
Sci 493 Without Paper II and III
143
Mannan-hydrolysis by hemicellulases Enzyme-polysaccharide interaction of a modular β-mannanase Per Hägglund Department of Biochemistry Lund University Sweden 2002 AKADEMISK AVHANDLING som för avläggande av filosofie doktorsexamen vid Matematisk-Naturvetenskapliga fakulteten vid Lunds Universitet offentligt kommer att försvaras i hörsal C, Kemicentrum, Getingevägen 60, fredagen den 31:a maj 2002, kl 10.15 Fakultetsopponent: Prof. R. Anthony J. Warren, Department of Microbiology and Immunology, University of British Columbia, Vancouver, BC, Canada
-
Upload
faradisa-anindita -
Category
Documents
-
view
20 -
download
5
description
nn
Transcript of Sci 493 Without Paper II and III

Mannan-hydrolysis by hemicellulasesEnzyme-polysaccharide interaction of a modular β-mannanase
Per Hägglund
Department of BiochemistryLund University
Sweden2002
AKADEMISK AVHANDLING
som för avläggande av filosofie doktorsexamen vid Matematisk-Naturvetenskapliga fakulteten vidLunds Universitet offentligt kommer att försvaras i hörsal C, Kemicentrum,
Getingevägen 60, fredagen den 31:a maj 2002, kl 10.15
Fakultetsopponent: Prof. R. Anthony J. Warren, Department of Microbiology and Immunology,University of British Columbia, Vancouver, BC, Canada


Denna avhandling tillägnas min Mormor Alice Rönnberg


Enzyme-polysaccharide interaction of a modular β-mannanase
Per Hägglund
Department of BiochemistryLund University
Sweden2002
Mannan-hydrolysis by hemicellulases

2002 Per HägglundLund UniversityDep. of BiochemistryCenter for Chemistry and Chemical engineeringLund UniversityP.O. Box 124S-221 00 LUNDSweden
ISBN: 91-628-5203-5LUNKDL/(NKBK-1074)/ 1-176 /2002Printed by JaBe Offset, Lund, Sweden

Publication List ………………………………………………………………………………………………………………………… 5Abbreviations …………………………………………………………………………………………………………………………... 6Summary ……………………………………………………………………………………………………………………………………… 7Introduction …………………………………………………………………………………………………………………………….… 9General background ………………………………………………………………………………………………………….… 13
Mannan-based polysaccharides ………………………………………………………………… ………….…..…… 13Mannans in wood: hemicelluloses …………………………………………………………………………….. 13
Algal mannans ………………………………………………………………………………………………………………… 14
Mannan-based storage polysaccharides ……………………………………………………………………… 14
Crystalline mannans ………………………………………………………………………………………………….…… 15
Polysaccharide-degrading enzymes: an introduction …………………………………………… 15Polysaccharide-degrading microorganisms ……………………………………………………………....… 15
Industrially important fungi ……………………………………………………………………………………….…… 16
Modularity of polysaccharide-degrading enzymes ………………………………………………..…… 16
Glycoside hydrolases ……………………………………………..………………………………………………………….… 16General catalytic features …………………………………………………………………………………………….… 16
Family classification …………………………………………………………………………………………………..….… 17
Structural features ………………………………………………………………………………………………………..…… 17
Carbohydrate-binding modules ………………………………………..………………………………………..…… 18Family classification …………………………………………………………………………………………………...…… 18
Structure and function ………………………………………………………………………………………………..….… 18
Cellulose-binding CBMs ………………………………………………………………………………………………… 19
Galactoglucomannan-degrading enzymes ………………………………………………………….……… 21β-Mannanase ………………………………………………………………………………………………………………...…… 21
Biochemical properties ……………………………………………………………………………………………….…… 21
Occurrence and regulation ……………………………………………………………………………………………… 23
Family classification and structural determination ………………………………………………..…… 23
Modular β-mannanases ………………………………………………………………………………………...………… 24
Applications ………………………………………………………………………………………………………………….…… 25
Exo-acting enzymes …………………………………………………………………………………………………..…… 25β-Mannosidase ……………………………………………………………………………………………………………….… 25
Other exo-acting enzymes ………………………………………………………………………………………….…… 26
Present investigation …………………………………………………………………………………………………….………. 29
Outline …………………………………………………………………………………………………..………………………………….… 29
3
Table of contents

Mannan-degrading enzymes ………………………………………………………………......………….…………… 30Aspergillus niger β-mannosidase, AnMan2A ……………………………………………………………… 30
The M. edulis β-mannanase, MeMan5A ……………………………………………………………………… 31
The modular T. reesei β-mannanase, TrMan5A …………………………………………..……………… 32
The CBM of TrMan5A ………………………………………………………………………………….....…….………… 33Sequence and structure …………………………………………………………………………………………………… 33
Binding properties of the CBM ……………………………………………………………….………………...… 34
Effect on hydrolysis ……………………………………………………………………………………………………….... 35
Why a cellulose-binding CBM on a β-mannanase? …………………………………………………… 36
The catalytic module of TrMan5A ………………………………………………………………………………… 37The active site cleft …………………………………………………………………………………………………………… 37
Characterisation of mutants ………………………………………………………………………..…………………… 37
Mutant Glu169Ala ………………………………………………………………………………………………….………… 38
Mannan-hydrolysis …………………………………………………………………………………………………………..…… 38Degradation of unsubstituted mannan ………………………………………………………………………….. 39
Crystallisation of mannan ………………………………………………………………………………………...……... 39
Degradation of mannan crystals ………………………………………………………………………………...….. 39
Degradation of heteromannans ……………………………………………………………………………...…..…… 40
Conclusions & future perspectives …………………………………………………………………. 43Acknowledgements ………………………………………………………………………………………………..………... 45References ………………………………………………………………………………………………………………………………… 55Papers I-VII
4

Publication List
This thesis is based on the following papers, referred to in the text by their Roman numerals.
I. Ademark, P., Lundqvist, J., Hägglund, P., Tenkanen, M., Torto, N., Tjerneld, F.and Stålbrand, H. (1999) Hydrolytic properties of a β-mannosidase purified fromAspergillus niger. J. Biotechnol. 75, 281-289. 1
II. Ademark, P., de Vries, R. P., Hägglund, P., Stålbrand, H. and Visser, J. (2001)Cloning and characterization of Aspergillus niger genes encoding an α-galactosi-dase and a β-mannosidase involved in galactomannan degradation. Eur. J.Biochem. 268, 2982-2990. 2
III. Hägglund, P., Sabini, E., Boisset, C., Wilson, K., Chanzy, H. and Stålbrand, H.(2001) Degradation of mannan I and II crystals by fungal endo-β-1,4-mannanasesand a β-1,4-mannosidase studied with transmission electron microscopy.Biomacromolecules 2, 694-699. 3
IV. Hägglund, P., Eriksson, T., Collén, A., Nerinckx, W., Claeyssens, M. andStålbrand, H. (2002) A cellulose-binding module of the Trichoderma reesei β-mannanase Man5A increases the mannan-hydrolysis of complex substrates. Sub-mitted for publication in J. Biotechnol.
V. Xu, B., Hägglund, P., Stålbrand, H. and Janson, J. C. (2002) endo-β-1,4-Mannanases from blue mussel, Mytilus edulis: purification, characterization, andmode of action. J. Biotechnol. 92, 267-277. 1
VI. Lundqvist, J., Hägglund, P., Eriksson. T., Persson, P., Stoll, D., Siika-aho, M.,Gorton, L. and Stålbrand, H. Degradation of glucomannan and O-acetyl-galactoglucomannan by mannoside- and glucoside-hydrolases. Manuscript.
VII. Hägglund, P., Anderson, L. and Stålbrand, H. The active site cleft of Trichodermareesei β-mannanase Man5A: analysis of mutants of the catalytic glutamates andarginine 171 positioned in the +2 subsite. Manuscript.
1 Reprinted with permission from Elsevier Science, Ltd.2 Reprinted with permission from Blackwell Publishing3 Reproduced with permission from Biomacromolecules. Copyright American Chemical Society.
5

Abbreviations
3D Three-dimensionalA. niger Aspergillus nigerAnMan2A A. niger β-mannosidaseArg ArginineC. fimi Cellulomonas fimiCBM Carbohydrate-binding moduleCfMan26A C. fimi β-mannanaseDP Degree of PolymerisationE. coli Escherichia coliEC Enzyme CommissionGH-A Glycoside Hydrolase clan AGln GlutamineGlu GlutamategpdA glyceraldehyde-3-phosphate dehydrogenaseLBG Locust Bean GumLys LysineM. edulis Mytilus edulisMeMan5A M. edulis β-mannanaseO-acetyl-GGM O-acetyl-galactoglucomannanP. cellulosa Pseudomonas cellulosaP. pastoris Pichia pastorisS. cerevisae Saccharomyces cerevisaeSLH S-Layer Homologysp. SpeciesT. reesei Trichoderma reeseiTrMan5A T. reesei β-mannanaseTrMan5A∆CBM Catalytic module of TrMan5ATrp TrypophanTyr Tyrosine
6

Summary
The enzymatic degradation of plant polysaccharides is a process of fundamental impor-tance in nature. Furthermore, polysaccharide-degrading enzymes are very important inmany industrial processes. Therefore, the study of these enzymes is an important field ofresearch. The degradation of the plant cell wall is a complex process that involves a widerange of enzymes, mainly produced by microorganisms. These enzymes include thosewhich degrade cellulose and hemicellulose – two of the main components in plant cellwalls. Such enzymes are often composed of two or several separated modules whichperform different functions. Carbohydrate-binding modules (CBMs) are frequently presentand are known to be important for efficient hydrolysis of cellulose. However, the role ofCBMs in hemicellulose degradation is less clear.
In this work, the structure and function of hemicellulose-degrading enzymes was inves-tigated. The focus was on enzymes which degrade mannan and heteromannans such asgalactoglucomannan, the major softwood hemicellulose. The main enzyme studied wasa β-mannanase (TrMan5A) produced by the filamentous fungus Trichoderma reesei.This enzyme is composed of a catalytic module and a CBM. In order to study the func-tion of the two modules, a mutant lacking the C-terminal CBM was constructed (PaperIV). By use of this mutant and the full-length enzyme, the binding properties of theCBM and its effect on hydrolysis of different substrates was investigated. The results ofthis study demonstrate that the CBM of TrMan5A has a positive influence on the hy-drolysis of complex mannan substrates containing cellulose. Furthermore, this increasein hydrolysis could be linked to binding of the CBM to the substrate. Binding studiesrevealed that the CBM binds to cellulose, but not to mannan.
The enzyme-polysaccharide interaction in the active site cleft of the catalytic module ofTrMan5A was also studied in this work (Paper VII). Several mutants of specific aminoacids were designed, based on the previously solved structure of the catalytic module.The enzymatic activity of the catalytic residue mutants was very low or abolished. Incontrast, a mutant of Arg171 (Arg171Lys) displayed activity in the same range as thewild-type enzyme. Interestingly, this mutant also appears to have a more alkaline pH-optimum than the wild-type. However, the Arg171Lys mutant was impaired in hydroly-sis of small substrates.
In addition to TrMan5A, the properties of a β-mannanase (MeMan5A) from blue mussel(Mytilus edulis) and a β-mannosidase (AnMan2A) from the fungus Aspergillus niger,were studied in this work (Papers I, II and V). MeMan5A belongs to the same enzymefamily as TrMan5A. Also, the β-mannosidase is related to the β-mannanases since theyare members of the same enzyme clan (GH-A). Studies on the catalytic properties of theenzymes showed that all three enzymes are capable of degrading polymeric mannan
7

(Papers I, V and VI). Furthermore, TrMan5A and AnMan2A also degraded highly crys-talline mannan, which was visualised by transmission electron microscopy (Paper III).
Also included in the present study is a comparative investigation of the enzymatic deg-radation of heteromannans (Paper VI). Here, glucomannan and galactoglucomannan weredegraded by several polysaccharide-degrading enzymes involved in the breakdown ofcellulose and hemicellulose. The results show that these substrates can be hydrolysed byboth mannoside- and glucoside-hydrolases.
In conclusion, this work showed that the enzyme-polysaccharide interaction in the twomodules of TrMan5A is important in determining overall enzymatic efficiency andspecificity in the hydrolysis of complex substrates. Altogether, the results presented dem-onstrate the need to use complex substrates in order to reveal the mechanisms of plantpolysaccharide degradation.
8

Introduction
Carbohydrates are essential for life on earth.They function as long-term storage depots of the energy captured in photosynthesis, and as inte-gral parts of genetic information carried in DNA. Furthermore, carbohydrates play many otherimportant roles in nature: for example, they are involved in intercellular communication and host/pathogen recognition. Moreover, carbohydrates in the form of polysaccharides are the main struc-tural elements of plants.
In terms of biomass, cellulose and hemicellulose are the most abundant polysaccharides on earthand are synthesised in huge amounts: it has been estimated that 1012 tonnes of cellulose are pro-duced per annum [60]. Thus, these polymers are powerful renewable resources. However, inorder to maintain a balance in the ecosystem, these polysaccharides must eventually be degraded.Even though the spontaneous degradation of polysaccharides under physiological conditions isthermodynamically favourable, it is exceedingly slow; it has been estimated that the half-life ofcellulose is almost 5 million years [282]. In nature, the rate of turnover of plant polysaccharides isenhanced by polysaccharide-degrading enzymes, produced mainly by various soil-livingdecomposers which degrade decaying plant material (Figure 1).
The plant cell wall is mainly composed of tightly associated cellulose, hemicellulose and lignin.Due to this complex structural composition, the degradation of the plant cell wall is a difficulttask. Accordingly, a complex mixture of enzymes is required in order to degrade the cell wallcomponents. Many of these enzymes have modular structures with separate carbohydrate-bind-ing modules (CBMs) which anchor the enzymes to different components in the cell wall [278].
Figure 1. Synthesis and degradation of plant polysaccharides.
9

In several cases, these CBMs are very important for efficient substrate hydrolysis. For example,generally cellulose-binding CBMs linked to cellulases mediate an increase in rate of cellulosedegradation [268]. However, cellulose-binding CBMs are also found in many hemicellulose-degrading enzymes and their function in these enzymes is more elusive [95].
Hemicelluloses comprise a family of diverse polysaccharides. Generally, hemicelluloses have acomplex chemical structure and are often referred to as mannans, xylans and galactans on thebasis of the predominant sugar type in the main chain. One of the most common mannans is O-acetyl-galactoglucomannan which comprises up to 25 % of the dry weight in softwood [263].However, a range of other mannan-type polysaccharides are synthesised by a wide variety ofplants, and are found in different types of plant tissue [185]. Their main role is often to function asstructural polysaccharides and/or as reserve energy.
Due to the complex structure of hemicellulose, several different hemicellulose-degrading en-zymes (usually referred to as an enzyme system) are produced for the complete degradation ofthese polymers into their monomeric components [26, 61]. Such a system often includes a com-bination of endo- and exo-acting enzymes. Two of the major endo-acting enzymes involved indegradation of hemicellulose are β-mannanase and β-xylanase. In the case of O-acetyl-galactoglucomannan, β-mannanase is the major depolymerising enzyme. In addition, the exo-acting enzymes β-mannosidase, α-galactosidase and β-glucosidase are needed for a completedegradation of galactoglucomannan.
In addition to their importance in nature, hemicellulases are important in many industrial applica-tions [183, 280, 284]. Polysaccharide-degrading enzymes in general are the second largest groupof commercially produced enzymes [100]. Furthermore, cellulases and hemicellulases accountfor approximately 20 % of the world enzyme market [193]. In particular, hemicellulases haveseveral existing and potential uses in the pulp and paper industry [280]. Environmental concernshave been raised against the use of large quantities of chlorinated compounds in pulp bleachingprocesses. Treatment of pulp with β-xylanases prior to bleaching reduces the amount of bleach-ing chemicals needed in the process, thus providing an environment-friendly alternative [280]. Inaddition, β-mannanases also have potential uses in pulp-bleaching [45, 249].
From prehistoric time, wood have been used by humans as building material and as an energysource. The more refined industrial uses of wood have been focused on the cellulose component.However, in recent years it has been realised that hemicelluloses also have many potential indus-trial applications [89]. As environmental problems increase, it is likely that recyclablepolysaccharides will be attractive for industrial applications in the future [16]. In order to studythe structure of hemicelluloses, and to modify their properties, hemicellulose-degrading enzymesare important tools [158]. Thus, research in these enzymes is of vital interest.
The general aim of the current work was to increase the understanding of the molecular mecha-nisms in mannan-degrading hemicellulases. In particular the enzyme-polysaccharide interactionin a number of mannan-degrading enzymes was investigated. The main enzyme studied was amodular β-mannanase (TrMan5A) from Trichoderma reesei, which contains a catalytic moduleand a CBM. In this work, the binding properties of the CBM and its effect on the overall activityof this enzyme was studied (Paper IV). Moreover, the substrate interaction in the active site cleftof the catalytic module of TrMan5A was investigated (Paper VII).
10

In addition, the molecular properties of a β-mannosidase (Papers I and II) from the fungus As-pergillus niger and a β-mannanase (Paper V) from the blue mussel (Mytilus edulis)were studied. Furthermore, the activity of these enzymes and TrMan5A were investigated, withan emphasis on their ability to degrade mannan polymers (Papers I, III and VI).
This thesis is divided into two major parts (see Table of contents, page 3)
In the first part of this thesis (General background), the mannan-degrading enzymes are de-scribed in the context of plant cell-wall degradation. First, the mannan-containing polysaccharidesare presented. Then, an overview of polysaccharide-degrading enzymes is given. Finally, themannan-degrading enzymes are introduced, with an emphasis on β-mannanases.
In the second part (Present investigation), the results described in Papers I-VII (listed on page 5)are presented and discussed.
11

12
General background
Mannan-based polysaccharides
Mannans in wood: hemicelluloses
Wood comprises the bulk of the tree trunk and is essentially composed of the cell walls of xylemcells. The major constituents of these cell walls are cellulose, hemicellulose and lignin [83]. Therelative amounts of cellulose, hemicellulose and lignin can vary to some degree, depending onthe species (Table 1). Furthermore, the chemical composition of wood depends on the tissue, celltype and growth rate [226].
Table 1. Composition of woods from some representative species. Data from Timell [263]
Species Cellulose (%) Hemicellulose (%) Lignin (%)Acer rubrum 45 29 24
Betula papyrifera 42 38 19
Fagus grandifora 45 29 22
Pinus strobus 41 27 29
Picea glauca 41 31 27
The concept of hemicellulose was introduced by Schulze in 1891, to define alkali-extractableplant polysaccharides [83]. Hemicelluloses are a group of heteropolysaccharides with a degree ofpolymerisation (DP) around 100-200 [263]. They are built up by a main chain composed of oneor several types of sugar monomers. In addition, different types of side-groups are frequentlyattached to the main chain [234].
As seen in Table 2, different hemicelluloses are found in wood derived from gymnosperms(softwoods) and angiosperms (hardwoods) [263]. The amounts of the different hemicellulosescan also vary considerably depending on the cell type and the stage in development. In softwoods,the major hemicelluloses are O-acetyl-galactoglucomannan and arabino-4-O-methylglucuronoxylan, an exception being larchwood where arabinogalactan is the predominanthemicellulose [263]. In hardwoods, the major hemicellulose component is O-acetyl-4-O-methylglucuronoxylan, but a smaller amount of glucomannan is also found (Table 2).
13

Table 2. Hemicelluloses in hardwoods and softwoods. Data from Timell [263].
Hemicellulose Amount (% of wood ) Degree of polymerisationHardwood
Glucomannan 3-5 >70
O-acetyl-4-O-methylglucuronoxylan 10-35 ~200
Softwood
Arabino-4-O-methylglucuronoxylan 10-15 >120
O-acetyl-galactoglucomannan 15-25 >100
Arabinogalactana 10-20 ~220a In larchwood only
Hardwood glucomannan is built up by a linear chain of β-(1→4)-linked mannose and glucoseresidues. The mannose/glucose ratio in glucomannans is typically in the range 1/1-2/1 [263].Galactoglucomannan is built up by a glucomannan main chain, but also includes α-(1→6)-linkedgalactosyl side-groups attached at some mannose residues (Figure 2). In softwoodgalactoglucomannan, acetyl groups have been reported to be attached at the C-2 and C-3 posi-tions of some mannose residues [150, 158], in an apparently random fashion [139].O-acetyl galactoglucomannan can be divided in two fractions: one soluble in water, which has agalactose/glucose/mannose ratio of 1/1/3 and one soluble in aqueous alkali, which has a galac-tose/glucose/mannose ratio of 0.1/1/3 [263].
Figure 2. A schematic view of O-acetyl-galactoglucomannan.
Algal mannans
A range of different polysaccharides are produced by algae [208], some of which have industrialimportance [79]. In vivo, two of the main roles of these polysaccharides in the algae are to func-tion as reserve energy and as structural material. Linear β-(1→4) mannan is present in the cellwalls of several siphonaceous green algae in the families Acetabularia, Codium and Halicoryne[88, 126, 157]. Furthermore, mannan is also found in some red algae, such as Porphyra umbilicalis
14

[135]. In some of these algae, mannan is the main structural polymer and displays a microfibrillarmorphology [51, 166]. Some algal mannans display a high degree of polydispersity: the mannanfrom Codium fragile has a degree of polymerisation between 20 and 10 000 [167].
Mannan-based storage polysaccharides
Besides amylose and amylopectin, which are the most widespread storage polysaccharides inplants, there is a diverse group of mannan-based storage polysaccharides found in the seeds,roots, bulbs and tubers of various plants [185]. These include the mannans, galactomannans andglucomannans which are discussed below.
Mannan: Linear chains of β-(1→4) mannan are found in the plant seed endosperms of certainplant species [18, 184, 283]. Mannan has been isolated from ivory nut (Phytelephas macrocarpa),date (Phoenix dactylifera) and green coffee bean (Coffea arabica). In most cases, thesepolysaccharides are highly insoluble in water and very dense. Accordingly, it has been suggestedthat the mannan forms the molecular basis for the hardness which is characteristic for palm ker-nels, such as the ivory nut. In the cell wall of the seed endosperm of ivory nut, mannan is themajor component and it has been characterised in some detail. Based on their solubility in alkali,two different fractions of mannan have been isolated from the ivory nut [162]. These fractionsdiffer mainly in their DP [18, 184] and morphology [51, 184].
Galactomannan: Galactomannans are reserve polysaccharides in the seed endosperm of legu-minous plants (Leguminosae) [217]. In contrast to unsubstituted mannans, the galactomannansare water soluble and can imbibe water, thus providing a water-holding function for the seed[217]. They are composed of β-(1→4)-linked mannan chains with α-(1→6)-linked galactosylside groups [179]. Both the solubility and the viscosity of the galactomannans are influenced bythe mannose/galactose ratio, which can vary from 1 to 5 [217]. Furthermore, the distribution ofthe substituents can vary considerably [67], which also affects the physical properties ofgalactomannans [73]. Two of the most well characterised galactomannans are those found inlocust bean gum and guar gum, isolated from the seeds of Ceratonia siliqua and Cyanaposistetragonolobus, respectively [101, 220]. Locust bean gum galactomannan has a mannose/galac-tose ratio of approximately 5/1 and a molecular weight of 310 000 [220]; guar gum galactomannanhas a mannose/galactose ratio of 2/1 and a molecular weight of 220 000 [101]. In combinationwith other polysaccharides, these galactomannans have strong gelling properties and are thusused as thickeners in the food and feed industries. Galactomannans and their derivatives are alsoused in paper making, mining and in the textile industry [101, 220].
Glucomannan: Some glucomannans are found as storage polysaccharides in the seeds of certainannual plants, for example some lilies (Liliaceae) and irises (Iridaceae) [185]. Furthermore,glucomannans are found in the bulbs, roots and tubers of several other types of plants [185].Many of these glucomannans are water soluble and have the same general structure asglucomannans found in wood: they are composed of a β-(1→4)-linked mannan chain with inter-spersed glucose residues in the main chain and are often acetylated [185]. The mannose/glucoseratio ranges from 4/1 to below 1/1 [185]. One of the most thoroughly characterised of theseglucomannans is the so-called konjac mannan – isolated from the tubers of Amorphophalluskonjac [198]. This polysaccharide has a mannose/glucose ratio of 1.6/1 and a degree of polymeri-sation above 6000. [198].
15

Crystalline mannans
Polysaccharides in nature can be organised in more or less regular structures; ranging from ir-regular or amorphous structures to highly organised and crystalline structures. The most abun-dant polysaccharide in nature, cellulose, is partially crystalline. In addition to cellulose, X-rayanalysis has yielded information about the structures of a number of other polysaccharides suchas chitin, xylan, amylose and mannan [169].
Crystalline linear β-(1→4) D-mannan has been found in the cell walls of ivory nuts and in thealgae Acetabularia crenulata and Codium fragile [51, 88, 184]. Two polymorphs – mannans Iand II – have been observed in these cell walls. The morphologies of mannan I and mannan II aregranular and fibrillar, respectively [51]. Crystalline glucomannan from wood can be obtained,but only after partial degradation or modification [262]. After dissolution of glucomannan inalkali it can be recrystallised as mannan I or mannan II, depending on the crystallisation condi-tions [50]. Furthermore both mannan and glucomannan can be recrystallised onto cellulose fibers,yielding the so-called shish/kebab (cellulose/mannan) morphology [50]. Native glucomannanand O-acetyl-galactoglucomannan appear to be mostly non-crystalline in nature, probably due totheir more complex structures [169].
Polysaccharide-degrading enzymes: an introduction
Polysaccharide-degrading microorganisms
Bacteria and fungi thriving on decaying plant material constitute an important part of the ecosys-tem. These microorganisms decompose polysaccharides and other plant materials, thus recyclingorganic and inorganic material in the atmosphere and biosphere. Many of these polysaccharide-degrading microorganisms are soil- or water-living. However, some polysaccharide degradinganaerobes degrade plant polysaccharides in the stomach of ruminants [55]. For severaldecomposers, plant cell wall polysaccharides are the principal carbon and energy source. Accord-ingly, these organisms usually produce secreted enzymes which degrade the polysaccharides intomono- and oligosaccharides which can be further metabolised in the cell.
Industrially important fungi
The ability of some soil-living fungi to produce large amounts of polysaccharide-degrading en-zymes and other groups of extracellular enzymes (e.g. proteinases and lipases) makes them at-tractive for use in industrial enzyme production. Fungi from the genera Aspergillus and Trichode-rma are examples of two families of industrially important fungi. Notably, Trichoderma reesei isa potent producer of several cellulases and hemicellulases which are widely used in industrialapplications [279, 280]. Besides its high enzyme production capacity, T. reesei has the advantageof being non-toxic and non-pathogenic which is important in large scale fermentation processes[196]. In addition to endogenous enzymes, strains of both Aspergillus and Trichoderma have also
16

been used as hosts for expression of foreign eukaryotic proteins [142, 206]. For example, T.reesei has been used for the production of single chain antibodies and mammalian interleukin[206].
Modularity of polysaccharide-degrading enzymes
Polysaccharide-degrading enzymes often have a modular structure. By definition, modular en-zymes are composed of two or more independently folded modules – each formed from a con-tiguous sequence [37]. Furthermore, each module has a distinct structure which is related to itsfunction. The overall structures and the functionally important residues are often conserved amongstsimilar modules. A modular architecture has been found amongst lipases, endonucleases, peptidesynthases and several other classes of enzymes [141]. Modular polysaccharide-degrading en-zymes are commonly composed of a catalytic module connected to one or more additional cata-lytic or non-catalytic module [278]. In many cases, these modules are connected by linker se-quences [96]. The most common modules are the catalytic O-glycoside hydrolases and the non-catalytic carbohydrate-binding modules (CBM), which are discussed below. However, severaladditional non-catalytic modules have also been described [268]. Thermostabilizing moduleshave been found in some xylanases [114], and S-layer homology (SLH) modules, involved incell-surface attachment, have been described in several types of polysaccharide-degrading en-zymes [268]. Dockerin modules, found in polysaccharide-degrading enzymes from anaerobicbacteria and fungi, attach the enzymes to the “cellulosome” – a large multi-enzyme complexcomposed of several different enzymatic activities [230]. This cellulosome is composed of a corestructure called a scaffold, onto which several types of enzymes can be attached.
Glycoside hydrolases
General catalytic features
The O-glycoside hydrolases constitute the main group of enzymes which participate in the degra-dation of plant polysaccharides. The diversity of carbohydrate structures is reflected in the widerange of substrate specificity found amongst glycoside hydrolases. The structure and function ofglycoside hydrolases has been intensively studied: the first enzyme structure solved by X-raydiffraction was a glycoside hydrolase (hen egg-white lysozyme), solved in 1965 [30]. Based onkinetic data and later on structural information, a general mechanism for glycoside hydrolaseshas been proposed [70, 144, 233]. According to this theory, two main mechanisms for glycosidehydrolases exist. These are called retaining and inverting mechanisms, referring to the stere-ochemical outcome at the anomeric carbon in the product.
In both mechanisms, the hydrolysis of the glycosidic bond proceeds through general acid/basecatalysis involving two carboxylates (glutamates or aspartates) positioned in the active site [281].The inverting mechanism proceeds through a single step reaction involving the direct attack by anucleophilic water on the anomeric carbon, and the simultaneous protonation of the glycosidic
17
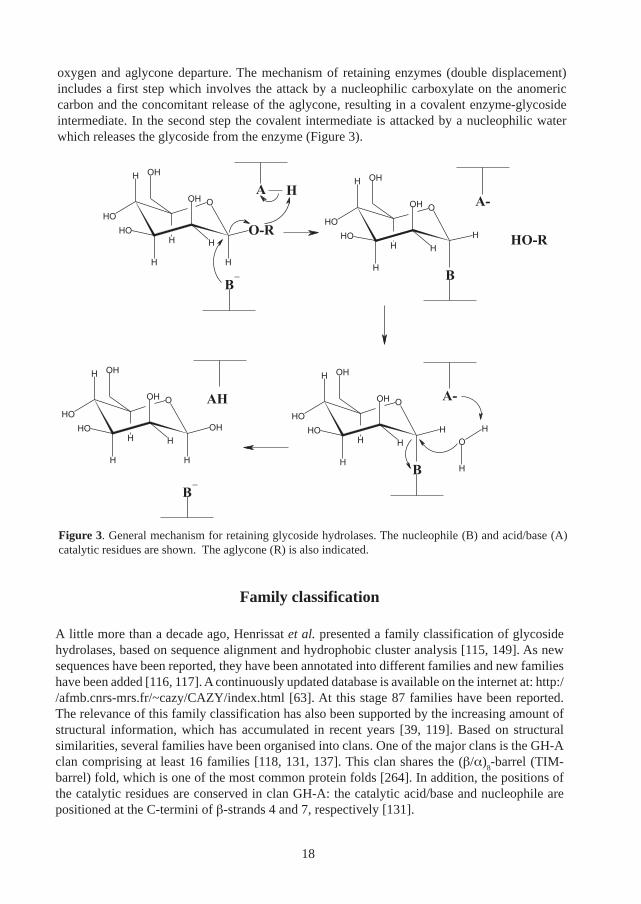
oxygen and aglycone departure. The mechanism of retaining enzymes (double displacement)includes a first step which involves the attack by a nucleophilic carboxylate on the anomericcarbon and the concomitant release of the aglycone, resulting in a covalent enzyme-glycosideintermediate. In the second step the covalent intermediate is attacked by a nucleophilic waterwhich releases the glycoside from the enzyme (Figure 3).
Figure 3. General mechanism for retaining glycoside hydrolases. The nucleophile (B) and acid/base (A)catalytic residues are shown. The aglycone (R) is also indicated.
Family classification
A little more than a decade ago, Henrissat et al. presented a family classification of glycosidehydrolases, based on sequence alignment and hydrophobic cluster analysis [115, 149]. As newsequences have been reported, they have been annotated into different families and new familieshave been added [116, 117]. A continuously updated database is available on the internet at: http://afmb.cnrs-mrs.fr/~cazy/CAZY/index.html [63]. At this stage 87 families have been reported.The relevance of this family classification has also been supported by the increasing amount ofstructural information, which has accumulated in recent years [39, 119]. Based on structuralsimilarities, several families have been organised into clans. One of the major clans is the GH-Aclan comprising at least 16 families [118, 131, 137]. This clan shares the (β/α)
8-barrel (TIM-
barrel) fold, which is one of the most common protein folds [264]. In addition, the positions ofthe catalytic residues are conserved in clan GH-A: the catalytic acid/base and nucleophile arepositioned at the C-termini of β-strands 4 and 7, respectively [131].
18

Structural features
Glycoside hydrolases have been found to have one of two major cleavage preferences: exo-actingenzymes hydrolyse residues at the chain ends and endo-acting enzymes hydrolyse glycosidicbonds internally in the polysaccharide chain [233]. For several structurally determined enzymesthe cleavage preference is reflected by the active site architecture (Figure 4) [69]. Endo-actingenzymes, such as endoglucanase and β-mannnanase, often have cleft shaped active site surfaces[76, 223]. On the other hand, pocket-shaped active site structures are found amongst several exo-acting enzymes which release monosaccharides from non-reducing ends, such as β-galactosidaseand glucoamylase [8, 136]. A tunnel-shaped active surface is found in cellobiohydrolases, whichrelease cellobiose from the reducing or non-reducing ends of cellulose [75]. However, it shouldbe pointed out that the architectures of the active sites of endo- and exo-acting enzymes does notnecessarily reflect the observed cleavage preference – discrepancies may exist. Furthermore, theboundaries between exo- and endo-acting enzymes are not absolute: enzymes which are mainlyendo-active may show exo-activivity and vice versa [241, 266].
In glycoside hydrolases, binding of several glycoside residues in the active site region is oftenrequired. Accordingly the substrate binding surface can be divided into several subsites. Accord-ing to the recommended nomenclature [72], these are numbered +4, +3, +2, +1, -1, -2 etc., fromthe non-reducing end to the reducing end, with the O-glycosidic bond to be cleaved being posi-tioned between subsite +1 and –1 (Figure 5).
Figure 5. A schematic overview of the sugar residue binding subsites in the active site region in glycosidehydrolases. The arrow indicates the point of cleavage.
Figure 4. Examples of active site topologies in glycoside hydrolases: (a) the pocket (glucoamylase fromAspergillus awamori). (b) The cleft (endoglucanase from Clostridium thermocellum). (c) The tunnel(cellobiohydrolase Cel6A from Trichoderma reesei). Adapted from [222].
19

Carbohydrate-binding modules
Family classification
Carbohydrate-binding sites in general are common and are found in several types of proteins,such as toxins, sugar transporters, lectins, antibodies and glycolytic enzymes [214]. Carbohy-drate-binding modules (CBMs) however, are found mostly in polysaccharide-degrading enzymes.CBMs have been widely studied in cellulose-degrading enzymes which bind tightly to cellulose[257, 267, 269, 276]. These binding entities were originally referred to as “cellulose-bindingdomains (CBDs)” [269]. However, more recently CBMs which bind to chitin, starch, xylan, mannanand other polysaccharides have been described, not only in cellulases but also in other polysac-charide-degrading enzymes [278]. In order to encompass a broader binding specificity, the con-cept of the carbohydrate-binding module (CBM) was introduced [36]. By analogy to the glyco-side hydrolases, CBMs are classified into families based on sequence similarities and when pos-sible, 3D-structures [36, 39, 95, 269]. This classification is continuously updated and is availableat: http://afmb.cnrs-mrs.fr/~cazy/CAZY/index.html [63]. Presently, 30 families of CBMs havebeen reported, some of which are listed in Table 3. A number of these families are closely relatedand consequently were recently grouped into a superfamily [66, 247].
Structure and function
Depending on their binding properties, three types of CBMs have been identified [36]. Type ACBMs bind to insoluble polysaccharides, such as cellulose and chitin with high affinity [155].These types of CBMs, are found in e.g. family I, II, III and V. Type B binding modules, found ine.g. family II and IV, bind soluble polysaccharides and oligosaccharides, and in some cases alsoamorphous, insoluble polysaccharides [265]. In addition to poly- and oligosaccharides, type CCBMs also have relatively high affinity for monosaccharides [34]. These CBMs are found in e.g.family IX and XIII.
The structure of CBMs from several families has been solved by X-ray crystallography and NMRspectroscopy [39, 95]. In most cases, the structures are built up mainly of β-sheets, often with ananti-parallel β-strand topology [39]. In general, CBMs which bind crystalline polysaccharides(type A) display a flat ligand-binding surface with exposed aromatic residues [145, 288] (Figure6). On the other hand, several modules which bind amorphous polysaccharides and oligosaccharides(Type B) display a groove-shaped binding surface lined with polar residues [132, 143]. Recently,the structure of a Type C CBM from family IX was solved and it was found to have a slot-shapedbinding site, “large enough to accommodate a disaccharide” [34, 202].
Analyses of mutational and chemical modifications have demonstrated the importance of thearomatic residues found on the flat surfaces of type A CBMs in ligand binding [41, 152, 218]. Inat least some type B CBMs, both aromatic and polar residues have been shown to contributesignificantly to ligand binding [143, 285]. The significance of aromatic and polar residues inCBMs is not surprising; the presence of these types of residues is a recurring theme in protein-carbohydrate interactions [214]. A general scheme in protein-carbohydrate associations is that
20

aromatic residues are aligned face-to-face with the hydrophobic face of the sugar ring, thus pro-viding stacking interactions. Polar residues, on the other hand, are thought to make hydrogenbonds with the hydrophilic hydroxyl groups. Thermodynamic analyses of CBM binding haveshown that, generally, binding of soluble polysaccharides and oligosaccharides is promoted mainlyby a decrease in enthalpy, resulting from several hydrogen bonding contacts between the proteinand the ligand [34, 35, 42, 133, 250]. In contrast, binding of a type A CBM to crystalline cellulosehas been shown to be driven by an increase in entropy, partly due to dehydration of the ligand-binding surface [64].
Cellulose-binding CBMs
Some of the most studied cellulose-binding CBMs are those from the fungus T. reesei, and fromthe bacteria Cellulomonas fimi and Pseudomonas cellulosa [36, 114, 155, 269]. The cellulose-binding CBMs from T. reesei are relatively small (30-40 residues) and classified into family I. Amajority of the family I CBMs are connected to cellulases of fungal origin [155, 257]. However,a few fungal hemicellulases, including the T. reesei β-mannanase also carry family I CBMs [171,244]. In T. reesei cellulases it has been shown that family I CBMs are important for efficienthydrolysis of cellulose [257, 267, 276]. Two of the most studied T. reesei CBMs are those fromthe endoglucanase Cel7B and the cellobiohydrolase Cel7A [151, 152, 154]. The solved 3D struc-tures of these CBMs reveal that they share a similar overall fold [145, 174]. In addition, structuraland mutational work has shown that the three exposed aromatic residues on the flat bindingsurfaces of the CBMs are important for binding [152, 173, 175, 218, 219]. However, the twoCBMs show different binding affinities for cellulose and it has been proposed that this differencedepends to a large degree on a single amino acid substitution on the flat binding surface [151].Furthermore, substitution of the CBM in Cel7A with the CBM from Cel7B demonstrated that anincreased binding to cellulose of the hybrid enzyme could be linked to an increase in cellulosehydrolysis [236].
The CBMs from C. fimi are mainly classified into family II and IV [36, 269]. The modules inthese families are found only in bacteria and are larger than those in family I (100-150 residues).One of the most studied CBMs is the family II CBM from the C. fimi enzyme CfXyn10A which
Figure 6. Examples of structures of type A (a) and type B (b) CBMs. a. The family I CBM from the T. reeseiendoglucanase TrCel7A [145]. b. The family IV CBM from the C. fimi endoglucanase CfCel9B [42].Residues implicated in binding are shown in red.
21
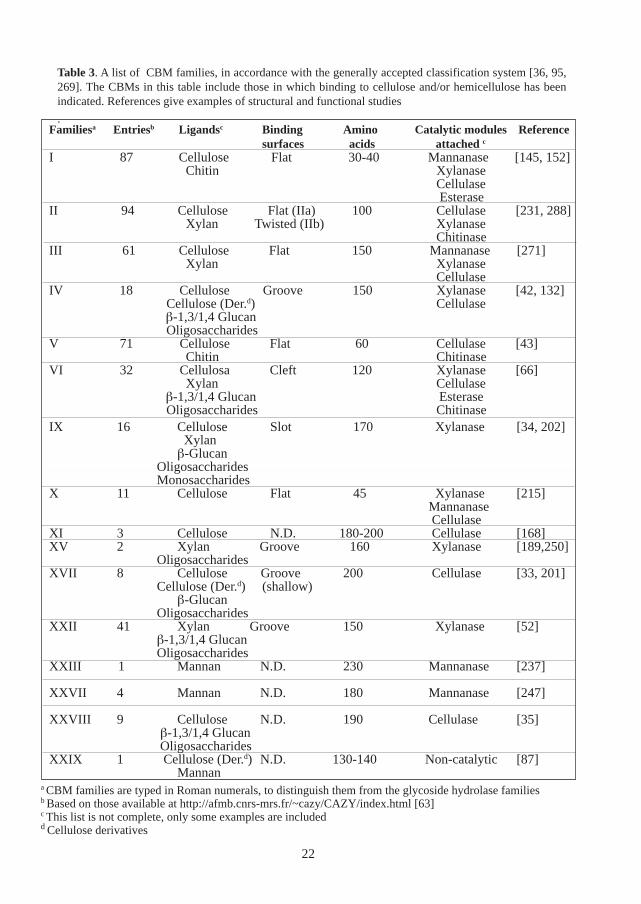
Table 3. A list of CBM families, in accordance with the generally accepted classification system [36, 95,269]. The CBMs in this table include those in which binding to cellulose and/or hemicellulose has beenindicated. References give examples of structural and functional studies
.
22
Familiesa Entriesb Ligandsc Binding Amino Catalytic modules Referencesurfaces acids attached c
I 87 Cellulose Flat 30-40 Mannanase [145, 152]Chitin Xylanase
Cellulase Esterase
II 94 Cellulose Flat (IIa) 100 Cellulase [231, 288]Xylan Twisted (IIb) Xylanase
ChitinaseIII 61 Cellulose Flat 150 Mannanase [271]
Xylan Xylanase Cellulase
IV 18 Cellulose Groove 150 Xylanase [42, 132] Cellulose (Der.d) Cellulase
β-1,3/1,4 Glucan Oligosaccharides
V 71 Cellulose Flat 60 Cellulase [43]Chitin Chitinase
VI 32 Cellulosa Cleft 120 Xylanase [66]Xylan Cellulase
β-1,3/1,4 Glucan Esterase Oligosaccharides Chitinase
IX 16 Cellulose Slot 170 Xylanase [34, 202] Xylanβ-Glucan
Oligosaccharides Monosaccharides
X 11 Cellulose Flat 45 Xylanase [215] Mannanase Cellulase
XI 3 Cellulose N.D. 180-200 Cellulase [168]XV 2 Xylan Groove 160 Xylanase [189,250]
OligosaccharidesXVII 8 Cellulose Groove 200 Cellulase [33, 201]
Cellulose (Der.d) (shallow)β-Glucan
OligosaccharidesXXII 41 Xylan Groove 150 Xylanase [52]
β-1,3/1,4 Glucan Oligosaccharides
XXIII 1 Mannan N.D. 230 Mannanase [237]
XXVII 4 Mannan N.D. 180 Mannanase [247]
XXVIII 9 Cellulose N.D. 190 Cellulase [35] β-1,3/1,4 Glucan Oligosaccharides
XXIX 1 Cellulose (Der.d) N.D. 130-140 Non-catalytic [87]Mannan
a CBM families are typed in Roman numerals, to distinguish them from the glycoside hydrolase familiesb Based on those available at http://afmb.cnrs-mrs.fr/~cazy/CAZY/index.html [63]c This list is not complete, only some examples are includedd Cellulose derivatives

Galactoglucomannan-degrading enzymes
Due to the complex chemical structure of hemicelluloses, multiple enzymes are often needed forthe complete degradation of these polymers. In the case of O-acetyl-galactoglucomannan, fourdifferent glycoside hydrolases (β-mannanase, β-mannosidase, α-galactosidase and β-glucosidase)and acetyl esterase are required for the complete conversion of the polymer into unsubstitutedmonosaccharides (Figure 7). β-Mannanase is the major endo-acting enzyme involved ingalactoglucomannan degradation [26, 180, 243]. This enzyme degrades the polymer into smalleroligosaccharides, which are further hydrolysed to monosaccharides by β-mannosidase, α-galac-tosidase and β-glucosidase.
display mixed glucanase/xylanase activity [97, 98, 288]. This CBM displays a ligand-bindingarea somewhat resembling the flat surface in family I CBMs [288], and it has been shown to bindtightly to cellulose [97]. Furthermore the hydrolysis of cellulose by CfXyn10A increases in thepresence of the attached CBM [98]. In constrast, another family II CBM from C. fimi module hasbeen proposed to promote non-hydrolytic disruption of cellulose surfaces, independent of thecatalytic module [74]. Family II has been divided into two subfamilies, on the basis of the pre-ferred binding ligand: family IIa modules bind to cellulose and family IIb modules bind to xylan[97, 231]. Remarkably, site-directed mutagenesis of a single amino acid residue on a family IIbCBM changed its binding specificity from xylan to cellulose [232]. The two tandem family IVCBMs in the C. fimi endoglucanase Cel9B bind to amorphous cellulose and oligosaccharides[133, 265].
Cellulose-binding CBMs from family II and family X are ubiquitous among polysaccharide-degrading enzymes from P. cellulosa, and are found in both cellulases and several types ofhemicellulases [114]. In P. cellulosa, cellulose-binding CBMs from both cellulases andhemicellulases have been shown to influence hydrolytic activity [28, 29, 109]. In some cases, twoCBMs are connected to the same enzyme. For example, the P. cellulosa xylanase Xyl10A con-tains one family II CBM and one family X CBM which bind to cellulose apparently independentof each other [99].
23
Figure 7. Schematic overview of the enzymes involved in degradation of O-acetyl-galactoglucomannan.

β-Mannanase
Biochemical properties
Endo-β-1,4-D-mannanase (β-mannanase; EC 3.2.1.78) catalyses the random hydrolysis of manno-glycosidic bonds in mannan-based polysaccharides. Most β-mannanases degrade manno-oligosaccharides down to a DP of 4 [26, 180, 243]. In addition, some β-mannanases are alsoactive on mannotriose, although at a much lower rate, thus indicating the presence of at least 4subsites in several β-mannanases [7, 112]. However, hydrolysis of oligosaccharides by some β-mannanases from anaerobic fungi and bacteria has indicated that binding is required over at least6 subsites [82, 110, 190]. The main end-products of mannan hydrolysis by β-mannanase are oftenmannobiose and mannotriose [3, 57, 225, 245, 273], although minor amounts of mannose andmannotetraose also are produced in some cases [273]. In the degradation of heteromannans, thepattern of released oligosaccharides is often more complex, probably due to hindrance of theenzymatic hydrolysis caused by the substituents [182, 210, 240, 259]. It has also been shown thatat least some β-mannanases are capable of degrading crystalline mannan (Paper III).
In addition to hydrolysis, several β-mannanases can also perform transglycosylation [62, 107,111]. For example, the T. reesei β-mannanase has been shown to form transglycosylation prod-ucts with either mannose or mannobiose as glycosidic bond acceptors [111].
The pH and temperature optima of some β-mannanases are shown in Table 4. Generally, β-mannanases have moderate temperature optima (40-70 °C), except some β-mannanases fromthermophiles which have higher temperature optima [93, 205, 209, 248]. The pH optima of mostβ-mannanases are found in the neutral or acidic region. Commonly, the molecular weights of β-mannanases are in the range of 30-80 kDa (Table 4).
However, some modular β-mannanases have molecular weights near or above 100 kDa [46, 239,248]. Typically, most β-mannanases have isoelectric points between 4 to 8. Frequently, multipleβ-mannanases with different isoelectric points and/or molecular weights are found in the sameorganism [4, 125, 172, 190, 245, 270]. In some cases these enzymes are produced from differentgenes [172, 190, 199], and in other cases, the enzymes are isoforms produced from the same gene[4, 244]. These isoforms may be caused by differences in post-translational modifications.
24
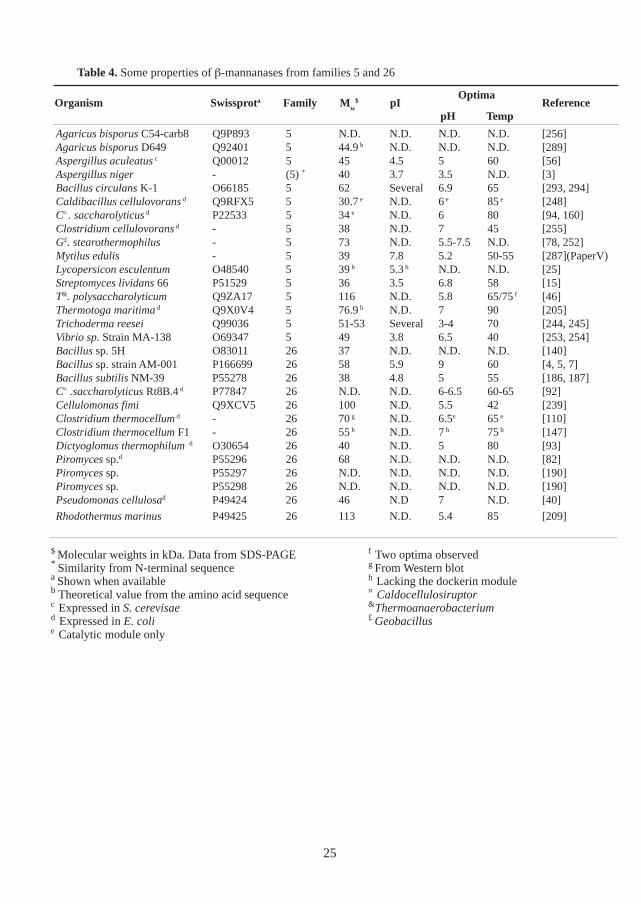
Table 4. Some properties of β-mannanases from families 5 and 26
pH Temp
Agaricus bisporus C54-carb8 Q9P893 5 N.D. N.D. N.D. N.D. [256]Agaricus bisporus D649 Q92401 5 44.9 b N.D. N.D. N.D. [289]Aspergillus aculeatus c Q00012 5 45 4.5 5 60 [56]Aspergillus niger - (5) * 40 3.7 3.5 N.D. [3]Bacillus circulans K-1 O66185 5 62 Several 6.9 65 [293, 294]Caldibacillus cellulovorans d Q9RFX5 5 30.7 e N.D. 6 e 85 e [248]C¤ . saccharolyticus d P22533 5 34 e N.D. 6 80 [94, 160]Clostridium cellulovorans d - 5 38 N.D. 7 45 [255]G£. stearothermophilus - 5 73 N.D. 5.5-7.5 N.D. [78, 252]Mytilus edulis - 5 39 7.8 5.2 50-55 [287](PaperV)Lycopersicon esculentum O48540 5 39 b 5.3 b N.D. N.D. [25]Streptomyces lividans 66 P51529 5 36 3.5 6.8 58 [15]T&. polysaccharolyticum Q9ZA17 5 116 N.D. 5.8 65/75 f [46]Thermotoga maritima d Q9X0V4 5 76.9 b N.D. 7 90 [205]Trichoderma reesei Q99036 5 51-53 Several 3-4 70 [244, 245]Vibrio sp. Strain MA-138 O69347 5 49 3.8 6.5 40 [253, 254]Bacillus sp. 5H O83011 26 37 N.D. N.D. N.D. [140]Bacillus sp. strain AM-001 P166699 26 58 5.9 9 60 [4, 5, 7]Bacillus subtilis NM-39 P55278 26 38 4.8 5 55 [186, 187]C¤ .saccharolyticus Rt8B.4 d P77847 26 N.D. N.D. 6-6.5 60-65 [92]Cellulomonas fimi Q9XCV5 26 100 N.D. 5.5 42 [239]Clostridium thermocellum d - 26 70 g N.D. 6.5e 65 e [110]Clostridium thermocellum F1 - 26 55 h N.D. 7 h 75 h [147]Dictyoglomus thermophilum d O30654 26 40 N.D. 5 80 [93]Piromyces sp.d P55296 26 68 N.D. N.D. N.D. [82]Piromyces sp. P55297 26 N.D. N.D. N.D. N.D. [190]Piromyces sp. P55298 26 N.D. N.D. N.D. N.D. [190]Pseudomonas cellulosad P49424 26 46 N.D 7 N.D. [40]
Rhodothermus marinus P49425 26 113 N.D. 5.4 85 [209]
Organism Swissprota Family Mw$ pI Reference
Optima
$ Molecular weights in kDa. Data from SDS-PAGE* Similarity from N-terminal sequencea Shown when availableb Theoretical value from the amino acid sequencec Expressed in S. cerevisaed Expressed in E. colie Catalytic module only
f Two optima observedg From Western bloth Lacking the dockerin module¤ Caldocellulosiruptor&Thermoanaerobacterium£ Geobacillus
25

Occurrence and regulation
β-Mannanases have been isolated from a wide range of organisms, including bacteria [7, 40,239], fungi [3, 56, 245], plants [25, 172] and animals (Paper V) [53]. Amongst bacteria, β-mannanases have been found amongst aerobes, anaerobes and different extremophiles such asthermophiles, halophiles and psycrophiles [205, 274, 296]. Most β-mannanases are extracellular,however some appear to remain attached to the cell [91]. In plants, β-mannanase activity hasbeen correlated with seed germination [77, 199, 200], and in some cases also with fruit ripening[24, 38]. Some β-mannanases from molluscs have been isolated from their digestive tract (PaperV) [85].
Expression of many microbial β-mannanases is induced by growth on mannan or galactomannan[3, 13, 221]. β-Mannanases from some microbes, including T. reesei, have also been produced bygrowth on cellulose [221, 242]. However, expression of the T. reesei β-mannanase is repressed onglucose and several other monosaccharides [170, 228]. In plants, production of β-mannanase hasbeen shown to be regulated by plant hormones, such as gibbrellins and abscisic acid [10, 104]
Family classification and structural determination
All β-mannanases for which the genes have been cloned have been classified into families 5 or 26of glycoside hydrolases [115, 119]. Both of these families are included in the GH-A clan ofglycoside hydrolases [32, 131] and the retaining mechanism has been confirmed for family 5 andfamily 26 β-mannanases [3, 32, 112]. Both bacterial and eukaryotic β-mannanase have beenannotated to family 5. This family also includes endo- and exoglucanases, xylanases andendoglycoceramidases. An alignment of the amino acid sequences of some family 5 β-mannanasesmainly from eukaryotic sources is shown in Figure 8.
With the exception of a few anaerobic fungi, the β-mannanases in family 26 are of bacterialorigin. Besides β-mannanases, some endoglucanases and a few β-1,3-xylanases are also found inthis family. In some cases, β-mannanases from the same genus have been classified in differentfamilies; β-mannanases from different strains of Caldocellulosiruptor saccharolyticus have beenclassified in both families 5 and 26 [92, 94], and those from different Bacillus species are alsofound in both families [5, 187].
26
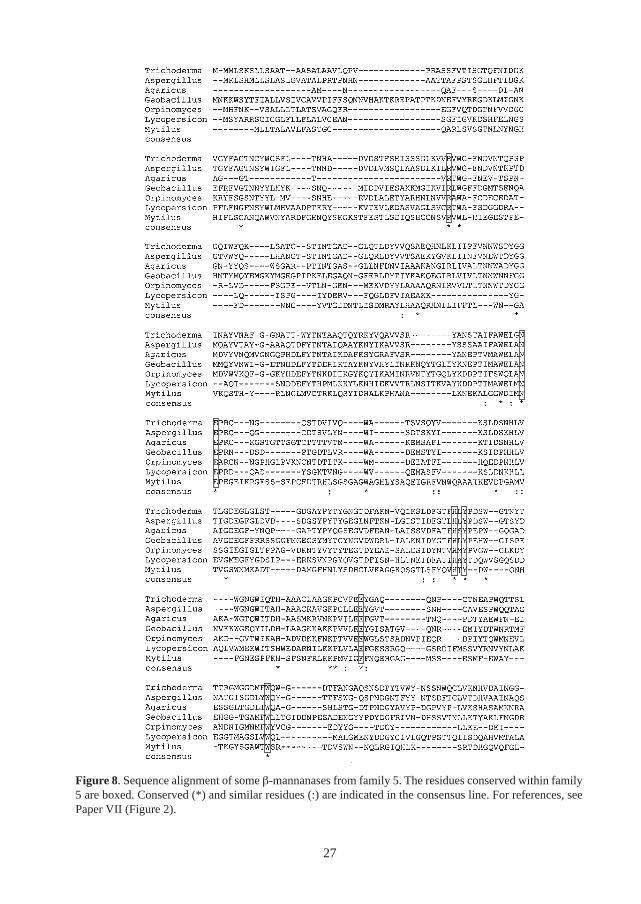
Figure 8. Sequence alignment of some β-mannanases from family 5. The residues conserved within family5 are boxed. Conserved (*) and similar residues (:) are indicated in the consensus line. For references, seePaper VII (Figure 2).
27

The structures of two family 5 β-mannanases – those from T. reesei and T. fusca – have beendetermined by X-ray crystallography [121, 223]. The structures of the T. reesei and T. fusca β-mannanases share the overall (β/α)
8 fold, which is conserved in clan GH-A (Figure 9). Interest-
ingly, the T. reesei β-mannanase also contained two additional β-sheets which are not conservedin other (β/α)
8 enzymes. Furthermore, four glycosylation sites are occupied with N-
acetylglucosamine residues (Figure 9).
The active site in the β-mannanase structures is located in a shallow cleft exposed to the solvent.Within the active site, at least seven amino acids conserved in other family 5 enzymes (Figure 8)are present. In the T. reesei β-mannanase these correspond to Arg54, Asn168, Glu169, His241,Tyr243, Glu276 and Trp306 [223, 244]. Structures of the T. fusca and T. reesei enzymes witholigosaccharides bound their active site clefts have also been solved [121, 223]. These structuresreveal the presence of several substrate-enzyme interactions in the -2 and -3 (T. fusca) and +1 and+2 (T. reesei) subsites. The structure of the family 26 β-mannanase ManA from Pseudomonascellulosa has also been solved recently [124]. As expected, this β-mannanase also exhibits the (β/α)
8 fold and the conserved positions of the catalytic residues in clan GH-A. Very recently, the
crystallisation of the family 5 β-mannanase (studied in Paper V) from blue mussel was alsoreported [286].
Figure 9. The structure of the catalytic module of family 5 β-mannanase (TrMan5A) from T. reesei [223]. α-Helices are shown in gold, β-sheets in light blue. The predicted acid/base and nucleophile residues areshown in red and blue, respectively. The four N-acetylglucosamine residues are shown in green.
28

Modular β-mannanases
Several β-mannanases display a modular architecture (Figure 10). Amongst β-mannanases fromaerobic fungi, the enzymes from T. reesei and A. bisporus are composed of a family 5 catalyticmodule linked to a family I CBM [244, 256, 289]. Some bacterial β-mannanases from families 5and 26 have more complex structures: the C. fimi β-mannanase Man26A contains both a mannan-binding family XXIII CBM, a putative SLH-module and a module of unknown function [237,239, 240]. The family 5 β-mannanase from Thermoanaerobacterium polysaccharolyticum alsocontain an SLH-module and, in addition, two internal family XVI CBMs [46]. Several β-mannanases from anaerobic bacteria and fungi contain dockerin modules which attach the β-mannanases to multienzyme complexes [82, 110, 190, 255]. A family 26 β-mannanase fromCaldocellulosiruptor saccharolyticus contains two family XXVII mannan-binding modules [247].
Figure 10. A schematic picture of the modular organisation in some β-mannananses. Catalytic modules areshown as boxes, CBMs as ellipses. Other modules are shown as pentagons. The β-mannanases are thosefrom Trichoderma reesei [244], Agaricus bisporus [256], Cellulomonas fimi [239] Thermoanaerobacteriumpolysaccharolyticum [46], Caldocellulosiruptor saccharolyticus [247] and Caldibacillus cellulovorans [248].
29

Applications
β-Mannanases have several existing and potential industrial applications. They have been shownto be effective in increasing the brightness of pulps in bleaching experiments [45, 192, 249, 280],most notably in combination with xylanases [45, 58]. In the food and feed industries, β-mannanasesare used in the production of fruit juices and soluble coffee [102, 108, 224], and also in thepreparation of poultry diets [128]. β-Mannanases have also been shown to have a strong potentialas viscosity reducers of hydraulic fracturing fluids used in oil and gas production [183]. Further-more, β-mannanases have potential applications in recycling of copra and coffee wastes [216].
Exo-acting enzymes
β-Mannosidase
β-Mannosidase (EC 3.2.1.25; β-1,4-D-mannoside mannohydrolase) catalyses the hydrolysis ofmannose units from the non-reducing end of mannosides. However, some β-mannosidases areactive both on glucosides and mannosides [21, 80]. The most commonly employed substrate foranalysis of β-mannosidase activity is a chromogenic monosaccharide. In addition, several β-mannosidases are also capable of degrading longer manno-oligosaccharides, with DP over 4 [6,12, 113]. However, only a few β-mannosidases have been shown to release mannose from thenon-reducing end of mannan-based polymers [14, 122, 146] (Papers I, III and VI).
β-Mannosidases have been isolated from widely different types of organisms, including eubacteria,archaebacteria, plants, fungi and animals [12, 21, 53, 176, 239]. β-mannosidase appears to carryout different functions, depending on the producing organism. β-Mannosidases from microbesare often employed in the degradation of mannans and heteromannanas from decaying plant ma-terial for nutritional purposes. β-Mannosidases in plants are involved in the release of storagepolysaccharides in the seed endosperm during germination [178]. In contrast, mammalian β-mannosidases appear to function mainly as lysosomal enzymes involved in degradation of pro-tein-linked glycans. β-Mannosidosis is a congenital disorder, which results from the lack of afunctional β-mannosidase activity. This disease was first found in ruminants, but has more re-cently also been described in humans [134].
Despite their functional difference, many β-mannosidases are related to each other and are classi-fied in family 2 of glycoside hydrolases, which is included in the GH-A clan [115, 119, 131]. Themolecular weights of most β-mannosidases, as determined from SDS-PAGE, are in the range 50-130 kDa (Table 5). However, some β-mannosidases appear to consist of several subunits [21,204]. The isoelectric points of most β-mannosidases are in the acidic range, except for somebacterial enzymes which have isoelectric points near neutrality (Table 5). Most β-mannosidasesshow maximal activity at acidic or neutral pH, and with the exception of some thermophilicenzymes, most β-mannosidases show their maximal activity in the temperature range 40-70 °C.
No three-dimensional structure of a β-mannosidase has been determined at this stage although thecrystallisation of a T. reesei β-mannosidase has been reported [11]. However, the structures of two
30
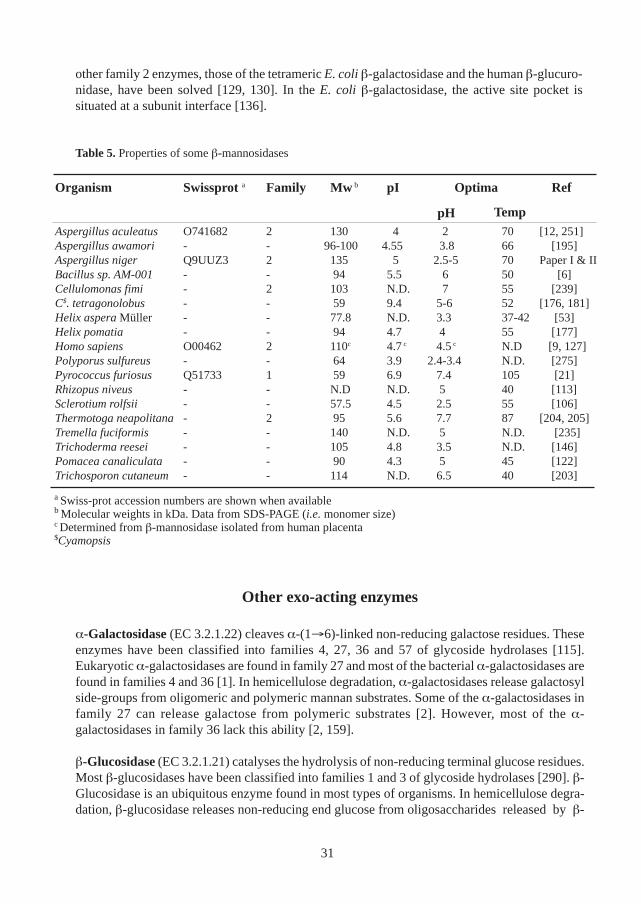
Table 5. Properties of some β-mannosidases
Other exo-acting enzymes
α-Galactosidase (EC 3.2.1.22) cleaves α-(1→6)-linked non-reducing galactose residues. Theseenzymes have been classified into families 4, 27, 36 and 57 of glycoside hydrolases [115].Eukaryotic α-galactosidases are found in family 27 and most of the bacterial α-galactosidases arefound in families 4 and 36 [1]. In hemicellulose degradation, α-galactosidases release galactosylside-groups from oligomeric and polymeric mannan substrates. Some of the α-galactosidases infamily 27 can release galactose from polymeric substrates [2]. However, most of the α-galactosidases in family 36 lack this ability [2, 159].
β-Glucosidase (EC 3.2.1.21) catalyses the hydrolysis of non-reducing terminal glucose residues.Most β-glucosidases have been classified into families 1 and 3 of glycoside hydrolases [290]. β-Glucosidase is an ubiquitous enzyme found in most types of organisms. In hemicellulose degra-dation, β-glucosidase releases non-reducing end glucose from oligosaccharides released by β-
other family 2 enzymes, those of the tetrameric E. coli β-galactosidase and the human β-glucuro-nidase, have been solved [129, 130]. In the E. coli β-galactosidase, the active site pocket issituated at a subunit interface [136].
Organism Swissprot a Family Mw b pI Optima Ref
pHAspergillus aculeatus O741682 2 130 4 2 70 [12, 251]Aspergillus awamori - - 96-100 4.55 3.8 66 [195]Aspergillus niger Q9UUZ3 2 135 5 2.5-5 70 Paper I & IIBacillus sp. AM-001 - - 94 5.5 6 50 [6]Cellulomonas fimi - 2 103 N.D. 7 55 [239]C$. tetragonolobus - - 59 9.4 5-6 52 [176, 181]Helix aspera Müller - - 77.8 N.D. 3.3 37-42 [53]Helix pomatia - - 94 4.7 4 55 [177]Homo sapiens O00462 2 110c 4.7 c 4.5 c N.D [9, 127]Polyporus sulfureus - - 64 3.9 2.4-3.4 N.D. [275]Pyrococcus furiosus Q51733 1 59 6.9 7.4 105 [21]Rhizopus niveus - - N.D N.D. 5 40 [113]Sclerotium rolfsii - - 57.5 4.5 2.5 55 [106]Thermotoga neapolitana- 2 95 5.6 7.7 87 [204, 205]Tremella fuciformis - - 140 N.D. 5 N.D. [235]Trichoderma reesei - - 105 4.8 3.5 N.D. [146]Pomacea canaliculata - - 90 4.3 5 45 [122]Trichosporon cutaneum - - 114 N.D. 6.5 40 [203]
Temp
a Swiss-prot accession numbers are shown when availableb Molecular weights in kDa. Data from SDS-PAGE (i.e. monomer size)c Determined from β-mannosidase isolated from human placenta$Cyamopsis
31

mannanase. In a few cases it has been shown to release glucose residues from polymericglucomannan (Paper VI). β-Glucosidase is also important in the degradation of cellulose; it de-grades cellobiose released by cellobiohydrolase and endoglucanase
In addition to glycoside hydrolases, acetyl esterases also participate in the degradation ofacetylated heteromannans. Acetyl esterase catalyses the hydrolysis of acetyl groups from varioussubstrates. In the context of hemicellulose degradation, most studies on deacetylation have beenconducted with acetylxylan esterase [26]. However, a few examples of acetyl esterases active onacetylated mannans have been reported [210, 260, 261].
32

Present investigation
Outline
In this text, the results from Papers I-VII of the thesis are presented and discussed (see List ofpapers, page 5). The text is divided into four parts:
In the first part (Mannan-degrading enzymes), the major enzymes studied in this work arepresented. The enzymatic characterisation (Paper I) and cloning (Paper II) of a β-mannosidase(AnMan2A) is presented. Next, the characterisation of a β-mannanase (MeMan5A) from bluemussel is described (Paper V). After this, the modular organisation of the β-mannanase (TrMan5A)from T. reesei is presented (Papers IV and VII). Finally, the expression of TrMan5A in T. reesei(Paper IV) and Pichia pastoris (Paper VII) is described.
In the second part of the text (The CBM of TrMan5A) , the characterisation of the CBM fromTrMan5A is presented (Paper IV). In this work, the binding properties of the CBM was studiedand its influence on the catalytic performance of this enzyme was investigated. Furthermore thepossible role of cellulose-binding modules in β-mannanases is discussed.
The third part (The catalytic module of TrMan5A) describes the mutagenesis of specific aminoacids in the active site cleft of the TrMan5A catalytic module (Paper VII).
In the fourth part (Mannan-hydrolysis), the specificities in mannan-hydrolysis of severalhemicellulases are discussed (Papers I, III, VI, V and VI). The hydrolysis of mannan andheteromannans by AnMan2A is presented (Papers I and VI). In addition, the hydrolysis of crys-talline mannan by β-mannanase and β-mannosidase is described (Paper III). Finally, the degrada-tion of different heteromannans by hemicellulases and cellulases is compared (Paper VI).
33

Mannan-degrading enzymes
Aspergillus niger β-mannosidase, AnMan2A
Aspergillus niger produces several enzymes involved in hemicellulose degradation [1, 86]. Inthis work a β-mannosidase isolated from A. niger was purified and characterised (Paper I). Inaccordance with the recommended nomenclature for glycoside hydrolases [120], this enzyme isreferred to as AnMan2A (it has earlier been called Mnd2A). The investigation of the molecularorganisation and the enzymatic specificity of AnMan2A which has been conducted in this work(Papers I, II, III and VI) reveals some interesting features which are discussed in this section andlater (see pages 45, 46 and 48).
In this work, AnMan2A was purified to homogeneity in three steps (Paper I). The molecularweight of the purified enzyme was analysed by gel filtration and SDS-PAGE. Interestingly, theresults suggest that the β-mannosidase is a homodimer. This is an unusual observation since mostother β-mannosidases analysed to date are monomeric. Only in a few previous cases have oligo-meric β-mannosidases been described [21, 204]. One example is the tetrameric β-mannosidasefrom Pyrococcus furiosus [21].
Furthermore, an investigation of the enzymatic properties of AnMan2A showed that this enzymeis able to degrade manno-oligosaccharides (Paper I). The standard substrate used in many β-mannosidase studies is a small substrate composed of a chromophore linked to a mannosyl resi-due. In this work we showed that oligosaccharides up to a DP of 6 are degraded by AnMan2A(Paper I). An assessment of the activity of AnMan2A on longer saccharides was not made. How-ever, as will be described later (see page 45) AnMan2A was also active on polymeric mannansubstrates. Thus, it can be speculated that AnMan2A probably also degrades longeroligosaccharides. AnMan2A was also active on galactosyl-substituted manno-oligosaccharides(Papers I and II). However, galactosyl groups appear to pose some restrictions on AnMan2A,since hydrolysis is blocked at the point of the first substituent. Degradation of oligosaccharidesabove DP 3 and galactosyl substituted substrates has previously only been reported for a few β-mannosidases [6, 12, 113, 240].
Further in this work, the gene encoding AnMan2A was cloned (Paper II). Most β-mannosidasegenes analysed previously have been isolated from mammals (Bos taurus, Mus musculus, Homosapiens, Capra hircus) and bacteria (C. fimi, Thermotoga maritima, Thermotoga neapolitana) [9,22, 54, 148, 205, 239]. Apart from the β-mannosidase from the archaeon Pyrococcus furiosuswhich has been assigned to family 1 [21], all β-mannosidases with known sequences have beenclassified into family 2. In the present case, AnMan2A was assigned to family 2 of glycosidehydrolases on the basis of sequence analysis (Paper II). Besides man2A, the only previouslydescribed gene encoding a fungal β-mannosidase was isolated from Aspergillus aculeatus [251].
The catalytic acid and nucleophile of AnMan2A were predicted to be Glu479 and Glu584, re-spectively (Paper II). The only experimentally identified catalytic residue among β-mannosidasesis the catalytic nucleophile (Glu519) in the β-mannosidase from C. fimi, which was identified bymass-spectrometry using a mannosyl fluoride inhibitor [238]. Analysis of the AnMan2A sequence
34

also revealed 13 putative N-glycosylation sites (Paper II). The presence of N-linked glycans wasalso indicated by enzymatic deglycosylation of AnMan2A, which yielded a decrease in molecu-lar weight of the enzyme (Paper I).
The M. edulis β-mannanase, MeMan5A
Both in terms of species and individuals, the molluscs (Phylum Mollusca) constitute one of thelargest groups of animals and display a wide diversity in terms of feeding habits [65]. Filter-feeding mussels (Polysyringia) are a group of molluscs which thrive on microscopic algae anddissolved organic matter. As part of their digestive system, these organisms produce digestiveenzymes which are secreted into the stomach from a style sac [212]. Several polysaccharide-degrading enzymes, including cellulases and amylases, have been found in the digestive tract offilter-feeding mussels [212], but there has been no previous report on β-mannanases in theseorganisms. In this work (Paper V), β-mannanase from the common blue mussel (Mytilus edulis)was purified and characterised. Two β-mannanase variants with approximately similar molecularweights and isoelectric points were identified. Both variants also had similar pH and temperatureoptima (Paper V). Furthermore, the N-terminal sequences of both variants were identical andshowed significant similarity to two unclassified β-mannanases from the molluscs Littorinabrevicula and Pomacea insularus [291, 292].
Very recently the corresponding β-mannanase gene was cloned [287]. Sequence analysis revealedsimilarities to family 5 of glycoside hydrolases and the encoded gene product will hereafter bereferred to as MeMan5A. Furthermore, since the gene was isolated from a tissue separate fromthe digestive tract, possible contaminations from organisms inhabiting these organs were avoidedand unequivocal proofs of the endogenous nature of the β-mannanase was gained. Variants of β-mannanases apparently encoded by one gene have been observed in several other organisms,including T. reesei [244]. For other organisms different β-mannanases appear to be encoded byseveral genes [172, 190].
The hydrolysis of manno-oligosaccharides by MeMan5A was analysed in this work (Paper V).MeMan5A showed no activity on manno-oligosaccharides of DP up to 3. Moreover, mannotetraosewas degraded at a lower rate than mannopentaose. Thus, it was concluded that binding in at leastfive subsites is probably required for optimal activity. It was recently reported by others that crystalsof this β-mannanase has been obtained [286]. A 3D structure of this enzyme will possibly providesome information about the specific enzyme-polysaccharide interactions in the active site cleft.
Mannan-degrading enzymes have previously been found in several other types of molluscs. Amajority of these have been isolated from snails [53, 85, 122, 177, 246]. Due to the variability ofmolluscs in terms of nutrition, it is tempting to believe that these enzymes are used for differentpurposes. In the case of M. edulis it could be speculated that the β-mannanase participate in thedegradation of mannan from algal cell walls [88, 156, 208].
The modular T. reesei β-mannanase, TrMan5A
The filamentous fungus T. reesei is a potent producer of hemicellulose-degrading enzymes in-volved in the degradation of different mannans and xylans [26]. One of the major hemicellulases
35

produced by this fungus is a β-mannanase (TrMan5A), which is expressed in media containingcellulose and galactomannan [221, 242]. Previously it was shown that several β-mannanaseisoforms with different molecular weights and isoelectric points are produced by T. reesei [245].Later, a β-mannanase encoding gene (man1) was isolated from T. reesei and it was concluded thatthe different isoforms are products of the same gene [242, 244]. Thus, it was suggested that theisoforms are likely to differ mainly in their patterns of glycosylation or other types of post-trans-lational modifications. Analysis of the β-mannanase sequence suggested that it is a modular enzyme comprised of a N-terminal catalytic module and a C-terminal CBM connected by a Ser/Thr/Pro rich linker sequence. On the basis of sequence alignment and hydrophobic cluster analy-sis the catalytic module was classified into family 5 of glycosyl hydrolases according to theclassification by Henrissat et al [115]. The C-terminal module showed extensive sequence simi-larity with CBMs from T. reesei cellulases and was classified [244] into family I according to theclassification of Tomme et al. [269].
TrMan5A appears to be specific for mannosidic linkages [244, 245]. Furthermore, in the presentwork, no hydrolysis was observed when TrMan5A was incubated with cellulose or cello-oligosaccharides (unpublished results).
In this study (Papers III, IV, VI and VII), the functions of the two modules of the T. reesei β-mannanase TrMan5A were investigated. In order to study the properties of the modules, a mutantlacking the C-terminal CBM (TrMan5A∆CBM), was constructed, and expressed in T. reesei asdescribed below (Paper IV). The full-length enzyme was also expressed under the same condi-tions and both enzymes were purified to electrophoretic homogeneity. Furthermore, several mu-tants of specific amino acids in the active site of TrMan5A∆CBM were expressed in Pichiapastoris and characterised, along with the wild type TrMan5A∆CBM (Paper VII).
Homologous expression in T. reesei. As stated earlier, T. reesei is a very efficient producer ofsecreted enzymes and was thus an attractive candidate as a host for gene expression of TrMan5A.Furthermore, since T. reesei is the native host of TrMan5A, correct processing of the enzyme ismore likely. Since T. reesei is utilised as a host for heterologous expression [206], several methodsfor transformation of foreign genes into T. reesei have been developed [164], which can be em-ployed also for the expression of homologous proteins. The main problem posed for expression ofmutated homologous enzymes, such as TrMan5A∆CBM, is to avoid the expression of the endog-enous chromosomal gene. However, it has been shown that β-mannanase expression in T. reesei isrepressed when it is grown in a medium with glucose as the sole carbon source [170]. Furthermore,it is known that genes under the control of the Aspergillus nidulans glyceraldehyde phosphate dehy-drogenase (gpdA) promoter are constitutively expressed in the presence of glucose [211].
Thus, in this work, the gpdA promoter was utilised for expression of mutants of T. reesei β-mannanase (Paper IV), as has been done previously with other glucose-repressed enzymes [59,207]. The expression of TrMan5A and TrMan5A∆CBM in T. reesei under the control of the gpdApromoter was successful and enzyme yields of approximately 5 mg/ml were obtained (Paper IV).Furthermore, the culture filtrate was composed of a limited number of proteins and protein puri-fication was thus facilitated. Pure enzyme preparations were isolated in one or sometimes twochromatographic steps.
Expression in Pichia pastoris: Traditionally, Saccharomyces cerevisae has been the modelorganism for yeast expression and also used for heterologous expression. In this specific case,
36

TrMan5A was previously expressed in S. cerevisae but the enzyme yield was very low [244]. Themolecular weight of TrMan5A expressed in S. cerevisae was higher (75 kDa) than that of the T.reesei expressed enzyme (52-54 kDa) and it was suggested that the enzyme was likely to beoverglycosylated [242]. Expression of a β-mannanase from Aspergillus aculeatus also resulted ina higher molecular weight compared to the enzyme expressed in the wild-type organism [229].Expression in the methylotrophic yeast P. pastoris shares many of the advantages of S. cerevisae,such as high expression yields of extracellular enzymes and simple techniques for moleculargenetic manipulations. Furthermore, P. pastoris has been reported to attach glycans of lowermolecular weight than those found in S. cerevisae [103].
In the present case, an expression system which utilises the alcohol oxidase promoters in P. pastoriswas used. This promoter is strongly induced when the organisms are grown on methanol as solecarbon source. In this work, TrMan5A∆CBM and a number of mutants thereof were expressedand secreted by P. pastoris using the native signal sequence from the T. reesei enzyme (PaperVII). The expression levels were approximately 10 mg/ml and pure enzyme preparations wereachieved after two chromatographic steps
The CBM of TrMan5A
Sequence and structure
Family I CBMs are common among polysaccharide-degrading enzymes from T. reesei and otherfungi. A majority of the T. reesei cellulases carry family I CBMs either at their N- or C-termini[155, 257]. Amongst T. reesei hemicellulases, family I CBMs have been found only in TrMan5A[244] and an acetyl xylan esterase [171]. However, family I CBMs have also been found in somexylanases from Humicola insolens [68] and in β-mannanases from Agaricus bisporus [256, 289].The C-terminal family I CBM of TrMan5A is proposed to be composed of 37 amino acids [242].Figure 11 shows a sequence alignment of the CBM from TrMan5A with several other family ICBMs derived from T. reesei endoglucanases and cellobiohydrolases. The three aromatic residueswhich are exposed on the flat binding surface in the structure of TrCel7A (Figure 12a) are con-served in this alignment – in TrMan5A these residues correspond to Tyr378, Trp403 and Tyr404.The importance of these conserved aromatic residues in several family I CBMs linked to T. reeseicellulases has been extensively studied by site-directed mutagenesis [151-153, 218, 219].
Figure 11. Sequence alignment of family I CBMs from TrMan5A and a number of T. reesei cellulases. Thethree conserved aromatic residues are boxed. Identical residues are also indicated (¤). The numbers of theamino acids at the N- and C-termini are shown. Adapted from [244].
37

Based on the structure of the CBM from TrCel7A [145], the structures of the CBMs of the T.reesei cellulases TrCel6A, TrCel7B, TrCel45A and TrCel5A were studied by molecular dynamicsand were predicted to display a relatively similar structure [123]. Subsequently, the structure ofthe CBM from TrCel7B was solved and the similarity with the CBM from TrCel7A was validated[174]. In the current study, a model of the CBM from TrMan5A was generated using the programSwiss-PdbViewer [105]. The modelling was based on the structure of the CBM from TrCel7A[145]. As seen in Figure 12b, the flat surface from the CBM of TrCel7A is preserved in thismodel. Interestingly, two bulky tyrosines protrude from the surface on the rough side of the CBMmodel. One of these tyrosines (Tyr399) is present in a roughly similar position in the TrCel7Bstructure [174]. The other tyrosine (Tyr402) appears to be unique for TrMan5A as no counterpartcould be found in an extensive sequence alignment [222]. However, it should be pointed out thatthis is a preliminary model. The positions of these two residues and their possible involvement inpolysaccharide binding need to be further studied.
Binding properties of the CBM
Binding to cellulose and kraft fibers with TrMan5A has been observed previously [3, 90, 258],and it was postulated that this binding is mediated by the C-terminal CBM [242]. In this work, thequantitative and qualitative binding properties of the CBM were studied (Paper IV). TrMan5Aand TrMan5A∆CBM were incubated with cellulose and the amount of free enzyme was ana-lysed. Only TrMan5A adsorbed significantly to cellulose and it could thus be inferred that thebinding of TrMan5A to cellulose is indeed mediated by its C-terminal CBM.
The binding of TrMan5A to cellulose was also visualised by immuno-gold labelling (Figure 13).Again, TrMan5A and TrMan5A∆CBM were incubated with cellulose. After several washes andincubations with primary antibodies and protein A conjugated to gold-particles, the cellulosesamples were analysed by transmission electron microscopy. As seen in Figure 13, cellulosemicrofibrils incubated with TrMan5A were more strongly labelled in comparison with those in-cubated with TrMan5A∆CBM.
38

Figure 12. a. Structure of the CBM from TrCel7A. b. Model of the CBM from TrMan5A, based on thestructure of the CBM from TrCel7A. The numbers of the aromatic residues on the flat sides of the CBMfrom TrCel7A and the model of the TrMan5A CBM are indicated. The numbers of the tyrosines on therough side of the model of the CBM from TrMan5A are also indicated.
39
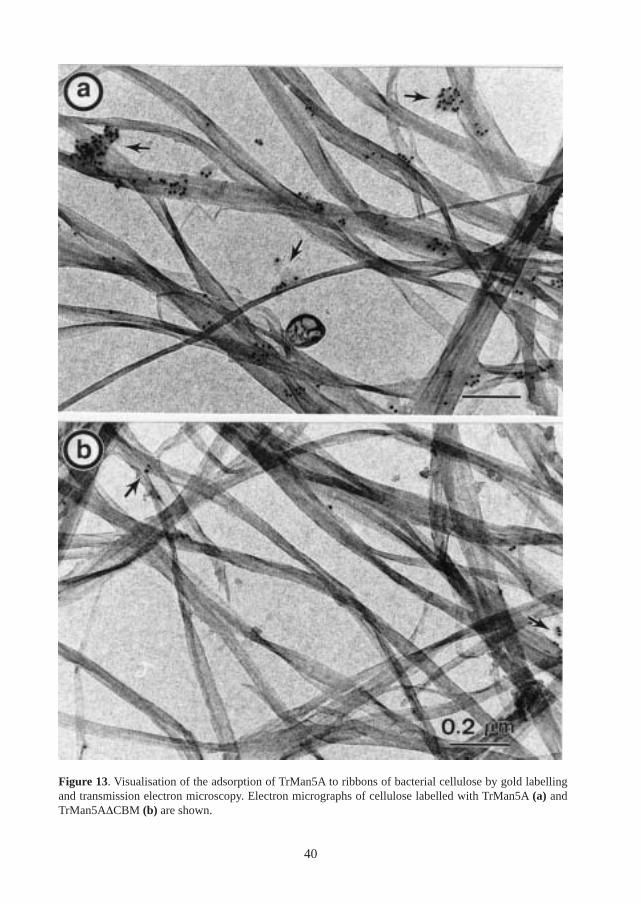
Figure 13. Visualisation of the adsorption of TrMan5A to ribbons of bacterial cellulose by gold labellingand transmission electron microscopy. Electron micrographs of cellulose labelled with TrMan5A (a) andTrMan5A∆CBM (b) are shown.
40

A quantitative comparison of cellulose binding with a T. reesei cellulase carrying a family I CBM,TrCel7B, was made (Paper IV, Figure 4). Analysis of the binding permitted an estimation of therelative binding constant (K
r) for the two enzymes. The results showed that the values of this
constant were fairly similar with TrCel7B (1.05) and TrMan5A (0.84). This similarity indicatesthat the C-terminal CBM is a true cellulose-binding CBM. Thus, it can be suggested that TrMan5Ashares some of the properties which have been observed in other family I CBMs.
An investigation of the binding of TrMan5A toward mannan-based polysaccharides was alsoconducted (Paper IV). Earlier, a Clostridium thermocellum β-mannanase was shown to bind in-soluble mannan [110]. Furthermore, the β-mannanase Man26A from Cellulomonas fimi and a β-mannanase from Caldicellulosiruptor saccharolyticus were shown to bind to soluble galactomannanvia mannan-binding modules from families XXIII and XXVII, respectively [237, 247]. In con-trast, no binding of TrMan5A to insoluble mannan, soluble galactomannan and crystalline mannanwas detected in the current work (Paper IV). However, TrMan5A did bind to a mannan samplewhich contained some cellulose (Paper IV, Figure 6b). This mannan/cellulose complex had beenisolated from ivory nut and contained approximately 15 % cellulose. Further binding analysiswith an in vitro mixture of mannan and cellulose, suggested that TrMan5A binds to the cellulosecomponent in this mannan/cellulose complex (Paper IV).
Effect on hydrolysis
In cellulose degradation, cellulose-binding CBMs are very important for efficient hydrolysis [98,109, 267, 276]. In particular, several family I CBMs have been shown to increase the activitytowards insoluble cellulose substrates [155, 257]. The effect of hemicellulase-linked CBMs hasbeen investigated only to a limited extent and most studies have been related to xylan degrada-tion. The presence of cellulose-binding CBMs have been shown to influence the hydrolysis ofcomplex cellulose/xylan substrates by a xylanase and an arabinofuranosidase from P. cellulosa[28, 29, 99]. Also, xylan-binding CBMs from family VI and XXII have been shown to potentiatethe hydrolysis of insoluble but not soluble xylan by xylanases from Clostridium thermocellum[52, 84]. Amongst β-mannanases, the mannan-binding CBM of Man26A from Clostridiumthermocellum has been shown to increase the activity towards insoluble ivory nut mannan [110].Furthermore, hydrolysis of soluble galactomannan by a family 5 β-mannanase fromCaldicellulosiruptor saccharolyticus was reduced in the absence of the two internal family IIICBMs [92].
In the present case, the possible influence of the family I CBM of TrMan5A on the hydrolysis ofmannan substrates was investigated by comparing the catalytic activity of TrMan5A andTrMan5A∆CBM on a range of mannan-containing substrates (Paper IV). Analysis of the hydroly-sis of ivory nut mannan and galactomannan showed that the CBM had no effect on degradation ofeither insoluble or soluble mannan. On the contrary, in the hydrolysis of a mannan/cellulose com-plex, an increased hydrolysis rate was observed for TrMan5A in comparison with TrMan5A∆CBM(Paper IV, Figure 5b). Thus, it was concluded that the CBM has a positive influence on the degra-dation of this substrate under the conditions used.
Interestingly, the results from these hydrolysis experiments fit well with the data from the bindingexperiments: binding of the CBM to the mannan/cellulose complex could be correlated to the
41

increase in activity towards this substrate (Paper IV, Figures 5b and 6b). In the cases where nobinding was detected, there was no influence on hydrolysis (Paper IV, Figures 5a and 6a). Thelack of influence on hydrolysis of soluble substrates agrees with what has been observed withfamily I CBMs linked to cellulases where no major influence of the CBMs on hydrolysis ofsoluble cellulose derivatives has been observed [155].
Why a cellulose-binding CBM on a β-mannanase?
In cellulose-degrading enzymes, the presence of cellulose-binding CBMs makes clear sense sincethe CBM binds to the preferred substrate of the enzyme, thus increasing the effective substrateconcentration. The rationale behind the presence of cellulose-binding CBMs in hemicellulose-degrading enzymes is more ambigious. Previous results have indicated that some cellulose-bind-ing CBMs may promote non-hydrolytic disruption of cellulose surfaces [74]. It could be specu-lated that such activities of a CBM in a plant cell wall could reveal previously inaccessible partsof the hemicellulose to mannan-degrading enzymes.
On the contrary, results with some other CBMs have shown that, when isolated, these CBMscannot enhance hydrolysis independently of their catalytic modules [31, 191]. Hence, it has beensuggested that the increased hydrolysis obtained with CBMs is due to an increase in effectivesubstrate concentration [31]. Furthermore, it has been postulated that cellulose functions as ageneral receptor for CBMs of both cellulases and hemicellulases [191]. Since cellulose is a majorcomponent in the plant cell wall it might be an easy available target for CBMs. Moreover, sincecellulose has a uniform chemical composition it is possible that it displays a large number ofsimilar binding sites for CBMs.
Several studies have shown that mannan and cellulose are closely associated in the plant cell wall[227, 297]. Therefore, it could be speculated that the binding to cellulose of the CBM in TrMan5Apositions the catalytic module in the close vicinity of accessible mannan chains. There are someindications that at least some CBMs bind to different sites on cellulose [47]. However, morestudies need to be carried out in order to assess any possible cellulose binding preferences of theCBM from TrMan5A.
It is instructive to compare the modular structure of TrMan5A with those in other β-mannanases.CBMs have been predicted for several β-mannanases from family 5 and family 26 (Figure 10).However, only in a few cases has the binding been experimentally confirmed. In addition toTrMan5A, family I CBMs have been indicated in the fungal β-mannanases from Agaricus bisporus[256, 289]. However, the β-mannanases from Aspergillus aculeatus and A. niger appear to lackcellulose-binding CBMs [3, 56].
Some other family 5 β-mannanases have CBMs which have been classified into family III andXVI (Figure 10). Moreover, some family 26 β-mannanases, have family XXIII and XXVII CBMswhich have been shown to bind to mannan [237, 247]. However, many β-mannanases from bothfamilies 5 and 26 appear to be sole catalytic modules. Thus, at present, it is difficult to establishany obvious patterns in modularity among β-mannanases. It clearly appears as if several differentstrategies in mannan-hydrolysis exists.
42

The catalytic module of TrMan5A
The active site cleft
The specificity of catalytic modules is governed mostly by its interaction with the substrate in theactive site cleft. Thus, an understanding of the function of specific amino acid residues positionedat the active site or in its vicinity, is essential to an in-depth understanding of the function of anenzyme. In order to pinpoint the function of a given amino acid, site-directed mutagenesis is apowerful method which allows for comparative analyses with the wild-type enzyme. In the presentstudy, a number of mutants (listed in Paper VII, Table 1) of specific amino acids in the active siteof the catalytic module of TrMan5A were designed (Paper VII). The basis for the choices ofmutants was the solved structure of the catalytic module of TrMan5A, in complex with mannobiose[223].
Figure 1 (Paper VII) shows a close up of the active site of TrMan5A, highlighting the catalyticresidues and one of the residues involved in mannobiose binding. As predicted from sequenceanalysis [244], the catalytic residues (Glu169 and Glu276) are positioned in the active site in thepositions conserved within family 5 of glycoside hydrolases [71, 76, 121]. Also shown in Figure1 (Paper of VII), is the mannobiose molecule positioned in subsites +1 and +2 [223]. Interest-ingly, the mannose residue in subsite +2 is predicted to hydrogen bond to an arginine residue(Arg171) [223]. The donor in this hydrogen bond is the C-2 hydroxyl of the mannose residue. Theconformation of the C-2 hydroxyl (which is axial in mannose) is the only conformatory differ-ence from its C-2 epimer – glucose (in which the C-2 hydroxyl is equatorial). Thus, the hydrogenbond between Arg171 and the mannose residue in the +2 subsite, can potentially be an interactionwhich is important in determining mannan specificity.
Previous kinetic studies on oligosaccharide hydrolysis with TrMan5A has indicated that the +2subsite is likely to be occupied in substrate binding, at least in the case of saccharides of DPhigher than 3 [112]. Furthermore, analysis of the transglycosylation pattern of TrMan5A indi-cates that the enzyme can produce transglycosylation products of DP n+2 after incubation with asubstrate of DP n=5 [111]. Thus, it can be envisaged that the interaction between mannose in the+2 subsite and Arg171 influences the rate of transglycosylation and potentially also the hydro-lytic activity of the enzyme.
In summary, Arg171 is likely to be involved in substrate binding and was thus a strong candidatefor mutagenesis. In order to maintain the charge balance, a lysine mutant was constructed(Arg171Lys). Furthermore, mutants of the catalytic residues were constructed in order to confirmtheir role in catalysis. The aim was also to construct an inactive mutant for use in crystallisationof an enzyme/substrate complex.
Characterisation of mutants
The analysis of the catalytic rate for the active site mutants constructed in this work (Paper VII)gave interesting results. Comparison with the wild-type enzyme indicated that the Arg171Lys
43

mutant was almost equally active on polymeric galactomannan: kcat
values of 262 and 241 s-1
were obtained for Arg171Lys and the wild-type, respectively (Paper VII, Table 2). However,the apparent K
M value for the mutant (1.3 g/l) was somewhat higher than the corresponding
value for the wild-type (0.59 g/l). Furthermore, a reduction in hydrolysis rate of this mutantwas observed on an oligomeric substrate (mannopentaose). It could be speculated that the cata-lytic activity of the mutant is impaired only on shorter substrates, and that a certain DP higherthan 5 is required for this mutant to show activity in a range close to the wild-type enzyme.Thus, it would be interesting to study the rate of degradation of a range of oligosaccharideswith different DP.
Interestingly, an indication of a shift in the activity optimum of Arg171Lys toward a more alka-line pH was detected (Paper VII). Alkaline conditions are preferred in several industrial processesin which cellulases and hemicellulases can be used [44]. Accordingly, there is an interest to engi-neering these enzymes to increase their pH optima. Several previous studies have focused onmutating charged or polar residues in the close vicinity of the catalytic residues [23, 81]. In thepresent case, the mutated residue is located at the +2 subsite. However, it is also at hydrogen bonddistance to Glu205, which is situated closer to the catalytic acid/base (Paper VII, Figure 1). How-ever, further kinetic studies need to be carried out in order to elucidate the mechanisms behind theobserved activity pH-optimum shift.
The activities of the mutants of the predicted catalytic residues (Glu169 and Glu276) were largelyreduced (Paper VII). No activity at all was detected for these mutants, with the exception of amutant of the acid/base (Glu169Gln) which did display activity, albeit very low (a 25-fold de-crease in k
cat (Paper VII, Table 2)). The low or abolished activities of these mutants support the
postulated involvement of Glu169 and Glu276 in the catalytic mechanism. In accordance withthe current observations, previous site-directed mutagenesis of either the nucleophile or the cata-lytic acid/base residue of glycoside hydrolases has often resulted in a dramatic decrease in activ-ity and frequently no activity was detected [161]. In several family 5 enzymes, the role of cata-lytic residues has been established by site-directed mutagenesis [20, 48, 194, 213] or by massspectrometry, using covalently linked inhibitors [165, 277].
Mutant Glu169Ala
A 3D-structure of an enzyme can provide important information concerning the overall structureof the enzyme and the position of its active site. However, in order to gain knowledge aboutspecific substrate interactions, it is often necessary to achieve a structure of the enzyme in acomplex with a substrate analogue. As stated earlier, TrMan5A has previously been crystallisedwith a substrate in subsites +1 and +2 [223]. In order to get more information about the substrateinteractions in the active site cleft, it would be of interest to crystallise TrMan5A in the presenceof a substrate which covers more subsites. In particular, a structure with a substrate residue boundin the –1 subsite might provide information about the catalytic mechanisms of mannoside hy-drolysis. However, due to the hydrolytic activity of the wild-type enzyme, there is a big risk thatthe substrate is degraded in the crystallisation process. Therefore, it was of interest to construct acatalytically inactive mutant. In this work, one of the mutants (Glu169Ala) of the catalytic acid/base residue which did not display any catalytic activity (Paper VII) was chosen as a candidatefor crystallisation.
44

Mannan-hydrolysis
Degradation of unsubstituted mannan
In this work, the hydrolysis of unsubstituted mannan by different mannoside-hydrolases was inves-tigated (Papers I, IV and V). MeMan5A, TrMan5A and AnMan2A were incubated with mannan andthe amount of hydrolysis was analysed. Mannan was degraded by TrMan5A (Paper IV), MeMan5A(Paper V) and, interestingly also by AnMan2A (Paper I). The mannan used in these studies was theivory nut mannan extracted from seeds of Phytelephas macrocarpa. This mannan has been reportedto have a DP of 20 [184] and was shown in the current work to be free of oligosaccharides of DPlower than 7 (Paper I). AnMan2A was capable of releasing approximately 25 % of the mannoseresidues in ivory nut mannan from the non-reducing end (Paper I). The ability of β-mannosidases todegrade polymeric mannan has been studied only to a limited extent and very few previous reportshave revealed the hydrolysis of mannan by β-mannosidases [14].
The main products formed after extensive hydrolysis of mannan with TrMan5A were mannobioseand mannotriose (Paper IV). This product pattern agrees with previous reports on TrMan5A [245],and other fungal β-mannanases [3, 229]. MeMan5A also degraded mannan to mainly mannobioseand mannotriose, but also produced significant amounts of mannotetraose (Paper V). Productionof mannotetraose has previously been reported after limited hydrolysis of ivory nut mannan witha β-mannanase from A. niger [272, 273].
Crystallisation of mannan
As stated earlier (page 16), two polymorphs of crystalline mannan are found in the native state –mannan I and mannan II [51]. These two crystal forms can also be obtained upon recrystallisationof mannan from solution. Recrystallisation of mannan in either polymorph is dependant on thecrystallisation conditions: a high molecular weight of the polysaccharide, a high polarity of thecrystallisation medium and a low temperature favour the formation of mannan II crystals, and thereverse conditions favour the formation of mannan I [50, 51]. In this study, mannans from ivorynut and Acetabularia crenulata were crystallised as mannan I and mannan II, respectively (PaperIII). The morphologies of mannan I and mannan II crystals are strikingly different. Crystallisa-tion in the mannan I form yields lozenge-shaped, laminated single crystals (Paper III, Figure 1c).Mannan II, on the other hand, shows a ribbon-like morphology (Paper III, Figure 3).
The molecular structure of mannan I has been studied with electron diffraction and X-ray diffrac-tion [19, 49, 88, 197]. Mannan I crystals are composed of anti-parallel sheets of mannan chainspacked with their molecular axes perpendicular to the base plane of the crystal (Figure 14). Themolecular structure of mannan II is less well known. However, based on X-ray diffraction analy-sis a model has been proposed [188, 295], in which the mannan chains are packed anti-parallelwithin sheets which themselves are anti-parallel to neighbouring sheets.
45

Figure 14. Structure of mannan I viewed as a projection of the unit cell onto the ab plane. The unit celldimensions are a=8.92 Å, b=7.21 Å and c=10.27 Å. Adapted from [222].
Degradation of mannan crystals
In the present study (Paper III), crystals of mannan I and II were degraded by TrMan5A,TrMan5A∆CBM and AnMan2A. In addition, mannan I crystals were also degraded by the A.niger β-mannanase (AnMan5A). The enzymatic degradation of the crystals was studied withtransmission electron microscopy, electron diffraction and sugar analysis. The results show thatboth mannan I and mannan II crystals can be degraded with the enzymes used in this study.However, only a limited degradation was seen with AnMan2A, especially in the case of mannanII. Based on the results from these enzymatic degradations and the established structure of mannanI crystals, a model for degradation of mannan I crystals by the mannan-degrading enzymes wasproposed (Paper III, Figure 6). According to this model, the initial sites of degradation by themannan-degrading enzymes are the corners of the crystal lamellae. After the initial degradationof the corners, the endo-acting β-mannanase will continue to hydrolyse the interior of the lamel-lae. β-Mannosidase, on the other hand, will be unable to degrade the crystals after the initialattack on the corner chains. Due to the antiparallel chain packing of the mannan I crystal everyother layer of mannan chains will have its non-reducing end buried in the interior of the crystaland thus is unavailable for attack by β-mannosidase (Paper III, Figure 6).
46

The proposed enzymatic attack at the crystal corners was further supported by immuno-goldlabelling with β-mannanase (Figure 15). TrMan5A and TrMan5A∆CBM were incubated withmannan I crystals and subsequently analysed by transmission electron microscopy. As seen inFigure 15, the labelling appears to be clustered at the crystal corners to some extent.
Figure 15. Visualisation of the adsorption of TrMan5A to mannan I crystals by gold labelling and transmis-sion electron microscopy. Electron micrographs of crystals labelled with TrMan5A (a) and TrMan5A∆CBM(b) are shown.
47

The degradation of mannan II crystals by TrMan5A∆CBM altered the morphology of the crystalsamples (Paper III, Figure 4). After prolonged incubation, the ribbon-like crystallites disappearedand a granular matrix remained. However, the electron diffraction pattern of the degraded sampleis similar to the pattern prior to degradation which is typical of crystalline mannan II. Thus, itseems possible that the obtained granular matrix consists of undegraded mannan II crystals.
Degradation of heteromannans
In this study, the degradation of different types of heteromannans by mannoside-hydrolases wasstudied (Papers I, V, VI and VII). These investigations revealed several interesting features con-cerning the catalytic properties of mannoside-hydrolases, in particular regarding β-mannosidase.AnMan2A was able to liberate mannose from galactomannan, glucomannan andgalactoglucomannan (Paper I and VI). Furthermore, AnMan2A also released acetyl-substitutedmannose from the non-reducing end of O-acetyl-galactoglucomannan (O-acetyl-GGM) (PaperVI). The hydrolysis of heteromannans by β-mannosidase has previously only been studied to avery limited extent [106, 122, 146], and this is the first indication that a β-mannosidase have theability to liberate acetyl-substituted mannose. However, as was described in Paper I, AnMan2A isrestricted by galactosyl substituents.
Degradation of heteromannans also enabled some comparisons between several β-mannanases.In these studies TrMan5A, MeMan5A and the β-mannanase from C. fimi (CfMan26A) were used(Papers V, VI and VII). Hydrolysis of LBG galactomannan by TrMan5A and MeMan5A underassay conditions permitted an estimation of the Michaelis constant (K
M) for the two enzymes
(Paper V and Paper VII). The apparent KM value for LBG obtained with MeMan5A was 3.95 g/l
(Paper V). This was considerably higher than the corresponding value obtained with TrMan5A0.6-0.7 g/l (Paper VII, Table 2). Interestingly, the K
M values for LBG in previous reports on β-
mannanases from molluscs are generally considerably higher than those in reports on fungal β-mannanases. The K
M values for three β-mannanases from snails lie in the range (1.4-3 g/l) [53,
85, 180]. In constrast, reports on fungal β-mannanases from Trichoderma and Aspergillus havegiven values between 0.0015-0.7 g/l [17, 56, 57, 229].
The degradation of glucomannan by CfMan26A and TrMan5A was also compared in the presentstudy (Paper VII, Figure 2). Analysis of the DP of the products formed after prolonged hydrolysisrevealed relatively similar patterns with the two enzymes; in both cases, mannobiose andmannotriose were the major products formed. Slightly more mannotriose was formed withCfMan26A in comparison with TrMan5A. Thus, no major difference in the product formation ofglucomannan degradation was observed under the conditions used. This may indicate that thetwo β-mannanases from families 5 and 26 have similar binding requirements with respect tomannose and glucose residues. However, it must be stated that the products were analysed onlywith respect to their DP. In order to further assess any possible differences/similarities in substratebinding requirements between family 5 and family 26 β-mannanases, a larger number of en-zymes should be included and a more detailed product analysis should be made.
In the context of mannan degradation, investigations on the hydrolysis of isolated glucomannansand galactoglucomannans have previously been conducted, which have provided informationabout the specific enzyme-polysaccharide interactions of β-mannanases. In particular, the subsite
48

specificity of the T. reesei and A. niger β-mannanases has been investigated to some extent [182,259]. In both these enzymes glucose is allowed in the +1 subsite, but not in the -1 site.
In the present study, a comparison of hydrolysis of glucomannan and O-acetyl-GGM by TrMan5Awas made (Paper VII). Analysis of the hydrolysis products showed that the DP profiles weredifferent for O-acetyl-GGM (mainly DP 2-5) and glucomannan (mainly DP 2-3). This indicatesthat some restrictions exist for the β-mannanase with regard to either galactosyl- or acetyl-substituents. The restrictions of galactosyl-substituents for TrMan5A [259] and the A. niger β-mannanase [182], has previously been observed and it has been concluded that galactosesubstituents can be accommodated in the –1 subsite, but not in the +1 subsite [182, 259]. Moreo-ver, Puls et al. [210] demonstrated that acetyl-groups appears to limit hydrolysis ofgalactoglucomannan by the A. niger β-mannanase. However, a more detailed study on the re-strictions of acetyl-substituents on β-mannanase activity has not yet been made. The degradationof O-acetyl-GGM by β-mannanases is important, not only from an enzymologic point of view.Previous work in our group has shown that β-mannanases can be used as tools to analyse thestructure of heteromannans [158].
Interestingly, results from analysis of O-acetyl-GGM hydrolysis also indicated that anendoglucanase (TrCel7B) from T. reesei has a limited depolymerisation activity on this polymer(Paper VII). It has previously been shown that TrCel7B and several other T. reesei cellulases areactive on glucomannan [138]. Furthermore, it has been reported that TrCel7B is active on xylan[27]. In addition, the endoglucanase TrCel5A from T. reesei display activity on galactomannan[163]. These cross-activities of cellulases are interesting in view of the close association betweencellulose and hemicellulose in the plant cell wall. A more detailed product analysis will possiblyreveal more information about specific enzyme-polysaccharide interactions in the degradation ofO-acetyl-GGM by TrCel7B. Of special interest here is to detect the specificity of interactions inthe -1 subsite.
49

50

Conclusions & future perspectives
In this thesis, new results concerning the enzyme-substrate interaction of mannan-degrading en-zymes have been presented and discussed. In particular, novel information has been put forwardregarding the function of a modular β-mannanase (TrMan5A). It was shown that the CBM ofTrMan5A binds to cellulose and potentiate the hydrolysis of complex substrates. Thus, the substratepreference of this enzyme is influenced by interactions in both modules. An interesting fact isthat, in the present case, the binding of the CBM is not directed to the mannan-substrate per se.However, these results may be a reflection of the tight and complex organisation of cellulose andhemicellulose in the plant cell wall.
This observation opens up new questions regarding the role of CBMs in β-mannanases and otherhemicellulases. It would be interesting to investigate if the cellulose-binding CBMs in theseenzymes bind to cellulose at specific positions where the cellulose and hemicellulose are closelyassociated. It would also be interesting to compare the T. reesei β-mannanase with other modularβ-mannanases and other hemicellulases. CfMan26A, which carries a family XXIII mannan-bindingCBM, is an example of an interesting candidate for such a study.
The results presented here concerning the roles of specific amino acid in a β-mannanase is some-what unique in the area of β-mannanases. Few studies of this kind have been carried out withthese enzymes. The results presented here have given some hints about the influence of Arg171,positioned in the +2 subsite, on hydrolysis. Therefore, it would be interesting to study this residueand further assess its role in substrate recognition. Furthermore, a new complex-structure with aninactive mutant will potentially provide new information about the specific enzyme/carbohy-drate interactions in the active site cleft.
This work has also included studies on mannan hydrolysis by different mannan-degrading en-zymes. The investigation of degradation patterns from hydrolysis of different heteromannans hasprovided new insights into the specificity of different enzymes on these substrates. Interestingly,a β-mannosidase was capable of degrading both mannans and heteromannans. A limited amountof degradation of mannan crystals was also detected with this β-mannosidase. These observa-tions indicate that the A. niger β-mannosidase can participate in the direct enzymatic attack onpolymeric substrates. However, the nature of the enzyme-polysaccharide interaction remains tobe elucidated.
Due to the complex nature of the plant cell wall and mannan-based substrates, several issues needto be addressed in order to achieve a better understanding of mannan-degradation. Firstly, theenzyme-polysaccharide interaction of mannoside-hydrolases in the degradation of the more com-plex heteromannans, like O-acetyl-galactoglucomanan, should be studied in more detail. In par-ticular, the influence of different substituents on the rate of hydrolysis needs to be investigatedfurther. Secondly, a larger comparative study of mannan-degrading enzymes from different en-zyme families would possibly reveal any differences or similarities in substrate specificity. Ingeneral, improved methods for the separation and detection of polysaccharides and oligosaccharideswould be very useful in these types of investigations.
51

Furthermore, the results from this work indicate that an endoglucanase from T. reesei candepolymerise galactoglucomannan. This observation is interesting to discuss in the framework ofmodular enzymes. Several modular enzymes possess both cellulase and hemicellulase catalyticmodules. Furthermore, modular enzymes may contain multiple CBMs with affinity for both cel-lulose and hemicellulose. Thus, it appears that, at least in some cases, the enzymatic degradationsof cellulose and hemicellulose are interlinked with each other. These connections between cata-lytic modules and CBMs with affinity for cellulose and hemicellulose are in accordance with theobserved enzyme-polysaccharide interaction of the modular β-mannanase from T. reesei and clearlyillustrate the complexity of the plant cell wall.
In the study of complex substrates, integrated approaches including several other types of cellwall-degrading enzymes, might prove to be useful. Since the results in this work showed that anendoglucanase is capable of depolymerising isolated galactoglucomannans, it would be interest-ing to assess if the enzyme also perform this activity in the plant cell wall. Possibly, such anactivity may facilitate the degradation of mannans by hemicellulases.
In summary, the work presented here has provided new information about enzyme-polysaccha-ride interaction in the modular β-mannanase from T. reesei which will be valuable for furtherstudies on this enzyme and other modular enzymes. Furthermore, hydrolysis of mannans bymannoside- and glycoside-hydrolases has opened up new questions regarding the substratespecificity in the subsites of these enzymes. Hopefully, the work presented in this thesis will be anincitement for continued studies on the specific polysaccharide interactions in modular enzymes.
52

Acknowledgements
This work would of course not have been possible without the help and support of others.
First, I would like to thank my supervisor Henrik Stålbrand for his great support and guid-ance during these years. You have been a very ambitious supervisor, which I appreciate, andyou have always had time to spare for talks and discussions. It has been a pleasure workingwith you.
I would also like to acknowledge all the collaborators in the different projects. Special thanksto: Henri Chanzy, who is a great scientist and an enormous knowledge which I admire.Elisabetta Sabini, who is good at crystallography and also a nice person. Claire Boisset, whointroduced me to immuno-gold labelling. Keith Wilson , who has provided fruitful discussionson the mannanase structure. I would also like to thank Jan-Christer Janson and Bingze Xu,for the work on the mussel mannanase and Marc Claeyssens and Wim Nerinckx for provid-ing interesting substrates.
I also want to thank all the people at the department who I have had the pleasure to work with:Jon and Torny (Järngänget), we have been working together a lot and we have also had a lot offun – Stureplan nästa! Lars, who did an excellent masters thesis. I am glad you took the rightdecision and continued in our group. It’s been very nice working with you. Pia, who I workedwith on the β-mannosidase. Thanks also to Anna for help with Trichoderma transformation.My acknowledgements also go to all other past and present members of the groups of HenrikStålbrand and Folke Tjerneld.
Diploma students and project-workers: Magnus, who did a very good job on the Pichia systemduring his master thesis. Ulrika and Esther who worked with the β-mannosidase.
I would also like to thank everybody at the department for providing a relaxed and stimulatingatmosphere. A special thank to the football and innebandy players: Ravi, Johan B, Henrik P,Tomas and others. Thank you to everybody who has met up at the pub and other social events:Jonathan, Irene, Johan K, Jörgen, Mia and many others.
A special thank to Jonathan Park for proof reading the text.
Gavelin klanen: Alf, Birgitta, Nicklas, Jonas, Helen, Ulf, Anna, Tove och Simon. Tack föralla goda minnen vi delat i Malmö och i Vitemölla.
Min syster Pernilla, du vet vad det här innebär. Du har verkligen varit en förebild. Tack förditt stöd och din uppmuntran. Torbjörn , som varit som en storebror under delar av min upp-växt. Adam och Amanda som är ljusglimtar i tillvaron. Generöse Torsten.
Mamma och pappa: tack för allt stöd ni har gett mig genom åren.
Slutligen vill jag tacka min älskade Erika . Tack för att du finns och för att du har stått ut medalla sena kvällar och helger.
53

54

References
55
1. Ademark, P. 2000. Galactoglucomannan-degrading enzymes from Aspergillus niger. Doctoral thesis.Lund University, Lund.2. Ademark, P., M. Larsson, F. Tjerneld, and H. Stålbrand. 2001. Multiple α-galactosidase fromAspergillus niger: purification, characterization, and substrate specificities. Enzyme Microb. Technol. 29:441-448.3. Ademark, P., A. Varga, J. Medve, V. Harjunpää, T. Drakenberg, F. Tjerneld, and H. Stålbrand.1998. Softwood hemicellulose-degrading enzymes from Aspergillus niger: purification and properties of aβ-mannanase. J. Biotechnol. 63(3): 199-210.4. Akino, T., C. Kato, and K. Horikoshi. 1989. The cloned β-mannanase gene from alkalophilic Bacillussp. AM-001 produces two β-mannanases in Escherichia coli. Arch. Microbiol. 152(1): 10-15.5. Akino, T., C. Kato, and K. Horikoshi. 1989. Two Bacillus β-mannanases having different COOHtermini are produced in Escherichia coli carrying pMAH5. Appl. Environ. Microbiol. 55(12): 3178-3183.6. Akino, T., N. Nakamura, and K. Horikoshi. 1988. Characterization of β-mannosidase of anAlkalophilic Bacillus sp. Agric. Biol. Chem. 52(6): 1459-1464.7. Akino, T., N. Nakamura, and K. Horikoshi. 1988. Characterization of three β-mannanases of analkalophilic Bacillus sp. Agric. Biol. Chem. 52(3): 773-779.8. Aleshin, A. E., L. M. Firsov, and R. B. Honzatko. 1994. Refined structure for the complex of acarbosewith glucoamylase from Aspergillus awamori var. X100 to 2.4-Å resolution. J. Biol. Chem. 269(22): 15631-15639.9. Alkhayat, A. H., S. A. Kraemer, J. R. Leipprandt, M. Macek, W. J. Kleijer, and K. H. Friderici.1998. Human β-mannosidase cDNA characterizatioon and first identificatioon of a mutation associatiedwith human β-mannosidosis. Hum. Mol. Genet. 7: 75-83.10. Alvarado, V., H. Nonogaki, and K. J. Bradford. 2000. Expression of endo-β-mannanase and SNF-related protein kinase genes in true potato seeds in relation to dormancy, gibberellin and abscisic acid, p. 347-364. In J. D. Viémont and J. Crabbé (ed.), Dormancy in Plants. C. A. B. International, Wallingford, U. K.11. Aparicio, R., E. V. Eneiskaya, A. A. Kulminskaya, A. N. Savel'ev, A. M. Golubev, K. N. Neustroev,J. Kobarg, and I. Polikarpov. 2000. Crystallization and preliminary X-ray study of β-mannosidase fromTrichoderma reesei. Acta Crystallogr., Sect. D: Biol. Crystallogr. 56(Pt 3): 342-343.12. Arai, M., H. Fujimoto, T. Ooi, S. Ogura, and S. Murao. 1995. Purification and properties of β-mannosidase from Aspergillus aculeatus. Oyo Toshitsu Kagaku. 42: 49-51.13. Araujo, A., and O. P. Ward. 1990. Extracellular mannanases and galactanases from selected fungi. J.Ind. Microbiol. 6(3): 171-178.14. Araujo, A., and O. P. Ward. 1990. Purification and some properties of the mannanases from Thielaviaterrestris. J. Ind. Microbiol. 6: 269-274.15. Arcand, N., D. Kluepfel, F. W. Paradis, R. Morosoli, and F. Shareck. 1993. β-mannanase ofStreptomyces lividans 66: cloning and DNA sequence of the manA gene and characterization of the enzyme.Biochem. J. 290: 857-863.16. Arima, T. 2001. Recycling of Wood and Fiber Products, p. 849-858. In D. N. S. Hon and N. Shiraishi(ed.), Wood and Cellulosic Chemistry, 2nd ed. Marcel Dekker, Inc., New York.17. Arisan-Atac, I., R. Hodits, D. Kristufek, and C. P. Kubicek. 1993. Purification and characterizationof a β-mannanase of Trichoderma reesei C-30. Appl. Microbiol. Biotechnol. 39(1): 58-62.18. Aspinall, G. O., E. L. Hirst, E. G. V. Percival, and I. R. Williamson. 1953. The Mannans of IvoryNut (Phytelephas macrocarpa). Part I. The Methylation of Mannan A and Mannan B. J. Chem. Soc. PartIII : 3184-3188.19. Atkins, E. D. T., S. Farnell, C. Burden, W. Mackie, and B. Sheldrick. 1988. Crystalline structureand packing of mannan I. Biopolymers. 27(7): 1097-1105.20. Baird, S. D., M. A. Hefford, D. A. Johnson, W. L. Sung, M. Yaguchi, and V. L. Seligy. 1990. TheGlu residue in the conserved Asn-Glu-Pro sequence of two highly divergent endo-β-1,4-glucanases is es-sential for enzymatic activity. Biochem. Biophys. Res. Commun. 169(3): 1035-1039.21. Bauer, M. W., E. J. Bylina, R. V. Swanson, and R. M. Kelly. 1996. Comparison of a β-glucosidaseand a β-mannosidase from the hyperthermophilic archaeon Pyrococcus furiosus. Purification, characteriza-tion, gene cloning and sequence analysis. J. Biol. Chem. 271: 23749-23755.

56
22. Beccari, T., L. Bibi, S. Stinchi, J. L. Stirling, and A. Orlacchio. 2001. Mouse β-mannosidase: cDNAcloning, expression, and chromosomal localization. Biosci. Rep. 21(3): 315-323.23. Becker, D., C. Braet, H. Brumer III, M. Claeyssens, C. Divne, B. R. Fagerstrom, M. Harris, T. A.Jones, G. J. Kleywegt, A. Koivula, S. Mahdi, K. Piens, M. L. Sinnott, J. Ståhlberg, T. T. Teeri, M.Underwood, and G. Wohlfahrt. 2001. Engineering of a glycosidase Family 7 cellobiohydrolase to morealkaline pH optimum: the pH behaviour of Trichoderma reesei Cel7A and its E223S/ A224H/L225V/T226A/D262G mutant. Biochem. J. 356(Pt 1): 19-30.24. Bewley, J. D., M. Banik, R. Bourgault, J. A. Feurtado, P. Toorop, and H. W. Hilhorst. 2000. Endo-β-mannanase activity increases in the skin and outer pericarp of tomato fruits during ripening. J. Exp. Bot.51(344): 529-538.25. Bewley, J. D., R. A. Burton, Y. Morohashi, and G. B. Fincher. 1997. Molecular cloning of a cDNAencoding a (1->4)-β-mannan endohydrolase from the seeds of germinated tomato (Lycopersicon esculentum).Planta. 203(4): 454-459.26. Biely, P., and M. Tenkanen. 1998. Enzymology of hemicellulose degradation, p. 25-47. In G. E.Harman and C. P. Kubicek (ed.), Trichoderma & Gliocladium, vol. 2. Taylor and Francis Ltd., London.27. Biely, P., M. Vrsanska, and M. Claeyssens. 1991. The endo-1,4-β-glucanase I from Trichodermareesei. Action on β-1,4-oligomers and polymers derived from D-glucose and D-xylose. Eur. J. Biochem.200(1): 157-163.28. Black, G. W., J. E. Rixon, J. H. Clarke, G. P. Hazlewood, L. M. Ferreira, D. N. Bolam, and H. J.Gilbert. 1997. Cellulose binding domains and linker sequences potentiate the activity of hemicellulasesagainst complex substrates. J. Biotechnol. 57(1-3): 59-69.29. Black, G. W., J. E. Rixon, J. H. Clarke, G. P. Hazlewood, M. K. Theodorou, P. Morris, and H. J.Gilbert. 1996. Evidence that linker sequences and cellulose-binding domains enhance the activity ofhemicellulases against complex substrates. Biochem. J. 319(Pt 2): 515-520.30. Blake, C. C. F., D. F. Koenig, G. A. Mair, A. C. T. North, D. C. Phillips, and V. R. Sarma. 1965.Structure of hen egg-white lysozyme — a three-dimensional Fourier synthesis at 2 Å resolution. Nature.206(4986): 757-761.31. Bolam, D. N., A. Ciruela, S. McQueen-Mason, P. Simpson, M. P. Williamson, J. E. Rixon, A.Boraston, G. P. Hazlewood, and H. J. Gilbert. 1998. Pseudomonas cellulose-binding domains mediatetheir effects by increasing enzyme substrate proximity. Biochem. J. 331(Pt 3): 775-781.32. Bolam, D. N., N. Hughes, R. Virden, J. H. Lakey, G. P. Hazlewood, B. Henrissat, K. L. Braithwaite,and H. J. Gilbert. 1996. Mannanase A from Pseudomonas fluorescens ssp. cellulosa is a retaining glycosylhydrolase in which E212 and E320 are the putative catalytic residues. Biochemistry. 35(50): 16195-16204.33. Boraston, A. B., P. Chiu, R. A. Warren, and D. G. Kilburn. 2000. Specificity and affinity of substratebinding by a family 17 carbohydrate-binding module from Clostridium cellulovorans cellulase 5A. Bio-chemistry. 39(36): 11129-11136.34. Boraston, A. B., A. L. Creagh, M. M. Alam, J. M. Kormos, P. Tomme, C. A. Haynes, R. A. War-ren, and D. G. Kilburn. 2001. Binding specificity and thermodynamics of a family 9 carbohydrate-bindingmodule from Thermotoga maritima xylanase 10A. Biochemistry. 40(21): 6240-6247.35. Boraston, A. B., M. Ghaffari, R. A. Warren, and D. G. Kilburn. 2002. Identification and glucan-binding properties of a new carbohydrate-binding module family. Biochem. J. 361(Pt 1): 35-40.36. Boraston, A. B., B. W. McLean, J. M. Kormos, M. Alam, N. R. Gilkes, C. A. Haynes, P. Tomme,D. G. Kilburn, and R. A. J. Warren. 1999. Carbohydrate-binding modules: diversity of structure andfunction, p. 202-211. In H. J. Gilbert, G. J. Davies, B. Henrissat, and B. Svensson (ed.), Recent Advances inCarbohydrate Bioengineering, vol. 246. Royal Society of Chemistry, Cambridge.37. Bork, P., A. K. Downing, B. Kieffer, and I. D. Campbell. 1996. Structure and distribution of mod-ules in extracellular proteins. Q. Rev. Biophys. 29(2): 119-167.38. Bourgault, R., J. D. Bewley, A. Alberici, and D. Decker. 2001. Endo-β-mannanase activity in to-mato and other ripening fruits. HortScience. 36(1): 72-75.39. Bourne, Y., and B. Henrissat. 2001. Glycoside hydrolases and glycosyltransferases: families andfunctional modules. Curr. Opin. Struct. Biol. 11(5): 593-600.40. Braithwaite, K. L., G. W. Black, G. P. Hazlewood, B. R. Ali, and H. J. Gilbert. 1995. A non-modular endo-β-1,4-mannanase from Pseudomonas fluorescens subspecies cellulosa. Biochem. J. 305(Pt3): 1005-1010.41. Bray, M. R., P. E. Johnson, N. R. Gilkes, L. P. McIntosh, D. G. Kilburn, and R. A. Warren. 1996.

57
Probing the role of tryptophan residues in a cellulose-binding domain by chemical modification. ProteinSci. 5(11): 2311-2318.42. Brun, E., P. E. Johnson, A. L. Creagh, P. Tomme, P. Webster, C. A. Haynes, and L. P. McIntosh.2000. Structure and binding specificity of the second N-terminal cellulose-binding domain from Cellulomonasfimi endoglucanase C. Biochemistry. 39(10): 2445-2458.43. Brun, E., F. Moriaud, P. Gans, M. J. Blackledge, F. Barras, and D. Marion. 1997. Solution struc-ture of the cellulose-binding domain of the endoglucanase Z secreted by Erwinia chrysanthemi. Biochemis-try. 36(51): 16074-16086.44. Buchert, J., T. Oksanen, J. Pere, M. Siika-Aho, A. Suurnakki, and L. Viikari. 1998. Applicationsof Trichoderma reesei enzymes in the pulp and paper industry, p. 343-363. In G. E. Harman and C. P.Kubicek (ed.), Trichoderma Gliocladium, vol. 2. Taylor and Francis Ltd., London.45. Buchert, J., J. Salminen, M. Siika-aho, M. Ranua, and L. Viikari. 1993. The role of Trichodermareesei xylanase and mannanase in the treatment of softwood kraft pulp prior to bleaching. Holzforschung.47(6): 473-478.46. Cann, I. K. O., S. Kocherginskaya, M. R. King, B. A. White, and R. I. Mackie. 1999. MolecularCloning, Sequencing, and Expression of a Novel Multidomain Mannanase Gene from Thermoanaerobac-terium polysaccharolyticum. J. Bacteriol. 181(5): 1643-1651.47. Carrard, G., A. Koivula, H. Söderlund, and P. Beguin. 2000. Cellulose-binding domains promotehydrolysis of different sites on crystalline cellulose. Proc. Natl. Acad. Sci. U. S. A. 97(19): 10342-10347.48. Chambers, R. S., A. R. Walden, G. S. Brooke, J. F. Cutfield, and P. A. Sullivan. 1993. Identifica-tion of a putative active site residue in the exo-β-(1,3)-glucanase of Candida albicans. FEBS Lett. 327(3):366-369.49. Chanzy, H., S. Pérez, D. P. Miller, G. Paradossi, and W. T. Winter. 1987. An electron diffractionstudy of the mannan I crystal and molecular structure. Macromolecules. 20(10): 2407-2013.50. Chanzy, H. D., A. Grosrenaud, J. P. Joseleau, M. Dubé, and R. H. Marchessault. 1982. Crystalli-zation behavior of glucomannan. Biopolymers. 21(2): 301-319.51. Chanzy, H. D., A. Grosrenaud, R. Vuong, and W. Mackie. 1984. The crystalline polymorphism ofmannan in plant cell walls and after recrystallization. Planta. 161(4): 320-329.52. Charnock, S. J., D. N. Bolam, J. P. Turkenburg, H. J. Gilbert, L. M. Ferreira, G. J. Davies, andC. M. Fontes. 2000. The X6 “thermostabilizing” domains of xylanases are carbohydrate-binding modules:structure and biochemistry of the Clostridium thermocellum X6b domain. Biochemistry. 39(17): 5013-5021.53. Charrier, M., and C. Rouland. 2001. Mannan-degrading enzymes purified from the crop of thebrown garden snail Helix aspersa Müller (Gastropoda pulmonata). J. Exp. Zool. 290(2): 125-135.54. Chen, H., J. R. Leipprandt, C. E. Traviss, B. L. Sopher, M. Z. Jones, K. T. Cavanagh, and K. H.Friderici. 1995. Molecular cloning and characterization of bovine β-mannosidase. J. Biol. Chem. 270(8):3841-3848.55. Chesson, A., and C. W. Forsberg. 1988. Polysaccharide Degradation by Rumen Microorganisms, p.251-284. In P. N. Hobson (ed.), The Rumen Microbial Ecosystem. Elsevier Science Publishers Ltd., London.56. Christgau, S., S. Kauppinen, J. Vind, L. V. Kofod, and H. Dalboge. 1994. Expression cloning,purification and characterization of a β-1,4-mannanase from Aspergillus aculeatus. Biochem. Mol. Biol. Int.33(5): 917-925.57. Civas, A., R. Eberhard, P. Le Dizet, and F. Petek. 1984. Glycosidases induced in Aspergillus tamarii.Secreted α-D-galactosidase and β-D-mannanase. Biochem. J. 219(3): 857-863.58. Clarke, J. H., K. Davidson, J. E. Rixon, J. R. Halstead, M. P. Fransen, H. J. Gilbert, and G. P.Hazlewood. 2000. A comparison of enzyme-aided bleaching of softwood paper pulp using combinations ofxylanase, mannanase and α-galactosidase. Appl. Microbiol. Biotechnol. 53(6): 661-667.59. Collén, A., M. Ward, F. Tjerneld, and H. Stålbrand. 2001. Genetically engineered peptide fusionsfor improved protein partitioning in aqueous two-phase systems. Effect of fusion localization on endoglucanaseI of Trichoderma reesei. J. Chromatogr. A. 910(2): 275-284.60. Colvin, J. R. 1980. The biosynthesis of cellulose, p. 543-570. In P. K. Stumpf and E. E. Conn (ed.),The Biochemisty of Plants, vol. 3. Academic Press, London.61. Coughlan, M. P., M. G. Touhy, E. X. F. Filho, J. Puls, M. Claeyssens, M. Vrsanska, and M. M.Hughes. 1993. Enzymological aspects of microbial hemicellulases with emphasis on fungal systems, p. 53-84. In M. P. Coughlan and G. P. Hazlewood (ed.), Hemicellulose and Hemicellulases, vol. 4. Portland PressRes. Monogr., London.

58
62. Coulombel, C., S. Clermont, M. J. Foglietti, and F. Percheron. 1981. Transglycosylation reactionscatalysed by two β-mannanases. Biochem. J. 195(1): 333-335.63. Coutinho, P. M., and B. Henrissat. 1999. Carbohydrate-active enzymes: an integrated databaseapproach, p. 3-12. In H. J. Gilbert, G. J. Davies, B. Henrissat, and B. Svensson (ed.), Recent Advances inCarbohydrate Bioengineering, vol. 246. Royal Society of Chemistry, Cambridge.64. Creagh, A. L., E. Ong, E. Jervis, D. G. Kilburn, and C. A. Haynes. 1996. Binding of the cellulose-binding domain of exoglucanase Cex from Cellulomonas fimi to insoluble microcrystalline cellulose isentropically driven. Proc. Natl. Acad. Sci. U. S. A. 93(22): 12229-12234.65. Curtis, H., and N. S. Barnes. 1989. Biology, 5th ed. Worth publishers Inc., New York.66. Czjzek, M., D. N. Bolam, A. Mosbah, J. Allouch, C. M. Fontes, L. M. Ferreira, O. Bornet, V.Zamboni, H. Darbon, N. L. Smith, G. W. Black, B. Henrissat, and H. J. Gilbert. 2001. The location ofthe ligand-binding site of carbohydrate-binding modules that have evolved from a common sequence is notconserved. J. Biol. Chem. 276(51): 48580-48587.67. Daas, P. J., H. A. Schols, and H. H. de Jongh. 2000. On the galactosyl distribution of commercialgalactomannans. Carbohydr. Res. 329(3): 609-619.68. Dalboge, H., and H. P. Heldt-Hansen. 1994. A novel method for efficient expression cloning offungal enzyme genes. Mol. Gen. Genet. 243(3): 253-260.69. Davies, G., and B. Henrissat. 1995. Structures and mechanisms of glycosyl hydrolases. Structure.3(9): 853-859.70. Davies, G., M. L. Sinnott, and S. G. Withers. 1998. Glycosyl transfer, p. 119-208. In M. L. Sinnott(ed.), Comprehensive biological catalysis, vol. 1. Academic Press, New York.71. Davies, G. J., M. Dauter, A. M. Brzozowski, M. E. Bjornvad, K. V. Andersen, and M. Schulein.1998. Structure of the Bacillus agaradherans family 5 endoglucanase at 1.6 A and its cellobiose complex at2.0 A resolution. Biochemistry. 37(7): 1926-1932.72. Davies, G. J., K. S. Wilson, and B. Henrissat. 1997. Nomenclature for sugar-binding subsites inglycosyl hydrolases. Biochem. J. 321(Pt 2): 557-559.73. Dea, I. C. M., A. H. Clark, and B. V. McCleary. 1986. Effect of the molecular fine structure ofgalactomannans on their interaction properties - the role of unsubstituted sides. Food Hydrocolloids. 1(2):129-140.74. Din, N., N. R. Gilkes, B. Tekant, R. C. Miller, A. J. Warren, and D. G. Kilburn. 1991. Non-Hydrolytic Disruption of Cellulose Fibers By the Binding Domain of a Bacterial Cellulase. Bio/Technol.9(11): 1096-1099.75. Divne, C., J. Ståhlberg, T. Reinikainen, L. Ruohonen, G. Pettersson, J. K. Knowles, T. T. Teeri,and T. A. Jones. 1994. The three-dimensional crystal structure of the catalytic core of cellobiohydrolase Ifrom Trichoderma reesei. Science. 265(5171): 524-528.76. Dominguez, R., H. Souchon, S. Spinelli, Z. Dauter, K. S. Wilson, S. Chauvaux, P. Beguin, and P.M. Alzari. 1995. A common protein fold and similar active site in two distinct families of β-glycanases. Nat.Struct. Biol. 2(7): 569-576.77. Dutta, S., K. J. Bradford, and D. J. Nevins. 1997. Endo-β-mannanase activity present in cell wallextracts of lettuce endosperm prior to radicle emergence. Plant Physiol. 113(1): 155-161.78. Ethier, N., G. Talbot, and J. Sygusch. 1998. Gene cloning, DNA sequencing, and expression of ther-mostable β-mannanase from Bacillus stearothermophilus. Appl. Environ. Microbiol. 64(11): 4428-4432.79. Evans, L. V., and D. M. Butler. 1988. Seaweed biotechnology - current status and future prospects,p. 335-350. In L. J. Rogers and J. R. Gallon (ed.), Biochemistry of the Algae and Cyanobacteria, vol. 28.Oxford University Press, Oxford.80. Ezaki, S., K. Miyaoku, K.-I. Nishi, T. Tanaka, S. Fujiwara, M. Takagi, H. Atomi, and T. Imanaka.1999. Gene analysis and enzymatic properties of thermostable β-glycosidase from Pyrococcus kodakaraensisKOD1. J. Biosci. Bioeng. 88(2): 130-135.81. Fang, T. Y., and C. Ford. 1998. Protein engineering of Aspergillus awamori glucoamylase to in-crease its pH optimum. Protein Eng. 11(5): 383-388.82. Fanutti, C., T. Ponyi, G. W. Black, G. P. Hazlewood, and H. J. Gilbert. 1995. The conservednoncatalytic 40-residue sequence in cellulases and hemicellulases from anaerobic fungi functions as a pro-tein docking domain. J. Biol. Chem. 270(49): 29314-29322.83. Fengel, D., and G. Wegener. 1989. Wood: Chemistry, Ultrastructure, Reactions. Walter de Gruyter,New York.

59
84. Fernandes, A. C., C. M. Fontes, H. J. Gilbert, G. P. Hazlewood, T. H. Fernandes, and L. M.Ferreira. 1999. Homologous xylanases from Clostridium thermocellum: evidence for bi-functional activity,synergism between xylanase catalytic modules and the presence of xylan-binding domains in enzyme com-plexes. Biochem. J. 342(Pt 1): 105-110.85. Flari, V., M. Matoub, and C. Rouland. 1995. Purification and characterization of a β-mannanasefrom the digestive tract of the edible snail Helix lucorum L. Carbohydr. Res. 275(1): 207-213.86. Fogarty, W. M. 1994. Enzymes of the Genus Aspergillus, p. 177-218. In J. E. Smith (ed.), Aspergillus,vol. 7. Plenum Press, New York.87. Freelove, A. C., D. N. Bolam, P. White, G. P. Hazlewood, and H. J. Gilbert. 2001. A novel carbo-hydrate-binding protein is a component of the plant cell wall-degrading complex of Piromyces equi. J. Biol.Chem. 276(46): 43010-43017.88. Frei, E., and R. D. Preston. 1968. Noncellulosic structural polysaccharides in algal cell walls. III.Mannan in siphoneous green algae. Proc. R. Soc. London, Ser. B. 169(1015): 127-145.89. Gabrielii, I. 1998. Hydrogels based on hardwood hemicelluloses.Licentiate thesis. Chalmers Univer-sity of Technology, Göteborg.90. Gerber, P. J., J. A. Heitmann, T. W. Joyce, J. Buchert, and M. Siika-aho. 1999. Adsorption ofhemicellulases onto bleached kraft fibers. J. Biotechnol. 67(1): 67-75.91. Gherardini, F. C., and A. A. Salyers. 1987. Characterization of an outer membrane mannanase fromBacteroides ovatus. J. Bacteriol. 169(5): 2031-2037.92. Gibbs, M. D., A. U. Elinder, R. A. Reeves, and P. L. Bergquist. 1996. Sequencing, cloning andexpression of a β-1,4-mannanase gene, manA, from the extremely thermophilic anaerobic bacterium,Caldicellulosiruptor Rt8B.4. FEMS Microbiol. Lett. 141(1): 37-43.93. Gibbs, M. D., R. A. Reeves, A. Sunna, and P. L. Bergquist. 1999. Sequencing and expression of a β-mannanase gene from the extreme thermophile Dictyoglomus thermophilum Rt46B.1, and characteristics ofthe recombinant enzyme. Curr. Microbiol. 39(6): 351-357.94. Gibbs, M. D., D. J. Saul, E. Luthi, and P. L. Bergquist. 1992. The β-mannanase from “Caldocellumsaccharolyticum” is part of a multidomain enzyme. Appl. Environ. Microbiol. 58(12): 3864-3867.95. Gilbert, H. J., D. N. Bolam, L. Szabo, H. Xie, M. P. Williamson, P. J. Simpson, S. Jamal, A. B.Boraston, D. G. Kilburn, and R. A. J. Warren. 2002. An update on carbohydrate binding modules, p. 89-98. In T. Teeri, B. Svensson, H. J. Gilbert, and T. Freizi (ed.), Carbohydrate Bioengineering: Interdiscipli-nary approaches, vol. 275. Royal Society of Chemistry, Cambridge.96. Gilkes, N. R., B. Henrissat, D. G. Kilburn, R. C. Miller, Jr., and R. A. Warren. 1991. Domains inmicrobial β-1, 4-glycanases: sequence conservation, function, and enzyme families. Microbiol. Rev. 55(2):303-315.97. Gilkes, N. R., E. Jervis, B. Henrissat, B. Tekant, R. C. Miller, Jr., R. A. Warren, and D. G.Kilburn. 1992. The adsorption of a bacterial cellulase and its two isolated domains to crystalline cellulose.J. Biol. Chem. 267(10): 6743-6749.98. Gilkes, N. R., R. A. Warren, R. C. Miller, Jr., and D. G. Kilburn. 1988. Precise excision of thecellulose binding domains from two Cellulomonas fimi cellulases by a homologous protease and the effecton catalysis. J. Biol. Chem. 263(21): 10401-10407.99. Gill, J., J. E. Rixon, D. N. Bolam, S. McQueen-Mason, P. J. Simpson, M. P. Williamson, G. P.Hazlewood, and H. J. Gilbert. 1999. The type II and X cellulose-binding domains of Pseudomonas xylanaseA potentiate catalytic activity against complex substrates by a common mechanism. Biochem. J. 342(Pt 2):473-480.100. Godfrey, T., and S. I. West. 1996. Introduction to industrial enzymology, p. 1-8. In T. Godfrey and S.I. West (ed.), Industrial Enzymology, 2nd ed. Stockton press, New York.101. Goldstein, A. M., E. N. Alter, and J. K. Seaman. 1973. Guar gum, p. 303-321. In R. Whistler and J.M. BeMiller (ed.), Industrial Gums. Academic Press, New York.102. Grassin, C., and P. Fauquembergue. 1996. Fruit juices, p. 225-264. In T. Godfrey and S. I. West(ed.), Industrial Enzymology, 2nd ed. Stockton Press, New York.103. Grinna, L. S., and J. F. Tschopp. 1989. Size Distribution and General Structural Features of N-Linked Oligosaccharides from the Methylotrophic Yeast, Pichia pastoris. Yeast. 5: 107-115.104. Groot, S. P. C., B. Kieliszewska-Rokicka, E. Vermeer, and C. M. Karssen. 1988. Gibberellin-in-duced hydrolysis of endosperm cell walls in gibberellin-deficient tomato seeds prior to radicle protrusion.Planta. 174(4): 500-504.

60
105. Guex, N., and M. C. Peitsch. 1997. Swiss-model and the Swiss-PdbViewer: An environment forcomparative protein modelling. Electrophoresis. 18: 2714-2723.106. Gübitz, G. M., M. Hayn, M. Sommerauer, and W. Steiner. 1996. Mannan-Degrading Enzymesfrom Sclerotium rolfsii: Characterisation and synergism of two Endo β-Mannanases and a β-Mannosidase.Bioresour. Technol. 58: 127-135.107. Gübitz, G. M., M. Hayn, G. Urbanz, and W. Steiner. 1996. Purification and properties of an acidicβ-mannanase from Sclerotium rolfsii. J. Biotechnol. 45(2): 165-172.108. Gübitz, G. M., A. Sachslehner, and D. Haltrich. 2001. Microbial Mannanases: Substrates, Produc-tion and Applicantions, p. 236-262. In M. E. Himmel, J. O. Baker, and M. E. Himmel (ed.), GlycosylHydrolases for Biomass Conversion, vol. 769. American Chemical Society, Washington D. C.109. Hall, J., G. W. Black, L. M. Ferreira, S. J. Millward-Sadler, B. R. Ali, G. P. Hazlewood, and H. J.Gilbert. 1995. The non-catalytic cellulose-binding domain of a novel cellulase from Pseudomonas fluorescenssubsp. cellulosa is important for the efficient hydrolysis of Avicel. Biochem. J. 309(Pt 3): 749-756.110. Halstead, J. R., P. E. Vercoe, H. J. Gilbert, K. Davidson, and G. P. Hazlewood. 1999. A family 26mannanase produced by Clostridium thermocellum as a component of the cellulosome contains a domainwhich is conserved in mannanases from anaerobic fungi. Microbiology (Reading U.K.). 145(Pt 11): 3101-3108.111. Harjunpää, V., J. Helin, A. Koivula, M. Siika-aho, and T. Drakenberg. 1999. A comparative studyof two retaining enzymes of Trichoderma reesei: transglycosylation of oligosaccharides catalysed by thecellobiohydrolase I, Cel7A, and the β-mannanase, Man5A. FEBS Lett. 443(2): 149-153.112. Harjunpää, V., A. Teleman, M. Siika-aho, and T. Drakenberg. 1995. Kinetic and stereochemicalstudies of manno-oligosaccharide hydrolysis catalysed by β-mannanases from Trichoderma reesei. Eur. J.Biochem. 234: 278-283.113. Hashimoto, Y., and J. Fukumoto. 1969. Enzymic treatment of coffee beans. II. Purification andproperties of β-mannosidase of Rhizopus niveus. J. Agric. Chem. Soc. Jpn. 43(8): 564-569.114. Hazlewood, G. P., and H. J. Gilbert. 1998. Structure and function analysis of Pseudomonas plantcell wall hydrolases. Prog. Nucleic. Acid Res. Mol. Biol. 61: 211-241.115. Henrissat, B. 1991. A classification of glycosyl hydrolases based on amino acid sequence similarities.Biochem. J. 280(Pt 2): 309-316.116. Henrissat, B., and A. Bairoch. 1993. New families in the classification of glycosyl hydrolases basedon amino acid sequence similarities. Biochem. J. 293(Pt 3): 781-788.117. Henrissat, B., and A. Bairoch. 1996. Updating the sequence-based classification of glycosyl hydrolases.Biochem. J. 316(Pt 2): 695-696.118. Henrissat, B., I. Callebaut, S. Fabrega, P. Lehn, J. P. Mornon, and G. Davies. 1995. Conservedcatalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases. Proc.Natl. Acad. Sci. U. S. A. 92: 7090-7094.119. Henrissat, B., and G. Davies. 1997. Structural and sequence-based classification of glycosidehydrolases. Curr. Opin. Struct. Biol. 7(5): 637-644.120. Henrissat, B., T. T. Teeri, and R. A. J. Warren. 1998. A scheme for designating enzymes that hydrolyzethe polysaccharides in the cell walls of plants. FEBS Lett. 425(2): 352-354.121. Hilge, M., S. M. Gloor, W. Rypniewski, O. Sauer, T. D. Heightman, W. Zimmermann, K.Winterhalter, and K. Piontek. 1998. High-resolution native and complex structures of thermostable beta-mannanase from Thermomonospora fusca - substrate specificity in glycosyl hydrolase family 5. Structure.6(11): 1433-1444.122. Hirata, K., Y. Aso, and M. Ishiguro. 1998. Purification and some properties of β-mannosidase, β-N-acetylglucosaminidase, and β-galactosidase from apple snails (Pomacea canaliculata). J. Fac. Agric., KyushuUniv. 42(3-4): 463-472.123. Hoffrén, A. M., T. T. Teeri, and O. Teleman. 1995. Molecular dynamics simulation of fungal cellu-lose-binding domains: differences in molecular rigidity but a preserved cellulose binding surface. ProteinEng. 8(5): 443-450.124. Hogg, D., E.-J. Woo, D. N. Bolam, V. A. McKie, H. J. Gilbert, and R. W. Pickersgill. 2001. Crystalstructure of mannanase 26A from Pseudomonas cellulosa and analysis of residues involved in substratebinding. J. Biol. Chem. 276(33): 31186-31192.125. Hossain, M. Z., J.-i. Ab, and S. Hizukuri. 1996. Multiple forms of β-mannanase from Bacillus sp.KK01. Enzyme Microb. Technol. 18(2): 95-98.

61
126. Ikiri, Y., and T. Miwa. 1960. Chemical Nature of the cell wall of the green algae, Codium, Acetabulariaand Halicoryne. Nature. 185: 178-179.127. Iwasaki, Y., A. Tsuji, K. Omura, and Y. Suzuki. 1989. Purification and characterization of β-mannosidase from human placenta. J. Biochem. (Tokyo). 106(2): 331-335.128. Jackson, M. E., D. W. Fodge, and H. Y. Hsiao. 1999. Effects of β-mannanase in corn-soybean mealdiets on laying hen performance. Poult. Sci. 78(12): 1737-1741.129. Jacobson, R. H., X. J. Zhang, R. F. DuBose, and B. W. Matthews. 1994. Three-dimensional struc-ture of β-galactosidase from E. coli. Nature. 369(6483): 761-766.130. Jain, S., W. B. Drendel, Z. W. Chen, F. S. Mathews, W. S. Sly, and J. H. Grubb. 1996. Structure ofhuman β-glucuronidase reveals candidate lysosomal targeting and active-site motifs. Nat. Struct. Biol. 3(4):375-381.131. Jenkins, J., L. Lo Leggio, G. Harris, and R. Pickersgill. 1995. β-glucosidase, β-galactosidase, fam-ily A cellulases, family F xylanases and two barley glycanases form a superfamily of enzymes with 8-fold β/α architecture and with two conserved glutamates near the carboxy-terminal ends of β-strands four andseven. FEBS Lett. 362(3): 281-285.132. Johnson, P. E., M. D. Joshi, P. Tomme, D. G. Kilburn, and L. P. McIntosh. 1996. Structure of theN-terminal cellulose-binding domain of Cellulomonas fimi CenC determined by nuclear magnetic reso-nance spectroscopy. Biochemistry. 35(45): 14381-14394.133. Johnson, P. E., P. Tomme, M. D. Joshi, and L. P. McIntosh. 1996. Interaction of solublecellooligosaccharides with the N-terminal cellulose-binding domain of Cellulomonas fimi CenC 2. NMRand ultraviolet absorption spectroscopy. Biochemistry. 35(44): 13895-13906.134. Johnson, W. G. 1997. Disorders of Glycoprotein Degradation: Sialidosis, Fucosidosis, α-Mannosidosis,β-Mannosidosis, and Aspartylglycosaminuria, p. 355-369. In R. N. Rosenberg (ed.), Molecular and geneticbasis of neurological disease. Butterworth-Heinemann, Boston.135. Jones, J. K. N. 1950. Structure of the mannan present in Porphyra umbilicalis. J. Chem. Soc.: 3292-3295.136. Juers, D. H., T. D. Heightman, A. Vasella, J. D. McCarter, L. Mackenzie, S. G. Withers, and B. W.Matthews. 2001. A structural view of the action of Escherichia coli (lacZ) β-galactosidase. Biochemistry.40(49): 14781-14794.137. Juers, D. H., R. E. Huber, and B. W. Matthews. 1999. Structural comparisons of TIM barrel proteinssuggest functional and evolutionary relationships between β-galactosidase and other glycohydrolases. Pro-tein Sci. 8(1): 122-136.138. Karlsson, J. 2000. Fungal Cellulases: Study of hydrolytic properties of endoglucanases from Tri-choderma reesei and Humicola insolens.Doctoral thesis. Lund University, Lund.139. Kenne, L., K.-G. Rosell, and S. Svensson. 1975. Studies on the distribution of the O-acetyl groups inpine glucomannan. Carbohydr. Res. 44: 69-76.140. Khanongnuch, C., T. Ooi, and S. Kinoshita. 1999. Cloning and nucleotide sequence of β-mannanaseand cellulase genes from Bacillus sp. 5H. World J. Microbiol. Biotechnol. 15(2): 221-228.141. Khosla, C., and P. B. Harbury. 2001. Modular enzymes. Nature. 409(6817): 247-252.142. Kinghorn, J. R., and S. E. Unkles. 1994. Molecular Genetics and Expression of Foreign Proteins in theGenus Aspergillus, p. 65-100. In J. E. Smith (ed.), Aspergillus, vol. 7. Plenum Press, New York.143. Kormos, J., P. E. Johnson, E. Brun, P. Tomme, L. P. McIntosh, C. A. Haynes, and D. G. Kilburn.2000. Binding site analysis of cellulose binding domain CBD(N1) from endoglucanase C of Cellulomonasfimi by site-directed mutagenesis. Biochemistry. 39(30): 8844-8852.144. Koshland, D. E. J. 1953. Stereochemistry and the mechanism of enzymic reactions. Biol. Revs. Cam-bridge Phil. Soc. 28: 416-436.145. Kraulis, J., G. M. Clore, M. Nilges, T. A. Jones, G. Pettersson, J. Knowles, and A. M. Gronenborn.1989. Determination of the three-dimensional solution structure of the C-terminal domain of cellobiohydrolaseI from Trichoderma reesei. A study using nuclear magnetic resonance and hybrid distance geometry-dy-namical simulated annealing. Biochemistry. 28(18): 7241-7257.146. Kulminskaya, A. A., E. V. Eneiskaya, L. S. Isaeva-Ivanova, A. N. Savel’ev, I. A. Sidorenko, K.Shabalin, A., A. M. Golubev, and K. N. Neustroev. 1999. Enzymatic activity and β-galactomannan bin-ding property of β-mannosidase from Trichoderma reesei. Enzyme Microb.Technol. 25: 372-377.147. Kurokawa, J., E. Hemjinda, T. Arai, S. Karita, T. Kimura, K. Sakka, and K. Ohmiya. 2001.Sequence of the Clostridium thermocellum mannanase gene man26B and characterization of the translatedproduct. Biosci. Biotechnol. Biochem. 65(3): 548-554.

62
148. Leipprandt, J. R., S. A. Kraemer, B. E. Haithocock, H. Chen, J. L. Dyme, K. T. Cavanagh, K. H.Friderici, and M. Z. Jones. 1996. Caprine β-mannosidase: sequencing and characterization of the cDNAand identification of the molecular defect of caprine β-mannosidosis. Genomics. 37: 51-56.149. Lemesle-Varloot, L., B. Henrissat, C. Gaboriaud, V. Bissery, A. Morgat, and J. P. Mornon. 1990.Hydrophobic cluster analysis: procedures to derive structural and functional information from 2-D-repre-sentation of protein sequences. Biochimie. 72(8): 555-574.150. Lindberg, B., K.-G. Rosell, and S. Svensson. 1973. Position of the O-acetyl groups in pineglucomannan. Svensk Papperstidning. 76: 383-384.151. Linder, M., G. Lindeberg, T. Reinikainen, T. T. Teeri, and G. Pettersson. 1995. The difference inaffinity between two fungal cellulose-binding domains is dominated by a single amino acid substitution.FEBS Lett. 372(1): 96-98.152. Linder, M., M. L. Mattinen, M. Kontteli, G. Lindeberg, J. Ståhlberg, T. Drakenberg, T.Reinikainen, G. Pettersson, and A. Annila. 1995. Identification of functionally important amino acids inthe cellulose-binding domain of Trichoderma reesei cellobiohydrolase I. Protein Sci. 4(6): 1056-1064.153. Linder, M., T. Nevanen, and T. T. Teeri. 1999. Design of a pH-dependent cellulose-binding domain.FEBS Lett. 447(1): 13-16.154. Linder, M., I. Salovuori, L. Ruohonen, and T. T. Teeri. 1996. Characterization of a double cellulose-binding domain. Synergistic high affinity binding to crystalline cellulose. J. Biol. Chem. 271(35): 21268-21272.155. Linder, M., and T. T. Teeri. 1997. The roles and function of cellulose-binding domains. J. Biotechnol.57: 15-28.156. Loos, E., and D. Meindl. 1982. Composition of the cell wall of Chlorella fusca. Planta. 156(3): 270-273.157. Love, J., and E. Percival. 1964. The polysaccharides of the green seaweed Codium fragile Part III. Aβ-1,4-linked mannan. J. Chem. Soc.: 3345-3350.158. Lundqvist, J., A. Teleman, L. Junel, G. Zaachi, O. Dahlman, F. Tjerneld, and H. Stålbrand. 2002.Isolation and characterization of galactoglucomannan from Spruce (Picea abies). Carbohydr. Polym. 48(1):29-39.159. Luonteri, E., M. Tenkanen, and L. Viikari. 1998. Substrate Specificities of Penicillium simplicissimumα-galactosidases. Enzyme Microb. Technol. 22: 192-198.160. Luthi, E., D. R. Love, J. McAnulty, C. Wallace, P. A. Caughey, D. Saul, and P. L. Bergquist. 1990.Cloning, sequence analysis, and expression of genes encoding xylan-degrading enzymes from the ther-mophile “Caldocellum saccharolyticum”. Appl. Environ. Microbiol. 56(4): 1017-1024.161. Ly, H. D., and S. G. Withers. 1999. Mutagenesis of glycosidases. Annu. Rev. Biochem. 68: 487-522.162. Lüdtke, M. 1927. Zur Kenntnins der pflanzlichen Zellmembran. Über die Kohlenhydrate desSteinnußsamens. Ann. Chem. 456: 201-224.163. Macarrón, R., C. Acebal, M. P. Castillón, and M. Claeyssens. 1996. Mannanase activity ofEndoglucanase III from Trichoderma reesei QM9414. Biotechnol. Lett. 18(5): 599-602.164. Mach, R.L., and S. Zeilinger. 1998. Genetic transformation of Trichoderma and Gliacladium, p. 225-241. In C.P. Kubicek and G. E. Harman (ed.), Trichoderma & Gliocladikum, vol. 1. Taylor & Francis Ltd.,London.165. Mackenzie, L. F., G. S. Brooke, J. F. Cutfield, P. A. Sullivan, and S. G. Withers. 1997. Identifica-tion of Glu-330 as the catalytic nucleophile of Candida albicans exo-β-(1,3)-glucanase. J. Biol. Chem.272(6): 3161-3167.166. Mackie, W., and R. D. Preston. 1968. The occurrence of mannan microfibrils in the green algaeCodium fragile and Acetabularia crenulata. Planta. 79: 249-253.167. Mackie, W., and D. B. Sellen. 1969. Degree of polymerization and polydispersity of mannan from thecell wall of the green seaweed Codium fragile. Polymer. 10(8): 621-632.168. Malburg, S. R., L. M. Malburg, Jr., T. Liu, A. H. Iyo, and C. W. Forsberg. 1997. Catalytic proper-ties of the cellulose-binding endoglucanase F from Fibrobacter succinogenes S85. Appl. Environ. Microbiol.63(6): 2449-2453.169. Marchessault, R. H., and A. Sarko. 1967. X-ray structure of polysaccharides. Advan. Carbohyd.Chem. Biochem. 22: 421-482.170. Margolles-Clark, E., M. Ilmén, and M. Penttilä. 1997. Expression patterns of ten hemicellulase genesof the filamentous fungus Trichoderma reesei on various carbon sources. J. Biotechnol. 57(1-3): 167-179.

63
171. Margolles-Clark, E., M. Tenkanen, H. Söderlund, and M. Penttilä. 1996. Acetyl xylan esterasefrom Trichoderma reesei contains an active-site serine residue and a cellulose-binding domain. Eur. J. Biochem.237(3): 553-560.172. Marraccini, P., W. J. Rogers, C. Allard, M. L. Andre, V. Caillet, N. Lacoste, F. Lausanne, and S.Michaux. 2001. Molecular and biochemical characterization of endo-β-mannanases from germinating cof-fee (Coffea arabica) grains. Planta. 213(2): 296-308.173. Mattinen, M. L., M. Kontteli, J. Kerovuo, M. Linder, A. Annila, G. Lindeberg, T. Reinikainen,and T. Drakenberg. 1997. Three-dimensional structures of three engineered cellulose-binding domains ofcellobiohydrolase I from Trichoderma reesei. Protein Sci. 6(2): 294-303.174. Mattinen, M. L., M. Linder, T. Drakenberg, and A. Annila. 1998. Solution structure of the cellulose-binding domain of endoglucanase I from Trichoderma reesei and its interaction with cello-oligosaccharides.Eur. J. Biochem. 256(2): 279-286.175. Mattinen, M. L., M. Linder, A. Teleman, and A. Annila. 1997. Interaction between cellohexaose andcellulose binding domains from Trichoderma reesei cellulases. FEBS Lett. 407(3): 291-296.176. McCleary, B. V. 1982. Purification and properties of a β-D-mannoside mannohydrolase from guar.Carbohydr. Res. 101(1): 75-92.177. McCleary, B. V. 1983. β-D-Mannosidase from Helix pomatia. Carbohydr. Res. 111(2): 297-310.178. McCleary, B. V. 1983. Enzymic interactions in the hydrolysis of galactomannan in germinating guar:the role of exo-β-mannanase. Phytochemistry. 22(3): 649-658.179. McCleary, B. V. 1985. The Fine Structure of Carob and Guar Galactomannans. Carbohydr. Res. 139:237-260.180. McCleary, B. V. 1988. β-D-Mannanse. Methods Enzymol. 160: 596-610.181. McCleary, B. V. 1988. Exo-β-D-mannanase from Cyamopsis tetragonolobus guar seed. MethodsEnzymol. 160(Biomass, Pt. A): 589-595.182. McCleary, B. V., and N. K. Matheson. 1983. Action Patterns and Substrate-Binding Requirements ofβ-D-Mannanase with Mannosaccharides and Mannan-type Polysaccharides. Carbohydr. Res. 119: 191-219.183. McCutchen, C. M., G. D. Duffaud, P. Leduc, A. R. H. Petersen, A. Tayal, S. A. Khan, and R. M.Kelly. 1996. Characterization of extremely thermostable enzymic breakers (α-1,6-galactosidase and β-1,4-mannanase) from the hyperthermophilic bacterium Thermotoga neapolitana 5068 for hydrolysis of guargum. Biotechnol. Bioeng. 52(2): 332-339.184. Meier, H. 1958. On the structure of cell walls and cell wall mannans from ivory nuts and from dates.Biochim. Biophys. Acta. 28: 229-240.185. Meier, H., and J. S. G. Reid. 1982. Reserve Polysaccharides Other Than Starch in Higher Plants, p.418-471. In F. A. Loewus and W. Tanner (ed.), Encyclopaedia of Plant Physiology New Ser., vol. 13A.Springer Verlag, Berlin.186. Mendoza, N. S., M. Arai, T. Kawaguchi, T. Yoshida, and L. M. Joson. 1994. Purification and prop-erties of mannanase from Bacillus subtilis. World J. Microbiol. Biotechnol. 10(5): 551-555.187. Mendoza, N. S., M. Arai, K. Sugimoto, M. Ueda, T. Kawaguchi, and L. M. Joson. 1995. Cloning andsequencing of β-mannanase gene from Bacillus subtilis NM-39. Biochim. Biophys. Acta. 1243(3): 552-554.188. Millane, R. P., and T. L. Hendrixson. 1994. Crystal structures of mannan and glucomannans. Carbohydr.Polym. 25(4): 245-251.189. Millward-Sadler, S. J., K. Davidson, G. P. Hazlewood, G. W. Black, H. J. Gilbert, and J. H. Clarke.1995. Novel cellulose-binding domains, NodB homologues and conserved modular architecture in xylanasesfrom the aerobic soil bacteria Pseudomonas fluorescens subsp. cellulosa and Cellvibrio mixtus. Biochem. J.312(Pt 1): 39-48.190. Millward-Sadler, S. J., J. Hall, G. W. Black, G. P. Hazlewood, and H. J. Gilbert. 1996. Evidencethat the Piromyces gene family encoding endo-1,4-mannanases arose through gene duplication. FEMSMicrobiol. Lett. 141(2-3): 183-188.191. Millward-Sadler, S. J., D. M. Poole, B. Henrissat, G. P. Hazlewood, J. H. Clarke, and H. J. Gil-bert. 1994. Evidence for a general role for high-affinity non-catalytic cellulose binding domains in microbialplant cell wall hydrolases. Mol. Microbiol. 11(2): 375-382.192. Montiel, M. D., M. Hernandez, J. Rodriguez, and M. E. Arias. 2002. Evaluation of an endo-β-mannanase produced by Streptomyces ipomoea CECT 3341 for the biobleaching of pine kraft pulps. Appl.Microbiol. Biotechnol. 58(1): 67-72.193. Mäntylä, A., M. Paloheimo, and P. Suominen. 1998. Industrial mutants and recombinant strains of

64
Trichoderma reesei, p. 291-309. In G. E. Harman and C. P. Kubicek (ed.), Trichoderma & Gliocladium, vol.2. Taylor & Francis Ltd., London.194. Navas, J., and P. Beguin. 1992. Site-directed mutagenesis of conserved residues of Clostridiumthermocellum endoglucanase CelC. Biochem. Biophys. Res. Commun. 189(2): 807-812.195. Neustroev, K. N., A. S. Krylov, L. M. Firsov, and O. N. Abroskina. 1991. Isolation and properties ofβ-mannosidase from Aspergillus awamori. Biokhimiya. 56: 1406-1412.196. Nevalainen, H., and D. Neethling. 1998. The Safety of Trichoderma and Gliocladium, p. 193-205. InC. P. Kubicek and G. E. Harman (ed.), Trichoderma & Gliocladium, vol. 1. Taylor & Francis Ltd., London.197. Nieduszynski, I., and R. H. Marchessault. 1972. Crystal Structure of Poly-β,D-(1 ->4')-mannose:Mannan I. Can. J. Chem. 50(13): 2130-2138.198. Nishinari, K., P. A. Williams, and G. O. Phillips. 1992. Review of the physico-chemical characteris-tics and properties of konjac mannan. Food Hydrocolloids. 6(2): 199-222.199. Nonogaki, H., O. H. Gee, and K. J. Bradford. 2000. A germination-specific endo-β-mannanase gene isexpressed in the micropylar endosperm cap of tomato seeds. Plant. Physiol. 123(4): 1235-1246.200. Nonogaki, H., and Y. Morohashi. 1999. Temporal and spatial pattern of the development of endo-β-mannanase activity in germinating and germinated lettuce seeds. J. Exp. Bot. 50(337): 1307-1313.201. Notenboom, V., A. B. Boraston, P. Chiu, A. C. J. Freelove, D. G. Kilburn, and D. R. Rose. 2001.Recognition of Cello-oligosaccharides by a Family 17 Carbohydrate-binding Module: An X-RayCrystallographic, Thermodynamic and Mutagenic Study. J. Mol. Biol. 314(4): 797-806.202. Notenboom, V., A. B. Boraston, D. G. Kilburn, and D. R. Rose. 2001. Crystal structures of thefamily 9 carbohydrate-binding module from Thermotoga maritima xylanase 10A in native and ligand-boundforms. Biochemistry. 40(21): 6248-6256.203. Oda, Y., and K. Tonomura. 1996. Characterization of β-mannanase and β-mannosidase secretedfrom the yeast Trichosporon cutaneum JCM 2947. Lett. Appl. Microbiol. 22(2): 173-178.204. Parker, K. N., S. Chhabra, D. Lam, M. A. Snead, E. J. Mathur, and R. M. Kelly. 2001. β-Mannosidase from Thermotoga species. Methods Enzymol. 330: 238-246.205. Parker, K. N., S. R. Chhabra, D. Lam, W. Callen, G. D. Duffaud, M. A. Snead, J. M. Short, E. J.Mathur, and R. M. Kelly. 2001. Galactomannanases Man2 and Man5 from Thermotoga species: growthphysiology on galactomannans, gene sequence analysis, and biochemical properties of recombinant en-zymes. Biotechnol. Bioeng. 75(3): 322-333.206. Penttilä, M. 1998. Heterologous protein production in Trichoderma, p. 365-382. In G. E. Harman andC. P. Kubicek (ed.), Trichoderma & Gliocladium, vol. 2. Taylor & Francis Ltd., London.207. Penttilä, M., H. Nevalainen, M. Rättö, E. Salminen, and J. Knowles. 1987. A versatile transforma-tion system for the cellulolytic filamentous fungus Trichoderma reesei. Gene. 61(2): 155-164.208. Percival, E. 1970. Algal polysaccharides, p. 537-568. In A. Herp (ed.), The Carbohydrates, vol. 2B.Academic Press, New York.209. Politz, O., M. Krah, K. K. Thomsen, and R. Borriss. 2000. A highly thermostable endo-β-mannanasefrom the marine bacterium Rhodothermus marinus. Appl. Microbiol. Biotechnol. 53(6): 715-721.210. Puls, J., and J. Schuseil. 1993. Chemistry of hemicelluloses: relationship between hemicellulosestructure and enzymes required for hydrolysis, p. 1-27. In M. P. Coughlan and G. P. Hazlewood (ed.), Hemi-cellulose and Hemicellulases, vol. 4. Portland Press Res. Monogr., London.211. Punt, P. J., M. A. Dingemanse, A. Kuyvenhoven, R. D. Soede, P. H. Pouwels, and C. A. van denHondel. 1990. Functional elements in the promoter region of the Aspergillus nidulans gpdA gene encodingglyceraldehyde-3-phosphate dehydrogenase. Gene. 93(1): 101-109.212. Purchon, R. D. 1977. The biology of the mollusca, 2nd ed, vol. 57. Pergamon Press Ltd., Oxford.213. Py, B., I. Bortoli-German, J. Haiech, M. Chippaux, and F. Barras. 1991. Cellulase EGZ of Erwiniachrysanthemi: structural organization and importance of His98 and Glu133 residues for catalysis. ProteinEng. 4(3): 325-333.214. Quiocho, F. A., and N. K. Vyas. 1999. Atomic Interactions Between Proteins/Enzymes and Carbohy-drates, p. 441-457. In M. Hecht (ed.), Bioorganic chemistry: Carbohydrates, vol. 1. Oxford UniversityPress, Oxford.215. Raghothama, S., P. J. Simpson, L. Szabo, T. Nagy, H. J. Gilbert, and M. P. Williamson. 2000.Solution structure of the CBM10 cellulose binding module from Pseudomonas xylanase A. Biochemistry.39(5): 978-984.216. Regalado, C., B. E. Garcia-Almendarez, L. M. Venegas-Barrera, A. Tellez-Jurado, G. Rodriguez-Serrano, S. Huerta-Ochoa, and J. R. Whitaker. 2000. Production, partial purification and properties of β-

65
mannanases obtained by solid substrate fermentation of spent soluble coffee wastes and copra paste usingAspergillus oryzae and Aspergillus niger. J. Sci. Food Agric. 80(9): 1343-1350.217. Reid, J. S. G. 1985. Galactomannans, p. 265-288. In P. M. Dey and R. A. Dixon (ed.), Biochemistry ofStorage Carbohydrates in Green Plants. Academic Press, London.218. Reinikainen, T., L. Ruohonen, T. Nevanen, L. Laaksonen, P. Kraulis, T. A. Jones, J. K. Knowles,and T. T. Teeri. 1992. Investigation of the function of mutated cellulose-binding domains of Trichodermareesei cellobiohydrolase I. Proteins. 14(4): 475-482.219. Reinikainen, T., O. Teleman, and T. T. Teeri. 1995. Effects of pH and high ionic strength on theadsorption and activity of native and mutated cellobiohydrolase I from Trichoderma reesei. Proteins. 22(4):392-403.220. Rol, F. 1973. Locust bean gum, p. 323-327. In R. Whistler and J. M. BeMiller (ed.), Industrial gums.Academic Press, New York.221. Rättö, M., and K. Poutanen. 1988. Production of Mannan-Degrading Enzymes. Biotechnol. Lett.10(9): 661-664.222. Sabini, E. 2001. Structural Investigation of hemicellulose degrading enzymes. Doctoral thesis. Uni-versity of York, York.223. Sabini, E., H. Schubert, G. Murshudov, K. S. Wilson, M. Siika-Aho, and M. Penttilä. 2000. Thethree-dimensional structure of a Trichoderma reesei β-mannanase from glycoside hydrolase family 5. ActaCrystallogr., Sect. D: Biol. Crystallogr. 56(Pt 1): 3-13.224. Sachslehner, A., G. Foidl, N. Foidl, G. Gübitz, and D. Haltrich. 2000. Hydrolysis of isolated coffeemannan and coffee extract by mannanases of Sclerotium rolfsii. J. Biotechnol. 80(2): 127-134.225. Sachslehner, A., and D. Haltrich. 1999. Purification and some properties of a thermostable acidic endo-β-1,4-D-mannanase from Sclerotium (Athelia) rolfsii. FEMS Microbiol. Lett. 177(1): 47-55.226. Saka, S. 2001. Chemical Composition and distribution, p. 51-82. In D. N. S. Hon and N. Shiraishi(ed.), Wood and Cellulosic Chemistry, 2nd ed. Marcel Dekker Inc., New York.227. Salmén, L., and A.-M. Olsson. 1998. Interaction Between Hemicelluloses, Lignin and Cellulose:Structure-Property Relationships. J. Pulp Pap. Sci. 24(3): 99-103.228. Saloheimo, A., M. Ilmén, N. Aro, E. Margolles-Clark, and Penttilä. 1998. Regulatory MechanismsInvolved in Expression of Extracellular Hydrolytic Enzymes of Trichoderma reesei, p. 267-273. In M.Claeyssens, W. Nerinckx, and P. K. (ed.), Carbohydrases from Trichoderma reesei and other microorgan-isms: Structure, Biochemistry, Genetics and Applications, vol. 219. Royal Society of Chemistry, Cambridge.229. Setati, M. E., P. Ademark, W. H. van Zyl, B. Hahn-Hagerdal, and H. Stålbrand. 2001. Expressionof the Aspergillus aculeatus endo-beta-1,4-mannanase encoding gene (man1) in Saccharo myces cerevisiaeand characterization of the recombinant enzyme. Protein Expr. Purif. 21(1): 105-114.230. Shoham, Y., R. Lamed, and E. A. Bayer. 1999. The cellulosome concept as an efficient microbialstrategy for the degradation of insoluble polysaccharides. Trends Microbiol. 7(7): 275-281.231. Simpson, P. J., D. N. Bolam, A. Cooper, A. Ciruela, G. P. Hazlewood, H. J. Gilbert, and M. P.Williamson. 1999. A family IIb xylan-binding domain has a similar secondary structure to a homologousfamily IIa cellulose-binding domain but different ligand specificity. Structure Fold. Des. 7(7): 853-864.232. Simpson, P. J., H. Xie, D. N. Bolam, H. J. Gilbert, and M. P. Williamson. 2000. The structural basisfor the ligand specificity of family 2 carbohydrate-binding modules. J. Biol. Chem. 275(52): 41137-41142.233. Sinnott, M. L. 1990. Catalytic Mechanism of Enzymic Glycosyl Transfer. Chem. Rev. 90: 1171-1202.234. Sjöström, E. 1993. Wood chemistry, fundamentals and applications, 2nd ed. Academic Press, SanDiego.235. Sone, Y., and A. Misaki. 1978. Purification and characterization of a β-D-mannosidase and β-N-acetyl-D-hexosaminidase of Tremella fuciformis. J. Biochem. 83: 1135-1144.236. Srisodsuk, M., J. Lehtio, M. Linder, E. Margolles-Clark, T. Reinikainen, and T. T. Teeri. 1997.Trichoderma reesei cellobiohydrolase I with an endoglucanase cellulose-binding domain: action on bacte-rial microcrystalline cellulose. J. Biotechnol. 57(1-3): 49-57.237. Stoll, D., A. Boraston, H. Stålbrand, B. W. McLean, D. G. Kilburn, and R. A. Warren. 2000.Mannanase Man26A from Cellulomonas fimi has a mannan-binding module. FEMS Microbiol. Lett. 183(2):265-269.238. Stoll, D., S. He, S. G. Withers, and R. A. J. Warren. 2000. Identification of Glu-519 as the catalyticnucleophile in β-mannosidase 2A from Cellulomonas fimi. Biochem. J. 351(3): 833-838.

66
239. Stoll, D., H. Stålbrand, and R. A. Warren. 1999. Mannan-degrading enzymes from Cellulomonasfimi. Appl. Environ. Microbiol. 65(6): 2598-2605.240. Stoll, D., H. Stålbrand, and R. A. J. Warren. 2000. Hydrolysis of mannans by Cellulomons fimi, p.91-98. In M. E. Himmel, J. O. Baker, and J. N. Saddler (ed.), Glycosyl Hydrolases for Biomass Conversion,vol. 769. American Chemical Society, Washington, D. C.241. Ståhlberg, J., G. Johansson, and G. Pettersson. 1993. Trichoderma reesei has no true exo-cellulase:all intact and truncated cellulases produce new reducing end groups on cellulose. Biochim. Biophys. Acta.1157(1): 107-113.242. Stålbrand, H. 1995. Hemicellulose-degrading enzymes from fungi: Characterization of β−mannanaseand the man1 gene of Trichoderma reesei. Doctoral thesis. Lund University, Lund.243. Stålbrand, H. 2002. Enzymology of endo-1,4-β-Mannanases, p. 961-969. In Whitaker, Voragen, andWong (ed.), Handbook of Food Enzymology. Marcel Dekker, Inc., New York.244. Stålbrand, H., A. Saloheimo, J. Vehmaanpera, B. Henrissat, and M. Penttilä. 1995. Cloning andexpression in Saccharomyces cerevisiae of a Trichoderma reesei β-mannanase gene containing a cellulosebinding domain. Appl. Environ. Microbiol. 61(3): 1090-1097.245. Stålbrand, H., M. Siika-aho, M. Tenkanen, and L. Viikari. 1993. Purification and characterizationof two β-mannanases from Trichoderma reesei. J. Biotechnol. 29: 229-242.246. Sugahara, K., T. Okumura, and I. Yamashina. 1972. Purification of β-mannosidase from snail,Achatina fulica and its action on glycopeptides. Biochim. Biophys. Acta. 268: 488-496.247. Sunna, A., M. D. Gibbs, and P. L. Bergquist. 2001. Identification of novel β-mannan- and β-glucan-binding modules: evidence for a superfamily of carbohydrate-binding modules. Biochem. J. 356(Pt 3): 791-798.248. Sunna, A., M. D. Gibbs, C. W. Chin, P. J. Nelson, and P. L. Bergquist. 2000. A gene encoding anovel multidomain β-1,4-mannanase from Caldibacillus cellulovorans and action of the recombinant en-zyme on kraft pulp. Appl. Environ. Microbiol. 66(2): 664-670.249. Suurnäkki, A., T. Clark, R. Allison, J. Buchert, and L. Viikari. 1996. Mannanase aided bleaching ofsoft-wood kraft pulp, p. 69-74. In K. Messner and E. Srebotnik (ed.), Biotechnology in Pulp and PaperIndustry - Advances in Applied and Fundamental Research. WUA Universitätsverlag, Vienna.250. Szabo, L., S. Jamal, H. Xie, S. J. Charnock, D. N. Bolam, H. J. Gilbert, and G. J. Davies. 2001.Structure of a family 15 carbohydrate-binding module in complex with xylopentaose: Evidence that xylanbinds in an approximate three-fold helical conformation. J. Biol. Chem. 11: 49061-49065.251. Takada, G., T. Kawaguchi, T. Kaga, J. Sumitani, and M. Arai. 1999. Cloning and sequencing of β-mannosidase gene from Aspergillus aculeatus NO. F-50. Biosci. Biotechnol. Biochem. 63: 206-209.252. Talbot, G., and J. Sygusch. 1990. Purification and characterization of thermostable β-mannanase andα-galactosidase from Bacillus stearothermophilus. Appl. Environ. Microbiol. 56(11): 3505-3510.253. Tamaru, Y., T. Araki, H. Amagoi, H. Mori, and T. Morishita. 1995. Purification and characteriza-tion of an extracellular β-1,4-mannanase from a marine bacterium, Vibrio sp. strain MA-138. Appl. Environ.Microbiol. 61(12): 4454-4458.254. Tamaru, Y., T. Araki, T. Morishita, T. Kimura, K. Sakka, and K. Ohmiya. 1997. Cloning, DNAsequencing, and expression of the β-1,4-mannanase gene from a marine bacterium, Vibrio sp. strain MA-138. J. Ferment. Bioeng. 83(2): 201-205.255. Tamaru, Y., and R. H. Doi. 2000. The engL gene cluster of Clostridium cellulovorans contains a genefor cellulosomal ManA. J. Bacteriol. 182(1): 244-247.256. Tang, C. M., L. D. Waterman, M. H. Smith, and C. F. Thurston. 2001. The cel4 gene of Agaricusbisporus encodes a β-mannanase. Appl. Environ. Microbiol. 67(5): 2298-2303.257. Teeri, T., T. Reinikainen, L. Ruohonen, T. A. Jones, and J. K. C. Knowles. 1992. Domain Functionin Trichoderma reesei Cellobiohydrolases. J. Biotechnol. 24(2): 169-176.258. Tenkanen, M., J. Buchert, and L. Viikari. 1995. Binding of hemicellulases on isolated polysaccha-ride substrates. Enzyme Microb. Technol. 17: 499-505.259. Tenkanen, M., M. Makkonen, M. Perttula, L. Viikari, and A. Teleman. 1997. Action of Trichodermareesei mannanase on galactoglucomannan in pine kraft pulp. J. Biotechnol. 57(1-3): 191-204.260. Tenkanen, M., J. Puls, M. Rattö, and L. Viikari. 1993. Enzymic deacetylation of galactoglucomannans.Appl. Microbiol. Biotechnol. 39(2): 159-165.261. Tenkanen, M., J. Thornton, and L. Viikari. 1995. An acetylglucomannan esterase of Aspergillusoryzae; purification, characterization and role in the hydrolysis of O-acetyl-galactoglucomannan. J. Biotechnol.42(3): 197-206.

67
262. Timell, T. E. 1965. Wood Hemicelluloses: Part II. Adv. Carbohydr. Chem. 20: 409-483.263. Timell, T. E. 1967. Recent Progress in the chemistry of Wood Hemicellulose. Wood Sci. Technol. 1:45-70.264. Todd, A. E., C. A. Orengo, and J. M. Thornton. 1999. Evolution of protein function, from a struc-tural perspective. Curr. Opin. Chem. Biol. 3(5): 548-556.265. Tomme, P., A. L. Creagh, D. G. Kilburn, and C. A. Haynes. 1996. Interaction of polysaccharideswith the N-terminal cellulose-binding domain of Cellulomonas fimi CenC. 1. Binding specificity and calori-metric analysis. Biochemistry. 35(44): 13885-13894.266. Tomme, P., E. Kwan, N. R. Gilkes, D. G. Kilburn, and R. A. J. Warren. 1996. Characterization ofCenC, an enzyme from Cellulomonas fimi with both endo- and exoglucanase activities. J. Bacteriol. 178(14):4216-4223.267. Tomme, P., H. Van Tilbeurgh, G. Pettersson, J. Van Damme, J. Vandekerckhove, J. Knowles, T.Teeri, and M. Claeyssens. 1988. Studies of the cellulolytic system of Trichoderma reesei QM 9414. Analy-sis of domain function in two cellobiohydrolases by limited proteolysis. Eur. J. Biochem. 170(3): 575-581.268. Tomme, P., R. A. Warren, and N. R. Gilkes. 1995. Cellulose hydrolysis by bacteria and fungi. Adv.Microb. Physiol. 37: 1-81.269. Tomme, P., R. A. J. Warren, R. C. J. Miller, D. G. Kilburn, and N. R. Gilkes. 1995. Cellulose-Binding Domains: Classification and Properties, p. 142-163. In J. M. Saddler and M. Penner (ed.), EnzymaticDegradation of Insoluble Polysaccharides. American Chemical Society, Washington, DC.270. Toorop, P. E., A. C. van Aelst, and H. W. Hilhorst. 2000. The second step of the biphasic endospermcap weakening that mediates tomato (Lycopersicon esculentum) seed germination is under control of ABA.J. Exp. Bot. 51(349): 1371-1379.271. Tormo, J., R. Lamed, A. J. Chirino, E. Morag, E. A. Bayer, Y. Shoham, and T. A. Steitz. 1996.Crystal structure of a bacterial family-III cellulose-binding domain: a general mechanism for attachment tocellulose. EMBO J. 15(21): 5739-5751.272. Torto, N., T. Buttler, L. Gorton, G. Marko-Varga, H. Stålbrand, and F. Tjerneld. 1995. Monitor-ing of enzymic hydrolysis of ivory nut mannan using online microdialysis sampling and anion-exchangechromatography with integrated pulsed electrochemical detection. Anal. Chim. Acta. 313(1-2): 15-24.273. Torto, N., G. Marko-Varga, L. Gorton, H. Stålbrand, and F. Tjerneld. 1996. Online quantitation ofenzymic mannan hydrolyzates in small-volume bioreactors by microdialysis sampling and column liquid chro-matography-integrated pulsed electrochemical detection. J. Chromatogr. A. 725(1): 165-175.274. Waino, M., and K. Ingvorsen. 1999. Production of halostable β-mannanase and β-mannosidase bystrain NN, a new extremely halotolerant bacterium. Appl. Microbiol. Biotechnol. 52(5): 675-680.275. Wan, C., J. Muldry, Y. Li, and Y. Lee. 1976. β-Mannosidase from the mushroom Polyporous sulfureus.J. Biol. Chem. 251: 4384-4388.276. van Tilbeurgh, H., P. Tomme, M. Claeyssens, R. Bhikhabhai, and G. Pettersson. 1986. Limitedproteolysis of the cellobiohydrolase I from Trichoderma reesei. FEBS. 204(2): 223-227.277. Wang, Q., D. Tull, A. Meinke, N. R. Gilkes, R. A. Warren, R. Aebersold, and S. G. Withers. 1993.Glu280 is the nucleophile in the active site of Clostridium thermocellum CelC, a family A endo-β-1,4-glucanase. J. Biol. Chem. 268(19): 14096-14102.278. Warren, R. A. 1996. Microbial hydrolysis of polysaccharides. Annu. Rev. Microbiol. 50: 183-212.279. Viikari, L., J. Pere, A. Suurnäkki, T. Oksanen, and J. Buchert. 1998. Use of cellulases in pulp andpaper applications, p. 245-254. In M. Claeyssens, W. Nerinckx, and K. Piens (ed.), Carbohydrases fromTrichoderma reesei and other microorganisms: Structure, Biochemistry, Genetics and Applications, vol.219. Royal Society of Chemistry, Cambridge.280. Viikari, L., M. Tenkanen, J. Buchert, M. Rättö, M. Bailey, M. Siika-aho, and M. Linko. 1993.Hemicellulases for Industrial Applications, p. 131-182. In J. N. Saddler (ed.), Bioconversion of forest andagricultural plant residues, vol. 9. C. A. B. International, Wallingford, UK.281. Withers, S. G. 2001. Mechanisms of glycosyl transferases and hydrolases. Carbohydr. Polym. 44(4):325-337.282. Wolfenden, R., X. Lu, and G. Young. 1998. Spontaneous Hydrolysis of Glycosides. J. Am. Chem.Soc. 120(27): 6814-6815.283. Wolfrom, M. L., M. L. Layer, and D. L. Patin. 1961. Carbohydrates of the coffee bean. II. Isolationand characterization of a mannan. J. Org. Chem. 26: 4533-4535.284. Wong, K. K. Y., and J. N. Saddler. 1993. Applications of hemicellulases in the food, feed, and pulp

68
and paper industries, p. 127-143. In M. P. Coughlan and G. P. Hazlewood (ed.), Hemicellulose andHemicellulases, vol. 4. Portland Press Res. Monogr., London.284. Wong, K. K. Y., and J. N. Saddler. 1993. Applications of hemicellulases in the food, feed, and pulpand paper industries, p. 127-143. In M. P. Coughlan and G. P. Hazlewood (ed.), Hemicellulose andHemicellulases, vol. 4. Portland Press Res. Monogr., London.285. Xie, H., D. N. Bolam, T. Nagy, L. Szabo, A. Cooper, P. J. Simpson, J. H. Lakey, M. P. Williamson,and H. J. Gilbert. 2001. Role of hydrogen bonding in the interaction between a xylan binding module andxylan. Biochemistry. 40(19): 5700-5707.286. Xu, B., I. I. Munoz, J. C. Janson, and J. Ståhlberg. 2002. Crystallization and X-ray analysis ofnative and selenomethionyl β-mannanase Man5A from blue mussel, Mytilus edulis, expressed in Pichiapastoris. Acta Crystallogr., Sect D: Biol. Crystallogr. 58(Pt 3): 542-545.287. Xu, B., D. Sellos, and J. C. Janson. 2002. Cloning and expression in Pichia pastoris of a blue mussel(Mytilus edulis) β-mannanase gene. Eur. J. Biochem. 269(6): 1753-1760.288. Xu, G. Y., E. Ong, N. R. Gilkes, D. G. Kilburn, D. R. Muhandiram, M. Harris-Brandts, J. P.Carver, L. E. Kay, and T. S. Harvey. 1995. Solution structure of a cellulose-binding domain fromCellulomonas fimi by nuclear magnetic resonance spectroscopy. Biochemistry. 34(21): 6993-7009.289. Yagüe, E., M. Mehak-Zunic, L. Morgan, D. A. Wood, and C. F. Thurston. 1997. Expression ofCEL2 and CEL4, two proteins from Agaricus bisporus with similarity to fungal cellobiohydrolase I and β-mannanase, respectively, is regulated by the carbon source. Microbiology (Reading, U. K.). 143(Pt 1): 239-244.290. Yamamoto, K., S.-C. Li, and Y.-T. Li. 2000. Microbial glycosidases. Carbohydr. Chem. Biol. 3: 497-511.291. Yamaura, I., and T. Matsumoto. 1993. Purification and some properties of endo-1,4-β−D-mannanasefrom a mud snail, Pomacea insularus (de Ordigny). Biosci. Biotechnol. Biochem. 57(8): 1316-1319.292. Yamaura, I., Y. Nozaki, T. Matsumoto, and T. Kato. 1996. Purification and some properties of anendo-1,4-β-D-mannanase from a marine mollusc, Littorina brevicula. Biosci. Biotechnol. Biochem. 60(4):674-676.293. Yoshida, S., Y. Sako, and A. Uchida. 1998. Cloning, sequence analysis, and expression in Escherichiacoli of a gene coding for an enzyme from Bacillus circulans K-1 that degrades guar gum. Biosci. Biotechnol.Biochem. 62(3): 514-520.294. Yosida, S., Y. Sako, and A. Uchida. 1997. Purification, properties, and N-terminal amino acid se-quences of guar gum-degrading enzyme from Bacillus circulans K-1. Biosci. Biotechnol. Biochem. 61(2):251-255.295.Yui, T., K. Ogawa, and A. Sarko. 1992. Packing analysis of carbohydrates and polysaccharides. Part18. Molecular and crystal structure of konjac glucomannan in the mannan II polymorphic form. Carbohydr.Res. 229(1): 41-55.296. Zakaria, M. M., M. Ashiuchi, S. Yamamoto, and T. Yagi. 1998. Optimization for β-mannanase produc-tion of a psychrophilic bacterium, Flavobacterium sp. Biosci. Biotechnol. Biochem. 62(4): 655-660.297. Åkerholm, M., and L. Salmén. 2001. Interactions between wood polymers studied by dynamic FT-IR spectroscopy. Polymer. 42(3): 963-969.

I
69

Journal of Biotechnology 75 (1999) 281–289
Hydrolytic properties of a b-mannosidase purified fromAspergillus niger
Pia Ademark a, Jon Lundqvist a, Per Hagglund a, Maija Tenkanen b,Nelson Torto c, Folke Tjerneld a, Henrik Stalbrand a,*
a Department of Biochemistry, Center for Chemistry and Chemical Engineering, Lund Uni6ersity, PO Box 124,S-221 00 Lund, Sweden
b VTT Biotechnology and Food Research, PO Box 1500, FIN-02044 VTT, Espoo, Finlandc Department of Analytical Chemistry, Center for Chemistry and Chemical Engineering, Lund Uni6ersity, PO Box 124,
S-221 00 Lund, Sweden
Received 25 January 1999; received in revised form 1 July 1999; accepted 6 July 1999
Abstract
A b-mannosidase was purified to homogeneity from the culture filtrate of Aspergillus niger. A specific activity of500 nkat mg−1 and a 53-fold purification was achieved using ammonium sulfate precipitation, anion-exchangechromatography, and gel filtration. The isolated enzyme has an isoelectric point of 5.0 and appears to be a dimercomposed of two 135-kDa subunits. It is a glycoprotein and contains 17% N-linked carbohydrate by weight. Maximalactivity was observed at pH 2.4–5.0 and at 70°C. The b-mannosidase hydrolyzed b-1,4-linked manno-oligosaccha-rides of degree of polymerization (DP) 2–6 and also released mannose from polymeric ivory nut mannan andgalactomannan. The Km and Vmax values for p-nitrophenyl-b-D-mannopyranoside were 0.30 mM and 500 nkat mg−1,respectively. Hydrolysis of D-galactose substituted manno-oligosaccharides showed that the b-mannosidase was ableto cleave up to, but not beyond, a side group. An internal peptide sequence of 15 amino acids was highly similar tothat of an Aspergillus aculeatus b-mannosidase belonging to family 2 of glycosyl hydrolases. © 1999 Elsevier ScienceB.V. All rights reserved.
Keywords: Aspergillus niger ; b-Mannosidase; Manno-oligosaccharides; Hydrolysis
www.elsevier.com/locate/jbiotec
1. Introduction
b-Mannosidase (b-D-mannoside mannohydro-lase, EC 3.2.1.25) catalyzes the successive removal
of D-mannose residues from the nonreducing endof various b-1,4-linked manno-oligosaccharides. Itis essential for the complete hydrolysis of plantpolysaccharides such as galacto(gluco)mannanand mannan, and readily converts the manno-oligosaccharides produced by b-mannanase (EC3.2.1.78) to mannose (Reese and Shibata, 1965).Thus, b-mannosidase is important in the enzy-
* Corresponding author. Tel.: +46-46-222-8202; fax: +46-46-222-4534.
E-mail address: [email protected] (H.Stalbrand)
0168-1656/99/$ - see front matter © 1999 Elsevier Science B.V. All rights reserved.
PII: S 0168 -1656 (99 )00172 -8

P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289282
matic saccharification of hemicellulose tomonomeric sugars for further conversion tochemicals and fuels. It also cleaves b-1,4-manno-sidic linkages in the carbohydrate chains of gly-coproteins and is frequently used for structuralstudies of these glycans. There is a great interestin using b-mannosidase and related enzymes inthe synthesis of oligosaccharides for medical andother purposes (Kobata, 1993). A b-mannosi-dase from Cellulomonas fimi was recently mutag-enized in order to improve the enzyme forsynthetic applications (Stoll, 1998), similarly ashas been done for other glycosidases (McKenzieet al., 1998). The lysosomal storage disease b-mannosidosis, which is caused by insufficient b-mannosidase activity, has also attractedattention to this enzyme (Neufeld, 1991).
Relatively few microbial b-mannosidases havebeen purified and characterized (reviewed by Vi-ikari et al., 1993), even though they have beenreported to occur in many microorganisms,plants, and animal tissues (Dey, 1978). Purifica-tion of a b-mannosidase from Aspergillus nigerwas first described by Elbein et al. (1977) andBouquelet et al. (1978). In the present worksome biochemical properties of a highly purifiedb-mannosidase from Aspergillus niger are de-scribed. Its activity towards different substrates,especially side group substituted oligosaccharidesand polymers, was investigated.
2. Materials and methods
2.1. Enzyme source
A. niger ATCC-46890 was obtained from theQM Culture Collection, Department of Botany,University of Massachusetts, Amherst, USA.The fungus was cultivated on galactomannan asdescribed previously (Ademark et al., 1998) andthe culture fluid was used as the source of b-mannosidase.
2.2. Acti6ity assays
b-Mannosidase, a-galactosidase, and b-glucosi-
dase activities were assayed at pH 4.5 with 2mM p-nitrophenyl-b-D-mannopyranoside (SigmaN-1268), 4 mM p-nitrophenyl-a-D-galactopyra-noside (Sigma N-0877), and 5 mM p-nitro-phenyl-b-D-glucopyranoside (Sigma N-7006),respectively, as described by Ratto andPoutanen (1988). All enzyme activities were ex-pressed in SI units (katals). A plate assay tech-nique with locust bean gum galactomannan assubstrate and Congo red staining (Stalbrand etal., 1993) was used to detect b-mannanaseactivity.
2.3. Gel electrophoresis
Sodium dodecylsulfate-polyacrylamide gelelectrophoresis (SDS-PAGE) under both reduc-ing (with b-mercaptoethanol) and nonreducingconditions and isoelectric focusing (IEF) wereperformed using the PhastSystem (PharmaciaBiotech, Uppsala, Sweden) in accordance withthe manufacturer’s instructions. Precast gels(PhastGel 4-15 and PhastGel IEF 4-6.5) wereobtained from Pharmacia, and proteins were vi-sualized with silver staining. Marker proteins(Low pI Kit, HMW-SDS and LMW CalibrationKits) from Pharmacia were run in parallel. Two-dimensional gel electrophoresis (IEF followed bySDS-PAGE) was carried out as described by Yuet al. (1994).
2.4. Protein assay
Protein concentrations were measured by theMicro BCA Protein Assay (Pierce, Rockford,IL), using bovine serum albumin (BSA) as stan-dard. All chromatographic runs were monitoredfor protein by absorbance at 280 nm.
2.5. Deglycosylation
The carbohydrate content of the b-mannosi-dase was estimated by treating it with N-glycosi-dase F (PNGase F, New England Biolabs,USA) as described by the manufacturer. Themolecular mass of the N-deglycosylated enzymewas subsequently determined by SDS-PAGE.

P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289 283
2.6. Internal amino acid sequence determination
An internal amino acid sequence determinationwas done by Innovagen AB (Lund, Sweden). Theb-mannosidase was digested with endoproteinaseLys C and the amino acid sequence of one of thepeptide fragments was determined by Edmandegradation, using an ABI 476A instrument (Fos-ter City, CA).
2.7. Purification of the b-mannosidase
Solid ammonium sulfate was added to 100 mlof the culture filtrate under slow stirring to obtain80% saturation. The precipitate was collected bycentrifugation and then dissolved in 20 ml sodiumacetate buffer, pH 4.5. Three ml of this solutionwas desalted and equilibrated with 20 mM ammo-nium acetate buffer, pH 7.5, by using a FastDesalting Column (Pharmacia). The sample wasthen directly loaded on a Resource™ Q 1 mlanion-exchange column (Pharmacia) equilibratedwith the same buffer. Proteins were eluted with alinear gradient (0–60%) of 1 M ammonium ac-etate, pH 7.5. Fractions of 0.25 ml were collectedand assayed for b-mannosidase, a-galactosidase,and b-mannanase activities. The eight fractionsmost active in b-mannosidase were pooled andfurther purified on a HiLoad™ 16/60 gel filtrationcolumn pre-packed with Superdex 200 pg (Phar-macia). Elution was achieved using 200 mM NaClin 100 mM sodium acetate buffer, pH 4.5, at aflow rate of 0.5 ml min−1. Fractions of 1 ml werecollected and assayed for b-mannosidase and a-galactosidase activities. Both ion-exchange chro-matography and gel filtration were performed atroom temperature using an FPLC System(Pharmacia).
2.8. pH and temperature optima and stabilities
The pH stability was investigated by incubatingthe purified b-mannosidase (2.5 mg ml−1) at dif-ferent pH values (pH 2.6–7: 50 mM citrate-phos-phate buffer, pH 7.5–8: 50 mM phosphate buffer)for 24 h at 50°C, and then measuring the residualactivity. The pH optimum was determined bymeasuring b-mannosidase activity at pH values
ranging from 2.6 to 8 using the same buffersystems as above. For determination of tempera-ture stability, purified b-mannosidase was incu-bated (2.5 mg ml−1) at different temperatures for24 h in citrate-phosphate buffer (50 mM, pH 5.0).The remaining activity was then measured at stan-dard conditions. Optimal temperature for b-man-nosidase activity was determined by assaying atdifferent temperatures. All samples were analyzedin duplicate.
2.9. Kinetic properties
The Michaelis–Menten constant (Km) and themaximal reaction velocity (Vmax) for the b-man-nosidase were determined by incubating it withp-nitrophenyl-b-D-mannopyranoside in concen-trations ranging from 0.1 to 4 mM. The reactionswere stopped after 5 min, when less than 5% ofthe substrate had been degraded, and the releaseof p-nitrophenol was measured at standard assayconditions. All assays were performed in dupli-cate. The linearity of the reaction was confirmedby measuring the amount of released p-nitrophe-nol at three different timepoints: 3, 5, and 7 min.Values for Km and Vmax were determined from aLineweaver–Burk plot.
2.10. Hydrolysis experiments
The ability of the b-mannosidase to hydrolyzedifferent oligosaccharides as well as polymericmannan and galactomannan was investigated.Linear b-1,4-linked manno-oligosaccharides ofDP 2 to 6, 61-a-D-galactosylmannotriose(GalMan3), 63,64-a-D-galactosylmannopentaose(Gal2Man5) and ivory nut (Phytelephas macro-carpa) mannan were purchased from Megazyme(Bray, Ireland). 61-a-D-galactosylmannobiose(GalMan2) and 4-b-D-glucosylmannose (GlcMan)were prepared as described in Tenkanen et al.(1997). Locust bean gum galactomannan (a linearb-1,4-linked mannan backbone substituted withgalactose side groups) was obtained from Sigma(St. Louis, MO). The substrate concentrationswere 2.5 mg ml−1 in 100 mM sodium acetatebuffer (pH 4.5), and the b-mannosidase dosagewas 2.5 mg ml−1 for locust bean gum hydrolysis
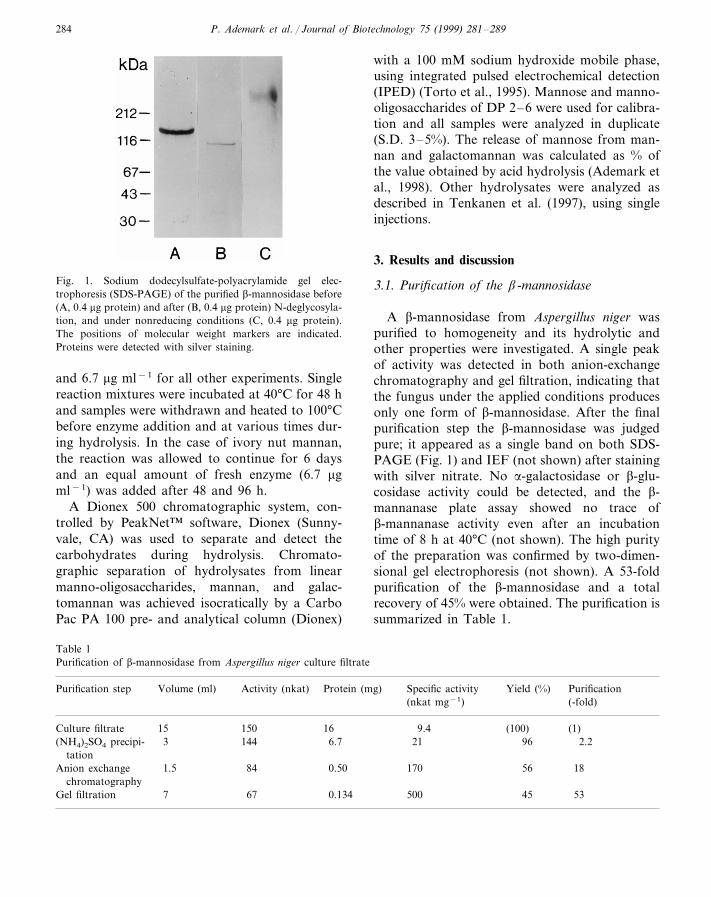
P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289284
Fig. 1. Sodium dodecylsulfate-polyacrylamide gel elec-trophoresis (SDS-PAGE) of the purified b-mannosidase before(A, 0.4 mg protein) and after (B, 0.4 mg protein) N-deglycosyla-tion, and under nonreducing conditions (C, 0.4 mg protein).The positions of molecular weight markers are indicated.Proteins were detected with silver staining.
with a 100 mM sodium hydroxide mobile phase,using integrated pulsed electrochemical detection(IPED) (Torto et al., 1995). Mannose and manno-oligosaccharides of DP 2–6 were used for calibra-tion and all samples were analyzed in duplicate(S.D. 3–5%). The release of mannose from man-nan and galactomannan was calculated as % ofthe value obtained by acid hydrolysis (Ademark etal., 1998). Other hydrolysates were analyzed asdescribed in Tenkanen et al. (1997), using singleinjections.
3. Results and discussion
3.1. Purification of the b-mannosidase
A b-mannosidase from Aspergillus niger waspurified to homogeneity and its hydrolytic andother properties were investigated. A single peakof activity was detected in both anion-exchangechromatography and gel filtration, indicating thatthe fungus under the applied conditions producesonly one form of b-mannosidase. After the finalpurification step the b-mannosidase was judgedpure; it appeared as a single band on both SDS-PAGE (Fig. 1) and IEF (not shown) after stainingwith silver nitrate. No a-galactosidase or b-glu-cosidase activity could be detected, and the b-mannanase plate assay showed no trace ofb-mannanase activity even after an incubationtime of 8 h at 40°C (not shown). The high purityof the preparation was confirmed by two-dimen-sional gel electrophoresis (not shown). A 53-foldpurification of the b-mannosidase and a totalrecovery of 45% were obtained. The purification issummarized in Table 1.
and 6.7 mg ml−1 for all other experiments. Singlereaction mixtures were incubated at 40°C for 48 hand samples were withdrawn and heated to 100°Cbefore enzyme addition and at various times dur-ing hydrolysis. In the case of ivory nut mannan,the reaction was allowed to continue for 6 daysand an equal amount of fresh enzyme (6.7 mgml−1) was added after 48 and 96 h.
A Dionex 500 chromatographic system, con-trolled by PeakNet™ software, Dionex (Sunny-vale, CA) was used to separate and detect thecarbohydrates during hydrolysis. Chromato-graphic separation of hydrolysates from linearmanno-oligosaccharides, mannan, and galac-tomannan was achieved isocratically by a CarboPac PA 100 pre- and analytical column (Dionex)
Table 1Purification of b-mannosidase from Aspergillus niger culture filtrate
Activity (nkat) PurificationPurification step Yield (%)Volume (ml) Specific activityProtein (mg)(-fold)(nkat mg−1)
1615015Culture filtrate (1)(100)9.42.296(NH4)2SO4 precipi- 216.71443
tation0.50 170Anion exchange 561.5 1884
chromatography0.1347 500 45 5367Gel filtration
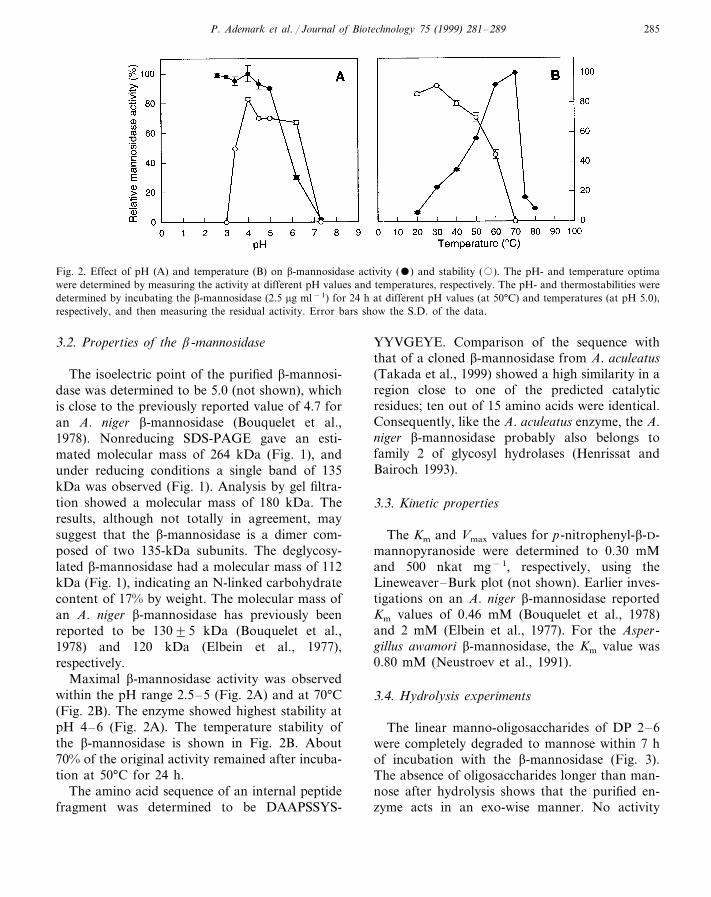
P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289 285
Fig. 2. Effect of pH (A) and temperature (B) on b-mannosidase activity () and stability (�). The pH- and temperature optimawere determined by measuring the activity at different pH values and temperatures, respectively. The pH- and thermostabilities weredetermined by incubating the b-mannosidase (2.5 mg ml−1) for 24 h at different pH values (at 50°C) and temperatures (at pH 5.0),respectively, and then measuring the residual activity. Error bars show the S.D. of the data.
3.2. Properties of the b-mannosidase
The isoelectric point of the purified b-mannosi-dase was determined to be 5.0 (not shown), whichis close to the previously reported value of 4.7 foran A. niger b-mannosidase (Bouquelet et al.,1978). Nonreducing SDS-PAGE gave an esti-mated molecular mass of 264 kDa (Fig. 1), andunder reducing conditions a single band of 135kDa was observed (Fig. 1). Analysis by gel filtra-tion showed a molecular mass of 180 kDa. Theresults, although not totally in agreement, maysuggest that the b-mannosidase is a dimer com-posed of two 135-kDa subunits. The deglycosy-lated b-mannosidase had a molecular mass of 112kDa (Fig. 1), indicating an N-linked carbohydratecontent of 17% by weight. The molecular mass ofan A. niger b-mannosidase has previously beenreported to be 13095 kDa (Bouquelet et al.,1978) and 120 kDa (Elbein et al., 1977),respectively.
Maximal b-mannosidase activity was observedwithin the pH range 2.5–5 (Fig. 2A) and at 70°C(Fig. 2B). The enzyme showed highest stability atpH 4–6 (Fig. 2A). The temperature stability ofthe b-mannosidase is shown in Fig. 2B. About70% of the original activity remained after incuba-tion at 50°C for 24 h.
The amino acid sequence of an internal peptidefragment was determined to be DAAPSSYS-
YYVGEYE. Comparison of the sequence withthat of a cloned b-mannosidase from A. aculeatus(Takada et al., 1999) showed a high similarity in aregion close to one of the predicted catalyticresidues; ten out of 15 amino acids were identical.Consequently, like the A. aculeatus enzyme, the A.niger b-mannosidase probably also belongs tofamily 2 of glycosyl hydrolases (Henrissat andBairoch 1993).
3.3. Kinetic properties
The Km and Vmax values for p-nitrophenyl-b-D-mannopyranoside were determined to 0.30 mMand 500 nkat mg−1, respectively, using theLineweaver–Burk plot (not shown). Earlier inves-tigations on an A. niger b-mannosidase reportedKm values of 0.46 mM (Bouquelet et al., 1978)and 2 mM (Elbein et al., 1977). For the Asper-gillus awamori b-mannosidase, the Km value was0.80 mM (Neustroev et al., 1991).
3.4. Hydrolysis experiments
The linear manno-oligosaccharides of DP 2–6were completely degraded to mannose within 7 hof incubation with the b-mannosidase (Fig. 3).The absence of oligosaccharides longer than man-nose after hydrolysis shows that the purified en-zyme acts in an exo-wise manner. No activity
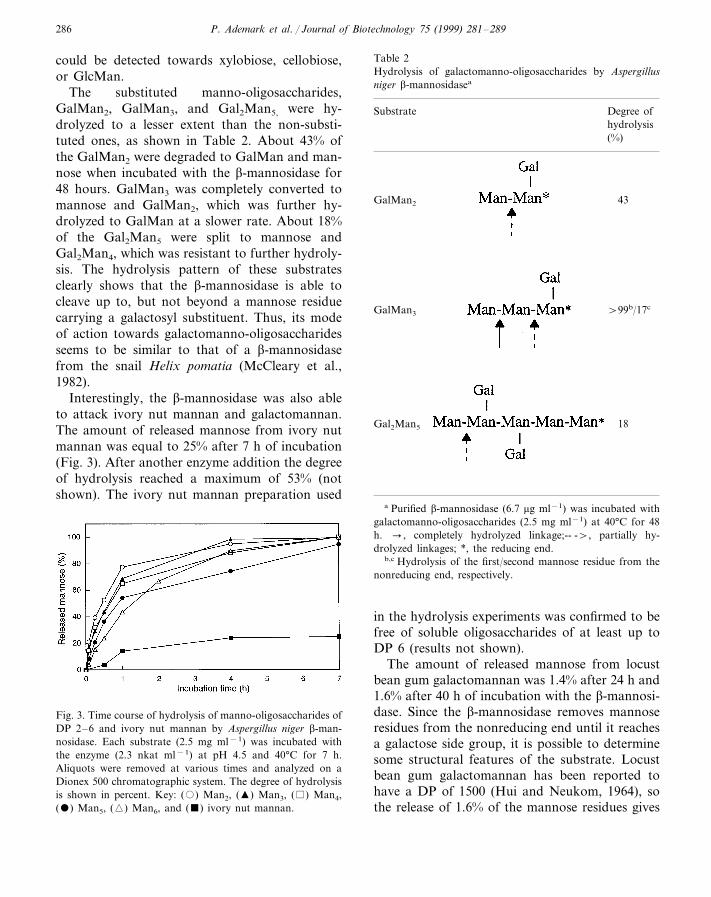
P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289286
could be detected towards xylobiose, cellobiose,or GlcMan.
The substituted manno-oligosaccharides,GalMan2, GalMan3, and Gal2Man5, were hy-drolyzed to a lesser extent than the non-substi-tuted ones, as shown in Table 2. About 43% ofthe GalMan2 were degraded to GalMan and man-nose when incubated with the b-mannosidase for48 hours. GalMan3 was completely converted tomannose and GalMan2, which was further hy-drolyzed to GalMan at a slower rate. About 18%of the Gal2Man5 were split to mannose andGal2Man4, which was resistant to further hydroly-sis. The hydrolysis pattern of these substratesclearly shows that the b-mannosidase is able tocleave up to, but not beyond a mannose residuecarrying a galactosyl substituent. Thus, its modeof action towards galactomanno-oligosaccharidesseems to be similar to that of a b-mannosidasefrom the snail Helix pomatia (McCleary et al.,1982).
Interestingly, the b-mannosidase was also ableto attack ivory nut mannan and galactomannan.The amount of released mannose from ivory nutmannan was equal to 25% after 7 h of incubation(Fig. 3). After another enzyme addition the degreeof hydrolysis reached a maximum of 53% (notshown). The ivory nut mannan preparation used
Table 2Hydrolysis of galactomanno-oligosaccharides by Aspergillusniger b-mannosidasea
Substrate Degree ofhydrolysis(%)
43GalMan2
GalMan3 \99b/17c
18Gal2Man5
a Purified b-mannosidase (6.7 mg ml−1) was incubated withgalactomanno-oligosaccharides (2.5 mg ml−1) at 40°C for 48h. � , completely hydrolyzed linkage;-- -\, partially hy-drolyzed linkages; *, the reducing end.
b,c Hydrolysis of the first/second mannose residue from thenonreducing end, respectively.
Fig. 3. Time course of hydrolysis of manno-oligosaccharides ofDP 2–6 and ivory nut mannan by Aspergillus niger b-man-nosidase. Each substrate (2.5 mg ml−1) was incubated withthe enzyme (2.3 nkat ml−1) at pH 4.5 and 40°C for 7 h.Aliquots were removed at various times and analyzed on aDionex 500 chromatographic system. The degree of hydrolysisis shown in percent. Key: (�) Man2, (�) Man3, ( ) Man4,() Man5, (�) Man6, and () ivory nut mannan.
in the hydrolysis experiments was confirmed to befree of soluble oligosaccharides of at least up toDP 6 (results not shown).
The amount of released mannose from locustbean gum galactomannan was 1.4% after 24 h and1.6% after 40 h of incubation with the b-mannosi-dase. Since the b-mannosidase removes mannoseresidues from the nonreducing end until it reachesa galactose side group, it is possible to determinesome structural features of the substrate. Locustbean gum galactomannan has been reported tohave a DP of 1500 (Hui and Neukom, 1964), sothe release of 1.6% of the mannose residues gives

P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289 287
that the estimated number of terminal, unsubsti-tuted mannose units at the nonreducing end is, inaverage, equal to 24.
The ability of the b-mannosidase to attack ivorynut mannan and galactomannan is an interestingproperty. It obviously has a role in the degradationof mannan and galacomannan polymers, but amore detailed kinetic study is needed to fullyunderstand the mode of action of this enzyme. Thiswork is currently in progress. Several microbialb-mannosidases have been shown to hydrolyzemanno-oligosaccharides (Reese and Shibata, 1965;Hashimoto and Fukumoto, 1969; Wan et al., 1976;Elbein et al., 1977; Sone and Misaki, 1978; Akinoet al., 1988; Oda and Tonomura, 1996), but fewpublications contain information on whether theenzyme is active on polymeric substrates. An A.niger b-mannosidase preparation was earlier shownto release reducing sugars from galactomannan(Elbein et al., 1977). However, this preparation alsocontained a-galactosidase and other activities. Ab-mannosidase from the mushroom Polyporus sul-fureus hydrolyzed mannobiose and mannotriose,but was not able to attack ivory nut mannan (Wanet al., 1976). Araki and Kitamikado (1982) reportedon an enzyme from Aeromonas sp. that hydrolyzedcodium and coffee mannans and manno-oligosac-charides of DP 3 or more. It released however onlymannobiose, and not mannose, from the substratesand did not act on p-nitrophenyl-b-D-mannopyra-noside or mannobiose. b-Xylosidases from thefungi Trichoderma reesei (Margolles-Clark et al.,1996) and Malbranchea pulchella var. sulfurea(Matsuo et al., 1977) have been reported to releasexylose from polymeric xylan.
The exo-wise mode of action of b-mannosidasehas made it a widely used tool for structuralanalysis of complex oligosaccharides. A b-man-nosidase preparation from A. niger has been usedtogether with Mortierella 6inacea a-galactosidaseto determine the structure of galactomanno-oligosaccharides derived from copra galactoman-nan (Kusakabe et al., 1990), and an A. nigerb-mannosidase was also used in studies of acetyl-glucomannan esterase (Tenkanen et al., 1995). Theb-mannosidase purified in the current work wasrecently used to determine the structure of oligosac-charides formed during T. reesei b-mannanase
treatment of pine kraft pulp (Tenkanen et al.,1997).
The ability to transglycosylate seems to be com-mon among b-mannosidases of microbial originand has been noted in for example Rhizopus ni6eus(Hashimoto and Fukumoto, 1969) and an alka-lophilic Bacillus sp. (Akino et al., 1988). A crudeb-mannosidase preparation from A. niger showedgood transmannosylation capacity and producedalkyl-b-mannosides when incubated with manno-biose and various alcohols (Holazo et al., 1992;Itoh and Kamiyama, 1995). No obvious signs oftransglycosylation activity were observed in thecurrent work, but this needs to be further investi-gated. Classified b-mannosidases belong to family1 and 2 of glycosyl hydrolases (Chen et al., 1995;Bauer et al., 1996; Leipprandt et al., 1996; Alkhayatet al., 1998; Stoll et al., 1999; Takada et al., 1999).The enzymes in both these families act by theretaining mechanism, a prerequisite for the abilityto perform transglycosylation, i.e. synthesis ofoligosaccharides. Cloning and characterization ofthe gene encoding the A. niger b-mannosidase isnow in progress, and will provide further informa-tion on the family classification and thus thehydrolytic mechanism of the enzyme.
Acknowledgements
We thank Dr Shi-Gui Yu for performing thetwo-dimensional electrophoresis, Marjukka Pert-tula for HPLC analysis of hydrolysates from thegalactomanno-oligosaccharides, and RiittaIsoniemi for performing part of the hydrolysisexperiments. This study was supported by a grantto FT from the Swedish National Board for Indus-trial and Technical Development (NUTEK) and agrant to HS from the Swedish Research Council forEngineering Sciences (TFR).
References
Ademark, P., Varga, A., Medve, J., Harjunpaa, V., Draken-berg, T., Tjerneld, F., Stalbrand, H., 1998. Softwood hemi-cellulose-degrading enzymes from Aspergillus niger :purification and properties of a b-mannanase. J. Biotech-nol. 63, 199–210.

P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289288
Akino, T., Nakamura, N., Horikoshi, K., 1988. Characteri-zation of b-mannosidase of an Alkalophilic Bacillus sp.Agric. Biol. Chem. 52, 1459–1464.
Alkhayat, A.H., Kraemer, S.A., Leipprandt, J.R., Macek,M., Kleijer, W.J., Friderici, K.H., 1998. Human b-man-nosidase cDNA characterization and first identificationof a mutation associated with human b-mannosidosis.Hum. Mol. Genet. 7, 75–83.
Araki, T., Kitamikado, M., 1982. Purification and character-ization of a novel exo-b-mannanase from Aeromonas sp.F-25. J. Biochem. 91, 1181–1186.
Bauer, M.W., Bylina, E.J., Swanson, R.V., Kelly, R.M.,1996. Comparison of a b-glucosidase and a b-mannosi-dase from the hyperthermophilic archaeon Pyrococcus fu-riosus. Purification, characterization, gene cloning, andsequence analysis. J. Biol. Chem. 271, 23749–23755.
Bouquelet, S., Spik, G., Montreuil, J., 1978. Properties of ab-D-mannosidase from Aspergillus niger. Biochim. Bio-phys. Acta 522, 521–530.
Chen, H., Leipprandt, J.R., Traviss, C.E., Sopher, B.L.,Jones, M.Z., Cavanagh, K.T., Friderici, K.H., 1995.Molecular cloning and characterization of bovine b-man-nosidase. J. Biol. Chem. 270, 3841–3848.
Dey, P.M., 1978. Biochemistry of plant galactomannans.Adv. Carbohydr. Chem. Biochem. 35, 341–376.
Elbein, A.D., Adya, S., Lee, Y.C., 1977. Purification andproperties of a b-mannosidase from Aspergillus niger. J.Biol. Chem. 252, 2026–2031.
Hashimoto, Y., Fukumoto, J., 1969. Studies on the enzymetreatment of coffee beans. Part II. Purification and prop-erties of b-mannosidase from Rhizopus ni6eus. NipponNogei Kagaku Kaishi 43, 564–569.
Henrissat, B., Bairoch, A., 1993. New families in the classifi-cation of glycosyl hydrolases based on amino-acid se-quence similarities. Biochem. J. 293, 781–788.
Holazo, A., Shinoyama, H., Kamiyama, Y., Yasui, T., 1992.Screening for b-mannosidases with transmannosidationcapacity. Biosci. Biotech. Biochem. 56, 822–824.
Hui, P.A., Neukom, H., 1964. Properties of galactomannans.Tappi 47, 39–42.
Itoh, H., Kamiyama, Y., 1995. Synthesis of alkyl b-man-nosides from mannobiose by Aspergillus niger b-mannosi-dase. J. Ferment. Bioeng. 80, 510–512.
Kobata, A., 1993. Glycobiology: an expanding research areain carbohydrate chemistry. Acc. Chem. Res. 26, 319–324.
Kusakabe, I., Kaneko, R., Takada, N., Zamora, A.F., Fer-nandez, W.L., Murakami, K., 1990. A simple method forelucidating structures of galactomanno-oligosaccharidesby sequential actions of b-mannosidase and a-galactosi-dase. Agric. Biol. Chem. 54, 1081–1083.
Leipprandt, J.R., Kraemer, S.A., Haithcock, B.E., Chen, H.,Dyme, J.L., Cavanagh, K.T., Friderici, K.H., Jones,M.Z., 1996. Caprine b-mannosidase: sequencing andcharacterization of the cDNA and identification of themolecular defect of caprine beta-mannosidosis. Genomics37, 51–56.
Margolles-Clark, E., Tenkanen, M., Nakari-Setala, T., Pent-tila, M., 1996. Cloning of genes encoding a-L-arabino-furanosidase and b-xylosidase from Trichoderma reesei byexpression in Saccharomyces cere6isiae. Appl. Environ.Microbiol. 62, 3840–3846.
Matsuo, M., Yasui, T., Kobayashi, T., 1977. Enzymaticproperties of b-xylosidase from Malbranchea pulchellavar. sulfurea No. 48. Agric. Biol. Chem. 41, 1601–1606.
McCleary, B.V., Taravel, F.R., Cheetham, N.W.H., 1982.Preparative-scale isolation and characterisation of 61-a-D-galactosyl-(1–4)-b-D-mannobiose and 62-a-D-galactosyl-(1–4)-b-D-mannobiose. Carbohydr. Res. 104, 285–297.
McKenzie, L.F., Wang, Q., Warren, R.A.J., Withers, S.G.,1998. Glycosynthases: mutant glycosidases for oligosac-charide synthesis. J. Am. Chem. Soc. 120, 5583–5584.
Neufeld, E.F., 1991. Lysosomal storage diseases. Annu. Rev.Biochem. 60, 257–280.
Neustroev, K.N., Krylov, A.S., Firsov, L.M., Abroskina,O.L., Khorlin, A.Ya., 1991. Isolation and properties ofb-mannosidase from Aspergillus awamori. Biokhimiya 56,1406–1412.
Oda, Y., Tonomura, K., 1996. Characterization of b-man-nanase and b-mannosidase secreted from the yeast Tri-chosporon cutaneum JCM 2947. Lett. Appl. Microbiol.22, 173–178.
Reese, E.T., Shibata, Y., 1965. b-mannanases of fungi. Can.J. Microbiol. 11, 167–183.
Ratto, M., Poutanen, K., 1988. Production of mannan-de-grading enzymes. Biotechnol. Lett. 10, 661–664.
Sone, Y., Misaki, A., 1978. Purification and characterizationof b-D-mannosidase and b-N-acetyl-D-hexosaminidase ofTremella fuciformis. J. Biochem. 83, 1135–1144.
Stoll, D. (1998) Characterization of the mannan-degradingsystem of Cellulomonas fimi, Thesis for Doctor of Philos-ophy, University of British Columbia, Vancouver.
Stoll, D., Stalbrand, H., Warren, R.A.J., 1999. Mannan-de-grading enzymes from Cellulomonas fimi. Appl. Environ.Microbiol. 65, 2598–2605.
Stalbrand, H., Siika-aho, M., Tenkanen, M., Viikari, L.,1993. Purification and characterization of two b-man-nanases from Trichoderma reesei. J. Biotechnol. 29, 229–242.
Takada, G., Kawaguchi, T., Kaga, T., Sumitani, J.-I., Arai,M., 1999. Cloning and sequencing of b-mannosidase genefrom Aspergillus aculeatus No. F-50. Biosci. Biotechnol.Biochem. 63, 206–209.
Tenkanen, M., Thornton, J., Viikari, L., 1995. An acetylglu-comannan esterase of Aspergillus oryzae ; purification,characterization and role in the hydrolysis of O-acetyl-galactoglucomannan. J. Biotechnol. 42, 197–206.
Tenkanen, M., Makkonen, M., Perttula, M., Viikari, L.,Teleman, A., 1997. Action of Trichoderma reesei man-nanase on galactoglucomannan in pine kraft pulp. J.Biotechnol. 57, 191–204.
Torto, N., Buttler, T., Gorton, L., Marko-Varga, G.,Stalbrand, H., Tjerneld, F., 1995. Monitoring of enzy-matic hydrolysis of ivory nut mannan using on-line mi-

P. Ademark et al. / Journal of Biotechnology 75 (1999) 281–289 289
crodialysis sampling and anion-exchange chromatographywith integrated pulsed electochemical detection. Anal. Chim.Acta 313, 15–24.
Viikari, L., Tenkanen, M., Buchert, J., Ratto, M., Bailey, M.,Siika-aho, M., Linko, M., 1993. Hemicellulases for indus-trial applications. In: Saddler, J.N. (Ed.), Biotechnology inAgriculture, No. 9. Bioconversion of Forest and Agricul-tural Plant Residues. C.A.B. International, Wallingford, pp.
131–182.Wan, C.C., Muldrey, J.E., Li, S.-C., Li, Y.-T., 1976. Mannosi-
dase from the mushroom Polyporus sulfureus. J. Biol. Chem.251, 4384–4388.
Yu, S.-G., Stef·nsson, H., Romanowska, E., Albertsson, P.-A, .,1994. Two dimensional electrophoresis of thylakoid mem-brane proteins and its application to microsequencing.Photosynth. Res. 41, 475–486.
.

II
81

III
93

IV
101

A cellulose-binding module of the Trichoderma reeseib-mannanase Man5A increases the mannan-hydrolysis of
complex substrates
Per Hagglund a, Torny Eriksson a, Anna Collen a, Wim Nerinckx b,Marc Claeyssens b, Henrik Stalbrand a,*
a Department of Biochemistry, Centre for Chemistry and Chemical Engineering, Lund University, PO Box 124, S-221 00 Lund, Swedenb Department of Biochemistry/Physiology/Microbiology, Ghent University, K.L. Ledeganckstraat 35, B-9000 Ghent, Belgium
Received 11 January 2002; received in revised form 29 August 2002; accepted 3 September 2002
Abstract
Endo-b-1,4-D-mannanases (b-mannanase; EC 3.2.1.78) are endohydrolases that participate in the degradation of
hemicellulose, which is closely associated with cellulose in plant cell walls. The b-mannanase from Trichoderma reesei
(Man5A) is composed of an N-terminal catalytic module and a C-terminal carbohydrate-binding module (CBM). In
order to study the properties of the CBM, a construct encoding a mutant of Man5A lacking the part encoding the CBM
(Man5ADCBM), was expressed in T. reesei under the regulation of the Aspergillus nidulans gpdA promoter. The wild-
type enzyme was expressed in the same way and both proteins were purified to electrophoretic homogeneity using ion-
exchange chromatography. Both enzymes hydrolysed mannopentaose, soluble locust bean gum galactomannan and
insoluble ivory nut mannan with similar rates. With a mannan/cellulose complex, however, the deletion mutant lacking
the CBM showed a significant decrease in hydrolysis. Binding experiments using activity detection of Man5A and
Man5ADCBM suggests that the CBM binds to cellulose but not to mannan. Moreover, the binding of Man5A to
cellulose was compared with that of an endoglucanase (Cel7B) from T. reesei .
# 2002 Elsevier Science B.V. All rights reserved.
Keywords: Carbohydrate-binding module; Hemicellulase; Cellulose; Hemicellulose; Endoglucanase
1. Introduction
Endo-b-1,4-D-mannanase (b-mannanase; EC
3.2.1.78) catalyses the random hydrolysis of
manno-glycosidic bonds in both mannans and
heteromannans. These polysaccharides are wide-
spread in nature and are found in several different
forms. Linear b-1,4-mannan polymers are found in
Abbreviations: CBM, carbohydrate-binding module; M2,
mannobiose; M3, mannotriose; DNS, 3,5-dinitrosalicylic acid.
* Corresponding author. Tel.: �/46-46-222-8202; fax: �/46-
46-222-4534.
E-mail address: [email protected] (H.
Stalbrand).
Journal of Biotechnology 101 (2003) 37�/48
www.elsevier.com/locate/jbiotec
0168-1656/02/$ - see front matter # 2002 Elsevier Science B.V. All rights reserved.
PII: S 0 1 6 8 - 1 6 5 6 ( 0 2 ) 0 0 2 9 0 - 0

certain plant species such as ivory nut (Phytelephas
macrocarpa ) and date (Phoenix dactylifera ; Meier,
1958). Glucomannans and galactomannans are
present in the endosperm of many plants as
storage polysaccharides (Meier and Reid, 1982).
Acetylated galactoglucomannan is the major hemi-
cellulose in softwood (Sjostrom, 1993). Further-
more, mannan can be found in two crystal forms,
which are referred to as mannan I and mannan II(Chanzy et al., 1984; Frei and Preston, 1968).
b-Mannanases have several existing and poten-
tial applications in, for example, the food and feed,
and the pulp and paper industries (Buchert et al.,
1998; Sachslehner et al., 2000; Viikari et al., 1993).
Previously, the b-mannanase from Trichoderma
reesei (Man5A) has been purified and the corre-
sponding gene has been cloned (Stalbrand et al.,1993, 1995). The hydrolysis patterns of Man5A on
oligosaccharides and branched mannan-polymers
have been studied (Tenkanen et al., 1997; Harjun-
paa et al., 1995), and the degradation of crystalline
mannan has been investigated (Hagglund et al.,
2001). Sequence alignment and hydrophobic clus-
ter analysis have indicated that Man5A is a two-
module enzyme, composed of an N-terminalcatalytic module and a C-terminal putative carbo-
hydrate-binding module (CBM) connected by a
linker region (Stalbrand et al., 1995). The catalytic
module of Man5A belongs to family 5 of glycoside
hydrolases (Henrissat, 1991), and it has the (a/b)8-
barrel fold characteristic for this enzyme family
(Sabini et al., 2000).
On the basis of sequence alignment, the Man5ACBM has been classified into family 1 of CBMs
(Tomme et al., 1995). A majority of the modules in
this family are connected to cellulases of fungal
origin, and several have been shown to bind to
cellulose (Linder and Teeri, 1997; Teeri et al.,
1992). The role of the CBMs in cellulolytic
enzymes has been thoroughly investigated and it
has been shown that they often play an importantrole in binding to, and increasing the hydrolysis of,
insoluble cellulose substrates (van Tilbeurgh et al.,
1986; Tomme et al., 1988; Gilkes et al., 1988).
Besides Man5A, CBMs have been found in
other b-mannanases (Sunna et al., 2001; Stoll et
al., 2000), and also in other types of hemicellulose-
degrading enzymes (Black et al., 1996; Millward-
Sadler et al., 1994). To this date, most studies ofhemicellulase-linked CBMs, and their effect on
hydrolysis have been carried out with xylanases
(Ali et al., 2001; Black et al., 1996, 1997; Millward-
Sadler et al., 1994; Gill et al., 1999). However,
Biely and Tenkanen (1998) have discussed the
possibility that the presence of a CBM on the T.
reesei b-mannanase is part of the reason for the
superior performance in pulp bleaching experi-ments for this enzyme compared with some other
b-mannanases (Suurnakki et al., 1996).
The binding of Man5A to cellulose and kraft
fibres, likely mediated by the CBM, has been
shown earlier (Gerber et al., 1999; Ademark et al.,
1998; Tenkanen et al., 1995), but the detailed
binding properties are unknown. In order to
further study the binding properties of Man5A,also on different mannans, and to investigate the
effect of the CBM on hydrolysis of different
substrates, we have constructed a mutant of
Man5A lacking the CBM (Man5ADCBM). We
have expressed the mutant and the wild-type
enzyme in T. reesei and compared the catalytic
activities and the binding properties of the purified
enzymes. Moreover, the binding of Man5A tocellulose was compared with that of a T. reesei
endoglucanase (Cel7B) carrying a family 1 CBM.
The results indicate that the C-terminal CBM of
Man5A binds specifically to cellulose. Further-
more, the results of the hydrolysis experiments
indicate that the CBM helps to increase the rate of
hydrolysis of insoluble mannan/cellulose com-
plexes.
2. Materials and methods
2.1. Strains, plasmids and media
T. reesei QM9414 was maintained on potato
dextrose agar slants at 30 8C. Escherichia coli
XL1 blue (Stratagene, La Jolla, CA, USA) wasused as a host for recombinant plasmids. E. coli
cells were cultured in luria broth medium at 37 8Cand ampicillin (100 mg ml�1) was added when
appropriate. The plasmids used in this study were
pHSM30 (Stalbrand et al., 1995), pAN52 (Punt et
al., 1990) and pTOC202. pAN52 was kindly
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/4838

provided by Dr Peter Punt at TNJ (Rijswijk, TheNetherlands), and pTOC202 was kindly provided
by Professor Merja Penttila, VTT Biotechnology
(Espoo, Finland).
2.2. DNA techniques
Plasmid DNA preparation, restriction analysis,
and DNA fragment isolation was performed asdescribed previously (Sambrook et al., 1989).
Mutagenesis was performed by PCR using Pfu
polymerase with the Quikchange† site-directed
mutagenesis kit (Stratagene). All PCR runs were
initiated with a denaturation step at 94 8C for 1
min, followed by cycles of denaturation, annealing
and extension. The number of cycles and the
temperatures in the different steps in each cyclevaried in the different runs. All runs were ended
with an extension step for 10 min. All constructs
were sequenced using ABI sequencing instruments
with cycle sequencing methodology at Innovagen
(Lund, Sweden).
2.3. Construction of plasmids
The SacII site in the man5A sequence (cDNAencoding T. reesei Man5A) on plasmid pHSM30
was removed by site-directed silent mutagenesis,
using PCR with the primers 5?-CCTCAGCGCT-
GGCGGCCGTACTGCAGCCTG-3? (primer 1)
and 5?-CAGGCTGCAGTACGGCCGCCAGC-
GCTGAGG-3? (primer 2). The PCR was run in
16 cycles, each composed of the following steps:
denaturation (94 8C), 30 s; annealing (62 8C), 1min; extension (70 8C), 14 min. The PCR product
was treated with DpnI (which cleaves methylated
DNA) in order to remove remaining template
plasmid DNA. The new plasmid was called
pPH001.
The coding sequence of man5A was amplified
from pPH001 by PCR using the primers 5?-CT-
CGAGCCGCGGACTGGCATCATGATGATG-CTCTCAAAG-3? (primer 3) and 5?-GGGCGC-
CTCGAGGATCCTCTAGACAAACAGGACT-
TAAG-3? (primer 4). The PCR was run in 27
cycles, each composed of the following steps:
denaturation (94 8C), 45 s; annealing (61 8C), 1
min; extension (70 8C), 4 min. The coding se-
quence of the catalytic module (bases 1�/1137 ofman5A) (man5ADCBM) was amplified from
pPH001 by PCR using primer 3 and 5?-GCCTCGAGGATCCCTAGGGAGGAG-
GAGTGGTTG-3? (primer 5). The PCR was run
in 27 cycles, each composed of the following steps:
denaturation (94 8C), 45 s; annealing (62 8C), 45
s; extension (72 8C), 3 min. The two amplified
fragments (man5A and man5ADCBM) were iso-lated from an agarose electrophoresis gel using
Jetsorb† (Genomed, Research Triangle Park, NC,
USA), subsequent to treatment of the PCR
products with SacII and BamHI. pPH002 was
constructed by ligation of BamHI and SacII
digested pAN52 with man5A. pPH003 was con-
structed by legation of BamHI and SacII digested
pAN52 with man5ADCBM.
2.4. Transformation of T. reesei
Transformation of plasmids into T. reesei was
performed essentially as described by Collen et al.
(2001b), a modification of the method described
by Penttila et al. (1987). pPH002 and pPH003 were
transformed into T. reesei QM9414 together withpTOC202, carrying the Aspergillus nidulans gene
encoding acetamidase (amdS ). Transformants of
T. reesei were selected by growing on plates
containing KH2PO4 (0.11 M), glucose (0.11 M),
sorbitol (1 M), acetamide (10 mM), CsCl (15 mM),
CaCl2 (4.1 mM), MgSO4 (2.4 mM), CaCl2 (5.4
mM), FeSO4 �/ 7H2O (0.18 mM), MnSO4 �/ H2O
(0.095 mM), ZnSO4 �/ 7H2O (0.049 mM), CoCl2 �/6H2O (0.16 mM), agar noble (18 g l�1). Trans-
formants were replated two to three times on a
medium as described above but with sorbitol
omitted and Triton X-100 (0.1% (v/v)) added.
Isolated transformants were tested for b-manna-
nase activity using the activity assay described
below.
2.5. Determination of specific activity
The activity of Man5A and Man5ADCBM on
soluble galactomannan was determined with the b-
mannanase assay using the 3,5-dinitrosalicylic acid
(DNS)-method with 0.5% locust bean gum (Sigma,
St. Louis, MO, USA) as substrate (Stalbrand et
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/48 39

al., 1993). Protein concentrations were determinedby analysis of UV absorbance as described by
Mach et al. (1992). The standard deviations in the
b-mannanase activity measurements and the pro-
tein concentration determinations were below 8%.
2.6. Expression and purification
Stable transformants were cultured in 1 lindented flasks at 30 8C and 160 rpm in a volume
of 100 ml of a medium containing KH2PO4 (0.22
M), K2HPO4 (0.046 M), (NH4)SO4 (0.015 M),
glucose (0.22 M), MgSO4 (5 mM), CaCl2 (5.4
mM), FeSO4 �/ 7H2O (0.18 mM), MnSO4 �/ H2O
(0.095 mM), ZnSO4 �/ 7H2O (0.049 mM), CoCl2 �/6H2O (0.16 mM). The glucose concentration was
monitored regularly by measuring reducing sugarswith DNS (Collen et al., 2001a) and maintained
above 1%. Supernatant from these cultures was
used to screen for b-mannanase activity using the
DNS-method with locust bean gum (0.5%) as
substrate.
Culture filtrates containing Man5A and Man5-
ADCBM were concentrated approximately ten
times by ultrafiltration using a Stirred Cell System(Pall Gelman Sciences, Ann Arbor, MI, USA).
The concentrated culture filtrate was equilibrated
in 10 mM Tris�/HCl buffer (pH 7.8) and loaded
onto a XK 26/20 column packed with 75 ml
Sepharose HP† (Amersham Pharmacia Biotech,
Uppsala, Sweden). A gradient from 0 to 50% 1 M
NaCl (in 10 mM Tris�/HCl buffer pH 7.8) was
created over 20 column volumes. Man5A waspurified further on a Mono-P chromatofocusing
column (Amersham Pharmacia Biotech) with 25
mM triethanolamine (pH 8.3) as equilibration
buffer. Elution was carried out with polybuffer
96 (30%) and polybuffer 74 (70%).
Cel7B was purified from a culture filtrate of T.
reesei QM 9414 according to Eriksson et al.
(2001). Cel7BDCBM (amino acid 1�/371), con-structed as described previously (Collen et al.,
2001b), was expressed under control by the gpdA
promoter from A. nidulans . Cel7BDCBM was
purified by anion-exchange chromatography with
Source Q (Amersham Pharmacia Biotech), using
20 mM Na-acetate (pH 4.5) as buffer A and 1 M
Na-acetate (pH 4.5) as buffer B. Cel7BDCBM waseluted with a linear gradient of buffer B.
2.7. Gel electrophoresis
SDS-PAGE electrophoresis was performed ac-
cording to Laemmli (1970) using precast gels
(Nupage† 4�/12% bis-Tris), from Novex (San
Diego, CA, USA) and the proteins were stainedwith a silver staining kit (SilverExpress†) from
Novex. Affinity gel electrophoresis using gels
including 0.2 mg l�1 locust bean gum galacto-
mannan was performed according to Stoll et al.
(2000).
2.8. Polysaccharides
In this work, polysaccharides isolated from theendosperm of ivory nut (P. macrocarpa ), were
used. P. macrocarpa contains mostly mannan but
also some cellulose (Chanzy et al., 1984; Timell,
1957; Meier, 1958). A commercially available
(Megazyme, Bray, Ireland) mannan preparation
(more than 99% mannan, see below) isolated from
ivory nut was used, here referred to as ‘ivory nut
mannan’. In addition, a preparation isolated fromivory nut, which is the residue after removal of low
molecular weight mannan with alkali treatment
(1.25 M KOH) according to Aspinall et al. (1953),
was used. Earlier investigations have shown that
such a residue is composed of mainly mannan, but
also contains some cellulose (Chanzy et al., 1984;
Meier, 1958), as was also confirmed here (see
below). In this text, this preparation is referred toas ‘mannan/cellulose complex’.
Acid hydrolysis of ivory nut mannan was
carried out in 0.4 M H2SO4 at 120 8C for 2 h.
Acid hydrolysis of the mannan/cellulose complex
from ivory nut was performed according to
Hagglund (1951). The monosaccharides released
upon acid hydrolysis were analysed using a
Dionex† P-500 chromatographic system withCarbo Pac PA-10 pre- and analytical columns.
Detection was performed with an electrochemical
detector (ED40). Separation was achieved isocra-
tically with 10 mM NaOH as eluent. Mannose,
galactose and glucose (Fluka) were used as stan-
dards. The monosaccharide composition analysis
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/4840

of ivory nut mannan yielded more than 99%mannose residues. The composition of the man-
nan/cellulose complex sample was determined to
be 81% mannose, 15% glucose and 0.4% galactose
residues. This sample also contained 3% material
which was insoluble under the conditions used.
Ivory nut mannan was also used for crystal-
lisation of mannan I crystals as described pre-
viously (Hagglund et al., 2001). Crystals ofmannan II were made from Acetabularia crenulata
mannan as described previously (Hagglund et al.,
2001).
2.9. Hydrolysis experiments
Mannopentaose (Megazyme) was used at a
concentration of 1 mM and the concentration ofivory nut mannan and the mannan/cellulose com-
plex was 2.5 mg ml�1. In all experiments, equal
amounts of Man5A and Man5ADCBM were
added based on the activity assay towards locust
bean gum. All hydrolysis experiments were carried
out in 50 mM Na-acetate buffer (pH 4.5) at 40 8Cand bovine serum albumin (BSA) (100 mg ml�1)
was included.Samples were withdrawn at different time points
during hydrolysis and heated to 100 8C in 2 min,
followed by centrifugation (20 000�/g , 10 min)
and filtration (0.22 mm filter) to remove insoluble
matter. The released oligosaccharides were ana-
lysed using a Dionex† P-500 chromatographic
system with Carbo Pac PA-100 pre- and analytical
columns. Detection was performed with an ED40.Separation was achieved isocratically with 100
mM NaOH as eluent. Enzymatic activity was
calculated on the basis of the amount of manno-
biose (M2) and mannotriose (M3) released; the
main products of hydrolysis by the b-mannanase.
M2 and M3 standards were from Megazyme.
2.10. Binding studies
Various amounts (see legend of Figs. 4 and 6) of
Man5A and Man5ADCBM were incubated with
Avicel† cellulose (Merck, Darmstadt, Germany; 2
mg ml�1), ivory nut mannan (2.5 mg ml�1),
mannan/cellulose complex (2.5 mg ml�1), mannan
I crystals (1 mg ml�1), mannan II crystals (1 mg
ml�1) in 50 mM Na-acetate buffer (pH 4.5) at4 8C for 1 h under continuous stirring. BSA (100
mg ml�1) was included in all incubations. After
incubation the polysaccharides were removed by
centrifugation (20 000�/g , 10 min) and residual
enzyme activity was assayed using the DNS-
method with locust bean gum (0.5%) as substrate
(Stalbrand et al., 1993). The residual activity was
compared with the activity of Man5A and Man5-ADCBM incubated under the same conditions, but
without polysaccharides. Binding of Cel7B and
Cel7BDCBM to Avicel cellulose was determined in
the same way, except that 50 mM Na-acetate
buffer (pH 4.8) was used and 0.5% carboxymethyl-
cellulose (ICN Biomedicals, Aurora, OH, USA)
was used as substrate in the assay. The relative
equilibrium association constants (Kr) for bindingof Man5A and Cel7B to Avicel cellulose were
determined as described by Gilkes et al. (1992),
assuming a one-site binding mode. Non-linear
regression analyses were made in Kaleidagraph†
3.5.
3. Results
3.1. Expression and purification of Man5A and
Man5ADCBM
The genes encoding the T. reesei b-mannanase
(man5A ) and the region encoding the catalytic
module (man5ADCBM (bases 1�/1137; amino acid
1�/352)) were inserted into pAN52 (Fig. 1). In this
plasmid, the transcription of man5A and man5-
ADCBM is controlled by the constitutive gpdA
promoter from A. nidulans (Punt et al., 1990). The
plasmids carrying man5A and man5ADCBM were
transformed into T. reesei QM9414 and grown in
a glucose-rich medium. Under these conditions,
expression of several other extracellular glycoside
hydrolases, including the endogenous Man5A, is
suppressed (Ilmen et al., 1997).To confirm the absence of expression of en-
dogenous b-mannanase activity, untransformed T.
reesei QM9414 was cultured under the same
conditions as the transformants and the b-manna-
nase activity was determined to be lower than 0.01
nkat ml�1. In contrast, the activity of the positive
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/48 41

transformants was between 20 and 30 nkat ml�1.Man5A and Man5ADCBM were expressed in T.
reesei and the enzymes were purified by anion-
exchange chromatography. The purified enzymes
appeared as single bands of expected mobility on
SDS-PAGE (Fig. 2).
3.2. Catalytic activity of Man5A and
Man5ADCBM on soluble substrates
The hydrolysis of mannopentaose and soluble
locust bean gum galactomannan by Man5A and
Man5ADCBM was compared. The specific activityon galactomannan was 330 kat mol�1 for Man5A
and 340 kat mol�1 for Man5ADCBM. Equal
amounts of both enzymes were incubated with
mannopentaose and samples were withdrawn at
several time points for analysis of released oligo-
saccharides. The results show that Man5A and
Man5ADCBM exhibited similar hydrolytic activ-
ity towards mannopentaose (Fig. 3). The rate ofmannopentaose hydrolysis was 10 mol mannopen-
taose per s mol�1 enzyme for Man5A and 12.6
mol mannopentaose per s mol�1 enzyme for
Man5ADCBM determined as the production of
M2 and M3 after 90 min.
3.3. Adsorption of Man5A and Man5ADCBM to
cellulose
Man5A and Man5ADCBM were incubated with
cellulose, and the amount of bound enzyme was
determined, as described in Section 2. As pre-dicted, cellulose binding is observed with Man5A,
but no significant binding is seen with Man5-
ADCBM (Fig. 4). As a comparison, Cel7B and the
catalytic module of this enzyme (Cel7BDCBM)
were incubated with cellulose in the same way and
the amount of binding to cellulose was analysed
(Fig. 4). In analogy to the b-mannanase, signifi-
cant binding was seen with the full-length enzymeonly.
In order to quantify the adsorption to cellulose,
the relative binding constants of Man5A and
Cel7B for this polysaccharide were determined as
described by Gilkes et al. (1992), using the
relationship:
[B]�[N0]Ka[F ]
1 � Ka[F ](1)
where [B ] is the amount of enzyme bound, [F ] isthe concentration of free enzyme, [N0] is the
concentration of binding sites and Ka is the
equilibrium association constant. Non-linear re-
gression of the binding data gives values for [N0]
and Ka. Unique solutions for [N0] and Ka can not
be obtained but the relative equilibrium associa-
tion constant, Kr, (which can be used to compare
the binding of different enzymes to the samecellulose preparation) can be obtained using the
relationship (Gilkes et al., 1992):
Kr� [N0]Ka (2)
Analysis of the binding data in this manner,
assuming a one-site binding mode, gives a slightly
higher Kr for Cel7B in comparison with Man5A
(1.05 and 0.84 l g�1, respectively).
3.4. Hydrolysis of ivory nut mannan and a mannan/
cellulose complex
To investigate whether the CBM has an influ-
ence on the hydrolysis of insoluble mannan-poly-
mers, Man5A and Man5ADCBM were incubated
with ivory nut mannan and a mannan/cellulose
complex isolated from ivory nut (P. macrocarpa ).
The ivory nut mannan is extracted from ivory nutwith alkali, and contains mannan only (confirmed
by acid hydrolysis which yielded more than 99%
mannose). The mannan/cellulose complex is the
residue when low molecular weight mannan is
Fig. 1. Schematic view of Man5A and Man5ADCBM. The C-
terminus (C) and N-terminus (N) of mature Man5A (Stalbrand
et al., 1995) are indicated. Numbers refer to the amino acid
number of the C- and N-terminal amino acids of Man5A and
Man5ADCBM.
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/4842
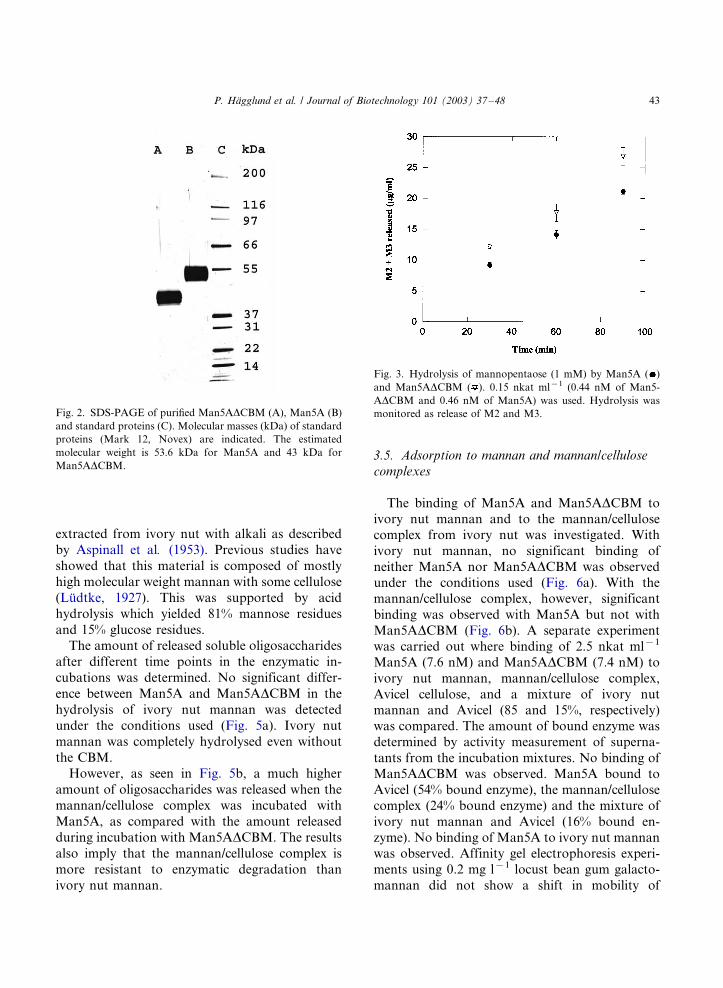
extracted from ivory nut with alkali as described
by Aspinall et al. (1953). Previous studies have
showed that this material is composed of mostly
high molecular weight mannan with some cellulose
(Ludtke, 1927). This was supported by acid
hydrolysis which yielded 81% mannose residues
and 15% glucose residues.
The amount of released soluble oligosaccharides
after different time points in the enzymatic in-
cubations was determined. No significant differ-
ence between Man5A and Man5ADCBM in the
hydrolysis of ivory nut mannan was detected
under the conditions used (Fig. 5a). Ivory nut
mannan was completely hydrolysed even without
the CBM.
However, as seen in Fig. 5b, a much higher
amount of oligosaccharides was released when the
mannan/cellulose complex was incubated with
Man5A, as compared with the amount released
during incubation with Man5ADCBM. The results
also imply that the mannan/cellulose complex is
more resistant to enzymatic degradation than
ivory nut mannan.
3.5. Adsorption to mannan and mannan/cellulose
complexes
The binding of Man5A and Man5ADCBM to
ivory nut mannan and to the mannan/cellulose
complex from ivory nut was investigated. With
ivory nut mannan, no significant binding of
neither Man5A nor Man5ADCBM was observed
under the conditions used (Fig. 6a). With the
mannan/cellulose complex, however, significant
binding was observed with Man5A but not with
Man5ADCBM (Fig. 6b). A separate experiment
was carried out where binding of 2.5 nkat ml�1
Man5A (7.6 nM) and Man5ADCBM (7.4 nM) to
ivory nut mannan, mannan/cellulose complex,
Avicel cellulose, and a mixture of ivory nut
mannan and Avicel (85 and 15%, respectively)
was compared. The amount of bound enzyme was
determined by activity measurement of superna-
tants from the incubation mixtures. No binding of
Man5ADCBM was observed. Man5A bound to
Avicel (54% bound enzyme), the mannan/cellulose
complex (24% bound enzyme) and the mixture of
ivory nut mannan and Avicel (16% bound en-
zyme). No binding of Man5A to ivory nut mannan
was observed. Affinity gel electrophoresis experi-
ments using 0.2 mg l�1 locust bean gum galacto-
mannan did not show a shift in mobility of
Fig. 2. SDS-PAGE of purified Man5ADCBM (A), Man5A (B)
and standard proteins (C). Molecular masses (kDa) of standard
proteins (Mark 12, Novex) are indicated. The estimated
molecular weight is 53.6 kDa for Man5A and 43 kDa for
Man5ADCBM.
Fig. 3. Hydrolysis of mannopentaose (1 mM) by Man5A ( )
and Man5ADCBM ( ). 0.15 nkat ml�1 (0.44 nM of Man5-
ADCBM and 0.46 nM of Man5A) was used. Hydrolysis was
monitored as release of M2 and M3.
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/48 43
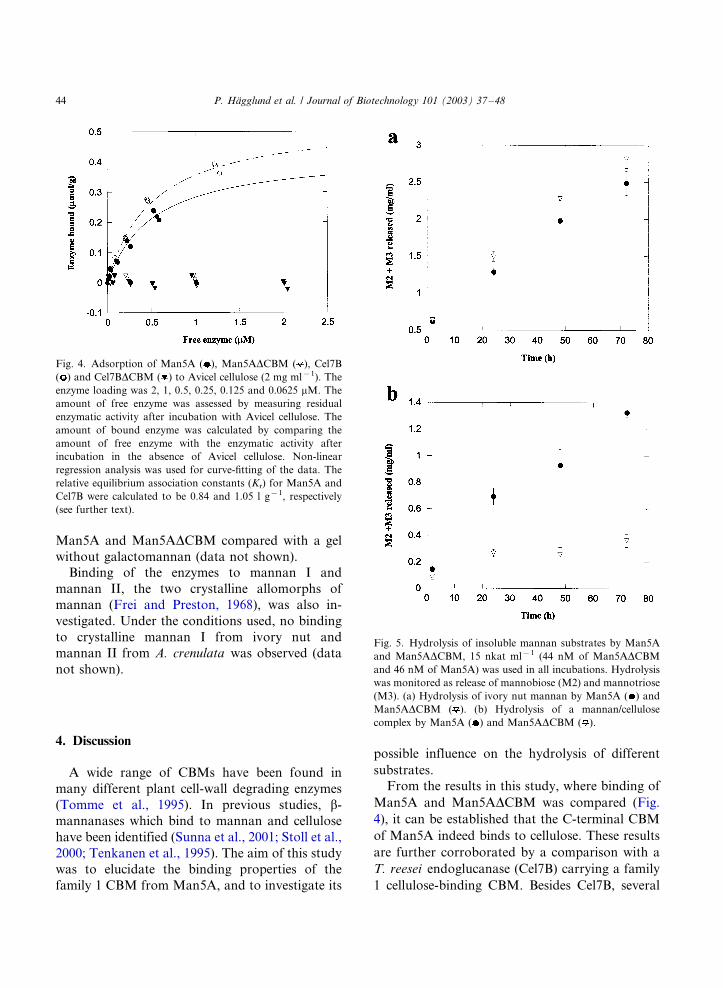
Man5A and Man5ADCBM compared with a gel
without galactomannan (data not shown).
Binding of the enzymes to mannan I and
mannan II, the two crystalline allomorphs of
mannan (Frei and Preston, 1968), was also in-
vestigated. Under the conditions used, no binding
to crystalline mannan I from ivory nut and
mannan II from A. crenulata was observed (data
not shown).
4. Discussion
A wide range of CBMs have been found in
many different plant cell-wall degrading enzymes
(Tomme et al., 1995). In previous studies, b-
mannanases which bind to mannan and cellulose
have been identified (Sunna et al., 2001; Stoll et al.,
2000; Tenkanen et al., 1995). The aim of this study
was to elucidate the binding properties of the
family 1 CBM from Man5A, and to investigate its
possible influence on the hydrolysis of different
substrates.
From the results in this study, where binding of
Man5A and Man5ADCBM was compared (Fig.
4), it can be established that the C-terminal CBM
of Man5A indeed binds to cellulose. These results
are further corroborated by a comparison with a
T. reesei endoglucanase (Cel7B) carrying a family
1 cellulose-binding CBM. Besides Cel7B, several
Fig. 5. Hydrolysis of insoluble mannan substrates by Man5A
and Man5ADCBM, 15 nkat ml�1 (44 nM of Man5ADCBM
and 46 nM of Man5A) was used in all incubations. Hydrolysis
was monitored as release of mannobiose (M2) and mannotriose
(M3). (a) Hydrolysis of ivory nut mannan by Man5A ( ) and
Man5ADCBM ( ). (b) Hydrolysis of a mannan/cellulose
complex by Man5A ( ) and Man5ADCBM ( ).
Fig. 4. Adsorption of Man5A ( ), Man5ADCBM ( ), Cel7B
( ) and Cel7BDCBM ( ) to Avicel cellulose (2 mg ml�1). The
enzyme loading was 2, 1, 0.5, 0.25, 0.125 and 0.0625 mM. The
amount of free enzyme was assessed by measuring residual
enzymatic activity after incubation with Avicel cellulose. The
amount of bound enzyme was calculated by comparing the
amount of free enzyme with the enzymatic activity after
incubation in the absence of Avicel cellulose. Non-linear
regression analysis was used for curve-fitting of the data. The
relative equilibrium association constants (Kr) for Man5A and
Cel7B were calculated to be 0.84 and 1.05 l g�1, respectively
(see further text).
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/4844
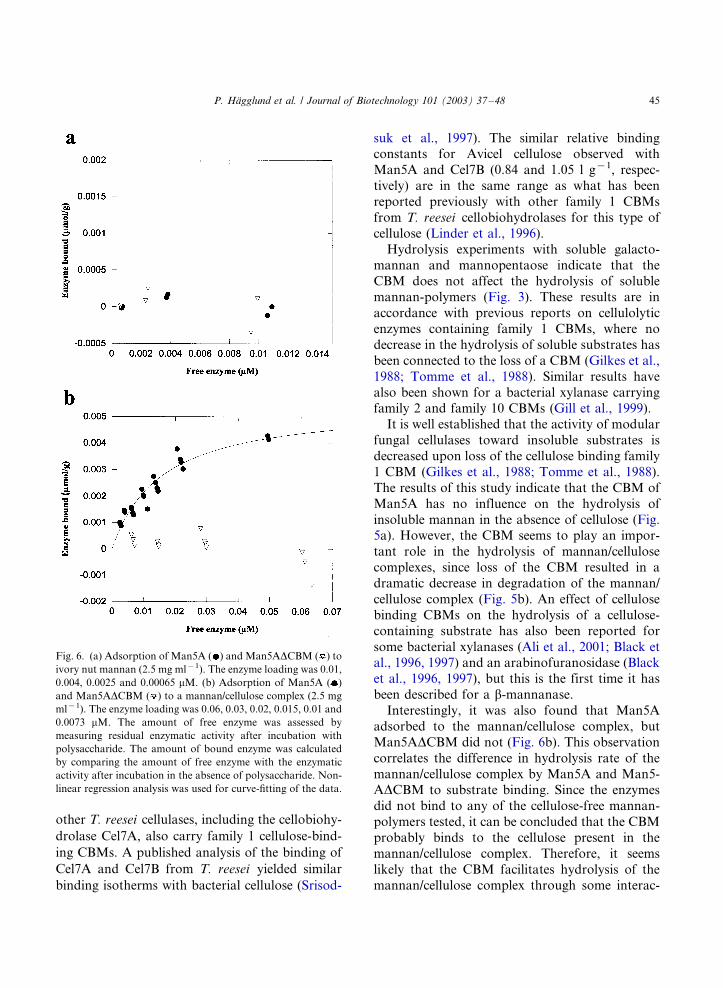
other T. reesei cellulases, including the cellobiohy-
drolase Cel7A, also carry family 1 cellulose-bind-
ing CBMs. A published analysis of the binding of
Cel7A and Cel7B from T. reesei yielded similar
binding isotherms with bacterial cellulose (Srisod-
suk et al., 1997). The similar relative bindingconstants for Avicel cellulose observed with
Man5A and Cel7B (0.84 and 1.05 l g�1, respec-
tively) are in the same range as what has been
reported previously with other family 1 CBMs
from T. reesei cellobiohydrolases for this type of
cellulose (Linder et al., 1996).
Hydrolysis experiments with soluble galacto-
mannan and mannopentaose indicate that theCBM does not affect the hydrolysis of soluble
mannan-polymers (Fig. 3). These results are in
accordance with previous reports on cellulolytic
enzymes containing family 1 CBMs, where no
decrease in the hydrolysis of soluble substrates has
been connected to the loss of a CBM (Gilkes et al.,
1988; Tomme et al., 1988). Similar results have
also been shown for a bacterial xylanase carryingfamily 2 and family 10 CBMs (Gill et al., 1999).
It is well established that the activity of modular
fungal cellulases toward insoluble substrates is
decreased upon loss of the cellulose binding family
1 CBM (Gilkes et al., 1988; Tomme et al., 1988).
The results of this study indicate that the CBM of
Man5A has no influence on the hydrolysis of
insoluble mannan in the absence of cellulose (Fig.5a). However, the CBM seems to play an impor-
tant role in the hydrolysis of mannan/cellulose
complexes, since loss of the CBM resulted in a
dramatic decrease in degradation of the mannan/
cellulose complex (Fig. 5b). An effect of cellulose
binding CBMs on the hydrolysis of a cellulose-
containing substrate has also been reported for
some bacterial xylanases (Ali et al., 2001; Black etal., 1996, 1997) and an arabinofuranosidase (Black
et al., 1996, 1997), but this is the first time it has
been described for a b-mannanase.
Interestingly, it was also found that Man5A
adsorbed to the mannan/cellulose complex, but
Man5ADCBM did not (Fig. 6b). This observation
correlates the difference in hydrolysis rate of the
mannan/cellulose complex by Man5A and Man5-ADCBM to substrate binding. Since the enzymes
did not bind to any of the cellulose-free mannan-
polymers tested, it can be concluded that the CBM
probably binds to the cellulose present in the
mannan/cellulose complex. Therefore, it seems
likely that the CBM facilitates hydrolysis of the
mannan/cellulose complex through some interac-
Fig. 6. (a) Adsorption of Man5A ( ) and Man5ADCBM ( ) to
ivory nut mannan (2.5 mg ml�1). The enzyme loading was 0.01,
0.004, 0.0025 and 0.00065 mM. (b) Adsorption of Man5A ( )
and Man5ADCBM ( ) to a mannan/cellulose complex (2.5 mg
ml�1). The enzyme loading was 0.06, 0.03, 0.02, 0.015, 0.01 and
0.0073 mM. The amount of free enzyme was assessed by
measuring residual enzymatic activity after incubation with
polysaccharide. The amount of bound enzyme was calculated
by comparing the amount of free enzyme with the enzymatic
activity after incubation in the absence of polysaccharide. Non-
linear regression analysis was used for curve-fitting of the data.
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/48 45

tion with the cellulose component of the complex.Since mannan and cellulose are believed to be
closely associated in plant cell walls (Akerholm
and Salmen, 2001), it is plausible that the binding
of Man5A to cellulose facilitates the degradation
of the mannan/cellulose complex by increasing the
substrate proximity, a mechanism which has been
suggested for some modular xylanases (Black et
al., 1996, 1997; Gill et al., 1999). Another possi-bility could be that the CBM disrupts the mannan/
cellulose complex, in analogy to the previously
demonstrated cell wall disruptive activity of a
CBM from a bacterial endoglucanase (Din et al.,
1991), thus making the substrate more available
for enzymatic degradation. However, at this stage,
it is not clear exactly how the CBM affects the
degradation of the mannan/cellulose complex.To conclude, the work presented here suggests
that the C-terminal CBM of Man5A binds to
cellulose but not to mannan. The fact that Man5A
appears to have similar binding properties as the
T. reesei cellulases, further strengthens the view
that the Man5A CBM is indeed a cellulose binding
module. Moreover, it was shown that the presence
of the Man5A CBM increases the hydrolysis rateof an insoluble mannan/cellulose complex. How-
ever, it can not be ruled out that the CBM binds
also to other polysaccharides besides cellulose in
natural complex substrates. Further experiments
will be carried out to investigate more closely how
the CBM facilitates the action of the b-mannanase
in degradation of such substrates.
Acknowledgements
Lars Anderson is thanked for help with affinity
gel electrophoresis. The Swedish Research Council
for Engineering Sciences (TFR), the Swedish
Research Council (VR) and Carl Tryggers Re-
search Foundation are gratefully thanked for
grants to Henrik Stalbrand.
References
Ademark, P., Varga, A., Medve, J., Harjunpaa, V., Draken-
berg, T., Tjerneld, F., Stalbrand, H., 1998. Softwood
hemicellulose-degrading enzymes from Aspergillus niger :
purification and properties of a b-mannanase. J. Biotechnol.
63, 199�/210.
.Akerholm, M., Salmen, L., 2001. Interactions between wood
polymers studied by dynamic FT-IR spectroscopy. Polymer
42, 963�/969.
Ali, M.K., Hayashi, H., Karita, S., Goto, M., Kimura, T.,
Sakka, K., Ohmiya, K., 2001. Importance of the
carbohydrate-binding module of Clostridium stercorarium
Xyn10B to xylan hydrolysis. Biosci. Biotechnol. Biochem.
65, 41�/47.
Aspinall, G.O., Hirst, E.L., Percival, E.G.V., Williamson, I.R.,
1953. The mannans of ivory nut (Phytelephas macrocarpa ).
Part I, the methylation of mannan A and mannan B. J.
Chem. Soc., 3184�/3188.
Biely, P., Tenkanen, M., 1998. Enzymology of hemicellulose
degradation. In: Harman, G.E., Kubicek, C.P. (Eds.),
Trichoderma Gliocladium, vol. 2. Taylor & Francis, Lon-
don, pp. 25�/47.
Black, G.W., Rixon, J.E., Clarke, J.H., Hazlewood, G.P.,
Theodorou, M.K., Morris, P., Gilbert, H.J., 1996. Evidence
that linker sequences and cellulose-binding domains en-
hance the activity of hemicellulases against complex sub-
strates. Biochem. J. 319, 515�/520.
Black, G.W., Rixon, J.E., Clarke, J.H., Hazlewood, G.P.,
Ferreira, L.M., Bolam, D.N., Gilbert, H.J., 1997. Cellulose
binding domains and linker sequences potentiate the activity
of hemicellulases against complex substrates. J. Biotechnol.
57, 59�/69.
Buchert, J., Oksanen, T., Pere, J., Siika-Aho, M., Suurnakki,
A., Viikari, L., 1998. Applications of Trichoderma reesei
enzymes in the pulp and paper industry. In: Harman, G.E.,
Kubicek, C.P. (Eds.), Trichoderma Gliocladium, vol. 2.
Taylor & Francis, London, pp. 343�/363.
Chanzy, H.D., Grosrenaud, A., Vuong, R., Mackie, W., 1984.
The crystalline polymorphism of mannan in plant cell walls
and after recrystallization. Planta 161, 320�/329.
Collen, A., Ward, M., Tjerneld, F., Stalbrand, H., 2001a.
Genetically engineered peptide fusions for improved protein
partitioning in aqueous two-phase systems. Effect of fusion
localization on endoglucanase I of Trichoderma reesei . J.
Chromatogr. A 910, 275�/284.
Collen, A., Ward, M., Tjerneld, F., Stalbrand, H., 2001b.
Genetic engineering of the Trichoderma reesei endogluca-
nase I (Cel7B) for enhanced partitioning in aqueous two-
phase systems containing thermoseparating ethylene oxide-
propylene oxide copolymers. J. Biotechnol. 87, 179�/191.
Din, N., Gilkes, N.R., Tekant, B., Miller, R.C., Warren, A.J.,
Kilburn, D.G., 1991. Non-hydrolytic disruption of cellulose
fibers by the binding domain of a bacterial cellulase.
Biotechnology 9, 1096�/1099.
Eriksson, T., Karlsson, J., Tjerneld, F., 2001. A model
explaining declining rate in hydrolysis of lignocellulose
substrates with cellobiohydrolase I (Cel7A) and endogluca-
nase I (Cel7B) of Trichoderma reesei . Appl. Biochem.
Biotechnol. 101, 41�/60.
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/4846

Frei, E., Preston, R.D., 1968. Noncellulosic structural poly-
saccharides in algal cell walls. III. Mannan in siphoneous
green algae. Proc. R. Soc. London, Ser. B 169, 127�/145.
Gerber, P.J., Heitmann, J.A., Joyce, T.W., Buchert, J., Siika-
aho, M., 1999. Adsorption of hemicellulases onto bleached
kraft fibers. J. Biotechnol. 67, 67�/75.
Gilkes, N.R., Warren, R.A., Miller, R.C., Jr, Kilburn, D.G.,
1988. Precise excision of the cellulose binding domains from
two Cellulomonas fimi cellulases by a homologous protease
and the effect on catalysis. J. Biol. Chem. 263, 10401�/
10407.
Gilkes, N.R., Jervis, E., Henrissat, B., Tekant, B., Miller, R.C.,
Jr, Warren, R.A., Kilburn, D.G., 1992. The adsorption of a
bacterial cellulase and its two isolated domains to crystalline
cellulose. J. Biol. Chem. 267, 6743�/6749.
Gill, J., Rixon, J.E., Bolam, D.N., McQueen-Mason, S.,
Simpson, P.J., Williamson, M.P., Hazlewood, G.P., Gilbert,
H.J., 1999. The type II and X cellulose-binding domains of
Pseudomonas xylanase A potentiate catalytic activity
against complex substrates by a common mechanism.
Biochem. J. 342, 473�/480.
Harjunpaa, V., Teleman, A., Siika-aho, M., Drakenberg, T.,
1995. Kinetic and stereochemical studies of manno-
oligosaccharide hydrolysis catalysed by b-mann-
anases from Trichoderma reesei . Eur. J. Biochem. 234,
278�/283.
Henrissat, B., 1991. A classification of glycosyl hydrolases
based on amino acid sequence similarities. Biochem. J. 280,
309�/316.
Hagglund, E., 1951. Chemistry of Wood. Academic Press, New
York.
Hagglund, P., Sabini, E., Boisset, C., Wilson, K., Chanzy, H.,
Stalbrand, H., 2001. Degradation of mannan I and II
crystals by fungal endo-b-1,4-mannanases and a b-1,4-
mannosidase studied with transmission electron micro-
scopy. Biomacromolecules 2, 694�/699.
Ilmen, M., Saloheimo, A., Onnela, M.L., Penttila, M.E., 1997.
Regulation of cellulase gene expression in the filamentous
fungus Trichoderma reesei . Appl. Environ. Microbiol. 63,
1298�/1306.
Laemmli, U.K., 1970. Cleavage of structural proteins during
the assembly of the head of bacteriophage T4. Nature 227,
680�/685.
Linder, M., Teeri, T.T., 1997. The roles and function of
cellulose-binding domains. J. Biotechnol. 57, 15�/28.
Linder, M., Salovuori, I., Ruohonen, L., Teeri, T.T., 1996.
Characterization of a double cellulose-binding domain.
Synergistic high affinity binding to crystalline cellulose. J.
Biol. Chem. 271, 21268�/21272.
Ludtke, M., 1927. Zur Kenntnins der pflanzlichen Zellmem-
bran. Uber die Kohlenhydrate des Steinnußsamens. Ann.
Chem. 456, 201�/224.
Mach, H., Middaugh, C.R., Lewis, R.V., 1992. Statistical
determination of the average values of the extinction
coefficients of tryptophan and tyrosine in native proteins.
Anal. Biochem. 200, 74�/80.
Meier, H., 1958. On the structure of cell walls and cell wall
mannans from ivory nuts and from dates. Biochim.
Biophys. Acta 28, 229�/240.
Meier, H., Reid, J.S.G., 1982. Reserve polysaccharides other
than starch in higher plants. Encyl. Plant Physiol. New Ser.
13A, 418�/471.
Millward-Sadler, S.J., Poole, D.M., Henrissat, B., Hazlewood,
G.P., Clarke, J.H., Gilbert, H.J., 1994. Evidence for a
general role for high-affinity non-catalytic cellulose binding
domains in microbial plant cell wall hydrolases. Mol.
Microbiol. 11, 375�/382.
Penttila, M., Nevalainen, H., Ratto, M., Salminen, E.,
Knowles, J., 1987. A versatile transformation system for
the cellulolytic filamentous fungus Trichoderma reesei . Gene
61, 155�/164.
Punt, P.J., Dingemanse, M.A., Kuyvenhoven, A., Soede, R.D.,
Pouwels, P.H., van den Hondel, C.A., 1990. Functional
elements in the promoter region of the Aspergillus nidulans
gpdA gene encoding glyceraldehyde-3-phosphate dehydro-
genase. Gene 93, 101�/109.
Sabini, E., Schubert, H., Murshudov, G., Wilson, K.S., Siika-
Aho, M., Penttila, M., 2000. The three-dimensional struc-
ture of a Trichoderma reesei b-mannanase from glycoside
hydrolase family 5. Acta Crystallogr. D Biol. Crystallogr.
56, 3�/13.
Sachslehner, A., Foidl, G., Foidl, N., Gubitz, G., Haltrich, D.,
2000. Hydrolysis of isolated coffee mannan and coffee
extract by mannanases of Sclerotium rolfsii . J. Biotechnol.
80, 127�/134.
Sambrook, J., Fritsch, E.F., Maniatis, T., 1989. Molecular
Cloning: A Laboratory Manual. Cold Spring Harbor
Laboratory Press, Plainview, NY.
Sjostrom, E., 1993. Wood Chemistry, Fundamentals and
Applications. Academic Press, San Diego.
Srisodsuk, M., Lehtio, J., Linder, M., Margolles-Clark, E.,
Reinikainen, T., Teeri, T.T., 1997. Trichoderma reesei
cellobiohydrolase I with an endoglucanase cellulose-binding
domain: action on bacterial microcrystalline cellulose. J.
Biotechnol. 57, 49�/57.
Stoll, D., Boraston, A., Stalbrand, H., McLean, B.W., Kilburn,
D.G., Warren, R.A., 2000. Mannanase Man26A from
Cellulomonas fimi has a mannan-binding module. FEMS
Microbiol. Lett. 183, 265�/269.
Stalbrand, H., Siika-aho, M., Tenkanen, M., Viikari, L., 1993.
Purification and characterization of two b-mannanases from
Trichoderma reesei . J. Biotechnol. 29, 229�/242.
Stalbrand, H., Saloheimo, A., Vehmaanpera, J., Henrissat, B.,
Penttila, M., 1995. Cloning and expression in Saccharo-
myces cerevisiae of a Trichoderma reesei b-mannanase gene
containing a cellulose binding domain. Appl. Environ.
Microbiol. 61, 1090�/1097.
Sunna, A., Gibbs, M.D., Bergquist, P.L., 2001. Identification of
novel b-mannan- and b-glucan-binding modules: evidence
for a superfamily of carbohydrate-binding modules. Bio-
chem. J. 356, 791�/798.
Suurnakki, A., Clark, T., Allison, R., Buchert, J., Viikari, L.,
1996. Mannanase aided bleaching of soft-wood Kraft pulp.
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/48 47

In: Messner, K., Srebotnik, E. (Eds.), Biotechnology in Pulp
and Paper Industry*/Advances in Applied and Funda-
mental Research. WUA Universitatsverlag, Vienna, pp. 69�/
74.
Teeri, T., Reinikainen, T., Ruohonen, L., Jones, T.A.,
Knowles, J.K.C., 1992. Domain function in Trichoderma
reesei cellobiohydrolases. J. Biotechnol. 24, 169�/176.
Tenkanen, M., Buchert, J., Viikari, L., 1995. Binding of
hemicellulases on isolated polysaccharide substrates. En-
zyme Microb. Technol. 17, 499�/505.
Tenkanen, M., Makkonen, M., Perttula, M., Viikari, L.,
Teleman, A., 1997. Action of Trichoderma reesei manna-
nase on galactoglucomannan in pine Kraft pulp. J. Bio-
technol. 57, 191�/204.
Timell, T.E., 1957. Vegetable ivory as a source of a mannan
polysaccharide. Can. J. Chem. 35, 333�/338.
Tomme, P., Van Tilbeurgh, H., Pettersson, G., Van Damme, J.,
Vandekerckhove, J., Knowles, J., Teeri, T., Claeyssens, M.,
1988. Studies of the cellulolytic system of Trichoderma
reesei QM 9414. Analysis of domain function in two
cellobiohydrolases by limited proteolysis. Eur. J. Biochem.
170, 575�/581.
Tomme, P., Warren, R.A.J., Miller, R.C.J., Kilburn, D.G.,
Gilkes, N.R., 1995. Cellulose-binding domains: classifica-
tion and properties. In: Saddler, J.M., Penner, M. (Eds.),
Enzymatic Degradation of Insoluble Polysaccharides.
American Chemical Society, Washington, DC, pp. 142�/163.
van Tilbeurgh, H., Tomme, P., Claeyssens, M., Bhikhabhai, R.,
Pettersson, G., 1986. Limited proteolysis of the cellobiohy-
drolase I from Trichoderma reesei . FEBS 204, 223�/227.
Viikari, L., Tenkanen, M., Buchert, J., Ratto, M., Bailey, M.,
Siika-aho, M., Linko, M., 1993. Hemicellulases for indus-
trial applications. In: Saddler, J.N. (Ed.), Bioconversion of
Forest and Agricultural Plant Residues, vol. 9. CAB
International, Wallingford, UK, pp. 131�/182.
P. Hagglund et al. / Journal of Biotechnology 101 (2003) 37�/4848

V
121

Journal of Biotechnology 92 (2002) 267–277
endo-�-1,4-Mannanases from blue mussel, Mytilus edulis :purification, characterization, and mode of action�
Bingze Xu a, Per Hagglund b, Henrik Stalbrand b, Jan-Christer Janson a,*a Center for Surface Biotechnology, Biomedical Center, Uppsala Uni�ersity, Box 577, SE-751 23 Uppsala, Sweden
b Department of Biochemistry, Center for Chemistry and Chemical Engineering, Lund Uni�ersity, Box 124,SE-221 00 Lund, Sweden
Received 29 January 2001; received in revised form 28 June 2001; accepted 6 July 2001
Abstract
Two variants of an endo-�-1,4-mannanase from the digestive tract of blue mussel, Mytilus edulis, were purified bya combination of immobilized metal ion affinity chromatography, size exclusion chromatography in the absence andpresence of guanidine hydrochloride and ion exchange chromatography. The purified enzymes were characterizedwith regard to enzymatic properties, molecular weight, isoelectric point, amino acid composition and N-terminalsequence. They are monomeric proteins with molecular masses of 39 216 and 39 265 Da, respectively, as measured byMALDI-TOF mass spectrometry. The isoelectric points of both enzymes were estimated to be around 7.8, howeverslightly different, by isoelectric focusing in polyacrylamide gel. The enzymes are stable from pH 4.0 to 9.0 and havetheir maximum activities at a pH about 5.2. The optimum temperature of both enzymes is around 50–55 °C. Theirstability decreases rapidly when going from 40 to 50 °C. The N-terminal sequences (12 residues) were identical forthe two variants. They can be completely renatured after denaturation in 6 M guanidine hydrochloride. The enzymesreadily degrade the galactomannans from locust bean gum and ivory nut mannan but show no cross-specificity forxylan and carboxymethyl cellulose. There is no binding ability observed towards cellulose and mannan. © 2002Elsevier Science B.V. All rights reserved.
Keywords: Mytilus edulis ; Blue mussel; Purification; �-Mannanase
www.elsevier.com/locate/jbiotec
1. Introduction
endo-�-1,4-Mannanases (�-mannanase, EC3.2.1.78) cleave randomly within the �-1,4-man-nan main chain of galactomannan, glucomannan,galactoglucomannan and mannan (Matheson andMcCleary, 1985; McCleary and Matheson, 1986).They are very important for the enzymatic diges-tion of hemicelluloses, one of the most abundantgroup of polymers in nature. �-Mannanases hy-
� Note : The novel N-terminal amino-acid sequence datapublished here has been submitted to the SWISS-PROTprotein sequence database and is available under accessionnumber P82801.
* Corresponding author. Tel.: +46-18-471-4011; fax: +46-18-555-016.
E-mail address: [email protected] (J.-C.Janson).
0168-1656/02/$ - see front matter © 2002 Elsevier Science B.V. All rights reserved.
PII: S 0168 -1656 (01 )00367 -4

B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277268
drolyze mannan yielding mannotriose and man-nobiose (Stalbrand et al., 1993). �-Mannanasesare used in the food and pharmaceutical industry(Christgau et al., 1994a; McCleary, 1990), as wellas in the paper industry (Paice and Jurasek, 1984).
�-Mannanases are primarily produced by bacte-ria and hemicellulolytic fungi (Stalbrand et al.,1993; Gubitz et al., 1996; Tamaru et al., 1995;Duffaud et al., 1997; Christgau et al., 1994b).�-Mannanases are classified in family 5 and fam-ily 26 of glycosyl hydrolases according to theclassification by Henrissat and Bairoch (1993). Afew �-mannanases have been purified and charac-terized from higher organisms, such as mollusca(Yamaura and Matsumoto, 1993; Yamaura et al.,1996; McCleary, 1988). However, no report hasbeen found in the literature on �-mannanasesisolated from blue mussel, Mytilus edulis. Some ofthese organisms may perhaps digest mannan fromsea-weed (Yamaura et al., 1996). The mussel di-gestive enzymes are secreted from a crystallinestyle which projects from a style-sac, a primitiveform of pancreas, into the lumen of the stomach(Purchon, 1977). We isolated crystalline stylesfrom both fresh and frozen blue mussel and foundhigh �-mannanase activity in the extracts fromthese. The preliminary purification and characteri-zation studies were performed with these extractsas starting material. However, for large scalepreparations, dissected whole digestive glandswere used as starting material.
This work describes for the first time the purifi-cation and characterization of �-mannanases fromblue mussel. No mollusc �-mannanase gene se-quence has yet been published. However, the N-terminal sequence data obtained in this workenabled comparison with those published for �-mannanases from two other molluscs.
2. Materials and methods
2.1. Materials
M. edulis used as raw material in this workoriginate from waters off the Swedish west coast.Larger quantities of frozen mussel were boughtdirectly from a whole seller, Lysekils Fryshus AB,
Lysekil, Sweden. STREAMLINE® Chelating, Su-perdex® 75 prep grade (60/600 prepackedcolumn), Mono® S HR 10/10 and HR 5/5, Su-perdex® 200 prep grade (16/60 prepackedcolumn), ExcelGel® SDS gradient gel (8–18),PhaseGel™ IEF 3–9 gel, Low Molecular WeightCalibration Kit (LMW) and Broad pI CalibrationKit were obtained from Amersham PharmaciaBiotech (Uppsala, Sweden). CarboPac PA-100Guard and Analytical columns were obtainedfrom Dionex. 3,5-Dinitrosalicylic acid (DNS) andlocust bean gum were from Sigma Chemical Co.(St. Louis, MO, USA). Linear �-1,4-linkedmanno-oligosaccharides and ivory nut mannanwere purchased from Megazyme. Coomassie Bril-liant Blue (R-250) was from Bio-Rad Laborato-ries. All other chemicals were standard analyticalgrade commercial products. FPLC® System, Mul-tiphor® II Electrophoresis System and PhastSys-tem™ were obtained from Amersham PharmaciaBiotech (Uppsala, Sweden). For the HPLC-analy-sis the following equipment from Dionex (Sunny-vale, CA) were used: AS 50 chromatographyCompartment, ED 40 Electrochemical Detector,GP 50 Gradient Pump and AS 50 Autosampler.
2.2. Purification procedure
In Table 1 are summarized all the steps in-volved in the purification procedure. All purifica-tion steps were carried out at room temperatureunless otherwise stated.
2.2.1. Enzyme extractionIn a typical purification process, 1.42 kg hepa-
topancreas (digestive glands) were excised from28.7 kg of whole frozen mussel. Phenylmethylsul-fonylfluoride (PMSF), dissolved in 5 ml ethanol,was added to a concentration of 1 mM followedby homogenisation in a meat blender cooled onice. Twenty millimoles of ice cold phosphatebuffer pH 7.0 was added to a total volume of 3.2l followed by extraction by stirring for 30 min.Insoluble materials were removed by centrifuga-tion at 10 000×g for 30 min. The supernatantwas collected including residues of a difficult-to-remove top layer of floating lipid material. Fi-nally, NaCl was added to a concentration of 1 M,
B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277 269
this material served as a starting material forpurification.
2.2.2. Batch adsorption to STREAMLINEChelating (Zn2+) and column elution
The presence of small amounts of lipid particlesin the crude extract supernatant after the centrifu-gation prevented the use of column adsorptionand this was why batch adsorption was used asthe first purification step. Thus 300 ml (sedi-mented volume) STREAMLINE Chelating, satu-rated with Zn2+ ions and rinsed with distilledwater, was added to the crude blue mussel extractcontaining 1 M NaCl in 20 mM sodium phos-phate buffer pH 7.0. After low speed propellerblade stirring for 2 h, the adsorbent particles wereallowed to sediment and the supernatant contain-ing non-adsorbed material and floating lipid parti-cles were decanted. After washing three times with500 ml, 20 mM sodium phosphate buffer pH 7.0containing 1 M NaCl, the adsorbent particlescontaining the target proteins were packed into aXK 50/30 column (Amersham Pharmacia Bio-tech). After further washing with the same bufferuntil the absorbance at A280 had reached thebase line, the less strongly bound proteins wereeluted with equilibration buffer containing 50 mMimidazole at a flow rate of 150 cm h−1. The targetprotein was eluted with equilibration buffer con-taining 50 mM EDTA at a flow rate of 50 cmh−1.
2.2.3. Size exclusion chromatographyThe fractions containing �-mannanase activity
were pooled (about 80 ml), further concentratedto 20 ml using an Amicon Ultrafilter PM10 andloaded to the 60×600 mm Superdex 75 pgcolumn equilibrated in 20 mM Na–phosphatebuffer pH 7.0 containing 0.2 M NaCl. The flow-rate was 30 cm h−1.
2.2.4. Ion exchange chromatographyThe �-mannanase active fractions were pooled
(about 120 ml) and changed into 20 mM sodiumacetate buffer pH 5.5 by gel filtration on Sep-hadex G-25. The sample was then applied to aMono S (HR10/10) column equilibrated in thesame buffer at a flow-rate of 300 cm h−1. Elutionwas performed by applying a 20-column volumes0–400 mM NaCl-gradient in the same buffer at aflow-rate of 150 cm h−1. Two �-mannanase activepeaks called ManA and ManB, respectively, ap-peared at about 200–250 mM NaCl.
2.2.5. Size exclusion chromatography in 6 MGu–HCl
SDS-PAGE analysis of the material in theManA and ManB peaks from the IEC on MonoS revealed several trace contaminants, not ob-served in native-PAGE. In addition, MALDI-TOF analysis data were difficult to interpret dueto a multitude of signals. This is why materialfrom the two �-mannanase active peaks were
Table 1Purification of �-mannanases from M. edulis
Total protein Total activityStep Yield (%)Specific activity (U mg−1) Purification (fold)(mg) (units)
32 275 n.d.aCrude extract n.c.b
n.c.IMAC eluate n.c.0.62204331248 159UF concentrate 0.64 1.03 77.9
7.2 100SEC eluate 13.9 22.4 49.20.89 43.6IEC eluate (ManA) 49.0 79.0 21.4
18.71.15 38.1 33.1 53.4IEC eluate (ManB)49.7 80.2 19.00.78IEC (ManA, after 38.8
Gu–HCl)32.4 52.3 16.01.01IEC (ManB, after 32.7
Gu–HCl)
a n.d., not determined due to high carbohydrate (reducing group) content of the extract.b n.c., not calculated. The total yield of ManA+ManB is 19.0+16.0=35%.

B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277270
subjected to further purification by size exclusionchromatography under denaturing conditions. Tothis end a HiLoad 16/60 Superdex 200 pg columnwas equilibrated in 50 mM phosphate buffer con-taining 6 M guanidine–hydrochloride (Gu–HCl)at pH 6.4. A 2-ml sample was applied and elutedat a flow-rate of 1 ml min−1.
2.2.6. Final ion exchange chromatographyThe main peak fractions collected from the
Superdex 200 pg experiment in 6 M Gu–HCl ofboth ManA and ManB were pooled (10 ml) andthe Gu–HCl was removed by gel filtration on aSephadex G-25 column equilibrated in 20 mMsodium acetate buffer pH 5.5. Finally the �-man-nanases were rerun on a Mono S HR 5/5 columnunder the same condition as described above.
2.3. Analytical methods
2.3.1. Polyacrylamide gel electrophoresis analysisSDS-PAGE on reduced and alkylated samples
was performed using Multiphor II ElectrophoresisSystem for purity check, protein compositionanalysis and for the estimation of molecular mass.Isoelectric focusing (IEF) and native-PAGE werecarried out using the Phast System™. The isoelec-tric points of ManA and ManB were estimatedusing PhastGel IEF 3–9.
2.3.2. Mass spectrometry and N-terminalsequence analysis
Samples from the second Mono S purificationstep of both ManA and ManB were analyzed byMALDI-MS (Kratos Kompact MALDI IV,Kratos, UK) and subjected to N-terminal se-quence analysis by Dr A� ke Engstrom at the De-partment of Medical Chemistry, UppsalaBiomedical Center, Uppsala, Sweden, using �-cy-ano-4-hydroxycinnaminic acid as matrix.
2.3.3. Amino acid composition analysisAmino acid composition analysis was per-
formed at the amino acid analysis laboratory atUppsala Biomedical Center. An LKB Alpha-Plusamino acid analyzer was used following standardhydrolysis procedures and applying the standardprotein hydrolysate system with ninhydrindetection.
2.3.4. Determination of enzyme acti�ityThe �-mannanase activity was routinely deter-
mined by adapting the dinitrosalicylic acid (DNS)method for reducing sugar analysis according toStalbrand et al. (1993). As substrate was usedlocust bean gum (LBG), the main component ofwhich is galactomannan with a backbone chain of�-1,4-linked mannosyl substituted with �-1,6-linked galactosyl side-groups. Enzyme assay wascarried out by mixing 50 �l of appropriately di-luted enzyme samples with 450 �l of 50 mMsodium citrate buffer, pH 5.5 containing 0.5%LBG. The mixture was incubated at 40 °C for 20min and the reaction was stopped by the additionof 1 ml DNS reagent. After 5 min in a boilingwater bath and quick cooling to room tempera-ture, the degree of enzymatic hydrolysis of theLBG was determined spectrophotometrically bymeasuring the absorbance at 540 nm. The reduc-ing sugars released were then determined against astandard curve obtained with mannose. One unitof �-mannanase activity is defined as the amountof enzyme that gives rise to reducing end groupscorresponding to 1 mg mannose under the currentexperimental conditions.
Xylanase activity was assayed as described byBailey et al. (1992) using 1.0% (w/v) birchwoodxylan (Sigma X-0502) and oat spelts xylan (SigmaX-0627) as substrates in 50 mM sodium citratebuffer, pH 5.3. Amylase activity was assayed asdescribed by Bernfeld (1955) using 1.0% (w/v)soluble starch (Sigma S-9765) as substrate in 50mM sodium acetate buffer. Endoglucanase activ-ity was assayed as described by Wood and Bhat(1988) using 1.0% (w/v) carboxymethyl cellulose(CMC, medium viscosity, Fluka) as substrate in50 mM sodium acetate buffer, pH 5.5. �-Galac-tosidase and �-mannosidase activities were as-sayed as described by Ratto and Poutanen (1988)using 4.0 mM p-nitrophenyl-�-D-galactopyra-noside (Sigma N-0877) as substrate in 50 mMsodium citrate buffer, pH 4.5 and 2.0 mM 4-nitro-phenyl-�-D-mannopyranoside (Sigma N-1268) assubstrate in 50 mM sodium citrate buffer, pH 5.5.The amount of p-nitrophenol released in the mix-ture was determined by measuring absorbance at405 nm.

B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277 271
2.3.5. Determination of protein concentrationProtein was determined using the Bio-Rad
Protein Assay according to the method of Brad-ford (1976), using bovine serum albumin (BSA) asa reference standard. All chromatographic runswere monitored for protein by absorbance at 280nm.
2.3.6. Determination of pH and temperatureoptima
The optimum pH was determined by incubatingaliquots of the enzymes (3.5 pmol in 50 �l) with450 �l of 0.5% substrate (LBG) at 40 °C atdifferent pH values (pH 3–6: 50 mM sodiumcitrate buffer, pH 6.5–7.0: 50 mM phosphatebuffer, pH 7.2–9.0: 50 mM Tris–HCl, pH 9.0–10: 50 mM glycine–NaOH buffer). The reactionswere stopped after 20 min by adding DNS reagentand the activities were assayed for as describedabove. The same procedure was used to determinethe optimum temperature by incubating the en-zymes with substrate at different temperatures (atoptimum pH) for 20 min and the activities wereassayed for as described above.
2.3.7. Determination of pH and temperaturestability
The pH stability was determined by incubatingthe enzyme solutions (in the presence of 0.1%BSA) at different pH values at room temperaturefor 24 h. The activity was measured as describedabove. The temperature stability measurementswere performed by incubating the enzyme solu-tions in the presence of 0.1% BSA at 10, 20, 30,40, 50 and 60 °C and samples were withdrawnafter different time intervals and immediatelycooled in an ice bath. The activities were assayedfor as described above.
2.3.8. Determination of kinetic propertiesThe Michaelis–Menten constant (Km) and the
maximum reaction velocity (Vmax) for ManA wasdetermined by incubating the enzyme at a concen-tration of 7.5 pmol with substrate concentrationsranging from 0.25 to 10 mg ml−1 LBG in 50 mMcitrate buffer, pH 5.5 at 40 °C for 10 min. Thereleased oligosaccharides were measured by theDNS method under the standard conditions.Woolf plot was used to determine parameters.
2.3.9. Determination of substrate specificity:mannopentaose and mannan hydrolysis
The �-mannanase substrate specificity was stud-ied using mannopentaose as substrate. An aliquotof ManA (3.5 pmol, 0.03 nkat) was incubatedwith 1 mM mannopentoase at 40 °C in 50 mMsodium citrate buffer, pH 5.5 containing 0.1%BSA. Samples were withdrawn after different timeintervals and immediately put in a boiling waterbath for 2 min to stop the reaction. The dena-tured proteins were removed by centrifugation at14 000×g for 10 min. The supernatants contain-ing the reaction products were analysed by HPLCon CarboPac PA-100 analytical column using 100mM NaOH as the mobile phase at a flow-rate of1 ml min−1. A Dionex 500 chromatographic sys-tem, controlled by PeakNet™ software (Dionex)was used to separate and detect the oligosaccha-rides. Mannose, mannobiose, mannotriose, man-notetraose and mannopentaose at concentrations1, 5, 10, 20 and 40 �g ml−1 were used asstandards.
In a separate experiment, purified ManA (3.5pmol) was added to 2.5 mg ml−1 suspensions ofivory nut mannan in 50 mM sodium citratebuffer, pH 5.5 containing 0.1% BSA. The ivorynut mannan used here is an insoluble linear poly-mer of �-1,4-linked mannosyl residues. The degreeof polymerisation has been reported to be in theorder of 20 (Meier, 1958). The hydrolysis wascarried out at 40 °C for 24 h during which periodthe ivory nut mannan was only partially dis-solved. After centrifugation the contents of thesupernatant were analysed by HPLC as describedabove.
2.3.10. Determination of cellulose and mannanbinding ability
Binding of the �-mannanases to mannan andcellulose was investigated by incubating the en-zymes with the polysaccharides at 4 °C for 1 hafter which the samples were centrifugated andthe residual enzymatic activity in the supernatantwas analysed. Here 3.5 pmol ManA and ManBwere mixed with 1% suspension of Avicel PH 101and Ivory nut mannan, respectively, in 50 mMsodium citrate buffer at pH 5.2.

B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277272
3. Results
3.1. Purification of the �-mannanases
SEC on the Superdex 75 pg 60/600 column ofthe �-mannanase active peak from the IMAC stepgave a 22-fold purification. The enzyme eluted asa narrow, symmetric peak. Due to the relativelyhigh pI (approximate 7.8) of the �-mannanases,the pooled active fractions from the SEC stepwere further purified on a Mono S HR10/10column at pH 5.5. Two active, well separated�-mannanase peaks, ManA and ManB, wereeluted, with purification factors 3.5 and 2.3,respectively.
The results of the purification procedure aresummarized in Table 1. An approximately 1300-fold purification was achieved for ManA with anoverall yield of 20%. For ManB, a 900-fold purifi-cation was achieved with a yield of 16%. Thepurities of the final products were analysed bySDS-PAGE and IEF using the silver stainingmethod. Typical chromatograms of each step areshown in Fig. 1.
3.2. Properties of the �-mannanases
In Fig. 2 is shown the result of the SDS-PAGEanalysis of samples from the different steps of thepurification process. The positions of the singlebands representing the pure �-mannanases afterthe final IEC step indicate a single polypeptidechain with a molecular weight around 39 000 Da.MALDI-TOF measurements resulted in molecu-lar weights 39 702 Da for ManA and 39 265 Dafor ManB, respectively (data not shown). IEFanalysis gave an estimated isoelectric point forManA of 7.8 (Fig. 3). A slightly higher isoelectricpoint was observed for ManB. SDS-PAGE didnot reveal any impurities, even at high sampleloading and using the silver staining method.
As shown in Table 1, there is a significantdifference in specific activity of the purified �-mannanase variants. Thus for ManA a value of49.7 U mg−1 and for ManB a value of 32.4 Umg−1 was obtained, respectively.
Amino acid composition analysis of the purifiedenzymes showed no significant difference between
the two varieties (data not shown). The �-man-nanases are rich in histidine. The N-terminal se-quence of ManA is: R-L-S-V-S-G-T-N-L-N-Y-N-G-H-H-I-F-L-S-G-A-N-Q-A-W-V-N-Y-A-R-D-F-G-H-N-Q-. The first 12 amino acid sequences atthe N-terminal were analysed for ManB andfound to be identical to that of ManA.
The N-terminal sequence alignment of the M.edulis �-mannanase with that of �-mannanasesfrom other higher organisms, P. insularus and themarine mollusc, L. bre�icula, shows significantsequence similarity (Fig. 4). Thus, a conservedregion of ten amino acids could be registered.
The effect of pH on the activity of both �-man-nanases was investigated (data not shown). Maxi-mum activity was obtained at pH 5.2 with acomparatively rapid decrease on both acid andalkaline sides. More than 80% of the activity wasfound in the range pH 4.0–6.5. The enzymes arestable from pH 4 to 9.5 (data not shown). Theoptimum temperature of the �-mannanase activitywas studied at optimum pH (data not shown).Both ManA and ManB showed maximum activityat about 50 °C. At 60 °C the activity was re-duced to 50% of maximum. In a separate temper-ature stability study in the presence of 0.1% BSAwe found that the enzymes are unstable at highertemperatures. At 60 °C the enzymes lost theiractivity within 1 min. At 50 °C the half-life ofboth ManA and ManB was 20 min. Both enzymeslost their activity completely after 60 min (datanot shown). However, the enzymes are stable forseveral hours at 30 and 40 °C.
3.3. Kinetic properties
The Km and Vmax values for ManA and LBGwere determined to be 3.95 mg ml−1 and 176.8nkat mg−1, respectively, using the Woolf plot(not shown).
3.4. Enzymatic hydrolysis
The purified ManA and ManB were tested forother enzymatic activities. No xylanase, amylase,cellulase, �-galactosidase and �-mannosidase ac-tivities were detected using an incubation time of
B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277 273
Fig. 1. Chromatograms of each step in the M. edulis �-mannanase purification process. Details of each experiment are described inSection 2. (A) IMAC on STREAMLINE Chelating. Arrows indicate where the buffer changes were made. (A) Equilibration buffercontaining 50 mM imidazole, (B) Equilibration buffer containing 50 mM EDTA. (B) SEC on Superdex 75. (C) IEC on Mono S HR10/10. (D) SEC on Superdex 200 under denaturing conditions. (E) IEC on Mono S HR 5/5. The solid lines in all chromatogramsrepresent the UV absorbance at 280 nm and in (A) and (B) the dashed lines represent the �-mannanase activity. For (D) and (E)similar curves were obtained for ManB.
4 h. No binding to cellulose or mannan could beobserved (data not shown).
The purified ManA and ManB show the samepattern of hydrolysis, thus they do not hydrolyse
mannobiose and mannotriose (data not shown).ManA cleaves mannopentaose mainly to manno-biose and mannotriose and mannotetraose (Fig.5). Mannotetraose can be hydrolyzed to mannose
B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277274
and mannotriose plus little mannobiose, but thehydrolysis rate is very low in comparison with thehydrolysis of mannopentaose. After 24 h incuba-tion, 73% of the mannotetraose still remains in-tact. The main products of hydrolysis of ivory nutmannan are similar to the hydrolysis pattern ofmannopentaose. The results of the hydrolysis ex-periments indicate that the enzymes are endo-�-mannanases in conformity with data from studiesof �-mannanases from other organisms.
4. Discussion
Due to the relatively high viscosity and turbid-ity of the crude extracts from the blue musseldigestive glands, we choose the large particle di-ameter (average dp 200 �m) STREAMLINEChelating, saturated with zinc ions, as an IMACabsorbent for the initial capture step in stirredbatch adsorption mode. The strong binding of the�-mannanases and the high sedimentation rate ofthis adsorbent made the washing procedure both
Fig. 3. Isoelectric focusing analysis in polyacrylamide gel of�-mannanases from the last IEC step. Lane 1: ManA; lane 2:ManB. 0.2 �g protein was loaded to the gel. The sample lanesare surrounded by lanes containing the pI calibration kitproteins (pH 3–10).
Fig. 2. SDS-PAGE of samples from the different purificationsteps. Lanes 1 and 9: LMW calibration kit proteins; lane 2:crude extract; lane 3: �-mannanase fraction after IMAC; lane4: �-mannanase fraction after SEC; lanes 5 and 6: ManA andManB after the first IEC step; lanes 7 and 8: ManA and ManBafter the second IEC step. Each lane contains 0.5 �g protein.The gel was stained with silver nitrate. The positions of theMw-standard proteins are marked at the side.
easier and faster. The use of the STREAMLINEChelating in expanded bed mode turned out to beless feasible due to clogging of the net in thecolumn bottom end piece.
Purity analysis of the material in the peaksfrom the IEC step using native-PAGE showedonly one strong band even when using the silverstaining method. However, SDS-PAGE analysisrevealed several low-molecular weight bands inaddition to the �-mannanase band. Our interpre-tation is that there are complex forming proteinspresent in the sample that causes this phe-nomenon. The problem was finally solved usingSEC on Superdex 200 16/60 pg in the presence of6 M Gu–HCl to remove the contaminatingproteins. It seems as if the denaturing conditiondissociates the �-mannanase–protein complexes.Fortunately, the �-mannanases can be easily rena-tured by removing the Gu–HCl by SEC on Sep-hadex G-25 without significant loss of activity.The remaining contaminants were then removedby a rerun on Mono S HR 5/5 column after
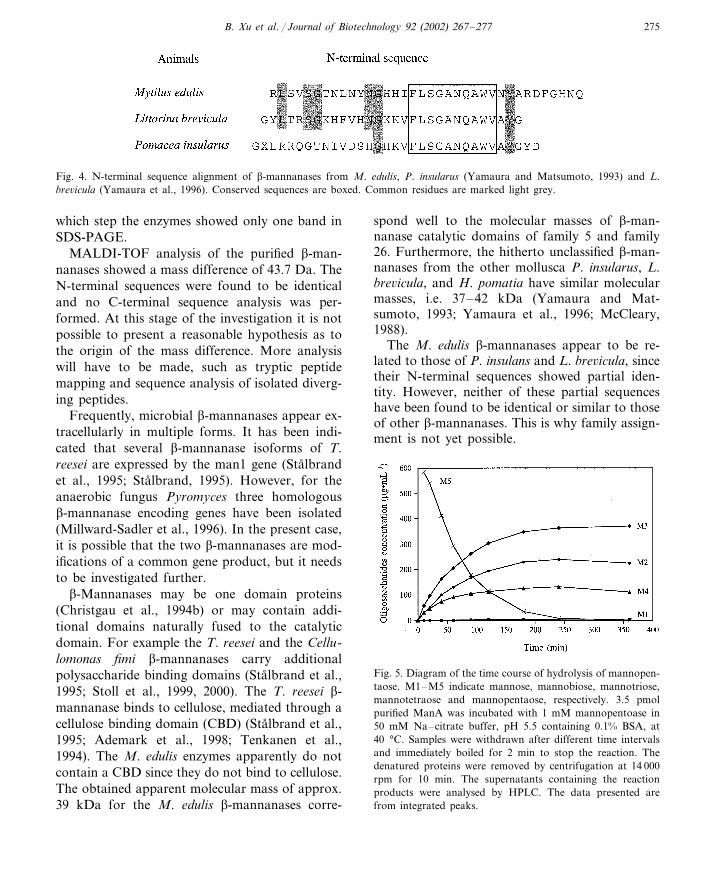
B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277 275
Fig. 4. N-terminal sequence alignment of �-mannanases from M. edulis, P. insularus (Yamaura and Matsumoto, 1993) and L.bre�icula (Yamaura et al., 1996). Conserved sequences are boxed. Common residues are marked light grey.
which step the enzymes showed only one band inSDS-PAGE.
MALDI-TOF analysis of the purified �-man-nanases showed a mass difference of 43.7 Da. TheN-terminal sequences were found to be identicaland no C-terminal sequence analysis was per-formed. At this stage of the investigation it is notpossible to present a reasonable hypothesis as tothe origin of the mass difference. More analysiswill have to be made, such as tryptic peptidemapping and sequence analysis of isolated diverg-ing peptides.
Frequently, microbial �-mannanases appear ex-tracellularly in multiple forms. It has been indi-cated that several �-mannanase isoforms of T.reesei are expressed by the man1 gene (Stalbrandet al., 1995; Stalbrand, 1995). However, for theanaerobic fungus Pyromyces three homologous�-mannanase encoding genes have been isolated(Millward-Sadler et al., 1996). In the present case,it is possible that the two �-mannanases are mod-ifications of a common gene product, but it needsto be investigated further.
�-Mannanases may be one domain proteins(Christgau et al., 1994b) or may contain addi-tional domains naturally fused to the catalyticdomain. For example the T. reesei and the Cellu-lomonas fimi �-mannanases carry additionalpolysaccharide binding domains (Stalbrand et al.,1995; Stoll et al., 1999, 2000). The T. reesei �-mannanase binds to cellulose, mediated through acellulose binding domain (CBD) (Stalbrand et al.,1995; Ademark et al., 1998; Tenkanen et al.,1994). The M. edulis enzymes apparently do notcontain a CBD since they do not bind to cellulose.The obtained apparent molecular mass of approx.39 kDa for the M. edulis �-mannanases corre-
spond well to the molecular masses of �-man-nanase catalytic domains of family 5 and family26. Furthermore, the hitherto unclassified �-man-nanases from the other mollusca P. insularus, L.bre�icula, and H. pomatia have similar molecularmasses, i.e. 37–42 kDa (Yamaura and Mat-sumoto, 1993; Yamaura et al., 1996; McCleary,1988).
The M. edulis �-mannanases appear to be re-lated to those of P. insulans and L. bre�icula, sincetheir N-terminal sequences showed partial iden-tity. However, neither of these partial sequenceshave been found to be identical or similar to thoseof other �-mannanases. This is why family assign-ment is not yet possible.
Fig. 5. Diagram of the time course of hydrolysis of mannopen-taose. M1–M5 indicate mannose, mannobiose, mannotriose,mannotetraose and mannopentaose, respectively. 3.5 pmolpurified ManA was incubated with 1 mM mannopentoase in50 mM Na–citrate buffer, pH 5.5 containing 0.1% BSA, at40 °C. Samples were withdrawn after different time intervalsand immediately boiled for 2 min to stop the reaction. Thedenatured proteins were removed by centrifugation at 14 000rpm for 10 min. The supernatants containing the reactionproducts were analysed by HPLC. The data presented arefrom integrated peaks.

B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277276
As for other mollusc �-mannanases, the M.edulis enzymes have a high content of basic aminoacids, and consequently relatively high isoelectricpoints, about 7.8. In contrast, many bacterial aswell as fungal �-mannanases have got acidic pIs(Viikari et al., 1993). The P. insularus and L.bre�icula �-mannanases are inhibited by Ag+
ions, whereas the M. edulis enzymes are not.The Km values for locust bean gum galactoman-
nan of 3.95 mg ml−1 for the M. edulis ManA isconsiderably higher than the values reported forthe H. pomatia enzyme (0.3 mg ml−1) and alsohigh compared to some previously reported bacte-rial and fungal �-mannanases (0.0015–1.1 mgml−1) (McCleary, 1988; Stoll et al., 1999; Arisan-Atac et al., 1993).
The acidic pH optimum (5.2) for the M. edulisenzymes is similar to, or higher than, those offungal �-mannanases that usually lie between 3.0and 5.5 (Stalbrand et al., 1993; Christgau et al.,1994b; Ademark et al., 1998; Viikari et al., 1993).However, it is lower than those of bacterial �-mannanases which have optima close to neutralpH (Viikari et al., 1993). The M. edulis enzymesdisplay a relatively low thermostability in com-parison to many �-mannanases from non-ther-mophilic microorganisms that are stable within aninterval of several pH units for several hours at50 °C (Stalbrand et al., 1993; Ademark et al.,1998). The �-mannanase of P. insularus was un-stable at a temperature as low as 8 °C whenincubated for 16 h at around pH 5 (McCleary,1988).
As for many �-mannanases, including the T.reesei and A. niger enzymes (Stalbrand et al.,1993; Ademark et al., 1998; Harjunpaa et al.,1995, 1999; Torto et al., 1995, 1996), the majorproducts from hydrolysis of mannopentaose andivory nut mannan by the M. edulis ManA aremannobiose and mannotriose. An accumulationof mannotetraose also after 24 h incubation withivory nut mannan was observed for the M. edulisenzyme. Such accumulation has also been ob-served, at similar enzyme loadings, when the T.reesei and A. niger enzymes were incubated withivory nut mannan (Torto et al., 1995, 1996). Therelease of significant amounts of mannotetraosecombined with the fact that only trace quantities
of mannose can be detected, indicates that the M.edulis �-mannanases are capable of performingtransglycosylation reactions. This is in accordancewith the fact that classified �-mannanases belongto family 5 and family 26 of glycoside hydrolases,both having the retaining mechanism. Transglyco-sylation has been shown for several �-man-nanases, including those from A. niger and T.reesei (Ademark et al., 1998; Harjunpaa et al.,1995, 1999). The overall pattern of hydrolysis bythe M. edulis �-mannanases appears to be similarto that of these enzymes. However, a more de-tailed kinetic study need to be carried out in orderto detect possible differences in the subsite sub-strate binding requirements.
Acknowledgements
We are most grateful to the R&D Managementof Amersham Pharmacia Biotech in Uppsala fortheir financial support of the adjunct professor-ship of J.-C. J. that made this work possible. H.S.thanks the Swedish Research Council for Engi-neering Sciences (TFR) for financial support.
References
Ademark, P., Varga, A., Medve, J., Harjunpaa, V., Draken-berg, T., Tjerneld, F., Stalbrand, H., 1998. Soft-woodhemicellulose-degrading enzymes from Aspergillus niger :purification and properties of a �-mannanase. J. Biotech-nol. 63, 199–210.
Arisan-Atac, I., Hodits, R., Kristufek, D., Kubicek, C.P.,1993. Purification and characterization of a �-mannanaseof Trichoderma reesei C-30. Appl. Microbiol. Biotechnol.39, 58–62.
Bailey, M.J., Biely, P., Poutanen, K., 1992. Interlaboratorytesting of methods for assay of xylanase activity. J. Bio-technol. 23, 257–270.
Bernfeld, P., 1955. Amylases, �- and �-. Methods Enzymol. 1,149–158.
Bradford, M.M., 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizingthe principle of protein–dye binding. Anal. Biochem. 72,248–254.
Christgau, S., Andersen, L.N., Kauppinen, S., Heldt-Hansen,H.P., Dalboege, H., 1994. Purified enzyme exhibiting man-nanase activity; application in oil, paper, pulp, fruit andvegetable juice industry and in carrageenan extraction,Patent Novo-Nordisk, 9425576, 10 November 1994.

B. Xu et al. / Journal of Biotechnology 92 (2002) 267–277 277
Christgau, S., Kauppinen, S., Vind, J., Kofod, L.V., Dalbøge,H., 1994b. Expression cloning, purification and characteri-zation of a �-1,4-mannanase from Aspergillus aculeatus.Biochem. Mol. Biol. Int. 33, 917–925.
Duffaud, G.D., McCutchen, C.M., Leduc, P., Parker, K.N.,Kelly, R.M., 1997. Purification and characterization ofextremely thermostable �-mannanase, �-mannosidase, and�-galactosidase from the hyperthermophilic eubacteriumThermotoga meapolitana 5068. Appl. Environ. Microbiol.63, 169–177.
Gubitz, G.M., Hayn, M., Urbanz, G., Steiner, W., 1996.Purification and properties of an acidic �-mannanase fromSclerotium rolfsii. J. Biotechnol. 45, 165–172.
Harjunpaa, V., Teleman, A., Siika-aho, M., Drakenberg, T.,1995. Kinetic and stereochemical studies of manno-oligosaccharide hydrolysis catalysed by �-mannanase fromTrichoderma reesei. Eur. J. Biochem. 234, 278–283.
Harjunpaa, V., Helin, J., Koivula, A., Siika-Aho, M., Draken-berg, T., 1999. A comparative study of two retainingenzymes of Trichoderma reesei : transglycosylation ofoligosaccharides catalysed by the cellobiohydrolase I,Cel7A, and the �-mannanase, Man5A. FEBS Lett. 443,149–153.
Henrissat, B., Bairoch, A., 1993. New families in the classifica-tion of glycosyl hydrolases based on amino-acid sequencesimilarities. Biochem. J. 293, 781–788.
Matheson, N.K., McCleary, B.V., 1985. In: Aspinall, G.O.(Ed.), The Polysaccharides, vol. 3. Academic Press, NewYork, p. 1.
McCleary, B.V., Matheson, N.K., 1986. Enzymic analysis ofpolysaccharide structure. In: Tipson, R.S., Horton, D.(Eds.), Adv. Carbohydr. Chem. Biochem. Academic Press,UK, pp. 147–276.
McCleary, B.V., 1988. �-D-Mannanase. Methods Enzymol.160, 596–614.
McCleary, B.V., 1990. Comparion of endolytic hydrolases thatdepolymerize 1,4-�-D-mannan, 1,5-�-L-arabinan and 1,4-�-D-galaktan. ACS Symp. Ser. 460, 437–449.
Meier, H., 1958. On the structure of cell walls and cell wallmannans from ivory nuts and from dates. Biochem. Bio-phys. Acta 28, 229–240.
Millward-Sadler, S.J., Hall, J., Black, G.W., Hazlewood, G.P.,Gilbert, H.J., 1996. Evidence that the Piromyces genefamily encoding endo-1,4-mannanases arose through geneduplication. FEMS Microbiol. Lett. 141, 183–188.
Paice, M.G., Jurasek, L., 1984. Removing hemicellulose frompulps by specific enzymatic hydrolysis. J. Wood Chem.Technol. 4, 187–198.
Purchon, R.D., 1977. The Biology of the Mollusca, second ed.Pergamon, Oxford.
Ratto, M., Poutanen, K., 1988. Production of mannan-degrad-ing enzymes. Biotechnol. Lett. 10, 661–664.
Stalbrand, S., Siika-aho, M., Tenkanen, M., Viikari, L., 1993.Purification and characterization of two �-mannanasesfrom Trichoderma reesei. J. Biotechnol. 29, 229–242.
Stalbrand, H., Saloheimo, A., Vehmaanpera, J., Henrissat, B.,Penttila, M., 1995. Cloning and expression in Saccha-romyces cere�isiae of a Trichoderma reesei �-mannanasegene containing a cellulose binding domain. Appl. Envi-ron. Microbiol. 61, 1090–1097.
Stalbrand, H., 1995. Hemicellulose-degrading enzymes fromfungi. Characterization of �-mannanase and the man1 geneof Trichoderma reesei. Department of Biochemistry, LundUniversity, Lund, Sweden Ph.D. thesis.
Stoll, D., Stalbrand, H., Warren, R.A.J., 1999. Mannan-de-grading enzymes from Cellulomonas fimi. Appl. Environ.Microbiol. 65, 2598–2605.
Stoll, D., Boraston, A., Stalbrand, H., McClean, B.W., Kil-burn, D.G., Warren, R.A.J., 2000. Mannanase Man 26Afrom Cellulomonas fimi has a mannan binding module.FEMS Microbiol. Lett. 183, 265–269.
Tamaru, Y., Araki, T., Amagoi, H., Mori, H., Morishita, T.,1995. Purification and characterization of an extracellular�-1,4-mannanase from a marine bacterium, Vibrio sp.Strain MA-138. Appl. Environ. Microbiol. 61, 4454–4458.
Tenkanen, M., Buchert, J., Viikari, L., 1994. Binding ofhemicellulases on isolated polysaccharide substrates. En-zyme Microb. Technol. 17, 499–505.
Torto, N., Buttler, T., Gorton, L., Marko-Varga, G.,Stalbrand, H., Tjerneld, F., 1995. Monitoring of enzymatichydrolysis of ivory nut mannan using on-line microdialysissampling and anion-exchange chromatography with inte-grated pulsed electrochemical detection. Anal. Chem. Acta313, 15–24.
Torto, N., Marko-Varga, G., Gorton, L., Stalbrand, H., Tjer-neld, F., 1996. On-line quantitation of enzymatic mannanhydrolysates in small-volume bioreactors by microdialysissampling and column liquid chromatography-integratedpulsed electrochemical detection. J. Chromatogr. 725, 165–175.
Viikari, L., Tenkanen, M., Buchert, J., Ratto, M., Bailey, M.,Siika-aho, M., Linko, M., 1993. Hemicellulases for indus-trial applications. In: Saddler, J.N. (Ed.), Bioconversion ofForest and Agricultural Plant Residues. CAB Interna-tional, Wallingford, pp. 131–182.
Wood, T.M., Bhat, K.M., 1988. Methods for measuring cellu-lase activities. Methods Enzymol. 160, 87–112.
Yamaura, I., Matsumoto, T., 1993. Purification and someproperties of endo-1,4-�-D-mannanase from a mud snail,Pomacea insularus (de Ordigny). Biosci. Biotech. Biochem.57, 1316–1319.
Yamaura, I., Nozaki, Y., Matsumoto, T., Kato, T., 1996.Purification and some properties of an endo-1,4-�-D-man-nanase from a marine mollusc, Littorina bre�icula. Biosci.Biotech. Biochem. 60, 674–676.

VI
135

Degradation of glucomannan and O-acetyl-galactoglucomannan by mannoside- and
glucoside-hydrolases
Jon Lundqvist1, Per Hägglund1*, Torny Eriksson1, Per Persson2, Dominik Stoll3,
Matti Siika-aho4, Lo Gorton2 and Henrik Stålbrand1
1Department of Biochemistry, Center for Chemistry and Chemical Engineering, Lund University,P.O. Box 124, S-221 00, Lund, Sweden
2Department of Analytical chemistry, Center for Chemistry and Chemical Engineering, Lund University,P.O. Box 124, S-221 00, Lund, Sweden
3Department of Microbiology and Immunology, The University of British Columbia, Vancouver,British Columbia, Canada
4VTT Biotechnology, P.O. Box 1500, Espoo FIN-02044, Finland
*Corresponding author: Department of Biochemistry, Centre for Chemistry and Chemical Engineering,University of Lund, Lund, P.O. Box 124, S-221 00, Lund, Sweden.
Tel: +46 46 222 9854 Fax: +46 222 4534. e-mail:[email protected]
Abstract
The plant cell wall is a tissue with cellulose, hemicellulose and lignin, tightly associated throughvarious interactions. Cellulases are therefore likely to be affected by the presence of thehemicellulose components, and in the same way hemicellulases are probably affected by cellulose.In this study we have analysed the degradation of mannan-based hemicelluloses (glucomannanand O-acetyl-galactoglucomannan) by hemicellulases and cellulases. The products were analysedby reducing sugar analysis, size-exclusion chromatography and HPLC analysis. As expected, theβ-mannanases used in this study degraded the mannans to oligosaccharides. More surprisingly,the endoglucanase Cel7B from Trichoderma reesei also displayed activity against bothglucomannan and O-acetyl-galactoglucomannan. Endoglucanase Cel45A from T. reesei was activeagainst glucomannan. Furthermore, β-mannosidase from Aspergillus niger released mannosefrom both glucomannan and O-acetyl-galactoglucomannan. Mass spectrometry analysis revealedthat acetylated mannose can also be released by this β-mannosidase.
137

Introduction
Mannans and heteromannans are among the most abundant biopolymers on earth. Linear b-(1Æ4)-D- mannans are found in the seed endosperms of numerous plants (Meier, 1958; Wolfrom et al.,1961), and also in the cell walls of some green algae (Frei & Preston, 1968; Ikiri & Miwa, 1960).Galactomannans, which are composed of a linear mannan chain with some a-(1Æ6)-linkedgalactosyl side-groups, are found mainly in the seeds of leguminous plants (Reid, 1985). Themost abundant mannan-based polysaccharides, in terms of biomass, are the O-acetyl-galactoglucomannans (O-acetyl-GGM) and glucomannans, which are found as hemicelluloses insoftwoods and hardwoods, respectively (Timell, 1967). Besides in wood, glucomannan are alsofound as reserve polysaccharides in the root and tubers of some annual plants (Meier & Reid,1982). Glucomannan is built up by a linear chain of b-(1Æ4)-linked mannose and glucose units,which may be acetylated. O-acetyl-GGM is built up by a glucomannan main chain with a-(1Æ6)-linked galactosyl branches (Figure 1). Acetyl groups may be attached at the C-2 and C-3 posi-tions of some mannose residues (Lindberg et al., 1973; Timell, 1967).
The complete degradation of the complex mannan-based hemicelluloses require the presence ofseveral hemicellulose-degrading enzymes. Endo-β-1,4-D-mannanase (β-mannanase; EC 3.2.1.78)is the major endo-hydrolase involved in mannan hydrolysis, degrading glucomannans andgalactoglucomannans into smaller soluble oligosaccharides. These oligosaccharides can subse-quently be further degraded by several exo-hydrolases, namely β-mannosidase (EC 3.2.1.25), β-glucosidase (EC 3.2.1.21) and α-galactosidase (EC 3.2.1.22). β-Mannosidase and β-glucosidaseattack the main chain of oligosaccharides from its non-reducing ends and release mannose andglucose residues, respectively. α-Galactosidase release galactosyl side-groups, apparently with apreference for either oligomeric or polymeric substrates depending on the enzyme source (Ademarket al., 2001a; Ademark et al., 2001b). Acetyl groups may be removed by acetyl (mannan) esterase(Tenkanen et al., 1995).
β-Mannanases have been purified from a wide range of microbes (Ademark et al., 1998; Stoll etal., 1999; Stålbrand et al., 1993; Sunna et al., 2000) and also from some higher organisms (Xu etal., 2002). Based on sequence similarities, all β-mannanases characterised this far have beenassigned to family 5 and 26 of glycoside hydrolases, according to the classification of Henrissatet al. (1991). β-Mannosidases have also been found in a wide range of organisms, and most β-mannosidases analysed have been classified in family 2 (Ademark et al., 2001a). Families 2, 5and 26 are all included in the GH-A clan, which share the (β/α)
8 barrel structural fold (Jenkins et
al., 1995). Included in the GH-A clan is also family 1, which includes some β-glucosidases. α-Galctosidases are found in family 4, 27 and 36.
The degradation patterns of the β-mannanases from Trichoderma reesei and Aspergillus niger ondifferent heteromannans have been studied previously (McCleary & Matheson, 1983; Puls &Schuseil, 1993; Rättö et al., 1993; Tenkanen et al., 1997). These studies have provided someinformation about the sub-site specificity of these enzymes, but the influence of acetyl groups onthe hydrolysis has not been examined in detail. We and others have previously shown that at leastsome β-mannosidases can degrade mannan polymers (Ademark et al., 1999; Kulminskaya et al.,1999), and we have also shown that the β-mannosidase Man2A from Aspergillus niger (previ-ously called Mnd2A) degrade even highly crystalline polymeric mannan (Hägglund et al., 2001).
138

However, only sparse information is available on the degradation of glucomannan by β-mannosidase (Gübitz et al., 1996; Hirata et al., 1998). We and others have previously reportedthat some endoglucanases depolymerise glucomannan (Chhabra et al., 2002; Karlsson, 2000).The rationality behind glucomannan degradation by endoglucanases is unclear and the detaileddegradation patterns have not yet been investigated.
Previously we have detected activity toward O-acetyl-GGM by β-mannanase and α-galactosi-dase (Lundqvist et al., 2002b), but otherwise little information is published on the degradation ofthis polysaccharide. In this study, we have analysed the degradation of glucomannan and O-acetyl-GGM by a series of different mannan-degrading enzymes and endoglucanases (Table 1).We have used an isolated fraction of O-acetyl-GGM from spruce (Lundqvist et al., 2002b) and aglucomannan from Amorphophallus konjac (so called konjac glucomannan (KM)) as substratesfor these enzymes and we have studied the products after prolonged degradation. In the nativestate KM has a molecular weight of 1000000 (Nishinari et al., 1992). Native O-acetyl-GGM hasa degree of polymerisation between 100-150 (Timell, 1967).
The results of this study indicate that the T. reesei endoglucanase Cel7B, can depolymerise O-acetyl-GGM to a limited extent. Furthermore, we present results which show that Man2A canrelease acetylated mannose residues from the non-reducing end of O-acetyl-GGM.
Materials and methods
SubstratesKonjac glucomannan (KM) from the tubers of Amorphophallus konjac plants was purchasedfrom Megazyme (Bray, Ireland). O-acetyl-galactoglucomannan (O-acetyl-GGM) from spruce waspreviously isolated in our group (Lundqvist et al., 2002b). The O-acetyl-GGM was isolated froma filtrate obtained by heat fractionation of spruce and has a molecular weight of 2600. It is mostlikely a fragment of the native polysaccharide and displayed a monomer composition similar tolow galactosyl-substituted O-acetyl-GGM (Lundqvist et al., 2002b). O-acetyl-GGM and KMwere solubilised in the appropriate buffer before the enzymatic treatment.
EnzymesThe catalytic module (amino acid 1-352) of the Trichoderma reesei β-mannanase Man5A (Stålbrandet al., 1995), was expressed in T. reesei under control by the gpdA promoter from Aspergillusnidulans and purified as described earlier (Hägglund et al., 2002). The catalytic module (aminoacid 1-371) of the T. reesei endoglucanase Cel7B (Penttilä et al., 1986), was expressed in T. reeseiunder control of the gpdA promoter from A. nidulans, as described previously (Collén et al.,2001). Cel7B was purified by anion-exchange chromatography with Source Q (AmershamPharmacia Biotech), using 20 mM Na-acetate (pH 4.5) as buffer A and 1 M Na-Acetate (pH 4.5)as buffer B. Cel7B was eluted with a linear gradient of buffer B. Man2A from A. niger wasoverexpressed in a multi-copy strain of A. niger as described by Ademark et al (Ademark et al.,2001a), and purified as described previously (Ademark et al., 1999). The A. niger α-galactosi-dase Agal II was purified according to Ademark (Ademark et al., 2001b).
139

The catalytic module (amino acid 1-182, including signal peptide) of Cel45A (Saloheimo et al.,1994) from T. reesei was purified from the culture filtrate of a T. reesei strain constructed forexpression of the protein in glucose-containg medium, as described by earlier (Nakari-Setäla &Penttilä, 1995). The culture filtrate was adjusted to pH 6.0 and applied to a column with DEAESepharose FF. The protein was eluted with a NaCl gradient from 0 - 0.2 M. The fractions contain-ing Cel45A were pooled and the buffer was adjusted to 0.02 M sodium acetate (pH 5.0) and 0.5 M(NH
4)
2SO
4. The pooled fractions were applied to a column with Phenyl Sepharose FF. For elution
a gradient of decreasing concentration of sodium acetate and (NH4)
2SO
4 was used where Cel45A
was eluted with 1 mM sodium acetate. The pooled fractions with Cel45A were adjusted to pH 3.0with 5 mM acetic acid/sodium acetate and applied to a SP Sepharose FF column. Cel45A waseluted with a NaCl gradient from 0 - 0.5 M.
The catalytic module (amino acid 1-464) of Cellulomonas fimi Man26A (Stoll et al., 1999) wasexpressed from a pET vector in Escherichia coli BL21 and purified essentially as describedpreviously (Stoll et al., 1999). Cells were ruptured in a French pressure cell and equilibrated in abinding buffer (5 mM imidazole, 500 mM NaCl, 20 mM Tris-HCl, pH 7.9). The equilibratedsample was applied on a 5 ml column packed with His Bind Resin (Novagen, Madison, WI,USA) at a flow rate of 1 ml/min and eluted by an stepwise increase in imidazole concentrationfrom 5 mM to 500 mM. A commercial preparation of A. niger β-glucosidase (β-Gluc) was used(Megazyme). All purified enzymes appeared as single bands on SDS-PAGE (Laemmli, 1970)after silver staining.
Enzyme activity assaysThe β-mannosidase/β-glucosidase activity ratio in the Man2A and β-Gluc preparations were as-sayed essentially as described previously (Rättö & Poutanen, 1988) at pH 4.5 using 2 mM p-nitrophenyl-β-D-glucopyranoside (Sigma, St Louis, MO, USA) and 2 mM p-nitrophenyl-β-D-mannopyranoside (Sigma).
Dot-blot analysisAll steps were performed at room temperature. Drops of 4 µl containing either Man5A or Cel7Bwere placed on a Biotrace NT blotting nitrocellulose membrane (Pall Gelman Sciences, AnnArbor, MI, USA) and allowed to dry for 15 min. The membrane was placed in blocking solution(TBS buffer (20 mM Tris, 137 mM NaCl, pH7.6), 0.1 % Tween and 1.5 % Bovine serum albumin(BSA)) for one hour with gentle shaking. The membrane was washed twice for 5 min in washingbuffer (TBS, 0.1 % Tween). New TBS buffer including polyclonal antibodies (diluted 1/1000)against Man5Araised in rabbit, was added and the mebrane was incubated for 1 h.
After two washes as described above, new TBS buffer containing secondary goat anti-rabbitantibodies (Bio-Rad (Hecules, CA, USA)) diluted 1/1500 was added, and the membrane wasincubated for 1 h. After two washes in Tris buffer (0.1 M Tris, 0.5 mM MgCl
2, pH9.5), Tris buffer
containing Nitroblue tetrazolium chloride (300 µg/ml) and AP Color Development Reagent (150µg/ml) (Bio-Rad, Hercules, CA, USA) solutions was added, and the mixture was incubated untilspots were clearly visible (approximately 60 minutes). The incubation was ended, by immersingthe membrane in distilled water for 10 min.
140

Enzymatic hydrolysis of konjac glucomannan and galactoglucomannanAll enzymatic incubations were carried out at 40 ºC in 50 mM ammonium acetate buffer, pH 4.8,except Man26A which was incubated in 50 mM ammonium acetate buffer, pH 6.2. In all enzymaticincubations BSA (100 µg/ml) was included and in all incubations the enzyme concentration was0.1 µM. Man5A, Man26A, Cel7B and Cel45A were incubated with KM (5 mg/ml) for 72 h andMan2A and β-glucosidase were incubated with KM (5 mg/ml) for 92 h. Man2A, Man5A, Cel7B,Agal II and β-Gluc were incubated with 10 mg/ml O-acetyl-GGM for 92 h.
In order to ensure that all incubations were hydrolysed to completion, samples were withdrawn atdifferent time points and subjected to reducing sugar analysis, as described below. The differencein amount of reducing sugars in the two last time points, which were separated by 24 h, did notexceed 12 % in any incubation. All enzymatic reactions were terminated by boiling the samplesfor 2 minutes.
Reducing sugar analysisSamples from the enzymatic incubations were withdrawn and analysed for reducing sugars using3,5-dinitrosalicylic acid (DNS), according to Sumner and Somers (1949), using a mannose (FlukaChemie AG, Buchs, Switzerland ) standard curve. Corrections were made for reducing sugarspresent in parallel control incubations of polysaccharides without enzyme.
Size exclusion chromatographyThe end products formed after hydrolysis of KM and O-acetyl-GGM by the β-mannanases andendoglucanases were separated using an FPLC-system (Pharmacia Biotech, Uppsala, Sweden)with refractive index (RI)- (Erma-inc, Tokyo, Japan) and ultra violet (UV)- (Pharmacia Biotech,280nm) detectors. Two size exclusion chromatography (SEC) systems were set up; SEC1 fordetermination the molecular weight distribution (MWD) of polysaccharides of and SEC2 foranalysis of oligosaccharides with degree of polymerization (DP) from 1-7.
SEC1: The separations (Molecular weight-range of 1000-40000) were performed using gel filtra-tion media from Pharmacia Biotech . In this SEC system three columns were connected in series,(i) a precolumn Superdex 75 (HR 5/5), (ii) a Superdex 75 (HR 10/30), and (iii) a Superdex 200(HR 10/30) column, (Pharmacia Biotech). The MWD was determined using O-acetyl-GGM iso-lated previously as standards (Lundqvist et al., 2002a). The molecular weight of these GGMstandards were determined previously by matrix assisted laser desorption/ionisation massspectrometry (Lundqvist et al., 2002a). A sample volume of 500 µl was loaded on the columnsand water was used as mobile phase at a flow rate of 0.5 ml/min in room temperature. Acetonewas used to determine the total volume of the columns (47 ml).
SEC2: The oligosaccharide analysis (DP 1-9, molecular weight 100-1500) was performed usingBio-Gel P-2 Gel, Fine, gel filtration media from Bio-Rad Laboratories. The media was packed ina Pharmacia column (HR 30/95) with a total volume of 670 ml. Manno-oligosaccharides (DP1-5)from Megazyme were used as calibration standards. A sample volume of 500 µl was loaded onthe column and water was used as mobile phase at a flow rate of 0.3 ml/min in room temperature.Standards were manno-oligosaccharides of DP 1-6 (Megazyme)
141

HPLC-analysisFor the determination of released monosaccharides after hydrolysis of the oligo- andpolysaccharides, High Performance Anionic Exchange Chromatography with Pulsed AmperometricDetection (HPAEC-PAD) (DIONEX, Sunnyvale Ca, USA) was used. The HPAEC-PAD con-sisted of an AS50 autosampler, a GP40 gradient pump and Carbo Pac Pa10 guard and analyticalcolumn and an ED40 electrochemical detector. The flow rate using water elution was 1 ml/minand the injection volume was 10 µl. A post-column pump (DIONEX) with 200 mM NaOH aseluent was used to enable detection of the sugars. D-mannose, D-xylose, D-glucose, D-galactose,L-arabinose (Fluka Chemie) was used as standards for the analysis.
MS-analysisThe mass spectrometry analysis of hexoses and acetylated hexoses was performed on an Esquire-LC ion-trap mass spectrometer equipped with an API-electrospray interface operated in positiveionisation mode (Bruker Daltonics, Bremen, Germany). The mass spectrometer was set to scan inthe mass-to-charge ratio (m/z) range of 50-310. Nitrogen was used as a drying gas and was pumpedinto the interface at a rate of 3 L/min and at a temperature of 350°C. Nitrogen was also used asnebuliser gas and was kept at 10 psi. The nitrogen gas was supplied by a nitrogen generator fromWhatman Inc. (Haverhill, MA, USA). The following voltages were used: HV capillary 4000 V,HV end plate offset -500 V, capillary exit offset 65 V, skimmer 1 35 V and skimmer 2 8 V. Thesample was continuously injected into the interface by a syringe pump (Cole Parmer 74900 se-ries, Cole Parmer Instrument Company, Vernon Hills, IL, USA). A 250 µl syringe (Hamilton,Reno, NV, USA) was used for infusion and the injection speed was 90 µl/h. Five parts of thesample was mixed with one part of a sodium acetate solution (3.8 mg/ml) prior to injection.
Results
In this study, konjac glucomannan (KM) and O-acetyl-galactoglucomannan (O-acetyl-GGM) havebeen hydrolysed by several hemicellulases and cellulases (Table 1) Two endoglucanases (Cel7Band Cel45A), two β-mannanases (Man5A and Man26A) and a β-mannosidase (Man2A). Degra-dation patterns, reducing ends and released monosaccharides have been established by size ex-clusion chromatography (SEC), reducing sugar assay, high performance anionic exchange chro-matography (HPAEC) and mass spectrometry (MS). Two SEC systems were used; SEC1 for theanalysis of products in the molecular weight-range of 1000-40000 (DP 6-250) and SEC2 for theanalysis of products in the molecular weight-range of 100-1500 (DP 1-9)
The properties of the substrates used have been determined previously (Lundqvist et al., 2002b;Nishinari et al., 1992). KM has a mannose:glucose ratio of 1.6:1 and an molecular weight
of
1000000. It has a degree of acetylation (DSAc
) of 0.07. The O-acetyl-GGM fraction used wasisolated as described previously (Lundqvist et al., 2002b). It is an isolated fragment of O-acetyl-GGM hemcellulose isolated from steam fractionated spruce. This preparation has agalactosyl:glucosyl:mannosyl ratio of 0.1:1:4 with a DS
Ac of 0.28 and a molecular weight of
2600. A schematic picture of O-acetyl-GGM is shown in Figure 1. The KM and O-acetyl-GGMused solubilised in the appropriate buffer before incubation with the enzymes.
142

Hydrolysis of KMKM (5 mg/ml) was hydrolyzed by Cel7B, Cel45A, and Man5A from Trichoderma reesei andMan26A from Cellulomonas fimi to completion. Reducing ends and mono- and oligosaccharidesobtained after hydrolysis were analysed. The reducing end concentrations estimated after degra-dation with Cel7B, Cel45A, Man5A and Man26A on KM were 2.7, 1.7, 2.5 and 2.2 mg/ml,respectively. The elution profile from SEC2 of the oligosaccharides formed after KM hydrolysisby the different enzymes are shown in Figure 2. The distribution of the products formed afterhydrolysis of KM by all the enzymes was between DP 1-6, except for Cel45A where it wasessentially DP 6 and higher (Figure 2B). Man5A and Man26A had the highest content of DP 2and 3 with minor production (< 10%) of DP 1 and 3-7 (Figure 2A). The hydrolysis with Cel7Bformed an even distribution of DP 1-7 with the highest production of DP 3 (Figure 2B). Cel45Adid not degrade KM in the same extent as the other enzymes.
A hydrolysis of KM (5 mg/ml) with Man2A and β-Gluc was also performed and the amount ofliberated reducing sugar was analysed. A release of 1.2 µg/ml (Man2A) and 1.6 µg/ml (β-Gluc)was detected when the enzymes were incubated individually. When Man2A and β-Gluc wereused in combination a release of 19 µg/ml reducing sugars was observed.
Hydrolysis of GGMAn hydrolysis of O-acetyl-GGM (10 mg/ml) with Cel7B, Man5A and Man2A was performedwith the objective to investigate the products formed after prolonged hydrolysis. The depolymerisedO-acetyl-GGM was analysed with SEC1 and SEC2. The resulting elution curves for Cel7B andMan5A are displayed in Figure 3 (SEC1) and Figure 4 (SEC2). For Cel7B, a moderate but signifi-cant shift of the chromatogram elution profile toward smaller apparent molecular weight is seen.This is an indication of depolymerising activity of Cel7B on O-acetyl-GGM. As seen in the SEC2chromatogram only small amounts of oligosaccharides of DP 5 or lower could be detected in thesample hydrolysed by Cel7B. The reducing end concentrations after hydrolysis with Cel7B was2.1 mg/ml. Man5A degrades O-acetyl-GGM to a higher extent with a reducing sugar concentra-tion of 4.3 mg/ml. This is also seen in the SEC1 elution profile for the products formed aftertreatment with Man5A which display a major peak of an approximate molecular weight of 1200(Figure 3). The elution profile established by SEC 2 showed that the products formed after treat-ment with Man5A were between DP 2-6 (Figure 4).
O-acetyl-GGM was also hydrolysed by Man5A in combination with Man2A, and by Man2A incombination with Agal II, and β-Gluc. The concentration of reducing ends after hydrolysis usingMan5A in combination with Man2A was slightly higher than for Man5A alone (Table 2). Thehydrolysis by Man5A in combination with Man2A also results in a new peak at molecular weight<500 in the chromatogram of SEC1, compared with incubation of Man5A alone (Figure 3). Thispeak correspond to monomeric products. The combined action of Man2A, Agal II and β-Gluc didnot result in a complete degradation of O-acetyl-GGM into monosaccharides (Table 2). In theincubation with Man2A and β-Gluc, Agal II released about 47 % of the galactosyl groups at-tached to the O-acetyl-GGM. The release of mannose was slightly enhanced compared to theaction with Man2A alone.
143

Dot-blot analysis of Cel7BThe preparation of T. reesei Cel7B used in this study appeared as single bands on SDS-PAGE andisoelectric focusing gels (data not shown). In order to ensure that the depolymerisation of O-acetyl-GGM with the Cel7B preparation was a true endoglucanase activity and not due to acontamination of β-mannanase, the purity of the Cel7B preparation was further assessed. A dot-blot analysis with antibodies directed against Man5A, the only known β-mannanase from T.reesei (Stålbrand et al., 1995), was performed (Figure 5). No visible antibody labeling of theCel7B sample was seen, even if a 10 000 fold more Cel7B (on a molar basis) was applied incorrelation to the amount of Man5A which gave a clearly visible spot.
The acetyl-groups attached to the mannosyl unitsO-acetyl-GGM (10 mg/ml) was incubated with Man2A for 92 h. A control incubation of Man2Awithout O-acetyl-GGM was also included. After the incubations electrospray ionization massspectrometry (ESI-MS) analyses was performed on the samples. The analytes were detected astheir sodium adducts with one positive charge, i.e. hexaose with a molar mass of 180 was de-tected at m/z 203 and the acetylated hexaose was detected at m/z 245. As seen in the MS spectrumof the hydrolyzed sample (Figure 6A), a peak at m/z 245 was detected, indicating that Man2A hasreleased acetylated mannose from O-acetyl-GGM. No significant peak at m/z 245 was detectedin a sample of O-acetyl-GGM incubated without enzyme (Figure 6B).
Discussion
The lignified plant cell wall is a complex mixture of cellulose, hemicellulose, lignin and minoramounts of pectic substances. Although the detailed organisation of these components in the cellwall is not fully understood, several reports have indicated a close association between mannanand cellulose (Åkerholm & Salmén, 2001). In this study, we have investigated the activity of anumber of mannan- and cellulose-degrading enzymes on glucomannan isolated fromAmorphophallus konjac (KM) and O-acetyl-GGM isolated from spruce.
It has been indicated earlier that several endoglucanases from T. reesei have the ability todepolymerise konjac glucomannan (Karlsson, 2000). However, the distribution of releasedoligosaccharides has not been analysed. Our results indicate a difference in product distributionbetween Cel7B and Cel45A. Degradation of KM by Man5A and Man26A yielded similar oli-gosaccharide distributions, indicating that these enzymes might have similar subsite binding prop-erties in glucomannan hydrolysis.
It has previously been demonstrated that Man2A can release mannose from mannan andgalactomannan (Ademark et al., 1999; Hägglund et al., 2001). In this study we also found thatMan2A was capable of liberating sugar from glucomannan. Furthermore the amount of releasedsugars was increased more than ten times when Man2A was incubated with glucomannan in thepresence of β-glucosidase, which may be explained by the following: β-mannosidase hydrolysethe polymeric main-chain until a non-reducing glucose monomer is exposed, which can be hy-drolysed by the β-glucosidase. In the same way the β-glucosidase can subsequently release
144

glucose residues. The activity of the Man2A on glucomannan indicates that a stretch of mannoseunits without interspersed glucose units are present at the non-reducing end of the polymer.Glucomannan degradation has been reported previously for β-mannosidases from Sclerotiumrolfsii, Bacillus subtilis and apple-snails (Gübitz et al., 1996; Hirata et al., 1998).
The action of Man5A and Cel7B from Trichoderma reesei on O-acetyl-GGM showed some inter-esting features. After hydrolysis with Man5A, the oligosaccharides formed were between DP 2-5.The extent of galactose units attached to the isolated O-acetyl-GGM was low (mannosyl:galactosylratio 4:0.1) and would probably not interfere in a higher extent on the action with Man5A. Tenkanenand co-workers have previously investigated the action of Man5A from Trichoderma reesei onnon-acetylated galactoglucomannan (Tenkanen et al., 1997). They showed a degradation patternof DP 2-4 with major extent of 2 and 3. Puls and Schuseil (Puls & Schuseil, 1993), hydrolyzedacetylated GGM from spruce with a β-mannanase from Aspergillus niger, and observed possiblesteric hindrance by acetyl groups. They showed a higher extent of sugar liberation with the β-mannanase using deacetylated GGM compared to acetylated GGM. These results indicate thatthe pattern of distribution of acetyl groups influence the action of some β-mannanases. Morestudies have to be made in order to investigate if acetyl-groups are sterical hinders for the actionof Trichoderma reesei Man5A. Further experiments will be conducted in order to analyse thepossible hindrance by acetyl substituents on Man5A.
The hydrolysis of O-acetyl-GGM by Man2A has not been analysed previously. The release ofacetylated mannose by Man2A established by EIS-MS was an interesting discovery and has notbeen found for any β-mannosidase before. The exact positions of the acetyl groups were notdetermined and further studies will be carried out to in order to determined if the acetyl substituentsare attached at the C-2 and/or C-3 positions on the released acetylmannose residues. Thus, thefamily 2 β-mannosidase studied appear to be able to recognise acetylated mannose residues.However, at least one β-mannanase (Puls & Schuseil, 1993) was clearly restricted by acetylsubstituents. This is an interesting reflection, since all known β-mannanases are classified infamily 5 and 26, and thus belong to the same (β/a)
8-barrel clan (GH-A) of glycoside hydrolases as
Man2A. Further studies will be conducted to reveal differences or similarities in the influence ofacetyl substituents in clan GH-A mannoside-hydrolases.
Cel5A is the only endoglucanase from Trichoderma reesei that previously has been reported toshow β-mannanase activity (Macarrón et al., 1996). Interestingly, we found that Cel7B had activ-ity against O-acetyl-GGM. Two peaks were formed after SEC 1 with slightly lower molecularweight than before hydrolysis. However, further investigations have to be made regarding theGGM activity to establish if Cel7B has β-mannanase activity or if it has the ability to catalyze thecleavage of bonds between –1 and +1 subsite having mannosyl or glucosyl units placed at the -1subsite. Furthermore, it would be interesting to explore the possible effect of substituents onendoglucanase hydrolysis of O-acetyl-GGM.
145

References
Ademark, P., de Vries, R. P., Hägglund, P., Stålbrand, H. & Visser, J. (2001a). Cloning and char-acterization of Aspergillus niger genes encoding an α-galactosidase and a β-mannosidaseinvolved in galactomannan degradation. Eur. J. Biochem 268(10), 2982-2990.
Ademark, P., Larsson, M., Tjerneld, F. & Stålbrand, H. (2001b). Multiple α-galactosidase fromAspergillus niger: purification, characterization, and substrate specificities. EnzymeMicrob. Technol. 29, 441-448.
Ademark, P., Lundqvist, J., Hägglund, P., Tenkanen, M., Torto, N., Tjerneld, F. & Stålbrand, H.(1999). Hydrolytic properties of a β-mannosidase purified from Aspergillus niger. J.Biotechnol. 75(2-3), 281-289.
Ademark, P., Varga, A., Medve, J., Harjunpää, V., Drakenberg, T., Tjerneld, F. & Stålbrand, H.(1998). Softwood hemicellulose-degrading enzymes from Aspergillus niger: purificationand properties of a β-mannanase. J. Biotechnol. 63(3), 199-210.
Chhabra, S. R., Shockley, K. R., Ward, D. E. & Kelly, R. M. (2002). Regulation of endo-actingglycosyl hydrolases in the hyperthermophilic bacterium Thermotoga maritima grownon glucan- and mannan-based polysaccharides. Appl. Environ. Microbiol 68(2), 545-554.
Collén, A., Ward, M., Tjerneld, F. & Stålbrand, H. (2001). Genetic engineering of the Trichode-rma reesei endoglucanase I (Cel7B) for enhanced partitioning in aqueous two-phasesystems containing thermoseparating ethylene oxide—propylene oxide copolymers. J.Biotechnol 87(2), 179-191.
Frei, E. & Preston, R. D. (1968). Noncellulosic structural polysaccharides in algal cell walls. III.Mannan in siphoneous green algae. Proc. R. Soc. London, Ser. B 169(1015), 127-145.
Gübitz, G. M., Hayn, M., Sommerauer, M. & Steiner, W. (1996). Mannan-Degrading Enzymesfrom Sclerotium rolfsii: Characterisation and synergism of two Endo β-Mannanases anda β-Mannosidase. Bioresour. Technol. 58, 127-135.
Henrissat, B. (1991). A classification of glycosyl hydrolases based on amino acid sequence simi-larities. Biochem. J. 280(Pt 2), 309-316.
Hirata, K., Aso, Y. & Ishiguro, M. (1998). Purification and some properties of β-mannosidase, β-N-acetylglucosaminidase, and β-galactosidase from apple snails (Pomacea canaliculata).J. Fac. Agric., Kyushu Univ. 42(3-4), 463-472.
Hägglund, P., Eriksson, T., Collén, A., Nerinckx, W., Claeyssens, M. & Stålbrand, H. (2002). Acellulose-binding module of the Trichoderma reesei b-mannanase Man5A increases themannan-hydrolysis of complex substrates. Submitted.
Hägglund, P., Sabini, E., Boisset, C., Wilson, K., Chanzy, H. & Stålbrand, H. (2001). Degradationof mannan I and II crystals by fungal endo-β-1,4-mannanases and a β-1,4-mannosidasestudied with transmission electron microscopy. Biomacromolecules 2(3), 694-699.
Ikiri, Y. & Miwa, T. (1960). Chemical Nature of the cell wall of the green algae, Codium,Acetabularia and Halicoryne. Nature 185, 178-179.
Jenkins, J., Lo Leggio, L., Harris, G. & Pickersgill, R. (1995). β-glucosidase, β-galactosidase,family A cellulases, family F xylanases and two barley glycanases form a superfamily ofenzymes with 8-fold β/α architecture and with two conserved glutamates near the carboxy-terminal ends of β-strands four and seven. FEBS Lett 362(3), 281-285.
Karlsson, J. (2000). Fungal Cellulases: Study of hydrolytic properties of endoglucanases fromTrichoderma reesei and Humicola insolens. Doctoral thesis, Lund University.
146

Kulminskaya, A. A., Eneiskaya, E. V., Isaeva-Ivanova, L. S., Savel’ev, A. N., Sidorenko, I. A.,Shabalin, K., A., Golubev, A. M. & Neustroev, K. N. (1999). Enzymatic activity and β-galactomannan binding property of β-mannosidase from Trichoderma reesei. EnzymeMicrob.Technol. 25, 372-377.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227(259), 680-685.
Lindberg, B., Rosell, K.-G. & Svensson, S. (1973). Position of the O-acetyl groups in pineglucomannan. Svensk Papperstidning 76, 383-384.
Lundqvist, J., Jacobs, A., Palm, M., Zacchi, G., Dahlman, O. & Stålbrand, H. (2002a).Characterization of galactoglucomannan extracted from spruce (Picea abies) by heatfractionation at different conditions. Submitted.
Lundqvist, J., Teleman, A., Junel, L., Zaachi, G., Dahlman, O., Tjerneld, F. & Stålbrand, H. (2002b).Isolation and characterization of galactoglucomannan from Spruce (Picea abies).Carbohydr. Polym. 48(1), 29-39.
Macarrón, R., Acebal, C., Castillón, M. P. & Claeyssens, M. (1996). Mannanase activity ofEndoglucanase III from Trichoderma reesei QM9414. Biotechnol. Lett. 18(5), 599-602.
McCleary, B. V. & Matheson, N. K. (1983). Action Patterns and Substrate-Binding Requirementsof β-D-Mannanase with Mannosaccharides and Mannan-type Polysaccharides.Carbohydr. Res. 119, 191-219.
Meier, H. (1958). On the structure of cell walls and cell wall mannans from ivory nuts and fromdates. Biochim. Biophys. Acta 28, 229-240.
Meier, H. & Reid, J. S. G. (1982). Reserve Polysaccharides Other Than Starch in Higher Plants.In Encyclopaedia of Plant Physiology New Ser. (Loewus, F. A. & Tanner, W., eds.), Vol.13A, pp. 418-471. Springer Verlag, Berlin.
Nakari-Setäla, T. & Penttilä, M. (1995). Production of Trichoderma reesei cellulases on glucose-containing media. Applied and Environmental Microbiology 61(10), 3650-3655.
Nishinari, K., Williams, P. A. & Phillips, G. O. (1992). Review of the physico-chemical charac-teristics and properties of konjac mannan. Food Hydrocolloids 6(2), 199-222.
Penttilä, M., Lehtovaara, P., Nevalainen, H., Bhikhabhai, R. & Knowles, J. (1986). Homologybetween cellulase genes of Trichoderma reesei: complete nucleotide sequence of theendoglucanase I gene. Gene 45, 253-263.
Puls, J. & Schuseil, J. (1993). Chemistry of hemicelluloses: relationship between hemicellulosestructure and enzymes required for hydrolysis. In Hemicellulose and Hemicellulases(Coughlan, M. P. & Hazlewood, G. P., eds.), Vol. 4, pp. 1-27. Portland Press Res. Monogr.,London.
Reid, J. S. G. (1985). Galactomannans. In Biochemistry of Storage Carbohydrates in Green Plants(Dey, P. M. & Dixon, R. A., eds.), pp. 265-288. Academic Press, London.
Rättö, M. & Poutanen, K. (1988). Production of Mannan-Degrading Enzymes. Biotechnol. Lett.10(9), 661-664.
Rättö, M., Siika-aho, J., Buchert, J., Valkeajärvi, A. & Viikari, L. (1993). Enzymatic hydrolysis ofisolated and fibre-bound galactoglucomannans from pine-wood and pine kraft pulp.Applied Microbiology and biotechnology 40, 449-454.
Saloheimo, A., Henrissat, B., Hoffrén, A. M., Teleman, O. & Penttilä, M. (1994). A novel, smallendoglucanase gene, egl5, from Trichoderma reesei isolated by expression in yeast.Mol. Microbiol. 13(2), 219-28.
Siika-aho, M., Karlsson, J., Nakari-Setälä, T., Saloheimo, A., Teleman, A., Tjerneld, F. & Pentitilä,M. (2002). EG V (Cel45) of Trichoderma reesei expression of the core protein, purifica-tion and characterization. Submitted..
147

Stoll, D., Stålbrand, H. & Warren, R. A. (1999). Mannan-degrading enzymes from Cellulomonasfimi. Appl. Environ. Microbiol. 65(6), 2598-2605.
Stålbrand, H., Saloheimo, A., Vehmaanpera, J., Henrissat, B. & Penttilä, M. (1995). Cloning andexpression in Saccharomyces cerevisiae of a Trichoderma reesei β-mannanase genecontaining a cellulose binding domain. Appl. Environ. Microbiol. 61(3), 1090-1097.
Stålbrand, H., Siika-aho, M., Tenkanen, M. & Viikari, L. (1993). Purification and characterizationof two β-mannanases from Trichoderma reesei. J. Biotechnol. 29, 229-242.
Sunna, A., Gibbs, M. D., Chin, C. W., Nelson, P. J. & Bergquist, P. L. (2000). A gene encoding anovel multidomain β-1,4-mannanase from Caldibacillus cellulovorans and action of therecombinant enzyme on kraft pulp. Appl. Environ. Microbiol. 66(2), 664-670.
Tenkanen, M., Makkonen, M., Perttula, M., Viikari, L. & Teleman, A. (1997). Action ofTrichoderma reesei mannanase on galactoglucomannan in pine kraft pulp. J. Biotechnol.57(1-3), 191-204.
Tenkanen, M., Thornton, J. & Viikari, L. (1995). An acetylglucomannan esterase of Aspergillusoryzae; purification, characterization and role in the hydrolysis of O-acetyl-galactoglucomannan. J. Biotechnol. 42(3), 197-206.
Timell, T. E. (1967). Recent Progress in the chemistry of Wood Hemicellulose. Wood Sci. Technol.1, 45-70.
Wolfrom, M. L., Layer, M. L. & Patin, D. L. (1961). Carbohydrates of the coffee bean. II. Isola-tion and characterization of a mannan. J. Org. Chem. 26, 4533-4535.
Xu, B., Hägglund, P., Stålbrand, H. & Janson, J. C. (2002). endo-β-1,4-Mannanases from bluemussel, Mytilus edulis: purification, characterization, and mode of action. J. Biotechnol.92(3), 267-277.
Åkerholm, M. & Salmén, L. (2001). Interactions between wood polymers studied by dynamic FT-IR spectroscopy. Polymer 42(3), 963-969.
148
Enzymes Reducing sugars (mg/ml)
Man5A+Man2A 4.92
Man2A 1.41
Man2A+β-Gluc+Agal II 2.14
Man5A 4.30
Cel7B 2.11
Name Activity E.C number Mr * References
Man5Aa Endo-1,4-β-mannanase 3.2.1.78 45 000 (Hägglund et al., 2002;Stålbrand et al., 1995)
Man26Ab Endo-1,4-β-mannanase 3.2.1.78 48 000 (Stoll et al., 1999)
Man2A Exo-1,4-β-mannosidase 3.2.1.25 135 000 (Ademark et al., 2001a;Ademark et al., 1999)
Cel7Bc Endo-1,4-β-glucanase 3.2.1.4 46 000 (Collén et al., 2001;Penttilä et al., 1986)
Cel45Ad Endo-1,4-β-glucanase 3.2.1.4 22 000 (Saloheimo et al., 1994;Siika-aho et al., 2002)
Agal II Exo-α-1,6-galactosidase 3.2.1.22 64 000 (Ademark et al., 2001b)
β-Gluc Exo-1,4-β-glucosidase 3.2.1.21 118 000 Megazyme$
Table 1. Enzymes used in this study.
* Mr values from SDS-PAGE, i.e. monomer molecular weight
$ Commercial preparation purchased from Megazymea Catalytic module of Man5A, residues 1-352b Catalytic module of Man26, residues 1-464c Catalytic module of Cel7B, residues 1-371d Catalytic module of Cel45A, residues 1-182
Table 2. Released reducing sugars after enzymatic hydrolysis of O-acetyl-GGM (10 mg/ml).
149
Figure 1. A schematic picture of O-acetyl-galactoglucomannan, built up by a main chain of β-(1→4)-linkedmannose (MAN) and glucose (GLC) residues, with α-(1→6)-linked galactose (GAL) residues attached tosome MAN residues. Some MAN residues are acetylated at C-2 (2 Ac) or C-3 (3 Ac).
150
Figure 2. SEC2 elution profiles of the products formed after enzymatic hydrolysis of KM. (A) KM treatedwith Man5A (dotted line) and Man26A (hatched line) (B) KM treated with Cel45A (dotted line) and Cel7B(hatched line). In both A and B the nonhydrolysed KM (solid line) is also shown. RI was used for thedetection of the products formed. The applied sample volume was 500µl. The arrows mark the elutionvolumes of manno-oligosaccharides (DP 1-6). V
0 was estimated to 170 ml.
151
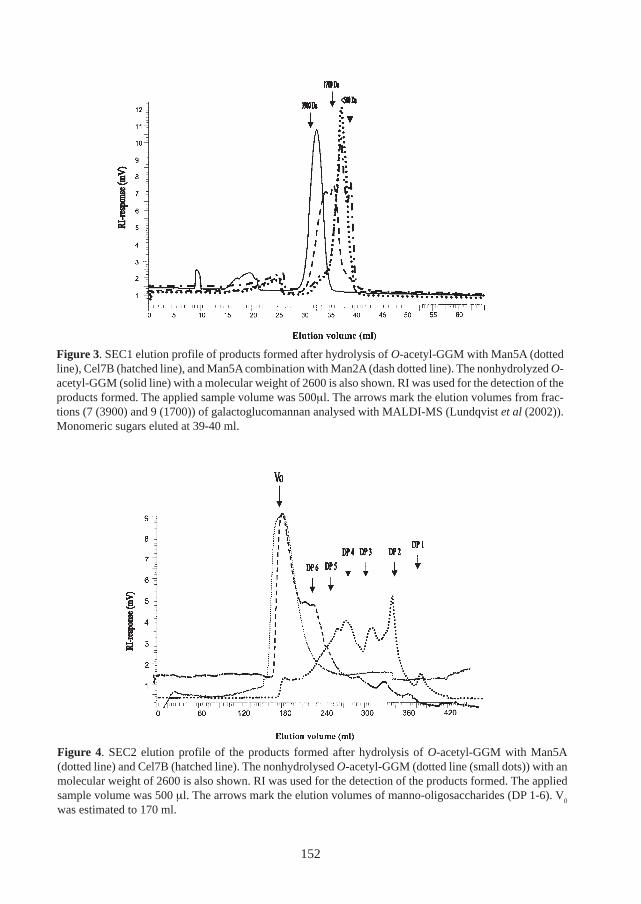
Figure 3. SEC1 elution profile of products formed after hydrolysis of O-acetyl-GGM with Man5A (dottedline), Cel7B (hatched line), and Man5A combination with Man2A (dash dotted line). The nonhydrolyzed O-acetyl-GGM (solid line) with a molecular weight of 2600 is also shown. RI was used for the detection of theproducts formed. The applied sample volume was 500µl. The arrows mark the elution volumes from frac-tions (7 (3900) and 9 (1700)) of galactoglucomannan analysed with MALDI-MS (Lundqvist et al (2002)).Monomeric sugars eluted at 39-40 ml.
Figure 4. SEC2 elution profile of the products formed after hydrolysis of O-acetyl-GGM with Man5A(dotted line) and Cel7B (hatched line). The nonhydrolysed O-acetyl-GGM (dotted line (small dots)) with anmolecular weight of 2600 is also shown. RI was used for the detection of the products formed. The appliedsample volume was 500 µl. The arrows mark the elution volumes of manno-oligosaccharides (DP 1-6). V
0
was estimated to 170 ml.
152
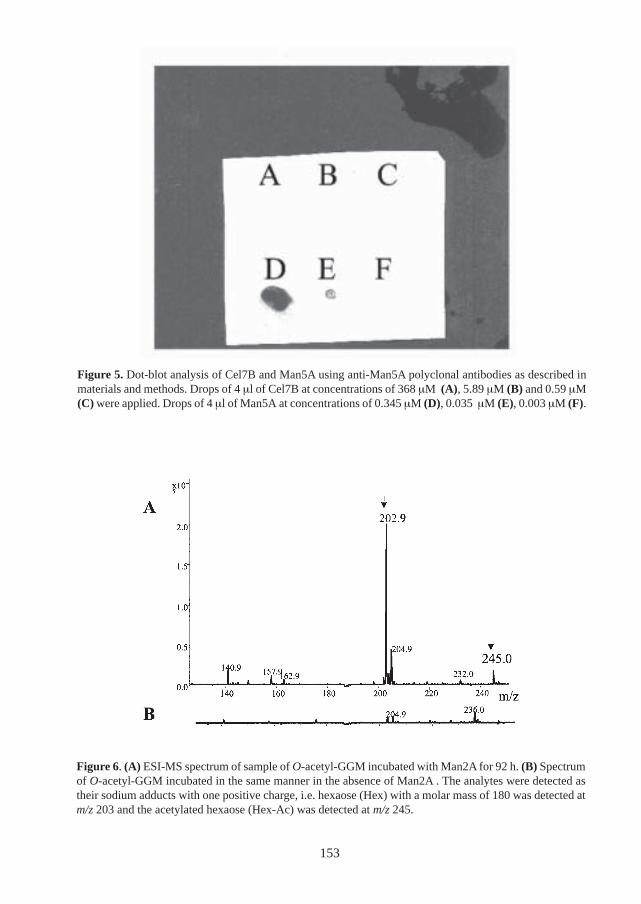
Figure 5. Dot-blot analysis of Cel7B and Man5A using anti-Man5A polyclonal antibodies as described inmaterials and methods. Drops of 4 µl of Cel7B at concentrations of 368 µM (A), 5.89 µM (B) and 0.59 µM(C) were applied. Drops of 4 µl of Man5A at concentrations of 0.345 µM (D), 0.035 µM (E), 0.003 µM (F).
Figure 6. (A) ESI-MS spectrum of sample of O-acetyl-GGM incubated with Man2A for 92 h. (B) Spectrumof O-acetyl-GGM incubated in the same manner in the absence of Man2A . The analytes were detected astheir sodium adducts with one positive charge, i.e. hexaose (Hex) with a molar mass of 180 was detected atm/z 203 and the acetylated hexaose (Hex-Ac) was detected at m/z 245.
153

154

VII
155

156

The Active Site Cleft of Trichoderma reeseiβ-Mannanase Man5A: Analysis of Mutantsof the Catalytic Glutamates and Arginine
171 Positioned in the +2 Subsite
Per Hägglund, Lars Anderson and Henrik Stålbrand
Department of Biochemistry, Center for Chemistry and Chemical Engineering,Lund University, PO Box 124, S-221 00, Lund, Sweden
Abstract
The β-mannanase (Man5A) from the filamentous fungus Trichoderma reesei is amodular enzyme composed of a catalytic module and a carbohydrate-bindingmodule. The N-terminal catalytic module is classified into family 5 of glycosidehydrolases and the previously solved structure of this module revealed that itshares the overall (β/α)
8 fold which is conserved in this family. Furthermore, the
two catalytic glutamates (Glu169 and Glu276) were also found in their conservedpositions. In addition, an arginine residue (Arg171), which is conserved in sev-eral other β-mannanases, is positioned in the +2 subsite. In order to study thefunction of Arg171, mutants of this residue were constructed and expressed inPichia pastoris. Mutants of the catalytic glutamates were also designed and ex-pressed in the same way. The hydrolytic properties of the mutants were studied.The results show that a mutant of Arg171 (R171K) display retained activity onpolymeric galactomannan but reduced activity on oligosaccharides. Moreover,this mutant appears to have a more alkaline activity pH-optimum than the wild-type enzyme. The mutants of the catalytic glutamates were essentially inactive,with the exception of one of the acid/base mutants (E169Q) for which a 25-folddecrease in k
cat was observed.
Abbreviations: Locust bean gum (LBG); 3,5-dinitrosalicylic acid (DNS)
157

Introduction
Polysaccharides with β-(1→4)-linked mannose residues are common in nature. They are foundin a variety of terrestrial plants and also in some algae (Frei & Preston, 1968; Meier, 1958; Timell,1967). O-acetyl-galactoglucomannan hemicellulose found in woody tissues from gymnospermsis one of the most abundant polysaccharides on earth. Galactomannans and glucomannans in theseeds and tubers of plants function mainly as storage polysaccharides (Meier & Reid, 1982).
Endo-1,4-β-D-mannanase (E.C. 3.2.1.78; β-mannanase) is the major depolymerising mannan-degrading enzyme: it catalyses the random hydrolysis of β-D-1,4-mannopyranosyl linkages withinthe main chain of different mannans. A majority of the characterised β-mannanases are producedby bacteria and fungi (Ademark et al., 1998; Stoll et al., 1999; Stålbrand et al., 1995), but β-mannanases have also been isolated from plants, algae and mollusks (Bewley et al., 1997; Loos& Meindl, 1985; Xu et al., 2002). The β-mannanases characterised this far, have been annotatedto family 5 and 26 of glycoside hydrolases in the classification according to Henrissat et al.(Henrissat, 1991; Henrissat & Bairoch, 1996). Enzymes from both of these families share the (β/α)
8 barrel fold and the structurally conserved catalytic residues found in clan GH-A of glycosyl
hydrolases (Henrissat et al., 1995; Jenkins et al., 1995).
The filamentous fungus Trichoderma reesei produces a β-mannanase (Man5A) (Stålbrand et al.,1995; Stålbrand et al., 1993). The corresponding gene has been cloned and the N-terminal catalyticmodule is classified in family 5 of glycoside hydrolases (Stålbrand et al., 1995). The C-terminalmodule belong to family I of carbohydrate-binding modules (CBMs) according to the classificationof Tomme et al. (Tomme et al., 1995). The CBM binds to cellulose and has been shown to affectthe hydrolysis of complex substrates by Man5A (Hägglund et al., 2002). The structure of thecatalytic module of Man5A (Man5A∆CBM) has been solved by X-ray crystallography (Sabini etal., 2000). Man5A∆CBM share the same overall fold conserved in other family 5 enzymes.
At least seven conserved residues in family 5 are also conserved in the Man5A structure.Included among these are the two residues predicted to be the catalytic residues: the acid/base(Glu169) and the nucleophile (Glu276). The influence of several of the residues conserved infamily 5 on hydrolysis has been investigated earlier (Belaich et al., 1992; Navas & Beguin, 1992;Py et al., 1991). In particular, the importance of the catalytic residues has been experimentallydemonstrated previously for several family 5 endo- and exoglucanases using mutational analysisand covalently trapped inhibitors (Macarron et al., 1993; Mackenzie et al., 1997; Wang et al.,1993). Interestingly, despite the conserved amino acids and similar structural folds in family 5,several specificities are included in this family. Beside β-mannanases, family 5 also include forexample endo- and exoglucanases.
The structure of Man5A∆CBM in complex with mannobiose reveals some interesting features(Sabini et al., 2000). In this complex, the mannobiose molecule is bound in the +1 and +2 subsitesof Man5A through several interactions with the enzyme. Specifically, it was indicated that Arg171 is important for substrate binding since it is positioned within hydrogen bond distance fromthe axial C2 hydroxyl of the +2 mannose residue (Figure 1). Since mannose and glucose onlydiffer in the position of the C2 hydroxyl group it is possible that the interaction with this group isimportant for determining the enzymes substrate specificity between mannan and cellulose.
158

Interestingly, sequence alignment of family 5 β-mannanases reveals that this arginine residueappears to be conserved in several eukaryotic β-mannanases (Figure 2).
Previous kinetic analyses of manno-oligosaccharide degradation with Man5A suggest that the +2subsite may be important for hydrolysis, at least in the case of substrates with a degree ofpolymerization of 4 or more (Harjunpää et al., 1995). Furthermore, product analysis suggests thatthe +2 subsite also may participate in aglycone binding in transglycosylation reactions (Harjunpääet al., 1999).
In order to investigate the specificity in the active site of Man5A, we have constructed mutants ofthe catalytic acid/base (Glu169) and nucleophile (Glu276). We have also mutated Arg171 positionedin +2 subsite. In accordance with the prediction, we found a pronounced reduction in activity forthe mutants of the catalytic residues. However, one of the Arg171 mutants (R171K) showedretained activity on polymeric, soluble galactomannan, but reduced activity on mannopentaose.Furthermore, our results indicate a shift toward a more alkaline activity pH optimum for R171K,in comparison with the wild-type enzyme.
Materials and Methods
Plasmids and strainsThe plasmid pHSM30 (Stålbrand et al., 1995) was maintained in Escherichia coli XL1 blue(Stratagene, La Jolla, CA, USA). All other plasmids were maintained in E. coli TOP10F’(Invitrogen, San Diego, CA, USA). All E. coli cells were cultured in luria broth medium at 37 °Cand Ampicillin (100 µg/ml) or Zeocin (25 µg/ml) was added when appropriate. The Pichiapastoris strain KM71H was purchased from Invitrogen and maintained on YPD plates (1 % YeastExtract, 2 % Peptone, 2 % Dextrose) according to the manufacturers recommendations.
Construction of mutantsMutagenesis was performed by PCR using Pfu polymerase with the Quikchange site-directedmutagenesis kit (Stratagene). Molecular biology techniques, including DNA isolation and prepa-ration, agarose gel electrophoresis, restriction enzyme analysis, and DNA fragment isolation wereperformed as described previously (Sambrook et al., 1989). The sequence encoding Man5A wasdigested from pHSM30 using XhoI and EcoRI and ligated into pPICZA which has been cut withthe same enzymes. The resulting plasmid, pPICZAMan5A, was used as a template for site-di-rected mutagenesis using Primer 1 and Primer 2 (Table 1), yielding a plasmid(pPICZAMan5A∆CBM) containing a truncated version of man5A, coding for the catalytic mod-ule only (man5A∆CBM; bases 1-1113 of man5A)). This plasmid was used as a template for allthe following site directed mutagenesis using the primers 3-16 (Table 1). All PCR runs wereinitiated with a denaturation step at 95 °C for two minutes, followed by 17 cycles of denaturation(95 °C, 30 seconds), annealing (62°C, 1 minute) and extension (72°C, 10 minutes). The PCRproducts were treated with Dpn I, to remove template plasmid and transformed into Escherichiacoli as described previously (Sambrook et al., 1989). The sequences of all mutants were analysedat the BM-unit, Lund University.
159

Pichia pastoris transformationThe constructed plasmids were purified using the JETstar Plasmid Midiprep (Genomed, ResearchTriangle Park, NC, USA), linearised using Sac I and subsequently purified by phenol extractionand ethanol precipitation as described previously (Sambrook et al., 1989). P. pastoris cells, wereelectroporated with 5-10 µg of linearised plasmid according to the supplier’s recommendations(Invitrogen). P. pastoris transformants were transferred to YPD plates containing sorbitol (1 M)and Zeocin (100 µg/ml). The plates were incubated for 3 days at 30°C. Positive transformantswere subsequently replated several times. Isolated transformants were screened for β-mannananseexpression using the β-mannanase activity plate assay and dot-blot analysis described below.
Expression in Pichia pastorisPositive transformants were inoculated in YPD media (50 ml in 1 l indented flasks) and culturedfor 48 h (30 ºC, 200 rpm). After harvesting by centrifugation (5000 rpm; 10 min) the cells wereresuspended in buffered minimal media (1.34 % yeast nitrogen base, 4x10-5 % biotin, 0.5 %methanol, 100 mM potassium phosphate buffer, pH 6) and cultured in a volume of 200 ml in a 2l indented flask (30 ºC, 250 rpm) for 4 days. Methanol was added every day to a final concentra-tion of 0.5 % to compensate for loss because of evaporation and consumption. After 4 days, cellswere pelleted by centrifugation and the supernatants were collected and stored for purification.
Enzyme purificationThe supernatants were concentrated using a ultrafiltration device with a membrane of 10 kDacut-off. The. The concentrated samples were equilibrated in 10 mM Tris pH:7.8 using a PD-10column (Amersham Pharmacia Biotech, Uppsala, Sweden). Chromatography purification wasperformed on a FPLC® System (Amersham Pharmacia Biotech). Equilibrated samples were ap-plied to a 6 ml Resource Q anion exchange column (Amersham Pharmacia Biotech). The proteinswere eluted in a NaCl gradient (0-0.5 M) over 20 column volumes and fractions of 2 ml werecollected. The flow was 1.0 ml/min. The proteins were further purified on a HiLoad® 16/60 gelfiltration column (Pharmacia). The eluent was 200 mM NaCl in 100 mM Na-Acetate buffer, pH4.5 and the flow rate was 0.5 ml/min. Fractions of 2 ml were collected. All purified enzymesappeared as single bands on SDS-PAGE after silver staining (Laemmli, 1970). The concentra-tions of purified proteins were determined by measuring absorbance at 280 nm, according to themethod of Mach (Mach et al., 1992).
Dot-blot analysisAll steps were performed at room temperature. Drops of 10 µl were placed on a Biotrace NTblotting nitrocellulose membrane (Pall Gelman Sciences, Ann Arbor, MI, USA) and allowed todry for 15 min. The membrane was placed in blocking solution (TBS buffer (20 mM Tris, 137mM NaCl, pH7.6), 0.1 % tween and 1.5 % bovine serum albumin (BSA)) for one hour withgentle shaking. The membrane was washed twice for 5 min with changes of fresh washing buffer(TBS, 0.1 % tween). New TBS buffer including polyclonal antibodies (diluted 1/1000) againstMan5A∆CBM raised in rabbit, was added and the mebrane was incubated for 1 h. After twowashes as described above, new TBS buffer containing secondary goat anti-rabbit antibodies(Bio-Rad, Hercules, CA, USA) diluted 1/1500 was added, and the membrane was incubated for 1h. After two washes in Tris buffer (0.1 M Tris, 0.5 mM MgCl
2, pH9.5), new Tris buffer containing
Nitroblue tetrazolium chloride (300 µg/ml) and AP Color Development Reagent (150 µg/ml)(Bio-Rad) solutions was added, and the mixture was incubated until spots were clearly visible.The incubation was ended, by immersing the membrane in distilled water for 10 min.
160

β-mannanase activity plate assayTransformants were screened for β-mannanase activity by a β-mannanase plate assay, in analogyto a previously described method (Stålbrand, 1995). The P. pastoris cells were grown on bufferedminimal media plates (1.34 % yeast nitrogen base, 4x10-5 % biotin, 0.5 % methanol, 100 mMpotassium phosphate buffer, pH 6). These plates also contained 0.1 % locust bean gumgalactomannan (LBG) (Sigma) as substrate. Transformants were also grown on plates containingglycerol instead of methanol as a non-induced control. The plates were incubated in 30 ºC for 2days. 100 µl methanol was added to the lid of the inverted plate every day to compensate for lossbecause of evaporation and consumption. After two days, the cells were removed from the platedby several washes in distilled water. A 10 µl drop of previously purified β-mannanase solutionwas placed on the plate as a positive control and the plates were incubated in 42ºC for 30 min.After this incubation, the plates were coloured with Congo Red (Sigma) overnight and destainedwith 1 M NaCl for 3 hours.
β-Mannanase activity assayScreening of β-mannanase activity in purified fractions was performed using the previously de-scribed DNS-stopping method with 0.5% LBG as substrate, in 50 mM Na-acetate buffer pH 4.5(Stålbrand et al., 1993). The reaction was stopped by adding 3,5-dinitrosalicylic acid (DNS). Thereducing ends of the sugars were detected by boiling for 10 min. The absorbance is measured at540 nm and the activity calculated from a mannose standard curve.
pH optimum of LBG hydrolysisMan5A∆CBM and R171K (28 nM) were incubated in 0.5 % LBG solutions at 40 °C, in 50 mMbuffers at different pH (Na-Citrate pH 3.5; Na-Acetate pH 4.5; Na-Citrate pH 5.3; Na-Citrate pH6.0). Prior to the LBG incubations the enzyme stock solutions were diluted in the buffer of appro-priate pH and pre-incubated for 30 min. The release of reducing sugars after the LBG incubationswas determined as in the β-mannanase activity assay described above using DNS.
Mannopentaose hydrolysisMan5A∆CBM and R171K (0.45 nM) were incubated with mannopentaose at 40 °C in 50 mMNa-Acetate pH 4.5. BSA (100 µg/ml) was included. Oligosaccharides released after differenttime points in the mannopentaose hydrolysis were analysed using high performance anionic ex-change chromatography with pulsed amperometric detection (HPAEC-PAD) with a Dionex CX500 System (Dionex, Sunnyvale CA, USA). The system included an AS50 autosampler, a GP40gradient pump and an ED40 electrochemical detector. A Carbo Pac Pa100 guard and analyticalcolumn was used for separation and the oligosaccharides were eluted with an isocratic eluent of100 mM NaOH. The flow rate was 1 ml/min and the injection volume was 10 µl.Mannooligosaccharides of degree of polymerisation of 1-6 (Megazyme, Bray, Ireland) were usedas standards. Mannobiose and mannotriose were the main products released.
Determination of kinetic parametersKinetic parameters for LBG hydrolysis were determined by incubating the enzymes in LBGsolutions of different concentrations (ranging from 0.03 to 0.5 %). The hydrolysis velocity wasplotted as a function of substrate concentration and values of K
M and k
cat were obtained from non-
linear regression analysis using Kaleidagraph 3.51 (Synergy Software).
161

Results and discussion
Expression of Man5A in Pichia pastorisThe catalytic module of the Trichoderma reesei β-mannanase (Man5A∆CBM; amino acid 1-346) (Stålbrand et al., 1995), and a number of constructed mutants, were (listed in Table 1),where expressed in the methylotrophic yeast, P. pastoris under control of the alcohol oxidasepromoter. The level of expression of Man5A∆CBM in P. pastoris was approximately 40 nkat/ml,estimated from activity measurements. This level is slightly higher than the expression levels of30 nkat/ml obtained from T. reesei cultures when Man5A∆CBM was expressed under the controlof the Aspergillus nidulans gpdA promoter (Hägglund et al., 2002). Kinetic analysis with LBG asthe substrate showed that the value of k
cat of the purified Man5A∆CBM (241 s-1) expressed in P.
pastoris was similar to T. reesei expressed Man5A∆CBM (285 s-1) (Table 2). Furthermore, thek
cat/K
M ratios were similar (Table 2).
Activity of mutants of the catalytic residuesThe catalytic activities of a number of mutants of the catalytic residues (Glu169 and Glu276), ofMan5A∆CBM were analysed. The activity of the mutants was assessed either by the β-mannanaseplate assay or by the β-mannanase activity assay described in materials and methods. In accordancewith the prediction of the catalytic residues, the activities of the mutants were greatly reduced orabolished. With the nucleophile mutants (E276Q, E276A and E276S), no activity was detected(Table 2). In addition, no activity was detected with the alanine mutant of the acid/base mutants(E169A). However, activity was detected for the E169Q mutant, but it was apparently largelyreduced (Table 2). Kinetic analysis of the hydrolysis of LBG by E169Q revealed that the reducedactivity was mainly due to an approximately 25-fold decrease in k
cat (Table 2). K
M was essentially
unchanged. Previous mutational analyses of other family 5 enzymes have also shown a dramatic
decrease in activity of mutants of the catalytic residues (Navas & Beguin, 1992; Py et al., 1991).As in the present case, a greater activity reduction for an acid/base alanine mutant in comparisonwith a glutamine mutant was also seen with the Clostridium thermocellum endoglucanase CelC(Navas & Beguin, 1992). Moreover, as in the present case, the K
m of the glutamine mutant was
similar to the wild-type.
Comparison of the R171K mutant and the wild-type enzymeTwo mutants of Arg171 were created (R171K and R171A) and transformed into P. pastoris. Withthe R171A mutant, P. pastoris transformants were obtained, but no β-mannanase activity was detected(Table 2). Furthermore, a negative dot-blot analysis using anti-Man5A∆CBM antibodies, indicatedabolished expression and/or secretion. In contrast, the R171K mutant showed activity on the plateassay, and was thus subjected to further analysis. Kinetic analysis showed that the activity of thismutant on LBG was essentially retained, in comparison with the wild-type (Table 2).
Since it was found that the R171K mutant was active, the hydrolytic properties of this mutant onLBG were further studied. Thus, the activity pH optimum profile of this mutant was investigatedand compared with the wild-type. As seen in the activity pH optima plots shown in Figure 3, amore alkaline pH optimum was obtained with the R171K mutant in comparison with the wild-type enzyme. This is a very interesting observation in terms of applications of fungal β-mannanases.Many applications in the pulp and paper industry demand enzymes with more alkaline pH optimathan most of the fungal wild-type enzymes have (Buchert et al., 1998).
162

In the current case, the increase in activity pH optimum is difficult to explain solely by enzyme/substrate interactions in the +2 subsite. Several previous reports have shown that charged/polaramino acid residues in the close vicinity of the catalytic residues can influence the enzymes pHoptima (Becker et al., 2001). Besides mannobiose, Arg171 is hydrogen bonded to Glu205 whichlies closer to the catalytic residues (Figure 1) (Sabini et al., 2000).
In order to evaluate the activity of the R171K mutant also on oligomeric substrates, mannopentaosewas hydrolysed by the wild-type and R171K and samples withdrawn at different time-pointswere analysed. In contrast to the hydrolysis of polymeric substrate LBG, a significant decrease inactivity was observed for the R171K mutant in mannopentaose hydrolysis in comparison with thewild-type enzyme (Figure 4). It may be possible that this decrease in activity results from areduced substrate affinity in the +2 subsite, possibly as a result of the lost hydrogen bond betweenArg171 and the C-2 hydroxyl of mannose. Previous kinetic analyses of mannopentaose hydrolysiswith Man5A indicate an involvement of the +2 subsite (Harjunpää et al., 1995). Continued kineticanalysis of mannopentaose degradation by R171K will further establish the role of R171K insubstrate binding and its influence on hydrolysis.
AcknowledgementMagnus Nilsson is thanked for performing part of the mutagenesis. The Swedish Research Coun-cil (VR), the Swedish Research Council for Engineering Sciences (TFR) and Carl Trygger Foun-dation (CTS) are thanked for research grants to Henrik Stålbrand.
References
Ademark, P., Varga, A., Medve, J., Harjunpää, V., Drakenberg, T., Tjerneld, F. & Stålbrand, H.(1998). Softwood hemicellulose-degrading enzymes from Aspergillus niger: purificationand properties of a β-mannanase. J. Biotechnol. 63(3), 199-210.
Becker, D., Braet, C., Brumer III, H., Claeyssens, M., Divne, C., Fagerstrom, B. R., Harris, M.,Jones, T. A., Kleywegt, G. J., Koivula, A., Mahdi, S., Piens, K., Sinnott, M. L., Ståhlberg, J.,Teeri, T. T., Underwood, M. & Wohlfahrt, G. (2001). Engineering of a glycosidase Family7 cellobiohydrolase to more alkaline pH optimum: the pH behaviour of Trichoderma reeseiCel7A and its E223S/ A224H/L225V/T226A/D262G mutant. Biochem. J 356(Pt 1), 19-30.
Belaich, A., Fierobe, H. P., Baty, D., Busetta, B., Bagnara-Tardif, C., Gaudin, C. & Belaich, J. P.(1992). The catalytic domain of endoglucanase A from Clostridium cellulolyticum: effectsof arginine 79 and histidine 122 mutations on catalysis. J Bacteriol 174(14), 4677-82.
Bewley, J. D., Burton, R. A., Morohashi, Y. & Fincher, G. B. (1997). Molecular cloning of acDNA encoding a (1->4)-β-mannan endohydrolase from the seeds of germinated tomato(Lycopersicon esculentum). Planta 203(4), 454-459.
Buchert, J., Oksanen, T., Pere, J., Siika-Aho, M., Suurnakki, A. & Viikari, L. (1998). Applicationsof Trichoderma reesei enzymes in the pulp and paper industry. In Trichoderma Gliocladium(Harman, G. E. & Kubicek, C. P., eds.), Vol. 2, pp. 343-363. Taylor and Francis Ltd., Lon-don.
Frei, E. & Preston, R. D. (1968). Noncellulosic structural polysaccharides in algal cell walls. III.Mannan in siphoneous green algae. Proc. R. Soc. London, Ser. B 169(1015), 127-145.
163

Harjunpää, V., Helin, J., Koivula, A., Siika-aho, M. & Drakenberg, T. (1999). A comparativestudy of two retaining enzymes of Trichoderma reesei: transglycosylation of oligosaccharidescatalysed by the cellobiohydrolase I, Cel7A, and the β-mannanase, Man5A. FEBS Lett443(2), 149-153.
Harjunpää, V., Teleman, A., Siika-aho, M. & Drakenberg, T. (1995). Kinetic and stereochemicalstudies of manno-oligosaccharide hydrolysis catalysed by β-mannanases from Trichodermareesei. Eur. J. Biochem. 234, 278-283.
Henrissat, B. (1991). A classification of glycosyl hydrolases based on amino acid sequencesimilarities. Biochem. J. 280(Pt 2), 309-316.
Henrissat, B. & Bairoch, A. (1996). Updating the sequence-based classification of glycosylhydrolases. Biochem. J 316(Pt 2), 695-696.
Henrissat, B., Callebaut, I., Fabrega, S., Lehn, P., Mornon, J. P. & Davies, G. (1995). Conservedcatalytic machinery and the prediction of a common fold for several families of glycosylhydrolases. Proc. Natl. Acad. Sci. U. S. A 92, 7090-7094.
Hägglund, P., Eriksson, T., Collén, A., Nerinckx, W., Claeyssens, M. & Stålbrand, H. (2002). Acellulose-binding module of the Trichoderma reesei b-mannanase Man5A increases themannan-hydrolysis of complex substrates. Submitted.
Jenkins, J., Lo Leggio, L., Harris, G. & Pickersgill, R. (1995). β-glucosidase, β-galactosidase,family A cellulases, family F xylanases and two barley glycanases form a superfamily ofenzymes with 8-fold β/α architecture and with two conserved glutamates near the carboxy-terminal ends of β-strands four and seven. FEBS Lett 362(3), 281-285.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head ofbacteriophage T4. Nature 227(259), 680-685.
Loos, E. & Meindl, D. (1985). Cell-wall-bound lytic activity in Chlorella fusca: function andcharacterization of an endo-mannanase. Planta 166(4), 557-562.
Macarron, R., van Beeumen, J., Henrissat, B., de la Mata, I. & Claeyssens, M. (1993). Identificationof an essential glutamate residue in the active site of endoglucanase III from Trichodermareesei. FEBS Lett 316(2), 137-40.
Mach, H., Middaugh, C. R. & Lewis, R. V. (1992). Statistical determination of the average valuesof the extinction coefficients of tryptophan and tyrosine in native proteins. Anal. Biochem.200(1), 74-80.
Mackenzie, L. F., Brooke, G. S., Cutfield, J. F., Sullivan, P. A. & Withers, S. G. (1997). Identifi-cation of Glu-330 as the catalytic nucleophile of Candida albicans exo-β-(1,3)-glucanase.J. Biol. Chem 272(6), 3161-3167.
Meier, H. (1958). On the structure of cell walls and cell wall mannans from ivory nuts and fromdates. Biochim. Biophys. Acta 28, 229-240.
Meier, H. & Reid, J. S. G. (1982). Reserve Polysaccharides Other Than Starch in Higher Plants.In Encyclopaedia of Plant Physiology New Ser. (Loewus, F. A. & Tanner, W., eds.), Vol.13A, pp. 418-471. Springer Verlag, Berlin.
Navas, J. & Beguin, P. (1992). Site-directed mutagenesis of conserved residues of Clostridiumthermocellum endoglucanase CelC. Biochem. Biophys. Res. Commun. 189(2), 807-812.
Py, B., Bortoli-German, I., Haiech, J., Chippaux, M. & Barras, F. (1991). Cellulase EGZ of Erwiniachrysanthemi: structural organization and importance of His98 and Glu133 residues forcatalysis. Protein Eng 4(3), 325-333.
Sabini, E., Schubert, H., Murshudov, G., Wilson, K. S., Siika-Aho, M. & Penttilä, M. (2000). Thethree-dimensional structure of a Trichoderma reesei β-mannanase from glycoside hydro-lase family 5. Acta Crystallogr., Sect. D: Biol. Crystallogr. 56(Pt 1), 3-13.
164

Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). Molecular cloning: a laboratory manual. 2ndedit, Cold Spring Harbor Laboratory Press, Plainview, N.Y.
Stoll, D., Stålbrand, H. & Warren, R. A. (1999). Mannan-degrading enzymes from Cellulomonasfimi. Appl. Environ. Microbiol. 65(6), 2598-2605.
Stålbrand, H., Saloheimo, A., Vehmaanpera, J., Henrissat, B. & Penttilä, M. (1995). Cloning andexpression in Saccharomyces cerevisiae of a Trichoderma reesei β-mannanase genecontaining a cellulose binding domain. Appl. Environ. Microbiol. 61(3), 1090-1097.
Stålbrand, H., Siika-aho, M., Tenkanen, M. & Viikari, L. (1993). Purification and characterizationof two β-mannanases from Trichoderma reesei. J. Biotechnol. 29, 229-242.
Timell, T. E. (1967). Recent Progress in the chemistry of Wood Hemicellulose. Wood Sci. Technol.1, 45-70.
Tomme, P., Warren, R. A. J., Miller, R. C. J., Kilburn, D. G. & Gilkes, N. R. (1995). Cellulose-Binding Domains: Classification and Properties. In Enzymatic Degradation of InsolublePolysaccharides (Saddler, J. M. & Penner, M., eds.), pp. 142-163. American Chemical So-ciety, Washington, DC.
Wang, Q., Tull, D., Meinke, A., Gilkes, N. R., Warren, R. A., Aebersold, R. & Withers, S. G.(1993). Glu280 is the nucleophile in the active site of Clostridium thermocellum CelC, afamily A endo-β-1,4-glucanase. J. Biol. Chem 268(19), 14096-14102.
Xu, B., Hägglund, P., Stålbrand, H. & Janson, J. C. (2002). endo-β-1,4-Mannanases from blue mussel, Mytilus edulis: purification, characterization, and mode of action. J. Biotechnol. 92(3), 267-277.
165
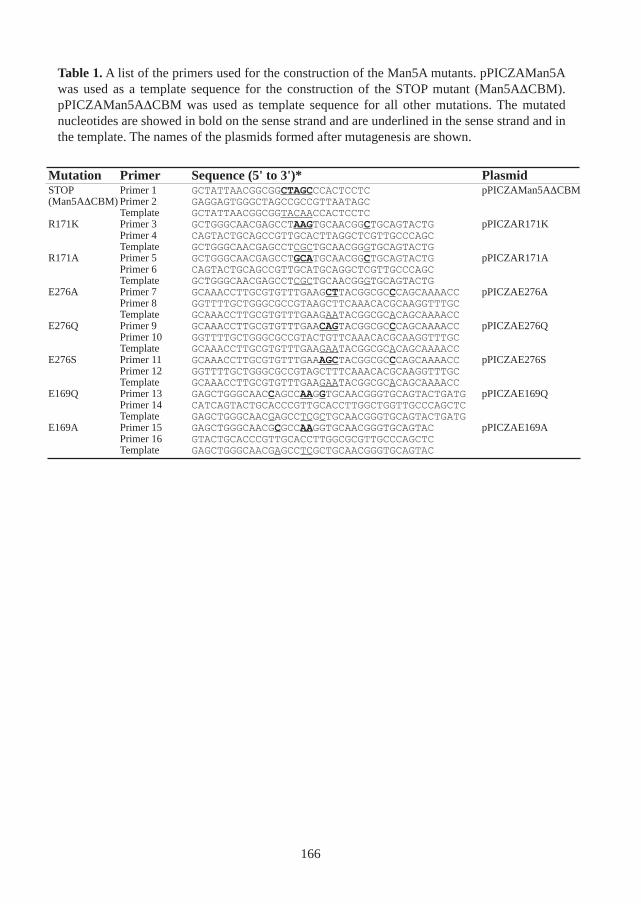
Table 1. A list of the primers used for the construction of the Man5A mutants. pPICZAMan5Awas used as a template sequence for the construction of the STOP mutant (Man5A∆CBM).pPICZAMan5A∆CBM was used as template sequence for all other mutations. The mutatednucleotides are showed in bold on the sense strand and are underlined in the sense strand and inthe template. The names of the plasmids formed after mutagenesis are shown.
166
Mutation Primer Sequence (5' to 3')* PlasmidSTOP Primer 1 GCTATTAACGGCGGCTAGCCCACTCCTC pPICZAMan5A∆CBM(Man5A∆CBM) Primer 2 GAGGAGTGGGCTAGCCGCCGTTAATAGC
Template GCTATTAACGGCGGTACAACCACTCCTCR171K Primer 3 GCTGGGCAACGAGCCTAAGTGCAACGGCTGCAGTACTG pPICZAR171K
Primer 4 CAGTACTGCAGCCGTTGCACTTAGGCTCGTTGCCCAGCTemplate GCTGGGCAACGAGCCTCGCTGCAACGGGTGCAGTACTG
R171A Primer 5 GCTGGGCAACGAGCCTGCATGCAACGGCTGCAGTACTG pPICZAR171APrimer 6 CAGTACTGCAGCCGTTGCATGCAGGCTCGTTGCCCAGCTemplate GCTGGGCAACGAGCCTCGCTGCAACGGGTGCAGTACTG
E276A Primer 7 GCAAACCTTGCGTGTTTGAAGCTTACGGCGCCCAGCAAAACC pPICZAE276APrimer 8 GGTTTTGCTGGGCGCCGTAAGCTTCAAACACGCAAGGTTTGCTemplate GCAAACCTTGCGTGTTTGAAGAATACGGCGCACAGCAAAACC
E276Q Primer 9 GCAAACCTTGCGTGTTTGAACAGTACGGCGCCCAGCAAAACC pPICZAE276QPrimer 10 GGTTTTGCTGGGCGCCGTACTGTTCAAACACGCAAGGTTTGCTemplate GCAAACCTTGCGTGTTTGAAGAATACGGCGCACAGCAAAACC
E276S Primer 11 GCAAACCTTGCGTGTTTGAAAGCTACGGCGCCCAGCAAAACC pPICZAE276SPrimer 12 GGTTTTGCTGGGCGCCGTAGCTTTCAAACACGCAAGGTTTGCTemplate GCAAACCTTGCGTGTTTGAAGAATACGGCGCACAGCAAAACC
E169Q Primer 13 GAGCTGGGCAACCAGCCAAGGTGCAACGGGTGCAGTACTGATG pPICZAE169QPrimer 14 CATCAGTACTGCACCCGTTGCACCTTGGCTGGTTGCCCAGCTCTemplate GAGCTGGGCAACGAGCCTCGCTGCAACGGGTGCAGTACTGATG
E169A Primer 15 GAGCTGGGCAACGCGCCAAGGTGCAACGGGTGCAGTAC pPICZAE169APrimer 16 GTACTGCACCCGTTGCACCTTGGCGCGTTGCCCAGCTCTemplate GAGCTGGGCAACGAGCCTCGCTGCAACGGGTGCAGTAC
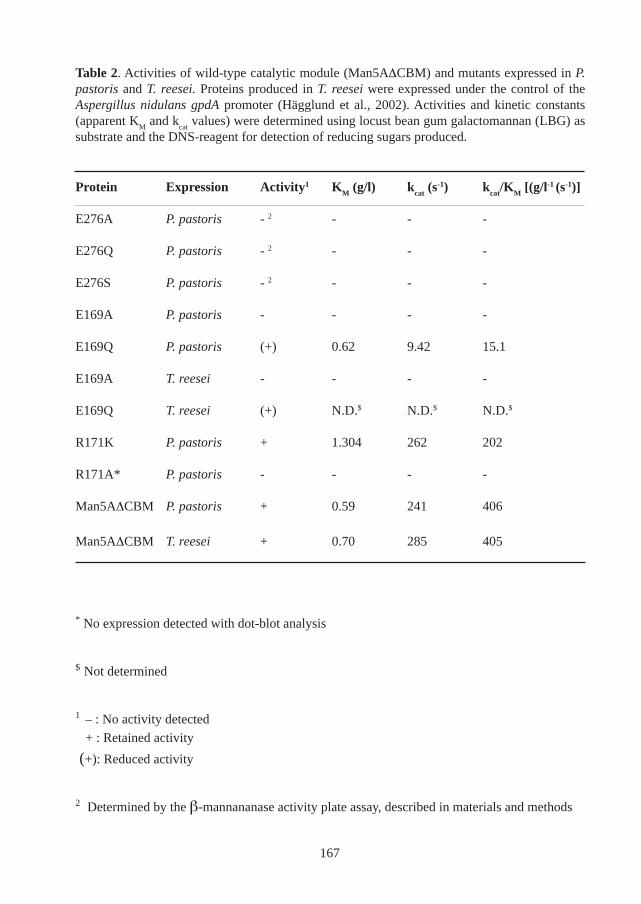
Table 2. Activities of wild-type catalytic module (Man5A∆CBM) and mutants expressed in P.pastoris and T. reesei. Proteins produced in T. reesei were expressed under the control of theAspergillus nidulans gpdA promoter (Hägglund et al., 2002). Activities and kinetic constants(apparent K
M and k
cat values) were determined using locust bean gum galactomannan (LBG) as
substrate and the DNS-reagent for detection of reducing sugars produced.
* No expression detected with dot-blot analysis
$ Not determined
1 – : No activity detected
+ : Retained activity
(+): Reduced activity
2 Determined by the β-mannananase activity plate assay, described in materials and methods
167
Protein Expression Activity1 KM (g/l) k
cat (s-1) k
cat/K
M [(g/l-1 (s-1)]
E276A P. pastoris - 2 - - -
E276Q P. pastoris - 2 - - -
E276S P. pastoris - 2 - - -
E169A P. pastoris - - - -
E169Q P. pastoris (+) 0.62 9.42 15.1
E169A T. reesei - - - -
E169Q T. reesei (+) N.D.$ N.D.$ N.D.$
R171K P. pastoris + 1.304 262 202
R171A* P. pastoris - - - -
Man5A∆CBM P. pastoris + 0.59 241 406
Man5A∆CBM T. reesei + 0.70 285 405

Figure 1. Close up of the active site of Man5A in complex with mannobiose, showing the residues selectedfor mutagenesis in this study. Possible hydrogen bonds are indicated by dotted lines. This figure was gener-ated in POV-Ray using data from the solved structure of Man5A∆CBM (PDB code: 1QNR) (Sabini et al.,2000)
Figure 2. Sequence alignment of family 5 β-mannanases, showing the regions around Arg171 (boxed) andGlu169 (boxed) in Man5A. The β-mannanases are from: Trichoderma reesei (Man5A), Swiss-Prot: Q99036;Aspergillus aculeatus, Swiss-Prot: Q00012; Agaricus bisporus, Swiss-Prot: Q92401; Geobacillusstearothermophilus, GenPept: AAC71692.1; Orpinomyces sp. PC-2, GenPept: AAL01213.1; Lycopersiconesculentum, Swiss-Prot: O48540; Mytilus edulis, GenPept: CAC81056.1. (*) = conserved residues, (:) =similar residue.
168
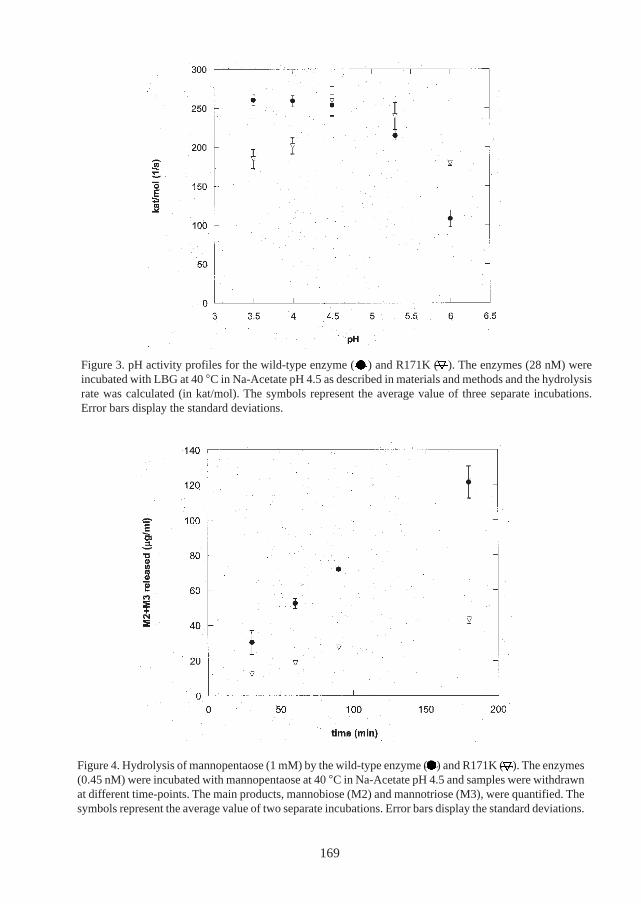
Figure 4. Hydrolysis of mannopentaose (1 mM) by the wild-type enzyme ( ) and R171K ( ). The enzymes(0.45 nM) were incubated with mannopentaose at 40 °C in Na-Acetate pH 4.5 and samples were withdrawnat different time-points. The main products, mannobiose (M2) and mannotriose (M3), were quantified. Thesymbols represent the average value of two separate incubations. Error bars display the standard deviations.
Figure 3. pH activity profiles for the wild-type enzyme ( ) and R171K ( ). The enzymes (28 nM) wereincubated with LBG at 40 °C in Na-Acetate pH 4.5 as described in materials and methods and the hydrolysisrate was calculated (in kat/mol). The symbols represent the average value of three separate incubations.Error bars display the standard deviations.
169









![a, arXiv:1906.06378v3 [cond-mat.mtrl-sci] 4 Aug 2019](https://static.fdocument.org/doc/165x107/61c0d52e1c1cea23c461e775/a-arxiv190606378v3-cond-matmtrl-sci-4-aug-2019.jpg)

![arXiv:1504.02216v4 [cond-mat.mtrl-sci] 14 Jul 2015](https://static.fdocument.org/doc/165x107/61bd302961276e740b1034c6/arxiv150402216v4-cond-matmtrl-sci-14-jul-2015.jpg)



![BIRKHOFF’S VARIETY THEOREM WITH AND WITHOUT FREE ALGEBRAS · 2005. 12. 18. · BIRKHOFF’S VARIETY THEOREM WITH AND WITHOUT FREE ALGEBRAS 429 (In [9] preconstant functors are called](https://static.fdocument.org/doc/165x107/60fce4a9a6fcce5a9f04a117/birkhoffas-variety-theorem-with-and-without-free-2005-12-18-birkhoffas.jpg)


