


Lectures 7-8: Charge and Energy Transfer, Photosynthesis, Biofules
32
Transcript of Lectures 7-8: Charge and Energy Transfer, Photosynthesis, Biofules
Organic Dyes – Organic Semiconductors
• Organic photovoltaics
• Photosynthesis
Common questions:
• How is energy transported?
• How are charges transferred between molecules?
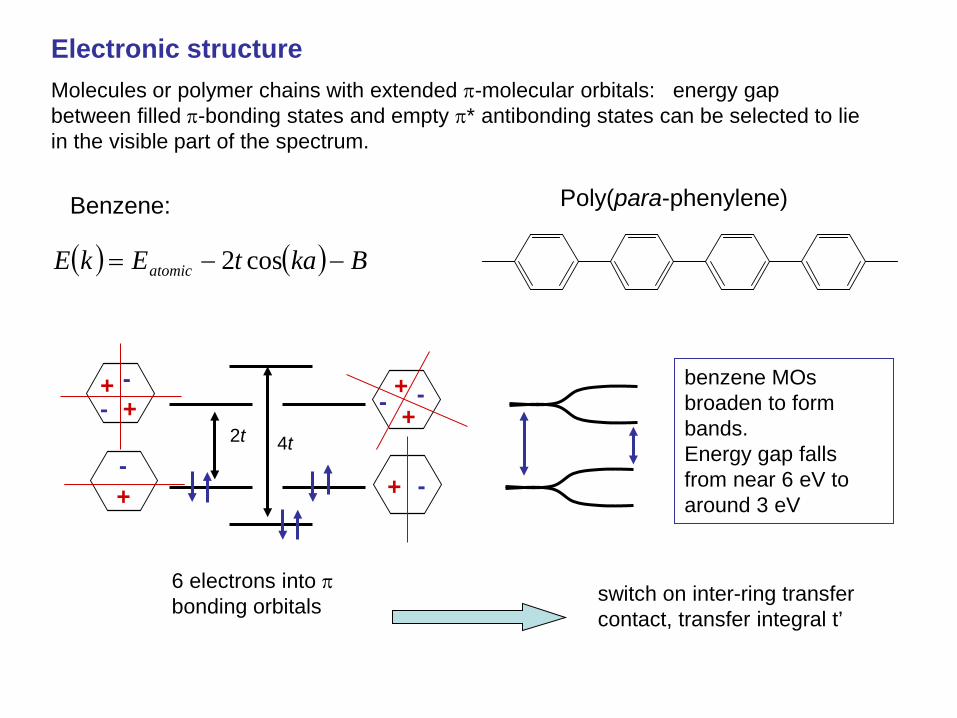
Benzene:
+ + -
- + +
- -
+ - + -
2t 4t
( ) ( ) BkatEkE atomic −−= cos2
6 electrons into π bonding orbitals
Poly(para-phenylene)
switch on inter-ring transfer contact, transfer integral t’
benzene MOs broaden to form bands. Energy gap falls from near 6 eV to around 3 eV
Electronic structure Molecules or polymer chains with extended π-molecular orbitals: energy gap between filled π-bonding states and empty π* antibonding states can be selected to lie in the visible part of the spectrum.

Excitons
( )eVmeEbinding 6.13/
42 220
4
×==ε
µπεε
µ
where µ is the reduced mass for the e-h system: ∗∗ +=
he mm111
µ
Recall for inorganic semiconductors: • optical excitation can produce a bound electron-hole state
• Typical energies 2-40 meV • Exciton radii: a few nm (many atomic sites)
Organics:
• Dielectric constant much lower (εr ~ 3 - 4) • Mott-Wannier model above (me*= mh* = me) gives Eb ~ 0.75 eV, r ~ 0.3 nm • Localised on one molecule – “Frenkel exciton” • Localisation of the exciton to relatively few bonds causes significant
coupling of the molecular geometry to the electronic configuration.
Difficult to generate free electrons and holes, even at room temperature
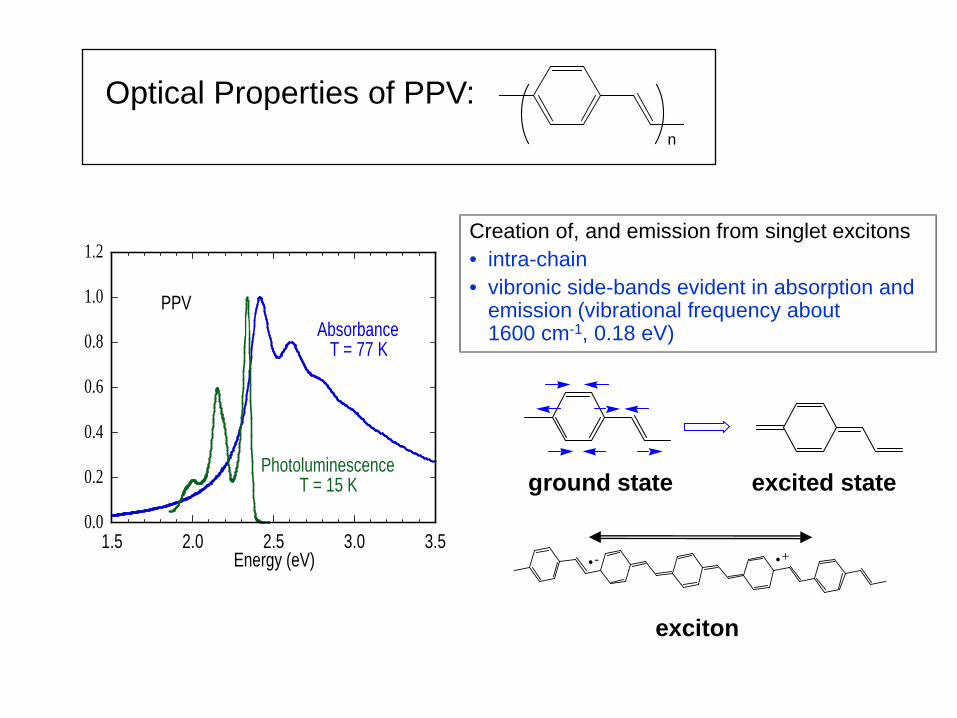
.- .+
ground state excited state
exciton
Optical Properties of PPV: n
Creation of, and emission from singlet excitons • intra-chain • vibronic side-bands evident in absorption and
emission (vibrational frequency about 1600 cm-1, 0.18 eV)
1.5 2.0 2.5 3.0 3.50.0
0.2
0.4
0.6
0.8
1.0
1.2
Energy (eV)
PPV(high structure) Absorbance
T = 77 K
PhotoluminescenceT = 15 K
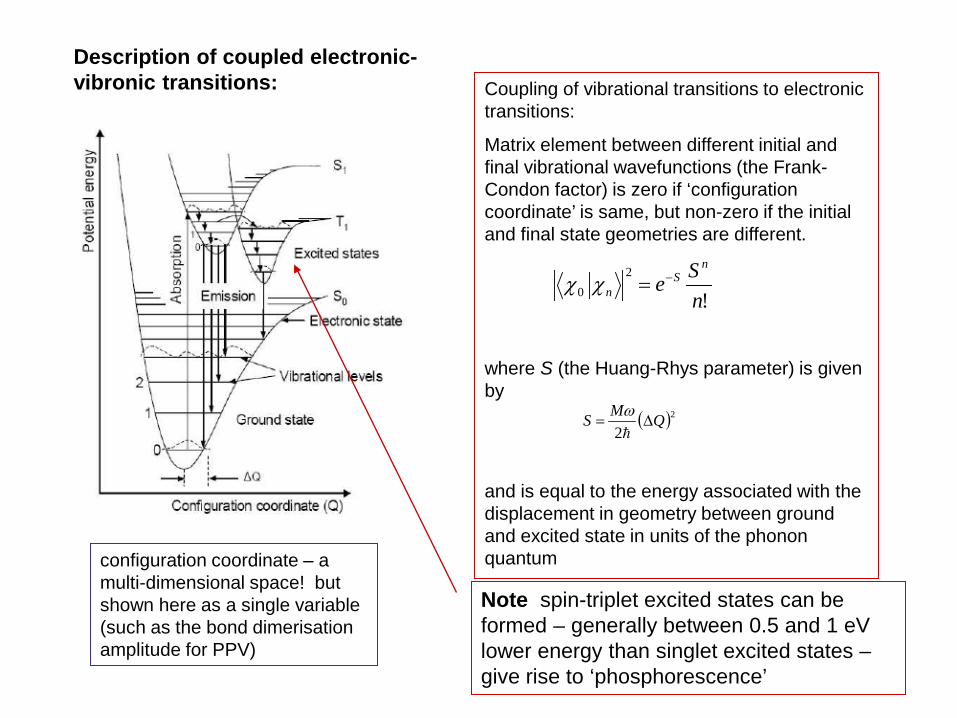
Coupling of vibrational transitions to electronic transitions:
Matrix element between different initial and final vibrational wavefunctions (the Frank-Condon factor) is zero if ‘configuration coordinate’ is same, but non-zero if the initial and final state geometries are different.
where S (the Huang-Rhys parameter) is given by
and is equal to the energy associated with the displacement in geometry between ground and excited state in units of the phonon quantum
Description of coupled electronic-vibronic transitions:
configuration coordinate – a multi-dimensional space! but shown here as a single variable (such as the bond dimerisation amplitude for PPV)
!2
0 nSe
nS
n−=χχ
( )2
2QMS ∆=
ω
Note spin-triplet excited states can be formed – generally between 0.5 and 1 eV lower energy than singlet excited states – give rise to ‘phosphorescence’
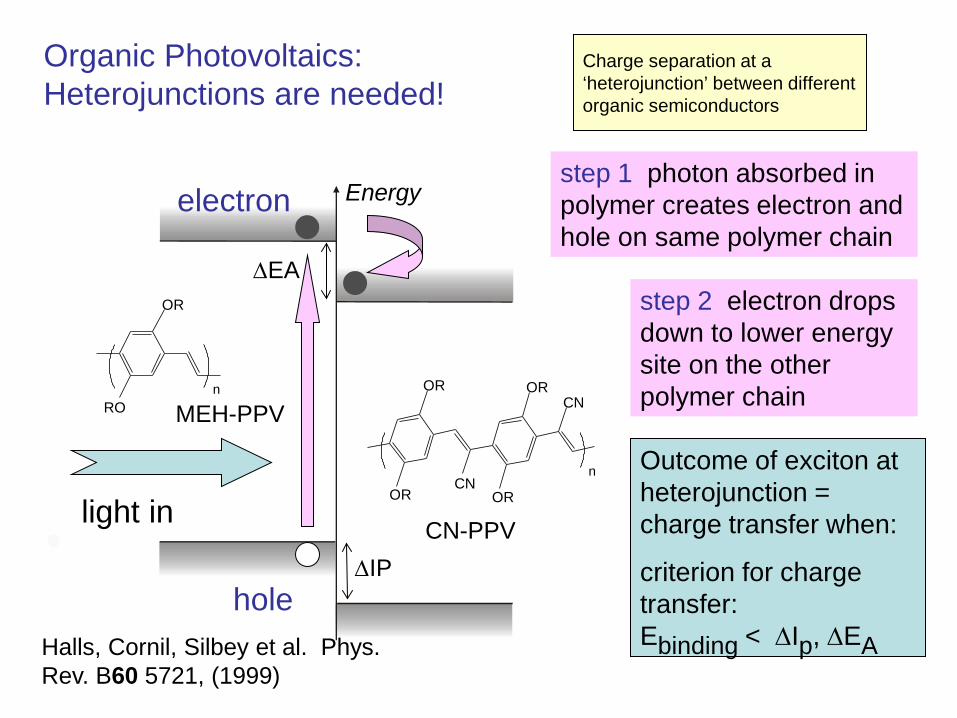
Charge separation at a ‘heterojunction’ between different organic semiconductors
CN-PPV
n
OR
OR OR
OR
CN
CN
Energy
n
OR
RO MEH-PPV
step 2 electron drops down to lower energy site on the other polymer chain
hole
electron step 1 photon absorbed in polymer creates electron and hole on same polymer chain
light in
Organic Photovoltaics: Heterojunctions are needed!
Halls, Cornil, Silbey et al. Phys. Rev. B60 5721, (1999)
Outcome of exciton at heterojunction = charge transfer when:
criterion for charge transfer: Ebinding < ∆Ip, ∆EA
∆IP
∆EA
-
+
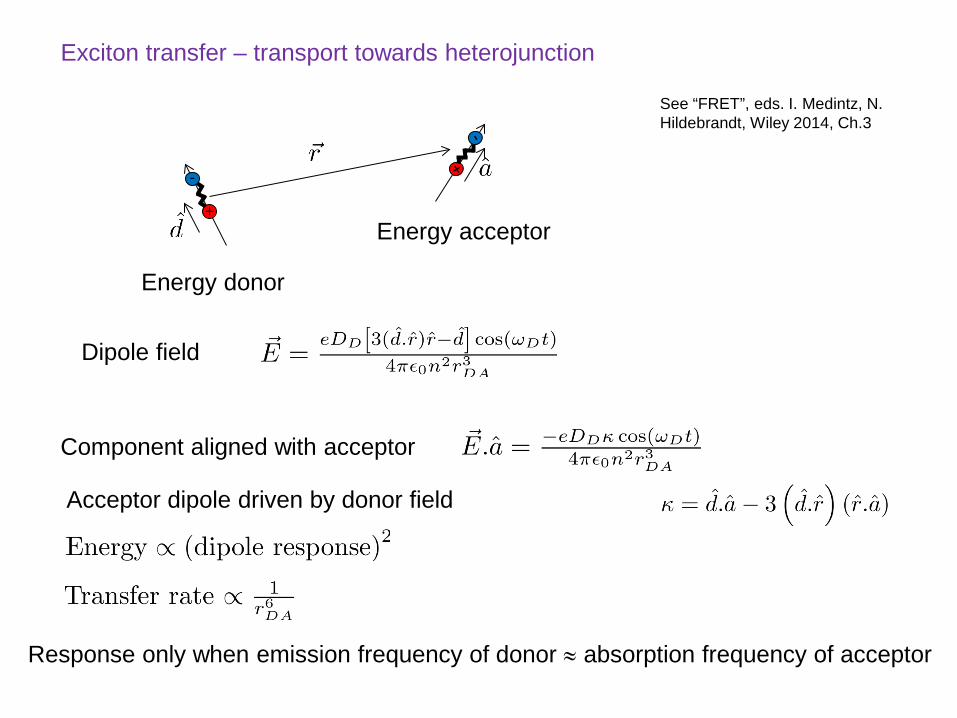
Exciton transfer – transport towards heterojunction
Energy donor
Energy acceptor
Dipole field
Component aligned with acceptor
Response only when emission frequency of donor ≈ absorption frequency of acceptor
Acceptor dipole driven by donor field
See “FRET”, eds. I. Medintz, N. Hildebrandt, Wiley 2014, Ch.3

Values of the Forster radius, R0, are in the range 2-4 nm.
NB. no photon is actually emitted and reabsorbed
Forster Transfer
donor lifetime
donor quantum efficiency
refractive index
Avogadro’s number
frequency
normalised donor emission spectrum
acceptor molar absorption coefficient

Exciton diffusion
• Exciton lifetime ~ 1ns • Hops between many sites (by Förster transfer) • Typical diffusion range 5 – 10 nm
• For efficient PV operation, must find interface within 5 – 10 nm • But, absorption depth still ~100 nm
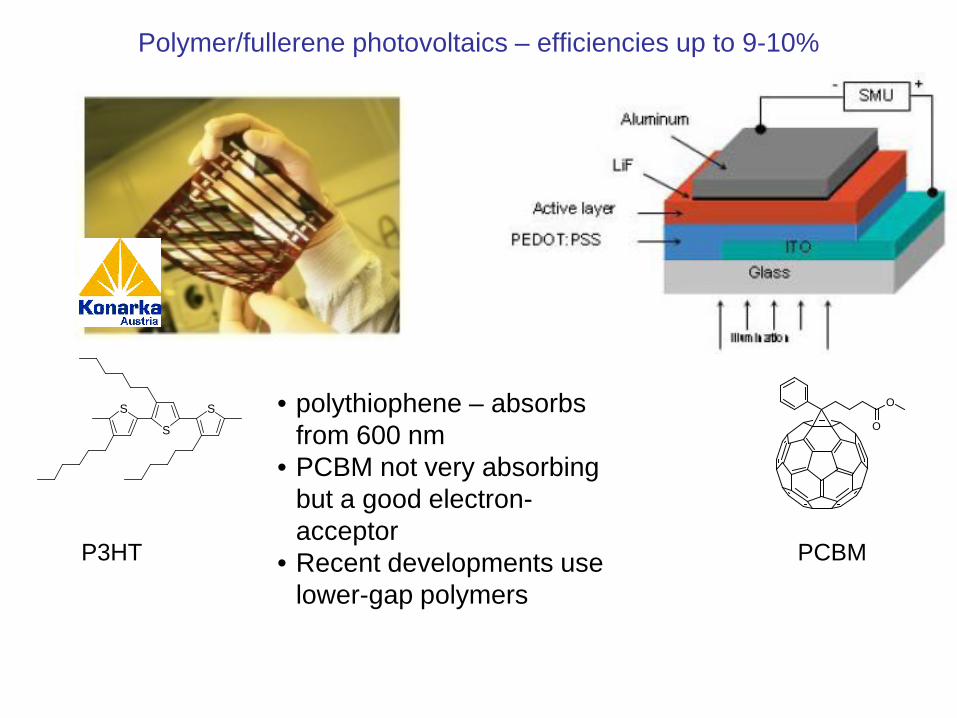
Polymer/fullerene photovoltaics – efficiencies up to 9-10%
• polythiophene – absorbs from 600 nm
• PCBM not very absorbing but a good electron-acceptor
• Recent developments use lower-gap polymers
S
S S O
O
P3HT PCBM
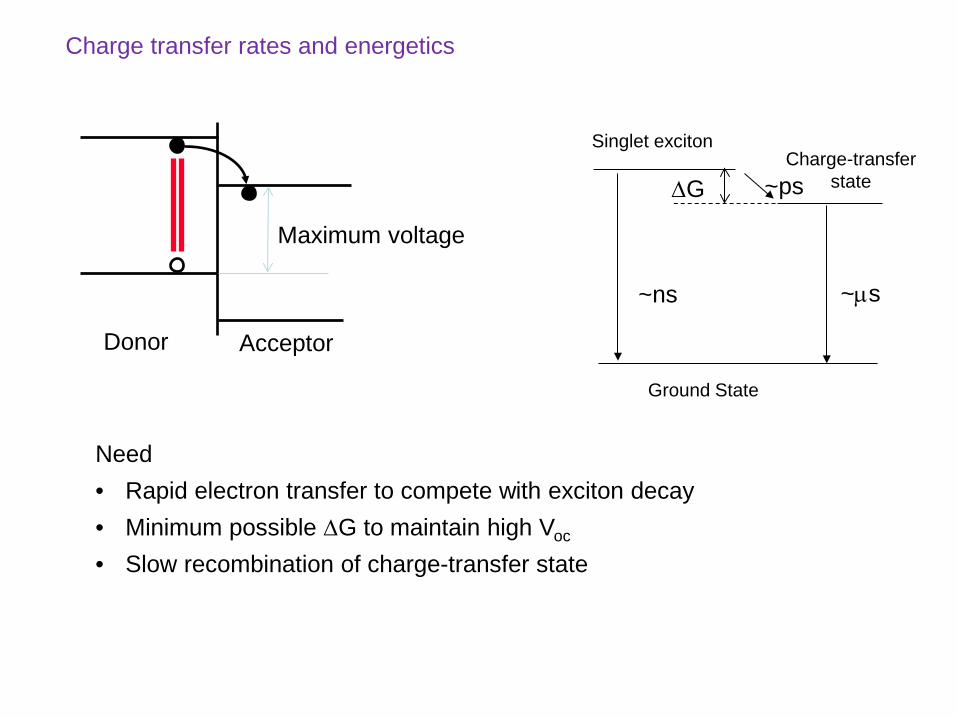
Donor Acceptor
Maximum voltage
Ground State
Singlet exciton Charge-transfer
state ∆G
~ns ~µs
Need • Rapid electron transfer to compete with exciton decay • Minimum possible ∆G to maintain high Voc
• Slow recombination of charge-transfer state
~ps
Charge transfer rates and energetics
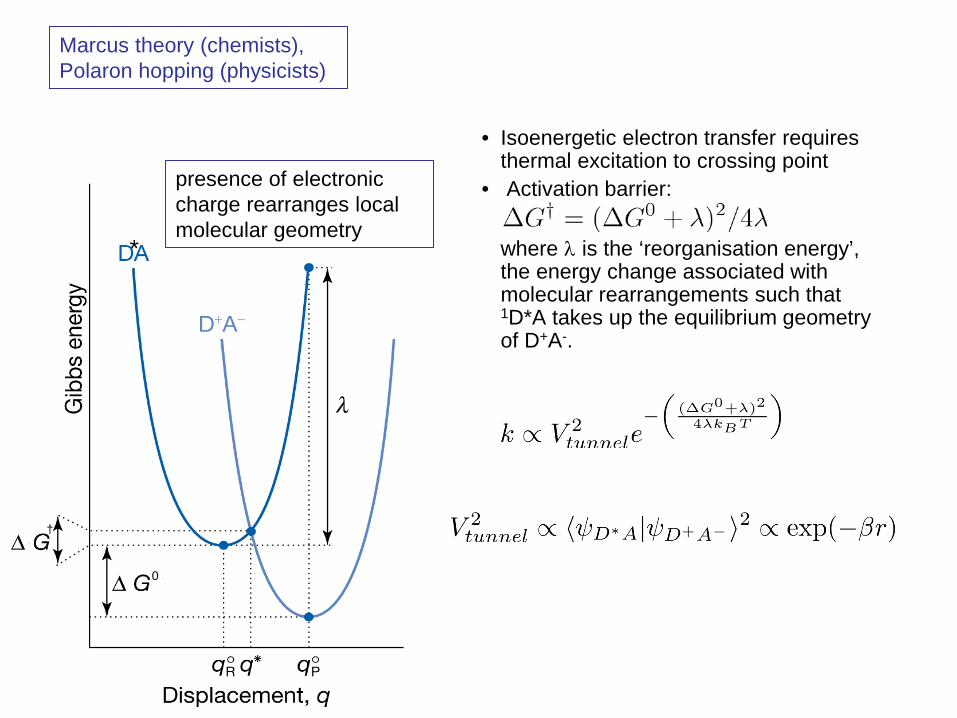
• Isoenergetic electron transfer requires thermal excitation to crossing point
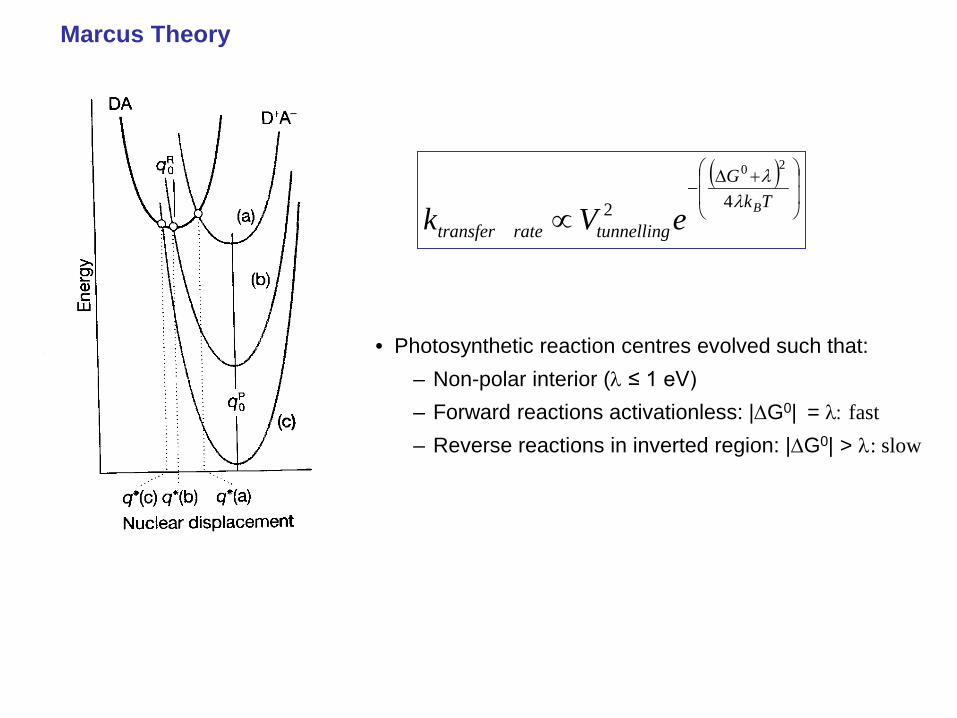
• Activation barrier: where λ is the ‘reorganisation energy’,
the energy change associated with molecular rearrangements such that 1D*A takes up the equilibrium geometry of D+A-.
1D*A
D+A-
Marcus theory (chemists), Polaron hopping (physicists)
presence of electronic charge rearranges local molecular geometry
†
0
*
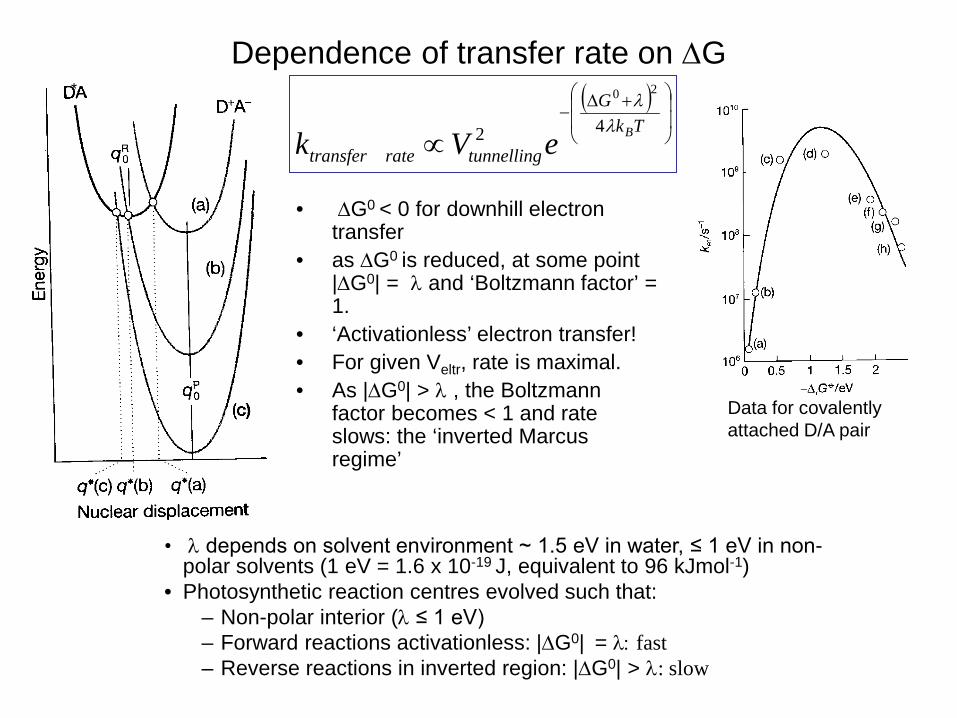
Dependence of transfer rate on ∆G
Data for covalently attached D/A pair
• λ depends on solvent environment ~ 1.5 eV in water, ≤ 1 eV in non-polar solvents (1 eV = 1.6 x 10-19 J, equivalent to 96 kJmol-1)
• Photosynthetic reaction centres evolved such that: – Non-polar interior (λ ≤ 1 eV) – Forward reactions activationless: |∆G0| = λ: fast – Reverse reactions in inverted region: |∆G0| > λ: slow
• ∆G0 < 0 for downhill electron transfer
• as ∆G0 is reduced, at some point
|∆G0| = λ and ‘Boltzmann factor’ = 1.
• ‘Activationless’ electron transfer! • For given Veltr, rate is maximal. • As |∆G0| > λ , the Boltzmann
factor becomes < 1 and rate slows: the ‘inverted Marcus regime’
( )
+∆−
∝Tk
G
tunnellingratetransferBeVk
λλ
42
20*

Photosynthesis:
• Capture of sunlight by light-absorbing ‘antennae’ molecules
• transfer of excitons to the ‘reaction centre’
• charge separation across a ‘semiconductor heterojunction’
• chemical reactions driven by net positive charge (e.g. oxidation product causes reduction of water to oxygen)
• chemical reactions driven by negative charge: (e.g. formation of hydroquinones from quinones as intermediate chemical feedstocks)
• generation of ATP from ADP and NADPH from NADP+
• ATP and NADPH used in the Calvin cycle to convert CO2 to glucoses
Many different systems:
not all of them are green… colour is selected according to available sunlight – bacterial systems come in many colours (as seen in coral reefs..)
not all of them produce oxygen: purple bacteria use a wide range of reductants such as hydrogen sulphide, other organic matter (sewage ponds..)
6CO2 + 6H2O + photons → C6H12O6 + 6O2
Useful book: Molecular Mechanisms of Photosynthesis, R. E. Blankenship

Efficiency losses in photosynthesis:
• Absorption only at wavelengths < 700 nm
• Shockley-Queisser limit (1.8 eV) ~ 24%
• Efficiency of converting 1.8 eV photons into glucose = 27% (9 - 10 photons required per molecule)
• 24% × 27% = 6.5%
• Not all photons absorbed (plants are green, not black)
⇒ net efficiency ~ 5%
Further losses reduce realised efficiencies to typically below 1%:
- shading
- seasonal growing conditions (winter, rainfall, excess sunlight)
Chloroplast
Separate question: Efficiency of capture of carbon in plant matter: constant CO2 concentration in the atmosphere implies steady state in the pre-industrial past?
Many mechanisms for re-generation of CO2. Plant respiration, plant decay, combustion etc.
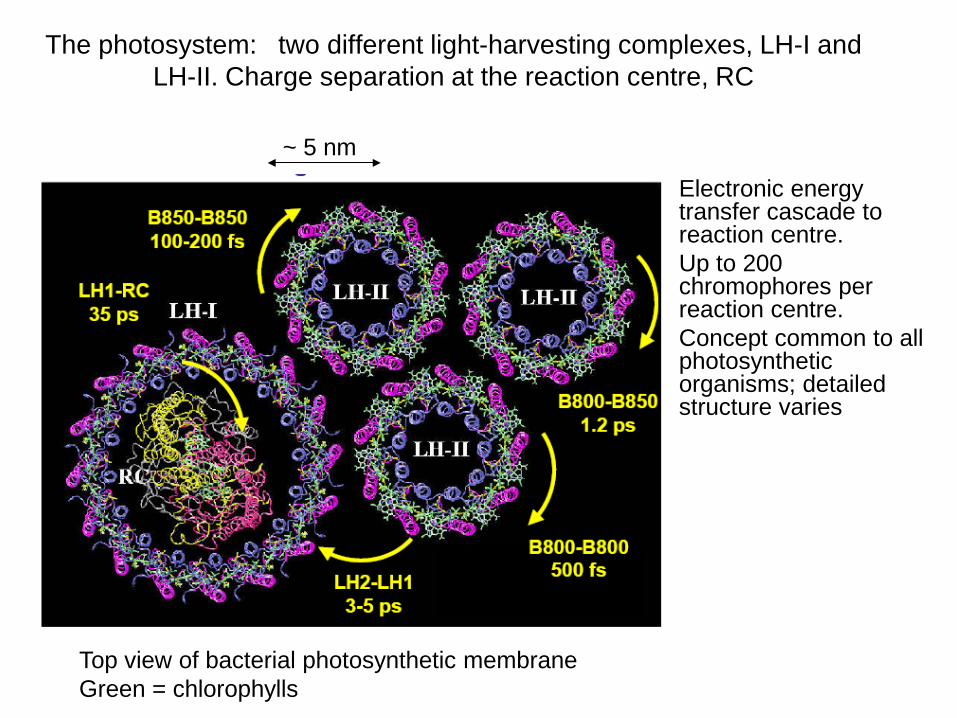
The photosystem: two different light-harvesting complexes, LH-I and LH-II. Charge separation at the reaction centre, RC
• Electronic energy transfer cascade to reaction centre.
• Up to 200 chromophores per reaction centre.
• Concept common to all photosynthetic organisms; detailed structure varies
Top view of bacterial photosynthetic membrane Green = chlorophylls
~ 5 nm
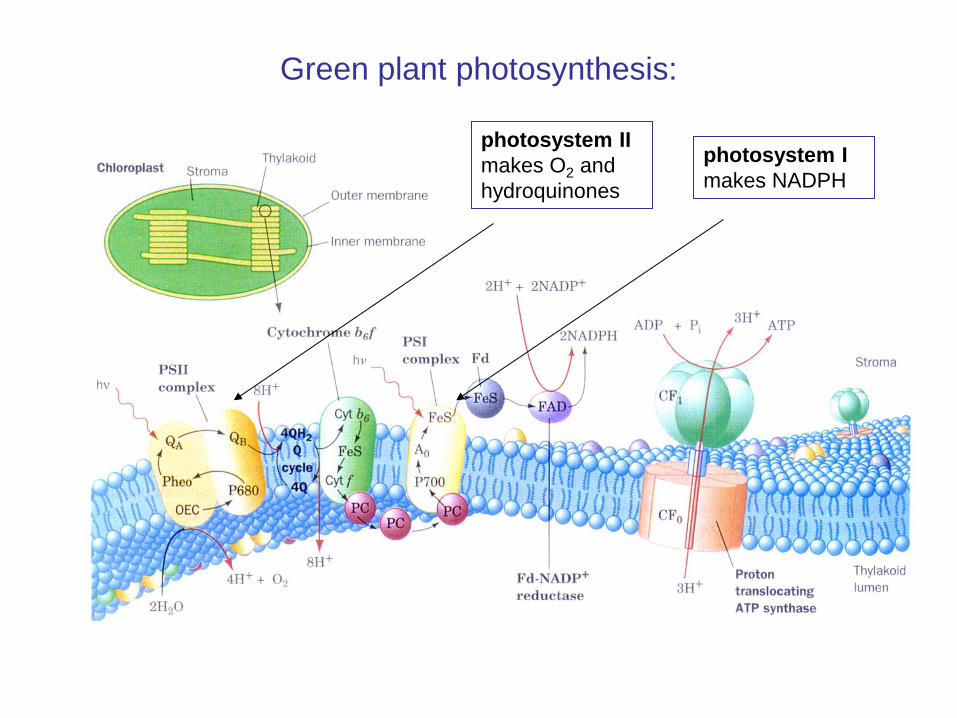
Green plant photosynthesis:
photosystem II makes O2 and hydroquinones
photosystem I makes NADPH
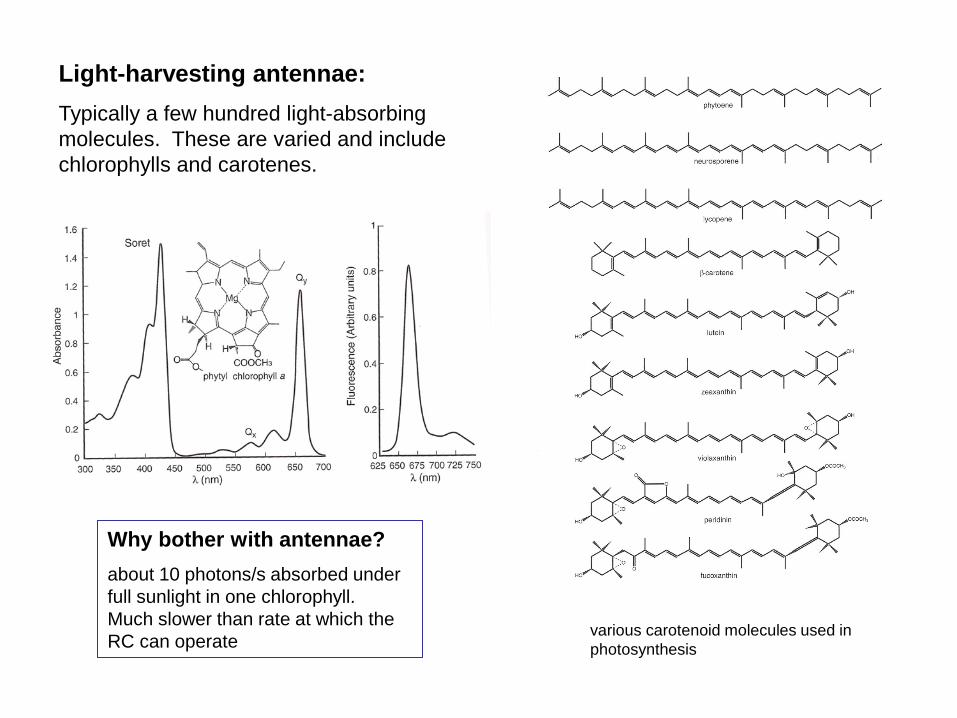
Light-harvesting antennae: Typically a few hundred light-absorbing molecules. These are varied and include chlorophylls and carotenes.
various carotenoid molecules used in photosynthesis
Why bother with antennae? about 10 photons/s absorbed under full sunlight in one chlorophyll. Much slower than rate at which the RC can operate
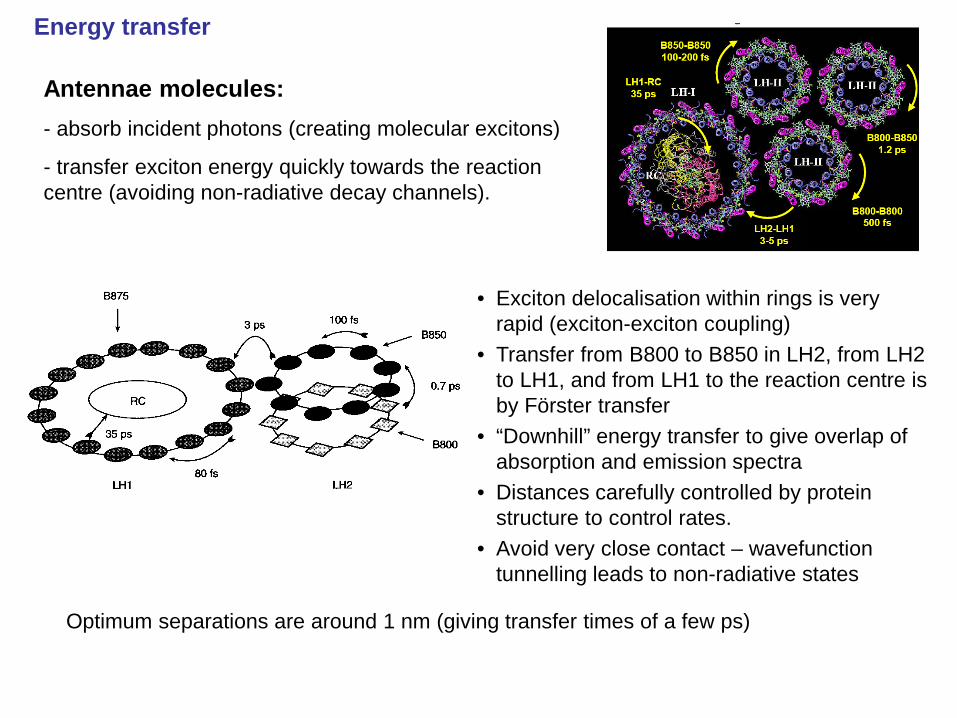
Energy transfer
• Exciton delocalisation within rings is very rapid (exciton-exciton coupling)
• Transfer from B800 to B850 in LH2, from LH2 to LH1, and from LH1 to the reaction centre is by Förster transfer
• “Downhill” energy transfer to give overlap of absorption and emission spectra
• Distances carefully controlled by protein structure to control rates.
• Avoid very close contact – wavefunction tunnelling leads to non-radiative states
Antennae molecules: - absorb incident photons (creating molecular excitons)
- transfer exciton energy quickly towards the reaction centre (avoiding non-radiative decay channels).
Optimum separations are around 1 nm (giving transfer times of a few ps)
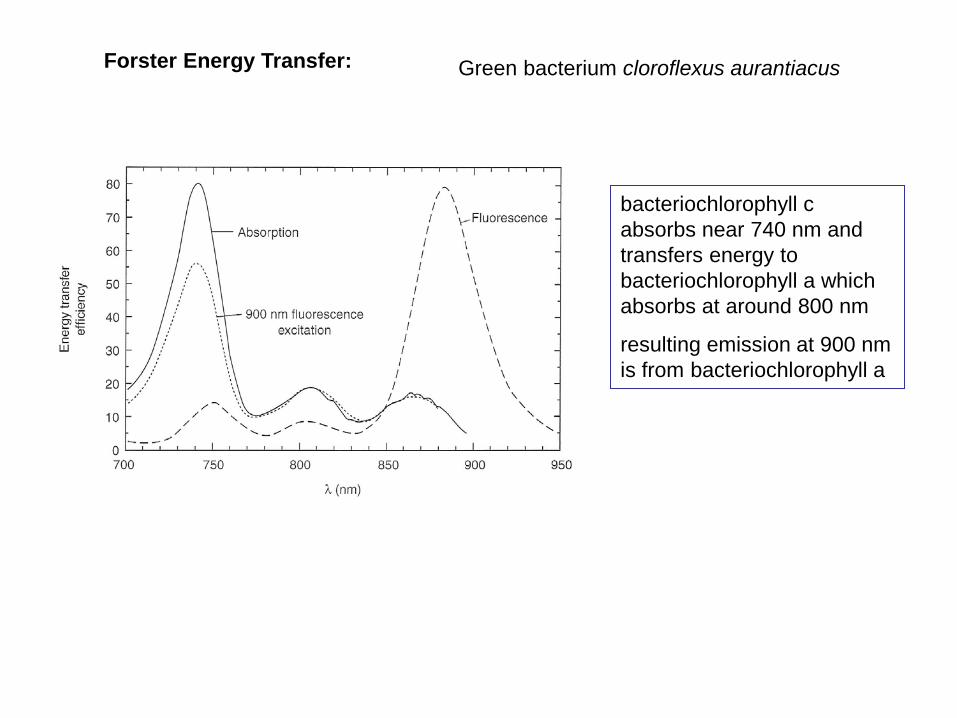
Forster Energy Transfer: Green bacterium cloroflexus aurantiacus
bacteriochlorophyll c absorbs near 740 nm and transfers energy to bacteriochlorophyll a which absorbs at around 800 nm
resulting emission at 900 nm is from bacteriochlorophyll a
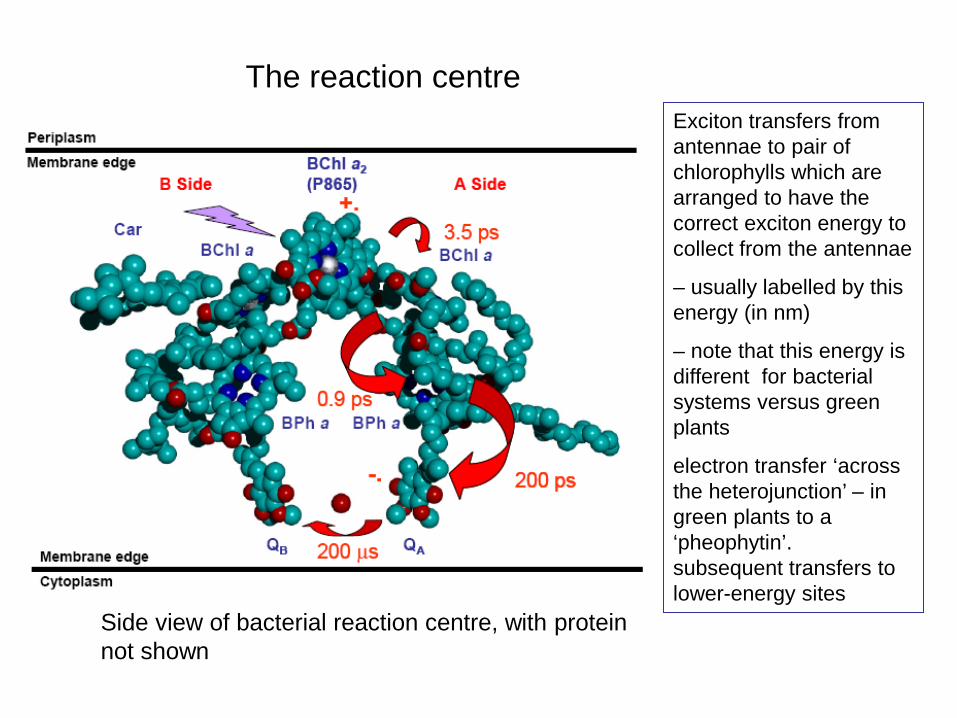
The reaction centre
Side view of bacterial reaction centre, with protein not shown
Exciton transfers from antennae to pair of chlorophylls which are arranged to have the correct exciton energy to collect from the antennae
– usually labelled by this energy (in nm)
– note that this energy is different for bacterial systems versus green plants
electron transfer ‘across the heterojunction’ – in green plants to a ‘pheophytin’. subsequent transfers to lower-energy sites
• Photosynthetic reaction centres evolved such that: – Non-polar interior (λ ≤ 1 eV) – Forward reactions activationless: |∆G0| = λ: fast – Reverse reactions in inverted region: |∆G0| > λ: slow
( )
+∆−
∝Tk
G
tunnellingratetransferBeVk
λλ
42
20
Marcus Theory
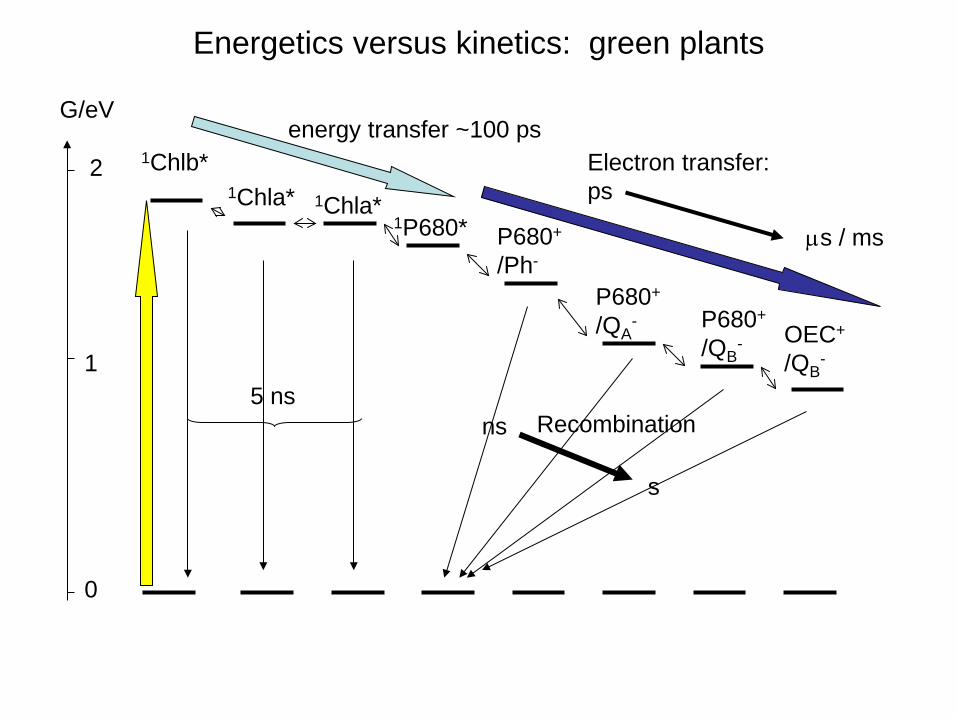
Energetics versus kinetics: green plants
1Chlb* 1Chla* 1Chla*
1P680* P680+ /Ph-
P680+ /QA
- P680+ /QB
- OEC+ /QB
-
ns
s
Recombination 5 ns
energy transfer ~100 ps Electron transfer: ps
µs / ms
G/eV
1
2
0
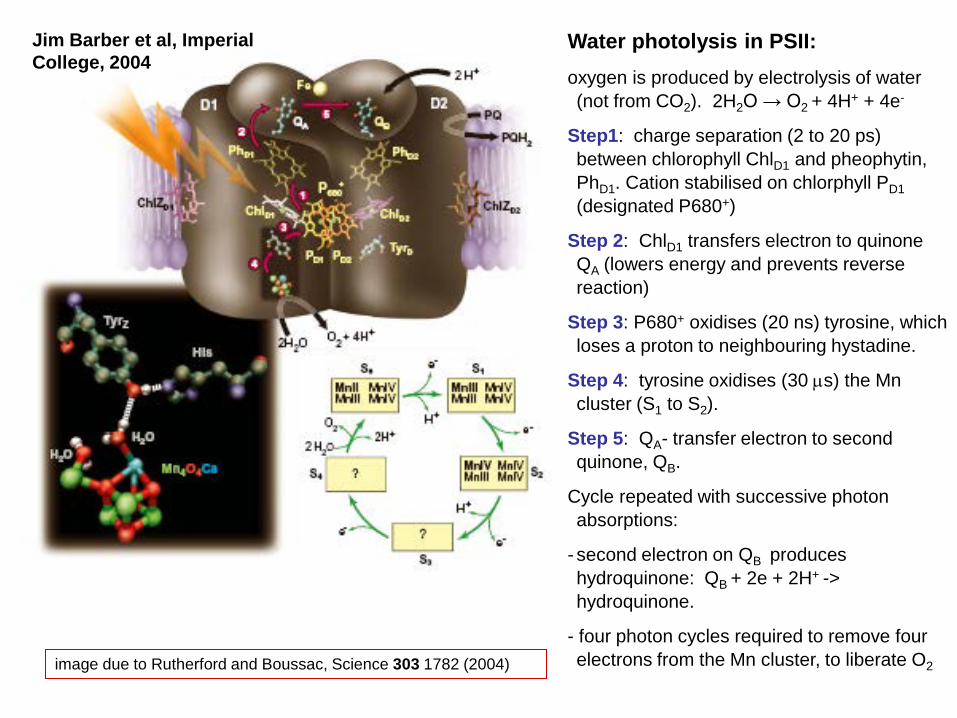
image due to Rutherford and Boussac, Science 303 1782 (2004)
Water photolysis in PSII: oxygen is produced by electrolysis of water (not from CO2). 2H2O → O2 + 4H+ + 4e-
Step1: charge separation (2 to 20 ps) between chlorophyll ChlD1 and pheophytin, PhD1. Cation stabilised on chlorphyll PD1 (designated P680+)
Step 2: ChlD1 transfers electron to quinone QA (lowers energy and prevents reverse reaction)
Step 3: P680+ oxidises (20 ns) tyrosine, which loses a proton to neighbouring hystadine.
Step 4: tyrosine oxidises (30 µs) the Mn cluster (S1 to S2).
Step 5: QA- transfer electron to second quinone, QB.
Cycle repeated with successive photon absorptions:
- second electron on QB produces hydroquinone: QB + 2e + 2H+ -> hydroquinone.
- four photon cycles required to remove four electrons from the Mn cluster, to liberate O2
Jim Barber et al, Imperial College, 2004
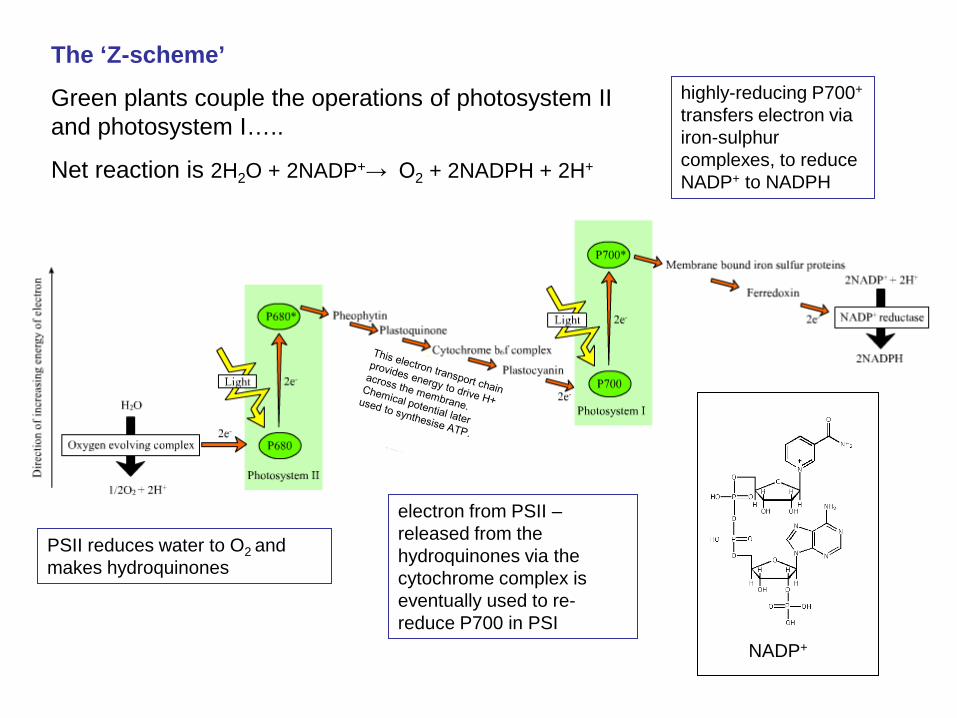
The ‘Z-scheme’
Green plants couple the operations of photosystem II and photosystem I…..
Net reaction is 2H2O + 2NADP+→ O2 + 2NADPH + 2H+
electron from PSII – released from the hydroquinones via the cytochrome complex is eventually used to re-reduce P700 in PSI
PSII reduces water to O2 and makes hydroquinones
highly-reducing P700+ transfers electron via iron-sulphur complexes, to reduce NADP+ to NADPH
NADP+
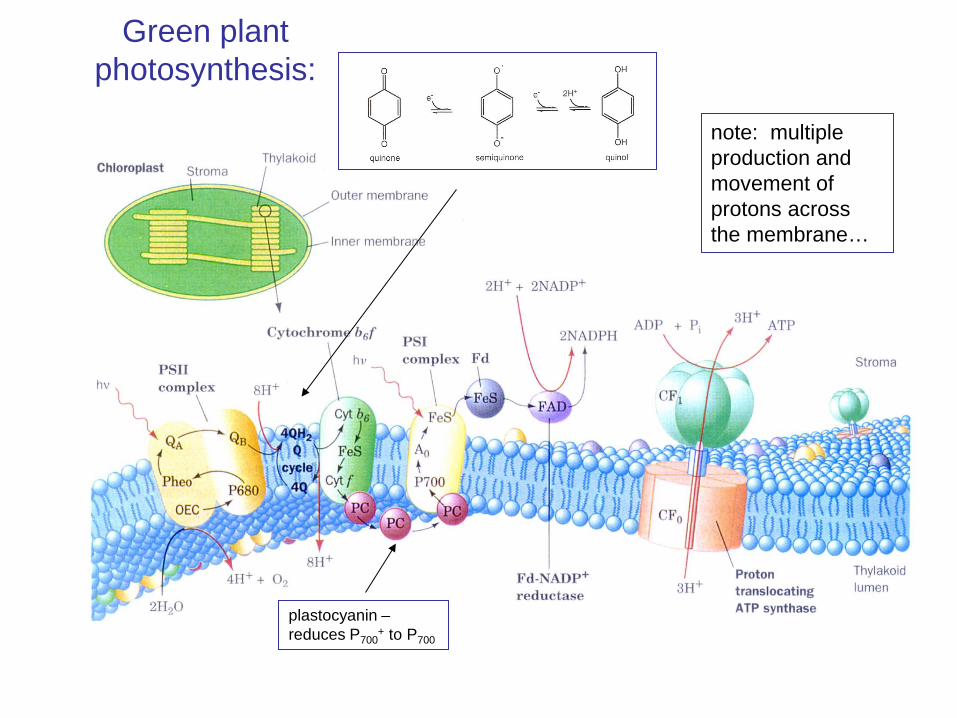
Green plant photosynthesis:
plastocyanin – reduces P700
+ to P700
note: multiple production and movement of protons across the membrane…
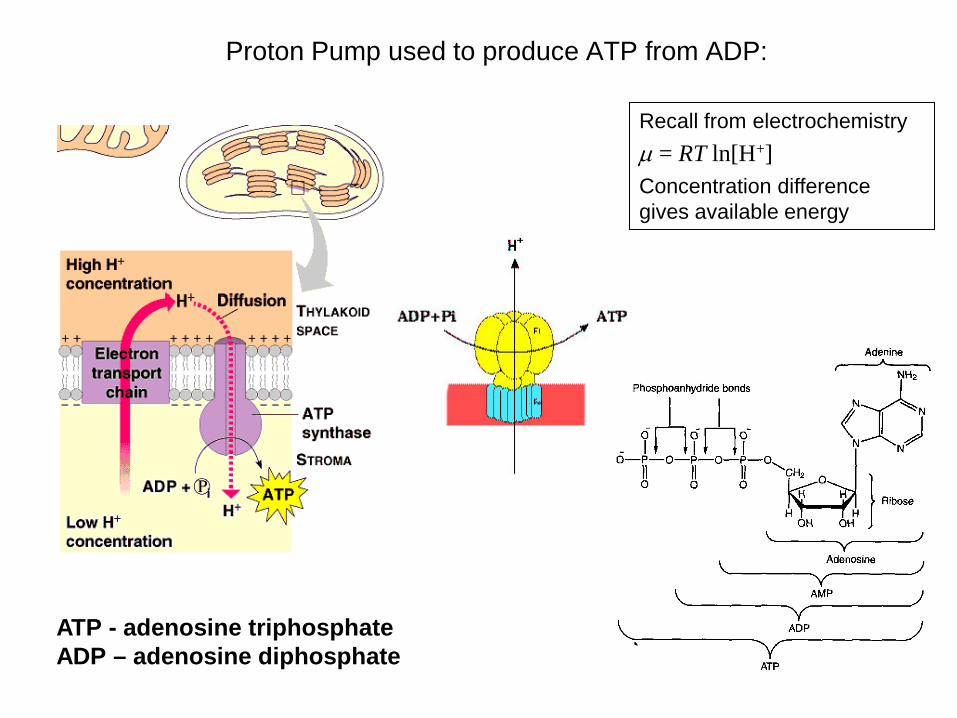
Proton Pump used to produce ATP from ADP:
ATP - adenosine triphosphate ADP – adenosine diphosphate
Recall from electrochemistry µ = RT ln[H+] Concentration difference gives available energy
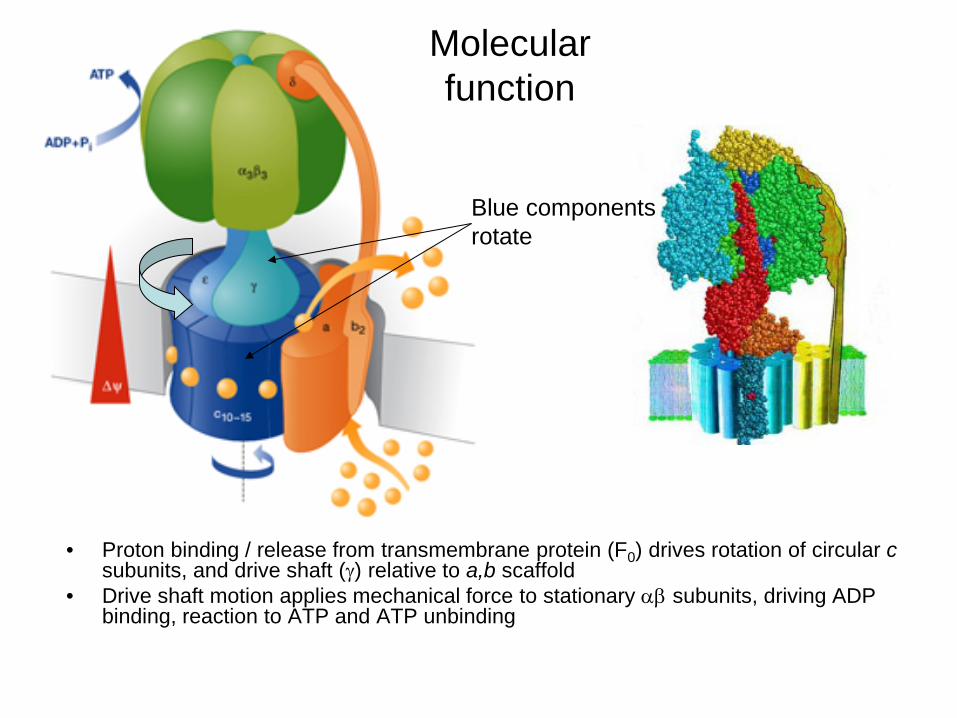
• Proton binding / release from transmembrane protein (F0) drives rotation of circular c subunits, and drive shaft (γ) relative to a,b scaffold
• Drive shaft motion applies mechanical force to stationary αβ subunits, driving ADP binding, reaction to ATP and ATP unbinding
Molecular function
Blue components rotate
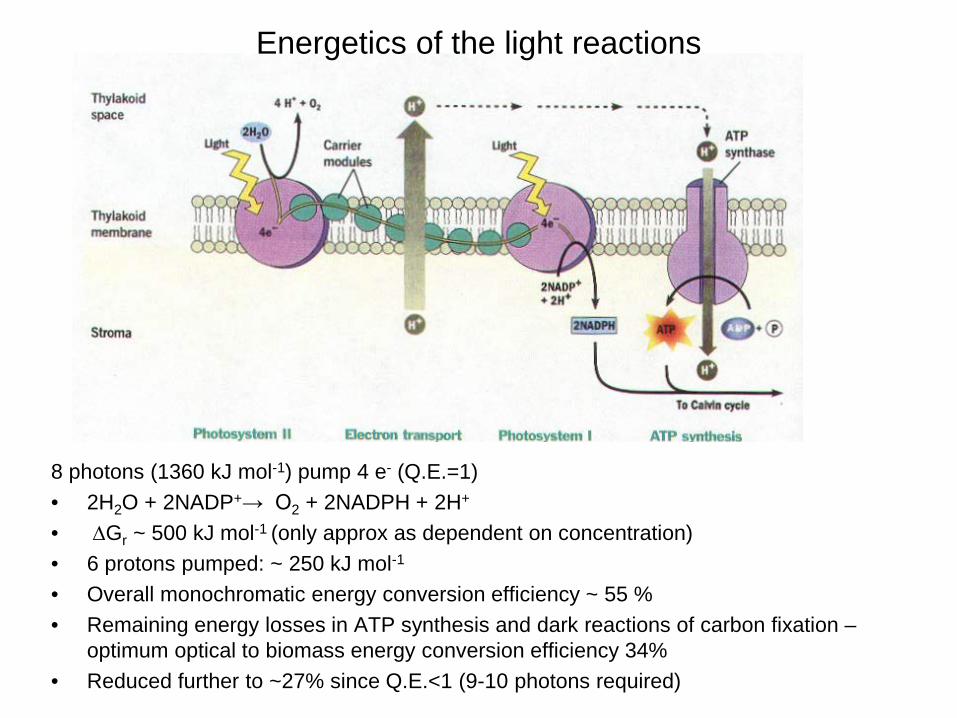
8 photons (1360 kJ mol-1) pump 4 e- (Q.E.=1) • 2H2O + 2NADP+→ O2 + 2NADPH + 2H+
• ∆Gr ~ 500 kJ mol-1 (only approx as dependent on concentration) • 6 protons pumped: ~ 250 kJ mol-1 • Overall monochromatic energy conversion efficiency ~ 55 % • Remaining energy losses in ATP synthesis and dark reactions of carbon fixation –
optimum optical to biomass energy conversion efficiency 34% • Reduced further to ~27% since Q.E.<1 (9-10 photons required)
Energetics of the light reactions
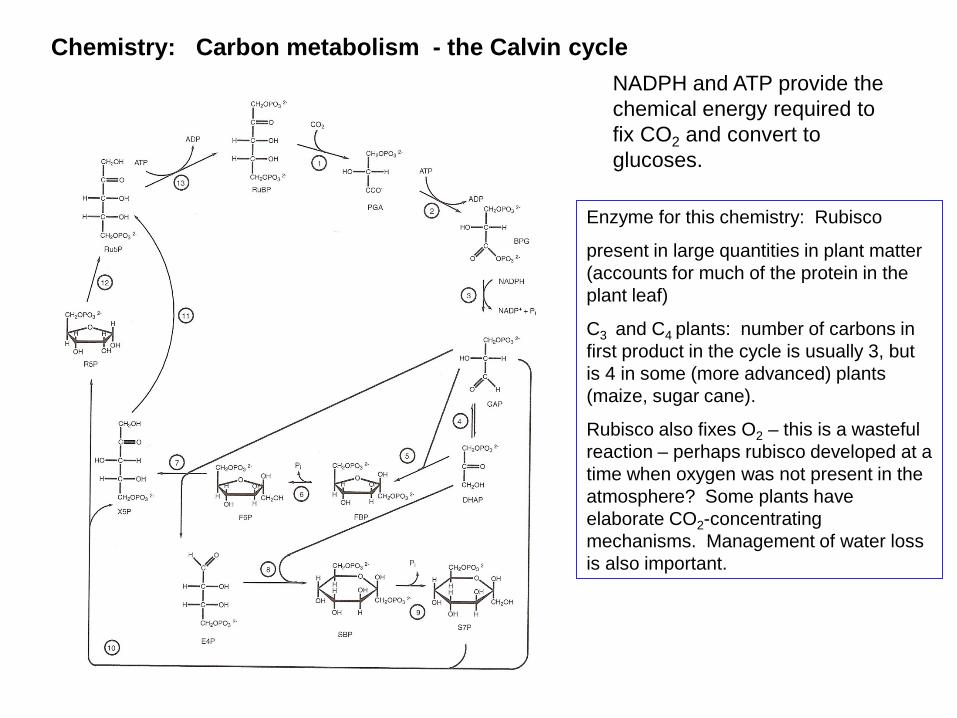
Chemistry: Carbon metabolism - the Calvin cycle
NADPH and ATP provide the chemical energy required to fix CO2 and convert to glucoses.
Enzyme for this chemistry: Rubisco
present in large quantities in plant matter (accounts for much of the protein in the plant leaf)
C3 and C4 plants: number of carbons in first product in the cycle is usually 3, but is 4 in some (more advanced) plants (maize, sugar cane).
Rubisco also fixes O2 – this is a wasteful reaction – perhaps rubisco developed at a time when oxygen was not present in the atmosphere? Some plants have elaborate CO2-concentrating mechanisms. Management of water loss is also important.


















