
Subchronic apocynin treatment attenuates methamphetamine-induced dopamine release and hyperactivity...
6
Subchronic apocynin treatment attenuates methamphetamine-induced dopamine release and hyperactivity in rats Dennis K. Miller a,b, ⁎, Clark E. Oelrichs a,c , Grace Y. Sun d , Agnes Simonyi d a Department of Psychological Sciences, University of Missouri, Columbia, MO, USA b Bond Life Sciences Center, University of Missouri, Columbia, MO, USA c Department of Statistics, University of Missouri, Columbia, MO, USA d Department of Biochemistry, University of Missouri, Columbia, MO, USA abstract article info Article history: Received 17 September 2013 Accepted 20 December 2013 Keywords: Methamphetamine Phenolic Apocynin Dopamine Behavior Aims: The effects of methamphetamine are linked to stimulation of dopaminergic neurons, which can be accompa- nied by production of reactive oxygen species (ROS). Apocynin (4-hydroxy-3-methoxy-acetophenone) is a nicotin- amide adenine dinucleotide phosphate (NADPH)-oxidase (NOX) inhibitor shown to mitigate oxidative stress in a number of models. The present study aimed at testing whether apocynin suppresses the dopamine-releasing and locomotor-activating properties of methamphetamine. Main methods: (1) Apocynin (0.01–100 μM) was applied to rat striatal slices preloaded with [ 3 H]dopamine and its efficacy to evoke [ 3 H]overflow and to alter methamphetamine (3 μM)-evoked [ 3 H]overflow was measured. (2) Groups of rats received apocynin (15 or 50 mg/kg/day) or vehicle injection for seven consecutive days, and the efficacy and potency of methamphetamine to evoke [ 3 H]overflow were determined. (3) Groups of apocynin-treated rats were administered methamphetamine (0.5 or 1 mg/kg) or saline to determine the effect of apocynin on stimulant-induced hyperactivity. Key findings: (1) Apocynin applied to striatal slices did not evoke [ 3 H]overflow or alter methamphetamine- evoked [ 3 H]overflow. (2) However, subchronic apocynin treatment significantly and dose-dependently decreased methamphetamine's potency and efficacy to evoke [ 3 H]overflow. (3) Subchronic apocynin treatment also decreased the locomotor activity evoked by methamphetamine. Significance: Subchronic apocynin treatment diminished methamphetamine induced dopamine-release and its locomotor-activating properties. The pattern of results indicates that apocynin is more effective after repeated, rather than after acute, treatment. The findings also suggest that NOX inhibitors or agents suppressing oxidative stress may constitute a new area for research to understand how methamphetamine produces its deleterious and neurotoxic outcomes in the brain. © 2014 Elsevier Inc. All rights reserved. Introduction Apocynin (4-hydroxy-3-methoxy-acetophenone, acetovanillone) is a phenolic compound originally isolated from the medicinal plant Picrorhiza kurroa and a variety of Apocynum species in North America and Asia. It has been investigated for its potential use as a prophylactic against the adverse neurochemical and behavioral outcomes of Parkinson's disease [1–3], a pathology characterized by degeneration of striatal dopamine neurons. The phenolic can improve functional outcomes from damage when administered systemically (e.g., peroral infusion or parenteral injection), and its efficacy is typically greater when applied before, rather than after, the insult or induction of the disease state [1,2]. A significant, causative factor in Parkinson's disease and its neural degeneration is initiation of oxidative stress and the production of reac- tive oxygen species (ROS) [4,5]. Apocynin has antioxidant properties and is a selective inhibitor (IC 50 value ≈ 10 μM) of nicotinamide adenine dinucleotide phosphate (NADPH)-oxidase (NOX) [6], an enzyme responsible for a major source of ROS production in the brain [2,7]. The mechanism through which apocynin inhibits NOX is unclear, although one candidate is interference with the assembly of NOX com- ponents (e.g., the cytosolic subunit p47phox) [7]. Apocynin-induced NOX inhibition has been shown to ameliorate the deleterious impact of dopaminergic neurons, including 6-hydroxydopamine (6-OHDA) [8], rotenone [9], 1-methyl-4-phenylpyridinium (MPP + ) [10] and para- quat [11]. Park and colleagues recently reported that psychostimulant methamphetamine (10 μM) induced an increase in phosphorylation of the p47phox NOX subunit and subsequently enhanced NOX activity Life Sciences 98 (2014) 6–11 ⁎ Corresponding author at: Department of Psychological Sciences, 203 McAlester Hall, University of Missouri, Columbia, MO 65211, USA. Tel.: +1 573 884 8141; fax: +1 573 882 7710. E-mail address: [email protected] (D.K. Miller). 0024-3205/$ – see front matter © 2014 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.lfs.2013.12.031 Contents lists available at ScienceDirect Life Sciences journal homepage: www.elsevier.com/locate/lifescie
Transcript of Subchronic apocynin treatment attenuates methamphetamine-induced dopamine release and hyperactivity...

Life Sciences 98 (2014) 6–11
Contents lists available at ScienceDirect
Life Sciences
j ourna l homepage: www.e lsev ie r .com/ locate / l i fesc ie
Subchronic apocynin treatment attenuates methamphetamine-induceddopamine release and hyperactivity in rats
Dennis K. Miller a,b,⁎, Clark E. Oelrichs a,c, Grace Y. Sun d, Agnes Simonyi d
a Department of Psychological Sciences, University of Missouri, Columbia, MO, USAb Bond Life Sciences Center, University of Missouri, Columbia, MO, USAc Department of Statistics, University of Missouri, Columbia, MO, USAd Department of Biochemistry, University of Missouri, Columbia, MO, USA
⁎ Corresponding author at: Department of PsychologicUniversity of Missouri, Columbia, MO 65211, USA. Tel.: +882 7710.
E-mail address: [email protected] (D.K. Miller).
0024-3205/$ – see front matter © 2014 Elsevier Inc. All rihttp://dx.doi.org/10.1016/j.lfs.2013.12.031
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 17 September 2013Accepted 20 December 2013Keywords:MethamphetaminePhenolicApocyninDopamineBehavior
Aims: The effects of methamphetamine are linked to stimulation of dopaminergic neurons, which can be accompa-nied by production of reactive oxygen species (ROS). Apocynin (4-hydroxy-3-methoxy-acetophenone) is a nicotin-amide adenine dinucleotide phosphate (NADPH)-oxidase (NOX) inhibitor shown to mitigate oxidative stress in anumber of models. The present study aimed at testing whether apocynin suppresses the dopamine-releasing andlocomotor-activating properties of methamphetamine.Main methods: (1) Apocynin (0.01–100 μM) was applied to rat striatal slices preloaded with [3H]dopamine andits efficacy to evoke [3H]overflow and to alter methamphetamine (3 μM)-evoked [3H]overflow was measured.(2) Groups of rats received apocynin (15 or 50 mg/kg/day) or vehicle injection for seven consecutive days,and the efficacy and potency of methamphetamine to evoke [3H]overflow were determined. (3) Groups of
apocynin-treated rats were administered methamphetamine (0.5 or 1 mg/kg) or saline to determine the effectof apocynin on stimulant-induced hyperactivity.Key findings: (1) Apocynin applied to striatal slices did not evoke [3H]overflow or alter methamphetamine-evoked [3H]overflow. (2) However, subchronic apocynin treatment significantly and dose-dependentlydecreased methamphetamine's potency and efficacy to evoke [3H]overflow. (3) Subchronic apocynin treatmentalso decreased the locomotor activity evoked by methamphetamine.Significance: Subchronic apocynin treatment diminished methamphetamine induced dopamine-release and itslocomotor-activating properties. The pattern of results indicates that apocynin is more effective after repeated,rather than after acute, treatment. The findings also suggest that NOX inhibitors or agents suppressing oxidativestress may constitute a new area for research to understand how methamphetamine produces its deleteriousand neurotoxic outcomes in the brain.© 2014 Elsevier Inc. All rights reserved.
Introduction
Apocynin (4-hydroxy-3-methoxy-acetophenone, acetovanillone) isa phenolic compound originally isolated from the medicinal plantPicrorhiza kurroa and a variety of Apocynum species in North Americaand Asia. It has been investigated for its potential use as a prophylacticagainst the adverse neurochemical and behavioral outcomes ofParkinson's disease [1–3], a pathology characterized by degenerationof striatal dopamine neurons. The phenolic can improve functionaloutcomes from damage when administered systemically (e.g., peroralinfusion or parenteral injection), and its efficacy is typically greater
al Sciences, 203 McAlester Hall,1 573 884 8141; fax: +1 573
ghts reserved.
when applied before, rather than after, the insult or induction of thedisease state [1,2].
A significant, causative factor in Parkinson's disease and its neuraldegeneration is initiation of oxidative stress and the production of reac-tive oxygen species (ROS) [4,5]. Apocynin has antioxidant propertiesand is a selective inhibitor (IC50 value ≈ 10 μM) of nicotinamideadenine dinucleotide phosphate (NADPH)-oxidase (NOX) [6], anenzyme responsible for a major source of ROS production in the brain[2,7]. The mechanism through which apocynin inhibits NOX is unclear,although one candidate is interference with the assembly of NOX com-ponents (e.g., the cytosolic subunit p47phox) [7]. Apocynin-inducedNOX inhibition has been shown to ameliorate the deleterious impactof dopaminergic neurons, including 6-hydroxydopamine (6-OHDA)[8], rotenone [9], 1-methyl-4-phenylpyridinium (MPP+) [10] and para-quat [11]. Park and colleagues recently reported that psychostimulantmethamphetamine (10 μM) induced an increase in phosphorylationof the p47phox NOX subunit and subsequently enhanced NOX activity

7D.K. Miller et al. / Life Sciences 98 (2014) 6–11
in endothelial cells [12]. The impact of apocynin on methamphetamineand NOX activity in the brain has not been investigated.
Methamphetamine induces the release of dopamine from intra- andextraneuronal stores in the brain [13,14]. Elevated intracellular dopa-mine levels can lead to ROSproduction and severe neurotoxicity, similarto that observed in Parkinson's disease, especially after repeated and/orhigh-dose methamphetamine administration [15,16]. However,methamphetamine's efficacy to increase dopamine in the brain isalso associated with its behavior-activating, reinforcing, and addictiveproperties [17,18]. Whether apocynin can ameliorate the dopamine re-lease evokedbymethamphetaminehas not been investigated. Apocynin'sreported ability to diminish the effects of dopamine-related ligands in thebrain [8–11] andmethamphetamine's NOX activation in endothelial cells[12], suggest that apocynin shouldminimizemethamphetamine's effects.The goal of the present study was to determine if acute and subchronicadministration of apocynin could alter the efficacy of methamphetamineto evoke [3H]dopamine release from rat striatal slices. As methamphet-amine increases locomotor activity after acute administration to rodents[17–19] and based on the present neurochemical findings, the effect ofsubchronic apocynin treatment onmethamphetamine-induced hyperac-tivity in rats also was determined.
Materials and methods
Subjects
Male Sprague–Dawley rats (Harlan, Indianapolis, IN; ~200 g uponarrival to the laboratory) were used for all experiments. The colonywas maintained under a 12-hour/12-hour light/dark cycle and the ex-periments were conducted during the light phase of the cycle. Ratswere housed 2 rats per cage and received ad libitum access to standardrodent chow and tap water. All animal procedures were approved bythe Institutional Animal Care and Use Committee of the University ofMissouri.
Drugs and chemicals
Apocynin and (±)-methamphetamine hydrochloride were pur-chased from Sigma-Aldrich (St. Louis, MO). Ascorbic acid, glucoseand pargyline hydrochloride were purchased from Acros Organics(Fairlawn, NJ). Radiolabeled dopamine (dihydroxyphenylethylamine3,4-[ring-2,5,6-3H]; specific activity ~25 Ci/mmol) was purchased fromPerkinElmer Life Sciences (Boston, MA). All other chemicals were pur-chased from Fisher Scientific (Pittsburgh, PA).
Throughout the study, all drug concentrations and doses representthe free base weight. For the behavioral experiments, apocynin andmethamphetamine were prepared in saline (0.9% w/v). All injectionswere administered at a volume of 1 ml solution/kg body weight.Apocynin was administered i.p. and methamphetamine was adminis-tered s.c.
Effect of apocynin to evoke [3H]overflow
This experiment determined the intrinsic activity of a range ofapocynin concentrations to evoke [3H]overflow from rat striatal slicespreloaded with [3H]dopamine. This experiment used a within-subjectsdesign, such that the concentration–response for apocynin was deter-mined in slices from a single rat (n = 5 rats).
Rats were euthanized via rapid decapitation and dorsal striata weredissected and sliced (750 μm thick slices). Slices were incubated inoxygenated (95% 02/5% CO2) buffer (in mM, 108 NaCl, 25 NaHCO3,11.1 glucose, 4.7 KCl, 1.3 CaCl2, 1.2 MgSO4, 1.0 Na2HPO4, 0.11 ascorbicacid, 0.005 EDTA; pH 7.4) in a metabolic shaker at 37 °C for 30 min.Slices were transferred to fresh buffer, [3H]dopamine (0.1 μM) wasadded, and slices were incubated for an additional 30 min. Each slicewas then transferred to a reaction chamber (0.2 ml) bounded by glass
microfiberfilters (GF/B,Whatman,Madistoine, England) in an automatedsystem (Suprafusion 2500, Brandel, Gaithersburg, MD). Pargyline(10 μM) was added to the buffer that was used for the remainder of theexperiment to inhibit monoamine oxidase (MAO) [20]. Slices weresuperfused at a rate of 0.75 ml/min for 60 min, a period that our prelim-inary experiments indicate is required to achieve consistent and stablebasal [3H]overflow.
After the 60 min of preliminary superfusion, sample collection com-menced at a rate of 1 sample/3 min and the time course for the samplecollection period is presented in panel A of Fig. 1. Slices were superfusedwith only buffer for 9 min. Apocynin (0.01–100 μM)was added to bufferand slices were superfused for 9 min. The apocynin concentration rangeincludes those that inhibit (IC50 value ≈ 10 μM) NOX activity [6]. Oneslice was superfused with buffer that did not contain apocynin as a con-trol condition. All slices were then superfused with buffer that did notcontain apocynin for an additional 9 min. Slices and filterswere removedfrom the chambers and solubilized (0.6N ScintiGest) for at least 24 h.Radioactivity in superfusate samples and the slices/filters was measuredby liquid scintillation (LS 6500 Scintillation Counter, Beckman-Coulter,Fullerton, CA; counting efficiency ≈ 60%).
Fractional release for each 3-minute superfusate sample was calcu-lated by dividing the [3H] collected in each sample by the overall [3H]present in the tissue at the time of sample collection. Basal [3H]overflowwas calculated from the average of fractional release in the three sam-ples before the addition of apocynin. The fractional samples after the ad-dition of apocynin that were greater than basal were summed, and thenbasal outflow was subtracted to determine the dependent measure,total [3H]overflow. A one-way repeated measures analysis of variance(ANOVA) was performed (IBM SPSS, version 20) on total [3H]overflowvalues with apocynin concentration as a within-subjects factor. Tukeypost hoc tests were performed when appropriate (p b 0.05).
Effect of apocynin on methamphetamine-evoked [3H]overflow
The effect of apocynin, applied to slices, on methamphetamine-evoked [3H]overflow was determined. This experiment used a within-subjects design, such that the effect of methampethamine acrossapocynin concentrations was determined in slices from a single rat(n = 6 rats).
Slices and buffer were prepared as described for the previous ex-periment. After 60 min of preliminary superfusion, sample collectioncommenced and the time course for the sample collection period ispresented in panel B of Fig. 1. Slices were superfused with bufferfor 9 min. Apocynin (0.01–100 μM) was added to buffer and sliceswere superfused for 6 min. Methamphetamine (3 μM) was addedand slices were superfused with buffer containing apocynin andmethamphetamine for 9 min. The methamphetamine concentrationwas selected based on our previous work as a concentration that issubject to inhibition and enhancement by dopamine-regulating ligands[21–24]. As control conditions, one slicewas superfusedwith only bufferand one slice was superfused with only methamphetamine. All sliceswere then superfused with buffer that did not contain apocynin ormethamphetamine for an additional 9 min.
Basal [3H]overflowwas calculated from the average of fractional re-lease in thefive samples before the addition ofmethamphetamine. Total[3H]overflowduring superfusionwithmethamphetaminewas analyzedvia one-way repeatedmeasures ANOVAwith apocynin concentration asa within-subjects factor.
Effect of subchronic apocynin treatment on methamphetamine-evoked[3H]overflow
To determine if subchronic apocynin treatment altersmethamphetamine-evoked [3H]overflow, rats were administered 15or 50 mg/kg apocynin or saline vehicle once-daily for 7 consecutivedays (days 1–7). These parameters were selected from previous work

Fig. 1. Time course for the sample collection periods in the three [3H]overflow experiments. The solid lines indicate the periods when apocynin or methamphetamine was included.
8 D.K. Miller et al. / Life Sciences 98 (2014) 6–11
demonstrating reduced ROS activity without adverse effects in rodents[25–27]. Three apocynin treatment groups (n = 6–9 rats/group) wereformed — vehicle, a low (15 mg/kg) apocynin dose, and a high(50 mg/kg) apocynin dose. The [3H]overflow component of the experi-ment was a within-subjects design, such that the concentration–re-sponse formethamphetaminewas determined in slices froma single rat.
On the day (day 8) following the last apocynin injection, slices andbuffer were prepared as described for the previous experiments. After60 min of preliminary superfusion, sample collection commenced. Thetime course for the sample collection period is presented in panel C ofFig. 1. Slices were superfused with buffer for 9 min. Methamphetamine(0.03–60 μM)was added to buffer and sliceswere superfused for 9 min.One slice from each rat was superfused with only buffer as a controlcondition. All slices were then superfused with buffer that did not con-tain methamphetamine for an additional 9 min.
Basal [3H]overflow was calculated from the average of fractionalrelease in the three samples before the addition of methamphetamine.Total [3H]overflow during superfusion with methamphetamine wasanalyzed via two-way repeated measures ANOVA with apocynin doseas a between-groups factor and methamphetamine concentration as awithin-subjects factor. Methamphetamine dose–response curves fromeach rat were determined via nonlinear regression and EC50 valueswere calculated. The log-transformed EC50 values were analyzed viaone-way ANOVA with apocynin dose as a between-groups factor.
Effect of subchronic apocynin treatment on methamphetamine-inducedchanges in locomotor activity
To determine the effect of subchronic apocynin treatment onmethamphetamine-induced hyperactivity, rats were administered 15or 50 mg/kg apocynin or saline vehicle once-daily for 7 consecutivedays (days 1–7). On days 6 and 7, rats were acclimated to standardautomated activity monitors (Med Associates, St. Albans, VT) describedpreviously [19]. On day 8, rats were placed in the monitor for 15 min,injectedwithmethamphetamine (0.5 or 1 mg/kg) or saline, and returnedto the monitor for 75 min. The methamphetamine doses were selectedbased on our previous study [19] as those that produce a transient
increase in locomotor activity without stereotypy or other untoward ef-fects (e.g., seizures). In the design of the experiment nine treatmentgroups were formed—Vehicle–Saline, Vehicle–Methamphetamine (0.5and 1 mg/kg), Apocynin (15 and 50 mg/kg)–Saline, and Apocynin (15and 50 mg/kg)–Methamphetamine (15 and 50 mg/kg).
Distance traveled (in cm) was calculated by the monitoring systemand evaluated in 5-minute intervals over the 90-minute session. Datafrom the 75-minute period after methamphetamine injection wereanalyzed via three-way repeated measures ANOVA with apocynindose and methamphetamine dose as between-group factors and time(5-mintue intervals) as a within-subjects factor.
Results
Acute administration of apocynin does not evoke [3H]overflow or altermethamphetamine-evoked [3H]overflow
Fig. 2 presents total [3H]overflow from superfusion with apocynin inthe absence (0 μM methamphetamine) or presence of methamphet-amine (3 μM). Apocynin did not have intrinsic activity to evoke [3H]overflow, relative to the [3H]overflow observed in the absence ofapocynin.
Methamphetamine induced an increase in [3H]overflow, relativeto the [3H]overflow observed in the absence of methamphetamine.However, none of the apocynin concentrations significantly alteredmethamphetamine-evoked [3H]overflow [F(5,20) = 0.94, p = 0.48].
Subchronic apocynin treatment attenuates methamphetamine-evoked[3H]overflow
Fig. 3 presents methamphetamine-evoked [3H]overflow from ratsthat received subchronic apocynin or vehicle treatment. Methamphet-amine produced a concentration-dependent increase in [3H]overflow, asexpected [F(9,171) = 54.26, p b 0.001]. Subchronic apocynin treatmentdecreased in methamphetamine-evoked [3H]overflow [F(18,171) =2.80, p b 0.01].
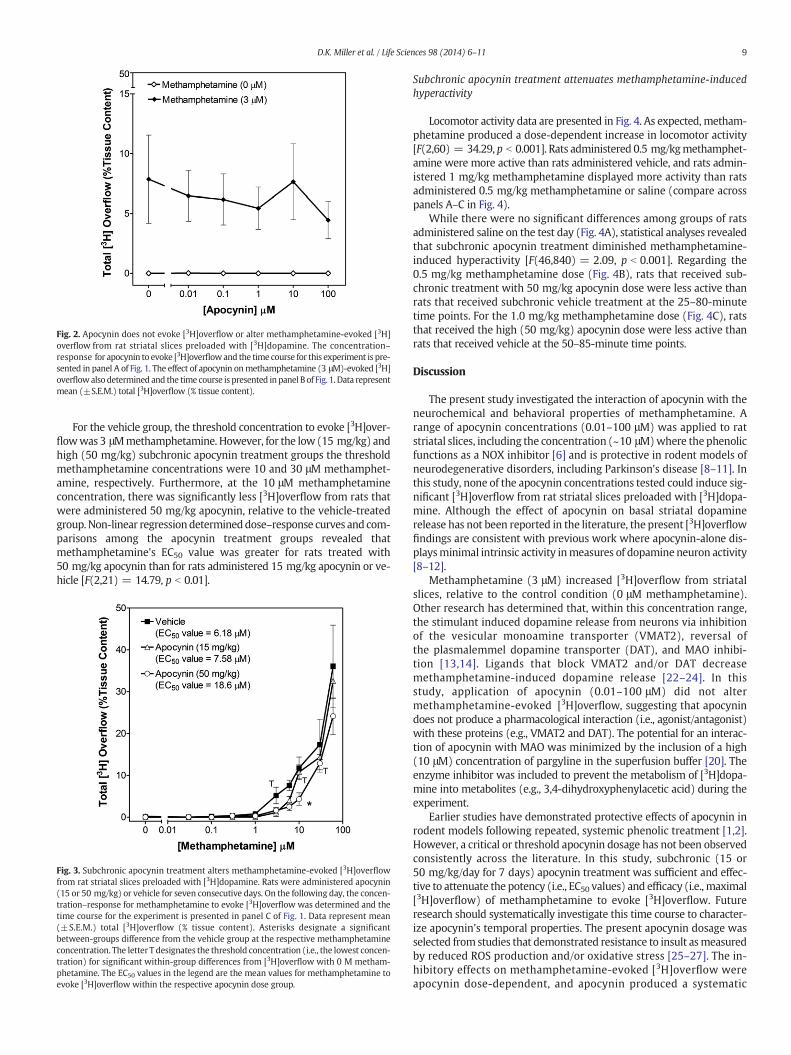
Fig. 2. Apocynin does not evoke [3H]overflow or alter methamphetamine-evoked [3H]overflow from rat striatal slices preloaded with [3H]dopamine. The concentration–response for apocynin to evoke [3H]overflowand the time course for this experiment is pre-sented in panel A of Fig. 1. The effect of apocynin onmethamphetamine (3 μM)-evoked [3H]overflowalso determined and the time course is presented in panel B of Fig. 1. Data representmean (±S.E.M.) total [3H]overflow (% tissue content).
9D.K. Miller et al. / Life Sciences 98 (2014) 6–11
For the vehicle group, the threshold concentration to evoke [3H]over-flowwas 3 μMmethamphetamine. However, for the low (15 mg/kg) andhigh (50 mg/kg) subchronic apocynin treatment groups the thresholdmethamphetamine concentrations were 10 and 30 μM methamphet-amine, respectively. Furthermore, at the 10 μM methamphetamineconcentration, there was significantly less [3H]overflow from rats thatwere administered 50 mg/kg apocynin, relative to the vehicle-treatedgroup.Non-linear regressiondetermineddose–response curves and com-parisons among the apocynin treatment groups revealed thatmethamphetamine's EC50 value was greater for rats treated with50 mg/kg apocynin than for rats administered 15 mg/kg apocynin or ve-hicle [F(2,21) = 14.79, p b 0.01].
Fig. 3. Subchronic apocynin treatment alters methamphetamine-evoked [3H]overflowfrom rat striatal slices preloaded with [3H]dopamine. Rats were administered apocynin(15 or 50 mg/kg) or vehicle for seven consecutive days. On the following day, the concen-tration–response for methamphetamine to evoke [3H]overflow was determined and thetime course for the experiment is presented in panel C of Fig. 1. Data represent mean(±S.E.M.) total [3H]overflow (% tissue content). Asterisks designate a significantbetween-groups difference from the vehicle group at the respective methamphetamineconcentration. The letter T designates the threshold concentration (i.e., the lowest concen-tration) for significant within-group differences from [3H]overflow with 0 M metham-phetamine. The EC50 values in the legend are the mean values for methamphetamine toevoke [3H]overflow within the respective apocynin dose group.
Subchronic apocynin treatment attenuates methamphetamine-inducedhyperactivity
Locomotor activity data are presented in Fig. 4. As expected, metham-phetamine produced a dose-dependent increase in locomotor activity[F(2,60) = 34.29, p b 0.001]. Rats administered 0.5 mg/kgmethamphet-amine were more active than rats administered vehicle, and rats admin-istered 1 mg/kg methamphetamine displayed more activity than ratsadministered 0.5 mg/kg methamphetamine or saline (compare acrosspanels A–C in Fig. 4).
While there were no significant differences among groups of ratsadministered saline on the test day (Fig. 4A), statistical analyses revealedthat subchronic apocynin treatment diminished methamphetamine-induced hyperactivity [F(46,840) = 2.09, p b 0.001]. Regarding the0.5 mg/kg methamphetamine dose (Fig. 4B), rats that received sub-chronic treatment with 50 mg/kg apocynin dose were less active thanrats that received subchronic vehicle treatment at the 25–80-minutetime points. For the 1.0 mg/kg methamphetamine dose (Fig. 4C), ratsthat received the high (50 mg/kg) apocynin dose were less active thanrats that received vehicle at the 50–85-minute time points.
Discussion
The present study investigated the interaction of apocynin with theneurochemical and behavioral properties of methamphetamine. Arange of apocynin concentrations (0.01–100 μM) was applied to ratstriatal slices, including the concentration (~10 μM)where the phenolicfunctions as a NOX inhibitor [6] and is protective in rodent models ofneurodegenerative disorders, including Parkinson's disease [8–11]. Inthis study, none of the apocynin concentrations tested could induce sig-nificant [3H]overflow from rat striatal slices preloaded with [3H]dopa-mine. Although the effect of apocynin on basal striatal dopaminerelease has not been reported in the literature, the present [3H]overflowfindings are consistent with previous work where apocynin-alone dis-playsminimal intrinsic activity inmeasures of dopamineneuron activity[8–12].
Methamphetamine (3 μM) increased [3H]overflow from striatalslices, relative to the control condition (0 μM methamphetamine).Other research has determined that, within this concentration range,the stimulant induced dopamine release from neurons via inhibitionof the vesicular monoamine transporter (VMAT2), reversal ofthe plasmalemmel dopamine transporter (DAT), and MAO inhibi-tion [13,14]. Ligands that block VMAT2 and/or DAT decreasemethamphetamine-induced dopamine release [22–24]. In thisstudy, application of apocynin (0.01–100 μM) did not altermethamphetamine-evoked [3H]overflow, suggesting that apocynindoes not produce a pharmacological interaction (i.e., agonist/antagonist)with these proteins (e.g., VMAT2 and DAT). The potential for an interac-tion of apocynin with MAO was minimized by the inclusion of a high(10 μM) concentration of pargyline in the superfusion buffer [20]. Theenzyme inhibitor was included to prevent the metabolism of [3H]dopa-mine into metabolites (e.g., 3,4-dihydroxyphenylacetic acid) during theexperiment.
Earlier studies have demonstrated protective effects of apocynin inrodent models following repeated, systemic phenolic treatment [1,2].However, a critical or threshold apocynin dosage has not been observedconsistently across the literature. In this study, subchronic (15 or50 mg/kg/day for 7 days) apocynin treatment was sufficient and effec-tive to attenuate the potency (i.e., EC50 values) and efficacy (i.e., maximal[3H]overflow) of methamphetamine to evoke [3H]overflow. Futureresearch should systematically investigate this time course to character-ize apocynin's temporal properties. The present apocynin dosage wasselected from studies that demonstrated resistance to insult asmeasuredby reduced ROS production and/or oxidative stress [25–27]. The in-hibitory effects on methamphetamine-evoked [3H]overflow wereapocynin dose-dependent, and apocynin produced a systematic
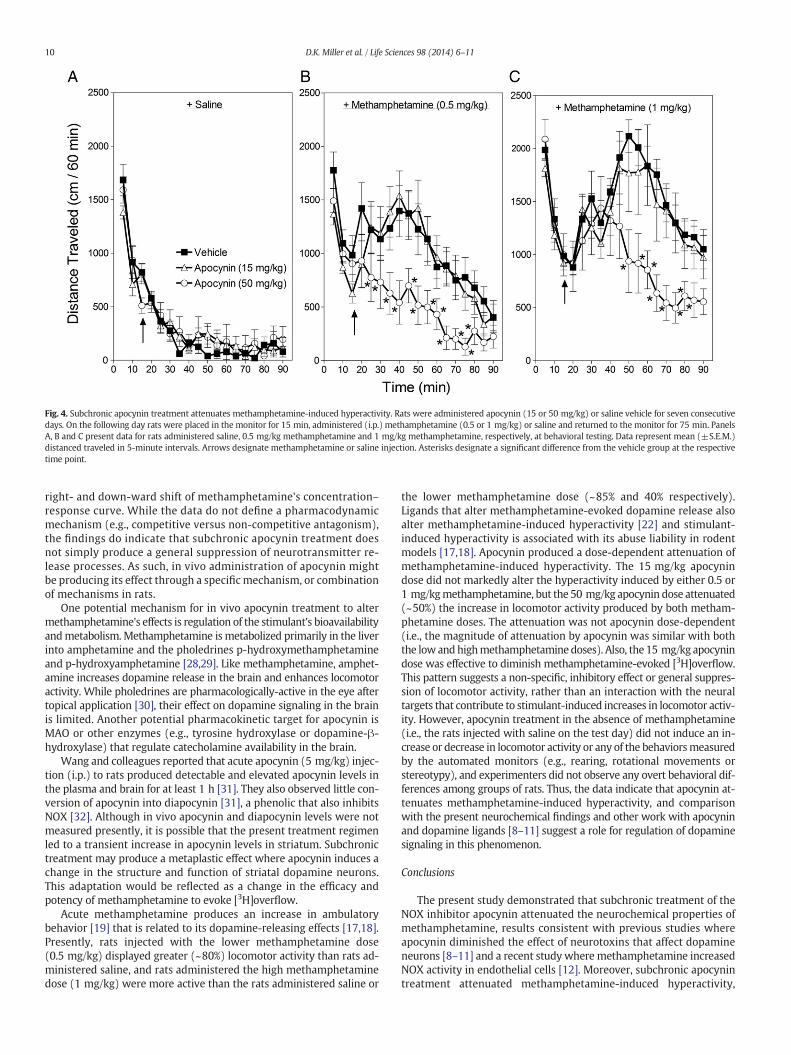
Fig. 4. Subchronic apocynin treatment attenuates methamphetamine-induced hyperactivity. Rats were administered apocynin (15 or 50 mg/kg) or saline vehicle for seven consecutivedays. On the following day rats were placed in the monitor for 15 min, administered (i.p.) methamphetamine (0.5 or 1 mg/kg) or saline and returned to the monitor for 75 min. PanelsA, B and C present data for rats administered saline, 0.5 mg/kg methamphetamine and 1 mg/kg methamphetamine, respectively, at behavioral testing. Data represent mean (±S.E.M.)distanced traveled in 5-minute intervals. Arrows designate methamphetamine or saline injection. Asterisks designate a significant difference from the vehicle group at the respectivetime point.
10 D.K. Miller et al. / Life Sciences 98 (2014) 6–11
right- and down-ward shift of methamphetamine's concentration–response curve. While the data do not define a pharmacodynamicmechanism (e.g., competitive versus non-competitive antagonism),the findings do indicate that subchronic apocynin treatment doesnot simply produce a general suppression of neurotransmitter re-lease processes. As such, in vivo administration of apocynin mightbe producing its effect through a specific mechanism, or combinationof mechanisms in rats.
One potential mechanism for in vivo apocynin treatment to altermethamphetamine's effects is regulation of the stimulant's bioavailabilityandmetabolism.Methamphetamine is metabolized primarily in the liverinto amphetamine and the pholedrines p-hydroxymethamphetamineand p-hydroxyamphetamine [28,29]. Like methamphetamine, amphet-amine increases dopamine release in the brain and enhances locomotoractivity. While pholedrines are pharmacologically-active in the eye aftertopical application [30], their effect on dopamine signaling in the brainis limited. Another potential pharmacokinetic target for apocynin isMAO or other enzymes (e.g., tyrosine hydroxylase or dopamine-β-hydroxylase) that regulate catecholamine availability in the brain.
Wang and colleagues reported that acute apocynin (5 mg/kg) injec-tion (i.p.) to rats produced detectable and elevated apocynin levels inthe plasma and brain for at least 1 h [31]. They also observed little con-version of apocynin into diapocynin [31], a phenolic that also inhibitsNOX [32]. Although in vivo apocynin and diapocynin levels were notmeasured presently, it is possible that the present treatment regimenled to a transient increase in apocynin levels in striatum. Subchronictreatment may produce a metaplastic effect where apocynin induces achange in the structure and function of striatal dopamine neurons.This adaptation would be reflected as a change in the efficacy andpotency of methamphetamine to evoke [3H]overflow.
Acute methamphetamine produces an increase in ambulatorybehavior [19] that is related to its dopamine-releasing effects [17,18].Presently, rats injected with the lower methamphetamine dose(0.5 mg/kg) displayed greater (~80%) locomotor activity than rats ad-ministered saline, and rats administered the high methamphetaminedose (1 mg/kg) were more active than the rats administered saline or
the lower methamphetamine dose (~85% and 40% respectively).Ligands that alter methamphetamine-evoked dopamine release alsoalter methamphetamine-induced hyperactivity [22] and stimulant-induced hyperactivity is associated with its abuse liability in rodentmodels [17,18]. Apocynin produced a dose-dependent attenuation ofmethamphetamine-induced hyperactivity. The 15 mg/kg apocynindose did not markedly alter the hyperactivity induced by either 0.5 or1 mg/kgmethamphetamine, but the 50 mg/kg apocynin dose attenuated(~50%) the increase in locomotor activity produced by both metham-phetamine doses. The attenuation was not apocynin dose-dependent(i.e., the magnitude of attenuation by apocynin was similar with boththe low andhighmethamphetamine doses). Also, the 15 mg/kg apocynindose was effective to diminish methamphetamine-evoked [3H]overflow.This pattern suggests a non-specific, inhibitory effect or general suppres-sion of locomotor activity, rather than an interaction with the neuraltargets that contribute to stimulant-induced increases in locomotor activ-ity. However, apocynin treatment in the absence of methamphetamine(i.e., the rats injected with saline on the test day) did not induce an in-crease or decrease in locomotor activity or any of the behaviorsmeasuredby the automated monitors (e.g., rearing, rotational movements orstereotypy), and experimenters did not observe any overt behavioral dif-ferences among groups of rats. Thus, the data indicate that apocynin at-tenuates methamphetamine-induced hyperactivity, and comparisonwith the present neurochemical findings and other work with apocyninand dopamine ligands [8–11] suggest a role for regulation of dopaminesignaling in this phenomenon.
Conclusions
The present study demonstrated that subchronic treatment of theNOX inhibitor apocynin attenuated the neurochemical properties ofmethamphetamine, results consistent with previous studies whereapocynin diminished the effect of neurotoxins that affect dopamineneurons [8–11] and a recent study where methamphetamine increasedNOX activity in endothelial cells [12]. Moreover, subchronic apocynintreatment attenuated methamphetamine-induced hyperactivity,

11D.K. Miller et al. / Life Sciences 98 (2014) 6–11
demonstrating translation in efficacy from the brain to behavior. Sincethe neurotoxic and addictive properties of methamphetamine areclosely related to its ability to increase dopamine levels in the brainand increase hyperactivity [15–18], results from present study suggest aregulatory role for apocynin in methamphetamine neuropsycho-pharmacology. Although much additional research is required in thisline, these results suggest the potential of apocynin and possibly otherNOX inhibitors towards the development of pharmacotherapies formethamphetamine addiction and intoxication.
Role of funding source
This research was partially supported by a grant from the NationalInstitutes of Health (AT003859). It had no further roles in the studydesign; in the collection, analysis and interpretation of data; in thewritingof the report; or in the decision to submit the paper for publication.
Conflict of interest
The authors have no other funding sources or conflicts of interest to report.
Acknowledgments
The authors appreciate the assistance of Andrew S. Sage andDouglasKeightley in the conduct of this research.
References
Simonyi A, Serfozo P, Lehmidi TM, Cui J, Gu Z, LubahnDB, et al. The neuroprotective effectsof apocynin. Front Biosci (Elite. Ed) 2012];4:2183–93.
Stefanska J, Pawliczak R. Apocynin: molecular aptitudes. Mediat Inflamm 2008];2008:106507.
SunAY,WangQ, Simonyi A, SunGY. Botanical phenolics and brain health. NeuromolecularMed 2008];10:259–74.
Adibhatla RM, Hatcher JF. Lipid oxidation and peroxidation in CNS health and disease:from molecular mechanisms to therapeutic opportunities. Antioxid Redox Signal2010];12:125–69.
Sorce S, KrauseKH. NOXenzymes in the central nervous system: from signaling to disease.Antioxid Redox Signal 2009];11:2481–504.
Simons JM, Hart BA, Vai Ching TRIp, Van DH, Labadie RP. Metabolic activation of naturalphenols into selective oxidative burst agonists by activated human neutrophils.Free Radic Biol Med 1990];8:251–8.
Stolk J, Hiltermann TJ, Dijkman JH, Verhoeven AJ. Characteristics of the inhibition ofNADPH oxidase activation in neutrophils by apocynin, amethoxy-substituted catechol.Am J Respir Cell Mol Biol 1994];11:95–102.
Rodriguez-Pallares J, Parga JA, Munoz A, Rey P, Guerra MJ, Labandeira-Garcia JL. Mecha-nism of 6-hydroxydopamine neurotoxicity: the role of NADPH oxidase andmicroglialactivation in 6-hydroxydopamine-induced degeneration of dopaminergic neurons. JNeurochem 2007];103:145–56.
Kumar A, Singh BK, Ahmad I, Shukla S, Patel DK, Srivastava G, et al. Involvement ofNADPH oxidase and glutathione in zinc-induced dopaminergic neurodegenerationin rats: similarity with paraquat neurotoxicity. Brain Res 2012];1438:48–64.
Anantharam V, Kaul S, Song C, Kanthasamy A, Kanthasamy AG. Pharmacological inhibi-tion of neuronal NADPH oxidase protects against 1-methyl-4-phenylpyridinium
(MPP+)-induced oxidative stress and apoptosis in mesencephalic dopaminergicneuronal cells. Neurotoxicol 2007];28:988–97.
Cristovao AC, Choi DH, Baltazar G, Beal MF, Kim YS. The role of NADPH oxidase 1-derivedreactive oxygen species in paraquat-mediated dopaminergic cell death. AntioxidRedox Signal 2009];11:2105–18.
Park M, Hennig B, Toborek M. Methamphetamine alters occludin expression via NADPHoxidase-induced oxidative insult and intact caveolae. J Cell Mol Med 2012];16:362–75.
Seiden LS, Sabol KE, Ricaurte GA. Amphetamine: effects on catecholamine systems andbehavior. Ann Rev Pharmacol Toxicol 1993];33:639–77.
Sulzer D, Sonders MS, Poulsen NW, Galli A. Mechanism of neurotransmitter release byamphetamines: a review. Prog Neurobiol 2005];75:406–33.
Krasnova IN, Cadet JL. Methamphetamine toxicity andmessengers of death. Brain Res Rev2009];60:379–407.
Quinton MS, Yamamoto BK. Causes and consequences of methamphetamine and MDMAtoxicity. AAPS J 2006];8:E337–47.
Robinson TE, Berridge KC. The neural basis of drug craving: an incentive-sensitizationtheory of addiction. Brain Res Brain Res Rev 1993];18:247–91.
Wise RA, Bozarth MA. A psychomotor stimulant theory of addiction. Psychol Rev1987];94:469–92.
Sage AS, Miller DK. What can rodent activity tell us about methamphetamine abuse. In:Ornoy J, He X, editors. Methamphetamines: Abuse, Health Effects and TreatmentOptions. New York: Nova Science Publishers; 2012]. p. 47–82.
Zumstein A, KarduckW, Starke K. Pathways of dopaminemetabolism in the rabbit caudatenucleus in vitro. Naunyn Schmiedebergs Arch Pharmacol 1981];316:205–17.
Dopheide MM, Morgan RE, Rodvelt KR, Schachtman TR, Miller DK. Modafinil evokesstriatal [3H]dopamine release and alters the subjective properties of stimulants. EurJ Pharmacol 2007];568:112–23.
Miller DK, Crooks PA, Teng L, Witkin JM, Munzar P, Goldberg SR, et al. Lobeline inhibitsthe neurochemical and behavioral effects of amphetamine. J Pharmacol Exp Ther2001];296:1023–34.
Beckmann JS, Siripurapu KB, Nickell JR, Horton DB, Denehy ED, Vartak A, et al. The novel pyr-rolidine nor-lobelane analog UKCP-110 [cis-2,5-di-(2-phenethyl)-pyrrolidine hydro-chloride] inhibits VMAT2 function, methamphetamine-evoked dopamine release, andmethamphetamine self-administration in rats. J Pharmacol Exp Ther 2010];335:841–51.
GeldenhuysWJ, Bezuidenhout LM, Dluzen DE. Effects of a novel dopamine uptake inhibitorupon extracellular dopamine from superfused murine striatal tissue. Eur J Pharmacol2009];619:38–43.
Paliege A, Pasumarthy A, Mizel D, Yang T, Schnermann J, Bachmann S. Effect of apocynintreatment on renal expression of COX-2, NOS1, and renin in Wistar-Kyoto and spon-taneously hypertensive rats. Am J Physiol Regul Integr Comp Physiol 2006];290:R694–700.
Hougee S, Hartog A, Sanders A, Graus YM, Hoijer MA, Garssen J, et al. Oral administration ofthe NADPH-oxidase inhibitor apocynin partially restores diminished cartilage proteo-glycan synthesis and reduces inflammation in mice. Eur J Pharmacol 2006];531:264–9.
Cotter MA, Cameron NE. Effect of the NAD(P)H oxidase inhibitor, apocynin, on peripheralnerve perfusion and function in diabetic rats. Life Sci 2003];73:1813–24.
Kanamori T, Tsujikawa K, Ohmae Y, Iwata YT, Inoue H, Kishi T, et al. A study of themetab-olism of methamphetamine and 4-bromo-2,5-dimethoxyphenethylamine (2C-B) inisolated rat hepatocytes. Forensic Sci Int 2005];148:131–7.
Shima N, Kamata HT, Katagi M, Tsuchihashi H. Urinary excretion of the mainmetabolites of methamphetamine, including p-hydroxymethamphetamine-sulfateand p-hydroxymethamphetamine-glucuronide, in humans and rats. Xenobiotica2006];36:259–67.
Wilhelm H, Wilhelm B, Kriegbaum C. Interaction of the indirectly acting topical sympa-thomimetics cocaine and pholedrine. Ger J Ophthalmol 1996];5:168–70.
WangQ, Smith RE, Luchtefeld R, Sun AY, Simonyi A, Luo R, et al. Bioavailability of apocyninthrough its conversion to glycoconjugate but not to diapocynin. Phytomedicine2008];15:496–503.
Kanegae MP, Condino-Neto A, Pedroza LA, de Almeida AC, Rehder J, da Fonseca LM, et al.Diapocynin versus apocynin as pretranscriptional inhibitors of NADPH oxidase andcytokine production by peripheral blood mononuclear cells. Biochem Biophys ResCommun 2010];393:551–4.


















