
Regulación de la expresión del Receptor Tipo Toll 4...
47
UNIVERSIDAD DE CHILE Facultad de Ciencias Químicas y Farmacéuticas Departamento de Bioquímica y Biología Molecular Regulación de la expresión del Receptor Tipo Toll 4 (TLR4) por el Factor de Necrosis Tumoral α y Glucocorticoides Ramón Daniel Pérez Núñez Memoria para optar al título de Bioquímico Profesor Patrocinante: Dr. Javier Puente Piccardo Departamento de Bioquímica y Biología Molecular Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile Directora de Memoria: Dra. Marcela Hermoso Ramello Laboratorio de Inmunidad Innata Programa Disciplinario de Inmunología Facultad de Medicina Universidad de Chile Santiago-Chile 2008
-
Upload
nguyenmien -
Category
Documents
-
view
218 -
download
0
Transcript of Regulación de la expresión del Receptor Tipo Toll 4...

UNIVERSIDAD DE CHILE
Facultad de Ciencias Químicas y Farmacéuticas Departamento de Bioquímica y Biología Molecular
Regulación de la expresión del Receptor Tipo Toll 4 (TLR4) por el Factor de Necrosis
Tumoral α y Glucocorticoides
Ramón Daniel Pérez Núñez Memoria para optar al título de Bioquímico
Profesor Patrocinante: Dr. Javier Puente Piccardo Departamento de Bioquímica y Biología Molecular Facultad de Ciencias Químicas y Farmacéuticas Universidad de Chile
Directora de Memoria: Dra. Marcela Hermoso Ramello Laboratorio de Inmunidad Innata Programa Disciplinario de Inmunología Facultad de Medicina Universidad de Chile
Santiago-Chile 2008

Esta memoria esta dedicada con todo mi cariño a mis padres, Ramón y María…
ii

AGRADECIMIENTOS
Creo que nunca voy a poder terminar de agradecer a todas aquellas personas
que estuvieron junto a mí, entregándome su apoyo y orientación en cada momento.
Por ello, la satisfacción de todo este trabajo lo quiero compartir con todos ustedes.
Agradeciendo en primer lugar a mi profesora guía, la Dra. Marcela Hermoso,
que me otorgó desde el principio la confianza y sabiduría necesaria para el buen
desarrollo de esta memoria.
De la misma manera agradecer a todos los que forman y formaron parte del
Laboratorio de Inmunidad Innata, lugar que me acogió y consideré como mi segundo
hogar, integrado por personas con las cuales compartí muy gratos momentos, que
conservaré por siempre, gracias…Rodrigo, Enzo, Frano, Alexis, Sergio, Bendelin,
Sofía, Alejandra, Lucía, Patricia, Paulina, Caroll, Salvador, David, José, Carolina,
María José, Luis, Benjamín…sin poder dejar de nombrar a una persona que considero
mucho y que me entregó permanentemente su amistad, bondad y energía, cuando
mas la necesitaba, gracias a Ud. Don Roli.
Al pilar fundamental que me ha sostenido toda la vida, mi familia, la cual nunca
se ha puesto limites para darme todo el apoyo y cariño que he necesitado, infinitas
gracias a mi papá, mi mamá y mis hermanitas Josefina, Ana y Gabriela…además de
una hermosa persona que considero parte de mi familia, que siempre está
entregándome su amor en la cosas mas simples de la vida, a mi niña linda,
Gabicienta…
iii

INDICE GENERAL
Página AGRADECIMIENTOS iii INDICE GENERAL iv INDICE DE FIGURAS vi ABREVIATURAS vii RESUMEN viii ABSTRACT ix 1. INTRODUCCION 1
1.1 Sistema Inmune 1 1.2 Receptores de Reconocimiento de Patógenos 2 1.3 Receptores Tipo Toll (TLRs) 3 1.4 Glucocorticoides y el Sistema Inmune 4 1.5 Regulación de la expresión de TLR4 por moléculas
pro- y anti-inflamatorias 6 2. HIPOTESIS 7
2.1 Objetivo General 7 2.2 Objetivos Específicos 7 2.3 Diseño Experimental 8
3. MATERIALES Y METODOS 10
3.1 Reactivos y Anticuerpos 10 3.2 Células 10 3.3 Transfección Transitoria y Plasmidios 11
3.4 PCR en Tiempo Real 12 3.5 Ensayo de Gen Reportero Luciferasa 13 3.6 Inmunoblot 14
iv

3.7 Citometría de Flujo 15 3.8 Estadística 16
4. RESULTADOS 17
4.1 Los glucocorticoides revierten la inducción de la expresión del mRNA de TLR4 inducido por TNFα 17
4.2 Los glucocorticoides modulan el efecto de TNFα sobre la actividad transcripcional del promotor del TLR4 21
4.2.1 Análisis bioinformático de los elementos de respuesta putativos del promotor de TLR4 murino 21
4.2.2 Glucocorticoides contrarrestan el efecto de TNFα sobre la actividad del promotor de TLR4 22 4.3 Los glucocorticoides revierten el aumento de la expresión
de TLR4 a nivel proteico inducida por TNFα 24
4.3.1 Dexametasona antagoniza la inducción de TLR4 mediada por TNFα a nivel de proteína total 24
4.3.2 Efecto de glucocorticoides sobre la expresión del TLR4 a nivel de superficie inducida por TNFα 26
5. DISCUSION 28
5.1 Efecto de TNFα y Dexametasona sobre la expresión del mRNA de TLR4 en células A549 28
5.2 Efecto de TNFα sobre la actividad transcripcional del promotor del TLR4 en presencia o ausencia de Dexametasona 29
5.3 Efecto de los glucocorticoides sobre la expresión de la proteína de TLR4 inducida por TNFα 30
5.4 Regulación de la expresión de TLR4 por glucocorticoides inducida por TNFα 31
6. CONCLUSIONES 33 7. REFERENCIAS 34
8. ANEXO 38
v

INDICE DE FIGURAS
Página Figura 1. Curva estándar de PCR en Tiempo Real para TLR4 y diagrama del plasmidio pCMV-Flag-hTLR4. 18 Figura 2. Curva estándar de PCR en Tiempo Real para G3DP. 19 Figura 3. Dexametasona revierte el efecto de TNFα sobre la expresión del mRNA de TLR4. 20
Figura 4. Promotor de TLR4 murino. 21 Figura 5. Actividad transcripcional del promotor de TLR4. 23 Figura 6. Curso temporal de la expresión de TLR4 inducido por TNFα. 25 Figura 7. Curso temporal de la expresión de TLR4 total en células expuestas a Dexametasona. 25 Figura 8. Los glucocorticoides revierten el efecto inductor de TNFα sobre a la expresión de la proteína de TLR4. 26
Figura 9. Expresión de TLR4 en la membrana plasmática de células epiteliales humanas. 27
Figura 10. Modelo de la regulación de la expresión de TLR4 por glucocorticoides inducida por TNFα. 32
vi

ABREVIATURAS A549: Línea celular, derivada de un adenocarcinoma de pulmón ACTH: Hormona adrenocorticotropina AP-1: Factor de transcripción Proteína activadora 1 CD14: Co-receptor de LPS C3H/HeJ y C57BL/10ScCr: Cepas Murinas Mutantes del gen de tlr4 Dex: Dexametasona DEPC: Dietilpirocarbonato DMEM: Medio de cultivo celular (Dulbecco´s modified Eagle´s médium) DNA: Acido desoxirribonucleico dNTPs: deoxinuleosidos trifosfatos mRNA: RNA mensajero DTT: Ditiotreitol EDTA: Acido dietilendiamintetraacetico ERK: Quinasa reguladas por señales extracelulares F12: Medio de cultivo celular FBS: Suero fetal bovino G3DP: Gliceraldehido-3-fosfo deshidrogenasa GCs: Glucocorticoides GR: Receptor de glucocorticoides GRE: Elemento de respuesta a glucocorticoides HPA: Eje Hipotálamo-Hipófisis-Glándula adrenal hsp: Proteínas de shock térmico IL-x: Interleuquinas INF-β: Interfreron β IκB: Proteína inhibidora κappa B LBP: Proteína de unión a LPS LDB: Tampón de baja concentración de detergente LPS: Lipopolisacarido LRRs: Repeticiones ricas en Leucina MD-2: Proteína de unión al dominio extracelular de TLR4 MHC: Complejo mayor de histocompatibilidad MyD88: Proteína 88 de diferenciación mieloide NFκB: Factor nuclear κappa B Oligo(dT): Partidor inespecífico poli-T(15) PAMPs: Patrones moleculares asociados a patógenos PCR: Reacción de polimerasa en cadena PBS: Tampón salino a base de fosfato phRL: Plasmidio control de transfección con el gen de Renilla pGL3: Plasmidio básico de clonamiento PRRs: Receptores de reconocimiento de patrones SDS: Dodecil sulfato de sodio STAT1: Factor transcripcional de la señal transductora y activadora transcripcional RLU: Unidad Relativa de Luciferasa RRU: Unidad Relativa de Renilla RT: Transcripción reversa TBS: Tampón salino a base de Tris TIRAP: Proteína adaptadora que contiene dominio TIR TIR: Dominio intracelular, homologo al receptor de IL-1 y de TLR TLRs: Receptores tipo Toll TNFα: Factor de necrosis tumoral α TRIF: Molécula Adaptadora que contiene un dominio TIR y que induce INF-β
vii

RESUMEN
Durante la inflamación de las mucosas, las células epiteliales y los macrófagos
expresan el receptor de tipo Toll 4 (TLR4) y secretan citoquinas, las que potencian el
cuadro. Sin embargo, hasta ahora existe escasa información acerca de la regulación
de la expresión de TLR4 por la combinación de moléculas pro-inflamatorias y anti-
inflamatorias.
Utilizando la línea celular de epitelio pulmonar, las células A549, se evaluó la
expresión del TLR4 a nivel de transcrito, proteína y actividad transcripcional de
mutantes con deleciones en la región promotora del receptor, posterior a la exposición
al factor de necrosis tumoral α (TNFα) en presencia o ausencia del glucocorticoide
sintético Dexametasona (Dex), mediante PCR en tiempo real, inmunoblot o citometría
de flujo, y ensayos del gen reportero luciferasa, respectivamente.
El aumento de la expresión de TLR4 inducido por TNFα, tanto a nivel del
mRNA como de la proteína, es revertido por la adición de Dex. La regulación de la
actividad transcripcional del promotor de TLR4 por estas moléculas fue similar a la
observada a nivel de proteína, y el efecto inhibitorio de Dex estaría comandado por al
menos un elemento de respuesta del receptor de glucocorticoides, GR (GRE).
En conjunto, estos resultados demuestran un efecto antagónico de
Dexametasona sobre la expresión de TLR4 inducida por TNFα, aparentemente por un
mecanismo que involucra al menos la regulación de GR a nivel del promotor de TLR4.
Se evidenció claramente en estas células los fuertes efectos antagónicos que ejercen
sobre la expresión de TLR4, moléculas pro-inflamatorias como es TNFα y anti-
inflamatorias como los glucocorticoides.
viii

ABSTRACT
“Regulation of The Toll-Like Receptor 4 expression by Tumor Necrosis Factor α and Dexamethasone”
During mucosal inflammation, epithelial cells and macrophages express TLR4
as well as cytokines that might induce its expression. Up to now, there is scarce
information about TLR4 regulation by the combination of pro-inflammatory and anti-
inflammatory molecules.
We used A549 cell line derived from lung epithelium treated with Tumor
necrosis factor alpha TNFα, in the presence or absence of the synthetic glucocorticoid
Dexamethasone (Dex). TLR4 expression was assessed at the transcript and protein
levels by real-time PCR, immuneblot or flow cytometry, respectively, as well as
evaluating the transcriptional activity of promoter deletion mutants by the luciferase
reporter assay.
The increase in TLR4 protein and mRNA levels upon TNFα exposure was
reverted by Dex. This effect may be mediated by the glucocortocoid receptor, GR.
Analysis of transcriptional activity indicate that at least one GR response element
(GRE) is required for the inhibitory effects of Dex over the TNFα response.
Taken together, these results indicate that there is an antagonist effect of TNFα
and Dex at the TLR4 expression. The strong effect of pro-inflammatory as TNFα and
anti-inflammatory as Dex is clearly evidenced in this cell line.
ix

1. INTRODUCCIÓN 1.1 Sistema Inmune El sistema inmune comprende la respuesta inmune innata y adquirida, ambas
integradas por un conjunto de células y moléculas con roles especializados en la
defensa contra infecciones provocadas por agentes patógenos, que difieren en cuanto
a su estructura y funcionalidad, respectivamente (1). El sistema inmune innato
corresponde a la primera barrera defensiva física, tal como la piel y las mucosas, y
está integrado por células, tales como basófilos, eosinófilos, y natural killer, y otras con
función fagocítica, tales como neutrófilos, monocitos y macrófagos, o de barrera física
como las células epiteliales. Estas células producen mediadores inflamatorios, como
las citoquinas que modulan la respuesta inmune innata (2), sumado a otros
componentes moleculares tales como el sistema del complemento (3).
Por otra parte, la respuesta inmune adaptativa esta constituida por dos clases
de células especializadas, los linfocitos T y los linfocitos B, que despliegan un único
receptor producido somáticamente durante el desarrollo celular, aportando una amplia
diversidad en su repertorio de reconocimiento de antígenos, a diferencia de la
inmunidad innata, mediada por receptores codificados por líneas germinales y de
especificidad predeterminada genéticamente (1, 4).
El sistema inmune innato es activado inmediatamente después de la invasión
de un patógeno, el cual interactúa posteriormente con el sistema inmune adaptativo,
para inducir una expansión clonal de sus tipos celulares (5).
1

Los receptores de los linfocitos reconocen a su ligando, mediado por células
presentadoras de antígeno, tales como células dendríticas o macrófagos, las que
exponen péptidos derivados del patógeno previamente procesado, en el contexto del
complejo mayor de histocompatibilidad (MHC), modulando y controlando la activación
de la respuesta inmune adaptativa (6).
En la fase inicial del reconocimiento de los patógenos participan receptores
especializados para censarlos, centrándose el trabajo de esta memoria en la
regulación de uno de los participantes este evento crucial de activación de la
respuesta inmune innata.
1.2 Receptores de Reconocimiento de Patógenos
Los receptores de reconocimiento de patrones (Pattern Recognition Receptors,
PRRs) identifican patrones moleculares asociados a patógenos (Pathogen-Associated
Molecular Patterns, PAMPs), los cuales corresponden a motivos conservados
esenciales para la sobrevida y patogenicidad del microorganismo (7).
Un tipo de PRRs son aquellos que activan vías de transducción intracelulares o
señalizadores, que conducen a la expresión de una variedad de genes de la respuesta
inmune, incluyendo citoquinas pro-inflamatorias. Destaca entre estos PRRs la familia
de los Receptores de tipo Toll (TLRs), homólogos a los Toll de Drosophila (8),
identificados a inicios de la década de los 90 y vinculados con el desarrollo
embrionario dorso-ventral y posteriormente por su papel determinante en la respuesta
inmune contra infecciones fúngicas (9).
2

1.3 Receptores Tipo Toll (TLRs)
Los TLRs son receptores de transmembrana de tipo I que juegan un papel
clave en la respuesta inmune innata (10), representando la primera línea de defensa
contra patógenos. Los TLRs poseen un dominio intracelular TIR (homólogo al del
receptor de IL-1, Toll/IL-1R), crucial en la activación de la vía de señalización, y un
dominio extracelular, que contiene repeticiones ricas en leucina (Leucine Rich
Repeats, LRR) involucradas en el reconocimiento de los PAMPs (11). Hasta el
momento, 11 TLRs han sido identificados en humanos y 13 en ratón los que, de
acuerdo al patógeno que reconocen, se pueden agrupar en dos categorías: TLRs de
PAMPs bacterianos (TLR2, 4 y 5) o virales (TLR3, 7/8 y 9), los que se localizan
principalmente en la superficie celular o requieren su internalización a compartimientos
endosomales, respectivamente (12).
Este trabajo se ha centrado en el estudio de la regulación de la expresión de un
integrante de esta familia de receptores, el TLR4, que reconoce el lipopolisacárido
(LPS), un componente glicolipídico de la membrana externa de bacterias gram-
negativas (13).
La resistencia al shock endotóxico que poseen los ratones C3H/HeJ y
C57BL/10ScCr es consecuencia de una mutación en el gen tlr4, que disminuye la
respuesta a LPS (14). Para la unión del ligando, el TLR4 requiere de otras moléculas
adicionales: inicialmente el LPS se une a una proteína presente en el suero (LBP;
Lipopolysaccharide-Binding Protein), y posteriormente son reconocidas por el receptor
CD14 que está anclado a la membrana por glucosilfosfatidilinositol. EL CD14 se
expresa en forma preferencial en monocitos, macrófagos y neutrófilos. Adicionalmente,
a este complejo se incorpora a la proteína secretada MD-2, que se asocia con la
porción extracelular del TLR4, estabilizando el complejo y la respuesta pro-inflamatoria
3

a LPS (15). La vía de señalización del receptor TLR4 se inicia en el dominio TIR, por la
asociación de moléculas adaptadoras, tales como: MyD88 (myeloid differentiation
primary response protein 88), TIRAP (TIR domain-contanining adaptor protein) y TRIF
(TIR domain-containing adaptor inducing IFN-β), activando factores de transcripción,
tales como NFκB (Factor Nuclear κappa B) (16), y conduciendo a la producción de
mediadores pro-inflamatorios, tales como TNFα (Factor de Necrosis Tumoral α), IL-12
(Interleuquina -12) (17; 18).
El TNFα, que en su forma soluble pesa 17 KDa, aumenta la expresión del gen
tlr4, mediante un mecanismo que involucra la unión a su receptor de membrana
plasmática, desencadenando una cascada de señales intracelulares que activan a
NFκB y la Proteína Activadora 1 (AP-1), aumentando la producción de citoquinas pro-
inflamatorias y de TLR4 (19).
1.4 Glucocorticoides y el Sistema Inmune
Las hormonas esteroidales han sido reconocidas como moléculas moduladoras
de diversos procesos celulares esenciales, tales como transducción de señales,
comunicación intercelular y metabolismo intermedio de carbohidratos, proteínas y
lípidos, regulando la glicemia, y el procesamiento de aminoácidos y ácidos grasos
libres, entre otros (20). Los glucocorticoides (GCs) son sintetizados en la zona
fasciculada/reticulada de la corteza adrenal y liberados a la circulación en respuesta a
estímulos estresantes. La secreción de los GCs es activada por el eje hipotálamo-
hipófisis-glándula adrenal (HPA), mediante la secreción hipotalámica de la hormona
liberadora de corticotropina (CRH), que activa la liberación de la hormona
adrenocorticotropina (ACTH) hipofisiaria, la cual finalmente, activa receptores
específicos en la glándula adrenal que inducen la secreción de cortisol (21).
4

Los GCs circulan en el plasma asociados a moléculas transportadoras, tales
como la globulina y la albúmina, que unen ligandos endógenos y/o sintéticos,
respectivamente (22). Los GCs regulan la función del sistema cardiovascular, tal como
la tensión vascular, y nervioso central, protegiendo al cerebro de las respuestas
inmune descontroladas, disminuyendo la expresión de moléculas de la inmunidad
adaptativa (23; 24).
Los GCs son moléculas lipofílicas que atraviesan la membrana celular
mediante difusión simple y se unen a un receptor citoplasmático (GR), translocándose
posteriormente al núcleo, donde el complejo hormona-receptor interacciona con una
región conservada dentro de promotores de determinados genes, los elemento de
respuesta a glucocorticoides (GRE) (25). Dependiendo de la composición de bases del
GRE, este puede ser activador o represor, y además del tipo celular en que se
encuentre, activa la expresión de genes, tales como el de IκB (proteína inhibidora IκB)
que inhibe la actividad transcripcional de NFκB. Mediante la interacción entre ambas
proteínas, IκB mantiene secuestrado a NFκB en el citoplasma, impidiendo su
translocación al núcleo (26). En ausencia de GCs, el GR se encuentra inactivo en el
citoplasma asociado a proteínas de shock térmico 56, 70 y 90 (hsp56, hsp70 y hsp90),
presentando una gran afinidad por el ligando (27).
Dada su propiedad anti-inflamatoria (28; 29), se han desarrollado GCs
sintéticos, tales como la Dexametasona (Dex), utilizados en el área clínica, para el
tratamiento de una amplia gama de patologías relacionadas con procesos
inflamatorios, tales como Lupus Eritematoso Sistémico, Enfermedad Inflamatoria
Intestinal, Psoriasis, Asma y Artritis Reumatoidea, entre otras (30).
5

1.5 Regulación de la expresión de TLR4 por moléculas pro- y anti-inflamatorias
La expresión de los TLRs puede ser modulada por diversos factores
involucrados en vías pro- o anti-inflamatorias, tales como citoquinas y GCs,
respectivamente. Estudios recientes con uno de los integrantes de la familia de los
TLRs, el TLR2, indican que el GC sintético Dexametasona, en combinación con TNFα,
aumenta sinérgicamente la expresión inducida por la citoquina, en células de epitelio
pulmonar (31). Estos resultados proponen mecanismos de acción de los GCs en la
respuesta inmune innata, que clásicamente han sido vinculados con la inhibición de la
expresión de moléculas de la inmunidad adaptativa (32).
Hasta la fecha, no existen evidencias sobre el efecto conjunto de los estímulos
antagónicos anteriormente citados, sobre la expresión y función del TLR4, tanto a nivel
genético como de la proteína. Por esta razón, en esta memoria se estudió el papel que
ejercen los GCs sobre la expresión de TLR4 inducida por TNFα.
Conociéndose los cambios en la expresión del gen tlr4, que ocurren en
respuesta a la combinación de TNFα y Dexametasona, se podrían estructurar mejores
tratamientos para diversas patologías que cursan con un cuadro inflamatorio en
respuesta a infecciones gram-negativas, tales como shock séptico (33; 34). En este
trabajo en particular, se utilizó el modelo celular de epitelio pulmonar humano (células
A549), que expresa tanto el TLR4 como el GR, como una aproximación experimental
al estudio de patologías respiratorias donde el componente bacteriano tienen un papel
central en ellas (35).
6

2. HIPÓTESIS
En base a los antecedentes indicados, surge el interés de evaluar la regulación
de la expresión del TLR4, dada su relevancia dentro del sistema inmune innato, por lo
que planteamos la siguiente hipótesis en esta memoria para optar al título de
Bioquímico:
“El Factor de Necrosis Tumoral α y los Glucocorticoides regulan
antagónicamente la expresión del Receptor Tipo Toll 4 en células de
epitelio pulmonar humano”
2.1 Objetivo General
Determinar el efecto del TNFα en presencia o ausencia de glucocorticoides
sobre la expresión del Receptor tipo Toll 4 (TLR4) en células de epitelio pulmonar.
2.2 Objetivos Específicos 1. Determinar el efecto de TNFα en presencia o ausencia del glucocorticoide
sintético Dexametasona, sobre la expresión del mRNA del TLR4 en una línea celular
de epitelio pulmonar humano, las células A549.
2. Evaluar el efecto de TNFα sobre la actividad transcripcional del promotor del
TLR4, en presencia o ausencia de Dexametasona, en células A549.
3. Determinar la expresión de la proteína total y de superficie del TLR4 en
células A549 expuestas a TNFα en presencia o ausencia de Dexametasona.
7

2.3 Diseño Experimental
Para desarrollar el objetivo específico 1: “Determinar el efecto de TNFα en
presencia o ausencia del glucocorticoide sintético Dexametasona (Dex), sobre la
expresión del mRNA del TLR4 en una línea celular de epitelio pulmonar humano,
las células A549”.
Se evaluó la expresión del transcrito de TLR4 por PCR en tiempo real, para lo
cual se confeccionó en primer lugar la curva estándar para TLR4, utilizando un
plasmidio que contiene la secuencia codificante para TLR4 humano (pCMV-Flag-
hTLR4). La expresión del mRNA de TLR4 se normalizó con el gen constitutivo
gliceraldehido 3-fosfo deshidrogenasa (G3DP). Para determinar la expresión del
transcrito de G3DP se generó una curva estándar con el producto amplificado a través
de una PCR convencional con partidores específicos. Para ello, se determinó la
concentración del amplicón de G3DP en un gel de agarosa mediante análisis
densitométrico de las bandas obtenidas en relación al estándar de peso molecular.
Posteriormente se determinó la expresión del mRNA de TLR4 y de G3DP con
los programas de PCR y partidores específicos, previamente definidas. Los resultados
del producto amplificado y su cuantificación se realizaron a través del programa
Opticon Monitor 3.
Para desarrollar el objetivo específico 2: “Evaluar el efecto de TNFα sobre la
actividad transcripcional del promotor del TLR4, en presencia o ausencia de
Dexametasona en células A549”.
Se abordó siguiendo las estrategias experimentales que se indican a
continuación:
8

a) Diseñar un diagrama teórico de los sitios de unión putativos de los factores
de transcripción presentes en la región del promotor del gen tlr4. Para ello, se
utilizaron los programas Genomatix y TFsearch de acceso en
http://www.genomatix.de/index.html y http://www.cbrc.jp/research/db/TFSEARCH.html,
respectivamente, que aplican algoritmos determinados con valores variables para
identificar cada uno de los elementos o dominios.
b) Expresar transitoriamente el vector pGL3 que posee el gen reportero
Luciferasa y el DNA del promotor de TLR4, de 2715 pb (pGL3-2715) o una serie de
deleciones del mismo (pGL3-1864, -1183, -336 y -144 bp) y el plasmidio control de
transfección con el gen de Renilla (phRL) en las células A549. Para ello, las células
fueron transfectadas con cada uno de estos constructos por separado y
posteriormente se estimularon con TNFα en presencia o ausencia de Dexametasona.
Posteriormente, se determinó la actividad de luciferasa (unidades relativas de
luciferasa, relative luciferase units; RLU) y renilla (unidades relativas de renilla, relative
renilla units; RRU), utilizando un Luminómetro de placa (TopCount NXT, PerkinElmer,
USA); la razón entre RLU/RRU fue proporcional a la actividad transcripcional de TLR4.
Para desarrollar el objetivo específico 3: “Determinar la expresión de la
proteína total y de superficie del TLR4 en células A549 expuestas a TNFα en
presencia o ausencia de Dexametasona”.
Se determinó la expresión de la proteína de TLR4 mediante inmunoblot de
extractos proteicos totales de células A549 expuestas a TNFα en presencia y ausencia
de Dexametasona.
Además, se determinó la expresión superficial de la proteína de TLR4, en
células expuestas a los estímulos de estudio, mediante citometría de flujo.
9

3. MATERIALES y MÉTODOS
3.1 Reactivos y Anticuerpos
TNFα recombinante humano proviene de R&D Systems; Dexametasona
obtenida de Steraloids (Wilton, NH, USA), SYBR Green I Platinum SYBER Green
qPCR SuperMix UDG proviene de Invitrogen. Las enzimas Taq polimerasa y
Transcriptasa Reversa provienen de Promega, DNasa (Ambion). Para los ensayos de
gen reportero luciferasa se utilizó el kit Dual-Luciferase® Reporter Assay System de
Promega, Reactivo de Bradford (BioRad) para la cuantificación de proteínas. Para los
inmunoblots se contó con el anticuerpo anti-TLR4 humano, en tanto que para la
citometría de flujo, con el anticuerpo conjugado anti-TLR4-PE, ambos producidos en
eBioscience (San Diego, CA, USA). Se usaron además anticuerpos secundarios
conjugados con peroxidasa y el reactivo quimio-luminiscente ECL, generados por la
empresa Amersham Pharmacia Biotech (Piscataway, NJ, USA).
3.2 Células
El modelo utilizado fue la línea celular A549 proveniente de un adenocarcinoma
pulmonar, las cuales expresan TLR4 y el receptor de GC (31). Las células se
cultivaron en medio DMEM (Dulbecco´s modified Eagle´s medium)/F12 (1:1)
suplementado con 5% de suero fetal bovino (SFB), 100 IU de penicilina por ml y 100
mg/ml de estreptomicina (Pen/Str), el que fue cambiado cada 48 horas. Los cultivos
celulares fueron mantenidos en una incubadora a 37ºC con 5% de CO2 y atmósfera
humedecida.
10

3.3 Transfección Transitoria y Plasmidios
Las transfecciones se realizaron con el reactivo Fugene® (lípido catiónico)
según especifica el fabricante (Roche, Indianapolis, IN, USA). En primer lugar, se
realizó una mezcla que incluyó: Optimen, el plasmidio conteniendo las diferentes
construcciones del promotor del TLR4, el plasmidio control de transfección con el gen
de Renilla (phRL) y Fugene, incubados por 40 min (para permitir la formación óptima
de las micelas con el lípido y los DNA). Luego se agregó la mezcla a las células,
estimulándolas a las 24 horas con TNFα en presencia o ausencia de Dex.
Finalmente, luego de 24 horas con los estímulos, los lisados celulares de cada
condición se exponen al sustrato de la luciferasa (luciferina), cuyo metabolito es
identificado con un Luminómetro de placa (TopCount NXT, PerkinElmer, USA).
Para estandarizar las condiciones de los ensayos de PCR en tiempo real, se
utilizó el plasmidio pCMV-Tag1 que contiene la secuencia codificante para TLR4
(TLR4-Flag; donado por el Dr. Mantovani) (36).
Para los experimentos de actividad transcripcional se contó con el plasmidio
pGL3-Vector Basic que contiene el gen reportero luciferasa, y donde se han insertado
las deleciones del promotor de TLR4 (pGL3-2715, -1864, -1183, -336y -144 bp),
donados por el Dr. Thierry Roger (Universidad de Vaudois, Suiza) (37). Como control
de transfección se utilizó el vector pGL3-hRL, que codifica para la enzima luciferasa de
Renilla reniformes (renilla reporter vectors; Promega Corp., Madison, WI).
Se utilizaron además plasmidios controles para cada estímulo, como el vector
3xMHC-Luc (que posee tres sitios de unión a NFκB, correspondiente al promotor del
complejo mayor de histocompatibilidad humano y que codifica para la enzima
luciferasa de la luciérnaga Photinus pyralis) para determinar la actividad transcripcional
de NFκB, activada a través de TNFα. Además del vector MMTV-Luc (38), generado en
el laboratorio del Dr. J.A. Cidlowski, que posee elementos de respuesta a
11

glucocorticoides (GREs), con la finalidad de determinar la actividad del factor
transcripcional del GR, inducido por dexametasona (39).
3.4 PCR en Tiempo Real
Se determinó la expresión del mRNA de TLR4 y del gen constitutivo G3DP por
medio de la técnica PCR en Tiempo Real, donde la detección por fluorescencia mide
la cantidad de ADN sintetizado en cada momento de la reacción (40). La emisión de
fluorescencia producida en la reacción fue proporcional a la cantidad de ADN
amplificado. Como sistema de detección de fluorescencia se utilizó un agente
intercalante de ADN de doble hebra, denominado SYBR Green I Platinum SYBER
Green qPCR SuperMix UDG (SYBR Green I, 60 U/ml Platinum Taq DNA polymerase,
40 mM Tris-HCl pH 4.8 ,100 mM KCl, 6 mM MgCl2, 400 μM dGTP, 400 μM dATP, 400
μM dCTP, 400 μM dUTP, 40 U/ml UDG, and stabilizers) Invitrogen.
Para la obtención de RNA total de las células A549 se utilizó el método de
extracción de Trizol, de acuerdo a protocolos del fabricante. Las células se lisaron y
sometieron a centrifugación con cloroformo. Se recuperó la fase acuosa y el RNA se
precipitó con isopropanol. El precipitado de RNA obtenido de la centrifugación se lavó
con etanol y luego se resuspendió en 30 µl de agua tratada con dietilpirocarbonato
(DEPC). El RNA extraído fue tratado con DNasa y se determinó la concentración por
absorbancia a 260 nm.
En la trascripción reversa se utilizó 5 μg de RNA total, tampón transcriptasa
reversa 5X, partidor inespecífico oligo(dT) (500 μg/ml), deoxinucleosidos trifosfatos
(dNTP) 10 mM, 50 U de M-MLV Reverse Transcriptase. La denaturación inicial de las
estructuras secundarias del RNA se realizó a 72ºC por 10 minutos, seguido por el
12

alineamiento y transcripción del templado a 42ºC, durante 50 minutos. La temperatura
posteriormente se elevó a 72ºC por 10 minutos para inactivar la RT.
Cada reacción individual de PCR en tiempo real se realizó para un volumen
final de 25 μl, conteniendo 12,5 μl de Platinum SYBER Green qPCR SuperMix UDG, 1
μl de cada partidor específico (10 μM) tanto para TLR4 5`-TAC GTT TCC TTA TAA
GTG TCT GA-3` (sense) y 5`-TTA TCT GAA GGT GTT GCA CAT TCC-3` (antisense)
como para G3DP 5’-CGG ATT TGG TCG TAT TGG G-3’ (sense) y 5’-CAC AGT CTT
CTG GGT GGC-3’ (antisense), 8,5 μl de H2O y 2 μl de muestra.
El programa de PCR para TLR4 consiste en una primera etapa de activación a
95ºC por 10 minutos, luego a 94ºC por 15 segundos (denaturación), el alineamiento a
53ºC por 20 segundos y la elongación a 72ºC por 30 segundos, por 40 ciclos esta
primera etapa. Luego se realizó la curva de disociación, donde se incubó a 72ºC por
10 minutos, realizándose un gradiente de temperatura hasta los 92ºC, con una lectura
de la caída de la luminiscencia cada 0,2ºC, incubándose el producto restante a 20ºC,
lo mismo se hizo para el gen constitutivo G3DP donde el programa de PCR fue de 1
ciclo de 95ºC por 10 minutos, 40 ciclos de 95ºC por 30 segundos, 59ºC por 30
segundos y 72ºC por 30 segundos, realizándose finalmente la respectiva curva de
disociación, utilizándose el termociclador Chromo4™ Multicolor Real-Time PCR
Detection System (MJ Research). El análisis de los resultados del producto
amplificado y su cuantificación se realizaron a través del programa Opticon Monitor 3.
3.5 Ensayo de Gen Reportero Luciferasa
Las transfecciones fueron realizadas con el reactivo Fugene® (Roche,
Indianapolis, IN, USA), donde en primer lugar, se subcultivaron células A549 hasta
obtener un 70% de confluencia. Al segundo día, se cambió el medio de mantención
13

DMEN/F12, 5% SFB, Pen/Str por el medio de transfección OPTIMEM (Life Sciences,
Inc., St. Petersburg, FL, USA).
La transfección se inicia con la preparación de la mezcla de 200 µl de
Optimen/ml de medio, 2 µg de DNA y 5 µl de Fugene®/1 µg de DNA, que se incubó
por 40 minutos a temperatura ambiente y se aplicó en los pocillos suavemente,
manteniéndose a 37ºC por 24 horas. Al tercer día, se repuso el medio de cultivo
DMEM/F12, 5%SFB, Pen/Str y se realizaron los respectivos tratamientos.
Para la determinación de la actividad transcripcional de TLR4, las células
fueron cultivadas y posteriormente transfectadas con alguno de los constructos del
promotor de TLR4-Luc y phRL (que contiene el gen reportero Renilla como control de
transfección). A las 24 horas siguientes a la estimulación con TNFα en presencia o
ausencia de Dexametasona, se determinó la actividad de luciferasa utilizando un
luminómetro de placa (TopCount NXT PerkinElmer, USA). Los resultados se
expresaron como la inducción/inhibición relativa de la actividad luciferasa, que
corresponde a la razón entre los valores absolutos de luciferasa y de renilla (phRL), y
estos normalizados a la condición control.
3.6 Inmunoblot
Para realizar los inmunoblots se utilizaron 1x106 células por ml, las que fueron
tratadas con TNFα, en presencia o ausencia de Dexametasona. Posteriormente, se
removió el medio y las células se lavaron con PBS 1% y se lisaron en un tampón LDB
(LDB: low detergent buffer, cuya composición en mM es: 20 Tris-HCl, pH 7.5, 2 EDTA,
150 NaCl, y 0.5% Triton X-100, tabletas de inhibidores de proteasas (Roche®) e
inhibidores de fosfatasas (100 nM de ortovanadato de sodio, pirofosfato de sodio y
14

fluoruro de sodio), para luego ser homogenizadas con un sonicador de vástago
(Microson) a 14 Watts por 10 segundos.
La concentración de proteínas fue determinada con el método de Bradford
utilizando el reactivo de Bio-Rad, siguiendo las especificaciones del fabricante (Bio-
Rad Laboratories, Inc., Hercules, CA, USA). A una cantidad equivalente de proteínas
se le agregó tampón de muestra conteniendo SDS y β-mercaptoetanol, para luego ser
denaturadas por calor (5 minutos a 100ºC). Posteriormente, 80 μg de proteína de cada
muestra se cargaron y corrieron en un gel de poliacrilamida con SDS (SDS-PAGE) al
10%, utilizando la solución de tampones para la porción concentradora y separadora
del gel descrita por Laemmli (41). Las proteínas fueron transferidas a una membrana
de nitrocelulosa y, luego, teñidas con una solución de rojo Ponceau S al 1%. La
membrana se lavó con una solución de TBS-Tween-20 0,1% (5M NaCl, 1M Tris/HCl
pH 7,5 y 0,1% Tween-20) y fue bloqueada toda la noche con leche descremada al 5%
en TBS-Tween-20 al 0,1%. La membrana se incubó con los anticuerpos primarios y
secundarios diluidos en TBS-Tween-20 al 0,1% por 2 y 1 hora, respectivamente,
lavándose la membrana con TBS-Tween-20 0,1% por 30 minutos entre y después de
la incubación con los anticuerpos. Finalmente, se reveló la membrana con el reactivo
quimio-luminiscente ECL de Amersham Pharmacia Biotech (Piscataway, NJ, USA). El
análisis densitométrico de las bandas inmunoreactivas fue realizado con el software
Gel Pro Analyzer 4.0 de MediaCybernetics, Inc.
3.7 Citometría de Flujo
Para determinar la expresión de TLR4 en la superficie de células A549
estimuladas con TNFα y Dexametasona, se utilizaron 5x105 células en placas de 6
15

cm. Las cuales fueron estimuladas durante 24 horas y posteriormente recuperadas
con una solución de EDTA 25 mM en PBS y recolectadas con PBS.
Las células se lavaron 2 veces con una solución de PBS y luego se incubaron
por 30 minutos con PBS conteniendo 10% suero de ratón (PBS/SR), con el propósito
de bloquear los sitios de unión inespecífica. Luego, las células se lavaron con PBS/SR
y se agregó el anticuerpo anti-TLR4-PE también diluido en PBS/SR, usando 40 µl por
cada 106 células y se incubó en oscuridad a 4ºC por una hora. Finalizada la
incubación, las células se lavaron dos veces con la solución de PBS. Luego de lavar,
las células se fijaron en p-formaldehído al 2% diluido en PBS.
Las muestras se adquirieron en un citómetro FACSort (Becton Dickinson) y se
analizaron usando el programa WinMD versión 2.9. Los análisis se realizaron con al
menos 10.000 eventos a partir de la población total y se determinó el porcentaje de
células que expresan TLR4 dentro de esta población. Todos lo datos fueron corregidos
por los controles de autofluorescencia y por la unión inespecífica de anticuerpos de
isotipo adecuados.
3.8 Estadística
Todos los pares de grupos tratados con ambos estímulos, en conjunto y por
separado, como también las células transfectadas con los distintos constructos fueron
comparados con Mann-Whitney test. Al menos tres experimentos independientes
fueron realizados y las muestras fueron significativas cuando *, p < 0,05. Los análisis
fueron realizados con el programa GraphPad Prism 4 (GraphPad Software, Inc).
16

4. RESULTADOS
4.1 Los glucocorticoides revierten la inducción de la expresión del mRNA de TLR4 inducido por TNFα
El efecto de TNFα en presencia o ausencia de Dexametasona sobre la
expresión del mRNA de TLR4 en células A549, se evaluó a través de PCR en tiempo
real. El sistema de detección utilizado comprende la medición de la fluorescencia
emitida por un fluoróforo que se intercala en el DNA de doble hebra, SYBR Green I,
que emite a una longitud de onda de 520 nm (42).
Para la estandarización de la PCR en tiempo real para TLR4 se confeccionó
una curva estándar, utilizando diluciones seriadas del plasmidio pCMV-Flag que
contiene la secuencia codificante para TLR4 (pCMV-Flag-hTLR4), entre 10 ng y 1 fg. A
partir del ensayo de PCR se obtuvo la ecuación y pendiente en base al Ct o ciclo
umbral de cada reacción (Figura 1). El Ct corresponde al punto de intersección de la
curva de amplificación de cada producto con la línea base de detección que fija el
propio programa, detectando con ello el ciclo donde se inicia el aumento significativo
de la fluorescencia, siendo esto inversamente proporcional a la cantidad de ADN inicial
de tenía cada muestra (43).
17

a) b)
y = -0,2944x + 5,7936R2 = 0,9956
0
1
2
3
4
5
6
3 6 9 12 15 18 2
C(t) Cycle
Log
Qua
ntity
1
Figura 1. Curva estándar de PCR en Tiempo Real para TLR4 y diagrama del plasmidio pCMV-Flag-hTLR4. (a) Curva estándar y ecuación de la recta para TLR4, donde se interpolaron los Ct de los tratamientos con los diferentes estímulos, determinando la concentración del mRNA de TLR4. (b) Estructura del plasmidio pCMV-Flag-hTLR4, que posee la secuencia codificante para TLR4, y fue el que se utilizó para realizar la curva estándar.
Para confeccionar la curva estándar de G3DP en PCR en tiempo real, se
realizó primero un PCR semicuantitativo con los partidores específicos y el producto
amplificado se corrió en un gel de agarosa. La banda del producto amplificado se
cuantificó por análisis densitométrico, y se determinó su concentración con respecto al
estándar de peso molecular y así obtener la curva estándar de G3DP (Figura 2).
Interpolando en ella los Ct de las diferentes condiciones, es posible determinar la
expresión del mRNA de G3DP y así normalizar los valores de expresión de TLR4 con
respecto a este gen.
18

y = -0,3288x + 8,5161R2 = 0,9993
01234567
8 11 14 17 20 23 26
C(t) Cycle
Log
Qua
ntity
Figura 2. Curva estándar de PCR en Tiempo Real para G3DP. Curva estándar y ecuación de la recta para G3DP, obtenida a través de PCR en tiempo real.
Una vez definidas las condiciones de trabajo para el PCR en tiempo real, se
determinó el efecto de TNFα y Dexametasona sobre la expresión del mRNA de TLR4.
Para ello, las células A549 fueron estimuladas con TNFα (10 ng/ml) en presencia o
ausencia de Dexametasona (100 nM) por 6 hrs., para luego extraer el RNA total de las
células, el cual se trató con DNasa, se cuantificó y se obtuvo el cDNA mediante la
reacción de transcriptasa reversa (RT). EL cDNA, se utilizó para la PCR en tiempo real
amplificando el TLR4 con los partidores específicos.
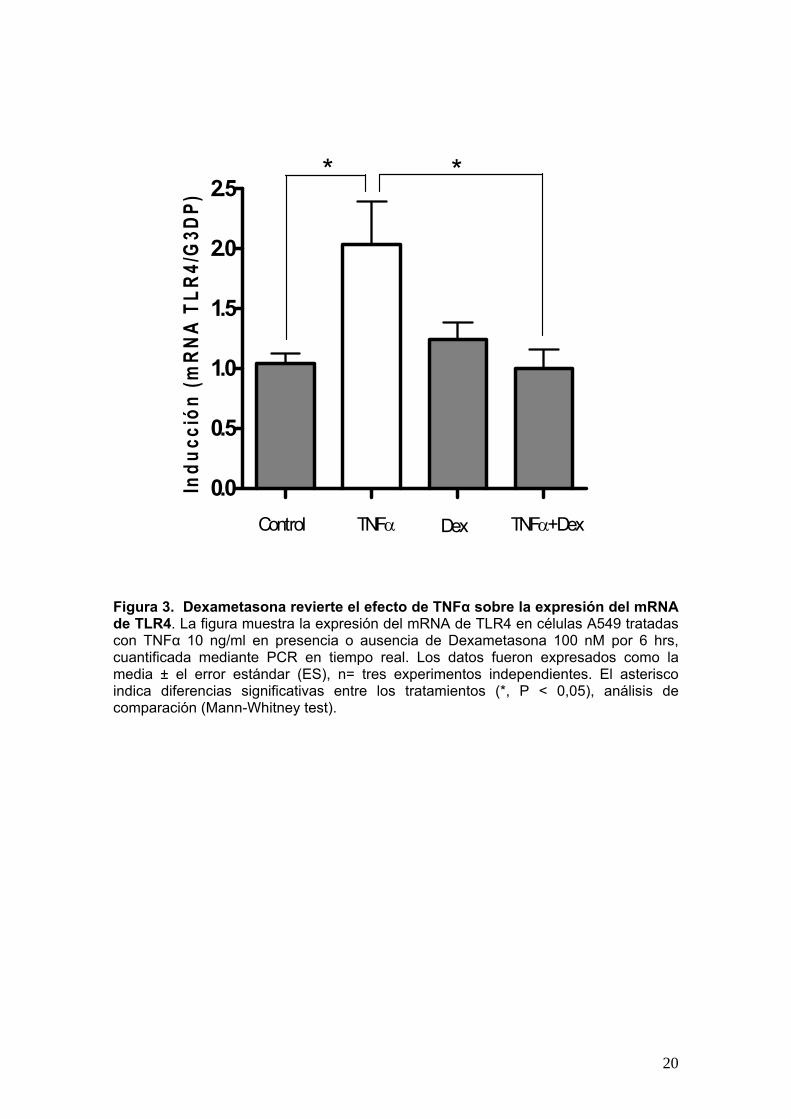
En la Figura 3 se observa que el TNFα aumentó significativamente la expresión
del mRNA de TLR4, con respecto al control. El estímulo con Dexametasona no
modificó la expresión de TLR4 con respecto al control. En cuanto a la co-estimulación
con ambas moléculas, Dexametasona revirtió significativamente el efecto de TNFα,
aportando una primera evidencia relacionada al efecto de los GCs sobre la expresión
del mRNA de TLR4 inducido por TNFα.
19
0.0
0.5
1.0
1.5
2.0
2.5
TNFαControl Dex TNFα+Dex
Indu
cció
n (m
RN
A T
LR4/
G3D
P)* *
Figura 3. Dexametasona revierte el efecto de TNFα sobre la expresión del mRNA de TLR4. La figura muestra la expresión del mRNA de TLR4 en células A549 tratadas con TNFα 10 ng/ml en presencia o ausencia de Dexametasona 100 nM por 6 hrs, cuantificada mediante PCR en tiempo real. Los datos fueron expresados como la media ± el error estándar (ES), n= tres experimentos independientes. El asterisco indica diferencias significativas entre los tratamientos (*, P < 0,05), análisis de comparación (Mann-Whitney test).
20

4.2 Los glucocorticoides modulan el efecto de TNFα sobre la actividad transcripcional del promotor del TLR4
4.2.1 Análisis bioinformático de los elementos de respuesta
putativos del promotor de TLR4 murino
Se diseñó un diagrama del promotor de TLR4 murino con los sitios de unión de
los diferentes factores de transcripción potencialmente involucrados en las rutas de
señalización de los estímulos utilizados en esta memoria, tales como AP-1, NFκB,
STAT1 y GR (Figura 4) (44; 45). Para identificar la posición de los sitios putativos de
interés en el promotor de TLR4, se usaron los programas computacionales Genomatix
y TFsearch, que aplican algoritmos determinados, según la base de datos que poseen
para los diferentes motivos consenso de los sitios de unión al promotor por parte de
los factores de transcripción.
Figura 4. Promotor de TLR4 murino El diagrama muestra los sitios de unión putativos para factores de transcripción potencialmente involucrados en las rutas de señalización de TNFα y Dexametasona. El número que aparece sobre cada sitio de unión indica la posición que ocupan en el promotor, río arriba con respecto al inicio de la transcripción (+1); Luc = Luciferasa. Se indican además, las diferentes deleciones de la construcción del promotor de TLR4 con el tamaño correspondiente en el extremo 5`.
21

4.2.2 Glucocorticoides contrarrestan el efecto de TNFα sobre
la actividad del promotor de TLR4 Los factores de transcripción, tales como NFκB, AP-1 e integrantes de la familia
STAT (46), forman parte de la ruta de señalización de TNFα en la regulación de la
transcripción de genes inflamatorios (47).
En este trabajo se evaluó el efecto de los GCs sobre la actividad transcripcional
del promotor de TLR4. Para ello se utilizaron distintas construcciones del promotor de
TLR4-Luc las que se transfectaron junto con el plasmidio control Renilla (pGL3-hRL)
en células A549, y se determinó posteriormente la expresión de los genes reporteros
luciferasa y renilla, siendo el primero un indicador de la activación transcripcional del
promotor de TLR4. En todos los experimentos se utilizaron las construcciones,
3xMHC-Luc y MMTV-Luc como controles para la respuesta a TNFα y Dexametasona,
respectivamente.
En la Figura 5 se muestra que TNFα induce la expresión de luciferasa de dos
de las deleciones del promotor de TLR4 (pGL3-2715 y pGL3-1864), mientras que
Dexametasona por si sola no la modificó. El co-tratamiento de Dexametasona con
TNFα revirtió la inducción de la expresión del gen reportero inducida por TNFα. Al
evaluar la expresión del reportero luciferasa en las deleciones del promotor pGL3-1183
y pGL3-336, TNFα no la modificó, tanto en presencia o en ausencia de
Dexametasona. Sin embargo, la actividad transcripcional de la deleción del promotor
de TLR4, pGL3-144, fue significativamente inducida con Dexametasona. En el inserto
de la Figura 5 se indican la activación de los plasmidios controles, 3xMHC-Luc y
MMTV-Luc, lo que sugiere que las células responden a los estímulos en estudio. Estos
resultados muestran que la regulación de la activación transcripcional de TLR4 por los
GCs involucra varios GREs descritos en el modelo teórico (Figura 4), sin embargo su
efecto activador o represor aun no se ha descrito (48).
22

2715 1864 1183 336 1440
2
4
ControlTNFαDEXTNFα+DEX
8
10
12
14
* *
*
* *
Deleciones de la Construcción del promotor de TLR4
Indu
cció
n R
elat
iva
de la
Act
ivid
ad L
ucife
rasa
p4-
Luc
CONTROL TNFα CONTROL DEX0
102030405060708090
3XMHC MMTV
Indu
cció
n R
elat
iva
de la
Act
ivid
ad L
ucife
rasa
TLR
Figura 5. Actividad transcripcional del promotor de TLR4. Las células A549 fueron co-transfectadas con las distintas construcciones del promotor de TLR4-Luc y con pGL3-hRL. Las células fueron tratadas por 24 horas con 10 ng/ml de TNFα, 100 nM de Dexametasona o la combinación de ambos, determinándose posteriormente la actividad de luciferasa utilizando un sustrato quimioluminiscente. Se consideraron aquellos experimentos donde se observó la inducción de la expresión de luciferasa de los vectores 3xMHC-Luc y MMTV-Luc para TNFα y Dexametasona, respectivamente (inserto). Las barras representan la razón entre las unidades de luciferasa (RLU) y las unidades de renilla (RRU) normalizadas al respectivo control. Todas las muestras fueron analizadas en duplicado y los valores significan ± S.E. de 3 experimentos. El asterisco indica diferencias significativas entre los tratamientos (*, P < 0,05), por análisis de comparación de pares (Mann-Whitney test) para cada condición control.
23

4.3. Los glucocorticoides revierten el aumento de la expresión de TLR4 a nivel proteico inducida por TNFα 4.3.1. Dexametasona antagoniza la inducción de TLR4 a nivel
de proteína total mediada por TNFα El paso siguiente fue evaluar la expresión de TLR4 a nivel de proteína total,
verificando si los cambios inducidos por TNFα y GC obedecen a lo observado a nivel
genómico.
La determinación de los cambios en la expresión de la proteína de TLR4 en
respuesta a los estímulos de interés se realizó mediante inmunoblot de lisados
celulares. Primero, se evaluó el efecto de TNFα (10 ng/ml) a diferentes tiempos (0, 6,
18 y 24 hrs.), observándose una mayor expresión a las 6 hrs. post-estimulo (Figura 6),
como una banda inmunoreactiva de 105 KDa que corresponde al receptor anclado a
membrana, y las otras de alrededor 75 y 50 kDa (TLR4s1 y TLR4s2) a isoformas
solubles de TLR4. Estos resultados reflejan una clara dependencia de la inducción de
la expresión de todas las isoformas del receptor con el tiempo de exposición a TNFα.
Luego se determinó el efecto de Dexametasona (100 nM) a los mismos tiempos (0, 6,
12 y 18 hrs.), no observándose variaciones significativas (Figura 7). El tratamiento de
las células con ambos estímulos, TNFα y Dexametasona por 6 hrs., indica que esta
última revierte el efecto inductor de TNFα sobre la expresión de TLR4 (Figura 8).
24

Figura 6. Curso temporal de la expresión de TLR4 inducida por TNFα. Las células fueron tratadas con TNFα (10 ng/ml) por 6, 18 y 24 hrs. y posteriormente lisadas. La figura muestra tres bandas inmunorreactivas con diferentes pesos moleculares: 105, 75 y 50 KDa, correspondiente al receptor anclado a la membrana (TLR4), y las formas solubles del receptor (TLR4s1 y TLR4s2), respectivamente, cuya expresión es dependiente del tiempo de exposición a TNFα. La inmunoreactividad de la isoforma TLR4s2 no se observa en lisados de células expuestas a la citoquina por 18-24 hrs. Los valores (*) representan la razón de inducción con respecto al control, el cual se realizó por análisis densitométrico de las bandas normalizadas a la β-actina en relación a la banda de TLR4 (105 KDa).
Figura 7. Curso temporal de la expresión de TLR4 total en células expuestas a Dexametasona. Las células fueron tratadas con Dexametasona (100 nM) por 6, 12 y 18 hrs. La figura muestra dos bandas inmunorreactivas de 105 y 75 KDa, correspondientes al receptor anclado a la membrana (TLR4) y la forma soluble (TLR4s1). La razón de inducción (*) con respecto al control se realizó por análisis densitométrico de las bandas normalizadas a la β-actina en relación a la banda de 105 KDa.
25

TLR4
TLR4s1
TLR4s2
β-actina
TNFα (10 ng/ml)
Dex (100 nM)
(*) 1 1,6 1,1 0,9
-- -
-++
++
Figura 8. Los glucocorticoides revierten el efecto inductor de TNFα sobre la expresión de la proteína de TLR4. La figura muestra la inmunoreactividad para TLR4 en células expuestas a TNFα (10 ng/ml), Dexametasona (100 nM) y el co-tratamientos de ambos por 6 hrs. La razón de inducción (*) con respecto al control se realizó por análisis densitométrico de las bandas normalizadas a la β-actina en relación a la banda inmunoreactiva de TLR4.
4.3.2 Efecto de glucocorticoides sobre la expresión del TLR4 a
nivel de superficie inducida por TNFα
La expresión de TLR4 en la membrana plasmática de las células A549, se
realizó mediante citometría de flujo. La expresión del receptor en este modelo celular
es menor con respecto a otros tipos celulares del sistema inmune innato, como los
macrófagos, sin embargo este estudio apoyará en la compresión de los mecanismos
involucrados en la regulación de la expresión de TLR4.
26

Las células se trataron con TNFα (10 ng/ml) en presencia o ausencia de
Dexametasona (100 nM) durante 24 horas. Los resultados obtenidos muestran que
TNFα induce la expresión de TLR4 a nivel de superficie, efecto que también es
revertido por Dexametasona. Sin embargo, esta última por sí sola no modificó la
expresión de TLR4 (Figura 9). Estos datos indican que los estímulos estudiados
modifican la expresión de la proteína de TLR4 a nivel de superficie como un
mecanismo de alerta frente en cuadros inflamatorios.
Control TNFα Dex TNFα + Dex
0
5
10
15
* *
% c
élul
as T
LR4+
Figura 9. Expresión de TLR4 en la membrana plasmática de células epiteliales humanas. La figura muestra el porcentaje de células positivas para TLR4 en función del tratamiento con TNFα (10ng/ml) en presencia o ausencia de Dexametasona (100 nM) por 24 horas. Los datos fueron expresados como la media ± el error estándar (ES), n= cuatro experimentos independientes. El asterisco indica diferencias significativas entre los tratamientos (*, P < 0,05), análisis de comparación (Mann-Whitney test).
27

5. DISCUSIÓN
El papel que juegan los Receptores Tipo Toll en el sistema inmune innato es
crucial, ya que censan la presencia de PAMPs específicos, desencadenando con esto
la activación de señales intracelulares que se traducen finalmente en la expresión de
moléculas vinculadas a procesos pro-inflamatorios (49).
Se han realizado numerosos estudios sobre la afinidad y especificidad de los
TLRs por sus ligandos (50), como también sobre la regulación de su expresión por
moléculas de diverso origen y en diferentes tipos celulares (19).
En esta memoria se propuso dilucidar el efecto de moléculas que ejercen
respuestas antagónicas en el sistema inmune, sobre la expresión de uno de estos
TLRs en particular, el TLR4. Por una parte, se estudió el efecto de una citoquina de
gran relevancia en procesos inflamatorios, como es TNFα y por otro lado los
glucocorticoides, que poseen propiedades anti-inflamatorias.
5.1 Efecto de TNFα y Dexametasona sobre la expresión del mRNA de TLR4 en células A549
Inicialmente se estudió la expresión del gen de tlr4, mediante PCR en tiempo
real, que permite cuantificar el mRNA sintetizado, donde TNFα indujo un aumento
significativo en células A549 de la expresión del mRNA de TLR4. Dexametasona por
si solo no afectó la expresión del mRNA de TLR4, sin embargo revirtió
significativamente el efecto de TNFα, lo que demuestra el gran poder que poseen los
glucocorticoides en este caso, de contrarrestar casi por completo el efecto que tiene
TNFα sobre la síntesis del mRNA de TLR4, lo que finalmente se traduce en una menor
28

presencia de este receptor para censar a su ligando específico y con ello no
desencadenar la expresión de otras moléculas pro-inflamatorias que exacerbarían este
cuadro.
Además de este importante resultado, el haber podido detectar mediante una
técnica sensible, los diferentes comportamientos de expresión génica que producen
moléculas antagónicas en células epiteliales pulmonares, es también la primera
evidencia que muestra el efecto de los glucocorticoides sobre la expresión de TLR4,
inducido por un estímulo inflamatorio, y que ayudará posteriormente a entender mejor
el comportamiento del sistema inmune innato, en relación a los patrones de expresión
de estos tipos de receptores.
5.2 Efecto de TNFα sobre la actividad transcripcional del promotor del TLR4 en presencia o ausencia de Dexametasona
El papel que juegan los distintos factores transcripcionales sobre la expresión
genética es crucial para la sobrevida celular (51), y en este caso en particular para el
gen de tlr4, depende de los elementos de respuesta que posea en su región promotora
(52).
La actividad transcripcional del promotor de TLR4 murino, que posee una alta
homología al humano (con una identidad del 85%), se analizó en diferentes deleciones
mutantes del mismo.
Los resultados obtenidos muestran la misma tendencia a nivel del transcrito, en
el que Dexametasona revierte el efecto de TNFα; este resultado fue observado
exclusivamente para el constructo del promotor completo y la siguiente en tamaño, las
cuales poseen sitios de unión putativos para AP-1 y GR (GRE). El sitio de unión para
29

AP-1 tendría mayor relevancia en la inducción de la transcripción del gen tlr4 por TNFα
y el GRE podría ser responsable del efecto inhibidor de Dexametasona sobre TNFα.
Las deleciones mutantes del promotor de TLR4 de menor tamaño (-336 y -144)
confirman que los efectos observados a nivel de mRNA y actividad transcripcional del
constructo completo dependen de, al menos, algunos de estos factores de
transcripción. Además, el análisis en silico del promotor con los programas descritos
identificó la presencia de tres mitades de GRE (mGRE) en las posiciones -269, -235 y
-41 río arriba con respecto al inicio de la transcripción. Estos mGRE están
conformados por las bases más conservadas, información que ayudaría a comprender
el nivel de actividad transcripcional de estas últimas deleciones. Dependiendo de la
cantidad y tipo (activador o represor) de GREs que posea un determinado promotor,
van a ejercer un patrón de expresión génica característico. Además el GR puede
ejercer su efecto inhibidor a través de la interacción proteína-proteína con otros
factores de transcripción (53).
5.3 Efecto de los glucocorticoides sobre la expresión de la proteína de TLR4 inducida por TNFα
Para detectar la presencia de la proteína se realizaron inmunoblots y citometría
de flujo, identificando con esto la proteína total y la asociada a membrana,
respectivamente. Las células A549 fueron estimuladas con TNFα y Dexametasona,
determinándose el efecto de cada uno de los estímulos sobre la expresión de TLR4 en
el tiempo. La máxima inducción de TLR4 por TNFα se observó a las 6 horas, tiempo
en el cual Dexametasona revirtió su efecto. Corroborándose el mismo patrón de
respuesta a nivel del TLR4 de superficie por citometría de flujo pero a las 24 horas.
30

5.4 Regulación de la expresión de TLR4 por glucocorticoides inducido por TNFα
En este trabajo se evidenció claramente el efecto de moléculas que ejercen
efectos antagónicos, tales como TNFα y Dexametasona, sobre la expresión del TLR4
(Figura 10). La figura muestra que la unión de TNFα a su receptor de membrana
activa factores transcripcionales, tales como NFκB, AP-1 y STAT1, que conducen a un
aumento en la expresión de TLR4. Por otro lado, la Dexametasona, un GC sintético
que por su naturaleza lipídica permea la membrana y se une a su receptor
citoplasmático el cual transloca al núcleo el cual revierte el efecto inducido por TNFα
sobre la expresión de TLR4, ya sea por interacción directa con GRE presentes en el
promotor del gen o por inhibición de AP-1 o NFκB mediante interacción proteína-
proteína. Estos resultados evidencian el papel de factores de transcripción activados
por vías pro y anti inflamatorias sobre la actividad transcripcional de TLR4 que
conducen a cambios en la expresión proteica en estados inflamatorios o infecciosos.
Los estudios que se proyectan de este trabajo están focalizados a dilucidar el
mecanismo molecular que estarían ejerciendo los glucocorticoides sobre la expresión
del TLR4, los que muy probablemente estén ocurriendo a nivel genómico. La
identificación de la secuencia de los GRE en el promotor del TLR4, ha sido un valioso
aporte para profundizar en el tipo de respuesta que puedan evocar diferentes tipos de
elementos de respuesta.
31

Figura 10. Modelo de la regulación de la expresión de TLR4 por glucocorticoides inducida por TNFα. La figura representa a modo general los factores que estarían involucrados en la regulación de la expresión del TLR4 por TNFα y glucocorticoides en células epiteliales de vías respiratorias. Las moléculas antagónicas TNFα y glucocorticoides son reconocidas por los respectivos receptores, uno localizado en la membrana plasmática y el otro a nivel citoplasmático, respectivamente, para posteriormente desencadenar la translocación de factores de trascripción específicos de cada vía los cuales regularían directamente la expresión génica a través de la regulación transcripcional de TLR4.
32

6. CONCLUSIONES
En este trabajo se demostró que TNFα incrementa la expresión de TLR4, tanto
a nivel del transcrito, de la proteína y de la actividad transcripcional del promotor en
células epiteliales A549. Por primera vez, este trabajo demuestra que los
glucocorticoides revierten el efecto de TNFα sobre la expresión de TLR4.
Los resultados obtenidos a nivel del transcrito y proteína de TLR4 evidencian el
gran efecto inhibidor de Dexametasona sobre TNFα y, los ensayos de actividad
transcripcional indican los sitios de unión y/o factores de transcripción presentes en el
promotor de TLR4 que tendrían un papel relevante en la respuesta antagónica de
estos compuestos.
Finalmente, los hallazgos presentados en este trabajo demuestran el poderoso
efecto que estarían ejerciendo los GCs, sobre la expresión de TLR4, uno de los más
importantes y estudiados miembros de la familia de los TLRs.
Queda aún por dilucidar los mecanismos moleculares específicos por los
cuales estas moléculas regularían la expresión de TLR4, para comprender más
cabalmente el efecto de estas moléculas sobre algunos integrantes de la inmunidad
innata.
33

7. REFERENCIAS
1. Delves, P. J., and Roitt I. M. 2000. The immune system. First of two parts. The New England journal of Medicine 343:37-49.
2. Medzhitov, R., and Janeway, C. Jr. 2000. Innate immunity. The New England
Journal of Medicine 343:338-344. 3. Medzhitov, R., and Janeway, C. A. Jr. 1997. Innate immunity: impact on the
adaptive immune response. Current opinion in Immunology 9:4-9. 4. Delves, P. J., and Roitt I. M.. 2000. The immune system. Second of two parts.
The New England Journal of Medicine 343:108-117. 5. Itano, A. A., and Jenkins M. K. 2003. Antigen presentation to naive CD4 T cells
in the lymph node. Nature immunology 4:733-739. 6. Allan, R. S., Waithman J., Bedoui S., Jones C. M., Villadangos J. A., Zhan Y.,
Lew A. M., Shortman K., Heath W. R., and Carbone F. R.. 2006. Migratory dendritic cells transfer antigen to a lymph node-resident dendritic cell population for efficient CTL priming. Immunity 25:153-162.
7. Duez, C., Gosset P., and Tonnel A. B. 2006. Dendritic cells and toll-like
receptors in allergy and asthma. Eur Journal Dermatology 16:12-16. 8. Hashimoto, C., Hudson K. L., and Anderson K. V.. 1988. The Toll gene of
Drosophila, required for dorsal-ventral embryonic polarity, appears to encode a transmembrane protein. Cell 52:269-279.
9. West, A. P., Koblansky A. A., and Ghosh S.. 2006. Recognition and signaling
by toll-like receptors. Annual Review of Cell and Developmental Biology 22:409-437.
10. MacLeod, H., and Wetzler L. M.. 2007. T cell activation by TLRs: a role for
TLRs in the adaptive immune response. Sci STKE 2007:pe48. 11. Takeda, K., and Akira S.. 2004. TLR signaling pathways. Seminars in
Immunology 16:3-9. 12. Miggin, S. M., and O'Neill L. A.. 2006. New insights into the regulation of TLR
signaling. Journal of Leukocyte Biology 80:220-226. 13. Van Amersfoort, E. S., Van Berkel T. J., and Kuiper J.. 2003. Receptors,
mediators, and mechanisms involved in bacterial sepsis and septic shock. Clinical Microbiology Reviews 16:379-414.
14. Poltorak, A., He X., Smirnova I., Liu M. Y., Van Huffel C., Du X., Birdwell D.,
Alejos E., Silva M., Galanos C., Freudenberg M., Ricciardi-Castagnoli P., Layton B., and Beutler B.. 1998. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science (New York, N.Y 282:2085-2088.
34

15. Abreu, M. T., Arnold E. T., Thomas L. S., Gonsky R., Zhou Y., Hu B., and Arditi
M.. 2002. TLR4 and MD-2 expression is regulated by immune-mediated signals in human intestinal epithelial cells. The Journal of Biological Chemistry 277:20431-20437.
16. Akira, S. 2003. Toll-like receptor signaling. The Journal of Biological Chemistry
278:38105-38108. 17. Nunez Miguel, R., Wong J., Westoll J. F., Brooks H. J., O'Neill L. A., Gay N. J.,
Bryant C. E., and Monie T. P.. 2007. A dimer of the Toll-like receptor 4 cytoplasmic domain provides a specific scaffold for the recruitment of signalling adaptor proteins. PLoS ONE 2:e788.
18. Dunne, A., and O'Neill L. A.. 2003. The interleukin-1 receptor/Toll-like receptor
superfamily: signal transduction during inflammation and host defense. Sci STKE 2003:re3.
19. Gutierrez-Canas, I., Juarranz Y., Santiago B., Arranz A., Martinez C., Galindo
M., Paya M., Gomariz R. P., and Pablos J. L.. 2006. VIP down-regulates TLR4 expression and TLR4-mediated chemokine production in human rheumatoid synovial fibroblasts. Rheumatology (Oxford, England) 45:527-532.
20. Brann, D. W., Hendry L. B., and Mahesh V. B.. 1995. Emerging diversities in
the mechanism of action of steroid hormones. The Journal of Steroid Biochemistry and Molecular Biology 52:113-133.
21. Hermoso, M. A., and Cidlowski J. A.. 2003. Putting the brake on inflammatory
responses: the role of glucocorticoids. IUBMB life 55:497-504. 22. Breuner, C. W., and Orchinik M.. 2002. Plasma binding proteins as mediators of
corticosteroid action in vertebrates. The Journal of Endocrinology 175:99-112. 23. Munck, A., Guyre P. M., and Holbrook N. J.. 1984. Physiological functions of
glucocorticoids in stress and their relation to pharmacological actions. Endocrine Reviews 5:25-44.
24. Glezer, I., and Rivest S.. 2004. Glucocorticoids: protectors of the brain during
innate immune responses. Neuroscientist 10:538-552. 25. Reichardt, H. M., Kaestner K. H., Tuckermann J., Kretz O., Wessely O., Bock
R., Gass P., Schmid W., Herrlich P., Angel P., and Schutz G.. 1998. DNA binding of the glucocorticoid receptor is not essential for survival. Cell 93:531-541.
26. Barnes, P. J. 2006. Corticosteroid effects on cell signalling. Eur Respiratory
Journal 27:413-426. 27. Smith, D. F., and Toft D. O.. 1993. Steroid receptors and their associated
proteins. Molecular Endocrinology 7:4-11. 28. Clark, A. R. 2007. Anti-inflammatory functions of glucocorticoid-induced genes.
Molecular and Cellular Endocrinology 275:79-97.
35

29. Galon, J., Franchimont D., Hiroi N., Frey G., Boettner A., Ehrhart-Bornstein M., O'Shea J. J., Chrousos G. P., and Bornstein. S. R. 2002. Gene profiling reveals unknown enhancing and suppressive actions of glucocorticoids on immune cells. Faseb Journal 16:61-71.
30. Rhen, T., and Cidlowski J. A.. 2005. Antiinflammatory action of glucocorticoids--
new mechanisms for old drugs. The New England Journal of Medicine 353:1711-1723.
31. Hermoso, M. A., Matsuguchi T., Smoak K., and Cidlowski J. A.. 2004.
Glucocorticoids and tumor necrosis factor alpha cooperatively regulate toll-like receptor 2 gene expression. Molecular and Cellular Biology 24:4743-4756.
32. Nissen, R. M., and Yamamoto K. R.. 2000. The glucocorticoid receptor inhibits
NFkappaB by interfering with serine-2 phosphorylation of the RNA polymerase II carboxy-terminal domain. Genes & Development 14:2314-2329.
33. Holmes, C. L., Russell J. A., and Walley K. R.. 2003. Genetic polymorphisms in
sepsis and septic shock: role in prognosis and potential for therapy. Chest 124:1103-1115.
34. Silverstein, R., and Johnson D. C.. 2003. Endogenous versus exogenous
glucocorticoid responses to experimental bacterial sepsis. Journal of Leukocyte Biology 73:417-427.
35. Basu, S., and Fenton M. J.. 2004. Toll-like receptors: function and roles in lung
disease. American Journal of Physiology 286:L887-892. 36. Muzio, M., Natoli G., Saccani S., Levrero M., and Mantovani A.. 1998. The
human toll signaling pathway: divergence of nuclear factor kappaB and JNK/SAPK activation upstream of tumor necrosis factor receptor-associated factor 6 (TRAF6). The Journal of Experimental Medicine 187:2097-2101.
37. Roger, T., David J., Glauser M. P., and Calandra T.. 2001. MIF regulates innate
immune responses through modulation of Toll-like receptor 4. Nature 414:920-924.
38. Ostrowski, M. C., Richard-Foy H., Wolford R. G., Berard D. S., and Hager G. L..
1983. Glucocorticoid regulation of transcription at an amplified, episomal promoter. Molecular and Cellular Biology 3:2045-2057.
39. Yudt, M. R., Jewell C. M., Bienstock R. J., and Cidlowski J. A.. 2003. Molecular
origins for the dominant negative function of human glucocorticoid receptor beta. Molecular and Cellular Biology 23:4319-4330.
40. Valasek, M. A., and Repa J. J.. 2005. The power of real-time PCR. Advances in
Physiology Education 29:151-159. 41. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 227:680-685. 42. Lekanne Deprez, R. H., Fijnvandraat A. C., Ruijter J. M., and Moorman A. F..
2002. Sensitivity and accuracy of quantitative real-time polymerase chain reaction using SYBR green I depends on cDNA synthesis conditions. Analytical Biochemistry 307:63-69.
36

43. Costa, J. 2004. [Real-time PCR]. Enfermedades Infecciosas y Microbiologia
Clinica 22:299-304. 44. Rogatsky, I., and Ivashkiv L. B.. 2006. Glucocorticoid modulation of cytokine
signaling. Tissue Antigens 68:1-12. 45. Wesemann, D. R., and Benveniste E. N.. 2003. STAT-1 alpha and IFN-gamma
as modulators of TNF-alpha signaling in macrophages: regulation and functional implications of the TNF receptor 1:STAT-1 alpha complex. Journal Immunology 171:5313-5319.
46. Reich, N. C., and Liu L.. 2006. Tracking STAT nuclear traffic. Nature Reviews
6:602-612. 47. Barrett, E. G., Johnston C., Oberdorster G., and Finkelstein J. N.. 1998. Silica-
induced chemokine expression in alveolar type II cells is mediated by TNF-alpha. The American Journal of Physiology 275:L1110-1119.
48. Dostert, A., and Heinzel T.. 2004. Negative glucocorticoid receptor response
elements and their role in glucocorticoid action. Current Pharmaceutical Design 10:2807-2816.
49. Trinchieri, G., and Sher A.. 2007. Cooperation of Toll-like receptor signals in
innate immune defence. Nature Reviews 7:179-190. 50. Beutler, B. 2004. Inferences, questions and possibilities in Toll-like receptor
signalling. Nature 430:257-263. 51. Brivanlou, A. H., and Darnell, Jr J. E.. 2002. Signal transduction and the control
of gene expression. Science 295:813-818. 52. Pedchenko, T. V., Park G. Y., Joo M., Blackwell T. S., and Christman J. W..
2005. Inducible binding of PU.1 and interacting proteins to the Toll-like receptor 4 promoter during endotoxemia. American Journal of Physiology 289:L429-437.
53. Karin, M. 1998. New twists in gene regulation by glucocorticoid receptor: is
DNA binding dispensable? Cell 93:487-490.
37

8. ANEXO
Como parte del trabajo realizado durante esta memoria de título, se generaron
las siguientes presentaciones:
Trabajos presentados en congresos nacionales
Regulación negativa endógena de la Respuesta Inmune Innata mediada por los Toll-like Receptors (TLRs): Papel de los glucocorticoides. Frano Malinarich, Rodrigo Valenzuela, Ramón Pérez, Lucia Núñez, Sofía Sepúlveda,
Enzo Candia, Rodrigo Quera y Marcela A. Hermoso.
Presentación ICBM, Facultad de Medicina, Universidad de Chile.
Santiago, Chile, Septiembre 2007.
Regulación de la Expresión del Receptor Tipo Toll 4 (TLR4) por el Factor de Necrosis Tumoral α y Dexametasona Ramón Pérez, Rodrigo Valenzuela, Enzo Candia y Marcela Hermoso.
XXI Reunión Anual de la Sociedad de Biología Celular de Chile.
Pucón, Chile, Octubre 2007.
Trabajos presentados en congresos internacionales Regulation of The Toll-Like Receptor 4 expression by Tumor Necrosis Factor α and Dexamethasone.
R. D. Pérez, R. Valenzuela, A. Paredes, M. A. Hermoso.
Congreso Internacional de Inmunología
Rio de Janeiro, Brasil, Agosto 2007
38






![EL MÉTODO DE LAS ὑποθέσεις EN PHÆDOrepositorio.uchile.cl/tesis/uchile/2005/caro_s/sources/caro_s.pdf · que, para Platón, la ἰδέα es el carácter de todas las [cosas]](https://static.fdocument.org/doc/165x107/5be1c3da09d3f2f02d8b4c38/el-metodo-de-las-en-pha-que-para-platon-la-.jpg)



