
Obtinerea de enzime mutante New mutant enzymes · Atasarea proteinelor pe unele situsuri de legare...
44
Obtinerea de enzime mutante https://www.neb.com/ β-lactamaza 1982 New mutant enzymes β-lactamase 1982 Mutații punctiforme Site-directed mutagenesis
Transcript of Obtinerea de enzime mutante New mutant enzymes · Atasarea proteinelor pe unele situsuri de legare...
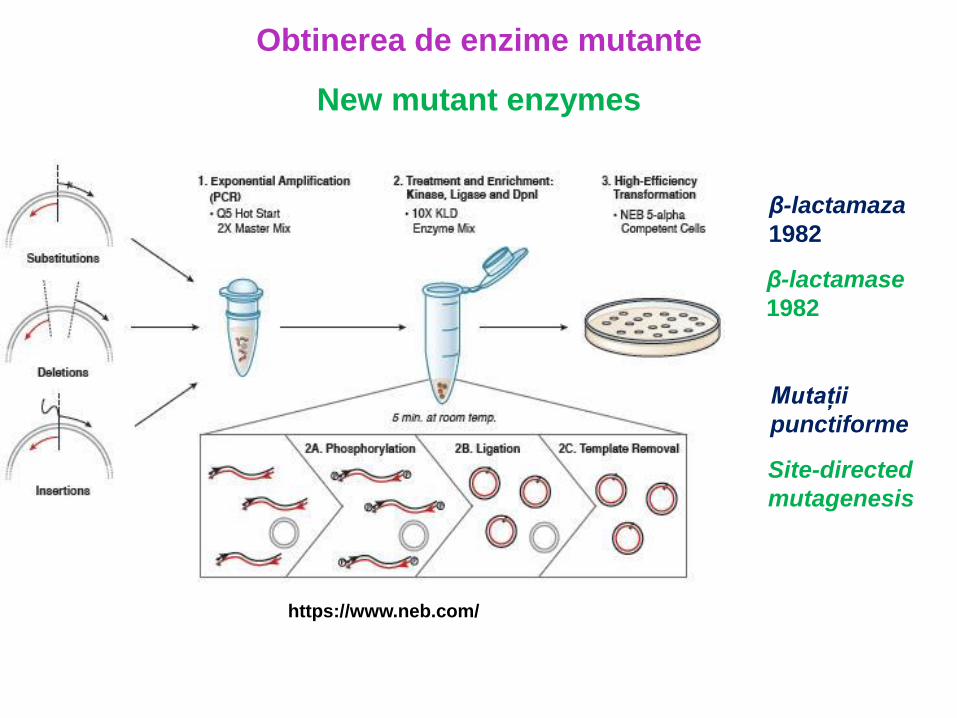
Obtinerea de enzime mutante
https://www.neb.com/
β-lactamaza
1982
New mutant enzymes
β-lactamase
1982
Mutații
punctiforme
Site-directed
mutagenesis
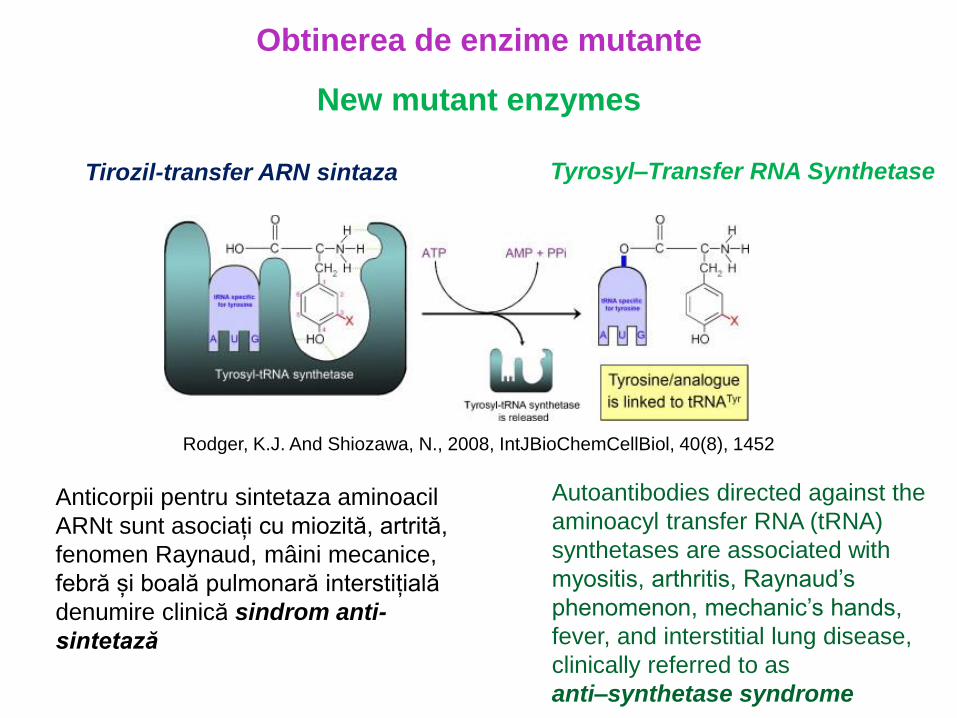
Obtinerea de enzime mutante
New mutant enzymes
Tirozil-transfer ARN sintaza Tyrosyl–Transfer RNA Synthetase
Rodger, K.J. And Shiozawa, N., 2008, IntJBioChemCellBiol, 40(8), 1452
Autoantibodies directed against the
aminoacyl transfer RNA (tRNA)
synthetases are associated with
myositis, arthritis, Raynaud’s
phenomenon, mechanic’s hands,
fever, and interstitial lung disease,
clinically referred to as
anti–synthetase syndrome
Anticorpii pentru sintetaza aminoacil
ARNt sunt asociați cu miozită, artrită,
fenomen Raynaud, mâini mecanice,
febră și boală pulmonară interstițială
denumire clinică sindrom anti-
sintetază
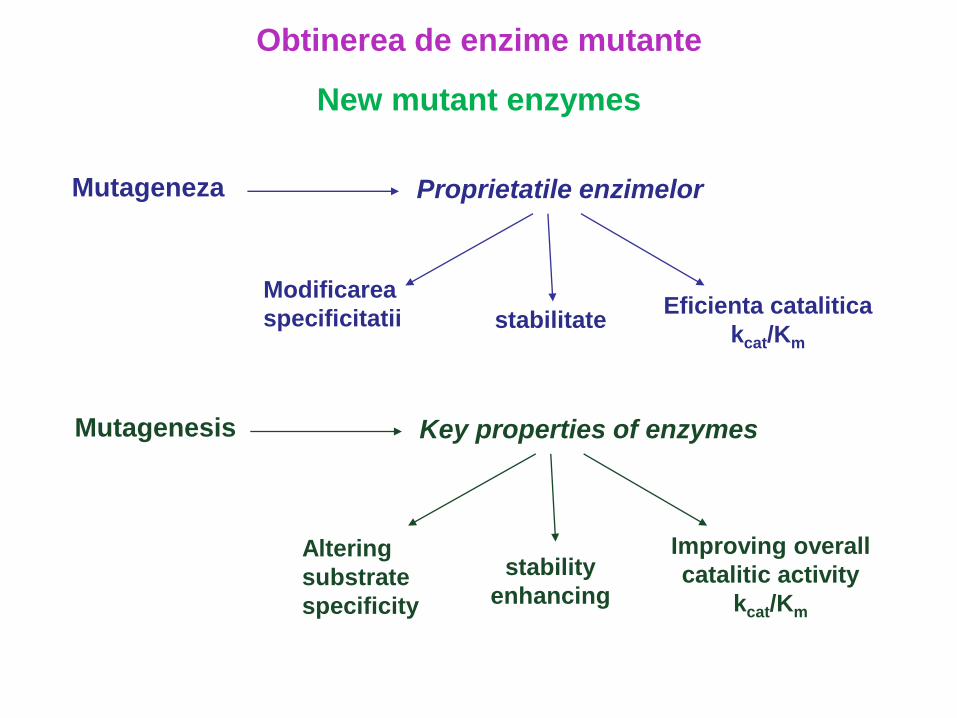
Mutageneza Proprietatile enzimelor
stabilitate
Modificarea
specificitatiiEficienta catalitica
kcat/Km
Obtinerea de enzime mutante
New mutant enzymes
Mutagenesis Key properties of enzymes
stability
enhancing
Altering
substrate
specificity
Improving overall
catalitic activity
kcat/Km

Stabilizarea enzimelor
Utilizate în aplicații industriale
Enzyme Stability
Wide range of industrial applications
Îmbunătațirea performanțelor
în condițiile de lucru
To improve enzyme performance
under process conditions
Condiții extreme
temperatura
presiune
pH
alți parametri
Extreme conditions
temperature
pressure
pH
other conditions
Enzime hiperstabile
(condiții extreme de mediu)
Hyperstable enzymes
(extreme environmental
conditions)
2 strategii de stabilizare
prezenta unor suprafețe
mari cu interacții electrostatice
tendință de multimerizare
2 stabilizing strategies
large surface networks of
electrostatic interaction
tendency to be multimeric

Stabilizarea enzimelor
O serie de strategii-mutații
Enzyme Stability
Several approaches-mutations
o Reducerea entropiei
proteinelor despachetate
punți S-S
↑nr. Pro (bucle mobile)
o To reduce the enthropy
of unfolded proteins
disulfide bridges
↑no Pro (mobile loops)
o Cresterea nr de -helixuri
Gly→Ala
impachetare
centru hidrofob
o To increase the number of -helices
Gly→Ala
well-packed
hydropobic cores
o Inginerie de punți de sare o Engineering of salt bridges
o Analiza filogenetica
proiectare logică
AA preferati in
organsimele termofile
o Engineering of salt bridges
Rational design
favoring common residues
(thermophiles)
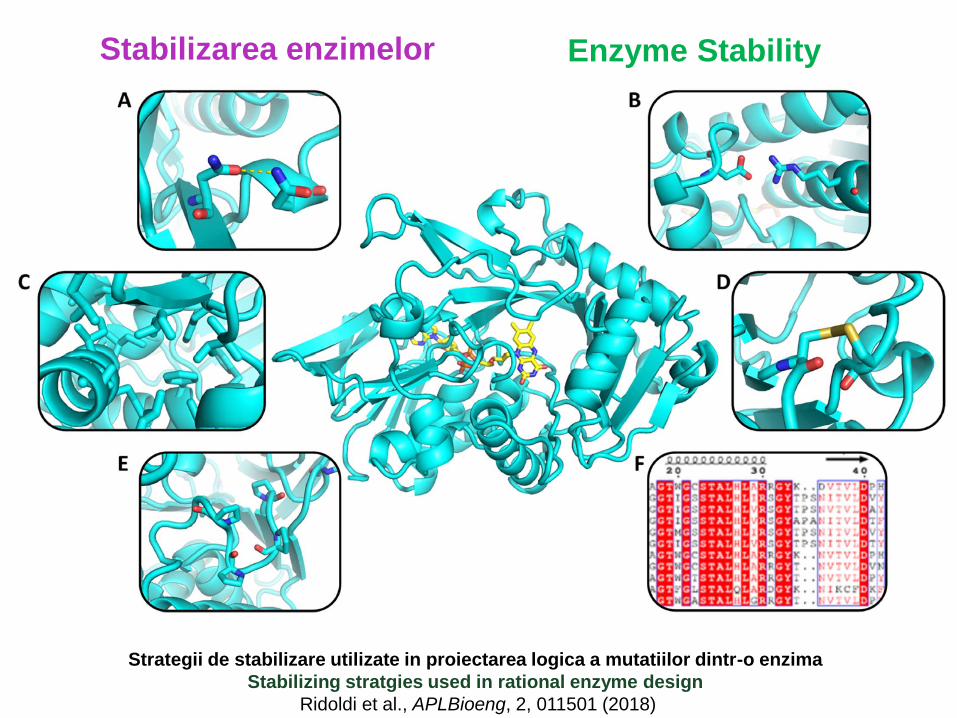
Stabilizarea enzimelor Enzyme Stability
Strategii de stabilizare utilizate in proiectarea logica a mutatiilor dintr-o enzima
Stabilizing stratgies used in rational enzyme design
Ridoldi et al., APLBioeng, 2, 011501 (2018)
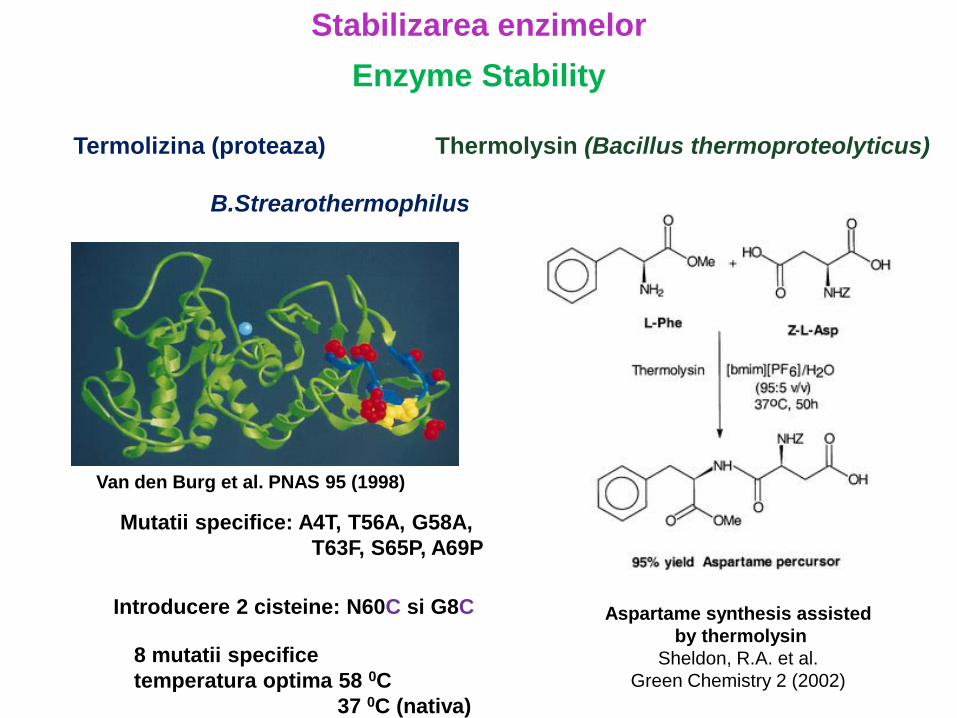
Termolizina (proteaza)
B.Strearothermophilus
Mutatii specifice: A4T, T56A, G58A,
T63F, S65P, A69P
Introducere 2 cisteine: N60C si G8C
Van den Burg et al. PNAS 95 (1998)
8 mutatii specifice
temperatura optima 58 0C
37 0C (nativa)
Stabilizarea enzimelor
Enzyme Stability
Thermolysin (Bacillus thermoproteolyticus)
Aspartame synthesis assisted
by thermolysin
Sheldon, R.A. et al.
Green Chemistry 2 (2002)
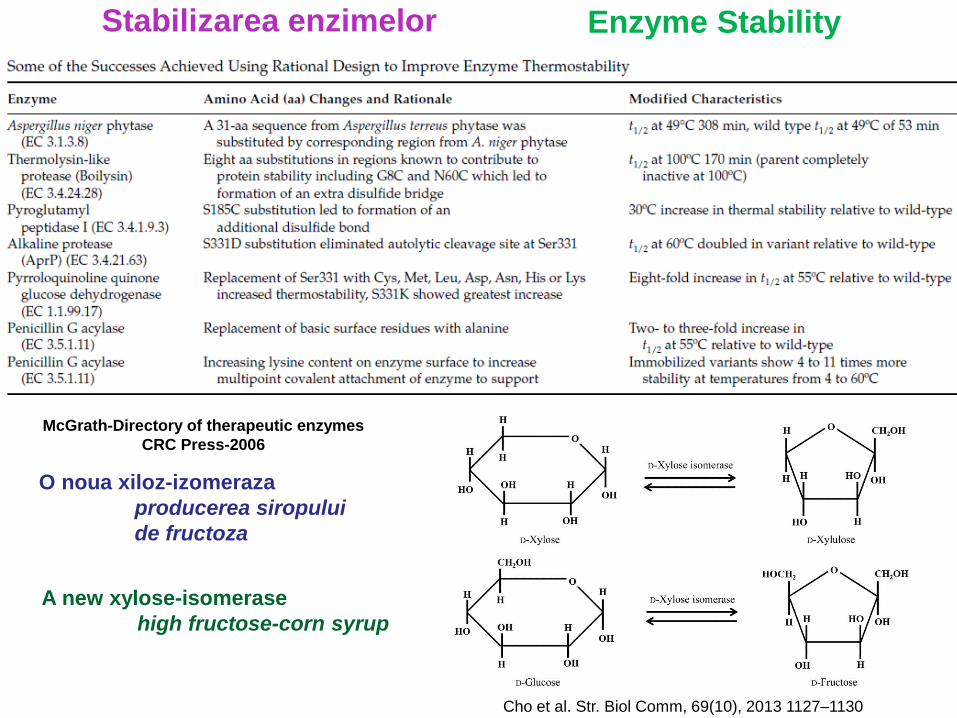
Stabilizarea enzimelor
McGrath-Directory of therapeutic enzymes
CRC Press-2006
O noua xiloz-izomeraza
producerea siropului
de fructoza
Enzyme Stability
A new xylose-isomerase
high fructose-corn syrup
Cho et al. Str. Biol Comm, 69(10), 2013 1127–1130
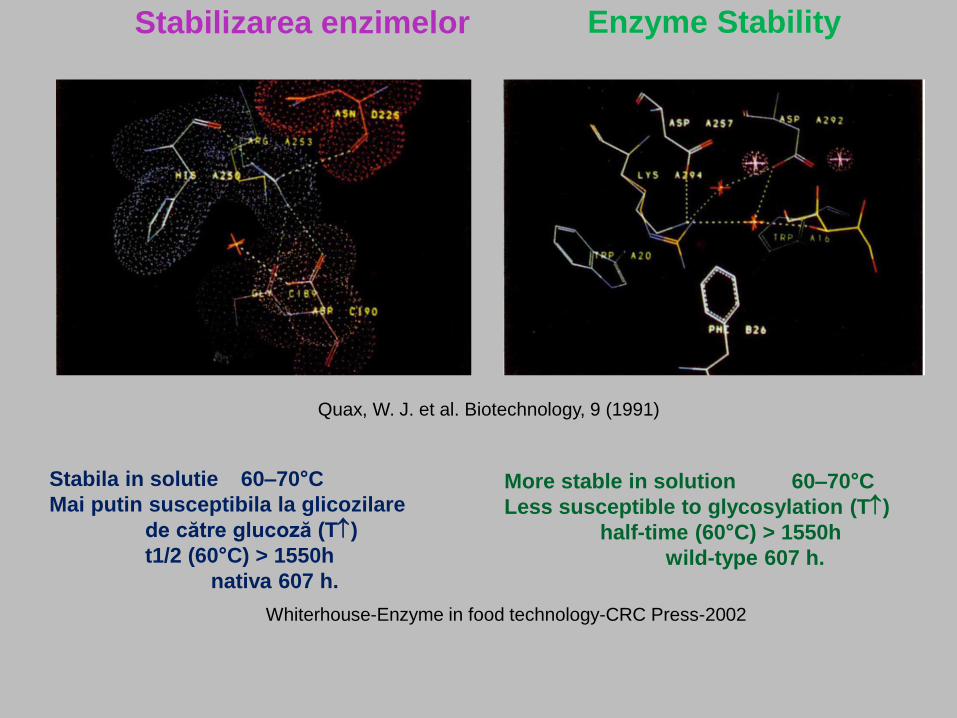
Stabilizarea enzimelor Enzyme Stability
Quax, W. J. et al. Biotechnology, 9 (1991)
Stabila in solutie 60–70°C
Mai putin susceptibila la glicozilare
de către glucoză (T)
t1/2 (60°C) > 1550h
nativa 607 h.
Whiterhouse-Enzyme in food technology-CRC Press-2002
More stable in solution 60–70°C
Less susceptible to glycosylation (T)
half-time (60°C) > 1550h
wild-type 607 h.
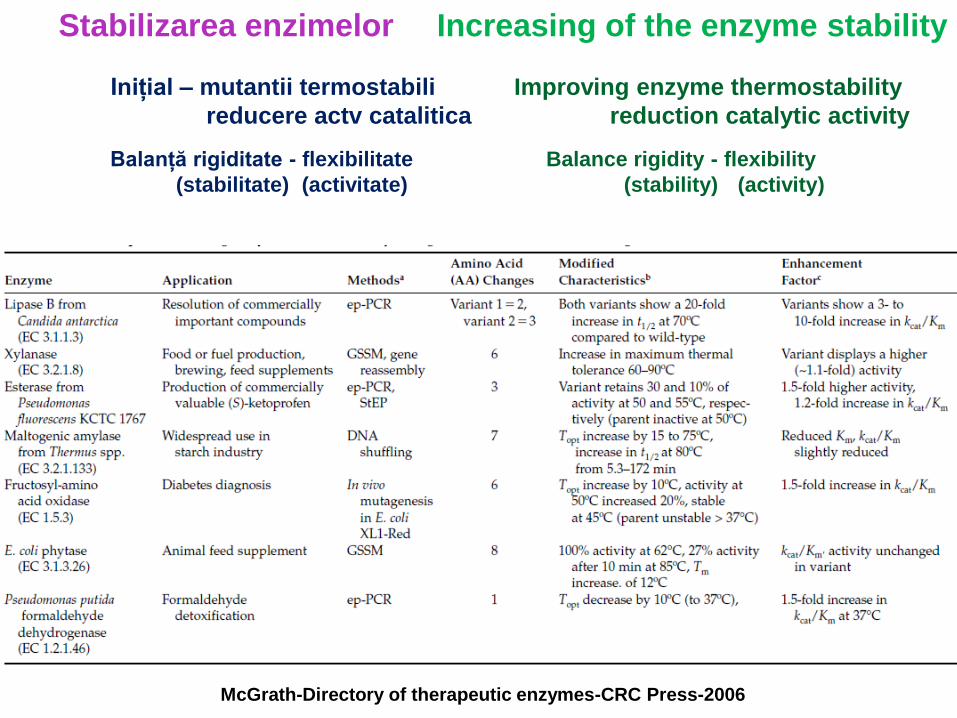
McGrath-Directory of therapeutic enzymes-CRC Press-2006
Stabilizarea enzimelor Increasing of the enzyme stability
Inițial – mutantii termostabili
reducere actv catalitica
Improving enzyme thermostability
reduction catalytic activity
Balanță rigiditate - flexibilitate
(stabilitate) (activitate)
Balance rigidity - flexibility
(stability) (activity)

Structurile enzimelor pot fi
stabilizate prin reticulare
intramoleculara
Enzyme structures can be stabilized
by forming intracellular crosslinks
Stabilizarea enzimelor Increasing of the enzyme stability
glutaraldehida glutaraldehyde
Cristale enzimatice reticulate
cu glutaraldehidaCrosslinked enzyme crystals
Glucoamilaza, lipaza
Subtilizina, termolizina
Glucoamylase, lipases
subtilizin, thermolysin
Atasarea proteinelor pe unele
situsuri de legare ale polimerilor
Enzyme attachment to multiple sites
of a suitable polymers
Invertaza / Chitosan sau pectina65 grade 5 min → 5h, 5 min → 2 zile
Invertase / Chitosan and pectin65 degree 5 min → 5h, 5 min → 2 days
DAAO instabila > 55 grade
atasare pe dextranstabila pana la 75 grade
DAAO unstable > 55 degree
DAAO/dextranstability up to 75 degree
Glucoz oxidaza- biosenzori glucozaInd. alimentara, fermentatie
GO / bile de sticla
stabila pana la 75 grade
crestere cu 180% activitate
Glucose Oxidase – glucose biosensorsfood and fermentation processes
GO / Glass beads
stability up to 75 degree
180% increase of relative activity
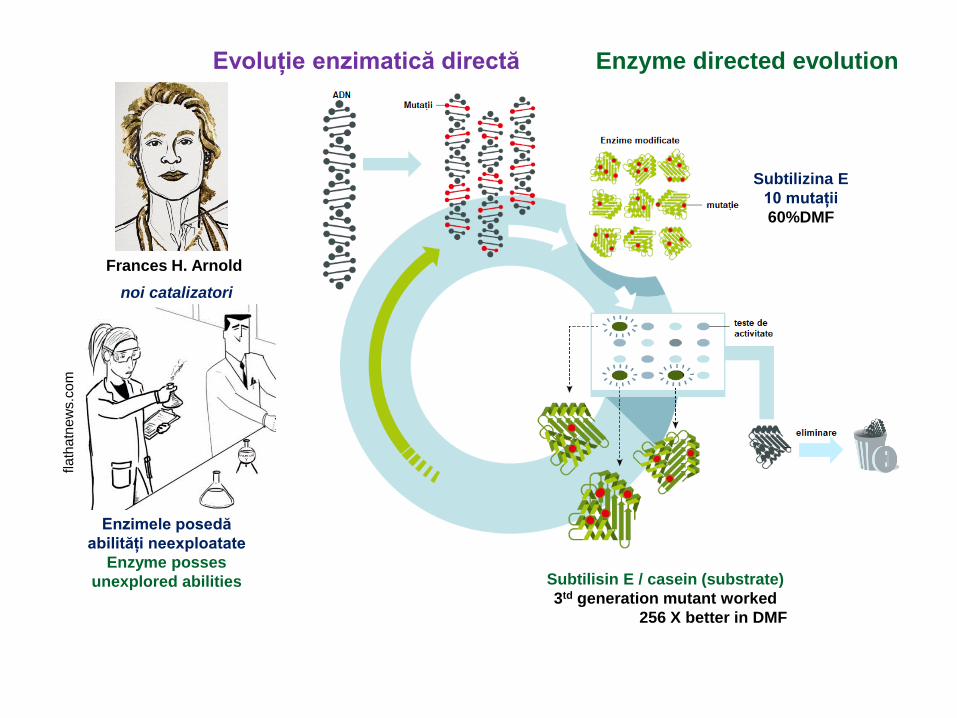
Frances H. Arnold
Evoluție enzimatică directă Enzyme directed evolution
noi catalizatori
Enzimele posedă
abilități neexploatate
Enzyme posses
unexplored abilities
fla
tha
tne
ws.c
om
Subtilizina E
10 mutații
60%DMF
Subtilisin E / casein (substrate)
3td generation mutant worked
256 X better in DMF

Evoluție enzimatică directă Enzyme directed evolution
Metodologia se bazează pe A methodology relying on
1) Identificarea unei enzime
adecvate pentru cerința
1) Identification of a suitable starting
enzymes for the chosen task
2) Construcția unui set de
secvențe ADN ce acoperă
subsetul ales din secvență
2) DNA-sequence library construction
to cover well-chosen subset
of sequence space
3) Identificarea
unor criterii de selectie pentru
îmbunătățiri/ funcții noi
metodele de selectie a
versiunilor de mutanti
optimizate
3) Identification
selection criteria that will lead
to enhanced/ new function
methods for selection of optimized
enzyme variants
4) Rediversificarea genelor pentru
a crea noi librarii de ADN pe baza
secvențelor din prima selectie
4) Re-diversification of the genes
to create new DNA-sequence libraries
arround the sequences from
the first selection
5) Configurarea criteriilor de selecție
pentru stabilirea numarului de
runde necesare pentru a atinge
nivelul țintă al performanței enzimelor
5) Setup of selection criteria with
increased stringency, for as many rounds
as needed to reach the target level
of enzyme performance
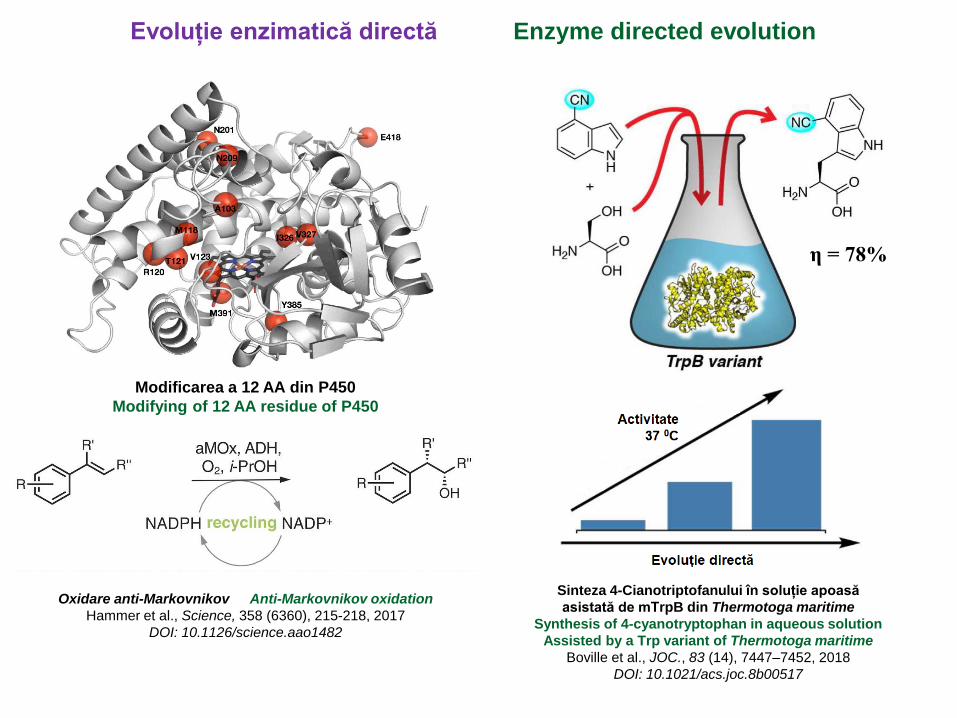
Oxidare anti-Markovnikov Anti-Markovnikov oxidation
Hammer et al., Science, 358 (6360), 215-218, 2017
DOI: 10.1126/science.aao1482
Modificarea a 12 AA din P450
Modifying of 12 AA residue of P450
Sinteza 4-Cianotriptofanului în soluție apoasă
asistată de mTrpB din Thermotoga maritime
Synthesis of 4-cyanotryptophan in aqueous solution
Assisted by a Trp variant of Thermotoga maritime
Boville et al., JOC., 83 (14), 7447–7452, 2018
DOI: 10.1021/acs.joc.8b00517
η = 78%
Evoluție enzimatică directă Enzyme directed evolution
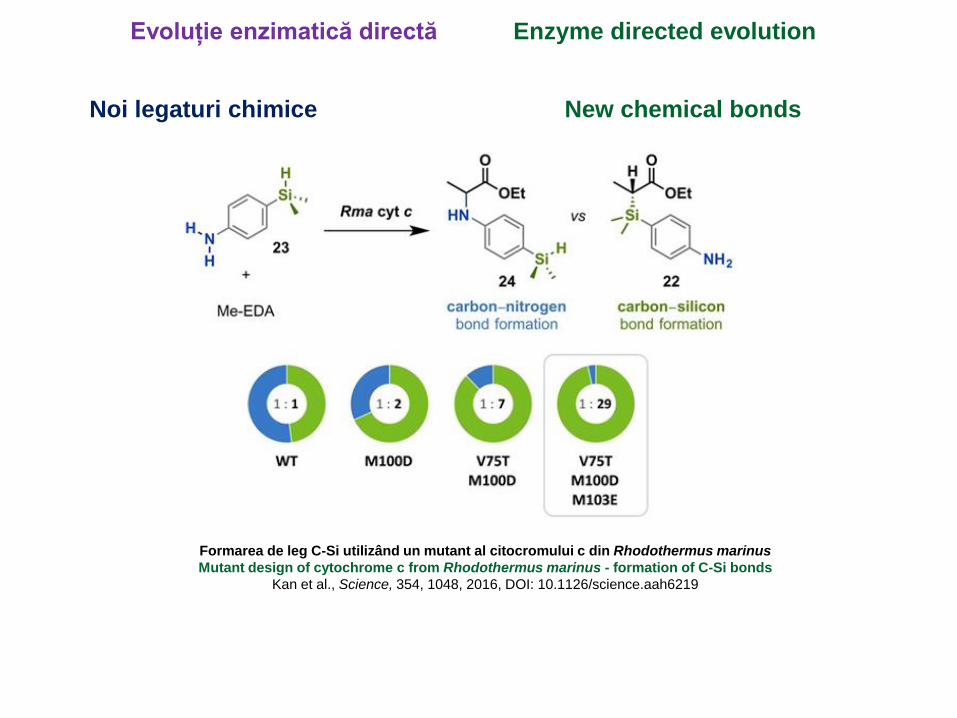
Formarea de leg C-Si utilizând un mutant al citocromului c din Rhodothermus marinus
Mutant design of cytochrome c from Rhodothermus marinus - formation of C-Si bonds
Kan et al., Science, 354, 1048, 2016, DOI: 10.1126/science.aah6219
Evoluție enzimatică directă Enzyme directed evolution
Noi legaturi chimice New chemical bonds
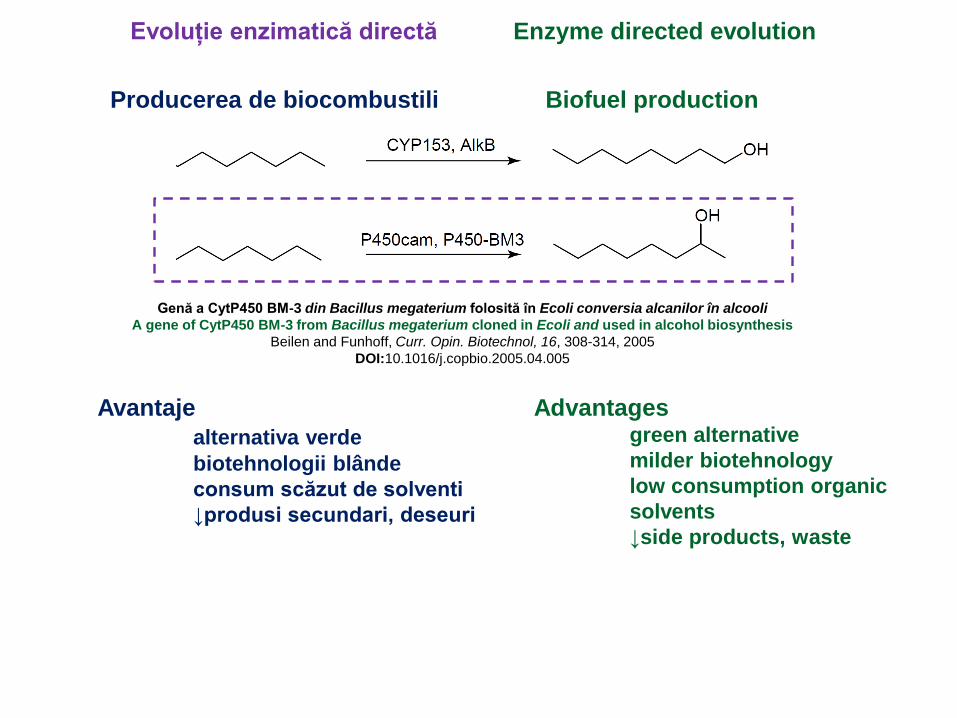
Genă a CytP450 BM-3 din Bacillus megaterium folosită în Ecoli conversia alcanilor în alcooli
A gene of CytP450 BM-3 from Bacillus megaterium cloned in Ecoli and used in alcohol biosynthesis
Beilen and Funhoff, Curr. Opin. Biotechnol, 16, 308-314, 2005
DOI:10.1016/j.copbio.2005.04.005
Evoluție enzimatică directă Enzyme directed evolution
Producerea de biocombustili Biofuel production
Avantaje
alternativa verde
biotehnologii blânde
consum scăzut de solventi
↓produsi secundari, deseuri
Advantagesgreen alternative
milder biotehnology
low consumption organic
solvents
↓side products, waste
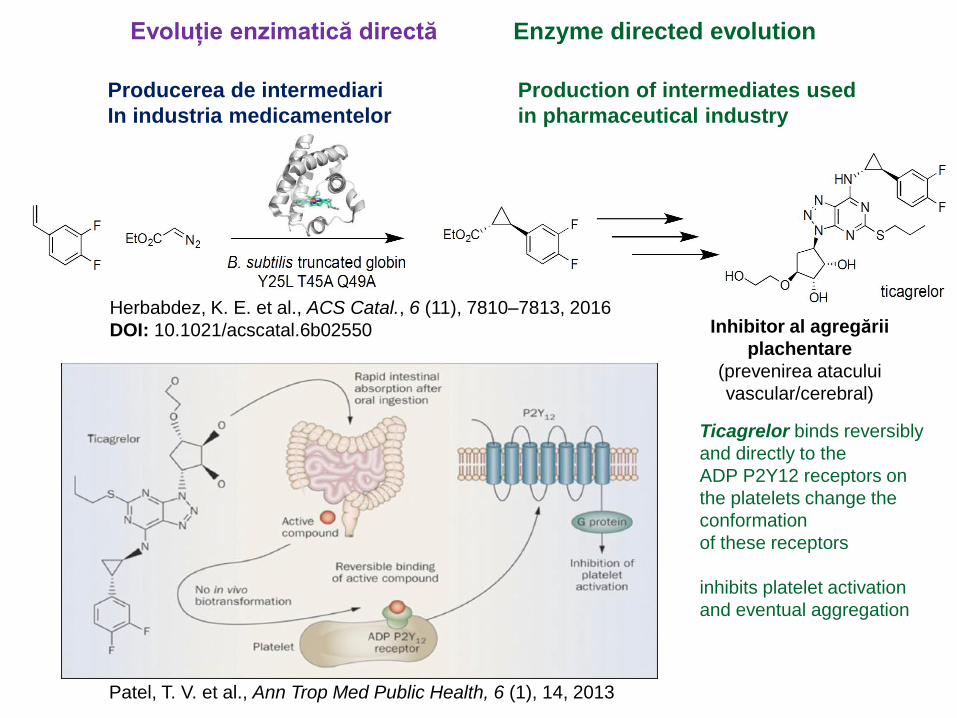
Herbabdez, K. E. et al., ACS Catal., 6 (11), 7810–7813, 2016
DOI: 10.1021/acscatal.6b02550 Inhibitor al agregării
plachentare
(prevenirea atacului
vascular/cerebral)
Evoluție enzimatică directă Enzyme directed evolution
Producerea de intermediari
In industria medicamentelor
Production of intermediates used
in pharmaceutical industry
Patel, T. V. et al., Ann Trop Med Public Health, 6 (1), 14, 2013
Ticagrelor binds reversibly
and directly to the
ADP P2Y12 receptors on
the platelets change the
conformation
of these receptors
inhibits platelet activation
and eventual aggregation
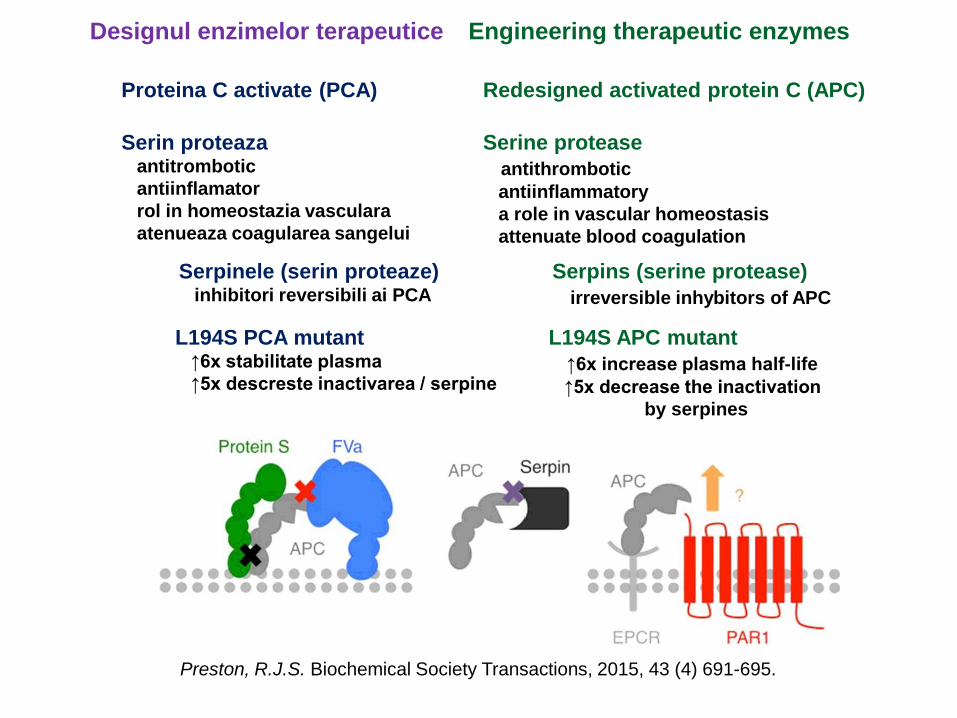
Designul enzimelor terapeutice Engineering therapeutic enzymes
Proteina C activate (PCA) Redesigned activated protein C (APC)
Serin proteazaantitrombotic
antiinflamator
rol in homeostazia vasculara
atenueaza coagularea sangelui
Serine protease
antithrombotic
antiinflammatory
a role in vascular homeostasis
attenuate blood coagulation
Serpinele (serin proteaze)inhibitori reversibili ai PCA
Serpins (serine protease)
irreversible inhybitors of APC
Preston, R.J.S. Biochemical Society Transactions, 2015, 43 (4) 691-695.
L194S PCA mutant↑6x stabilitate plasma
↑5x descreste inactivarea / serpine
L194S APC mutant
↑6x increase plasma half-life
↑5x decrease the inactivation
by serpines
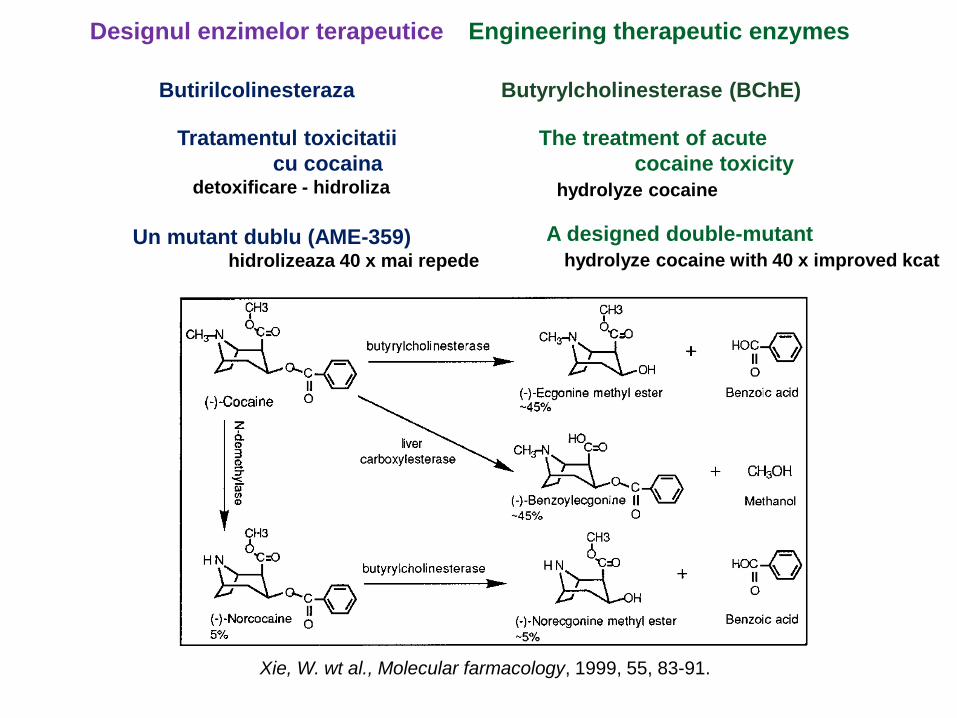
Designul enzimelor terapeutice Engineering therapeutic enzymes
Butirilcolinesteraza Butyrylcholinesterase (BChE)
Tratamentul toxicitatii
cu cocainadetoxificare - hidroliza
The treatment of acute
cocaine toxicity
hydrolyze cocaine
Un mutant dublu (AME-359)hidrolizeaza 40 x mai repede
A designed double-mutant
hydrolyze cocaine with 40 x improved kcat
Xie, W. wt al., Molecular farmacology, 1999, 55, 83-91.
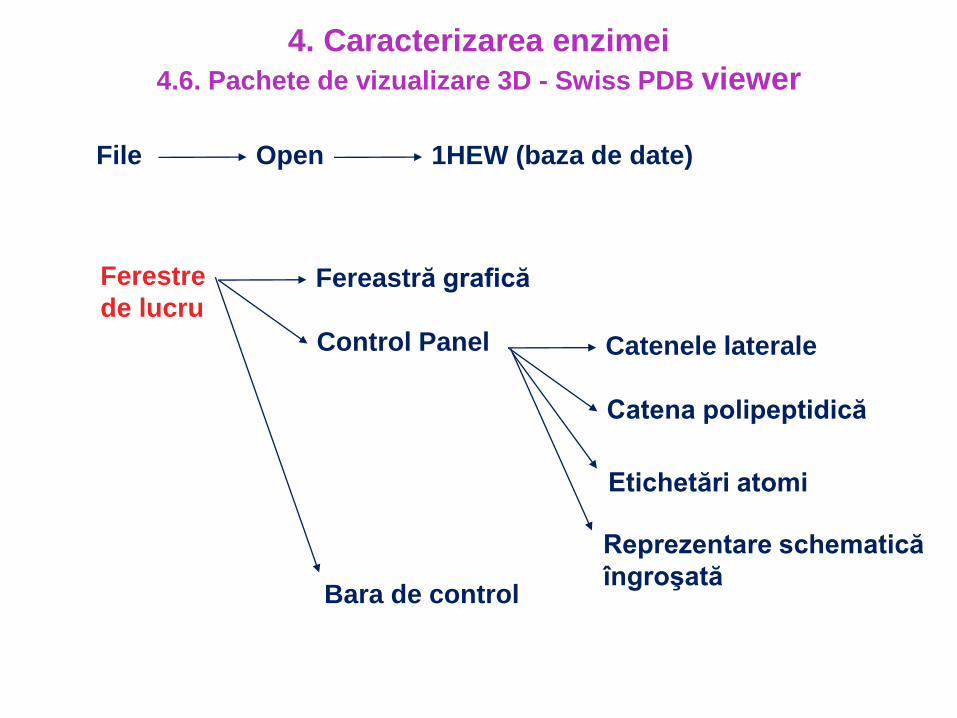
4. Caracterizarea enzimei
4.6. Pachete de vizualizare 3D - Swiss PDB viewer
File Open 1HEW (baza de date)
Ferestre
de lucru Fereastră grafică
Control Panel Catenele laterale
Catena polipeptidică
Etichetări atomi
Reprezentare schematică
îngroşatăBara de control
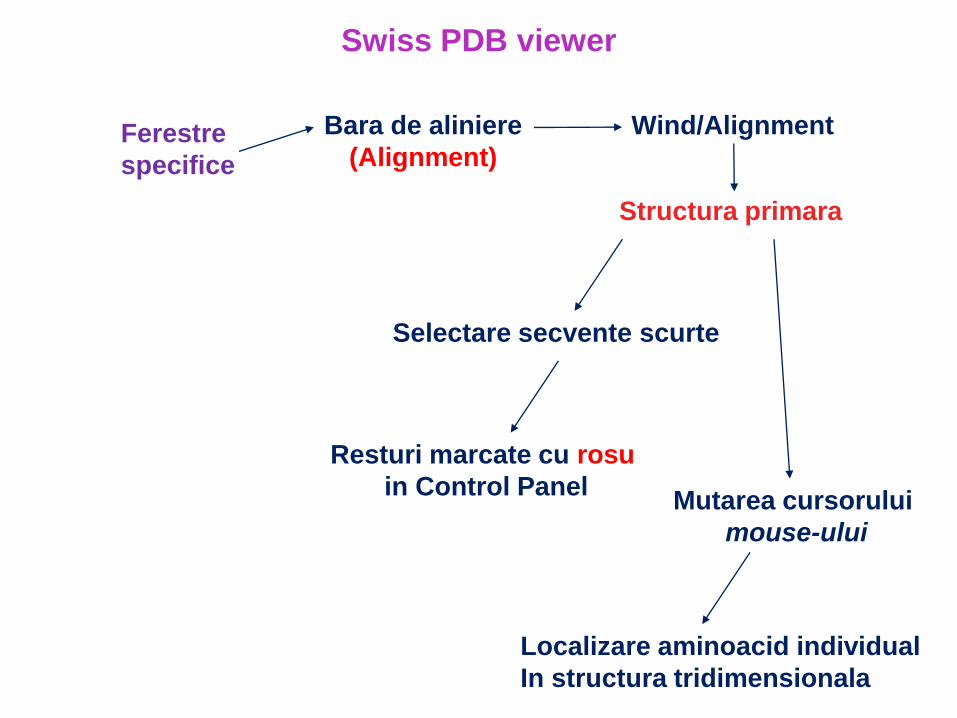
Bara de aliniere
(Alignment)Ferestre
specifice
Wind/Alignment
Structura primara
Swiss PDB viewer
Selectare secvente scurte
Resturi marcate cu rosu
in Control PanelMutarea cursorului
mouse-ului
Localizare aminoacid individual
In structura tridimensionala
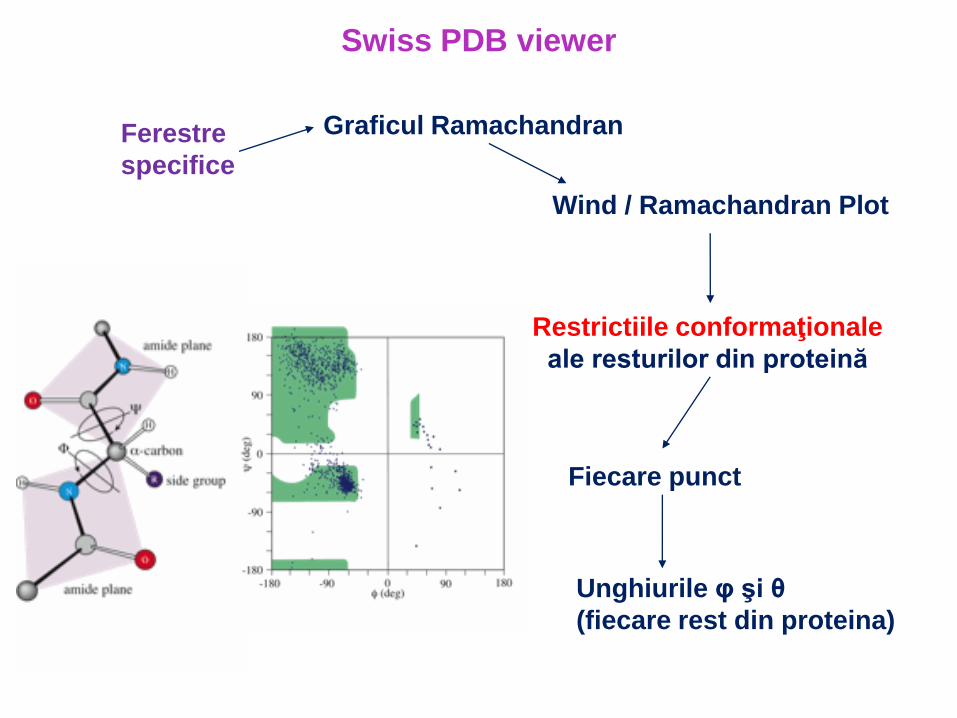
Graficul RamachandranFerestre
specifice
Wind / Ramachandran Plot
Swiss PDB viewer
Restrictiile conformaţionale
ale resturilor din proteină
Fiecare punct
Unghiurile φ şi θ
(fiecare rest din proteina)
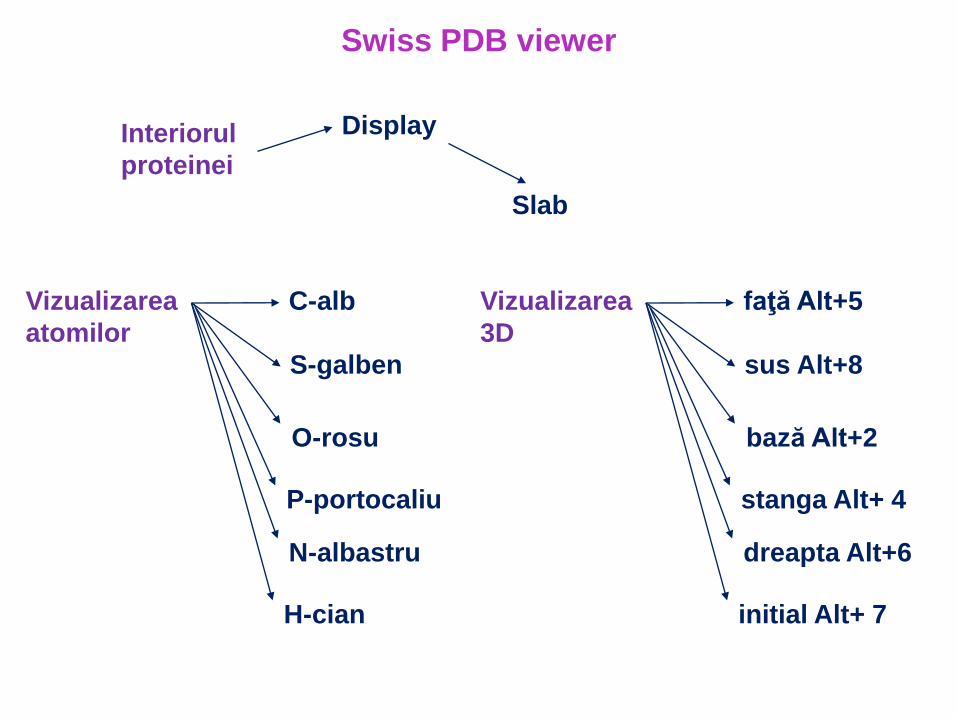
DisplayInteriorul
proteinei
Slab
Swiss PDB viewer
Vizualizarea
atomilor
C-alb
S-galben
O-rosu
P-portocaliu
N-albastru
H-cian
Vizualizarea
3D
faţă Alt+5
sus Alt+8
bază Alt+2
stanga Alt+ 4
dreapta Alt+6
initial Alt+ 7
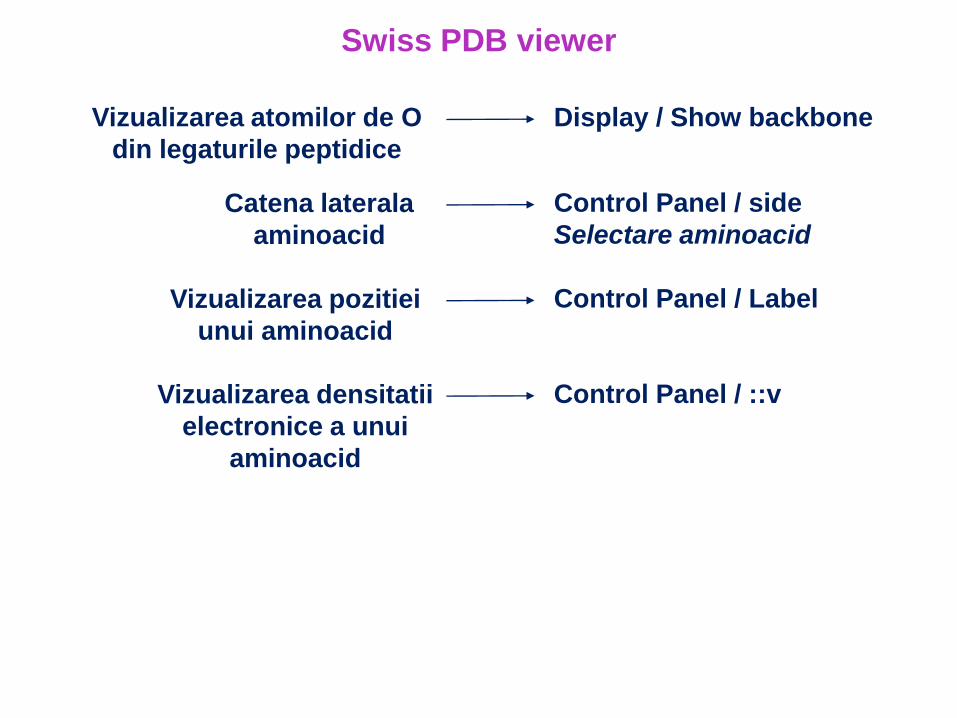
Swiss PDB viewer
Vizualizarea atomilor de O
din legaturile peptidice
Display / Show backbone
Catena laterala
aminoacid
Control Panel / side
Selectare aminoacid
Vizualizarea pozitiei
unui aminoacid
Control Panel / Label
Vizualizarea densitatii
electronice a unui
aminoacid
Control Panel / ::v


Motivele structurale
4. Caracterizarea enzimei4.4. Determinarea structurii secundare/supersecundare
Scop
http://scop2.mrc-lmb.cam.ac.uk/0.html
Relatiile dintre proteine
Clase structurale
Tipuri de proteine
Elemente de evolutie structurala
Clase structurale
Impachetarea proteinelor in
functie de continutul
structurii secundare
Motivele structurale
4. Caracterizarea enzimei4.4. Determinarea structurii secundare/supersecundare
Scop
Protein Relationships
Other Relationships
Motif
CGxCxxCx(n)C motiv legare ion metalic
N - terminal ADH
Motivul Rosmann element de conexiune în buclă
GX[GS]XX[GSTA]
GS - contact cu pirofosfat (nucleotidă)

Motivele structurale
4. Caracterizarea enzimei4.4. Determinarea structurii secundare/supersecundare
MOTIF Search
http://www.genome.jp/tools/motif/

Motivele structurale
MOTIF Search http://www.genome.jp/tools/motif/
pepsinogen A
MKWLLLLGLVALSECIMYKVPLIRKKSFRRTLSERGLLKDFLKKHNLNPARKY
FPQWKAPTLVDEQPLEN YLDMEYFGTIGIGTPAQDFTVLFDTGSSNLWVPSV
YCSSLACTNHNRFNPEDSSTYQSTSETVSITYGTG SMTGILGYDTVQVGGISD
TNQIFGLSETEPGSFLYYAPFDGILGLAYPSISSSGATPVFDNIWNQGLVSQ DL
FSVYLSADDQSGSVVIFGGIDSSYYTGSLNWVPVTVEGYWQITVDSITMNGEA
IACAEGCQAIVDTGT SLLTGPTSPIANIQSDIGASENSDGDMVVSCSAISSLPDI
VFTINGVQYPVPPSAYILQSEGSCISGFQG MNLPTESGELWILGDVFIRQYFTV
FERANNQVGLAPVA

Motivele structurale
MOTIF Search
HUMAN Tyrosine-protein phosphatase non-receptor type 6 Homo sapiens MVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDF
YDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQA
ETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGG
LETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYYATRVNAADIENRVLELNKKQESEDTA
KAGFWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSD
YINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNK
CVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDH
GVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDC
DIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQFIETTKKKLEVLQSQKGQESEYGNIT
YPPAMKNAHAKASRTSSKHKEDVYENLHTKNKREEKVKKQRSADKEKSKGSLKR
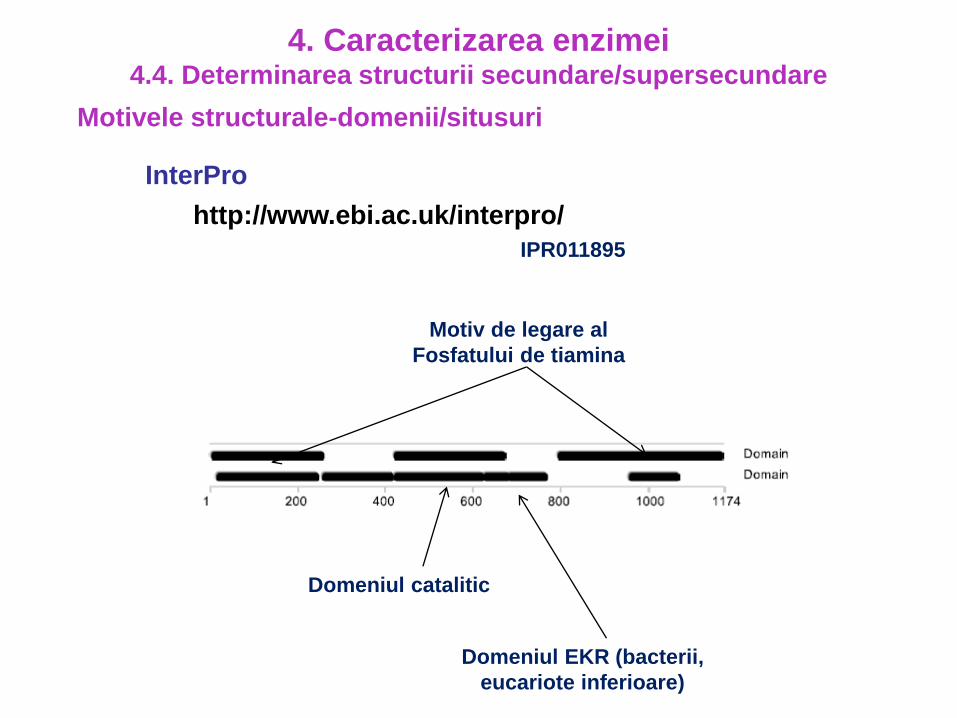
Motivele structurale-domenii/situsuri
4. Caracterizarea enzimei4.4. Determinarea structurii secundare/supersecundare
InterPro
http://www.ebi.ac.uk/interpro/
IPR011895
Domeniul catalitic
Motiv de legare al
Fosfatului de tiamina
Domeniul EKR (bacterii,
eucariote inferioare)
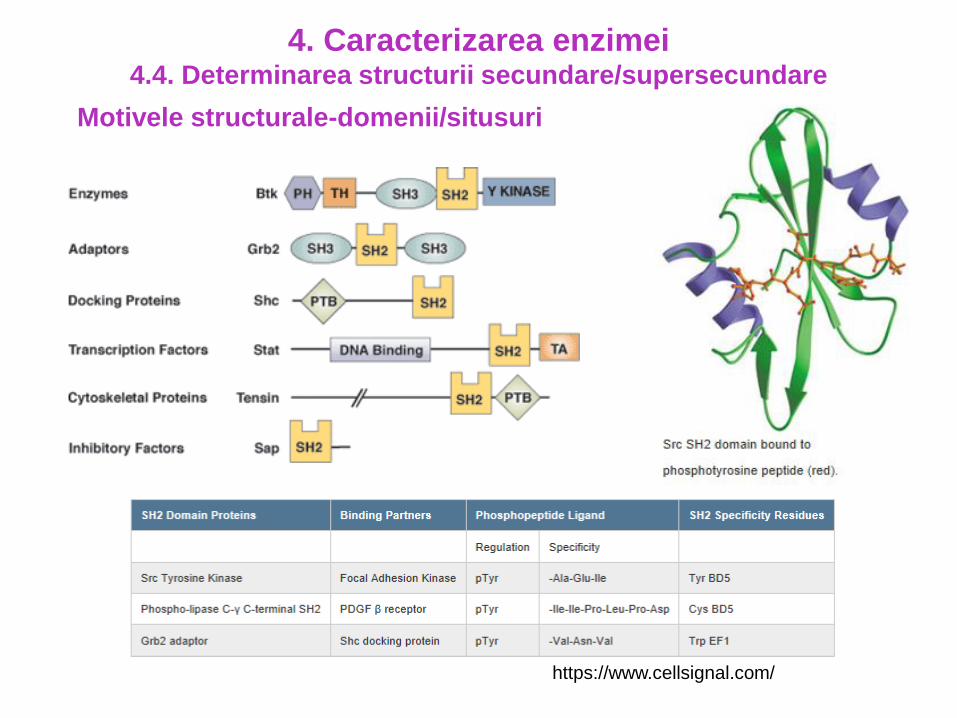
Motivele structurale-domenii/situsuri
4. Caracterizarea enzimei4.4. Determinarea structurii secundare/supersecundare
https://www.cellsignal.com/

Motivele structurale-domenii/situsuri
4. Caracterizarea enzimei4. 4. Determinarea structurii secundare/supersecundare
CATH
http://www.cathdb.info/
95 milioane de domenii proteice
6,199 superfamilii
Superfamilia
3.30.70.330Domeniul
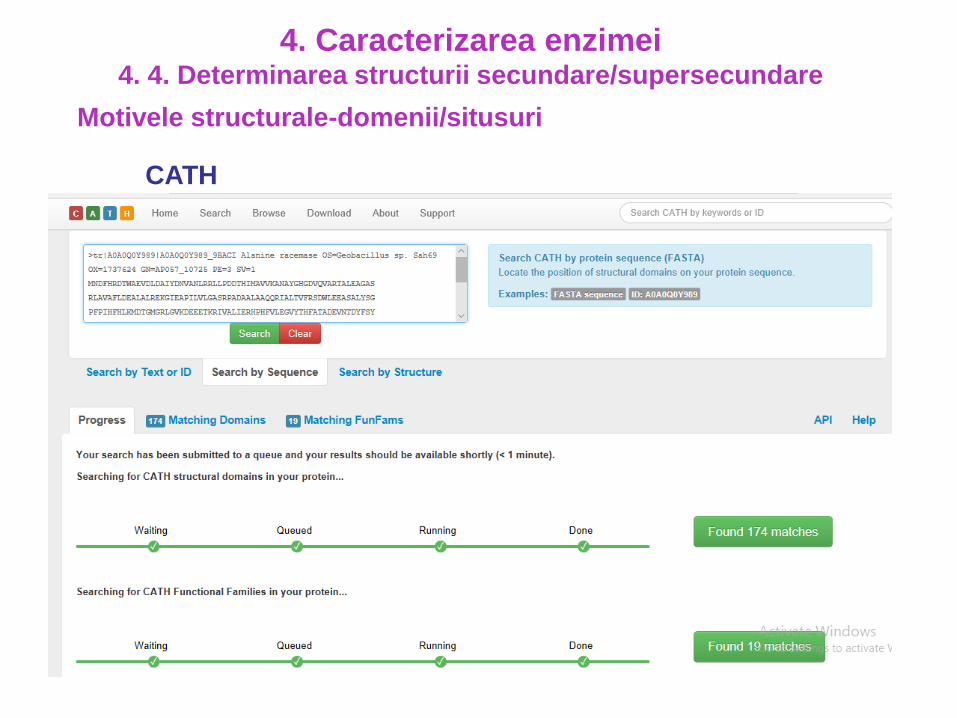
Motivele structurale-domenii/situsuri
4. Caracterizarea enzimei4. 4. Determinarea structurii secundare/supersecundare
CATH
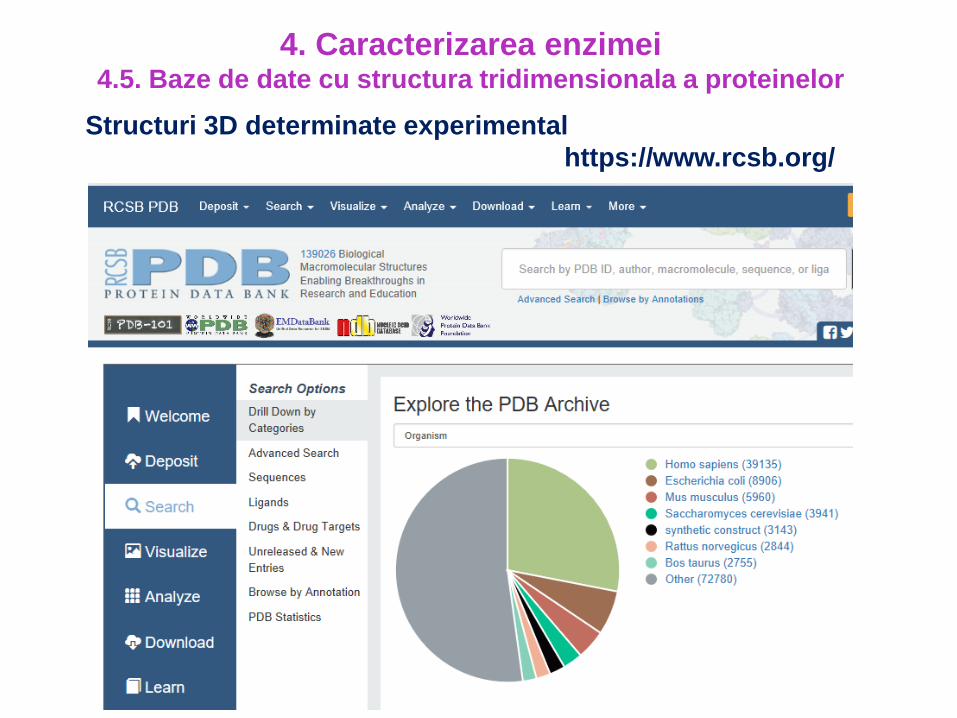
Structuri 3D determinate experimental
https://www.rcsb.org/
4. Caracterizarea enzimei4.5. Baze de date cu structura tridimensionala a proteinelor
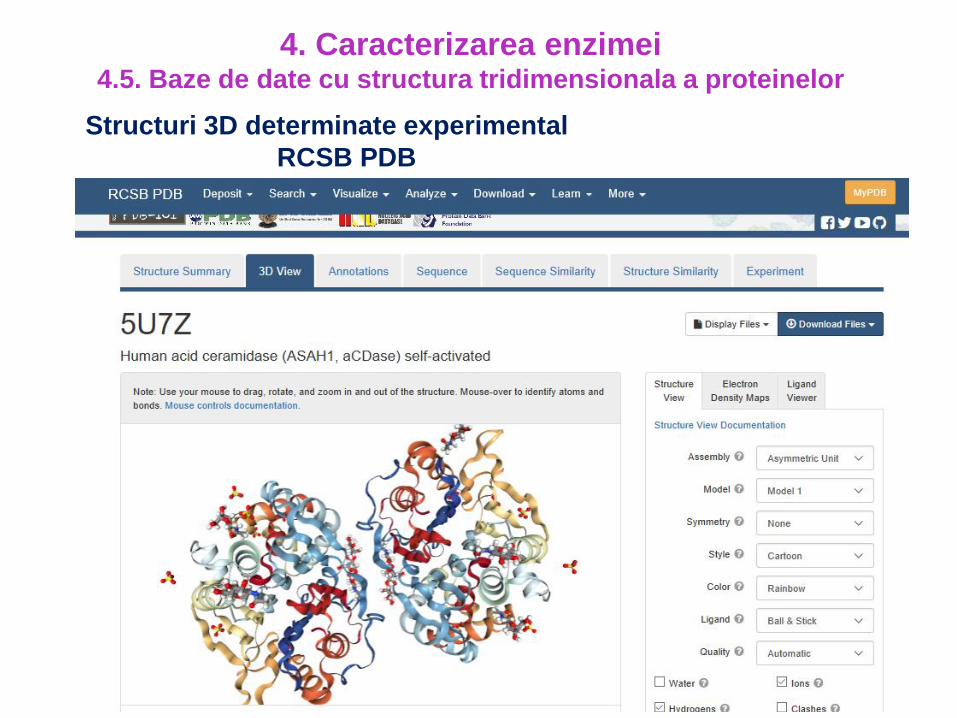
Structuri 3D determinate experimental
RCSB PDB
4. Caracterizarea enzimei4.5. Baze de date cu structura tridimensionala a proteinelor

http://www.expasy.org/proteomics
Expert Protein Analysis System ExPASy
4. Caracterizarea enzimei4.5. Baze de date cu structura tridimensionala a proteinelor
https://www.proteinmodelportal.org/

http://www.uniprot.org/ (pir.georgetown.edu)
5 milioane proteine – conexiune cu alte str 3D
UniProt
4. Caracterizarea enzimei4.5. Baze de date cu structura tridimensionala a proteinelor
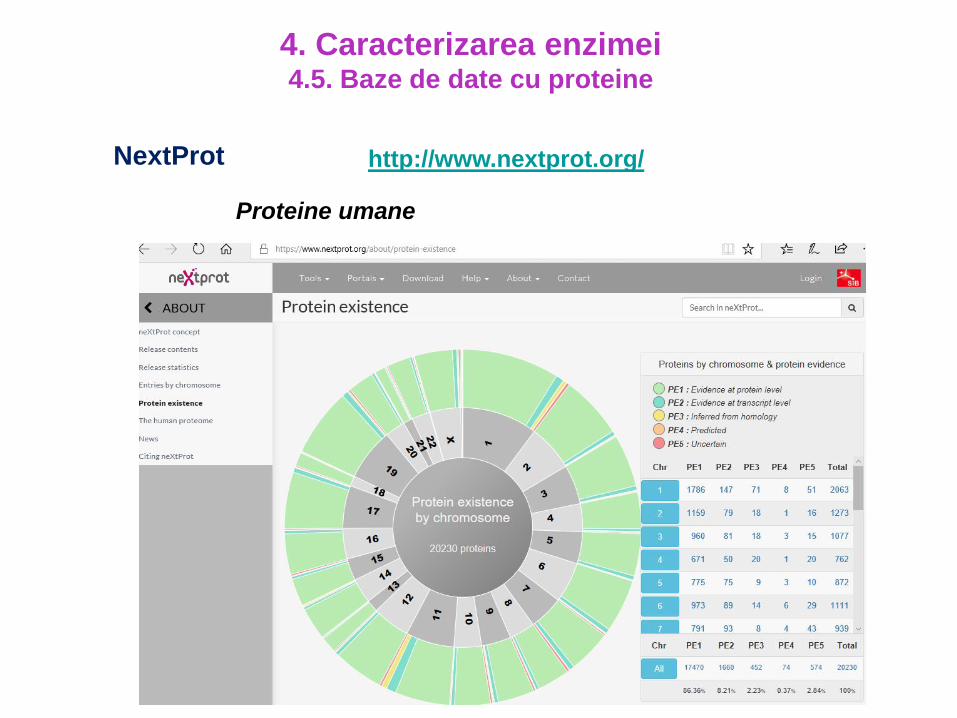
http://www.nextprot.org/
Proteine umane
NextProt
4. Caracterizarea enzimei4.5. Baze de date cu proteine
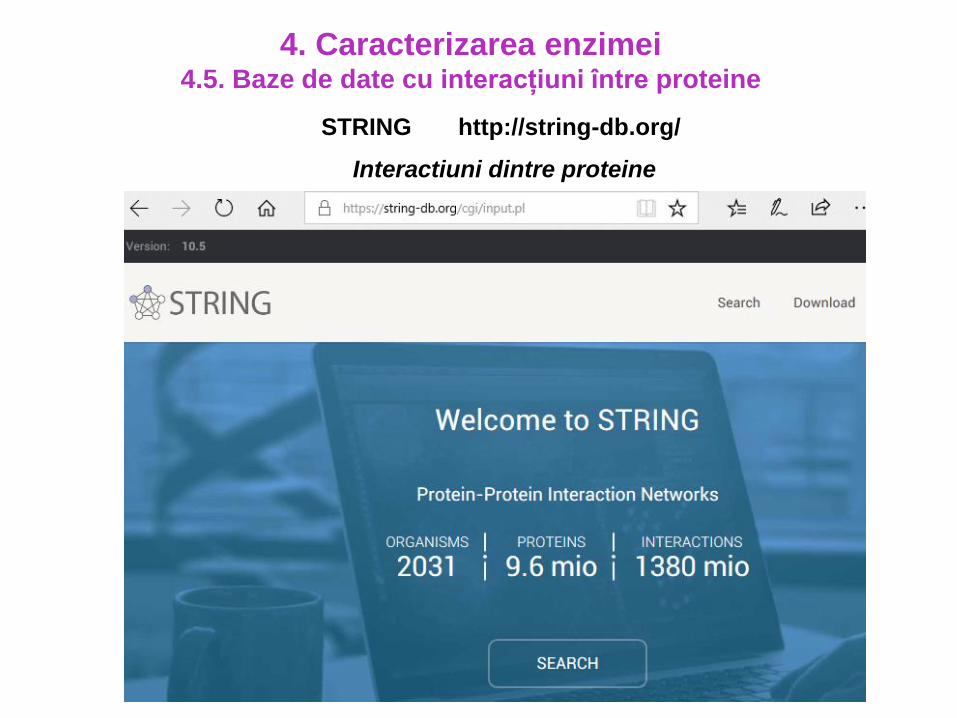
STRING http://string-db.org/
Interactiuni dintre proteine
4. Caracterizarea enzimei4.5. Baze de date cu interacțiuni între proteine

Comparari structuri
3D proteine
Dali
http://ekhidna.biocenter.
helsinki.fi/dali/
4. Caracterizarea
enzimei

Entrez Databases
http://www.ncbi.nlm.nih.gov/Class/MLACourse/Original8Hour/Entrez/
4. Caracterizarea enzimei
4.5. Baze de date cu structura tridimensionala a
proteinelor


Baze de date cu nomenclatura enzimelor
UniProtKB/Swiss-Prot http://enzyme.expasy.org/







![MANUAL DE INSTRUC}IUNI · Respecta]i toate regurile de prevenire a accidentelor. Produsul trebuie fixat bine pentru a rezista la ridic\ri [i manipulari. Unele pompe electrice au beciuge](https://static.fdocument.org/doc/165x107/5e36cbc7c380f77b2b623af7/manual-de-instruciuni-respectai-toate-regurile-de-prevenire-a-accidentelor-produsul.jpg)








