Nuclear targeting of β-catenin and p120ctn during thrombin ... 6.pdfisolation, cDNA synthesis and...
22
Nuclear targeting of β-catenin and p120ctn during thrombin-induced endothelial barrier dysfunction. Cora M.L. Beckers 1 , Juan J. García-Vallejo 2 , Victor W.M. van Hinsbergh 1 and Geerten P. van Nieuw Amerongen 1 6 VU University medical center, Institute for Cardiovascular Research, Department for Physiology 1 , Department of Molecular Cell Biology & Immunology 2 , Amsterdam, the Netherlands. Cardiovascular Research, 2008 Abstract Cytosolic and nuclear localization of β-catenin was observed in leaky vessels and in tumors. Several lines of evidence indicate that nuclear β- catenin facilitates angiogenesis. We hypothesized, that nuclear β-catenin liberated from endothelial junctional complexes marks the transition from hyperpermeability to angiogenesis. The aim of this study was therefore, to investigate the fate of β-catenin and the related catenin p120ctn, during disruption of the endothelial barrier function in human umbilical vein endothelial cells. The hyperpermeability-inducer thrombin caused a Rho kinase dependent redistribution of β-catenin from the membrane to the cytosol as evidenced by western blot analysis of membrane and cytosol fractions and by immunohistochemistry. Glycogen synthase kinase 3β, which phosphorylates cytosolic β-catenin and thereby facilitates its proteasomal degradation, was inhibited by thrombin. Analysis of nuclear extracts demonstrated a thrombin- induced nuclear accumulation of β-catenin as well as p120ctn. Thrombin stimulation activated β-catenin-mediated transcriptional activity as evidenced by reporter assays. Finally, real time-PCR revealed increased mRNA levels of several β-catenin target genes. Thrombin induced a cytosolic stabilization of membrane-liberated β-catenin, which, together with p120ctn, subsequently translocated to the nucleus where it induces several β-catenin target genes. This supports the suggestion that membrane-liberated β-catenin and p120ctn contribute to angiogenic responses of endothelial cells following episodes of vascular leakage. 121
Transcript of Nuclear targeting of β-catenin and p120ctn during thrombin ... 6.pdfisolation, cDNA synthesis and...

Nuclear targeting of β-catenin and p120ctn during thrombin-induced endothelial barrier dysfunction.
Cora M.L. Beckers1, Juan J. García-Vallejo2, Victor W.M. van Hinsbergh1 and Geerten P. van Nieuw Amerongen1
6 VU University medical center, Institute for Cardiovascular Research, Department for Physiology1, Department of Molecular Cell Biology & Immunology2, Amsterdam, the Netherlands. Cardiovascular Research, 2008 Abstract Cytosolic and nuclear localization of β-catenin was observed in leaky vessels and in tumors. Several lines of evidence indicate that nuclear β-catenin facilitates angiogenesis. We hypothesized, that nuclear β-catenin liberated from endothelial junctional complexes marks the transition from hyperpermeability to angiogenesis. The aim of this study was therefore, to investigate the fate of β-catenin and the related catenin p120ctn, during disruption of the endothelial barrier function in human umbilical vein endothelial cells. The hyperpermeability-inducer thrombin caused a Rho kinase dependent redistribution of β-catenin from the membrane to the cytosol as evidenced by western blot analysis of membrane and cytosol fractions and by immunohistochemistry. Glycogen synthase kinase 3β, which phosphorylates cytosolic β-catenin and thereby facilitates its proteasomal degradation, was inhibited by thrombin. Analysis of nuclear extracts demonstrated a thrombin-induced nuclear accumulation of β-catenin as well as p120ctn. Thrombin stimulation activated β-catenin-mediated transcriptional activity as evidenced by reporter assays. Finally, real time-PCR revealed increased mRNA levels of several β-catenin target genes. Thrombin induced a cytosolic stabilization of membrane-liberated β-catenin, which, together with p120ctn, subsequently translocated to the nucleus where it induces several β-catenin target genes. This supports the suggestion that membrane-liberated β-catenin and p120ctn contribute to angiogenic responses of endothelial cells following episodes of vascular leakage.
121

Nuclear targeting of β-catenin
Introduction The endothelial cells (ECs) form a barrier between the blood and the surrounding tissue and actively control the exchange of nutrients and proteins between these compartments.1, 2 During inflammation or exposure to ischemia and other angiogenic stimuli the endothelial cell junctions become loosened, which might result in the generation of focal intercellular gaps that facilitate paracellular passage of fluids, solutes, and proteins. Subsequent to loosening of the cell junctions ECs can become involved in angiogenesis. In most types of ECs the adherens junctions (AJs) play a dominant role in the endothelial barrier function.2 The disassembly of junctional complexes of the AJs is accompanied by liberation of β-catenin.3 Interestingly, β-catenin can also induce transcriptional activation in the nucleus of various types of cells.4, 5 In Caenorhabditis elegans, the adhesion and the signaling function of β-catenin are performed by two different β-catenin homologues, leaving the possibility that separate pools of β-catenin exist in the mammalian system.6 To date, it is unknown whether disintegration of AJs in ECs is accompanied by nuclear translocation of β-catenin to the nucleus.7 The intercellular bridges in the AJs of neighboring ECs are formed by homotypic interactions of the transmembrane protein vascular endothelial (VE)-cadherin.8, 9 β-Catenin participates in the adhesion complex by connecting the F-actin cytoskeleton to VE-cadherin, an interaction that also involves α-catenin.10, 11 A third catenin, p120catenin (p120ctn), of which four isoforms have been identified, is also present in AJs of ECs and stabilizes the adherens junction complex.12 β-Catenin is also a key player in the Wnt signaling pathway. Here it drives gene transcription upon nuclear translocation.4, 13 Cytosolic β-catenin is constitutively bound to a destruction complex containing axin and adenomatosis poliposis coli (APC) as well as casein kinase (CK)-1 and glycogen synthase kinase (GSK)-3β. The latter two kinases phosphorylate β-catenin, thereby inactivating it and targeting it for ubiquitination and subsequent degradation by proteasomes.14 Due to this rapid degradation, there are very low levels of non-phosphorylated (active) β-catenin in the cytosol under normal non-stimulated conditions.15, 16 In the presence of Wnt signaling and as a response to growth factor signaling, cytosolic β-catenin phosphorylation and degradation is prevented via inhibition of GSK3β.17 GSK3β phosphorylation can be stimulated in human umbilical vein endothelial cells (HUVECs).18 In other cells it has been shown that
122

Nuclear targeting of β-catenin
stabilization of β-catenin enables β-catenin to translocate to the nucleus and to induce transcription.19 It is still unknown how β-catenin shuttles between the cytosol and nucleus,7 but once β-catenin accumulates in the nucleus it serves as a co-transcription factor to enhance T cell factor / Leukocyte Enhancing Factor (Tcf/LEF) inducible gene transcription.4, 20 Among β-catenin target genes that have been linked to cancer and angiogenesis are c-myc,21 cyclinD1,22, 23 cox-2,24, 25 IL-826, 26 and endothelin-1.27 Evidence is accumulating that β-catenin-mediated gene transcription only occurs if p120ctn binds, and thereby inhibits, the β-catenin target gene repressor Kaiso.28, 29 β-Catenin is an important factor in the development of organs, and in angiogenesis.30 In the adult, angiogenesis is mainly associated with pathological conditions, such as inflammation and ischemia.31 Blankesteijn et al.16 reported that β-catenin accumulated in the cytosol, and partially in the nucleus of vascular cells, after myocardial ischemia. In contrast, cells of normal adult vasculature rarely accumulate β-catenin in the cytoplasm or nucleus.15, 16 Vascular leakage and angiogenesis are closely related processes.32 Disassembly of the AJs is a requirement for angiogenesis and remodeling of the vascular bed.33 The liberated β-catenin might provide a pool for nuclear translocation and subsequent gene transcription,4, 7, 30 and thus contribute to angiogenesis. To clarify whether in HUVECs junction disassembly and nuclear accumulation of β-catenin reflect one pool of β-catenin, we studied whether upon stimulation by thrombin, which induces junction disintegration,34 membrane-localized β-catenin and p120ctn can translocate and stabilize in the cytosol. Subsequently, we investigated whether stabilization of β-catenin was accompanied by translocation to the nucleus and whether this relocalisation activated the β-catenin reporter Topflash and enhanced β-catenin target gene expression. Materials & Methods Sources of reagents can be found in the expanded Materials and Methods section in the online data supplement (CD chapter 6). Human umbilical vein endothelial cells (HUVECs) were cultured as previously described35 and transfected with the TCF reporter constructs Topflash and Fopflash using the Amaxa technology. Cytosol and membrane fractions were obtained as described by Kouklis et al.,36 while nuclear fractions were obtained as described by Nilsson et al.37 For quantification of Western blots AIDA Image
123

Nuclear targeting of β-catenin
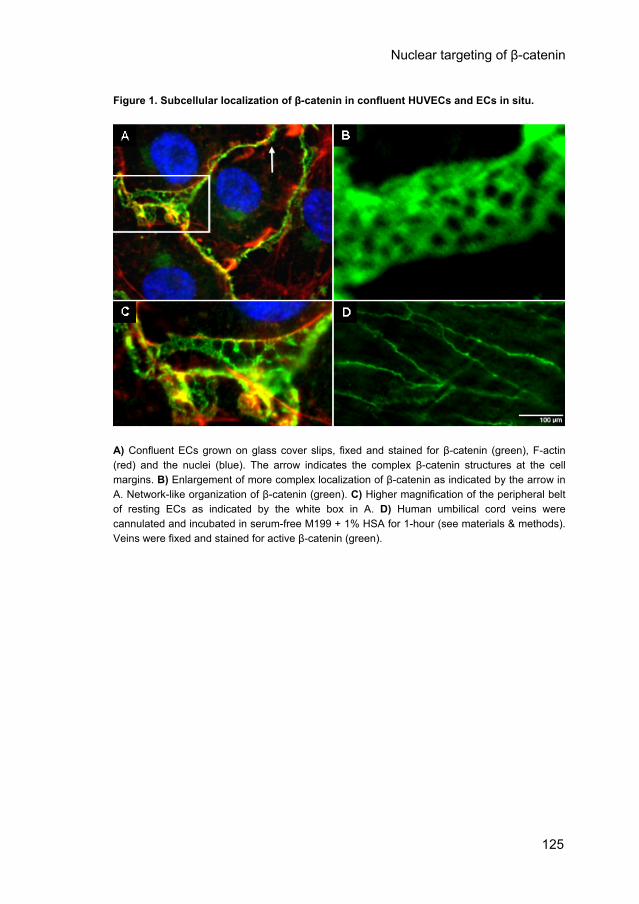
Analyzer software was used. 3D-Digital fluorescence imaging microscopy was performed with a Zeiss Axiovert 200 MarianasTM inverted microscope under the control of Slidebook software as described before.38 mRNA isolation, cDNA synthesis and real time-PCR were performed as previously described.39 Primers are described in table 1 in the Materials and Methods section of the online data supplement (CD chapter 6). Data were compared by paired sample t-tests. Probability values of less than 0.05 were considered to be significant. Results Subcellular localization of β-catenin in confluent HUVECs and ECs in situ. To investigate the fate of β-catenin before and after stimulation of endothelial permeability, we determined the subcellular distribution of β-catenin in confluent HUVECs, which were counter-stained for F-actin. Using a general β-catenin antibody (Sigma C2206), β-catenin was encountered at the cell margins, where it formed a continuous lining (Fig. 1A, green). A complex staining pattern of β-catenin was locally observed (Fig. 1A, arrow). Detailed analysis of such regions revealed a network-like structure (Fig. 1B) that co-localized with VE-cadherin (data not shown). Furthermore, at specific sites (box in Fig. 1A, enlarged in Fig. 1C) β-catenin was curled around actin strands of the peripheral belt of F-actin filaments. Under basal conditions β-catenin was not detectable in the nucleus, and only tiny amounts of immunostaining were present in the cytosol. We verified, in freshly isolated umbilical cord veins in situ, the presence of β-catenin by using the general β-catenin antibody and the antibody αABC that recognizes active β-catenin, as shown for the latter in figure 1D. In both cases β-catenin was mainly located at the cell-margins of the ECs. 3D-deconvolution microscopy combined with 3D-reconstruction confirmed the network-like structure of β-catenin at the cell-margins in situ (data not shown).
124
Nuclear targeting of β-catenin
Figure 1. Subcellular localization of β-catenin in confluent HUVECs and ECs in situ.
A) Confluent ECs grown on glass cover slips, fixed and stained for β-catenin (green), F-actin (red) and the nuclei (blue). The arrow indicates the complex β-catenin structures at the cell margins. B) Enlargement of more complex localization of β-catenin as indicated by the arrow in A. Network-like organization of β-catenin (green). C) Higher magnification of the peripheral belt of resting ECs as indicated by the white box in A. D) Human umbilical cord veins were cannulated and incubated in serum-free M199 + 1% HSA for 1-hour (see materials & methods). Veins were fixed and stained for active β-catenin (green).
125

Nuclear targeting of β-catenin
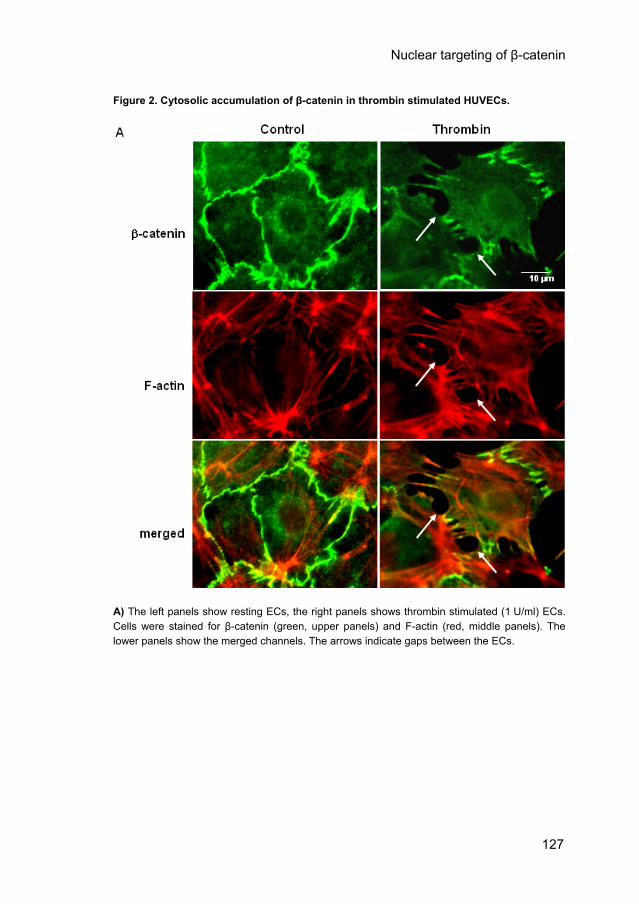
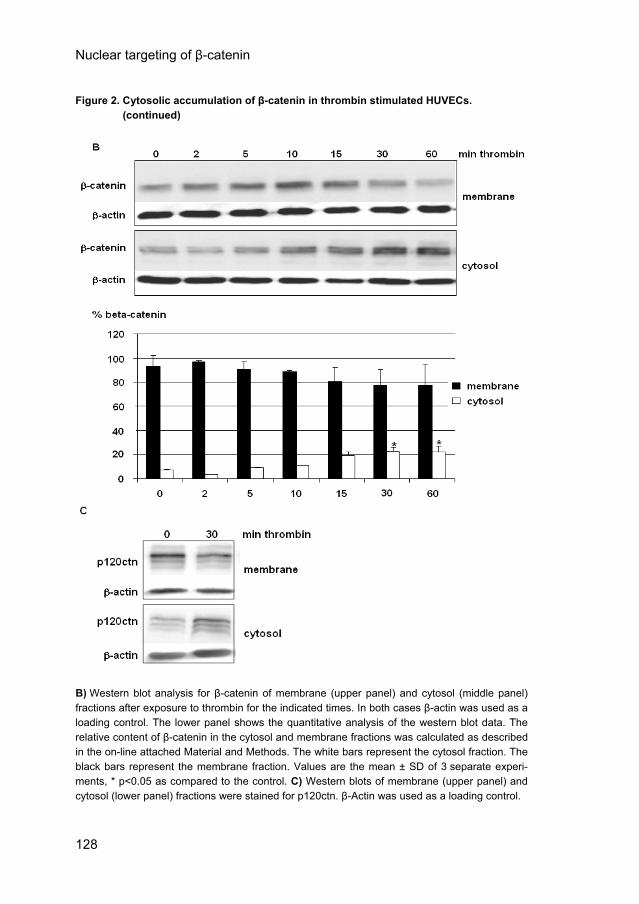
Cytosolic accumulation of β-catenin in thrombin-stimulated HUVECs. To investigate whether β-catenin re-distributes during hyperpermeability responses, confluent HUVECs were stimulated by thrombin. Thirty minutes after stimulation by thrombin (1 U/ml) β-catenin (green) was present at the end of the stress fibers (red), but had disappeared at sites where gaps between the ECs had been formed (Fig. 2A; arrows mark inter-endothelial gaps). In the cytosol β-catenin did no longer co-localize with VE-cadherin (data not shown). Subsequently, we determined whether β-catenin re-distributed from the membrane to the cytosol during thrombin stimulation of confluent HUVECs by quantifying the amounts of β-catenin in isolated membrane and cytosol fractions (Fig. 2B). Western blot analysis demonstrated that in untreated cells 93 ± 9% (n=3) of the β-catenin is present in the membrane and 7 ± 1% (n=3) in the cytosol (Fig. 2B, first lane). During a 1-hour stimulation by thrombin the amount of β-catenin in the membrane fraction slowly decreased, while it increased simultaneously in the cytosol fraction. The cytosolic β-catenin was significantly increased after stimulation with thrombin for 30 and 60 minutes (from 7 ± 1% to 22 ± 4% and 22 ± 5%, respectively, n=3 p<0.05) as compared to the untreated cells (Fig. 2B). Similarly, p120ctn increased in the cytosol after thrombin stimulation (Fig. 2C). Concurrent with our previous observation that thrombin-induced hyperpermeability was inhibited by the Rho kinase inhibitor Y-27632, pretreatment of the ECs with Y-27632 inhibited thrombin-induced stress fiber and gap formation (supplementary Fig. IA, CD chapter 6), as well as the redistribution of β-catenin towards the cytosol of thrombin-stimulated ECs as shown by Western blot analysis (supplementary Fig. IB, CD chapter 6). While thrombin increased cytosolic β-catenin to 120% of control values, the β-catenin content of the cytosol dropped to 74% of control values in thrombin-stimulated cells that had been preincubated with Y27632. Thus, thrombin-induced β-catenin redistribution in confluent ECs is dependent on the activation of Rho kinase.
126
Nuclear targeting of β-catenin
Figure 2. Cytosolic accumulation of β-catenin in thrombin stimulated HUVECs.
A) The left panels show resting ECs, the right panels shows thrombin stimulated (1 U/ml) ECs. Cells were stained for β-catenin (green, upper panels) and F-actin (red, middle panels). The lower panels show the merged channels. The arrows indicate gaps between the ECs.
127
Nuclear targeting of β-catenin
Figure 2. Cytosolic accumulation of β-catenin in thrombin stimulated HUVECs. Figure 2. (continued)
B) Western blot analysis for β-catenin of membrane (upper panel) and cytosol (middle panel) fractions after exposure to thrombin for the indicated times. In both cases β-actin was used as a loading control. The lower panel shows the quantitative analysis of the western blot data. The relative content of β-catenin in the cytosol and membrane fractions was calculated as described in the on-line attached Material and Methods. The white bars represent the cytosol fraction. The black bars represent the membrane fraction. Values are the mean ± SD of 3 separate experi-ments, * p<0.05 as compared to the control. C) Western blots of membrane (upper panel) and cytosol (lower panel) fractions were stained for p120ctn. β-Actin was used as a loading control.
128

Nuclear targeting of β-catenin
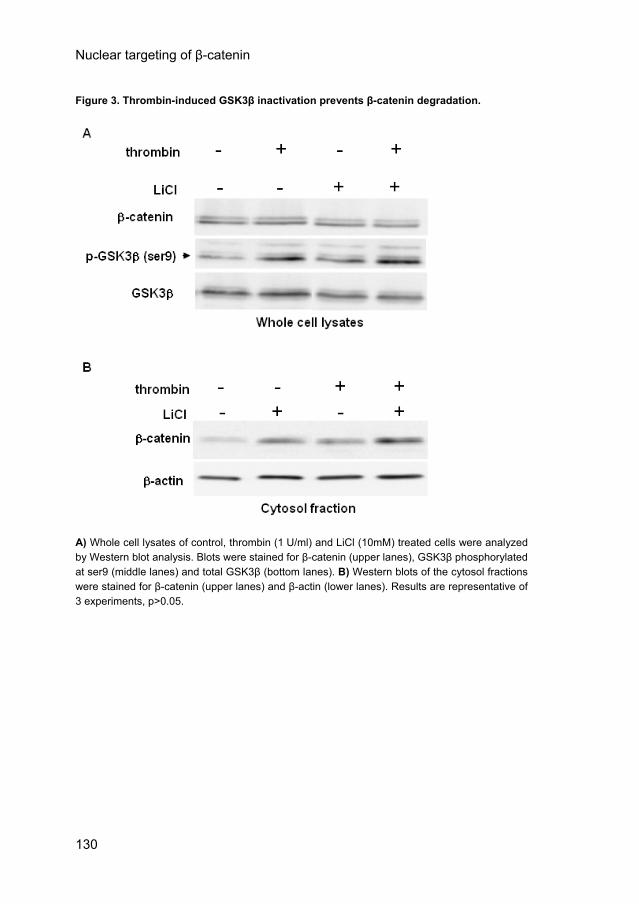
Thrombin-induced GSK3β inactivation prevents β-catenin degradation. The cytoplasmic β-catenin accumulation may be solely due to liberation of β-catenin from the adhesion complexes or additionally by protection against proteolytic degradation in the cytosol, which is facilitated by GSK3β phosphorylation of β-catenin. Thrombin may reduce this degradation by inhibitory phosphorylation of GSK3β on Ser9.18 Western blot analysis of whole cell lysates revealed that thrombin increased the phosphorylation of GSK3β on Ser9 in confluent ECs as compared to their control counterparts (Fig. 3A, middle panel). Simultaneously, β-catenin levels increased in the cytosol fraction (Fig. 3B, upper panel). The GSK3β inhibitor LiCl (10mM) was used as a positive control for GSK3β phosphorylation and its effect on the stabilization of cytosolic β-catenin (Fig. 3). Thrombin induced GSK3β phosphorylation was not inhibited by Wortmannin (50nM) or by Akt inhibitor VII (50μM) (data not shown). Thrombin induced a PI3K/Akt-independent GSK3β phosphorylation, which prevents GSK3β mediated β-catenin degradation and contributes to enhance cytosolic β-catenin levels.
129
Nuclear targeting of β-catenin
Figure 3. Thrombin-induced GSK3β inactivation prevents β-catenin degradation.
A) Whole cell lysates of control, thrombin (1 U/ml) and LiCl (10mM) treated cells were analyzed by Western blot analysis. Blots were stained for β-catenin (upper lanes), GSK3β phosphorylated at ser9 (middle lanes) and total GSK3β (bottom lanes). B) Western blots of the cytosol fractions were stained for β-catenin (upper lanes) and β-actin (lower lanes). Results are representative of 3 experiments, p>0.05.
130

Nuclear targeting of β-catenin
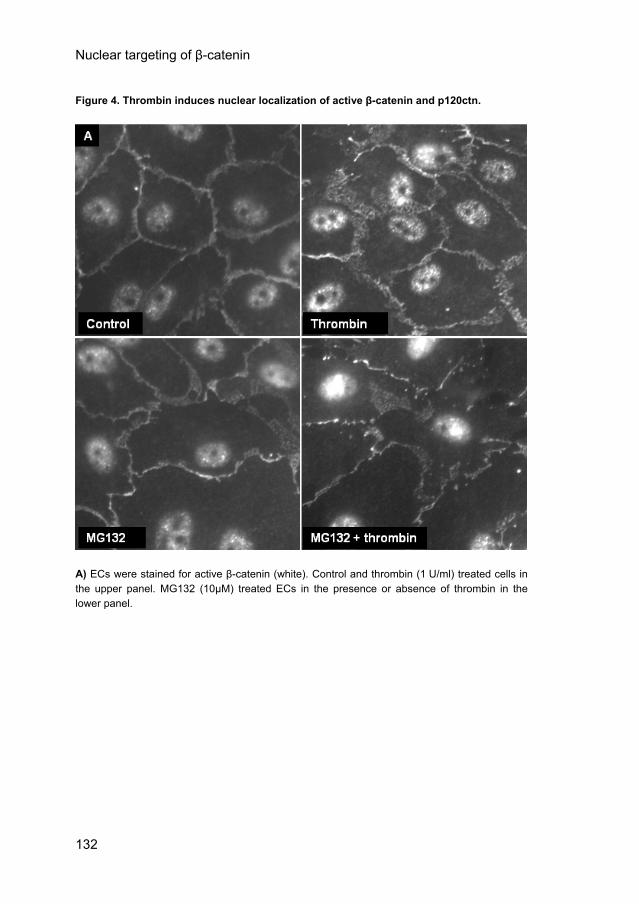
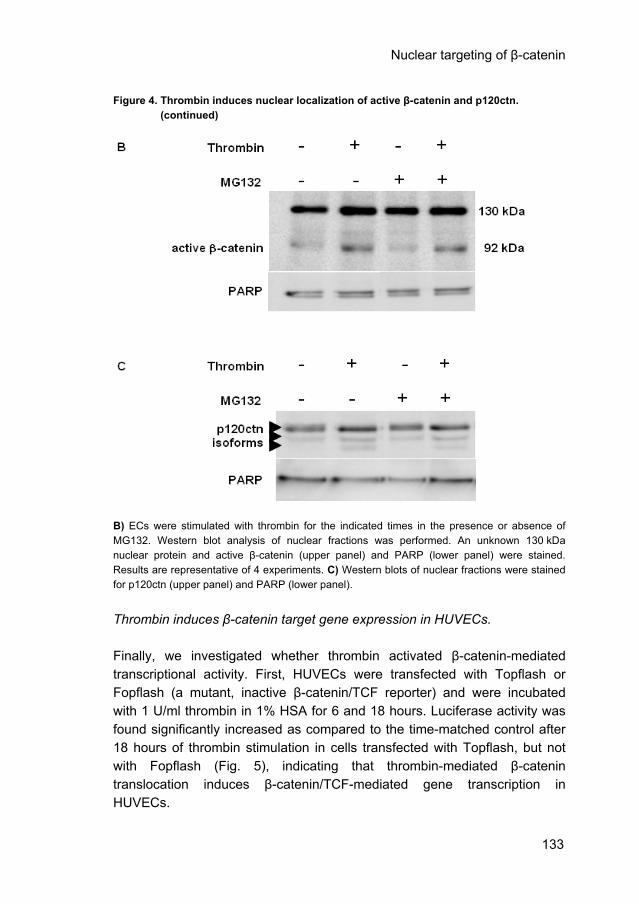
Thrombin induces nuclear localization of active β-catenin and p120ctn. Next we investigated whether thrombin (1 U/ml) induced translocation of β-catenin and p120ctn to the nucleus of confluent HUVECs. To that end, we used the antibody αABC that specifically recognizes the active, non-phosphorylated form of β-catenin. Figure 4A shows that this antibody recognized some activity in the nucleus of non-stimulated confluent ECs. Thrombin stimulation for 30 minutes increased the degree of immunoreactivity for nuclear β-catenin. However, Western blot analysis of nuclear fractions (Fig. 4B, lanes 1 and 2) showed that in addition to the 92 kDa active β-catenin the antibody also recognized an unidentified protein of 130 kDa. This protein was similarly enhanced by thrombin as the active β-catenin protein. Although the immunostaining may be flawed by the 130 kDa protein, the Western blots clearly demonstrate that nuclear β-catenin is enhanced after stimulation of the cells with thrombin (2.8 ± 0.8 fold-increase as compared to control, n=4, p<0.05). These results were verified using a NE-PER kit (# 78833) from Pierce to obtain cytosol and nuclear fractions (2.3-fold increase, data not shown). Pre-incubation of the proteasome inhibitor MG132 did not affect or only slightly enhanced basal β-catenin levels (Fig. 4A). Subsequent thrombin stimulation increased the nuclear localization of β-catenin (Fig. 4A) as was conformed by Western blot analysis (Fig. 4B, lanes 3 and 4). In addition to β-catenin, the nuclear amount of p120ctn also increased (Fig. 4C). Proteasome inhibition by MG132 also did not affect the amount of nuclear p120ctn. These data show that thrombin stimulation of ECs induces membrane liberated β-catenin and p120ctn translocation to the nucleus, but that inhibition of β-catenin degradation alone is not sufficient to increase the nuclear β-catenin levels.
131
Nuclear targeting of β-catenin
Figure 4. Thrombin induces nuclear localization of active β-catenin and p120ctn.
A) ECs were stained for active β-catenin (white). Control and thrombin (1 U/ml) treated cells in the upper panel. MG132 (10μM) treated ECs in the presence or absence of thrombin in the lower panel.
132
Nuclear targeting of β-catenin
Figure 4. Thrombin induces nuclear localization of active β-catenin and p120ctn. Figure 4. (continued)
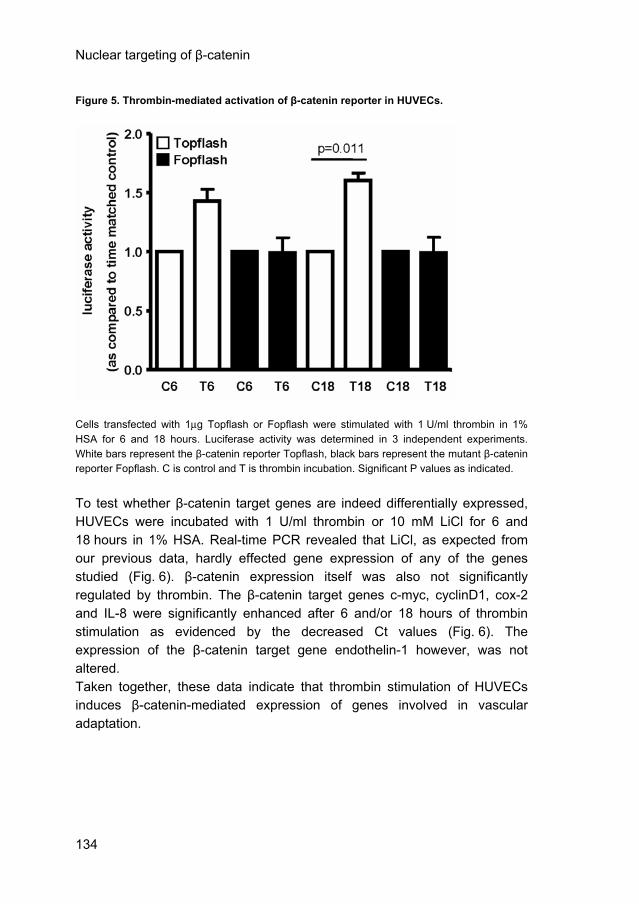
B) ECs were stimulated with thrombin for the indicated times in the presence or absence of MG132. Western blot analysis of nuclear fractions was performed. An unknown 130 kDa nuclear protein and active β-catenin (upper panel) and PARP (lower panel) were stained. Results are representative of 4 experiments. C) Western blots of nuclear fractions were stained for p120ctn (upper panel) and PARP (lower panel). Thrombin induces β-catenin target gene expression in HUVECs. Finally, we investigated whether thrombin activated β-catenin-mediated transcriptional activity. First, HUVECs were transfected with Topflash or Fopflash (a mutant, inactive β-catenin/TCF reporter) and were incubated with 1 U/ml thrombin in 1% HSA for 6 and 18 hours. Luciferase activity was found significantly increased as compared to the time-matched control after 18 hours of thrombin stimulation in cells transfected with Topflash, but not with Fopflash (Fig. 5), indicating that thrombin-mediated β-catenin translocation induces β-catenin/TCF-mediated gene transcription in HUVECs.
133
Nuclear targeting of β-catenin
Figure 5. Thrombin-mediated activation of β-catenin reporter in HUVECs.
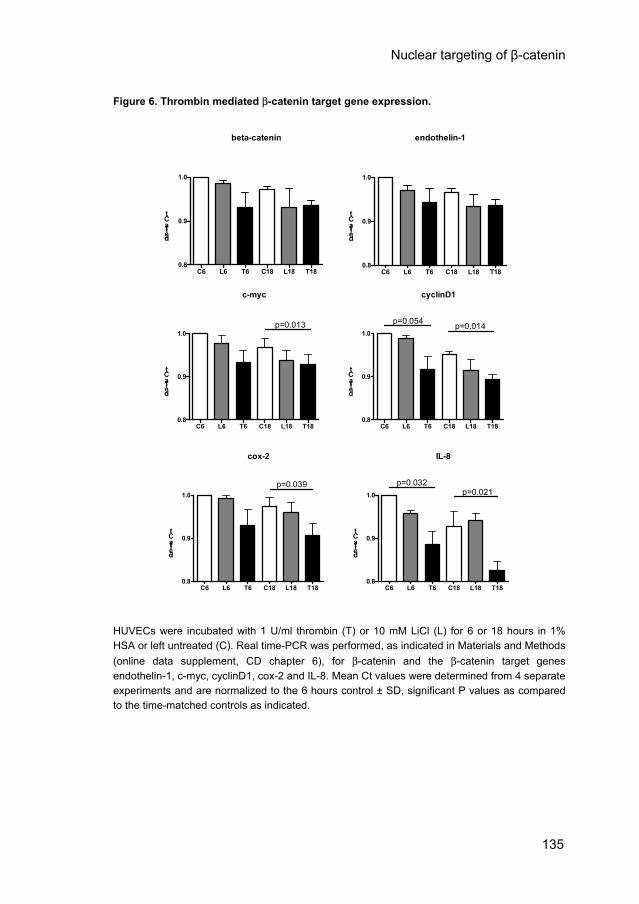
Cells transfected with 1μg Topflash or Fopflash were stimulated with 1 U/ml thrombin in 1% HSA for 6 and 18 hours. Luciferase activity was determined in 3 independent experiments. White bars represent the β-catenin reporter Topflash, black bars represent the mutant β-catenin reporter Fopflash. C is control and T is thrombin incubation. Significant P values as indicated. To test whether β-catenin target genes are indeed differentially expressed, HUVECs were incubated with 1 U/ml thrombin or 10 mM LiCl for 6 and 18 hours in 1% HSA. Real-time PCR revealed that LiCl, as expected from our previous data, hardly effected gene expression of any of the genes studied (Fig. 6). β-catenin expression itself was also not significantly regulated by thrombin. The β-catenin target genes c-myc, cyclinD1, cox-2 and IL-8 were significantly enhanced after 6 and/or 18 hours of thrombin stimulation as evidenced by the decreased Ct values (Fig. 6). The expression of the β-catenin target gene endothelin-1 however, was not altered. Taken together, these data indicate that thrombin stimulation of HUVECs induces β-catenin-mediated expression of genes involved in vascular adaptation.
134
Nuclear targeting of β-catenin
Figure 6. Thrombin mediated β-catenin target gene expression.
c-myc
C6 L6 T6 C18 L18 T180.8
0.9
1.0p=0.013
delta Ct
cyclinD1
C6 L6 T6 C18 L18 T180.8
0.9
1.0p=0.014p=0.054
delta Ct
cox-2
C6 L6 T6 C18 L18 T180.8
0.9
1.0p=0.039
delta Ct
IL-8
C6 L6 T6 C18 L18 T180.8
0.9
1.0
p=0.032p=0.021
delta Ct
beta-catenin
C6 L6 T6 C18 L18 T180.8
0.9
1.0
delta Ct
endothelin-1
C6 L6 T6 C18 L18 T180.8
0.9
1.0
delta Ct
HUVECs were incubated with 1 U/ml thrombin (T) or 10 mM LiCl (L) for 6 or 18 hours in 1% HSA or left untreated (C). Real time-PCR was performed, as indicated in Materials and Methods (online data supplement, CD chapter 6), for β-catenin and the β-catenin target genes endothelin-1, c-myc, cyclinD1, cox-2 and IL-8. Mean Ct values were determined from 4 separate experiments and are normalized to the 6 hours control ± SD, significant P values as compared to the time-matched controls as indicated.
135

Nuclear targeting of β-catenin
Discussion The present study shows that thrombin, an enhancer of the endothelial barrier dysfunction, induces targeting of membrane-localized β-catenin and p120ctn to the nucleus, resulting in enhanced β-catenin target gene expression. In particular, thrombin induces the release of β-catenin from the membrane, a process that involves Rho kinase-mediated changes of the F-actin cytoskeleton. Thrombin-mediated inactivation of GSK3β stabilizes the membrane-liberated β-catenin in the cytoplasm and subsequently allows its translocation to the nucleus. Based on these data we propose a model in which both the release of β-catenin from the AJs as well as the stabilization of β-catenin in the cytosol are required for nuclear targeting of β-catenin after thrombin stimulation. In non-stimulated confluent ECs β-catenin is recognized as a continuous lining at the membrane, which is well known from epithelial and endothelial cells.7, 40 The non-phosphorylated β-catenin is here bound to VE-cadherin.41 We observed a network-like organization of β-catenin at areas where the membranes between the ECs overlapped, similar as the structures recently shown by Birukova et al.42 These network-like structures of AJs may enforce the tethering forces between adjacent ECs once they become challenged. In addition, we encountered β-catenin in a new structure, namely a curled network around the peripheral belt of F-actin filaments. In non-stimulated confluent ECs little β-catenin was observed in the cytosol and no active β-catenin was present in their nuclei. This indicates that either no cytosolic β-catenin pool is formed at all, or that cytosolic β-catenin is efficiently targeted for ubiquitination and degradation. Similarly, little p120ctn was encountered in the cytosol and nucleus of non-stimulated ECs. After stimulation with thrombin we found relocalization of β-catenin from the membrane to the cytosol. Upon VEGF-mediated disruption of the AJs, VE-cadherin was shown to be internalized via endocytosis43 and subsequently degraded or recycled to the cell membrane.44 Notwithstanding this, Konstantoulaki et al.41 reported that VE-cadherin was relocalized but remained at the plasma membrane after thrombin stimulation of human pulmonary arterial ECs. While β-catenin co-localized with VE-cadherin in the AJs, in agreement with previous reports, this co-localization was largely lost in the cytoplasm.41 Dissociation of β-catenin and VE-cadherin rapidly occurs after phosphorylation of Tyr654 of β-catenin.45 In this respect it is of interest to note that protein tyrosine phosphorylation is one of the factors that contribute to barrier dysfunction induced by thrombin, together with an
136

Nuclear targeting of β-catenin
increase in cytosolic calcium ion concentration and activation of RhoA/Rho kinase and PKC.41, 46 While protein tyrosine phosphorylation may be sufficient to liberate β-catenin from the AJs, our data on the inhibition of Rho kinase indicate that Rho kinase activity was required for the accumulation of β-catenin in the cytosol. This in contrast to the non-canonical Wnt pathway in early development, in which RhoA signaling was inhibited without affecting nuclear β-catenin signaling.47 The thrombin-mediated regulation of β-catenin accumulation appears therefore different from the mechanisms observed in early development. The GSK3β-mediated degradation of β-catenin occurs when the N-terminal part of β-catenin by phosphorylation of its Tyr142 and subsequent structural changes becomes sensitive to serine phosphorylations by CK-1 and GSK-3β.48 The CK-1 phosphorylation facilitates subsequent phosphorylations by GSK3β, which make β-catenin prone to ubiquitination and subsequent degradation by the proteasomes.14 In agreement with earlier data, we demonstrated that thrombin mediates inactivation of GSK3β which depends on PKC activation41 and proceeds independent of the PI3K/Akt pathway.49 The antibody αABC recognizes specifically the active, non-phosphorylated form of β-catenin, the form that enters the nucleus and causes gene transcription.20 In addition, it targeted an unidentified 130 kDa protein which was also reported in the original manuscript by van Noort et al.17 Nonetheless, we provide clear evidence that thrombin mediates a transient relocalization of β-catenin as well as p120ctn to the nucleus. Nuclear β-catenin has been associated with EC survival, proliferation and angiogenesis.5, 50 Accumulating evidence suggests that p120ctn is a key mediator in this process.28, 51, 52 Therefore, β-catenin and p120ctn might form a link between the initial breakdown of the EC barrier and the subsequent angiogenic processes. Here, we show that thrombin-mediated permeability in HUVECs induces activation of the TCF reporter Topflash as well as the expression of the cancer and angiogenesis related β-catenin target genes c-myc, cyclinD1, cox-2 and IL-8. We found no effect on endothelin-1 or β-catenin expression itself. Our data are in line with resent data from Ichikawa et al.,53 who showed that prolonged incubation of HUVECs with MMP7 induced disassembly of the AJs, nuclear β-catenin translocation and increased MMP7 and cox-2, but not cyclinD1 mRNA expression. Of these genes MMP7 and cyclinD1 contain a Kaiso binding site. In several human cancer cell types MMP7 is indeed regulated in a Kaiso/p120ctn mediated manner in
137

Nuclear targeting of β-catenin
vivo.52 Our data show nuclear accumulation of p120ctn, which strongly suggests that also in ECs p120ctn plays a role in the β-catenin-mediated gene expression. In conclusion, in human ECs the adhesive and signaling functions of β-catenin are interconnected. The hyperpermeability inducer thrombin stimulates targeting of membrane-localized β-catenin and p120ctn to the nucleus. Both the release of β-catenin from the adherens junctions as well as thrombin-mediated β-catenin stabilization in the cytosol is required for nuclear targeting. In sharp contrast to the canonical Wnt signaling, where nuclear localization of β-catenin is due to the stabilization of the cytosolic pool of β-catenin, our data strongly suggest that upon thrombin stimulation, it is the pool of active-β-catenin coming from the membrane that relocates to the nucleus. Nuclear accumulation of β-catenin and p120ctn induced transcription of several β-catenin target genes. Future studies have to reveal whether this contributes to an angiogenic response of ECs following episodes of vascular leakage.
Acknowledgements: We thank Martin Kramer and Ing. Clarissa Jungerius for her pleasant company and excellent technical assistance, and Dr. Rene J.P. Musters for sharing his knowledge on 3D wide field microscopy. Sources of funding: Dr. Geerten P. van Nieuw Amerongen was supported by The Netherlands Heart Foundation (The Hague, grant 2003T032). Our laboratory was supported by the EU (EVGN contract LSHM-2003-503254). References (1) Aird W.C. Phenotypic heterogeneity of the endothelium: I. Structure, function, and
mechanisms. Circ Res 2007 February 2; 100 (2): 158-73. (2) Dejana E. Endothelial cell-cell junctions: Happy together. Nature Reviews Molecular
Cell Biology 2004 April; 5 (4): 261-70. (3) Potter M.D., Barbero S., Cheresh D.A. Tyrosine phosphorylation of VE-cadherin
prevents binding of p120- and β-catenin and maintains the cellular mesenchymal state. Journal of Biological Chemistry 2005 September 9; 280 (36): 31906-12.
(4) Behrens J., von Kries J.P., Kuhl M., Bruhn L., Wedlich D., Grosschedl R., Birchmeier W. Functional interaction of β-catenin with the transcription factor LEF-1. Nature 1996 August 15; 382 (6592): 638-42.
(5) Kim K.I., Cho H.J., Hahn J.Y., Kim T.Y., Park K.W., Koo B.K., Shin C.S., Kim C.H., Oh B.H., Lee M.M., Park Y.B., Kim H.S. β-Catenin overexpression augments angiogenesis and skeletal muscle regeneration through dual mechanism of vascular endothelial growth factor-mediated endothelial cell proliferation and progenitor cell
138

Nuclear targeting of β-catenin
mobilization. Arteriosclerosis Thrombosis and Vascular Biology 2006 January; 26 (1): 91-8.
(6) Korswagen H.C., Herman M.A., Clevers H.C. Distinct β-catenins mediate adhesion and signaling functions in C-elegans. Nature 2000 August 3; 406 (6795): 527-32.
(7) Clevers H. Wnt/β-catenin signaling in development and disease. Cell 2006 November 3; 127 (3): 469-80.
(8) Telo P., Breviario F., Huber P., Panzeri C., Dejana E. Identification of a novel cadherin (vascular endothelial cadherin-2) located at intercellular junctions in endothelial cells. Journal of Biological Chemistry 1998 July 10; 273 (28): 17565-72.
(9) Lampugnani M.G., Resnati M., Raiteri M., Pigott R., Pisacane A., Houen G., Ruco L.P., Dejana E. A Novel Endothelial-Specific Membrane-Protein Is A Marker of Cell Cell Contacts. Journal of Cell Biology 1992 September; 118 (6): 1511-22.
(10) Aberle H., Schwartz H., Kemler R. Cadherin-catenin complex: protein interactions and their implications for cadherin function. J Cell Biochem 1996 June 15; 61 (4): 514-23.
(11) Hirano S., Nose A., Hatta K., Kawakami A., Takeichi M. Calcium-dependent cell-cell adhesion molecules (cadherins): subclass specificities and possible involvement of actin bundles. J Cell Biol 1987 December; 105 (6 Pt 1): 2501-10.
(12) Reynolds A.B., Herbert L., Cleveland J.L., Berg S.T., Gaut J.R. p120, a novel substrate of protein tyrosine kinase receptors and of p60v-src, is related to cadherin-binding factors β-catenin, plakoglobin and armadillo. Oncogene 1992 December; 7 (12): 2439-45.
(13) Molenaar M., van de Wetering M., Oosterwegel M., Peterson Maduro J., Godsave S., Korinek V., Roose J., Destree O., Clevers H. XTcf-3 transcription factor mediates β-catenin-induced axis formation in Xenopus embryos. Cell 1996 August 9; 86 (3): 391-9.
(14) Aberle H., Bauer A., Stappert J., Kispert A., Kemler R. β-Catenin is a target for the ubiquitin-proteasome pathway. EMBO J 1997 July 1; 16 (13): 3797-804.
(15) Eberhart C.G., Tihan T., Burger P.C. Nuclear localization and mutation of β-catenin in medulloblastomas. Journal of Neuropathology and Experimental Neurology 2000 April; 59 (4): 333-7.
(16) Blankesteijn W.M., van Gijn M.E., Essers-Janssen Y.P., Daemen M.J., Smits J.F. β-Catenin, an inducer of uncontrolled cell proliferation and migration in malignancies, is localized in the cytoplasm of vascular endothelium during neovascularization after myocardial infarction. Am J Pathol 2000 September; 157 (3): 877-83.
(17) van Noort M., Meeldijk J., van der Zee R., Destree O., Clevers H. Wnt signaling controls the phosphorylation status of β-catenin. Journal of Biological Chemistry 2002 May 17; 277 (20): 17901-5.
(18) Eto M., Kouroedov A., Cosentino F., Luscher T.F. Glycogen synthase kinase-3 mediates endothelial cell activation by tumor necrosis factor-alpha. Circulation 2005 August 30; 112 (9): 1316-22.
(19) Couffinhal T., Dufourcq P., Duplaa C. β-Catenin nuclear activation: common pathway between Wnt and growth factor signaling in vascular smooth muscle cell proliferation? Circ Res 2006 December 8; 99 (12): 1287-9.
(20) Gottardi C.J., Gumbiner B.M. Distinct molecular forms of β-catenin are targeted to adhesive or transcriptional complexes. J Cell Biol 2004 October 25; 167 (2): 339-49.
(21) He T.C., Sparks A.B., Rago C., Hermeking H., Zawel L., da Costa L.T., Morin P.J., Vogelstein B., Kinzler K.W. Identification of c-MYC as a target of the APC pathway. Science 1998 September 4; 281 (5382): 1509-12.
(22) Tetsu O., McCormick F. β-Catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999 April 1; 398 (6726): 422-6.
(23) Shtutman M., Zhurinsky J., Simcha I., Albanese C., D'Amico M., Pestell R., Ben Ze'ev A. The cyclin D1 gene is a target of the β-catenin/LEF-1 pathway. Proc Natl Acad Sci U S A 1999 May 11; 96 (10): 5522-7.
(24) Haertel-Wiesmann M., Liang Y., Fantl W.J., Williams L.T. Regulation of cyclooxygenase-2 and periostin by Wnt-3 in mouse mammary epithelial cells. J Biol Chem 2000 October 13; 275 (41): 32046-51.
139

Nuclear targeting of β-catenin
(25) Howe L.R., Subbaramaiah K., Chung W.J., Dannenberg A.J., Brown A.M. Transcriptional activation of cyclooxygenase-2 in Wnt-1-transformed mouse mammary epithelial cells. Cancer Res 1999 April 1; 59 (7): 1572-7.
(26) Masckauchan T.N., Shawber C.J., Funahashi Y., Li C.M., Kitajewski J. Wnt/β-catenin signaling induces proliferation, survival and interleukin-8 in human endothelial cells. Angiogenesis 2005; 8 (1): 43-51.
(27) Kim T.H., Xiong H., Zhang Z., Ren B. β-Catenin activates the growth factor endothelin-1 in colon cancer cells. Oncogene 2005 January 20; 24 (4): 597-604.
(28) Park J.I., Ji H., Jun S., Gu D., Hikasa H., Li L., Sokol S.Y., McCrea P.D. Frodo links Dishevelled to the p120-catenin/Kaiso pathway: distinct catenin subfamilies promote Wnt signals. Dev Cell 2006 November; 11 (5): 683-95.
(29) Soubry A., van Hengel J., Parthoens E., Colpaert C., van Marck E., Waltregny D., Reynolds A.B., van Roy F. Expression and nuclear location of the transcriptional repressor Kaiso is regulated by the tumor microenvironment. Cancer Res 2005 March 15; 65 (6): 2224-33.
(30) Goodwin A.M., D'Amore P.A. Wnt signaling in the vasculature. Angiogenesis 2002; 5 (1-2): 1-9.
(31) Carmeliet P., Jain R.K. Angiogenesis in cancer and other diseases. Nature 2000 September 14; 407 (6801): 249-57.
(32) van Nieuw Amerongen G.P., Koolwijk P., Versteilen A., van Hinsbergh V.W.M. Involvement of RhoA/Rho kinase signaling in VEGF-induced endothelial cell migration and angiogenesis in vitro. Arterioscler Thromb Vasc Biol 2003 February 1; 23 (2): 211-7.
(33) Cavallaro U., Liebner S., Dejana E. Endothelial cadherins and tumor angiogenesis. Experimental Cell Research 2006 March 10; 312 (5): 659-67.
(34) van Nieuw Amerongen G.P., Draijer R., Vermeer M.A., van Hinsbergh V. Transient and prolonged increase in endothelial permeability induced by histamine and thrombin: role of protein kinases, calcium, and RhoA. Circ Res 1998 November 30; 83 (11): 1115-23.
(35) Jaffe E.A., Nachman R.L., Becker C.G., Minick C.R. Culture of human endothelial cells derived from umbilical veins. Identification by morphologic and immunologic criteria. J Clin Invest 1973 November; 52 (11): 2745-56.
(36) Kouklis P., Konstantoulaki M., Vogel S., Broman M., Malik A.B. Cdc42 regulates the restoration of endothelial barrier function. Circ Res 2004 February 6; 94 (2): 159-66.
(37) Nilsson I., Shibuya M., Wennstrom S. Differential activation of vascular genes by hypoxia in primary endothelial cells. Exp Cell Res 2004 October 1; 299 (2): 476-85.
(38) van der Sar A.M., Musters R.J.P., van Eeden F.J.M., Appelmelk B.J., van den Broucke-Grauls C.M.J., Bitter W. Zebrafish embryos as a model host for the real time analysis of Salmonella typhimurium infections. Cellular Microbiology 2003 September; 5 (9): 601-11.
(39) Garcia-Vallejo J.J., van het Hof B., Robben J., van Wijk J.A., van Die I., Joziasse D.H., van Dijk W. Approach for defining endogenous reference genes in gene expression experiments. Anal Biochem 2004 June 15; 329 (2): 293-9.
(40) Liebner S., Cavallaro U., Dejana E. The multiple languages of endothelial cell-to-cell communication. Arterioscler Thromb Vasc Biol 2006 July; 26 (7): 1431-8.
(41) Konstantoulaki M., Kouklis P., Malik A.B. Protein kinase C modifications of VE-cadherin, p120, and β-catenin contribute to endothelial barrier dysregulation induced by thrombin. Am J Physiol Lung Cell Mol Physiol 2003 August; 285 (2): L434-L442.
(42) Birukova A.A., Malyukova I., Poroyko V., Birukov K.G. Paxillin - β-Catenin interactions are involved in Rac/Cdc42-mediated endothelial barrier-protective response to oxidized phospholipids. Am J Physiol Lung Cell Mol Physiol 2007 May 18; 293: L199-L211.
(43) Gavard J., Gutkind J.S. VEGF controls endothelial-cell permeability by promoting the β-arrestin-dependent endocytosis of VE-cadherin. Nat Cell Biol 2006 November; 8 (11): 1223-34.
(44) Xiao K., Garner J., Summers S., Buckley K.M., Dejana E., Faundez V., Kowalczyk A.P. p120 catenin regulates clathrin-dependent endocytosis of VE-cadherin. Journal of Investigative Dermatology 2005 April; 124 (4): A30.
140

Nuclear targeting of β-catenin
141
(45) Piedra J, Martinez D., Castano J., Miravet S., Dunach M., de Herreros A.G. Regulation of β-catenin structure and activity by tyrosine phosphorylation. J Biol Chem 2001 June 8; 276 (23): 20436-43.
(46) van Nieuw Amerongen G.P., van Delft S., Vermeer M.A. and van Hinsbergh V.W.M. Activation of RhoA by thrombin in endothelial hyperpermeability: role of Rho kinase and protein tyrosine kinases. Circ Res 2000 August 18; 87 (4): 335-40.
(47) Habas R., Kato Y., He X. Wnt/Frizzled activation of Rho regulates vertebrate gastrulation and requires a novel Formin homology protein Daam1. Cell 2001 December 28; 107 (7): 843-54.
(48) Liu C., Li Y., Semenov M., Han C., Baeg G.H., Tan Y., Zhang Z., Lin X., He X. Control of β-catenin phosphorylation/degradation by a dual-kinase mechanism. Cell 2002 March 22; 108 (6): 837-47.
(49) Thors B., Halldorsson H., Clarke G.D., Thorgeirsson G. Inhibition of Akt phosphorylation by thrombin, histamine and lysophosphatidylcholine in endothelial cells. Differential role of protein kinase C. Atherosclerosis 2003 June; 168 (2): 245-53.
(50) Skurk C., Maatz H., Rocnik E., Bialik A., Force T., Walsh K. Glycogen-synthase kinase3β / β-catenin axis promotes angiogenesis through activation of vascular endothelial growth factor signaling in endothelial cells. Circulation Research 2005 February 18; 96 (3): 308-18.
(51) Kondapalli J., Flozak A.S., Albuquerque M.L. Laminar shear stress differentially modulates gene expression of p120 catenin, Kaiso transcription factor, and vascular endothelial cadherin in human coronary artery endothelial cells. J Biol Chem 2004 March 19; 279 (12): 11417-24.
(52) Spring C.M., Kelly K.F., O'Kelly I., Graham M., Crawford H.C., Daniel J.M. The catenin p120ctn inhibits Kaiso-mediated transcriptional repression of the β-catenin/TCF target gene matrilysin. Exp Cell Res 2005 May 1; 305 (2): 253-65.
(53) Ichikawa Y., Ishikawa T., Momiyama N., Kamiyama M., Sakurada H., Matsuyama R., Hasegawa S., Chishima T., Hamaguchi Y., Fujii S., Saito S., Kubota K., Hasegawa S., Ike H., Oki S., Shimada H. Matrilysin (MMP-7) degrades VE-cadherin and accelerates accumulation of β-catenin in the nucleus of human umbilical vein endothelial cells. Oncol Rep 2006 February; 15 (2): 311-5.

142









![Standard specifications - Nachi Robotics · Standard specifications MZ07-02 ... [rad] = 180 /π[°], 1[N・m ... - The specification and externals described in this specifications](https://static.fdocument.org/doc/165x107/5b15cdf47f8b9a5e798b477d/standard-specifications-nachi-standard-specifications-mz07-02-rad-.jpg)