
LEZIONE DEL 20/04/2017 BIOENERGETICA I. LO … · lezione del 20/04/2017 bioenergetica i. lo schema...
18
LEZIONE DEL 20/04/2017 BIOENERGETICA I. LO SCHEMA GENERALE DEL CATABOLISMO; II. QUATTRO MOLECOLE MOLTO ENERGETICHE; III. ΔG°' E LE REAZIONI ACCOPPIATE; IV. LA PIROFOSFATASI INORGANICA; V. L'ACETIL-CoA (primo cofattore enzimatico). I. BIOENERGETICA "CELLULARE" (Mario Ageno): Energia "pregiata" - negli zuccheri e nei lipidi - che può essere immagazzinata e non si disperde, con resa elevatissia, non produce, una volta espulsa, scorie, non innalza la temperatura. Per mandare avanti i processi biochimici serve energia: 1) "contante" (immediatamente pronta all'uso); 2) "in banca" (di deposito, la cui accumulazione richiede più sforzi). Tale energia è coinvolta nelle: 1) reazioni di sintesi: l'entropia si abbassa, aumenta l'ordine: - ΔS (variazione di entropia) < 0 (ordine); - ΔH (variazione di entalpia) < 0 (formazione di legame); 2) reazioni di degradazione: - ΔS >> 0 (produzione di calore e di scarti). QUATTRO MACROMOLECOLE FONDAMENTALI NEL CATABOLISMO E LA PRODUZIONE DI ENERGIA 1. TRIGLICERIDI; 1
Transcript of LEZIONE DEL 20/04/2017 BIOENERGETICA I. LO … · lezione del 20/04/2017 bioenergetica i. lo schema...

LEZIONE DEL 20/04/2017
BIOENERGETICA
I. LO SCHEMA GENERALE DEL CATABOLISMO;
II. QUATTRO MOLECOLE MOLTO ENERGETICHE;
III. ΔG°' E LE REAZIONI ACCOPPIATE;
IV. LA PIROFOSFATASI INORGANICA;
V. L'ACETIL-CoA (primo cofattore enzimatico).
I. BIOENERGETICA "CELLULARE"
(Mario Ageno): Energia "pregiata" - negli zuccheri e nei lipidi - che può essere immagazzinata e non si disperde, con resa elevatissia, non produce, una volta espulsa, scorie, non innalza la temperatura. Per mandare avanti i processi biochimici serve energia:
1) " contante " (immediatamente pronta all'uso);
2) " in banca " (di deposito, la cui accumulazione richiede più sforzi).
Tale energia è coinvolta nelle:
1) reazioni di sintesi : l'entropia si abbassa, aumenta l'ordine:
- ΔS (variazione di entropia) < 0 (ordine);
- ΔH (variazione di entalpia) < 0 (formazione di legame);
2) reazioni di degradazione :
- ΔS >> 0 (produzione di calore e di scarti).
QUATTRO MACROMOLECOLE FONDAMENTALI NEL CATABOLISMO E LA PRODUZIONE DI ENERGIA
1. TRIGLICERIDI;
1
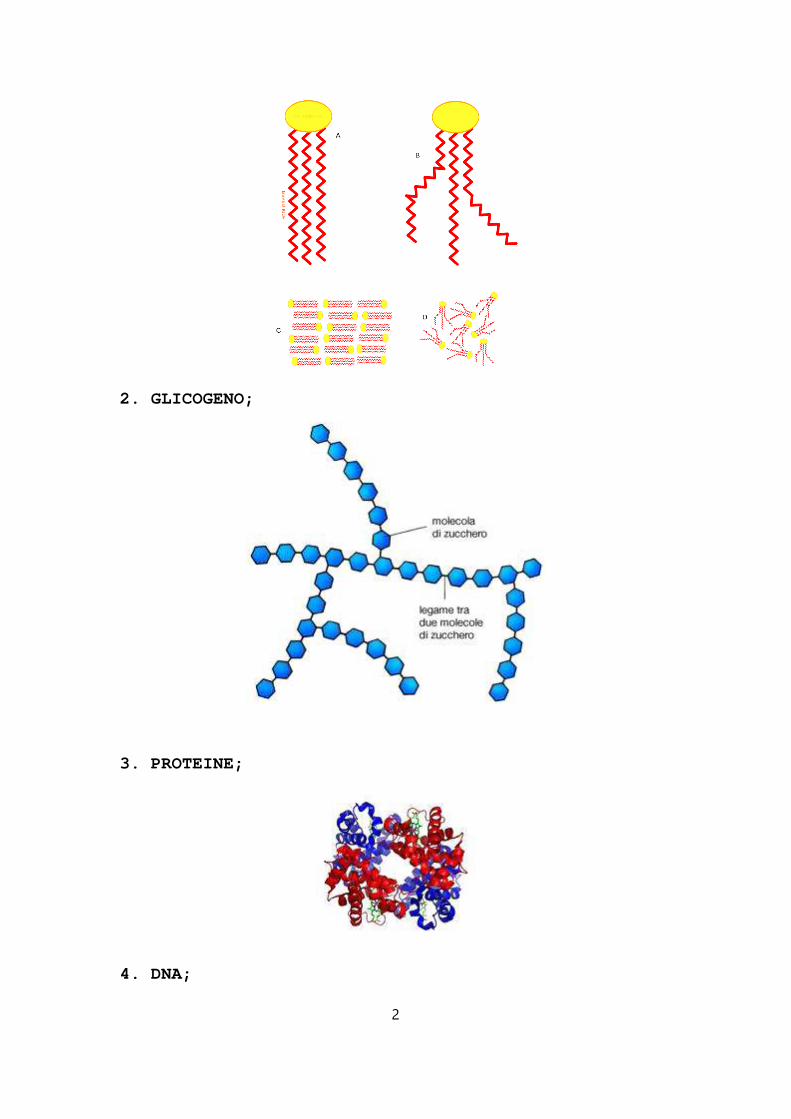
2. GLICOGENO;
3. PROTEINE;
4. DNA;
2

RISPETTIVI MONOMERI DELLE 4 MACROMOLECOLE E CATABOLISMO
1) GLICEROLO + 3 ACIDI : la molecola di trigliceride si compone partendo dal glicerolo a cui si legano t re catene di acidi a residui idrocarburici molto lungh i uguali o diversi tra loro.
[ Non scriveremo mai l'acido non carico perché stia mo a PH 7, inoltre dovremmo rappresentarlo con l'elettrone delocalizzato tra i due ossigeni legati al carbonio carbossilico dell'acido ];
2) GLUCOSIO: β-ossidazione, glicolisi;
3) AMMINOACIDI : le proteine introdotte nel corpo umano vengono fatte a pezzi per poi essere ricorstruite rispettando la struttura del corpo umano. Le proteine non si accumulano, esse non vengono utilizzate come carburanti;
4) NUCLEOTIDI.
1) Gli acidi grassi vanno nel mitocondrio , dove sono decomposti per produrre molta energia sotto forma d i due derivati vitaminici, trasportori di elettroni ( NADH e FADH2) e acetil-CoA . Questo sistema funziona come un circuito separatore di cariche: si prendono le cariche negative inglobate nei trasportori che i l mitocondio separa e si emette come residuo soltanto acqua (per produrre energia);
2) Il glicerolo va nel citosol , dove avviene la
3

glicolisi, si brucia e produce energia sotto forma di NADH, Acetil-CoA e ATP;
3) Gli amminoacidi devono essere sintetizzati da piante e batteri. La loro particolarità nell'avere l'azoto come componente. Il problema è che nell'atmosfera l'azoto si trova in forma molecolare ( :N≡N: ). Solo i batteri fanno rompere il triplo legame precedente: gli amminoacidi si decompongono con riluttanza. Tuttavia, sottoposti alla necessità di farlo, la parte carbonilica, separatasi dal complesso amminoacidico, andrebbe eliminata col cic lo per l'urea passando per il mitocondrio, la parte co n l'azoto passerebbe per altre vie.
In seguito, l'acetil-CoA entra nel ciclo di Krebs, in cui produce i suoi FADH 2 e NADH. Tutto questo si ricongiunge nella parte interna della membrana del mitocondrio, dai trasportori si prendono le cariche per formare ATP.
[ Abbiamo discusso del catabolismo degli acidi grassi, del glucosio e degli amminoacidi. Ci concentreremo nell'avvenire sull'anabolismo del glucosio e degli acidi grassi ].
II. QUATTRO MOLECOLE ENERGETICHE E LA METAFORA MEIA NA DEI FANTASTICI 4
1) FOSFOENOLPIRUVATO o PEP(dal piruvato
):
4
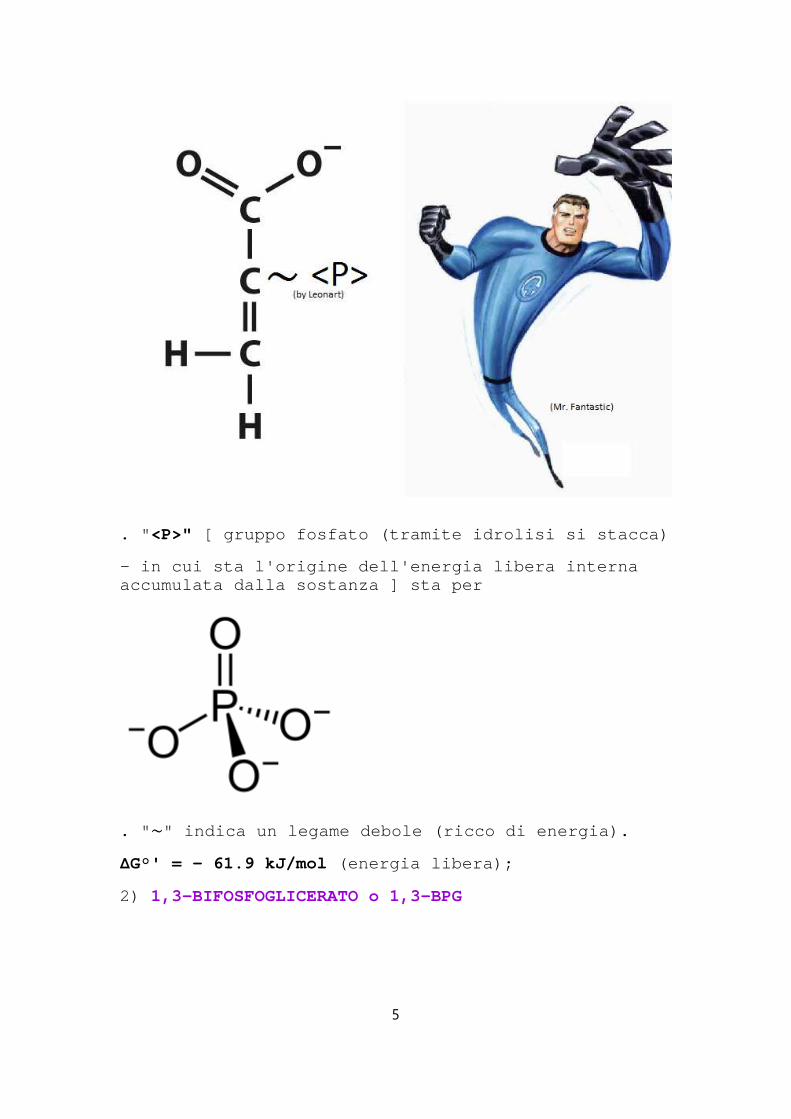
. " <P>" [ gruppo fosfato (tramite idrolisi si stacca)
- in cui sta l'origine dell'energia libera interna accumulata dalla sostanza ] sta per
. " ∼" indica un legame debole (ricco di energia).
ΔG°' = - 61.9 kJ/mol (energia libera);
2) 1,3-BIFOSFOGLICERATO o 1,3-BPG
5

Il legame tra <P> e C1 è più debole a causa dell'elettronegatività dell'ossigeno e dal doppio legame che attrae il C1 e lo "distrae" dal legarsi col gruppo fosfato.
ΔG°' = -49.3 kJ/mol;
[ PEP e 1,3-BPG (Mr. Fantastic e Torcia Umana) sono coinvolti nella glicolisi e sono simili a livello conformazionale al glicerolo :
3) FOSFOCREATINA o PCr
6
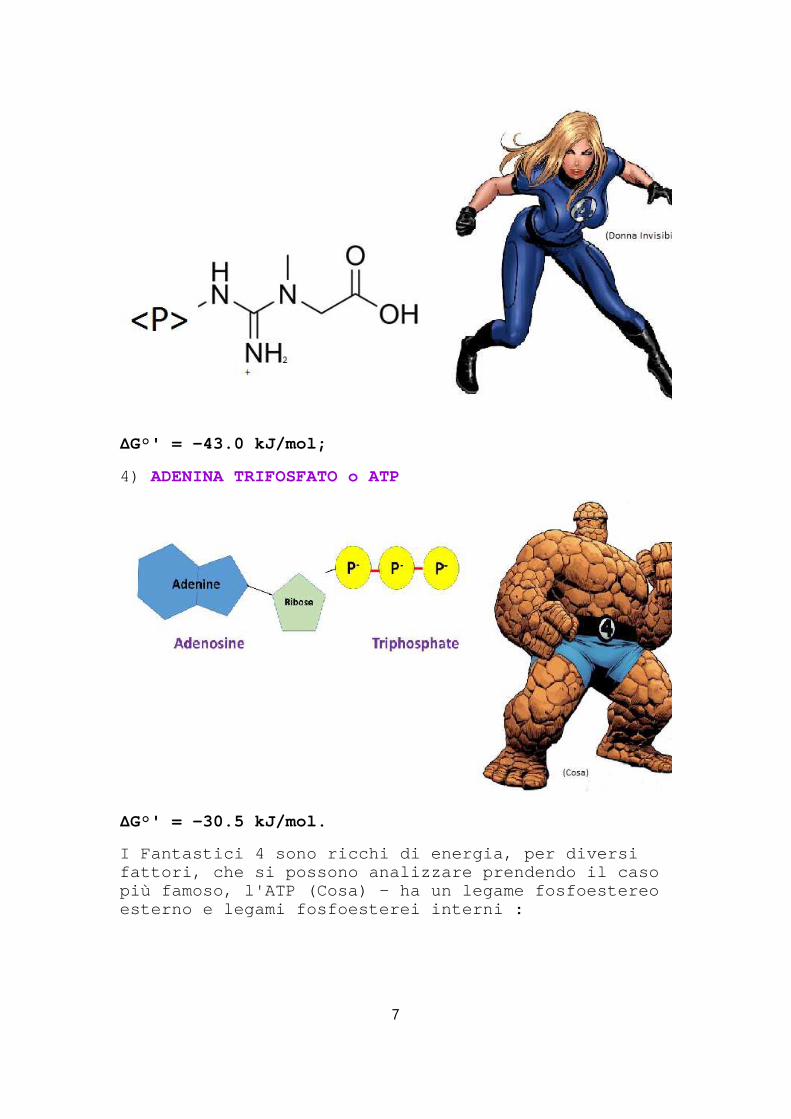
ΔG°' = -43.0 kJ/mol;
4) ADENINA TRIFOSFATO o ATP
ΔG°' = -30.5 kJ/mol.
I Fantastici 4 sono ricchi di energia, per diversi fattori, che si possono analizzare prendendo il cas o più famoso, l'ATP (Cosa) - ha un legame fosfoestereo esterno e legami fosfoesterei interni :
7
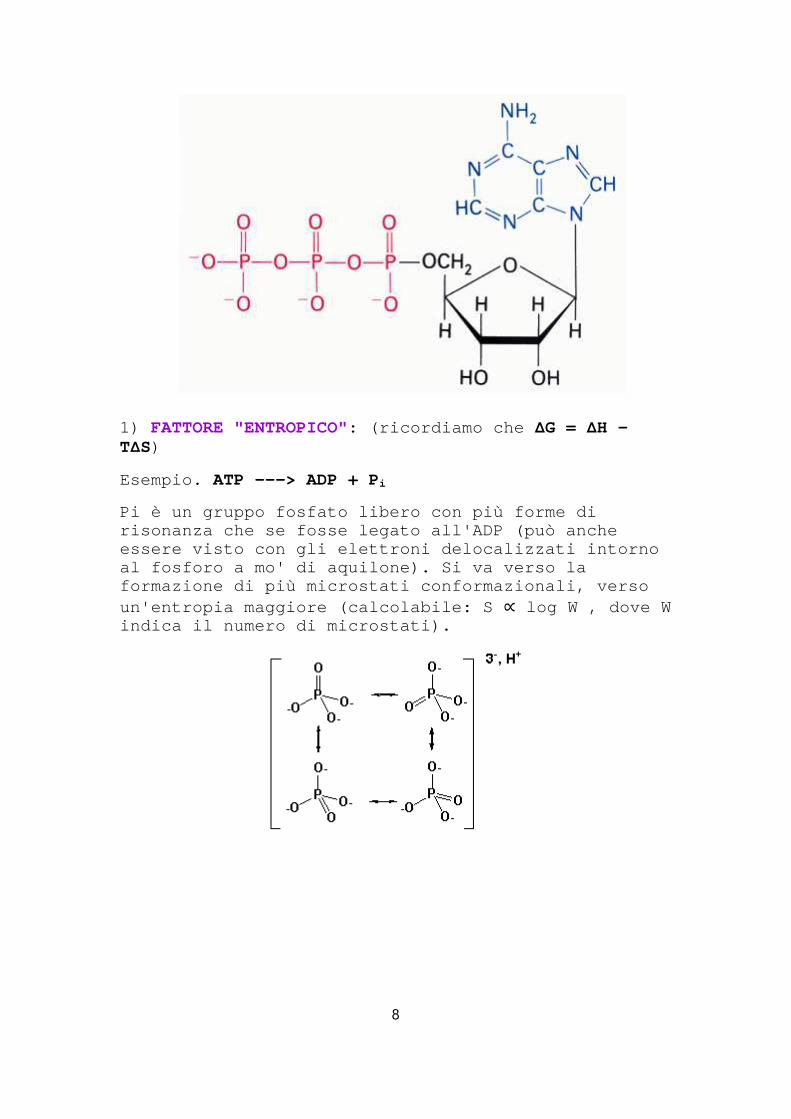
1) FATTORE "ENTROPICO": (ricordiamo che ΔG = ΔH -TΔS)
Esempio. ATP ---> ADP + P i
Pi è un gruppo fosfato libero con più forme di risonanza che se fosse legato all'ADP (può anche essere visto con gli elettroni delocalizzati intorn o al fosforo a mo' di aquilone). Si va verso la formazione di più microstati conformazionali, verso un'entropia maggiore (calcolabile: S ∝ log W , dove W indica il numero di microstati).
8
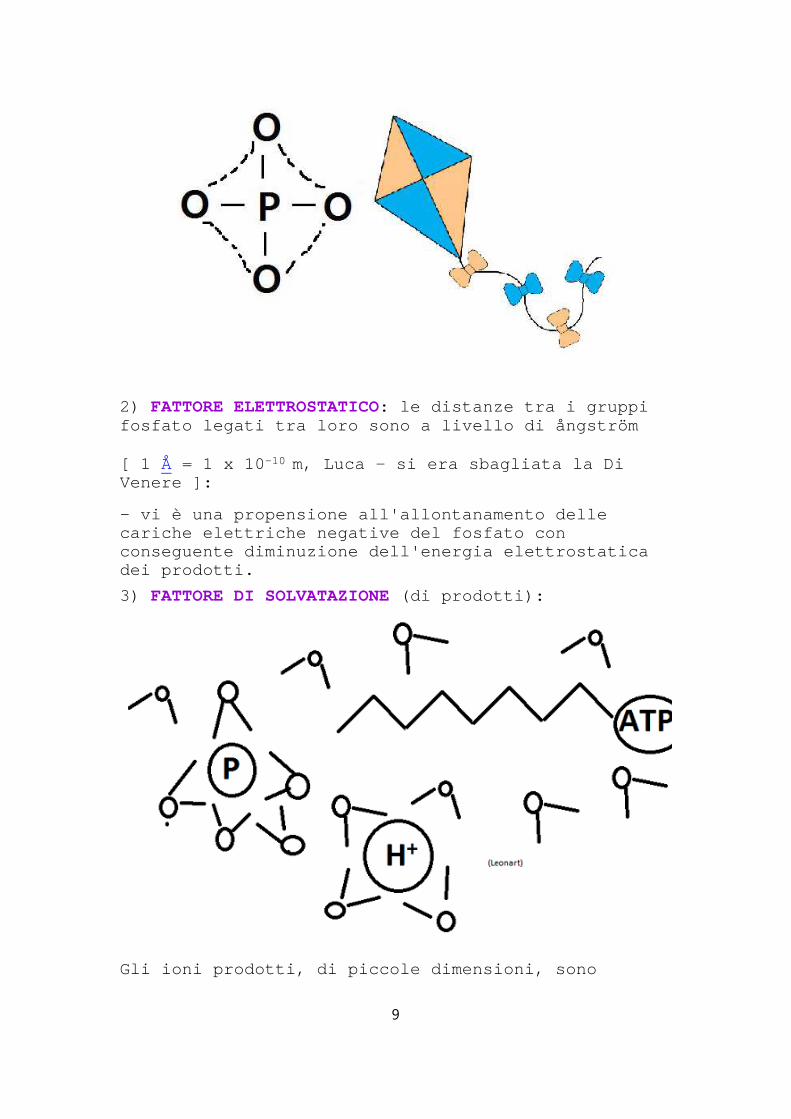
2) FATTORE ELETTROSTATICO: le distanze tra i gruppi fosfato legati tra loro sono a livello di ångström
[ 1 Å = 1 x 10 -10 m, Luca - si era sbagliata la Di Venere ]:
- vi è una propensione all'allontanamento delle cariche elettriche negative del fosfato con conseguente diminuzione dell'energia elettrostatica dei prodotti.
3) FATTORE DI SOLVATAZIONE (di prodotti):
Gli ioni prodotti, di piccole dimensioni, sono
9

stabilizzati in acqua;
4) FATTORE DI DISSOCIAZIONE (di ATP a PH7).
Si possono "fattorizzare" anche gli altri tre fantastici (PEP, 1,3-BPG e PCr).
III. OSS.1 SU ΔG°'
- In chimica ΔG° indica la variazione di energia libera in " condizioni standard ", cioé
T = 25° C, C oncentrazione = 1 M, pH = 0 ;
- In biochimica lo stato standard è riferito alle stesse condizioni, ad eccezione del pH, in questo caso pari a 7.0 . Il ΔG°' , riferendosi a delle condizioni standard, è una costante, specifica per ogni processo biochimico.
[Nella cellula, le concentrazioni sono molto divers e da 1 M che abbiamo scelto come standard;
ricordiamo che G = H - TS ].
ΔGvero = ΔG°' + RTln([prodotti]/[substrati])
Keq = [prodotti]/[substrati] :
in una reazione A + B ---> C + D :
10

Keq = ([C][D])/([A][B])
se K << 1 => ΔG < 0 (reazione esoergonica ).
Esercizio. ATP negli eritrociti - idrolisi di ATP:
ATP ---> ADP + P i
[ATP] = 2.25 mM (intracellulare);
[P i ] = 1.65 mM;
[ADP] = 0.25 mM.
(l'ADP viene rifosforilato immediatamente in quanti tà ingenti - serve che l'ATP sia subito riformato)
Idrolisi ATP:
ΔGvero = -30.5 kJ/mol - 22kJ/mol
= ΔG°' + RTln(K)
= [il grosso dell'energia] + [il piccolo dell'energia, non affato trascurabile]
= [Obelix] + [Asterix]
=
OSS.2 : ΔG E' UNA GRANDEZZA SCALARE ADDITIVA
1) REAZIONI SEQUENZIALI :
A ---> B ---> C
ΔGTOT = ΔGA->B + ΔGB->C
2) REAZIONI ACCOPPIATE:
-[ A ---> B
11
-[ C ---> D
A + C ---> B + D
ΔGTOT = ΔGA->B + ΔGC->D
VANTAGGI / SVANTAGGI
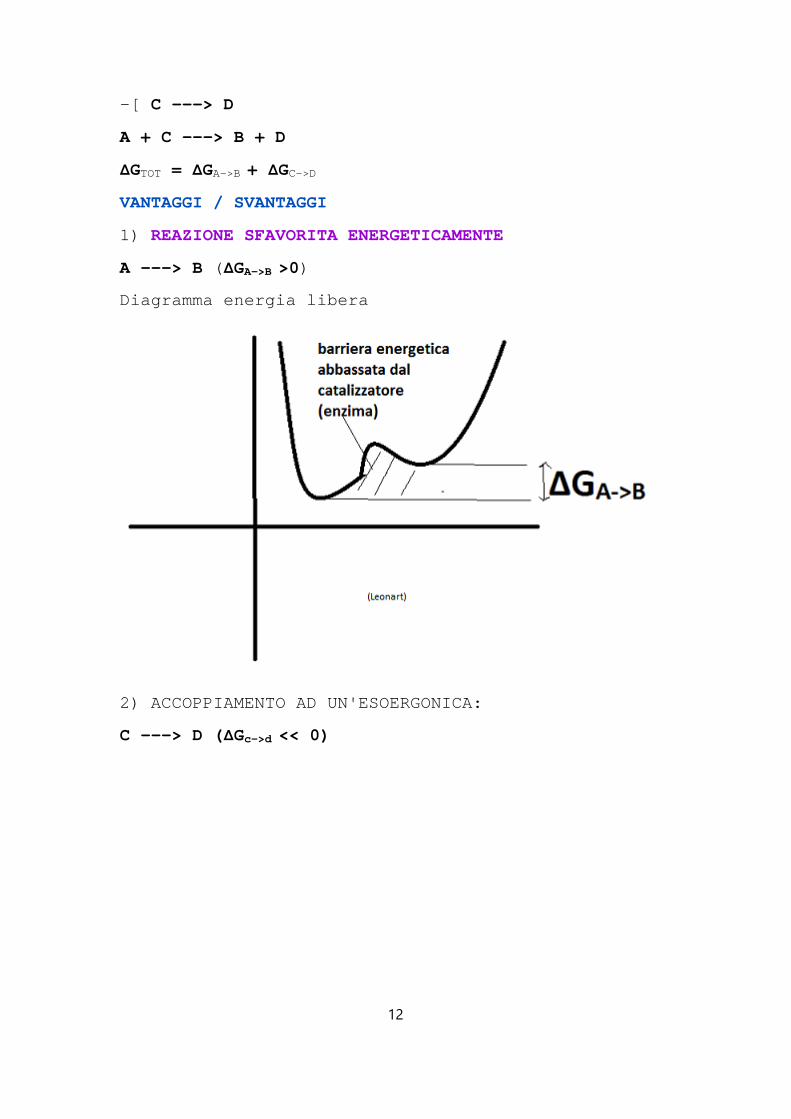
1) REAZIONE SFAVORITA ENERGETICAMENTE
A ---> B ( ΔGA->B >0)
Diagramma energia libera
2) ACCOPPIAMENTO AD UN'ESOERGONICA:
C ---> D ( ΔGc->d << 0)
12
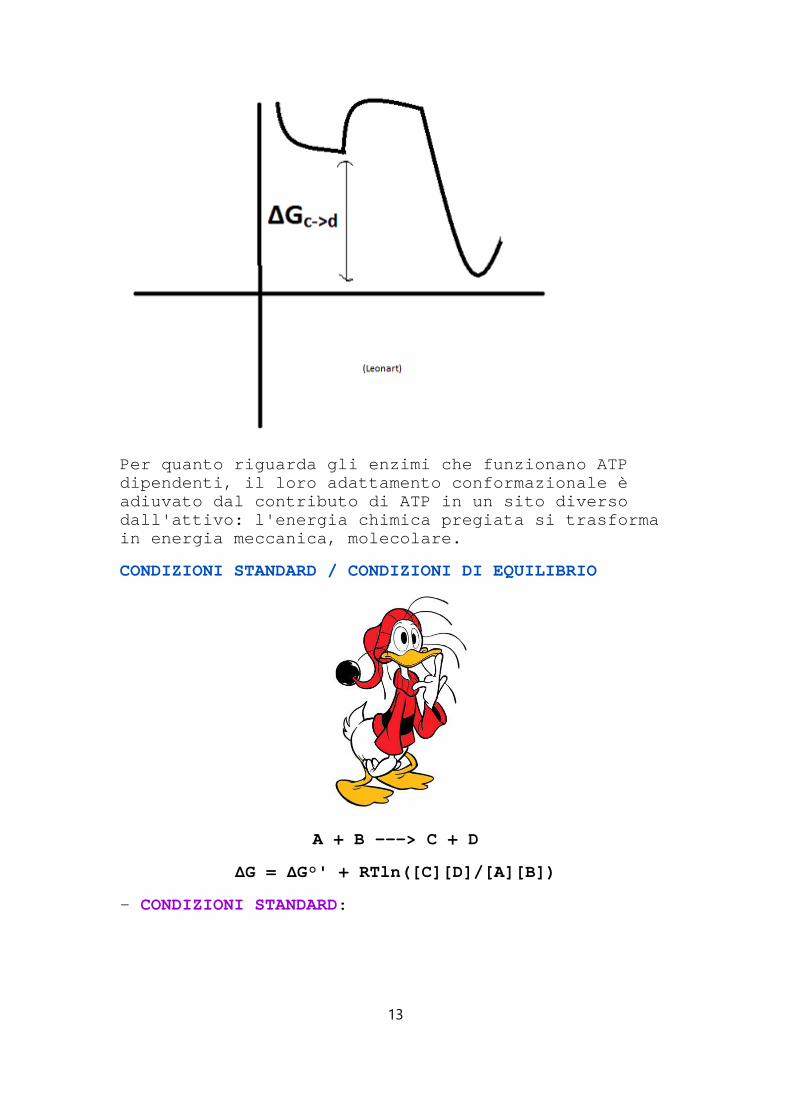
Per quanto riguarda gli enzimi che funzionano ATP dipendenti, il loro adattamento conformazionale è adiuvato dal contributo di ATP in un sito diverso dall'attivo: l'energia chimica pregiata si trasform a in energia meccanica, molecolare.
CONDIZIONI STANDARD / CONDIZIONI DI EQUILIBRIO
A + B ---> C + D
ΔG = ΔG°' + RTln([C][D]/[A][B])
- CONDIZIONI STANDARD:
13

Tutti i soluti e i prodotti sono 1 M
( [A] = [B] = [C] = [D] = 1 M )
=> ln(1) = 0 -> ΔG ≡ ΔG°'
- CONDIZIONI DI EQUILIBRIO:
ΔG = 0 => ΔG°' = -RTln([C][D]/[A][B])
All'equilibrio le concentrazioni delle diverse spec ie molecolari determinano un particolare valore di K,. ..
ΔG°' = -RTln(K eq)
ESEMPIO #1: PRIMA REAZIONE DELLA GLICOLISI
Glc + ATP ---> G6P + ADP
Calcolare la ΔG°' TOT a 25° C (R = 8.31 J/mol°C)
- ΔG°' idrolisi ATP = -30.5 kJ/mol
(ATP ---> ADP + P i )
- ΔG° idrolisi G6P = -14.0 kJ/mol
(G6P ---> Glc + P i )
A me interessa il processo inverso all'esoergonicit à
14
espressa nelle variazioni di entalpia:
ΔG°' TOT = ΔG°' idrolisi ATP - ΔG°' idrolisi G6P
ΔG°' TOT = -16500 kJ/mol
Keq = e -( ΔG°'/RT) = 800
IV.ALTRO NUMERO DI MAGIA DI ATP: LA DONNA SEGATA IN DUE
Bisogna formare una molecola più energetica dell'AT P. Come ci riescono le cellule?
IDROLISI ATP ad AMP :
ATP ---> AMP + PP i
dove PP i è il pirofosfato
Interviene poi la pirofosfatasi inorganica (l'enzima compagno) che taglia in due il gruppo fosfato. A questa azione fa generalmente seguito quella esoergonica.
ΔG°' = -19 kJ/mol (della pirofosfatasi inorganica)
[ Per intenderci, la pirofosfatasi inorganica è la maga Circe, il mágos che fa la magia della donna
15
segata in due con la ninfa di Leucò (il pirofosfato ), in uno dei loro fantastici dialoghi narrati da Cesa re Pavese... Trovate la scena raffigurata anche in "Topolino": scusatemi ma non ho il numero che la contiene, quindi non posso riportarvi la foto! ]
PPi + H2O ---> (p.inorg.) Pi + P i
Staccato PP i , non c'è più possibilità di riformare ATP (la reazione è fortemente spostata verso destra , esoergonica, fortemente irreversibile). Per riforma re eventualmente ATP da AMP due gruppi fosfato si dovrebbero rilegare all'AMP uno alla volta e ciò è molto sfavorito dal punto di vista energetico.
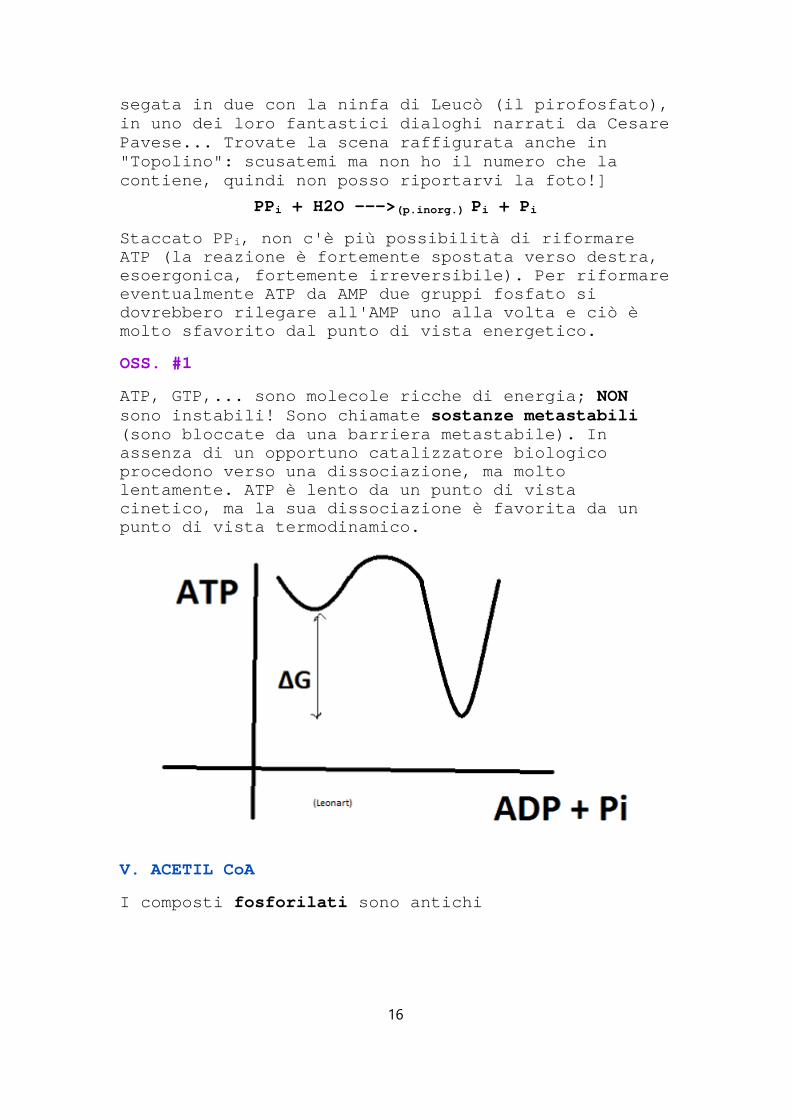
OSS. #1
ATP, GTP,... sono molecole ricche di energia; NONsono instabili! Sono chiamate sostanze metastabili (sono bloccate da una barriera metastabile). In assenza di un opportuno catalizzatore biologico procedono verso una dissociazione, ma molto lentamente. ATP è lento da un punto di vista cinetico, ma la sua dissociazione è favorita da un punto di vista termodinamico.
V. ACETIL CoA
I composti fosforilati sono antichi
16

L'approccio con cui venivano sintetizzati era per via anaerobica (ancora in uso) -fosforilazione a livello di substrato .
Una di queste sostanze primitive è una molecolona ubiquitaria che tuttora svolge un compito centrale in tutti i viventi:
17
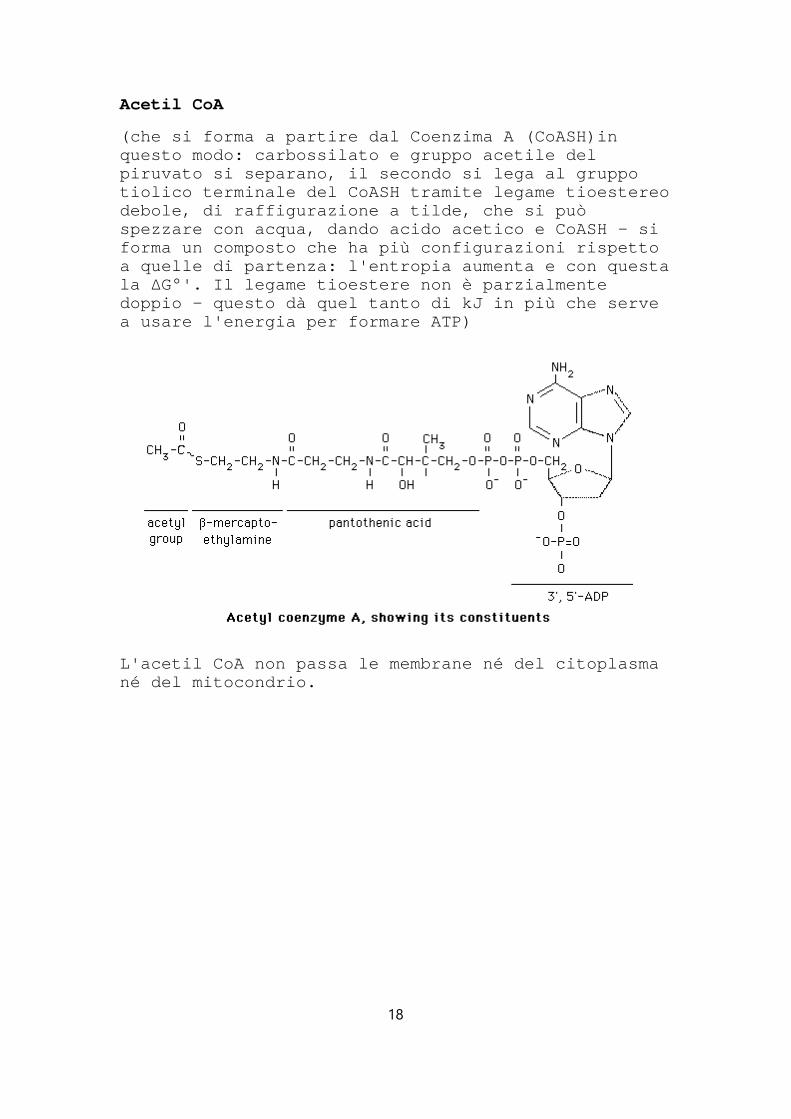
Acetil CoA
(che si forma a partire dal Coenzima A (CoASH)in questo modo: carbossilato e gruppo acetile del piruvato si separano, il secondo si lega al gruppo tiolico terminale del CoASH tramite legame tioester eo debole, di raffigurazione a tilde, che si può spezzare con acqua, dando acido acetico e CoASH - si forma un composto che ha più configurazioni rispett o a quelle di partenza: l'entropia aumenta e con ques ta la ΔG°'. Il legame tioestere non è parzialmente doppio - questo dà quel tanto di kJ in più che serve a usare l'energia per formare ATP)
L'acetil CoA non passa le membrane né del citoplasm a né del mitocondrio.
18


















