

Src kinase activity is essential for osteoclast function€¦ · · 2004-01-22Department of Cell...
44
Src kinase activity is essential for osteoclast function Tsuyoshi Miyazaki*, Archana Sanjay*, Lynn Neff *, Sakae Tanaka ‡ , William C. Horne* and Roland Baron* *Department of Cell Biology and Orthopaedics, Yale University School of Medicine, New Haven, Connecticut 06510, USA ‡ Department of Orthopaedic Surgery, Faculty of Medicine, The University of Tokyo, 7- 3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan --------------------------------------------------------------- ---------------------------------------- T. M. and A. S. equally contributed to this work. Corresponding author: Roland Baron Department of Cell Biology and Orthopaedics, Yale University School of Medicine, P.O Box 208044 New Haven, Connecticut 06510, USA Tel: 203-785-5986 Fax: 203-785-2744 E-mail: [email protected] Running title: Osteoclast function requires Src kinase activity Key words: Pyk2, Cbl, phosphatidylinositol 3-kinase, bone resorption, apoptosis, Csk 1 JBC Papers in Press. Published on January 22, 2004 as Manuscript M311032200 Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc. by guest on June 13, 2018 http://www.jbc.org/ Downloaded from
Transcript of Src kinase activity is essential for osteoclast function€¦ · · 2004-01-22Department of Cell...

Src kinase activity is essential for osteoclast function
Tsuyoshi Miyazaki*, Archana Sanjay*, Lynn Neff *, Sakae Tanaka ‡, William C. Horne*
and Roland Baron*
*Department of Cell Biology and Orthopaedics, Yale University School of Medicine,
New Haven, Connecticut 06510, USA
‡Department of Orthopaedic Surgery, Faculty of Medicine, The University of Tokyo, 7-
3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan
---------------------------------------------------------------
----------------------------------------
T. M. and A. S. equally contributed to this work.
Corresponding author: Roland Baron
Department of Cell Biology and Orthopaedics, Yale University School of Medicine,
P.O Box 208044 New Haven, Connecticut 06510, USA
Tel: 203-785-5986 Fax: 203-785-2744 E-mail: [email protected]
Running title: Osteoclast function requires Src kinase activity
Key words: Pyk2, Cbl, phosphatidylinositol 3-kinase, bone resorption, apoptosis, Csk
1
JBC Papers in Press. Published on January 22, 2004 as Manuscript M311032200
Copyright 2004 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

SUMMARY
Deletion of the c-src gene impairs osteoclast bone resorbing activity, causing
osteopetrosis. Although it has been concluded that restoring only Src’s adaptor function
at least partly rescues the cell attachment and skeletal phenotypes, the contribution of
Src’s kinase activity remains controversial. Src forms a complex with Pyk2 and Cbl
following adhesion-induced stimulation of αVβ3 integrin. To demonstrate the
importance of the Pyk2-Src association in osteoclasts and to distinguish the contributions
of Src’s adaptor and kinase activities in cytoskeletal organization and osteoclast function,
we expressed mutants of Src and Pyk2 in osteoclasts using adenovirus vectors.
Eliminating the Src-binding site on Pyk2 (Pyk2Y402F) markedly inhibited bone
resorption by osteoclast-like cells (OCLs), while kinase-dead Pyk2 had little effect.
Kinase-dead Src, unlike kinase-dead Pyk2, markedly inhibited the bone-resorbing
activity of wild type osteoclasts and failed to significantly restore bone-resorbing activity
to Src-/- OCLs. Activation of Src kinase by overexpressing kinase-dead Csk failed to
reverse the inhibitory effect of Pyk2Y402F, suggesting that osteoclastic bone resorption
requires both c-Src kinase activity and the targeting of Src kinase by Pyk2. Src-
catalyzed phosphorylation of Cbl on Y731 is reported to induce the activation and
recruitment of phosphatidylinositol 3-kinase to the cell membrane in a signaling pathway
2
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

that is critical for osteoclast function. Expressing the CblY731F mutant in osteoclasts
markedly reduced their bone resorbing activity, suggesting that phosphorylation of
CblY731 and the subsequent recruitment and activation of phosphatidylinositol 3-kinase may
be critical signaling events downstream of Src in osteoclasts.
3
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

INTRODUCTION
Targeted disruption of the src gene in mice induces osteopetrosis, a disorder
characterized by decreased bone resorption, without showing any obvious morphological or
functional abnormalities in other tissues or cells (1). The osteopetrotic phenotype of src-/-
mice is cell-autonomous and results from defects in mature osteoclasts, which normally
express high levels of Src protein (2,3). The mobility of Src- osteoclasts is reduced by 60%,
which is likely to account for at least some of the deficit in bone resorbing activity (4). It
remains uncertain, however, if catalytically active Src is required for normal osteoclast
function. Some investigators have reported the genetic rescue of the src-/- osteopetrosis by
kinase-deficient mutants of Src (5). Together with the findings of Felsenfeld et al. that the
adapter function of Src promotes fibroblasts motility (6), these results suggest that the
predominant role of Src in osteoclasts might be as an adaptor molecule, recruiting other
proteins that are necessary for the modulation of integrin-cytoskeletal links, cell attachment
and migration. On the other hand, there are reports that indicate that Src kinase activity is
required for normal osteoclastic bone resorption (7-9) leaving the question of the
contribution of Src kinase activity unsettled.
The identity of the Src target(s) in the signaling pathways that are required for normal
osteoclast mobility and resorption also remains to be resolved. We previously reported that
Cbl’s basal phosphotyrosine content in osteoclasts is markedly Src-dependent, in contrast to
4
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

several other known Src substrates (10), and that Src forms an adhesion-induced complex
with Cbl and the calcium-dependent tyrosine kinase Pyk2 in osteoclasts (4). Replating Src+
osteoclasts on vitronection receptor induced the phosphorylation of all three proteins,
whereas only Pyk2 was phosphorylated in replated Src-/- osteoclasts, indicating that Src
phosphorylates Cbl in this complex. In a reconstituted model system (HEK 293 cells
transfected with the vitronection and Pyk2), Src kinase activity was required for adhesion-
induced Pyk2-dependent tyrosine phosphorylation of Cbl (4). However, we did not
determine if the absence of Src kinase activity affected OC function in vitro.
We have therefore extended our investigation to examine the requirement for Src
kinase activity in osteoclast function, taking advantage of the ability of adenoviral vectors to
transduce foreign genes into osteoclasts (8,11-13). We found that both Src kinase activity
and the Src SH2-dependent formation of the Pyk2/Src complex are required for bone
resorption. Furthermore, we showed that inhibiting Src leads to a marked reduction in the
phosphorylation of Y731 in Cbl, the third component in the Py2-Src complex (4), and that
mutating Cbl Y731 markedly inhibits bone resorption, possibly identifying Cbl as a key
substrate of Src downstream of the Pyk2-Src complex in osteoclasts.
5
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

EXPERIMENTALPROCEDURES
Antibodies and Chemicals
Minimum essential medium, alpha modification (αMEM), Dulbecos modified
essential medium (DMEM), fetal bovine serum (FBS) and PP2 were purchased from
GIBCO BRL. Collagen was obtained from Nitta Gelatin Co. (Osaka, Japan). Bacterial
collagenase and dispase were purchased from Calbiochem. Mouse monoclonal anti-
Pyk2, anti-MEK1, anti-Cbl and anti-Csk antibodies were obtained from Transduction
Laboratories. Mouse monoclonal anti-Src antibody (Ab-1) was purchased from
Oncogene Research Products. Anti-avian Src antibody was obtained from Upstate
Biotechnology, rabbit polyclonal anti-Src antibody and anti-myc antibody from Santa
Cruz Biotechnology, and anti-Xpress antibody from Invitrogen. An anti-Pyk2
phosphotyrosine residue 402 antibody was obtained from Biosource International. An
anti-Cbl phosphotyrosine 731 antibody was obtained from Cell Signaling Technologies.
Constructs and Gene Transduction
Adenovirus vectors carrying the various forms of Pyk2, Src and Cbl were
constructed as reported (14). The recombinant adenovirus vector carrying constitutively
active MEK1 (S218E and S222E, AxMEKCA), wild type Csk (AxCskWT), or kinase-
dead Csk (K222R, AxCskKD) genes were constructed as previously described (8,12).
6
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Infection of OCLs with adenovirus vectors was carried out following the method
previously described (8).
7
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Cells and Cell Cultures
Mouse primary osteoblastic cells were obtained from 1-day-old mouse calvaria
by enzymatic digestion, and bone marrow cells were from tibiae of 6-week-old Src+/? or
Src-/- mice. Osteoblastic cells (5 x 105 cells/dish) were co-cultured with bone marrow
cells (1 x 107 cells/dish) on 10 cm tissue culture dishes or collagen gel-coated dishes in
the presence of 10 nM 1α, 25-dihydroxy vitamin D3 and 1µM prostaglandin E2. For
biochemical assays, osteoclast-like cells (OCLs) were purified as reported elsewhere (8).
RAW 264.7 cells were cultured in DMEM supplemented with 10% FBS and 1%
penicillin and streptomycin. Prior to experiments, RAW264.7 cells were starved for 18 h
in medium containing 0.1% FBS and treated with 10uM PP2 for 30 min.
Coimmunoprecipitation
Cell cultures were washed twice with ice-cold PBS and lysed in buffer containing
20 mM Hepes-NaOH (pH7.4), 150 mM NaCl, 0.05% NP-40, 10% glycerol, 10 mM
EDTA, 1 mM Na3VO4, 10 µg/ml leupeptin, 10 µg/ml aprotinin, and 1 mM PMSF. After
30 min on ice, cell lysates were cleared by centrifugation at 12,000-x g for 20 min.
Immunoprecipitation was performed by incubating 500 µg of total cell lysate protein with
2 µg of antibody for 2 h on ice, then adding 20 µl of protein-G agarose. After incubation
for 1 h at 4°C with end-over-end mixing, the immune complex was recovered by
8
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

centrifugation and washed four times with buffer containing 20 mM Hepes-NaOH
(pH7.4), 150 mM NaCl, 0.02% IGEPAL CA-630, 10 mM EDTA, 1 mM Na3VO4, 10
µg/ml leupeptin, 10 µg/ml aprotinin, and 1 mM PMSF. The immunoprecipitates were then
subjected to SDS-PAGE and Western blotting.
In Vitro Kinase Assay
In vitro kinase activity was assayed as previously reported (8). Briefly, Src
immunoprecipitates were resuspended in 60 µl of kinase buffer (20 mM HEPES-NaOH,
pH 7.4, and 10 mM MgCl2) with 5 µCi of (γ-32P) ATP in the presence of 1 µg of acid-
treated enolase, then incubated for 20 minutes at 30°C. The reaction was stopped by
adding 20 µl of 4 x sample buffer (250 mM Tris-HCl, pH 6.8, 8 mM EDTA, 12% SDS,
500 mM 2-mercaptoethanol, 15% glycerol, and 0.01% bromophenol blue). The samples
were subjected to SDS-PAGE under reducing condition, followed by autoradiography.
Immunofluorescence Microscopy
For immunofluorescence analysis, cells were plated on sterile FBS-coated glass
coverslips. OCLs were fixed in PBS containing 3.7% formaldehyde for 10 min, and then
permeabilized with ice-cold acetone for 5 min. Coverslips for actin labeling were
incubated in a 1:40 dilution (in PBS) of rhodamine phalloidin stock solution (Molecular
9
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Probes) for 20 min. Cells were sequentially stained with appropriate primary antibody
followed by fluorescein-conjugated secondary antibody as previously reported (4). Cells
were examined using a confocal imaging system (MRC-600; Bio-Rad Laboratories).
Pit Formation Assay
Functionally active OCLs were formed in co-cultures carried out on collagen gel
as described elsewhere (8). An aliquot of the crude OCL preparation was transferred
onto dentine slices and cultured for an additional 12 h. The resorbed area was measured
using an image analysis system linked to a light microscope (Osteometrics).
Survival of OCLs
The survival rate was measured as previously reported (12). OCLs were purified
3 days after the infection and some of the cultures were subjected to tartrate-resistant
acid phosphatase (TRAP) staining. Other cultures were further incubated for 24 h, and
then the number of living OCLs was counted. Cell viability/survival at the different time
points is the number of morphologically intact TRAP-positive multinucleated cells
expressed as a percentage of the number of cells at time 0.
Statistical Analysis
Each series of experiments was repeated at least three times. The results obtained
10
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

from a typical experiment were expressed as the means ± standard deviation (SD).
Significant differences were determined using factorial analysis of variance (ANOVA).
11
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

RESULTS
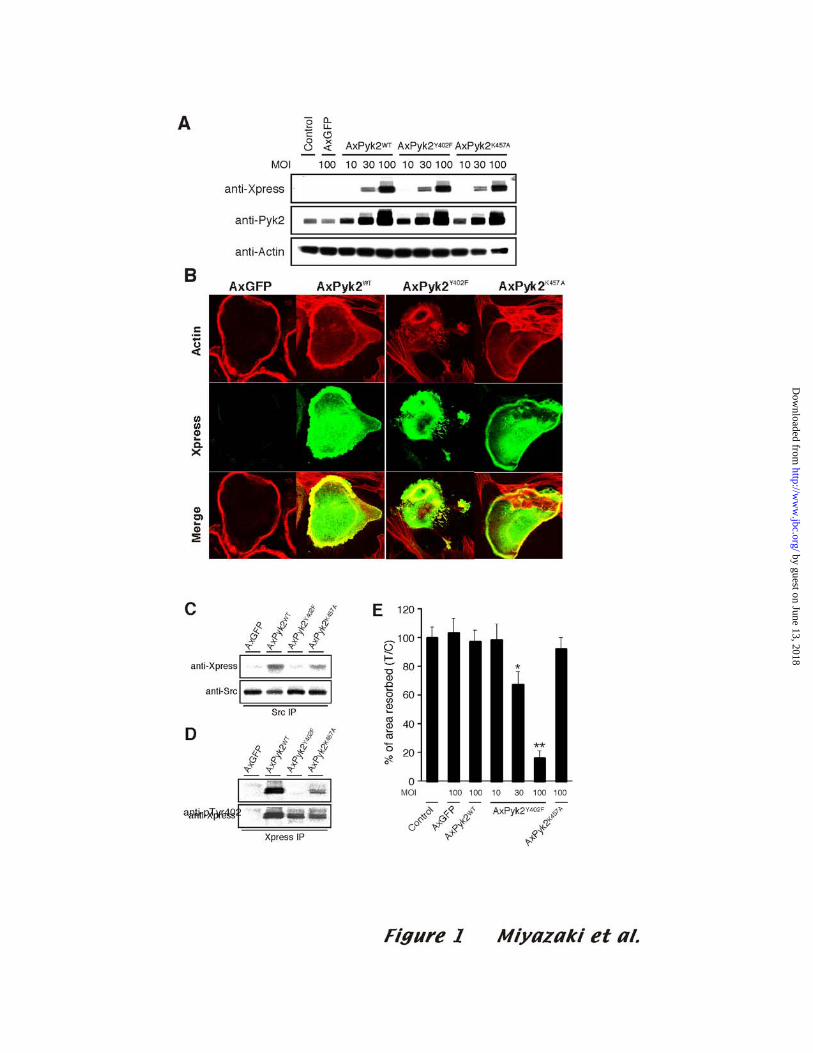
Pyk2Y402F overexpression inhibits the pit-forming activity of osteoclast-like cells
Pyk2 is highly expressed in the podosomes of osteoclasts (4,15) and is
autophosphorylated in response to integrin engagement (4). To investigate the roles of
Pyk2 kinase activity and the autophosphorylation of Pyk2Y402 in osteoclast function, we
constructed adenoviruses containing Xpress-tagged wild type Pyk2 (AxPyk2WT),
autophosphorylation site-mutated Pyk2 (AxPyk2Y402F), or kinase-dead Pyk2
(AxPyk2K457A). We used a replication-deficient adenovirus vector that contains a reporter gene
encoding GFP (AxGFP) as a control vector. The protein expression of the purified OCLs
from cocultures infected with the recombinant viruses at different MOI’s was examined
after three days of infection by Western blotting with anti-Xpress antibody or anti-Pyk2
antibody. Equal amount of the three Pyk2 proteins were expressed in infected OCLs in a
MOI-dependent manner (Fig. 1A). Cells infected with AxPyk2WT or AxPyk2K457A
exhibited in a single large actin ring similar to the ring seen in the cells infected with the
control AxGFP. In contrast, Pyk2Y402F-expressing OCLs contained actin patches and
one or more small actin rings (Fig. 1B).
Pyk2Y402 is autophosphorylated following cell attachment and binds to Src’s SH2
domain (4,15,16). Consistent with these reports, endogenous Src bound to Xpress-tagged
12
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Pyk2WT and Pyk2K457A, but not to Pyk2Y402F (Fig. 1C). As expected, no phospho-
Y402 was detected in the cells expressing Pyk2Y402F. In contrast, a low level of
phosphorylation of the Y402 of Pyk2K457A was detected (Fig. 1D), indicating either that
this residue can be phosphorylated by some other kinase in osteoclasts, or that the small
residual kinase activity of mutated Pyk2 is sufficient to catalyze a low level of
phosphorylation.
To investigate the functional consequences of overexpressing Pyk2WT,
Pyk2Y402F, or Pyk2K457A, the bone resorbing activity of OCLs was quantified by measuring
the areas of pits formed on dentine slices. As shown in Fig.1E, the pit-forming activity
of OCLs was strongly inhibited in proportion to Pyk2Y402F expression. In contrast,
overexpression of Pyk2WT or Pyk2K457A had no effect on bone resorption. There was
no significant difference in osteoclast survival among OCLs expressing the different
Pyk2 proteins (data not shown). Thus, these data demonstrate that the interaction of Src
with Pyk2, mediated by the SrcSH2 domain and the Pyk2 pTyr402, is required for the
normal organization of the osteoclast actin cytoskeleton and for bone resorption.
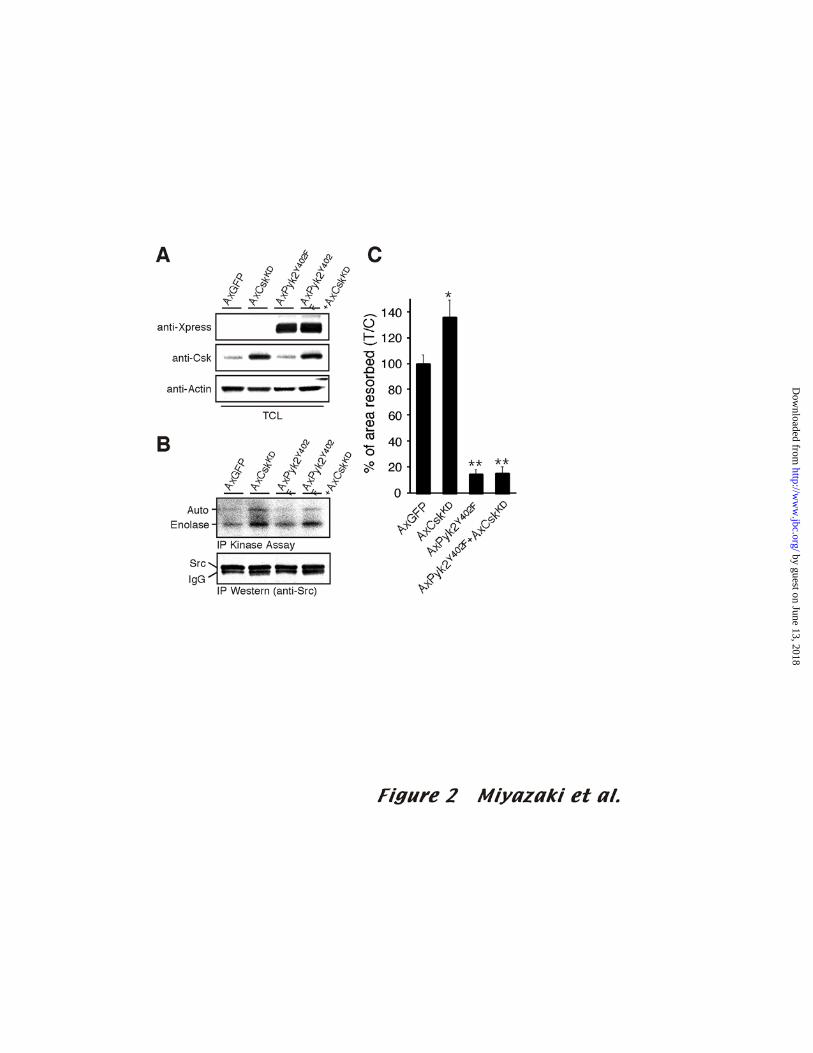
The inhibitory effect of Pyk2Y402Foverexpression on bone resorption is not reversed by
up-regulating Src kinase activity
13
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

The effect of overexpressing Pyk2Y402F suggests that the recruitment of Src to
autophosphorylated Pyk2 at adhesion sites is a key event in bone resorption, but it does
not address the question of whether Src kinase activity is required or if the adaptor
functions of Src’s SH2 and SH3 domains are sufficient. Indeed, Schwartzberg and
colleagues concluded from their in vivo results that the kinase activity of Src may be
dispensable (5). To address this question, we examined the effect of modulating Src
kinase activity independently of the binding of Src to Pyk2. We first examined whether
or not the up-regulation of Src kinase activity was sufficient to rescue the decreased pit-
forming activity of Pyk2Y402F-expressing OCLs. For this purpose, we co-infected
OCLs with AxPyk2Y402F and adenovirus carrying kinase-dead C-terminal Src family
kinase (AxCskKD), which prevents the phosphorylation of the negative regulatory Y527
and increases Src kinase activity in OCLs (8). Co-infected OCLs expressed Pyk2Y402F
and CskKD at levels similar to the cells infected with either Pyk2Y402F or CskKD alone
(Fig. 2A). As shown in Fig. 2B, Src kinase activity in OCLs was up-regulated when
CskKD was overexpressed alone or when it was co-expressed with Pyk2Y402F. The
inhibitory effect of AxPyk2Y402F on osteoclast function was, however, not rescued by
co-infection with AxCskKD (Fig. 2C), indicating that high Src kinase activity alone is
not sufficient to rescue the decreased bone resorbing activity in the Pyk2Y402F-
14
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

expressing OCLs and that the Pyk2-dependent recruitment of Src is necessary for bone
resorption.
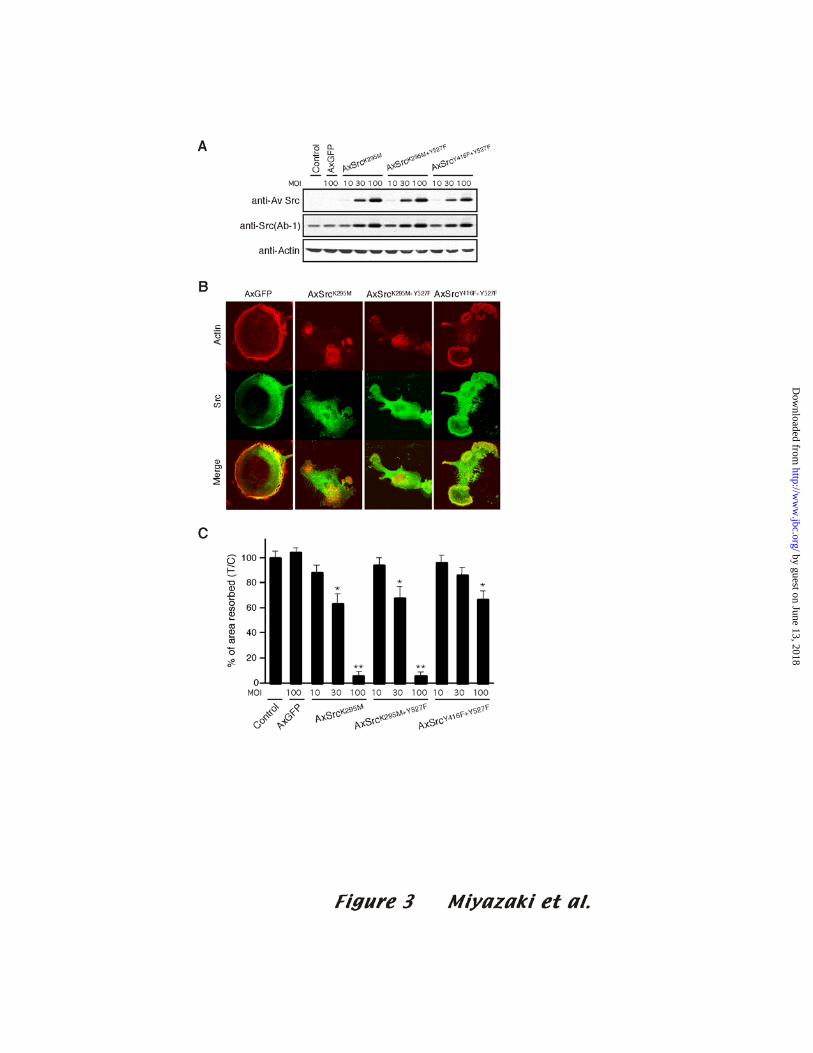
Src kinase activity is required for bone resorption
We then examined the role of Src kinase activity in situations where Pyk2-
dependent recruitment of Src was not altered. For this purpose, we constructed an
adenovirus carrying kinase-dead Src (SrcK295M); kinase-dead Src with open
conformation (SrcK295M+Y527F), or autophosphorylation site-mutated Src with open
conformation (SrcY416F+Y527F). The open conformation allows that the adaptor
domains of Src are available to bind other proteins. As shown in the upper and middle
panels of Fig. 3A, the Src mutant proteins were expressed in infected OCLs in a MOI-
dependent manner. SrcK295M-expressing OCLs do not display the typical rounded
appearance of AxGFP-infected OCLs, and the actin ring was disrupted in >90% of
AxSrcK295M-infected OCLs (Fig. 3B). This morphology of SrcK295M-expressing
OCLs is similar to that of Src-/- OCLs (4). In addition, pit-forming activity was also
inhibited, albeit never completely blocked, in proportion to the expression level of
SrcK295M protein (Fig. 3C).
These results strongly suggest that it is the presence of the kinase activity of Src in
15
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

the osteoclast adhesion structures, and not just the Src protein, that is necessary for actin
ring formation and bone resorption. However, the autophosphorylation of Src promotes
the adoption of the fully open active conformation (17,18), and thus SrcK295M might not
be an effective adaptor. To rule out the possibility that the dominant negative effects of
SrcK295M could be due to a loss of adaptor function, OCLs were infected with
AxSrcK295M+Y527F, which prevents the phosphorylation of the negative regulatory Y527 and
promotes the availability of the SH2 and SH3 domains to bind other proteins. To ensure
that the mutant Src proteins were indeed binding to Pyk2 in OCLs, the Src proteins were
immunoprecipitated from OCLs lysates and the immune complexes were blotted to
demonstrate the presence of Pyk2 (data not shown). The morphology and bone-resorbing
activity of SrcK295M+Y527F-expressing OCLs was similar to that of OCLs infected
with AxSrcK295M (Fig. 3B and C), indicating that it is the absence of kinase activity, not
the possible failure to bind to Pyk2 and/or other proteins in the adhesion complex, that
causes the loss of bone-resorbing activity.
To further confirm the importance of Src kinase activity, we investigated the
functional changes in OCLs infected with AxSrcY416F+Y527F. The Y416F mutation
reduces kinase activity about 50% by preventing the phosphorylation of a tyrosine on the
activation loop of Src (19,20). In spite of its open conformation, SrcY416F+Y527F
16
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

partially inhibited osteoclast function, consistent with its reduced kinase activity.
AxSrcY416F+Y527F-infected OCLs exhibited a few small actin rings (Fig. 3B) and still retained
partial bone-resorbing activity (about 70 % of control) at an MOI of 100 (Fig. 3C).
These results strongly suggest that the localization of catalytically active Src in adhesion
structures is required for actin ring formation and bone resorption.
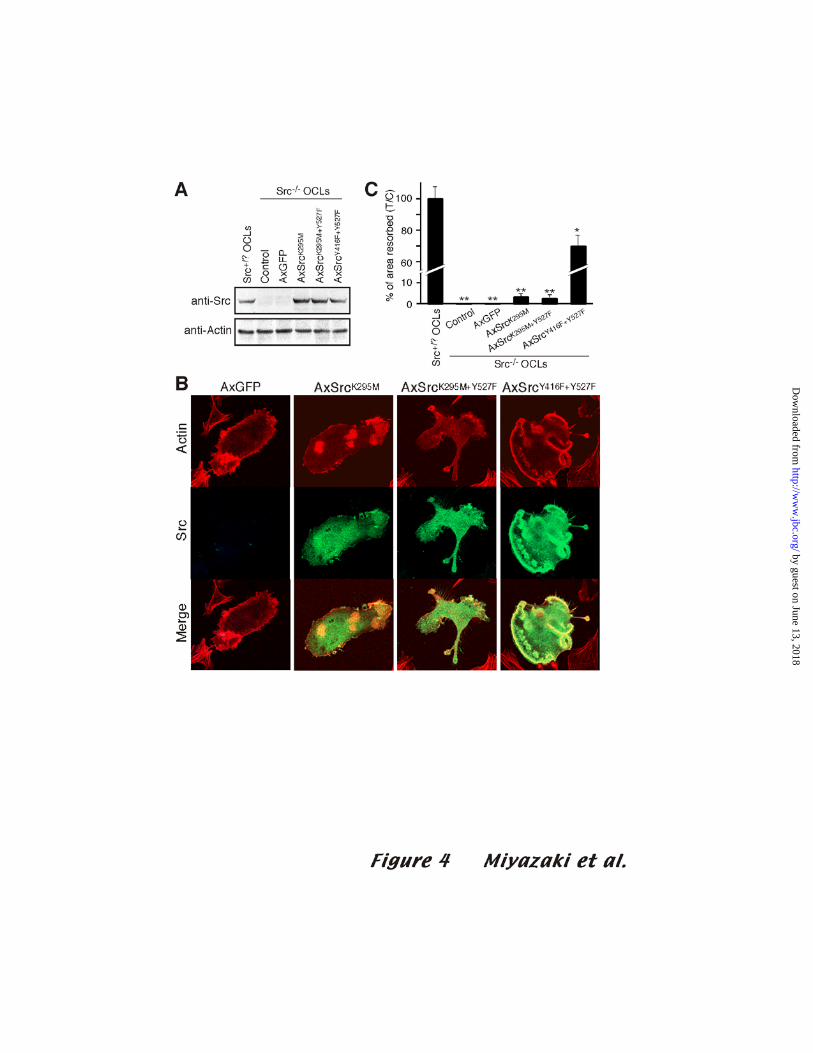
Src kinase activity is required for the efficient rescue of the Src-/- osteoclast phenotype
To further confirm the importance of Src kinase activity for osteoclastic bone
resorption, and to determine the influence of endogenous Src levels in our previous
experiments, we introduced Src mutants into Src-/- osteoclasts. As shown in Fig. 4A,
the mutant Src proteins were expressed in infected Src-/- OCLs. Under these
experimental conditions, Src-/- OCLs were completely unable to form resorption pits in
our assay. Interestingly, SrcK295M and SrcK295M+Y527F, which did not notably affect
the cytoskeletal organization of OCLs, restored, albeit extremely minimally (less than
3%), some bone-resorbing activity in Src-/- osteoclasts (p<0.001). This minimal effect
could be due to the adaptor function of Src, or to the very low residual kinase activity in
these mutants. In contrast, the expression of SrcY416F+Y527F, which retains about 50%
kinase activity, induced the formation of actin rings and very significantly increased
17
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

bone-resorbing activity to approximately 70% of control (Fig. 4B and C). These results
clearly show that Src kinase activity is indeed required to rescue the disruptive effect of
Src deficiency on the cytoskeletal organization and bone-resorbing activity of
osteoclasts.
Identification of Src kinase-dependent signaling events
Having determined that both the Src adaptor function (binding to Pyk2) and Src
kinase activity are required for bone resorption, we sought to identify downstream
elements in Src-dependent signaling mechanism(s). We previously showed that Cbl is
phosphorylated in a Src-dependent manner in osteoclasts and that bone resorption by
OCLs in vitro is inhibited by both Src-and Cbl-specific antisense oligonucleotides
(4,10), suggesting that tyrosine phosphorylation of Cbl by Src is essential for bone
resorption. Of the 22 tyrosines in Cbl, those at positions 700, 731, and 774 are the
principal sites of phosphorylation by Src family kinases (21). Tyr731 occurs within the
sequence YEAM, which has been shown to provide a docking site for the SH2 domain(s)
of the p85 regulatory subunit of phosphatidylinositol kinase (PI3-K) (22-24). Since PI3-
K plays an important role in osteoclast function (25-27), phosphorylation of Cbl Tyr731
by Src and the consequent binding of PI3-K to Cbl could be a critical signaling event.
To confirm that Src catalyzes the phosphorylation of Cbl Tyr731, RAW264.7 cells were
18
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

treated with PP2, a Src family kinase inhibitor. The phosphorylation of Tyr731 was
examined by Western blotting with anti phospho-Tyr731 antibodies. Treatment of the
cells with PP2 significantly reduced the total phosphotyrosine content as well as
phosphorylation of Y731 (Fig. 5A). We next examined how mutating Cbl Y731 affects
the OC function. Osteoclasts were infected with adenovirus carrying Cbl in which Y731
was mutated to phenylalanine (AxCblY731F), with AxCblWT or with Ax GFP. The
protein expression of infected OCLs was examined after three days of infection by
Western blotting (Fig. 5B). Infection with AxCblY731F at an MOI of 100 inhibited pit
formation by 70%, while AxCblWT had no effect (Fig. 5C). Thus, the Src-catalyzed
phosphorylation of Cbl Y731 appears to play an important role in the signaling leading to
normal osteoclastic bone resorption.
Src kinase activity is not involved in osteoclast survival
We then determined whether Src kinase activity was also required for other
functional aspects of the osteoclasts. The activation of Akt via a Src-dependent signaling
pathway has been reported to be important for RANKL-induced osteoclast survival
(9,28). On the other hand, the Ras/ERK signaling pathways also appears to be involved
in osteoclast survival, since inhibiting ERK activity by overexpressing dominant negative
19
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Ras (RasDN) rapidly induces the apoptosis of OCLs, whereas activating ERK by
overexpressing constitutively active MEK1 (MEKCA) markedly reduces spontaneous
apoptosis (12). To examine the role of c-Src kinase in regulating osteoclast survival, we
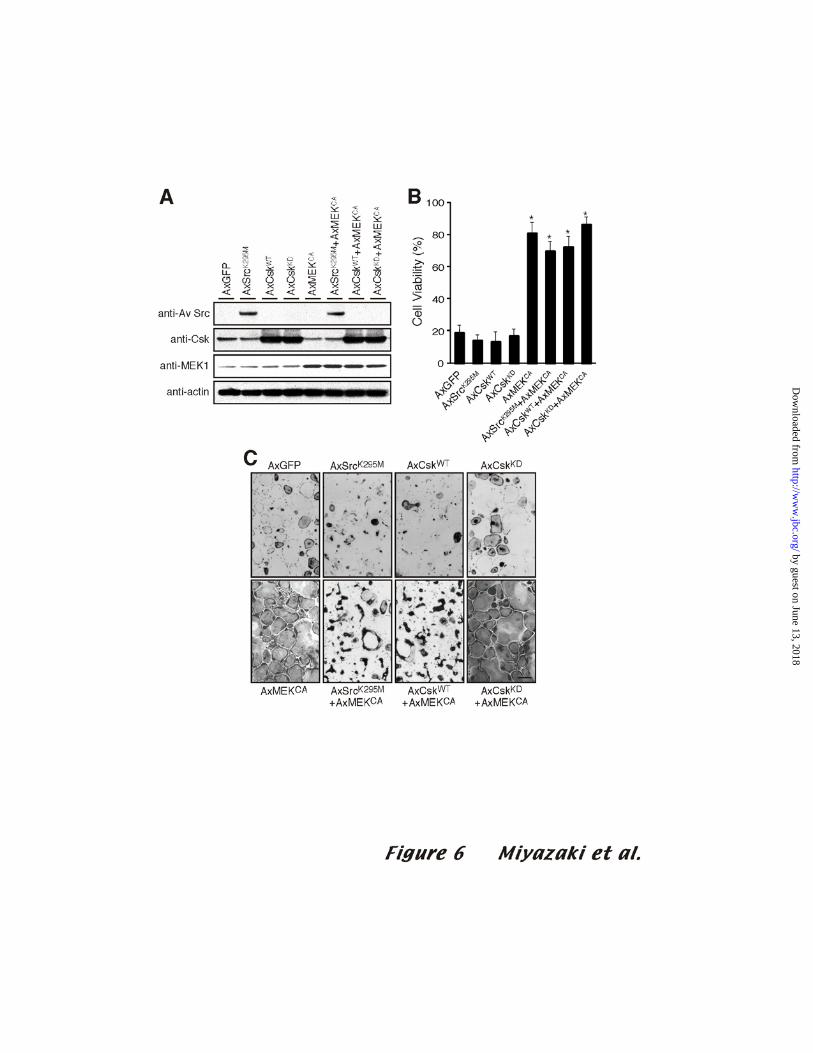
infected OCLs with AxSrcK295M, AxCskWT, or AxCskKD, either alone or together
with AxMEKCA. The OCLs infected with AxMEKCA expressed SrcK295M, CskWT,
or CskKD proteins at similar levels as the cells without AxMEKCA (Fig. 6A). Neither
SrcK295M, CskWT, nor CskKD had any effect on the survival of OCLs, regardless of
whether the OCLs were infected with AxMEKCA (Fig. 6B and C). More than 70% of
OCLs infected with AxSrcK295M or AxCskWT were still alive even 24 hours after
purification when infected by AxMEKCA. These results indicate that c-Src kinase
activity is important for cytoskeletal organization and bone resorption, but not for the
survival of osteoclasts.
20
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

DISCUSSION
We have previously established that activating osteoclast vitronectin receptors by
cross-linking or replating induces the formation of signaling complex that includes Pyk2,
Src and Cbl (4). The primary goal of the present study was to determine if Pyk2-
associated Src kinase activity is required for efficient osteoclast bone resorption or
conversely, if the adaptor function of Src is sufficient to restore bone resorbing activity to
Src-/- osteoclasts, as suggested by other investigators (5). We demonstrate that both Src
kinase activity and the localization of active Src to the complex via an interaction with
Pyk2’s phosphotyrosine 402 are essential for osteoclastic bone resorption, whereas Pyk2
kinase activity is not.
We first examined the importance of the binding of Src to Pyk2. The mutation of
Pyk2 Tyr402 inhibited both the Pyk2-Src interaction and bone resorption. In contrast,
the kinase-dead Pyk2K457A, which was phosphorylated on Tyr402, albeit to a lesser
degree than the wild type Pyk2, bound Src and had little effect on bone resorption (Fig. 1
C and D). The phosphorylation of Tyr402 of kinase-dead Pyk2 is likely catalyzed by
another kinase, although a role for low residual kinase activity cannot be ruled out.
Duong, et al., have reported that preventing the synthesis of Pyk2 protein with Pyk2
antisense oligonucleotides inhibited osteoclast spreading, actin ring formation, and bone
resorption (13), and Lakkakorpi, et al. recently showed that Pyk2 phosphorylation but not
21
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

its kinase activity is required for the interaction with Src, osteoclast spreading and bone
resorption (29), consistent with our results. Thus, the ability of phosphorylated Pyk2 to
serve as a docking site for Src is critically important for the organization of the osteoclast
actin cytoskeleton and attachment sites and for bone resorption. The reputed inhibition of
osteoclast activity by small molecules that bind to the Src SH2 domain (30) may be due
to the prevention of the Pyk2-Src interaction.
While these and our earlier results suggested that recruitment of Src to adhesion
structures is crucial in osteoclasts, as in other cell types (31), we also tested whether
simply increasing Src kinase activity while preventing its binding to Pyk2 would affect
bone resorption. The introduction of CskKD failed to reverse the inhibitory effect of
AxPyk2Y402F on bone resorption (Fig. 2C), further establishing that the formation of a
Pyk2/c-Src complex occurs after integrin binding (4) but also that the formation of this
complex is indispensable for osteoclastic bone resorption.
As noted earlier, the importance of Src kinase activity in bone resorption has been
controversial. Schwartzberg et al., previously reported that osteoclast-specific
expression of the kinase-dead SrcK295M mutant partially restored normal trabecular
bone volume and tooth eruption in the Src-/- mice and suggested that Src’s key function
in osteoclastic bone resorption might be as an adaptor molecule rather than tyrosine
kinase (5). On the other hand, others have reported that inhibiting Src kinase reduces
22
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

bone resorption in vivo and in vitro (7,8) and in the Schwartzberg study, Src kinase
activity was required to restore the F-actin-rich ring of podosome attachment structures
that is believed to be critically important for normal osteoclastic bone resorption. Thus,
the central issue addressed by this study was the requirement for Src kinase activity in
resorbing osteoclast. The data from our study clearly show that at the level of the
individual osteoclast, the kinase activity of Src is indeed required for bone resorption.
Overexpressing kinase-dead SrcK295M in OCLs that express endogenous c-Src
disrupted actin ring formation, mimicking the cytoskeletal alterations observed in Src-/-
osteoclasts, and strongly inhibited the pit-forming activity (Fig. 3C). Furthermore,
promoting the open conformation of Src by mutating Y527 did not alter the inhibitory
effect of SrcK295M (Fig. 3C), indicating that Src’s adaptor function alone is not
sufficient to support osteoclastic bone resorption. Overexpressing the partially active
SrcY416F+Y527F mutant only partially inhibited the bone resorbing activity, further
indicating need for full Src kinase activity in osteoclastic bone resorption.
These results were confirmed in Src-/- OCLs, where the potential interference of
endogenously expressed Src is avoided. Most interestingly, while Src-/- OCLs were
100% devoid of resorbing activity, cells reconstituted with SrcK295M demonstrated a
very minimal (about 3%) but significant (p<0.001) level of pit forming activity (Fig. 4C),
23
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

which was still much less than the approximately 70% restoration of pit formation by
OCLs reconstituted with SrcY416F+Y527F. This minimal restoration of activity by
SrcK295M suggests a possible explanation of the apparent discrepancy between our results
with individual OCLs and Schwartzberg’s findings in animals. It is known that there are
several-fold more osteoclasts in Src-/- than in wild type animals, and the degree of
osteopetrosis in these animals is moderate (32). Even a small increase in the activity of
the larger number of osteoclasts acting over a longer time than our in vitro bone resorbing
assay could induce sufficient additional bone resorption to account for the partial rescue
reported by Schwartzberg and colleagues (5). Schwartzberg et al. did not note the time
required for tooth eruption, which might be longer in the mice expressing the mutant Src
proteins. It should also be noted that the Schwartzberg study found that SrcY416 rescued
the osteopetrotic phenotype more efficiently than SrcK295M. This is consistent with our
findings that both Src’s adaptor function, which allows Src to be properly recruited to
Pyk2 at the adhesion sites, and Src kinase activity are indispensable for efficient
cytoskeletal organization and osteoclastic bone resorption.
Elucidation of the role of Src in OC function requires the identification of the
signaling elements whose activities might to be modulated by the recruitment of active
Src kinase to Pyk2. We previously reported that the tyrosine phosphorylation of Cbl
appears to be particularly dependant on the presence of Src (10). Others have reported
24
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

that Src phosphorylates CblY731 (33) creating a binding site for the regulatory p85
subunit of PI3-K (22,24). PI3-K in turn is involved in a reorganization of the
cytoskeleton that results in cell spreading and migration of several cell types. In
osteoclast PI3-K activity is important for osteoclast activity (25). Our confirmation that
Src catalyzes the phosphorylation of CblY731 and demonstration that the mutation of that
residue to phenylalanine, thereby preventing the Src-catalyzed phosphorylation inhibits
bone resorption further supports our previous suggestion that Cbl is important substrate
of Src in osteoclasts and focus our attention on PI3-K as a likely downstream effector of
the Pyk2/Src/Cbl complex.
Src-dependant Akt activation is important for RANKL-induced osteoclast
survival (28), and Schwartzberg and colleagues reported that the expression of Src251, a
truncated Src molecule that lacks the kinase domain, decreases Akt activity and osteoclast
survival in transgenic mice (9). We therefore considered whether the requirement for Src
kinase activity could be related to the regulation of osteoclast survival. Src can also
contribute to the activation of Ras/ERK signaling pathway (34,35), which has also been
implicated in promoting osteoclast survival. However, our finding that down-regulation
of c-Src kinase activity had no effect on the survival of OCLs, together with the fact that
there is no significant difference between the survival of wild type OCLs and c-Src-/-
OCLs (28) suggests that Src makes little contribution to enhancing the survival of
25
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

osteoclasts.
In conclusion, our study demonstrates that both the formation of a Pyk2/Src
complex, presumably in the podosomes, and the kinase activity of Src are required for
bone resorption by osteoclasts. Src kinase activity may be required in part to
phosphorylate Cbl, thereby creating a binding site for PI3-K and recruiting it to the
Pyk2/Src/Cb complex. These results provide further insight into how the attachment of
osteoclasts to the bone surface regulates bone resorption and how Src participates in this
regulation.
26
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

ACKNOWLEDGEMENTS
We thank Y. Tanaka for assistance in making adenoviruses; K. Ford for
maintaining the mouse colony; K. Uematsu and A. Miyazaki for consistent support. This
work was supported by National Institutes of Health grant DE-04724 and AR-42927 (to
R.B.).
27
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

FOOTNOTES
1Abbreviations: Csk, C-terminal Src Family kinase; ERK, extracellular signal-regulated
kinase; GFP, green fluorescent protein; MEK, MAPK/ERK kinase; MOI, multiplicity of
infection; OCLs, osteoclast-like cells.
28
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

FIGURE LEGENDS
Figure 1. Adenovirus vector-dependent expression of Xpress-tagged Pyk2 Y402F in
OCLs inhibits bone resorption
(A) OCLs formed on culture dishes were infected with AxGFP, AxPyk2WT,
AxPyk2Y402F, or AxPyk2KD, at the indicated MOI’s. Total cell lysates were Western blotted
with anti-Xpress (top), then stripped and reprobed with anti-Pyk2 (middle) and anti-
actin (lower) antibodies. (B) Double immunofluorescence staining of F-actin (red) and
Xpress-tagged Pyk2 mutants (green) in OCLs infected with AxGFP, AxPyk2WT,
AxPyk2Y402F, or AxPyk2KD at an MOI of 100. (C) Interaction between c-Src and
Pyk2 mutants in OCLs. Src was immunoprecipitated with anti-Src antibody (rabbit
polyclonal) from total cell lysates of OCLs expressing individual Pyk2 mutants at an
MOI of 100, and the immune complexes were immunoblotted with anti-Xpress antibody
(top). Membranes were stripped and reprobed anti-Src antibody (bottom). (D)
Phosphorylation of Y402 of different Pyk2 constructs in OCLs. Xpress–tagged Pyk2
proteins were immunoprecipitated from total cell lysate of OCLs expressing Pyk2
mutants at an MOI of 100, and the immune complexes were immunoblotted with an
antibody specific for Pyk2 phosphorylated on Y402 (top). Membranes were stripped and
reprobed with anti-Xpress antibody (bottom). (E) Dentine-resorbing activity of OCLs
29
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

expressing the Pyk2 mutants. OCLs generated in co-culture was transferred onto dentine
and further cultured for 12 h, as described in Materials and Methods. The values are
means ± SD (n=8; *p<0.05, ** p<0.01 compared to non-infected OCLs).
30
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 2. The effect of Pyk2Y402F-overexpression on bone resorption is not rescued by
overexpression of CskKD
(A) Three days after infection, OCLs co-infected with AxCskKD and AxPyk2Y402F
were purified by enzymatic digestion as described in Experimental Procedures. Co-
infected OCLs expressed AxCskKD and AxPyk2Y402F at levels similar to those of the
cells infected with either AxCskKD or AxPyk2Y402F. (B) Src kinase activity in OCLs
co-infected with AxCskKD and AxPyk2Y402F was measured by in vitro kinase assay
using enolase as a substrate. Positions of endogenous Src and enolase are indicated on the
left. The amount of immunoprecipitated Src protein was assayed by immunoblotting
with anti-Src antibody. (C) Dentine-resorbing activity of OCLs co-infected with
AxCskKD and AxPyk2Y402F was measured as described in Fig. 1E. The values are means ± SD
(n=8; *p<0.05, ** p<0.01 compared to AxGFP-infected OCLs).
31
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 3. Effects of adenovirus vector-dependent expression of Src mutants in c-Src+/?
OCLs
(A) OCLs formed on culture dishes were infected with AxGFP, AxSrcK295M,
AxSrcK295M+Y427, or AxSrcY416F+Y527F at the indicated MOI’s. Total cell lysates were
immunoblotted with anti avian-Src antibodies (top), and then the membrane was stripped
and reprobed with anti-Src (middle) and anti-actin (bottom) antibodies. (B) Double
immunofluorescence staining of F-actin (red) and Src (green) in OCLs infected with
AxGFP, AxSrcK295M, AxSrcK295M+Y527F, or AxSrcY416F+Y527F at an MOI of
100. Colocalization of the proteins is seen as yellow. (C) Dentine-resorbing activity of
OCLs expressing the Src mutants. The values are means ± SD (n=8; *p<0.01, **
p<0.001 compared to non-infected OCLs).
32
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 4. Effects of adenovirus vector-dependent expression of Src mutants in Src-/-
OCLs
(A) Western blot of total cell lysate from Src+/? OCLs, uninfected Src-/- OCLs, and
Src-/- OCLs infected with AxGFP, AxSrcK295M, AxSrcK295M+Y427, or
AxSrcY416F+Y527F at an MOI of 100. Membranes were immunoblotted with anti-Src (Ab-1)
antibody (top), then stripped and reprobed with anti-actin (bottom) antibody. (B) Double
immunofluorescence staining of F-actin (red) and Src (green) in Src-/- OCLs infected
with AxGFP, AxSrcK295M, AxSrcK295M+Y527F, or AxSrcY416F+Y527F at an MOI
of 100. (C) Dentine-resorbing activity of Src-/- OCLs expressing the Src mutants. The
values are means ± SD (n=8; *p<0.05, ** p<0.01 compared to Src+/? OCLs).
33
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 5. Src-mediated phosphorylation of Cbl Y731is required for bone resorption
(A) Phosphorylation of CblY731. RAW 264.7 cells were treated either with or
without PP2 as indicated. Cbl was immunoprecipitated from lysates and the immune
complex were immunoblotted with anti-phosphoY731 antibodies (top). Membranes
were stripped and reprobed with anti-Cbl antibodies (bottom). (B) Western blot of
total cell lysates from Src+/- OCLs infected with AxGFP, AxCblWT and
AxCblY731F at an MOI of 100. Membranes were immunoblotted with anti-myc (top), then
stripped and reprobed with anti-Cbl (middle) and anti-actin antibodies (bottom). (C)
Dentine-resorbing activity of OCLs expressing Cbl proteins. The values are mean +
SD (**p <0.001 compared to AxGFP infected OCLs).
34
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Figure 6. Src kinase activity is not required for osteoclast survival
(A) Mouse co-cultures were infected with AxGFP, AxSrcK295M, AxCskWT, or
AxCskKD and/or AxMEKCA as indicated at an MOI of 100. OCLs co-infected with
AxMEKCA expressed SrcK2995M, CskWT, or CskKD protein at levels similar to those of the cells
infected without AxMEKCA. (B) After purification, OCLs were incubated in
αMEM/10%FBS for 24 h. The number of viable cells remaining 24 h after purification is
shown as a percentage of the cells at time 0. Modulation of c-Src kinase activity did not
significantly affect osteoclast survival in the presence or absence of MEKCA. * p<0.01
(C) Typical TRAP staining of OCLs 24 h after purification in the experiment quantified in
(B). Bar: 100 µm
35
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

References
1. Soriano, P., Montgomery, C., Geske, R., and Bradley, A. (1991) Cell 64, 693-7022. Horne, W. C., Neff, L., Chatterjee, D., Lomri, A., Levy, J. B., and Baron, R.
(1992) J. Cell Biol. 119, 1003-10133. Lowe, C., Yoneda, T., Boyce, B. F., Chen, H., Mundy, G. R., and Soriano, P.
(1993) Proc Natl Acad Sci U S A 90, 4485-4489.4. Sanjay, A., Houghton, A., Neff, L., Didomenico, E., Bardelay, C., Antoine, E.,
Levy, J., Gailit, J., Bowtell, D., Horne, W. C., and Baron, R. (2001) J. Cell Biol.152, 181-195
5. Schwartzberg, P. L., Xing, L., Hoffmann, O., Lowell, C. A., Garrett, L., Boyce, B. F., and Varmus, H. E. (1997) Genes Dev 11, 2835-2844.
6. Felsenfeld, D. P., Schwartzberg, P. L., Venegas, A., Tse, R., and Sheetz, M. P. (1999) Nat Cell Biol 1, 200-206.
7. Missbach, M., Jeschke, M., Feyen, J., Muller, K., Glatt, M., Green, J., and Susa, M. (1999) Bone 24, 437-449
8. Miyazaki, T., Takayanagi, H., Isshiki, M., Takahashi, T., Okada, M., Fukui, Y., Oda, H., Nakamura, K., Hirai, H., Kurokawa, T., and Tanaka, S. (2000) J Bone Miner Res 15, 41-51.
9. Xing, L., Venegas, A. M., Chen, A., Garrett-Beal, L., Boyce, B. F., Varmus, H. E., and Schwartzberg, P. L. (2001) Genes Dev 15, 241-253.
10. Tanaka, S., Amling, M., Neff, L., Peyman, A., Uhlmann, E., Levy, J. B., and Baron, R. (1996) Nature 383, 528-531
11. Tanaka, S., Takahashi, T., Takayanagi, H., Miyazaki, T., Oda, H., Nakamura, K., Hirai, H., and Kurokawa, T. (1998) J Bone Miner Res 13, 1714-1720
12. Miyazaki, T., Katagiri, H., Kanegae, Y., Takayanagi, H., Sawada, Y., Yamamoto, A., Pando, M. P., Asano, T., Verma, I. M., Oda, H., Nakamura, K., and Tanaka, S. (2000) J Cell Biol 148, 333-342
13. Duong, L. T., Nakamura, I., Lakkakorpi, P. T., Lipfert, L., Bett, A. J., and Rodan, G. A. (2001) J Biol Chem 276, 7484-7492
14. He, T. C., Zhou, S., da Costa, L. T., Yu, J., Kinzler, K. W., and Vogelstein, B. (1998) Proc Natl Acad Sci U S A 95, 2509-2514.
15. Duong, L. T., Lakkakorpi, P. T., Nakamura, I., Machwate, M., Nagy, R. M., and
36
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Rodan, G. A. (1998) J Clin Invest 102, 881-89216. Dikic, I., Tokiwa, G., Lev, S., Courtneidge, S. A., and Schlessinger, J. (1996)
Nature 383, 547-550.17. Xu, W., Harrison, S. C., and Eck, M. J. (1997) Nature 385, 595-60218. Xu, W., Doshi, A., Lei, M., Eck, M. J., and Harrison, S. C. (1999) Mol. Cell 3,
629-638.19. Kmiecik, T. E., and Shalloway, D. (1987) Cell 49, 65-73.20. Piwnica-Worms, H., Saunders, K. B., Roberts, T. M., Smith, A. E., and Cheng, S.
H. (1987) Cell 49, 75-82.21. Thien, C. B. F., and Langdon, W. Y. (2001) Nat. Rev. Mol. Cell. Biol. 2, 294-
305.22. Ueno, H., Sasaki, K., Honda, H., Nakamoto, T., Yamagata, T., Miyagawa, K.,
Mitani, K., Yazaki, Y., and Hirai, H. (1998) Blood 91, 46-5323. Hunter, S., Burton, E. A., Wu, S. C., and Anderson, S. M. (1999) J Biol Chem
274, 2097-210624. Kassenbrock, C. K., Hunter, S., Garl, P., Johnson, G. L., and Anderson, S. M.
(2002) J Biol Chem 277, 24967-2497525. Lakkakorpi, P. T., Wesolowski, G., Zimolo, Z., Rodan, G. A., and Rodan, S. B.
(1997) Exp Cell Res 237, 296-306.26. Palacio, S., and Felix, R. (2001) Eur J Endocrinol 144, 431-44027. Grey, A., Chen, Y., Paliwal, I., Carlberg, K., and Insogna, K. (2000)
Endocrinology 141, 2129-213828. Wong, B. R., Besser, D., Kim, N., Arron, J. R., Vologodskaia, M., Hanafusa, H.,
and Choi, Y. (1999) Mol Cell 4, 1041-1049.29. Lakkakorpi, P. T., Bett, A. J., Lipfert, L., Rodan, G. A., and Duong le, T. (2003) J
Biol Chem 278, 11502-1151230. Violette, S. M., Shakespeare, W. C., Bartlett, C., Guan, W., Smith, J. A., Rickles,
R. J., Bohacek, R. S., Holt, D. A., Baron, R., and Sawyer, T. K. (2000) Chem Biol7, 225-235
31. Parsons, J. T., Martin, K. H., Slack, J. K., Taylor, J. M., and Weed, S. A. (2000)Oncogene 19, 5606-5613.
32. Lowell, C. A., Niwa, M., Soriano, P., and Varmus, H. E. (1996) Blood 87, 1780-1792.
33. Feshchenko, E. A., Langdon, W. Y., and Tsygankov, A. Y. (1998) J. Biol. Chem.273, 8323-8331
34. Naor, Z., Benard, O., and Seger, R. (2000) Trends Endocrinol Metab 11, 91-9935. Migliaccio, A., Castoria, G., Di Domenico, M., de Falco, A., Bilancio, A.,
Lombardi, M., Barone, M. V., Ametrano, D., Zannini, M. S., Abbondanza, C., and Auricchio, F. (2000) Embo J 19, 5406-5417.
37
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Roland BaronTsuyoshi Miyazaki, Archana Sanjay, Lynn Neff, Sakae Tanaka, William C. Horne and
Src kinase activity is essential for osteoclast function
published online January 22, 2004J. Biol. Chem.
10.1074/jbc.M311032200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on June 13, 2018http://w
ww
.jbc.org/D
ownloaded from

![· Web viewglycolysis and tumor growth[22]. PKM2 is essential for TGF-induced EMT in several human cancers [16, 23]. The HIF-1α and c-Myc-hnRNP cascades are essential mediators](https://static.fdocument.org/doc/165x107/5e63c210f9d8e019e876dc5f/web-view-glycolysis-and-tumor-growth22-pkm2-is-essential-for-tgf-induced-emt.jpg)
















