
Studies on the mechanism of action of somatostatin on renal gluconeogenesis: Evidence for the...
8
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 213, No. 1, January, pp. 169-176, 1982 Studies on the Mechanism of Action of Somatostatin on Renal Gluconeogenesis: Evidence for the Involvement of cx,-Adrenergic Stimuli K. N. DILEEPAN,’ A. M. KHAWAJA, AND S. R. WAGLE Department of Pharmacology, Indiana University School of Medicine, Indianapolis, Indiana 46223 Received June 15,1981, and in revised form August 11, 1981 The details of the mode of action of somatostatin on renal gluconeogenesis was inves- tigated in cortical slices prepared from rats fed ad Zibitum Somatostatin-stimulated glucose synthesis from lactate and pyruvate and the increase in incorporation of [%]bicarbonate and [2-‘*C]pyruvate into glucose were inhibited by the (I~ antagonist, prazosin, but not by the LYE antagonist, yohimbine. The inhibition of somatostatin-induced gluconeogenesis by prazosin was dose dependent and was not reversed by excess so- matostatin. Treatment of the animals with reserpine prior to the experiments did not abolish the stimulatory effect of somatostatin indicating that catecholamine release may not mediate the enhanced gluconeogenic activity. The enhancement in renal gluconeo- genesis exerted by somatostatin was found to be blocked by insulin. Tissue depletion and the omission of calcium from the incubation system abolished somatostatin-induced glu- cose synthesis. It is proposed that the stimulatory effect by somatostatin may be via ai receptors or those which functionally resemble (pi receptors and that an increased influx of calcium may be the causative factor in carrying out the stimulus. In recent years rapid progress has been made in the isolation and characterization of various bioactive peptides from the gas- trointestinal tract and brain. These bioac- tive peptides are now known to share, in some instances, common amino acid se- quences and the ability to interact with receptors in many tissues. Although the physiological processes subserved by each of these are not clear, structure-activity and phylogenetic studies suggest that a variety of metabolic functions may be reg- ulated selectively by these peptides. So- matostatin, the hypothalamic release in- hibiting factor, has been implicated in the regulation of the release of growth hor- mone (1, 2), insulin (3, 4), and glucagon (5), and it abolishes glucagon-induced changes in glycogenolysis, gluconeogene- sis, and cyclic AMP production in isolated liver preparations (6-8). However, so- matostatin by itself does not cause met- abolic changes in the liver. In earlier 1 Present address: Enzymology Research Labora- tory, Veterans Administration Center, Kansas City, Missouri 64128. studies, somatostatin stimulated gluco- neogenesis in rat renal cortical slices (9). This effect was blocked by the cy-adren- ergic antagonist, phentolamine, but not by the fl-adrenergic antagonist, propranolol (10). Thus, the stimulatory effect of so- matostatin on gluconeogenesis is via ff- adrenergic stimuli. The present study was designed to differentiate the subtype of the a-adrenergic response (11-13) and to elucidate the details of the mechanism of action of somatostatin on gluconeogenesis in rat renal cortex. MATERIALS AND METHODS Male Sprague-Dawley rats (200-250 g) which were fed ad lib&urn were used in this study. The animals were killed by stunning and decapitation. Kidney pairs were quickly removed, rinsed in normal saline, and blotted dry. Cortical slices were prepared with a Stadie-Riggs tissue slicer. Gluconeogenic activity was determined by measuring the net glucose pro- duction from lactate unless otherwise specified. Re- actions for gluconeogenesis were carried out in 50- ml Erlenmeyer flasks containing 4 ml Krebs-Ringer bicarbonate buffer (pH 7.4,1.2 mM calcium) (14), one slice of the renal cortex (equivalent to l-2 mg dry wt) and 10 mM substrate. Reaction mixtures were 169 0003-9861/82/010169-08$02.00/O Copyright 0 1982 by Academic Press, Inc. All rights of reproduction in any form reserved.
-
Upload
kn-dileepan -
Category
Documents
-
view
212 -
download
0
Transcript of Studies on the mechanism of action of somatostatin on renal gluconeogenesis: Evidence for the...

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 213, No. 1, January, pp. 169-176, 1982
Studies on the Mechanism of Action of Somatostatin on Renal Gluconeogenesis: Evidence for the Involvement of cx,-Adrenergic Stimuli
K. N. DILEEPAN,’ A. M. KHAWAJA, AND S. R. WAGLE
Department of Pharmacology, Indiana University School of Medicine, Indianapolis, Indiana 46223
Received June 15,1981, and in revised form August 11, 1981
The details of the mode of action of somatostatin on renal gluconeogenesis was inves- tigated in cortical slices prepared from rats fed ad Zibitum Somatostatin-stimulated glucose synthesis from lactate and pyruvate and the increase in incorporation of [%]bicarbonate and [2-‘*C]pyruvate into glucose were inhibited by the (I~ antagonist, prazosin, but not by the LYE antagonist, yohimbine. The inhibition of somatostatin-induced gluconeogenesis by prazosin was dose dependent and was not reversed by excess so- matostatin. Treatment of the animals with reserpine prior to the experiments did not abolish the stimulatory effect of somatostatin indicating that catecholamine release may not mediate the enhanced gluconeogenic activity. The enhancement in renal gluconeo- genesis exerted by somatostatin was found to be blocked by insulin. Tissue depletion and the omission of calcium from the incubation system abolished somatostatin-induced glu- cose synthesis. It is proposed that the stimulatory effect by somatostatin may be via ai receptors or those which functionally resemble (pi receptors and that an increased influx of calcium may be the causative factor in carrying out the stimulus.
In recent years rapid progress has been made in the isolation and characterization of various bioactive peptides from the gas- trointestinal tract and brain. These bioac- tive peptides are now known to share, in some instances, common amino acid se- quences and the ability to interact with receptors in many tissues. Although the physiological processes subserved by each of these are not clear, structure-activity and phylogenetic studies suggest that a variety of metabolic functions may be reg- ulated selectively by these peptides. So- matostatin, the hypothalamic release in- hibiting factor, has been implicated in the regulation of the release of growth hor- mone (1, 2), insulin (3, 4), and glucagon (5), and it abolishes glucagon-induced changes in glycogenolysis, gluconeogene- sis, and cyclic AMP production in isolated liver preparations (6-8). However, so- matostatin by itself does not cause met- abolic changes in the liver. In earlier
1 Present address: Enzymology Research Labora- tory, Veterans Administration Center, Kansas City, Missouri 64128.
studies, somatostatin stimulated gluco- neogenesis in rat renal cortical slices (9). This effect was blocked by the cy-adren- ergic antagonist, phentolamine, but not by the fl-adrenergic antagonist, propranolol (10). Thus, the stimulatory effect of so- matostatin on gluconeogenesis is via ff- adrenergic stimuli. The present study was designed to differentiate the subtype of the a-adrenergic response (11-13) and to elucidate the details of the mechanism of action of somatostatin on gluconeogenesis in rat renal cortex.
MATERIALS AND METHODS
Male Sprague-Dawley rats (200-250 g) which were fed ad lib&urn were used in this study. The animals were killed by stunning and decapitation. Kidney pairs were quickly removed, rinsed in normal saline, and blotted dry. Cortical slices were prepared with a Stadie-Riggs tissue slicer. Gluconeogenic activity was determined by measuring the net glucose pro- duction from lactate unless otherwise specified. Re- actions for gluconeogenesis were carried out in 50- ml Erlenmeyer flasks containing 4 ml Krebs-Ringer bicarbonate buffer (pH 7.4,1.2 mM calcium) (14), one slice of the renal cortex (equivalent to l-2 mg dry wt) and 10 mM substrate. Reaction mixtures were
169 0003-9861/82/010169-08$02.00/O Copyright 0 1982 by Academic Press, Inc. All rights of reproduction in any form reserved.
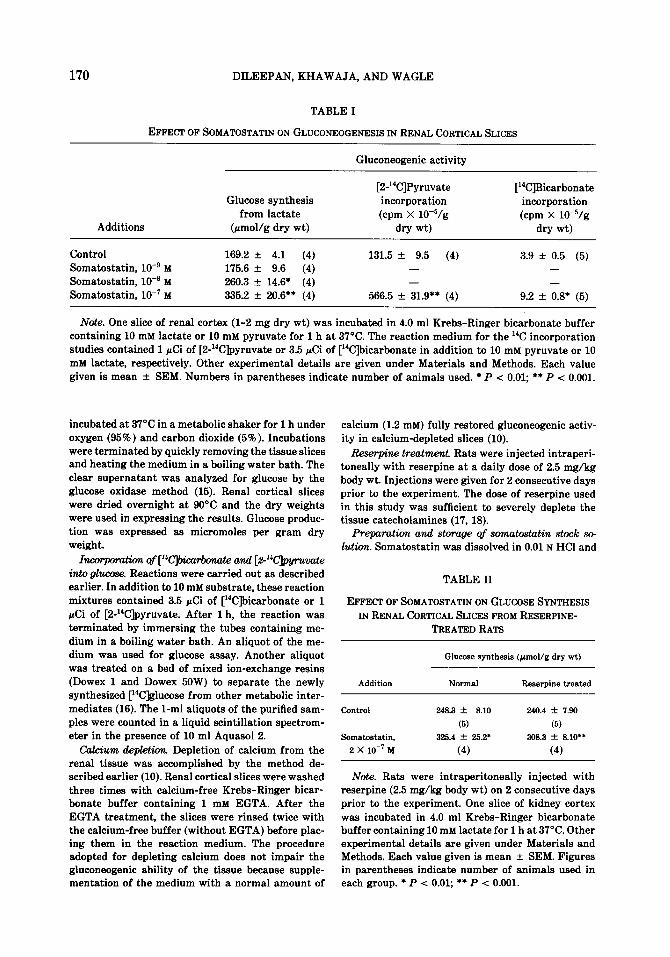
170 DILEEPAN, KHAWAJA, AND WAGLE
TABLE I
Gluconeogenic activity
Additions
Glucose synthesis from lactate
(pmol/g dry wt)
[2-r4C]Pyruvate incorporation (cpm X 1O-5/g
dry wt)
[“ClBicarbonate incorporation (cpm X 10m5/g
dry wt)
Control Somatostatin, lo-’ M Somatostatin, lo-’ M Somatostatin, lo-’ M
169.2 + 4.1 (4) 131.5 f 9.5 (4) 3.9 + 0.5 (5) 175.6 * 9.6 (4) - - 260.3 k 14.6* (4) - - 335.2 + 26.6** (4) 566.5 + 31.9** (4) 9.2 + o.B?3* (5)
Note. One slice of renal cortex (l-2 mg dry wt.) was incubated in 4.0 ml Krebs-Ringer bicarbonate buffer containing 10 mM lactate or 10 mM pyruvate for 1 h at 37°C. The reaction medium for the r4C incorporation studies contained 1 CCi of [2-r4C]pyruvate or 3.5 PCi of [“CJbicarbonate in addition to 10 mM pyruvate or 10 mM lactate, respectively. Other experimental details are given under Materials and Methods. Each value given is mean + SEM. Numbers in parentheses indicate number of animals used. * P < 0.01; **P < 0.001.
incubated at 37°C in a metabolic shaker for 1 h under calcium (1.2 mM) fully restored gluconeogenic activ- oxygen (95%) and carbon dioxide (5%). Incubations ity in calcium-depleted slices (10). were terminated by quickly removing the tissue slices Reserpin~ treatment. Rats were injected intraperi- and heating the medium in a boiling water bath. The toneally with reserpine at a daily dose of 2.5 mg/kg clear supernatant was analyzed for glucose by the body wt. Injections were given for 2 consecutive days glucose oxidase method (15). Rena1 cortical slices prior to the experiment. The dose of reserpine used were dried overnight at 90°C and the dry weights in this study was sufficient to severely deplete the were used in expressing the results. Glucose produc- tissue catecholamines (17, 18). tion was expressed as micromoles per gram dry Preparation and storage of somatostatin stock SC+ weight. lution. Somatostatin was dissolved in 0.01 N HCl and
hwrpomttion of [‘4cjbica*te and [2-“(&fruv& into glucose Reactions were carried out as described earlier. In addition to 10 mM substrate, these reaction mixtures contained 3.5 pCi of [“Clbicarbonate or 1 pCi of [2-14Clpyruvate. After 1 h, the reaction was terminated by immersing the tubes containing me- dium in a boiling water bath. An aliquot of the me- dium was used for glucose assay. Another aliquot was treated on a bed of mixed ion-exchange resins (Dowex 1 and Dowex 50W) to separate the newly synthesized [14CJglucose from other metabolic inter- mediates (16). The l-ml aliquots of the purified sam- ples were counted in a liquid scintillation spectrom- eter in the presence of 10 ml Aquasol 2.
TABLE II
EFFE~TOFSOMATOSTATINONGLUCOSESYNTHESIS INRENALCORTICALSLICESFROMRESERPINE-
TREATEDRATS
Glucose synthesis ($mol/g dry wt)
Addition Reserpine treated
Calcium depletion. Depletion of calcium from the renal tissue was accomplished by the method de- scribed earlier (10). Rena1 cortical slices were washed three times with calcium-free Krebs-Ringer bicar- bonate buffer containing 1 mM EGTA. After the EGTA treatment, the slices were rinsed twice with the calcium-free buffer (without EGTA) before plac- ing them in the reaction medium. The procedure adopted for depleting calcium does not impair the gluconeogenic ability of the tissue because supple- mentation of the medium with a normal amount of
Control 248.3 + 8.10 240.4 f 7.90
(5) (5) Somatostatin, 325.4 + 25.2. 308.3 f s.10**
2 X 106 M (4) (4)
Note. Rats were intraperitoneally injected with reserpine (2.5 mg/kg body wt) on 2 consecutive days prior to the experiment. One slice of kidney cortex was incubated in 4.0 ml Krebs-Ringer bicarbonate buffer containing 10 m?d lactate for 1 h at 37°C. Other experimental details are given under Materials and Methods. Each value given is mean + SEM. Figures in parentheses indicate number of animals used in each group. * P < 0.01; ** P -z 0.001.

LYE STIMULI IN SOMATOSTATIN-STIM~ATED RENAL GLUCON~OGENESIS 171
TABLE III
EFFECT OF INSULIN ON SOMATOSTATIN- AND GLUCAGON-INDUCED GLUCOSE SYNTHESIS
IN RENAL CORTICAL SLICES
Glucose synthesis (pmol/g dry wt)
Additions +Insulin
-Insulin (150 ru
Control 191.0 +- 11.1 143.0 + 19.6 Somatostatin, lo-* M 214.4 + 6.9 182.8 + 9.0 Somatostatin, 10-r M 242.6 + 8.1 181.1 * 7.5 ~matostatin, lo-” M 297.8 + 5.7 187.3 i 8.4 Glucagon, 10-l’ M 224.3 + 14.5 194.0 i 3.0 Glucagon, lo-’ M 267.3 * 39.0 208.0 -e 18.9 Glucagon, low7 M 288.3 z!z 19.8 213.0 f 9.1
Note. One slice of kidney cortex was incubated in 4.0 ml Krebs-Ringer bicarbonate buffer in the pres- ence of lactate (10 mM). The incubation was carried out for 1 h at 37°C. Other details of the experiment are given under Materials and Methods. Each value given is mean f SEM of the results from five ani- mals.
stored in plastic tubes in small quantities at -70°C. As such, potency was retained for several months. Our experience indicates that the pH of the solvent and storage conditions are critical factors for re- taining the potency of somatostatin. Substantial loss of potency of somatostat~n was often noticed if proper precautions were not taken in preparing the stock solution. Required dilutions were made in Kreb’s-Ringer bicarbonate buffer just before each experiment.
Materials. Somatostatin was purchased from Boeh- ringer-Mannheim Biochemicals, Indianapolis, Indi- ana. Insulin and glucagon were gifts from Dr. W. Shaw of Eli Lilly and Company, Indiana~lis, Indi- ana. [2-‘4C]Pyruvic acid (sodium salt) and [‘*@odium bicarbonate were supplied by ICN Chemical and Ra- dioisotope Division, Irvine, California. Prazosin (a product of Pfizer), was a gift from Dr. Larry Jones, Department of Pharmacology, Indiana University School of Medicine, Indianapolis, Indiana. Reserpine was obtained from the CIBA Pharmaceutical Com- pany, Summit, New Jersey. All other chemicals used in this study were purchased from Sigma Chemicals Company, St. Louis, Missouri. Animals used in the study were supplied by Harlan Sprague-Dawley, In- dianapolis, Indiana.
RESULTS
Somatostatin (1O-8-1O-7 M) increased glucose synthesis form lactate and the in-
corporation of radioactivity from [Z- l*C]pyruvate and [14C]bicarbonate into glu- cose (Table 1).
However, depletion of catecholamine stores by prior treatment of the animals with reserpine did not abolish the stimu- latory effect of somatostatin (Table II).
The addition of insulin (150 ~IJi4.0 ml reaction mixture) completely inhibited so- matostatin-induced enhancement in glu- cose synthesis (Table III). The enhance- ment in glucose production caused by glucagon was also abolished by insulin, although at higher concentrations of glu- cagon insulin failed to revert the full stim- ulatory effect.
The involvement of calcium as a possible second messenger in somatostatin action in the renal tissue was investigated by measuring glucose synthesis from lactate in calcium-depleted slices of renal cortex. The depletion of tissue calcium by EGTA treatment and the ommission of calcium in the reaction mixture considerably re- duced gluconeogenic activity in the slices (by approximately 75%) (Table IV). In the calcium-free system, somatostatin failed
TABLE IV
SOMATOSTATIN EFFECT ON GLUCONEOGENESIS IN CALCIUM-DEPLETED RENAL CORTICAL SLICES
Glucose synthesis (pmol/g dry wt)
Additions Normal Calcium depleted
Control
Somatostatin, 2 X 10-l M
260.0 It 10.2 72.2 r 5.9 (10) (5)
455.2 rfi 13.0* 80.3 f 11.1 (10) (5)
Note. Renal cortical slices were depleted of calcium by washing with calcium-free Krebs-Ringer bicar- bonate buffer containing 1 m&i EGTA as mentioned under Materials and Methods. One slice of calcium- depleted kidney cortex (l-2 mg dry wt) was incubated in 4.0 ml calcium-free Krebs-Ringer bicarbonate buffer in the presence of lactate (10 mM) for 1 h at 37% Each value given is mean zt SEM. Figures in parentheses indicate number of animals used. *P < 0.001.

172 DILEEPAN, KflAWAJA, AND WAGLE
NO ANTAGONIST
FIG. 1. Effects of prazosin and yohimbine on somatostatin-stimulated glucose synthesis from (A) 10 mM lactate (B) 10 mM pyruvate. Incubations were carried out in 4.0 ml Krebs-Ringer bicarbonate buffer for 1 h at 37°C as mentioned under Materials and Methods. Somatostatin con- centration was 2 X lo-’ M. Each value given is mean k SEM of the results obtained from four to five animals.
to cause any stimulation of glucose syn- thesis. The addition of normal concentra- tion of calcium (1.2 mM) to the calcium- free system restored normal glucose syn- thesis as well as the stimulatory effect of somatostatin (10).
Since a-adrenergic stimuli can be sub- classified into cq and a2 types, we at- tempted to determine which of the two subtypes is involved in mediating the so- matostatin effect. Somatostatin-mediated increases in glucose synthesis from lactate (Panel A) and pyruvate (Panel B) were blocked by the q antagonist, prazosin, whereas the enhancement in glucose syn- thesis was not blocked by the (~2 antago- nist, yohimbine (Fig. 1). Prazosin com- pletely abolished the somatostatin-induct
increase in the incorporation of [2- 14C]pyruvate (Panel A) and [‘4C]bicar- bonate (Panel B) into glucose (Fig. 2). The same concentration of yohimbine, on the other hand, did not inhibit the somato- statin-stimulated incorporation of 14C pre- cursors. Prazosin or yohimbine alone (lo- M) did not cause any inhibition in glucose production.
The blockade of somatostatin-stimu- lated glucose synthesis by prazosin was dose dependent (Fig. 3). Whereas lo-‘M prazosin inhibited 98% of the stimulated glucose synthesis, yohimbine was ineffee- tive as an inhibitor of somatostatin-in- duced glucose production, even at lop4 M. In the presence of lo-’ M prazosin, increas- ing the ~oneentration of somatostatin to

ai STIMULI IN SOMATOSTATIN-STIMULATED RENAL GLUCONEOGENESIS 173
NO ANTAGONIST
T T
FIG. 2. Effects of prazosin and yohimhine on somatstatin-stimulated increase in ‘“C incorporation into glucose from (A) [2-‘%]pyruvate and (B) [‘%]bicarbonate. Reaction mixture contained 4.0 ml Krebs-Ringer bicarbonate, one slice of kidney cortex, 10 mM pyruvate, and 1 &i of [2-‘%]pyruvate (Panel A) or 10 mM lactate and 3.5 pCi of [Wlbicarbonate (Panel B). Somatostatin concentration used in this experiment was 2 X lo-’ M. Incubations were carried out for 1 h at 37°C. After ter- mination of reaction the medium was processed for radioactivity determination as described under Materials and Methods. Each value given is mean + SEM of four to five observations.
10e6 M did not reverse the inhibitory effect of the q antagonist (Table V).
DISCUSSION
A variety of hormones and other bioac- tive peptides exert their effects through the release of catecholamines. Guder and Rupprecht (19) have demonstrated that norepinephrine stimulates gluconeogene- sis in renal tubules from fasted rats and that the effect is mediated by a-adrenergic receptors. Further evidence for the stim- ulation of renal gluconeogenesis by an CY- adrenergic mechanism has been provided by the work of McDonald and Saggerson
(20). In the present study, prior treatment of the animals with reserpine did not alter the gluconeogenic response to somato- statin in renal cortical slices (Table II). Therefore, it is likely that the somato- statin effect is, indeed, an independent phenomenon, and noradrenaline release may not be a causative factor for the en- hanced gluconeogenesis.
Data presented in this report (Table III) demonstrate that insulin is capable of re- versing somatostatin- and glucagon-in- duced enhancement of glucose production. Blackmore et al. (21) have recently dem- onstrated that insulin inhibits a-adren- ergic responses to phenylephrine and,

174
170
160
=i 150
s
iz fj 140
s
El F 130
Y
8 tE 120
ii
8
2 110
100
0
I /I I I I I I I 1
0 ” 10-J 10-g 10-E 10-t 10-e 10-S lo-4
ANTAGONIST CONCENTRATION fMi
FIG. 3. Dose dependence of prazosin and yohimbine on the inhibition of somatostatin-stimulated renal gluconeogenesis. The reaction contained one slice of renal cortex (1-2 mg dry wt), 10 mM lactate, 2 X lo-’ M somatostatin, and the specified concentrations of prazosin or yohimbine. Incu- bations were carried out for 1 h at 37°C. Each point represents mean of four observations.
thereby, blocks phenylephrine-induced glucose release in hepatocytes. Because the action of somatostatin, in this regard, is similar to that of other Lu-adrenergic agonists, it may be inferred that the in- hibitory effect of insulin on the somato- statin response may be exerted on an ad- renergic receptor.
a-Adrenergie receptors have been sub- divided into LY~ and LYE subclasses based on the type of function and the second mes- senger involved. This subclassification is achieved based on studies using selective agonists and antagonists (11-13). Prazosin is an antagonist selective for the cq recep-
tors whereas yohimbine is selective for the az receptors. The finding that somato- statin-induced gluconeogenesis in renal cortex was blocked by prazosin but was not blocked by yohimbine suggests that q receptors may mediate the effect of so- matostatin. The failure to reverse the in- hibition of prazosin by a lo-fold higher concentration of somatostatin suggests that the inhibitor may have been tightly bound to its specific receptor (22).
In 1967, Robison et al. (23) suggested that cr-adrenergic stimuli may influence cellular function by lowering the intra- cellular cyclic AMP concentration. In re-

a1 STIMULI IN SOMATOSTATIN-STIMULATED RENAL GLUCONEOGENESIS 175
TABLE V
EFFECT OF SOMATOSTATIN CONCENTRATION ON PRAZOSIN INHIBITION
American Cancer Society (In 46T) to K.N.D. The au- thors wish to thank Dr. Lynn R. Willis for critical reading of this manuscript.
REFERENCES
Additions
Glucose synthesis
(pmol/g dry wt)
Control Somatostatin, lo-? M Somatostatin, lo-’ M
+ prazosin lo-? M Somatostatin, lo+ M Somatostatin, 10m6 M
+ prazosin 10m7 M
139.4 -+ 9.5 270.3 f 28.7*
154.5 f 14.3 313.0 + 19.4**
142.5 + 4.6
1. VALE, W., BRAZEAU, P., RIVIER, C., BROWN, M., Boss, B., RIVIER, J., BURGUS, R., LING, N., AND GUILAEMIN, R. (1975) Ret Prog. Harm Res 31, 365-397.
2. SCHALLY, A. V., DUPONT, A., ARIMURA, A., RED- DING, T. W., NISHI, N., LINTHICIJM, G. L., AND SCHLESINGER, D. H. (1976) Biochemistry 15, 509-515.
Note. One slice of kidney cortex was incubated in 4.0 ml Krebs-Ringer bicarbonate buffer in the pres- ence of lactate (10 mM) and required amounts of so- matostatin and prazosin. Incubations were carried out for 1 h at 37°C. Each value given in mean + SEM of four observations. * P < 0.01; ** P < 0.001.
3. CURRY, D. L., BENNEXT, L. L., AND LI, C. H. (1976) Biochem Biophys. Res. Commun 58,885-892.
4. OLIVER, J. R., AND WAGLE, S. R. (1975) Biochem Biophys Res. Commun 62.772-777.
5. KOERKER, D. J., RUCH, W., CHIDECKEL, E., PALMER, J., GOODNER, C. J., ENSINCK, J., AND GALE, C. C. (1974) science 184.482-484.
6. OLIVER, J. R., LONG, K., WAGLE, S. R., AND ALLEN, D. 0. (1976) Proc Sot Exp. Biol Med 153,367- 370.
cent years evidence has been furnished by many laboratories demonstrating the in- volvement of calcium in the cu-adrenergic response (24-26). In the case of a-adren- ergic activation of hepatic gluconeogene- sis, Tolbert and Fain (27) have demon- strated an absolute requirement for calcium. The hypothesis that cxl effects are due to elevation of cytosolic calcium levels and a2 effects are due to adenylate cyclase inhibition has been suggested by Wikberg (28). Sabol and Nirenberg (29) also have speculated that 1y2 effects might be due to inhibition of adenylate cyclase. The dif- ferentiation of CY~ and o2 receptors is, therefore, believed to be functional rather than anatomic. Thus, the blockade of so- matostatin-induced renal gluconeogenesis by an cxl antagonist, and the failure by somatostatin to stimulate glucose synthe- sis in the absence of calcium, suggest that the effect occurs via CX~ adrenergic recep- tors, or those which functionally resemble ffl receptors.
7. SACKS, H., WALIGORA, K., MATHEWS, J., AND PIMSTONE, B. Endocrindogy (1977) 101. 1751- 1759.
8. IGNARRO, L. J.,AND GROSS, R. A. (1978) Fed Proc 37, 339.
9. LUPIANEZ, J. A., DILEEPAN, K. N., AND WAGLE, S. R. (1979) Bioch,em. Biophys Res. Cwrnmun 89, 735-742.
lo. DILEEPAN, K. N., LUPIANEZ, J. A., AND WAGLE, S. R. (1980) Life Sci 26.20672072.
11. LANGER, S. Z. (1977) Brit J. Phurmucd 60,481- 497.
12. WESTFW, T. C. (1977) Physiol Rev. 57,659-728.
13. FAIN, J. N., AND GARCIA-SAINZ, J. A. (1980) Life Sci 26, 1183-1194.
14. DELUCA, H. F., AND COHEN, P. 0. (1964) In Man- nometric Techniques (Umbreit, W. W., Burris, R. H., and Stauffer, J. F., eds.), p. 131, Burgess Publishing Company, Minneapolis.
15. HUGGET, A. S. G., AND NIXON, D. A. (1957) Luti 2,368.
ACKNOWLEDGMENT
16. CORREDOR, C., BRANDEL, K., AND BRESSLER, R. (1967) Proc Nat. Ad Sci USA 58,22X)-2306.
17. CARLSSON, A. (1966) Pharmud Rev. 18,541~549. 18. RAMACHANDRAN, C. K., DILEEPAN, K. N., SINGH,
V. N., AND VENKITASUBRAMANIAN, T. A. (1974) Environ PhysioL Biochem 4, 89-96.
19. GUDER, W. G., AND RUPPRECHT, A. (1975) Eur. J. B&hem 52,283-290.
This investigation was supported by a grant from 20. MCDONALD, D. W. R., AND SAGGERSON, E. D. Biomedical Research Committee of Indiana Univer- (1977) B&hem J. 168.33-42. sity to S.R.W. and an Institutional grant from the 21. BLACKMORE, P. F., ASSIMACOPOULOS-JEANET, F.,

176 DILEEPAN, KHAWAJA, AND WAGLE
CHAN, T. M., AND EXTON, J. H. (1979) J. Bid Chem 254.2828-2834.
22. BARNES, P., KARLINER, J., HAMILTON, C., AND DOLLERY, C. (1979) L& Sti 25, 1207-1214.
23. ROBISON, G. A., BUTCHER, R. W., AND SUTHER- LAND, E. W. (1967) Ann N. Y Ad Sci 139. 703-723.
24. KAMINSKY, N. I., BROADUS, A. E., HARDMAN, J. G., JONES, D. J., BALL,, J. H., SUTHERLAND, E. W., AND LIDDLE, G. W. (1970) J. Clin Invest. 49,2387-2395.
25. GOLDBERG, N. D., O’DEA, R. F., AND HADDOX, M. K. (1973) Advan CycL NucL Res. 3,155-223.
26. SELINGER, Z., BATZRI, S., EIMERL, S., AND SCHRAMM, M. (1973) J. BioL Chem. 248, 369- 372.
27. TOLBERT, M. E. M., AND FAIN, J. N. (1974) J. Biol Chem 249, 1162-1166.
28. WIKBERG, J. E. S. (1979) Acta PhysioL Scud Sup~l. 468, l-99.
29. SABOL, S. L., AND NIRENBERG, M. (1979) J. Biol Chem 254, 1913-1920.





![Exploring the Involvement of NLRP3 and IL-1β in …downloads.hindawi.com/journals/mi/2019/2363460.pdfhand osteoarthritis [26] were recruited in the Rheumatol-ogy Unit of Siena Hospital](https://static.fdocument.org/doc/165x107/5f93c90a1258491ec9221a4a/exploring-the-involvement-of-nlrp3-and-il-1-in-hand-osteoarthritis-26-were-recruited.jpg)












