
PARTICIPACIÓN DEL FACTOR DE NECROSIS...
54
UNIVERSIDAD DE CHILE Facultad de Ciencias Químicas y Farmacéuticas Departamento de Bioquímica y Biología Molecular PARTICIPACIÓN DEL FACTOR DE NECROSIS TUMORAL ALFA DE TRANSMEMBRANA (mTNF-α) EN LA LIPOGÉNESIS Memoria para optar al Título profesional de Bioquímico. Por PAULA LORETO FIGUEROA MOYA Profesor guía: Dra. Mireya Fernández Gálvez. Laboratorio de Biología Celular, INTA, Universidad de Chile. Profesor Patrocinante: Dr. Javier Puente P. Santiago, Chile 2007
Transcript of PARTICIPACIÓN DEL FACTOR DE NECROSIS...

UNIVERSIDAD DE CHILE
Facultad de Ciencias Químicas y Farmacéuticas Departamento de Bioquímica y Biología Molecular
PARTICIPACIÓN DEL FACTOR DE NECROSIS TUMORAL ALFA D E
TRANSMEMBRANA (mTNF- α) EN LA LIPOGÉNESIS
Memoria para optar al Título profesional de
Bioquímico.
Por
PAULA LORETO FIGUEROA MOYA
Profesor guía: Dra. Mireya Fernández Gálvez.
Laboratorio de Biología Celular, INTA,
Universidad de Chile.
Profesor Patrocinante: Dr. Javier Puente P.
Santiago, Chile
2007

Dedico esta tesis a mi hija Ignacia, a mi esposo Carlos,
a mis hermanos Daniela, María Paz y Alfredo, y a mis Padres.

AGRADECIMIENTOS
Agradezco a mis directoras de tesis, Dra. Mireya Fernández y Dra. María Pía
De la Maza, por recibirme en el laboratorio, confiar en mí y apoyarme en los buenos y
en los malos momentos. A todas las personas que conforman el laboratorio de Biología
Celular, especialmente a María José, por enseñarme todo lo que sabía.
A mi esposo Carlos, por su apoyo incondicional, a mi hija Ignacia por alentarme
cada día, a toda mi familia (Pablo, Mónica, Daniela, María Paz, Alfredo, Abuelita
Mónica y Rosita) por su permanente preocupación y cariño.
A Abuelita Adriana y Tío Andrés, que aunque ya no estén presentes, siempre
me brindaron todo su apoyo y cariño.
A todos ellos, muchísimas gracias.

i
ÍNDICE DE CONTENIDOS
Contenidos Página
ÍNDICE DE FIGURAS...........................................................................................ii
LISTA DE ABREVIATURAS ................................................................................iV
RESUMEN............................................................................................................V
ABSTRACT ..........................................................................................................Vi
INTRODUCCIÓN..................................................................................................1
HIPÓTESIS...........................................................................................................10
OBJETIVOS ..........................................................................................................10
MATERIALES .......................................................................................................11
REACTIVOS.........................................................................................................11
MÉTODOS............................................................................................................15
RESULTADOS .....................................................................................................23
I. EXPRESIÓN DE LA TACE EN CÉLULAS PRECURSORAS DE
ADIPOCITOS HUMANOS ........................................................................23
II. CAMBIO EN LA EXPRESIÓN DE TNF- αααα DE MEMBRANA
(mTNF-αααα) EN CPA AL INHIBIR LA TACE ........................................................25
ii Efecto de Ilomastat ...................................................................................... 26
ii Efecto de TACEINH-1 ....................................................................................27
III. EVALUACIÓN DE LA ACUMULACIÓN DE LIPIDOS EN CPA AL
ESTIMULAR CON INHIBIDOR ESPECÍFICO PARA LA TACE ..........................28
IV. EVALUACIÓN DE LA ACUMULACIÓN DE LÍPIDOS EN CPA
ESTIMULADAS CON ANTICUERPO MONOCLONAL Anti-TNF- αααα....................30

ii
V. EVALUACIÓN DE LA FOSFORILACIÓN DE ERKS PRODUC TO DE LA
ESTIMULACIÓN DEL mTNF- αααα CON Anti TNF- α..............................................32
DISCUSIÓN..........................................................................................................35
RESUMEN DE RESULTADOS ............................................................................41
CONCLUSIÓN......................................................................................................41
BIBLIOGRAFÍA ....................................................................................................42
ÍNDICE DE FIGURAS
FIGURA 1: Esquema de diferenciación de adipocitos..........................................3
FIGURA 2: Generación de la forma soluble de TNF-α (sTNF-α) a partir de su
precursor de transmembrana (mTNF-α)...............................................................6
FIGURA 3: Evaluación por inmunoflouorescencia de la expresión proteica de la
TACE en la superficie celular................................................................................24
FIGURA 4: Expresión de la TACE en CPA...........................................................24
FIGURA 5: Análisis por Western Blot del efecto de Ilomastat sobre la
expresión de mTNF-α........................................................................................26
FIGURA 6: Efecto del inhibidor de la TACE (TACEINH-1) en la expresión
proteica de mTNF-α...............................................................................................27
FIGURA 7: Evaluación por tinción con Oil Red O del efecto de TACEINH-1
sobre la acumulación de lípidos neutros en CPA..................................................29
FIGURA 8: Análisis por citometría de flujo de la acumulación de lípidos
en CPA estimuladas con TACEINH-1...................................................................29

iii
FIGURA 9: Evaluación por tinción con Oil Red O del efecto del anticuerpo
monoclonal anti TNF-α sobre la acumulación de lípidos neutros
intracelulares........................................................................................................30
FIGURA 10: Análisis por citometría de flujo de CPA marcadas con
Nile Red luego del estímulo con el anticuerpo anti-TNFα.....................................31
FIGURA 11: Activación de la vía de señalización de MAPK/ERK en CPA
estimuladas con anticuerpo anti TNF-α.................................................................33
FIGURA 12: Análisis por citometría de flujo del efecto de PD98059 en la acumulación
de lípidos................................................................................................................34
FIGURA 13: Modelo propuesto..............................................................................40

iv
LISTA DE ABREVIATURAS
ADAM17: A Desintegrin And Metalloprotease N°17
AGL: Ácidos Grasos Libres.
C/EBP: CAAT/Enhancer Binding Protein.
CPA: Células Precursoras de Adipocitos.
DMEM: Dulbecco`s modified eagle medium.
ERKs: Extracellular-Signal-Regulated-Kinases.
ES: Error Standard.
HSL: Lipasa sensible a hormonas.
IF: Inmunofluorescencia.
IL-1: Interleuquina 1
IL-4: Interleuquina 4
IL-6: Interleuquina 6
IMC: Indice de Masa Corporal.
IMF: Intensidad Media de Fluorescencia.
INF-γ: Interferon gama.
LPL: Lipoproteína lipasa.
MAPKs: Mitogen-activated protein kinases.
mTNF-α: Factor de Necrosis tumoral alfa de Membrana.
mRNA: Messenger Ribonucleic Acid
PPAR: Peroxisome Proliferator activated receptor.
Pp: Preperitoneal.
Sc: Subcutáneo.
sTNF-α: Factor de Necrosis Tumoral alfa Soluble.
TA: Tejido Adiposo.
TAG: Triacilglicerol.
TACE: TNF-α Converting Enzime (ADAM 17).
TNF-α: Factor de Necrosis tumoral alfa.
TNFR: Receptor del Factor de Necrosis Tumoral alfa.
WB: Western Blot.

v
RESUMEN
El Factor de Necrosis Tumoral alfa (TNF-α) es una citoquina proinflamatoria
sintetizada y secretada por el tejido adiposo (TA). TNF-α es producida como una
proteína transmembrana de 26 kDa (mTNF-α) y procesada por una metaloproteasa de
matriz (TNF-α converting enzyme, TACE) dando origen a la forma soluble de TNF-α
(sTNF-α), de 17 kDa. Se ha descrito que en el TA sTNF-α modula procesos
relacionados con el mantenimiento del balance energético. Sin embargo, poco se sabe
acerca de las funciones que cumpliría mTNF- α en este tejido.
El objetivo de esta tesis fue estudiar la expresión de la TACE en células
precursoras de adipocito (CPA) humanos diferenciadas y no diferenciadas. Evaluar el
cambio en la expresión de mTNF-α en estas células, en presencia de un inhibidor de
la TACE, asociándolo a la acumulación de lípidos intracelulares. Y por último, evaluar
la acumulación de lípidos en CPA, como respuesta a la estimulación del mTNF-α con
un anticuerpo monoclonal específico.
Nuestros resultados indican que las CPA expresan TACE y dicha expresión no
cambia al inducir diferenciación. Por otra parte, la inhibición de la TACE determina un
aumento tanto en la expresión de mTNF-α como en la acumulación lipídica en estas
células. Encontramos también que la estimulación directa de mTNF-α con un
anticuerpo monoclonal específico, se asocia a un aumento en la acumulación de
lípidos en CPA.
Por lo tanto, dado que un aumento en la expresión de mTNF-α así como su
estimulación, se asocian a un aumento en la acumulación lipídica proponemos que a
diferencia de sTNF-α, mTNF-α podría tener un efecto lipogénico en CPA.

vi
ABSTRACT
Participation of transmembrane form of tumor necros is factor alpha
(mTNF-α) in lypogenesis
Tumor necrosis factor alpha (TNF-α) is one of the molecules produced and
secreted by adipose tissue (AT). TNF- α is a proinflammatory cytokine derived from the
cleavage of active 26 kDa membrane-bound precursor, by the matrix metalloproteinase
(TNF-α converting enzyme, TACE), to release a soluble 17 kDa form (sTNF-α). In AT,
soluble TNF-α (sTNF- α) has been described as a modulator of energy balance.
However, the role of membrane TNF (mTNF-α) in this tissue is already unknown.
The aims of this thesis was to study the expression of TACE in humans
adipocyte precursors cells (APC), both differentiated and non differentiated. To study
expression of mTNF-α in these cells, when treated with a TACE inhibitor, relating it to
intracellular lipid accumulation. And lastly, to evaluate intracellular lipid accumulation in
APC, after stimulation of mTNF-α whith a specific monoclonal antibody.
Our results indicate that APC express TACE, irrespective of cell differentiation.
On the other hand, the inhibition of TACE is associated with an increase in mTNF-α
expression in parallel whith intracellular lipid accumulation. Also, the direct stimulation
of mTNF-α whith a specific monoclonal antibody, is correlated with an increase in lipid
accumulation in APC.
In conlusion, both enhanced expression of mTNF- α and its stimulation, are
associated with an increase in lipid accumulation, therefore we propose that, unlike
sTNF- α , mTNF-α could have a lipogenic effect in APC.

1
INTRODUCCIÓN
TEJIDO ADIPOSO
El Tejido Adiposo (TA) constituye el principal reservorio energético del
organismo en mamíferos (Large y col, 2004). Las células del TA almacenan energía en
forma de triacilglicerol (TAG), el cual puede ser hidrolizado a ácidos grasos libres en
períodos de restricción calórica, lo que le otorga un papel central en la homeostasis
energética (Moreno y Martínez, 2002). Sin embargo, las funciones de este tejido son
mucho más variadas y complejas que sólo constituir un reservorio de grasa.
Actualmente se considera al TA como un órgano endocrino / secretor con un
importante rol en la regulación del balance energético (Warne, 2003; Large y col, 2004;
Kershaw and Flier, 2004).
El TA se define como un órgano heterogéneo, que está formado por diversos
tipos celulares. Uno de los principales componentes celulares del TA son los adipocitos
maduros, células esféricas que contienen gran cantidad de triglicéridos. El resto del
tejido esta compuesto por células endoteliales y células precursoras de adipocitos
(CPA), con distintos grados de diferenciación (Moreno y Martínez, 2002).
El proceso por el cual las CPA, derivadas de una célula troncal
mesenquimática, se diferencian para formar adipocitos maduros se denomina
adipogénesis (Gregorie, y col, 1998; McDoguald and Lane, 1995). Durante el proceso
de diferenciación, las CPA sufren cambios morfológicos asociados a eventos celulares
y moleculares. Desde el punto de vista morfológico las CPA cambian de un fenotipo
fibroblastoide a uno esférico con inclusiones lipídicas evidentes (figura 1) (Warne y

2
col, 2003; Gregorie, 2002). Estos cambios morfológicos son acompañados por cambios
en la composición de la matriz extracelular y el citoesqueleto (Gregoire, 2001).
Los eventos moleculares y celulares implicados en la diferenciación de CPA
han sido ampliamente estudiados in vitro. A nivel molecular, el programa de
diferenciación se desarrolla a través de la activación de genes específicos, que actúan
cooperativa y secuencialmente para expresar y mantener el fenotipo del adipocito
(McDoguald and Lane, 1995). Este programa se inicia con la expresión de los factores
de transcripción CCAAT / enhancer-binding protein β (C/EBPβ) y C/EBPδ, los que a su
vez activan a C/EBPα y PPARγ (peroxisome proliferator-activated receptor γ),
responsables finales de la expresión de varias proteínas relacionadas con la
acumulación de lípidos (triglicéridos) en los adipocitos, tales como la proteína ligante
de ácidos grasos aP2, la proteína transportadora de ácidos grasos (FATP),la acil-CoA
sintetasa (ACS), la lipoproteína lipasa (LPL) y la enzima málica (ME) (Mandrup and
Lane 1997; Uauy y col, 2000; Gregoire y col, 2001).
La lipogénesis es el proceso mediante el cual los ácidos grasos libres (AGL)
son reesterificados para formar triacilglicerol (TAG). Los AGL provienen principalmente
de hidrólisis de los quilomicrones provenientes de la absorción de lípidos dietarios o de
las lipoproteínas de muy baja densidad VLDL (very low density lipoproteins)
sintetizadas en el hígado. Los TAG de las lipoproteínas circulantes en el plasma, son
hidrolizados hasta AGL por la enzima lipoproteina lipasa (LPL), que se encuentra en la
pared de los capilares de los adipocitos. Una vez en el interior de los adipocitos, los
AGL son reesterificados para formar TAG (Moreno y Martínez, 2002; Large,y col,
2004).
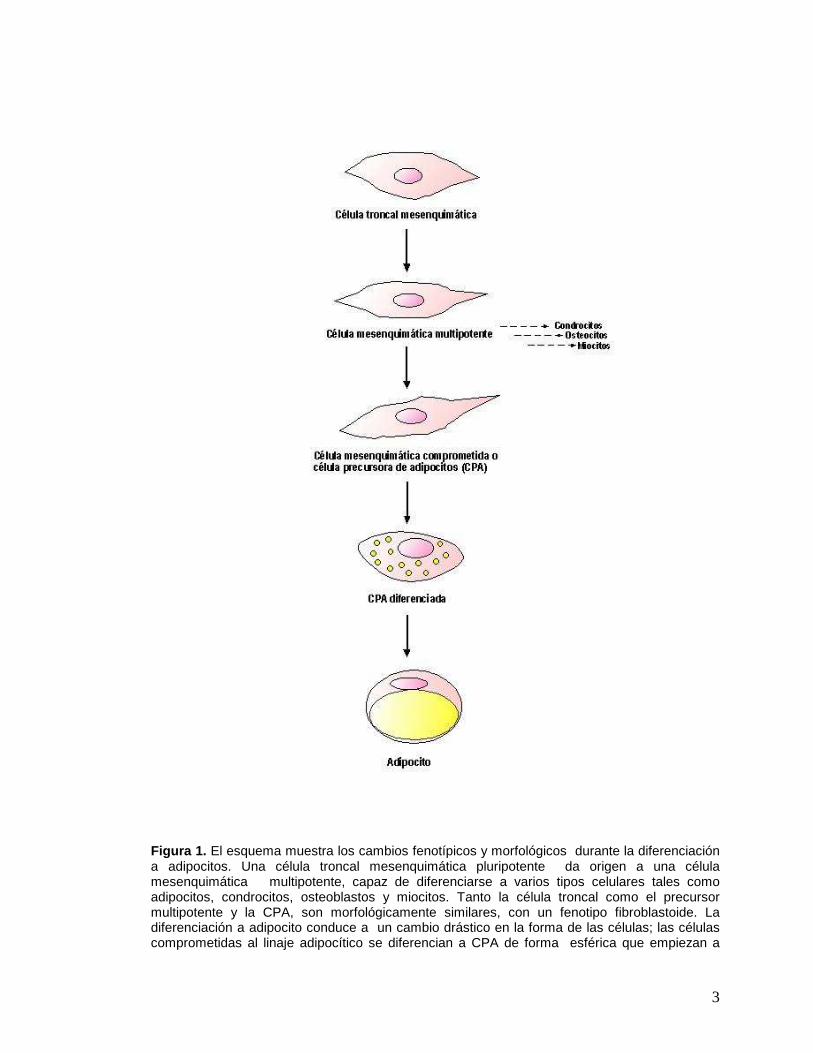
3
Figura 1. El esquema muestra los cambios fenotípicos y morfológicos durante la diferenciación a adipocitos. Una célula troncal mesenquimática pluripotente da origen a una célula mesenquimática multipotente, capaz de diferenciarse a varios tipos celulares tales como adipocitos, condrocitos, osteoblastos y miocitos. Tanto la célula troncal como el precursor multipotente y la CPA, son morfológicamente similares, con un fenotipo fibroblastoide. La diferenciación a adipocito conduce a un cambio drástico en la forma de las células; las células comprometidas al linaje adipocítico se diferencian a CPA de forma esférica que empiezan a

4
acumular lípidos y que van adquiriendo progresivamente las características morfológicas y bioquímicas propias de los adipocitos maduros. (Figura modificada de Gregoire 1998).
Los adipocitos son las únicas células capaces de cambiar drásticamente de
tamaño en condiciones fisiológicas. Esto se debe principalmente a que durante la
diferenciación, aumenta notablemente su capacidad de almacenar lípidos. Se ha
establecido que los cambios producidos en la matriz extracelular influencian los
programas de crecimiento y diferenciación celular. La activación de la vía de
señalización de ERKs (Extracellular-Signal-Regulated-Kinases) se asocia a dichos
programas (Bordeau & Jones, 1999). En estudios en TA de ratón, se ha demostrado
que la vía de señalización intracelular de las ERKs juega un rol importante en la
adaptación de las funciones adiposas al cambio de tamaño celular, producido por la
acumulación de lípidos (Farnier, 2003). Estudios en “stem cells” provenientes de
embriones de ratón, muestran que la activación de la vía de señalización de ERKs
inducida por ácido retinoico es necesaria para la diferenciación de estas células hacia
la línea adipocítica (Bost y col, 2002,2005).
Como se dijo anteriormente, el TA participa activamente en la regulación del
balance energético. Este control lo ejerce a través de la secreción de un conjunto de
factores con función endocrina, autocrina y paracrina (Moreno y Martinez, 2002). Estos
factores, que se denominan actualmente adipocitoquinas, incluyen a un conjunto de
moléculas producidas y secretadas por el TA, entre las cuales podemos mencionar la
leptina, la adiponectina, IL-6, IL-1 y TNF-α (Guzik y col, 2006; Kershaw and Flier,
2004). Además de adipocitoquinas, el TA también secreta una serie de receptores
solubles, entre los cuales está el receptor de insulina, el receptor de IL-6; el receptor de
leptina y los receptores para TNF-α (Coppack. 2001; Kershaw and Flier, 2004).

5
Dentro de la gran variedad de citoquinas secretadas por el TA, recibe particular
atención el factor de necrosis tumoral α (TNF-α) debido a su rol regulador de la masa
de tejido adiposo (Warne, 2003).
TNF-αααα EN EL TEJIDO ADIPOSO
El Factor de Necrosis Tumoral alfa (TNF-α) es una de las citoquinas
producidas y secretadas por el tejido adiposo (Warne, 2003; Hauner, 2004).
Inicialmente es producido como una proteína transmembrana de 26 kDa (mTNF-α)
(Sewter y col, 1999; Xu y col 2002; Voros y col, 2004). Esta forma es procesada
(shedding) por la metaloproteasa de matriz TACE (Tumor necrosis factor α converting
enzyme), dando origen a la forma soluble de TNF-α (sTNF-α), de 17 kDa (figura 2)
(Sewter y col, 1999; Xu y col 2002; Voros y col, 2004; Black, 2001). TACE es miembro
de la familia de las desintegrinas y metaloproteasas, o familia ADAM (ADAM17) y
corresponde a una proteína de transmembrana tipo I de 80 kDa (Black, 2001;
Sholondorff y col, 2000). EL “shedding” es el mecanismo por el cual una proteína
transmembrana es cortada proteolíticamente para liberar su dominio extracelular
desde la membrana celular (Mezyk y col, 2003). Este proceso puede ser de importante
impacto en las funciones biológicas de una proteína, ya que determina el cambio de
una actividad juxtacrina de la proteína transmembrana a una paracrina, en la forma
soluble (Mezyk y col, 2003).

6
Figura 2. El corte proteolítico del precursor transmembrana de TNF-α (mTNF-α) de 26 kDa, producido por la metaloproteasa de matriz extracelular, TACE , permite la generación de la forma soluble de TNF-α (sTNF-α) de 17 kDa (Figura modificada de Wajant H. 2003).
Se ha demostrado que tanto sTNF-α como mTNF-α son activos y se expresan
en el tejido adiposo, pudiendo interactuar con sus receptores: TNFR1 de 80 kDa y
TNFR2 de 60 kDa (Coppack, 2001). Estos dos receptores poseen una baja homología
en la zona de unión del ligando, lo que sugiere que son capaces de inducir distintas
respuestas celulares (Xu, 2001). Se ha visto que la acción antilipogénica de sTNF-α es
mediada por el TNFR1, proponiendo a mTNF-α como el principal ligando de TNFR2
(Grell y col ,1995; Xu y col, 2001; Waetzig, 2004).
Está bien establecido que, en el tejido adiposo, sTNF-α promueve lipólisis al
inducir la expresión de la lipasa sensible a hormona (HSL), enzima que cataliza la
hidrólisis de las moléculas de TAG almacenadas en el TA para convertirlas en AGL y
glicerol. Al mismo tiempo sTNF-α inhibe la expresión de factores involucrados en la
lipogénesis, como la lipoproteina lipasa (LPL). sTNF-α también es capaz de inhibir la
diferenciación de CPA, al reprimir la expresión de factores de transcripción tales como
C/EBPα y PPARγ , ambos indispensables para dar paso al proceso de adipogénesis
(Coppack y col, 1994; Gregoire y col,1998: Warne, 2003). Debido a esto, se cree que
sTNF-α tendría un rol modulador importante en muchos de los procesos relacionados

7
con el mantenimiento del balance energético. Otros antecedentes señalan a sTNF-α
como un regulador de la masa de tejido adiposo a través de su efecto anti adipogénico
(Coppack, 2001). Por otra parte, se ha relacionado con disfuncionalidades metabólicas
como la obesidad (Warne, 2003; Sewter y col, 1999; Xu y col 2002; Voros y col, 2004).
Se ha descrito en TA de individuos obesos, que existe un aumento del mRNA para
sTNF-α, sin embargo los niveles plasmáticos de esta citoquina no se correlacionan con
adipogénesis (Hotamisligil y col, 1995).
A diferencia de lo reportado para sTNF-α, existe escasa evidencia acerca de
las posibles funciones del mTNF-α . En cuanto al tejido adiposo, existen evidencias, en
líneas celulares de preadipocitos murinos que expresan la forma no procesable de
TNF-α, que demuestran que mTNF-α es biológicamente activo y llevaría a cabo sus
funciones mediante el contacto célula-célula (Xu y col, 1999). Estudios en explantes de
tejido adiposo de ratones y de humanos obesos, mostraron que la presencia de
mTNF-α es mayor en estos individuos que en los individuos controles. También se
observó que mTNF-α predomina sobre la forma soluble en individuos obesos (Xu y col,
2002). Por otra parte se ha descrito, en ratones transgénicos que expresan mTNF-α
resistente al procesamiento, que la ganancia de peso en ellos es mayor y que sus
adipocitos son de mayor tamaño que en los ratones controles que expresan mTNF-α
normal, cuando se sometieron a una dieta rica en grasa (Voros y col, 2004).
Resultados preliminares obtenidos por nuestro grupo de investigación, señalan
en cultivos de CPA humanos, que la expresión de mTNF-α aumenta al inducir
diferenciación (4 días con medio de diferenciación) y que este incremento se produce
antes que las células comienzan a acumular lípidos (7 a 11 días con medio de
diferenciación). Del mismo modo, el tratamiento de los cultivos de CPA con un

8
inhibidor no específico de metaloproteasas de matriz incrementa la acumulación de
lípidos intracelulares, una característica de la diferenciación. Respecto a la enzima
TACE, se ha encontrado expresión de su mRNA en muchos tejidos humanos, tales
como corazón, placenta, músculo esquelético, páncreas, bazo, testículos, ovarios,
entre otros (Black, y col, 1997). Sin embargo, la regulación de la actividad de la TACE
ha sido menos estudiada. Estudios en timocitos aislados de ratones transgénicos que
no expresan la TACE, mostraron una reducción de hasta un 90% en la liberación de la
forma soluble de TNF-α, demostrando el importante rol de la TACE en la regulación de
sTNF-α/mTNF-α (Killar y Col, 1999). Antecedentes en líneas celulares adipocíticas,
señalan que no hay una disminución en la expresión del mRNA de la TACE que
justifique el incremento de mTNF-α en células diferenciadas, aunque si se observó una
disminución de su actividad en células diferenciadas en comparación con células no
diferenciadas. Asimismo, los autores no encontraron una deficiencia evidente del
mRNA de TACE en tejido adiposo de ratones obesos. (Xu y col, 2002). Otros
antecedentes en líneas celulares de preadipocitos murinos, mostraron que la expresión
del mRNA de TACE no cambió significativamente cuando las células fueron sometidas
a diferenciación (Voros y col, 2003).
Además de los antecedentes descritos en TA, existen evidencias acerca del
posible rol de mTNF-α en otros modelos de estudio. Se ha descrito que los ligandos
integrados a membrana poseen la capacidad de recibir señales, actuando como
receptores (Eissner,y col,2004). Bajo este contexto, estudios en monocitos describen
que mTNF-α tendría la capacidad de inducir señalización intracelular, actuando en
sentido reverso, al ser estimulado por sus propios receptores, a través del contacto
célula-célula. (Grell y col, 1995; Eissner y col, 2000). Estudios posteriores en monocitos

9
estimulados con anticuerpo anti TNF- α revelaron que este es capaz de inducir
resistencia al LPS mediante señalización reversa a través del mTNF- α (Eissner y Col,
2004). A su vez, existen antecedentes en linfocitos que señalan que la estimulación de
mTNF-α con un anticuerpo anti TNF-α es capaz de proveer una señal coestimulatoria
para CD3 que termina en la activación de la transcripción de los genes de INF-γ e IL-4.
(Ferran y col, 1994). Estudios en células T de ratones transgénicos que sólo expresan
la forma transmembrana de TNF-α, demuestran que mTNF-α es capaz de estimular la
actividad inmune en respuesta a la infección con M.Tubercuosis (Saunders y col,
2005).Por otra parte, estudios en líneas celulares de macrófagos, han permitido
identificar que existe un alto grado de conservación de residuos de serina en el
dominio intracelular del mTNF-α (Watts y Col, 1999). En un estudio en donde
mTNF- α fue aislado de monocitos estimulados con LPS se encontró la presencia de
residuos de serina fosforilados en la porción citoplasmática de la molécula (Eissner y
Col 2004). En estudios posteriores con monocitos, se observó que existe una relación
entre la estimulación de la señalización reversa a través de mTNF- α y la activación de
la vía de las ERK/MAPK (Kirchner y Col 2003).
Las evidencias anteriormente expuestas sugieren que existe una relación entre
mTNF-α y la acumulación de lípidos intracelulares en células precursoras de
adipocitos. Además de plantear la posibilidad de que dicha molécula esté actuando
como un ligando-receptor capaz de desencadenar la acumulación de lípidos en
respuesta a un estímulo. En este trabajo evaluamos, en células precursoras de
adipocitos obtenidas de tejido adiposo humano, si un aumento en la cantidad de
mTNF-α y/o su estimulación con un anticuerpo monoclonal se correlaciona con un
aumento de los lípidos intracelulares.

10
HIPÓTESIS:
El factor de necrosis tumoral alfa de transmembrana (mTNF-α) determina un aumento
de los lípidos en células precursoras de adipocitos humanos (CPA).
OBJETIVOS a desarrollar para validar la hipótesis:
En CPA humanas:
(i) Determinar la expresión de la enzima convertidora de TNF-α (TACE).
(ii) Evaluar la acción del Inhibidor de la TACE en la expresión de mTNF-α y en
la acumulación de lípidos intracelulares.
(iii) Estudiar el efecto de la estimulación de mTNF-α con un anticuerpo
monoclonal específico en la acumulación de lípidos intracelulares. Evaluar
la participación de la vía de MAPK/ERK en respuesta al estímulo.

11
MATERIALES
Instrumentos y equipos.
- Campana de bioseguridad forma Scientific modelo 1129.
- Incubador Lab-Line CO2 automático.
- Cámaras de electroforesis y transferencia Bio-Rad.
- Centrifuga Biofuge prime R Heraeuns.
- Microscopio de Fluorescencia Nikon labophot-2.
- Microscopio Invertido de Contraste de fase Nikon modelo TMS-F
- Lector de Elisa modelo Elx-806.
- Cámara Kodak Digital Science DC120.
- Cámara Digital Nikon CoolPix 4500.
REACTIVOS
Anticuerpos.
- Anticuerpo Monoclonal anti human TNF-α, IgG 0.51 µg/µl. Purificado por Dr. J. C. Aguillón.
Laboratorio de Inmunología, Facultad de Medicina, Universidad de Chile.
-Anticuerpo Monoclonal anti-IgG de ratón, conjugado a peroxidasa, Rockland.
-Anticuerpo Monoclonal anti human TACE, R&D sistems.
-Anticuerpo Monoclonal anti-β actina, SIGMA.
- Anticuerpo Monoclonal anti-IgG de ratón, conjugado a FITC, SIGMA.
-Anticuerpo Monoclonal anti p-ERK 1,2 ratón IgG 2a, Santa Cruz.
-Anticuerpo Policlonal anti ERK 1,2 conejo IgG, Santa Cruz.
-Anticuerpo Policlonal anti IgG de conejo, conjugado a peroxidasa, Rockland.

12
Inhibidores.
- GM6001 Ilomastat 2.5mM, Chemicon Cat. Nº CC1010
-Inhibidor de la TACE “TACEINH-1” Gytognos.
-Inhibidor de ERKs PD98059, Calbiochem.
Reactivos para cultivo celular.
- Medio DMEM, GIBCO BRL 12800-017
- Medio F12, GiBCO BRL 21700-075
- Gentamicina 80mg/2ml, Laboratorio Andrómaco.
- Suero Fetal Bovino, GIBCO BRL 16000-036
- Agua bidestilada Sanderson; Laboratorio Sanderson S.A.
- Penicilina-Estreptomicina 1000U y 1000µg respectivamente por ml, GIBCO BRL
15140-122
Proteínas.
- Aprotinina, USBiological, Cat. A2300
- Albúmina Sérica de Bovino (BSA) fracción V, SIGMA A-2153.
- Leupeptina, USBiological, Cat.L2050.
- Pepstatina A, USBiological, Cat.P3280
- Colagenasa tipo 1, Worthington, Cat. 4196
- Tripsina, SIGMA, Cat No. T-4799.
Reactivos Químicos.
- Ácido clorhídrico fumante 37% p.a., Merck 317.
- Acrilamida, SIGMA, No. A-9099.
- Azul de bromofenol, USB, US12370.

13
- Azul Tripan, tinción 0,4%, GIBCO BRL 15250-061.
- β-Mercaptoetanol, Merck 15433.
- Bis-acrilamida, SIGMA, No. M-7279.
- Cloruro de Calcio dihidratado, p.a., SIGMA, Nº. C-3881
- Cloruro de magnesio hexahidrato, Merck 5833
- Cloruro de potasio, p.a., Merck 4936.1000.
- Cloruro de sodio, p.a., Vetec analytical reagents 1.0106-09
- Deoxicolato de Sodio (DOC), SIGMA, No.D-6750.
- Dexametasona, SIGMA, No. D-4902.
- Dimetilsulfóxido (DMSO), SIGMA, No. D-5879.
- D-glucosa, Merk 8337
- Dodecil sulfato de sodio (SDS), Calbiochem, Cat. 428023.
- Etiléndiamino tetraacético disódico dihidratado (EDTA), SIGMA, No. E-4884.
- Fluoruro de fenilmetilsulfonilo (PMSF), SIGMA, No. P-7626.
- Formaldehído 37%, Merck 4003.
- Fosfato dihidrogeno de potasio p.a., Merck 4871.1000
- Fosfato dibásico de sodio heptahidrato, p.a., Winkler. BM-1670
- Fosfato monobásico de sodio, p.a., Merck 6343.
- Glicerol, Winkler Cat. BM-0800.
- Glicina, SIGMA, No. G-7126.
-Hematoxilina, Merck 1.09252.0100.
- Hidróxido de sodio, p.a., Merck 6498.
- Indometacina, SIGMA, No. 17378-5G.
- Isobutilmetilxantina (IBMX), Calbiochem, Cat. 410957.

14
- Isopropanol, Merck 1.09634.2500.
- Marcador de peso molecular, Prestained Protein Marker, BioLabs P7708S
- Metanol, TCL.
- N-etilmaleimida (NEM), SIGMA, No. E-1271.
- Nile Red, SIGMA, No. N-3013.
- Nonidet (NP-40), SIGMA, No. N-6507.
- OIL RED O, SIGMA, No. O-0625.
-Ortovanadato de Sodio, SIGMA, No. S-6508.
- Sodio Bicarbonato, TCL Cat. SO-1410.
- Sulfato de Magnesio Heptahidratado, Winkler Cat. BM-1670.
- TEMED, SIGMA, No. T-9281.
- Tris, p.a., Merck 8382.
- Tween 20, Winkler Cat. BM-2031.
Otros
- Biomax Film MR, Kodak, Cat.8701302.
- Bio-Rad Dc Protein assay, Reactivos A, B y S. Cat. 500-0113/14/15
- ECL, Amersham Life Science biotech, RPN 2108.
- Inmuno-Blot Membrane, BioRad, Cat. 162-0115.
- Solución de montaje DAKO Fluorescent S3023.
- Programa Kodak Digital Science, 1D Image Análisis Software versión 3.0.
- Programa GraphPad Prism 4

15
MÉTODOS
Origen de las muestras
Se obtuvieron muestras de tejido adiposo de los depósitos grasos subcutáneo
y/o preperitoneal de pacientes hombres no obesos (IMC promedio 27,39±1,45) que
ingresaron a centros hospitalarios para herniorrafía electiva. Esta investigación fue
financiada por el Proyecto Fondecyt número 1030597/2003-2005 y su realización fue
aprobada por el comité de ética del Instituto de Nutrición y Tecnología de los Alimentos
(INTA) de la Universidad de Chile.
Obtención de células precursoras de adipocitos
La muestra de tejido adiposo subcutáneo o preperitoneal se lavó con HBSS
(CaCl·2H2O 1,2M; KCl 5,4mM; KH2PO4 44mM; MgCl2·6H2O 40mM; NaCl 0,1M;
NaHCO3 4,2mM; Na2HPO4·7H2O 22mM; D-glucosa 5,5mM; pH 7,2), se limpió de
capilares y luego se cortó en pequeños trozos de alrededor de 1mm2 en condiciones
estériles, en campana de flujo laminar. Luego se incubó con 1mg/mL de colagenasa
tipo I, por 1 hora a 37ºC en agitador de balancín, de manera de obtener las células
presentes en la muestra. Luego se filtró con gasa estéril. El filtrado obtenido se
centrifugó 7 minutos a 355 g; en el pellet se encuentran las células precursoras de
adipocitos. Estas células se resuspendieron en medio de cultivo DMEM-F12 (1:1)/10%
suero fetal bovino (SFB) con 1% de penicilina/streptomicina (100 U/100 mg/mL) y se
incubaron en placas de cultivo a 37ºC en una atmósfera controlada con 5% de CO2.

16
Expansión de CPA in Vitro
Las células se dejaron proliferar hasta que formaron una monocapa en
confluencia. Luego, fueron desprendidas de la placa con tripsina/EDTA (0.05% y 0.02%
p/v respectivamente) durante 5 minutos a 37ºC. Las células fueron removidas de las
placas y centrifugadas a 400 g por 5 minutos. A continuación fueron resuspendidas en
medio DMEM/F12 10% de suero fetal. La concentración celular se determinó en una
alícuota de 10 µl de la muestra a la que se le adicionó 10 µl de solución de Azul Tripán
al 0,4%. Concentraciones conocidas de células se sembraron en placas de 35mm o de
60mm y se dejaron proliferar hasta lograr la cantidad de células necesarias para
realizar los diferentes ensayos.
Diferenciación de células precursoras de adipocitos
Las CPA se sembraron en números conocidos y en placas de distinto tamaño,
dependiendo del ensayo. Ahí se llevaron a confluencia en medio DMEM/F12 10% SFB.
Una vez confluentes fueron inducidas a diferenciarse con DMEM-F12 / 10% SFB que
contenía dexametasona 10mM, isobutilmetilxantina 0.1 mg/ml, indometacina 0,1mM e
insulina 10,5 µg/ml (medio de diferenciación). Las células se dejaron en medio de
diferenciación por 4, 7 y 11 días y luego se utilizaron para los ensayos deseados. El
medio se renovó cada dos días.Se escogieron estos tres tiempos debido a que marcan
tres etapas en la diferenciación: a los 4 días las células no han acumulado lípidos ,
aunque sí existe un aumento en la expresión de mTNF-α; a los 7 días comienzan a
aparecer las gotas de lípidos y a los 11 días la acumulación lipídica ya es
completamente evidente.

17
Evaluación de la acumulación de lípidos neutros por tinción con Oil Red O
Se sembraron 2x104 células precursoras, depositando gotas de la suspensión
celular sobre cubreobjetos de vidrio de 12 mm de diámetro previamente esterilizados,
colocados en placas de 4 pozos. A las 24h de siembra, las células se lavaron con
HBSS y se dejaron con medio de cultivo DMEM/F12 10% SFB. Al día subsiguiente las
células fueron incubadas con medio de diferenciación o inhibidor de la TACE o anti
TNF-α por 4, 7 y 11 días. Las células fueron lavadas 2 veces con HBSS y luego fueron
teñidas con una solución saturada de Oil Red O en isopropanol al 60% durante 30
minutos a temperatura ambiente, en los días indicados. Luego las células se lavaron 3
veces con PBS (0,1M NaCl; 3mM KCl; 10mM Na2HPO4·7H2O; 2mM KH2PO4; pH 7.2) y
se agregó una solución de hematoxilina al 20% por 5 minutos a temperatura ambiente
como contratinción. Finalmente, se lavaron con agua destilada para eliminar el exceso
de hematoxilina. A continuación los cubreobjetos fueron montados en porta objetos con
solución de montaje DAKO y se dejaron secar. Finalmente las células fueron
fotografiadas con un aumento de 40X, con cámara digital Nikon montada sobre un
microscopio invertido NIKON.
Efecto de la inhibición de la TACE sobre la expresi ón de mTNF- α
Se sembraron 1x105 células precursoras en placas de 35 mm2 y se llevaron a
confluencia en medio de cultivo DMEM/F12 10% SFB. Para los experimentos se
utilizaron dos tipos de inhibidores: uno genérico para metaloproteasas de matriz
(Ilomastat) y uno específico para la TACE (TACEINH-1). Una vez que las CPA
alcanzaron confluencia, se trataron con uno u otro inhibidor (Ilomastat en
concentración 25 µM y TACEINH-1 en concentraciones 12 y 24 µM), en medio de

18
cultivo DMEM/F12 10% SFB. Para ambos casos, el medio de cultivo se renovó cada 2
días. Las células fueron lisadas para ensayos de western blot a los 4 días de estímulo,
tanto en el caso de Ilomastat como en el de TACEINH-1.
Efecto del anticuerpo anti TNF- α en la fosforilación de ERKs
Se sembraron 1x105 células precursoras en placas de 35 mm2 y se llevaron a
confluencia en medio de cultivo DMEM/F12 10% SFB. A continuación fueron
incubadas con 2µg/ml de anticuerpo monoclonal anti TNF-α, durante 15, 30 y 60
minutos para experimento a tiempos cortos, y 7 y 11 días a tiempos largos. En el
segundo caso, el medio se renovó cada 2 días. Al finalizar los tiempos establecidos de
incubación, las células fueron lisadas para ensayos de western blot.
Lisados celulares
Las células se lavaron con HBSS dos veces y se les agregó buffer de lisis ripa
(150 mM NaCl; 0.1% NP-40, 0.5% DOC; 0.1% SDS; 50mM tris pH 8; 5 µg/mL
aprotinina; 0.1 mg/mL PMSF; 1 µg/mL pepstatina; 20 µg/mL leupeptina; 1mM NEM; 0.1
mM O-vanadato), luego se lisaron las células a 4ºC utilizando un rastrillo (cell scraper).
Se estimó la concentración proteica de los lisados utililzando el kit de BioRad (Bio-Rad
Dc Protein assay), para lo cual se tomó una alícuota de 5 µL del lisado y se le agregó
25 µL de una mezcla de los reactivos A y S en la proporción 50 A: 1 S y luego 200 µL
del reactivo B, agregados en pozos de una placa de 96. La placa se analizó en un
lector de ELISA a 630 nm de longitud de onda. Los lisados se guardaron
a -20ºC.

19
Western Blot
Una alícuota de 25 µg de proteína de los lisados se sometió a denaturación en
condiciones reductoras con buffer de carga (40% SDS, 20 %Glicerol, 0.012% azul de
Bromofenol, 500 mM Tris-HCl) en presencia de β- mercaptoetanol (4%) por 5 minutos a
60ºC. Se prepararon los geles de poliacrilamida al 15% para mTNF-α y al 10% para la
TACE y p-ERKs. En todos los casos, se cargó 25 µg de proteína proveniente de los
lisados celulares por cada carril. La electroforesis se realizó a temperatura ambiente a
80V hasta que el frente pasó el gel concentrador y luego se aumentó a 120V hasta que
el frente salió del gel. El buffer de corrida contiene 50 mM Tris, 190 mM glicina y 0,1%
SDS.
A continuación de la electroforesis, se realizó la transferencia de las proteínas
desde los geles a membranas de nitrocelulosa. El buffer de transferencia contiene
50mM Tris, 190mM glicina, 20% metanol y 0,1% SDS. La transferencia se realizó a
20V, toda la noche a 4ºC. Posteriormente, se bloquearon los sitios de unión
inespecífica en las membranas con una solución de albúmina sérica de bovino (BSA) al
5% en TBS-Tween (25mM Tris; 0.1% Tween-20) por 1 hora a temperatura ambiente.
Las membranas se ensayaron para la identificación de mTNF-α, TACE, y ERK
fosforilada y no fosforilada por incubación durante 1 hora a temperatura ambiente con
el anticuerpo primario correspondiente (anti TNFα, anti TACE o anti pERK ). Los
anticuerpos primarios se usaron a una dilución de 1:1000 en BSA al 5% en TBS-
0,05% Tween 20. Se realizaron luego tres lavados, de 10 minutos cada uno, con TBS-
0,05% Tween 20. Luego se incubó con el anticuerpo secundario, anti-IgG de ratón
conjugado a peroxidasa a una dilución de 1:5000 en TBS-0,05% Tween 20, por 45
minutos a temperatura ambiente y con agitación. Luego se lavó cuatro veces con TBS-

20
0,05% Tween 20 por 10 minutos cada vez. Se reveló la membrana usando un kit de
quimioluminiscencia (ECL) y exponiendo posteriormente a film fotográfico durante uno
a diez minutos.
Las bandas obtenidas fueron cuantificadas por análisis densitométrico usando
el programa Kodak Digital Science, 1D Image 3.0. Los resultados se expresaron como
la intensidad relativa con respecto al control, todos los datos fueron corregidos por la
intensidad de β-actina como referencia de carga para el caso de mTNF-α y TACE; y
por la intensidad de ERK total para el caso de pERK.
Stripping (Lavado de anticuerpos)
Una vez que las membranas se revelaron, se sometieron a un “stripping” para
eliminar la unión del anticuerpo con la proteína de interés y ser incubadas con otro
anticuerpo contra una proteína de referencia para obtener la carga (β-actina, ERK-2).
Las membranas se lavaron en TBS/Tween y luego se colocaron en tubos con solución
de stripping (50mM Tris pH 7.2, 10% SDS, 0.07% β-mercaptoetanol) en una estufa a
65 ºC con agitación por 20 minutos. Se lavaron dos veces con PBS por 15 minutos y se
continuó con el protocolo para western blot desde la etapa de bloqueo.
Inmunofluorescencia
Se sembraron 2x104 células precursoras en medio de cultivo DMEM/F12 10%
SFB, sobre cubreobjetos de vidrio de 12 mm de diámetro previamente esterilizados y
colocados en placas de 4 pozos. Luego de 24h se retiró el medio y se lavó 2 veces con
HBSS y se mantuvieron con medio de cultivo DMEM/F12 10% SFB hasta confluencia.
Posteriormente, las células se incubaron con medio de diferenciación durante 11 días.

21
Al término del tiempo estimado para la diferenciación, las células se lavaron con HBSS
y se fijaron con PBS-paraformaldehído 2% por 10 minutos a temperatura ambiente. Se
saturó la placa con PBS/BSA 4% durante 20 minutos a 37ºC. Luego se incubó con un
primer anticuerpo monoclonal anti-human TACE (1:100 en BSA 1%) por 45 min a 37ºC.
Luego se lavó 3 veces con PBS y se incubó en cámara oscura con el anticuerpo
secundario anti-IgG de ratón conjugado a FITC por 45 minutos a 37ºC. Luego se
lavaron las células 3 veces con PBS. Para descartar marca fluorescente inespecífica,
las células fueron incubadas sólo con el anticuerpo secundario unido a FITC. Los
cristales fueron montados con una solución de montaje DAKO. Se analizaron las
muestras con un microscopio de fluorescencia y se fotografiaron con una cámara
Kodak Digital Science DC120, con un aumento de 40X.
Citometría de Flujo
Se sembraron 1x105 células en placas de 35 mm2 y se llevaron a confluencia en
medio DMEM/F12 10% SFB. Una vez confluentes se estimularon con TACEINH-1 en
concentración 24 µM por 11 días o con anti TNF-α 2 µg/ml o con anti TNF-α 2 µg/ml +
PD98059 [25µM] por 7 días. Para ambos casos se usaron células incubadas con
medio de diferenciación como control positivo. Luego de esto las células fueron
desprendidas de la placa con tripsina/EDTA (0.05 y 0.02% p/v), se lavaron con HBSS y
se dejaron en PBS. Posteriormente se les agregó una solución de Nile Red 1µg/mL,
que marca los lípidos e inmediatamente se cuantificó la fluorescencia en el citómetro
de flujo.

22
Análisis estadístico
Los resultados fueron presentados como el promedio de la intensidad de las
bandas ± error estándar. El análisis estadístico fue hecho con el programa GraphPad
Prism 4. Los resultados de la expresión de ERK se analizaron con pruebas de ANOVA
de dos vías. Para los cambios de expresión del mTNF-α, se realizaron pruebas de
ANOVA de una vía. La significancia de los resultados se representa por un valor de
p<0.05.

23
RESULTADOS
I) EXPRESIÓN DE LA TACE EN CÉLULAS PRECURSORAS DE
ADIPOCITOS HUMANOS
Para conocer si las CPA de tejido adiposo humano expresan la TACE,
evaluamos mediante ensayos de inmunofluorescencia la presencia de dicha proteína
en células no permeabilizadas. Las células fueron incubadas con o sin medio de
diferenciación durante 11 días, ya que a este tiempo se evidencia claramente la
acumulación lipídica en estas células, y por tanto se considera que se ha inducido
diferenciación. Como se observa en la figura 3 tanto las CPA incubadas con medio de
diferenciación como las no diferenciadas, evidencian presencia de la TACE en la
membrana.
Por ensayos de Western Blot (WB) se evaluó la cantidad de la TACE. Las
células se incubaron con medio de diferenciación durante 4, 7 y 11 días. Como se
aprecia en la figura 4, no existe un cambio significativo en la expresión de la proteína
en CPA diferenciadas y controles (sin diferenciar) a los tres tiempos estudiados.
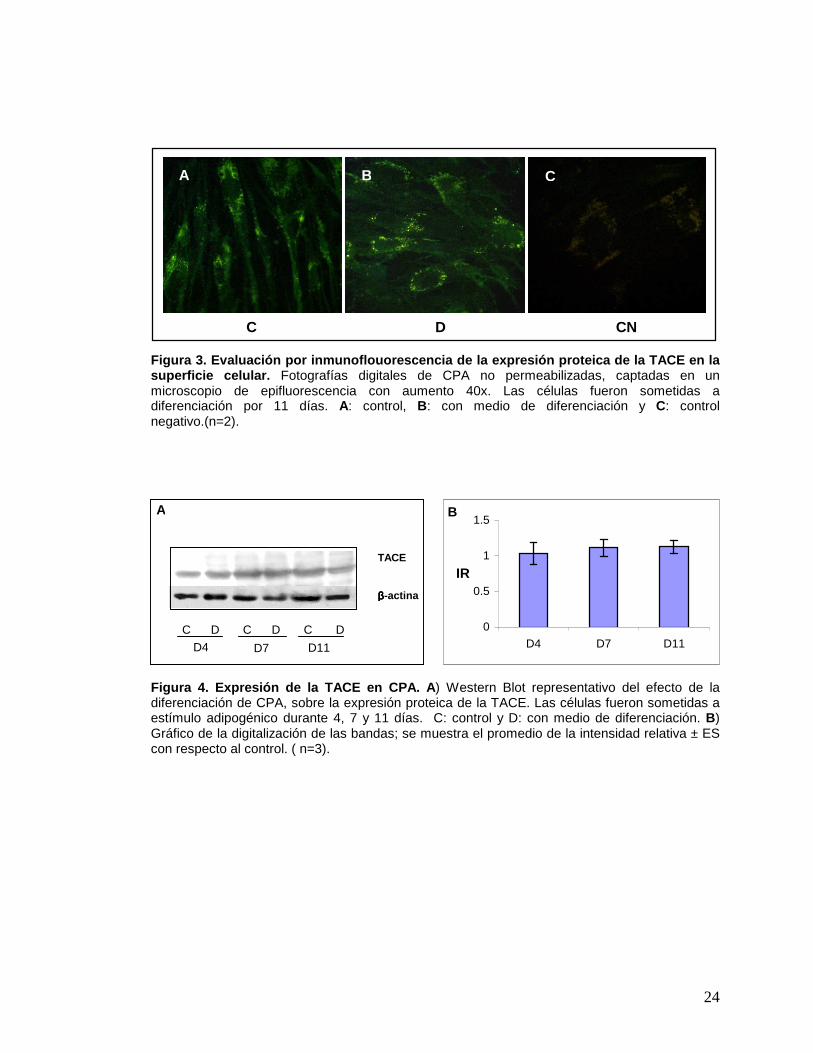
24
C D CN
Figura 3. Evaluación por inmunoflouorescencia de la expresión proteica de la TACE en la superficie celular. Fotografías digitales de CPA no permeabilizadas, captadas en un microscopio de epifluorescencia con aumento 40x. Las células fueron sometidas a diferenciación por 11 días. A: control, B: con medio de diferenciación y C: control negativo.(n=2).
Figura 4. Expresión de la TACE en CPA. A ) Western Blot representativo del efecto de la diferenciación de CPA, sobre la expresión proteica de la TACE. Las células fueron sometidas a estímulo adipogénico durante 4, 7 y 11 días. C: control y D: con medio de diferenciación. B) Gráfico de la digitalización de las bandas; se muestra el promedio de la intensidad relativa ± ES con respecto al control. ( n=3).
A C B
TACE
ββββ-actina
C D C D C D
D4 D7 D11
A
0
0.5
1
1.5
D4 D7 D11
IR
B

25
II) CAMBIO EN LA EXPRESIÓN DE TNF- αααα DE MEMBRANA (mTNF- αααα) EN
CPA AL INHIBIR LA TACE
Para conocer el efecto de la inhibición de la TACE sobre la expresión de
mTNF-α, las células, tanto preperitoneales como subcutáneas, se incubaron con dos
tipos de inhibidores: un inhibidor genérico de mataloproteasas de matriz (Ilomastat) y
uno específico para la TACE (TACEINH-1). Posteriormente se evaluó la expresión de
mTNF-α por ensayos de Western Blot.
i. Efecto de Ilomastat: En la figura 5, se muestra el cambio en la expresión de
mTNF-α cuando las células (CPA-Sc y Pp) fueron incubadas con Ilomastat. Se observa
que al tratar las CPA durante 4 días con Ilomastat, la expresión de m-TNFα aumenta
con respecto al control. También se aprecia un aumento del mTNF-α en las células
incubadas con medio de diferenciación.
ii. Efecto de TACEINH-1: Las CPA, tanto Sc como Pp, fueron incubadas con
dos concentraciones distintas del inhibidor específico para la TACE, TACEINH-1. En la
figura 6, se observa un aumento significativo en la expresión de mTNF-α para ambos
casos, al utilizar cualquiera de las dos concentraciones del inhibidor, con respecto al
control. Este resultado indica que la presencia de ambos inhibidores determina un
aumento importante en la expresión de mTNF-α en CPA provenientes de ambos
depósitos grasos, lo que sugiere una inhibición de su procesamiento.
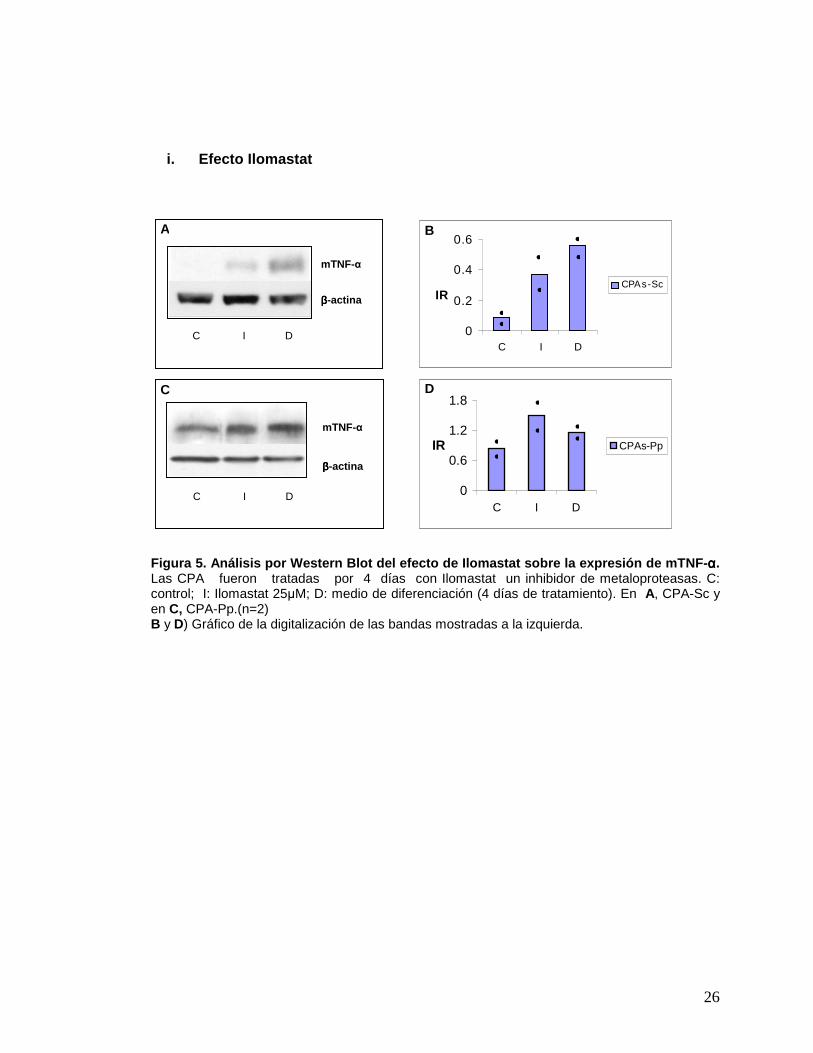
26
i. Efecto Ilomastat
Figura 5. Análisis por Western Blot del efecto de I lomastat sobre la expresión de mTNF- αααα. Las CPA fueron tratadas por 4 días con Ilomastat un inhibidor de metaloproteasas. C: control; I: Ilomastat 25µM; D: medio de diferenciación (4 días de tratamiento). En A, CPA-Sc y en C, CPA-Pp.(n=2) B y D) Gráfico de la digitalización de las bandas mostradas a la izquierda.
A
mTNF-α
ββββ-actina
C I D
C
mTNF-α
ββββ-actina
C I D
0
0.2
0.4
0.6
C I D
IRCPAs-Sc
B
0
0.6
1.2
1.8
C I D
IR CPAs-Pp
D
.
.
.
..
.
....
..
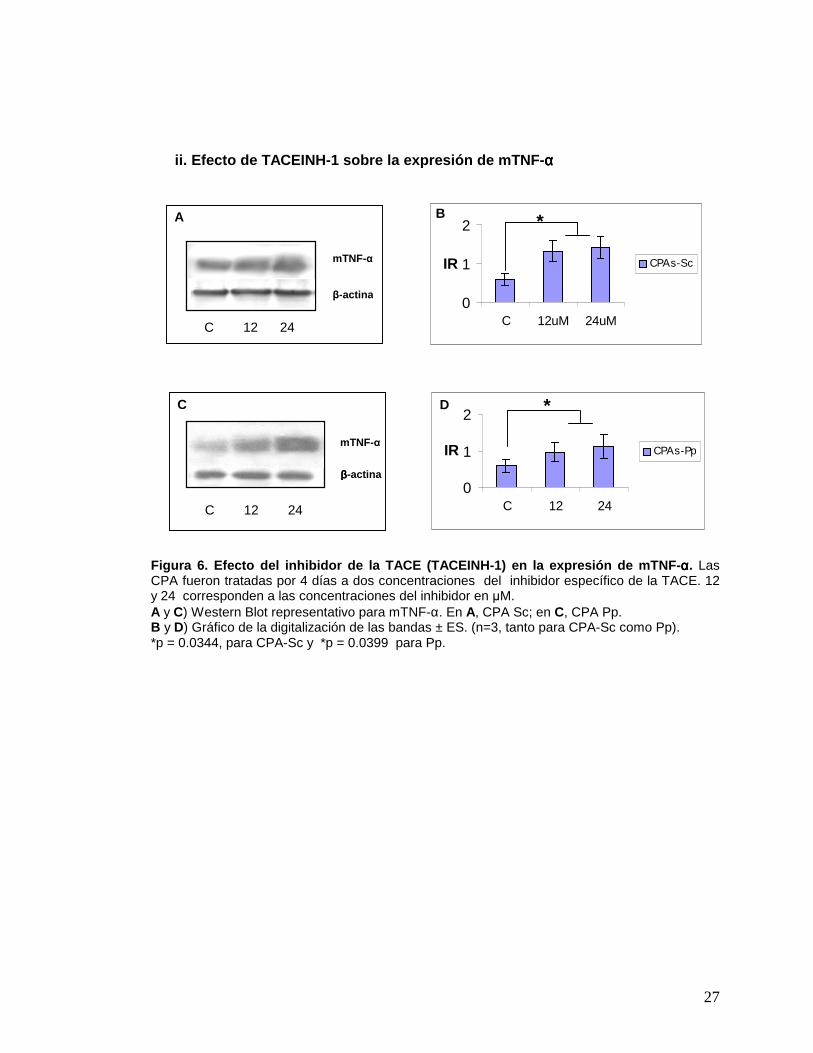
27
ii. Efecto de TACEINH-1 sobre la expresión de mTNF- αααα
Figura 6. Efecto del inhibidor de la TACE (TACEINH- 1) en la expresión de mTNF- αααα. Las CPA fueron tratadas por 4 días a dos concentraciones del inhibidor específico de la TACE. 12 y 24 corresponden a las concentraciones del inhibidor en µM. A y C) Western Blot representativo para mTNF-α. En A, CPA Sc; en C, CPA Pp. B y D) Gráfico de la digitalización de las bandas ± ES. (n=3, tanto para CPA-Sc como Pp). *p = 0.0344, para CPA-Sc y *p = 0.0399 para Pp.
B
*
0
1
2
C 12 24
IR CPAs-Pp
D
* B
mTNF-α
ββββ-actina
C 12 24
C
A
C 12 24
β-actina
mTNF-α
0
1
2
C 12uM 24uM
IR CPAs-Sc
* B
*

28
III) EVALUACIÓN DE LA ACUMULACIÓN DE LÍPIDOS EN CPA AL
ESTIMULAR CON INHIBIDOR ESPECÍFICO PARA LA TACE.
Para determinar si existe una posible correlación entre el aumento en la
expresión de mTNF-α y la aparición de gotas de lípidos intracelulares en CPA, se
evaluó el efecto del inhibidor de la TACE, TACEINH-1, por tinción con Oil Red O y
citometría de flujo. Las CPA Sc y Pp se incubaron con TACEINH-1 o con medio de
diferenciación durante 11 días, para luego evidenciar el patrón de acumulación de
lípidos por tinción de los cultivos celulares con Oil Red O, un colorante liposoluble que
tiñe los lípidos neutros. Como se muestra en la figura 7, al tratar las CPA con el
inhibidor, se produce un aumento evidente en el contenido de lípidos neutros
intracelulares, en comparación con el control, aunque menor al observado en células
tratadas con medio de diferenciación. La cuantificación de los lípidos intracelulares se
realizó mediante citometría de flujo utilizando Nile Red, un colorante fluorescente que
tiñe los depósitos de lípidos neutros. Como se muestra en la figura 8, cuando las
células son incubadas con el inhibidor de la TACE el porcentaje de células con marca
positiva para Nile Red, es 2.5 veces mayor que en la condición control.
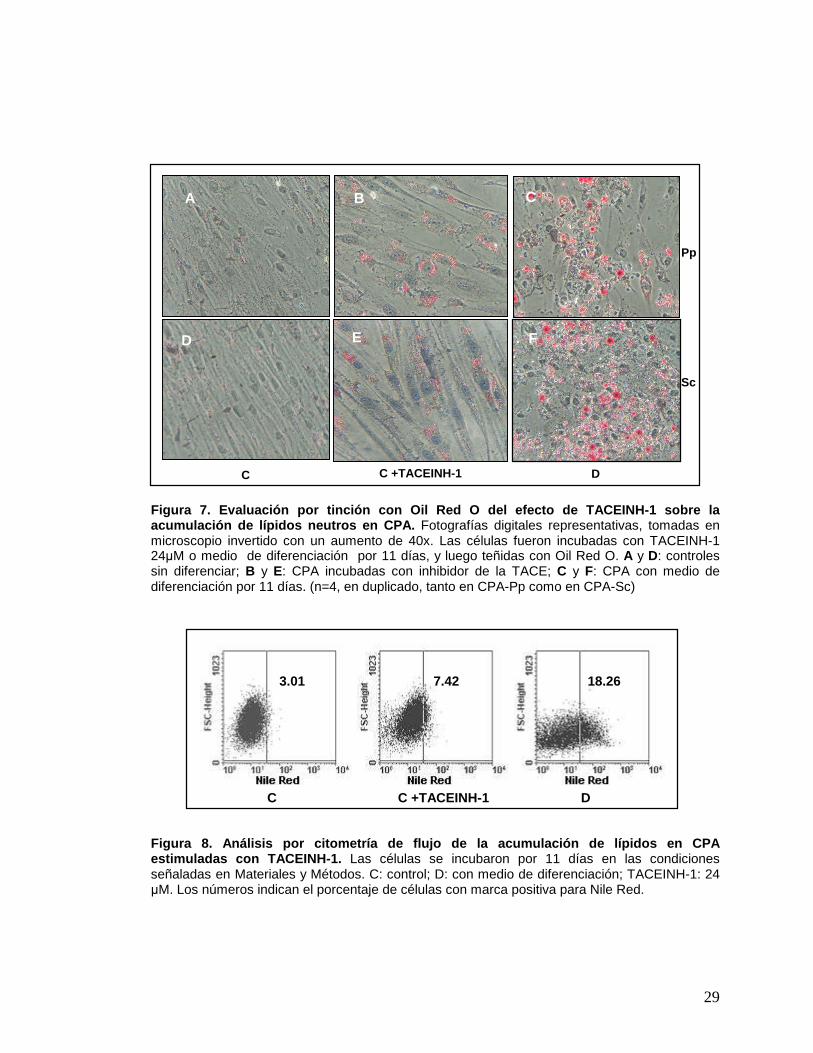
29
Figura 7. Evaluación por tinción con Oil Red O del efecto de TACEINH-1 sobre la acumulación de lípidos neutros en CPA. Fotografías digitales representativas, tomadas en microscopio invertido con un aumento de 40x. Las células fueron incubadas con TACEINH-1 24µM o medio de diferenciación por 11 días, y luego teñidas con Oil Red O. A y D: controles sin diferenciar; B y E: CPA incubadas con inhibidor de la TACE; C y F: CPA con medio de diferenciación por 11 días. (n=4, en duplicado, tanto en CPA-Pp como en CPA-Sc)
Figura 8. Análisis por citometría de flujo de la ac umulación de lípidos en CPA estimuladas con TACEINH-1. Las células se incubaron por 11 días en las condiciones señaladas en Materiales y Métodos. C: control; D: con medio de diferenciación; TACEINH-1: 24 µM. Los números indican el porcentaje de células con marca positiva para Nile Red.
C C +TACEINH-1 D
Pp
Sc
C C +TACEINH-1 D
3.01 7.42 18.26
A
D
C
F
B
E

30
IV) EVALUACIÓN DE LA ACUMULACIÓN DE LÍPIDOS EN CPA
ESTIMULADAS CON ANTICUERPO MONOCLONAL Anti TNF- αααα.
Para conocer que efectos podría tener una estimulación directa de
mTNF-α en la acumulación de lípidos, utilizamos un anticuerpo monoclonal anti TNF-α,
el cual sabemos que interactúa directamente con mTNF-α. Para esto, evaluamos por
tinción con Oil Red O, el patrón de acumulación de lípidos de CPA en respuesta a la
incubación con el anticuerpo. Las células fueron incubadas con un anticuerpo
monoclonal anti TNF-α durante 7 días. En la figura 9 observamos que existe un
aumento en la acumulación de lípidos neutros intracelulares cuando se incuba las
CPA con el anticuerpo anti TNF-α. Al cuantificar los lípidos mediante citometría de flujo
(Figura 10), podemos apreciar que cuando las células son estimuladas con el
anticuerpo se observa un aumento significativo en el porcentaje de lípidos marcados
con Nile Red respecto al control, muy cercano al de las células con medio de
diferenciación.
Figura 9. Evaluación por tinción con Oil Red O del efecto del anticuerpo monoclonal anti TNF-α sobre la acumulación de lípidos neutros intracelul ares. Fotografías digitales representativas tomadas en microscopio invertido con un aumento de 40x. Las células fueron estimuladas con anticuerpo anti TNF-α durante 7 días y luego teñidas con Oil Red O. Condiciones: A control; B 2µg/ml de anti TNF-α; C con medio de diferenciación. (n=2, en duplicado tanto en CPA-Pp, como en Sc).
B A C B
C C + Ac D
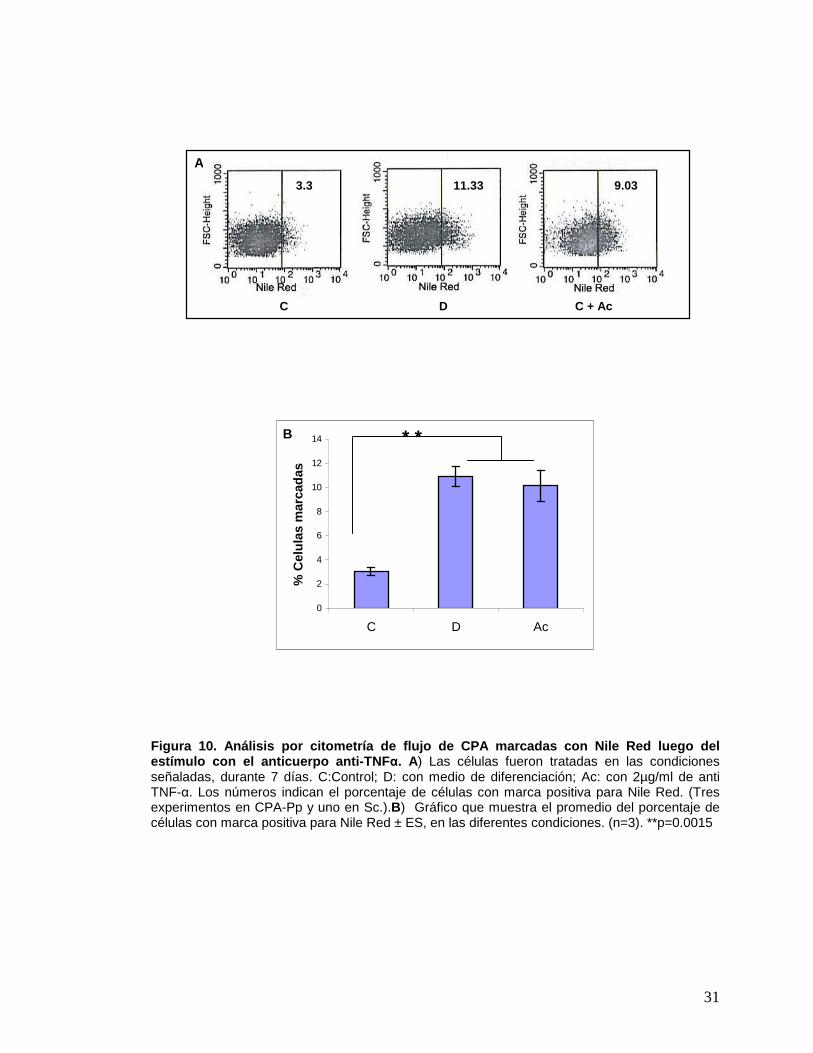
31
0
2
4
6
8
10
12
14
C D Ac
% C
elul
as m
arca
das
Figura 10. Análisis por citometría de flujo de CPA marcadas con Nile Red luego del estímulo con el anticuerpo anti-TNF α. A) Las células fueron tratadas en las condiciones señaladas, durante 7 días. C:Control; D: con medio de diferenciación; Ac: con 2µg/ml de anti TNF-α. Los números indican el porcentaje de células con marca positiva para Nile Red. (Tres experimentos en CPA-Pp y uno en Sc.).B) Gráfico que muestra el promedio del porcentaje de células con marca positiva para Nile Red ± ES, en las diferentes condiciones. (n=3). **p=0.0015
A
3.3 11.33 9.03
* *
C D C + Ac
B

32
V) EVALUACIÓN DE LA FOSFORILACIÓN DE ERKS PRODUCTO DE LA
ESTIMULACIÓN DEL mTNF- αααα CON Anti TNF- α
Sabemos por antecedentes que la activación de la vía de señalización
intracelular de las MAPK/ERK se relaciona con procesos de diferenciación celular. Con
el fin de evaluar la participación de la vía MAPK/ERK en respuesta a la estimulación
del mTNF-α, estudiamos la expresión de ERKs en CPA como consecuencia del
estímulo con el anticuerpo anti TNF-α. Las células fueron estimuladas con el anticuerpo
anti TNF-α por tiempos cortos. La expresión de ERKs se midió por Western Blot a tres
tiempos distintos. En la figura 11, observamos el cambio en la expresión de ERK
fosforilada a tiempos cortos de 15, 30 y 60 minutos luego de agregado el estímulo. Se
aprecia un aumento significativo de la fosforilación de ERKs cuando se agrega el
anticuerpo. Para conocer el efecto del anticuerpo en la fosforilación a tiempos largos,
se estimuló a las CPA con anti TNF-α o medio de diferenciación durante 7 y 11 días.
Esta vez, no se observó una activación de la fosforilación de ERKs en las células
estimuladas con el anticuerpo con respecto al control. Aunque si se apreció una
disminución en la fosforilación de ERKs en las células incubadas con medio de
diferenciación.
Una vez probado el efecto del anticuerpo sobre la activación de ERKs y sobre la
lipogénesis, evaluamos el efecto de un inhibidor de esta vía (PD98059) en la
acumulación lipídica en CPA. Para esto, las células fueron estimuladas ya sea con anti
TNF-α+PD98059, anti TNF-α, o con medio de diferenciación, durante 7 días. Al
analizar las células por citometría de flujo (figura 12), observamos que el porcentaje
de lípidos marcados con Nile Red disminuye a un valor cercano al del control cuando
33
agregamos el inhibidor de MAPK/ERK junto con el anticuerpo comparado a la
condición en que las células están siendo estimuladas sólo con el anticuerpo.
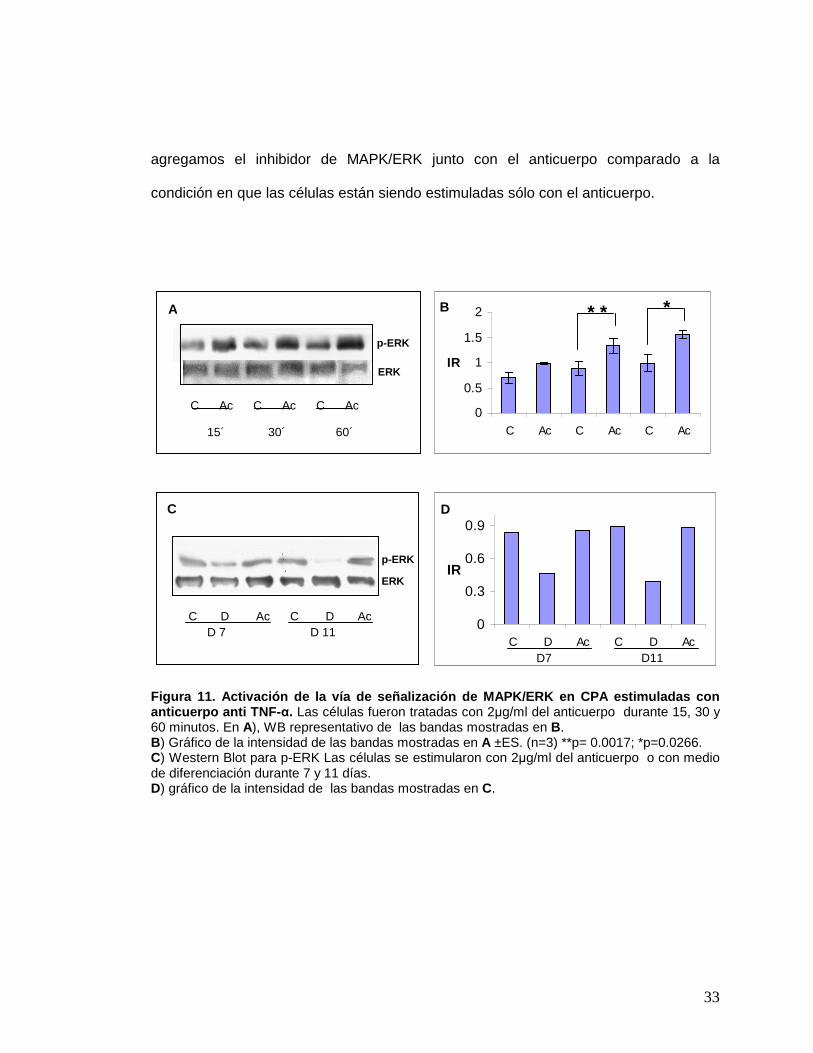
Figura 11. Activación de la vía de señalización de MAPK/ERK en CPA estimuladas con anticuerpo anti TNF- α. Las células fueron tratadas con 2µg/ml del anticuerpo durante 15, 30 y 60 minutos. En A), WB representativo de las bandas mostradas en B. B) Gráfico de la intensidad de las bandas mostradas en A ±ES. (n=3) **p= 0.0017; *p=0.0266. C) Western Blot para p-ERK Las células se estimularon con 2µg/ml del anticuerpo o con medio de diferenciación durante 7 y 11 días. D) gráfico de la intensidad de las bandas mostradas en C.
p-ERK
ERK
C Ac C Ac C Ac
15´ 30´ 60´
A
0
0.3
0.6
0.9
C D Ac C D Ac
IRp-ERK
ERK
C D Ac C D Ac D 7 D 11
C D
D7 D11
15´ 30´ 60´
0
0.5
1
1.5
2
C Ac C Ac C Ac
IR
B * * *
34
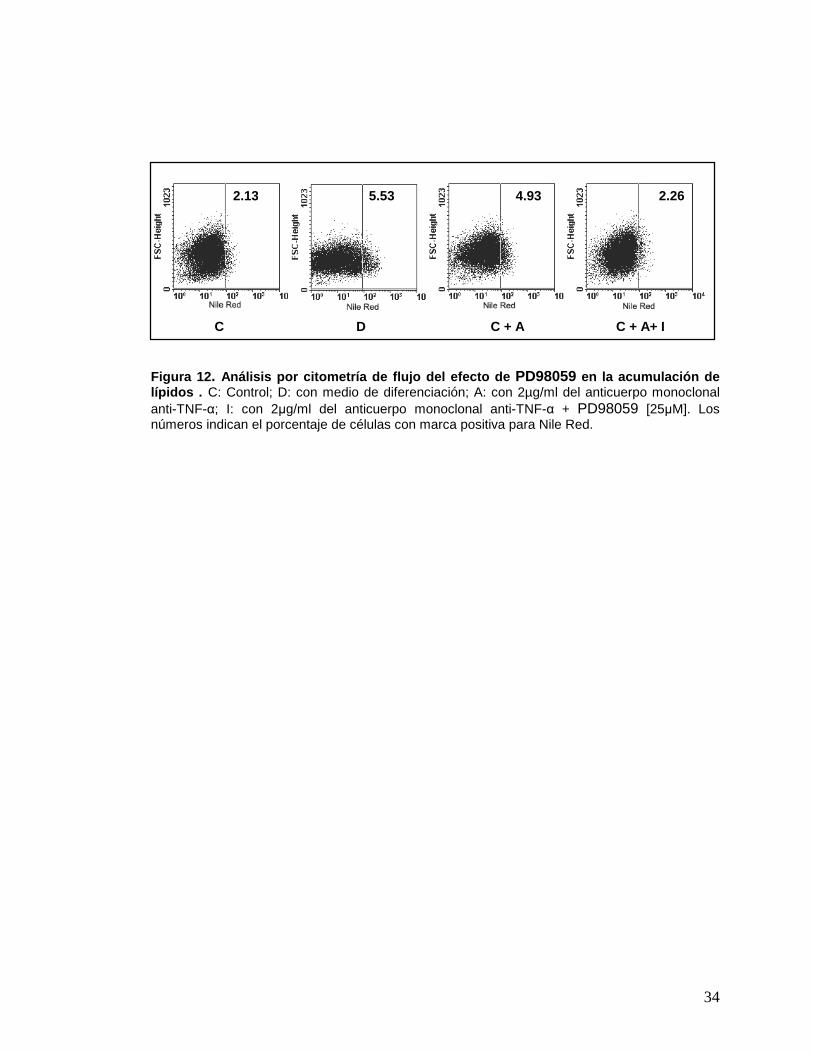
Figura 12 . Análisis por citometría de flujo del efecto de PD98059 en la acumulación de lípidos . C: Control; D: con medio de diferenciación; A: con 2µg/ml del anticuerpo monoclonal anti-TNF-α; I: con 2µg/ml del anticuerpo monoclonal anti-TNF-α + PD98059 [25µM]. Los números indican el porcentaje de células con marca positiva para Nile Red.
C D C + A C + A+ I
2.13 5.53 4.93 2.26

35
DISCUSIÓN
El estudio de los mecanismos celulares y moleculares que regulan la
proliferación y diferenciación de los preadipocitos así como el proceso de lipogénesis
es relevante para el entendimiento del desarrollo del tejido adiposo.
En esta tesis se estudió la participación del mTNF-α en la lipogénesis en
Células Precursoras de Adipocito, obtenidas de tejido adiposo humano. Para ello,
inicialmente se determinó la expresión de la TACE en CPA, se evaluó tanto la acción
de un inhibidor genérico de metaloproteasas como la de uno específico para la TACE
en la expresión de mTNF-α y en la acumulación de lípidos intracelulares. Se evaluó
también el efecto de la estimulación de mTNF-α con un anticuerpo monoclonal
específico en la acumulación de lípidos intracelulares y por último se estudió la
participación de la vía de las MAPK/ERK como respuesta intracelular desencadenada
al estimular el mTNF-α.
mTNF-α es cortado proteolíticamente por la metaloproteasa de matriz TACE,
dando origen a la forma soluble sTNF- α. Se ha observado expresión del mRNA de la
TACE en tejido adiposo murino, en preadipocitos murinos y en líneas celulares de
preadipocitos (Voros y col, 2003, Wang y col, 2006), sin embargo no existen evidencias
de la expresión proteica de la TACE en células provenientes de tejido adiposo
humano. Aunque en esta tesis no se estudió expresión del mensajero de la TACE,
nuestros resultados constituyen el primer hallazgo de la expresión proteica de la TACE
en células provenientes de tejido adiposo humano de sujetos no obesos. Observamos
que tanto las células controles como las células diferenciadas expresan una cantidad
similar de la proteína TACE. Estos resultados se relacionan con los obtenidos por Xu y

36
col, 2002 y Voros, y col 2003 en un estudio con líneas celulares de tejido adiposo
murino, donde se observó que la expresión del mRNA de TACE no cambió
significativamente durante la diferenciación. Sin embargo, el grupo de Xu y col, 2002
evaluó la actividad de TACE, observando una disminución de ésta cuando las células
fueron sometidas a diferenciación. Por otra parte, antecedentes obtenidos por nuestro
grupo de investigación señalan que en CPA obtenidas a partir de tejido adiposo
humano, la expresión de mTNF-α aumenta al inducir diferenciación. A partir de lo
anterior podemos sugerir que dado que la expresión de la TACE se mantiene
constante en las células diferenciadas y no diferenciadas, el aumento de mTNF-α
observado a los 4 días de tratamiento de las CPA con medio de diferenciación podría
estar relacionado con un cambio en la actividad de la enzima.
Sabemos por antecedentes de la literatura, que una forma de regular la
cantidad de mTNF-α es inhibiendo su procesamiento. (Xu y col, 2002; Sewter y col,
1999). Al respecto, observamos en CPA, un aumento en la expresión de mTNF-α
cuando las células se incubaron durante 4 días con el inhibidor genérico de
metaloproteasas “Ilomastat”. Para corroborar que el efecto en la expresión de mTNF-
α se debió a la inhibición de la TACE y no a la de otra metaloproteasa, tratamos las
CPA con el inhibidor específico para la TACE, TACEINH-1, y también observamos un
aumento significativo en la expresión de mTNF-α.
Nuestros resultados indican que la inhibición de la TACE es capaz de modular
la expresión de mTNF-α en CPA de tejido adiposo humano.
Considerando resultados previos de nuestro grupo que muestran que la
aparición de gotas de lípidos producto de la diferenciación de las CPA es un evento
posterior al aumento en la expresión de mTNF-α en CPA incubadas con medio de

37
diferenciación, quisimos evaluar si un aumento en la expresión de mTNF-α producto
de la inhibición de la TACE, tenía un efecto en la acumulación de lípidos intracelulares
en CPA. Para esto tratamos las CPA con el inhibidor TACEINH-1 y evaluamos por
tinción con Oil Red O y por citometría de flujo, la formación de lípidos. Por tinción con
Oil Red O observamos un aumento en la acumulación de lípidos neutros en presencia
del inhibidor con respecto al control, aunque este aumento fue menor que en las
células estimuladas con medio de diferenciación. El análisis por citometría de flujo,
mostró un aumento en el porcentaje de células positivas para Nile Red en presencia de
TACEINH-1, aunque menor que el de las células con medio de diferenciación. Si
consideramos que un aumento en la acumulación lipídica conlleva a un cambio en la
morfología y tamaño de estas células (Gregoire 1998), podemos relacionar este último
resultado con el obtenido por Voros y col, 2004, en donde observaron en ratones
transgénicos que expresan mTNF-α resistente al procesamiento, que sus adipocitos
son de mayor tamaño que en los ratones controles que expresan mTNF-α normal,
cuando se sometieron a una dieta rica en grasa. Otros antecedentes en tejido adiposo
proveniente de modelos murinos y de humanos obesos, señalan que la expresión de
mTNF-α es elevada en la obesidad (Xu y col, 2002). Nuestros resultados nos permiten
establecer una relación entre el aumento en la expresión del mTNF-α y el aumento en
la acumulación de lípidos intracelulares en CPA obtenidas de tejido adiposo de sujetos
no obesos.
Existen evidencias acerca de la capacidad de miembros de la superfamilia de
TNF de actuar como receptores de señales extracelulares (X. Wan y col, 2002; Eissner
y col, 2004). Estudios en monocitos muestran que su estimulación con anticuerpo anti
TNF-α es capaz de inducir una señal reversa a través del TNF-α de transmembrana

38
(Georg, y col, 2004). Otros estudios en líneas celulares de macrófagos, han permitido
identificar que existe un alto grado de conservación de residuos de serina en el
dominio intracelular del mTNF-α (Watts y Col, 1999). En un estudio en donde mTNF-
α fue aislado de monocitos estimulados con LPS se encontró la presencia de residuos
de serina fosforilados en la porción citoplasmática de la molécula (Eissner y Col 2004).
La presencia de estos dominios citoplasmáticos altamente conservados en la molécula
de mTNF-α, sumado el hecho de que se encuentren fosforilados en respuesta a un
estímulo, constituyen la evidencia de una posible vía de señalización intracelular en la
señalización reversa. Tomando en cuenta estos antecedentes consideramos evaluar si
la estimulación de mTNF-α con un anticuerpo monoclonal específico, sería capaz de
generar un estímulo inductor de la formación de gotas de lípidos en CPA. Bajo este
contexto, observamos por tinción con Oil Red O, que al estimular las células con el
anticuerpo monoclonal anti TNF-α se produce un aumento evidente en la cantidad de
lípidos intracelulares. Al evaluar la acumulación de lípidos por citometría de flujo, se
observó un aumento estadísticamente significativo en la cantidad de células con marca
positiva para Nile Red. Estos resultados sugieren que mTNF-α podría estar actuando
como una señal lipogénica en estas células, desencadenada por la estimulación
directa de mTNF-α con un anticuerpo específico.
Sabemos por la literatura que la vía de señalización de las ERKs está
involucrada en procesos de proliferación y diferenciación celular (Bost, y col,
2002,2005). Se ha visto, en tejido adiposo de rata, que la vía de señalización de ERKs
juega un rol importante en la adaptación de los adipocitos al cambio de tamaño
producto del almacenaje de lípidos (Farnier, y col, 2003). Además se ha reportado en
líneas celulares adipocíticas que en etapas avanzadas de la diferenciación, la

39
actividad de ERK decrece (Bordeau and Jones, 1999; Bost y col, 2005; Kim, 2007).
Con el fin de profundizar en el mecanismo molecular responsable de la aparición de
lípidos intracelulares como respuesta a la activación de mTNF-α, evaluamos la
participación de la vía de señalización MAPK/ERK en este evento. Observamos un
aumento de la forma fosforilada de ERK producto de la estimulación de CPA con el
anticuerpo a tiempos cortos de 15, 30 y 60 minutos. Sin embargo a tiempos largos de 7
y 11 días de estímulo, la intensidad relativa de las bandas obtenidas para la condición
con el anticuerpo es similar al control. Cuando se incubó las células simultáneamente
con el anticuerpo y PD98059, un inhibidor específico de la ruta de señalización
MAPK/ERK, el número de células con marca positiva para Nile Red, fue similar al
control; en cambio en ausencia del inhibidor el número de células con marca positiva
para Nile red fue el doble que en la condición control sin el anticuerpo. Estos datos
sugieren que la activación de ERKs producto de la estimulación del mTNF-α con el
anticuerpo se relaciona con un aumento en la acumulación de lípidos intracelulares en
CPA. Sabemos por antecedentes de la literatura que la activación de la vía de
señalización de MAPK/ERK regula la transcripción de una serie genes necesarios para
el crecimiento y diferenciación celular; sin embargo, para seguir adelante con el
proceso de diferenciación es necesario que la activación de ERKs sea inhibida
(Bordeau and Jones, 1999). Nosotros también observamos que al exponer las células
al medio de diferenciación por 7 y 11 días, la forma fosforilada de ERK decrece por
debajo del control, lo que está de acuerdo con lo reportado en la literatura (Bost y col
,2005; Kim y col, 2007). Lo anterior sugiere que la acumulación de lípidos como
resultado de la estimulación directa del mTNF-alpha tiene lugar a través de una(s)
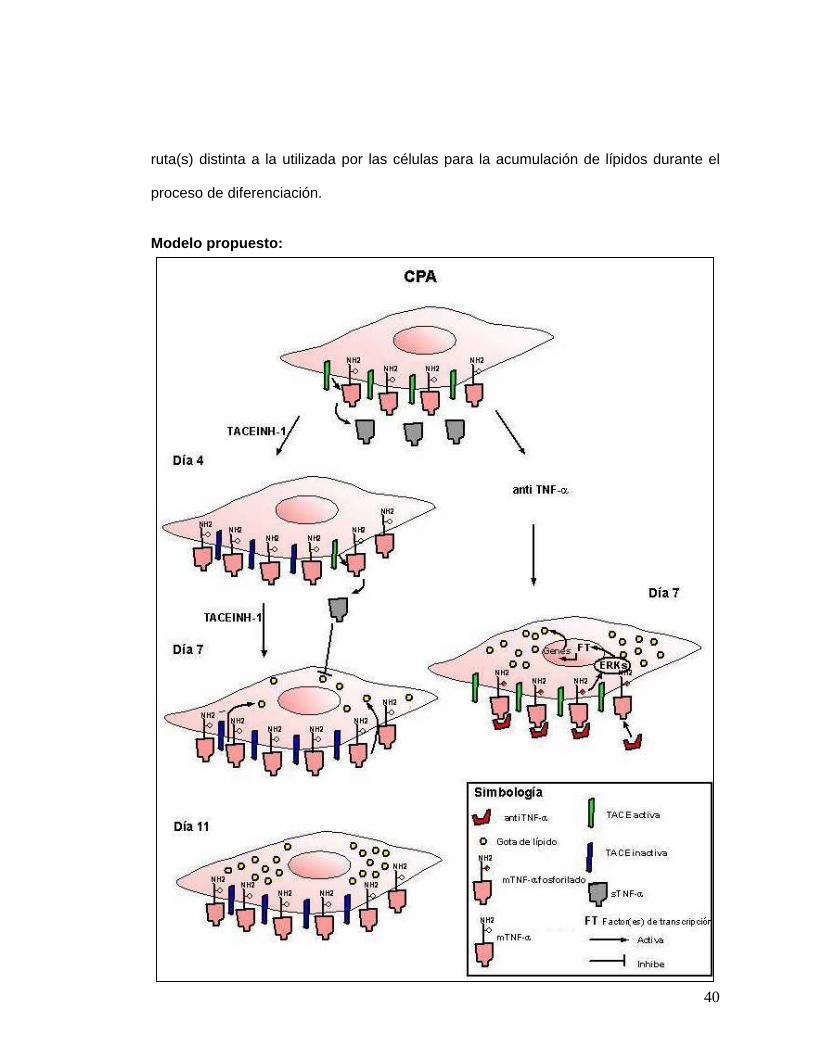
40
ruta(s) distinta a la utilizada por las células para la acumulación de lípidos durante el
proceso de diferenciación.
Modelo propuesto:

41
RESUMEN DE RESULTADOS
•Las CPA expresan la proteína TACE y su expresión no cambia durante
la diferenciación.
•La inhibición de la TACE determina un aumento en la expresión de mTNF-α lo
que se correlaciona con un aumento en la acumulación de lípidos intracelulares.
•La estimulación del mTNF- α con un anticuerpo monoclonal específico activa
la vía de las MAPK/ERKs y desencadena un aumento en la acumulación de lípidos
en estas células.
CONCLUSIÓN
Los resultados obtenidos en esta tesis sugieren que la forma
transmembrana del factor de necrosis tumoral α (mTNF-α) expresada en células de
preadipocitos (CPA) de tejido adiposo humano, juega un rol en la aparición de gotas de
lípidos (lipogénesis) en estas células. Dicho evento ocurre tanto en CPA en las que se
ha aumentado la cantidad proteica de mTNF-α utilizando un inhibidor de la enzima que
lo procesa, como en CPA estimuladas con un anticuerpo monoclonal anti TNF-α
específico.
Proponemos que la razón de mTNF-α/sTNF- α podría tener un importante
rol en el metabolismo lipídico en el tejido adiposo.

42
BIBLIOGRAFÍA
Black Roy. (2001) ”Tumor necrosis factor- α converting enzyme”. The International Journal of Biochemistry & Cell Biology. 34:1-5. Bordreau Nancy J. and Jones Peter Lloyd. (1999) “ Extracellular matrix and integrin signalling : the shape of things to come”. Journal of Biochemistry 339, 481-488. Bost Frederic, Caron Leslie, Marchettti Irene, Dani Christian, Le Marchand-Brustel Yannick and Binetruy Bernard. (2002) “Retinoic acid activation of the ERK pathway is required for embryonic stem cell commitment into the adipocyte lineage”. Biochemical Journal 361:621±627. Bost F., Aouadi M., Caron L., Binétruy B. (2005) “The role of MAPKs in adipocyte differentiation and obesity”. Biochimie 87 51–56. Coppack S., Jensen M. and Miles J. (1994) “ In vivo regulation of lipolysis in humans” Journal of Lipid Research. 35:177-193. Coppack S. W. (2001) “ Pro-inflammatory cytokines and adipose tissue”. Proceedings Of The Nutrition Society 60:349-356. Eissner Gûnter, Kolch Walter, Scheurich Peter (2004 ). “ Ligands working as a receptors: reverse signalling by members of the TNF superfamily anhance the plasticity of the immune system”. Citokine & Grouth Factor Reviews 15:353-366. Farnier C., Blache K., Diot-Dupuy F., Mory G., Ferr e P. and Bazin R. (2003) “ Adipocyte functions are modulated by cell size change: potential involvement of an integrin/ERK signalling pathway”. International Journal of Obesity. 27, 1178–1186. Ferran C., Dautry F., Merite S., Sheehan K., Schrei ber R., Grau G., Bach J-F. and Chatenoud L. (1994) “Anti-Tumor Necrosis Factor Modulates Anti-CD3-triggered T Cell Cytokine Gene Expression In Vivo”. Journal of Clinical Investigation. 93:2189-2196. Gregoire Francine M., Smas Cynthia M., and Sook Sul Hei (1998). “Understanding Adipocyte Differentiation”. Physiological Reviews 78 (3). Gregoire Francine M. (2001) “Adipocyte Differentiation: From Fibroblast to Endocrine Cell” Experimental Biological Medicine. Vol. 226(11):997–1002. Grell M., Douni E., Wajant H. , Lohden M., Clauss M., Maxeiner B., Georgopoulos S., Lesslauer W., Kollias G., Pfizenmaier K. and Sc heurich P. (1995). “The Transmembrane Form of Tumor Necrosis Factor Is the Prime Activating Ligand of the 80 kDa Tumor Necrosis Factor Receptor”. Cell 83: 793-802.

43
Guzik T. J. , Mangalat D., Korbut R. (2006) “ Adipocytokines - Novel Link Between Inflammation And Vascular Function”. Journal Of Physiology and Pharmacology 57 (4):505-508. Hauner H, (2004) “The new concepts of adipose tissue function”. Physiol Behav 83: 653-658. Hotamisligil G. Arner P., Caro J., Atkinson R, Spie gelman B.(1995) “ Increased Adiposse Tissue Expresion of Tumor Necrosis Factor –α in Human Obesity and Insuline Resistance”. Journal of chemical investigation.95:2409-2415. Kershaw Erin and Flier Jeffrey (2004). “Adipose Tissue as an Endocrine Organ”. The Journal of Clinical Endocrinology & Metabolism 89(6):2548–2556. Killar L, White J, Black R, Peschon J.(1999). “ Adamalysins. A family of metzincins including TNF-alpha converting enzyme (TACE)”. Annals of the New York Acedemy of Science 30;878:442-452. Kirkegaard T., Pedrsen G., Saermark t. and Brynskov J.(2004) “ Tumor Necrosis Factor-α converting enzyme (TACE) activity in human colonic epithelial cells”.Clinical experimental immunology. 153, 146-153. Kirchner S., Boldt S., Kolch W., Haffner S., Kazak S., Janosch P., Holler E., Andreesen R. and Eissner G. (2004). “LPS resistance in monocytic cells caused by reverse signaling through transmembrane TNF(mTNF) is mediated by the MAPK/ERK pathway” Journal of Leukocyte Biology. 75,324-331. Kyung-Ah Kim, Jung-Hyun Kim,Yuhui Wang, and Hei Soo k Sul. (2007) “ Pref-1 (Preadipocyte Factor 1) Activates the MEK/Extracellular Signal-Regulated Kinase Pathway To Inhibit Adipocyte Differentiation”. Molecular and Cellular Biology. 27:2294–2308. Large V, Peroni O, Letexier D, Ray H, Beylot M . (2 004)” Metabolism of lipids in human white adipocyte”. Diabetes Metabolism. 30,294-309. Mandrup Sussane and Lane Daniel (1997). “Regulating Adipogenesis”. The Journal of Biological Chemistry 272 (9):5367-5370. Mezyk Renata, Bzowska Monika and Bereta Joanna (200 3). “ Structure and functions of tumor necrosis factor- converting enzyme” Acta Biochimica Polonica 50:3. Moreno M.J., Martínez J.A. (2002) “ Adipose tissue: a storage and secretory organ”. Departamento de Fisiología y Nutrición. Universidad de Navarra. 25(Supl):29-39. Warne J. P. , (2003) “ Tumor necrosis factor alfa: a Key regulator of adipose tissue mass”. Journal of Endocrinology. 177: 351-355.

44
Saunders Bernadette M., Tran Stephen, Ruuls Sigrid, Sedgwick Jonathon D., Briscoe Helen and Britton Warwick J. (2005) “Transmembrane TNF Is Sufficient to Initiate Cell Migration and Granuloma Formation and Provide Acute, but Not Long-Term, Control of Mycobacterium tuberculosis Infection1”. The Journal of Immunology., 174: 4852–4859. Sewter C.P.; Digby J.E.; Blows F.; Prins J.; O’Rahi lly S. (1999) “Regulation of tumour necrosis factor-alpha release from human adipose tissue in vitro”. J. Endocrinol. 163: 33-38. Uauy D, R., Martinez A, J. y Rojas B, C. “Nutrición molecular, papel del sistema PPAR en el metabolismo lipídico y su importancia en obesidad y diabetes mellitus: regulation of lipid metabolism by peroxisome proliferator activated receptors (PPAR). Their relatioship to obesity and diabetes mellitus”. Rev. méd. Chile, abr. 2000, vol.128, no.4, p.437-446. ISSN 0034-9887. Voros G., Maquoi E., Collen D., Lijnen H.R. (2004) “Influence of membrane-bound tumor necrosis factor (TNF)-α on obesity and glucose metabolism”. J. Thromb. Haemost. 2: 507-513. Voros G., Maquoi E., Collen D., Lijnen R.H. (2003) “Differential expresión of plasminogen activator inhibitor-1, tumor necrosis factor-α , TNF-α converting enzyme and ADAMTS family members in murine fat territories”. Biochimica et Biophysica Acta 1625:36–42. Waetzig G., Rosenstiel P., Arlt A., Till A., Bräuti gam K., Schäfer H., Rose-John S., Seegert D. and Schreiber S. (2004). “Soluble tumor necrosis factor (TNF) receptor-1 induces apoptosis via reverse TNF signaling and autocrine transforming growth factor-β1”. The FASEB Journal Wan X., Zhang J., Luo H., Shi G., Kapnik E., Kim S. et al. (2002) “A TNF family member LIGHT transduces costimulatory signals into human T cells”. J. Immunol. 169 6813–6821. Watts A., Hunt H., Wanigasekara Y., Bloomfield G., Wallach D., Roufogalis B. and Chaudhri G. (1999) “A casein kinase I motif present in the cytoplasmatic domain of members of the tumor necrosis factor ligand family is implicated in reverse signaling”. The EMBO journal 8: 2119-2126. Xu H., Uysal K.T., Becherer J.D., Arner P., Hotamis ligil G.S. (2002) , “Altered Tumor Necrosis Factor-α (TNF-α) Processing in Adipocytes and Increased Expression of Transmembrane TNF-α in Obesity.” Diabetes 51: 1876-1883. Xu H., Hotamisliguil G. (2001) ” Signaling pathways utilized by tumor necrosis factor receptor 1 in adipocytes to suppress differenciation”. FEBS letters.506:97-102.

45
Xu H., Sethi J. and Hotamisliguil G. (1999) “ Transmembrane form of tumor necrosis factor α inhibits adipocyte differenciation by selectively activating TNF receptor “. The Journal Of Biological Chemistry. 274, 37: 26287-26295. Yoshiyuki Tanabe, Masaru Koga, Maki Saito, Yumi Mat sunaga and Koichi Nakayama. (2004) “ Inhibition of adipocyte differentiation by mechanical stretching through ERK-mediated downregulation of PPARy". Journal of Cell Science. 117: 3605-3614.





![EL MÉTODO DE LAS ὑποθέσεις EN PHÆDOrepositorio.uchile.cl/tesis/uchile/2005/caro_s/sources/caro_s.pdf · que, para Platón, la ἰδέα es el carácter de todas las [cosas]](https://static.fdocument.org/doc/165x107/5be1c3da09d3f2f02d8b4c38/el-metodo-de-las-en-pha-que-para-platon-la-.jpg)
