose is already used successfully as an anti-diabetic drug and is
mass-produced, its biosynthesis is not completely discovered yet[6].
The biosynthesis can be separated into the productions of two parts: the
cyclitol sub-site unit and the deoxy sugar rest, which are reassembled quite
lately.
The first step of the synthesis of the cyclitol sub-site unit is the cyclization of
sedoheptulose-7-phosphate. Sedoheptulose-7-phosphate is the product from
the pentose phosphate pathway, a process which generates NADH and
converts various Carbohydrates into each other, for almost every creature [7].
The cyclization is catalyzed by the enzyme AcbC, 2-epi-5-epi-Valiolon is formed
from the reaction. This molecule undergoes the phosporylation in the C7-
position with the help of the enzyme AcbM. Next steps are the epimerization
in the C2-position, catalyzed by the enzyme AcbO, and the reduction of the
ketone group (C1) to the hydroxyl group with the help of the enzyme AcbL.
AcbN now catalyzes the 5,6-elimination of water from 2-epi-5-epi-Valiolon.
The OH-group at C1-positon is again phosphorylated, whereby 1,7-diphospho-
1-epi-valienol is formed. The enzyme for this reaction still remains unknown.
At last, the phosphate group at C1 is replaced by a nucleoside phosphate (the
reaction is catalyzed by the enzyme AcbR). NDP-1-epi-Valienol-7-phosphate is
thus formed.
The deoxy sugar, the second important part of the biosyntheis, is synthesized
from D-glucose. First of all D-glucose is phosphorylated to D-glucose-1-
phosphate. With the help of the enzyme AcbA, the phosphate rest is now
replaced by deoxythymidine-diphosphate. In the next step, AcbB catalyzes the
oxidation of the hydroxyl group at the C4-position, and the removing of OH-
group at C6. As a result, dTDP-4-keto-deoxy-D-Glucose is formed. Finally, the
ketone group is replaced by an amino group (enzyme: AcbV), which contains
the nitrogen atom that decides the activity of acarbose. The synthesis has now
stopped at dTDP-4-amino-4,6-dideoxy-D-Glucose.

ACARBOSE
DaMocles – the students project 9
The two, until now parallel going synthesises finally meet each other: AcbS or
AcbI (still debated) catalyzes the coupling of NDP-1-epi-Valienol-7-phosphate
and dTDP-4-amino-4,6-dideoxy-D-Glucose to dTDP-Acarviose-7-phosphat,
which is the direct precursor of acarbose. With the help of AcbQI or AcbSJ (also
debated), the missing maltosyl rest is now added, and the synthesis of
acarbose comes to its end.
Fig.3: Postulated biosynthetic pathway of acarbose in Actinoplanes sp.SE50[1]

ACARBOSE
DaMocles – the students project 10
The acb gene cluster from Actinoplanes sp.SE50 (responsible for the production of acarbose) is fully analyzed. It is realized that glucose inhibits the production of acarbose, while maltose and maltooligodextrin induces it. Upstream of some key genes, carbohydrate-regulated operator sites are found.
Fig.4: The acb-cluster of Actinoplanes sp.SE50[1]

ACARBOSE
DaMocles – the students project 11
A slightly different mechanism is proposed by Taifo Mahmud and colleges, University of Washington, using H-NMR spectroscopy to study the biosynthetic mechanism. The biosynthesis begins with the condensation between 2-epi-5-epi-valiolone (the direct precursor of the Valienamine moiety) and the amino group of dTDP-4-amino-4,6-dideoxy-D-glucose. The newly formed Schiff base undergoes a few modifications to become a pseudo-disaccharide. Much like the former mechanism, two further glucose units are now added, finishing the biosynthesis process.
Fig. 5: an alternative biosynthetic pathway of acarbose[8]
ACARBOSE
DaMocles – the students project 12
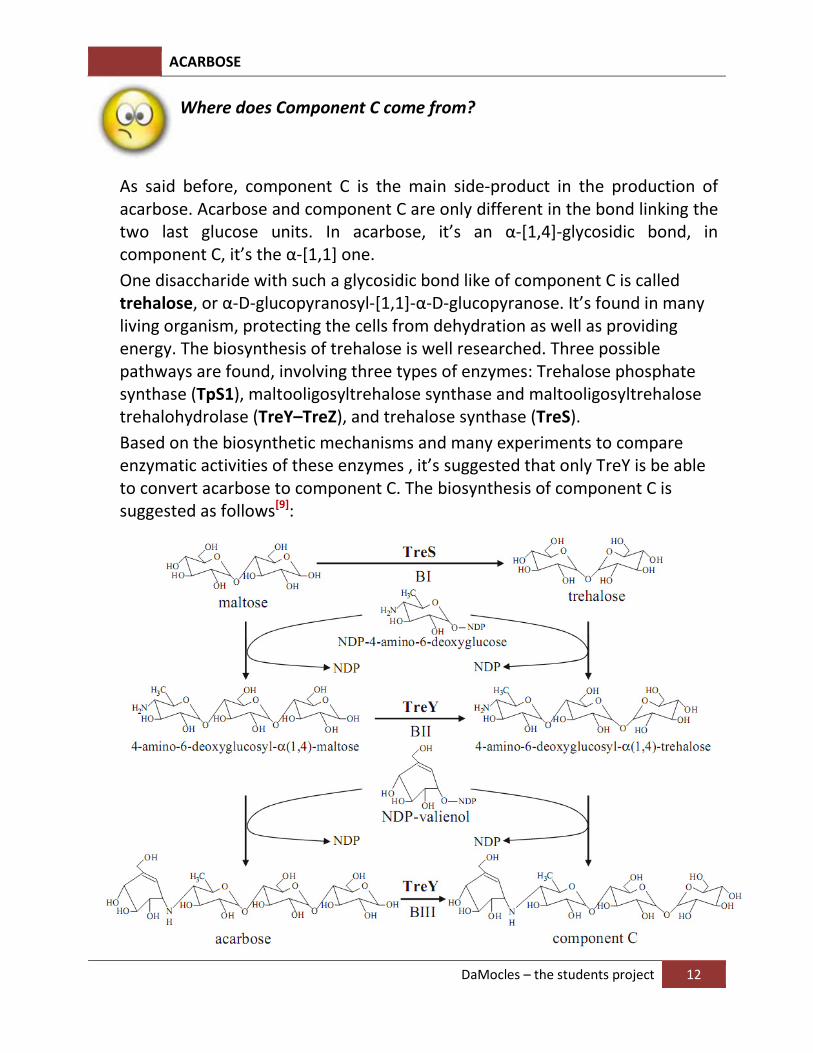
Where does Component C come from?
As said before, component C is the main side-product in the production of acarbose. Acarbose and component C are only different in the bond linking the two last glucose units. In acarbose, it’s an α-[1,4]-glycosidic bond, in component C, it’s the α-[1,1] one.
One disaccharide with such a glycosidic bond like of component C is called trehalose, or α-D-glucopyranosyl-[1,1]-α-D-glucopyranose. It’s found in many living organism, protecting the cells from dehydration as well as providing energy. The biosynthesis of trehalose is well researched. Three possible pathways are found, involving three types of enzymes: Trehalose phosphate synthase (TpS1), maltooligosyltrehalose synthase and maltooligosyltrehalose trehalohydrolase (TreY–TreZ), and trehalose synthase (TreS).
Based on the biosynthetic mechanisms and many experiments to compare enzymatic activities of these enzymes , it’s suggested that only TreY is be able to convert acarbose to component C. The biosynthesis of component C is suggested as follows[9]:
ACARBOSE
DaMocles – the students project 13
4. Total synthesis of acarbose[10]
Acarbose is a valuable α-glucosidase-inhibitor used widely in treatments of
diabetes type II. Acarbose is industrially produced from the fermentation of
developed strains of Actinomycetes. Due to its importance, acarbose is
studied extensively in both terms of mechanisms of action as well as structure
and synthesis.
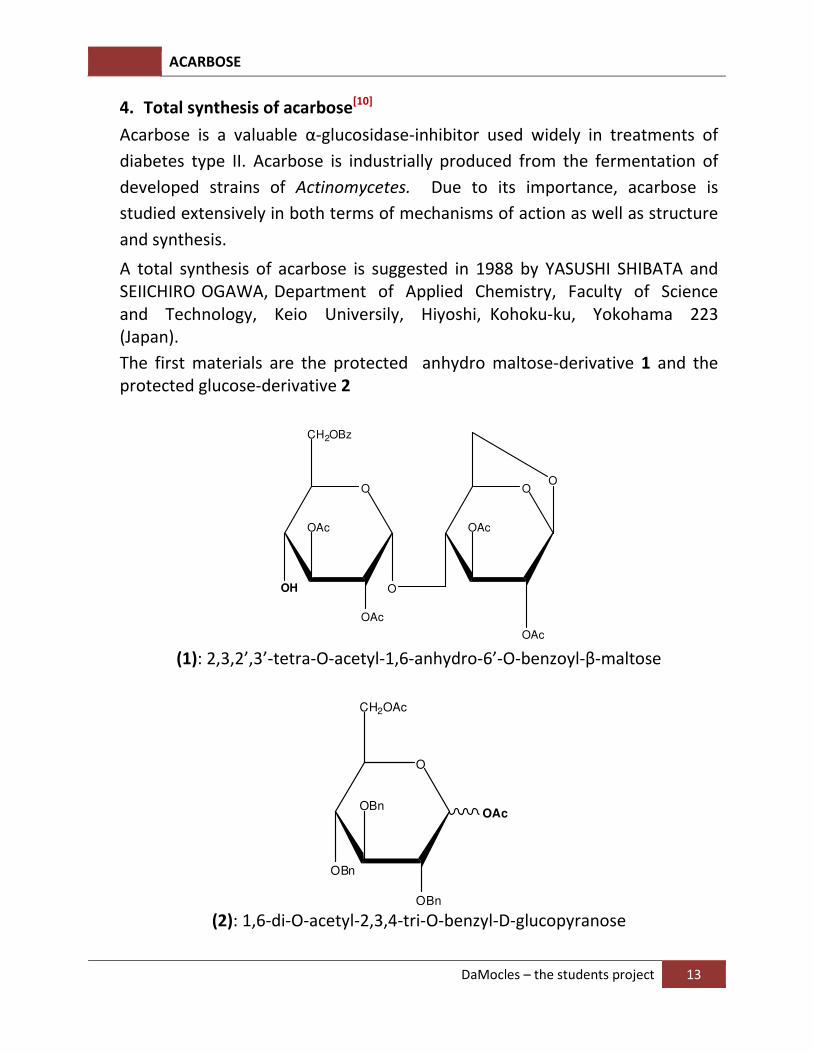
A total synthesis of acarbose is suggested in 1988 by YASUSHI SHIBATA and SEIICHIRO OGAWA, Department of Applied Chemistry, Faculty of Science and Technology, Keio Universily, Hiyoshi, Kohoku-ku, Yokohama 223 (Japan).
The first materials are the protected anhydro maltose-derivative 1 and the protected glucose-derivative 2
O
CH2OBz
OH
OAc
O
O
OAc
OAc
O
OAc
(1): 2,3,2’,3’-tetra-O-acetyl-1,6-anhydro-6’-O-benzoyl-β-maltose
O
CH2OAc
OBn
OBnOAc
OBn
(2): 1,6-di-O-acetyl-2,3,4-tri-O-benzyl-D-glucopyranose
ACARBOSE
DaMocles – the students project 14
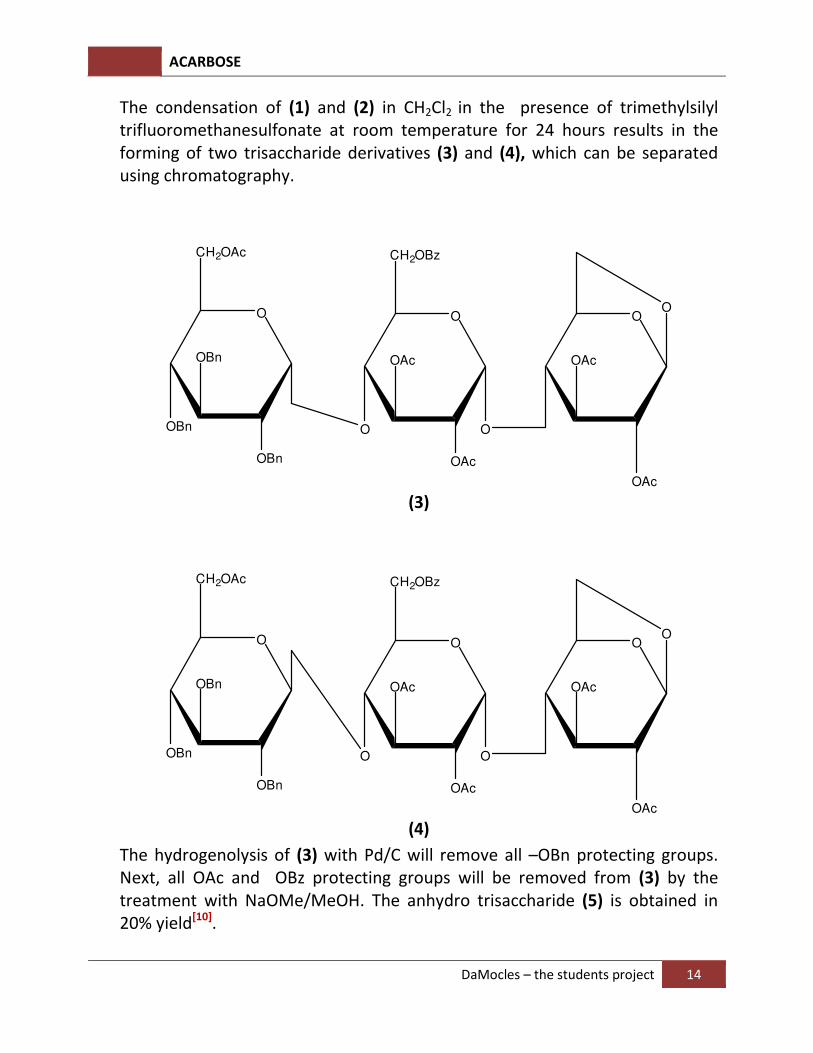
The condensation of (1) and (2) in CH2Cl2 in the presence of trimethylsilyl
trifluoromethanesulfonate at room temperature for 24 hours results in the forming of two trisaccharide derivatives (3) and (4), which can be separated using chromatography.
O
CH2OBz
O
OAc
O
O
OAc
OAc
OO
CH2OAc
OBn
OBn
OBn OAc
(3)
O
CH2OBz
O
OAc
O
O
OAc
OAc
OO
CH2OAc
OBn
OBn
OBn OAc
(4)
The hydrogenolysis of (3) with Pd/C will remove all –OBn protecting groups. Next, all OAc and OBz protecting groups will be removed from (3) by the treatment with NaOMe/MeOH. The anhydro trisaccharide (5) is obtained in 20% yield[10].
ACARBOSE
DaMocles – the students project 15
O
CH2OH
O
OH
O
O
OH
OH
OO
CH2OH
OH
OH
OH OH
(5)
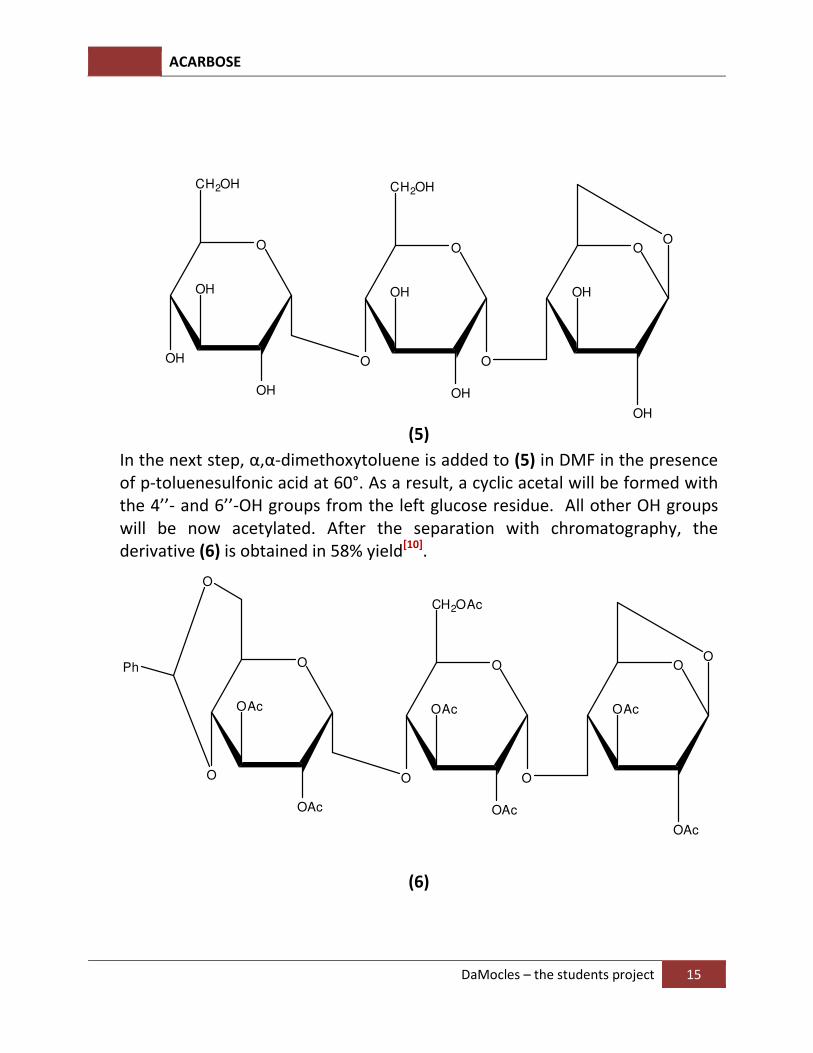
In the next step, α,α-dimethoxytoluene is added to (5) in DMF in the presence of p-toluenesulfonic acid at 60°. As a result, a cyclic acetal will be formed with the 4’’- and 6’’-OH groups from the left glucose residue. All other OH groups will be now acetylated. After the separation with chromatography, the derivative (6) is obtained in 58% yield[10].
O
CH2OAc
O
OAc
O
O
OAc
OAc
OO
O
OAc
OAc OAc
O
Ph
(6)
ACARBOSE
DaMocles – the students project 16
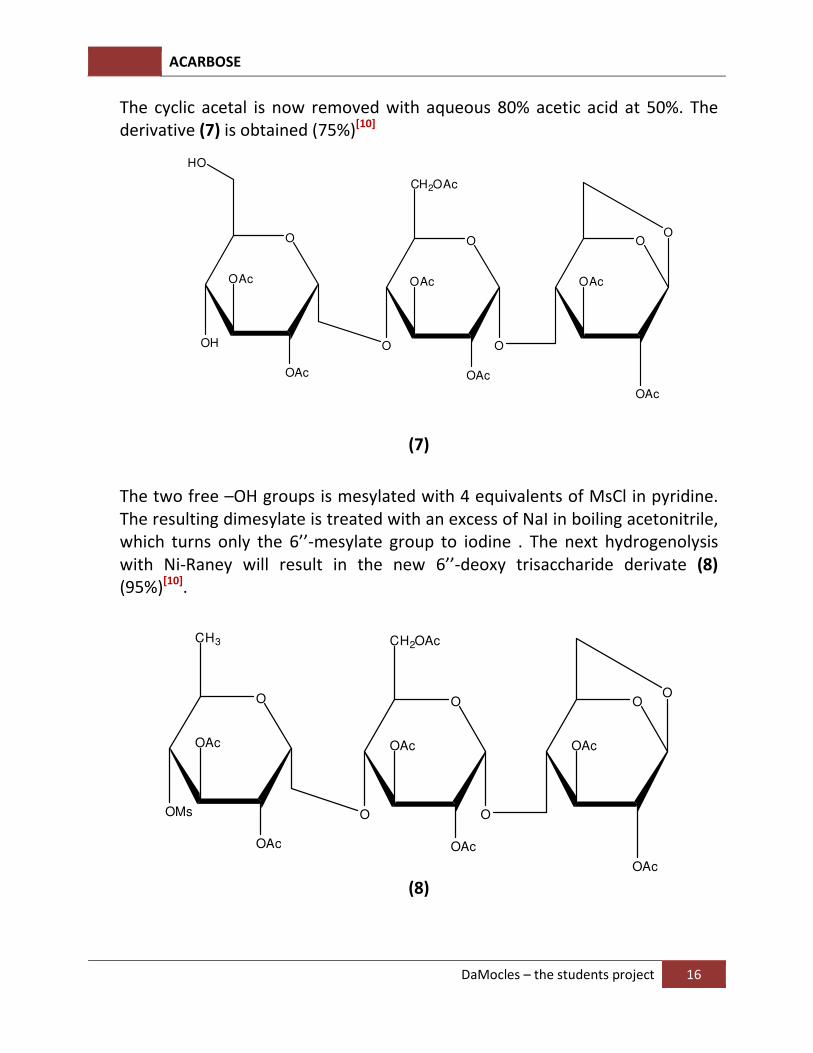
The cyclic acetal is now removed with aqueous 80% acetic acid at 50%. The derivative (7) is obtained (75%)[10]
O
CH2OAc
O
OAc
O
O
OAc
OAc
OO
OH
OAc
OAc OAc
HO
(7)
The two free –OH groups is mesylated with 4 equivalents of MsCl in pyridine. The resulting dimesylate is treated with an excess of NaI in boiling acetonitrile, which turns only the 6’’-mesylate group to iodine . The next hydrogenolysis with Ni-Raney will result in the new 6’’-deoxy trisaccharide derivate (8) (95%)[10].
O
CH2OAc
O
OAc
O
O
OAc
OAc
OO
CH3
OMs
OAc
OAc OAc
(8)
ACARBOSE
DaMocles – the students project 17
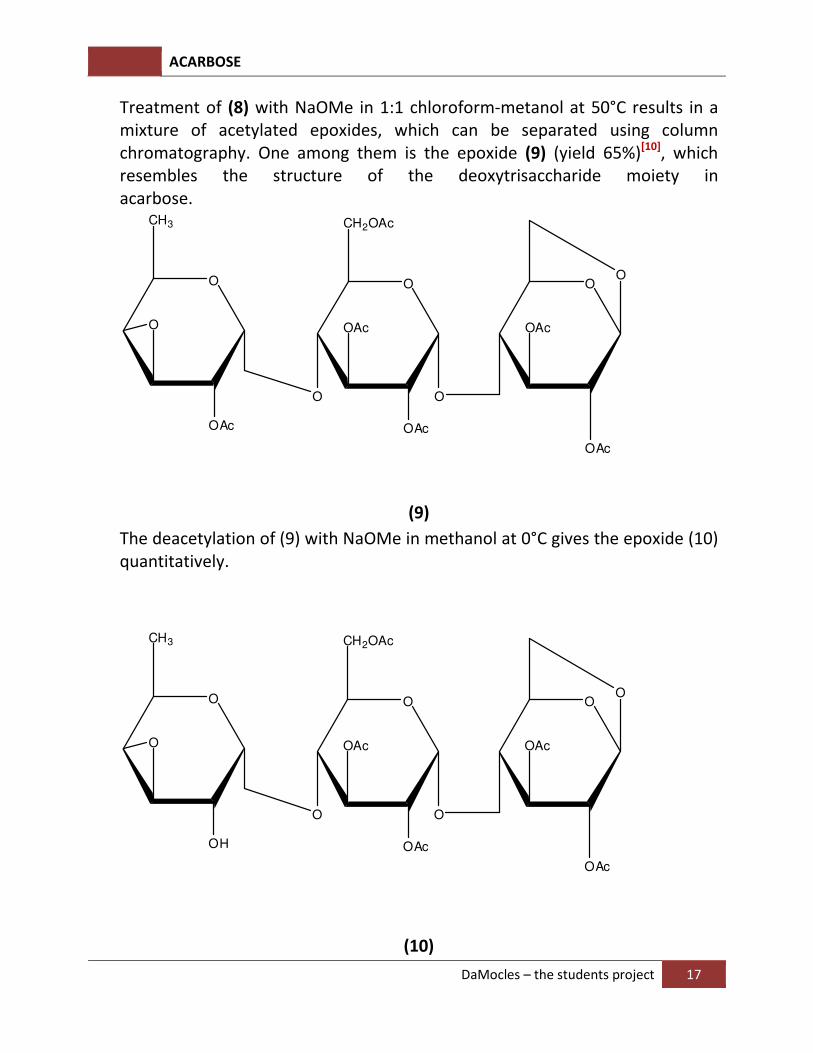
Treatment of (8) with NaOMe in 1:1 chloroform-metanol at 50°C results in a mixture of acetylated epoxides, which can be separated using column chromatography. One among them is the epoxide (9) (yield 65%)[10], which resembles the structure of the deoxytrisaccharide moiety in acarbose.
O
CH2OAc
O
OAc
O
O
OAc
OAc
OO
CH3
O
OAc OAc
(9)
The deacetylation of (9) with NaOMe in methanol at 0°C gives the epoxide (10) quantitatively.
O
CH2OAc
O
OAc
O
O
OAc
OAc
OO
CH3
O
OH OAc
(10)
ACARBOSE
DaMocles – the students project 18
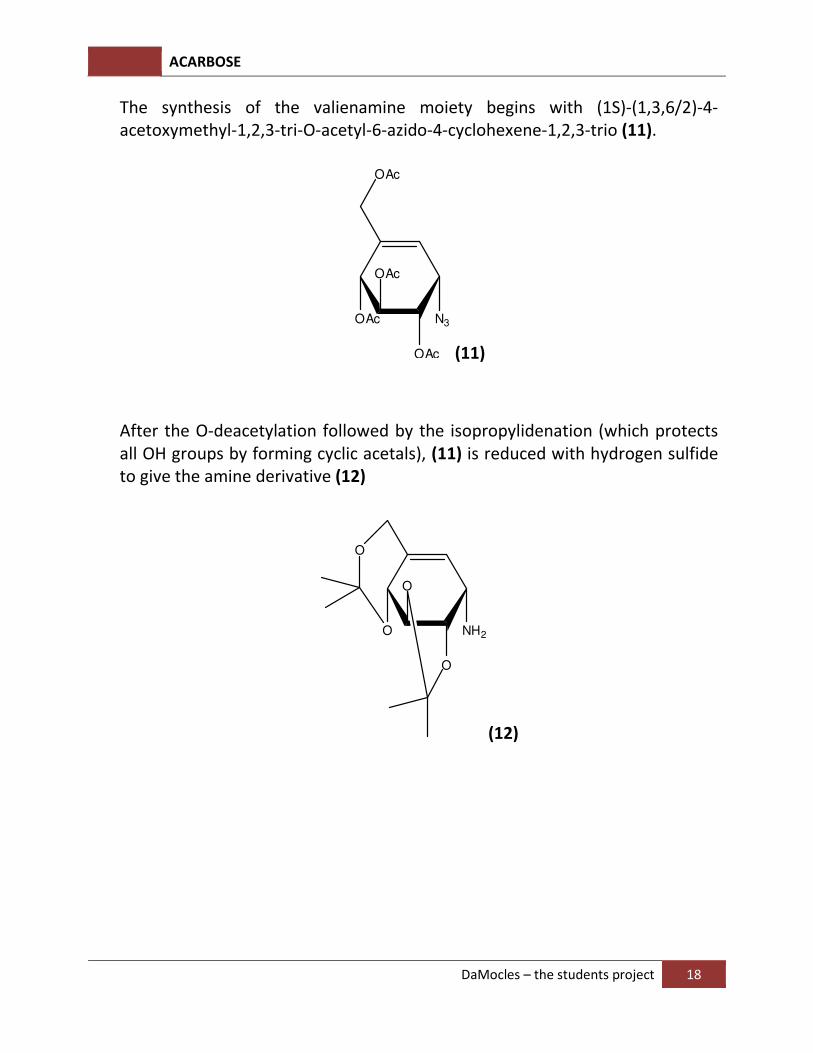
The synthesis of the valienamine moiety begins with (1S)-(1,3,6/2)-4-acetoxymethyl-1,2,3-tri-O-acetyl-6-azido-4-cyclohexene-1,2,3-trio (11).
OAc
OAc
OAc
OAc
N3
(11)
After the O-deacetylation followed by the isopropylidenation (which protects all OH groups by forming cyclic acetals), (11) is reduced with hydrogen sulfide to give the amine derivative (12)
O
O
O
NH2
O
(12)
ACARBOSE
DaMocles – the students project 19
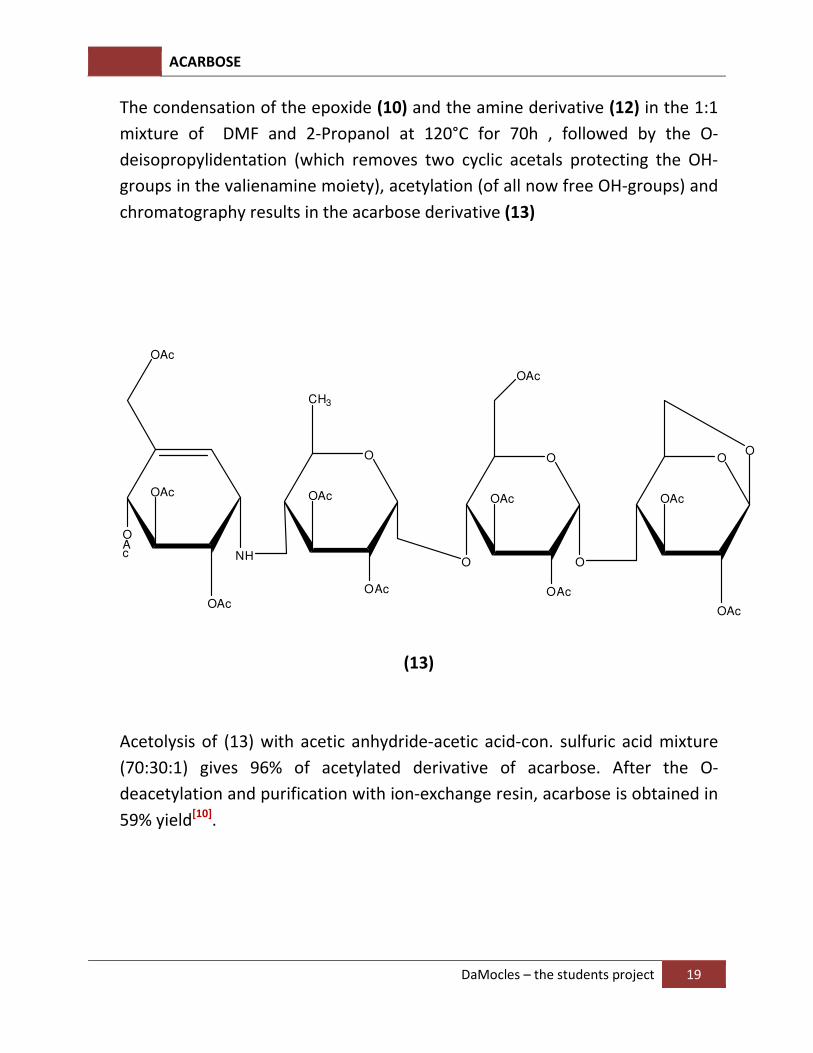
The condensation of the epoxide (10) and the amine derivative (12) in the 1:1
mixture of DMF and 2-Propanol at 120°C for 70h , followed by the O-
deisopropylidentation (which removes two cyclic acetals protecting the OH-
groups in the valienamine moiety), acetylation (of all now free OH-groups) and
chromatography results in the acarbose derivative (13)
O
O
OAc
O
O
OAc
OAc
OO
CH3
OAc
OAc OAc
OAc
OAc
OAc
OAc
NH
OAc
(13)
Acetolysis of (13) with acetic anhydride-acetic acid-con. sulfuric acid mixture
(70:30:1) gives 96% of acetylated derivative of acarbose. After the O-
deacetylation and purification with ion-exchange resin, acarbose is obtained in
59% yield[10].

ACARBOSE
DaMocles – the students project 20
5.Introduction of α-Glucosidase and α-Glucosidase-inhibitors
5.1. Glucosidase enzymes:
Glucosidase are enzymes that catalyze the cleavage of glycosidic bonds in oligosaccharides and glycoconjugates. As their names suggest, α-glucosidase enzymes cut the α-glycosidic bonds, and β-glucosidase enzymes cut β-glycosidic bonds. Due to the nature of the uptaken polysaccharides, human have α-glucosidase, while some other animals which can consume cellulose have β-glucosidase.
The activities of glucosidase enzymes play a crucial role in several important biochemical processes, such as: (i) degradation of diet polysaccharides to furnish monosaccharide units, which are then able to be metabolically absorbed and used by the organism, (ii) lysosomal glycoconjugate catabolism and glycoprotein processing, and (iii) biosynthesis of oligosaccharide units in glycoproteins or glycolipids[11].
In some diseases, especially diabetes, due to defects in the digestion machinery, the degradation of polysaccharides goes wrong, causing malicious effects. One of the widely used treatments for these diseases is the inhibition of α-glucosidase. Currently, three drugs are therapeutically used as anti-glucosidases: acarbose (Precose© or Glucobay©), miglitol (Glyset©), and N-butyl-1-deoxynojirimycin (Zavesca©). The first two drugs are used in the treatment of non-insulin-dependent diabetes (diabetes type II), while the third is employed for the control of Gaucher’s disease, related to disturbed lysosomal storage.
5.2. Glucosidase’s mechanism of action:
The widely accepted mechanism for retaining hydrolysis of glycosidic bonds involves a double-displacement mechanism originally proposed by Koshland (1953) and recently reviewed by McCarter & Withers (1994) and Davies & Henrissat (1995). The first step is the protonation of the anomeric O-atom(which requires a proton donor, normally a Glu-residue of the enzyme itself), followed by the cleavage of glycosidic bond and the formation of the oxocarbonium ion. Alternatively, this oxocarbonium ion can be stabilized by the forming of covalent bond with the side-chain of an Asp residue. A third mechanism involving the opening of the glucosidic ring is also suggested, but it remains controversial due to the lack of experimental supports.

ACARBOSE
DaMocles – the students project 21
Fig.6 The general mechanism of action of glucosidase enzymes
[11]
The next step is the attack of the water molecule and the forming of the new carbohydrate. It’s proposed that both steps operate via transition states with substantial oxocarbonium ion character(Sinnott, 1990), which implies the half-chair conformation. As a result, compounds containing the same type of structure (for example, acarbose) are good candidates for α-glucosidase inhibitons.
The detailed mechanisms of specific enzymes with specific substrates are of course slightly different from each other. They’ll be discussed later.
5.3. Some examples of α-glucosidase-inhibitors[11]
:
α-glucosidase inhibitors can be sorted into several important categories:
5.3.1. Disaccharides (text extracted from [11]
):
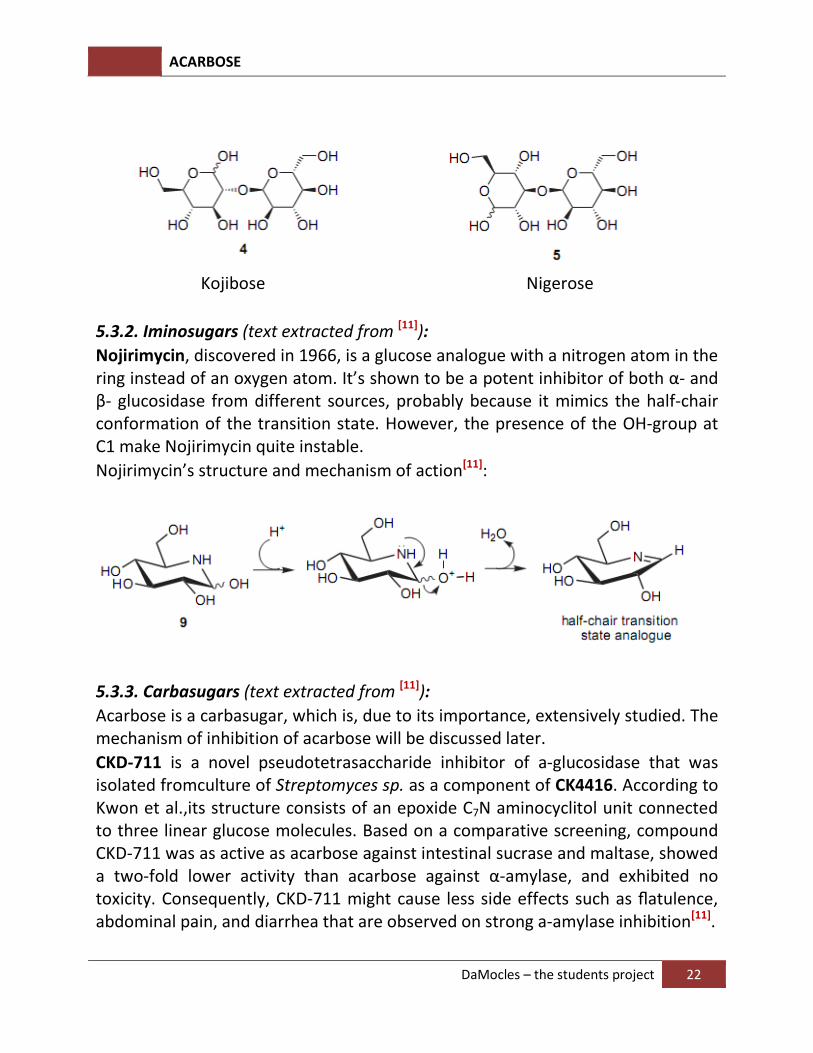
The isolation of kojibiose and nigerose, inhibitors of α-D-glucosidase I and a-D-glucosidase II, respectively, opened up new perspectives for the development of novel drugs, especially of the pseudodisaccharide class, for the treatment of HIV infections. Kojibiose containing α-(1->2) glycosidic bonds was isolated in 1953 from sake extracts and also from its primary source, koji, a product related to rice fermentation by Aspergillus oryzae.
On the other hand, acid hydrolysis of amylopectin produced nigerose, which was shown to have an α-(1->3) linkage.The importance of nigerose and nigerosylmalto oligosaccharides has also been shown to influence the immune function and quality of life in the healthy elderly person as a supplemental syrup on food.
ACARBOSE
DaMocles – the students project 22
Kojibose Nigerose
5.3.2. Iminosugars (text extracted from [11]
):
Nojirimycin, discovered in 1966, is a glucose analogue with a nitrogen atom in the ring instead of an oxygen atom. It’s shown to be a potent inhibitor of both α- and β- glucosidase from different sources, probably because it mimics the half-chair conformation of the transition state. However, the presence of the OH-group at C1 make Nojirimycin quite instable.
Nojirimycin’s structure and mechanism of action[11]:
5.3.3. Carbasugars (text extracted from [11]
):
Acarbose is a carbasugar, which is, due to its importance, extensively studied. The mechanism of inhibition of acarbose will be discussed later.
CKD-711 is a novel pseudotetrasaccharide inhibitor of a-glucosidase that was isolated fromculture of Streptomyces sp. as a component of CK4416. According to Kwon et al.,its structure consists of an epoxide C7N aminocyclitol unit connected to three linear glucose molecules. Based on a comparative screening, compound CKD-711 was as active as acarbose against intestinal sucrase and maltase, showed a two-fold lower activity than acarbose against α-amylase, and exhibited no toxicity. Consequently, CKD-711 might cause less side effects such as flatulence, abdominal pain, and diarrhea that are observed on strong a-amylase inhibition[11].

ACARBOSE
DaMocles – the students project 23
CKD-711 CK4416
5.3.4. Thiosugars (text extracted from [11]
)
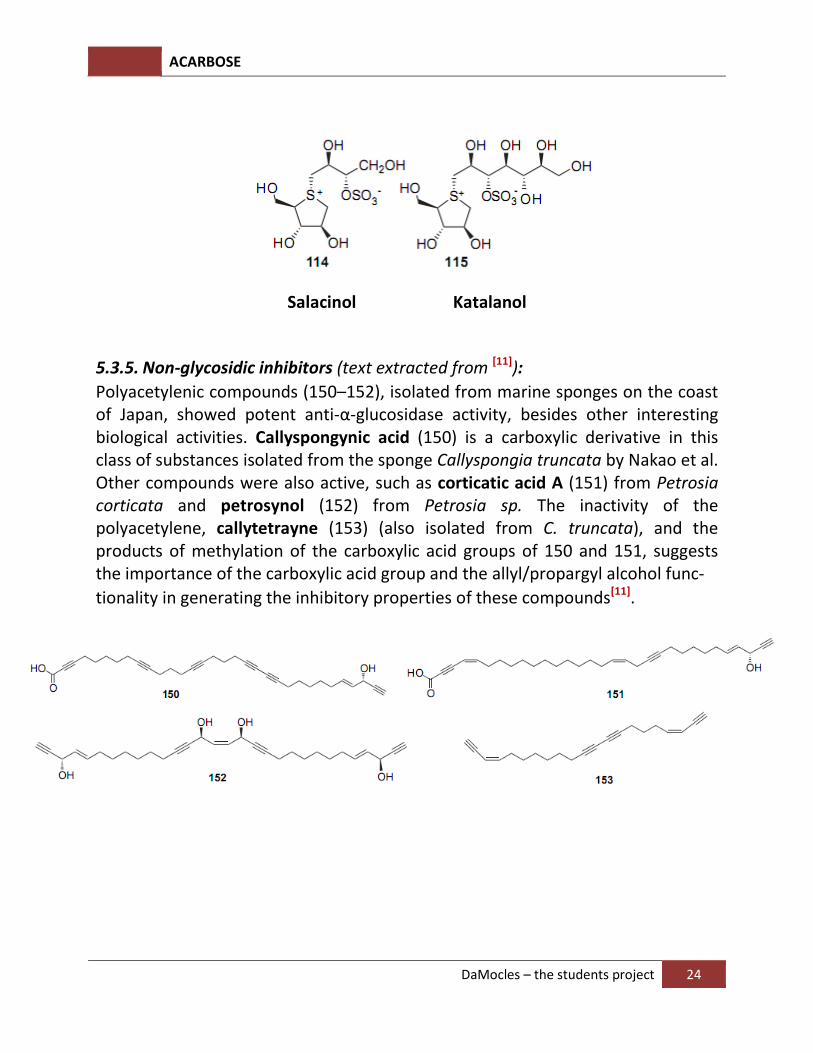
Replacement of the oxygen atom in the ring of a carbohydrate by sulfur affords thiosugars, a class of compounds comprising derivatives with strong anti-α-glucosidase activity. The naturally occurring salacinol is a remarkable member of this class, consisting of a thiocyclopentane with a trivalent sulfur atom in the ring (sulfonium ion) and an O-sulfate as part of an erythritol side chain, which is able to act as a counterion. Its structure was established by X-ray crystallographic analysis, showing it to have a spiro-like configuration with the 1-deoxy-4-thioarabinofuranosyl cation, bearing a resemblance with the iminoalditol
linked through sulfur to a 10-deoxyerythrosyl-30-sulfate anion. Salacinol was isolated by Yoshikawa et al. from an aqueous extract of the roots and stems of Salacia reticulata Wight, traditionally used in India and Sri Lanka for the treatment of diabetes, and it has been synthesized. Salacinol produced a strong inhibition for the increase of serum glucose levels in in-vivo screening, along with competitive inhibition against intestinal α-glucosidases such as maltase, sucrase, and isomaltase, in which the activity against isomaltase was higher than that of acarbose . Kotalanol, a derivative of 1,2,3-trihydroxy-propyl-salacinol, also consists of an inner salt sulfonium-sulfate structure and has been isolated from Salacia reticulate, and showed more potent inhibitory activity against sucrase than salacinol and acarbose[11].
ACARBOSE
DaMocles – the students project 24
Salacinol Katalanol
5.3.5. Non-glycosidic inhibitors (text extracted from [11]
):
Polyacetylenic compounds (150–152), isolated from marine sponges on the coast of Japan, showed potent anti-α-glucosidase activity, besides other interesting biological activities. Callyspongynic acid (150) is a carboxylic derivative in this class of substances isolated from the sponge Callyspongia truncata by Nakao et al. Other compounds were also active, such as corticatic acid A (151) from Petrosia
corticata and petrosynol (152) from Petrosia sp. The inactivity of the polyacetylene, callytetrayne (153) (also isolated from C. truncata), and the products of methylation of the carboxylic acid groups of 150 and 151, suggests the importance of the carboxylic acid group and the allyl/propargyl alcohol func-
tionality in generating the inhibitory properties of these compounds[11].

ACARBOSE
DaMocles – the students project 25
6. α-Glucosidase inhibitions by acarbose
6.1. Inhibition of CGTase by acarbose:
6.1.1. Introduction (text extracted from [12]
).
Cyclodextrin glycosyltransferases (CGTases; EC 2.4.1.19) form a class of proteins which convert starch into cyclodextrins and related α-(1->4)-linked glucose polymers (French, 1957). Cyclodextrins (CDs) are cyclic compounds consisting of six, seven, or eight α-(1->4)-linked D-glucopyranose units: alpha-CD, beta-CD, and gamma-CD, respectively. CGTases generally produce a mixture of these three types of cyclodextrins, and, according to their most preferred product, they are divided into alpha-, beta-, and gamma-CGTases. The beta-CGTases are especially of interest since beta-cyclodextrin is most widely applied (Schmid, 1989). However, the industrially used CGTases suffer from product inhibition and their product specificity is not very high.
Acarbose is a effective α-Glucosidase-inhibitor. Its structure is very similar to the structure of tetramaltose. The structure of acarbose as well as its differences from tetramaltose are shown in the following figure[12]:

ACARBOSE
DaMocles – the students project 26
The X-ray structure of acarbose’s complex with wild-type CGTase from B. circulans is studied.
6.1.2. Results and discussions.
6.1.2.1. Structure of acarbose in the complex with CGTase
The following picture shows the electron density of acarbose bound in the active site of CGTase. All oxygen-atoms are shown with their index and the residue where they come from[12].
The picture shows clearly, acarbose is twisted in the complex with CGTase, making a sharp bend at the glycosidic bond between the B and C glucose residues. In contrast, the torsion angles between the A and B, as well as C and D residues are found to be close to the angles between the two glucose units of maltose. The torsion angles imply that, while the OH- (and -NH) groups of A and B, C and D can interact with each other using hydrogen bonds, there is no internal hydrogen bond between B and C. Thus, the resulting acarbose structure may be considered to consist of two disaccharides with normal maltose-like conformations (A-B, respectively C-D), linked by a conformationally unusual glycosidic bond between residues B and C, that is not stabilized by an internal hydrogen bond[12].
6.1.2.2. Conformation changes of CGTase upon the binding of acarbose.
The structure study shows that, the binding of acarbose causes only a few changes in conformation of a few active sites, most significantly the side-chain of the Glu257 residue. In the native structure, it’s found linking with the side-chain of the Asp328 residue, using hydrogen bond. Upon binding of acarbose,

ACARBOSE
DaMocles – the students project 27
the distance between them increase from 2,5 to 3,6 Å, implying that the hydrogen bond is broken. The data of torsion angles also shows that, the side-chain of Glu257 has moved away. Several site-directed mutagenesis experiments prove the importance of both residues for the catalytic activity of CGTase. Another important residue is Asp229, but no change in conformation of this residue is found upon binding of acarbose[12].
6.1.2.3. Interaction of residues B and C of acarbose with the active site
The facts that, two important residues (Glu257, Asp238) of the enzyme are found bound to the B and C residues, as well as the unusual bond between them, imply the importance of B and C residues in the inhibition activity of acarbose.
Analysis of the hydrogen bonding network shows that residue B makes the largest number of hydrogen bonds with the nearby active site residues, including two of the three catalytic residues (Glu257, Asp328) and the very conserved His327 residue (Svensson, 1988). Further binding interactions of the sugar ring come from a stacking contact with the aromatic ring of Tyr100 which is approximately parallel to the plane of the sugar ring and the van der Waals contacts with the very conserved His140.

ACARBOSE
DaMocles – the students project 28
The residue C of acarbose is linked with His233 and Glu257 residues. Furthermore, there is a weak interaction between the hydroxyl group of Tyr195 and the O5 atom (4.1 Å) [12].
6.1.2.4. CGTase’s mechanism of actions
The catalytic mechanism of CGTase is thought to be similar to the general mechanism of α-glucosidase shown earlier. Glu257 is the proton donor, and Asp238 helps stabilizing the oxocarbonium ion. From the study of 3D-structure of both enzyme and enzyme-inhibitor complex, the mechanism is suggested as follows:
In native CGTase, the carboxylic groups of Glu257 and Asp238 are found binding strongly (the distance between them is 2,5Å), indicating that one of them is protonated (most probably Glu257). Upon the binding of acarbose, there is a conformation change which move Asp238 toward the OH-groups at C2 and C3 of the residue B, disrupts the old bond between Asp238 and Glu257. As the result, Glu257 moves away to a new position. The substrate binding is expected to cause the same conformation change, which brings the Glu257 to the position, from which it can donate the proton and initiate the catalytic reaction[12].
What should be the function of Asp229? It is found important for the catalytic activity of the enzyme, but due to the close proximity of Asp229 and Asp238, it’s somewhat difficult to clarify the situation. From the 3D-structure of the complex with acarbose, where Asp229 is found close (2,8Å) to the C1 atom of the residue B, it’s suggested that, Asp acts like a nucleophile stabilizing the intermediate by forming new covalent bond.
It’s originally thought that, acarbose is transition state analogue, because it has the residue A with the half-chair conformation. But neither the residue A is found bound to the active site, nor any other residues of acarbose is found distorted toward this conformation. Therefore, it’s most likely that, in CGTase, acarbose is rather a substrate analogue
[12].
The last question remains is, how can the glycosidic bond between B and C residues be not cut by the enzyme itself? The answer may be the deoxy C6-rest of the residue B. There are at least three explanations for this, all emphasizing the importance of the deoxy nature of C6. Apparently, the lack of the O-atom prevents the enzyme to bring acarbose into position, where the glycosidic bond between B and C residues can be cut[12].

ACARBOSE
DaMocles – the students project 29
6.2. Inhibition of barley α-amylase by acarbose
6.2.1. Introduction (text extracted from[13]
):
α–amylases (α -1,4-glucan-4-glucanohydrolase,EC 3.2.1.1) are monomeric enzymes (enzymes consisting of only one monomer) that catalyse the hydrolysis of internal α -D-(1,4) glucosidic linkages in starch and related oligo- and polysaccharides with release of malto-oligosaccharides and glucose in the α-anomeric form. The α-amylases and related amylolytic enzymes representing different specificities are widely occurring, multi-domain proteins that contain a catalytic (β/α)8-barrel. In barley seeds, high and low pI a-amylase isozymes (different enzymes for the same chemical reaction),encoded by two multigene families, are synthesized during germination in response to the phytohormone gibberellic acid. Amino acid sequences deduced from these
gene families, designated AMY1 (low pI) and AMY2 (high pI), consist of 414 and 403 residues, respectively, and show about 80% identity. AMY1 and AMY2, however, display several differences with respect to, for example, affinity for starch granules and soluble substrates, sensitivity to the endogenous α-amylase/subtilisin inhibitor (BASI), Ca2+ affinity and influence on activity, as well as stability at low pH and at elevated temperature[13].
6.2.2. Structure study of AMY2 and its complex with acarbose:
6.2.2.1. Overall structure:
The overall structure of AMY2 upon binding of acarbose is shown in the following figure[13]:
Fig.7: 3D-structure of acarbose-
AMY2-Komplex

ACARBOSE
DaMocles – the students project 30
In the figure, all helices are red, all beta-strands are green and the looping structres are yellow. The calcium ions are white and acarbose is showed with ball-and-stick model. The AMY2 enzyme consists of 3 domains (A,B,C).The carboxy-terminal domain C is at the lower end, consisting of 5 green anti-parallel beta strands. The biggest domain A is at the center of the model, consisting of 8 alpha helices and 8 beta strands (thus the name beta/alpha 8) positioning alternatively along the peptide backbone. This structural motif is also called TIM barrel, after the enzyme triosephosphate isomerase.
(For more information, see: http://en.wikipedia.org/wiki/TIM_barrel)
A small irregular loop domain (domain B) protrudes from domain A connecting strand β3 and helix α3. In AMY2, domain B contains 64 residues and binds three calcium ions, which are most probably critical for proper folding and conformational stability.
Two fragments of acarbose are also visible. One containing 3 rings stays in the middle, at the active binding-site. The other containing 2 rings is on the periphere. The fragmentation is probably caused by the hydrolysis cleavage of acarbose by the enzyme[13]. Both fragments are fixed by many hydrogen bonds with the enzyme’s residues.
Fig.8 Hydrogen bonds between acarbose fragments and active binding-site of
AMY2[13]

ACARBOSE
DaMocles – the students project 31
6.2.2.2. Proposed mechanism.
The widely accepted mechanism for retaining hydrolysis of glycosidic bonds involves a double-displacement mechanism originally proposed by Koshland (1953) and recently reviewed by McCarter & Withers (1994) and Davies &
Henrissat (1995). In this mechanism, proton donor, transition states with half-chair conformation and the intermediate-stabilizing residue (either by the free electron pair or by forming a new covalent bond as a nucleophile) play a fundamental role.
Glu204 and Asp179 are excellent candidates to act as general acid catalyst and catalytic nucleophile, respectively. This interpretation is supported by the distances between the O-atom of Glu204 and the N-atom of acarbose being 2.7 Å and from the O-atom of Asp179 to C1-A atom of acarbose being only 3.2Å . The distance between Glu204 and Asp179 is 5.2 Å, which is a typical distance for a retaining mechanism[13].
Other important residues for the catalytic activity are Asp87,His92, Arg177, , His288 and Asp289, which form hydrogen bonds stabilizing the substrate-enzyme-complex.
The valienamine ring (ring A) of acarbose is considered to be crucial in the inhibition mechanism of α-glucosidases, α-amylases and other amylolytic enzymes. Its unsaturated structure and half-chair conformation are reminiscent of a planar oxocarbonium ion, proposed along the hydrolytic pathway of glycosidases as either a transition state or an intermediate. Of equal importance, its imino group between rings A and B forms a salt-link to the catalytic carboxylate groups. It’s suggested that acarbose is a transition
state analogue[13].

ACARBOSE
DaMocles – the students project 32
7. Medical effects of acarbose
Acarbose is able to prevent dental caries by binding a bacterially produced enzyme, which plays a significant role in initiating dental caries. The bacteria mutans streptococci is an essential initiator of dental caries. It produces extracellular glucosyltransferases (GTF) in large quantities. GTF makes use of the high free energy of the sucrose glycosidic bond to synthezise long-chain glucans. Glucan causes smooth enamel surfaces, which lay the foundation for dental caries. [14]
Via the synthesis of the long-chain glucans both mutans streptococci and plaque get its “cleavability”. [15] Glucan also assists in piling microorganisms in the oral cavity by providing a protected environment for further reproduction for the bacteria. It achieves a fixed anchoring on the tooth with the help of microorganisms which induce harmful plaque. [16] Acarbose inhibits glucosyltransferase by binding specificity at the active site of the enzyme. Thus mutans streptococci cannot initiate harmful plaque and eventually dental caries will not be produced. [17]
Acarbose is also able to lower the blood glucose level of diabetics type 2 by decelerating the breaking down of oligosaccharids. After having a meal the blood glucose level increases. In the pancreas the beta cells of the islets of Langerhans react to it by releasing insulin into the blood. [18]
Insulin is a peptide hormone, which is able to lower the blood sugar concentration. [19] In a healthy organism insulin is accumulated in golgi-vesicles which are able to release it quickly when needed. This process happens in two steps. First insulin is released from the golgi-vesicles in large quantities, which quickly lowers the blood sugar concentration. Then it comes to a finer adjustment in which insulin is released slowly and in the required quantity until the level of blood sugar is decreased to its standard concentration. [20]
In type.2 diabetics, the secretion of insulin is inhibited because the accumulation function of the golgi-vesicles is defective. Thus the glucose level can be controlled only to a small extent and the adjustment takes longer. [21] To avoid this, Acarbose (Glucobay) is administered to the patient.

ACARBOSE
DaMocles – the students project 33
In the duodenum and the jejunum, amylases break down starch and other complex carbohydrates to oligosaccharides. Because oligosaccharides are difficult to resorb, they need to be broken down to monosaccharides. Acid alpha-glucosidases in the intestinal mucosa of the small intestine are responsible for this. Acarbose inhibits these acid alpha-glucosidases as a competitive and reversible inhibitor. Instead of the oligosaccharide, Acarbose is binding at the active site of the enzyme. Thus the oligosaccharides will not break down to monosaccharides and therefore will not be resorbed. Now this happens in the lower small intestine section. Acarbose has got an acarviosyl sub-site unit, which is the significant component in the inhibiting effect. In contrast to alpha-1,4-O-glycosidic bonds of sugars, the enzyme acid alpha-glucosidase is not able to break the N-glycosidic bond of the acarviosyl sub-site unit. [22] Acarbose can thus compensate the defective golgi-vesicles since the quick increase of the blood glucose level after a meal is decelerated and the absorption of glucose is adapted to the defective release of insulin. As a result, the surplus of insulin in the blood is lowered and the patient has got less sugar in the blood. [23]
Even after taking Acarbose for several years its effect on diabetes type 2 does not decline. [24]

ACARBOSE
DaMocles – the students project 34
Preference:
[1]. Biotechnology and molecular biology of the alpha-glucosidase inhibitor
acarbose.
U. F. Wehmeier and W. Piepersberg
Chemische Mikrobiologie, Bergische Universitaet Wuppertal, Gauß-Straße 20,
42097 Wuppertal, Germany
[2].http://www.drugbank.ca/drugs/DB00284
[3].http://www.bikudo.com/product_search/details/27884/acarbose.html
[4]. Acarbose-Metabolismus in Actinoplanes sp. SE50/110; Dissertation; Holger
Thomas; Universität Wuppertal; November 2010
[5]. weiße Biotechnologie bei Bayer HealthCare Product Supply: Mehr als 30 Jahre
Erfahrung; Chemie Ingenieur Technik 2006, 78, No.4; Michael Schedel
[6].http://de.wikipedia.org/wiki/pentosephosphatweg ; 12.6.2010
[7]. Acarbose, ein therapeutisch eingesetzter Wirkstoff: Biosynthese und
Funktion; BIOspektrum 1/04; Udo Wehmeier
[8]. Biosynthetic Studies on the R-Glucosidase Inhibitor Acarbose in Actinoplanes
sp.: 2-epi-5-epi-Valiolone Is the Direct Precursor of the Valienamine Moiety
Taifo Mahmud, Ingo Tornus, Erin Egelkrout, Eckardt Wolf, Charmaine Uy, Heinz G.
Floss, and Sungsook Lee
Contribution from the Department of Chemistry, Box 351700, UniVersity of
Washington,Seattle, Washington 98195-1700
[9]. Three trehalose synthetic pathways in the acarbose-producing Actinoplanes
sp. SN223/29 and evidence for the TreY role in biosynthesis of component C
Jin-Sook Lee & Tran Hai & Hermann Pape &Tae-Jong Kim & Joo-Won Suh
Appl Microbiol Biotechnol (2008) 80:767–778
DOI 10.1007/s00253-008-1582-3
[10].TOTAL SYNTHESIS OF ACARBOSE AND ADIPOSIN-2
YASUSHI SHIBATA AND SEIICHlRO OGAWA
Department of Applied Chemistry, Faculty of Science and Technology, Keio
Universily, Hiyoshi, Kohoku-ku, Yokohama 223 (Japan)
Carbohydrate Research, 189 (1989) 309-322

ACARBOSE
DaMocles – the students project 35
© 1989 Elsevier Science Publishers B.V.
[11]. α- and β-Glucosidase inhibitors: chemical structure and biological activity
Eduardo Borges de Melo, Adriane da Silveira Gomes and Ivone Carvalhob
©2006 Elsevier Ltd.
[12]. X-ray Structure of Cyclodextrin Glycosyltransferase Complexed with
Acarbose. Implications for the Catalytic Mechanism of Glycosidase
Boris Strokopytov Dirk Penninga,Henriette J. Rozeboom,Kor H. Kalk Lubbert
Dijkhuizen,/ and Bauke W. Dijkstra
BIOSON Research Institute and Laboratory of Biophysical Chemistry, University of
Groningen, Nijenborgh 4, 9747 AG
Groningen, The Netherlands, and Department of Microbiology, University of
Groningen, Kerklaan 30, 9751 NN Groningen, The Netherlands
Received August I I, 1994; Revised Manuscript Received November 14, 1994©
[13]. Molecular Structure of a Barley α -Amylase-Inhibitor Complex: Implications
for Starch Binding and Catalysis
Anders Kadziola , Morten Sùgaard , Birte Svensson and Richard Haser
J. Mol. Biol. (1998) 278, 205-217
[14]. Acarbose binding specificity with oral bacterial glucosyltransferase; Kumari
Devulapalle and Gregory Mooser; School of Dentistry, University of Southern
California; p.1288; 17.08.2000
[15].Biochemische Zusammensetzung der in vivo gebildeten Pellikel in
verschiedenen Lokalisationen der Mundhöhle; Dissertation; Roland R. Stigler;
p.12; 2003
[16].Morphologische Studien der Primärkolonisation von pellikelbeschichteten
Schmelzoberflächen am Beispiel Streptococcus mutans; Dissertation; Andrea
Kordes; p.14; 2007
[17].Acarbose binding specificity with oral bacterial glucosyltransferase; Kumari
Devulapalle and Gregory Mooser; School of Dentistry, University of Southern
California; p.1288; 17.08.2000
[18]. http://www.uni-heidelberg.de/uni/presse/rc9/4.html

ACARBOSE
DaMocles – the students project 36
[19]. http://www.3sat.de/nano/glossar/insulin.html
[20]. Diabetes mellitus- Acarbose- Leitfaden für die Praxis; R. Zick, F. Schnitger; S.
9-11; 2. Auflage 2004
[21]. Diabetes mellitus- Acarbose- Leitfaden für die Praxis; R. Zick, F. Schnitger; S.
13; 2. Auflage 2004
[22].http://deposit.ddb.de/cgi-bin/dokserv?idn=964175738&dok_var=d1&dok
_ext=pdf&filename=964175738.pdf 22.06.2010
[23].Diabetes mellitus- Acarbose- Leitfaden für die Praxis; R. Zick, F. Schnitger; S.
38-40; 2. Auflage 2004
[24].Diabetes mellitus- Acarbose- Leitfaden für die Praxis; R. Zick, F. Schnitger; S.
41; 2. Auflage 2004