
RESPOSTAS FISIOLÓGICAS E AVALIAÇÃO DE ... Figura 8. Potencial da água na folha (Ψw) de quatro...
148
INSTITUTO AGRONÔMICO CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA TROPICAL E SUBTROPICAL RESPOSTAS FISIOLÓGICAS E AVALIAÇÃO DE PROGÊNIES DE IRMÃOS GERMANOS INTERPOPULACIONAIS DE MILHO COM ÊNFASE EM TOLERÂNCIA À SECA PAULA DE SOUZA GUIMARÃES Orientadora: Dra Maria Elisa Ayres Guidetti Zagatto Paterniani Co-orientador: Dr. Rafael Vasconcelos Ribeiro Tese submetida como requisito parcial para obtenção do grau de Doutora em Agricultura Tropical e Subtropical. Área de Concentração em Genética, Melhoramento Vegetal e Biotecnologia. Campinas, SP Junho, 2013
Transcript of RESPOSTAS FISIOLÓGICAS E AVALIAÇÃO DE ... Figura 8. Potencial da água na folha (Ψw) de quatro...

INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
RESPOSTAS FISIOLÓGICAS E AVALIAÇÃO DE
PROGÊNIES DE IRMÃOS GERMANOS
INTERPOPULACIONAIS DE MILHO COM ÊNFASE
EM TOLERÂNCIA À SECA
PAULA DE SOUZA GUIMARÃES
Orientadora: Dra Maria Elisa Ayres Guidetti Zagatto Paterniani
Co-orientador: Dr. Rafael Vasconcelos Ribeiro
Tese submetida como requisito parcial para
obtenção do grau de Doutora em Agricultura
Tropical e Subtropical. Área de Concentração em
Genética, Melhoramento Vegetal e Biotecnologia.
Campinas, SP
Junho, 2013

AGRADECIMENTOS
Agradeço a Deus pelas alegrias e momentos de experiências ao longo da minha caminhada;
Ao Instituto Agronômico de Campinas (IAC), onde tive o privilégio de absorver conhecimento
durante o mestrado e doutorado;
À Fundação de Amparo à Pesquisa (FAPESP), pela concessão da bolsa de estudo;
Aos meus amados pais Jorge Coelho Guimarães e Divina de Souza Guimarães pela vida de muitas
alegrias, amor incondicional, educação e carinho a mim depositados. Sem o apoio e incentivo
desses anjos não seria possível à realização deste sonho;
Aos meus “segundos pais” José Marlindo e Joana por me receberem de braços abertos e
amenizarem a saudade da minha casa, deixando-me fazer parte das suas vidas;
Aos meus queridos irmãos Marcia, Valquiria e Cassiano pela amizade, companheirismo e carinho.
Não tenho nem palavras para expressar o amor que sinto por vocês que sempre estiveram comigo
em todos os momentos;
Ao meu eterno namorado e companheiro de todos os momentos Marlindo de Souza Melo pela
paciência, apoio e amor;
As minhas cunhadas Izabel, Cristina, Eliane, Maria do Carmo e Ana Paula. Aos meus cunhados
Bruno, Ricardo e Magno;
Aos sobrinhos maravilhosos Cássio, Bianca, Maria Eduarda, Luis Felipe e Fernanda por me fazer a
tia coruja mais feliz desse mundo;
Aos meus amados amigos Allan Henrique pelos momentos de alegrias e conversas proveitosas,
Cristiani Bernini pela amizade e sempre disponibilidade em ajudar nas minhas avaliações de campo,
Lílian Galdino pelos momentos felizes, Renata Hanashiro pela ajuda nas avaliações, Sara Rovaris

pela amizade, ajuda nas avaliações de campo e por me fazer rir nos momentos mais inusitados e
Andrea Morais pelos momentos de descontração;
Ao meu querido “amigo irmão” João Guilherme Gonçalves pela excelente convivência e pelos
momentos de alegrias durante o doutorado;
À Profa. Dr
a. Maria Elisa Ayres Guidetti Zagatto Paterniani que me norteou desde o Mestrado, cujo
valor e contribuição em minha formação são incalculáveis. Professora você me mostrou que para
orientar, antes de tudo, é necessário ser amiga, ter muita paciência e confiança. Muito obrigada!!!
Ao Prof. Dr. Rafael Vasconcelos Ribeiro pela ajuda indispensável durante a excecução do projeto.
Muito obrigada pela paciência e apoio;
À Agência Paulista de Tecnologia dos Agronegócios – APTA, da cidade de Mococa - SP, pela
instalação e toda infra-estrutura para realização do experimento. Quero deixar meus sinceros
agradecimentos ao Pesquisador Paulo Gallo e aos funcionários Ronaldo e Sr. Geraldo;
Às alunas do Centro de Fisiologia do Instituto Agronômico Fernanda Villela, Daniela Favero e
Karina Silva e aos alunos Paulo Marquiori e André Luiz pela ajuda indispensável na realização das
avaliações fisiológicas;
Ao Centro de Fisiologia do IAC por disponibilizar os equipamentos e o laboratório;
Aos membros da Pré-banca Dr. Eduardo Caruso Machado, Dra. Tammy Kiihl e Dr. João Andrade
pela ajuda indispensável ao trabalho.

i
SUMÁRIO
ÍNDICE DE FIGURAS E TABELAS.............................................................. iii
LISTA DE ABREVIATURAS.......................................................................... x
RESUMO............................................................................................................ 1
ABSTRACT........................................................................................................ 3
1 INTRODUÇÃO.................................................................................................. 5
2 REVISÃO DE LITERATURA........................................................................ 7
2.1 Déficit hídrico...................................................................................................... 7
2.2 Déficit hídrico na cultura do milho...................................................................... 8
2.3 Parâmetros fisiológicos em milho submetido ao défict hídrico........................... 11
2.4 Caracteres primários e secundários relacionados à tolerância do déficit hídrico
em milho............................................................................................................... 14
2.5 Seleção recorrente................................................................................................ 18
2.6 Seleção recorrente recíproca com irmãos germanos de milho............................. 19
3 MATERIAL E MÉTODOS............................................................................... 21
3.1 Material vegetal e delineamento estatístico......................................................... 21
3.2 Condições de cultivo............................................................................................ 23
3.3 Avaliações fisiológicas em casa de vegetação..................................................... 24
3.4 Avaliações biométricas em casa de vegetação..................................................... 27
3.5 Análise dos dados................................................................................................. 28
3.6 Melhoramento para tolerância à seca................................................................... 28
3.6.1 Obtenção da população tolerante ao déficit hídrico............................................. 28
3.6.2 Obtenção da população sensível ao déficit hídrico.............................................. 29
3.6.3 Obtenção das progênies de irmãos germanos e S1 de milho................................ 29
3.6.4 Avaliação em campo: progênies de irmãos germanos em Campinas e Mococa.. 29
3.6.5 Análise agrupada e obtenção de parâmetros genéticos ....................................... 32
3.6.6 Avaliação e discriminação das melhores e piores progênies de irmãos
germanos quanto à tolerância à seca em casa de vegetação................................. 34
4 RESULTADOS E DISCUSSÃO....................................................................... 36
4.1 RESULTADOS: Parâmetros fisiológicos.......................................................... 36
4.1.1 Déficit hídrico no estádio vegetativo................................................................... 36
4.1.2 Déficit hídrico no estádio de florescimento......................................................... 43
4.1.3 Déficit hídrico no estádio de enchimento de grãos.............................................. 51
4.2 DISCUSSÃO: Parâmetros fisiológicos............................................................... 56
4.2.1 Déficit hídrico no estádio vegetativo................................................................... 56
4.2.2 Déficit hídrico no estádio de florescimento......................................................... 60
4.2.3 Déficit hídrico no estádio de enchimento de grãos.............................................. 64
4.3 Seleção recorrente interpopulacional: Análise de variância agrupada das
progênies de irmãos germanos. Campinas, 2012................................................. 65
4.4 Análise de variância agrupada das progênies de irmãos germanos. Mococa,
2012...................................................................................................................... 68
4.5 Análise de variância conjunta de locais e agrupada das progênies de irmãos
germanos, Campinas e Mococa, 2012.................................................................. 70
4.6 Avaliação e discriminação das melhores e piores progênies de irmãos
germanos quanto à tolerância à seca em casa de vegetação.................................
82
4.6.1 Déficit hídrico no estádio vegetativo................................................................... 82
4.6.2 Déficit hídrico no estádio de florescimento......................................................... 88

ii
5 CONCLUSÕES.................................................................................................. 93
6 REFERÊNCIAS BIBLIOGRÁFICAS............................................................. 95
7 ANEXOS............................................................................................................. 106

iii
ÍNDICE DE FIGURAS
Figura 1. Disposição do experimento realizado em casa de vegetação. Controle:
parcelas com irrigação durante todo o ciclo da cultura; vegetativo: parcelas
representando déficit hídrico somente no estádio vegetativo; florescimento:
parcelas representando déficit hídrico somente no estádio de florescimento e
enchimento de grãos: parcelas representando déficit hídrico somente no estádio de
enchimento de grãos. Foto tirada quando as plantas apresentavam-se no estádio V3.
Campinas, 2011............................................................................................................ 23
Figura 2. Gráfico de temperatura e de precipitação média da localidade de
Campinas no período de 01/03/2012 a 01/08/2012...................................................... 30
Figura 3. Gráfico de temperatura e de precipitação média da localidade de Mococa
no período de 23/03/2012 a 01/08/2012....................................................................... 31
Figura 4. Potencial da água na folha (Ψw) de quatro híbridos de milho avaliados
sob condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5).
Avaliações realizadas às 05h30 após 40 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas
indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos
na mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídricos..................................................................... 36
Figura 5. Condutância estomática (gs, em a) e concentração intercelular de CO2
(Ci, em b) de quatro híbridos de milho avaliados sob condição irrigada e sob déficit
hídrico a partir do estádio vegetativo (V5). Avaliações realizadas às 13h30 após 40
dias sob déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio
padrão. Letras minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras
maiúsculas distintas representam diferenças estatísticas entre os tratamentos
hídricos......................................................................................................................... 37
Figura 6. Transpiração diurna (Ei, em a), assimilação diurna de CO2 (Ai, em b) e
eficiência do uso da água (EUA, em c) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5).
Avaliações realizadas após 40 dias sob déficit hídrico. Cada histograma representa
o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam
diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na
mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídricos..................................................................... 39
Figura 7. Teores de açúcares solúveis (AS, em a), sacarose (SAC, em b), amido
(AM, em c) e carboidrato total não estrutural (CTNE, em d) de quatro híbridos de
milho avaliados sob condição irrigada e sob déficit hídrico a partir do estádio
vegetativo (V5). Folhas coletadas às 08h30 após 40 dias sob déficit hídrico. Cada
histograma representa o valor médio (n=3) ± desvio padrão. Letras minúsculas
distintas indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os
híbridos na mesma condição hídrica e letras maiúsculas distintas representam
diferenças estatísticas entre os tratamentos hídricos....................................................
41

iv
Figura 8. Potencial da água na folha (Ψw) de quatro híbridos de milho avaliados
sob condição irrigada e sob déficit hídrico a partir do estádio de florescimento (R1).
Avaliações realizadas às 05h30 após 75 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas
indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos
na mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídricos.....................................................................
44
Figura 9. Condutância estomática (gs, em a) e concentração intercelular de CO2
(Ci, em b) de quatro híbridos de milho avaliados sob condição irrigada e sob déficit
hídrico a partir do estádio de florescimento (R1). Avaliações realizadas às 13h30
após 75 dias sob déficit hídrico. Cada histograma representa o valor médio (n=3) ±
desvio padrão. Letras minúsculas distintas indicam diferenças estatísticas (P<0,05)
pelo teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras
maiúsculas distintas representam diferenças estatísticas entre os tratamentos
hídricos......................................................................................................................... 45
Figura 10. Transpiração diurna (Ei, em a), assimilação diurna de CO2 (Ai, em b) e
eficiência do uso da água (EUA, em c) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio de florescimento (R1).
Avaliações realizadas após 75 dias sob déficit hídrico. Cada histograma representa
o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam
diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na
mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídricos.....................................................................
46
Figura 11. Teores de açúcares solúveis (AS, em a), sacarose (SAC, em b), amido
(AM, em c) e carboidrato total não estrutural (CTNE) de quatro híbridos de milho
avaliados sob condição irrigada e sob déficit hídrico a partir do estádio de
florescimento (VT). Folhas coletadas às 08h30 após 75 dias sob déficit hídrico.
Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras
minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo teste de Scott-
Knott entre os híbridos na mesma condição hídrica e letras maiúsculas distintas
representam diferenças estatísticas entre os tratamentos hídricos............................... 49
Figura 12. Potencial da água na folha (Ψw) de quatro híbridos de milho avaliados
sob condição irrigada e sob déficit hídrico a partir do estádio de enchimento de
grãos (R3). Avaliações realizadas às 05h30 após 52 dias sob déficit hídrico. Cada
histograma representa o valor médio (n=3) ± desvio padrão. Letras minúsculas
distintas indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os
híbridos na mesma condição hídrica e letras maiúsculas distintas representam
diferenças estatísticas entre os tratamentos hídricos.................................................... 51
Figura 13. Condutância estomática (gs, em a) e concentração intercelular de CO2
(Ci, em b) de quatro híbridos de milho avaliados sob condição irrigada e sob déficit
hídrico a partir do estádio de enchimento de grãos (R3). Avaliações realizadas às
13h30 após 52 dias sob déficit hídrico. Cada histograma representa o valor médio
(n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças estatísticas
(P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição hídrica e
letras maiúsculas distintas representam diferenças estatísticas entre os tratamentos
hídricos.........................................................................................................................
52
Figura 14. Transpiração diurna (Ei, em a), assimilação diurna de CO2 (Ai, em b) e

v
eficiência do uso da água (EUA, em c) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio de enchimento de grãos
(R3). Avaliações realizadas após 52 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas
indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos
na mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídrico.......................................................................
53
Figura 15. Representação gráfica da redução relativa da altura da planta (AP),
diâmetro do colmo (DIAM), massa de grãos (MG), massa seca total (MST),
condutância estomática (gs), concentração intercelular de CO2 (Ci), assimilação
diurna de CO2 (Ai), transpiração diurna (Ei), teor de clorofila total (Chl), eficiência
quântica potencial do fotossistema II (Fv/Fm), coeficiente de dissipação não-
fotoquímica da fluorescência (NPQ), coeficiente de dissipação fotoquímica (qP),
taxa aparente de transporte de elétrons (ETR), açúcares solúveis (AS), sacarose
(SAC), amido (AM), carboidrato total não estrutural (CTNE) e eficiência do uso da
água (EUA) de quatro híbridos de milho submetidos a dois tratamentos hídricos a
partir do estádio vegetativo (V5). Valores maiores que 1 indicam aumento na
variável específica sob déficit hídrico..........................................................................
60
Figura 16. Representação gráfica da redução relativa da altura da planta (AP),
diâmetro do colmo (DIAM), massa de grãos (MG), massa seca total (MST),
condutância estomática (gs), concentração intercelular de CO2 (Ci), assimilação
diurna de CO2 (Ai), transpiração diurna (Ei), teor de clorofila total (Chl), eficiência
quântica potencial do fotossistema II (Fv/Fm), coeficiente de dissipação não-
fotoquímica da fluorescência (NPQ), coeficiente de dissipação fotoquímica (qP),
taxa aparente de transporte de elétrons (ETR), açúcares solúveis (AS), sacarose
(SAC), amido (AM), carboidrato total não estrutural (CTNE) e eficiência do uso da
água (EUA) de quatro híbridos de milho submetidos a dois tratamentos hídricos a
partir do estádio de florescimento (R1). Valores maiores que 1 indicam aumento na
variável específica sob déficit hídrico.......................................................................... 64
Figura 17. Potencial da água na folha (Ψw) de seis genótipos de milho avaliados
sob condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5).
Avaliações realizadas às 05h30 após 25 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas
indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos
na mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídricos..................................................................... 82
Figura 18. Condutância estomática (gs, em a) e assimilação de CO2 (A, em b) de
seis genótipos de milho avaliados sob condição irrigada e sob déficit hídrico a
partir do estádio vegetativo (V5). Avaliações realizadas às 13h30 após 25 dias sob
déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão.
Letras minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo teste de
Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas representam diferenças estatísticas entre os tratamentos hídricos.................
83
Figura 19. Representação gráfica da redução relativa da altura da planta (AP),
massa de grãos (MG), massa seca total (MST), condutância estomática (gs),
assimilação de CO2 (A), eficiência quântica potencial do fotossistema II (Fv/Fm),
coeficiente de dissipação não-fotoquímica da fluorescência (NPQ), coeficiente de
dissipação fotoquímica (qP) e taxa aparente de transporte de elétrons (ETR) de seis

vi
genótipos de milho submetidos a dois tratamentos hídricos a partir do estádio
vegetativo (V5). Valores maiores que 1 indicam aumento na variável específica sob
déficit hídrico...............................................................................................................
87
Figura 20. Potencial da água na folha (Ψw) de seis genótipos de milho avaliados
sob condição irrigada e sob déficit hídrico a partir do estádio de florescimento (R1).
Avaliações realizadas às 05h30 após 37 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas
indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos
na mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídricos..................................................................... 88
Figura 21. Condutância estomática (gs, em a) e assimilação de CO2 (A, em b) de
seis genótipos de milho avaliados sob condição irrigada e sob déficit hídrico a
partir do estádio de florescimento (R1). Avaliações realizadas às 13h30 após 37
dias sob déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio
padrão. Letras minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras
maiúsculas distintas representam diferenças estatísticas entre os tratamentos
hídricos.........................................................................................................................
89
Figura 22. Representação gráfica da redução relativa da altura da planta (AP),
massa de grãos (MG), massa seca total (MST), condutância estomática (gs),
assimilação de CO2 (A), eficiência quântica potencial do fotossistema II (Fv/Fm),
coeficiente de dissipação não-fotoquímica da fluorescência (NPQ), coeficiente de
dissipação fotoquímica (qP) e taxa aparente de transporte de elétrons (ETR) de seis
genótipos de milho submetidos a dois tratamentos hídricos a partir do estádio de
florescimento (R1). Valores maiores que 1 indicam aumento na variável específica
sob déficit hídrico......................................................................................................... 92
ÍNDICE DE TABELAS
Tabela 1. Híbridos de milho utilizados em experimento de casa de vegetação para
fins de avaliação de parâmetros fisiológicos e biométricos sob déficit hídrico
imposto em diferentes estádios fenológicos da cultura. Campinas,
2011.............................................................................................................................. 22
Tabela 2. Período de déficit hídrico, temperatura média dentro da casa de
vegetação e potencial matricial do solo do tratamento submetido ao déficit hídrico
imposto em três estádios fenológicos da cultura..........................................................
24
Tabela 3. Período de déficit hídrico, temperatura média dentro da casa de
vegetação e potencial matricial do solo do tratamento submetido ao déficit hídrico
imposto em dois estádios fenológicos da cultura......................................................... 35
Tabela 4. Teor de clorofila total (Chl), eficiência quântica potencial do
fotossistema II (Fv/Fm), coeficientes de extinção fotoquímica (qP) e não
fotoquímica (NPQ) da fluorescência e transporte aparente de elétrons (ETR) de
quatro híbridos de milho avaliados sob condição irrigada e sob déficit hídrico no

vii
estádio vegetativo (V5). Campinas, 2011.................................................................... 40
Tabela 5. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG)
e massa seca total (MST) de quatro híbridos de milho avaliados sob condição
irrigada e sob déficit hídrico no estádio vegetativo (V5). Campinas,
2011.............................................................................................................................. 43
Tabela 6. Teor de clorofila total (Chl), eficiência quântica potencial do
fotossistema II (Fv/Fm), coeficientes de extinção fotoquímica (qP) e não
fotoquímica (NPQ) da fluorescência e transporte aparente de elétrons (ETR) de
quatro híbridos de milho avaliados sob condição irrigada e sob déficit hídrico no
estádio de florescimento (VT). Campinas, 2011.......................................................... 47
Tabela 7. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG)
e massa seca total (MST) de quatro híbridos avaliados de milho sob condição
irrigada e sob déficit hídrico no estádio de florescimento (VT). Campinas,
2011.............................................................................................................................. 50
Tabela 8. Teor de clorofila total (Chl), eficiência quântica potencial do
fotossistema II (Fv/Fm), coeficientes de extinção fotoquímica (qP) e não
fotoquímica (NPQ) da fluorescência e transporte aparente de elétrons (ETR) de
quatro híbridos de milho avaliados sob condição irrigada e sob déficit hídrico no
estádio de enchimento de grãos (R3). Campinas, 2011............................................... 54
Tabela 9. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG)
e massa seca total (MST) de quatro híbridos de milho avaliados sob condição
irrigada e sob déficit hídrico no estádio de enchimento de grãos (R3). Campinas,
2011.............................................................................................................................. 55
Tabela 10. Quadrados médios da análise de variância agrupada para florescimento
masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF),
stay green (SG), prolificidade (PROL), número de ramificações de pendão (NRP) e
comprimento de pendão (CP), referentes a 150 progênies de irmãos germanos
interpopulacionais de milho. Campinas, 2012............................................................. 65
Tabela 11. Quadrados médios da análise de variância agrupada para altura da
planta (AP), altura da espiga (AE), comprimento da espiga (CE), diâmetro da
espiga (DE), número de fileiras de grãos na espiga (NFG) e massa de grãos (MG),
referentes a 150 progênies de irmãos germanos interpopulacionais de milho.
Campinas, 2012............................................................................................................
65
Tabela 12. Quadrados médios da análise de variância agrupada para florescimento
masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF),
stay green (SG), prolificidade (PROL), número de ramificações de pendão (NRP) e
comprimento de pendão (CP), referentes a 150 progênies de irmãos germanos
interpopulacionais de milho. Mococa, 2012................................................................
69
Tabela 13. Quadrados médios da análise de variância agrupada para altura da
planta (AP), altura da espiga (AE), comprimento da espiga (CE), diâmetro da
espiga (DE), número de fileiras de grãos na espiga (NFG) e massa de grãos (PG),
referentes a 150 progênies de irmãos germanos interpopulacionais de milho.
Mococa, 2012............................................................................................................... 69

viii
Tabela 14. Quadrados médios da análise de variância conjunta agrupada de
florescimento masculino (FM), florescimento feminino (FF), intervalo entre
florescimentos (IF), stay green (SG), prolificidade (PROL), número de ramificação
de pendão (NRP), comprimento de pendão (CP), referentes a 150 progênies de
irmãos germanos interpopulacionais de milho. Campinas e Mococa, 2012................ 71
Tabela 15. Quadrados médios da análise conjunta agrupada de variância de altura
da planta (AP), altura da espiga (AE), comprimento da espiga (CE), diâmetro da
espiga (DE), número de fileiras de grãos na espiga (NFG) e massa de grãos (MG),
referentes a 150 progênies de irmãos germanos interpopulacionais de milho.
Campinas e Mococa, 2012............................................................................................ 71
Tabela 16. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), stay green (SG), altura da planta (AP), altura da
espiga (AE), massa de grãos (MG) e porcentagem de massa de grãos (MG%),
referentes às 30 progênies de irmãos germanos interpopulacionais selecionadas e de
quatro testemunhas comerciais de milho. Campinas e Mococa, 2012......................... 73
Tabela 17. Médias de massa de grãos (MG) em kg ha-1
, comprimento de espiga
(CE), diâmetro da espiga (DE), número de fileiras de grãos na espiga (NFG),
prolificidade (PROL), número de ramificação de pendão (NRP) e comprimento de
pendão (CP), referentes às 30 progênies de irmãos germanos interpopulacionais
selecionadas e de quatro testemunhas comerciais de milho. Campinas e Mococa,
2012.............................................................................................................................. 74
Tabela 18. Estimativas de variância de progênies ( 2
pσ ), ambiental ( 2
Eσ ) e da
interação progênies por locais ( 2
a x pσ ), da herdabilidade em nível de médias ( 2h ),
ganho genético (Gs e Gs%), índice de variação (b), coeficiente de variação genética
(CVg %) e de coeficiente de variação experimental (CV%), relativas aos caracteres
avaliados nas 150 progênies de irmãos germanos interpopulacionais de milho.
Campinas e Mococa, 2012........................................................................................... 78
Tabela 19. Estimativas de variância de progênies ( 2
pσ ), ambiental ( 2
Eσ ) e da
interação progênies por locais ( 2
a x pσ ), da herdabilidade em nível de médias ( 2h ),
ganho genético (Gs e Gs%), índice de variação (b), coeficiente de variação genética
(CVg %) e de coeficiente de variação experimental (CV%), relativas aos caracteres
avaliados nas 150 progênies de irmãos germanos interpopulacionais de milho.
Campinas e Mococa, 2012...........................................................................................
79
Tabela 20. Valores e significâncias dos coeficientes de correlação fenotípica (rf)
entre os caracteres avaliados nas 20% progênies de irmãos germanos selecionadas
em Campinas e Mococa. Safrinha 2012....................................................................... 81
Tabela 21. Eficiência quântica potencial do fotossistema II (Fv/Fm), coeficientes
de extinção fotoquímica (qP) e não fotoquímica (NPQ) da fluorescência e
transporte aparente de elétrons (ETR) de seis genótipos de milho avaliados sob
condição irrigada e sob déficit hídrico no estádio vegetativo (V5). Campinas, 2013.
85

ix
Tabela 22. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos
(MG) e massa seca total (MST) de seis genótipos de milho avaliados sob condição
irrigada e sob déficit hídrico no estádio vegetativo (V5). Campinas, 2013................. 86
Tabela 23. Eficiência quântica potencial do fotossistema II (Fv/Fm), coeficientes
de extinção fotoquímica (qP) e não fotoquímica (NPQ) da fluorescência e
transporte aparente de elétrons (ETR) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico no estádio de florescimento (VT). Campinas,
2013.............................................................................................................................. 90
Tabela 24. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos
(MG) e massa seca total (MST) de seis genótipos de milho avaliados sob condição
irrigada e sob déficit hídrico no estádio de florescimento (VT). Campinas, 2013...... 91

x
LISTA DE ABREVIATURAS
Ai – Assimilação diurna de CO2
AM – Amido
AS – Açúcares solúveis
Ci – Concentração intercelular de CO2
Chl – Teor de clorofila total
CTNE – Carboidrato total não estrutural
Ei – Transpiração diurna
ETR – Taxa aparente de transporte de elétrons
EUA – Eficiência do uso da água
Fm – Fluorescência máxima
Fv – Fluorescência variável
Fv/Fm – Eficiência quântica potencial do fotossistema II
gs – Condutância estomática
qP – Coeficiente de extinção fotoquímica da fluorescência
NPQ – Coeficiente de extinção não-fotoquímica da fluorescência
SAC - Sacarose
Ψw – Potencial da água na folha
Ψsolo – Potencial matricial do solo
AP – Altura da planta
AE – Altura da espiga
CE – Comprimento da espiga
CP – Comprimento do pendão
DE – Diâmetro da espiga
FM – Florescimento masculino
FF – Florescimento feminino
IF – Intervalo entre florescimentos
MG – Massa de grãos
MST – Massa seca total
NFG – Número de fileiras de grãos na espiga
NRP – Número de ramificações do pendão
PROL - Prolificidade
SG – Stay green

1
GUIMARÃES, P.S. Respostas fisiológicas e avaliação de progênies de irmãos
germanos interpopulacionais de milho com ênfase em tolerância à seca. 2013. 135p.
Tese (Doutorado) – Instituto Agronômico – IAC, Campinas.
RESUMO
A elucidação de possíveis respostas fisiológicas responsáveis pelo comportamento
diferencial de genótipos de milho sob condição de déficit hídrico pode facilitar o
processo de seleção de genótipos tolerantes à seca. Os objetivos deste trabalho foram: a)
elucidar os mecanismos fisiológicos de tolerância à seca em híbridos de milho
contrastantes, em casa de vegetação; b) confirmar o comportamento diferencial dos
híbridos e identificar padrões de tolerância e sensibilidade à seca; c) identificar
caracteres fisiológicos que possam melhorar a eficiência na seleção de genótipos
tolerantes à seca; d) estimar parâmetros genéticos de progênies de irmãos germanos
interpopulacionais de milho e verificar a existência de variabilidade genética para
tolerância à seca e e) validar a eficiência de caracteres fisiológicos relacionados à
tolerância ao déficit hídrico na seleção de progênies superiores, em programas de
melhoramento de milho. Foram avaliados em casa de vegetação o potencial de água na
folha, trocas gasosas, atividade fotoquímica, teor de clorofila, teor de carboidratos e
caracteres biométricos dos híbridos DAS2B707, DAS2B710, FT510 e AS1522. O
delineamento experimental em casa de vegetação foi o de blocos casualizados, com
parcelas subdivididas e três repetições, com deficiência hídrica em três estádios
fenológicos da cultura. Em condições de campo foram avaliadas 150 progênies de
irmãos germanos interpopulacionais de milho, oriundas do cruzamento de uma
população tolerante com uma sensível à seca, em Campinas e Mococa, na safrinha de
2012. Foram avaliados os caracteres florescimento masculino (FM) e feminino (FF),
intervalo entre florescimentos (IF), stay green (SG), número de ramificações (NRP) e
comprimento (CP) do pendão, altura da planta (AP) e da espiga (AE), prolificidade
(PROL), comprimento (CE) e diâmetro de espiga (DE), número de fileiras de grãos na
espiga (NFG) e massa de grãos (MG). O delineamento empregado foi o de blocos
casualizados com três repetições. Foram estimados parâmetros genéticos de
herdabilidade no sentido amplo, ganho de seleção e a correlação fenotípica dos
caracteres. Pelos resultados de MG obtidos nos experimentos de campo, selecionaram-
se as duas melhores e duas piores progênies contrastantes à seca, que foram semeadas

2
em casa de vegetação, a fim de estudar os parâmetros fisiológicos e biométricos. Os
resultados obtidos dos híbridos e das progênies em casa de vegetação indicam que o
déficit hídrico reduz o potencial da água na folha (Ψw), condutância estomática (gs),
assimilação diurna de CO2 (Ai), transpiração diurna (Ei), eficiência quântica potencial
do fotossistema II (Fv/Fm), taxa aparente de transporte de elétrons (ETR), coeficiente
de dissipação fotoquímica (qP) e aumenta o coeficiente de dissipação não fotoquímica
(NPQ), principalmente quando a seca ocorre nos estádios vegetativo e de florescimento.
O híbrido DAS2B707 foi confirmado como tolerante à seca no estádio vegetativo
devido à maior eficiência dos processos fotossintéticos, enquanto o AS1522 foi
considerado tolerante à seca no estádio de florescimento devido ao elevado teor de
carboidratos foliar obtido em condições de déficit hídrico. Verificou-se que a progênie
9Bx7A é tolerante à seca e a progênie 58Bx65A foi confirmada como padrão de
sensibilidade ao déficit hídrico. Não foi possível eleger um parâmetro fisiológico
eficiente na seleção e discriminação de progênies, para futura utilização em programas
de melhoramento de milho. Obtiveram-se altos valores de herdabilidade para NFG,
NRP, FM, FF, DE e AE, demonstrando a possibilidade de se obter altos ganhos com a
seleção entre progênies. As médias de produtividade e a magnitude das estimativas dos
parâmetros genéticos indicam que existe variabilidade genética para ser explorada nos
próximos ciclos de seleção recorrente recíproca para tolerância à seca em progênies de
milho. Estimativas da correlação fenotípica indicaram que a seleção para prolificidade
(PROL) pode indiretamente aumentar a massa de grãos (MG) sob condições de déficit
hídrico.
Palavras chave: Zea mays L., seleção recorrente recíproca, parâmetros fisiológicos,
herdabilidade, melhoramento para déficit hídrico.

3
GUIMARÃES, P.S. Physiological responses and evaluation of interpopulation full-sib
progenies of maize with emphasis in drought tolerance. 2013. 135p. Agronomic
Institute – IAC, Campinas.
ABSTRACT
The elucidation of physiological responses responsible to differential behavior of maize
genotypes under water deficit conditions can facilitate the process of selection of
drought tolerant genotypes. The objectives of this study were: a) elucidate the
physiological mechanisms of drought tolerance in maize hybrids, in greenhouse; b)
confirm the differential behavior of hybrids and identify patterns of tolerance and
sensitivity to drought; c) identify physiological characters that may improve the
efficiency in the selection of drought tolerant genotypes; d) evaluate full sib progenies,
estimate genetic parameters and check for interpopulational genetic variability for
tolerance to drought and e) validate the efficiency of physiological characters related to
tolerance to water deficit in the selection of progenies, in maize breeding programs.The
DAS2B707, DAS2B710, FT510 and AS1522 hybrids were evaluated in a greenhouse
for the leaf water potential, gas exchange, photochemical activity, chlorophyll content,
carbohydrate content and biometric parameters.The experiment in the greenhouse was
in randomized blocks design in a split plot schemme with three replicates, with water
deficiency in three phenological stages. Under field conditions were evaluated 150 full
sib progenies, obtained from crosses of a drought tolerant with a to drought sensitive
population in Campinas and Mococa, during the second crop of 2012. The traits male
(FM) and female flowering (FF), interval between flowering (IF), stay green (SG),
tassel branche number (NRP) and tassel length (CP), plant (AP) and ear (AE) height,
prolificacy (PROL), ear length (CE) and diameter (DE), number of kernels rows per ear
(NFG) and grain mass (MG) were evaluated. The design used was randomized blocks
with three replications. Broad sense heritability, gain selection and phenotypic
correlation of characters were estimated. Through the MG results obtained in field
experiments, two of the best and of the worst progenies tolerant to drought were
selected and sown in greenhouse to study the physiological and biometric parameters.
The results of the hybrids and progenies in the greenhouse conditions indicated that the
water deficit reduced leaf water potential (Ψw), stomatal conductance (gs), CO2
assimilation (Ai), transpiration (Ei), potential quantum efficiency of photosystem II
(Fv/Fm), apparent electron transport rate (ETR), photochemical quenching coefficient
(qP) and increased the non-photochemical quenching (NPQ) when drought occured
during vegetative and flowering stages. The hybrid DAS2B707 was confirmed as
drought tolerant in the vegetative stage due to higher efficiency of photosynthetic
processes, while the AS1522 was considered tolerant to drought in the flowering stage
due to the high content of leaf carbohydrates obtained in water deficit conditions. The
9Bx7A was considered drought tolerant and 58Bx65A was confirmed as a water deficit

4
sensitivity pattern. It was not possible to elect a physiological efficient parameter for
selection and discrimination of progenies for future use in corn breeding programs. The
high values of heritability to NFG, NRP, FM, FF and AE demonstrate the possibility of
obtaining high gains with the selection among progenies. The productivity and the
magnitude of the estimates of genetic parameters indicate that there is genetic variability
to be explored in the next cycles of reciprocal recurrent selection for drought tolerance
in progenies of corn. Phenotypic correlation estimates indicated that the selection for
prolificacy (PROL) may indirectly increase the grain mass (MG) under water deficit
conditions.
Keywords: Zea mays L., reciprocal recurrent selection, physiological parameters,
heritability, water deficit breeding.

5
1 INTRODUÇÃO
O milho (Zea mays L.) é considerado um dos mais importantes produtos do setor
agrícola no mundo, devido à sua versatilidade de uso como fonte de carboidratos na
alimentação humana e animal e pelo aspecto social na geração de empregos.
Atualmente seu uso se ampliou com a produção de amido, adoçantes, óleos e também
surge como fonte de biocombustíveis (AGRIANUAL, 2010).
A evolução da produção mundial de milho vem sendo expressiva nas últimas
duas décadas, passando de 453 milhões de toneladas no final da década de 80 para os
atuais 860,1 milhões na safra 2011/12, o que corresponde a um aumento de 90% no
período. No Brasil a área semeada com milho em 2012 foi de 14,85 milhões de hectares
e a produção nacional ficou em torno de 73 milhões de toneladas (CONAB, 2013). No
entanto, a produtividade da cultura é afetada por condições adversas, como a de déficit
hídrico que atinge praticamente todas as áreas produtoras de milho durante algum
estádio de desenvolvimento da cultura.
As previsões ambientais apontam para o aumento do aquecimento global nas
próximas décadas e situações de secas acompanharão esse evento. De acordo com o
Painel Intergovernamental das Mudanças Climáticas – IPCC, muitos ecossistemas
naturais estão sendo afetados pelas mudanças climáticas, principalmente pela elevação
da temperatura (IPCC, 2007).
O emprego de cultivares tolerantes à seca é uma estratégia eficiente para a
redução dos riscos na produção em áreas sujeitas a períodos constantes de déficit
hídrico. Entretanto, a tolerância à seca é uma característica bastante complexa. Algumas
plantas conseguem suportar a falta de água por meio de diferentes mecanismos,
envolvendo adaptações morfofisiológicas que permitem à planta sobreviver e produzir
satisfatoriamente (XOCONOSTLE-CAZARES et al., 2010). Assim, características
diferentes têm sido propostas como critérios de seleção visando maior tolerância à seca.
De fato, não há um único parâmetro que por si só possa ser indicativo da
tolerância à seca, sendo o ideal avaliar caracteres que possam ser medidos em diferentes
fases de desenvolvimento da cultura, em condições de casa de vegetação e em
experimento de campo (PIMENTEL & ROSSIELO, 1995). A seleção de genótipos de

6
milho tolerantes ao déficit hídrico e com elevado rendimento de grãos seria mais
eficiente se os atributos que propiciam o mesmo sob condições limitantes de água
pudessem ser identificados e utilizados como critérios de seleção (DURÃES et al.,
1997). A identificação destes genótipos pode ser realizada pela análise de características
fisiológicas, auxiliando o melhoramento genético na busca de caracteres que otimizam o
crescimento e conferem maior tolerância ao estresse.
Atualmente, pesquisas sobre melhoramento de milho para a tolerância à seca
vêm avançando nas instituições de pesquisa, demonstrando resultados satisfatórios na
obtenção de genótipos tolerantes ao déficit hídrico. Entretanto, o conhecimento sobre os
mecanismos fisiológicos para a tolerância ainda é considerado incipiente para a cultura
do milho, necessitando de mais estudos. Diante disso, a caracterização dos genótipos,
bem como a elucidação dos possíveis mecanismos responsáveis pelo desempenho
diferencial dos genótipos sob condição de déficit hídrico, pode facilitar o processo de
obtenção de novos materiais, além de contribuir para o desenvolvimento de técnicas de
seleção capazes de reduzir o tempo e o trabalho para avaliar fontes de genes para
tolerância à seca (QUEIROZ, 2010).
Uma vez selecionados os genótipos que atendam às exigências do programa de
melhoramento voltado para tolerância à seca, pode-se realizar o método de seleção
recorrente recíproca para aumentar a freqüência de alelos favoráveis gradativamente por
meio de repetidos ciclos de seleção. Tal método é utilizado em duas populações
simultaneamente, sem que haja perda da variabilidade genética de ambas as populações
(PINTO, 1995). A falta de informação sobre trabalhos desse tipo para obtenção de
genótipos tolerantes a fatores abióticos, como a seca, revela a importância de estudos
relacionados com esse tema. Por esta razão, a seleção de caracteres relacionados à
tolerância à seca deve ser enfatizada em programas de melhoramento de milho, visando
fornecer informações para o desenvolvimento de genótipos de milho capazes de tolerar
períodos de déficit hídrico sem que a produtividade seja substancialmente prejudicada.
Diante disso, os objetivos do trabalho foram: a) elucidar os mecanismos
fisiológicos de tolerância à seca em híbridos de milho contrastantes, em casa de
vegetação; b) confirmar o comportamento diferencial dos híbridos e identificar padrões
de tolerância e sensibilidade à seca; c) identificar caracteres fisiológicos que possam
melhorar a eficiência na seleção de genótipos tolerantes à seca; d) estimar parâmetros
genéticos de progênies de irmãos germanos interpopulacionais de milho e verificar a
existência de variabilidade genética para tolerância à seca e e) validar a eficiência de

7
caracteres fisiológicos relacionados à tolerância ao déficit hídrico na seleção de
progênies superiores, em programas de melhoramento de milho.
2 REVISÃO DE LITERATURA
2.1 Déficit hídrico
O déficit hídrico ocorre quando o conteúdo de água de um tecido ou célula está
abaixo do conteúdo de água mais alto exibido no estado de maior hidratação (TAIZ &
ZEIGER, 2009). Geralmente é causado por um fator externo, que exerce influência
desvantajosa sobre a planta, provocando desvio significativo das condições ótimas para
a vida, os quais são reversíveis a princípio, mas podem se tornar permanentes
(LARCHER, 2004). A deficiência hídrica é um dos estresses que causa maiores danos
nos processos fisiológicos e metabólicos das plantas, acarretando reduções na
produtividade (PIMENTEL, 2004).
Em condições adversas de umidade, TAIZ & ZEIGER (2009) mencionam a
ocorrência de três linhas de defesa das plantas para amenizar os efeitos do déficit
hídrico sobre o crescimento vegetal. A primeira linha de defesa é a redução da área
foliar, devido à diminuição do turgor das células e o aumento da senescência das folhas,
o que provoca uma redução da perda de água por transpiração. A segunda linha é o
aprofundamento das raízes no solo úmido, pois a atividade fotossintética da planta em
condições de estresse é menos atingida que a expansão foliar, o que permite maior
acúmulo de assimilados vegetais distribuídos ao sistema radicular. A terceira linha de
defesa refere-se ao fechamento dos estômatos, principalmente em condições onde o
início do estresse é mais rápido ou onde a planta atingiu sua área foliar plena, evitando a
perda de água por transpiração.
Há também os mecanismos de resistência à deficiência hídrica, que podem ser
divididos em escape, retardo e tolerância. No primeiro, as plantas adotam uma estratégia
de “fuga”, na qual apresentam rápido desenvolvimento fenológico e alto grau de
plasticidade, sendo capazes de completar seu ciclo de vida antes que a falta de água
torne-se severa o bastante para provocar danos fisiológicos. O retardo da desidratação
corresponde à manutenção do turgor e volume celular, tanto pela absorção de água por
um sistema radicular abundante quanto pela redução da perda por transpiração por

8
intermédio do fechamento estomático ou por vias não estomáticas como a cutícula. E a
tolerância à seca é um mecanismo que permite à planta manter o metabolismo, mesmo
com a redução do potencial hídrico dos tecidos, devido principalmente ao acúmulo de
solutos compatíveis ou osmólitos, proteínas osmoprotetoras e à capacidade antioxidante
(VERSLUES et al., 2006).
Segundo TAIZ & ZEIGER (2009), a tolerância das plantas frente a um estresse
pode ser por aclimatação ou por adaptação. Se a tolerância aumenta como consequência
de exposição anterior ao estresse, diz-se que a planta está aclimatada, mas se ocorre
tolerância determinada geneticamente adquirida por processo de seleção durante muitas
gerações, a planta está adaptada.
Em condição de déficit hídrico, o milho apresenta uma série de alterações
morfofisiológicas, causando assim, distúrbios em seu metabolismo. Dentre as alterações
apresentadas pelas plantas de milho submetidas à deficiência hídrica podem-se destacar:
alterações na altura da planta, no comprimento e na largura das folhas, na condutância
estomática, transpiração, na eficiência quântica do fotossistema II, no potencial de água
da folha, no conteúdo de clorofila, na atividade fotossintética, que visam reduzir os
efeitos deletérios da baixa disponibilidade hídrica, constituindo, portanto, mecanismos
de tolerância à seca (PIMENTEL, 2004).
2.2 Déficit hídrico na cultura do milho
O rendimento da cultura do milho é extremamente dependente das condições
hídricas durante o desenvolvimento da planta. Segundo BÄZINGER et al. (2000), em
condições de déficit hídrico, a produtividade de grãos é substancialmente afetada, sendo
os estádios de florescimento e enchimento de grãos considerados as fases mais críticas
da cultura. Entretanto, devido à ocorrência de variabilidade genética entre os genótipos
de milho para adaptação quanto à seca, deve-se avaliar o comportamento de diferentes
materiais em condições de déficit hídrico para recomendação de cultivo (BLUM, 1997).
No Brasil, a maioria das áreas de cultivo de milho não empregam irrigação e
mesmo em anos regulares de precipitação pluvial observam-se, normalmente, perdas na
produção devido a períodos de estiagem denominados “veranicos” (QUEIROZ, 2010).
As oscilações nas safras de milho das principais regiões produtoras do Brasil estão
associadas à disponibilidade de água, sobretudo nos estádios mais críticos da cultura.

9
Diante disso, torna-se essencial avaliar os efeitos da disponibilidade hídrica durante
todo o ciclo fenológico da cultura.
O milho é uma planta de metabolismo fotossintético C4, onde o primeiro produto
estável da fotossíntese, formado durante a fixação da molécula de dióxido de carbono
(CO2), é o ácido oxalacético que é reduzido a ácido málico e ácido aspártico, ambos
com quatro moléculas de carbono ainda mais estáveis. Além da presença da enzima
rubisco nas células da bainha Kranz, as plantas consideradas C4 contam com a presença
da enzima fosfoenolpiruvato carboxilase nas células do mesófilo foliar, responsável por
aumentar a capacidade de fixação de CO2 devido a sua alta afinidade por esta molécula.
A compartimentação espacial das duas enzimas faz com que o CO2 fixado pela
fosfoenolpiruvato carboxilase se transloque, via malato e aspartato, até a bainha dos
feixes vasculares, onde ocorre a descarboxilação com a entrada do carbono no ciclo de
Calvin-Benson (TAIZ & ZEIGER, 2009).
Por ser uma espécie de metabolismo C4, o milho tende a expressar sua elevada
produtividade quando a máxima área foliar coincide com a maior disponibilidade de
radiação solar, desde que não haja déficit hídrico. Essa condição permite a máxima
fotossíntese possível, porém aumenta a necessidade hídrica da cultura, já que o elevado
fluxo energético incidente também eleva a evapotranspiração (BERGAMASCHI et al.,
2004).
Sabe-se que a cultura é cultivada em regiões cuja precipitação varia de 300 a
5000 mm anuais, sendo que a quantidade de água consumida por uma planta, durante o
seu ciclo, é de aproximadamente 600 mm (ALDRICH et al., 1982). Logo, o milho
sofrerá redução da produção quando ocorrer déficit hídrico nos principais estádios
fenológicos da cultura. No estádio vegetativo caracterizado por V3 a V5 (com três a
cinco folhas completamente desenvolvidas) ocorre sensibilidade à limitação hídrica.
Durante o estádio V3 estão sendo formadas todas as folhas e espigas que a planta
eventualmente irá produzir. Pode-se dizer, portanto, que o número máximo de grãos ou
a produção potencial estão sendo definidos nesse estádio. No estádio V5, tanto a
iniciação das folhas como das espigas estará completa e a iniciação do pendão já pode
ser vista microscopicamente na extremidade de formação do caule, logo abaixo da
superfície do solo (MAGALHÃES et al., 1994). Durante esse período vegetativo, o
deficit hídrico também reduz o crescimento do milho em função da redução da área
foliar e da biomassa (BERGAMASCHI et al., 2006).

10
Segundo EDMEADES et al. (1999), períodos de estiagem durante o
florescimento provocam aumentos consideráveis no intervalo entre o florescimento
masculino e feminino, devido ao retardamento do florescimento feminino e também
provoca o abortamento de grãos durante duas a três semanas após o florescimento
feminino. A falta de água nesse período, além de afetar o sincronismo pendão-espiga,
pode reduzir a chance de aparecimento de uma segunda espiga em materiais prolíficos.
Na fase de enchimento de grãos, em especial quando os grãos apresentam
aparência amarelada e no seu interior um fluido de cor leitosa, ocorre o início da
transformação dos açúcares em amido, contribuindo para o incremento do peso seco dos
grãos. Esse incremento ocorre devido à translocação dos fotoassimilados presentes nas
folhas e no colmo para a espiga e grãos em formação. A eficiência dessa translocação,
além de ser importante para a produção, é extremamente dependente de água
(MAGALHÃES et al., 1998). O rendimento final depende do número de grãos em
enchimento (desenvolvimento) e do tamanho final (período de enchimento) que eles
alcançarão. Portanto, um déficit hídrico nessa fase pode afetar a produção. Com o
processo de maturação dos grãos, o potencial de redução na produção final de grãos
devido ao déficit hídrico vai diminuindo. Embora, nesse período, a planta deva
apresentar considerável teor de sólidos solúveis prontamente disponíveis, objetivando a
evolução do processo de formação de grãos, a fotossíntese mostra-se imprescindível.
QUEIROZ (2010) afirma que dois dias de déficit hídrico no florescimento
reduzem o rendimento em mais de 20%, enquanto que, quatro a oito dias no estádio de
enchimento de grãos diminuem a produção em mais de 50%. Segundo BÄZINGER et
al. (2000), quando o período de déficit hídrico se estende num intervalo entre a
emergência do pendão e o início do enchimento de grãos, pode ocorrer a completa
esterilidade das plantas de milho. BERGONCI et al. (2001) e BERGAMASCHI et al.
(2004 e 2006) indicam que o período mais crítico da cultura vai do pendoamento ao
início do enchimento de grãos. De acordo com MAGALHÃES et al. (2002), o aumento
no acúmulo de matéria seca nos grãos está intimamente relacionado à fotossíntese e,
uma vez que o estresse afeta este processo, reduz a produção de carboidratos,
implicando em menor acúmulo de matéria seca nos grãos. Por isso, para seu
desenvolvimento, o milho necessita de quantidades substanciais de água para uma
adequada produtividade em campo.
Diante desse fato, fica evidente que períodos prolongados de déficit hídrico na
cultura do milho podem causar impactos significativos no crescimento, na

11
produtividade e na qualidade dos grãos, sendo a habilidade das plantas em tolerar
períodos de seca uma importante estratégia para o desenvolvimento do agronegócio em
qualquer país.
Um exemplo de significativa redução da produtividade de grãos devido a
período de estiagem ocorreu nos Estados Unidos no ano de 2012, provocando aumentos
significativos dos preços dos alimentos, principalmente nos locais onde o milho se
constitui em alimento básico na dieta da população. O mais recente relatório do
Departamento de Agricultura Americano (USDA) revelou que a situação da cultura no
chamado Cinturão do Milho foi considerada crítica, uma vez que as principais áreas
produtoras decretaram situação de emergência devido à escassez de água (USDA,
2012). O impacto prolongado de seca nos EUA influenciou diretamente o preço do
milho no Brasil, provocando redução na produção de aves e suínos nas principais
regiões produtoras, uma vez que ocorreu significativo aumento do preço da ração
animal e, consequentemente, elevou o valor da carne e dos derivados desses animais,
que foram repassados aos consumidores. Portanto, estudos relacionados à tolerância à
seca envolvendo a cultura do milho podem trazer benefícios em relação ao crescimento
e no rendimento da cultura em regiões com limitação hídrica.
2.3 Parâmetros fisiológicos em milho submetido ao déficit hídrico
O crescimento celular é considerado o processo mais sensível a baixa
disponibilidade de água no solo, sendo a divisão e a expansão celular diretamente
inibidas pelo déficit hídrico (ZHU, 2001; SAUSEN, 2007). Esta sensibilidade se deve
ao impacto da falta de água sobre a taxa de expansão das células, devido à perda de
turgor (SAUSEN, 2007, KUNZ et al., 2007). Entretanto, a mudança mais visível no
dossel vegetativo da cultura de milho sob déficit hídrico é o enrolamento das folhas,
cuja intensidade está relacionada com o potencial da água da folha (MOULIA, 2000).
As medições do potencial da água da folha podem fornecer informações que
permitem identificar o estado hídrico em que as plantas se encontram. Para a maioria
das espécies, o potencial da água da folha acompanha as variações diurnas da demanda
evaporativa da atmosfera, atingindo um valor máximo logo antes do nascer do sol.
Nesse momento, as plantas encontram-se com a máxima turgescência possível para uma
dada condição hídrica no solo (KRAMER & BOYER, 1995). Dessa forma, o potencial
hídrico avaliado na antemanhã tem sido considerado um indicativo do estado hídrico

12
das plantas, bem como da quantidade de água disponível no solo (LARCHER, 2004 e
TAIZ & ZEIGER, 2009).
Para LARCHER (2004) uma das primeiras respostas para manter o nível hídrico
no interior da planta é fechamento dos estômatos. No entanto, quando isso acontece, se
restringe às trocas gasosas entre o interior da folha e a atmosfera, o que provoca a
redução no fluxo de CO2. Esse controle da perda de água na planta, realizado
principalmente pelos estômatos, é regulado geralmente pela diferença na pressão de
vapor entre o ambiente e a planta e por sinalizadores endógenos, como o ácido abscísico
(JIA et al., 1996). Esses reguladores endógenos modulam a condutância estomática, a
temperatura da planta e promovem alterações como o murchamento e o enrolamento das
folhas.
Diante disso, o controle das trocas gasosas é considerado um processo
complexo, pois a ligeira diminuição de turgescência celular, suficiente para causar o
fechamento dos estômatos, torna a absorção de CO2 difícil, diminuindo sensivelmente a
assimilação de carbono. As plantas necessitam abrir os estômatos para a entrada do CO2
e também fechá-los para evitar a perda de água, porém a tendência é favorecer a
assimilação fotossintética (PIMENTEL, 1998; ANGELOCCI, 2002).
Em condições de restrição hídrica, a fotossíntese pode ser reduzida por
limitações metabólicas pelo aumento do efeito fotoinibitório (GLAZ et al., 2004). Por
isso, o uso de parâmetros de fluorescência tem sido difundido principalmente no estudo
de fotossíntese por ser um método que permite a análise da absorção e aproveitamento
da energia luminosa através do fotossistema II e possíveis relações com a capacidade
fotossintética (KRAUSE & WEIS, 1991). Esta técnica tem permitido um aumento no
conhecimento dos processos fotoquímicos e não fotoquímicos que ocorrem na
membrana dos tilacóides dos cloroplastos (ROHÁCEK, 2002), além de possibilitar o
estudo de características relacionadas à capacidade de absorção e transferência da
energia luminosa na cadeia de transporte de elétrons, sendo possível também estudos
das mudanças conformacionais dos tilacóides (KRAUSE & WEIS, 1991).
A eficiência quântica potencial do fotossistema II (Fv/Fm) obtida a partir da
fluorescência da clorofila a, pode indicar o nível energético de excitação dos pigmentos
que dirigem a fotossíntese. A habilidade em manter elevadas razões Fv/Fm sob déficit
hídrico pode ser um indicativo de eficiência no uso da radiação pela fotoquímica
(TESTER & BACIC, 2005). Desta forma, as medidas de trocas gasosas e de
fluorescência da clorofila a podem ser utilizadas como ferramentas para diagnosticar a

13
integridade do aparato fotossintético frente às adversidades ambientais (TORRES
NETTO et al., 2005). Há evidências de que os efeitos do déficit hídrico sobre o
fotossistema II podem ser mediados pela produção e acumulação de espécies reativas de
oxigênio (LAWLOR, 1995), provocando danos às membranas, desencadeando
processos peroxidativos de lipídios, com perda de eletrólitos pela célula (QUEIROZ et
al., 1998) e queda na atividade fotossintética (SMIRNOFF, 1993).
Outras formas das plantas de milho conseguirem tolerar a deficiência hídrica é o
mecanismo de acumulação de solutos osmoticamente ativos na célula (MORGAN,
1984). Esse fenômeno é chamado de ajustamento osmótico e permite a manutenção da
turgescência, o crescimento da planta, a manutenção da abertura estomática e a
fotossíntese sob condições de baixo potencial da água da folha (CHAVES, 1991) e sob
baixos valores de potencial da água do solo (PATAKAS et al., 2002; VIEIRA JUNIOR
et al., 2007). A maioria das modificações que as plantas desenvolvem visa manter o
crescimento e a reprodução da planta em ambientes com limitações na disponibilidade
de água. Durante o período de seca, plantas superiores acumulam açúcares, ácidos
orgânicos e íons no citosol para diminuir o potencial osmótico e o potencial hídrico da
planta mais rapidamente que o do solo e, conseqüentemente, conseguem manter o
potencial hídrico e o turgor de suas células próximo do nível ótimo (TURNER &
JONES, 1980).
De acordo com KRAMER (1974), as plantas se adaptam ao estresse utilizando-
se de mecanismos de escape e tolerância podendo, assim, sofrer adaptações
morfológicas e fisiológicas. Essas respostas dependem da duração, da intensidade e da
velocidade do déficit hídrico (CARLESSO et al., 1996). MAGALHÃES (1995) afirma
que plantas de milho submetidas à restrição hídrica acumulam açúcares solúveis nas
folhas e reduzem a quantidade de amido. De acordo com o mesmo autor, a maior
mobilização de sacarose pelas plantas ocorre, mas não de modo generalizado, durante a
noite, quando a planta esta relativamente mais hidratada e sujeita às temperaturas mais
moderadas.
De modo geral, o emprego de genótipos tolerantes à deficiência hídrica tem sido
indicado como a melhor ferramenta para o aumento da produtividade em condições
onde existe dificuldade no controle do ambiente devido à variação pluviométrica.
Porém, para se desenvolver cultivares tolerantes ao estresse, é necessário o uso de
estratégias de melhoramento apropriadas e a aplicação de conhecimentos no manejo dos
ambientes para maximizar o ganho genético dos caracteres desejados.

14
2.4 Caracteres primários e secundários relacionados à tolerância ao déficit
hídrico em milho
Caracterizar o estresse hídrico em trabalhos de melhoramento genético vegetal é
um problema amplo, variando de um local para o outro e do período da safra. Além
disso, fica difícil distinguir os efeitos diretos de déficit hídrico de outros fatores como
fungos, baixa fertilidade do solo e altas temperaturas. Por isso, colocar no mercado
genótipos de milho produtivos e com características de tolerância à seca é um desafio
contínuo para os programas de melhoramento dessa cultura, pois o déficit hídrico é a
maior fonte de instabilidade do rendimento de grãos em áreas tropicais. Agrega-se a isto
o fato da herdabilidade para a característica produção de grãos ser baixa. Portanto,
genótipos melhor adaptados e com maior rendimento poderiam ser mais eficientes se
atributos que conferem rendimento sob condições limitantes de água pudessem ser
identificados e usados como critério de seleção (DURÃES et al., 2004).
Caracterizar o ambiente e identificar caracteres primários e secundários de
plantas visando à obtenção, seleção e uso de genótipos responsivos a fatores ambientais
subótimos, além de realizar melhorias nas práticas de manejo, são tarefas complexas e
requerem métodos adequados. De acordo com DURÃES et al. (2004) os caracteres
primários referem-se ao rendimento de grãos e aos componentes do rendimento,
possuindo baixa herdabilidade. Os secundários são os caracteres que possuem rápido e
baixo custo de mensuração e alta herdabilidade, estando relacionadas com rendimento
de grãos, ângulo foliar, stay green, tamanho pequeno de pendão e baixo intervalo entre
florescimentos feminino e masculino (EDMEADES et al., 1998).
Segundo CÂMARA et al. (2007) a seleção para produção de grãos sob déficit
hídrico tem sido considerada ineficiente em vista da alta proporção da variância
ambiental em relação à genética, o que reduz a herdabilidade do caráter e dificulta a
seleção de genótipos superiores. Nessas condições, o emprego de caracteres secundários
pode melhorar a eficiência da seleção se estes possuírem valor adaptativo ao estresse,
alta herdabilidade, correlação genética alta e significativa com a produção de grãos e
que sejam de fácil mensuração (BÄNZINGER et al., 2000; KAMARA et al., 2003).
BÄNZINGER et al. (2000) afirmam que o caráter prolificidade é provavelmente
o de maior importância em programas de melhoramento de milho para tolerância à seca,
uma vez que possui grande associação com produção de grãos sob condições de estresse
(LI et al., 2003). Genótipos de milho prolíficos têm apresentado melhor capacidade de

15
adaptação em ambientes estressantes como, por exemplo, ao adensamento de semeadura
e a tolerância a déficits hídricos (BOLAÑOS & EDMEADES, 1993a), o que minimiza
os riscos de redução da produtividade. Segundo CÂMARA (2006), a maior tolerância
ao défict hídrico nos genótipos prolíficos pode estar relacionado a um sistema radicular
mais desenvolvido, enquanto que, VARGA et al. (2004) relataram que híbridos
prolíficos possuem melhor eficiência no uso dos recursos ambientais e maior
estabilidade de produção quando submetidos a uma situação de estresse.
Alguns trabalhos relacionados ao tema demostram que sob condições de déficit
hídrico a herdabilidade para a prolificidade se mantém constante ou até mesmo aumenta
com o estresse (BOLAÑOS & EDMEADES, 1996), diferentemente da produção de
grãos. De maneira geral, o caráter apresenta herdabilidade de média a alta, apresentando
estimativas de herdabilidade em torno de 70% (AGUIAR, 2003; CÂMARA et al.,
2007). Estudando o caráter prolificidade, SOARES FILHO (1987) verificou que o
controle genético parece ser predominantemente aditivo. Em alguns trabalhos a
variância genética aditiva explicou cerca de 75% da variância genética total para
prolificidade (ALVES et al., 2002), enquanto que SILVA (2002) obteve estimativa da
variância aditiva três vezes maior que a de dominância para este caráter.
Frequentemente na literatura encontra-se relatos da associação positiva entre
produção de grãos e prolificidade (SOUZA JUNIOR et al., 1985; ZAIDI et al., 2004),
com valores de correlação superiores a 0,5 (SOUZA JUNIOR et al., 1985; AGUIAR,
2003; KAMARA et al., 2005; MEDICI et al., 2005). Devido à prolificidade possuir alta
herdabilidade e estar correlacionada positivamente com a produção de grãos, a seleção
direta para o caráter tende a ser eficiente e aumenta indiretamente a produtividade.
Assim como a prolificidade, o intervalo entre o florescimento feminino e
masculino também é considerado um dos mais importantes caracteres a serem
selecionados para a obtenção de genótipos de milho tolerantes à seca (BÄZINGER et
al., 2000; XIAO et al., 2004), uma vez que a protandria no milho é acentuada sob
condições de déficit hídrico devido ao maior retardamento do florescimento feminino
em relação ao masculino. Desse modo, dependendo da extensão do período de seca, a
produção de grãos tende a ser afetada em função da assincronia entre liberação de pólen
e dos estilo-estigmas.
BOLAÑOS & EDMEADES (1993b), avaliando ciclos avançados de seleção
baseando-se na produção de grãos e no intervalo entre florescimento (IF) em plantas de
milho submetidas à diferentes tratamentos de regime hídrico, verificaram uma redução

16
de 8,7% na produção por dia de acréscimo em IF. Como a cultura do milho é
considerada sensível à seca nos estádios de florescimento e de enchimento de grãos, a
falta de água nessas fases de desenvolvimento provoca séria instabilidade de produção
(KAMARA et al., 2003). Portanto, em progamas de melhoramento visando a formação
de genótipos com maior tolerância ao déficit hídrico, faz-se necessária uma redução no
intervalo entre florescimentos (BOLAÑOS & EDMEADES, 1993a).
Em condições de déficit hídrico têm-se observado estimativas de herdabilidade
constantes para florescimento feminino e intervalos entre florescimentos (BOLAÑOS &
EDMEADES, 1996), diferentemente da herdabilidade para produção de grãos que reduz
em condições de seca, dificultando a seleção de genótipos superiores (KAMARA et al.,
2003).
Resultados observados na literatura demostram que o intervalo entre
florescimentos apresenta coeficiente de herdabilidade de magnitude média a alta,
indicando que o caráter deve responder em níveis adequados à seleção (CÂMARA,
2006). Estimativas de herdabilidade para intervalo entre florescimentos apresenta
variação de 0,51 a 0,78 (BOLAÑOS & EDMEADES, 1996; JIANG et al., 1999).
Normalmente existe elevada e significativa correlação entre IF e produção de
grãos (KAMARA et al., 2003) e essa correlação aumenta principalmente em condições
de déficiência hídrica. Sob condições de estresse intemediário a severo, RIBAULT et al.
(1997) constataram significativa correlação fenotípica entre esses dois caracteres,
ficando em torno de -0,40. Os mesmos autores verificaram que a seleção para redução
do IF em variedades de milho tropicais de polinização aberta aumenta o rendimento de
grãos sob déficit hídrico.
Estudando a herança do caráter IF, SILVA (2002) observou que a estimativa de
variância genética aditiva apresentou valor seis vezes maior que a de dominância. Essa
tendência também foi verificada para o florescimento feminino e masculino.
MICKELSON et al. (2002), afirmam que o pendão reduz a produção de grãos
devido a diminuição na inteceptação de luz pela copa da planta e também pela
competição por fotoassimilados. Dentre os mecanismos responsáveis pela competição
por fotoassimilados está a produção de ácido indol acético (AIA) no pendão que
proporciona uma dominância apical por nutientes, inibindo o desenvolvimento da
espiga. Portanto, para a expressão da prolificidade, é necessário uma redução na
quantidade de AIA, que pode ser conseguida pela diminuição no tamanho e no número
de ramificações do pendão (CÂMARA, 2006). Assim, mais de uma espiga pode se

17
desenvolver simultaneamente em um menor espaço de tempo, induzindo a coincidência
da antese com a emergência dos estilos-estigmas, reduzindo o intervalo entre
florescimentos feminino e masculino (SOUZA JUNIOR et al., 1985). Portanto, o caráter
número de ramificações de pendão (NRP) e comprimento do pendão (CP) influenciam
de forma direta a prolificidade e o intervalo entre florescimentos. Alguns trabalhos têm
evidenciado que o número de ramificação de pendão apresenta de média a alta
herdabilidade, com estimativas de coeficiente de herdabilidade variando de 46% a 90%
(AGUIAR, 2003 e FLINT-GARCIA et al., 2005).
O caráter stay green ou senescência retardada de folhas e colmo vem sendo
observado em algumas espécies de importância econômica como o milho. Esse caráter
confere ao genótipo um melhor aproveitamento da fotossíntese a qual irá fornecer
carboidratos para colmos, folhas e raízes, e conseqüentemente maior resistência das
plantas a estresses bióticos e abióticos como déficit hídrico (JIANG et al., 2004;
CARMO et al., 2007). Segundo COSTA et al. (2008), a contribuição dos efeitos gênicos
aditivos é maior que a dos efeitos gênicos não aditivos na expressão fenotípica do
caráter stay green em milho. Em estudos de correlação, BETRÁN et al. (2003),
verificarm estimativas de correlação fenotípica entre stay green e produção de grãos sob
estresse hídrico severo variando de -0,38 a -0,48. ZAIDI et al. (2004), observaram
correlação significativa entre produção de grãos e stay green em condições de déficit
hídrico.
De maneira geral, uma das maiores limitações para o melhoramento genético
visando a tolerância à seca em plantas é o conhecimento insuficiente sobre as bases
fisiológicas, moleculares e genéticas das respostas das plantas ao déficit hídrico
(DEDEMO, 2006). Uma resposta fisiológica específica ao déficit hídrico representa na
verdade combinações de eventos moleculares que são ativados ou desativados pela
percepção do estresse. Compreender como tais eventos interagem representa um
importante passo no desenvolvimento de plantas com maior tolerância à seca.
Importante ressaltar também que a falta de informações sobre trabalhos de
progênies de irmãos germanos para obtenção de genótipos tolerantes à seca mostra a
importância de estudos relacionados com esse tema. CÂMARA et al. (2007) estudando
progênies F2:3 quanto a parâmetros genéticos de caracteres relacionados à tolerância à
seca em milho tropical, demonstraram que o desenvolvimento de cultivares com alta
produção de grãos e boa tolerância ao estresse poderia ser obtida pela seleção de

18
caracteres como prolificidade, intervalo entre florescimento, stay green e produção de
grãos sob condições normais de umidade.
Sendo assim, torna-se importante a identificação e a caracterização de genótipos,
bem como os estudos sobre a interação e sobreposição de mecanismos. A elucidação
desses mecanismos facilitará, por certo, o processo de formação de novos materiais,
além de contribuir para o desenvolvimento de técnicas de seleção que podem reduzir o
tempo e o trabalho para avaliação de fontes genéticas de tolerância a estresses abióticos,
como seca.
2.5 Seleção recorrente
A seleção recorrente é um processo cíclico de seleção que visa aumentar de
forma progressiva e contínua a freqüência dos alelos favoráveis e, simultaneamente,
manter a variabilidade genética em níveis adequados para os próximos ciclos seletivos.
Esse método de melhoramento foi primeiramente empregado por HULL (1945)
como sendo uma re-seleção, geração após geração, com intercruzamento entre os
materiais selecionados, visando obter a recombinação gênica para elevar a freqüência de
alelos favoráveis e manter a endogamia em baixo nível, assegurando um alto grau de
variabilidade genética (PINTO, 1995).
Segundo HALLAUER & MIRANDA FILHO (1988) cada ciclo do método
envolve três fases de condução, de acordo com esta sequência, exceto para o método de
seleção massal: 1) formação de progênies (meios-irmãos, irmãos germanos, S1, S2); 2)
avaliação das progênies em experimentos com repetições, visando seleção das melhores
com base nos caracteres de interesse e 3) intercruzamento das progênies selecionadas
para formar a nova população para o próximo ciclo de seleção.
As seleções são repetidas até que a freqüência de alelos favoráveis na população
atinja níveis satisfatórios. A seleção recorrente pode ser conduzida no melhoramento
das populações per se (intrapopulacional) ou da geração F1 (híbrido interpopulacional),
resultante do cruzamento entre duas populações (seleção recorrente recíproca ou
interpopulacional).
Segundo MIRANDA FILHO & NASS (2001), o sucesso de um programa de
melhoramento depende da herdabilidade, da escolha do método de melhoramento e da
intensidade de seleção dos caracteres desejados. Por isso, a escolha dos genótipos

19
superiores visa explorar ao máximo a variância genética aditiva, com finalidade de
atribuir um maior potencial aos caracteres selecionados, no próximo ciclo de seleção.
De acordo com BORÉM (2001), a população inicial à qual será aplicada a
seleção recorrente, pode ser uma variedade de polinização aberta, variedade sintética,
gerações F2 de híbrido ou clone, mas é de fundamental importância que tais populações
possuam um elevado comportamento médio e variabilidade genética suficiente para
assegurar um progresso contínuo durante os ciclos de seleção.
2.6 Seleção recorrente recíproca com irmãos germanos de milho
Proposta por COMSTOCK et al. (1949), a seleção recorrente recíproca visa
melhorar simultaneamente duas populações geneticamente distintas que possuem
elevado potencial agronômico. Os genótipos das duas populações são avaliados em
cruzamentos recíprocos e a recombinação dos genótipos superiores é realizada dentro de
cada população, mantendo suas identidades (SOUZA JÚNIOR, 2001). Portanto, na
seleção recorrente recíproca, se torna necessário o desenvolvimento de dois tipos de
progênies, as interpopulacionais para avaliação e as intrapopulacionais para
recombinação, buscando, dessa forma, o melhoramento das populações “per se” e da
heterose que se manifesta nos cruzamentos.
Com a seleção recorrente recíproca, teoricamente, se tira vantagem tanto dos
efeitos aditivos, pela concentração dos alelos favoráveis em ambas populações
empregadas no experimento, bem como dos desvios de dominância uma vez que se
mantém a distância entre as populações, permitindo explorar o fenômeno da heterose
por meio do cruzamento entre as populações e/ou de linhagens oriundas das mesmas
(SANTOS et al., 2003). Neste método os genótipos que apresentarem melhor
capacidade de combinação com a população recíproca são recombinados podendo,
assim avaliar tanto a capacidade geral quanto a capacidade específica de combinação,
aumentando a resposta heterótica no cruzamento entre as duas populações (PINTO,
1995).
O procedimento original da seleção recorrente recíproca envolve as seguintes
etapas: 1) autofecundação de plantas da população A e cruzamento simultâneo com
plantas da população B. Da mesma maneira plantas da população B são autofecundadas
e cruzadas com plantas da população A. Para os cruzamentos, a planta autofecundada é
cruzada com quatro a cinco plantas da outra população; 2) avaliação em ensaios de

20
produção das progênies formadas pelos cruzamentos e 3) recombinação das melhores
progênies S1 de A de acordo com os resultados dos ensaios. O mesmo é feito com as
melhores progênies S1 de B, obtendo-se, assim, as duas populações de primeiro ciclo,
AI e BI. Um novo ciclo pode ser iniciado, repetindo-se as etapas apresentadas.
PATERNIANI & VENCOVSKY (1977) apontaram algumas limitações do
método como: trabalho oneroso na realização de autofecundações e cruzamentos
simultâneos, além do baixo número de genótipos avaliados; falta de representatividade
do testador, uma vez que a amostragem de quatro a cinco plantas cruzadas com a planta
autofecundada pode não ser uma amostra adequada do testador, o que reduz a precisão
da avaliação; falta de recombinação adequada entre as progênies S1 selecionadas, a qual
seria obtida por pelo menos uma geração a mais após os cruzamentos entre as progênies
S1; e grande intervalo de tempo entre os ciclos resultando em menos ganho por ano e
menor oportunidades de selecionar para os anos médios.
HALLAUER & EBEHART (1970) propuseram a metodologia da seleção
recorrente recíproca de famílias de irmãos germanos obtidos em plantas prolíficas,
permitindo assegurar ao mesmo tempo ganhos genéticos diretos (nas populações “per
se”) e indiretos (nas populações em cruzamento). Esse método consiste em se cruzar
plantas S0 aos pares (plantas da população A com plantas da população B),
autofecundando simultaneamente os indivíduos cruzados. Tal método é baseado em
pares de plantas, o que torna necessário realizar anotações e a perda de uma espiga
autofecundada causa a perda do par do cruzamento, reduzindo a quantidade de material
avaliado.
De acordo com SANTOS et al. (2003), o método de melhoramento por seleção
recorrente recíproca com base em famílias de irmãos germanos parece ser uma boa
opção para a cultura do milho, pois ao final do processo, pode-se optar por utilizar
populações melhoradas para a obtenção de linhagens ou para a formação de um híbrido
intervarietal entre tais populações.
Geralmente na cultura do milho realiza-se a autofecundação da segunda espiga
de cada planta, sendo que a primeira espiga (superior) é polinizada com pólen de outra
planta. As sementes dessas espigas correspondem às progênies de irmãos germanos (A
x B), que são avaliadas em ensaios para identificar as combinações (progênies) mais
promissoras. Cada ciclo é completado quando as progênies S1 de cada população dos
melhores cruzamentos são recombinadas, produzindo assim as populações melhoradas
A1 e B1.

21
Segundo PATERNIANI & MIRANDA FILHO (1981), a seleção recorrente
recíproca apresenta a vantagem de possibilitar o desenvolvimento simultâneo de
híbridos de linhagens. Assim depois de avaliadas as progênies de irmãos germanos, as
sementes S1 das melhores progênies são semeadas, efetuando-se a autofecundação das
plantas envolvidas nos cruzamentos para produzir, após algumas gerações de
autofecundação, linhagens potenciais para híbridos de elevada produção.
PINTO (1995) afirma que o método é apropriado para produzir linhagens
visando a obtenção de híbridos, uma vez que, plantas da população A e B tornam-se
cada vez mais produtivas “per se” e em cruzamentos entre si. Ao mesmo tempo, os
cruzamentos de linhagens extraídas de A e B tornam-se cada vez mais heteróticos à
medida que avança o número de ciclos de seleção.
Na maioria dos trabalhos que utilizam a seleção recorrente recíproca, empregam-
se populações obtidas a partir de um grande número de parentais (sintéticos, variedades
e compostos). No entanto, a seleção recorrente recíproca tem apresentado eficiência em
trabalhos que utilizam populações oriundas de poucos parentais como aquelas derivadas
de híbridos simples comerciais (REIS, 2009). Segundo esse mesmo autor, essas
populações apresentam vantagens por apresentarem ampla adaptação e alto potencial
produtivo, uma vez que tais híbridos são recomendados somente se apresentarem alto
rendimento em vários ambientes. Outra vantagem da utilização desses híbridos
heteróticos é devido à formação de populações S0 contendo grande variabilidade
genética.
3 MATERIAL E MÉTODOS
3.1 Material vegetal e delineamento estatístico
Foi realizado um experimento em casa de vegetação no Centro Experimental
Central do Instituto Agronômico – IAC, localizado na Fazenda Santa Elisa (22° 53’S,
47° 04’W e altitude de 600 m) situado no município de Campinas, SP.
Foram utilizados quatro híbridos de milho contrastantes quanto à tolerância à
seca: DAS2B707 e DAS2B710, considerados tolerantes segundo informações da
empresa e resultados de experimentos de avaliação de cultivares e FT510 e AS1522,

22
considerados sensíveis pelas observações em experimentos de Safrinha no Estado de
São Paulo.
Os híbridos de milho, foram semeados diretamente no solo em casa de vegetação
(Tabela 1). O solo coletado em casa de vegetação, localizada na Fazenda Santa Elisa,
foi classificado como Latossolo Vermelho-Amarelo distrófico típico com textura
argilosa.
Tabela 1. Híbridos de milho utilizados em experimento de casa de vegetação para fins
de avaliação de parâmetros fisiológicos e biométricos sob déficit hídrico imposto em
diferentes estádios fenológicos da cultura. Campinas, 2011.
Híbridos Tipo Ciclo Porte Aparência/cor
dos grãos Finalidade Empresa
DAS2B707 simples precoce médio/
alto
semi-duro/
alaranjado grão
Dow
AgroSciences
DAS2B710 simples precoce baixo semi-dentado/
alaranjado grão
Dow
AgroSciences
FT510 simples superprecoce alto duro/alaranjado grão FT Sementes
AS1522 simples precoce alto semi-duro/
amarelado grão
Agroeste
Sementes
O delineamento empregado foi o de blocos casualizados, com parcelas
subdivididas e três repetições. As parcelas constituíram-se nos tratamentos hídricos
impostos (a) controle: plantas irrigadas durante todo o ciclo; (b) déficit hídrico apenas
no estádio vegetativo: quando as plantas apresentavam cinco folhas totalmente
expandidas (V5); (c) déficit hídrico apenas no estádio de florescimento: quando 50%
das plantas na parcela apresentavam estilo-estigma (R1) e (d) déficit hídrico apenas no
estádio de enchimento de grãos: quando as espigas apresentavam grãos com aspecto
leitoso (R3). As subparcelas foram constituídas pelos quatro híbridos. Cada parcela foi
constituída por uma linha de 3,5 m com espaçamento de 1,2 m entre linhas e 0,5 m entre
plantas e duas plantas por cova, totalizando 14 plantas na parcela (Figura 1).

23
Figura 1. Disposição do experimento realizado em casa de vegetação. Controle:
parcelas com irrigação durante todo o ciclo da cultura; vegetativo: parcelas
representando déficit hídrico somente no estádio vegetativo; florescimento: parcelas
representando déficit hídrico somente no estádio de florescimento e enchimento de
grãos: parcelas representando déficit hídrico somente no estádio de enchimento de
grãos. Foto tirada quando as plantas apresentavam-se no estádio V3. Campinas, 2011.
3.2 Condições de cultivo
Os tratos culturais foram realizados de acordo com as necessidades da cultura e
a irrigação fornecida por um sistema de gotejamento até o momento de imposição do
déficit hídrico bem como no período posterior ao estresse. O potencial matricial da água
no solo foi medido por tensiômetros de coluna de mercúrio, instalados a 45 cm de
profundidade em cada parcela. Foram fornecidas de duas a três irrigações semanais com
duração de dez minutos quando as plantas se encontravam no estádio vegetativo e de
vinte minutos no estádio de florescimento e de enchimento de grãos com vazão de 3,75
L h-1
. O fornecimento de água no tratamento irrigado foi realizado no momento em que
os tensiômetros indicavam potencial matricial do solo ao redor de -0,10 MPa.
controle vegetativo
floresc.
ench. grãos

24
Os períodos de déficit hídrico, a temperatura média dentro da casa de vegetação
e a média dos potenciais matriciais do solo em cada estádio fenológico estão
apresentados na Tabela 2. No momento da avaliação dos caracteres fisiológicos a
temperatura da casa de vegetação e do potencial matricial do solo indicavam valores de
28ºC e -0,80 MPa no estádio vegetativo, 32ºC e -1,10 MPa no florescimento e de 30°C e
-0,60 MPa na fase de enchimento de grãos. O dia de máximo déficit hídrico foi
determinado pelo aspecto visual das plantas, ou seja, quando as plantas apresentavam
sintomas de murcha acentuada no início do dia.
Vale ressaltar que as plantas irrigadas durante todo o ciclo (controle) foram
utilizadas para fins de comparação e desta forma, avaliadas concomitantemente com os
tratamentos sob déficit hídrico nos três estádios fenológicos da cultura. Posteriormente
ao déficit hídrico, a irrigação foi reestabelecida até o término do ciclo da cultura para
avaliações dos parâmetros biométricos.
Tabela 2. Período de déficit hídrico, temperatura média dentro da casa de vegetação e a
média do potencial matricial do solo do tratamento submetido ao déficit hídrico imposto
em três estádios fenológicos da cultura.
Estádio fenológico Período de déficit
hídrico
Temperatura
média
Potencial matricial
do solo (Ψs)*
Vegetativo 40 dias
(08/05 a 17/06/2011) 22,4ºC -0,69 Mpa
Florescimento 75 dias
(02/06 a 16/08/2011) 23,4ºC -0,75 Mpa
Enchimento de
grãos
52 dias
(11/07 a 01/09/2011) 24,8ºC -0,40 Mpa
(*) Média de três leituras dos tensiômetros de coluna de mercúrio instalados a 45 cm de profundidade no
solo em cada parcela durante o déficit hídrico.
3.3 Avaliações fisiológicas em casa de vegetação
No dia do máximo déficit hídrico, as medidas de parâmetros fisiológicos foram
realizadas em folhas completamente expandidas localizadas no terço superior da planta
quando essas estavam no estádio vegetativo, enquanto que, no estádio de florescimento
e no de enchimento de grãos, empregou-se a folha próxima à espiga principal. Foram
avaliados os seguintes parâmetros fisiológicos:

25
a) Potencial da água na folha (Ψw): medido com uma câmara de pressão modelo
3005 (SoilMoisture, EUA). As medidas foram realizadas na antemanhã (05:00 às
06:00h) em uma planta por repetição, totalizando três medidas de cada híbrido;
b) Trocas gasosas: avaliadas com um analisador de gases por radiação infravermelha
modelo Li-6400F (Licor, EUA). As variáveis estudadas foram: condutância
estomática (gs), concentração intercelular de CO2 (Ci), transpiração (E) e
assimilação de CO2 (A). Os dados da assimilação de CO2 (A) e da transpiração (E)
foram medidos ao longo do dia, às 08:30, 10:30, 13:30 e 16:00 horas. No dia de
máximo déficit hídrico nos três estádios fenológicos estudados, as quantidades de
CO2 assimilado e de vapor d’água transpirado entre 08:30 e 16:00 h foram
integradas. Optou-se por apresentar os valores de condutância estomática e
concentração intercelular de CO2 no horário de maior demanda atmosférica (13:30
h). As avaliações foram realizadas com concentração de CO2 no ar constante (380
μmol mol-1
) e radiação de 2100 μmol m-2
s-1
, controlados respectivamente pelo
misturador de gases e pela fonte de luz do Li-6400F. As avaliações foram
registradas quando o coeficiente de variação (CV) total foi inferior a 1%.
c) Medidas da emissão de fluorescência da clorofila a: realizadas com um fluorômetro
modulado (6400-40 LCF) integrado ao Li-6400F. As variáveis determinadas foram:
eficiência quântica potencial do fotossistema II (Fv/Fm), coeficientes de extinção
fotoquímica (qP) e não-fotoquímica (NPQ) da fluorescência e o transporte aparente
de elétrons (ETR) (ROHACEK, 2002). Os valores de Fv e Fm indicam,
respectivamente, a fluorescência variável e máxima determinada após 30 minutos
de adaptação ao escuro. Optou-se também por apresentar os valores dessas
variáveis fotoquímicas apenas no horário de maior demanda atmosférica;
d) Por meio da razão entre a assimilação diurna de CO2 (Ai) e transpiração diurna (Ei)
determinou-se a eficiência do uso da água (EUA);
e) Clorofila total (Chl): as avaliações do teor de clorofila a e b foram realizadas com o
equipamento ClorofiLOG (CFL1030-FALKER) em cinco plantas por repetição. A
média das cinco plantas avaliadas em cada parcela serviu para representar uma
repetição de cada tratamento, totalizando três medidas de cada híbrido. As
avaliações foram realizadas no momento de máximo déficit hídrico em cada
tratamento hídrico imposto nos três estádios fenológicos da cultura;
f) Teor de carboidratos nas folhas: Nos dias de máximo déficit hídrico, foram
coletadas amostras de folhas no horário das 08:30 h para realizar as quantificações

26
de carboidratos. As folhas coletadas em cada parcela foram secas em liofilizador
até obtenção de peso constante, sendo posteriormente maceradas e armazenadas até
o momento das análises. Os carboidratos mensurados foram: açúcares solúveis
(AS), sacarose (SAC), amido (AM) e carboidrato total não estrutural (CTNE).
Para a extração dos açúcares solúveis (AS) nas folhas foi utilizada uma solução
composta por metanol, clorofórmio e água (MCW) na proporção de 12:5:3 v/v
(BIELESKI & TURNER, 1966). Foram colocados aproximadamente 75 mg de matéria
seca e 3 mL de MCW em um tubo Falcon. Realizou-se agitação da solução
(aproximadamente 30 segundos em vórtex) e, posteriormente, as amostras foram
armazenadas por três dias sob refrigeração. Após, adicionou-se 1,2 mL de água e 1,8
mL de clorofórmio e, então, esta solução foi novamente armazenada sob refrigeração
por mais dois dias. Após esta etapa, o sobrenadante foi coletado e concentrado em
banho-maria (MA184, Marconi, Brasil) a 50 ºC por aproximadamente três horas e 30
minutos. O volume obtido foi mensurado e o extrato utilizado na determinação de AS e
SAC.
A fração AS foi determinada pelo método fenol-sulfúrico (DUBOIS et al.,
1956). Em 10 μL de amostra, foram adicionados 490 μL de água destilada, 500 μL de
fenol 5% e 2 mL de ácido sulfúrico p.a., agitando-se a solução em vórtex. Após o
resfriamento da solução, realizou-se a leitura da absorbância a 490 nm, em
espectrofotômetro modelo B342II (Micronal, Brasil), em duplicata. Os valores foram
transformados em teor de AS com o auxílio de uma reta-padrão obtida com
concentrações variadas de glicose (0, 5, 10, 20, 30, 40 e 50 μg).
A fração SAC foi determinada pelo método proposto por VAN HANDEL
(1968). Em tubos de ensaio, foram adicionados 15 μL de amostra, 485 μL de água e 500
μL de solução de hidróxido de potássio 30%. Os tubos foram vedados e incubados a
95ºC por 10 minutos em banho-maria. Posteriormente, foram adicionados 500 μL de
fenol 5% e 2 mL de ácido sulfúrico p.a. A solução foi agitada em vórtex e após o
resfriamento procedeu-se a leitura da absorbância a 490 nm, em espectrofotômetro, em
duplicata. Os valores foram transformados em teor de SAC com o auxílio de uma reta-
padrão obtida com concentrações variadas de sacarose (0, 5, 10, 20, 30, 40 e 50 μg).
Para determinação de amido (AM) utilizou-se o método enzimático proposto por
AMARAL et al. (2007). Amostras de 12 mg de matéria seca foram depositadas em
microtubos de 2 mL. Para obtenção do precipitado foram feitas quatro extrações com
500 μL de etanol 80% com incubação em banho-maria por 20 minutos a 80ºC. O

27
precipitado foi seco durante cinco dias a temperatura ambiente. Adicionou-se ao
precipitado 500 μL (110 U mL-1
) de α-amilase (EC 2.3.1.1) termoestável de Bacillus
licheniformis (cód. E-ANAAM, Megazyme, Irlanda), diluída em tampão MOPS 10 mM
e pH 6,5. As amostras foram incubadas a 75 ºC (em banho-maria) por 30 minutos. Este
passo foi repetido mais uma vez, totalizando 120 unidades da enzima. Em seguida
adicionou-se 500 μL (30 U mL-1
) de amiloglucosidase (EC 3.2.1.3) de Aspergillus niger
(cód. E-AMGPU, Megazyme, Irlanda) em tampão acetato de sódio 100 mM e pH 4,5.
As amostras foram incubadas a 50 ºC por 30 minutos. Este passo foi repetido mais uma
vez, totalizando 30 unidades da enzima, sendo em seguida acrescentados 100 μL de
ácido perclórico 0,8 M.
A quantificação de amido foi realizada em 50 μL da amostra adicionados a 750
μL de glicose PAP Liquiform (Labtest Diagnóstica S.A., Brasil), com posterior
incubação a 30 ºC por 15 minutos. A absorbância foi avaliada com um leitor de
microplacas modelo EL307C (Bio-Tek Instruments, EUA) a 490 nm, em triplicata. Os
valores foram transformados em teor de amido a partir da reta-padrão obtida com as
leituras de soluções contendo 0, 5, 10, 15, 20, 25 e 30 μg de glicose.
O conteúdo de carboidrato total não estrutural (CTNE) foi determinado
indiretamente pela soma do teor de açúcares solúveis totais com o amido
(CTNE=AS+AM).
3.4 Avaliações biométricas em casa de vegetação
Após 160 dias da semeadura, avaliaram-se os seguintes caracteres nas plantas de
cada parcela submetidas aos tratamentos hídricos em cada estádio fenológico avaliado:
a) Altura de planta (AP) – medida tomada do nível do solo até a inserção da folha
bandeira em cinco plantas de cada parcela, em centímetros (cm). A média das
cinco plantas avaliadas em cada parcela serviu para representar uma repetição de
cada tratamento, totalizando três medidas de cada híbrido;
b) Diâmetro do caule (DIAM) – medida realizada no meio do primeiro entrenó
acima do nível do solo em cinco plantas de cada parcela, em milímetros (mm).
A média das cinco plantas avaliadas em cada parcela serviu para representar
uma repetição de cada tratamento, totalizando três medidas de cada híbrido;

28
c) Massa de grãos (MG) - pesagem dos grãos resultantes da debulha das espigas de
sete plantas de cada parcela, em g planta-1
. Os dados foram corrigidos para 14 %
de umidade;
d) Massa seca total (MST) – pesagem das frações folha, caule, espigas e produção
de grãos de sete plantas de cada parcela, em g planta-1
. Essas amostras foram
coletadas e desidratadas em estufa de circulação forçada a 60˚C até a obtenção
da massa seca constante.
3.5 Análise dos dados
Os dados fisiológicos e biométricos foram submetidos à análise de variância e
os valores médios foram comparados pelo teste de Scott-Knott a 5% de probabilidade,
usando o programa estatístico SISVAR (FERREIRA, 2000).
3.6 Melhoramento para tolerância à seca
3.6.1 Obtenção da população tolerante ao déficit hídrico
A obtenção da população tolerante ao déficit hídrico foi realizada pelo “método
Irlandês” empregando-se os híbridos comerciais DAS2B707, DAS2B710 e DAS2B587.
Esses híbridos foram semeados de maneira que cada fileira de plantas empregadas como
parental feminino fosse representada pelos híbridos, enquanto que as plantas usadas
como parental masculino fossem formadas por linhas com as misturas dos três híbridos.
No período de florescimento da cultura foi realizado o despendoamento dos
híbridos que constituíam o parental feminino, deixando essas estruturas florais apenas
nas fileiras onde fora realizada a mistura dos três híbridos, ou seja, dos parentais
masculinos.
A semeadura do campo ocorreu em setembro de 2009, seguida de adubações e
tratos culturais conforme a exigência da cultura. Foram semeadas seis linhas de
parentais femininos e quatro linhas de parentais masculinos, ambas contendo 20 m de
comprimento, espaçamento de 0,80 cm entre linhas e 0,20 cm entre plantas.
Após a polinização livre, as sementes foram colhidas, misturadas e semeadas em
março de 2010. Nesse lote foram semeadas 20 linhas com aproximadamente 18 m cada,
com espaçamento de 0,80 cm entre linhas e de 0,20 cm entre plantas. Após o
intercruzamento ao acaso das plantas, o que permitiu o equilíbrio de Hardy-Weinberg,

29
desconsiderando os genes ligados, as sementes foram colhidas e misturadas para
compor a população tolerante ao défict hídrico (POP TOL).
3.6.2 Obtenção da população sensível ao déficit hídrico
Para a obtenção da população sensível ao déficit hídrico foram empregados os
híbridos comerciais FT510 e AS1522. Tais híbridos foram semeados em setembro de
2009, constituindo em oito linhas de 18 m cada, sendo quatro linhas do híbrido FT510 e
quatro linhas do híbrido AS1522, semeadas alternadamente.
Tais híbridos foram intercruzados ao acaso e as sementes obtidas foram
coletadas, misturadas e semeadas no mês de fevereiro de 2010 em 10 linhas de 20 m,
com espaçamento entre linhas de 0,80 cm e de 0,20 cm entre plantas, em um campo de
polinização aberta para que a população sensível (POP SENS) entrasse em equilíbrio,
desconsiderando os genes ligados. As adubações e tratos culturais foram realizados
conforme a necessidade da cultura do milho.
3.6.3 Obtenção das progênies de irmãos germanos e S1
A população tolerante ao déficit hídrico POP TOL e a população sensível ao
déficit hídrico POP SENS foram levadas a campo no mês de setembro de 2010 para,
posteriormente, serem realizados os cruzamentos direcionados entre elas, visando a
retirada de progênies. As plantas prolíficas de ambas as populações tiveram suas espigas
superiores cruzadas aos pares com plantas da população recíproca, gerando uma
progênie de irmãos germanos interpopulacional em cada planta. As espigas inferiores
foram autofecundadas, formando progênies S1. As sementes das progênies de irmãos
germanos foram utilizadas para avaliação e as sementes das progênies S1 foram
guardadas para compor a unidade de recombinação.
3.6.4 Avaliação em campo das progênies de irmãos germanos em Campinas e
Mococa
Para obtenção dos resultados de melhoramento genético visando à tolerância ao
déficit hídrico foram realizados experimentos de campo no Centro Experimental Central
do Instituto Agronômico – IAC em Campinas (22° 53’S, 47° 04’W e altitude de 600 m)
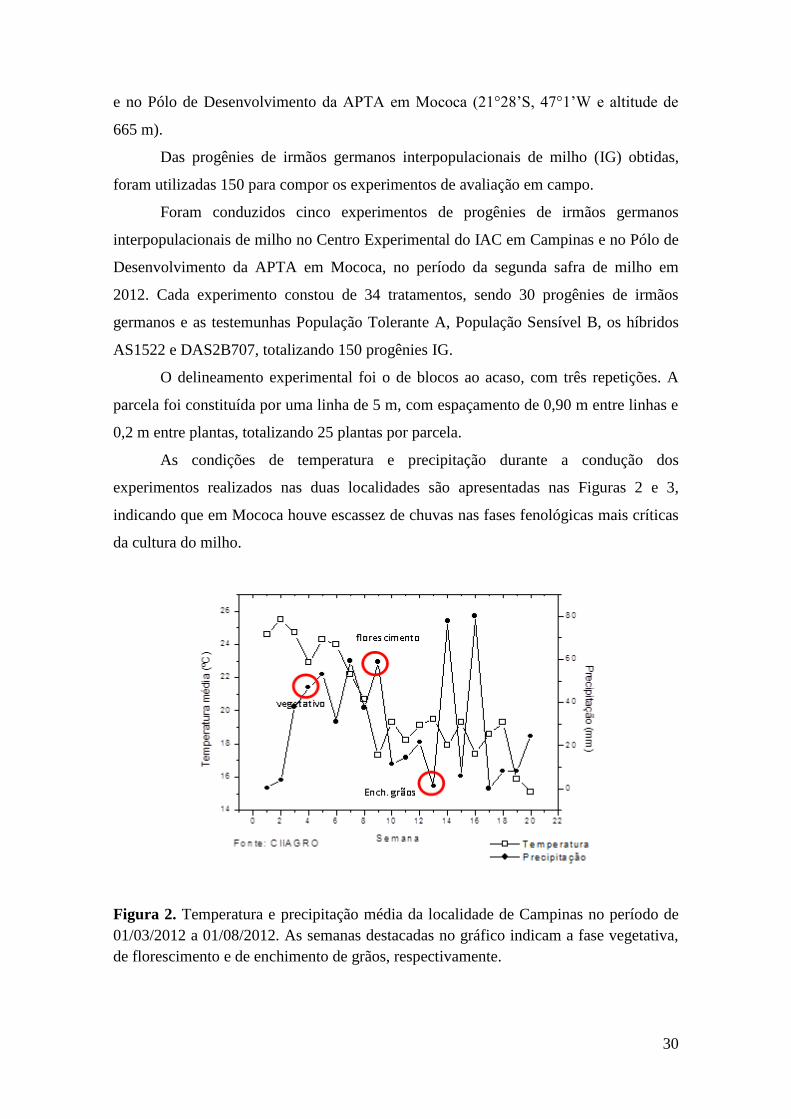
30
e no Pólo de Desenvolvimento da APTA em Mococa (21°28’S, 47°1’W e altitude de
665 m).
Das progênies de irmãos germanos interpopulacionais de milho (IG) obtidas,
foram utilizadas 150 para compor os experimentos de avaliação em campo.
Foram conduzidos cinco experimentos de progênies de irmãos germanos
interpopulacionais de milho no Centro Experimental do IAC em Campinas e no Pólo de
Desenvolvimento da APTA em Mococa, no período da segunda safra de milho em
2012. Cada experimento constou de 34 tratamentos, sendo 30 progênies de irmãos
germanos e as testemunhas População Tolerante A, População Sensível B, os híbridos
AS1522 e DAS2B707, totalizando 150 progênies IG.
O delineamento experimental foi o de blocos ao acaso, com três repetições. A
parcela foi constituída por uma linha de 5 m, com espaçamento de 0,90 m entre linhas e
0,2 m entre plantas, totalizando 25 plantas por parcela.
As condições de temperatura e precipitação durante a condução dos
experimentos realizados nas duas localidades são apresentadas nas Figuras 2 e 3,
indicando que em Mococa houve escassez de chuvas nas fases fenológicas mais críticas
da cultura do milho.
Figura 2. Temperatura e precipitação média da localidade de Campinas no período de
01/03/2012 a 01/08/2012. As semanas destacadas no gráfico indicam a fase vegetativa,
de florescimento e de enchimento de grãos, respectivamente.
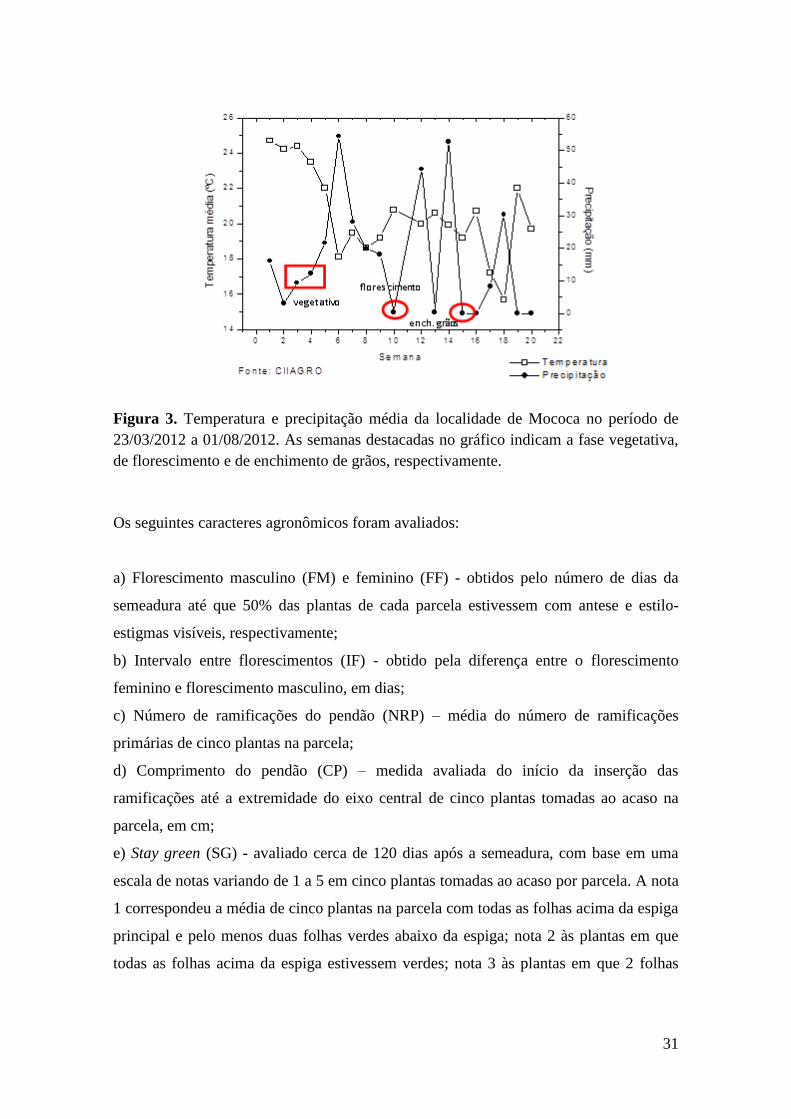
31
Figura 3. Temperatura e precipitação média da localidade de Mococa no período de
23/03/2012 a 01/08/2012. As semanas destacadas no gráfico indicam a fase vegetativa,
de florescimento e de enchimento de grãos, respectivamente.
Os seguintes caracteres agronômicos foram avaliados:
a) Florescimento masculino (FM) e feminino (FF) - obtidos pelo número de dias da
semeadura até que 50% das plantas de cada parcela estivessem com antese e estilo-
estigmas visíveis, respectivamente;
b) Intervalo entre florescimentos (IF) - obtido pela diferença entre o florescimento
feminino e florescimento masculino, em dias;
c) Número de ramificações do pendão (NRP) – média do número de ramificações
primárias de cinco plantas na parcela;
d) Comprimento do pendão (CP) – medida avaliada do início da inserção das
ramificações até a extremidade do eixo central de cinco plantas tomadas ao acaso na
parcela, em cm;
e) Stay green (SG) - avaliado cerca de 120 dias após a semeadura, com base em uma
escala de notas variando de 1 a 5 em cinco plantas tomadas ao acaso por parcela. A nota
1 correspondeu a média de cinco plantas na parcela com todas as folhas acima da espiga
principal e pelo menos duas folhas verdes abaixo da espiga; nota 2 às plantas em que
todas as folhas acima da espiga estivessem verdes; nota 3 às plantas em que 2 folhas

32
acima da espiga estivessem secas e as demais verdes; nota 4 às plantas em que 2 folhas
no ápice da planta estivessem verdes e 5 às plantas com todas as folhas secas;
f) Altura da planta (AP) e da espiga (AE) - obtidas pela medida tomada do nível do solo
até a inserção da folha bandeira e da espiga principal em cinco plantas na parcela,
respectivamente, em cm;
g) Componentes de produção - comprimento da espiga (CE), diâmetro da espiga (DE) e
número de fileiras de grãos na espiga (NFG) avaliados como média de cinco espigas,
tomadas aleatoriamente na parcela;
h) Prolificidade (PROL) - medida pela relação do número de espigas com número de
plantas da parcela, corrigido para estande ideal do experimento;
i) Massa de grãos (MG) - medido em gramas por planta (g/planta), ajustado para
umidade de 14% e estande ideal de 25 plantas na parcela pelo método proposto por
VENCOVSKY & CRUZ (1991).
j) Massa de grãos porcento (MG%) – porcentagem de peso de grãos de cada genótipo
em relação à testemunha tolerante à seca, calculado com base nas médias dos
tratamentos.
3.6.5 Análise agrupada e obtenção de parâmetros genéticos
Pelos resultados das análises individuais dos experimentos referentes às
progênies de irmãos germanos conduzidas em cinco experimentos em Campinas e em
Mococa, realizou-se a análise agrupada pela somatória das Somas de Quadrados e dos
Graus de Liberdade de cada experimento.
A análise de variância conjunta foi realizada conforme o delineamento em
blocos as acaso segundo o modelo estatístico:
Yijk = µ + (b/l)kj + pi +lj + plij + ɛijk
Onde:
Yijk = observação da i-ésima progênie no k-ésimo bloco e j-ésimo local;
µ = média geral do ensaio;
(b/l)kj = efeito do bloco k dentro do local j;
pi = efeito da progênie i;
lj = efeito do local j;
plij = efeito da interação entre a progênie i e o local j;

33
ɛijk = erro aleatório associado à observação ijk.
Posteriormente, pelas Esperanças dos Quadrados médios, foram estimados os
componentes de variância genética entre progênies ( 2
pσ ) e variância ambiental ( 2
Eσ )
para todos os caracteres avaliados, na análise conjunta.
No Quadro 1 é apresentado o esquema da análise de variância conjunta, com as
respectivas esperanças de quadrados médios, sendo que, com exceção de locais, as
demais fontes de variação foram consideradas aleatórias.
Quadro 1. Análise de variância conjunta e esperança dos quadrados médios,
considerando o local como sendo de efeito fixo e os demais efeitos aleatórios.
FV GL QM E (QM) F
Blocos/Ambientes (r-1)a QMB 2
E + p 2
bσ
Ambientes (A) a-1 QMA 2
E + rℓ 2
paσ + p 2
bσ + prϕa QMPAQMB
QMRQMA
Progênies (P) p-1 QMP 2
E + ar 2
pσ QMR
QMP
P x A (a-1)(p-1) QMPA 2
E + rℓ 2
paσ
QMR
QMPA
Resíduo (p-1)(r-1)a QMR 2
E
ℓ=a/(a-1)
Foram estimados os coeficientes de herdabilidade em nível de média de
progênies ( 2h ), com base na análise conjunta, pela seguinte fórmula:
2h = )/( 22
2
p
arEp
Os ganhos genéticos com a seleção entre progênies (Gs) foram calculados por:
Gs = K x Fσ x 2h , ou seja, Gs = F
K
2
p. em que k é função da intensidade de seleção
aplicada entre progênies (intensidade de seleção estandardizada) e Fσ é o desvio padrão
fenotípico em nível de médias. A intensidade de seleção foi de 20% (k = 1,4).
O índice de variação (b) foi determinado pela relação CVg/CV%, em que CVg é
o coeficiente de variação genético e CV% é o coeficiente de variação ambiental.

34
Realizou-se a correlação fenotípica de Pearson entre os caracteres por meio do
programa estatístico GENES (CRUZ, 2001).
3.6.6 Avaliação e discriminação das melhores e piores progênies de irmãos
germanos quanto à tolerância à seca em casa de vegetação
De acordo com os resultados obtidos de massa de grãos dos experimentos de
campo com as progênies de irmãos germanos interpopulacionais realizados em
Campinas e Mococa, foram selecionados dois pares de progênies contrastantes quanto a
tolerância à seca. As progênies 58Bx65A e 121Bx116A foram consideradas sensíveis à
seca, enquanto as progênies 9Bx7A e 156Ax150B foram selecionadas como tolerantes
ao déficit hídrico. Essas quatro progênies foram semeadas em casa de vegetação no dia
27 de novembro de 2012 no Centro Experimental Central do Instituto Agronômico –
IAC na Fazenda Santa Elisa. A População Tolerante (POP TOL) e a População Sensível
(POP SENS) foram utilizadas como testemunhas.
Empregou-se o delineamento de blocos casualizados, com parcelas subdivididas
e duas repetições. As parcelas constituíram-se nos tratamentos hídricos impostos (a)
controle: plantas irrigadas durante todo o ciclo; (b) déficit hídrico apenas no estádio
vegetativo: quando as plantas apresentavam cinco folhas totalmente expandidas (V5) e
(c) déficit hídrico apenas no estádio de florescimento: quando 50% das plantas na
parcela apresentavam estilo-estigma (R1). As subparcelas foram formadas pelos seis
genótipos empregados. Cada parcela foi constituída por uma linha de 3,5 m, com
espaçamento de 1,2 m entre linhas e 0,5 m entre plantas e duas plantas por cova,
totalizando 14 plantas na parcela.
O potencial matricial da água no solo foi medido por tensiômetros de coluna de
mercúrio, instalados a 45 cm de profundidade em cada parcela. O fornecimento de água
no tratamento irrigado foi realizado no momento em que os tensiômetros indicavam
potencial matricial do solo ao redor de -0,10 MPa.
Os períodos de déficit hídrico, temperatura média dentro da casa de vegetação e
os potenciais matriciais do solo em cada estádio fenológico estão apresentados na
Tabela 3. No momento da avaliação a temperatura máxima e o potencial matricial do
solo indicavam valores de 35°C e -1,00 MPa no estádio vegetativo e de 37,5°C e -1,20
MPa na fase de florescimento. O dia de máximo déficit hídrico foi determinado pelo

35
aspecto visual das plantas, ou seja, quando as plantas apresentavam sintomas de murcha
acentuada no início do dia.
Tabela 3. Período de déficit hídrico, temperatura média dentro da casa de vegetação e
potencial matricial do solo do tratamento submetido ao déficit hídrico imposto em dois
estádios fenológicos da cultura.
Estádio fenológico Período de déficit
hídrico
Temperatura
média
Potencial matricial
do solo (Ψs)*
Vegetativo 25 dias
(27/12 a 22/01/2013) 32,50 ºC -0,70 Mpa
Florescimento 37 dias
(20/01 a 26/02/2013) 35,7 ºC -0,75 Mpa
(*) Média de três leituras dos tensiômetros de coluna de mercúrio instalados a 45 cm de profundidade no
solo em cada parcela durante o déficit hídrico.
No dia do máximo déficit hídrico foram realizadas as medidas dos parâmetros
fisiológicos: potencial da água na folha (Ψw), condutância estomática (gs), transpiração
(E), assimilação de CO2 (A), eficiência quântica potencial do fotossistema II (Fv/Fm),
coeficientes de extinção fotoquímica (qP) e não-fotoquímica (NPQ) da fluorescência e
da taxa aparente do transporte de elétrons (ETR).
Após 140 dias da semeadura, avaliaram-se os seguintes caracteres biométricos:
altura da planta (AP) e diâmetro do colmo (DIAM) avaliadas em cinco plantas em cada
parcela, enquanto que para a massa de grãos (MG) e massa seca total (MST) foram
avaliadas sete plantas em cada parcela.
Os dados das variáveis fisiológicas e biométricas foram submetidos à análise de
variância e os valores médios foram comparados pelo teste de Scott-Knott a 5% de
probabilidade.

36
4 RESULTADOS E DISCUSSÃO
4.1 RESULTADOS: Parâmetros fisiológicos
4.1.1 Déficit hídrico no estádio vegetativo
Em medidas realizadas na antemanhã, verificou-se redução significativa do
potencial da água na folha (Ψw) dos híbridos devido ao déficit hídrico. Houve diferença
significativa (P<0,05) entre híbridos após o período de 40 dias sem irrigação no estádio
vegetativo. O híbrido DAS2B707 se destacou por apresentar maior Ψw (-0,30 MPa) em
relação aos demais híbridos avaliados sob déficit hídrico. O DAS2B710 foi o híbrido
com maior redução de Ψw (-0,99 MPa) devido à suspensão da rega. Não foi possível
verificar diferença entre os híbridos em plantas sob condição irrigada, verificando que
todos os híbridos apresentaram valores de Ψw próximos de zero, o que indica uma
adequada hidratação nessas plantas no tratamento irrigado (Figura 4).
Figura 4. Potencial da água na folha (Ψw) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5). Avaliações
realizadas às 05h30 após 40 dias sob déficit hídrico. Cada histograma representa o valor
médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças estatísticas
(P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras
maiúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos.
2B707 2B710 FT510 AS1522
-1,2
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
irrigado
déficit hídrico
bBbB
cB
aB
aAaAaA
aA
Po
ten
cia
l d
a á
gu
a n
a f
olh
a (
MP
a)
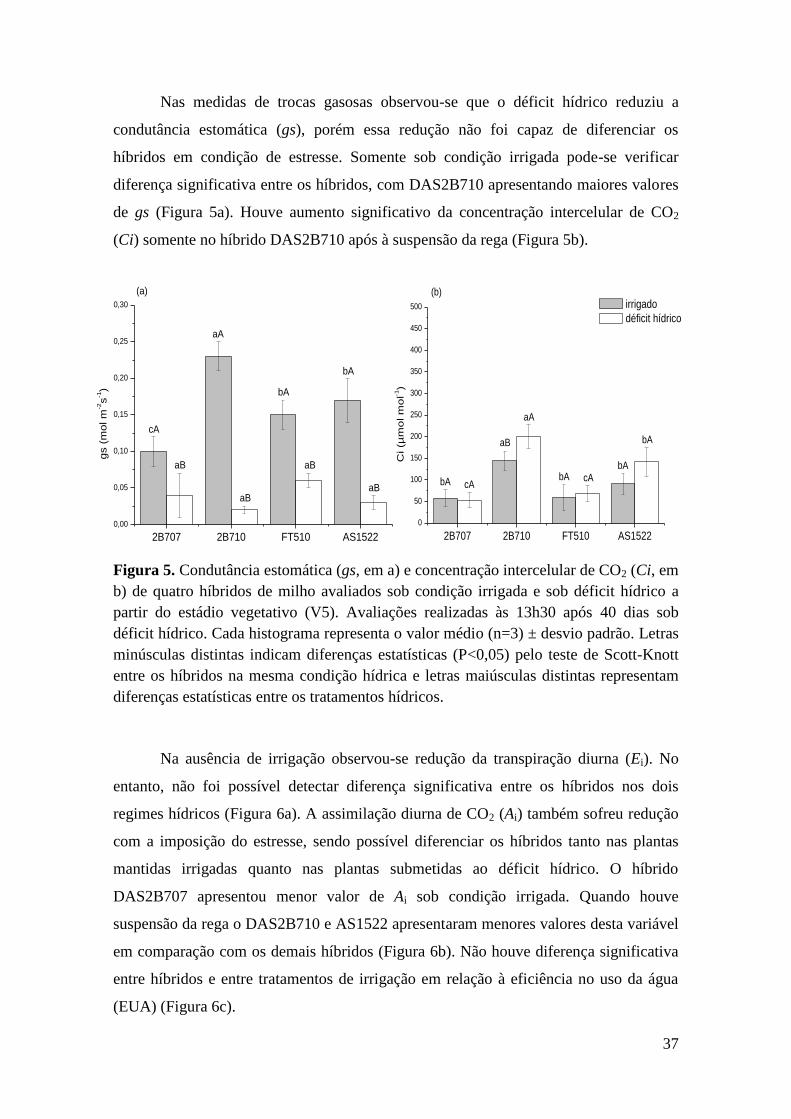
37
Nas medidas de trocas gasosas observou-se que o déficit hídrico reduziu a
condutância estomática (gs), porém essa redução não foi capaz de diferenciar os
híbridos em condição de estresse. Somente sob condição irrigada pode-se verificar
diferença significativa entre os híbridos, com DAS2B710 apresentando maiores valores
de gs (Figura 5a). Houve aumento significativo da concentração intercelular de CO2
(Ci) somente no híbrido DAS2B710 após à suspensão da rega (Figura 5b).
Figura 5. Condutância estomática (gs, em a) e concentração intercelular de CO2 (Ci, em
b) de quatro híbridos de milho avaliados sob condição irrigada e sob déficit hídrico a
partir do estádio vegetativo (V5). Avaliações realizadas às 13h30 após 40 dias sob
déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras
minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott
entre os híbridos na mesma condição hídrica e letras maiúsculas distintas representam
diferenças estatísticas entre os tratamentos hídricos.
Na ausência de irrigação observou-se redução da transpiração diurna (Ei). No
entanto, não foi possível detectar diferença significativa entre os híbridos nos dois
regimes hídricos (Figura 6a). A assimilação diurna de CO2 (Ai) também sofreu redução
com a imposição do estresse, sendo possível diferenciar os híbridos tanto nas plantas
mantidas irrigadas quanto nas plantas submetidas ao déficit hídrico. O híbrido
DAS2B707 apresentou menor valor de Ai sob condição irrigada. Quando houve
suspensão da rega o DAS2B710 e AS1522 apresentaram menores valores desta variável
em comparação com os demais híbridos (Figura 6b). Não houve diferença significativa
entre híbridos e entre tratamentos de irrigação em relação à eficiência no uso da água
(EUA) (Figura 6c).
2B707 2B710 FT510 AS1522
0
50
100
150
200
250
300
350
400
450
500 irrigado
déficit hídrico
(b)
bA
cA
aA
cA
bA
aB
bAbA
Ci (µ
mo
l m
ol-1
)
2B707 2B710 FT510 AS1522
0,00
0,05
0,10
0,15
0,20
0,25
0,30
(a)
aB
aB
aB
aB
cA
bA
bA
aA
gs (
mo
l m
-2s
-1)

38
Os teores de clorofila total (Chl) dos quatro híbridos foram influenciados pelo
déficit hídrico, apresentando reduções de 16%, 21%, 16% e 24% para DAS2B707,
DAS2B710, FT510 e AS1522, respectivamente (Tabela 4).
Quanto à atividade fotoquímica, não foram observados comportamentos
diferenciados entre híbridos e entre regimes hídricos na eficiência quântica potencial do
fotossistema II (Fv/Fm). No entanto, pode-se constatar diferenças entre os híbridos para
os coeficientes de extinção fotoquímica (qP) e não fotoquímica (NPQ) da fluorescência
devido ao déficit hídrico. O híbrido DAS2B707 apresentou maior valor de qP em
comparação com os demais híbridos avaliados sob condição de déficit hídrico. Este
mesmo híbrido e o AS1522 obtiveram menores valores de NPQ após período de
deficiência hídrica. O transporte aparente de elétrons (ETR) também foi influenciado
pelo déficit hídrico. O DAS2B707 apresentou maior valor de ETR no tratamento sob
déficit hídrico e menor valor quando submetido a condição irrigada (Tabela 4).
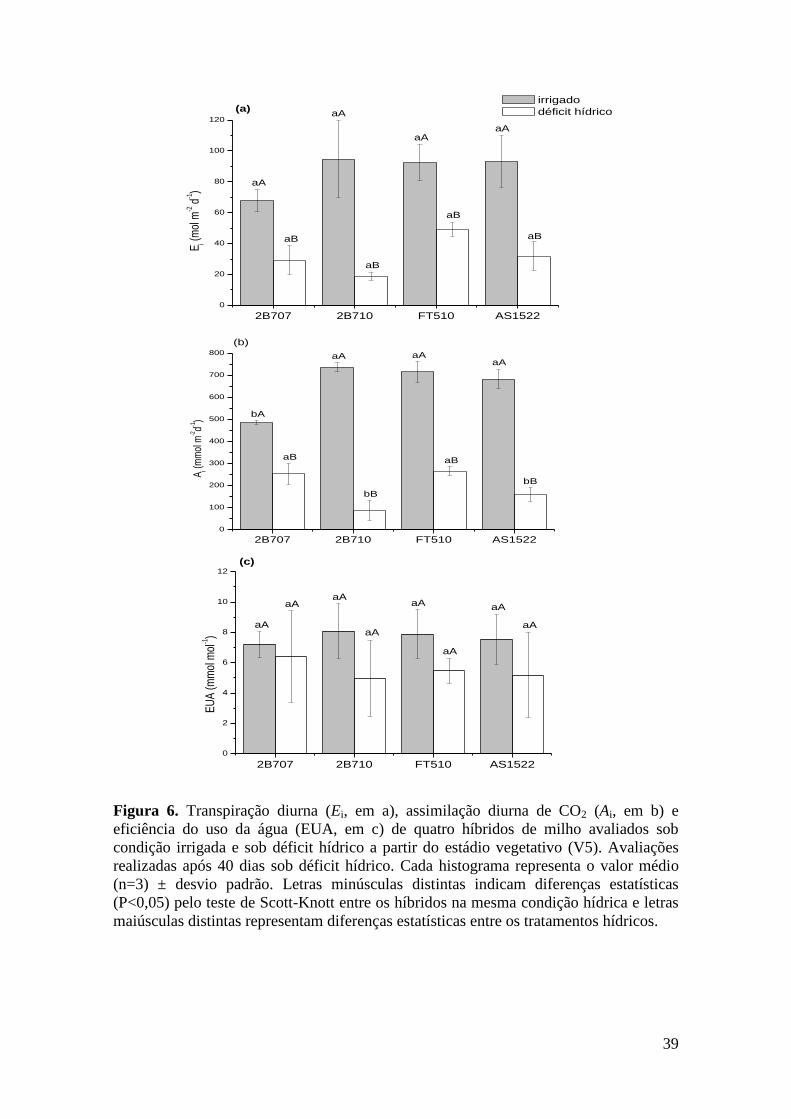
39
Figura 6. Transpiração diurna (Ei, em a), assimilação diurna de CO2 (Ai, em b) e
eficiência do uso da água (EUA, em c) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5). Avaliações
realizadas após 40 dias sob déficit hídrico. Cada histograma representa o valor médio
(n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças estatísticas
(P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras
maiúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos.
2B707 2B710 FT510 AS1522
0
20
40
60
80
100
120
irrigado
déficit hídrico(a)
aB
aB
aB
aB
aAaA
aA
aA
Ei (m
ol m
-2 d
-1)
2B707 2B710 FT510 AS1522
0
2
4
6
8
10
12
(c)
aA
aA
aA
aA aAaA
aA
aA
EU
A (
mm
ol m
ol-1)
2B707 2B710 FT510 AS1522
0
100
200
300
400
500
600
700
800
(b)
bB
bB
aB aB
bA
aAaAaA
Ai (m
mol
m-2d-1
)
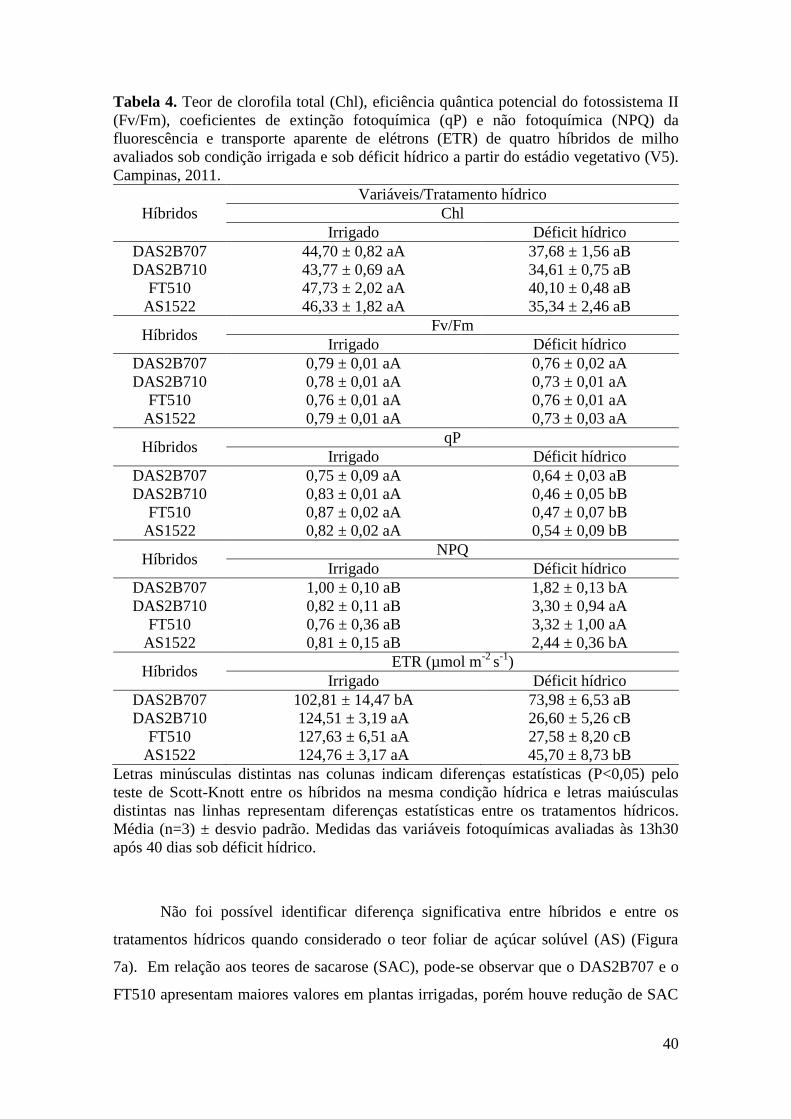
40
Tabela 4. Teor de clorofila total (Chl), eficiência quântica potencial do fotossistema II
(Fv/Fm), coeficientes de extinção fotoquímica (qP) e não fotoquímica (NPQ) da
fluorescência e transporte aparente de elétrons (ETR) de quatro híbridos de milho
avaliados sob condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5).
Campinas, 2011.
Híbridos
Variáveis/Tratamento hídrico
Chl
Irrigado Déficit hídrico
DAS2B707 44,70 ± 0,82 aA 37,68 ± 1,56 aB
DAS2B710 43,77 ± 0,69 aA 34,61 ± 0,75 aB
FT510 47,73 ± 2,02 aA 40,10 ± 0,48 aB
AS1522 46,33 ± 1,82 aA 35,34 ± 2,46 aB
Híbridos Fv/Fm
Irrigado Déficit hídrico
DAS2B707 0,79 ± 0,01 aA 0,76 ± 0,02 aA
DAS2B710 0,78 ± 0,01 aA 0,73 ± 0,01 aA
FT510 0,76 ± 0,01 aA 0,76 ± 0,01 aA
AS1522 0,79 ± 0,01 aA 0,73 ± 0,03 aA
Híbridos qP
Irrigado Déficit hídrico
DAS2B707 0,75 ± 0,09 aA 0,64 ± 0,03 aB
DAS2B710 0,83 ± 0,01 aA 0,46 ± 0,05 bB
FT510 0,87 ± 0,02 aA 0,47 ± 0,07 bB
AS1522 0,82 ± 0,02 aA 0,54 ± 0,09 bB
Híbridos NPQ
Irrigado Déficit hídrico
DAS2B707 1,00 ± 0,10 aB 1,82 ± 0,13 bA
DAS2B710 0,82 ± 0,11 aB 3,30 ± 0,94 aA
FT510 0,76 ± 0,36 aB 3,32 ± 1,00 aA
AS1522 0,81 ± 0,15 aB 2,44 ± 0,36 bA
Híbridos ETR (µmol m
-2 s
-1)
Irrigado Déficit hídrico
DAS2B707 102,81 ± 14,47 bA 73,98 ± 6,53 aB
DAS2B710 124,51 ± 3,19 aA 26,60 ± 5,26 cB
FT510 127,63 ± 6,51 aA 27,58 ± 8,20 cB
AS1522 124,76 ± 3,17 aA 45,70 ± 8,73 bB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas das variáveis fotoquímicas avaliadas às 13h30
após 40 dias sob déficit hídrico.
Não foi possível identificar diferença significativa entre híbridos e entre os
tratamentos hídricos quando considerado o teor foliar de açúcar solúvel (AS) (Figura
7a). Em relação aos teores de sacarose (SAC), pode-se observar que o DAS2B707 e o
FT510 apresentam maiores valores em plantas irrigadas, porém houve redução de SAC
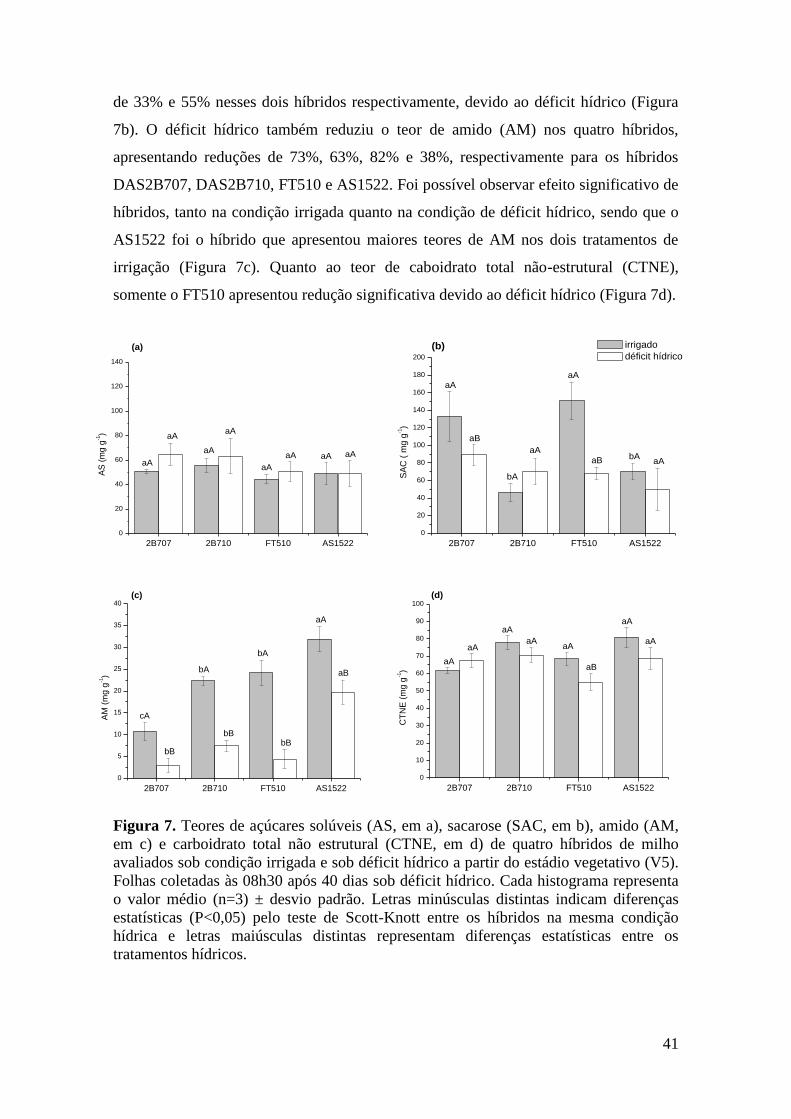
41
de 33% e 55% nesses dois híbridos respectivamente, devido ao déficit hídrico (Figura
7b). O déficit hídrico também reduziu o teor de amido (AM) nos quatro híbridos,
apresentando reduções de 73%, 63%, 82% e 38%, respectivamente para os híbridos
DAS2B707, DAS2B710, FT510 e AS1522. Foi possível observar efeito significativo de
híbridos, tanto na condição irrigada quanto na condição de déficit hídrico, sendo que o
AS1522 foi o híbrido que apresentou maiores teores de AM nos dois tratamentos de
irrigação (Figura 7c). Quanto ao teor de caboidrato total não-estrutural (CTNE),
somente o FT510 apresentou redução significativa devido ao déficit hídrico (Figura 7d).
Figura 7. Teores de açúcares solúveis (AS, em a), sacarose (SAC, em b), amido (AM,
em c) e carboidrato total não estrutural (CTNE, em d) de quatro híbridos de milho
avaliados sob condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5).
Folhas coletadas às 08h30 após 40 dias sob déficit hídrico. Cada histograma representa
o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças
estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição
hídrica e letras maiúsculas distintas representam diferenças estatísticas entre os
tratamentos hídricos.
2B707 2B710 FT510 AS1522
0
20
40
60
80
100
120
140
(a)
aAaA
aAaA
aA
aA
aA
aA
AS
(m
g g
-1)
2B707 2B710 FT510 AS1522
0
20
40
60
80
100
120
140
160
180
200
irrigado
déficit hídrico(b)
aAaBaA
aB
bA
bA
aAaA
SA
C (
mg
g-1)
2B707 2B710 FT510 AS1522
0
5
10
15
20
25
30
35
40(c)
aB
bBbB
bB
aA
bA
bA
cAAM
(m
g g
-1)
2B707 2B710 FT510 AS1522
0
10
20
30
40
50
60
70
80
90
100(d)
aA
aB
aAaA
aA
aA
aA
aA
CT
NE
(m
g g
-1)

42
Na Tabela 5 são apresentados os resultados da altura da planta (AP), diâmetro do
colmo (DIAM), massa de grãos (MG) e massa seca total (MST) referente aos híbridos
avaliados em condição irrigada e sob défcit hídrico. O déficit hídrico causou uma
redução da AP de 47% no híbrido DAS2B707, 41% no DAS2B710, 26% no FT510 e
27% no AS1522 em comparação ao tratamento irrigado. Houve diferença significativa
entre híbridos apenas na condição de estresse, indicando que os híbridos DAS2B707 e
DAS2B710 apresentaram menores AP sob suspensão da rega.
O caráter DIAM também foi afetado pelo estresse, exceto no híbrido DAS2B710
que não diferiu estatisticamente do tratamento irrigado. Houve diferença entre híbridos
nos dois tratamentos hídricos, com FT510 apresentando maior DIAM tanto no
tratamento irrigado quanto no tratamento submetido ao déficit hídrico.
Verificou-se alteração da massa de grãos (MG) quando houve suspensão da
rega. Reduções de magnitude de 18%, 42%, 51% e de 40% foram encontradas para os
híbridos DAS2B707, DAS2B710, FT510 e AS1522, respectivamente. No entanto, o
híbrido DAS2B707 não apresentou redução significativa de MG devido ao déficit
hídrico, indicando estabilidade de produção quando submetido à seca no estádio
vegetativo. Esse mesmo híbrido apresentou melhor desempenho produtivo em relação
aos demais híbridos avaliados sob condição de déficit hídrico.
O período de déficit hídrico também reduziu a massa seca total (MST) dos
quatro híbridos (Tabela 5), apresentando reduções de 33%, 48%, 50% e 41% para os
híbridos DAS2B707, DAS2B710, FT510 e AS1522, respectivamente. Em cada regime
hídrico, não foram observadas diferenças significativas entre os híbridos.
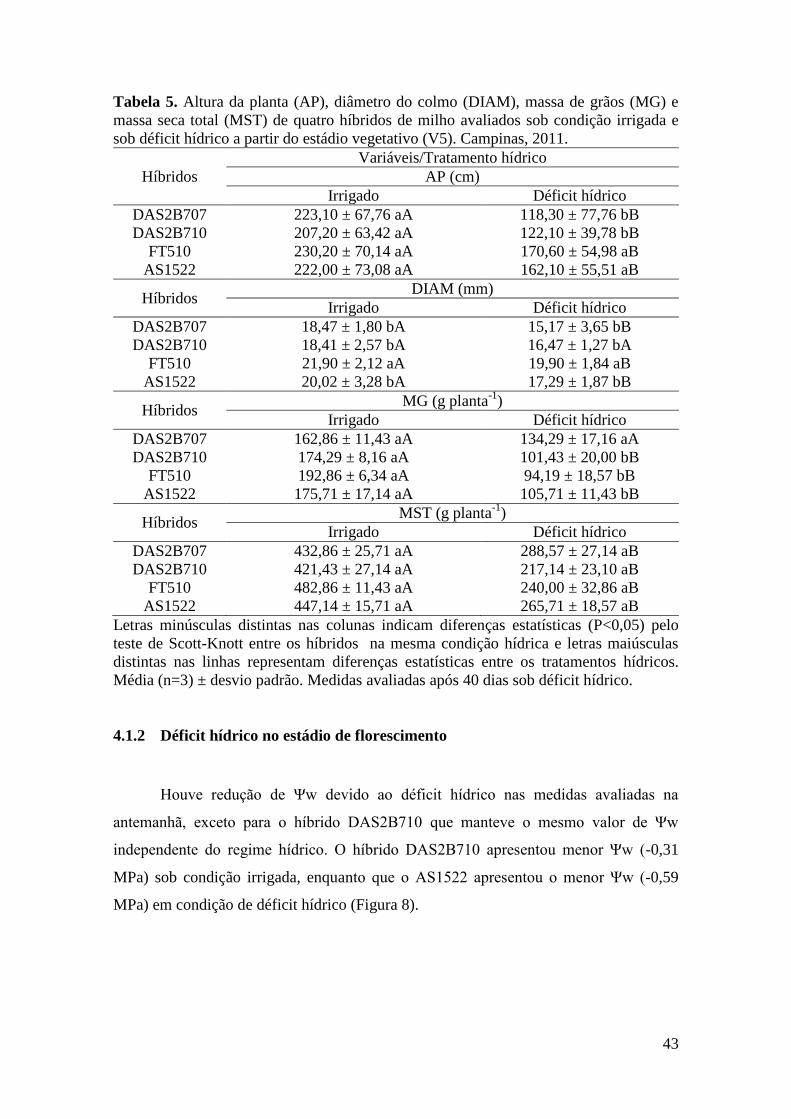
43
Tabela 5. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG) e
massa seca total (MST) de quatro híbridos de milho avaliados sob condição irrigada e
sob déficit hídrico a partir do estádio vegetativo (V5). Campinas, 2011.
Híbridos
Variáveis/Tratamento hídrico
AP (cm)
Irrigado Déficit hídrico
DAS2B707 223,10 ± 67,76 aA 118,30 ± 77,76 bB
DAS2B710 207,20 ± 63,42 aA 122,10 ± 39,78 bB
FT510 230,20 ± 70,14 aA 170,60 ± 54,98 aB
AS1522 222,00 ± 73,08 aA 162,10 ± 55,51 aB
Híbridos DIAM (mm)
Irrigado Déficit hídrico
DAS2B707 18,47 ± 1,80 bA 15,17 ± 3,65 bB
DAS2B710 18,41 ± 2,57 bA 16,47 ± 1,27 bA
FT510 21,90 ± 2,12 aA 19,90 ± 1,84 aB
AS1522 20,02 ± 3,28 bA 17,29 ± 1,87 bB
Híbridos MG (g planta
-1)
Irrigado Déficit hídrico
DAS2B707 162,86 ± 11,43 aA 134,29 ± 17,16 aA
DAS2B710 174,29 ± 8,16 aA 101,43 ± 20,00 bB
FT510 192,86 ± 6,34 aA 94,19 ± 18,57 bB
AS1522 175,71 ± 17,14 aA 105,71 ± 11,43 bB
Híbridos MST (g planta
-1)
Irrigado Déficit hídrico
DAS2B707 432,86 ± 25,71 aA 288,57 ± 27,14 aB
DAS2B710 421,43 ± 27,14 aA 217,14 ± 23,10 aB
FT510 482,86 ± 11,43 aA 240,00 ± 32,86 aB
AS1522 447,14 ± 15,71 aA 265,71 ± 18,57 aB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas avaliadas após 40 dias sob déficit hídrico.
4.1.2 Déficit hídrico no estádio de florescimento
Houve redução de Ψw devido ao déficit hídrico nas medidas avaliadas na
antemanhã, exceto para o híbrido DAS2B710 que manteve o mesmo valor de Ψw
independente do regime hídrico. O híbrido DAS2B710 apresentou menor Ψw (-0,31
MPa) sob condição irrigada, enquanto que o AS1522 apresentou o menor Ψw (-0,59
MPa) em condição de déficit hídrico (Figura 8).

44
Figura 8. Potencial da água na folha (Ψw) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio de florescimento (R1).
Avaliações realizadas às 05h30 após 75 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam
diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma
condição hídrica e letras maiúsculas distintas representam diferenças estatísticas entre
os tratamentos hídricos.
Houve redução dos valores de gs nos híbridos devido ao déficit hídrico, exceto
para o DAS2B710 que manteve sua condutância estomática independente do tratamento
hídrico empregado. Verificou-se diferença entre híbridos somente quando as plantas
encontravam-se sob condição irrigada, com DAS2B707 e FT510 apresentando maiores
valores de condutância estomática (Figura 9a). Houve aumento significativo de Ci
quando os híbridos foram submetidos ao estresse. Pode-se observar diferença entre
híbridos nos dois tratamentos hídricos empregados, com o DAS2B707 apresentando
menor concentração intercelular de CO2 no tratamento irrigado e o AS1522 com menor
Ci no tratamento submetido ao déficit hídrico (Figura 9b).
2B707 2B710 FT510 AS1522
-1,2
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
cB
aBaA
bB
cA
bA bA
aA
Po
ten
cia
l d
a á
gu
a n
a f
olh
a (
MP
a)
irrigado
déficit hídrico
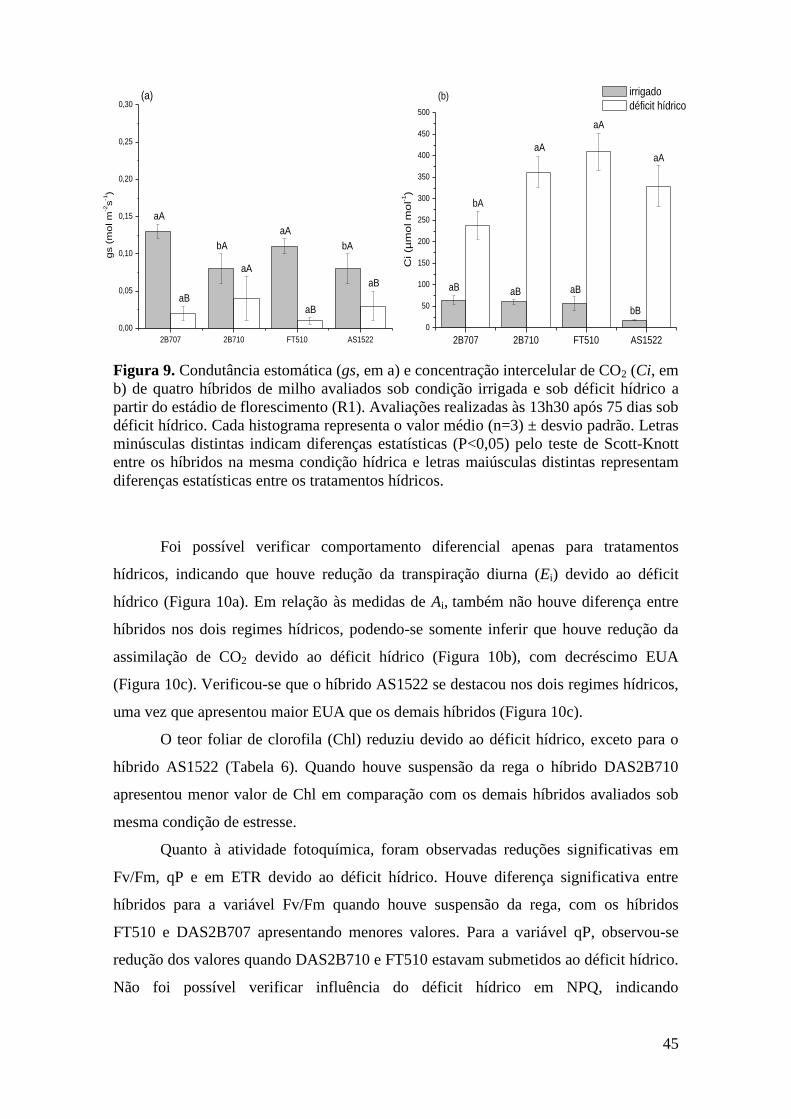
45
Figura 9. Condutância estomática (gs, em a) e concentração intercelular de CO2 (Ci, em
b) de quatro híbridos de milho avaliados sob condição irrigada e sob déficit hídrico a
partir do estádio de florescimento (R1). Avaliações realizadas às 13h30 após 75 dias sob
déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras
minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott
entre os híbridos na mesma condição hídrica e letras maiúsculas distintas representam
diferenças estatísticas entre os tratamentos hídricos.
Foi possível verificar comportamento diferencial apenas para tratamentos
hídricos, indicando que houve redução da transpiração diurna (Ei) devido ao déficit
hídrico (Figura 10a). Em relação às medidas de Ai, também não houve diferença entre
híbridos nos dois regimes hídricos, podendo-se somente inferir que houve redução da
assimilação de CO2 devido ao déficit hídrico (Figura 10b), com decréscimo EUA
(Figura 10c). Verificou-se que o híbrido AS1522 se destacou nos dois regimes hídricos,
uma vez que apresentou maior EUA que os demais híbridos (Figura 10c).
O teor foliar de clorofila (Chl) reduziu devido ao déficit hídrico, exceto para o
híbrido AS1522 (Tabela 6). Quando houve suspensão da rega o híbrido DAS2B710
apresentou menor valor de Chl em comparação com os demais híbridos avaliados sob
mesma condição de estresse.
Quanto à atividade fotoquímica, foram observadas reduções significativas em
Fv/Fm, qP e em ETR devido ao déficit hídrico. Houve diferença significativa entre
híbridos para a variável Fv/Fm quando houve suspensão da rega, com os híbridos
FT510 e DAS2B707 apresentando menores valores. Para a variável qP, observou-se
redução dos valores quando DAS2B710 e FT510 estavam submetidos ao déficit hídrico.
Não foi possível verificar influência do déficit hídrico em NPQ, indicando
2B707 2B710 FT510 AS1522
0
50
100
150
200
250
300
350
400
450
500
(b) irrigado
déficit hídrico
aA
aA
aA
bA
bB
aB aBaB
Ci (µ
mo
l m
ol-1
)
2B707 2B710 FT510 AS1522
0,00
0,05
0,10
0,15
0,20
0,25
0,30(a)
aB
aB
aA
aB
bA
aA
bA
aA
gs (
mo
l m
-2s
-1)
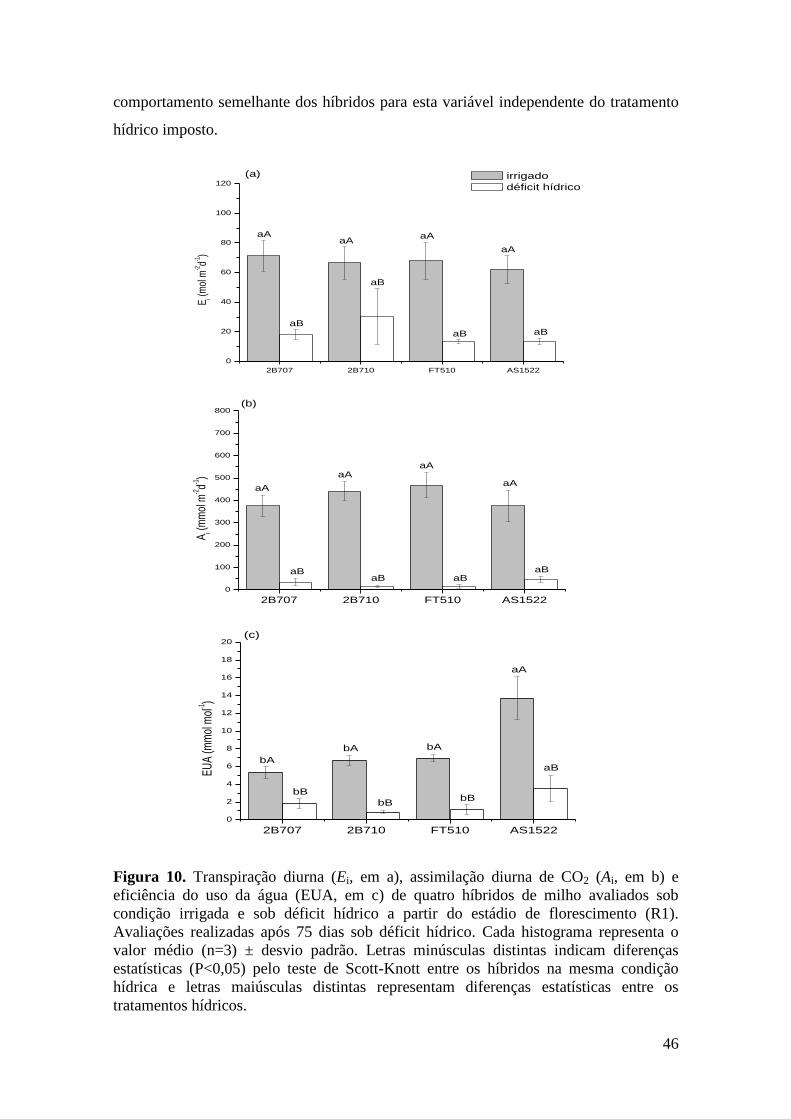
46
comportamento semelhante dos híbridos para esta variável independente do tratamento
hídrico imposto.
Figura 10. Transpiração diurna (Ei, em a), assimilação diurna de CO2 (Ai, em b) e
eficiência do uso da água (EUA, em c) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio de florescimento (R1).
Avaliações realizadas após 75 dias sob déficit hídrico. Cada histograma representa o
valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças
estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição
hídrica e letras maiúsculas distintas representam diferenças estatísticas entre os
tratamentos hídricos.
2B707 2B710 FT510 AS1522
0
100
200
300
400
500
600
700
800(b)
aBaBaB
aA
aAaA
aB
aA
Ai (m
mol
m-2d-1
)
2B707 2B710 FT510 AS1522
0
2
4
6
8
10
12
14
16
18
20(c)
aB
bBbB
bB
aA
bA bA
bA
EU
A (m
mol
mol
-1)
2B707 2B710 FT510 AS1522
0
20
40
60
80
100
120
(a)
aBaB
aB
aB
aA
aAaA
aA
Ei (m
ol m
-2d-1
)
irrigado
déficit hídrico
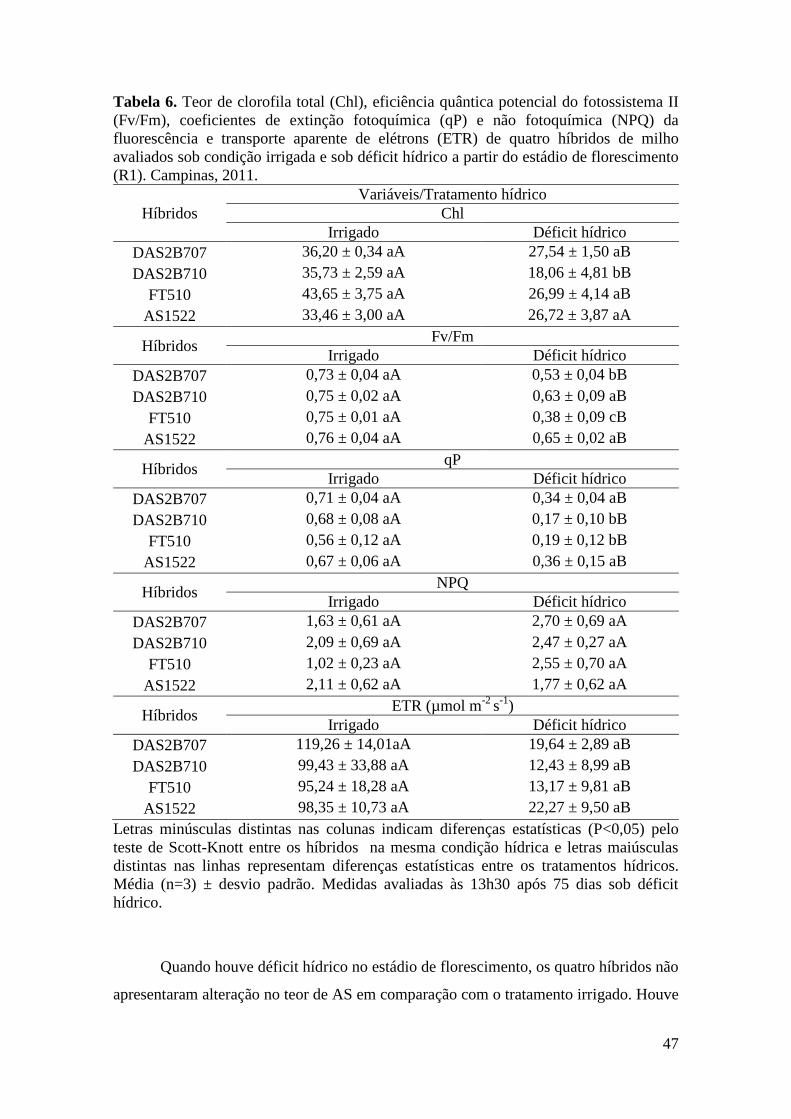
47
Tabela 6. Teor de clorofila total (Chl), eficiência quântica potencial do fotossistema II
(Fv/Fm), coeficientes de extinção fotoquímica (qP) e não fotoquímica (NPQ) da
fluorescência e transporte aparente de elétrons (ETR) de quatro híbridos de milho
avaliados sob condição irrigada e sob déficit hídrico a partir do estádio de florescimento
(R1). Campinas, 2011.
Híbridos
Variáveis/Tratamento hídrico
Chl
Irrigado Déficit hídrico
DAS2B707 36,20 ± 0,34 aA 27,54 ± 1,50 aB
DAS2B710 35,73 ± 2,59 aA 18,06 ± 4,81 bB
FT510 43,65 ± 3,75 aA 26,99 ± 4,14 aB
AS1522 33,46 ± 3,00 aA 26,72 ± 3,87 aA
Híbridos Fv/Fm
Irrigado Déficit hídrico
DAS2B707 0,73 ± 0,04 aA 0,53 ± 0,04 bB
DAS2B710 0,75 ± 0,02 aA 0,63 ± 0,09 aB
FT510 0,75 ± 0,01 aA 0,38 ± 0,09 cB
AS1522 0,76 ± 0,04 aA 0,65 ± 0,02 aB
Híbridos qP
Irrigado Déficit hídrico
DAS2B707 0,71 ± 0,04 aA 0,34 ± 0,04 aB
DAS2B710 0,68 ± 0,08 aA 0,17 ± 0,10 bB
FT510 0,56 ± 0,12 aA 0,19 ± 0,12 bB
AS1522 0,67 ± 0,06 aA 0,36 ± 0,15 aB
Híbridos NPQ
Irrigado Déficit hídrico
DAS2B707 1,63 ± 0,61 aA 2,70 ± 0,69 aA
DAS2B710 2,09 ± 0,69 aA 2,47 ± 0,27 aA
FT510 1,02 ± 0,23 aA 2,55 ± 0,70 aA
AS1522 2,11 ± 0,62 aA 1,77 ± 0,62 aA
Híbridos ETR (µmol m
-2 s
-1)
Irrigado Déficit hídrico
DAS2B707 119,26 ± 14,01aA 19,64 ± 2,89 aB
DAS2B710 99,43 ± 33,88 aA 12,43 ± 8,99 aB
FT510 95,24 ± 18,28 aA 13,17 ± 9,81 aB
AS1522 98,35 ± 10,73 aA 22,27 ± 9,50 aB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas avaliadas às 13h30 após 75 dias sob déficit
hídrico.
Quando houve déficit hídrico no estádio de florescimento, os quatro híbridos não
apresentaram alteração no teor de AS em comparação com o tratamento irrigado. Houve

48
diferença significativa entre híbridos tanto nas plantas mantidas sob irrigação quanto
nas plantas submetidas ao estresse. O híbrido AS1522 destacou-se nos dois tratamentos
hídricos, uma vez que apresentou valores elevados de AS, apresentando 86,8 e 97,6 mg
g-1
no tratamento irrigado e no de déficit hídrico, respectivamente (Figura 11a).
Em relação ao teor de SAC, o déficit hídrico aumentou o teor nos híbridos
DAS2B707 e AS1522, com aumentos de 38% e 47% de SAC, respectivamente. Em
condição irrigada, o híbrido DAS2B707 foi o que obteve menor valor, enquanto que,
em condição de déficit hídrico o AS1522 apresentou maior teor de SAC em comparação
aos demais híbridos (Figura 11b).
O teor de AM foi reduzido pelo déficit hídrico. Pôde-se verificar diferença entre
os híbridos somente na condição irrigada, sendo FT510 e AS1522 os híbridos que
obtiveram maiores teores de AM (Figura 11c).
Quanto ao teor de carboidrato total não-estrutural (CTNE), observou-se
diferença entre híbridos e entre tratamentos hídricos. Na condição irrigada o DAS2B707
apresentou menor valor de CTNE e, quando houve déficit hídrico, o AS1522 apresentou
maior conteúdo de CTNE (Figura 11d).
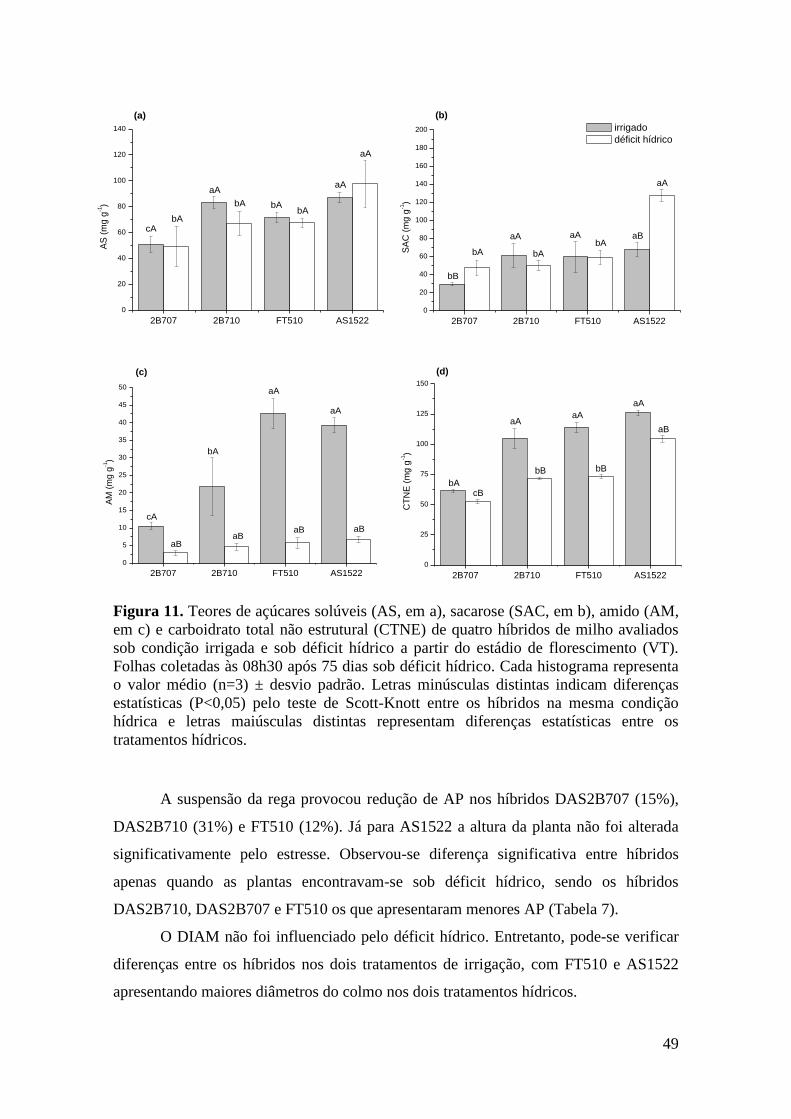
49
Figura 11. Teores de açúcares solúveis (AS, em a), sacarose (SAC, em b), amido (AM,
em c) e carboidrato total não estrutural (CTNE) de quatro híbridos de milho avaliados
sob condição irrigada e sob déficit hídrico a partir do estádio de florescimento (VT).
Folhas coletadas às 08h30 após 75 dias sob déficit hídrico. Cada histograma representa
o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças
estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição
hídrica e letras maiúsculas distintas representam diferenças estatísticas entre os
tratamentos hídricos.
A suspensão da rega provocou redução de AP nos híbridos DAS2B707 (15%),
DAS2B710 (31%) e FT510 (12%). Já para AS1522 a altura da planta não foi alterada
significativamente pelo estresse. Observou-se diferença significativa entre híbridos
apenas quando as plantas encontravam-se sob déficit hídrico, sendo os híbridos
DAS2B710, DAS2B707 e FT510 os que apresentaram menores AP (Tabela 7).
O DIAM não foi influenciado pelo déficit hídrico. Entretanto, pode-se verificar
diferenças entre os híbridos nos dois tratamentos de irrigação, com FT510 e AS1522
apresentando maiores diâmetros do colmo nos dois tratamentos hídricos.
2B707 2B710 FT510 AS1522
0
20
40
60
80
100
120
140
(a)
bAbA
bA
aA
cA
bA
aAaA
AS
(m
g g
-1)
2B707 2B710 FT510 AS1522
0
20
40
60
80
100
120
140
160
180
200 irrigado
déficit hídrico
(b)
aA
bA
bAbA
aBaA aA
bB
SA
C (
mg
g-1)
2B707 2B710 FT510 AS1522
0
5
10
15
20
25
30
35
40
45
50
(c)
aBaBaB
aB
aA
bA
aA
cA
AM
(m
g g
-1)
2B707 2B710 FT510 AS1522
0
25
50
75
100
125
150
(d)
aB
bBbB
cB
aA
aAaA
bA
CT
NE
(m
g g
-1)
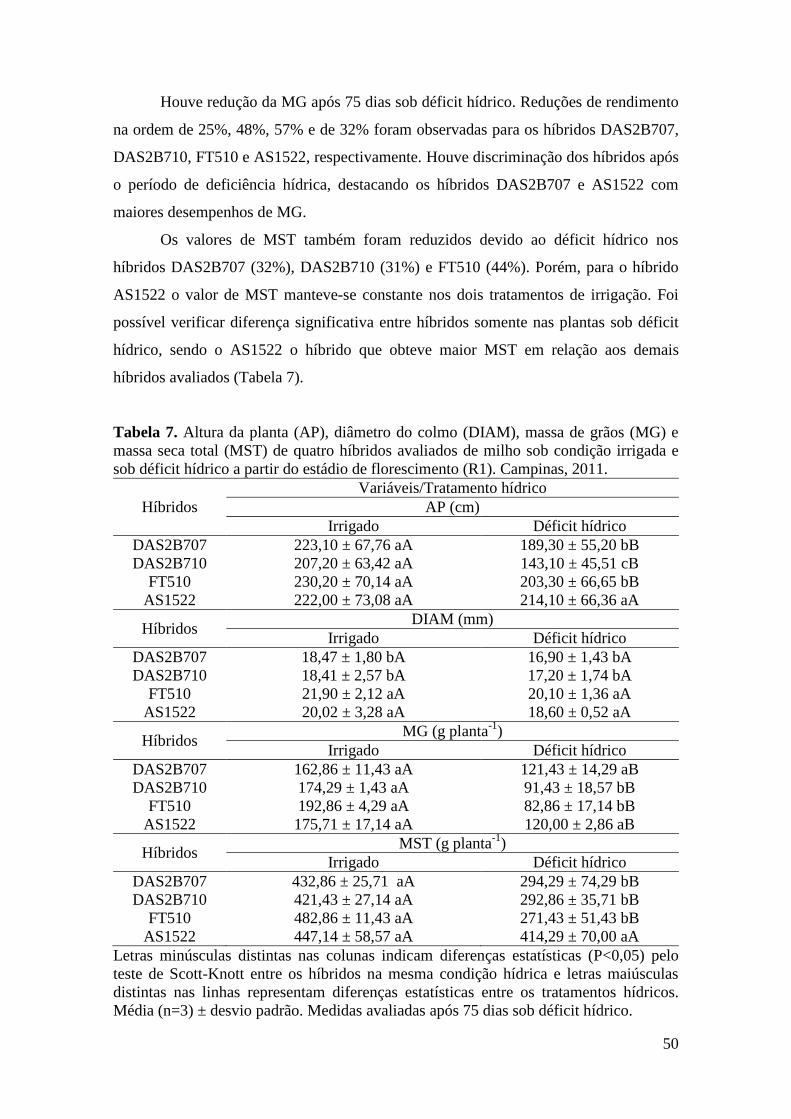
50
Houve redução da MG após 75 dias sob déficit hídrico. Reduções de rendimento
na ordem de 25%, 48%, 57% e de 32% foram observadas para os híbridos DAS2B707,
DAS2B710, FT510 e AS1522, respectivamente. Houve discriminação dos híbridos após
o período de deficiência hídrica, destacando os híbridos DAS2B707 e AS1522 com
maiores desempenhos de MG.
Os valores de MST também foram reduzidos devido ao déficit hídrico nos
híbridos DAS2B707 (32%), DAS2B710 (31%) e FT510 (44%). Porém, para o híbrido
AS1522 o valor de MST manteve-se constante nos dois tratamentos de irrigação. Foi
possível verificar diferença significativa entre híbridos somente nas plantas sob déficit
hídrico, sendo o AS1522 o híbrido que obteve maior MST em relação aos demais
híbridos avaliados (Tabela 7).
Tabela 7. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG) e
massa seca total (MST) de quatro híbridos avaliados de milho sob condição irrigada e
sob déficit hídrico a partir do estádio de florescimento (R1). Campinas, 2011.
Híbridos
Variáveis/Tratamento hídrico
AP (cm)
Irrigado Déficit hídrico
DAS2B707 223,10 ± 67,76 aA 189,30 ± 55,20 bB
DAS2B710 207,20 ± 63,42 aA 143,10 ± 45,51 cB
FT510 230,20 ± 70,14 aA 203,30 ± 66,65 bB
AS1522 222,00 ± 73,08 aA 214,10 ± 66,36 aA
Híbridos DIAM (mm)
Irrigado Déficit hídrico
DAS2B707 18,47 ± 1,80 bA 16,90 ± 1,43 bA
DAS2B710 18,41 ± 2,57 bA 17,20 ± 1,74 bA
FT510 21,90 ± 2,12 aA 20,10 ± 1,36 aA
AS1522 20,02 ± 3,28 aA 18,60 ± 0,52 aA
Híbridos MG (g planta
-1)
Irrigado Déficit hídrico
DAS2B707 162,86 ± 11,43 aA 121,43 ± 14,29 aB
DAS2B710 174,29 ± 1,43 aA 91,43 ± 18,57 bB
FT510 192,86 ± 4,29 aA 82,86 ± 17,14 bB
AS1522 175,71 ± 17,14 aA 120,00 ± 2,86 aB
Híbridos MST (g planta
-1)
Irrigado Déficit hídrico
DAS2B707 432,86 ± 25,71 aA 294,29 ± 74,29 bB
DAS2B710 421,43 ± 27,14 aA 292,86 ± 35,71 bB
FT510 482,86 ± 11,43 aA 271,43 ± 51,43 bB
AS1522 447,14 ± 58,57 aA 414,29 ± 70,00 aA
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas avaliadas após 75 dias sob déficit hídrico.
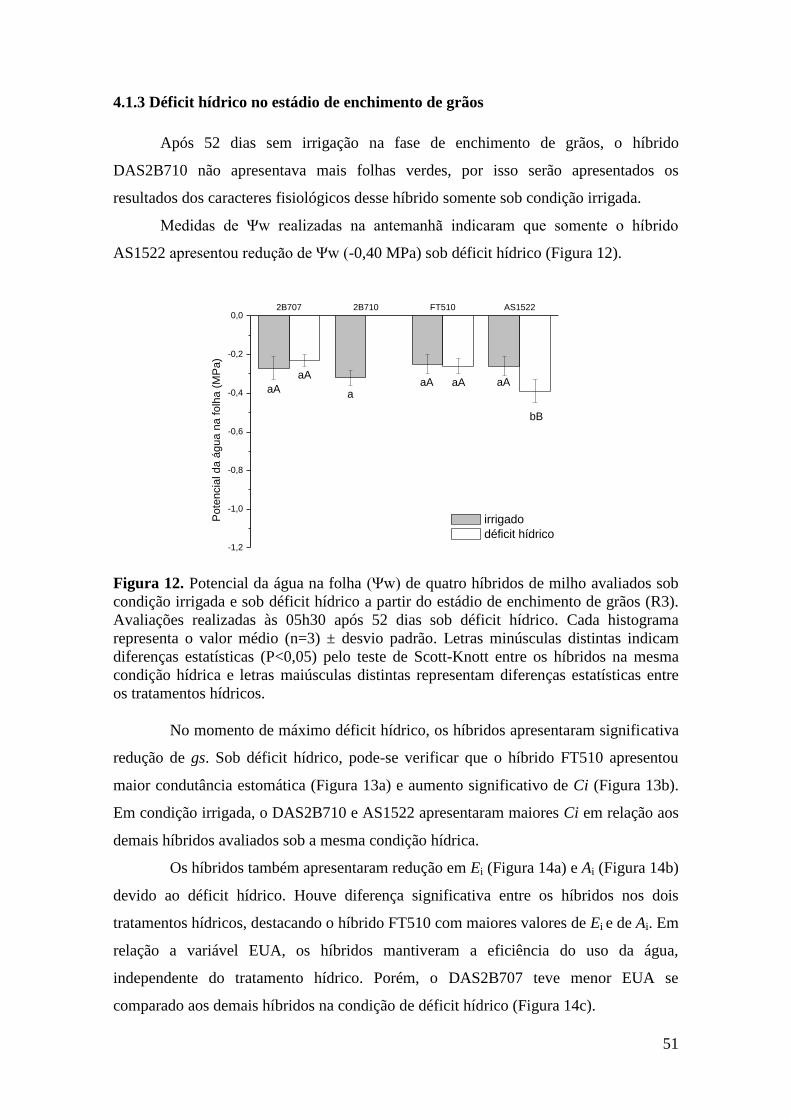
51
4.1.3 Déficit hídrico no estádio de enchimento de grãos
Após 52 dias sem irrigação na fase de enchimento de grãos, o híbrido
DAS2B710 não apresentava mais folhas verdes, por isso serão apresentados os
resultados dos caracteres fisiológicos desse híbrido somente sob condição irrigada.
Medidas de Ψw realizadas na antemanhã indicaram que somente o híbrido
AS1522 apresentou redução de Ψw (-0,40 MPa) sob déficit hídrico (Figura 12).
Figura 12. Potencial da água na folha (Ψw) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio de enchimento de grãos (R3).
Avaliações realizadas às 05h30 após 52 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam
diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma
condição hídrica e letras maiúsculas distintas representam diferenças estatísticas entre
os tratamentos hídricos.
No momento de máximo déficit hídrico, os híbridos apresentaram significativa
redução de gs. Sob déficit hídrico, pode-se verificar que o híbrido FT510 apresentou
maior condutância estomática (Figura 13a) e aumento significativo de Ci (Figura 13b).
Em condição irrigada, o DAS2B710 e AS1522 apresentaram maiores Ci em relação aos
demais híbridos avaliados sob a mesma condição hídrica.
Os híbridos também apresentaram redução em Ei (Figura 14a) e Ai (Figura 14b)
devido ao déficit hídrico. Houve diferença significativa entre os híbridos nos dois
tratamentos hídricos, destacando o híbrido FT510 com maiores valores de Ei e de Ai. Em
relação a variável EUA, os híbridos mantiveram a eficiência do uso da água,
independente do tratamento hídrico. Porém, o DAS2B707 teve menor EUA se
comparado aos demais híbridos na condição de déficit hídrico (Figura 14c).
2B707 2B710 FT510 AS1522
-1,2
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
bB
aAaAaAa
aA
aA
Po
ten
cia
l d
a á
gu
a n
a fo
lha
(M
Pa
)
irrigado
déficit hídrico
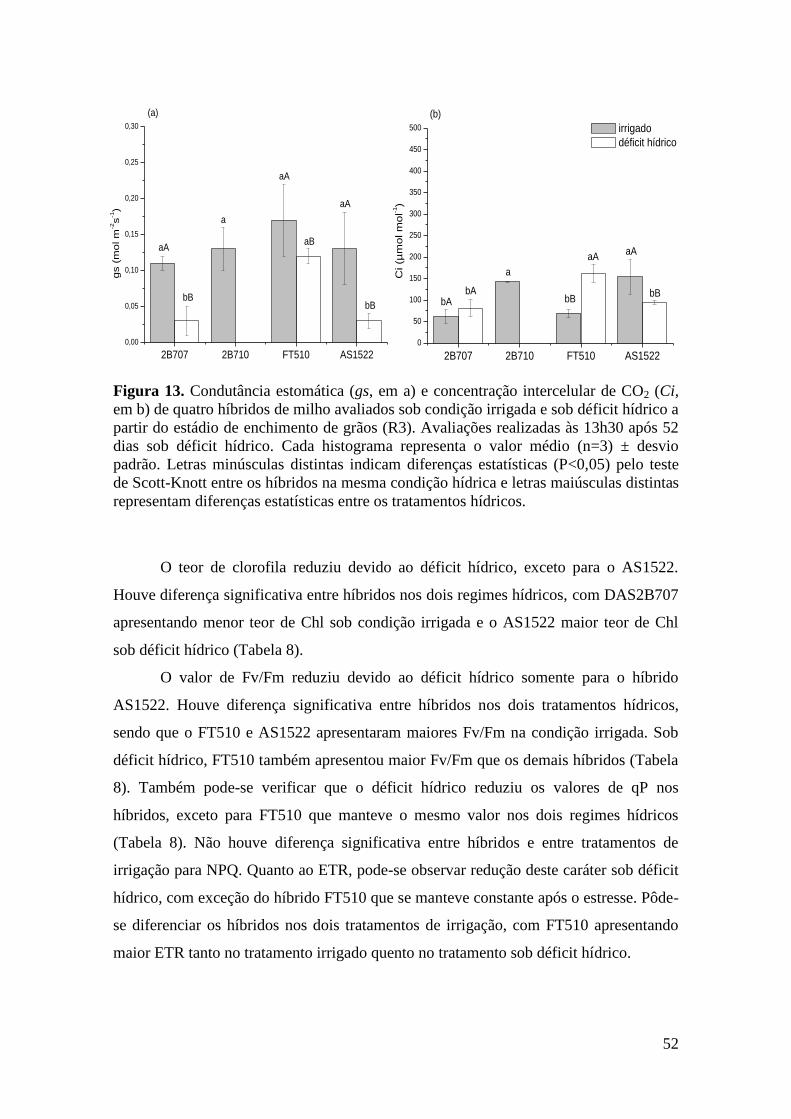
52
Figura 13. Condutância estomática (gs, em a) e concentração intercelular de CO2 (Ci,
em b) de quatro híbridos de milho avaliados sob condição irrigada e sob déficit hídrico a
partir do estádio de enchimento de grãos (R3). Avaliações realizadas às 13h30 após 52
dias sob déficit hídrico. Cada histograma representa o valor médio (n=3) ± desvio
padrão. Letras minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo teste
de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas distintas
representam diferenças estatísticas entre os tratamentos hídricos.
O teor de clorofila reduziu devido ao déficit hídrico, exceto para o AS1522.
Houve diferença significativa entre híbridos nos dois regimes hídricos, com DAS2B707
apresentando menor teor de Chl sob condição irrigada e o AS1522 maior teor de Chl
sob déficit hídrico (Tabela 8).
O valor de Fv/Fm reduziu devido ao déficit hídrico somente para o híbrido
AS1522. Houve diferença significativa entre híbridos nos dois tratamentos hídricos,
sendo que o FT510 e AS1522 apresentaram maiores Fv/Fm na condição irrigada. Sob
déficit hídrico, FT510 também apresentou maior Fv/Fm que os demais híbridos (Tabela
8). Também pode-se verificar que o déficit hídrico reduziu os valores de qP nos
híbridos, exceto para FT510 que manteve o mesmo valor nos dois regimes hídricos
(Tabela 8). Não houve diferença significativa entre híbridos e entre tratamentos de
irrigação para NPQ. Quanto ao ETR, pode-se observar redução deste caráter sob déficit
hídrico, com exceção do híbrido FT510 que se manteve constante após o estresse. Pôde-
se diferenciar os híbridos nos dois tratamentos de irrigação, com FT510 apresentando
maior ETR tanto no tratamento irrigado quento no tratamento sob déficit hídrico.
2B707 2B710 FT510 AS1522
0,00
0,05
0,10
0,15
0,20
0,25
0,30
(a)
bBbB
aB
aA
aA
a
aA
gs (
mo
l m
-2s
-1)
2B707 2B710 FT510 AS1522
0
50
100
150
200
250
300
350
400
450
500 irrigado
déficit hídrico
(b)
bB
aA
bA
aA
a
bBbA
Ci (µ
mo
l m
ol-1
)
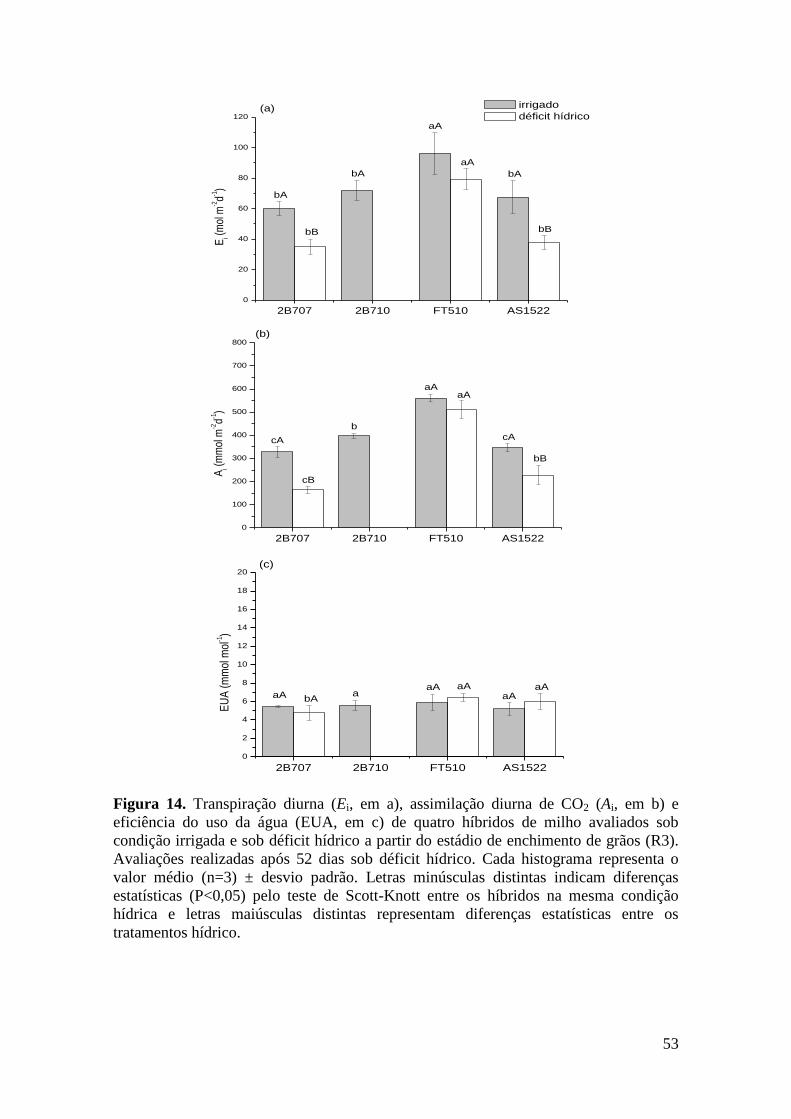
53
Figura 14. Transpiração diurna (Ei, em a), assimilação diurna de CO2 (Ai, em b) e
eficiência do uso da água (EUA, em c) de quatro híbridos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio de enchimento de grãos (R3).
Avaliações realizadas após 52 dias sob déficit hídrico. Cada histograma representa o
valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças
estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição
hídrica e letras maiúsculas distintas representam diferenças estatísticas entre os
tratamentos hídrico.
2B707 2B710 FT510 AS1522
0
20
40
60
80
100
120
irrigado
déficit hídrico(a)
bBbB
aA
bAbA
bA
aA
Ei (
mol
m-2d-1
)
2B707 2B710 FT510 AS1522
0
100
200
300
400
500
600
700
800(b)
cB
bB
aA
cA cA
b
aA
Ai (
mm
ol m
-2d-1
)
2B707 2B710 FT510 AS1522
0
2
4
6
8
10
12
14
16
18
20(c)
aAaA
bA aAaaA
aA
EU
A (
mm
ol m
ol-1)
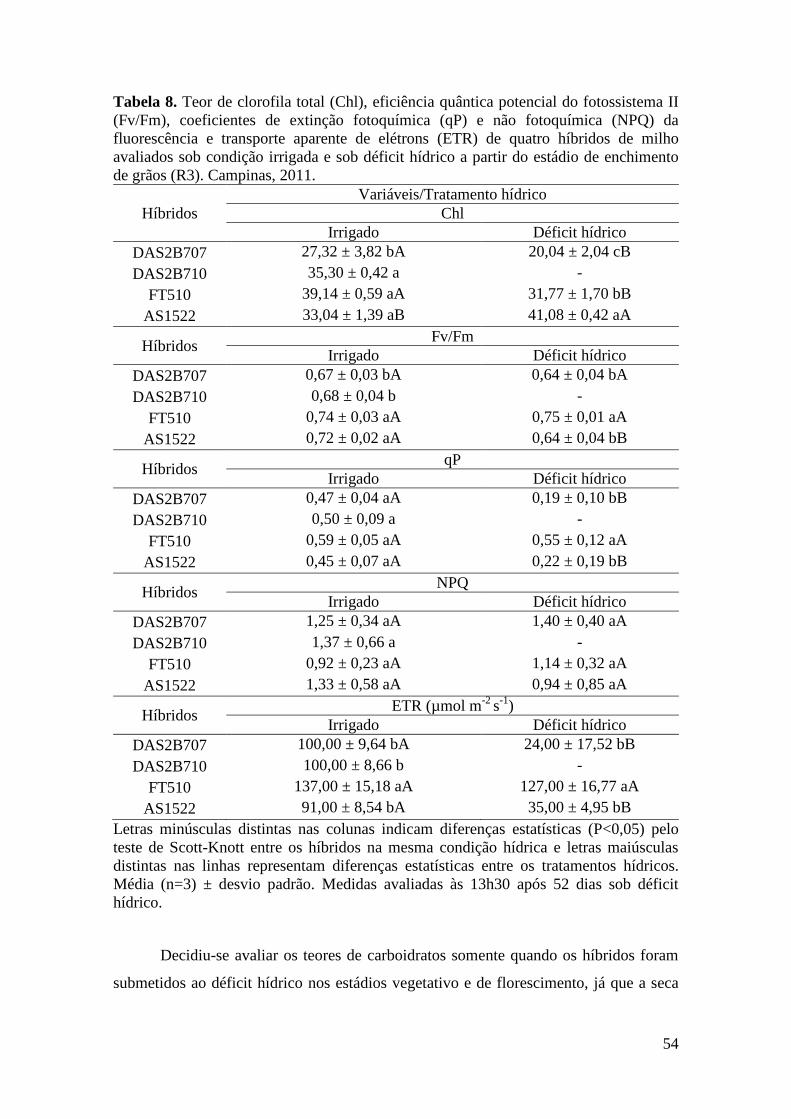
54
Tabela 8. Teor de clorofila total (Chl), eficiência quântica potencial do fotossistema II
(Fv/Fm), coeficientes de extinção fotoquímica (qP) e não fotoquímica (NPQ) da
fluorescência e transporte aparente de elétrons (ETR) de quatro híbridos de milho
avaliados sob condição irrigada e sob déficit hídrico a partir do estádio de enchimento
de grãos (R3). Campinas, 2011.
Híbridos
Variáveis/Tratamento hídrico
Chl
Irrigado Déficit hídrico
DAS2B707 27,32 ± 3,82 bA 20,04 ± 2,04 cB
DAS2B710 35,30 ± 0,42 a -
FT510 39,14 ± 0,59 aA 31,77 ± 1,70 bB
AS1522 33,04 ± 1,39 aB 41,08 ± 0,42 aA
Híbridos Fv/Fm
Irrigado Déficit hídrico
DAS2B707 0,67 ± 0,03 bA 0,64 ± 0,04 bA
DAS2B710 0,68 ± 0,04 b -
FT510 0,74 ± 0,03 aA 0,75 ± 0,01 aA
AS1522 0,72 ± 0,02 aA 0,64 ± 0,04 bB
Híbridos qP
Irrigado Déficit hídrico
DAS2B707 0,47 ± 0,04 aA 0,19 ± 0,10 bB
DAS2B710 0,50 ± 0,09 a -
FT510 0,59 ± 0,05 aA 0,55 ± 0,12 aA
AS1522 0,45 ± 0,07 aA 0,22 ± 0,19 bB
Híbridos NPQ
Irrigado Déficit hídrico
DAS2B707 1,25 ± 0,34 aA 1,40 ± 0,40 aA
DAS2B710 1,37 ± 0,66 a -
FT510 0,92 ± 0,23 aA 1,14 ± 0,32 aA
AS1522 1,33 ± 0,58 aA 0,94 ± 0,85 aA
Híbridos ETR (µmol m
-2 s
-1)
Irrigado Déficit hídrico
DAS2B707 100,00 ± 9,64 bA 24,00 ± 17,52 bB
DAS2B710 100,00 ± 8,66 b -
FT510 137,00 ± 15,18 aA 127,00 ± 16,77 aA
AS1522 91,00 ± 8,54 bA 35,00 ± 4,95 bB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas avaliadas às 13h30 após 52 dias sob déficit
hídrico.
Decidiu-se avaliar os teores de carboidratos somente quando os híbridos foram
submetidos ao déficit hídrico nos estádios vegetativo e de florescimento, já que a seca
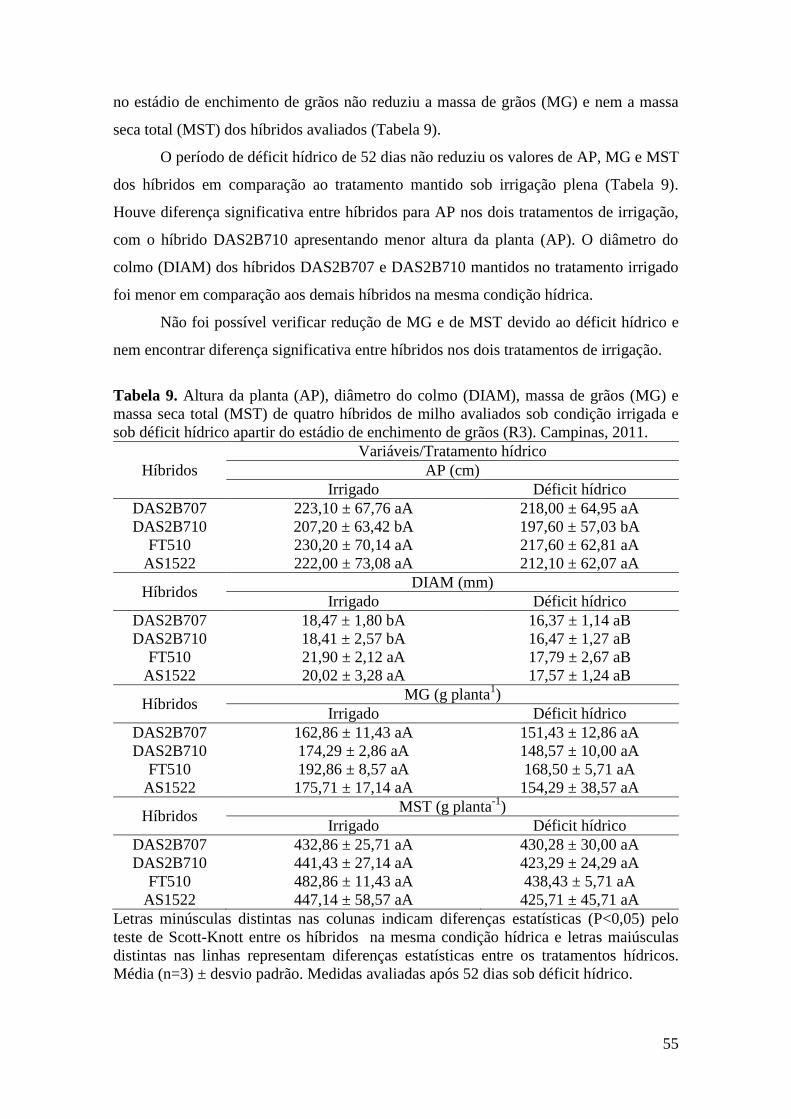
55
no estádio de enchimento de grãos não reduziu a massa de grãos (MG) e nem a massa
seca total (MST) dos híbridos avaliados (Tabela 9).
O período de déficit hídrico de 52 dias não reduziu os valores de AP, MG e MST
dos híbridos em comparação ao tratamento mantido sob irrigação plena (Tabela 9).
Houve diferença significativa entre híbridos para AP nos dois tratamentos de irrigação,
com o híbrido DAS2B710 apresentando menor altura da planta (AP). O diâmetro do
colmo (DIAM) dos híbridos DAS2B707 e DAS2B710 mantidos no tratamento irrigado
foi menor em comparação aos demais híbridos na mesma condição hídrica.
Não foi possível verificar redução de MG e de MST devido ao déficit hídrico e
nem encontrar diferença significativa entre híbridos nos dois tratamentos de irrigação.
Tabela 9. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG) e
massa seca total (MST) de quatro híbridos de milho avaliados sob condição irrigada e
sob déficit hídrico apartir do estádio de enchimento de grãos (R3). Campinas, 2011.
Híbridos
Variáveis/Tratamento hídrico
AP (cm)
Irrigado Déficit hídrico
DAS2B707 223,10 ± 67,76 aA 218,00 ± 64,95 aA
DAS2B710 207,20 ± 63,42 bA 197,60 ± 57,03 bA
FT510 230,20 ± 70,14 aA 217,60 ± 62,81 aA
AS1522 222,00 ± 73,08 aA 212,10 ± 62,07 aA
Híbridos DIAM (mm)
Irrigado Déficit hídrico
DAS2B707 18,47 ± 1,80 bA 16,37 ± 1,14 aB
DAS2B710 18,41 ± 2,57 bA 16,47 ± 1,27 aB
FT510 21,90 ± 2,12 aA 17,79 ± 2,67 aB
AS1522 20,02 ± 3,28 aA 17,57 ± 1,24 aB
Híbridos MG (g planta
1)
Irrigado Déficit hídrico
DAS2B707 162,86 ± 11,43 aA 151,43 ± 12,86 aA
DAS2B710 174,29 ± 2,86 aA 148,57 ± 10,00 aA
FT510 192,86 ± 8,57 aA 168,50 ± 5,71 aA
AS1522 175,71 ± 17,14 aA 154,29 ± 38,57 aA
Híbridos MST (g planta
-1)
Irrigado Déficit hídrico
DAS2B707 432,86 ± 25,71 aA 430,28 ± 30,00 aA
DAS2B710 441,43 ± 27,14 aA 423,29 ± 24,29 aA
FT510 482,86 ± 11,43 aA 438,43 ± 5,71 aA
AS1522 447,14 ± 58,57 aA 425,71 ± 45,71 aA
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas avaliadas após 52 dias sob déficit hídrico.

56
4.2 DISCUSSÃO: Parâmetros fisiológicos
4.2.1 Déficit hídrico no estádio vegetativo
O potencial da água na folha (Ψw) é um dos caracteres mais utilizados para
medir a intensidade do déficit hídrico em vegetais (FLEXAS et al., 2004) e quando
avaliado na antemanhã pode refletir a condição hídrica da planta em equilíbrio com o
solo (BERGONCI et al., 2000). A partir do estudo dessa variável foi possível verificar
uma redução significativa de Ψw em todos os híbridos avaliados devido ao déficit
hídrico imposto no estádio vegetativo (Figura 4), permitindo inferir que o Ψw pode ser
um bom indicador de déficit hídrico. Resultado semelhante foi obtido por BERGONCI
et al. (2000) que avaliando o Ψw em híbridos de milho submetidos a três níveis de
irrigação, desde a capacidade de campo até a ausência de irrigação, verificaram que nos
dias em que a disponibilidade de água no solo era baixa em plantas não irrigadas, o
potencial da água na folha era menor do que em plantas irrigadas.
Pelos resultados obtidos de Ψw no presente estudo, pode-se verificar que o
déficit hídrico no estádio vegetativo discriminou os híbridos de milho, com valores
variando de -0,30 (DAS2B707) a -0,99 MPa (DAS2B710). O alto valor do Ψw
observado no híbrido DAS2B707 sob condição de déficit hídrico está relacionado ao
fechamento parcial dos estômatos (Figura 5a). Além desse aspecto, a melhor hidratação
desse híbrido deve estar certamente associada a outra característica, tal como o aumento
da densidade e profundidade das raízes dessas plantas, garantindo maior exploração do
solo.
As medidas de condutância estomática (gs) apresentadas foram realizadas no
horário de maior demanda atmosférica (13:30 h) e indicam uma redução considerável da
abertura estomática em todos os híbridos submetidos ao déficit hídrico. Somente na
condição irrigada pôde-se discriminar os híbridos, sendo o DAS2B710 o híbrido que
apresentou maior gs (Figura 5a). A redução de gs poderia limitar a fotossíntese devido à
menor disponibilidade de CO2, sendo esse aspecto indicado pela redução de Ci.
Contudo, mesmo com o fechamento parcial dos estômatos, o híbrido DAS2B710
apresentou valor elevado de Ci (Figura 5b), demonstrando-se que havia CO2 disponível
para a realização de fotossíntese e, portanto, não foi este o fator limitante para a sua
realização.

57
Sabe-se que a deficiência hídrica reduz a capacidade fotossintética das plantas
(FLEXAS et al., 2004) e isso é atribuído em grande parte ao decréscimo da condutância
estomática (gs). O controle da abertura e fechamento dos estômatos é essencial para a
regulação do equilíbrio entre a perda de água pela transpiração (E) e a assimilação de
CO2 (A) pela planta (PIMENTEL, 2004). De fato, no presente estudo, o déficit hídrico
imposto na fase vegetativa implicou na redução de gs e, consequentemente, reduziu Ei e
Ai em todos os híbridos em comparação com as plantas mantidas irrigadas (Figuras 5a,
6a,b). Decréscimo na condutância estomática e na fotossíntese também foram
observados por XU et al. (2009) em uma variedade de milho chinesa e por
GHANNOUM et al. (2003) em quatro diferentes espécies de gramíneas sob déficit
hídrico.
Apesar da redução de gs e Ei devido ao período de restrição hídrica, os híbridos
DAS2B707, FT510 e AS1522 apresentaram os maiores valores de Ai. No caso dos
híbridos DAS2B707 e AS1522, os maiores valores de Ai podem estar relacionados à
maior atividade fotoquímica realizada por esses híbridos em condições de déficit
hídrico, obtendo maiores valores do coeficiente de extinção fotoquímica (qP) e da taxa
aparente do transporte de elétrons (ETR) em relação aos demais híbridos.
Para FT510 o alto valor de Ai sob déficit hídrico pode ser devido ao maior teor
de clorofila total (Tabela 4). Maiores teores de clorofila podem denotar aumentos na
longevidade das folhas e adequado funcionamento do fotossistema II, contribuindo para
melhoria na tolerância de híbridos de milho ao déficit hídrico e de nitrogênio
(TOLLENAAR & WU, 1999). A redução no conteúdo deste pigmento como
conseqüência do déficit hídrico tem sido relatado na cultura do milho (VIANA, 2002;
MACHADO, 2003) e corrobora com os resultados obtidos no presente estudo quando se
compara o tratamento sob déficit hídrico com o mantido sob irrigação.
A variável Fv/Fm é uma estimativa da eficiência quântica máxima da atividade
fotoquímica do fotossistema II, quando todos os centros de reação do FSII estão abertos
(BAKER & ROSENQVST, 2004). Essa relação tem sido empregada freqüentemente
para detectar perturbações no sistema fotossintético causada por estresses ambientais e
bióticos, visto que a redução desta indica inibição da atividade fotoquímica com
provável comprometimento da assimilação de CO2. Contudo, não foi possível detectar
diferença significativa entre híbridos e entre regimes hídricos para o caráter Fv/Fm
(Tabela 4). Esta habilidade em manter valores semelhantes de Fv/Fm sob déficit hídrico

58
em relação ao tratamento controle indica a alta resistência da atividade fotoquímica em
condições limitantes (SILVA et al., 2007).
Quando submetido ao déficit hídrico, os híbridos DAS2B710 e FT510
apresentaram aumento do coeficiente de extinção não fotoquímica da fluorescência
(NPQ) (Tabela 4). Esse aumento em NPQ indica que estas plantas apresentaram uma
maior dissipação da energia luminosa absorvida pela formação de calor, em detrimento
da atividade fotoquímica, ou seja, na produção de energia química na forma de ATP e
NADPH.
A deficiência hídrica provocou redução também na taxa aparente do transporte
de elétrons (ETR) em todos os híbridos em comparação ao tratamento irrigado (Tabela
4), indicando que o estresse inibe o transporte de elétrons no fotossistema II,
compromentendo o metabolismo de carbono nesses híbridos. Houve diferença
significativa entre híbridos no tratamento sob défcit hídrico, com os híbridos
DAS2B707 e AS1522 apresentando maiores valores de ETR após o estresse.
À medida que a água se torna menos disponível no solo pode haver o acúmulo
de solutos visando manter a pressão de turgor celular. Em plantas sob deficiência
hídrica, o aumento dos carboidratos solúveis totais pode estar relacionado com o
processo de ajustamento osmótico, mantendo a planta hidratada e, conseqüentemente,
retardando a desidratação dos seus tecidos (BRAY, 1997). Sabe-se que o metabolismo
de carboidratos na planta é outro fator diretamente relacionado ao crescimento e sua
dinâmica também é influenciada sob condições estressantes (CHAVES & OLIVEIRA,
2004). No presente estudo, o teor de açúcar solúvel (AS) não foi influenciado pelo
estresse e nenhum híbrido se destacou em relação aos tratamentos hídricos empregados
(Figura 7a).
Houve redução dos teores de sacarose (SAC) nas folhas dos híbridos DAS2B707
e FT510 quando esses foram submetidos ao déficit hídrico (Figura 7b). A redução da
sacarose se deve ao decréscimo da assimilação de CO2 e também pode estar associada a
exportação de sacarose para o crescimento do caule e desenvolvimento de raízes. Neste
trabalho, a sacarose pode ter sido empregada para o crescimento da planta do híbrido
FT510, enquanto que para o DAS2B707 foi utilizada para o desenvolvimento de raízes,
uma vez que, sob condições de déficit hídrico este híbrido obteve maior Ψw (Figura 4).
A deficiência hídrica também reduziu o teor de amido (AM) em todos os
híbridos, apresentando reduções de 73%, 63%, 82% e 38%, respectivamente para
DAS2B707, DAS2B710, FT510 e AS1522. Estas reduções podem estar ligadas a uma

59
menor síntese de amido uma vez que a assimilação de CO2 (Ai) reduziu sob déficit
hídrico (PINHEIRO & CHAVES, 2011) e também pode-se sugerir que o carbono
utilizado para manter o nível de açúcares solúveis (AST) nas folhas foi proveniente da
hidrólise do AM. De fato, o consumo de AM seria uma resposta inicial à seca
permitindo a manutenção do suprimento de carbono em uma condição de reduzida
fixação de CO2 atmosférico (KAKANI et al., 2011).
A concentração de carboidrato total não estrutural (CTNE) foi afetada pelo
déficit hídrico apenas no híbrido FT510. Nos demais híbridos o déficit hídrico manteve
o mesmo conteúdo deste carboidratado em relação ao tratamento irrigado, indicando
uma possível paralisação na exportação de triose-fosfato do cloroplasto associada à
redução da força de dreno e dos teores de SAC no tecido (AINSWORTH & BUSH,
2011).
Não houve redução da altura da planta (AP) quando os híbridos FT510 e
AS1522 foram submetidos ao tratamento sob déficit hídrico enquanto que, os híbridos
DAS2B707 e DAS2B710 apresentaram plantas de estaturas consideradas baixas após o
estresse (Tabela 5). Sabe-se que a redução na produção da fitomassa e os efeitos
adversos nos processos fisiológicos podem ser consequência do déficit hídrico, que
afeta diretamente a produção de grãos (MÜLLER et al., 2002). De fato, quando se
compara o tratamento sob déficit hídrico com o tratamento irrigado, verifica-se que os
híbridos DAS2B710, FT510 e AS1522 apresentam reduções de MG de 42%, 46% e
45%, respectivamente. Porém, o híbrido DAS2B707 apresenta estabilidade de produção
nos dois tratamentos hídricos impostos, apresentando redução de apenas 18% quando
houve restrição hídrica. Este resultado indica que este híbrido possui maior potencial
produtivo em condições de deficiência hídrica e pode ser considerado resistente ao
déficit hídrico no estádio vegetativo da cultura.
Assim como a variável MG, a produção de matéria seca total (MST) dos
híbridos foi significativamente afetada pelo déficit hídrico, porém para MST não foi
possível destacar nenhum híbrido nos regimes hídricos impostos (Tabela 5).
A redução relativa (em relação ao controle) em cada variável analisada foi
calculada e os resultados são apresentados na Figura 15. De acordo com essa análise,
pode-se inferir que o híbrido DAS2B707 foi menos afetado pelo déficit hídrico
considerando os valores de MG e de MST devido a maior eficiência na gs, Ai, NPQ, qP,
ETR, AS, CTNE e EUA (valores mais próximos de 1) em relação aos demais híbridos,
além de apresentar maior hidratação (Ψw) após o período de déficit hídrico.

60
Figura 15. Representação gráfica da redução relativa da altura da planta (AP), diâmetro
do colmo (DIAM), massa de grãos (MG), massa seca total (MST), condutância
estomática (gs), concentração intercelular de CO2 (Ci), assimilação diurna de CO2 (Ai),
transpiração diurna (Ei), teor de clorofila total (Chl), eficiência quântica potencial do
fotossistema II (Fv/Fm), coeficiente de dissipação não-fotoquímica da fluorescência
(NPQ), coeficiente de dissipação fotoquímica (qP), taxa aparente de transporte de
elétrons (ETR), açúcares solúveis (AS), sacarose (SAC), amido (AM), carboidrato total
não estrutural (CTNE) e eficiência do uso da água (EUA) de quatro híbridos de milho
submetidos a dois tratamentos hídricos a partir do estádio vegetativo (V5). Valores
maiores que 1 indicam aumento na variável específica sob déficit hídrico.
4.2.2 Déficit hídrico no estádio de florescimento
O período de déficit hídrico na fase de florescimento levou à maior redução de
Ψw no híbrido AS1522 com valor de -0,59 MPa (Figura 8). Valores de Ψw próximos a -
0,40 MPa afetam significativamente a expansão foliar em milho, embora a fotossíntese
máxima ocorra com potenciais foliares entre -0,60 a -0,80 MPa (BERGONCI et al.,
2000).
Para conservar água, nutrientes e carboidratos necessários para a sobrevivência,
as plantas apresentam modificações morfofisiológicas, como a redução da expansão
foliar e o fechamento dos estômatos. Quando houve suspensão da rega, pôde-se notar
redução significativa da condutância estomática (gs) nos híbridos, exceto para o

61
DAS2B710 que não apresentou diferença significativa para esta variável nos dois
tratamentos hídricos (Figura 9a). Contudo, não foi possível destacar nenhum híbrido sob
déficit hídrico em relação a gs.
Houve um aumento considerável de Ci em todos os híbridos quando submetidos
ao estresse (Figura 9b), indicando uma elevada concentração de substrato para serem
utilizados no processo de fotossíntese. No entanto, mesmo com essa elevada
concentração intercelular de CO2 (Ci) , o período de déficit hídrico reduziu os valores Ai
(Figura 10b) em todos os híbridos. Sabe-se que condição de déficit hídrico o Ci
geralmente permanece elevado ou com valores maiores do que aquele esperado quando
ocorre decréscimo na fotossíntese e na condutância estomática. Esse acréscimo no Ci,
frequentemente observado nos estágios mais avançados do déficit hídrico, indica a
predominância de limitações não estomáticas à fotossíntese à medida que o estresse se
torna mais severo (KAISER, 1987). No presente estudo, o decréscimo da taxa de
assimilação de CO2 foi devido não só ao aumento da resistência estomática, mas
também, ao efeito do estresse hídrico na fotossíntese, visto que um alto valor de Ci
associado a baixas condutâncias estomáticas indicaria um decréscimo na eficiência de
carboxilação (SCHULZE et al., 1987). Reduções na fotossíntese de plantas submetidas
à deficiência hídrica podem estar relacionadas à limitação da síntese de ribulose
bifosfato (RuBP), e não à inibição das enzimas no ciclo de Calvin. A limitação da
síntese de RuBP provavelmente está ligada à redução na síntese de ATP, por uma
progressiva inativação de fatores de acoplamento resultante do aumento da
concentração iônica (Mg+2
) e não à redução da capacidade do transporte de elétrons ou
de prótons (LAWLOR & CORNIC, 2002).
Houve redução também no teor de clorofila quando os híbridos foram
submetidos ao estresse por 75 dias na fase de florescimento, exceto para o AS1522 que
não difere estatisticamente do tratamento irrigado (Tabela 6). Entretanto essa maior
quantidade de Chl não levou ao aumento da assimilação de CO2 (Ai) neste híbrido no
tratamento sob estresse, o que pode induzir que houve comprometimento da integridade
do sistema fotossintético pelo déficit hídrico.
Verificou-se redução significativa de Fv/Fm em função do estresse causado pela
deficiência hídrica. Os menores valores de Fv/Fm alcançados foram de 0,53
(DAS2B707) e 0,38 (FT510). BOLHÀR-NORDENKAMPF et al. (1989), relatam que,
quando a planta está com seu aparelho fotossintético intacto, a razão Fv/Fm deve variar
entre 0,75 e 0,85 enquanto uma queda nesta razão reflete a presença de dano nos centros

62
de reação do PSII por algum estresse (BAKER, 1993). Os baixos valores de Fv/Fm
observados caracterizam danos que podem estar relacionados à redução da concentração
intercelular de CO2, em razão do fechamento estomático (BAKER,1993) e devido à
queda na atividade fotossintética (SMIRNOFF, 1993) em função da condição de
estresse causado pelo déficit hídrico. Mesmo apresentando valores maiores de Fv/Fm
sob deficiência hídrica, os híbridos DAS2B710 e AS1522 mostraram valores reduzidos
de Ai (Figura 10b), indicando que o estresse causou dano fotoquímico.
Pode-se observar decréscimos na capacidade da taxa aparente de transporte de
elétrons (ETR) do fotossistema II sob condições de estresse em todos os híbridos
avaliados. Esse resultado indica que a capacidade de transporte de elétrons foi alterada
em função do déficit hídrico.
O híbrido AS1522 destacou-se por apresentar maior quantidade de açúcares
solúveis (AS) tanto no tratamento irrigado quanto no tratamento submetido a restrição
hídrica (Figura 11a). É verificado que sob condições de baixa disponibilidade de água,
ocorre acúmulo de açúcares solúveis (MORGAN, 1984) e simultaneamente diminuição
no teor de amido. Esse acúmulo de AS em AS1522 pode ter ocorrido pela hidrólise do
amido (AM) e/ou devido a uma contínua produção de fotoassimilados pela fotossíntese,
que é menos afetada pelo déficit hídrico do que o crescimento (BOYER, 1970).
Para o teor de amido, verifica-se que houve redução significativa em todos os
híbridos avaliados após o déficit hídrico. Esta redução das concentrações de amido nas
folhas dos híbridos sob estresse está provavelmente relacionada com a redução da
fotossíntese e da degradação do amido, visando a formação de novos açúcares como a
sacarose para realizar o ajuste osmótico e/ou transporte para outros drenos preferenciais
e/ou a inativação da enzima chave na síntese de amido (KRAMER & BOYER, 1995).
Entretanto, houve aumento significativo de SAC nos híbridos DAS2B707 e AS1522
devido ao estresse, enquanto que os teores de SAC mantiveram-se constantes nos
híbridos DAS2710 e FT510 nos dois tratamentos hídricos empregados (Figura 11b). O
aumento das concentrações de sacarose nos híbridos DAS2B707 e AS1522 está ligado,
provavelmente, a quebra do amido pela ação das enzimas α e β-amilase em açúcares que
podem ser quebrados e, então, transportados para os diversos drenos na forma de
sacarose (KINGSTON-SMITH et al. 1999). Além disso, como os demais açúcares, a
sacarose pode se ligar às moléculas de água na folha com a finalidade de manter o nível
de água da folha, bem como induzir um ajustamento osmótico na planta. Por isso,
apresentou-se aumento de sacarose na ocorrência de estresse hídrico.

63
Dentro desse contexto, os resultados obtidos no presente trabalho, permitiram
concluir que o híbrido AS1522 apresenta um acúmulo de solutos como uma resposta ao
déficit hídrico, caracterizando um ajustamento osmótico. Este mecanismo é,
provavelmente, uma das estratégias que, atuando em conjunto com outros fatores,
permitem a planta sobreviver e reproduzir em condições estressantes.
Com relação ao desenvolvimento da planta, os híbridos DAS2B707 e
DAS2B710 apresentaram reduzida estatura quando houve déficit hídrico. No tratamento
sob condição de estresse o DAS2B707 e AS1522 obtiveram bom desempenho em
rendimento (MG), no entanto não foi possível verificar estabilidade de produção nesses
híbridos quando houve déficit hídrico, apresentando perdas significativas na produção
de 25% e 32%, respectivamente. Os híbridos FT510 e DAS2B710 apresentaram
maiores reduções no rendimento, com perdas de 57% e 48%, respectivamente (Tabela
7).
Para MST, verificou-se a ocorrência de estabilidade de produção de massa seca
total no híbrido AS1522 em relação ao tratamento irrigado mesmo após a deficiência
hídrica imposta no estádio de florescimento. Esse resultado pode estar relacionado ao
maior acúmulo de carboidratos na folha que esse híbrido produziu em condições de seca
(Figura 16), induzindo a uma maior biomassa neste híbrido.

64
Figura 16. Representação gráfica da redução relativa da altura da planta (AP), diâmetro
do colmo (DIAM), massa de grãos (MG), massa seca total (MST), condutância
estomática (gs), concentração intercelular de CO2 (Ci), assimilação diurna de CO2 (Ai),
transpiração diurna (Ei), teor de clorofila total (Chl), eficiência quântica potencial do
fotossistema II (Fv/Fm), coeficiente de dissipação não-fotoquímica da fluorescência
(NPQ), coeficiente de dissipação fotoquímica (qP), taxa aparente de transporte de
elétrons (ETR), açúcares solúveis (AS), sacarose (SAC), amido (AM), carboidrato total
não estrutural (CTNE) e eficiência do uso da água (EUA) de quatro híbridos de milho
submetidos a dois tratamentos hídricos a partir do estádio de florescimento (R1).
Valores maiores que 1 indicam aumento na variável específica sob déficit hídrico.
4.2.3 Déficit hídrico no estádo de enchimento de grãos
Somente no híbrido AS1522 apresentou redução do Ψw com valor de -0,40 MPa
após 52 dias sem irrigação (Figura 12), indicando que o défcit hídrico imposto no
estádio de enchimento de grãos foi considerado moderado. Em déficits não muito
severos, as plantas com sistema radicular mais profundo exploram o solo em locais que
ainda apresentam água passível de ser absorvida. Outra possível causa para uma maior
hidratação das plantas poderia ser o fluxo ascendente de água no solo à noite, o qual
seria suficiente para elevar o potencial de base.
O período de suspenção da rega levou ao fechamento estomático dos híbridos
DAS2B707 e AS1522, comportamento no qual pode estar associado a uma rápida
adaptação dessas plantas ao estresse hídrico (MENCUCCINI et al., 2000). Este fato
permite à planta transpirar menos (Figura 14a), controlando a perda de água e
diminuindo os possíveis danos à planta, apesar da redução da assimilação de CO2.
NABLE et al. (1999) também encontraram reduções nas taxas transpiratórias em sorgo
e cana-de-açúcar em condições de déficit hídrico, sendo que nesta última as reduções
foram mais severas, sempre que a fração de água disponível no solo decrescia. O
híbrido FT510 obteve maior gs (Figura 13a) e Ci (Figura 13b), indicando que não houve
restrição de substrato para a realização de fotossíntese devido ao déficit hídrico.
Em condições de ambientes estressantes ocorre o decréscimo característico na
eficiência quântica potencial do fotossistema II, podendo ser detectada pela queda na
relação Fv/Fm (KRAUSE & WEIS, 1991). Entretanto, redução de Fv/Fm devido déficit
hídrico foi verificada apenas no híbrido AS1522 (Tabela 8). O híbrido FT510 destacou-
se por apresentar maior relação Fv/Fm quando comparado com os outros híbridos
submetidos a 52 dias sob déficit hídrico. Seguindo essa mesma tendência, este híbrido

65
apresentou valores elevados de qP e de ETR, refletindo em metabolismo fotossintético
do carbono adequado mesmo quando as condições ambientais se encontravam adversas
devido a restrição hídrica.
Plantas submetidas a déficits hídricos acentuados, as quais estão com os
estômatos fechados, podem apresentar valores significantes de qP devido à reciclagem
interna de CO2. Nesta condição, a fluorescência pode fornecer informações sobre o
rendimento quântico, o fluxo de elétrons e sobre o estado de energia dos cloroplastos,
mesmo que nenhuma fixação do CO2 ou liberação do oxigênio ocorra. Neste caso qP
está relacionado com a estimativa da disponibilidade de dissipação de energia para o
metabolismo do carbono (BOLHÀR-NORDENKAMPF et al, 1989). Os valores de
NPQ permaneceram estáveis nos híbridos nos dois tratamentos hídricos impostos,
indicando ausência de danos no processo fotoquímico nessas plantas mesmo quando
submetidas ao estresse.
O déficit hídrico imposto no estádio de enchimento de grãos não possibilitou a
diferenciação dos híbridos em relação a massa de grãos (MG) e massa seca total (MST),
indicando que o período de estresse não foi considerado adequado para selecionar
híbridos contrastantes à seca (Tabela 9).
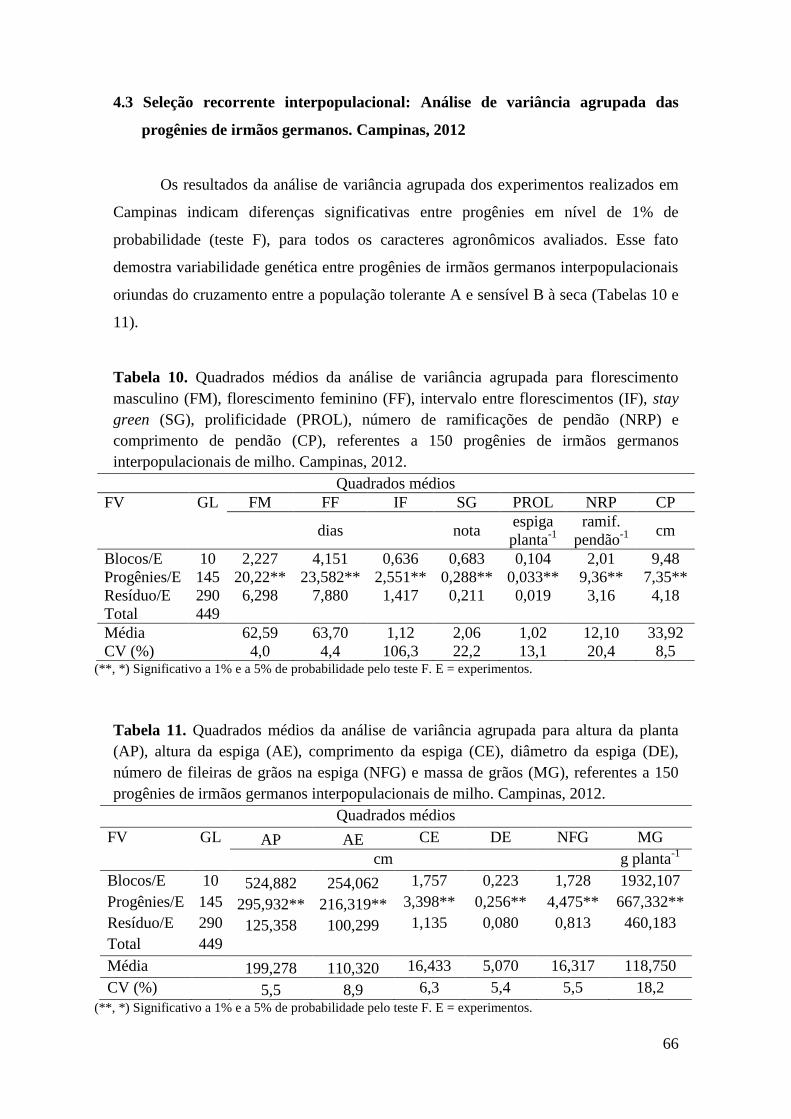
66
4.3 Seleção recorrente interpopulacional: Análise de variância agrupada das
progênies de irmãos germanos. Campinas, 2012
Os resultados da análise de variância agrupada dos experimentos realizados em
Campinas indicam diferenças significativas entre progênies em nível de 1% de
probabilidade (teste F), para todos os caracteres agronômicos avaliados. Esse fato
demostra variabilidade genética entre progênies de irmãos germanos interpopulacionais
oriundas do cruzamento entre a população tolerante A e sensível B à seca (Tabelas 10 e
11).
Tabela 10. Quadrados médios da análise de variância agrupada para florescimento
masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF), stay
green (SG), prolificidade (PROL), número de ramificações de pendão (NRP) e
comprimento de pendão (CP), referentes a 150 progênies de irmãos germanos
interpopulacionais de milho. Campinas, 2012.
Quadrados médios
FV GL FM FF IF SG PROL NRP CP
dias nota
espiga
planta-1
ramif.
pendão-1
cm
Blocos/E 10 2,227 4,151 0,636 0,683 0,104 2,01 9,48
Progênies/E 145 20,22** 23,582** 2,551** 0,288** 0,033** 9,36** 7,35**
Resíduo/E 290 6,298 7,880 1,417 0,211 0,019 3,16 4,18
Total 449
Média 62,59 63,70 1,12 2,06 1,02 12,10 33,92
CV (%) 4,0 4,4 106,3 22,2 13,1 20,4 8,5 (**, *) Significativo a 1% e a 5% de probabilidade pelo teste F. E = experimentos.
Tabela 11. Quadrados médios da análise de variância agrupada para altura da planta
(AP), altura da espiga (AE), comprimento da espiga (CE), diâmetro da espiga (DE),
número de fileiras de grãos na espiga (NFG) e massa de grãos (MG), referentes a 150
progênies de irmãos germanos interpopulacionais de milho. Campinas, 2012.
Quadrados médios
FV GL AP AE CE DE NFG MG
cm g planta-1
Blocos/E 10 524,882 254,062 1,757 0,223 1,728 1932,107
Progênies/E 145 295,932** 216,319** 3,398** 0,256** 4,475** 667,332**
Resíduo/E 290 125,358 100,299 1,135 0,080 0,813 460,183
Total 449
Média 199,278 110,320 16,433 5,070 16,317 118,750
CV (%) 5,5 8,9 6,3 5,4 5,5 18,2
(**, *) Significativo a 1% e a 5% de probabilidade pelo teste F. E = experimentos.

67
O intervalo entre os florescimentos feminino e masculino é considerado um dos
mais importantes caracteres a serem selecionados para a obtenção de genótipos de milho
tolerantes à seca (BÄNZINGER et al., 2000; LI et al., 2003 e XIAO et al., 2004). No
presente estudo as 150 progênies floresceram, em média, 63 e 64 dias após a semeadura
(DAS) para FM e para FF, respectivamente, resultando em baixo intervalo entre os
florescimentos (IF). Resultados semelhantes foram obtidos por PATERNIANI et al.
(2012) e GUIMARÃES et al. (2012) com progênies de meios-irmãos e irmãos
germanos, respectivamente, contendo baixos intervalos entre os florescimentos sob
condições de safrinha.
A média geral da nota de stay green foi de 2,06, sendo que a média do híbrido
considerado tolerante à seca (DAS2B707) foi 2,05. Uma vez que a nota 1 era dada à
planta com stay green e a nota 5 à planta sem stay green, as progênies apresentaram, em
média, um bom desempenho para o caráter, conferindo maior fotossíntese ao longo do
ciclo e fornecendo carboidratos para colmos, folhas e raízes. O stay green é um caráter
descrito em diversas espécies e vem sendo objeto de grande atenção por estar
relacionado à resistência ao estresse hídrico, ao aumento da tolerância a pragas e
doenças, redução do acamamento de plantas, tolerância ao maior adensamento, tendo
sido relacionado ao aumento da produtividade direta e indiretamente (DUVICK &
CASSMAN, 1999). Devido a isso, este caráter é considerado de muita importância nos
programas de melhoramento. De acordo com COSTA et al. (2008), os genótipos são
considerados com stay green quando as folhas e colmos permanecem verdes após o
estádio fenológico de enchimento de grãos. Dessa forma, a fotossíntese se prolonga
fornecendo carboidratos para as plantas, que apresentam maior resistência a estresses
abióticos como o de déficit hídrico (JIANG et al., 2004 e CARMO et al., 2007).
Segundo BOLAÑOS & EDMEADES (1993a), genótipos de milho prolíficos
demonstram melhor capacidade de adaptação a uma ampla gama de condições
estressantes como aumento da densidade de semeadura e a tolerância ao déficit hídrico.
A média das 150 progênies de IG interpopulacionais para o caráter PROL foi 1,02
espigas planta-1
, valor este maior que o encontrado por CÂMARA et al. (2007) que
obtiveram 0,70 a 0,83 espigas planta-1
em progênies F2:3 obtidas da população D e U,
respectivamente.
As médias de progênies de IG interpopulacionais para os caracteres NRP e CP
foram, respectivamente, 12,10 ramificações pendão-1
e 33,92 cm. Esses valores sugerem

68
que, em geral, as progênies apresentam média ramificação dos pendões e comprimento
de pendão considerado de tamanho médio.
Para massa de grãos (MG), as progênies se comportaram relativamente de forma
diferenciada, o que demonstra a possibilidade de selecionar híbridos promissores em
condições de safrinha. A média de produtividade das progênies de irmãos germanos foi
de 118,75 g planta-1
, resultado semelhante à média da testemunha tolerante à seca
DAS2B707, que foi 127,12 g planta-1
. De maneira geral, as progênies apresentaram
valores de MG acima das testemunhas empregadas nos experimentos, com amplitudes
de massa de grãos variando de 67,78 a 149,64 g planta-1
no Experimento I (Anexo 2), de
86,82 a 148,14 g planta-1
no Experimento II (Anexo 4), de 102,097 a 156,819 g planta-1
no Experimento III (Anexo 6), de 91,43 a 135,29 g planta-1
no Experimento IV (Anexo
8) e de 61,58 a 143,08 g planta-1
no Experimento V (Anexo 10).
Pôde-se verificar uma tendência das progênies mais produtivas apresentarem
AP, AE, CE, DE e NFG elevados em condições de safrinha.
4.4 Análise de variância agrupada das progênies de irmãos germanos. Mococa,
2012
Pela análise de variância foi possível verificar diferenças significativas entre
progênies (P<0,01) para todos os caracteres avaliados, exceto para IF. A existência de
significância indica que os cruzamentos realizados entre as duas populações
contrastantes quanto à tolerância à seca produziram progênies contendo alta
variabilidade, característica ideal para a condução de um programa de melhoramento
empregando o método da seleção recorrente recíproca, sendo possível que ganhos sejam
obtidos a partir do melhoramento dessas populações (Tabelas 12 e 13).
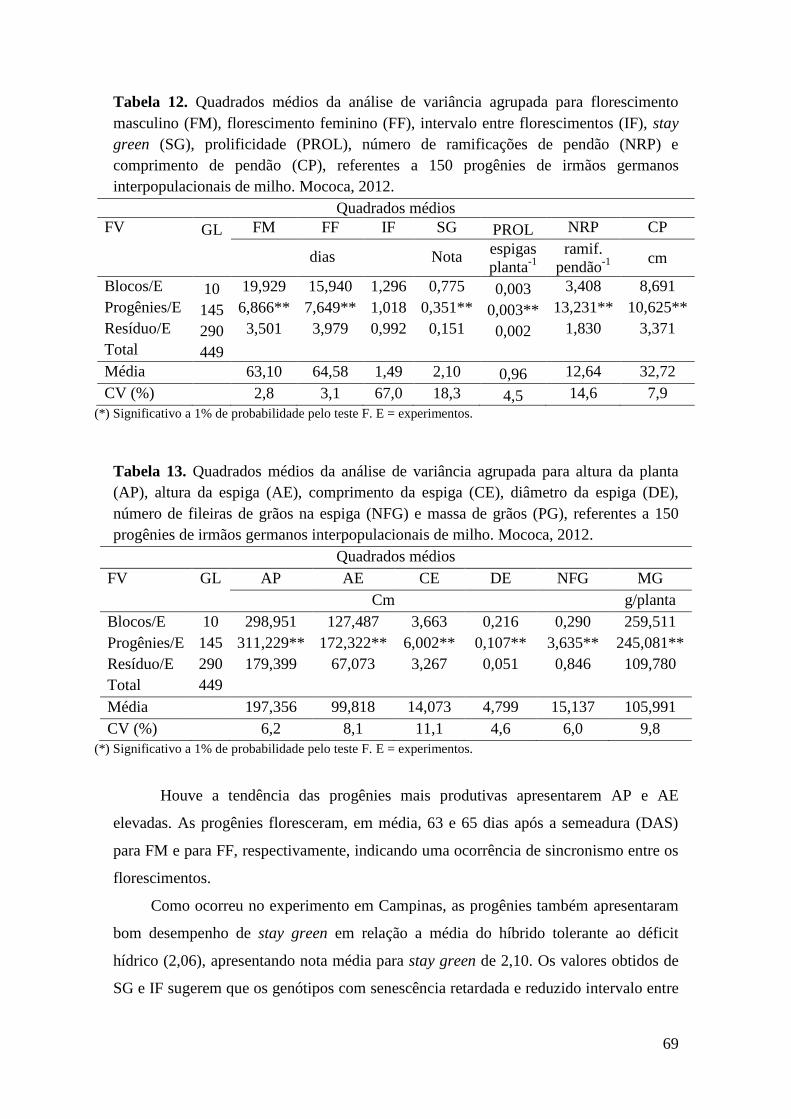
69
Tabela 12. Quadrados médios da análise de variância agrupada para florescimento
masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF), stay
green (SG), prolificidade (PROL), número de ramificações de pendão (NRP) e
comprimento de pendão (CP), referentes a 150 progênies de irmãos germanos
interpopulacionais de milho. Mococa, 2012.
Quadrados médios
FV GL FM FF IF SG PROL NRP CP
dias Nota espigas
planta-1
ramif.
pendão-1
cm
Blocos/E 10 19,929 15,940 1,296 0,775 0,003 3,408 8,691
Progênies/E 145 6,866** 7,649** 1,018 0,351** 0,003** 13,231** 10,625**
Resíduo/E 290 3,501 3,979 0,992 0,151 0,002 1,830 3,371
Total 449
Média 63,10 64,58 1,49 2,10 0,96 12,64 32,72
CV (%) 2,8 3,1 67,0 18,3 4,5 14,6 7,9
(*) Significativo a 1% de probabilidade pelo teste F. E = experimentos.
Tabela 13. Quadrados médios da análise de variância agrupada para altura da planta
(AP), altura da espiga (AE), comprimento da espiga (CE), diâmetro da espiga (DE),
número de fileiras de grãos na espiga (NFG) e massa de grãos (PG), referentes a 150
progênies de irmãos germanos interpopulacionais de milho. Mococa, 2012.
Quadrados médios
FV GL AP AE CE DE NFG MG
Cm g/planta
Blocos/E 10 298,951 127,487 3,663 0,216 0,290 259,511
Progênies/E 145 311,229** 172,322** 6,002** 0,107** 3,635** 245,081**
Resíduo/E 290 179,399 67,073 3,267 0,051 0,846 109,780
Total 449
Média 197,356 99,818 14,073 4,799 15,137 105,991
CV (%) 6,2 8,1 11,1 4,6 6,0 9,8
(*) Significativo a 1% de probabilidade pelo teste F. E = experimentos.
Houve a tendência das progênies mais produtivas apresentarem AP e AE
elevadas. As progênies floresceram, em média, 63 e 65 dias após a semeadura (DAS)
para FM e para FF, respectivamente, indicando uma ocorrência de sincronismo entre os
florescimentos.
Como ocorreu no experimento em Campinas, as progênies também apresentaram
bom desempenho de stay green em relação a média do híbrido tolerante ao déficit
hídrico (2,06), apresentando nota média para stay green de 2,10. Os valores obtidos de
SG e IF sugerem que os genótipos com senescência retardada e reduzido intervalo entre

70
florescimentos podem ser menos afetados pelo déficit hídrico, concordando com os
resultados obtidos por CHAPMAN & EDMEADES (1999) e PATERNIANI et al.
(2012).
As médias de progênies para os caracteres PROL, NRP e de CP foram,
respectivamente, 0,96 espigas planta-1
, 12,64 ramificações pendão-1
e 32,71 cm. Esses
resultados obtidos pelas progênies conduzidas em Mococa apresentam baixa
prolificidade, média ramificação dos pendões e médio comprimento de pendão.
Por meio dos resultados observados, verificou-se que houve o desenvolvimento de
progênies promissoras quanto a praticamente todas as características agronômicas
avaliadas, principalmente quanto à produtividade de grãos com valores médios de
105,99 g planta-1
, sendo que o híbrido DAS2B707 obteve em média produção de 111,93
g planta-1
. Os valores de massa de grãos (MG) variaram de 83,54 a 126,35 g planta-1
no
Experimento I (Anexo 12), de 90,64 a 118,42 g planta-1
no Experimento II (Anexo 14),
de 95,72 a 136,08 g planta-1
no Experimento III (Anexo 16), de 79,08 a 116,65 g planta-
1 no Experimento IV (Anexo 18) e de 93,08 a 121,35 g planta
-1 no Experimento V
(Anexo 20). Tal resultado indica a eficiência do cruzamento entre as populações
contrastantes quanto à tolerância à seca para o desenvolvimento de progênies de IG
interpopulacionais com características de tolerância ao déficit hídrico, em condições de
safrinha.
4.5 Análise de variância conjunta de locais e agrupada das progênies de irmãos
germanos, Campinas e Mococa, 2012
Na análise de variância conjunta agrupada dos cinco experimentos de Campinas e
Mococa, verificou-se diferença significativa (P<0,01) entre progênies para todos os
caracteres avaliados, o que revela a presença de variabilidade genética entre as
progênies de irmãos germanos. A fonte de variação ambientes não apresentou
significância somente para SG, confirmando a grande diferença edafoclimática entre
Campinas e Mococa. Foram constatados efeitos significativos da interação entre
progênies x ambientes para todos os caracteres avaliados, indicando que essas progênies
possuem desempenho diferenciado face às variações ambientais, o que caracteriza a
importância de realizar a seleção por local (Tabelas 14 e 15).
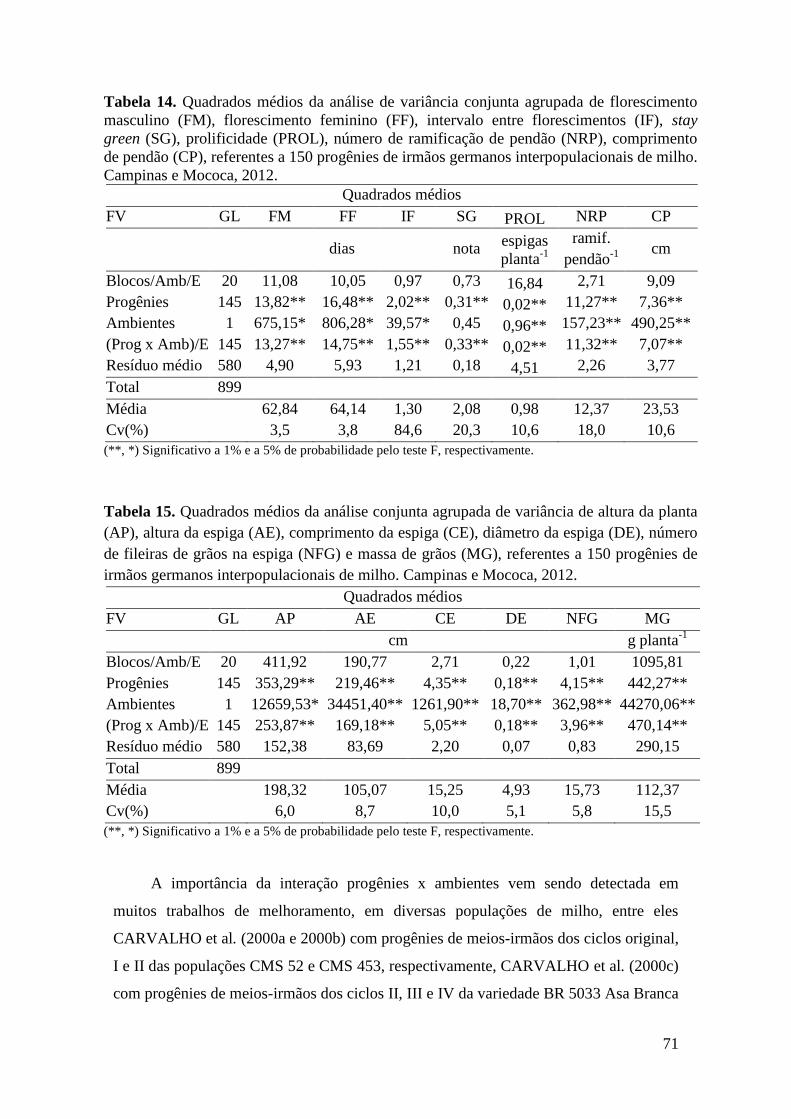
71
Tabela 14. Quadrados médios da análise de variância conjunta agrupada de florescimento
masculino (FM), florescimento feminino (FF), intervalo entre florescimentos (IF), stay
green (SG), prolificidade (PROL), número de ramificação de pendão (NRP), comprimento
de pendão (CP), referentes a 150 progênies de irmãos germanos interpopulacionais de milho.
Campinas e Mococa, 2012.
Quadrados médios
FV GL FM FF IF SG PROL NRP CP
dias nota espigas
planta-1
ramif.
pendão-1
cm
Blocos/Amb/E 20 11,08 10,05 0,97 0,73 16,84 2,71 9,09
Progênies 145 13,82** 16,48** 2,02** 0,31** 0,02** 11,27** 7,36**
Ambientes 1 675,15* 806,28* 39,57* 0,45 0,96** 157,23** 490,25**
(Prog x Amb)/E 145 13,27** 14,75** 1,55** 0,33** 0,02** 11,32** 7,07**
Resíduo médio 580 4,90 5,93 1,21 0,18 4,51 2,26 3,77
Total 899
Média 62,84 64,14 1,30 2,08 0,98 12,37 23,53
Cv(%) 3,5 3,8 84,6 20,3 10,6 18,0 10,6
(**, *) Significativo a 1% e a 5% de probabilidade pelo teste F, respectivamente.
Tabela 15. Quadrados médios da análise conjunta agrupada de variância de altura da planta
(AP), altura da espiga (AE), comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG) e massa de grãos (MG), referentes a 150 progênies de
irmãos germanos interpopulacionais de milho. Campinas e Mococa, 2012.
Quadrados médios
FV GL AP AE CE DE NFG MG
cm g planta-1
Blocos/Amb/E 20 411,92 190,77 2,71 0,22 1,01 1095,81
Progênies 145 353,29** 219,46** 4,35** 0,18** 4,15** 442,27**
Ambientes 1 12659,53* 34451,40** 1261,90** 18,70** 362,98** 44270,06**
(Prog x Amb)/E 145 253,87** 169,18** 5,05** 0,18** 3,96** 470,14**
Resíduo médio 580 152,38 83,69 2,20 0,07 0,83 290,15
Total 899
Média 198,32 105,07 15,25 4,93 15,73 112,37
Cv(%) 6,0 8,7 10,0 5,1 5,8 15,5
(**, *) Significativo a 1% e a 5% de probabilidade pelo teste F, respectivamente.
A importância da interação progênies x ambientes vem sendo detectada em
muitos trabalhos de melhoramento, em diversas populações de milho, entre eles
CARVALHO et al. (2000a e 2000b) com progênies de meios-irmãos dos ciclos original,
I e II das populações CMS 52 e CMS 453, respectivamente, CARVALHO et al. (2000c)
com progênies de meios-irmãos dos ciclos II, III e IV da variedade BR 5033 Asa Branca

72
em dois locais e CÂMARA et al. (2007) com progênies de meios-irmãos oriundas da
População D e U avaliadas em sete a nove locais.
Os coeficientes de variação (CV) das análises conjuntas agrupadas foram, em
geral, de magnitude baixa a mediana, segundo a classificação proposta por SCAPIM et
al. (1995). Esse resultado evidencia boa precisão na tomada dos dados dos
experimentos, o que é bastante desejável, porque permite a obtenção de melhor resposta
à seleção. Valores de CV menores que 10% foram observados para os caracteres FM,
FF, CP, AP, AE, CE, DE e NFG, enquanto que foram considerados medianos para SG
(20,3%), PROL (10,6%), NRP (18,0%) e para MG (15,1%). O elevado valor de CV
observado para IF (84,6%) é resultado da própria natureza do caráter cuja média,
formada por valores positivos, nulos e negativos, tende a ser muito baixa favorecendo
valor elevado de CV.
De maneira geral, a análise de variância conjunta confirmou a alta variabilidade
genética das 150 progênies de irmãos germanos para todos os caracteres avaliados. Esse
resultado indica ser possível obter ganhos com seleção nas progênies para os caracteres
estudados. Os ambientes foram apropriados para evidenciar o comportamento
diferenciado das progênies em face as oscilações ambientais, exceto para a variável SG.
Nas Tabelas 16 e 17 são apresentadas as 20% melhores progênies selecionadas em
relação à massa de grãos (MG).

73
Tabela 16. Médias conjuntas de florescimento masculino (FM), florescimento feminino
(FF), intervalo entre florescimentos (IF), stay green (SG), altura da planta (AP), altura
da espiga (AE), massa de grãos (MG) e porcentagem de massa de grãos (MG%),
referentes às 30 progênies de irmãos germanos interpopulacionais selecionadas e de
quatro testemunhas comerciais de milho. Campinas e Mococa, 2012.
Progênies FM FF IF SG AP AE MG MG
dias nota cm g planta-1
(%)
9B x 7A 61,00 62,33 1,33 2,10 208 115 134,55 112,58
140A x 153B 64,33 65,33 1,00 1,67 211 109 133,03 111,30
156A x 150B 63,33 64,83 1,50 2,07 209 117 131,88 110,34
20A x 24B 64,00 65,50 1,50 1,87 210 111 131,56 110,07
141A x 149B 61,83 63,50 1,67 1,77 204 107 131,38 109,92
127A x 110B 63,17 63,83 0,67 2,40 207 117 130,83 109,46
119A x 84B 64,17 65,33 1,17 2,17 203 117 127,85 106,97
19A x 37B 58,50 59,00 0,50 1,70 195 94 127,65 106,80
67B x 93A 64,17 64,50 0,33 1,57 202 104 127,47 106,65
73A x 41B 57,17 58,17 1,00 2,10 183 91 126,63 105,95
142A x 148B 62,33 63,50 1,17 2,40 205 110 126,27 105,65
97B x 58A 61,67 62,50 0,83 2,20 206 105 125,90 105,34
31B x 34A 62,33 63,67 1,33 2,20 199 99 125,63 105,11
148A x 135B 62,50 63,67 1,17 1,87 198 102 125,42 104,94
159B x 161A 62,50 63,00 0,50 1,93 208 109 125,37 104,89
97A x 81B 64,50 66,17 1,67 2,27 203 108 125,07 104,64
79B x 88A 63,50 64,33 0,83 2,00 210 118 124,97 104,56
51B x 27A 63,50 64,83 1,33 2,13 200 107 124,90 104,50
83A x 77B 62,33 64,00 1,67 1,90 204 109 124,73 104,36
52A x 49B 63,33 63,83 0,50 1,87 204 109 123,76 103,55
68A x 75B 63,33 64,83 1,50 2,10 195 103 123,69 103,49
176A x 171B 62,67 63,00 0,33 2,07 193 100 123,61 103,42
176B x 170A 62,50 64,00 1,50 2,20 197 110 123,13 103,02
138B x 118A 64,00 65,50 1,50 1,60 207 111 122,97 102,89
27A x 51B 62,67 63,00 0,33 2,63 190 104 122,58 102,56
12B x 11A 64,17 66,00 1,83 1,57 214 116 122,33 102,35
69A x 100B 61,50 61,83 0,33 2,03 205 110 122,01 102,08
163B x 158A 63,50 64,33 0,83 1,80 198 108 121,95 102,03
124B x 76A 65,00 65,50 0,50 1,73 213 120 121,87 101,97
47A x 168B 64,00 65,50 1,50 2,20 200 112 120,97 101,21
Média Prog. 62,78 63,84 1,06 2,00 203 108 126,00 105,42
Pop. tolerante 64,47 65,63 1,17 1,99 185 95 99,33 83,11
Pop. sensível 66,07 67,87 1,80 1,97 196 109 86,30 72,21
FT510 65,50 67,37 2,16 1,92 161 83 74,26 62,13
DAS2B707 63,80 64,60 0,80 1,79 192 99 119,52 100,00
Média Test. 64,96 66,37 1,48 1,92 184 97 94,85 79,36 *Porcentagem em relação à testemunha tolerante DAS2B707.
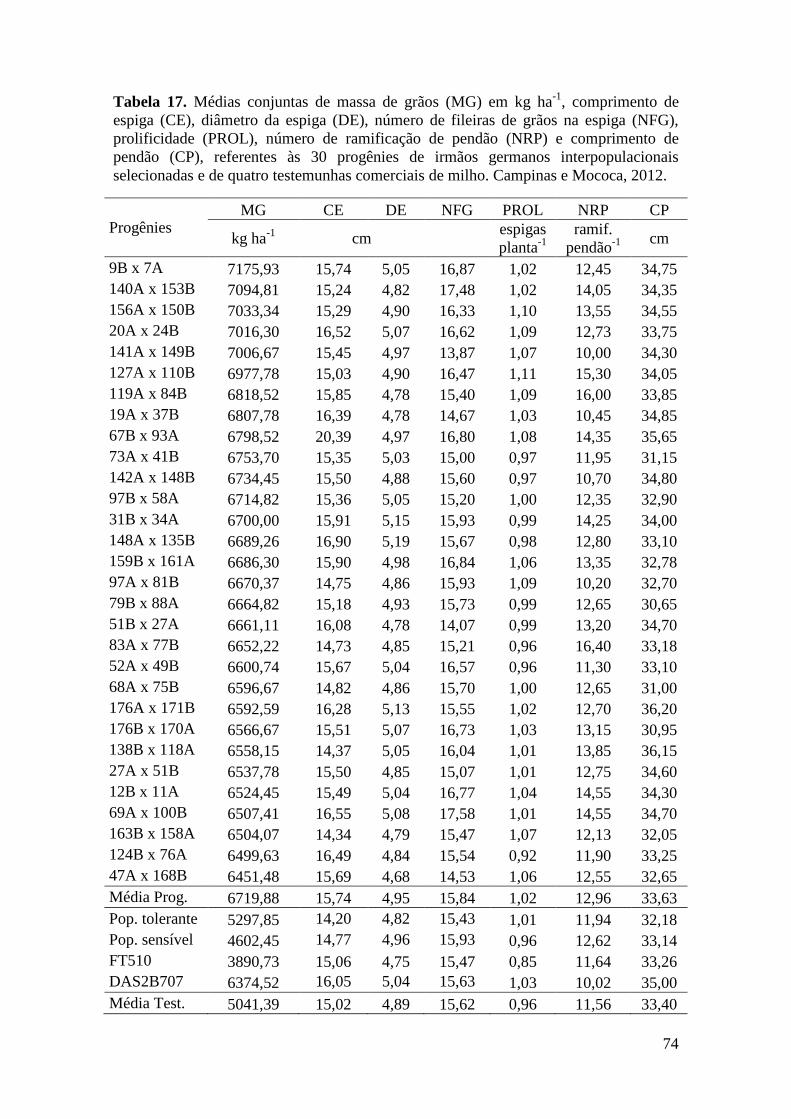
74
Tabela 17. Médias conjuntas de massa de grãos (MG) em kg ha-1
, comprimento de
espiga (CE), diâmetro da espiga (DE), número de fileiras de grãos na espiga (NFG),
prolificidade (PROL), número de ramificação de pendão (NRP) e comprimento de
pendão (CP), referentes às 30 progênies de irmãos germanos interpopulacionais
selecionadas e de quatro testemunhas comerciais de milho. Campinas e Mococa, 2012.
Progênies MG CE DE NFG PROL NRP CP
kg ha-1
cm espigas
planta-1
ramif.
pendão-1
cm
9B x 7A 7175,93 15,74 5,05 16,87 1,02 12,45 34,75
140A x 153B 7094,81 15,24 4,82 17,48 1,02 14,05 34,35
156A x 150B 7033,34 15,29 4,90 16,33 1,10 13,55 34,55
20A x 24B 7016,30 16,52 5,07 16,62 1,09 12,73 33,75
141A x 149B 7006,67 15,45 4,97 13,87 1,07 10,00 34,30
127A x 110B 6977,78 15,03 4,90 16,47 1,11 15,30 34,05
119A x 84B 6818,52 15,85 4,78 15,40 1,09 16,00 33,85
19A x 37B 6807,78 16,39 4,78 14,67 1,03 10,45 34,85
67B x 93A 6798,52 20,39 4,97 16,80 1,08 14,35 35,65
73A x 41B 6753,70 15,35 5,03 15,00 0,97 11,95 31,15
142A x 148B 6734,45 15,50 4,88 15,60 0,97 10,70 34,80
97B x 58A 6714,82 15,36 5,05 15,20 1,00 12,35 32,90
31B x 34A 6700,00 15,91 5,15 15,93 0,99 14,25 34,00
148A x 135B 6689,26 16,90 5,19 15,67 0,98 12,80 33,10
159B x 161A 6686,30 15,90 4,98 16,84 1,06 13,35 32,78
97A x 81B 6670,37 14,75 4,86 15,93 1,09 10,20 32,70
79B x 88A 6664,82 15,18 4,93 15,73 0,99 12,65 30,65
51B x 27A 6661,11 16,08 4,78 14,07 0,99 13,20 34,70
83A x 77B 6652,22 14,73 4,85 15,21 0,96 16,40 33,18
52A x 49B 6600,74 15,67 5,04 16,57 0,96 11,30 33,10
68A x 75B 6596,67 14,82 4,86 15,70 1,00 12,65 31,00
176A x 171B 6592,59 16,28 5,13 15,55 1,02 12,70 36,20
176B x 170A 6566,67 15,51 5,07 16,73 1,03 13,15 30,95
138B x 118A 6558,15 14,37 5,05 16,04 1,01 13,85 36,15
27A x 51B 6537,78 15,50 4,85 15,07 1,01 12,75 34,60
12B x 11A 6524,45 15,49 5,04 16,77 1,04 14,55 34,30
69A x 100B 6507,41 16,55 5,08 17,58 1,01 14,55 34,70
163B x 158A 6504,07 14,34 4,79 15,47 1,07 12,13 32,05
124B x 76A 6499,63 16,49 4,84 15,54 0,92 11,90 33,25
47A x 168B 6451,48 15,69 4,68 14,53 1,06 12,55 32,65
Média Prog. 6719,88 15,74 4,95 15,84 1,02 12,96 33,63
Pop. tolerante 5297,85 14,20 4,82 15,43 1,01 11,94 32,18
Pop. sensível 4602,45 14,77 4,96 15,93 0,96 12,62 33,14
FT510 3890,73 15,06 4,75 15,47 0,85 11,64 33,26
DAS2B707 6374,52 16,05 5,04 15,63 1,03 10,02 35,00
Média Test. 5041,39 15,02 4,89 15,62 0,96 11,56 33,40

75
O caráter intervalo entre florescimento feminino e masculino (IF) é considerado
indicador para selecionar genótipos com tolerância ao déficit hídrico (BOLAÑOS &
EDMEADES 1993b; DURÃES et al., 1997) e prestou-se para discriminar as progênies
estudadas. O baixo IF apresentado pelas progênies selecionadas, indica uma adaptação
ao déficit hídrico, evitando a característica de protandria no milho, que é acentuada sob
condições de déficit hídrico pelo maior retardamento do florescimento feminino em
relação ao masculino. Desse modo, dependendo da extensão do período de seca, a
produção de grãos tende a ser afetada em função da assincronia entre liberação de pólen
e dos estilo-estigmas, causando perda na produtividade.
As progênies também apresentaram bom desempenho de stay green, resultando
em um melhor aproveitamento da fotossíntese para o fornecimento de carboidratos para
as plantas que, provavelmente, possuem uma maior tolerância à seca.
O número de ramificação de pendão (NRP) das progênies foi considerado médio
a baixo, apresentando 12,4 ramificações. FISCHER et al. (1987), realiazando seis ciclos
de seleção recorrente em três populações, observaram que o NRP foi reduzido de 18
para 9 ramificações em média.
Quanto à média de massa de grãos (MG) das progênies, houve elevados valores
dessa variável em comparação com as quatro testemunhas empregadas. Os valores das
amplitudes de MG variaram de 84,96 a 124,73 g planta-1
no Experimento I (Anexo 22),
de 98,06 a 133,03 g planta-1
no Experimento II (Anexo 24), de 105,13 a 134,55 g planta-
1 no Experimento III (Anexo 26), de 89,74 a 120,97 g planta
-1 no Experimento IV
(Anexo 28) e de 83,24 a 126,63 g planta-1
no Experimento V (Anexo 30).
De modo geral, observou-se efeito significativo entre as progênies de irmãos
germanos para todas as características avaliadas, o que indica existência de
variabilidade genética na população. Outro resultado que reforça a existência de
variabilidade são as estimativas das herdabilidades, dos coeficientes de variação
genético e da relação entre o coeficiente de variação genética e o coeficiente de variação
experimental. A herdabilidade indica quanto da variação fenotípica é devida a efeitos
genéticos. Quanto mais próximo de 100%, menos o caráter é influenciado pelo ambiente
ou que a maior parte da variação é ocasionada por fatores genéticos com efeitos aditivos
(efeito médio dos genes) ou não-aditivos (efeitos de dominância e/ou epistasia).
Nas Tabelas 18 e 19 são apresentadas as estimativas dos componentes de
variância e da herdabilidade em nível de médias para os caracteres avaliados. Foram
obtidos elevados a médios coeficientes de herdabilidade no sentido amplo para NFG

76
(0,80), NRP (0,78), DE (0,65), FM (0,65), FF (0,64), AE (0,62), CP (0,58) e para AP
(0,57), indicando alto a médio controle genético nesses caracteres, condição favorável
para a seleção (FLINT-GARCIA et al., 2005). Para CE (0,49), SG (0,42), IF (0,40),
PROL (0,39) e MG (0,34) os coeficientes de herdabilidade indicam baixo controle
genético, indicando que estes caracteres são governados por muitos genes
(quantitativos) e sofrem muita influência do ambiente. As estimativas de herdabilidade
ampla de progênies apresentadas no presente trabalho são difícies de serem comparadas
com outras relatadas na literatura, uma vez que, as estimativas dos valores de
herdabilidade tendem a variar com o tipo de progênie, número de ambientes
empregados e números de repetições para a estimação do parâmetro. Além disso, na
grande maioria dos artigos as estimativas de herdabilidade são obtidas sob condições
favoráveis da cultura do milho, em semeadura de verão ou primeira safra. O baixo valor
de herdabilidade obtido para MG no presente trabalho pode ser em função do estresse
hídrico que ocorreu nas fases fenológicas mais críticas da cultura.
Segundo CÂMARA et al. (2007), é interessante empregar caracteres secundários
que apresentem herdabilidade elevada, com pouca alteração entre os diferentes níveis de
umidade e que estejam relacionados com a produtividade, pois estes caracteres podem
auxiliar na seleção. Os mesmos autores selecionando caracteres relacionados à
tolerância à deficiência hídrica em milho tropical, verificaram valores elevados de
herdabilidade em progênies de milho F2:3 para os caracteres massa de grãos (86,79 a
94,84% nas populações D e U, respectivamente), florescimento masculino (94,12%),
florescimento feminino (95,88%), número de ramificação de pendão (76,58%) e para
intervalo entre florescimento (76,58%), sendo portanto, caracteres considerados
adequados à seleção para tolerância à seca. Porém, destaca-se que o trabalho de
CÂMARA et al. (2007) foi desenvolvido na safra de verão, diferentemente do presente
trabalho, o qual foi conduzido em condições de safrinha.
A estimativa do componente da variância de progênies ( 2
pσ ) e o ganho genético
(Gs%) para MG foram considerados relativamente baixos. SOUZA JUNIOR & PINTO
(2000) obtiveram ganho de 7,2% para a característica produtividade empregando o
método de seleção recorrente em progênies de meios-irmãos, enquanto, TARDIN et al.
(2003) e GABRIEL (2009), verificaram ganhos de até 14,58% e 14,10% no oitavo e no
décimo ciclo de seleção recorrente em famílias de irmãos completos de milho,
respectivamente. REIS (2009), avaliando populações derivadas de híbridos simples

77
comerciais de milho empregando a seleção recorrente recíproca, mostraram que o
método foi eficiente para promover o progresso genético em 7,9% por ciclo para
produtividade de espigas despalhadas nos híbridos interpopulacionais obtidos a partir de
populações F2 de híbridos simples de milho. No presente estudo o ganho previsto para
MG foi de 1,25%. Ressalta-se que essas progênies foram semeadas em condições
adversas de irrigação (safrinha), enfrentando períodos de seca em pelo menos um dos
estádios fenológicos da cultura (Figura 2 e 3), principalmente na localidade de Mococa.
Observaram-se para 2
pσ valores bem próximos àqueles obtidos para a variância
da interação progênies x ambientes ( 2
a x pσ ) para alguns caracteres como FM, FF, PROL,
NRP, CP, DE, NEG e MG (Tabelas 18 e 19). No entanto, a variância de progênies foi
maior que a variância da interação genótipo por ambiente para IF, AP e AE. Essas
diferenças entre progênies para tais caracteres são decorrentes da constituição genética
diferencial. Entretanto, estimativas de 2
Eσ e de 2
a x pσ foram de magnitude elevada para
MG. Esse resultado está de acordo com o comportamento esperado para o caráter, em
que boa parte da sua variação fenotípica tem sido atribuída a influências ambientais. Os
caracteres SG, DE e IF tiveram, proporcionalmente, as menores estimativas de 2
a x pσ ,
indicando pouca influência das condições ambientais sobre eles.
A presença de variabilidade genética também pode ser confirmada e
quantificada pelo coeficiente de variação genética (CVg), que expressa a magnitude da
variação genética em relação à média do caráter (RESENDE, 1991). Segundo
SEBBENN et al. (1998), coeficientes de variação genética acima de 7% são
considerados altos. No presente estudo, os coeficientes de variação genética para os
caracteres NRP, IF e SG e foram considerados elevados (67,6, 28,3 e 7,1%,
respectivamente), indicando a existência de variação genética herdável na população, o
que possibilita maiores ganhos genéticos para tais caracteres. Os coeficientes de
variação genética para NFG (4,73%), AE (4,53%) e MG (4,48%) foram medianos e
baixos para CE (3,92%), AP (2,92%), DE (2,85%), CP (2,81%), FF (2,07%) e FM
(1,94%). A existência de variação genética indica a possibilidade de melhoramento para
esses caracteres com a obtenção de ganhos consideráveis com a seleção.
Somente para as variáveis NRP e NFG os valores de índice de seleção (b) foram
maiores e próximos a um (3,75 e 0,82, respectivamente). Segundo VENCOVSKY &
BARRIGA (1992), uma situação favorável para a seleção entre progênies de milho é
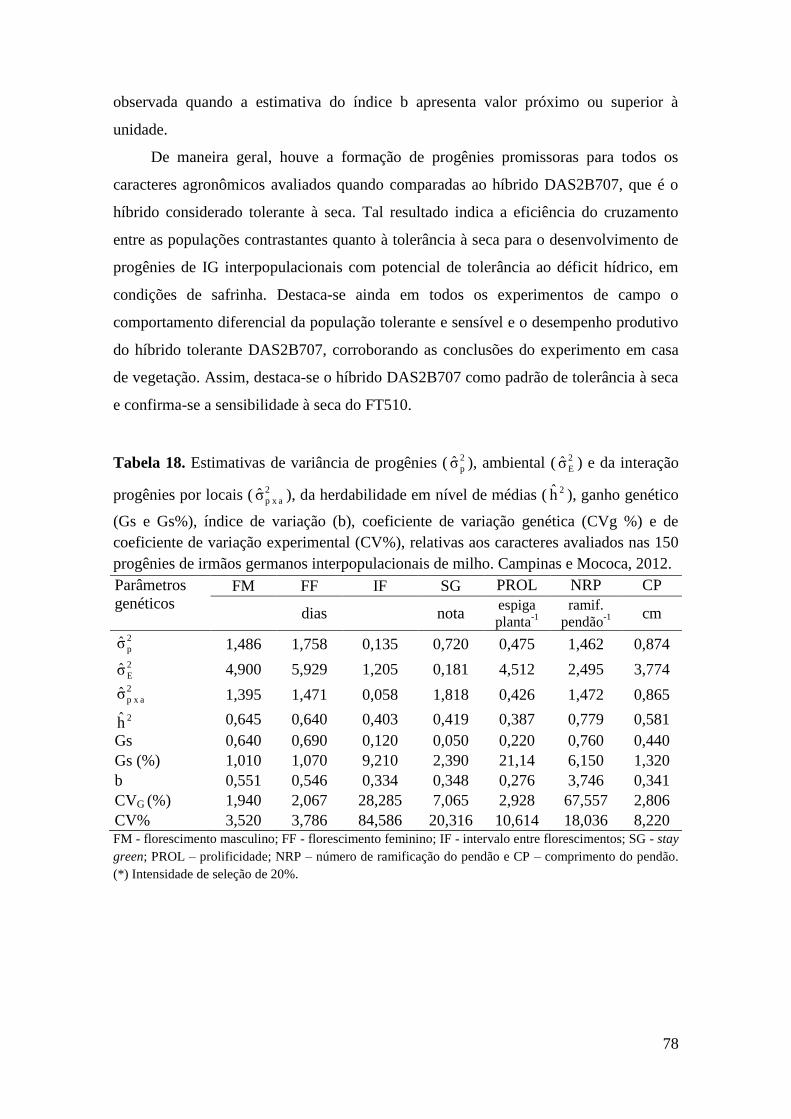
78
observada quando a estimativa do índice b apresenta valor próximo ou superior à
unidade.
De maneira geral, houve a formação de progênies promissoras para todos os
caracteres agronômicos avaliados quando comparadas ao híbrido DAS2B707, que é o
híbrido considerado tolerante à seca. Tal resultado indica a eficiência do cruzamento
entre as populações contrastantes quanto à tolerância à seca para o desenvolvimento de
progênies de IG interpopulacionais com potencial de tolerância ao déficit hídrico, em
condições de safrinha. Destaca-se ainda em todos os experimentos de campo o
comportamento diferencial da população tolerante e sensível e o desempenho produtivo
do híbrido tolerante DAS2B707, corroborando as conclusões do experimento em casa
de vegetação. Assim, destaca-se o híbrido DAS2B707 como padrão de tolerância à seca
e confirma-se a sensibilidade à seca do FT510.
Tabela 18. Estimativas de variância de progênies ( 2
pσ ), ambiental ( 2
Eσ ) e da interação
progênies por locais ( 2
a x pσ ), da herdabilidade em nível de médias ( 2h ), ganho genético
(Gs e Gs%), índice de variação (b), coeficiente de variação genética (CVg %) e de
coeficiente de variação experimental (CV%), relativas aos caracteres avaliados nas 150
progênies de irmãos germanos interpopulacionais de milho. Campinas e Mococa, 2012.
Parâmetros
genéticos FM FF IF SG PROL NRP CP
dias nota espiga
planta-1
ramif.
pendão-1
cm
2
pσ 1,486 1,758 0,135 0,720 0,475 1,462 0,874
2
Eσ 4,900 5,929 1,205 0,181 4,512 2,495 3,774 2
a x pσ 1,395 1,471 0,058 1,818 0,426 1,472 0,865
2h 0,645 0,640 0,403 0,419 0,387 0,779 0,581
Gs 0,640 0,690 0,120 0,050 0,220 0,760 0,440
Gs (%) 1,010 1,070 9,210 2,390 21,14 6,150 1,320
b 0,551 0,546 0,334 0,348 0,276 3,746 0,341
CVG (%) 1,940 2,067 28,285 7,065 2,928 67,557 2,806
CV% 3,520 3,786 84,586 20,316 10,614 18,036 8,220 FM - florescimento masculino; FF - florescimento feminino; IF - intervalo entre florescimentos; SG - stay
green; PROL – prolificidade; NRP – número de ramificação do pendão e CP – comprimento do pendão.
(*) Intensidade de seleção de 20%.
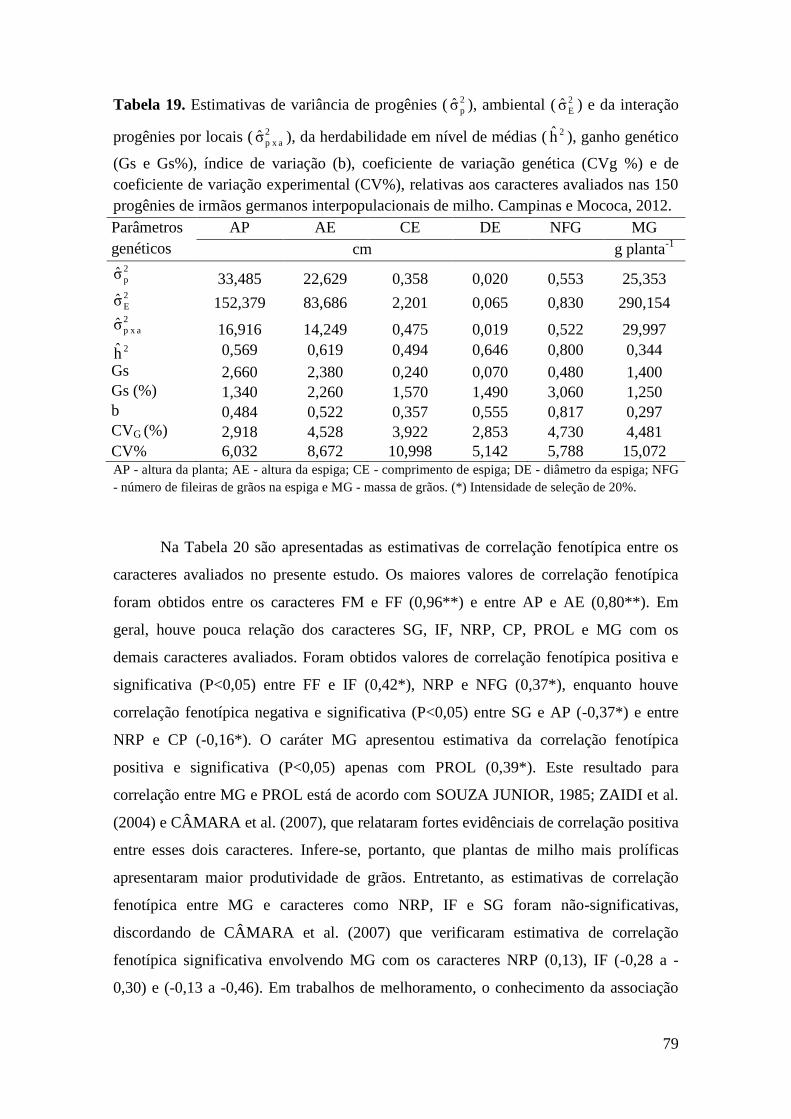
79
Tabela 19. Estimativas de variância de progênies ( 2
pσ ), ambiental ( 2
Eσ ) e da interação
progênies por locais ( 2
a x pσ ), da herdabilidade em nível de médias ( 2h ), ganho genético
(Gs e Gs%), índice de variação (b), coeficiente de variação genética (CVg %) e de
coeficiente de variação experimental (CV%), relativas aos caracteres avaliados nas 150
progênies de irmãos germanos interpopulacionais de milho. Campinas e Mococa, 2012.
Parâmetros
genéticos
AP AE CE DE NFG MG
cm g planta-1
2
pσ 33,485 22,629 0,358 0,020 0,553 25,353 2
Eσ 152,379 83,686 2,201 0,065 0,830 290,154 2
a x pσ 16,916 14,249 0,475 0,019 0,522 29,997 2h 0,569 0,619 0,494 0,646 0,800 0,344
Gs 2,660 2,380 0,240 0,070 0,480 1,400
Gs (%) 1,340 2,260 1,570 1,490 3,060 1,250
b 0,484 0,522 0,357 0,555 0,817 0,297
CVG (%) 2,918 4,528 3,922 2,853 4,730 4,481
CV% 6,032 8,672 10,998 5,142 5,788 15,072 AP - altura da planta; AE - altura da espiga; CE - comprimento de espiga; DE - diâmetro da espiga; NFG
- número de fileiras de grãos na espiga e MG - massa de grãos. (*) Intensidade de seleção de 20%.
Na Tabela 20 são apresentadas as estimativas de correlação fenotípica entre os
caracteres avaliados no presente estudo. Os maiores valores de correlação fenotípica
foram obtidos entre os caracteres FM e FF (0,96**) e entre AP e AE (0,80**). Em
geral, houve pouca relação dos caracteres SG, IF, NRP, CP, PROL e MG com os
demais caracteres avaliados. Foram obtidos valores de correlação fenotípica positiva e
significativa (P<0,05) entre FF e IF (0,42*), NRP e NFG (0,37*), enquanto houve
correlação fenotípica negativa e significativa (P<0,05) entre SG e AP (-0,37*) e entre
NRP e CP (-0,16*). O caráter MG apresentou estimativa da correlação fenotípica
positiva e significativa (P<0,05) apenas com PROL (0,39*). Este resultado para
correlação entre MG e PROL está de acordo com SOUZA JUNIOR, 1985; ZAIDI et al.
(2004) e CÂMARA et al. (2007), que relataram fortes evidênciais de correlação positiva
entre esses dois caracteres. Infere-se, portanto, que plantas de milho mais prolíficas
apresentaram maior produtividade de grãos. Entretanto, as estimativas de correlação
fenotípica entre MG e caracteres como NRP, IF e SG foram não-significativas,
discordando de CÂMARA et al. (2007) que verificaram estimativa de correlação
fenotípica significativa envolvendo MG com os caracteres NRP (0,13), IF (-0,28 a -
0,30) e (-0,13 a -0,46). Em trabalhos de melhoramento, o conhecimento da associação

80
entre caracteres é de grande importância, principalmente, se houver dificuldades na
seleção de um desses caracteres devido a baixa herdabilidade e/ou problemas na
identicação e medição. Neste caso, a seleção indireta pode levar progressos mais rápidos
que a seleção direta para o caráter desejado (CÂMARA et al., 2007). Diante dos
resultados obtidos, a seleção para prolificidade tenderia a ser mais precisa que para os
demais caracteres e poderia indiretamente aumentar a massa de grãos sob condições de
déficit hídrico.
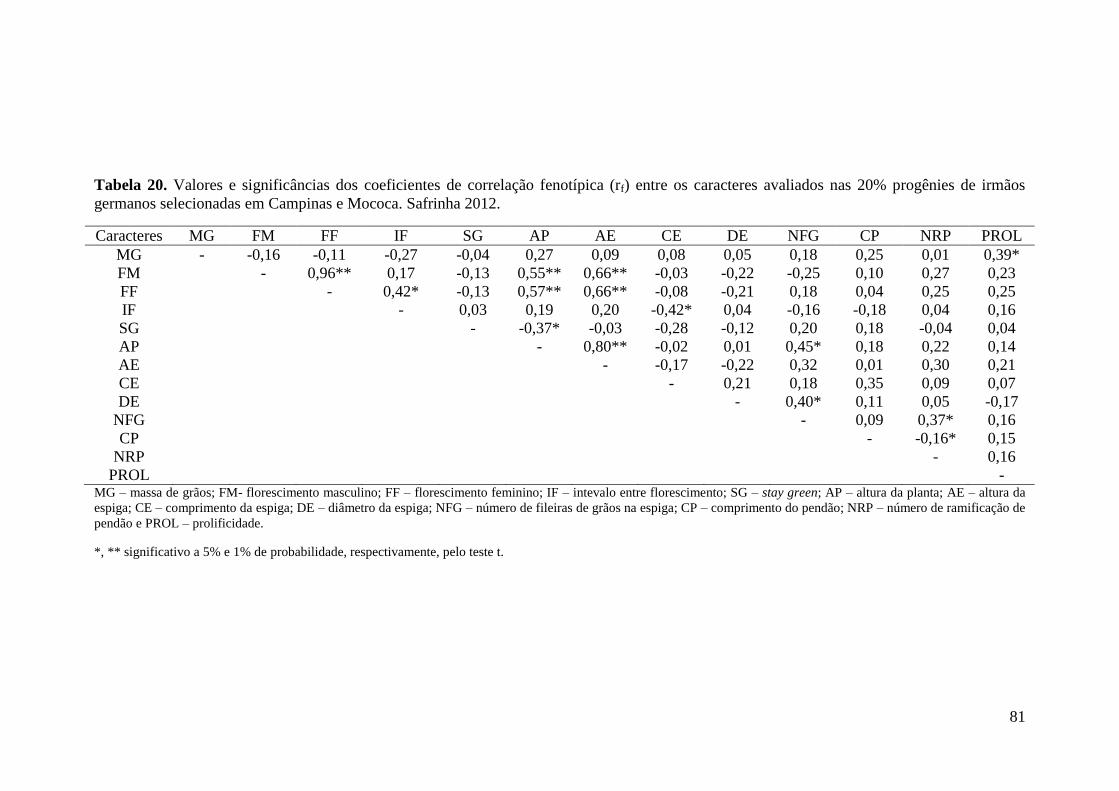
81
Tabela 20. Valores e significâncias dos coeficientes de correlação fenotípica (rf) entre os caracteres avaliados nas 20% progênies de irmãos
germanos selecionadas em Campinas e Mococa. Safrinha 2012.
Caracteres MG FM FF IF SG AP AE CE DE NFG CP NRP PROL
MG - -0,16 -0,11 -0,27 -0,04 0,27 0,09 0,08 0,05 0,18 0,25 0,01 0,39*
FM - 0,96** 0,17 -0,13 0,55** 0,66** -0,03 -0,22 -0,25 0,10 0,27 0,23
FF - 0,42* -0,13 0,57** 0,66** -0,08 -0,21 0,18 0,04 0,25 0,25
IF - 0,03 0,19 0,20 -0,42* 0,04 -0,16 -0,18 0,04 0,16
SG - -0,37* -0,03 -0,28 -0,12 0,20 0,18 -0,04 0,04
AP - 0,80** -0,02 0,01 0,45* 0,18 0,22 0,14
AE - -0,17 -0,22 0,32 0,01 0,30 0,21
CE - 0,21 0,18 0,35 0,09 0,07
DE - 0,40* 0,11 0,05 -0,17
NFG - 0,09 0,37* 0,16
CP - -0,16* 0,15
NRP - 0,16
PROL - MG – massa de grãos; FM- florescimento masculino; FF – florescimento feminino; IF – intevalo entre florescimento; SG – stay green; AP – altura da planta; AE – altura da
espiga; CE – comprimento da espiga; DE – diâmetro da espiga; NFG – número de fileiras de grãos na espiga; CP – comprimento do pendão; NRP – número de ramificação de
pendão e PROL – prolificidade.
*, ** significativo a 5% e 1% de probabilidade, respectivamente, pelo teste t.
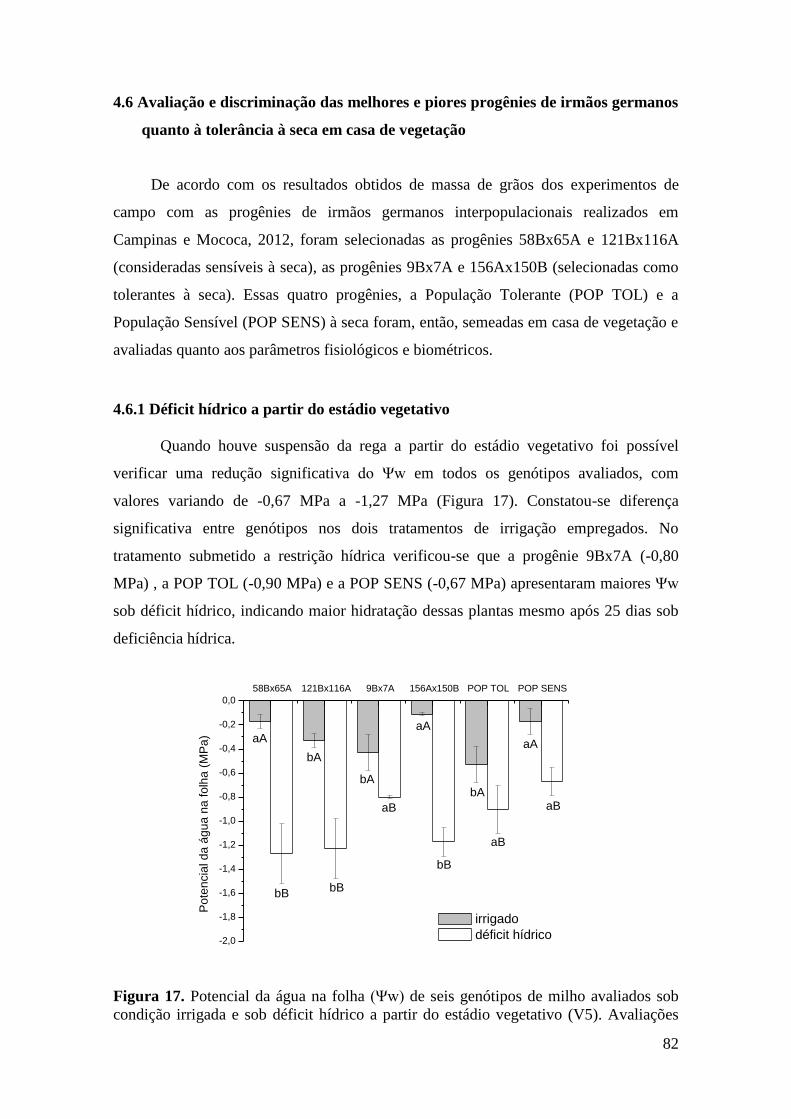
82
4.6 Avaliação e discriminação das melhores e piores progênies de irmãos germanos
quanto à tolerância à seca em casa de vegetação
De acordo com os resultados obtidos de massa de grãos dos experimentos de
campo com as progênies de irmãos germanos interpopulacionais realizados em
Campinas e Mococa, 2012, foram selecionadas as progênies 58Bx65A e 121Bx116A
(consideradas sensíveis à seca), as progênies 9Bx7A e 156Ax150B (selecionadas como
tolerantes à seca). Essas quatro progênies, a População Tolerante (POP TOL) e a
População Sensível (POP SENS) à seca foram, então, semeadas em casa de vegetação e
avaliadas quanto aos parâmetros fisiológicos e biométricos.
4.6.1 Déficit hídrico a partir do estádio vegetativo
Quando houve suspensão da rega a partir do estádio vegetativo foi possível
verificar uma redução significativa do Ψw em todos os genótipos avaliados, com
valores variando de -0,67 MPa a -1,27 MPa (Figura 17). Constatou-se diferença
significativa entre genótipos nos dois tratamentos de irrigação empregados. No
tratamento submetido a restrição hídrica verificou-se que a progênie 9Bx7A (-0,80
MPa) , a POP TOL (-0,90 MPa) e a POP SENS (-0,67 MPa) apresentaram maiores Ψw
sob déficit hídrico, indicando maior hidratação dessas plantas mesmo após 25 dias sob
deficiência hídrica.
Figura 17. Potencial da água na folha (Ψw) de seis genótipos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio vegetativo (V5). Avaliações
58Bx65A 121Bx116A 9Bx7A 156Ax150B POP TOL POP SENS
-2,0
-1,8
-1,6
-1,4
-1,2
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
irrigado
déficit hídrico
Po
ten
cia
l d
a á
gu
a n
a f
olh
a (
MP
a)
bBbB
bB
aB
aB
aB
aA
bA
bA
aA
bA
aA
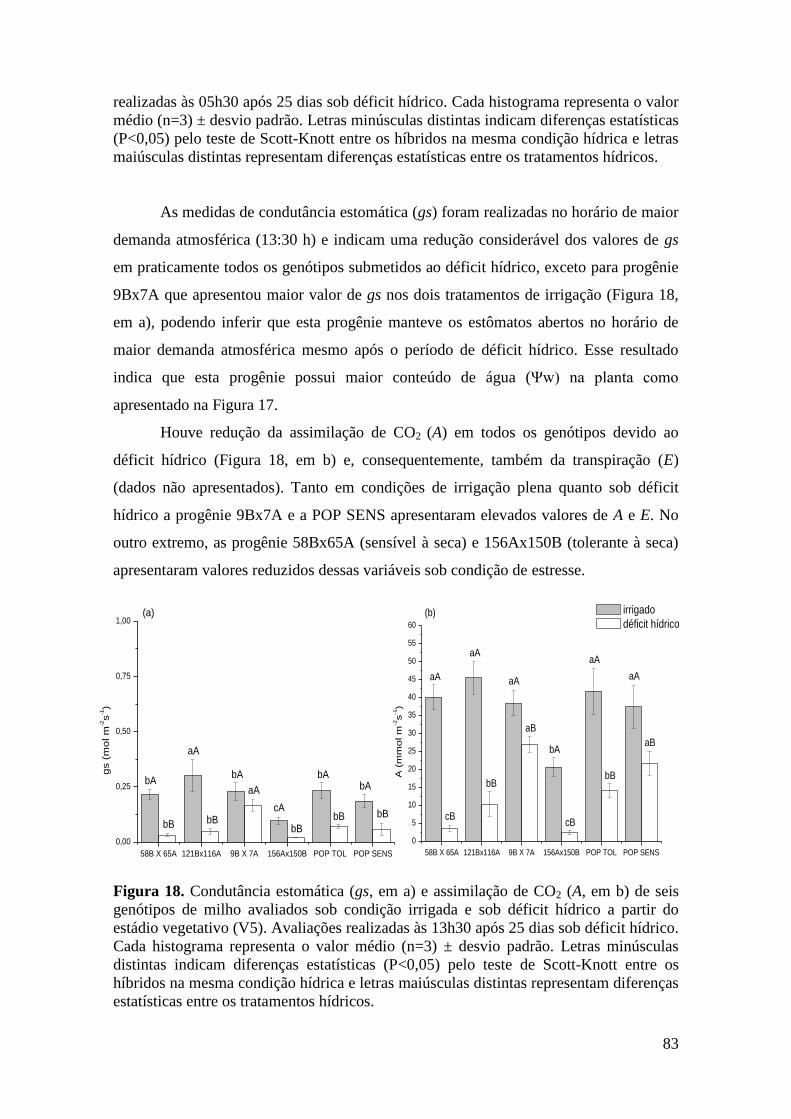
83
realizadas às 05h30 após 25 dias sob déficit hídrico. Cada histograma representa o valor
médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam diferenças estatísticas
(P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras
maiúsculas distintas representam diferenças estatísticas entre os tratamentos hídricos.
As medidas de condutância estomática (gs) foram realizadas no horário de maior
demanda atmosférica (13:30 h) e indicam uma redução considerável dos valores de gs
em praticamente todos os genótipos submetidos ao déficit hídrico, exceto para progênie
9Bx7A que apresentou maior valor de gs nos dois tratamentos de irrigação (Figura 18,
em a), podendo inferir que esta progênie manteve os estômatos abertos no horário de
maior demanda atmosférica mesmo após o período de déficit hídrico. Esse resultado
indica que esta progênie possui maior conteúdo de água (Ψw) na planta como
apresentado na Figura 17.
Houve redução da assimilação de CO2 (A) em todos os genótipos devido ao
déficit hídrico (Figura 18, em b) e, consequentemente, também da transpiração (E)
(dados não apresentados). Tanto em condições de irrigação plena quanto sob déficit
hídrico a progênie 9Bx7A e a POP SENS apresentaram elevados valores de A e E. No
outro extremo, as progênie 58Bx65A (sensível à seca) e 156Ax150B (tolerante à seca)
apresentaram valores reduzidos dessas variáveis sob condição de estresse.
Figura 18. Condutância estomática (gs, em a) e assimilação de CO2 (A, em b) de seis
genótipos de milho avaliados sob condição irrigada e sob déficit hídrico a partir do
estádio vegetativo (V5). Avaliações realizadas às 13h30 após 25 dias sob déficit hídrico.
Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras minúsculas
distintas indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os
híbridos na mesma condição hídrica e letras maiúsculas distintas representam diferenças
estatísticas entre os tratamentos hídricos.
58B X 65A 121Bx116A 9B X 7A 156Ax150B POP TOL POP SENS
0,00
0,25
0,50
0,75
1,00(a)
bBbBbB
bBbB
aA
cA
bAbA
bAbA
aA
gs (
mo
l m
-2s
-1)
58B X 65A 121Bx116A 9B X 7A 156Ax150B POP TOL POP SENS
0
5
10
15
20
25
30
35
40
45
50
55
60
irrigado
déficit hídrico(b)
cBcB
bBbB
aB
aB
aA
aA
aA
aA
aA
bA
A (
mm
ol m
-2s
-1)

84
Em relação a eficiência máxima potencial do fotossistema II (Fv/Fm), verificou-
se que os valores em 121Bx116A, 9Bx7A e POP TOL permaneceram constantes
mesmo após o déficit hídrico (Tabela 21), indicando alta eficiência no uso da radiação
nas reações primárias da fotossíntese (SILVA et al., 2007).
Houve redução significativa do coeficiente de extinção fotoquímica da
fluorescência (qP) em praticamente todos os genótipos devido ao déficit hídrico, exceto
para a progênie 9Bx7A, que não apresentou diferença significativa nos dois tratamentos
hídricos impostos. Essa mesma progênie apresentou maior valor desse parâmetro em
condições de seca quando comparado aos demais genótipos avaliados na mesma
condição de estresse, caracterizando uma maior dissipação de energia para o
metabolismo de carbono. Verificou-se que as progênies consideradas sensíveis à seca
58Bx65A, 121Bx116A e a POP SENS apresentaram menores valores de qP em
condições de deficiência hídrica.
O déficit hídrico aumentou o coeficiente de extinção não-fotoquímica da
fluorescência (NPQ) em todos os genótipos. A progênie 156Ax150B obteve maior valor
de NPQ no tratamento sob condição de estresse, indicando que houve maior dissipação
da energia luminosa absorvida na forma de energia térmica em detrimento ao
aproveitamento desta energia em efeito fotoquímico, ou seja, na produção de energia
química na forma de ATP e NADPH.
A deficiência hídrica provocou redução também na taxa aparente do transporte
de elétrons (ETR) em comparação ao tratamento irrigado, exceto para 9Bx7A e POP
TOL que não apresentaram diferença significativa entre os tratamentos hídricos
impostos. A redução de ETR indica que o estresse inibe o transporte de elétrons no
fotossistema II, compromentendo o metabolismo de carbano nesses genótipos. A POP
TOL e a progênie 9Bx7A apresentaram maiores valores de ETR no tratamento sob
déficit hídrico.

85
Tabela 21. Eficiência quântica potencial do fotossistema II (Fv/Fm), coeficientes de
extinção fotoquímica (qP) e não fotoquímica (NPQ) da fluorescência e transporte
aparente de elétrons (ETR) de seis genótipos de milho avaliados sob condição irrigada e
sob déficit hídrico a partir do estádio vegetativo (V5). Campinas, 2013.
Variáveis/Tratamento hídrico
Genótipos Fv/Fm
Irrigado Déficit hídrico
58Bx65A 0,74 ± 0,02 aA 0,64 ± 0,06 bB
121Bx116A 0,75 ± 0,03 aA 0,72 ± 0,03 aA
9Bx7A 0,74 ± 0,01 aA 0,74 ± 0,02 aA
156Ax150B 0,72 ± 0,03 aA 0,59 ± 0,08 bB
POP. TOL 0,76 ± 0,01 aA 0,71 ± 0,04 aA
POP. SENS 0,76 ± 0,01 aA 0,65 ± 0,05 bB
Genótipos qP
Irrigado Déficit hídrico
58Bx65A 0,53 ± 0,03 aA 0,33 ± 0,04 cB
121Bx116A 0,51 ± 0,02 aA 0,34 ± 0,04 cB
9Bx7A 0,53 ± 0,01 aA 0,51 ± 0,01 aA
156Ax150B 0,51 ± 0,03 aA 0,24 ± 0,08 dB
POP. TOL 0,49 ± 0,01 aA 0,44 ± 0,06 bB
POP. SENS 0,53 ± 0,02 aA 0,28 ± 0,02 dB
Genótipos NPQ
Irrigado Déficit hídrico
58Bx65A 0,74 ± 0,09 bB 1,84 ± 0,16 dA
121Bx116A 0,79 ± 0,13 bB 1,99 ± 0,08 bA
9Bx7A 0,86 ± 0,06 bB 2,09 ± 0,03 bA
156Ax150B 0,82 ± 0,05 bB 2,64 ± 0,07 aA
POP. TOL 0,95 ± 0,13 aB 2,03 ± 0,06 bA
POP. SENS 0,97 ± 0,07 aB 1,94 ± 0,05 cA
Genótipos ETR (µmol m
-2 s
-1)
Irrigado Déficit hídrico
58Bx65A 152,12 ± 10,17 aA 59,86 ± 3,27 bB
121Bx116A 178,01 ± 17,00 aA 68,02 ± 10,22 bB
9Bx7A 163,85 ± 8,12 aA 123,09 ± 10,53 aA
156Ax150B 135,94 ± 49,00 aA 32,25 ± 11,20 cB
POP. TOL 168,78 ± 10,48 aA 121,93 ± 13,41 aA
POP. SENS 145,27 ± 24,37 aA 79,15 ± 10,74 bB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas avaliadas após 25 dias sob déficit hídrico.
As progênies 58Bx65A e 156Ax150B sofreram redução na altura da planta (AP)
devido ao déficit hídrico, enquanto os demais genótipos não apresentaram diferença
significativa nos dois tratamentos hídricos empregados. O diâmetro do colmo (DIAM)
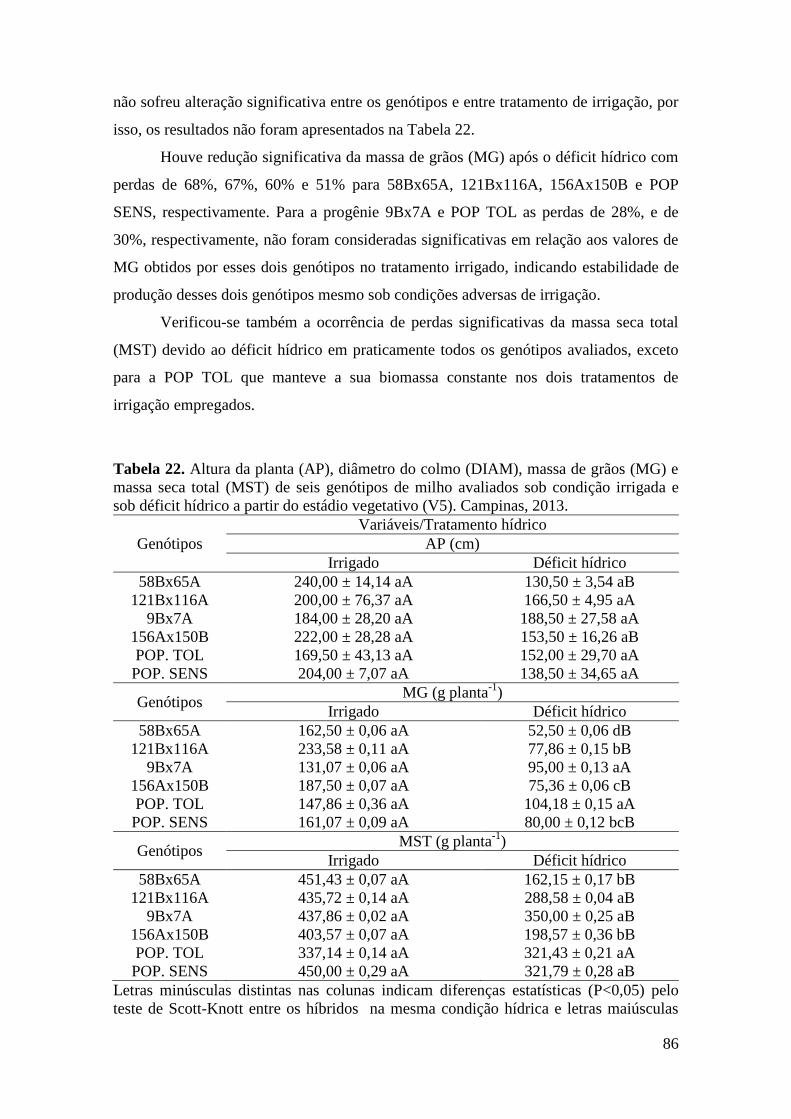
86
não sofreu alteração significativa entre os genótipos e entre tratamento de irrigação, por
isso, os resultados não foram apresentados na Tabela 22.
Houve redução significativa da massa de grãos (MG) após o déficit hídrico com
perdas de 68%, 67%, 60% e 51% para 58Bx65A, 121Bx116A, 156Ax150B e POP
SENS, respectivamente. Para a progênie 9Bx7A e POP TOL as perdas de 28%, e de
30%, respectivamente, não foram consideradas significativas em relação aos valores de
MG obtidos por esses dois genótipos no tratamento irrigado, indicando estabilidade de
produção desses dois genótipos mesmo sob condições adversas de irrigação.
Verificou-se também a ocorrência de perdas significativas da massa seca total
(MST) devido ao déficit hídrico em praticamente todos os genótipos avaliados, exceto
para a POP TOL que manteve a sua biomassa constante nos dois tratamentos de
irrigação empregados.
Tabela 22. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG) e
massa seca total (MST) de seis genótipos de milho avaliados sob condição irrigada e
sob déficit hídrico a partir do estádio vegetativo (V5). Campinas, 2013.
Genótipos
Variáveis/Tratamento hídrico
AP (cm)
Irrigado Déficit hídrico
58Bx65A 240,00 ± 14,14 aA 130,50 ± 3,54 aB
121Bx116A 200,00 ± 76,37 aA 166,50 ± 4,95 aA
9Bx7A 184,00 ± 28,20 aA 188,50 ± 27,58 aA
156Ax150B 222,00 ± 28,28 aA 153,50 ± 16,26 aB
POP. TOL 169,50 ± 43,13 aA 152,00 ± 29,70 aA
POP. SENS 204,00 ± 7,07 aA 138,50 ± 34,65 aA
Genótipos MG (g planta
-1)
Irrigado Déficit hídrico
58Bx65A 162,50 ± 0,06 aA 52,50 ± 0,06 dB
121Bx116A 233,58 ± 0,11 aA 77,86 ± 0,15 bB
9Bx7A 131,07 ± 0,06 aA 95,00 ± 0,13 aA
156Ax150B 187,50 ± 0,07 aA 75,36 ± 0,06 cB
POP. TOL 147,86 ± 0,36 aA 104,18 ± 0,15 aA
POP. SENS 161,07 ± 0,09 aA 80,00 ± 0,12 bcB
Genótipos MST (g planta
-1)
Irrigado Déficit hídrico
58Bx65A 451,43 ± 0,07 aA 162,15 ± 0,17 bB
121Bx116A 435,72 ± 0,14 aA 288,58 ± 0,04 aB
9Bx7A 437,86 ± 0,02 aA 350,00 ± 0,25 aB
156Ax150B 403,57 ± 0,07 aA 198,57 ± 0,36 bB
POP. TOL 337,14 ± 0,14 aA 321,43 ± 0,21 aA
POP. SENS 450,00 ± 0,29 aA 321,79 ± 0,28 aB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas

87
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=2) ± desvio padrão. Medidas avaliadas após 25 dias sob déficit hídrico.
A redução relativa (em relação ao controle) em cada variável analisada foi
calculada e os resultados são apresentados na Figura 19.
De acordo com essa análise, pode-se inferir que a progênie 9Bx7A foi menos
afetada pelo déficit hídrico (considerando MG e MST) devido à maior tolerância de gs,
A e ETR (valores mais próximos de 1), além de apresentar elevado Ψw (Figura 17),
confirmando sua resistência ao déficit hídrico. Por outro lado, a progênie 58Bx65A foi
comprovada como padrão de sensibilidade à seca devido as perdas significativas de MG
e de MST após o período de seca provocadas pela menor tolerância de gs e A, aliado ao
baixo valor de Ψw.
Figura 19. Representação gráfica da redução relativa da altura da planta (AP), massa de
grãos (MG), massa seca total (MST), condutância estomática (gs), assimilação de CO2
(A), eficiência quântica potencial do fotossistema II (Fv/Fm), coeficiente de dissipação
não-fotoquímica da fluorescência (NPQ), coeficiente de dissipação fotoquímica (qP) e
taxa aparente de transporte de elétrons (ETR) de seis genótipos de milho submetidos a
dois tratamentos hídricos a partir do estádio vegetativo (V5). Valores maiores que 1
indicam aumento na variável específica sob déficit hídrico.
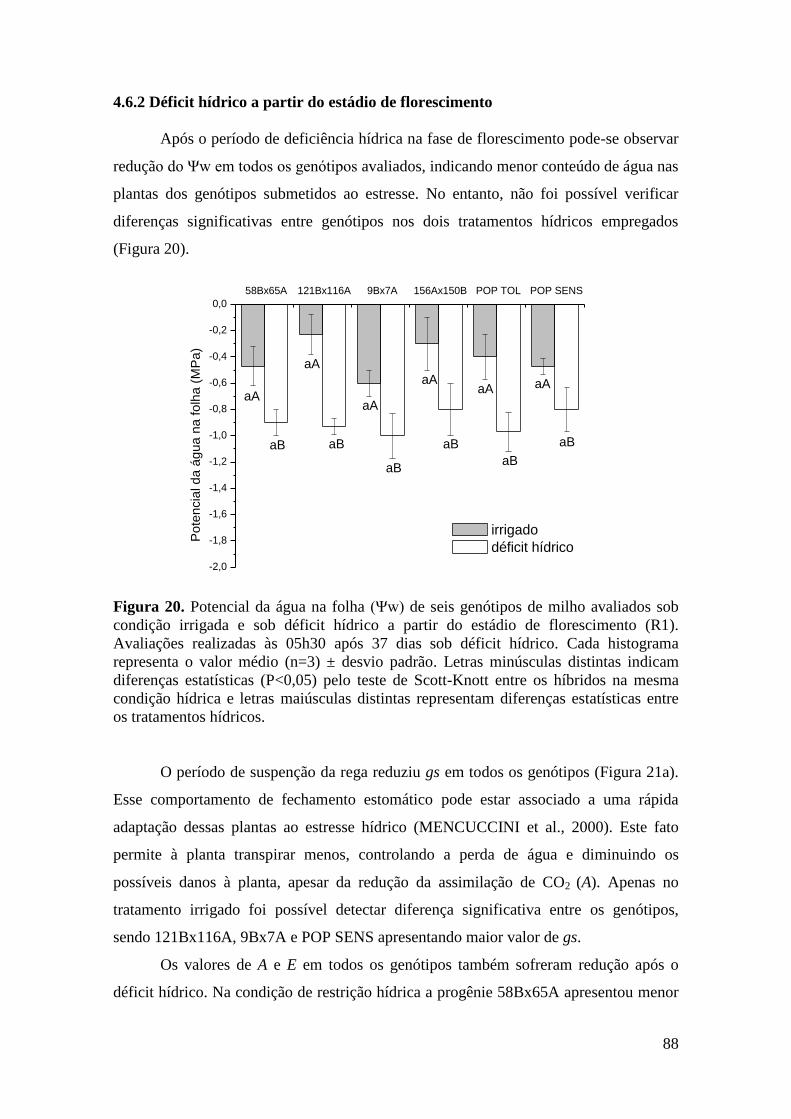
88
4.6.2 Déficit hídrico a partir do estádio de florescimento
Após o período de deficiência hídrica na fase de florescimento pode-se observar
redução do Ψw em todos os genótipos avaliados, indicando menor conteúdo de água nas
plantas dos genótipos submetidos ao estresse. No entanto, não foi possível verificar
diferenças significativas entre genótipos nos dois tratamentos hídricos empregados
(Figura 20).
Figura 20. Potencial da água na folha (Ψw) de seis genótipos de milho avaliados sob
condição irrigada e sob déficit hídrico a partir do estádio de florescimento (R1).
Avaliações realizadas às 05h30 após 37 dias sob déficit hídrico. Cada histograma
representa o valor médio (n=3) ± desvio padrão. Letras minúsculas distintas indicam
diferenças estatísticas (P<0,05) pelo teste de Scott-Knott entre os híbridos na mesma
condição hídrica e letras maiúsculas distintas representam diferenças estatísticas entre
os tratamentos hídricos.
O período de suspenção da rega reduziu gs em todos os genótipos (Figura 21a).
Esse comportamento de fechamento estomático pode estar associado a uma rápida
adaptação dessas plantas ao estresse hídrico (MENCUCCINI et al., 2000). Este fato
permite à planta transpirar menos, controlando a perda de água e diminuindo os
possíveis danos à planta, apesar da redução da assimilação de CO2 (A). Apenas no
tratamento irrigado foi possível detectar diferença significativa entre os genótipos,
sendo 121Bx116A, 9Bx7A e POP SENS apresentando maior valor de gs.
Os valores de A e E em todos os genótipos também sofreram redução após o
déficit hídrico. Na condição de restrição hídrica a progênie 58Bx65A apresentou menor
58Bx65A 121Bx116A 9Bx7A 156Ax150B POP TOL POP SENS
-2,0
-1,8
-1,6
-1,4
-1,2
-1,0
-0,8
-0,6
-0,4
-0,2
0,0
aB
aB
aB
aB
aB
aAaAaA
aA
aA
aA
aB
Po
ten
cia
l d
a á
gu
a n
a f
olh
a (
MP
a)
irrigado
déficit hídrico

89
assimilação de CO2 (Figura 21b). A mesma tendência também foi verificada para a
transpiração dessa progênie que apresentou reduzido valor de E após o déficit hídrico
(dados não apresentados).
As progênies 9Bx7A e 156Ax150B, consideradas tolerantes à seca,
apresentaram maiores valores de A e E nas condições de déficit hídrico.
Figura 21. Condutância estomática (gs, em a) e assimilação de CO2 (A, em b) de seis
genótipos de milho avaliados sob condição irrigada e sob déficit hídrico a partir do
estádio de florescimento (R1). Avaliações realizadas às 13h30 após 37 dias sob déficit
hídrico. Cada histograma representa o valor médio (n=3) ± desvio padrão. Letras
minúsculas distintas indicam diferenças estatísticas (P<0,05) pelo teste de Scott-Knott
entre os híbridos na mesma condição hídrica e letras maiúsculas distintas representam
diferenças estatísticas entre os tratamentos hídricos.
A variável Fv/Fm tem sido considerada uma boa referência para diagnosticar a
integridade do sistema fotossintético em diferentes espécies. No presente estudo, a
suspensão da irrigação levou a redução de Fv/Fm em praticamente todos os genótipos
avaliados, exceto para a progênie 9Bx7A e para a POP TOL (Tabela 23), o que indica
alta eficiência no uso da radiação nas reações primárias da fotossíntese nesses dois
genótipos. No tratamento sob déficit hídrico as progênies 58Bx65A e 121Bx116A,
consideradas sensíveis à seca, apresentaram valores reduzidos de Fv/Fm, indicando a
ocorrência de perturbações no sistema fotossintético causada pelo estresse, visto que a
redução desta variável indica inibição da atividade fotoquímica e, consequentemente,
compromete a assimilação de CO2.
Houve redução de qP em todos os genótipos devido ao déficit hídrico. Verificou-
se diferença significativa entre genótipos apenas no tratamento irrigado, sendo a POP
58B X 65A 121Bx116A 9B X 7A 156Ax150B POP TOL POP SENS
0,00
0,25
0,50
0,75
1,00
(a)
aBaB
aB
aB
aB
aB
bA
aA
bAbA
aAaA
gs (
mo
l m
-2s
-1)
58B X 65A 121Bx116A 9B X 7A 156Ax150B POP TOL POP SENS
0
5
10
15
20
25
30
35
40
45
50
55
60
irrigado
déficit hídrico(b)
bBbBbB
cB
aBaB
bAbA
bAaA
aA
aA
A (
mm
ol m
-2s
-1)
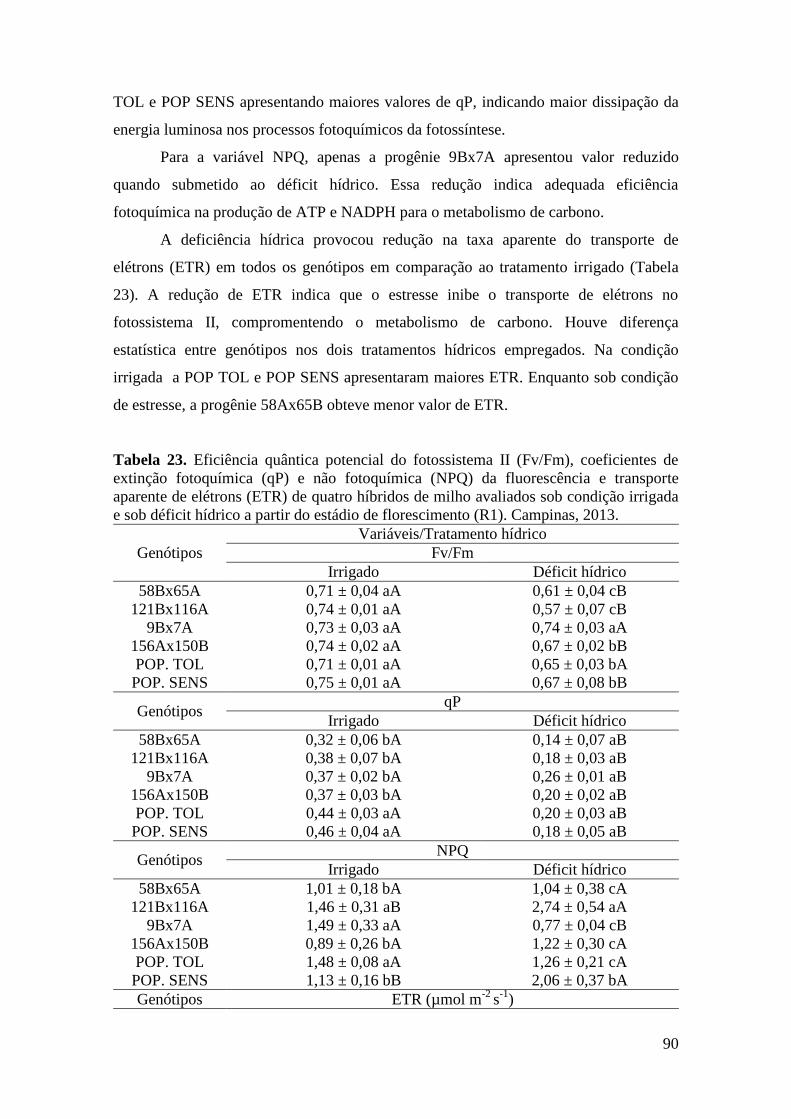
90
TOL e POP SENS apresentando maiores valores de qP, indicando maior dissipação da
energia luminosa nos processos fotoquímicos da fotossíntese.
Para a variável NPQ, apenas a progênie 9Bx7A apresentou valor reduzido
quando submetido ao déficit hídrico. Essa redução indica adequada eficiência
fotoquímica na produção de ATP e NADPH para o metabolismo de carbono.
A deficiência hídrica provocou redução na taxa aparente do transporte de
elétrons (ETR) em todos os genótipos em comparação ao tratamento irrigado (Tabela
23). A redução de ETR indica que o estresse inibe o transporte de elétrons no
fotossistema II, compromentendo o metabolismo de carbono. Houve diferença
estatística entre genótipos nos dois tratamentos hídricos empregados. Na condição
irrigada a POP TOL e POP SENS apresentaram maiores ETR. Enquanto sob condição
de estresse, a progênie 58Ax65B obteve menor valor de ETR.
Tabela 23. Eficiência quântica potencial do fotossistema II (Fv/Fm), coeficientes de
extinção fotoquímica (qP) e não fotoquímica (NPQ) da fluorescência e transporte
aparente de elétrons (ETR) de quatro híbridos de milho avaliados sob condição irrigada
e sob déficit hídrico a partir do estádio de florescimento (R1). Campinas, 2013.
Genótipos
Variáveis/Tratamento hídrico
Fv/Fm
Irrigado Déficit hídrico
58Bx65A 0,71 ± 0,04 aA 0,61 ± 0,04 cB
121Bx116A 0,74 ± 0,01 aA 0,57 ± 0,07 cB
9Bx7A 0,73 ± 0,03 aA 0,74 ± 0,03 aA
156Ax150B 0,74 ± 0,02 aA 0,67 ± 0,02 bB
POP. TOL 0,71 ± 0,01 aA 0,65 ± 0,03 bA
POP. SENS 0,75 ± 0,01 aA 0,67 ± 0,08 bB
Genótipos qP
Irrigado Déficit hídrico
58Bx65A 0,32 ± 0,06 bA 0,14 ± 0,07 aB
121Bx116A 0,38 ± 0,07 bA 0,18 ± 0,03 aB
9Bx7A 0,37 ± 0,02 bA 0,26 ± 0,01 aB
156Ax150B 0,37 ± 0,03 bA 0,20 ± 0,02 aB
POP. TOL 0,44 ± 0,03 aA 0,20 ± 0,03 aB
POP. SENS 0,46 ± 0,04 aA 0,18 ± 0,05 aB
Genótipos NPQ
Irrigado Déficit hídrico
58Bx65A 1,01 ± 0,18 bA 1,04 ± 0,38 cA
121Bx116A 1,46 ± 0,31 aB 2,74 ± 0,54 aA
9Bx7A 1,49 ± 0,33 aA 0,77 ± 0,04 cB
156Ax150B 0,89 ± 0,26 bA 1,22 ± 0,30 cA
POP. TOL 1,48 ± 0,08 aA 1,26 ± 0,21 cA
POP. SENS 1,13 ± 0,16 bB 2,06 ± 0,37 bA
Genótipos ETR (µmol m-2
s-1
)
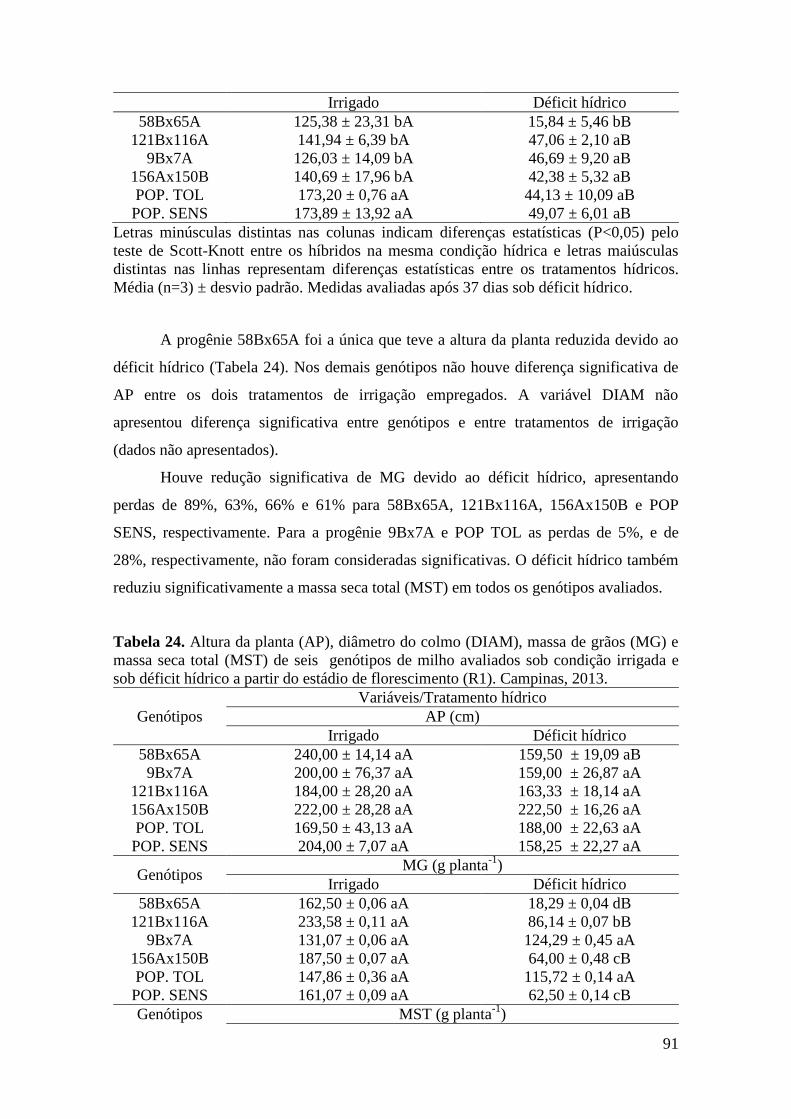
91
Irrigado Déficit hídrico
58Bx65A 125,38 ± 23,31 bA 15,84 ± 5,46 bB
121Bx116A 141,94 ± 6,39 bA 47,06 ± 2,10 aB
9Bx7A 126,03 ± 14,09 bA 46,69 ± 9,20 aB
156Ax150B 140,69 ± 17,96 bA 42,38 ± 5,32 aB
POP. TOL 173,20 ± 0,76 aA 44,13 ± 10,09 aB
POP. SENS 173,89 ± 13,92 aA 49,07 ± 6,01 aB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=3) ± desvio padrão. Medidas avaliadas após 37 dias sob déficit hídrico.
A progênie 58Bx65A foi a única que teve a altura da planta reduzida devido ao
déficit hídrico (Tabela 24). Nos demais genótipos não houve diferença significativa de
AP entre os dois tratamentos de irrigação empregados. A variável DIAM não
apresentou diferença significativa entre genótipos e entre tratamentos de irrigação
(dados não apresentados).
Houve redução significativa de MG devido ao déficit hídrico, apresentando
perdas de 89%, 63%, 66% e 61% para 58Bx65A, 121Bx116A, 156Ax150B e POP
SENS, respectivamente. Para a progênie 9Bx7A e POP TOL as perdas de 5%, e de
28%, respectivamente, não foram consideradas significativas. O déficit hídrico também
reduziu significativamente a massa seca total (MST) em todos os genótipos avaliados.
Tabela 24. Altura da planta (AP), diâmetro do colmo (DIAM), massa de grãos (MG) e
massa seca total (MST) de seis genótipos de milho avaliados sob condição irrigada e
sob déficit hídrico a partir do estádio de florescimento (R1). Campinas, 2013.
Genótipos
Variáveis/Tratamento hídrico
AP (cm)
Irrigado Déficit hídrico
58Bx65A 240,00 ± 14,14 aA 159,50 ± 19,09 aB
9Bx7A 200,00 ± 76,37 aA 159,00 ± 26,87 aA
121Bx116A 184,00 ± 28,20 aA 163,33 ± 18,14 aA
156Ax150B 222,00 ± 28,28 aA 222,50 ± 16,26 aA
POP. TOL 169,50 ± 43,13 aA 188,00 ± 22,63 aA
POP. SENS 204,00 ± 7,07 aA 158,25 ± 22,27 aA
Genótipos MG (g planta
-1)
Irrigado Déficit hídrico
58Bx65A 162,50 ± 0,06 aA 18,29 ± 0,04 dB
121Bx116A 233,58 ± 0,11 aA 86,14 ± 0,07 bB
9Bx7A 131,07 ± 0,06 aA 124,29 ± 0,45 aA
156Ax150B 187,50 ± 0,07 aA 64,00 ± 0,48 cB
POP. TOL 147,86 ± 0,36 aA 115,72 ± 0,14 aA
POP. SENS 161,07 ± 0,09 aA 62,50 ± 0,14 cB
Genótipos MST (g planta-1
)

92
Irrigado Déficit hídrico
58Bx65A 451,43 ± 0,07 aA 163,57 ± 0,10 aB
121Bx116A 435,72 ± 0,14 aA 271,79 ± 0,13 aB
9Bx7A 437,86 ± 0,02 aA 261,79 ± 0,18 aB
156Ax150B 403,57 ± 0,07 aA 280,72 ± 0,43 aB
POP. TOL 337,14 ± 0,14 aA 227,50 ± 0,46 aB
POP. SENS 450,00 ± 0,29 aA 209,29 ± 0,10 aB
Letras minúsculas distintas nas colunas indicam diferenças estatísticas (P<0,05) pelo
teste de Scott-Knott entre os híbridos na mesma condição hídrica e letras maiúsculas
distintas nas linhas representam diferenças estatísticas entre os tratamentos hídricos.
Média (n=2) ± desvio padrão. Medidas avaliadas após 37 dias sob déficit hídrico.
De acordo com a Figura 22 podemos inferir que a progênie 9Bx7A e a POP TOL
apresentaram adequada MG e MST devido a maiores valores de gs e A obtidos. A
progênie 58Bx65A apresentou perdas significativa de MG após o déficit hídrico, sendo
comprovada a sua sensibilidade à seca.
Figura 22. Representação gráfica da redução relativa da altura da planta (AP), massa de
grãos (MG), massa seca total (MST), condutância estomática (gs), assimilação de CO2
(A), eficiência quântica potencial do fotossistema II (Fv/Fm), coeficiente de dissipação
não-fotoquímica da fluorescência (NPQ), coeficiente de dissipação fotoquímica (qP) e
taxa aparente de transporte de elétrons (ETR) de seis genótipos de milho submetidos a
dois tratamentos hídricos a partir do estádio de florescimento (R1). Valores maiores que
1 indicam aumento na variável específica sob déficit hídrico.

93
Pelos resultados apresentados conclui-se que a POP TOL e a progênie 9Bx7A
são consideradas tolerantes ao déficit hídrico nos estádios vegetativo e florescimento da
cultura, enquanto que, a progênie 58Bx65A foi confirmada como padrão de
sensibilidade à seca nos dois estádios fenológicos avaliados. De maneira geral, as
progênies 121Bx116A e 156Ax150B apresentaram níveis intermediários de tolerância
ao déficit hídrico nas duas fases de desenvolvimento da cultura.
5 CONCLUSÕES
1. O híbrido DAS2B707 é considerado tolerante à seca no estádio vegetativo devido
à maior eficiência dos processos fotossintéticos após período de déficit hídrico,
enquanto o AS1522 mostra-se resistente ao déficit hídrico no estádio de
florescimento pela maior concentração de carboidratos na folha.
2. Os principais mecanismos fisiológicos de tolerância à seca são relacionados com
a fotossíntese (Ai, qP, ETR e NPQ) e manutenção do status hídrico ( Ψw e gs) da
planta.
3. Os estádios vegetativo e florescimento são considerados os mais críticos da
cultura do milho devido à maior redução da produtividade dos híbridos sob
déficit hídrico nesses estádios.
4. A progênie 9Bx7A confirma ser tolerante à seca e a 58Bx65A revela-se padrão
de sensibilidade ao déficit hídrico nos estádios vegetativo e florescimento.
5. Não foi possível eleger um parâmetro fisiológico eficiente na seleção e
discriminação de progênies, para futura utilização em programas de
melhoramento de milho.
6. Ressaltam-se progênies de IG interpopulacionais promissoras, com elevada
produtividade em condições de safrinha (déficit hídrico) e confirma-se a
importância de caracteres secundários (intervalo entre florescimentos,
prolificidade, número de ramificação de pendão e stay green) em progênies
tolerantes à seca.

94
7. Existe elevada herdabilidade para os caracteres número de fileiras de grãos na
espiga (NFG), número de ramificação de pendão (NRP), florescimento masculino
(FM) e feminino (FF), diâmetro da espiga (DE) e altura da espiga (AE),
demonstrando forte controle genético na herança destes caracteres e a
possibilidade de se obter altos ganhos com a seleção dentro de progênies.
8. As médias de produtividade e a magnitude das estimativas dos parâmetros
genéticos obtidos das progênies indicam que, de maneira geral, existe
variabilidade genética para ser explorada nos próximos ciclos de seleção
recorrente para tolerância à seca em milho.

95
6 REFERÊNCIAS BIBLIOGRÁFICAS
AGRIANUAL 2011. Anuário estatístico da agricultura brasileira. São Paulo: FNP,
p.496, 2010.
AGUIAR, A.M. Uso do Delineamento III com marcadores paraanálise genética da
produção de grãos e seus componentes em milho. 2003. 127p. Tese (Doutorado em
Genética e Melhoramento de Plnatas) – Escola Superior de Agricultura “Luiz de
Queiroz”, Universidade de São Paulo, Piracicaba.
AINSWORTH, E.A.; BUSH, D.R. Carbohydrate export from the leaf: a highly
regulated process and target to enhance photosynthesis and productivity. Plant
Physiology, v.155, p.64-69, 2011.
ALDRICH, S.R.; SCOTT, W.O.; LENG, E.R. Modern corn production. 2.ed.
Champaign: A & L Publication, p.371, 1982.
ALVES, G.F.; RAMALHO, M.A.P.; SOUZA, J.C. Alterações nas propriedades
genéticas da população CMS-39 submetida à seleção massal para a prolificidade.
Revista Brasileira de Milho e Sorgo, Sete Lagoas, v.1, n.3, p.89-101, 2002.
AMARAL, L.I.V.; GASPAR, M.; COSTA, P.M.F.; AIDAR, M.P.M.; BUCKERIDGE,
M.S. Novo método enzimático rápido e sensível de extração e dosagem de amido
em materiais vegetais. Hoehnea, São Paulo, v. 34, p. 425-431, 2007.
ANGELOCCI, L. R. Água na planta e trocas gasosas/energéticas com a atmosfera:
introdução ao tratamento biofísico. Piracicaba: FEALQ, p.272, 2002.
BAKER, N. R. Light-use efficiency and photoinhibition of photosynthesis in plants
under environmental stress. In: Smith JAC, Griffiths H (eds.) Water deficits plant
responses from cell to community. Bios Scientific Publisher, Oxford, p.221-235, 1993.
BAKER, N.R.; ROSENQVIST, E. Application of chlorophyll fluorescence can improve
crop production strategies: an examination of future possibilities. Journal of
Experimental Botany, v.55, p.1607-1621, 2004.
BÄNZINGER, M.; EDMEADES, G.O.; BECK, D.; BELLON, M. Breeding for
drought and nitrogen stress tolerance in maize: from theory to practice. Mexico,
D.F.: CIMMYT. 68p, 2000.
BERGAMASCHI, H.; DALMAGO, G.A.; BERGONCI, J.I.; BIANCHI, C.A.M.;
MÜLLER, A.G.; COMIRAN, F.; HECKLER, B.M.M. Distribuição hídrica no período
crítico do milho e produção de grãos. Pesquisa Agropecuária Brasileira, v.39, p.831-
839, 2004.
BERGAMASCHI, H.; DALMAGO, G.A.; COMIRAN, F.; BERGONCI, J.I.;
MÜLLER, A.G.; FRANÇA, S.; SANTOS, A.O.; RADIN, B.; BIANCHI, C.A.M.;

96
PEREIRA, P.G. Deficit hídrico e produtividade na cultura do milho. Pesquisa
Agropecuária Brasileira, v.41, p.243-249, 2006.
BERGONCI, J.I.; BERGAMASCHI, H.; BERLATO, M.A.; SANTOS, O.S. Potencial
da água na folha como um indicador de déficit hídrico em milho. Pesquisa
Agropecuária Brasileira, v.35, p.1531-1540, 2000.
BERGONCI, J.I.; BERGAMASCHI, H.; SANTOS, A.O.; FRANÇA, S.; RADIN, B.
Eficiência da irrigação em rendimento de grãos e matéria seca de milho. Pesquisa
Agropecuária Brasileira, v.36, p.949-956, 2001.
BETRÁN, F.J.; BECK, D.; BAZINGER, M.; EDMEADES, G.O. Secondary traits in
parental inbreds and hybrids under stress and non-stress environments in tropical maize.
Field Crops Research, Amsterdam, v.83, p.51-65, 2003.
BIELESKI, R.L.; TURNER, A. Separation and estimation of amino acids in crude plant
extracts by thin-layer electrophoresis and chromatography. Analytical Biochemistry,
New York, v.17, p. 278-293, 1966.
BLUM, A. Crop responses to drought and the interpretation of adaptation. In:
BELHASSEN, E. (Ed.). Drought tolerance in higher plants. Genetical, physiological
and molecular biological analysis. Dordrecht: Kluwer Academic, p.57-70, 1997.
BOYER, J. S. Leaf enlargement and metabolic rates in corn, soybean, and sunflower at
various leaf water potencials. Plant Physiology, 46, p. 233-235, 1970.
BOLAÑOS, J.; EDMEADES, G.O. Eight cycles of selection for drought tolerance in
lowland tropical maize. I. Responses in grain yield, biomass, and radiation utilization.
Field Crops Research, Amsterdam, v.31, p.233-252, 1993a.
BOLAÑOS, J.; EDMEADES, G. O. Eight cycles of selection for drought tolerance in
lowland tropical maize. II Responses in reproductive behaviour. Field Crops Research,
Amsterdam, v. 31, p. 253- 268, 1993b.
BOLAÑOS, J.; EDMEADES, G.O. The importance of the anthesis-silking interval in
breeding for drought tolerance in tropical maize. Field Crops Research, Amsterdam,
v.48, p.65-80, 1996.
BOLAÑOS, J.; EDMEADES, G. O.; MARTINEZ, L. Eight cycles of selection for
drought tolerance in lowland tropical maize. III. Responses in drought-adaptive
physiological and morphological traits. Field Crops Research, Amsterdam, v. 31, p.
269- 286, 1993.
BOLAÑOS, J.; EDMEADES, G.O. Maize breeding for drought tolerance. In:
INTERNATIONAL SYMPOSIUM ON ENVIRONMENTAL STRESS, 1992, Belo
Horizonte. Maize in perspective: proceedings. Sete Lagoas: Embrapa-CNPMS / Ciudad
del México: CIMMYT/UNDP, p.397-431, 1995.
BOLHÀR-NORDENKAMPF, H. R.; LONG, S. P.; BAKER, N. R.; ÖQUIST, G.;
SCHREIDER, U.; LECHNER E. G. Chlorophyll fluorescence as probe of the

97
photosynthetic competence of leaves in the field: A review of current instrument.
Functional Ecology, v.3, p.497- 514, 1989.
BORÉM, A. Melhoramento de Plantas, 3º edição. Viçosa: UFV. 453p., 2001.
BRAY, E. A. Plant responses to water deficit. Trend in Plant Science, London, v. 2, p.
48-54, 1997.
CÂMARA, T.M.M. Mapeamento de QTLs de caracteres relacionados à tolerância
ao estresse hídrico em milho tropical. 2006. 178p. Tese (Doutorado em Genética e
Melhoramento de Plantas). Escola Superior de Agricultura “Luiz de Queiroz” -
Universidade de São Paulo, Piracicaba.
CÂMARA, T.M.M.; VIEIRA BENTO, D.A.; ALVES, G.F.; SANTOS, M.F.;
MOREIRA, J.U.V.; SOUZA JUNIOR, C.L. de. Parâmetros genéticos de caracteres
relacionados à tolerância à deficiência hídrica em milho tropical. Bragantia, Campinas,
v.66, n.4, p.595-603, 2007.
CARLESSO, R.; PEITER, M. X.; JADOSKI, S. O.; MARCOLAN, A. Resposta do
milho irrigado submetido a diferentes frações de água disponível no solo. In:
Congresso Nacional de Milho e Sorgo, 21, 1996, Londrina. Resumos... Londrina:
IAPAR, p.196, 1996.
CARMO, S.L.M.; SANTOS, J.B.; HAGIWARA, W.E.; FERREIRA, J.L. Avaliação do
"stay green" em famílias segregantes de feijão (Phaseolus vulgaris L.) Ciência e
Agrotecnologia, v.31, p.953-957, 2007.
CARVALHO, H. W. L de.; GUIMARÃES, P. E.de O.; LEAL, M. de L da S.;
CARVALHO, P. C. L. de.; SANTOS, M. X. dos. Avaliação de progênies de meios-
irmãos da população de milho CMS 453 no Nordeste brasileiro. Pesquisa
Agropecuária Brasileira, Brasília, v.35, n.8, p.1577 - 1584, 2000b.
CARVALHO, H. W. L de.; LEAL, M. de L da S.; GUIMARÃES, P. E. de O.;
SANTOS, M. X. dos.; CARVALHO, P. C. L. de. Três ciclos de seleção entre e dentro
de progênies de meios-irmãos na população de milho CMS 52. Pesquisa Agropecuária
Brasileira, Brasília, v.35, n.8, p.1621-1628, 2000a.
CARVALHO, H. W. L de.; SANTOS, M. X. dos.; LEAL, M. de L da S.; CARAVLHO,
P. C. L. de. Melhoramento genético da cultivar de milho BR 5033-Asa Branca no
Nordeste brasileiro. Pesquisa Agropecuária Brasileira, Brasília, v.35, n.7, p.1417 -
1425, 2000c.
CHAPMAN, S.C., and G.O. EDMEADES. Selection improves tolerance to mid/late
season drought in tropical maize populations. II. Direct and correlated responses among
secondary traits. Crop Sci. 39 (In press), 1999.
CHAVES, M.M. Effects of water deficits on carbon assimilation. Journal of
Experimental Botany, v.42, p.1-16, 1991.

98
CHAVES, M.M.; OLIVEIRA , M.M. Mechanisms underlying plant resilience to water
deficits: prospects for water-saving agriculture. Journal of Experimental Botany,
v.55, p.2365-2384, 2004.
COMSTOCK, R.E.; ROBINSON, H.F.; HARVEY, P.H. A breeding procedure
designed to make maximum use of both general and specific combining ability.
Agronomy Journal, Madison, v.41, p.360-367, 1949.
CONAB – Companhia Nacional de Abastecimento (2013). Boletim informativo.
COSTA, E. F. N.; SANTOS, M.F.; MORO, G.V.; ALVES, G.F.; SOUZA JÚNIOR,
C.L. Herança da senescência retardada em milho. Pesquisa agropecuária Brasileira,
Brasília. v. 43, n.2, p. 207-213, 2008.
CRUZ, C.D. Programa Genes: Aplicativo Computacional em Genética e
Estatística. Universidade Federal de Viçosa, Viçosa-MG, 2001.
DEDEMO, G.C. Expressão gênica diferencial durante déficit hídrico em duas
cultivares de cana-de-açúcar. Dissertação (Mestrado em Agronomia – Genética e
Melhoramento de Plantas). Faculdade de Ciências Agrárias e Veterinárias – UNESP,
Botucatu.
DUBOIS, M.; GILLES, K.A.; HAMILTON, J.K.; REBERS, P.A.; SMITH, F.
Colorimetric method for determination of sugars and related substances. Analytical
Chemistry, London, v.28, p. 350-356, 1956.
DURÃES, F. O. M.; PAIVA, E.; MAGALHÃES, P. C.; SANTOS, M. X. DOS;
PEREIRA, J. J.; LABORY, C. R. G. Critérios morfofisiológicos utilizados para seleção
de genótipos de milho visando tolerância a seca. VI Congresso Brasileiro de
Fisiologia Vegetal. Belém, 1997.
DURÃES, F. O. M.; SANTOS, M. X. dos.; GOMES e GAMA; E. E.; MAGALHÃES,
P. C.; GUIMARÃES, C. T. Fenotipagem associada à tolerância à seca em milho para
uso em melhoramento, estudos genômicos e seleção assistida por marcadores. Sete
Lagoas: MAPA/Embrapa Milho e Sorgo, 2004. 18 p. (Circular Técnica, 39).
DUVICK, D.N.; CASSMAN, K.G. Post-green revolution trends in yield potential of
temperature maize in the North-Central United States. Crop Science, Madison,
v.39,p.1622-1630, 1999.
EDMEADES, G. O.; BOLAÑOS, J.; BÄNZIGER, M.; RIBAUT, J.M.; WHITE, J. W.;
REYNOLDS, M. P.; LAFITTE, H. R. Improving crop yields under water deficits in the
tropics. In: INTERNATIONAL CROP SCIENCE CONGRESS, 2., 1996, New
Delhi. Crop productivity and sustainability – Shaping the future: proceedings. New
Delhi: Oxford and IBH. p. 437-451, 1998.
EDMEADES, G.O.; BOLAÑOS, J.; CHAPMAN, S.C.; LAFITTE, H.R.; BÄNZIGER,
M. Selection improves tolerance to mid/late season drought in tropical maize

99
populations. I. Gains in biomass, grain yield and harvest index. Crop Science. v.39 (In
press), 1999.
FERREIRA, D. F. Manual do sistema Sisvar para análises estatísticas. Lavras:
UFLA, 2000. 66 p.
FLEXAS J, BOTA J, LORETO F, CORNIC G, SHARKEY TD. Diffusive and
metabolic limitations to photosynthesis under drought and salinity in C3 plants. Plant
Biology, v.6, p.269–279, 2004.
FLINT-GARCIA, S.A.; THUILLET, A.C.; YU, J.M.; PRESSOIR, G.; ROMERO,
S.M.; MICHELL, S.E.; DOEBLEY, J.; KRESOVICH, S.; GOODMAN, M.M.;
BUCKLER, E.S. Maize association population: a high-resolution platform for
quantitative trait locus dissection. Plant Journal, Oxon, v.44, p.1054-1064, 2005.
FISCHER, K.S.; EDMEADES, G.O.; JOHNSON, E.C. Recurrent selection for reduced
tassel branch number and reduced leaf-area density above the ear in tropical maize
populations. Crop Science, Madison,v.27, p.1150-1156, 1987.
GABRIEL, A.P.C. Seleção recorrente recíproca em famílias de irmãos completos
em milho comum (Zea mays L.) monitorada por marcadores moleculares: avanço
de gerações e avaliação do progresso genético. Tese (Doutorado em Genética e
Melhoramento de Plantas) Campos dos Goytacazes – UENF - RJ, 103p, 2009.
GHANNOUM, O.; CONROY, J. P.; DRISCOLL, S. P.; PAUL, M. J.; FOYER, C. H.;
LAWLOR, D. W. Nonstomatal limitations are responsible for drought-induced
photosynthetic inhibition in four C4 grasses. New Phytologist, v.159, p.599-608, 2003.
GLAZ, B.; MORRIS, D. R.; DAROUB, S. H. Sugarcane photosynthesis, transpiration,
and stomatal conductance due to flooding and water table. Crop Science, v.44, p.1633-
1641, 2004.
GUIMARÃES, P. S.; BERNINI, C. S.; PATERNIANI, M. E. A. G. Z.; GALLO, P.B. e
LODOVICO, F. A. Seleção de progênies de irmãos germanos interpopulacionais
visando à discriminação quanto à tolerância à seca em milho. In: 29 Congresso
Nacional de Milho e Sorgo, 2012, Campinas. Diversidade e Invações na era dos
trasngênicos. Campinas: IAC - Instituto Agronômico de Campinas, v.1. p.3115-3121,
2012.
HALLAUER, A.R.; EBERHART, S.A. Reciprocal full-sib selection. Crop.Science,
v.10, p.315-316, 1970.
HALLAUER, A.R.; MIRANDA FILHO, J.B. Quantitative genetics in maize
breeding. 2° ed. Ames. Iawa State University Press. 468p., 1988.
HULL, F.H. Recurrent selection for specific combining ability in corn. J. Am. Soc.
Agron., v.37, p.134-145, 1945.
IPCC. Climate Change 2007: Climate Change Impacts, Adaptation and
Vulnerability. Working Group II Contribution to the Intergovernmental Panel on

100
Climate Change. Fourth Assessment Report. Summary for Policymakers. Bonn,
Alemanha. 2007.
JIA, W.; ZHANG, J.; ZHANG, D.P. Metabolism of xylem-delivered ABA in relation to
ABA flux and concentration in leaves of maize and Commelina communis. Journal of
Experimental Botany, v.47, p.1085-1091, 1996.
JIANG, G.H.; EDMEADES, G.O.; ARMSTEAD, I.; LAFITTE, H.R.; HAYWARD,
M.D.; HOISINGTON, D. Genetic analysis of adaptation differences between highland
and lowland tropical maize using molecular markes. Theoretical and Applied
Genetics, v.99, p.1106-1119, 1999.
JIANG, G.H.; HE, Y.Q.; XU, C.G.; LI, X.H.; ZHANG, Q. The genetic basis of stay-
green in rice analyzed in a population of doubled haploid lines derived from an indica
by japonica cross. Theoretical and Applied Genetics, v.108, p.688-698, 2004.
KAISER, W.M. Effects of water deficit on photosynthetic capacity. Physiologia
Plantarum, Copenhagen, v.71, p.142-150, 1987.
KAKANI, V.G.; VU, J.C.V.; ALLEN Jr., L.H.; BOOTE, K.J. Leaf photosynthesis and
carbohydrates of CO2-enriched maize and grain sorghum exposed to a short period of
soil water deficit during vegetative development. Journal of Plant Physiology, v.168,
p.2169-2176, 2011.
KAMARA, A.Y.; MENKIR, A.; BADU-APRAKU, B.; IBIKUNLE, O. Reproductive
and stay-green trait responses of maize hybrids, improver open-pollinated varieties and
farmer’s local varieties to terminal drought stress. Maydica, Bergamo,v.48,p.29-37,
2003.
KAMARA, A.Y.; MENKIR, A.; AJALA, S.O.; KUREH, I. Performance of diverse
maize genotypes under nitrogen difeciency in the northern Guinea savanna of Nigeria.
Experimental Agriculture, New York, v.41, p.199-212, 2005.
KINGSTON-SMITH, A.H., WALKER, R.P. & POLLOCK, C.J. Invertase in leaves:
conundrum or control point? Journal of Experimental Botany, v50, p.735-743, 1999.
KRAMER, P. J. Fifty years of progress in water relations research. Plant Physiology,
v.54, p.463-471, 1974.
KRAMER, P. J.; BOYER J. S. Water relations of plants and soils. San Diego:
Academic Press, p.495, 1995.
KRAUSE, G.H.; WEIS, E. Chlorophyll fluorescence and photosynthesis: the basics.
Annual Review of Plant Physiology and Plant Molecular Biology, v.42, p.313-349,
1991.
KUNZ, J.H.; BERGONCI, J.I.; BERGAMASCHI, H.;DALMAGO, G.A.; HECKLER,
B.M.M; COMIRAN, F. Uso da radiação solar pelo milho sob diferentes preparos do
solo, espaçamento e disponibilidade hídrica. Pesquisa Agropecuária Brasileira,
Brasília, v.42, p.1511-1520, 2007.

101
LARCHER, W. Ecofiosiologia vegetal. São Carlos: Rima, p. 531, 2004.
LAWLOR, D.W. The effects of water deficit on photosynthesis. In: Smirnoff N (ed),
Environmental and plant metabolism-flexibility and acclimation, Bios Scientific
Publisher, Oxford, p.129-160, 1995.
LAWLOR, D. W.; CORNIC, G. Photosynthetic carbon assimilation and associated
metabolism in relation to water deficits in higher plants. Plant, Cell Environ., v.25,
p.275- 294, 2002.
LI, X.H.; LIU, X.D.; LI, M.S.; ZHANG, S.H. Identification of Quantitative Trait Loci
for Anthesis-Interval and Yield Components Under Drought Stress in Maize. Acta
Botanica Sinica, Beijing, v.45, n7, p.852-857, 2003.
MACHADO, R.A.F. Linhagens de milho contrastantes para tolerância à seca e
eficiência de utilização de nitrogênio. 2003. 90 p. Tese (Doutorado em Agronomia) –
Universidade Estadual Paulista "Júlio de Mesquita Filho", Botucatu, 2003.
MAGALHÃES, P. C.; DURÃES, F. O. M.; CARNEIRO, N. P.; PAIVA, E. Fisiologia
do milho. Sete Lagoas: MAPA/Embrapa Milho e Sorgo, 2002. 23 p. (Circular Técnica,
22).
MAGALHÃES, A. C. N. Fotossíntese, particionamento de assimilados e crescimento de
plantas sob condições de estresse: com destaque para o milho. In: Simpósio
Internacional sobre Estresse Ambiental, 1992, Belo Horizonte. O milho em
perspectiva.Sete Lagoas: EMBRAPA/CNPMS. México: CIMMYTIUNDP, p.195-221,
1995.
MAGALHÃES, P.C.; RESENDE, M.; OLIVEIRA, A. C. de; DURÃES, F.O.M.;
SANS, L. M. A. Caracterização morfológica de milho de diferentes ciclos. In:
CONGRESSO NACIONAL DE MILHO E SORGO, 20, 1994, Goiânia. Centro
Oeste-cinturao do milho e do sorgo no Brasil: resumos. Goiânia, ABMS, p. 190,
1994.
MAGALHÃES, P.C.; DURÃES, F.O.M.; OLIVEIRA, A. C. de. Efeitos do
quebramento do colmo no rendimento de grãos de milho. Ciência e Agrotecnologia,
Lavras, v. 22, p. 279-289, 1998.
MEDICI, L.O.; PEREIRA, M.B.; LEA, P.J.; AZEVEDO, R.A. Identification of maize
lines with contrasting responses to appliednitrogen. Journal of Plant Nutrition,
Philadelphia, v.28,p.903-915, 2005.
MENCUCCINI, M.; MAMBELLI, S.; COMSTOCK, J. Stomatal responsiveness to leaf
water status in common bean (Phaseolus vulgaris L.) is a function of time of day. Plant
Cell and Environment, v.23, p.1109-1118, 2000.
MICKELSON, S.M.; STUBER, C.S.; SENIOR, L.; KAEPPLER, S.M. Quantitative
trait loci controlling leaf and tassel traits in a B73 x MO17 population of maize. Crop
Science, Madison, v.42, p.1902-1909, 2002.

102
MIRANDA FILHO, J. B.; NASS, L. L. Hibridação no melhoramento. In: Recursos
Genéticos e Melhoramento - Plantas. Rondonópolis: Fundação MT, 2001. 1183 p.
MIRANDA, L.T.; MIRANDA, L.E.C.; SAWAZAKI, E. Genética ecológica e
melhoramento de milho. Campinas: Fundação Cargill, 1984. 30p.
MOULIA, B. Leaves as shell structures: double curvature, auto-stresses, and minimal
mechanical energy constraints on leaf rolling in grasses. Journal of Plant Growth
Regulation, v.19, p.19-30, 2000.
MORGAN, J.M. Osmoregulation and water stress in higher plants. Annual Review of
Plant Physiology, Bethesda, v.35, p.299-319, 1984.
MÜLLER, M.S.; FANCELLI, A.L.; DOURADO NETO, D. GARCIA, A.G.
OVEJERO, R.F.L. Produtividade do Panicum maximum cv. Mombaça irrigado, sob
pastejo rotacionado. Scientia Agrícola, v. 58, p.427-433. 2002.
NABLE, R. O.; ROBERTSON, M. J.; BERTHELSEN, S. Response of shoot growth
and transpiration to soil drying in sugarcane. Plant and Soil, v.207, p.59-65, 1999.
PATAKAS, A.; NIKOLAOU, N.; ZIOZIOU, E.; RADOGLOU, K.; NOITSAKI, B. The
role of organic solute and ion accumulation in osmotic adjustment in drought-stressed
grapevines. Plant Science, v.163, p.361-367, 2002.
PATERNIANI, M. E. A. G. Z.; BERNINI, C. S.; GUIMARÃES, P. S. e GALLO, P. B.
Seleção de progênies de meios-irmãos e estimativas de parâmetros genéticos da
população tolerante à seca de milho IAC Tol 1. In: 29 Congresso Nacional de Milho e
Sorgo, 2012, Campinas. Diversidade e Invações na era dos trasngênicos. Campinas:
IAC - Instituto Agronômico de Campinas, v.1. p.3126-3134, 2012.
PATERNIANI, E.; VENCOVSKY, R. Reciprocal recurrent selection in maize (Zea
mays L.) based on test crosses of half-sib families. Maydica, v.22, p.141-152, 1977.
PATERNIANI, E.; MIRANDA FILHO, J.B. Melhoramento de populações. In:
Paterniani, E (Ed.) Melhoramento e produção de milho no Brasil. ESALQ-
Piracicaba: Fundação Cargil, 1981.p.202-256.
PIMENTEL, C. Metabolismo de carbono na agricultura tropical. Seropédica:
EDUR, 1998. 159p.
PIMENTEL, C. A relação da planta com a água. Seropédica: UFRuralRJ, p.192,
2004.
PIMENTEL, C. Relações hídricas em dois híbridos de milho sob dois ciclos de
deficiência hídrica. Pesquisa Agropecuária Brasileira, v.34, p. 2021-2027, 1999.
PIMENTEL, C.; ROSSIELO, R.O.P. Entendimento sobre relações hídricas. In:
SIMPÓSIO INTERNACIONAL SOBRE ESTRESSE AMBIENTAL: O MILHO EM

103
PERSPECTIVA. Belo Horizonte, MG. Anais...EMBRAPA/CNPMS, 1995. v.1. 449p.
p.131-146, 1995.
PINHEIRO, C.; CHAVES, M.M. Photosynthesis and drought: can we make metabolic
connections from available data? Journal of Experimental Botany, v.62, p.869-882,
2011.
PINTO, R.J.B. Introdução ao Melhoramento Genético de Plantas. Maringá, 275p.,
1995.
QUEIROZ, C.G.S.; ALONSO, A.; MARES-GUIA, M.L.; MAGALHÃES, A.C.
Chilling-induced changes in membrane fluidity and antioxidant enzyme activities in
roots of (Coffea arabica L.) seedlings. Biol. Plant. v.41, p.403-413, 1998.
QUEIROZ, R.J.B. Resposta fisiológica e molecular de dois genótipos de milho à
limitação hídrica. 2010. 167p. Tese (Doutorado em Agronomia – Área de
concentração em Produção Vegetal). Faculdade de Ciências Agrárias e Veterinárias –
UNESP, Câmpus de Jaboticabal – SP, 2010.
REIS, M.C. Viabilidade da seleção recorrente recíproca em populações derivadas
de híbrido simples de milho. 2009. 81p. Tese (Doutorado em Genética e
Melhoramento de Plnatas). Universidade Federal de Lavras, 2009.
RESENDE, M.D.V. Correções nas expressões do progresso genético com seleção em
função da amostragem finita dentro de famílias de populações e implicações no
melhoramento florestal. Boletim Pesquisa Florestal, Colombo, n.22/23, p.61-77, 1991.
RIBAUT, J.M.; JIANG, C.; GONZALEZDELEON, D.; EDMEADES, G.O.;
HOISINGTON, D.A. Identification of quantitative trait loci under drought conditions in
tropical maize. Yield components and marker-assisted selection strategies. Theoretical
and Applied Genetics, New York, v.94, p. 887-896, 1997.
ROHÁCEK, K. Chlorophyll fluorescence parameters: the definitions, photosynthetic
meaning, and mutual relationships. Photosynthetica, Prague, v.40, p.13-29, 2002.
SANTOS, M.F.; AGUIAR, A.M.; FILHO, N.O. Efeitos da seleção recorrente recíproca
em um programa de milho. Anais do 2° Congresso Brasileiro de Melhoramento de
Plantas. Porto Seguro, Bahia, 2003.
SAUSEN, T. L. Respostas fisiológicas de Ricinuns communis à redução da
disponibilidade hídrica no solo. 87 f. Dissertacao (Mestrado) – Universidade Federal
do Rio Grande do Sul, Porto Alegre, 2007.
SCAPIM, C.A.; CARVALHO, C.G.P. de; CRUZ, C.D. Uma proposta de classificação
dos coeficientes de variação para a cultura do milho. Pesquisa Agropecuária
Brasileira, Brasília, v.30, n.5, p.683-686, maio 1995.
SEBBENN, A.M.; SIQUEIRA, A.C.M.F.; KAGEYAMA, P.Y.; MACHADO, J.A.R.
Parâmetros genéticos na conservação da cabreúva – Myroxylon peruiferum L.F.
Alemão. Scientia Forestalis, Piracicaba, n.53, p.31-38, 1998.

104
SILVA, A.R. Análise genética de caracteres quantitativos em milho com o
Delineamento III e marcadores moleculares. 2002. 143p. Tese (Doutorado em
Genética e Melhoramento de Plantas). Escola Superior de Agricultura “Luiz de
Queiroz”. Universidade de São Paulo, Piracicaba, 2002.
SILVA, M. de A.; JIFON, J. L.; SILVA, J. A. G. da; SHARMA, V. Use of
physiological parameters as fast tools to screen for drought tolerance in sugarcane.
Brazilian Journal of Plant Physiology, v.19, p.193-201, 2007.
SCHULZE, E.D.; ROBICHAUX, R.H.; GRACE, J.; RUNDEL, P.W & EHLERINGER,
J.R. Plant water balance. BioScience, v.37, p.30-37, 1987.
SMIRNOFF, N. The role of active oxygen in the response of plants to water deficit and
dessiccation. New Phytol., v.125, p.27-58, 1993.
SOARES FILHO, W.S. Características fenotípicas e genéticas das populações de
milho (Zea mays L.) braquítico Piranão – VD2B e Piranão – VF1B. 1987. 185p.
Tese (Doutorado em Genética e Melhoramento de Plantas). Escola Superior de
Agricultura “Luiz de Queiroz”. Universidade de São Paulo, Piracicaba, 1987.
SOUZA JUNIOR, C.L.; GERALDI, I.O.; ZINSLY, J.R. Influence of tassel size on the
expression of prolificacy in maize. Maydica, Bergamo,v.30, p.321-328, 1985.
SOUZA JUNIOR, C. L. Melhoramento de espécies alógamas. In: NASS, L. L.;
VALOIS, A. C. C.; MELO I. S.; VALADARESINGLIS, M. C. Recursos genéticos
e melhoramento de plantas. Rondonópolis: Fundação Rondonópolis, p.159-199, 2001.
SOUZA JUNIOR, C. L.; PINTO, R.M.C. Responses to a short-term reciprocal recurrent
selection procedure im maize. Maydica, Bergamo, v.45, p.21-28, 2000.
TAIZ, L. & ZEIGER, E. Fisiologia Vegetal. 4.ed. Porto Alegre: E.Atmed, p.819, 2009.
TESTER, M.; BACIC, A. Abiotic stress tolerance in grasses. From model plants to crop
plants. Plant Physiology, v.137, p.791-793, 2005.
TOLLENAAR, M.; WU, J. Yield improvement in temperate maize attributable to
greater stress tolerance. Crop Science, v.39, p.1597-1604, 1999.
TORRES NETTO, A.; CAMPOSTRINI, E.; OLIVEIRA, J. G. DE; BRESSAN SMITH,
R. E. Photosynthetic pigments, nitrogen, chlorophyll a fluorescence and SPAD-502
readings in coffee leaves. Scientia Horticulturae, v.104, p.199-209, 2005.
TURNER, N.C.; JONES, M.M. Turgor maintenance by osmotic adjustment: a
review and evaluation. In: TURNER, N.C.; KRAMER, P.J. Adaptation of plants to
water and higher temperature stress. New York: J. Wiley, p.87-103, 1980.
USDA: United States Department of Agriculture. Disponível em:
<http://www.usda.gov/wps/portal/usda/FOODNUTRITION&navtype> Acesso em: 17
de agosto de 2012.

105
VAN HANDEL, E. Direct microdetermination of sucrose. Anal. Biochem., 22:280-
283, 1968.
VARGA, B.; SVECNJAK, Z.; KNEZEVIC, M.;GRBESA, D. Performance of prolific
and nonprolific maize hybrids under reduced-input and high-input cropping systems.
Field Crops Research, Amsterdam, v.90, p.203-212, 2004.
VENCOVSKY, R.; BARRIGA, P. Genética biométrica no fitomelhoramento.
Ribeirão Preto: Sociedade Brasileira de Genética, p.496, 1992.
VENCOVSKY, R.; CRUZ, C.D. Comparação de métodos de correção do rendimento
de parcelas com estandes variados. I. Dados simulados. Pesquisa Agropecuária
Brasileira, Brasília, n.5, v.26, p.647-657, 1991.
VERSLUES, P. E.; AGARWAL, M.; KATIYAR-AGARWAL, S.; ZHU, J.; ZHU, J. K.
Methods and concepts in quantifying resistance to drought,salt and freezing, abiotic
stresses that affect plant water status. The Plant Journal, Blackwell, v. 45, p. 523–539,
2006.
VIANA, M.C.M. Déficit hídrico em genótipos de milho com tolerância diferencial à
seca. 2002. 75 p. Dissertação (Mestrado em Biologia Vegetal) – Universidade Federal
de Minas Gerais, Belo Horizonte, 2002.
VIEIRA JUNIOR, P. A.; DOURADO NETO, D.; FERRAZ DE OLIVEIRA, R.;
PERES, L.E.P.; MARTIN, T.N.; MANFRON, P.A.; BONNECARRÈRE. Relações
entre o potencial e a temperatura da folha de plantas de milho e sorgo submetidas a
estresse hídrico. Acta Scientiarum Agronomy, Maringá, v. 29, p. 555-561, 2007.
XIAO, Y.N.; LI, X.H.; ZHANG, S.H.; WANG, X.D.; LI, M.S.; ZHENG,Y.L.
Identification of quantitative trait loci (QTLs) for flowering time using SSR marker in
maize under water stress. Korean Journal of Genetics, Seoul, v.26, n.4, p.405-413,
2004.
XU, Z.; ZHOU, G.; SHIMIZU, H. Are plant growth and photosynthesis limited by
predrought following rewatering in grass? Journal of Experimental Botany, Oxford,
v.60, p. 3737-3749, 2009.
XOCONOSTLE-CAZARES, B.; RAMIREZ-ORTEGA, F.A.; FLORES-ELENES, L.
and RUIZ-MEDRANO. Drought tolerance in crop plants. Americam Journal Plant
Physiology, v.5, p.241-256, 2010.
ZAIDI, P.H.; SRINIVASAN, G.; CORDOVA, H.S.; SANCHEZ, C. Gains from
improvement for mid-season drought tolerance in tropical maize (Zea mays L.) Field
Crops Research, Amsterdam, v.89, p.135-152, 2004.
ZHU, J.K. Plant salt tolerance. Trends Plant Science. v.6, p.66–71, 2001.
106
7 ANEXOS
Anexo 1. Médias de florescimento masculino (FM), florescimento feminino (FF), intervalo
entre florescimentos (IF), stay green (SG), número de ramificação de pendão (NRP),
comprimento de pendão (CP), altura da planta(AP) e da altura da espiga (AE), referentes a 30
progênies de irmãos germanos interpopulacionais de milho e de quatro testemunhas do
Experimento 1. Campinas, 2012.
Progênies FM FF IF SG NRP CP AP AE
dias nota r. pendão-1
cm
102A x 89B 64,67 66,00 1,33 2,07 9,80 38,80 201 114
83A x 77B 60,33 61,67 1,33 2,07 17,10 31,85 216 124
96A x 91B 61,67 63,33 1,67 1,73 13,10 34,90 194 108
4B x 2A 63,33 64,67 1,33 2,13 12,00 32,10 210 107
10B x 13A 61,00 62,33 1,33 2,07 14,70 35,60 205 109
55B x 79A 68,67 69,33 0,67 2,20 14,50 30,90 210 116
54A x 115B 62,00 63,00 1,00 2,20 8,10 33,65 187 101
104B x 94A 61,67 61,33 -0,33 2,00 11,90 34,40 205 114
86B x 71A 60,00 60,33 0,33 1,93 10,60 32,40 224 123
11B x9A 56,00 56,33 0,33 1,93 11,30 35,40 217 114
155B x 157A 60,33 61,33 1,00 2,27 13,10 36,40 210 114
32A x 40B 59,00 61,67 2,67 1,53 10,10 34,70 202 117
84B x 119A 65,67 66,33 0,67 1,93 10,50 36,10 206 115
16B x 15A 66,00 65,00 -1,00 1,93 12,00 31,60 180 92
121B x116A 65,00 65,67 0,67 1,80 14,30 34,10 204 108
19B x 21A 62,33 63,00 0,67 1,47 14,80 35,90 208 110
152A x 126B 63,00 66,00 3,00 1,80 11,10 33,90 185 104
36B x 43A 63,00 64,00 1,00 1,73 12,50 36,90 210 117
18B x 17A 63,00 65,00 2,00 1,53 10,60 33,20 180 95
34B x 40A 61,67 63,33 1,67 2,67 11,20 33,40 206 128
126A x 127B 70,00 72,00 2,00 2,07 9,90 31,60 193 104
32A x 22B 63,33 64,00 0,67 1,73 13,70 33,80 209 124
82B x 60A 59,33 61,00 1,67 1,87 13,30 31,65 203 106
5A x 2B 59,67 61,33 1,67 1,87 11,90 36,90 211 105
120A x 95B 64,33 66,33 2,00 1,87 11,80 36,50 207 115
99B x 53A 67,00 68,00 1,00 2,47 8,20 35,05 192 104
90A x 78B 64,00 65,00 1,00 1,67 11,40 35,20 201 113
168B x 47A 66,33 67,00 0,67 1,93 12,50 38,10 215 122
16A x 6B 60,67 62,67 1,73 1,73 7,10 37,60 190 96
13B x 12A 63,67 63,67 0,00 2,40 10,70 36,60 201 103
Média prog. 62,89 64,02 1,13 1,95 11,79 34,64 203 111
Pop. tolerante 67,00 69,33 2,33 1,60 13,20 30,60 178 93
Pop. sensível 65,67 66,67 1,00 2,00 13,20 30,10 197 112
FT510 68,33 68,00 -0,33 2,47 12,10 30,03 167 105
DAS2B707 66,67 66,00 -0,67 1,93 9,40 36,40 201 111
Média test. 66,92 67,50 0,58 2,00 11,98 31,78 188 105
107
Anexo 2. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número de
fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 1. Campinas, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
102A x 89B 16,66 5,10 16,77 0,89 6262,96 117,43 118,99
83A x 77B 16,10 4,76 15,35 1,12 7980,74 149,64 151,63
96A x 91B 16,91 4,74 13,45 1,06 3898,52 73,10 74,07
4B x 2A 16,88 4,78 15,23 1,03 6180,00 115,88 117,42
10B x 13A 16,23 4,75 16,00 1,05 6159,26 115,49 117,02
55B x 79A 14,33 5,22 14,13 1,06 5332,59 99,99 101,32
54A x 115B 15,34 4,55 17,35 0,99 5039,26 94,49 95,74
104B x 94A 17,15 4,63 13,82 0,81 5899,26 110,61 112,08
86B x 71A 17,47 5,06 16,07 0,93 7094,07 133,01 134,78
11B x9A 16,86 5,89 14,88 1,05 3614,81 67,78 68,68
155B x 157A 18,17 5,20 13,50 0,94 5115,55 95,92 97,19
32A x 40B 16,40 5,77 14,78 1,01 6120,74 114,76 116,28
84B x 119A 17,48 5,48 14,95 1,01 5562,22 104,29 105,67
16B x 15A 16,70 4,99 15,87 0,72 5622,22 105,42 106,82
121B x116A 15,91 6,05 15,05 1,04 6945,93 130,24 131,97
19B x 21A 16,18 5,11 17,82 0,94 5070,37 95,07 96,33
152A x 126B 15,82 5,02 15,68 1,06 5194,81 97,40 98,69
36B x 43A 19,12 6,01 16,23 1,07 6622,22 124,17 125,82
18B x 17A 15,49 4,91 14,70 1,11 4758,52 89,22 90,40
34B x 40A 15,17 4,00 15,10 0,90 5993,33 112,38 113,87
126A x 127B 15,78 5,03 16,03 1,17 6078,52 113,97 115,48
32A x 22B 15,89 5,33 15,87 0,95 4710,37 88,32 89,49
82B x 60A 15,21 4,94 16,22 1,10 5249,63 98,43 99,74
5A x 2B 17,08 5,54 13,68 1,03 6651,85 124,72 126,38
120A x 95B 17,59 4,96 16,00 1,17 7500,74 140,64 142,51
99B x 53A 14,99 5,39 15,08 1,06 5508,15 103,28 104,65
90A x 78B 15,58 5,09 16,00 0,96 5950,37 111,57 113,05
168B x 47A 17,08 4,74 15,30 0,96 6700,00 125,63 127,30
16A x 6B 17,68 5,59 17,67 0,92 5988,15 112,28 113,77
13B x 12A 17,13 5,49 16,28 1,07 6086,67 114,13 115,64
Média prog. 16,48 5,14 15,50 1,01 5829,73 109,31 110,76
Pop. tolerante 16,68 4,94 15,73 1,02 6578,52 123,35 124,99
Pop. sensível 16,00 5,74 17,74 0,85 5060,00 94,88 96,14
FT510 15,40 4,81 15,33 0,94 4365,52 90,80 92,01
DAS2B707 16,23 5,50 16,43 1,06 5263,70 98,69 100,00
Média test. 16,08 5,25 16,31 0,97 5316,94 101,93 103,29
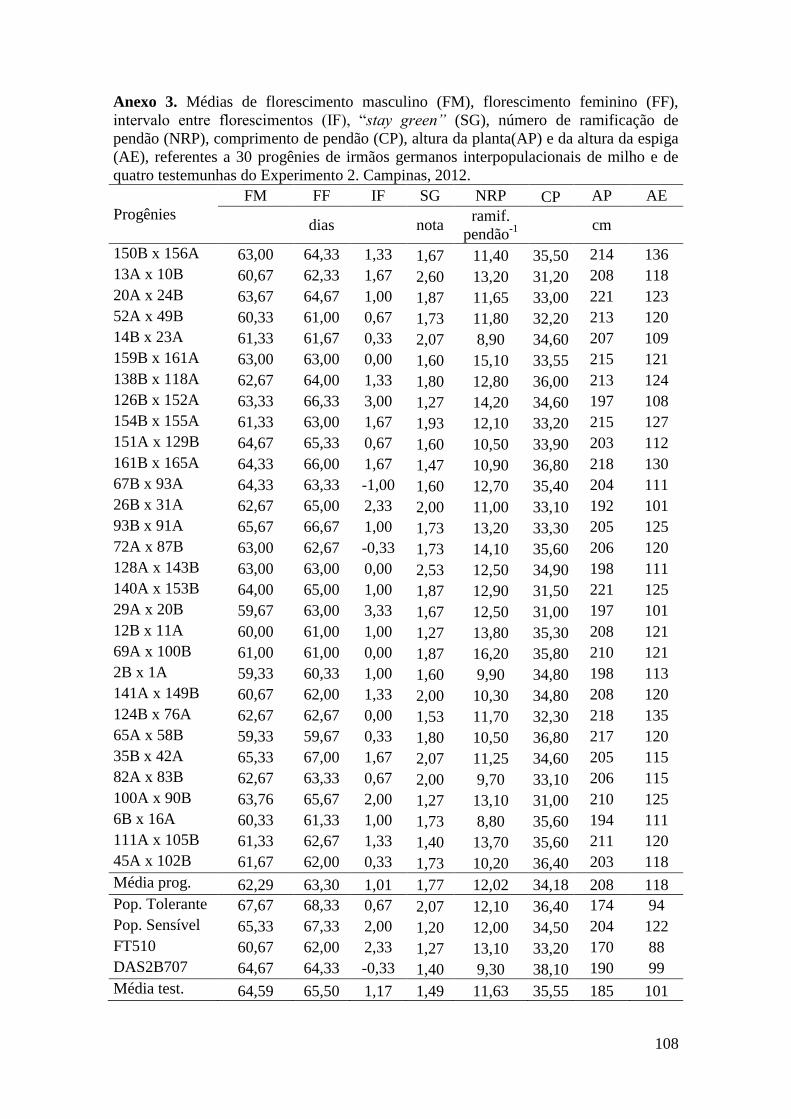
108
Anexo 3. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 2. Campinas, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
150B x 156A 63,00 64,33 1,33 1,67 11,40 35,50 214 136
13A x 10B 60,67 62,33 1,67 2,60 13,20 31,20 208 118
20A x 24B 63,67 64,67 1,00 1,87 11,65 33,00 221 123
52A x 49B 60,33 61,00 0,67 1,73 11,80 32,20 213 120
14B x 23A 61,33 61,67 0,33 2,07 8,90 34,60 207 109
159B x 161A 63,00 63,00 0,00 1,60 15,10 33,55 215 121
138B x 118A 62,67 64,00 1,33 1,80 12,80 36,00 213 124
126B x 152A 63,33 66,33 3,00 1,27 14,20 34,60 197 108
154B x 155A 61,33 63,00 1,67 1,93 12,10 33,20 215 127
151A x 129B 64,67 65,33 0,67 1,60 10,50 33,90 203 112
161B x 165A 64,33 66,00 1,67 1,47 10,90 36,80 218 130
67B x 93A 64,33 63,33 -1,00 1,60 12,70 35,40 204 111
26B x 31A 62,67 65,00 2,33 2,00 11,00 33,10 192 101
93B x 91A 65,67 66,67 1,00 1,73 13,20 33,30 205 125
72A x 87B 63,00 62,67 -0,33 1,73 14,10 35,60 206 120
128A x 143B 63,00 63,00 0,00 2,53 12,50 34,90 198 111
140A x 153B 64,00 65,00 1,00 1,87 12,90 31,50 221 125
29A x 20B 59,67 63,00 3,33 1,67 12,50 31,00 197 101
12B x 11A 60,00 61,00 1,00 1,27 13,80 35,30 208 121
69A x 100B 61,00 61,00 0,00 1,87 16,20 35,80 210 121
2B x 1A 59,33 60,33 1,00 1,60 9,90 34,80 198 113
141A x 149B 60,67 62,00 1,33 2,00 10,30 34,80 208 120
124B x 76A 62,67 62,67 0,00 1,53 11,70 32,30 218 135
65A x 58B 59,33 59,67 0,33 1,80 10,50 36,80 217 120
35B x 42A 65,33 67,00 1,67 2,07 11,25 34,60 205 115
82A x 83B 62,67 63,33 0,67 2,00 9,70 33,10 206 115
100A x 90B 63,76 65,67 2,00 1,27 13,10 31,00 210 125
6B x 16A 60,33 61,33 1,00 1,73 8,80 35,60 194 111
111A x 105B 61,33 62,67 1,33 1,40 13,70 35,60 211 120
45A x 102B 61,67 62,00 0,33 1,73 10,20 36,40 203 118
Média prog. 62,29 63,30 1,01 1,77 12,02 34,18 208 118
Pop. Tolerante 67,67 68,33 0,67 2,07 12,10 36,40 174 94
Pop. Sensível 65,33 67,33 2,00 1,20 12,00 34,50 204 122
FT510 60,67 62,00 2,33 1,27 13,10 33,20 170 88
DAS2B707 64,67 64,33 -0,33 1,40 9,30 38,10 190 99
Média test. 64,59 65,50 1,17 1,49 11,63 35,55 185 101
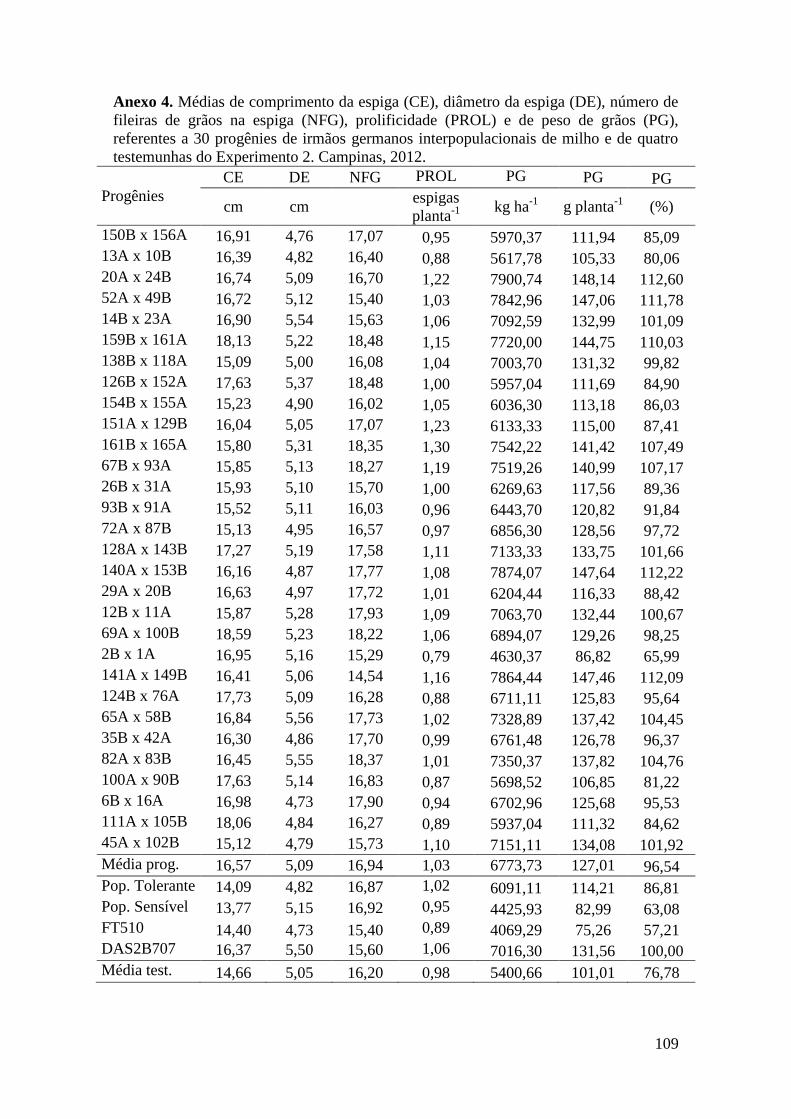
109
Anexo 4. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número de
fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 2. Campinas, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
150B x 156A 16,91 4,76 17,07 0,95 5970,37 111,94 85,09
13A x 10B 16,39 4,82 16,40 0,88 5617,78 105,33 80,06
20A x 24B 16,74 5,09 16,70 1,22 7900,74 148,14 112,60
52A x 49B 16,72 5,12 15,40 1,03 7842,96 147,06 111,78
14B x 23A 16,90 5,54 15,63 1,06 7092,59 132,99 101,09
159B x 161A 18,13 5,22 18,48 1,15 7720,00 144,75 110,03
138B x 118A 15,09 5,00 16,08 1,04 7003,70 131,32 99,82
126B x 152A 17,63 5,37 18,48 1,00 5957,04 111,69 84,90
154B x 155A 15,23 4,90 16,02 1,05 6036,30 113,18 86,03
151A x 129B 16,04 5,05 17,07 1,23 6133,33 115,00 87,41
161B x 165A 15,80 5,31 18,35 1,30 7542,22 141,42 107,49
67B x 93A 15,85 5,13 18,27 1,19 7519,26 140,99 107,17
26B x 31A 15,93 5,10 15,70 1,00 6269,63 117,56 89,36
93B x 91A 15,52 5,11 16,03 0,96 6443,70 120,82 91,84
72A x 87B 15,13 4,95 16,57 0,97 6856,30 128,56 97,72
128A x 143B 17,27 5,19 17,58 1,11 7133,33 133,75 101,66
140A x 153B 16,16 4,87 17,77 1,08 7874,07 147,64 112,22
29A x 20B 16,63 4,97 17,72 1,01 6204,44 116,33 88,42
12B x 11A 15,87 5,28 17,93 1,09 7063,70 132,44 100,67
69A x 100B 18,59 5,23 18,22 1,06 6894,07 129,26 98,25
2B x 1A 16,95 5,16 15,29 0,79 4630,37 86,82 65,99
141A x 149B 16,41 5,06 14,54 1,16 7864,44 147,46 112,09
124B x 76A 17,73 5,09 16,28 0,88 6711,11 125,83 95,64
65A x 58B 16,84 5,56 17,73 1,02 7328,89 137,42 104,45
35B x 42A 16,30 4,86 17,70 0,99 6761,48 126,78 96,37
82A x 83B 16,45 5,55 18,37 1,01 7350,37 137,82 104,76
100A x 90B 17,63 5,14 16,83 0,87 5698,52 106,85 81,22
6B x 16A 16,98 4,73 17,90 0,94 6702,96 125,68 95,53
111A x 105B 18,06 4,84 16,27 0,89 5937,04 111,32 84,62
45A x 102B 15,12 4,79 15,73 1,10 7151,11 134,08 101,92
Média prog. 16,57 5,09 16,94 1,03 6773,73 127,01 96,54
Pop. Tolerante 14,09 4,82 16,87 1,02 6091,11 114,21 86,81
Pop. Sensível 13,77 5,15 16,92 0,95 4425,93 82,99 63,08
FT510 14,40 4,73 15,40 0,89 4069,29 75,26 57,21
DAS2B707 16,37 5,50 15,60 1,06 7016,30 131,56 100,00
Média test. 14,66 5,05 16,20 0,98 5400,66 101,01 76,78
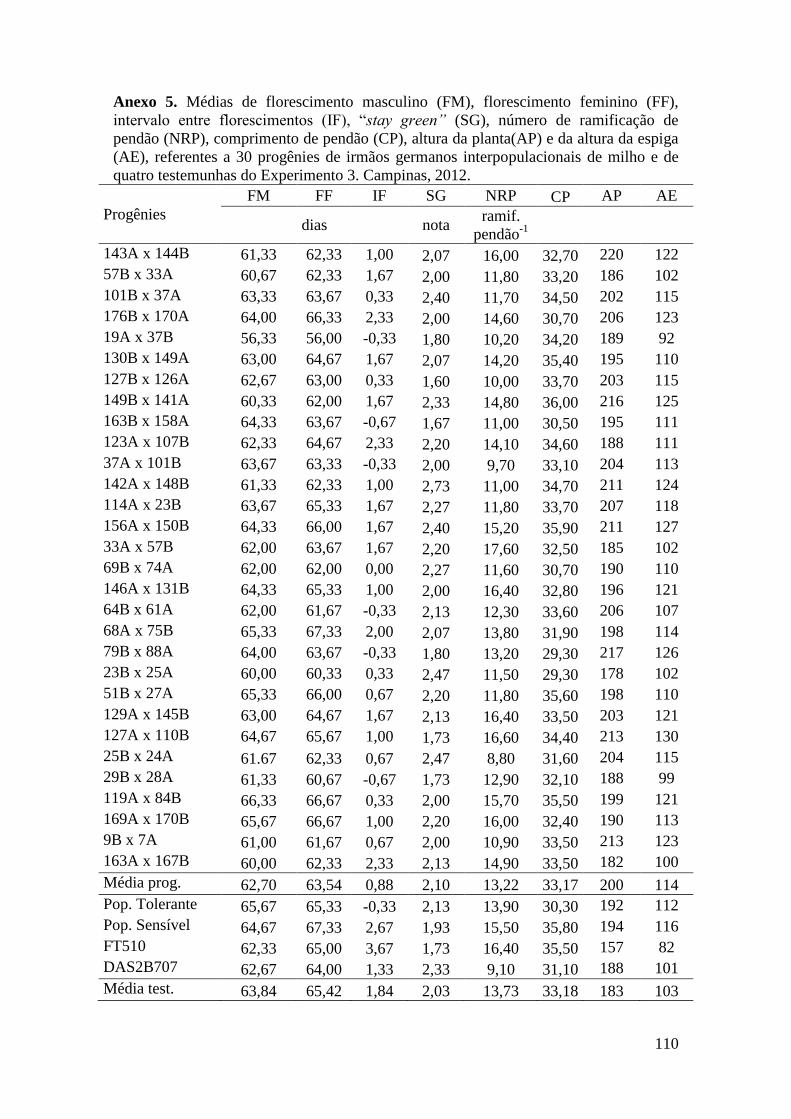
110
Anexo 5. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 3. Campinas, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
143A x 144B 61,33 62,33 1,00 2,07 16,00 32,70 220 122
57B x 33A 60,67 62,33 1,67 2,00 11,80 33,20 186 102
101B x 37A 63,33 63,67 0,33 2,40 11,70 34,50 202 115
176B x 170A 64,00 66,33 2,33 2,00 14,60 30,70 206 123
19A x 37B 56,33 56,00 -0,33 1,80 10,20 34,20 189 92
130B x 149A 63,00 64,67 1,67 2,07 14,20 35,40 195 110
127B x 126A 62,67 63,00 0,33 1,60 10,00 33,70 203 115
149B x 141A 60,33 62,00 1,67 2,33 14,80 36,00 216 125
163B x 158A 64,33 63,67 -0,67 1,67 11,00 30,50 195 111
123A x 107B 62,33 64,67 2,33 2,20 14,10 34,60 188 111
37A x 101B 63,67 63,33 -0,33 2,00 9,70 33,10 204 113
142A x 148B 61,33 62,33 1,00 2,73 11,00 34,70 211 124
114A x 23B 63,67 65,33 1,67 2,27 11,80 33,70 207 118
156A x 150B 64,33 66,00 1,67 2,40 15,20 35,90 211 127
33A x 57B 62,00 63,67 1,67 2,20 17,60 32,50 185 102
69B x 74A 62,00 62,00 0,00 2,27 11,60 30,70 190 110
146A x 131B 64,33 65,33 1,00 2,00 16,40 32,80 196 121
64B x 61A 62,00 61,67 -0,33 2,13 12,30 33,60 206 107
68A x 75B 65,33 67,33 2,00 2,07 13,80 31,90 198 114
79B x 88A 64,00 63,67 -0,33 1,80 13,20 29,30 217 126
23B x 25A 60,00 60,33 0,33 2,47 11,50 29,30 178 102
51B x 27A 65,33 66,00 0,67 2,20 11,80 35,60 198 110
129A x 145B 63,00 64,67 1,67 2,13 16,40 33,50 203 121
127A x 110B 64,67 65,67 1,00 1,73 16,60 34,40 213 130
25B x 24A 61.67 62,33 0,67 2,47 8,80 31,60 204 115
29B x 28A 61,33 60,67 -0,67 1,73 12,90 32,10 188 99
119A x 84B 66,33 66,67 0,33 2,00 15,70 35,50 199 121
169A x 170B 65,67 66,67 1,00 2,20 16,00 32,40 190 113
9B x 7A 61,00 61,67 0,67 2,00 10,90 33,50 213 123
163A x 167B 60,00 62,33 2,33 2,13 14,90 33,50 182 100
Média prog. 62,70 63,54 0,88 2,10 13,22 33,17 200 114
Pop. Tolerante 65,67 65,33 -0,33 2,13 13,90 30,30 192 112
Pop. Sensível 64,67 67,33 2,67 1,93 15,50 35,80 194 116
FT510 62,33 65,00 3,67 1,73 16,40 35,50 157 82
DAS2B707 62,67 64,00 1,33 2,33 9,10 31,10 188 101
Média test. 63,84 65,42 1,84 2,03 13,73 33,18 183 103
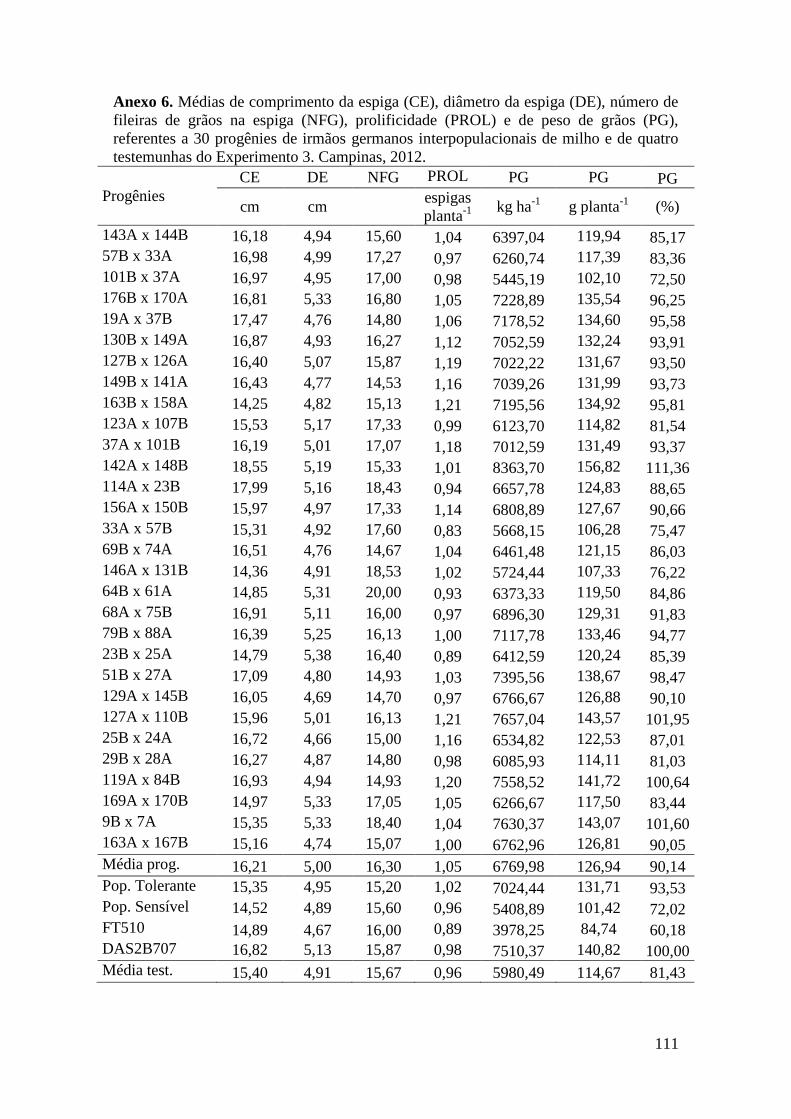
111
Anexo 6. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número de
fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 3. Campinas, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
143A x 144B 16,18 4,94 15,60 1,04 6397,04 119,94 85,17
57B x 33A 16,98 4,99 17,27 0,97 6260,74 117,39 83,36
101B x 37A 16,97 4,95 17,00 0,98 5445,19 102,10 72,50
176B x 170A 16,81 5,33 16,80 1,05 7228,89 135,54 96,25
19A x 37B 17,47 4,76 14,80 1,06 7178,52 134,60 95,58
130B x 149A 16,87 4,93 16,27 1,12 7052,59 132,24 93,91
127B x 126A 16,40 5,07 15,87 1,19 7022,22 131,67 93,50
149B x 141A 16,43 4,77 14,53 1,16 7039,26 131,99 93,73
163B x 158A 14,25 4,82 15,13 1,21 7195,56 134,92 95,81
123A x 107B 15,53 5,17 17,33 0,99 6123,70 114,82 81,54
37A x 101B 16,19 5,01 17,07 1,18 7012,59 131,49 93,37
142A x 148B 18,55 5,19 15,33 1,01 8363,70 156,82 111,36
114A x 23B 17,99 5,16 18,43 0,94 6657,78 124,83 88,65
156A x 150B 15,97 4,97 17,33 1,14 6808,89 127,67 90,66
33A x 57B 15,31 4,92 17,60 0,83 5668,15 106,28 75,47
69B x 74A 16,51 4,76 14,67 1,04 6461,48 121,15 86,03
146A x 131B 14,36 4,91 18,53 1,02 5724,44 107,33 76,22
64B x 61A 14,85 5,31 20,00 0,93 6373,33 119,50 84,86
68A x 75B 16,91 5,11 16,00 0,97 6896,30 129,31 91,83
79B x 88A 16,39 5,25 16,13 1,00 7117,78 133,46 94,77
23B x 25A 14,79 5,38 16,40 0,89 6412,59 120,24 85,39
51B x 27A 17,09 4,80 14,93 1,03 7395,56 138,67 98,47
129A x 145B 16,05 4,69 14,70 0,97 6766,67 126,88 90,10
127A x 110B 15,96 5,01 16,13 1,21 7657,04 143,57 101,95
25B x 24A 16,72 4,66 15,00 1,16 6534,82 122,53 87,01
29B x 28A 16,27 4,87 14,80 0,98 6085,93 114,11 81,03
119A x 84B 16,93 4,94 14,93 1,20 7558,52 141,72 100,64
169A x 170B 14,97 5,33 17,05 1,05 6266,67 117,50 83,44
9B x 7A 15,35 5,33 18,40 1,04 7630,37 143,07 101,60
163A x 167B 15,16 4,74 15,07 1,00 6762,96 126,81 90,05
Média prog. 16,21 5,00 16,30 1,05 6769,98 126,94 90,14
Pop. Tolerante 15,35 4,95 15,20 1,02 7024,44 131,71 93,53
Pop. Sensível 14,52 4,89 15,60 0,96 5408,89 101,42 72,02
FT510 14,89 4,67 16,00 0,89 3978,25 84,74 60,18
DAS2B707 16,82 5,13 15,87 0,98 7510,37 140,82 100,00
Média test. 15,40 4,91 15,67 0,96 5980,49 114,67 81,43
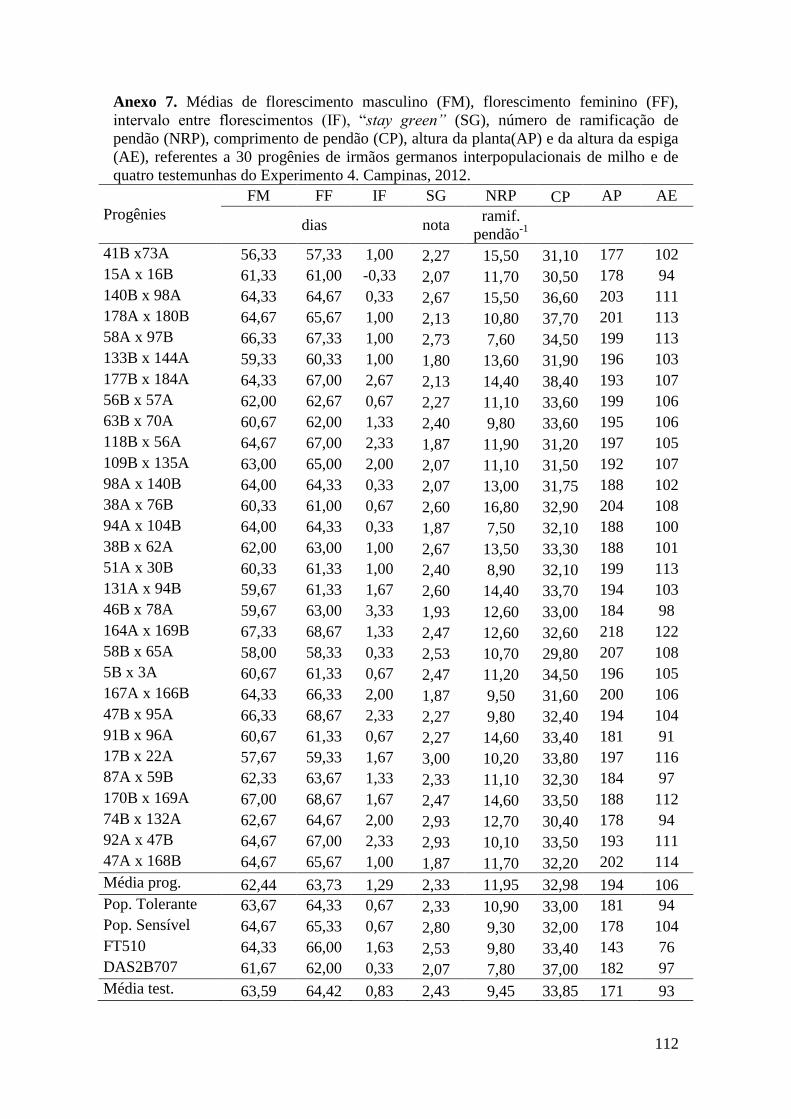
112
Anexo 7. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 4. Campinas, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
41B x73A 56,33 57,33 1,00 2,27 15,50 31,10 177 102
15A x 16B 61,33 61,00 -0,33 2,07 11,70 30,50 178 94
140B x 98A 64,33 64,67 0,33 2,67 15,50 36,60 203 111
178A x 180B 64,67 65,67 1,00 2,13 10,80 37,70 201 113
58A x 97B 66,33 67,33 1,00 2,73 7,60 34,50 199 113
133B x 144A 59,33 60,33 1,00 1,80 13,60 31,90 196 103
177B x 184A 64,33 67,00 2,67 2,13 14,40 38,40 193 107
56B x 57A 62,00 62,67 0,67 2,27 11,10 33,60 199 106
63B x 70A 60,67 62,00 1,33 2,40 9,80 33,60 195 106
118B x 56A 64,67 67,00 2,33 1,87 11,90 31,20 197 105
109B x 135A 63,00 65,00 2,00 2,07 11,10 31,50 192 107
98A x 140B 64,00 64,33 0,33 2,07 13,00 31,75 188 102
38A x 76B 60,33 61,00 0,67 2,60 16,80 32,90 204 108
94A x 104B 64,00 64,33 0,33 1,87 7,50 32,10 188 100
38B x 62A 62,00 63,00 1,00 2,67 13,50 33,30 188 101
51A x 30B 60,33 61,33 1,00 2,40 8,90 32,10 199 113
131A x 94B 59,67 61,33 1,67 2,60 14,40 33,70 194 103
46B x 78A 59,67 63,00 3,33 1,93 12,60 33,00 184 98
164A x 169B 67,33 68,67 1,33 2,47 12,60 32,60 218 122
58B x 65A 58,00 58,33 0,33 2,53 10,70 29,80 207 108
5B x 3A 60,67 61,33 0,67 2,47 11,20 34,50 196 105
167A x 166B 64,33 66,33 2,00 1,87 9,50 31,60 200 106
47B x 95A 66,33 68,67 2,33 2,27 9,80 32,40 194 104
91B x 96A 60,67 61,33 0,67 2,27 14,60 33,40 181 91
17B x 22A 57,67 59,33 1,67 3,00 10,20 33,80 197 116
87A x 59B 62,33 63,67 1,33 2,33 11,10 32,30 184 97
170B x 169A 67,00 68,67 1,67 2,47 14,60 33,50 188 112
74B x 132A 62,67 64,67 2,00 2,93 12,70 30,40 178 94
92A x 47B 64,67 67,00 2,33 2,93 10,10 33,50 193 111
47A x 168B 64,67 65,67 1,00 1,87 11,70 32,20 202 114
Média prog. 62,44 63,73 1,29 2,33 11,95 32,98 194 106
Pop. Tolerante 63,67 64,33 0,67 2,33 10,90 33,00 181 94
Pop. Sensível 64,67 65,33 0,67 2,80 9,30 32,00 178 104
FT510 64,33 66,00 1,63 2,53 9,80 33,40 143 76
DAS2B707 61,67 62,00 0,33 2,07 7,80 37,00 182 97
Média test. 63,59 64,42 0,83 2,43 9,45 33,85 171 93
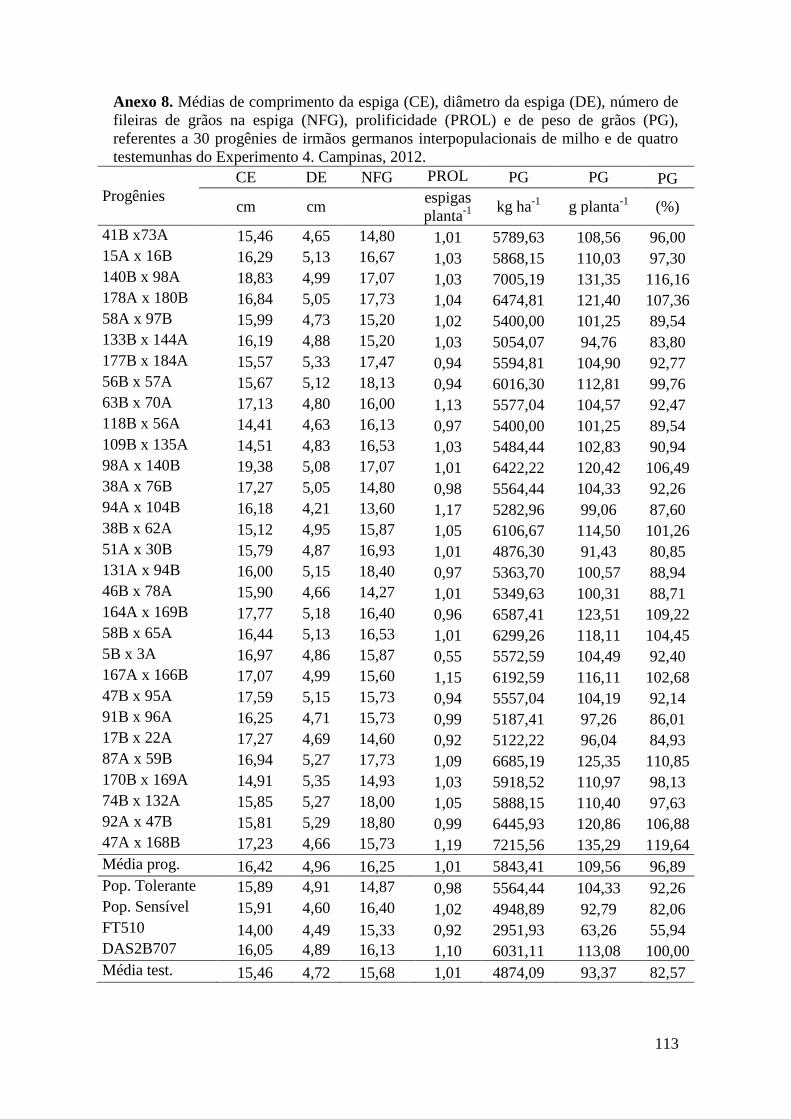
113
Anexo 8. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número de
fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 4. Campinas, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
41B x73A 15,46 4,65 14,80 1,01 5789,63 108,56 96,00
15A x 16B 16,29 5,13 16,67 1,03 5868,15 110,03 97,30
140B x 98A 18,83 4,99 17,07 1,03 7005,19 131,35 116,16
178A x 180B 16,84 5,05 17,73 1,04 6474,81 121,40 107,36
58A x 97B 15,99 4,73 15,20 1,02 5400,00 101,25 89,54
133B x 144A 16,19 4,88 15,20 1,03 5054,07 94,76 83,80
177B x 184A 15,57 5,33 17,47 0,94 5594,81 104,90 92,77
56B x 57A 15,67 5,12 18,13 0,94 6016,30 112,81 99,76
63B x 70A 17,13 4,80 16,00 1,13 5577,04 104,57 92,47
118B x 56A 14,41 4,63 16,13 0,97 5400,00 101,25 89,54
109B x 135A 14,51 4,83 16,53 1,03 5484,44 102,83 90,94
98A x 140B 19,38 5,08 17,07 1,01 6422,22 120,42 106,49
38A x 76B 17,27 5,05 14,80 0,98 5564,44 104,33 92,26
94A x 104B 16,18 4,21 13,60 1,17 5282,96 99,06 87,60
38B x 62A 15,12 4,95 15,87 1,05 6106,67 114,50 101,26
51A x 30B 15,79 4,87 16,93 1,01 4876,30 91,43 80,85
131A x 94B 16,00 5,15 18,40 0,97 5363,70 100,57 88,94
46B x 78A 15,90 4,66 14,27 1,01 5349,63 100,31 88,71
164A x 169B 17,77 5,18 16,40 0,96 6587,41 123,51 109,22
58B x 65A 16,44 5,13 16,53 1,01 6299,26 118,11 104,45
5B x 3A 16,97 4,86 15,87 0,55 5572,59 104,49 92,40
167A x 166B 17,07 4,99 15,60 1,15 6192,59 116,11 102,68
47B x 95A 17,59 5,15 15,73 0,94 5557,04 104,19 92,14
91B x 96A 16,25 4,71 15,73 0,99 5187,41 97,26 86,01
17B x 22A 17,27 4,69 14,60 0,92 5122,22 96,04 84,93
87A x 59B 16,94 5,27 17,73 1,09 6685,19 125,35 110,85
170B x 169A 14,91 5,35 14,93 1,03 5918,52 110,97 98,13
74B x 132A 15,85 5,27 18,00 1,05 5888,15 110,40 97,63
92A x 47B 15,81 5,29 18,80 0,99 6445,93 120,86 106,88
47A x 168B 17,23 4,66 15,73 1,19 7215,56 135,29 119,64
Média prog. 16,42 4,96 16,25 1,01 5843,41 109,56 96,89
Pop. Tolerante 15,89 4,91 14,87 0,98 5564,44 104,33 92,26
Pop. Sensível 15,91 4,60 16,40 1,02 4948,89 92,79 82,06
FT510 14,00 4,49 15,33 0,92 2951,93 63,26 55,94
DAS2B707 16,05 4,89 16,13 1,10 6031,11 113,08 100,00
Média test. 15,46 4,72 15,68 1,01 4874,09 93,37 82,57
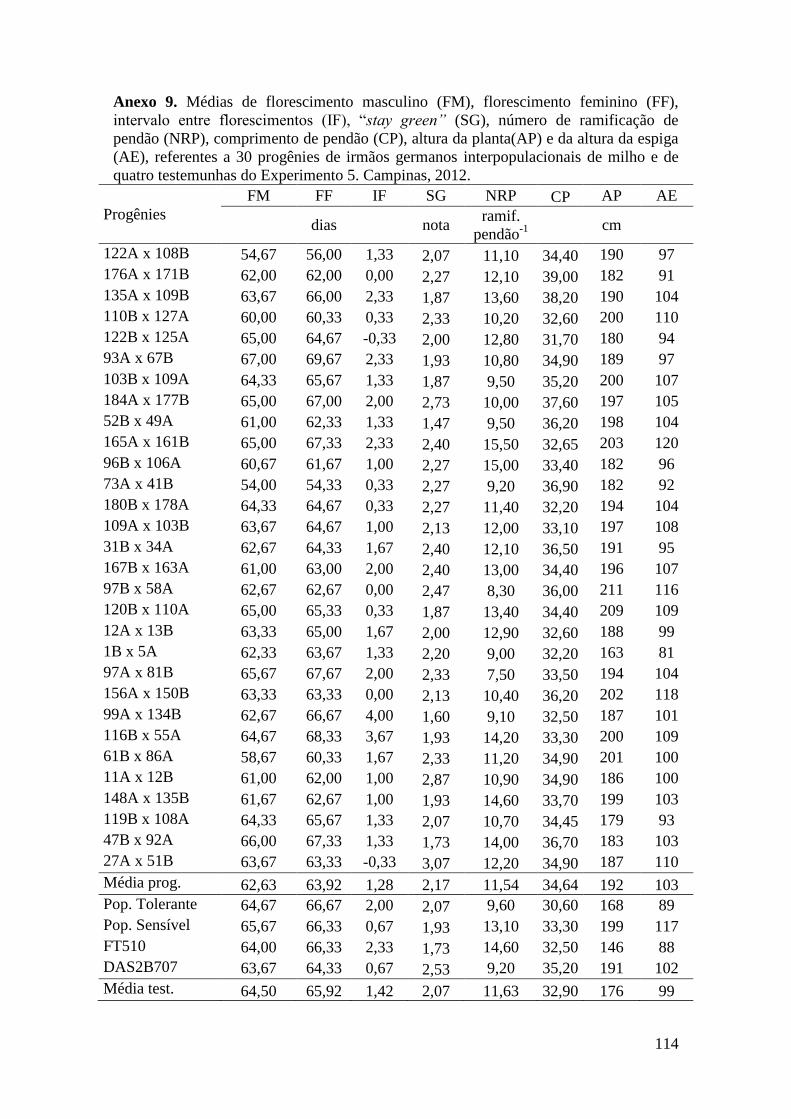
114
Anexo 9. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 5. Campinas, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
122A x 108B 54,67 56,00 1,33 2,07 11,10 34,40 190 97
176A x 171B 62,00 62,00 0,00 2,27 12,10 39,00 182 91
135A x 109B 63,67 66,00 2,33 1,87 13,60 38,20 190 104
110B x 127A 60,00 60,33 0,33 2,33 10,20 32,60 200 110
122B x 125A 65,00 64,67 -0,33 2,00 12,80 31,70 180 94
93A x 67B 67,00 69,67 2,33 1,93 10,80 34,90 189 97
103B x 109A 64,33 65,67 1,33 1,87 9,50 35,20 200 107
184A x 177B 65,00 67,00 2,00 2,73 10,00 37,60 197 105
52B x 49A 61,00 62,33 1,33 1,47 9,50 36,20 198 104
165A x 161B 65,00 67,33 2,33 2,40 15,50 32,65 203 120
96B x 106A 60,67 61,67 1,00 2,27 15,00 33,40 182 96
73A x 41B 54,00 54,33 0,33 2,27 9,20 36,90 182 92
180B x 178A 64,33 64,67 0,33 2,27 11,40 32,20 194 104
109A x 103B 63,67 64,67 1,00 2,13 12,00 33,10 197 108
31B x 34A 62,67 64,33 1,67 2,40 12,10 36,50 191 95
167B x 163A 61,00 63,00 2,00 2,40 13,00 34,40 196 107
97B x 58A 62,67 62,67 0,00 2,47 8,30 36,00 211 116
120B x 110A 65,00 65,33 0,33 1,87 13,40 34,40 209 109
12A x 13B 63,33 65,00 1,67 2,00 12,90 32,60 188 99
1B x 5A 62,33 63,67 1,33 2,20 9,00 32,20 163 81
97A x 81B 65,67 67,67 2,00 2,33 7,50 33,50 194 104
156A x 150B 63,33 63,33 0,00 2,13 10,40 36,20 202 118
99A x 134B 62,67 66,67 4,00 1,60 9,10 32,50 187 101
116B x 55A 64,67 68,33 3,67 1,93 14,20 33,30 200 109
61B x 86A 58,67 60,33 1,67 2,33 11,20 34,90 201 100
11A x 12B 61,00 62,00 1,00 2,87 10,90 34,90 186 100
148A x 135B 61,67 62,67 1,00 1,93 14,60 33,70 199 103
119B x 108A 64,33 65,67 1,33 2,07 10,70 34,45 179 93
47B x 92A 66,00 67,33 1,33 1,73 14,00 36,70 183 103
27A x 51B 63,67 63,33 -0,33 3,07 12,20 34,90 187 110
Média prog. 62,63 63,92 1,28 2,17 11,54 34,64 192 103
Pop. Tolerante 64,67 66,67 2,00 2,07 9,60 30,60 168 89
Pop. Sensível 65,67 66,33 0,67 1,93 13,10 33,30 199 117
FT510 64,00 66,33 2,33 1,73 14,60 32,50 146 88
DAS2B707 63,67 64,33 0,67 2,53 9,20 35,20 191 102
Média test. 64,50 65,92 1,42 2,07 11,63 32,90 176 99
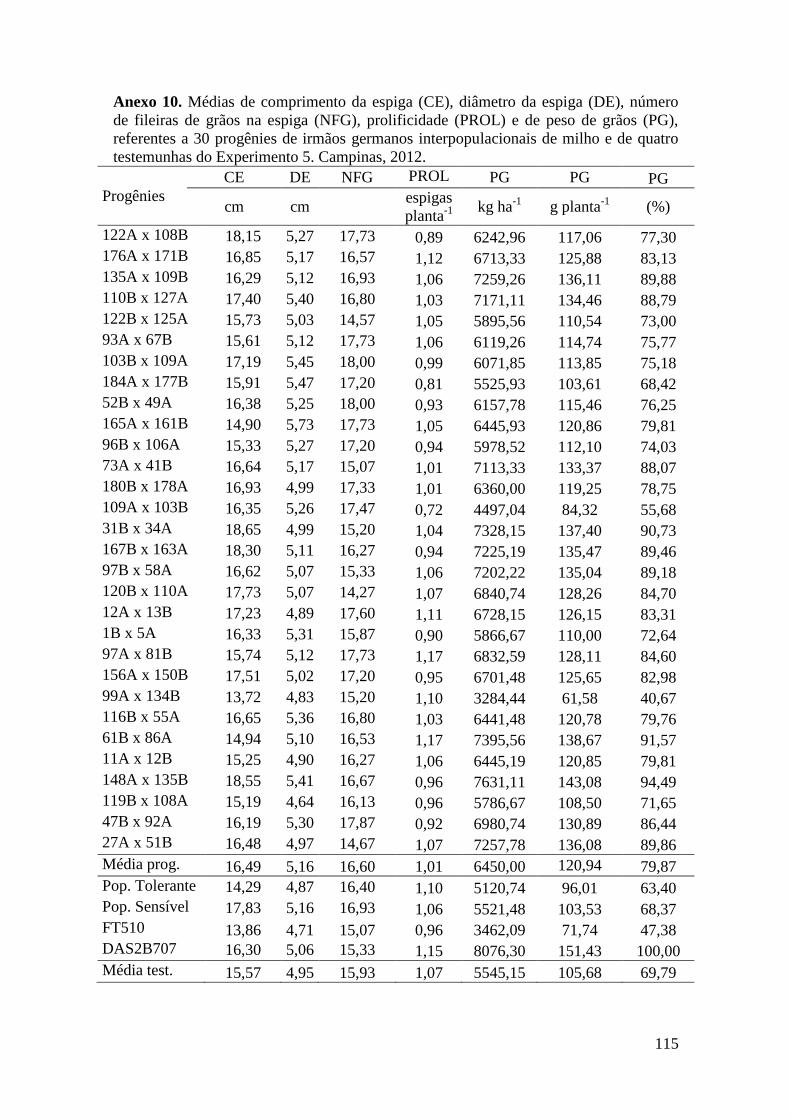
115
Anexo 10. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 5. Campinas, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
122A x 108B 18,15 5,27 17,73 0,89 6242,96 117,06 77,30
176A x 171B 16,85 5,17 16,57 1,12 6713,33 125,88 83,13
135A x 109B 16,29 5,12 16,93 1,06 7259,26 136,11 89,88
110B x 127A 17,40 5,40 16,80 1,03 7171,11 134,46 88,79
122B x 125A 15,73 5,03 14,57 1,05 5895,56 110,54 73,00
93A x 67B 15,61 5,12 17,73 1,06 6119,26 114,74 75,77
103B x 109A 17,19 5,45 18,00 0,99 6071,85 113,85 75,18
184A x 177B 15,91 5,47 17,20 0,81 5525,93 103,61 68,42
52B x 49A 16,38 5,25 18,00 0,93 6157,78 115,46 76,25
165A x 161B 14,90 5,73 17,73 1,05 6445,93 120,86 79,81
96B x 106A 15,33 5,27 17,20 0,94 5978,52 112,10 74,03
73A x 41B 16,64 5,17 15,07 1,01 7113,33 133,37 88,07
180B x 178A 16,93 4,99 17,33 1,01 6360,00 119,25 78,75
109A x 103B 16,35 5,26 17,47 0,72 4497,04 84,32 55,68
31B x 34A 18,65 4,99 15,20 1,04 7328,15 137,40 90,73
167B x 163A 18,30 5,11 16,27 0,94 7225,19 135,47 89,46
97B x 58A 16,62 5,07 15,33 1,06 7202,22 135,04 89,18
120B x 110A 17,73 5,07 14,27 1,07 6840,74 128,26 84,70
12A x 13B 17,23 4,89 17,60 1,11 6728,15 126,15 83,31
1B x 5A 16,33 5,31 15,87 0,90 5866,67 110,00 72,64
97A x 81B 15,74 5,12 17,73 1,17 6832,59 128,11 84,60
156A x 150B 17,51 5,02 17,20 0,95 6701,48 125,65 82,98
99A x 134B 13,72 4,83 15,20 1,10 3284,44 61,58 40,67
116B x 55A 16,65 5,36 16,80 1,03 6441,48 120,78 79,76
61B x 86A 14,94 5,10 16,53 1,17 7395,56 138,67 91,57
11A x 12B 15,25 4,90 16,27 1,06 6445,19 120,85 79,81
148A x 135B 18,55 5,41 16,67 0,96 7631,11 143,08 94,49
119B x 108A 15,19 4,64 16,13 0,96 5786,67 108,50 71,65
47B x 92A 16,19 5,30 17,87 0,92 6980,74 130,89 86,44
27A x 51B 16,48 4,97 14,67 1,07 7257,78 136,08 89,86
Média prog. 16,49 5,16 16,60 1,01 6450,00 120,94 79,87
Pop. Tolerante 14,29 4,87 16,40 1,10 5120,74 96,01 63,40
Pop. Sensível 17,83 5,16 16,93 1,06 5521,48 103,53 68,37
FT510 13,86 4,71 15,07 0,96 3462,09 71,74 47,38
DAS2B707 16,30 5,06 15,33 1,15 8076,30 151,43 100,00
Média test. 15,57 4,95 15,93 1,07 5545,15 105,68 69,79
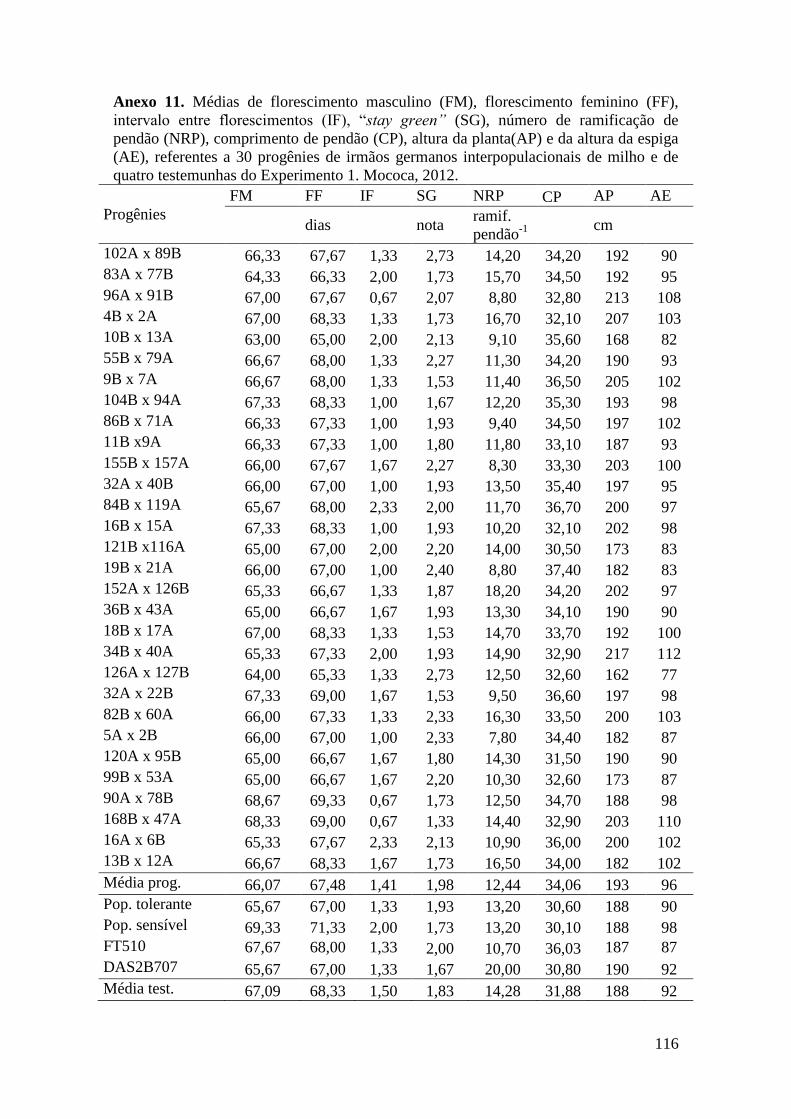
116
Anexo 11. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 1. Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
102A x 89B 66,33 67,67 1,33 2,73 14,20 34,20 192 90
83A x 77B 64,33 66,33 2,00 1,73 15,70 34,50 192 95
96A x 91B 67,00 67,67 0,67 2,07 8,80 32,80 213 108
4B x 2A 67,00 68,33 1,33 1,73 16,70 32,10 207 103
10B x 13A 63,00 65,00 2,00 2,13 9,10 35,60 168 82
55B x 79A 66,67 68,00 1,33 2,27 11,30 34,20 190 93
9B x 7A 66,67 68,00 1,33 1,53 11,40 36,50 205 102
104B x 94A 67,33 68,33 1,00 1,67 12,20 35,30 193 98
86B x 71A 66,33 67,33 1,00 1,93 9,40 34,50 197 102
11B x9A 66,33 67,33 1,00 1,80 11,80 33,10 187 93
155B x 157A 66,00 67,67 1,67 2,27 8,30 33,30 203 100
32A x 40B 66,00 67,00 1,00 1,93 13,50 35,40 197 95
84B x 119A 65,67 68,00 2,33 2,00 11,70 36,70 200 97
16B x 15A 67,33 68,33 1,00 1,93 10,20 32,10 202 98
121B x116A 65,00 67,00 2,00 2,20 14,00 30,50 173 83
19B x 21A 66,00 67,00 1,00 2,40 8,80 37,40 182 83
152A x 126B 65,33 66,67 1,33 1,87 18,20 34,20 202 97
36B x 43A 65,00 66,67 1,67 1,93 13,30 34,10 190 90
18B x 17A 67,00 68,33 1,33 1,53 14,70 33,70 192 100
34B x 40A 65,33 67,33 2,00 1,93 14,90 32,90 217 112
126A x 127B 64,00 65,33 1,33 2,73 12,50 32,60 162 77
32A x 22B 67,33 69,00 1,67 1,53 9,50 36,60 197 98
82B x 60A 66,00 67,33 1,33 2,33 16,30 33,50 200 103
5A x 2B 66,00 67,00 1,00 2,33 7,80 34,40 182 87
120A x 95B 65,00 66,67 1,67 1,80 14,30 31,50 190 90
99B x 53A 65,00 66,67 1,67 2,20 10,30 32,60 173 87
90A x 78B 68,67 69,33 0,67 1,73 12,50 34,70 188 98
168B x 47A 68,33 69,00 0,67 1,33 14,40 32,90 203 110
16A x 6B 65,33 67,67 2,33 2,13 10,90 36,00 200 102
13B x 12A 66,67 68,33 1,67 1,73 16,50 34,00 182 102
Média prog. 66,07 67,48 1,41 1,98 12,44 34,06 193 96
Pop. tolerante 65,67 67,00 1,33 1,93 13,20 30,60 188 90
Pop. sensível 69,33 71,33 2,00 1,73 13,20 30,10 188 98
FT510 67,67 68,00 1,33 2,00 10,70 36,03 187 87
DAS2B707 65,67 67,00 1,33 1,67 20,00 30,80 190 92
Média test. 67,09 68,33 1,50 1,83 14,28 31,88 188 92
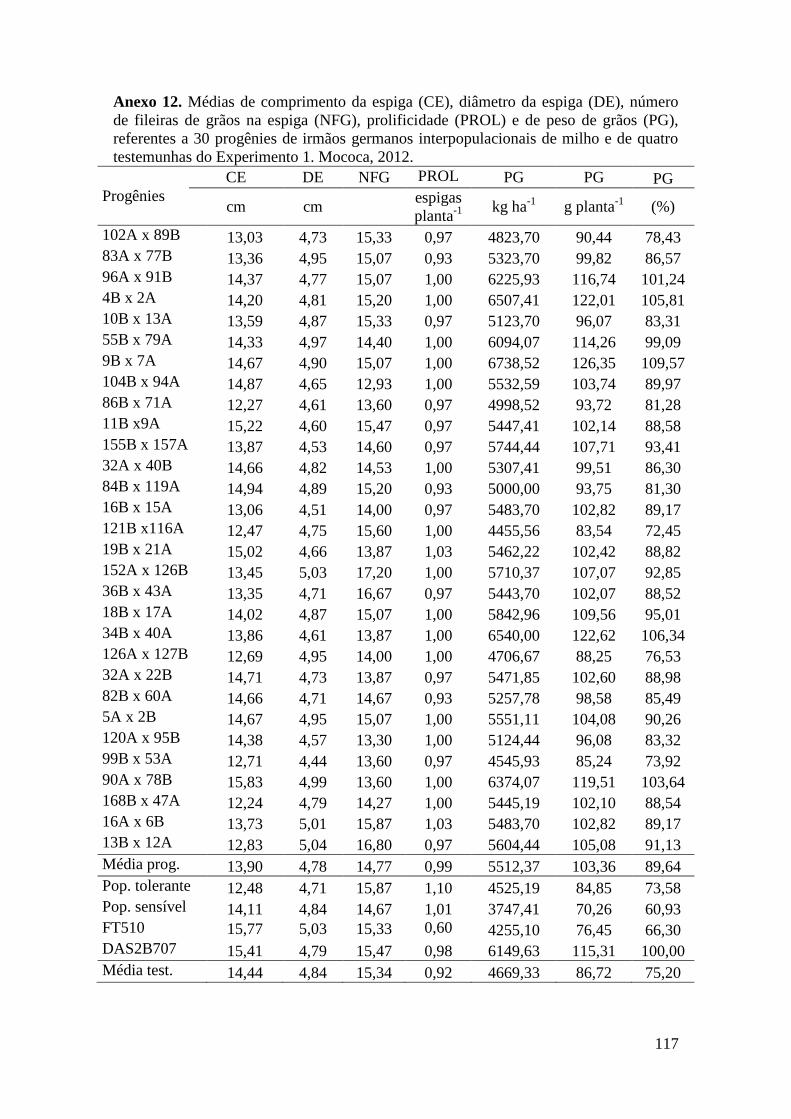
117
Anexo 12. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 1. Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
102A x 89B 13,03 4,73 15,33 0,97 4823,70 90,44 78,43
83A x 77B 13,36 4,95 15,07 0,93 5323,70 99,82 86,57
96A x 91B 14,37 4,77 15,07 1,00 6225,93 116,74 101,24
4B x 2A 14,20 4,81 15,20 1,00 6507,41 122,01 105,81
10B x 13A 13,59 4,87 15,33 0,97 5123,70 96,07 83,31
55B x 79A 14,33 4,97 14,40 1,00 6094,07 114,26 99,09
9B x 7A 14,67 4,90 15,07 1,00 6738,52 126,35 109,57
104B x 94A 14,87 4,65 12,93 1,00 5532,59 103,74 89,97
86B x 71A 12,27 4,61 13,60 0,97 4998,52 93,72 81,28
11B x9A 15,22 4,60 15,47 0,97 5447,41 102,14 88,58
155B x 157A 13,87 4,53 14,60 0,97 5744,44 107,71 93,41
32A x 40B 14,66 4,82 14,53 1,00 5307,41 99,51 86,30
84B x 119A 14,94 4,89 15,20 0,93 5000,00 93,75 81,30
16B x 15A 13,06 4,51 14,00 0,97 5483,70 102,82 89,17
121B x116A 12,47 4,75 15,60 1,00 4455,56 83,54 72,45
19B x 21A 15,02 4,66 13,87 1,03 5462,22 102,42 88,82
152A x 126B 13,45 5,03 17,20 1,00 5710,37 107,07 92,85
36B x 43A 13,35 4,71 16,67 0,97 5443,70 102,07 88,52
18B x 17A 14,02 4,87 15,07 1,00 5842,96 109,56 95,01
34B x 40A 13,86 4,61 13,87 1,00 6540,00 122,62 106,34
126A x 127B 12,69 4,95 14,00 1,00 4706,67 88,25 76,53
32A x 22B 14,71 4,73 13,87 0,97 5471,85 102,60 88,98
82B x 60A 14,66 4,71 14,67 0,93 5257,78 98,58 85,49
5A x 2B 14,67 4,95 15,07 1,00 5551,11 104,08 90,26
120A x 95B 14,38 4,57 13,30 1,00 5124,44 96,08 83,32
99B x 53A 12,71 4,44 13,60 0,97 4545,93 85,24 73,92
90A x 78B 15,83 4,99 13,60 1,00 6374,07 119,51 103,64
168B x 47A 12,24 4,79 14,27 1,00 5445,19 102,10 88,54
16A x 6B 13,73 5,01 15,87 1,03 5483,70 102,82 89,17
13B x 12A 12,83 5,04 16,80 0,97 5604,44 105,08 91,13
Média prog. 13,90 4,78 14,77 0,99 5512,37 103,36 89,64
Pop. tolerante 12,48 4,71 15,87 1,10 4525,19 84,85 73,58
Pop. sensível 14,11 4,84 14,67 1,01 3747,41 70,26 60,93
FT510 15,77 5,03 15,33 0,60 4255,10 76,45 66,30
DAS2B707 15,41 4,79 15,47 0,98 6149,63 115,31 100,00
Média test. 14,44 4,84 15,34 0,92 4669,33 86,72 75,20
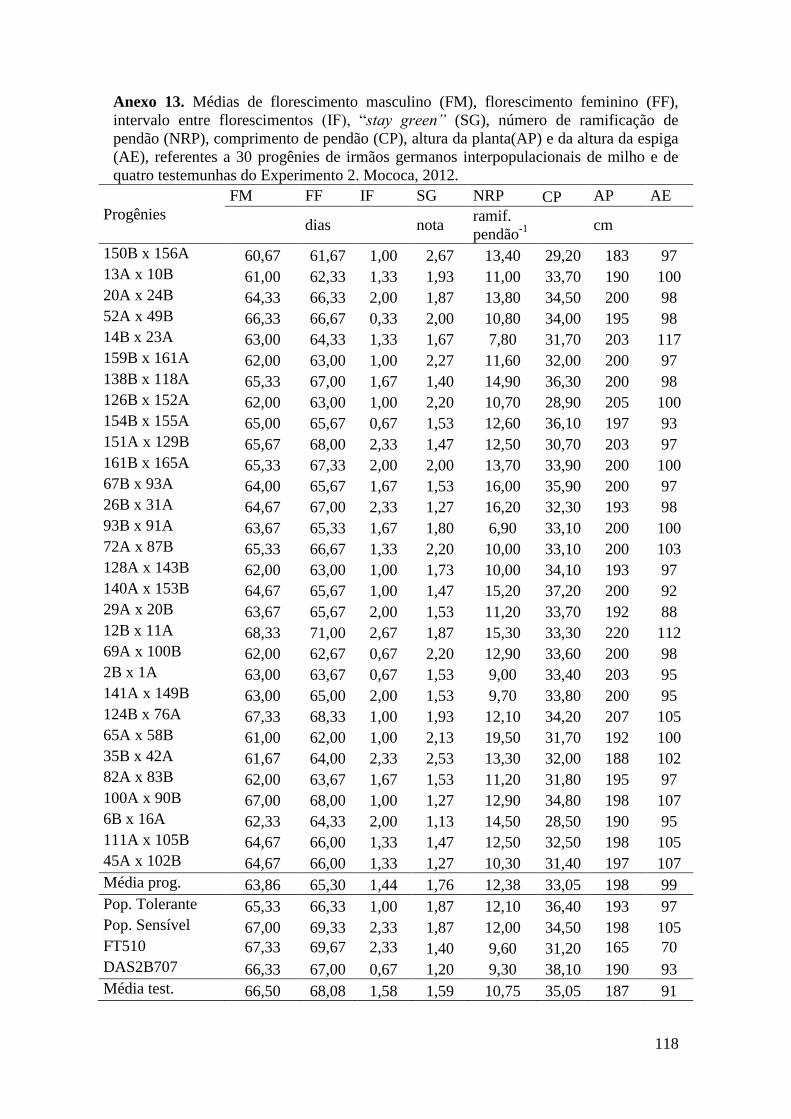
118
Anexo 13. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 2. Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
150B x 156A 60,67 61,67 1,00 2,67 13,40 29,20 183 97
13A x 10B 61,00 62,33 1,33 1,93 11,00 33,70 190 100
20A x 24B 64,33 66,33 2,00 1,87 13,80 34,50 200 98
52A x 49B 66,33 66,67 0,33 2,00 10,80 34,00 195 98
14B x 23A 63,00 64,33 1,33 1,67 7,80 31,70 203 117
159B x 161A 62,00 63,00 1,00 2,27 11,60 32,00 200 97
138B x 118A 65,33 67,00 1,67 1,40 14,90 36,30 200 98
126B x 152A 62,00 63,00 1,00 2,20 10,70 28,90 205 100
154B x 155A 65,00 65,67 0,67 1,53 12,60 36,10 197 93
151A x 129B 65,67 68,00 2,33 1,47 12,50 30,70 203 97
161B x 165A 65,33 67,33 2,00 2,00 13,70 33,90 200 100
67B x 93A 64,00 65,67 1,67 1,53 16,00 35,90 200 97
26B x 31A 64,67 67,00 2,33 1,27 16,20 32,30 193 98
93B x 91A 63,67 65,33 1,67 1,80 6,90 33,10 200 100
72A x 87B 65,33 66,67 1,33 2,20 10,00 33,10 200 103
128A x 143B 62,00 63,00 1,00 1,73 10,00 34,10 193 97
140A x 153B 64,67 65,67 1,00 1,47 15,20 37,20 200 92
29A x 20B 63,67 65,67 2,00 1,53 11,20 33,70 192 88
12B x 11A 68,33 71,00 2,67 1,87 15,30 33,30 220 112
69A x 100B 62,00 62,67 0,67 2,20 12,90 33,60 200 98
2B x 1A 63,00 63,67 0,67 1,53 9,00 33,40 203 95
141A x 149B 63,00 65,00 2,00 1,53 9,70 33,80 200 95
124B x 76A 67,33 68,33 1,00 1,93 12,10 34,20 207 105
65A x 58B 61,00 62,00 1,00 2,13 19,50 31,70 192 100
35B x 42A 61,67 64,00 2,33 2,53 13,30 32,00 188 102
82A x 83B 62,00 63,67 1,67 1,53 11,20 31,80 195 97
100A x 90B 67,00 68,00 1,00 1,27 12,90 34,80 198 107
6B x 16A 62,33 64,33 2,00 1,13 14,50 28,50 190 95
111A x 105B 64,67 66,00 1,33 1,47 12,50 32,50 198 105
45A x 102B 64,67 66,00 1,33 1,27 10,30 31,40 197 107
Média prog. 63,86 65,30 1,44 1,76 12,38 33,05 198 99
Pop. Tolerante 65,33 66,33 1,00 1,87 12,10 36,40 193 97
Pop. Sensível 67,00 69,33 2,33 1,87 12,00 34,50 198 105
FT510 67,33 69,67 2,33 1,40 9,60 31,20 165 70
DAS2B707 66,33 67,00 0,67 1,20 9,30 38,10 190 93
Média test. 66,50 68,08 1,58 1,59 10,75 35,05 187 91
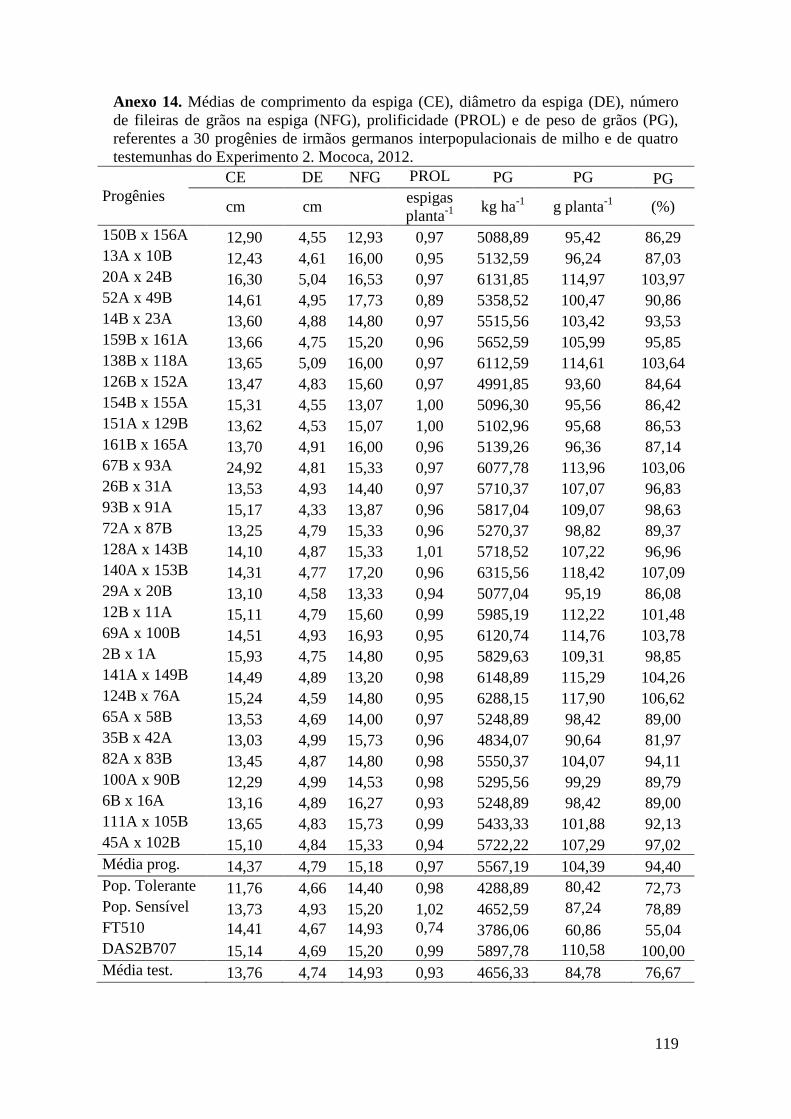
119
Anexo 14. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 2. Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
150B x 156A 12,90 4,55 12,93 0,97 5088,89 95,42 86,29
13A x 10B 12,43 4,61 16,00 0,95 5132,59 96,24 87,03
20A x 24B 16,30 5,04 16,53 0,97 6131,85 114,97 103,97
52A x 49B 14,61 4,95 17,73 0,89 5358,52 100,47 90,86
14B x 23A 13,60 4,88 14,80 0,97 5515,56 103,42 93,53
159B x 161A 13,66 4,75 15,20 0,96 5652,59 105,99 95,85
138B x 118A 13,65 5,09 16,00 0,97 6112,59 114,61 103,64
126B x 152A 13,47 4,83 15,60 0,97 4991,85 93,60 84,64
154B x 155A 15,31 4,55 13,07 1,00 5096,30 95,56 86,42
151A x 129B 13,62 4,53 15,07 1,00 5102,96 95,68 86,53
161B x 165A 13,70 4,91 16,00 0,96 5139,26 96,36 87,14
67B x 93A 24,92 4,81 15,33 0,97 6077,78 113,96 103,06
26B x 31A 13,53 4,93 14,40 0,97 5710,37 107,07 96,83
93B x 91A 15,17 4,33 13,87 0,96 5817,04 109,07 98,63
72A x 87B 13,25 4,79 15,33 0,96 5270,37 98,82 89,37
128A x 143B 14,10 4,87 15,33 1,01 5718,52 107,22 96,96
140A x 153B 14,31 4,77 17,20 0,96 6315,56 118,42 107,09
29A x 20B 13,10 4,58 13,33 0,94 5077,04 95,19 86,08
12B x 11A 15,11 4,79 15,60 0,99 5985,19 112,22 101,48
69A x 100B 14,51 4,93 16,93 0,95 6120,74 114,76 103,78
2B x 1A 15,93 4,75 14,80 0,95 5829,63 109,31 98,85
141A x 149B 14,49 4,89 13,20 0,98 6148,89 115,29 104,26
124B x 76A 15,24 4,59 14,80 0,95 6288,15 117,90 106,62
65A x 58B 13,53 4,69 14,00 0,97 5248,89 98,42 89,00
35B x 42A 13,03 4,99 15,73 0,96 4834,07 90,64 81,97
82A x 83B 13,45 4,87 14,80 0,98 5550,37 104,07 94,11
100A x 90B 12,29 4,99 14,53 0,98 5295,56 99,29 89,79
6B x 16A 13,16 4,89 16,27 0,93 5248,89 98,42 89,00
111A x 105B 13,65 4,83 15,73 0,99 5433,33 101,88 92,13
45A x 102B 15,10 4,84 15,33 0,94 5722,22 107,29 97,02
Média prog. 14,37 4,79 15,18 0,97 5567,19 104,39 94,40
Pop. Tolerante 11,76 4,66 14,40 0,98 4288,89 80,42 72,73
Pop. Sensível 13,73 4,93 15,20 1,02 4652,59 87,24 78,89
FT510 14,41 4,67 14,93 0,74 3786,06 60,86 55,04
DAS2B707 15,14 4,69 15,20 0,99 5897,78 110,58 100,00
Média test. 13,76 4,74 14,93 0,93 4656,33 84,78 76,67
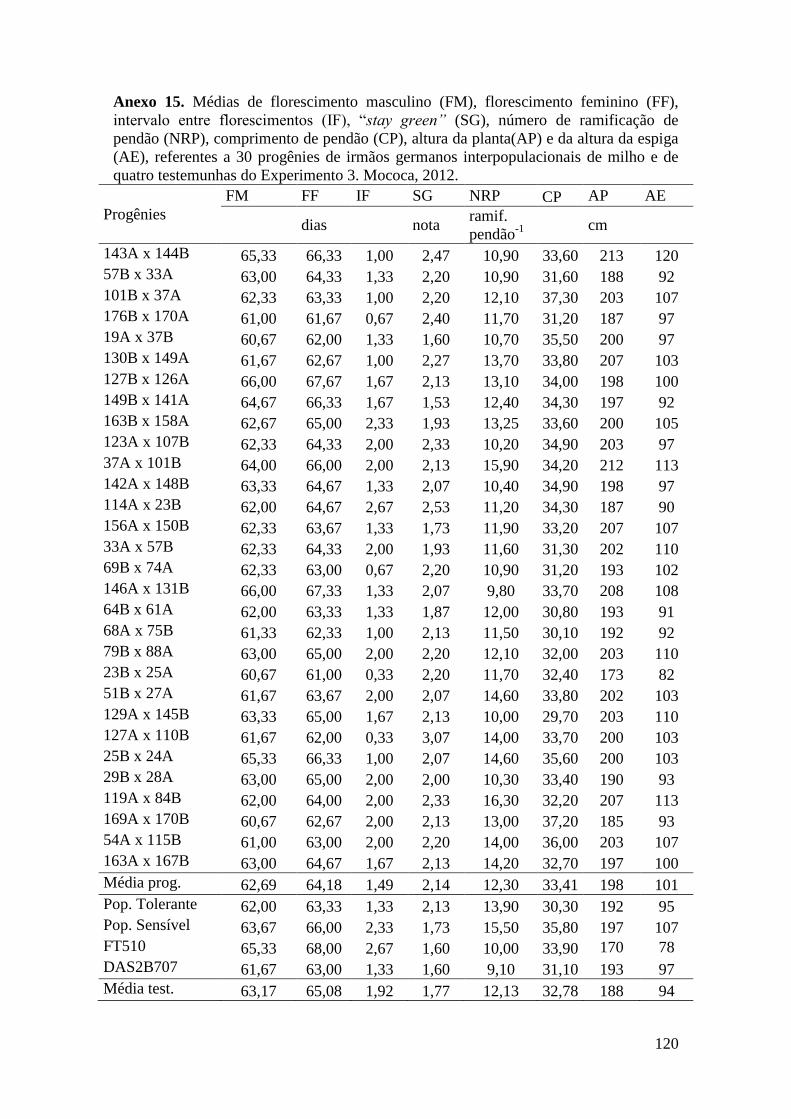
120
Anexo 15. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 3. Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
143A x 144B 65,33 66,33 1,00 2,47 10,90 33,60 213 120
57B x 33A 63,00 64,33 1,33 2,20 10,90 31,60 188 92
101B x 37A 62,33 63,33 1,00 2,20 12,10 37,30 203 107
176B x 170A 61,00 61,67 0,67 2,40 11,70 31,20 187 97
19A x 37B 60,67 62,00 1,33 1,60 10,70 35,50 200 97
130B x 149A 61,67 62,67 1,00 2,27 13,70 33,80 207 103
127B x 126A 66,00 67,67 1,67 2,13 13,10 34,00 198 100
149B x 141A 64,67 66,33 1,67 1,53 12,40 34,30 197 92
163B x 158A 62,67 65,00 2,33 1,93 13,25 33,60 200 105
123A x 107B 62,33 64,33 2,00 2,33 10,20 34,90 203 97
37A x 101B 64,00 66,00 2,00 2,13 15,90 34,20 212 113
142A x 148B 63,33 64,67 1,33 2,07 10,40 34,90 198 97
114A x 23B 62,00 64,67 2,67 2,53 11,20 34,30 187 90
156A x 150B 62,33 63,67 1,33 1,73 11,90 33,20 207 107
33A x 57B 62,33 64,33 2,00 1,93 11,60 31,30 202 110
69B x 74A 62,33 63,00 0,67 2,20 10,90 31,20 193 102
146A x 131B 66,00 67,33 1,33 2,07 9,80 33,70 208 108
64B x 61A 62,00 63,33 1,33 1,87 12,00 30,80 193 91
68A x 75B 61,33 62,33 1,00 2,13 11,50 30,10 192 92
79B x 88A 63,00 65,00 2,00 2,20 12,10 32,00 203 110
23B x 25A 60,67 61,00 0,33 2,20 11,70 32,40 173 82
51B x 27A 61,67 63,67 2,00 2,07 14,60 33,80 202 103
129A x 145B 63,33 65,00 1,67 2,13 10,00 29,70 203 110
127A x 110B 61,67 62,00 0,33 3,07 14,00 33,70 200 103
25B x 24A 65,33 66,33 1,00 2,07 14,60 35,60 200 103
29B x 28A 63,00 65,00 2,00 2,00 10,30 33,40 190 93
119A x 84B 62,00 64,00 2,00 2,33 16,30 32,20 207 113
169A x 170B 60,67 62,67 2,00 2,13 13,00 37,20 185 93
54A x 115B 61,00 63,00 2,00 2,20 14,00 36,00 203 107
163A x 167B 63,00 64,67 1,67 2,13 14,20 32,70 197 100
Média prog. 62,69 64,18 1,49 2,14 12,30 33,41 198 101
Pop. Tolerante 62,00 63,33 1,33 2,13 13,90 30,30 192 95
Pop. Sensível 63,67 66,00 2,33 1,73 15,50 35,80 197 107
FT510 65,33 68,00 2,67 1,60 10,00 33,90 170 78
DAS2B707 61,67 63,00 1,33 1,60 9,10 31,10 193 97
Média test. 63,17 65,08 1,92 1,77 12,13 32,78 188 94
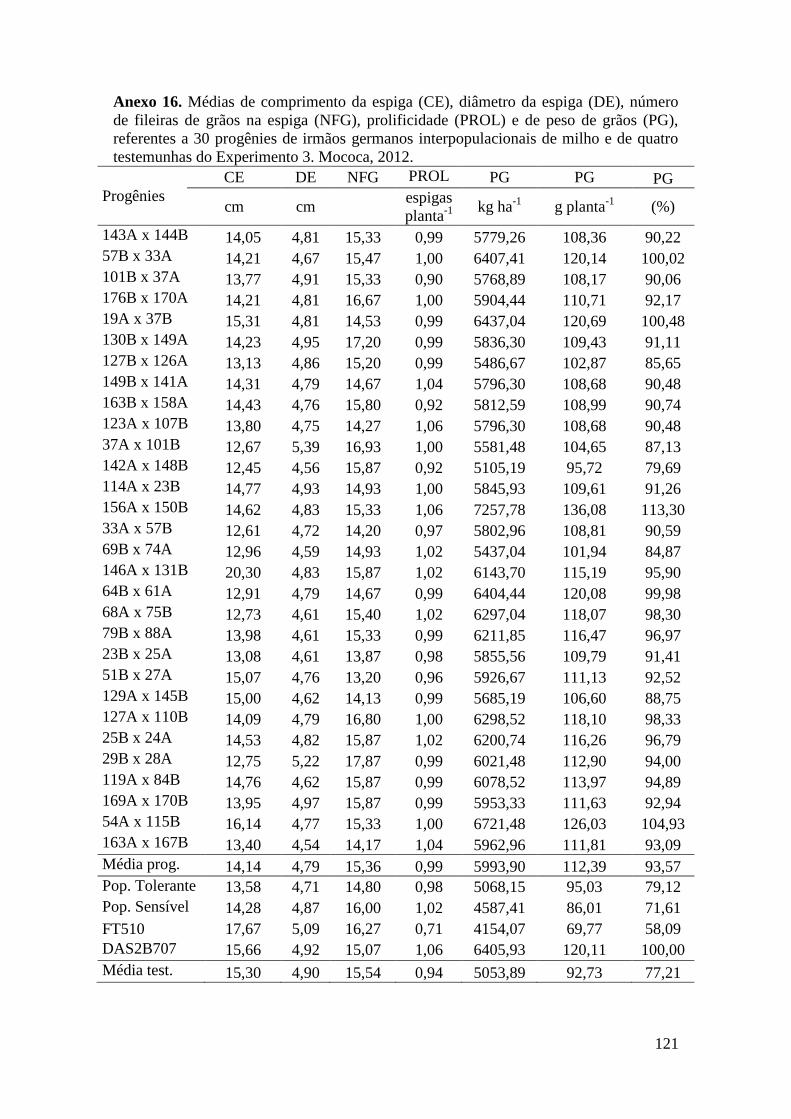
121
Anexo 16. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 3. Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
143A x 144B 14,05 4,81 15,33 0,99 5779,26 108,36 90,22
57B x 33A 14,21 4,67 15,47 1,00 6407,41 120,14 100,02
101B x 37A 13,77 4,91 15,33 0,90 5768,89 108,17 90,06
176B x 170A 14,21 4,81 16,67 1,00 5904,44 110,71 92,17
19A x 37B 15,31 4,81 14,53 0,99 6437,04 120,69 100,48
130B x 149A 14,23 4,95 17,20 0,99 5836,30 109,43 91,11
127B x 126A 13,13 4,86 15,20 0,99 5486,67 102,87 85,65
149B x 141A 14,31 4,79 14,67 1,04 5796,30 108,68 90,48
163B x 158A 14,43 4,76 15,80 0,92 5812,59 108,99 90,74
123A x 107B 13,80 4,75 14,27 1,06 5796,30 108,68 90,48
37A x 101B 12,67 5,39 16,93 1,00 5581,48 104,65 87,13
142A x 148B 12,45 4,56 15,87 0,92 5105,19 95,72 79,69
114A x 23B 14,77 4,93 14,93 1,00 5845,93 109,61 91,26
156A x 150B 14,62 4,83 15,33 1,06 7257,78 136,08 113,30
33A x 57B 12,61 4,72 14,20 0,97 5802,96 108,81 90,59
69B x 74A 12,96 4,59 14,93 1,02 5437,04 101,94 84,87
146A x 131B 20,30 4,83 15,87 1,02 6143,70 115,19 95,90
64B x 61A 12,91 4,79 14,67 0,99 6404,44 120,08 99,98
68A x 75B 12,73 4,61 15,40 1,02 6297,04 118,07 98,30
79B x 88A 13,98 4,61 15,33 0,99 6211,85 116,47 96,97
23B x 25A 13,08 4,61 13,87 0,98 5855,56 109,79 91,41
51B x 27A 15,07 4,76 13,20 0,96 5926,67 111,13 92,52
129A x 145B 15,00 4,62 14,13 0,99 5685,19 106,60 88,75
127A x 110B 14,09 4,79 16,80 1,00 6298,52 118,10 98,33
25B x 24A 14,53 4,82 15,87 1,02 6200,74 116,26 96,79
29B x 28A 12,75 5,22 17,87 0,99 6021,48 112,90 94,00
119A x 84B 14,76 4,62 15,87 0,99 6078,52 113,97 94,89
169A x 170B 13,95 4,97 15,87 0,99 5953,33 111,63 92,94
54A x 115B 16,14 4,77 15,33 1,00 6721,48 126,03 104,93
163A x 167B 13,40 4,54 14,17 1,04 5962,96 111,81 93,09
Média prog. 14,14 4,79 15,36 0,99 5993,90 112,39 93,57
Pop. Tolerante 13,58 4,71 14,80 0,98 5068,15 95,03 79,12
Pop. Sensível 14,28 4,87 16,00 1,02 4587,41 86,01 71,61
FT510 17,67 5,09 16,27 0,71 4154,07 69,77 58,09
DAS2B707 15,66 4,92 15,07 1,06 6405,93 120,11 100,00
Média test. 15,30 4,90 15,54 0,94 5053,89 92,73 77,21
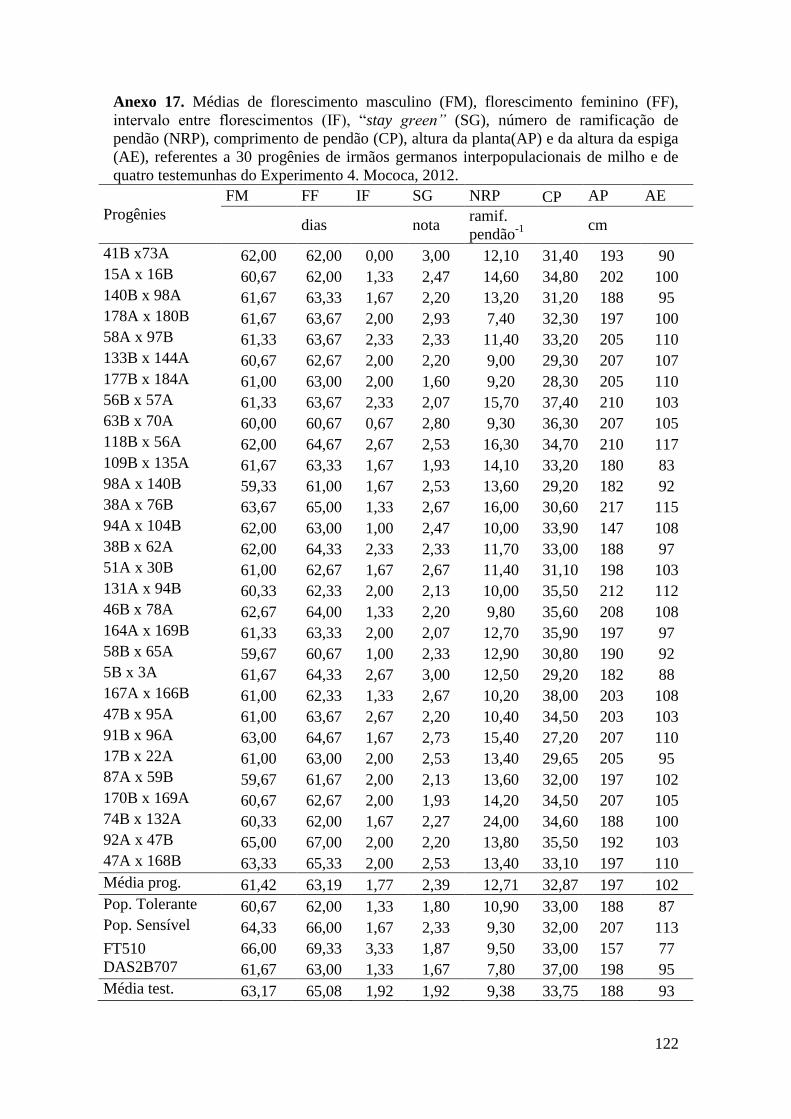
122
Anexo 17. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 4. Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
41B x73A 62,00 62,00 0,00 3,00 12,10 31,40 193 90
15A x 16B 60,67 62,00 1,33 2,47 14,60 34,80 202 100
140B x 98A 61,67 63,33 1,67 2,20 13,20 31,20 188 95
178A x 180B 61,67 63,67 2,00 2,93 7,40 32,30 197 100
58A x 97B 61,33 63,67 2,33 2,33 11,40 33,20 205 110
133B x 144A 60,67 62,67 2,00 2,20 9,00 29,30 207 107
177B x 184A 61,00 63,00 2,00 1,60 9,20 28,30 205 110
56B x 57A 61,33 63,67 2,33 2,07 15,70 37,40 210 103
63B x 70A 60,00 60,67 0,67 2,80 9,30 36,30 207 105
118B x 56A 62,00 64,67 2,67 2,53 16,30 34,70 210 117
109B x 135A 61,67 63,33 1,67 1,93 14,10 33,20 180 83
98A x 140B 59,33 61,00 1,67 2,53 13,60 29,20 182 92
38A x 76B 63,67 65,00 1,33 2,67 16,00 30,60 217 115
94A x 104B 62,00 63,00 1,00 2,47 10,00 33,90 147 108
38B x 62A 62,00 64,33 2,33 2,33 11,70 33,00 188 97
51A x 30B 61,00 62,67 1,67 2,67 11,40 31,10 198 103
131A x 94B 60,33 62,33 2,00 2,13 10,00 35,50 212 112
46B x 78A 62,67 64,00 1,33 2,20 9,80 35,60 208 108
164A x 169B 61,33 63,33 2,00 2,07 12,70 35,90 197 97
58B x 65A 59,67 60,67 1,00 2,33 12,90 30,80 190 92
5B x 3A 61,67 64,33 2,67 3,00 12,50 29,20 182 88
167A x 166B 61,00 62,33 1,33 2,67 10,20 38,00 203 108
47B x 95A 61,00 63,67 2,67 2,20 10,40 34,50 203 103
91B x 96A 63,00 64,67 1,67 2,73 15,40 27,20 207 110
17B x 22A 61,00 63,00 2,00 2,53 13,40 29,65 205 95
87A x 59B 59,67 61,67 2,00 2,13 13,60 32,00 197 102
170B x 169A 60,67 62,67 2,00 1,93 14,20 34,50 207 105
74B x 132A 60,33 62,00 1,67 2,27 24,00 34,60 188 100
92A x 47B 65,00 67,00 2,00 2,20 13,80 35,50 192 103
47A x 168B 63,33 65,33 2,00 2,53 13,40 33,10 197 110
Média prog. 61,42 63,19 1,77 2,39 12,71 32,87 197 102
Pop. Tolerante 60,67 62,00 1,33 1,80 10,90 33,00 188 87
Pop. Sensível 64,33 66,00 1,67 2,33 9,30 32,00 207 113
FT510 66,00 69,33 3,33 1,87 9,50 33,00 157 77
DAS2B707 61,67 63,00 1,33 1,67 7,80 37,00 198 95
Média test. 63,17 65,08 1,92 1,92 9,38 33,75 188 93
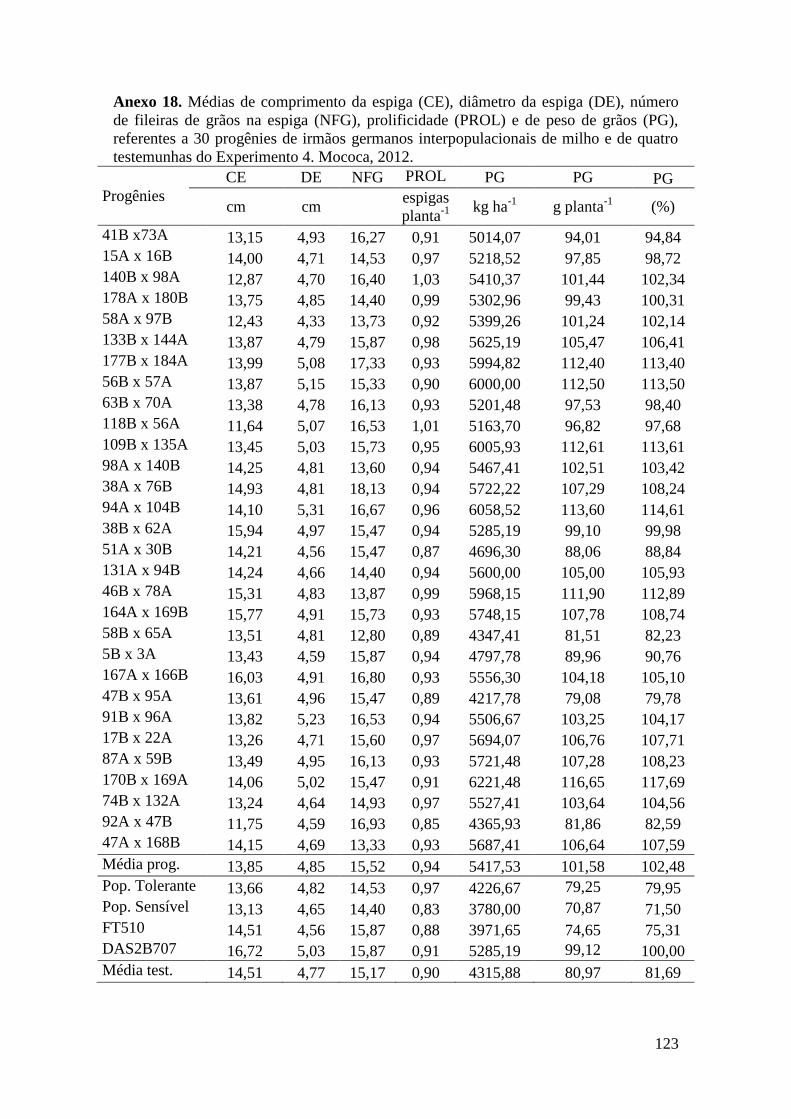
123
Anexo 18. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 4. Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
41B x73A 13,15 4,93 16,27 0,91 5014,07 94,01 94,84
15A x 16B 14,00 4,71 14,53 0,97 5218,52 97,85 98,72
140B x 98A 12,87 4,70 16,40 1,03 5410,37 101,44 102,34
178A x 180B 13,75 4,85 14,40 0,99 5302,96 99,43 100,31
58A x 97B 12,43 4,33 13,73 0,92 5399,26 101,24 102,14
133B x 144A 13,87 4,79 15,87 0,98 5625,19 105,47 106,41
177B x 184A 13,99 5,08 17,33 0,93 5994,82 112,40 113,40
56B x 57A 13,87 5,15 15,33 0,90 6000,00 112,50 113,50
63B x 70A 13,38 4,78 16,13 0,93 5201,48 97,53 98,40
118B x 56A 11,64 5,07 16,53 1,01 5163,70 96,82 97,68
109B x 135A 13,45 5,03 15,73 0,95 6005,93 112,61 113,61
98A x 140B 14,25 4,81 13,60 0,94 5467,41 102,51 103,42
38A x 76B 14,93 4,81 18,13 0,94 5722,22 107,29 108,24
94A x 104B 14,10 5,31 16,67 0,96 6058,52 113,60 114,61
38B x 62A 15,94 4,97 15,47 0,94 5285,19 99,10 99,98
51A x 30B 14,21 4,56 15,47 0,87 4696,30 88,06 88,84
131A x 94B 14,24 4,66 14,40 0,94 5600,00 105,00 105,93
46B x 78A 15,31 4,83 13,87 0,99 5968,15 111,90 112,89
164A x 169B 15,77 4,91 15,73 0,93 5748,15 107,78 108,74
58B x 65A 13,51 4,81 12,80 0,89 4347,41 81,51 82,23
5B x 3A 13,43 4,59 15,87 0,94 4797,78 89,96 90,76
167A x 166B 16,03 4,91 16,80 0,93 5556,30 104,18 105,10
47B x 95A 13,61 4,96 15,47 0,89 4217,78 79,08 79,78
91B x 96A 13,82 5,23 16,53 0,94 5506,67 103,25 104,17
17B x 22A 13,26 4,71 15,60 0,97 5694,07 106,76 107,71
87A x 59B 13,49 4,95 16,13 0,93 5721,48 107,28 108,23
170B x 169A 14,06 5,02 15,47 0,91 6221,48 116,65 117,69
74B x 132A 13,24 4,64 14,93 0,97 5527,41 103,64 104,56
92A x 47B 11,75 4,59 16,93 0,85 4365,93 81,86 82,59
47A x 168B 14,15 4,69 13,33 0,93 5687,41 106,64 107,59
Média prog. 13,85 4,85 15,52 0,94 5417,53 101,58 102,48
Pop. Tolerante 13,66 4,82 14,53 0,97 4226,67 79,25 79,95
Pop. Sensível 13,13 4,65 14,40 0,83 3780,00 70,87 71,50
FT510 14,51 4,56 15,87 0,88 3971,65 74,65 75,31
DAS2B707 16,72 5,03 15,87 0,91 5285,19 99,12 100,00
Média test. 14,51 4,77 15,17 0,90 4315,88 80,97 81,69
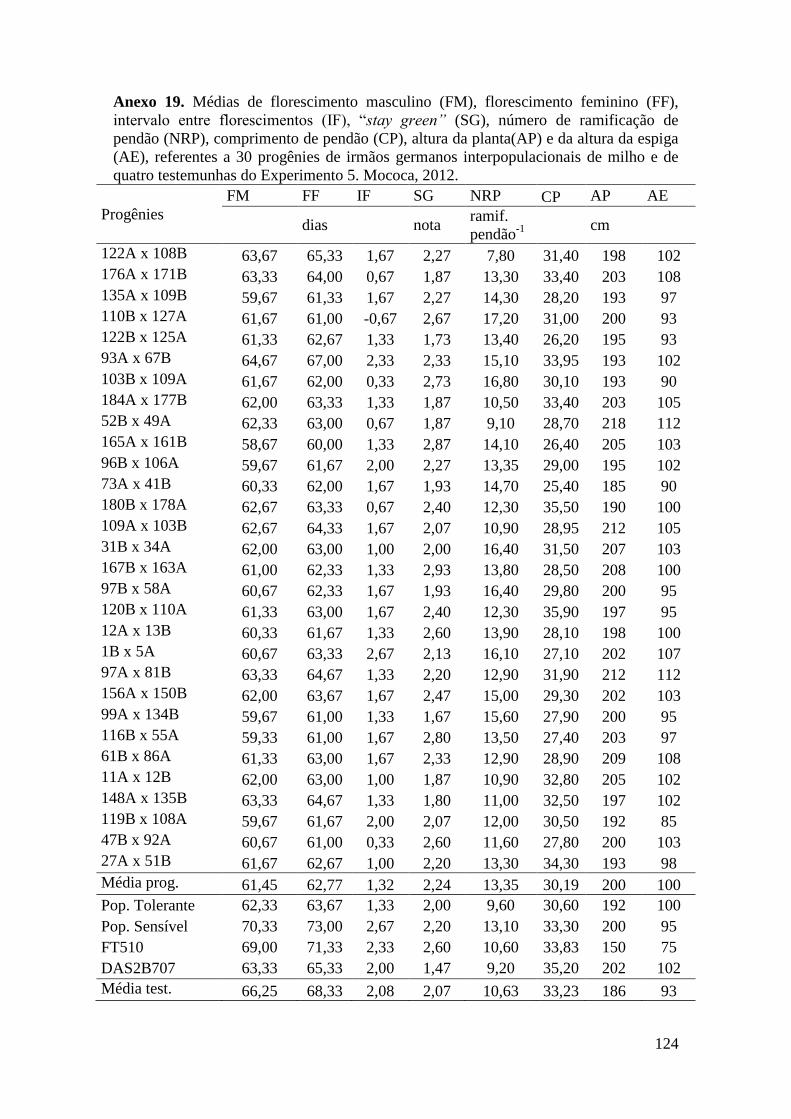
124
Anexo 19. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 5. Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
122A x 108B 63,67 65,33 1,67 2,27 7,80 31,40 198 102
176A x 171B 63,33 64,00 0,67 1,87 13,30 33,40 203 108
135A x 109B 59,67 61,33 1,67 2,27 14,30 28,20 193 97
110B x 127A 61,67 61,00 -0,67 2,67 17,20 31,00 200 93
122B x 125A 61,33 62,67 1,33 1,73 13,40 26,20 195 93
93A x 67B 64,67 67,00 2,33 2,33 15,10 33,95 193 102
103B x 109A 61,67 62,00 0,33 2,73 16,80 30,10 193 90
184A x 177B 62,00 63,33 1,33 1,87 10,50 33,40 203 105
52B x 49A 62,33 63,00 0,67 1,87 9,10 28,70 218 112
165A x 161B 58,67 60,00 1,33 2,87 14,10 26,40 205 103
96B x 106A 59,67 61,67 2,00 2,27 13,35 29,00 195 102
73A x 41B 60,33 62,00 1,67 1,93 14,70 25,40 185 90
180B x 178A 62,67 63,33 0,67 2,40 12,30 35,50 190 100
109A x 103B 62,67 64,33 1,67 2,07 10,90 28,95 212 105
31B x 34A 62,00 63,00 1,00 2,00 16,40 31,50 207 103
167B x 163A 61,00 62,33 1,33 2,93 13,80 28,50 208 100
97B x 58A 60,67 62,33 1,67 1,93 16,40 29,80 200 95
120B x 110A 61,33 63,00 1,67 2,40 12,30 35,90 197 95
12A x 13B 60,33 61,67 1,33 2,60 13,90 28,10 198 100
1B x 5A 60,67 63,33 2,67 2,13 16,10 27,10 202 107
97A x 81B 63,33 64,67 1,33 2,20 12,90 31,90 212 112
156A x 150B 62,00 63,67 1,67 2,47 15,00 29,30 202 103
99A x 134B 59,67 61,00 1,33 1,67 15,60 27,90 200 95
116B x 55A 59,33 61,00 1,67 2,80 13,50 27,40 203 97
61B x 86A 61,33 63,00 1,67 2,33 12,90 28,90 209 108
11A x 12B 62,00 63,00 1,00 1,87 10,90 32,80 205 102
148A x 135B 63,33 64,67 1,33 1,80 11,00 32,50 197 102
119B x 108A 59,67 61,67 2,00 2,07 12,00 30,50 192 85
47B x 92A 60,67 61,00 0,33 2,60 11,60 27,80 200 103
27A x 51B 61,67 62,67 1,00 2,20 13,30 34,30 193 98
Média prog. 61,45 62,77 1,32 2,24 13,35 30,19 200 100
Pop. Tolerante 62,33 63,67 1,33 2,00 9,60 30,60 192 100
Pop. Sensível 70,33 73,00 2,67 2,20 13,10 33,30 200 95
FT510 69,00 71,33 2,33 2,60 10,60 33,83 150 75
DAS2B707 63,33 65,33 2,00 1,47 9,20 35,20 202 102
Média test. 66,25 68,33 2,08 2,07 10,63 33,23 186 93
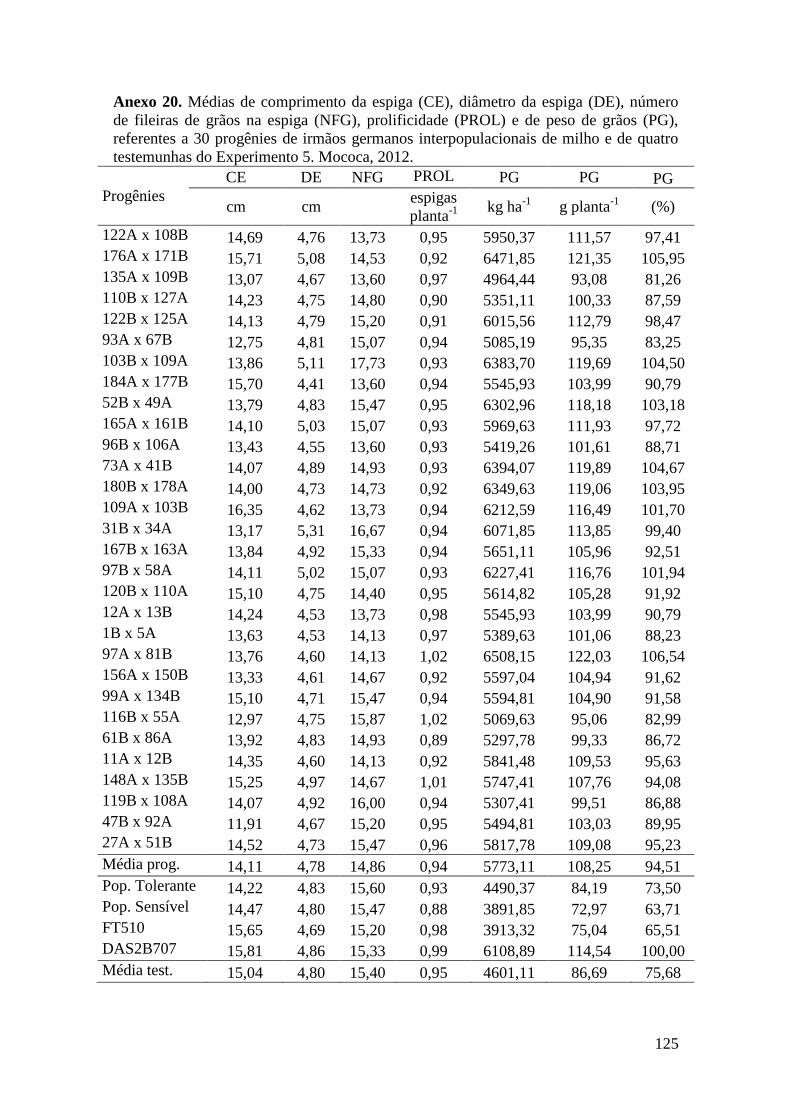
125
Anexo 20. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 5. Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
122A x 108B 14,69 4,76 13,73 0,95 5950,37 111,57 97,41
176A x 171B 15,71 5,08 14,53 0,92 6471,85 121,35 105,95
135A x 109B 13,07 4,67 13,60 0,97 4964,44 93,08 81,26
110B x 127A 14,23 4,75 14,80 0,90 5351,11 100,33 87,59
122B x 125A 14,13 4,79 15,20 0,91 6015,56 112,79 98,47
93A x 67B 12,75 4,81 15,07 0,94 5085,19 95,35 83,25
103B x 109A 13,86 5,11 17,73 0,93 6383,70 119,69 104,50
184A x 177B 15,70 4,41 13,60 0,94 5545,93 103,99 90,79
52B x 49A 13,79 4,83 15,47 0,95 6302,96 118,18 103,18
165A x 161B 14,10 5,03 15,07 0,93 5969,63 111,93 97,72
96B x 106A 13,43 4,55 13,60 0,93 5419,26 101,61 88,71
73A x 41B 14,07 4,89 14,93 0,93 6394,07 119,89 104,67
180B x 178A 14,00 4,73 14,73 0,92 6349,63 119,06 103,95
109A x 103B 16,35 4,62 13,73 0,94 6212,59 116,49 101,70
31B x 34A 13,17 5,31 16,67 0,94 6071,85 113,85 99,40
167B x 163A 13,84 4,92 15,33 0,94 5651,11 105,96 92,51
97B x 58A 14,11 5,02 15,07 0,93 6227,41 116,76 101,94
120B x 110A 15,10 4,75 14,40 0,95 5614,82 105,28 91,92
12A x 13B 14,24 4,53 13,73 0,98 5545,93 103,99 90,79
1B x 5A 13,63 4,53 14,13 0,97 5389,63 101,06 88,23
97A x 81B 13,76 4,60 14,13 1,02 6508,15 122,03 106,54
156A x 150B 13,33 4,61 14,67 0,92 5597,04 104,94 91,62
99A x 134B 15,10 4,71 15,47 0,94 5594,81 104,90 91,58
116B x 55A 12,97 4,75 15,87 1,02 5069,63 95,06 82,99
61B x 86A 13,92 4,83 14,93 0,89 5297,78 99,33 86,72
11A x 12B 14,35 4,60 14,13 0,92 5841,48 109,53 95,63
148A x 135B 15,25 4,97 14,67 1,01 5747,41 107,76 94,08
119B x 108A 14,07 4,92 16,00 0,94 5307,41 99,51 86,88
47B x 92A 11,91 4,67 15,20 0,95 5494,81 103,03 89,95
27A x 51B 14,52 4,73 15,47 0,96 5817,78 109,08 95,23
Média prog. 14,11 4,78 14,86 0,94 5773,11 108,25 94,51
Pop. Tolerante 14,22 4,83 15,60 0,93 4490,37 84,19 73,50
Pop. Sensível 14,47 4,80 15,47 0,88 3891,85 72,97 63,71
FT510 15,65 4,69 15,20 0,98 3913,32 75,04 65,51
DAS2B707 15,81 4,86 15,33 0,99 6108,89 114,54 100,00
Média test. 15,04 4,80 15,40 0,95 4601,11 86,69 75,68
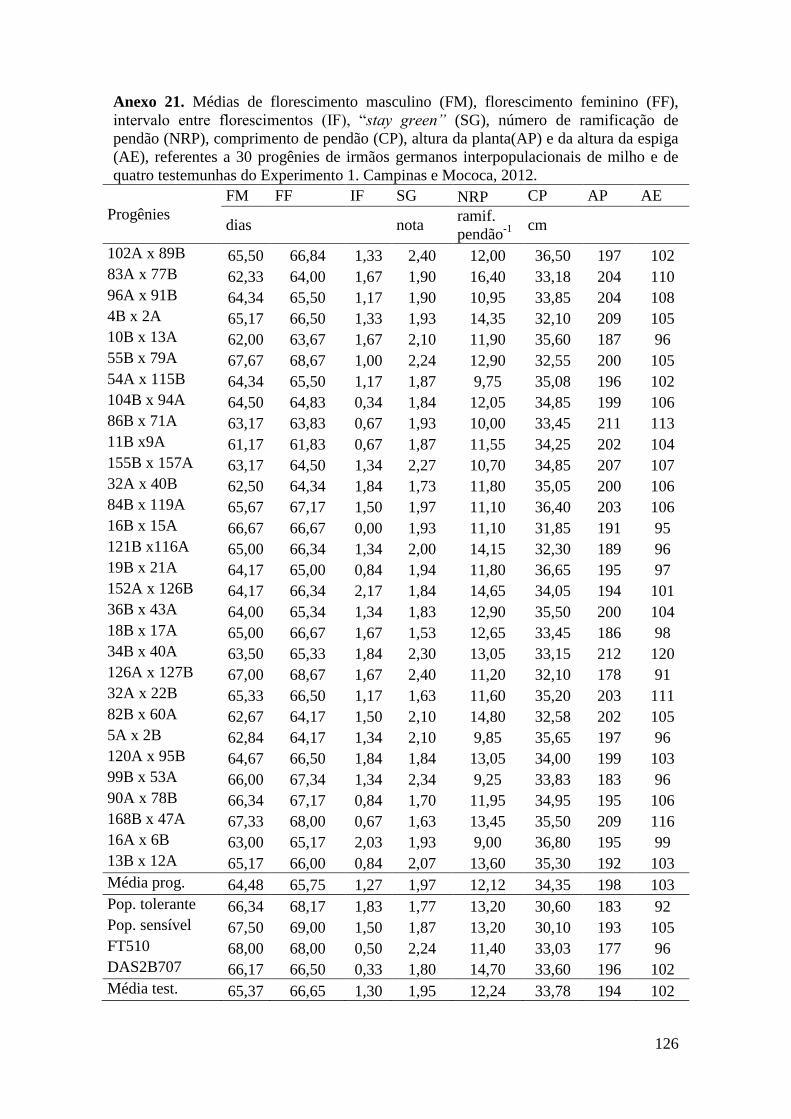
126
Anexo 21. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 1. Campinas e Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
102A x 89B 65,50 66,84 1,33 2,40 12,00 36,50 197 102
83A x 77B 62,33 64,00 1,67 1,90 16,40 33,18 204 110
96A x 91B 64,34 65,50 1,17 1,90 10,95 33,85 204 108
4B x 2A 65,17 66,50 1,33 1,93 14,35 32,10 209 105
10B x 13A 62,00 63,67 1,67 2,10 11,90 35,60 187 96
55B x 79A 67,67 68,67 1,00 2,24 12,90 32,55 200 105
54A x 115B 64,34 65,50 1,17 1,87 9,75 35,08 196 102
104B x 94A 64,50 64,83 0,34 1,84 12,05 34,85 199 106
86B x 71A 63,17 63,83 0,67 1,93 10,00 33,45 211 113
11B x9A 61,17 61,83 0,67 1,87 11,55 34,25 202 104
155B x 157A 63,17 64,50 1,34 2,27 10,70 34,85 207 107
32A x 40B 62,50 64,34 1,84 1,73 11,80 35,05 200 106
84B x 119A 65,67 67,17 1,50 1,97 11,10 36,40 203 106
16B x 15A 66,67 66,67 0,00 1,93 11,10 31,85 191 95
121B x116A 65,00 66,34 1,34 2,00 14,15 32,30 189 96
19B x 21A 64,17 65,00 0,84 1,94 11,80 36,65 195 97
152A x 126B 64,17 66,34 2,17 1,84 14,65 34,05 194 101
36B x 43A 64,00 65,34 1,34 1,83 12,90 35,50 200 104
18B x 17A 65,00 66,67 1,67 1,53 12,65 33,45 186 98
34B x 40A 63,50 65,33 1,84 2,30 13,05 33,15 212 120
126A x 127B 67,00 68,67 1,67 2,40 11,20 32,10 178 91
32A x 22B 65,33 66,50 1,17 1,63 11,60 35,20 203 111
82B x 60A 62,67 64,17 1,50 2,10 14,80 32,58 202 105
5A x 2B 62,84 64,17 1,34 2,10 9,85 35,65 197 96
120A x 95B 64,67 66,50 1,84 1,84 13,05 34,00 199 103
99B x 53A 66,00 67,34 1,34 2,34 9,25 33,83 183 96
90A x 78B 66,34 67,17 0,84 1,70 11,95 34,95 195 106
168B x 47A 67,33 68,00 0,67 1,63 13,45 35,50 209 116
16A x 6B 63,00 65,17 2,03 1,93 9,00 36,80 195 99
13B x 12A 65,17 66,00 0,84 2,07 13,60 35,30 192 103
Média prog. 64,48 65,75 1,27 1,97 12,12 34,35 198 103
Pop. tolerante 66,34 68,17 1,83 1,77 13,20 30,60 183 92
Pop. sensível 67,50 69,00 1,50 1,87 13,20 30,10 193 105
FT510 68,00 68,00 0,50 2,24 11,40 33,03 177 96
DAS2B707 66,17 66,50 0,33 1,80 14,70 33,60 196 102
Média test. 65,37 66,65 1,30 1,95 12,24 33,78 194 102

127
Anexo 22. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 1. Campinas e Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
102A x 89B 14,85 4,92 16,05 0,93 5543,33 103,94 97,14
83A x 77B 14,73 4,86 15,21 1,03 6652,22 124,73 116,57
96A x 91B 15,64 4,76 14,26 1,03 5062,23 94,92 88,71
4B x 2A 15,54 4,80 15,22 1,02 6343,71 118,95 111,17
10B x 13A 14,91 4,81 15,67 1,01 5641,48 105,78 98,86
55B x 79A 14,33 5,10 14,27 1,03 5713,33 107,13 100,12
54A x 115B 15,01 4,73 16,21 1,00 5888,89 110,42 103,20
104B x 94A 16,01 4,64 13,38 0,91 5715,93 107,18 100,17
86B x 71A 14,87 4,84 14,84 0,95 6046,30 113,37 105,95
11B x9A 16,04 5,25 15,18 1,01 4531,11 84,96 79,40
155B x 157A 16,02 4,87 14,05 0,96 5430,00 101,82 95,16
32A x 40B 15,53 5,30 14,66 1,01 5714,08 107,14 100,13
84B x 119A 16,21 5,19 15,08 0,97 5281,11 99,02 92,54
16B x 15A 14,88 4,75 14,94 0,85 5552,96 104,12 97,31
121B x116A 14,19 5,40 15,33 1,02 5700,75 106,89 99,90
19B x 21A 15,60 4,89 15,85 0,99 5266,30 98,75 92,29
152A x 126B 14,64 5,03 16,44 1,03 5452,59 102,24 95,55
36B x 43A 16,24 5,36 16,45 1,02 6032,96 113,12 105,72
18B x 17A 14,76 4,89 14,89 1,06 5300,74 99,39 92,89
34B x 40A 14,52 4,31 14,49 0,95 6266,67 117,50 109,81
126A x 127B 14,24 4,99 15,02 1,09 5392,60 101,11 94,50
32A x 22B 15,30 5,03 14,87 0,96 5091,11 95,46 89,21
82B x 60A 14,94 4,83 15,45 1,02 5253,71 98,51 92,07
5A x 2B 15,88 5,25 14,38 1,02 6101,48 114,40 106,92
120A x 95B 15,99 4,77 14,65 1,09 6312,59 118,36 110,62
99B x 53A 13,85 4,92 14,34 1,02 5027,04 94,26 88,09
90A x 78B 15,71 5,04 14,80 0,98 6162,22 115,54 107,98
168B x 47A 14,66 4,77 14,79 0,98 6072,60 113,87 106,42
16A x 6B 15,71 5,30 16,77 0,98 5735,93 107,55 100,51
13B x 12A 14,98 5,27 16,54 1,02 5845,56 109,61 102,44
Média prog. 15,19 4,96 15,13 1,00 5671,05 106,33 99,37
Pop. tolerante 14,58 4,83 15,80 1,06 5551,86 104,10 97,29
Pop. sensível 15,06 5,29 16,21 0,93 4403,71 82,57 77,17
FT510 15,59 4,92 15,33 0,77 4310,31 83,63 78,16
DAS2B707 15,82 5,15 15,95 1,02 5706,67 107,00 100,00
Média test. 15,10 4,97 15,26 0,99 5541,52 104,07 88,15
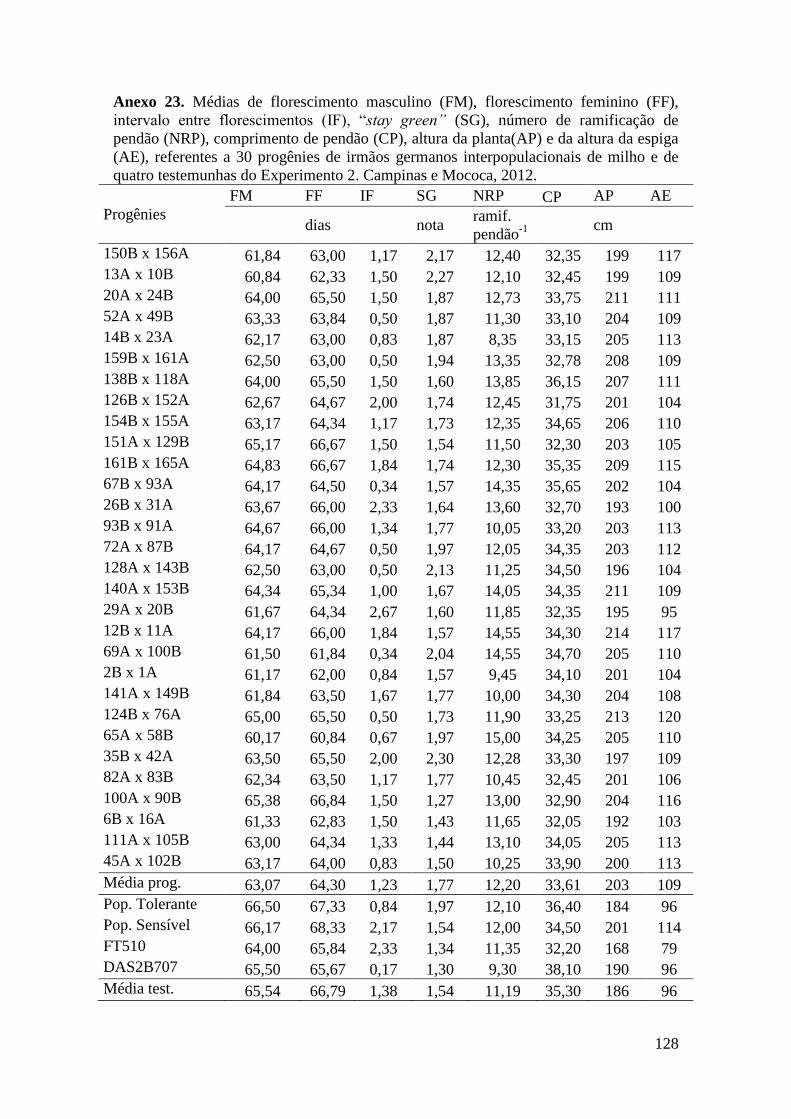
128
Anexo 23. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 2. Campinas e Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
150B x 156A 61,84 63,00 1,17 2,17 12,40 32,35 199 117
13A x 10B 60,84 62,33 1,50 2,27 12,10 32,45 199 109
20A x 24B 64,00 65,50 1,50 1,87 12,73 33,75 211 111
52A x 49B 63,33 63,84 0,50 1,87 11,30 33,10 204 109
14B x 23A 62,17 63,00 0,83 1,87 8,35 33,15 205 113
159B x 161A 62,50 63,00 0,50 1,94 13,35 32,78 208 109
138B x 118A 64,00 65,50 1,50 1,60 13,85 36,15 207 111
126B x 152A 62,67 64,67 2,00 1,74 12,45 31,75 201 104
154B x 155A 63,17 64,34 1,17 1,73 12,35 34,65 206 110
151A x 129B 65,17 66,67 1,50 1,54 11,50 32,30 203 105
161B x 165A 64,83 66,67 1,84 1,74 12,30 35,35 209 115
67B x 93A 64,17 64,50 0,34 1,57 14,35 35,65 202 104
26B x 31A 63,67 66,00 2,33 1,64 13,60 32,70 193 100
93B x 91A 64,67 66,00 1,34 1,77 10,05 33,20 203 113
72A x 87B 64,17 64,67 0,50 1,97 12,05 34,35 203 112
128A x 143B 62,50 63,00 0,50 2,13 11,25 34,50 196 104
140A x 153B 64,34 65,34 1,00 1,67 14,05 34,35 211 109
29A x 20B 61,67 64,34 2,67 1,60 11,85 32,35 195 95
12B x 11A 64,17 66,00 1,84 1,57 14,55 34,30 214 117
69A x 100B 61,50 61,84 0,34 2,04 14,55 34,70 205 110
2B x 1A 61,17 62,00 0,84 1,57 9,45 34,10 201 104
141A x 149B 61,84 63,50 1,67 1,77 10,00 34,30 204 108
124B x 76A 65,00 65,50 0,50 1,73 11,90 33,25 213 120
65A x 58B 60,17 60,84 0,67 1,97 15,00 34,25 205 110
35B x 42A 63,50 65,50 2,00 2,30 12,28 33,30 197 109
82A x 83B 62,34 63,50 1,17 1,77 10,45 32,45 201 106
100A x 90B 65,38 66,84 1,50 1,27 13,00 32,90 204 116
6B x 16A 61,33 62,83 1,50 1,43 11,65 32,05 192 103
111A x 105B 63,00 64,34 1,33 1,44 13,10 34,05 205 113
45A x 102B 63,17 64,00 0,83 1,50 10,25 33,90 200 113
Média prog. 63,07 64,30 1,23 1,77 12,20 33,61 203 109
Pop. Tolerante 66,50 67,33 0,84 1,97 12,10 36,40 184 96
Pop. Sensível 66,17 68,33 2,17 1,54 12,00 34,50 201 114
FT510 64,00 65,84 2,33 1,34 11,35 32,20 168 79
DAS2B707 65,50 65,67 0,17 1,30 9,30 38,10 190 96
Média test. 65,54 66,79 1,38 1,54 11,19 35,30 186 96
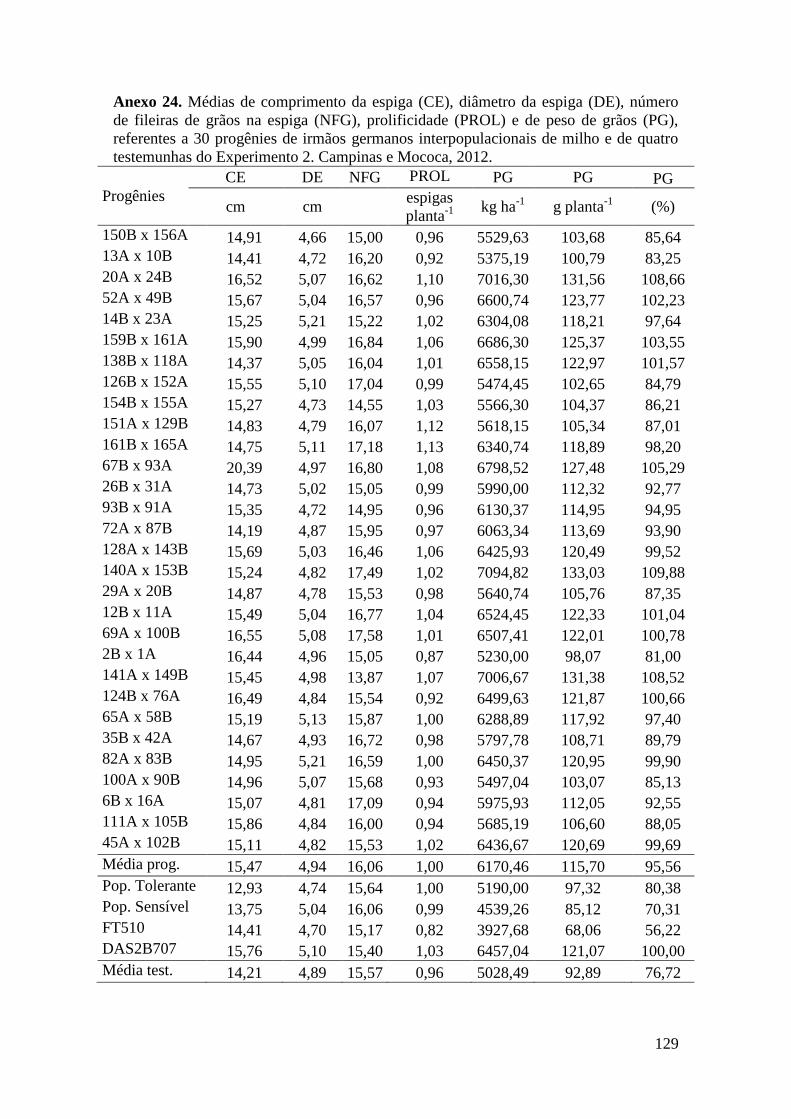
129
Anexo 24. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 2. Campinas e Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
150B x 156A 14,91 4,66 15,00 0,96 5529,63 103,68 85,64
13A x 10B 14,41 4,72 16,20 0,92 5375,19 100,79 83,25
20A x 24B 16,52 5,07 16,62 1,10 7016,30 131,56 108,66
52A x 49B 15,67 5,04 16,57 0,96 6600,74 123,77 102,23
14B x 23A 15,25 5,21 15,22 1,02 6304,08 118,21 97,64
159B x 161A 15,90 4,99 16,84 1,06 6686,30 125,37 103,55
138B x 118A 14,37 5,05 16,04 1,01 6558,15 122,97 101,57
126B x 152A 15,55 5,10 17,04 0,99 5474,45 102,65 84,79
154B x 155A 15,27 4,73 14,55 1,03 5566,30 104,37 86,21
151A x 129B 14,83 4,79 16,07 1,12 5618,15 105,34 87,01
161B x 165A 14,75 5,11 17,18 1,13 6340,74 118,89 98,20
67B x 93A 20,39 4,97 16,80 1,08 6798,52 127,48 105,29
26B x 31A 14,73 5,02 15,05 0,99 5990,00 112,32 92,77
93B x 91A 15,35 4,72 14,95 0,96 6130,37 114,95 94,95
72A x 87B 14,19 4,87 15,95 0,97 6063,34 113,69 93,90
128A x 143B 15,69 5,03 16,46 1,06 6425,93 120,49 99,52
140A x 153B 15,24 4,82 17,49 1,02 7094,82 133,03 109,88
29A x 20B 14,87 4,78 15,53 0,98 5640,74 105,76 87,35
12B x 11A 15,49 5,04 16,77 1,04 6524,45 122,33 101,04
69A x 100B 16,55 5,08 17,58 1,01 6507,41 122,01 100,78
2B x 1A 16,44 4,96 15,05 0,87 5230,00 98,07 81,00
141A x 149B 15,45 4,98 13,87 1,07 7006,67 131,38 108,52
124B x 76A 16,49 4,84 15,54 0,92 6499,63 121,87 100,66
65A x 58B 15,19 5,13 15,87 1,00 6288,89 117,92 97,40
35B x 42A 14,67 4,93 16,72 0,98 5797,78 108,71 89,79
82A x 83B 14,95 5,21 16,59 1,00 6450,37 120,95 99,90
100A x 90B 14,96 5,07 15,68 0,93 5497,04 103,07 85,13
6B x 16A 15,07 4,81 17,09 0,94 5975,93 112,05 92,55
111A x 105B 15,86 4,84 16,00 0,94 5685,19 106,60 88,05
45A x 102B 15,11 4,82 15,53 1,02 6436,67 120,69 99,69
Média prog. 15,47 4,94 16,06 1,00 6170,46 115,70 95,56
Pop. Tolerante 12,93 4,74 15,64 1,00 5190,00 97,32 80,38
Pop. Sensível 13,75 5,04 16,06 0,99 4539,26 85,12 70,31
FT510 14,41 4,70 15,17 0,82 3927,68 68,06 56,22
DAS2B707 15,76 5,10 15,40 1,03 6457,04 121,07 100,00
Média test. 14,21 4,89 15,57 0,96 5028,49 92,89 76,72
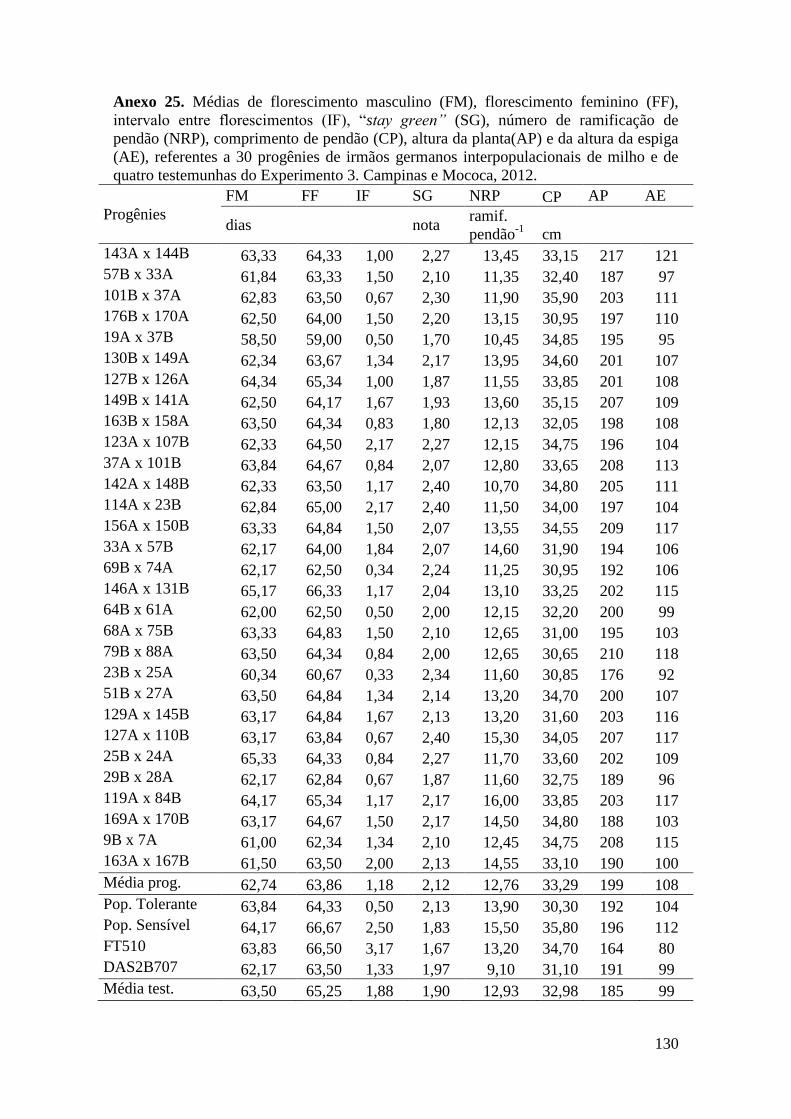
130
Anexo 25. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 3. Campinas e Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
143A x 144B 63,33 64,33 1,00 2,27 13,45 33,15 217 121
57B x 33A 61,84 63,33 1,50 2,10 11,35 32,40 187 97
101B x 37A 62,83 63,50 0,67 2,30 11,90 35,90 203 111
176B x 170A 62,50 64,00 1,50 2,20 13,15 30,95 197 110
19A x 37B 58,50 59,00 0,50 1,70 10,45 34,85 195 95
130B x 149A 62,34 63,67 1,34 2,17 13,95 34,60 201 107
127B x 126A 64,34 65,34 1,00 1,87 11,55 33,85 201 108
149B x 141A 62,50 64,17 1,67 1,93 13,60 35,15 207 109
163B x 158A 63,50 64,34 0,83 1,80 12,13 32,05 198 108
123A x 107B 62,33 64,50 2,17 2,27 12,15 34,75 196 104
37A x 101B 63,84 64,67 0,84 2,07 12,80 33,65 208 113
142A x 148B 62,33 63,50 1,17 2,40 10,70 34,80 205 111
114A x 23B 62,84 65,00 2,17 2,40 11,50 34,00 197 104
156A x 150B 63,33 64,84 1,50 2,07 13,55 34,55 209 117
33A x 57B 62,17 64,00 1,84 2,07 14,60 31,90 194 106
69B x 74A 62,17 62,50 0,34 2,24 11,25 30,95 192 106
146A x 131B 65,17 66,33 1,17 2,04 13,10 33,25 202 115
64B x 61A 62,00 62,50 0,50 2,00 12,15 32,20 200 99
68A x 75B 63,33 64,83 1,50 2,10 12,65 31,00 195 103
79B x 88A 63,50 64,34 0,84 2,00 12,65 30,65 210 118
23B x 25A 60,34 60,67 0,33 2,34 11,60 30,85 176 92
51B x 27A 63,50 64,84 1,34 2,14 13,20 34,70 200 107
129A x 145B 63,17 64,84 1,67 2,13 13,20 31,60 203 116
127A x 110B 63,17 63,84 0,67 2,40 15,30 34,05 207 117
25B x 24A 65,33 64,33 0,84 2,27 11,70 33,60 202 109
29B x 28A 62,17 62,84 0,67 1,87 11,60 32,75 189 96
119A x 84B 64,17 65,34 1,17 2,17 16,00 33,85 203 117
169A x 170B 63,17 64,67 1,50 2,17 14,50 34,80 188 103
9B x 7A 61,00 62,34 1,34 2,10 12,45 34,75 208 115
163A x 167B 61,50 63,50 2,00 2,13 14,55 33,10 190 100
Média prog. 62,74 63,86 1,18 2,12 12,76 33,29 199 108
Pop. Tolerante 63,84 64,33 0,50 2,13 13,90 30,30 192 104
Pop. Sensível 64,17 66,67 2,50 1,83 15,50 35,80 196 112
FT510 63,83 66,50 3,17 1,67 13,20 34,70 164 80
DAS2B707 62,17 63,50 1,33 1,97 9,10 31,10 191 99
Média test. 63,50 65,25 1,88 1,90 12,93 32,98 185 99
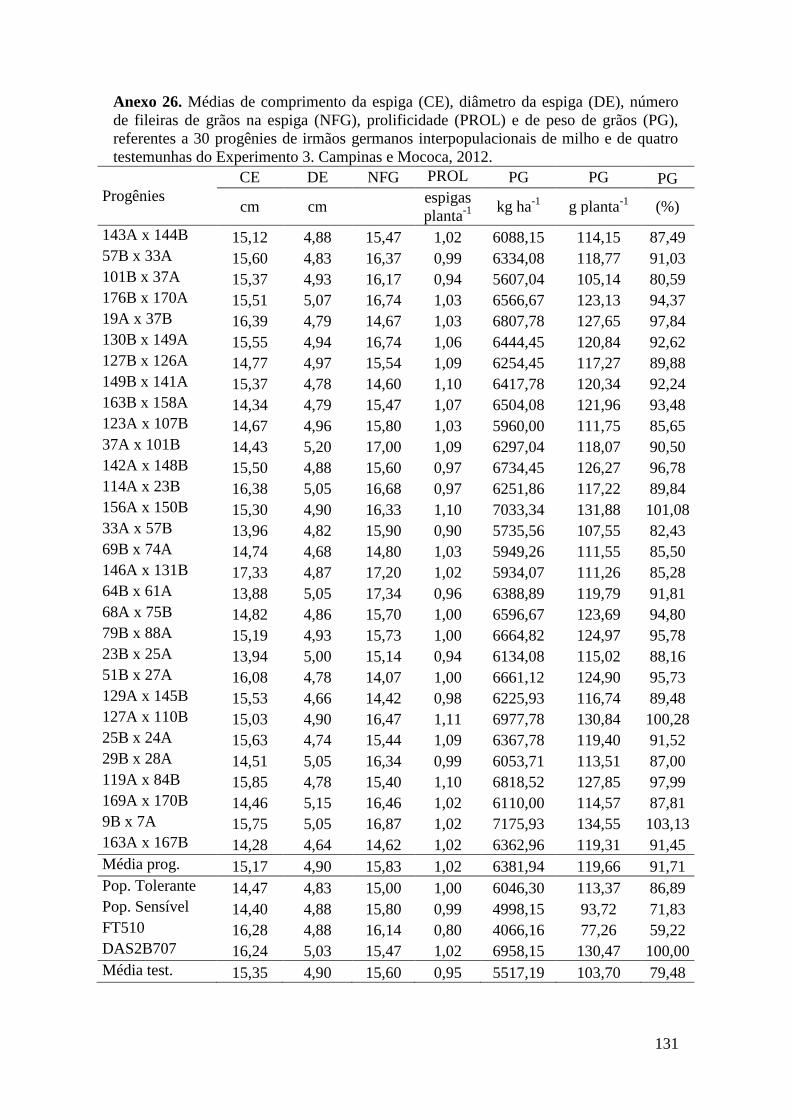
131
Anexo 26. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 3. Campinas e Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
143A x 144B 15,12 4,88 15,47 1,02 6088,15 114,15 87,49
57B x 33A 15,60 4,83 16,37 0,99 6334,08 118,77 91,03
101B x 37A 15,37 4,93 16,17 0,94 5607,04 105,14 80,59
176B x 170A 15,51 5,07 16,74 1,03 6566,67 123,13 94,37
19A x 37B 16,39 4,79 14,67 1,03 6807,78 127,65 97,84
130B x 149A 15,55 4,94 16,74 1,06 6444,45 120,84 92,62
127B x 126A 14,77 4,97 15,54 1,09 6254,45 117,27 89,88
149B x 141A 15,37 4,78 14,60 1,10 6417,78 120,34 92,24
163B x 158A 14,34 4,79 15,47 1,07 6504,08 121,96 93,48
123A x 107B 14,67 4,96 15,80 1,03 5960,00 111,75 85,65
37A x 101B 14,43 5,20 17,00 1,09 6297,04 118,07 90,50
142A x 148B 15,50 4,88 15,60 0,97 6734,45 126,27 96,78
114A x 23B 16,38 5,05 16,68 0,97 6251,86 117,22 89,84
156A x 150B 15,30 4,90 16,33 1,10 7033,34 131,88 101,08
33A x 57B 13,96 4,82 15,90 0,90 5735,56 107,55 82,43
69B x 74A 14,74 4,68 14,80 1,03 5949,26 111,55 85,50
146A x 131B 17,33 4,87 17,20 1,02 5934,07 111,26 85,28
64B x 61A 13,88 5,05 17,34 0,96 6388,89 119,79 91,81
68A x 75B 14,82 4,86 15,70 1,00 6596,67 123,69 94,80
79B x 88A 15,19 4,93 15,73 1,00 6664,82 124,97 95,78
23B x 25A 13,94 5,00 15,14 0,94 6134,08 115,02 88,16
51B x 27A 16,08 4,78 14,07 1,00 6661,12 124,90 95,73
129A x 145B 15,53 4,66 14,42 0,98 6225,93 116,74 89,48
127A x 110B 15,03 4,90 16,47 1,11 6977,78 130,84 100,28
25B x 24A 15,63 4,74 15,44 1,09 6367,78 119,40 91,52
29B x 28A 14,51 5,05 16,34 0,99 6053,71 113,51 87,00
119A x 84B 15,85 4,78 15,40 1,10 6818,52 127,85 97,99
169A x 170B 14,46 5,15 16,46 1,02 6110,00 114,57 87,81
9B x 7A 15,75 5,05 16,87 1,02 7175,93 134,55 103,13
163A x 167B 14,28 4,64 14,62 1,02 6362,96 119,31 91,45
Média prog. 15,17 4,90 15,83 1,02 6381,94 119,66 91,71
Pop. Tolerante 14,47 4,83 15,00 1,00 6046,30 113,37 86,89
Pop. Sensível 14,40 4,88 15,80 0,99 4998,15 93,72 71,83
FT510 16,28 4,88 16,14 0,80 4066,16 77,26 59,22
DAS2B707 16,24 5,03 15,47 1,02 6958,15 130,47 100,00
Média test. 15,35 4,90 15,60 0,95 5517,19 103,70 79,48
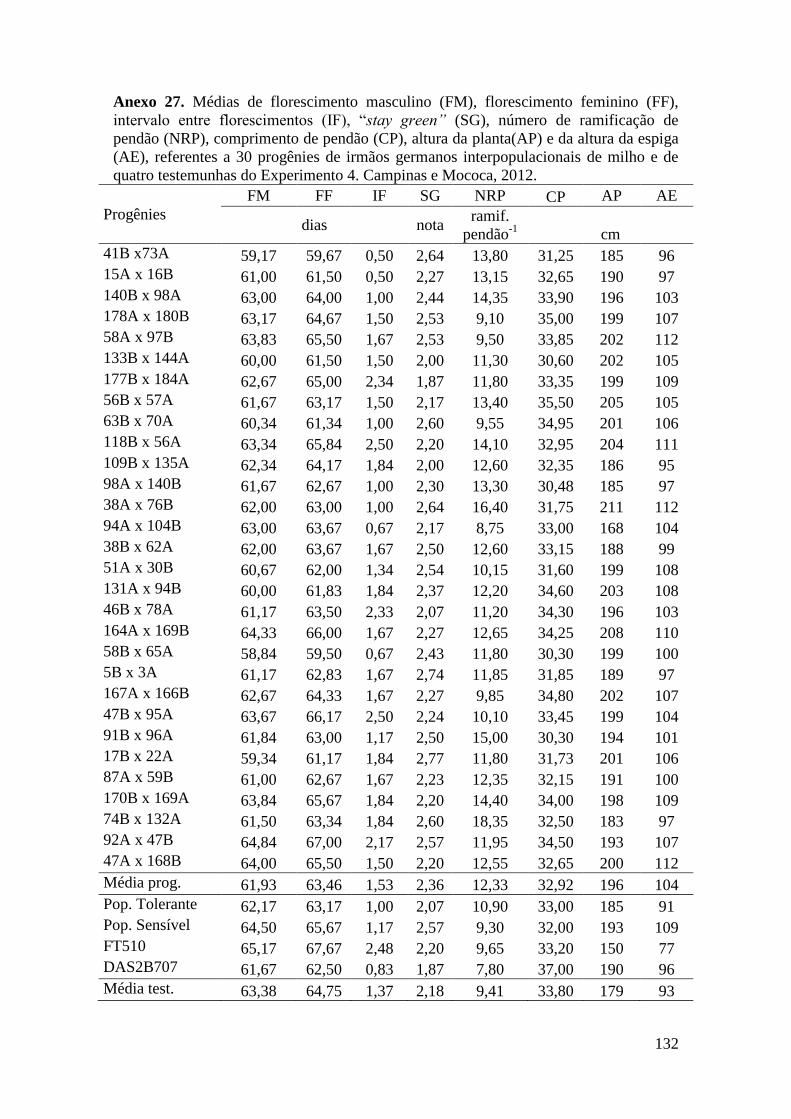
132
Anexo 27. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 4. Campinas e Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
41B x73A 59,17 59,67 0,50 2,64 13,80 31,25 185 96
15A x 16B 61,00 61,50 0,50 2,27 13,15 32,65 190 97
140B x 98A 63,00 64,00 1,00 2,44 14,35 33,90 196 103
178A x 180B 63,17 64,67 1,50 2,53 9,10 35,00 199 107
58A x 97B 63,83 65,50 1,67 2,53 9,50 33,85 202 112
133B x 144A 60,00 61,50 1,50 2,00 11,30 30,60 202 105
177B x 184A 62,67 65,00 2,34 1,87 11,80 33,35 199 109
56B x 57A 61,67 63,17 1,50 2,17 13,40 35,50 205 105
63B x 70A 60,34 61,34 1,00 2,60 9,55 34,95 201 106
118B x 56A 63,34 65,84 2,50 2,20 14,10 32,95 204 111
109B x 135A 62,34 64,17 1,84 2,00 12,60 32,35 186 95
98A x 140B 61,67 62,67 1,00 2,30 13,30 30,48 185 97
38A x 76B 62,00 63,00 1,00 2,64 16,40 31,75 211 112
94A x 104B 63,00 63,67 0,67 2,17 8,75 33,00 168 104
38B x 62A 62,00 63,67 1,67 2,50 12,60 33,15 188 99
51A x 30B 60,67 62,00 1,34 2,54 10,15 31,60 199 108
131A x 94B 60,00 61,83 1,84 2,37 12,20 34,60 203 108
46B x 78A 61,17 63,50 2,33 2,07 11,20 34,30 196 103
164A x 169B 64,33 66,00 1,67 2,27 12,65 34,25 208 110
58B x 65A 58,84 59,50 0,67 2,43 11,80 30,30 199 100
5B x 3A 61,17 62,83 1,67 2,74 11,85 31,85 189 97
167A x 166B 62,67 64,33 1,67 2,27 9,85 34,80 202 107
47B x 95A 63,67 66,17 2,50 2,24 10,10 33,45 199 104
91B x 96A 61,84 63,00 1,17 2,50 15,00 30,30 194 101
17B x 22A 59,34 61,17 1,84 2,77 11,80 31,73 201 106
87A x 59B 61,00 62,67 1,67 2,23 12,35 32,15 191 100
170B x 169A 63,84 65,67 1,84 2,20 14,40 34,00 198 109
74B x 132A 61,50 63,34 1,84 2,60 18,35 32,50 183 97
92A x 47B 64,84 67,00 2,17 2,57 11,95 34,50 193 107
47A x 168B 64,00 65,50 1,50 2,20 12,55 32,65 200 112
Média prog. 61,93 63,46 1,53 2,36 12,33 32,92 196 104
Pop. Tolerante 62,17 63,17 1,00 2,07 10,90 33,00 185 91
Pop. Sensível 64,50 65,67 1,17 2,57 9,30 32,00 193 109
FT510 65,17 67,67 2,48 2,20 9,65 33,20 150 77
DAS2B707 61,67 62,50 0,83 1,87 7,80 37,00 190 96
Média test. 63,38 64,75 1,37 2,18 9,41 33,80 179 93
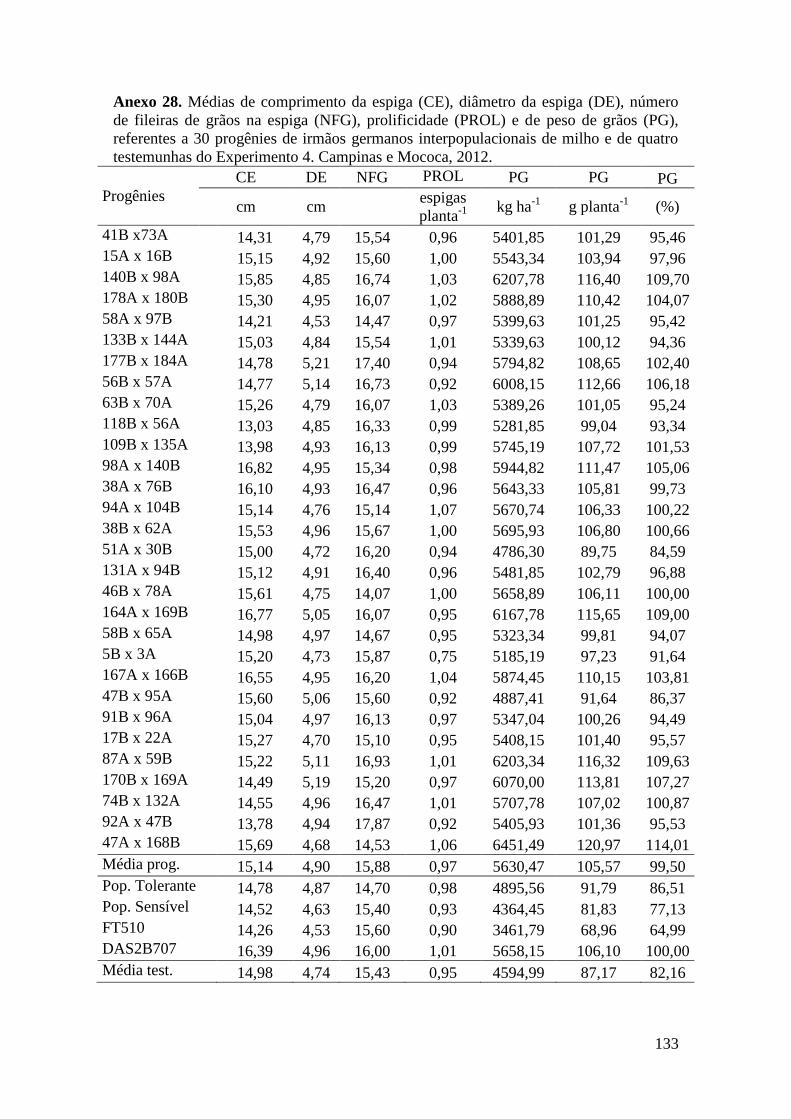
133
Anexo 28. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 4. Campinas e Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
41B x73A 14,31 4,79 15,54 0,96 5401,85 101,29 95,46
15A x 16B 15,15 4,92 15,60 1,00 5543,34 103,94 97,96
140B x 98A 15,85 4,85 16,74 1,03 6207,78 116,40 109,70
178A x 180B 15,30 4,95 16,07 1,02 5888,89 110,42 104,07
58A x 97B 14,21 4,53 14,47 0,97 5399,63 101,25 95,42
133B x 144A 15,03 4,84 15,54 1,01 5339,63 100,12 94,36
177B x 184A 14,78 5,21 17,40 0,94 5794,82 108,65 102,40
56B x 57A 14,77 5,14 16,73 0,92 6008,15 112,66 106,18
63B x 70A 15,26 4,79 16,07 1,03 5389,26 101,05 95,24
118B x 56A 13,03 4,85 16,33 0,99 5281,85 99,04 93,34
109B x 135A 13,98 4,93 16,13 0,99 5745,19 107,72 101,53
98A x 140B 16,82 4,95 15,34 0,98 5944,82 111,47 105,06
38A x 76B 16,10 4,93 16,47 0,96 5643,33 105,81 99,73
94A x 104B 15,14 4,76 15,14 1,07 5670,74 106,33 100,22
38B x 62A 15,53 4,96 15,67 1,00 5695,93 106,80 100,66
51A x 30B 15,00 4,72 16,20 0,94 4786,30 89,75 84,59
131A x 94B 15,12 4,91 16,40 0,96 5481,85 102,79 96,88
46B x 78A 15,61 4,75 14,07 1,00 5658,89 106,11 100,00
164A x 169B 16,77 5,05 16,07 0,95 6167,78 115,65 109,00
58B x 65A 14,98 4,97 14,67 0,95 5323,34 99,81 94,07
5B x 3A 15,20 4,73 15,87 0,75 5185,19 97,23 91,64
167A x 166B 16,55 4,95 16,20 1,04 5874,45 110,15 103,81
47B x 95A 15,60 5,06 15,60 0,92 4887,41 91,64 86,37
91B x 96A 15,04 4,97 16,13 0,97 5347,04 100,26 94,49
17B x 22A 15,27 4,70 15,10 0,95 5408,15 101,40 95,57
87A x 59B 15,22 5,11 16,93 1,01 6203,34 116,32 109,63
170B x 169A 14,49 5,19 15,20 0,97 6070,00 113,81 107,27
74B x 132A 14,55 4,96 16,47 1,01 5707,78 107,02 100,87
92A x 47B 13,78 4,94 17,87 0,92 5405,93 101,36 95,53
47A x 168B 15,69 4,68 14,53 1,06 6451,49 120,97 114,01
Média prog. 15,14 4,90 15,88 0,97 5630,47 105,57 99,50
Pop. Tolerante 14,78 4,87 14,70 0,98 4895,56 91,79 86,51
Pop. Sensível 14,52 4,63 15,40 0,93 4364,45 81,83 77,13
FT510 14,26 4,53 15,60 0,90 3461,79 68,96 64,99
DAS2B707 16,39 4,96 16,00 1,01 5658,15 106,10 100,00
Média test. 14,98 4,74 15,43 0,95 4594,99 87,17 82,16
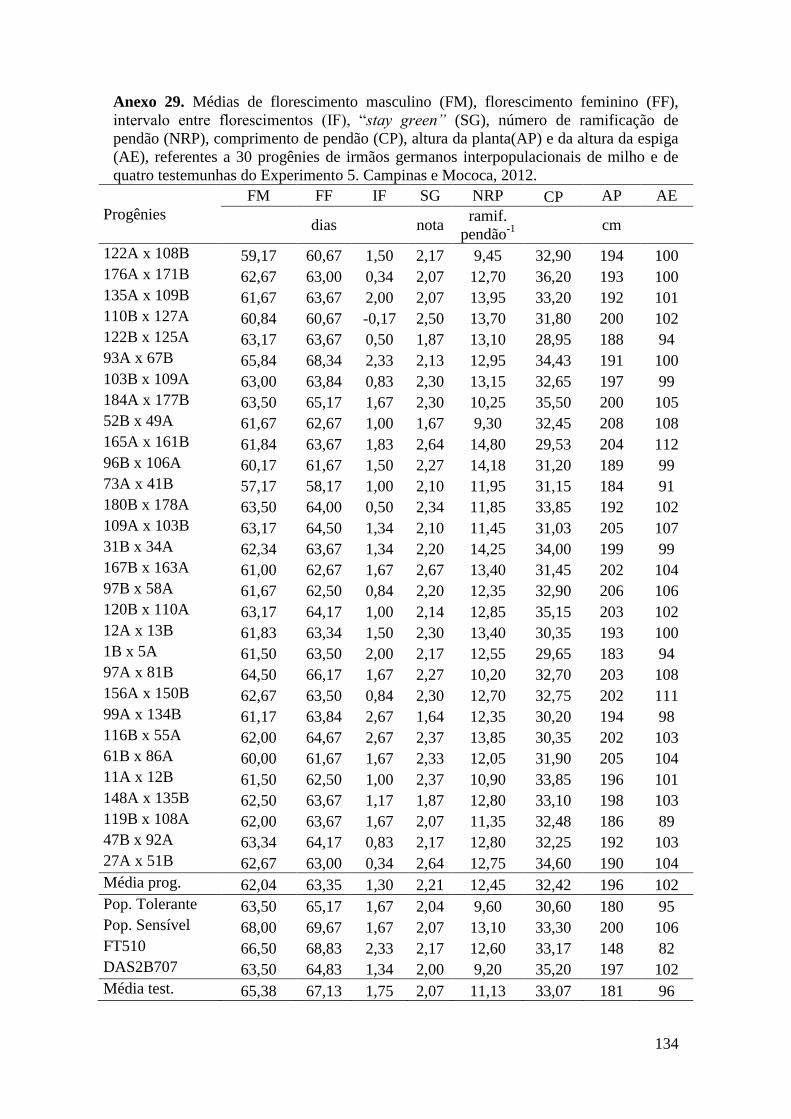
134
Anexo 29. Médias de florescimento masculino (FM), florescimento feminino (FF),
intervalo entre florescimentos (IF), “stay green” (SG), número de ramificação de
pendão (NRP), comprimento de pendão (CP), altura da planta(AP) e da altura da espiga
(AE), referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de
quatro testemunhas do Experimento 5. Campinas e Mococa, 2012.
Progênies
FM FF IF SG NRP CP AP AE
dias nota ramif.
pendão-1
cm
122A x 108B 59,17 60,67 1,50 2,17 9,45 32,90 194 100
176A x 171B 62,67 63,00 0,34 2,07 12,70 36,20 193 100
135A x 109B 61,67 63,67 2,00 2,07 13,95 33,20 192 101
110B x 127A 60,84 60,67 -0,17 2,50 13,70 31,80 200 102
122B x 125A 63,17 63,67 0,50 1,87 13,10 28,95 188 94
93A x 67B 65,84 68,34 2,33 2,13 12,95 34,43 191 100
103B x 109A 63,00 63,84 0,83 2,30 13,15 32,65 197 99
184A x 177B 63,50 65,17 1,67 2,30 10,25 35,50 200 105
52B x 49A 61,67 62,67 1,00 1,67 9,30 32,45 208 108
165A x 161B 61,84 63,67 1,83 2,64 14,80 29,53 204 112
96B x 106A 60,17 61,67 1,50 2,27 14,18 31,20 189 99
73A x 41B 57,17 58,17 1,00 2,10 11,95 31,15 184 91
180B x 178A 63,50 64,00 0,50 2,34 11,85 33,85 192 102
109A x 103B 63,17 64,50 1,34 2,10 11,45 31,03 205 107
31B x 34A 62,34 63,67 1,34 2,20 14,25 34,00 199 99
167B x 163A 61,00 62,67 1,67 2,67 13,40 31,45 202 104
97B x 58A 61,67 62,50 0,84 2,20 12,35 32,90 206 106
120B x 110A 63,17 64,17 1,00 2,14 12,85 35,15 203 102
12A x 13B 61,83 63,34 1,50 2,30 13,40 30,35 193 100
1B x 5A 61,50 63,50 2,00 2,17 12,55 29,65 183 94
97A x 81B 64,50 66,17 1,67 2,27 10,20 32,70 203 108
156A x 150B 62,67 63,50 0,84 2,30 12,70 32,75 202 111
99A x 134B 61,17 63,84 2,67 1,64 12,35 30,20 194 98
116B x 55A 62,00 64,67 2,67 2,37 13,85 30,35 202 103
61B x 86A 60,00 61,67 1,67 2,33 12,05 31,90 205 104
11A x 12B 61,50 62,50 1,00 2,37 10,90 33,85 196 101
148A x 135B 62,50 63,67 1,17 1,87 12,80 33,10 198 103
119B x 108A 62,00 63,67 1,67 2,07 11,35 32,48 186 89
47B x 92A 63,34 64,17 0,83 2,17 12,80 32,25 192 103
27A x 51B 62,67 63,00 0,34 2,64 12,75 34,60 190 104
Média prog. 62,04 63,35 1,30 2,21 12,45 32,42 196 102
Pop. Tolerante 63,50 65,17 1,67 2,04 9,60 30,60 180 95
Pop. Sensível 68,00 69,67 1,67 2,07 13,10 33,30 200 106
FT510 66,50 68,83 2,33 2,17 12,60 33,17 148 82
DAS2B707 63,50 64,83 1,34 2,00 9,20 35,20 197 102
Média test. 65,38 67,13 1,75 2,07 11,13 33,07 181 96
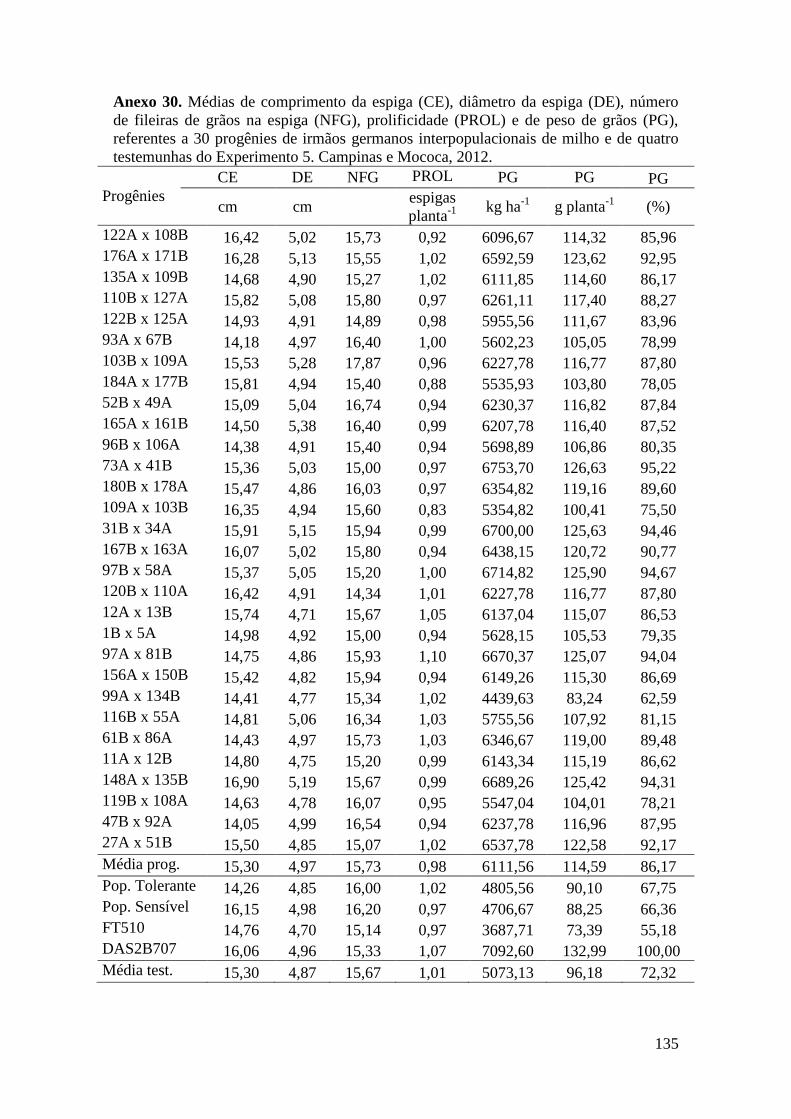
135
Anexo 30. Médias de comprimento da espiga (CE), diâmetro da espiga (DE), número
de fileiras de grãos na espiga (NFG), prolificidade (PROL) e de peso de grãos (PG),
referentes a 30 progênies de irmãos germanos interpopulacionais de milho e de quatro
testemunhas do Experimento 5. Campinas e Mococa, 2012.
Progênies
CE DE NFG PROL PG PG PG
cm cm espigas
planta-1
kg ha
-1 g planta
-1 (%)
122A x 108B 16,42 5,02 15,73 0,92 6096,67 114,32 85,96
176A x 171B 16,28 5,13 15,55 1,02 6592,59 123,62 92,95
135A x 109B 14,68 4,90 15,27 1,02 6111,85 114,60 86,17
110B x 127A 15,82 5,08 15,80 0,97 6261,11 117,40 88,27
122B x 125A 14,93 4,91 14,89 0,98 5955,56 111,67 83,96
93A x 67B 14,18 4,97 16,40 1,00 5602,23 105,05 78,99
103B x 109A 15,53 5,28 17,87 0,96 6227,78 116,77 87,80
184A x 177B 15,81 4,94 15,40 0,88 5535,93 103,80 78,05
52B x 49A 15,09 5,04 16,74 0,94 6230,37 116,82 87,84
165A x 161B 14,50 5,38 16,40 0,99 6207,78 116,40 87,52
96B x 106A 14,38 4,91 15,40 0,94 5698,89 106,86 80,35
73A x 41B 15,36 5,03 15,00 0,97 6753,70 126,63 95,22
180B x 178A 15,47 4,86 16,03 0,97 6354,82 119,16 89,60
109A x 103B 16,35 4,94 15,60 0,83 5354,82 100,41 75,50
31B x 34A 15,91 5,15 15,94 0,99 6700,00 125,63 94,46
167B x 163A 16,07 5,02 15,80 0,94 6438,15 120,72 90,77
97B x 58A 15,37 5,05 15,20 1,00 6714,82 125,90 94,67
120B x 110A 16,42 4,91 14,34 1,01 6227,78 116,77 87,80
12A x 13B 15,74 4,71 15,67 1,05 6137,04 115,07 86,53
1B x 5A 14,98 4,92 15,00 0,94 5628,15 105,53 79,35
97A x 81B 14,75 4,86 15,93 1,10 6670,37 125,07 94,04
156A x 150B 15,42 4,82 15,94 0,94 6149,26 115,30 86,69
99A x 134B 14,41 4,77 15,34 1,02 4439,63 83,24 62,59
116B x 55A 14,81 5,06 16,34 1,03 5755,56 107,92 81,15
61B x 86A 14,43 4,97 15,73 1,03 6346,67 119,00 89,48
11A x 12B 14,80 4,75 15,20 0,99 6143,34 115,19 86,62
148A x 135B 16,90 5,19 15,67 0,99 6689,26 125,42 94,31
119B x 108A 14,63 4,78 16,07 0,95 5547,04 104,01 78,21
47B x 92A 14,05 4,99 16,54 0,94 6237,78 116,96 87,95
27A x 51B 15,50 4,85 15,07 1,02 6537,78 122,58 92,17
Média prog. 15,30 4,97 15,73 0,98 6111,56 114,59 86,17
Pop. Tolerante 14,26 4,85 16,00 1,02 4805,56 90,10 67,75
Pop. Sensível 16,15 4,98 16,20 0,97 4706,67 88,25 66,36
FT510 14,76 4,70 15,14 0,97 3687,71 73,39 55,18
DAS2B707 16,06 4,96 15,33 1,07 7092,60 132,99 100,00
Média test. 15,30 4,87 15,67 1,01 5073,13 96,18 72,32


















