
L14. Sound Localization 2 - Cornell University...L14. Sound Localization 2 September 19, 2011...
9
9/18/2011 1 1 L14. Sound Localization 2 September 19, 2011 BioNB4240 C. D. Hopkins Linear Summation Delay-line Coincidence Detector delay + - dt h t f r ) ( ) ( ) ( Jeffress Model neural delay, ΔT coincidence detectors 3 Jeffres’ Model (1948) • Suppose Right & Left neural inputs from opposite sides converge.... • Delays, from neuronal conduction • Output cells (A-E) fire only if there is coincident inputs from both R and L • Delay-Line + coincidence detector produces a map of azimuth. Barn Owl 5 Barn Owls Roger Payne established that barn owls hunt in total darkness by using passive listening (Payne, 1971) * . Later work by Masakazu Konishi established the owl as an experimental model system in Neuroethology. * Payne, R. S. (1971) Acoustic location of prey by Barn owls (Tyto alba) 6 Owl can “localize” before leaving perch. Owl attack is precise within 2 degrees. Owl attacks sound source, not the mouse. •Conclusion: good localization in both vertical (elevation) and horizontal (azimuth) •How do they localize in the vertical?
Transcript of L14. Sound Localization 2 - Cornell University...L14. Sound Localization 2 September 19, 2011...

9/18/2011
1
1
L14. Sound Localization 2
September 19, 2011
BioNB4240 C. D. Hopkins
Linear Summation Delay-line Coincidence Detector
delay
+ -
dthtfr )()()(Jeffress Model
neural delay, ΔT
coincidence detectors
3
Jeffres’ Model (1948)
• Suppose Right & Left neural inputs from opposite sides converge....
• Delays, from neuronal conduction
• Output cells (A-E) fire only if there is coincident inputs from both R and L
• Delay-Line + coincidence detector produces a map of azimuth.
Barn Owl
5
Barn Owls
Roger Payne established that barn owls hunt in total darkness by using passive listening (Payne, 1971)*. Later work by Masakazu Konishi established the owl as an experimental model system in Neuroethology.
* Payne, R. S. (1971) Acoustic location of prey by
Barn owls (Tyto alba)
6
Owl can “localize” before leaving perch. Owl attack is precise within 2 degrees. Owl attacks sound source, not the mouse.
•Conclusion: good localization in both vertical (elevation) and horizontal (azimuth)
•How do they localize in the vertical?
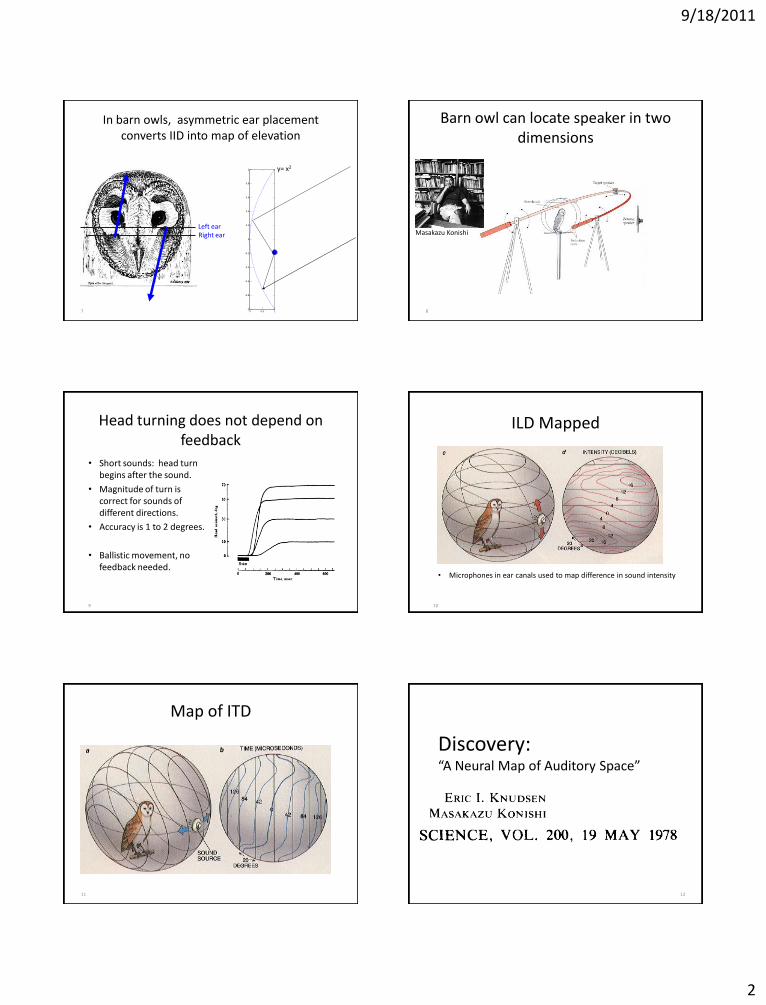
9/18/2011
2
7
In barn owls, asymmetric ear placement converts IID into map of elevation
Right ear Left ear
0 0.5 1-1
-0.8
-0.6
-0.4
-0.2
0
0.2
0.4
0.6
0.8
1 y= x2
8
Barn owl can locate speaker in two dimensions
Masakazu Konishi
9
Head turning does not depend on feedback
• Short sounds: head turn begins after the sound.
• Magnitude of turn is correct for sounds of different directions.
• Accuracy is 1 to 2 degrees.
• Ballistic movement, no feedback needed.
10
ILD Mapped
• Microphones in ear canals used to map difference in sound intensity
11
Map of ITD
12
Discovery: “A Neural Map of Auditory Space”
BioNB424
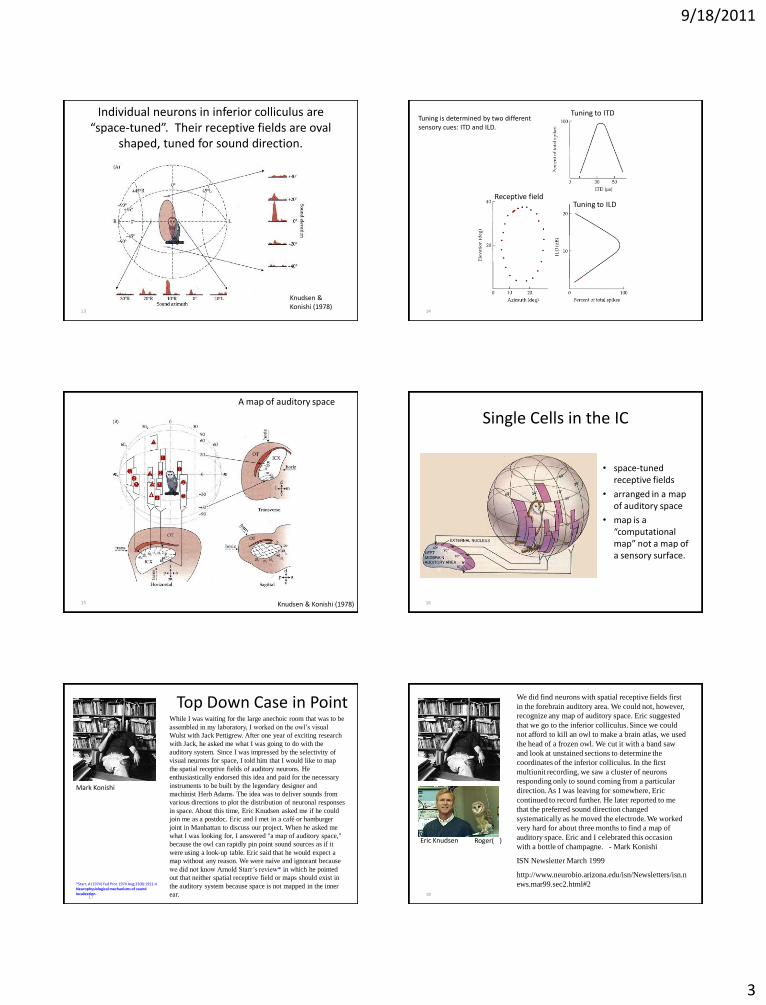
9/18/2011
3
13
Individual neurons in inferior colliculus are “space-tuned”. Their receptive fields are oval
shaped, tuned for sound direction.
Knudsen & Konishi (1978)
14
Receptive field
Tuning to ITD
Tuning to ILD
Tuning is determined by two different sensory cues: ITD and ILD.
15
A map of auditory space
Knudsen & Konishi (1978) 16
Single Cells in the IC
• space-tuned receptive fields
• arranged in a map of auditory space
• map is a “computational map” not a map of a sensory surface.
17
Top Down Case in Point While I was waiting for the large anechoic room that was to be
assembled in my laboratory, I worked on the owl’s visual
Wulst with Jack Pettigrew. After one year of exciting research
with Jack, he asked me what I was going to do with the
auditory system. Since I was impressed by the selectivity of
visual neurons for space, I told him that I would like to map
the spatial receptive fields of auditory neurons. He
enthusiastically endorsed this idea and paid for the necessary
instruments to be built by the legendary designer and
machinist Herb Adams. The idea was to deliver sounds from
various directions to plot the distribution of neuronal responses
in space. About this time, Eric Knudsen asked me if he could
join me as a postdoc. Eric and I met in a café or hamburger
joint in Manhattan to discuss our project. When he asked me
what I was looking for, I answered "a map of auditory space,"
because the owl can rapidly pin point sound sources as if it
were using a look-up table. Eric said that he would expect a
map without any reason. We were naive and ignorant because
we did not know Arnold Starr’s review* in which he pointed
out that neither spatial receptive field or maps should exist in
the auditory system because space is not mapped in the inner
ear.
Mark Konishi
*Starr, A (1974) Fed Proc 1974 Aug;33(8):1911-4 Neurophysiological mechanisms of sound localization. 18
We did find neurons with spatial receptive fields first
in the forebrain auditory area. We could not, however,
recognize any map of auditory space. Eric suggested
that we go to the inferior colliculus. Since we could
not afford to kill an owl to make a brain atlas, we used
the head of a frozen owl. We cut it with a band saw
and look at unstained sections to determine the
coordinates of the inferior colliculus. In the first
multiunit recording, we saw a cluster of neurons
responding only to sound coming from a particular
direction. As I was leaving for somewhere, Eric
continued to record further. He later reported to me
that the preferred sound direction changed
systematically as he moved the electrode. We worked
very hard for about three months to find a map of
auditory space. Eric and I celebrated this occasion
with a bottle of champagne. - Mark Konishi
ISN Newsletter March 1999
http://www.neurobio.arizona.edu/isn/Newsletters/isn.n
ews.mar99.sec2.html#2
Eric Knudsen Roger( )

9/18/2011
4
19
Behavioral Test of ILD
• Trained owls
• Right ear plugged: owl turns down
• Left ear plugged, turns up
20
Behavioral Test of ITD
• Owl presented with an artificial ITD from two earphones.
• Owl turns head after training to locate sounds at different azimuths
• Main cue: ongoing disparity of sound times (ITD)
21
ITD Test • Behavioral test, trained owl.
• ITD manipulated with stereo headphones.
• Owl turns toward time-advanced sound.
• 10 microsec sensitivity.
22
Hypothesis
• Vertical axis coded by IID
• Horizontal axis coded by ITD
23
Time pathway in
the owl (blue)
24
Cells in nucleus magnocellularis project to nucleus laminaris both ipsilaterally and contralerally. Ipsilateral axons enter dorsally; contralateral axons enter ventrally.
NL: azimuth maps onto depth in nucleus
NL analogous function to MSO in mammal
Carr, CE & Konishi, M 1988
1 mm
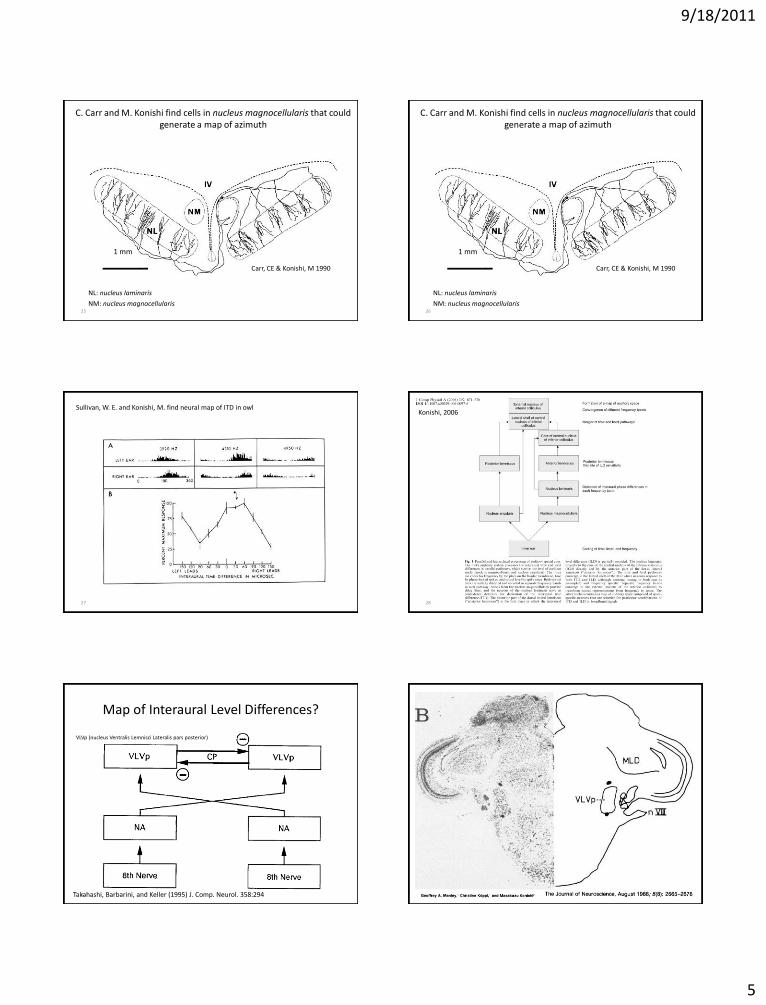
9/18/2011
5
25
C. Carr and M. Konishi find cells in nucleus magnocellularis that could generate a map of azimuth
NL: nucleus laminaris
NM: nucleus magnocellularis
Carr, CE & Konishi, M 1990
1 mm
26
C. Carr and M. Konishi find cells in nucleus magnocellularis that could generate a map of azimuth
NL: nucleus laminaris
NM: nucleus magnocellularis
Carr, CE & Konishi, M 1990
1 mm
27
Sullivan, W. E. and Konishi, M. find neural map of ITD in owl
28
Konishi, 2006
29
Map of Interaural Level Differences?
VLVp (nucleus Ventralis Lemnisci Lateralis pars posterior)
Takahashi, Barbarini, and Keller (1995) J. Comp. Neurol. 358:294 30
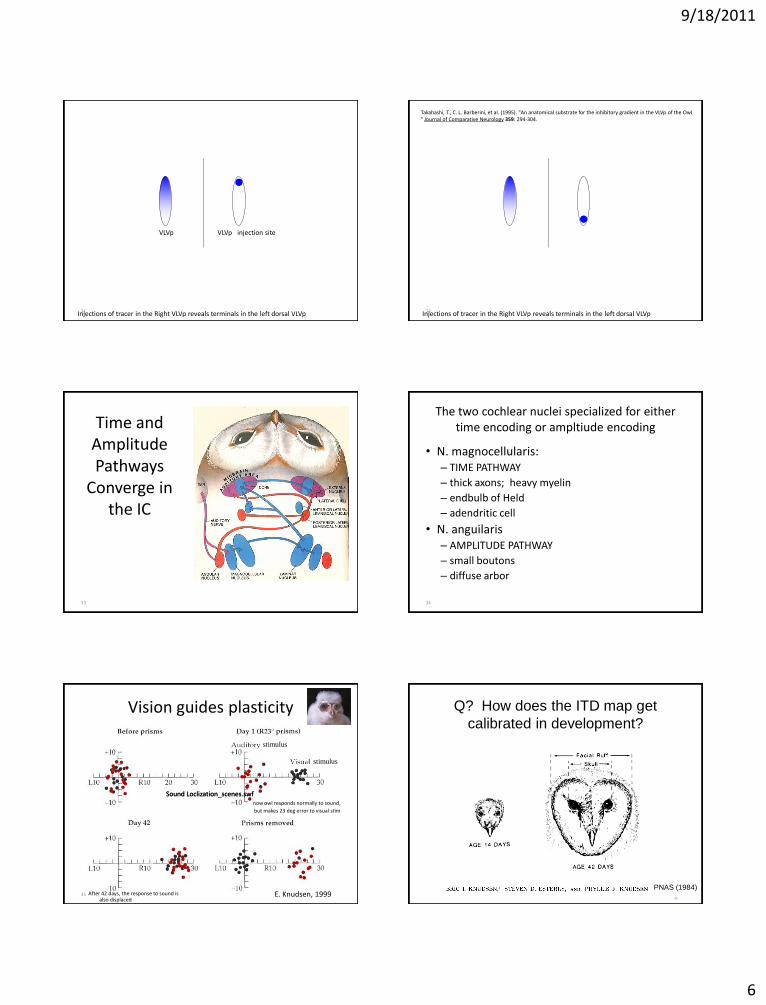
9/18/2011
6
31 Injections of tracer in the Right VLVp reveals terminals in the left dorsal VLVp
VLVp VLVp injection site
32 Injections of tracer in the Right VLVp reveals terminals in the left dorsal VLVp
Takahashi, T., C. L. Barberini, et al. (1995). "An anatomical substrate for the inhibitory gradient in the VLVp of the Owl " Journal of Comparative Neurology 359: 294-304.
33
Time and Amplitude Pathways
Converge in the IC
34
The two cochlear nuclei specialized for either time encoding or ampltiude encoding
• N. magnocellularis: – TIME PATHWAY
– thick axons; heavy myelin
– endbulb of Held
– adendritic cell
• N. anguilaris – AMPLITUDE PATHWAY
– small boutons
– diffuse arbor
35
Vision guides plasticity
stimulus
stimulus
E. Knudsen, 1999
now owl responds normally to sound,
but makes 23 deg error to visual stim
After 42 days, the response to sound is also displaced
Sound Loclization_scenes.swf Sound Loclization_scenes.swf
36
Q? How does the ITD map get
calibrated in development?
PNAS (1984)
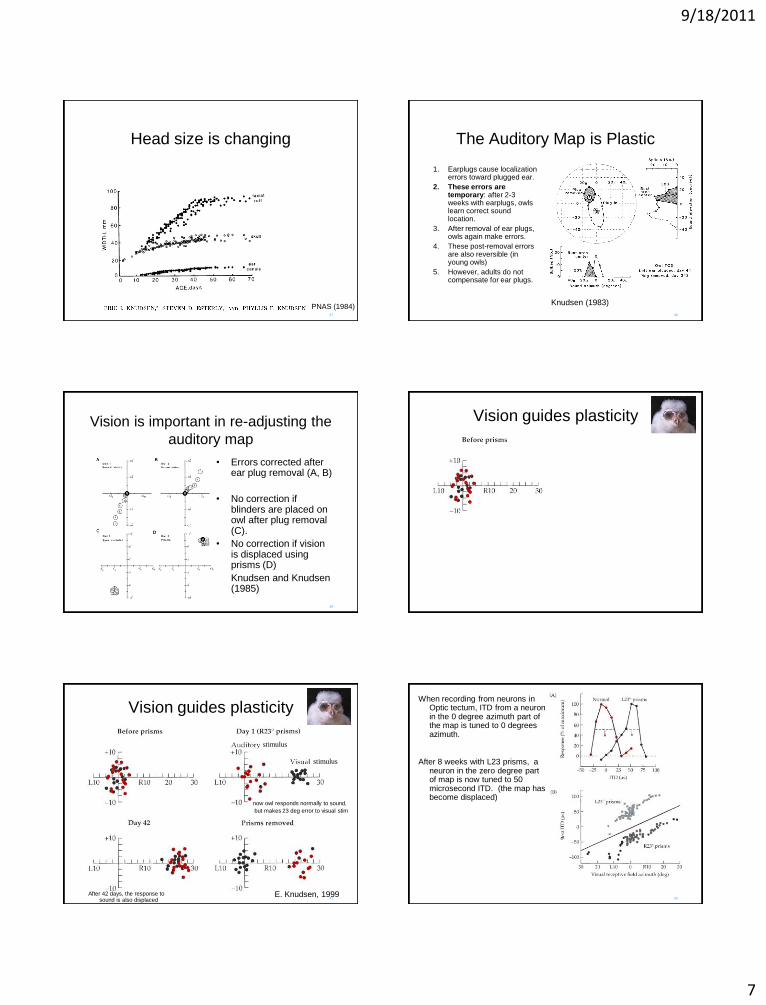
9/18/2011
7
37
Head size is changing
PNAS (1984) 38
The Auditory Map is Plastic
1. Earplugs cause localization errors toward plugged ear.
2. These errors are temporary: after 2-3 weeks with earplugs, owls learn correct sound location.
3. After removal of ear plugs, owls again make errors.
4. These post-removal errors are also reversible (in young owls)
5. However, adults do not compensate for ear plugs.
Knudsen (1983)
39
Vision is important in re-adjusting the
auditory map
• Errors corrected after ear plug removal (A, B)
• No correction if blinders are placed on owl after plug removal (C).
• No correction if vision is displaced using prisms (D)
Knudsen and Knudsen (1985)
40
Vision guides plasticity
stimulus
stimulus
E. Knudsen, 1999
now owl responds normally to sound,
but makes 23 deg error to visual stim
After 42 days, the response to sound is also displaced
41
Vision guides plasticity
stimulus
stimulus
E. Knudsen, 1999
now owl responds normally to sound,
but makes 23 deg error to visual stim
After 42 days, the response to sound is also displaced 42
When recording from neurons in Optic tectum, ITD from a neuron in the 0 degree azimuth part of the map is tuned to 0 degrees azimuth.
After 8 weeks with L23 prisms, a neuron in the zero degree part of map is now tuned to 50 microsecond ITD. (the map has become displaced)
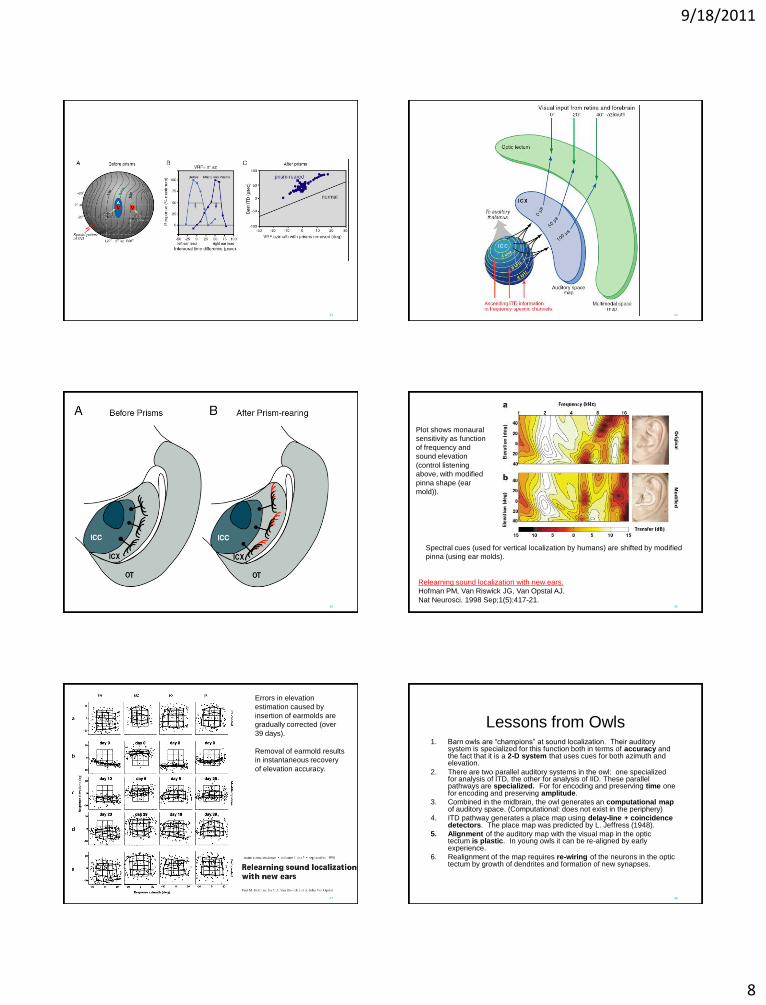
9/18/2011
8
43 44
45 46
Spectral cues (used for vertical localization by humans) are shifted by modified
pinna (using ear molds).
Relearning sound localization with new ears.
Hofman PM, Van Riswick JG, Van Opstal AJ.
Nat Neurosci. 1998 Sep;1(5):417-21.
Plot shows monaural
sensitivity as function
of frequency and
sound elevation
(control listening
above, with modified
pinna shape (ear
mold)).
47
Errors in elevation
estimation caused by
insertion of earmolds are
gradually corrected (over
39 days).
Removal of earmold results
in instantaneous recovery
of elevation accuracy.
48
Lessons from Owls 1. Barn owls are “champions” at sound localization. Their auditory
system is specialized for this function both in terms of accuracy and the fact that it is a 2-D system that uses cues for both azimuth and elevation.
2. There are two parallel auditory systems in the owl: one specialized for analysis of ITD, the other for analysis of IID. These parallel pathways are specialized. For for encoding and preserving time one for encoding and preserving amplitude.
3. Combined in the midbrain, the owl generates an computational map of auditory space. (Computational: does not exist in the periphery)
4. ITD pathway generates a place map using delay-line + coincidence detectors. The place map was predicted by L. Jeffress (1948).
5. Alignment of the auditory map with the visual map in the optic tectum is plastic. In young owls it can be re-aligned by early experience.
6. Realignment of the map requires re-wiring of the neurons in the optic tectum by growth of dendrites and formation of new synapses.

9/18/2011
9
Literature Carr, C. E., Fujita, I. and Konishi, M. (1989). Distribution of GABAergic neurons and terminals in the auditory system of
the barn owl. J Comp Neurol 286, 190-207.
Carr, C. E. and Konishi, M. (1988a). Axonal delay lines for time measurement in the owl's brainstem. Proc Natl Acad Sci
U S A 85, 8311-5.
Carr, C. E. and Konishi, M. (1988b). Axonal delay lines for time measurement in the owl's brainstem. Proc. Natl. Acad.
Sci. USA 85, 8311-8315.
Knudsen, E. I. (1975). Spatial aspects of electric fields generated by weakly electric fish. J. Comp. Physiol. 99, 193-118.
Knudsen, E. I. (1980). Sound localization in birds. In Compartive Studies of Hearing in Vertebrates, eds. A. N. Popper
and R. R. Fay), pp. 289-322. New York: Springer-Verlag.
Knudsen, E. I. (1982). Auditory and visual maps of space in the optic tectum of the owl. Journal of Neuroscience 2, 1177-
1194.
Knudsen, E. I., Knudsen, P. K. and Masino, T. (1993). Parallel pathways mediating both sound localization and gaze
control in the forebrain and midbrain of the barn owl. Journal of Neuroscience 13, 2837-2852.
Knudsen, E. I. and Konishi, M. (1978a). Center-surround organization of auditory receptive fields in the owl. Science 202,
778-80.
Knudsen, E. I. and Konishi, M. (1978b). A neural map of auditory space in the owl. Science 200, 795-7.
Knudsen, E. I. and Konishi, M. (1978c). Space and frequency are represented separately in auditory midbrain of the owl.
J Neurophysiol 41, 870-84.
Knudsen, E. I. and Konishi, M. (1980). Monaural occlusion shifts receptive-field locations of auditory midbrain units in the
owl. J Neurophysiol 44, 687-95.
Knudsen, E. I., Konishi, M. and Pettigrew, J. D. (1977). Receptive fields of auditory neurons in the owl. Science 198,
1278-80.
Konishi, M. (1977). Spatial localization of sound. In Recognition of Complex Acoustic Signals., (ed. T. H. Bullock). Berlin:
Dahlem Konferenzen.
Konishi, M. (1983). Neuroethology of acoustic prey localization in the barn owl. In Neuroethology and Behavioral
Physiology, eds. F. Huber and H. Markl), pp. 303-317. Berlin: Springer-Verlag.
Konishi, M. (1990). The neural algorithm for sound localization in the owl. Harvey Lect 86, 47-64.
Konishi, M. (1993a). Listening with two ears. Sci. American 268, 66-73.
Konishi, M. (1993b). Similar neural algorithms in owls and electric fish. Journal of Comparative Physiology A 173, 698-
700.
49
Konishi, M. (2003). Coding of auditory space. Annu. Rev. Neurosci. 26, 31-55.
Margoliash, D. and Konishi, M. (1985). Auditory representation of autogenous song in the song system of
white-crowned sparrows. Proc Natl Acad Sci U S A 82, 5997-6000.
Middlebrooks, J. C. and Knudsen, E. I. (1984). A neural code for auditory space in the cat's superior
colliculus. Journal of Neuroscience 4, 2621-2634.
Moiseff, A. and Konishi, M. (1981). Neuronal and behavioral sensitivity to binaural time differences in the
owl. J Neurosci 1, 40-8.
Moiseff, A. and Konishi, M. (1983). Binaural characteristics of units in the owl's brainstem auditory pathway:
precursors of restricted spatial receptive fields. J Neurosci 3, 2553-62.
Takahashi, T. and Konishi, M. (1986). Selectivity for interaural time difference in the owl's midbrain. J
Neurosci 6, 3413-22.
Takahashi, T., Moiseff, A. and Konishi, M. (1984). Time and intensity cues are processed independently in
the auditory system of the owl. J Neurosci 4, 1781-6.
Takahashi, T. T., Carr, C. E., Brecha, N. and Konishi, M. (1987). Calcium binding protein-like
immunoreactivity labels the terminal field of nucleus laminaris of the barn owl. J Neurosci 7, 1843-56.
Takahashi, T. T. and Konishi, M. (1988a). Projections of nucleus angularis and nucleus laminaris to the
lateral lemniscal nuclear complex of the barn owl. J Comp Neurol 274, 212-38.
Takahashi, T. T. and Konishi, M. (1988b). Projections of the cochlear nuclei and nucleus laminaris to the
inferior colliculus of the barn owl. J Comp Neurol 274, 190-211.
Volman, S. F. and Konishi, M. (1989). Spatial selectivity and binaural responses in the inferior colliculus of
the great horned owl. J Neurosci 9, 3083-96.
Volman, S. F. and Konishi, M. (1990). Comparative physiology of sound localization in four species of owls.
Brain Behav Evol 36, 196-215.
Wagner, H., Takahashi, T. and Konishi, M. (1987). Representation of interaural time difference in the central
nucleus of the barn owl's inferior colliculus. J Neurosci 7, 3105-16.
THE END


















