
Interação entre um péptido β-amilóide, responsável pela ... · Às minhas irmãs de...
100
Interação entre um péptido β-amilóide, responsável pela doença de Alzheimer, e modelos biomembranares Andreia Filipa Costa Cuco Dissertação para obtenção do Grau de Mestre em Bioengenharia e Nanossistemas Orientadoras: Doutora Benilde de Jesus Vieira Saramago e Doutora Ana Paula Valagão Amadeu do Serro Júri Presidente: Professor Doutor Luís Joaquim Pina da Fonseca Orientadora: Professora Doutora Benilde de Jesus Vieira Saramago Vogal: Professora Doutora Amélia Maria Pina Soares Gonçalves da Silva Novembro de 2014
Transcript of Interação entre um péptido β-amilóide, responsável pela ... · Às minhas irmãs de...

Interação entre um péptido β-amilóide, responsável pela
doença de Alzheimer, e modelos biomembranares
Andreia Filipa Costa Cuco
Dissertação para obtenção do Grau de Mestre em
Bioengenharia e Nanossistemas
Orientadoras: Doutora Benilde de Jesus Vieira Saramago e Doutora Ana Paula Valagão Amadeu do Serro
Júri
Presidente: Professor Doutor Luís Joaquim Pina da Fonseca
Orientadora: Professora Doutora Benilde de Jesus Vieira Saramago
Vogal: Professora Doutora Amélia Maria Pina Soares Gonçalves da Silva
Novembro de 2014

II

III
A mente que se abre a uma nova ideia, jamais volta ao seu tamanho inicial.
ALBERT EINSTEIN

IV

V
Agradecimentos
A elaboração desta dissertação só foi possível contando com a contribuição, apoio e cooperação de
diversas pessoas, às quais não poderia deixar de exprimir o meu mais sincero apreço e agradecimento.
Em primeiro lugar, quero agradecer à minha orientadora, Professora Doutora Benilde Vieira
Saramago, pelo incansável apoio e incentivo, pela disponibilidade e acompanhamento que sempre
demonstrou durante todo processo, por me acolher no seu grupo de trabalho.
À minha co-orientadora, Professora Doutora Ana Paula Serro, pelos conhecimentos transmitidos,
pela disponibilidade de auxílio e pela motivação transmitida ao longo de todo o trabalho.
À Professora Doutora Amélia Gonçalves da Silva, pela cedência do laboratório e equipamento, pela
enorme disponibilidade para apoiar e auxiliar nos longos dias de trabalho, pela ajuda na interpretação
dos resultados e pelo otimismo demonstrado.
À Professora Doutora Anabela Fernandes, por todo o apoio e disponibilidade, pela ajuda na
interpretação de resultados e pela cedência do laboratório e equipamento.
Ao Professor Doutor José Paulo Farinha pela cedência do laboratório e equipamento, pela
disponibilidade e ajuda na interpretação dos resultados.
Ao Professor Eduardo Melo pela disponibilidade, opiniões e troca de ideias.
Um enorme obrigado também ao José Restolho, Patrizia Paradiso, Sara Matias e Tânia Ribeiro pela
disponibilidade em auxiliar-me no trabalho dos diversos equipamentos e pelo apoio.
Mas, sem dúvida, que há muito a agradecer a outras pessoas que contribuíram para a elaboração
desta dissertação por outros motivos.
Um obrigado gigante aos meus colegas de laboratório: Ana Rita Valente, Andreia Pimenta, José
Restolho, Marina Esteves, Patrizia Paradiso e Vanessa Moreira, pelos incontáveis momentos de
gargalhadas, de descontração, pelo apoio quando começa tudo a parecer difícil.
Um obrigado particular a pessoas que foram mais do que colegas de mestrado, foram amigos: Ana
Rita Valente, Gonçalo Adriano e Mariana Pina, mesmo que os nossos caminhos não se voltem a cruzar,
levo-vos comigo, foram essenciais nestes dois anos no IST. Obrigado, por tudo!
Aos meus incansáveis amigos, um enorme obrigado. Em especial: Miguel Vidigal, pelo amigo que
é, pela preocupação, pela amizade incondicional, por mesmo longe estar sempre perto; ao “Jardel Luís”
(Luís Carlos), por mesmo do outro lado do mundo me transmitir um apoio incondicional; ao Henrique
Pernes, pela ajuda, pelo apoio, pelo incentivo e pela enorme paciência; ao Flávio Sá, por me tornar os
momentos mais aflitivos em momentos mais descontraídos.

VI
Às minhas irmãs de coração: Cláudia Azevedo Ferreira, Joana Pereira e Joana Rodrigues, pela
amizade incondicional, por nunca me deixarem ir a baixo, por acreditarem sempre em mim, pelas
gargalhadas, por cada momento nosso. “Os amigos nunca são para as ocasiões. São para sempre.”
Um obrigado especial à Joana Pereira, por seres a melhor amiga que podia ter, por seres a ‘irmã
de pais diferentes”, por saberes o que penso sem ser preciso falar, por mesmo longe nunca me deixares
sozinha: “Porque eu nunca tinha pedido uma irmã, mas a vida encarregou-se de te trazer”. És a certeza
do ‘para sempre’ e a família que eu escolhi.
À minha extraordinária família por todo o amor que sempre me transmitiram, por saberem que
sempre estarão lá para mim Por sempre acreditarem em mim.
À minha mãe, por ser um apoio inabalável no momento de aflição, por acreditar em mim, por nunca
me deixar desistir, por ser um porto de abrigo, por fazer tão bem o papel de mãe, por nunca me ter
deixado ir abaixo. Obrigado minha mãe!
Ao meu pai, para quem as palavras me parecem sempre escassas. Obrigado por todos estes anos
ter acreditado em mim e no meu sucesso, por me mostrar sempre que eu era capaz de ir mais além,
por me ter ensinado que com esforço e dedicação tudo se consegue. Obrigado por ser o melhor pai do
mundo, mas também por ser a melhor pessoa que conheço. “Não escolhi ser tua filha, apenas tive
sorte”. Obrigado pai, és o maior amor da minha vida! “Olha lá para cima, ela está a ver-nos.”.
E em conjunto, ao meu pai e à minha mãe por sempre me terem proporcionado o privilégio de viver
numa família e lar feliz. O que sinto por vocês é incondicional. Conseguimos, esta vitória é nossa!
E um agradecimento especial, a quem por infelicidade da vida não pode ouvir nem ler estas
palavras, mas a quem me deu amor incondicional em vida e, agora, me ilumina o caminho. À minha
estrela guia que me acompanha sempre, de quem as saudades são infinitas. À minha querida avó
Ricardina, a quem dedico esta dissertação.

VII

VIII
Resumo
A doença de Alzheimer caracteriza-se pela formação de placas no cérebro compostas por
agregados de péptido β-amilóide (Aβ). A neurotoxicidade de Aβ parece relacionar-se com a sua
interação com a membrana celular, que pode ser influenciada pela conformação do péptido ou pela
composição lipídica da membrana. As jangadas lipídicas são microdomínios ricos em esfingomielina
(SM) e colesterol (Chol) que têm sido associados a interações específicas com Aβ.
Usou-se uma mistura equimolar de 1-palmitoil-2-oleoil-sn-glicero-3-fosfocolina (POPC), SM e Chol
como um modelo de membrana com jangada lipídica para estudar a interação com o fragmento
peptídico Aβ25-35. Investigou-se o efeito do colesterol estudando a interação de Aβ25-35 com a mistura
POPC/SM (1:1) e o efeito do pH. As técnicas experimentais utilizadas foram: Análise de Localização
de Nanopartículas (NTA) e Dicroísmo Circular, para caracterização de Aβ25-35, e Microbalança de Cristal
de Quartzo com Dissipação (QCM-D), Calorimetria Diferencial de Varrimento (DSC) e Balança de
Langmuir, para caracterizar a interação lípido-péptido.
O Aβ25-35 a baixas concentrações adsorve à região polar da monocamada, afetando pouco a
organização dos lípidos. O aumento da concentração de Aβ25-35 conduz à formação de domínios ricos
em péptido, em ambas as misturas lipídicas, sendo mais forte na presença de colesterol e a pH=4
(agregados peptídicos de menor dimensão). Nos lipossomas suportados, a interação com Aβ25-35 é
favorecida em POPC/SM(1:1) e o aumento da concentração peptídica não afeta os lipossomas. Nos
lipossomas em suspensão, a baixas concentrações o péptido estabiliza os lipossomas enquanto o
aumento da concentração de Aβ25-35 conduz à sua destruição.
PALAVRAS-CHAVE: Jangada lipídica, Péptido β-amilóide, Lipossoma, Monocamada de
Langmuir, QCM-D, DSC

IX

X
Abstract
Alzheimer’s disease is characterized by plaque formation, in brain, composed by amyloid-β peptide
(Aβ) aggregates. The Aβ neurotoxicity seems to be related to its interaction with the cell membrane
which can be influenced by the peptide conformation or the lipid membrane composition. Lipid rafts are
microdomains rich in sphingomyelin (SM) and cholesterol (Chol), which have been associated with
specific interactions with Aβ peptide.
Used an equimolar mixture of 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphocoline (POPC), SM and
Chol as a membrane model for studying lipid raft interaction with Aβ25-35 peptide fragment, at different
concentrations. Investigated the effect of cholesterol studying the interaction between Aβ25-35 and
POPC/SM (1:1) mixture, and the pH effect by pH changes between 4 and 7. The experimental
techniques used were: Nanoparticle Tracking Analysis (NTA) and Circular Dichroism, to Aβ25-35
characterization, and Quartz Crystal Microbalance with Dissipation (QCM-D), Differential Scanning
Calorimetry (DSC) and Langmuir Monolayers, to characterize the lipid-peptide interaction.
At low Aβ25-35 peptide concentrations, the peptide is adsorbed in the polar region of the monolayers,
slightly affecting the organization of lipids. The increase of Aβ25-35 concentration leads to the formation
of peptide rich domains in both lipid mixtures, being stronger in the presence of Chol and at pH=4
(peptide aggregates smaller). In supported liposomes, the interaction with Aβ25-35 is favored POPC/SM
(1: 1) and increasing concentrations of peptide did not affect the liposomes. In suspended liposomes,
at low concentrations the peptide stabilizes the liposomes while increasing the Aβ25-35 concentration
leads to their destruction.
KEYWORDS: Lipid rafts, Amyloid β-Peptide, Liposome, Langmuir Monolayers, QCM-D, DSC

XI

XII
Índice
Lista de Quadros ................................................................................................................. XV
Lista de Figuras ................................................................................................................. XVII
Lista de Abreviaturas .......................................................................................................... XXI
Lista de Símbolos ............................................................................................................ XXIV
Capítulo I – Introdução .......................................................................................................... 1
1.1. Motivação ................................................................................................................ 1
1.2. Estado da Arte ........................................................................................................ 1
1.2.1. A doença de Alzheimer .................................................................................... 1
1.2.2. As membranas lipídicas e os péptidos β-amilóides .......................................... 2
1.3. Conteúdo ................................................................................................................ 4
Capítulo II – Fundamentos Teóricos ...................................................................................... 5
2.1. Membranas Celulares ............................................................................................. 5
2.2. Jangadas lipídicas ................................................................................................... 7
2.3. Modelos Biomembranares....................................................................................... 9
2.3.1. Modelos para jangadas lipídicas .....................................................................11
2.4. Péptido β-amilóide: fragmento (1-42) e fragmento (25-35) .....................................12
2.4.1. Agregação Peptídica .......................................................................................14
2.5. Interação entre modelos biomembranares e péptido Aβ ........................................14
2.6. Técnicas Experimentais .........................................................................................16
2.6.1. Microbalança de Cristal de Quartzo com Dissipação (QCM-D) .......................16
2.6.2. Balança de Langmuir ......................................................................................18
2.6.3. Análise de Localização de Nanopartículas ......................................................20
2.6.4. Calorimetria Diferencial de Varrimento (DSC) .................................................20
2.6.5. Dicroísmo Circular ...........................................................................................21

XIII
Capítulo III – Materiais e Métodos Experimentais .................................................................22
3.1. Materiais ................................................................................................................22
3.2. Preparação dos Lipossomas ..................................................................................23
3.3. Preparação de Soluções Peptídicas .......................................................................23
3.4. Preparação de Soluções Lipídicas .........................................................................25
3.5. QCM-D ...................................................................................................................25
3.6. Balança de Langmuir .............................................................................................26
3.7. NTA ........................................................................................................................28
3.8. DSC .......................................................................................................................28
3.9. Dicroísmo Circular ..................................................................................................29
Capítulo IV – Resultados e Discussão ..................................................................................30
4.1. Caracterização dos Modelos Lipídicos ...................................................................30
QCM-D ..........................................................................................................................30
Balança de Langmuir ....................................................................................................32
DSC ..............................................................................................................................37
4.2. Caracterização das soluções peptídicas ................................................................38
Balança de Langmuir ....................................................................................................38
NTA ...............................................................................................................................40
DSC ..............................................................................................................................42
Dicroísmo Circular .........................................................................................................39
4.3. Efeito do Péptido β-amilóide nas misturas lipídicas ................................................42
QCM-D ..........................................................................................................................42
Balança de Langmuir ....................................................................................................49
DSC ..............................................................................................................................55
Capítulo V – Conclusões e Trabalho Futuro .........................................................................57
Referências bibliográficas ....................................................................................................60

XIV

XV
Lista de Quadros
Quadro 1. Condições limite para estimar a viscosidade do filme, o módulo de distorção e a espessura no software QTools 3. ........................................................................................................................... 26 Quadro 2. Concentrações usadas para lípidos e misturas lipídicas nas isotérmicas 𝝅-A. ................... 27 Quadro 3. Valores de lift-off e do colapso das monocamadas dos lípidos puros (Colesterol, Esfingomielina e POPC), a pH=4 e pH=7 e 25ºC. ................................................................................ 36 Quadro 4. Valores médios e desvio padrão das propriedades viscoelásticas dos lipossomas POPC/SM (1:1) ao longo do processo de interação com 10 µM (cima) e 80 µM (baixo) de Aβ25-35 a 37ºC e pH=7, obtidos a partir da modelação por QTools 3. ........................................................................................ 48 Quadro 5. Valores médios e desvio padrão das propriedades viscoelásticas dos lipossomas POPC/SM/Chol (1:1:1) ao longo do processo de interação com 10 µM (cima) e 80 µM (baixo) de Aβ25-
35 a 37ºC e pH=7, obtidos a partir da modelação por QTools 3. ........................................................... 48

XVI

XVII
Lista de Figuras
Figura 1. Modelo de Mosaico Fluído para a membrana celular. Adaptado de [53]. ............................... 5
Figura 2. Estrutura química de um glicerofosfolípidos: fosfatidil colina (PC). Adaptado de [62]. ........... 6
Figura 3. Estrutura química da esfingomielina. Adaptado de [62]. ......................................................... 6
Figura 4. Estrutura química do colesterol. Adaptado de [62]. ................................................................. 7
Figura 5. Esquema representativo de jangadas lipídicas. As jangadas lipídicas podem ter diferentes
composições lipídicas e proteícas (jangada lipídica 1 e jangada lipídica 2). Adaptado de [81]. ............ 8
Figura 6. Representação esquemática da Bicamada Fosfolipídica Suportada, num substrato sólido.
Adaptado de [98]. .................................................................................................................................... 9
Figura 7. Representação esquemática de uma membrana lipídica negra. Adaptado de [98]. ............. 10
Figura 8. Representação esquemática de uma monocamada de Langmuir. Adaptado de [81]. .......... 10
Figura 9. Representação esquemática de SAMs, com um substrato de ouro. Adaptado de [98]. ....... 11
Figura 10. Representação esquemática de uma vesícula fosfolipídica (lipossoma). Adaptado de [107].
............................................................................................................................................................... 11
Figura 11. Diagrama de Fase do POPC, esfingomielina e colesterol a três importantes temperaturas
(23, 34 e 46ºC). Adaptado de [115]. ...................................................................................................... 12
Figura 12. Representação da sequência do péptido β-amilóide (1-42), (1-40) e (25-35), identificando os
terminais C e N. Adaptado de [129, 130]. ............................................................................................. 13
Figura 13. Exemplo de uma isotérmica 𝝅 –A de um fosfolípido, com os estados de organização
adotados pelas moléculas presentes na interface ar/líquido ao longo da compressão. Adaptado de
[178]. ...................................................................................................................................................... 19
Figura 14. Exemplo de um termograma típico de uma experiência DSC (DMPC Brain Lipid), onde se
mostra um pico correspondente a uma transição de fase. ................................................................... 21
Figura 15. Esquema das camadas dos cristais de quartzo a usar na QCM-D, com revestimento. 1) Ouro,
2) Crómico, 3) Quartzo. ......................................................................................................................... 25
Figura 16. Exemplo da variação da frequência (azul) e da dissipação (vermelho) da harmónica 3 para
uma experiência típica de adsorção de lipossomas no cristal de quartzo, a 37ºC e pH=7. Esquerda:
POPC/SM (1:1), 1.12 mM; Direita: POPC/SM/Chol (1:1:1), 1.12 mM. ................................................. 30
Figura 17. Valor médio e desvio padrão da variação da frequência (esquerda) e dissipação (direita) da
harmónica 3 para a mistura binária POPC/SM (1:1) e para a mistura ternária POPC/SM/Chol (1:1:1)
1.12 mM, a 37ºC e pH=7. ...................................................................................................................... 31
Figura 18. Espessura (esquerda) e viscosidade (direita) da bicamada lipídica de POPC/SM (1:1) e
POPC/SM/Chol (1:1:1), 1.12 mM, a 37ºC e pH=7. ............................................................................... 32
Figura 19. Isotérmicas -A a 25ºC, numa subfase aquosa, a pH=7. Lipidos Puros (curvas contínuas):
POPC (curva 1), esfingomielina (curva 2) e colesterol (curva 3). Misturas Lipídicas (curvas
descontínuas): POPC/SM (1:1) (curva 4) e POPC/SM/Chol (1:1:1) (curva 5). ..................................... 33

XVIII
Figura 20. Isotérmica -A experimental vs. Isotérmica -A teórica, a 25ºC e numa subfase aquosa a
pH=7. Lípidos Puros: POPC (curva 1) e esfingomielina (curva 2). Linha Descontínua: POPC/SM (1:1)
Teórica. .................................................................................................................................................. 34
Figura 21. Isotérmica -A experimental vs. Isotérmica -A teórica, a 25ºC e numa subfase aquosa a
pH=7. Lípidos Puros: POPC (curva 1), esfingomielina (curva 2) e colesterol (curva 3). Linha
Descontínua: POPC/SM/Chol (1:1:1) Teórica. ...................................................................................... 35
Figura 22. Efeito do pH nas isotérmicas -A dos lípidos puros, numa subfase aquosa: POPC (curva 1),
esfingomielina (curva 2) e colesterol (curva 3) (A.), e nas isotérmicas das misturas lipídicas POPC/SM
(1:1) (curva 4) e POPC/SM/Chol (1:1:1) (curva 5) (B.), a pH=4 (linha descontínua) e pH=7 (linha
contínua) e 25ºC. ................................................................................................................................... 36
Figura 23. Termograma dos lipossomas em suspensão da mistura binária POPC/SM (1:1) (cinzento
escuro) e da mistura ternária POPC/SM/Chol (1:1:1) (cinzento claro) 1.12 mM, em HEPES (pH=7). 37
Figura 24. Isotérmica -A do fragmento peptídico Aβ25-35, a 25ºC, numa subfase aquosa a pH=7, para
as concentrações de 1, 2 e 4 µM de péptido β-amilóide (25-35). ......................................................... 38
Figura 25. Espectro de dicroísmo circular do fragmento peptídico Aβ25-35 80 µM, a pH=4 (esquerda) e
pH=7 (direita). ........................................................................................................................................ 39
Figura 26. Efeito do pH no número de agregados de fragmento peptídico Aβ25-35 em solução a 1, 4 e 25
µM, a 22ºC. pH=4: Cinzento escuro; pH=7: Cinzento claro. ................................................................. 40
Figura 27. Efeito do pH na agregação peptídica a 1 µM (roxo), 4 µM (vermelho) e 25 µM (verde) de
fragmento peptídico Aβ25-35, a 22ºC. pH=4: Linha descontínua; pH=7: Linha contínua. ...................... 41
Figura 28. Termograma do péptido β-amilóide (25-35) 50µM, a pH=7. ............................................... 42
Figura 29. Exemplo de um ensaio de QCM-D da interação entre a mistura binária POPC/SM (1:1) com
80 µM de Aβ25-35, a 37ºC e pH=7. Esquerda: variação da frequência. Direita: variação da dissipação.
(1) Injeção de POPC/SM (1:1) 1.12 mM, durante 7 minutos; (2) Injeção de tampão de HEPES, para
enxaguamento, durante 10 minutos; (3) Injeção de Aβ25-35, durante 7 minutos; (4) Injeção de tampão de
HEPES, para enxaguamento, durante 10 minutos. .............................................................................. 43
Figura 30. Exemplo de um ensaio de QCM-D da interação entre a mistura ternária POPC/SM/Chol
(1:1:1) com 80µM de Aβ25-35, a 37ºC e pH=7. Esquerda: variação da frequência. Direita: variação da
dissipação. (1) Injeção de POPC/SM/Chol (1:1:1) 1.12 mM, durante 7 minutos; (2) Injeção de tampão
de HEPES, para enxaguamento, durante 10 minutos; (3) Injeção de Aβ25-35, durante 7 minutos; (4)
Injeção de tampão de HEPES, para enxaguamento, durante 10 minutos. .......................................... 43
Figura 31. Valores médios e desvio padrão da variação da frequência (esquerda) e da dissipação
(direita), em diferentes harmónicas, relativos à interação entre a mistura binária POPC/SM (1:1) 1.12
mM e 10 µM (azul), 25 µM (verde), 50 µM (laranja) e 80 µM (amarelo) de péptido Aβ25-35, a 37ºC e
pH=7. ..................................................................................................................................................... 44
Figura 32. Valores médios e desvio padrão da variação da frequência (esquerda) e da dissipação
(direita), em diferentes harmónicas, relativos à interação entre a mistura ternária POPC/SM/Chol (1:1:1)
1.12 mM e 10 µM (azul), 25 µM (verde), 50 µM (laranja) e 80 µM (amarelo) de péptido Aβ25-35, a 37ºC
e pH=7. .................................................................................................................................................. 45
Figura 33. Valores da variação da frequência (esquerda) e dissipação (direita) para a harmónica 3, a
37ºC e pH=7, após injeção de 1.12 mM de POPC/SM (1:1) (em cima) e POPC/SM/Chol (1:1:1) (em
baixo), enxaguamento com tampão de HEPES, injeção de 10, 25, 50 e 80 µM de Aβ25-35 e, novamente,

XIX
enxaguamento com tampão de HEPES. A intensidade do cinzento nas barras corresponde às injeções
indicadas nas caixas. ............................................................................................................................ 46
Figura 34. Iisotérmicas -A do efeito da concentração de Aβ25-35 em interação com POPC/SM (1:1), a
25ºC, numa subfase aquosa a pH=7. A seta vermelha significa o aumento da concentração do péptido
Aβ25-35 na subfase. ................................................................................................................................. 49
Figura 35. Exemplo de comportamento das isotérmicas -A do efeito da concentração de Aβ25-35 em
interação com POPC/SM/Chol (1:1:1), a 25ºC, numa subfase aquosa a pH=7. A seta vermelha significa
o aumento da concentração do péptido Aβ25-35 na subfase. ................................................................. 50
Figura 36. Isotérmica -A experimental vs. Isotérmica -A teórica, a 25ºC e numa subfase aquosa a
pH=7, da interação entre a monocamada lipídica POPC/SM/Chol (1:1:1) com 2 μM (A.) e 4 μM (B.) de
péptido Aβ25-35. ...................................................................................................................................... 51
Figura 37. Efeito do colesterol na formação da monocamada lipídica na presença de diferentes
concentrações de Aβ25-35, numa subfase aquosa a pH=7 e 25ºC, para pressões superficiais de 10 e 30
mN/m. .................................................................................................................................................... 52
Figura 38. Isotérmicas -A da mistura POPC/SM (1:1) (esquerda) e POPC/SM/Chol (1:1:1) (direita) em
interação com 1 e 4 µM de Aβ25-35, a 25ºC, numa subfase aquosa a pH=4 (linha descontínua) e pH=7
(linha contínua). ..................................................................................................................................... 54
Figura 39. Efeito do pH na variação da área da monocamada de POPC/SM (1:1) (esquerda) e
POPC/SM/Chol (1:1:1) (direita) em interação com 0.25, 0.5, 1, 2 e 4 µM de Aβ25-35, a 25ºC. Valores
médios para pressão superficial de 30mN/m. ....................................................................................... 54
Figura 40. Termogramas para POPC/SM (1:1) 1.12mM obtidos, a pH=7, com várias concentrações de
Aβ25-35. ................................................................................................................................................... 55
Figura 41. Temperatura de transição de fase da mistura POPC/SM (1:1) em função da concentração
de solução peptídica adicionada, a pH=7. ............................................................................................ 56

XX

XXI
Lista de Abreviaturas
Aβ péptido β-amilóide (amyloid-β peptide)
Aβ25-35 Fragmento peptídico β-amilóide (25-35)
Aβ1-40/42 Péptido β-amilóide (1-40) ou (1-42)
AFM Microscopia de força atómica (atomic force microscopy)
APP Proteína precursora amilóide
Chol Colesterol
DLPG 1,2-dilauroil-sn-glicero-3-fosfoglicerol
DMPA 1,2-ditetradecanoil-sn-glicero-3-fosfato
DMPC 1,2-dimiristoil-sn-glicero-3-fosfatidilcolina
DMPG 1,2-dimiristoil-sn-glicero-3-fosfoglicerol
DMPS 1,2-dimiristoil-sn-glicero-3-fosfatidilserina
DMSO Dimetilsulfóxido
DOPC 1,2-dioleoil-sn-glicero-3-fosfocolina
DPLC 1,2-dilauroil-sn-glicero-3-fosfatidilcolina
DPPC 1,2-dipalmitoil-sn-glicero-3-fosfocolina
DRM Membrana resistente a detergentes (detergent resistant membrane)
DSC Calorimetria diferencial de varrimento (differential scanning calorimetry)
FT-IR Espectroscopia de infravermelho por transformada de Fourier (Fourier transform infrared
spectroscopy)
GPI Glicofosfatidil inositol
GUV Vesículas unilamelares gigantes
HEPES ácido N-(2-hidroxietil)piperazina-N’-(2-etanossulfónico)
HFIP 1,1,1,3,3,3-Hexafluoro-2-propanol (hexafluorisopropanol)
LMV Vesículas multilamelares grandes

XXII
LUV Vesículas unilamelares grandes
PC Fosfatidil colina
PE Fosfatidil etanolamina
PG Fosfatidil glicerol
PI Fosfatidil inositol
PS Fosfatidil serina
POPC 1-palmitoil-2-oleoil-sn-glicero-3-fosfocolina
POPG 1-palmitoil-2-oleoil-sn-glicero-3-fosfoglicerol
PTFE Politetrafluoretileno
NMR Espectroscopia de ressonância magnética nuclear (nuclear magnetic ressonance
spectroscopy)
NTA Análise de localização de nanopartículas (nanoparticle tracking analysis)
QCM-D Microbalança de cristal de quartzo com dissipação (quartz crystal microbalance with
dissipation)
SAM Monocamada auto-montada
SDS Docedil sulfato de sódio
SLB Bicamada lipídica suportada
SM Esfingomielina
TFA Trifluoroacético
TFE Trifluoroetanol

XXIII

XXIV
Lista de Símbolos
Tt Temperatura de Transição de Fase
Lβ Fase líquida lamelar
Lα Fase líquida cristalina
LO Fase lamelar líquida-ordenada
Ld Fase lamelar líquida-desordenada
Lb Fase sólida ordenada
f Frequência de ressonância
D Dissipação de energia
Δf Variação da frequência de ressonância
ΔD Variação da dissipação de energia
Edis Energia dissipada
Est Energia armazenada
Δm Massa adsorvida
Cf Constante de sensibilidade característica do sensor
n Número da harmónica
ω Frequência angular
ρI Densidade do líquido
ɳI Viscosidade do líquido
ρf Densidade do filme
µf Elasticidade do filme
ɳf Viscosidade do filme
A0 Lift-off da monocamada
𝜋 Pressão superficial
ɣo Tensão superficial da água pura (ou seja, da subfase sem monocamada)

XXV
ɣ Tensão superficial da superfície ocupada pela monocamada
Cp Capacidade calorífica
N.A. Não avaliável

XXVI

1
Capítulo I – Introdução
1.1. Motivação
A doença de Alzheimer afeta cada vez mais a população mundial e é caracterizada pela formação
de placas no cérebro compostas por agregados de péptidos β-amilóides (Aβ). Alguns autores defendem
que a agregação destes péptidos é uma das causas para o desenvolvimento desta doença
neurodegenerativa. A neurotoxicidade dos péptidos Aβ na doença de Alzheimer relaciona-se com a
interação destes péptidos com os lípidos que constituem a membrana celular, afetando a sua
integridade e conduzindo a uma possível apoptose celular [1-3].
Existem microdomínios membranares que podem desempenhar um papel crucial no fenómeno de
agregação peptídica uma vez que estão associados a doenças neurodegenerativas. Estes
microdomínios são designados por jangadas lipídicas. A composição mais comum para um modelo de
jangadas lipídicas é uma mistura equimolar de 1-palmitoil-2-oleoil-sn-glicero-3-fosfocolina (POPC),
esfingomielina (SM) e colesterol (Chol) [4-6].
O principal objetivo desta investigação é compreender os fenómenos de interação existentes entre
os lípidos presentes na membrana celular e os péptidos Aβ, bem como entender o papel desta interação
na doença de Alzheimer. Deste modo, procurou-se estudar se a interação entre os péptidos Aβ e os
lípidos presentes na membrana celular ocorria preferencialmente em domínios ricos em colesterol
(jangadas lipídicas), em comparação com membranas mais fluídas sem a presença de colesterol. O
fragmento peptídico β-amilóide (25-35) (Aβ25-35) foi utilizado para estudar esta interação membrana
celular-péptido.
As técnicas experimentais utilizadas foram: a Microbalança de Cristal de Quartzo com Dissipação
(QCM-D), para deteção de mudanças na massa e nas propriedades viscoelásticas dos lipossomas
suportados na presença e ausência de colesterol, e a influência da concentração de péptido Aβ;
Balança de Langmuir, para estudar o efeito do colesterol e do pH na interação lípido-péptido; Análise
de Localização de Nanopartículas (NTA), para estudar o efeito da concentração de péptido Aβ na
agregação peptídica, e a influência do pH nesta agregação; Calorimetria Diferencial de Varrimento
(DSC), para detetar mudanças na transição de fase dos lípidos em interação com o péptido Aβ; e,
finalmente, Dicroísmo Circular, para estudar a conformação do péptido Aβ em solução, bem como a
influência do pH na sua conformação.
1.2. Estado da Arte
1.2.1. A doença de Alzheimer
A doença de Alzheimer é uma das doenças neurodegenerativas mais comuns, sendo caracterizada
por uma perda neuronal. A doença de Alzheimer consiste num distúrbio cerebral progressivo e fatal,
que conduz à atrofia do hipocampo e do córtex cerebral [7-9].
Uma das características desta doença neurodegenerativa é a formação de placas no cérebro
compostas principalmente por agregados de péptidos β-amilóides, exibindo uma estrutura de folha β;

2
a agregação peptídica é considerada como uma das causas para o desenvolvimento desta doença.
Considera-se, assim, que a inibição deste processo de agregação é uma potencial abordagem
terapêutica para o tratamento da doença de Alzheimer [1, 10]. As placas senis são compostas por uma
mistura de péptidos β-amilóides (1-40) (Aβ1-40) ou de péptidos β-amilóides (1-42) (Aβ1-42); contudo, o
fragmento peptídico Aβ1-42 predomina na doença de Alzheimer, apresentando este fragmento uma
maior tendência de agregação, comparativamente com o fragmento Aβ1-40 [11,12].
1.2.2. As membranas lipídicas e os péptidos β-amilóides
Diversos estudos indicam a existência de uma relação direta entre a composição e estrutura da
membrana celular e a tendência de agregação e neurotoxicidade dos péptidos β-amilóides [13-19].
Segundo alguns autores, a toxicidade (ou neurotoxicidade) dos péptidos β-amilóides, possivelmente
responsáveis pela doença de Alzheimer, pode estar relacionada com a interação destes péptidos com
a membrana celular, sendo importante compreender este fenómeno de interação [20, 21]. Segundo
Peters et al., a membrana celular é o primeiro local de interação entre o péptido β-amilóide e os
neurónios, sendo considerada o local onde a atividade neurotóxica destes péptidos se inicia [22]. Deste
modo, diversos são os mecanismos que têm sido apresentados como sendo possíveis causas para a
neurotoxicidade da interação do péptido β-amilóide com a membrana lipídica [23].
Vários autores afirmam que a neurotoxicidade da interação existente entre péptido β-amilóide e a
membrana lipídica pode estar associada a um aumento da permeabilidade da bicamada fosfolipídica,
à desregulação da homeostasia do cálcio e, ainda, à formação de canais iónicos devido aos agregados
de péptido Aβ. Kawahara et al., usando uma cultura de células neuronais, concluíram que as moléculas
de péptido Aβ1-40, em solução, podem interagir com a membrana celular, formando canais iónicos
seletivos [24]. A presença destes péptidos na membrana lipídica pode ainda conduzir à oxidação dos
lípidos presentes na membrana, conduzindo diretamente à perda da sua integridade. Além disso, vários
autores defendem que as ligações eletrostáticas dos péptidos Aβ ao grupo polar dos fosfolípidos pode
contribuir para o seu comportamento neurotóxico [19-21, 25-28].
Segundo diversos autores, a integridade da membrana lipídica é influenciada pela interação
existente entre a membrana lipídica e o péptido β-amilóide, conduzindo à diminuição da estabilidade e
aumento da fluidez da membrana celular e, eventualmente, à apoptose de células neuronais. Além
disso, estas interações membrana celular-péptido Aβ podem ser influenciadas pela conformação do
péptido, pelas propriedades físico-químicas da membrana celular e, ainda, pela sua composição lipídica
[29-31]. Estudos de modelação molecular sugeriram que o péptido β-amilóide in vivo é capaz de dar
origem a oligómeros simples ou a agregados de oligómeros. Aliados a estudos cristalográficos, estes
estudos de modelação molecular sugeriram que existe uma potencial interação entre os oligómeros e
a membrana celular [32]. D’Errico et al. e Dante et al., por sua vez, propuseram que a forma monomérica
do fragmento peptídico Aβ25-35 provoca menos alterações em membranas de 1-palmitoil-2-oleoil-sn-
glicero-3-fosfocolina/1-palmitoil-2-oleoil-sn-glicero-3-fosfoglicerol (POPC/POPG) e 1,2-dilauroil-sn-
glicero-3-fosfatidilcolina/1,2-dilauroil-sn-glicero-3-fosfoglicerol (DPLC/DLPG), contendo colesterol.
D’Errico et al. afirmaram, ainda, que a presença de DLPG na membrana lipídica não afeta a interação

3
desta com o péptido, concluindo que a inserção de péptidos na membrana lipídica é uma consequência
de interações hidrofóbicas [33, 34]. Outros autores concluíram que o péptido β-amilóide interatua
diretamente com os lípidos presentes na superfície da membrana lipídica, alterando as suas
propriedades mecânicas, as propriedades de coesão dos componentes membranares e a
permeabilidade da membrana lipídica, podendo este facto ser uma consequência da formação de
estruturas moleculares péptido-lípido na membrana celular [21, 35].
Como foi referido anteriormente, a presença de péptidos β-amilóides na membrana lipídica pode
conduzir à oxidação dos lípidos presentes na membrana. De facto, diversos autores estudaram, in vivo,
o papel do stress oxidativo na influência que os péptidos β-amilóides têm em doenças
neurodegenerativas, como a doença de Alzheimer. Segundo Betterfield et al. e Perluigi et al., os
péptidos Aβ1-42 estão diretamente associados à geração de radicais livres, que conduzem a danos
oxidativos dos lípidos presentes na membrana celular [36, 37]. Butterfield et al. demonstraram ainda
que o stress oxidativo não ocorre no cérebro in vivo, se o único resíduo de metionina na posição 35 do
péptido Aβ1-42 não estiver presente [38]. Veradarajan et al., por sua vez, demonstraram que o stress
oxidativo associado ao péptido Aβ1-42 em células neuronais é um processo que ocorre por via catalítica,
devido à reação em cadeia que é produzida na membrana lipídica [39].
A quantidade de péptido β-amilóide em interação com a membrana lipídica é também um fator a ter
em atenção. Quando a quantidade de péptido Aβ aumenta na membrana lipídica, estes péptidos
adotam a conformação de folha β, em vez de hélice α, conduzindo à geração de mais agregados
amilóides tóxicos no cérebro [40, 41]. De facto, Chakravarthy et al, usando tecidos do hipocampo e do
córtex frontal de cérebro humano pos-mortem, verificaram que um cérebro de um paciente com doença
de Alzheimer apresenta consideravelmente mais agregados de péptido Aβ1-42, que um cérebro de um
paciente sem esta doença. Estes autores utilizaram anticorpos monoclonais específicos para péptidos
Aβ para detetarem estes agregados [42].
De modo a estudar o fenómeno de interação existente entre os péptidos β-amilóides e a membrana
lipídica, diversos autores recorreram à utilização do fragmento Aβ25-35, uma vez que se trata de um
fragmento que apresenta um comportamento muito idêntico ao fragmento peptídico completo Aβ1-42 e
que, além disso, tem a capacidade de se movimentar na bicamada fosfolipídica, afetando fortemente a
sua dinâmica, principalmente na região apolar da membrana [31, 43-46]. Segundo Wood and Müller, o
fragmento peptídico Aβ25-35 representa a região biologicamente ativa do fragmento completo e agrega
mais facilmente do que outros fragmentos, mantendo o nível de toxicidade igual ao do péptido Aβ1-42.
Os efeitos do fragmento peptídico Aβ25-35 têm sido estudados por vários autores, usando modelos
biomembranares, como monocamadas ou bicamadas fosfolipídicas (exemplo: lipossomas) [47]. Por
sua vez, Mizkabekov et al. e Qi et al., demonstraram a capacidade do fragmento Aβ25-35 promover a
formação de canais iónicos em bicamadas fosfolipídicas planares e em membranas celulares,
respetivamente [48, 49]. Clementi et al, demonstrou ainda que o fragmento Aβ25-35, na forma não
agregada, é capaz de invadir as células neuronais do cérebro de um rato, conduzindo à sua morte [8].
Atualmente, a relação existente entre as características e mecanismos de ação dos péptidos β-
amilóides e a composição das bicamadas fosfolipídicas com as quais interatuam ainda não está

4
perfeitamente compreendida. Uma das razões é o facto de os resultados obtidos no estudo da interação
entre o fragmento peptídico Aβ25-35 e modelos biomembranares serem, por vezes, contraditórios.
Também o comportamento anfifílico destes péptidos tem sido considerado um importante fator na
interação péptido-lípido, existindo estudos que afirmam que as interações hidrofóbicas e eletrostáticas
são determinantes para os péptidos conduzirem a uma atividade disruptiva da membrana lipídica.
Torna-se, assim, necessário o desenvolvimento de mais estudos para compreender o verdadeiro papel
do fragmento peptídico β-amilóide (25-35) na doença de Alzheimer [8, 47].
1.3. Conteúdo
O presente trabalho é dividido em 4 capítulos principais. O primeiro capítulo, Fundamentos Teóricos,
tem como objetivo fornecer uma introdução em relação a vários tópicos: Membranas Celulares,
focando-se nos seus principais constituintes lipídicos; Jangadas Lipídicas, e o seu papel nas células;
Modelos Biomembranares, onde se apresenta os vários modelos para estudar as biomembranas e
modelos para estudar jangadas lipídicas; Péptido β-amilóide: fragmento (1-42) e fragmento (25-35), as
suas propriedades, funções, conformações e tendência de agregação peptídica; Interação entre
modelos biomembranares e péptidos Aβ, apresentando-se conclusões de diversos autores sobre esta
interação lípido-péptido; Técnicas Experimentais, onde são explicados os princípios de cada técnica
experimental utilizada na presente investigação.
O segundo capítulo, Materiais e Métodos, inclui os materiais e a descrição das técnicas
experimentais utilizadas para estudar as interações entre as membranas modelo e os péptidos Aβ, isto
é, Microbalança de Cristal de Quartzo com Dissipação (QCM-D), a Balança de Langmuir e a
Calorimetria Diferencial de Varrimento (DSC), bem como as técnicas de preparação das soluções
peptídicas, das soluções lipídicas e dos lipossomas, e, ainda, as técnicas utilizadas para estudar a
influência do pH na conformação do péptido Aβ e o efeito da concentração e pH na agregação
peptídica, isto é, Dicroísmo Circular e Análise de Localização de Nanopartículas (NTA), respetivamente.
No capítulo Resultados e Discussão apresenta-se os resultados da interação entre o fragmento
peptídico Aβ25-35 com os dois modelos biomembranares: lipossomas (suportados ou em suspensão) e
monocamadas de Langmuir de POPC/SM (1:1) e POPC/SM/Chol (1:1:1). Discutiu-se o efeito da
concentração do péptido Aβ nesta interação, bem como o efeito do colesterol. Além disso, discutiu-se
o efeito do pH na agregação peptídica e na interação péptido Aβ-modelos biomembranares, na
presença e ausência de colesterol, bem como o efeito do pH na conformação do fragmento peptídico
Aβ25-35 em solução.
O último capítulo, Conclusões e Trabalho Futuro, é o resumo das conclusões e suposições feitas de
acordo com os resultados obtidos através das técnicas experimentais utilizadas no presente estudo.
Neste capítulo sugere-se ainda possíveis direções para pesquisas futuras neste tema.

5
Capítulo II – Fundamentos Teóricos
2.1. Membranas Celulares
A zona primária de contacto entre uma dada célula e o seu ambiente envolvente é designada por
membrana celular. A membrana celular, também designada por membrana plasmática, membrana
citoplasmática ou membrana lipídica, define a estrutura externa de cada célula, separando o citoplasma
do ambiente envolvente, e funciona como uma barreira, regulando a saída e a entrada de material e
informação. As células eucariotas possuem ainda membranas subcelulares que dividem o citoplasma
em múltiplos compartimentos, designados por organelos, permitindo que cada função celular ocorra
eficientemente e simultaneamente em diferentes partes da célula. A conservação de energia da célula
e a comunicação célula-célula são outras duas propriedades da membrana celular; as funções
desempenhadas por esta membrana devem-se, essencialmente, às propriedades físicas que possui.
A membrana celular é flexível, o que permite a alteração da sua forma durante o crescimento e
movimento celular, e seletivamente permeável, armazenando compostos e iões importantes em
concentrações adequadas para a homeostasia celular; além disso, a membrana celular é uma estrutura
frágil e que se fecha sobre si mesma, permitindo que ocorram fusões com vesículas de exocitose e a
formação de vesículas de endocitose, sem perdas significativas de componentes celulares [50-52].
Figura 1. Modelo de Mosaico Fluído para a membrana celular. Adaptado de [53].
Atualmente, a estrutura da membrana celular é descrita pelo Modelo de Mosaico Fluído (Figura 1),
proposta por Singer e Nicholson, em 1972. De acordo com este modelo, a membrana celular consiste
numa bicamada fosfolipídica fluída, com proteínas a flutuar livremente. Segundo o Modelo de Mosaico
Fluído, a membrana celular é uma estrutura dinâmica e fluída, permitindo movimento; é
maioritariamente composta por fosfolípidos, formando a bicamada, com uma espessura de 5 a 8 nm,
onde se encontram diferentes moléculas distribuídas. As moléculas fosfolipídicas, que compõem a
membrana de uma célula eucariota, são moléculas anfipáticas, isto é, possuem uma terminação polar
e uma terminação apolar; enquanto a terminação polar é hidrofílica, preferindo o meio aquoso, a
terminação apolar é hidrofóbica, evitando-o. Assim, a bicamada fosfolipídica é uma estrutura estável
em meio aquoso, devido às interações hidrofílicas com este meio e, em especial, às interações
hidrofóbicas entre as cadeias alquílicas presentes no interior da bicamada [51, 54-56].

6
Cada tipo de membrana celular possui um grupo característico de proteínas, o que lhe confere
funções específicas. Na membrana celular, as proteínas mais comuns são as proteínas
transportadoras, que formam canais através da membrana celular, permitindo a passagem de solutos
orgânicos e de iões [55, 57]. Além destas, na membrana celular é possível encontrar proteínas
transmembranares e proteínas periféricas. As proteínas transmembranares ou integrais atravessam a
membrana de um lado ao outro, estando fortemente ligadas à membrana celular através de interações
hidrofóbicas. As proteínas periféricas, por seu lado, não atravessam a membrana, encontrando-se
numa das superfícies da membrana celular e são estabilizadas por interações electroestáticas e por
ligações de hidrogénio com o grupo polar dos fosfolípidos [58, 59].
Relativamente aos lípidos presentes na membrana celular, as principais classes lipídicas são:
glicerofosfolípidos e esfingolípidos [51]. Os glicerofosfolípidos (Figura 2) são compostos derivados do
ácido fosfatídico e podem ser designados, conforme o álcool presente na parte polar, por: fosfatidil
colina (PC), fosfatidil glicerol (PG), fosfatidil etanolamina (PE), fosfatidil inositol (PI) e fosfatidil serina
(PS) [51, 60, 61]
Figura 2. Estrutura química de um glicerofosfolípidos: fosfatidil colina (PC). Adaptado de [62].
Os esfingolípidos são estruturalmente semelhantes aos glicerofosfolípidos, mas, em vez de glicerol,
possuem esfingosina que é um aminoácido de cadeia longa. São uma família de lípidos anfipáticos que
derivam de ceramida; diferem na composição do grupo polar e são muito relevantes nas células
nervosas. A esfingomielina (Figura 3) é considerada um fosfolípido, uma vez que possui o grupo fosfato
[63].
Figura 3. Estrutura química da esfingomielina. Adaptado de [62].
Outro lípido presente nos tecidos biológicos é o colesterol (Figura 4), que pertence à família dos
esteróides. O colesterol é um dos constituintes da membrana celular das células eucariotas e é um
lípido anfipático, com um grupo hidroxilo polar. Este constituinte da membrana celular regula o estado
físico da bicamada fosfolipídica, aumenta a espessura da membrana celular e está envolvido,
diretamente, na formação de microdomínios na membrana, designados por jangadas lipídicas. O
colesterol não se encontra distribuído aleatoriamente na membrana celular e interatua com os
fosfolípidos. Quando estas interações ocorrem abaixo da temperatura de transição de fase (Tt),
correspondente à transição da fase de gel para a fase líquida cristalina, as cadeias de hidrocarbonetos
tornam-se mais fluídas, isto é, ocorre uma fluidificação da membrana lipídica; contrariamente, quando
estas interações ocorrem acima da Tt, as cadeias de hidrocarbonetos tornam-se mais rígidas, isto é,

7
ocorre um efeito condensante na membrana lipídica. O colesterol é vulnerável à oxidação, conduzindo
à formação de diversos produtos de oxidação, designados por COPs (Produtos da Oxidação do
Colesterol) [64-68].
Figura 4. Estrutura química do colesterol. Adaptado de [62].
A proporção relativa das três classes lipídicas presentes na membrana celular varia de acordo com
a espécie e o tipo de célula; tipicamente, os esfingolípidos representam 10 a 20% mol, o colesterol
representa cerca de 30 a 40% mol e os glicerofosfolípidos 40 a 60% mol do total dos lípidos presentes
na membrana celular [62].
A membrana lipídica apresenta uma distribuição assimétrica entre as duas monocamadas que a
compõem; a composição lipídica de cada monocamada é diferente. Tipicamente, a PC e a
esfingomielina encontram-se no lado exterior da membrana, enquanto PS, PG e PI se situam no interior
da membrana celular [62, 68, 69].
As interações entre as cadeias de acilo são fracas, sendo este o princípio básico da dinâmica da
membrana celular, permitindo o movimento dos lípidos e a formação de estruturas transitórias. Existem
diferentes formas de ordenar as cadeias de acilo dos lípidos, sendo estas responsáveis pelas
diferenças existentes na fluidez, estrutura e flexibilidade da membrana celular. O comportamento de
fase depende diretamente da temperatura e do tipo de lípido presente. Na fase gel lamelar (Lβ), as
cadeias de acilo encontram-se completamente estendidas e ordenadas e a espessura da bicamada
fosfolipídica é máxima; esta fase só ocorre em sistemas modelo. Na fase líquida cristalina (Lα), as
cadeias de acilo apresentam uma conformação desordenada e elevadas taxas de movimento intra e
intermolecular. A fase líquida ordenada é considerada uma fase intermédia entre as fases Lβ e Lα; esta
fase apresenta uma difusão rotacional livre das moléculas lipídicas, tal como acontece na fase Lα, e
cadeias de acilo completamente estendidas em configuração trans, como na fase Lβ [68, 62, 69, 70].
2.2. Jangadas lipídicas
A formação e a deteção de jangadas lipídicas em biomembranas têm sido alvo de atenção devido
ao possível papel que estas estruturas desempenham em diversos eventos celulares [71]. As jangadas
lipídicas são microdomínios da membrana celular, ricos em esfingomielina e colesterol, que possuem,
normalmente, um diâmetro inferior a 100 nm [72, 73]. Estes microdomínios membranares são um lugar
preferencial para interações lípido-lípido e interações lípido-péptido, criando uma plataforma de
sinalização que está envolvida em diversas funções neuronais, sendo as jangadas lipídicas
fundamentais para o desenvolvimento e funcionamento neuronal [71, 74, 75].

8
As principais funções das jangadas lipídicas estão associadas à mediação do transporte
membranar, transdução do sinal e comunicação com o citoesqueleto. As jangadas lipídicas
proporcionam ainda locais específicos para interações com toxinas, para a adesão celular, para
respostas imunitárias ou para a regulação do citoesqueleto e são, igualmente, vias importantes para o
processo de proliferação e apoptose [69, 71, 76, 77]. Alterações na estrutura e nas atividades das
jangadas lipídicas podem estar associadas a doenças neurodegenerativas, como a doença de
Alzheimer, uma vez que estes microdomínios membranares podem desempenhar um papel importante
na agregação peptídica [71, 76].
As proteínas periféricas ancoradas por glicofosfatidil inositol (GPI) e as proteínas sinalizadoras para
transdução do sinal através da membrana celular são frequentemente associadas às jangadas
lipídicas. Além destas, existem ainda proteínas transmembranares ancoradas a lípidos através de
interações fracas; a sua incorporação na membrana celular depende da afinidade específica em relação
às jangadas lipídicas [77-80]. No esquema da Figura 5, encontra-se a representação das moléculas
presentes nas jangadas lipídicas.
Figura 5. Esquema representativo de jangadas lipídicas. As jangadas lipídicas podem ter diferentes composições
lipídicas e proteícas (jangada lipídica 1 e jangada lipídica 2). Adaptado de [81].
A primeira evidência da existência de jangadas lipídicas in vivo foi a deteção, em células, de
microdomínios designados por Membranas Resistentes a Detergentes (DRMs). As DRMs são frações
de membrana ricas em colesterol e esfingomielina e são resistentes à solubilização por Triton X-100.
No entanto, este processo de extração pode ser problemático uma vez que pode causar modificações
no estado físico dos lípidos, e levar à remoção de algumas proteínas ou lípidos. Além disso, exige a
existência de marcadores internos, o controlo da depleção do colesterol e o uso de mais do que um
detergente. As DRMs são diferentes das jangadas lipídicas nativas em tamanho, composição e
abundância no organismo [69, 76, 79, 82-85].
A monitorização de jangadas lipídicas em células vivas pode ser feita recorrendo a diferentes
técnicas como a Espectroscopia de Ressonância Magnética Nuclear (NMR), que se baseia nas
mudanças de orientação dos grupos polares do lípido devido à formação de microdomínios, ou a
Calorimetria Diferencial de Varrimento, avaliando a existência de microdomínios através da
determinação da imiscibilidade de fase [86-88].
Jangada lipídica 1 Jangada lipídica 2

9
2.3. Modelos Biomembranares
O estudo do comportamento das membranas celulares faz-se frequentemente utilizando modelos
simplificados [89]. Os modelos membranares biomiméticos imitam a situação in vivo, e, de acordo com
Makky et. al., podem consistir em diferentes sistemas organizados, como monocamadas ou bicamadas
planares ou curvas (lipossomas) [90]. As membranas biomiméticas são formadas por misturas de
fosfolípidos, cuja composição se assemelha à composição real da membrana celular em causa. Através
destas é possível estudar processos biológicos in vitro, funções de proteínas, deposição de lipossomas,
entre outros estudos biofísicos [91-93].
a) Bicamadas Lipídicas Suportadas (SLBs) também conhecidas como bicamadas planares
suportadas (Figura 6), são um modelo biológico muito popular para mimetizar as membranas celulares,
podendo ser usadas para compreender as propriedades intrínsecas dos lípidos e para diferentes
estudos biomiméticos, biofísicos ou bioquímicos [94-96]. Estas bicamadas são suportadas em
substratos sólidos; entre a SLB e o substrato sólido encontra-se uma camada aquosa com 10 a 20 Å
de espessura [94, 97]. Os substratos usados para as SLBs depende da técnica a que a bicamada
lipídica se destina e são tipicamente de mica, vidro de borosilicato, óxido de titânio, ouro, platina, silício
oxidado e dióxido de silício (ou sílica). Existem, também, diversos métodos para a deposição dos lípidos
no substrato sólido e para a formação das SLBs: técnica de Lagmuir-Blodgett (LB) e técnica de Lagmuir-
Schaefer (LS), e a adsorção e fusão de vesículas lipídicas a partir de uma suspensão aquosa. Esta
última técnica aparenta ser o método mais simples e popular; a fusão é controlada pela composição,
pH, força iónica, pressão osmótica, tamanho e carga da superfície das vesículas. A partir deste método,
é possível obter uma bicamada estável, permitindo o uso de técnicas analíticas de superfície para
provar a existência de interações [95, 98].
Figura 6. Representação esquemática da Bicamada Fosfolipídica Suportada, num substrato sólido. Adaptado de
[98].
b) Bicamadas Fosfolipídicas Planares, também conhecidas como membranas lipídicas negras
(Figura 7), têm sido usadas para investigar diversos processos biofísicos. Trata-se de um modelo de
bicamada que envolve a formação de uma membrana ao longo de um orifício com 1 mm de diâmetro,
num material hidrofóbico, como o polietileno ou o teflon. Comparativamente às SLBs, as membranas
lipídicas negras não são suportadas por um substrato, encontrando-se em meio aquoso, onde está
presente um elétrodo de referência. O método mais popular para a formação deste modelo envolve a
deposição da solução lipídica sobre o orifício, e a formação de uma dupla camada com curvatura.
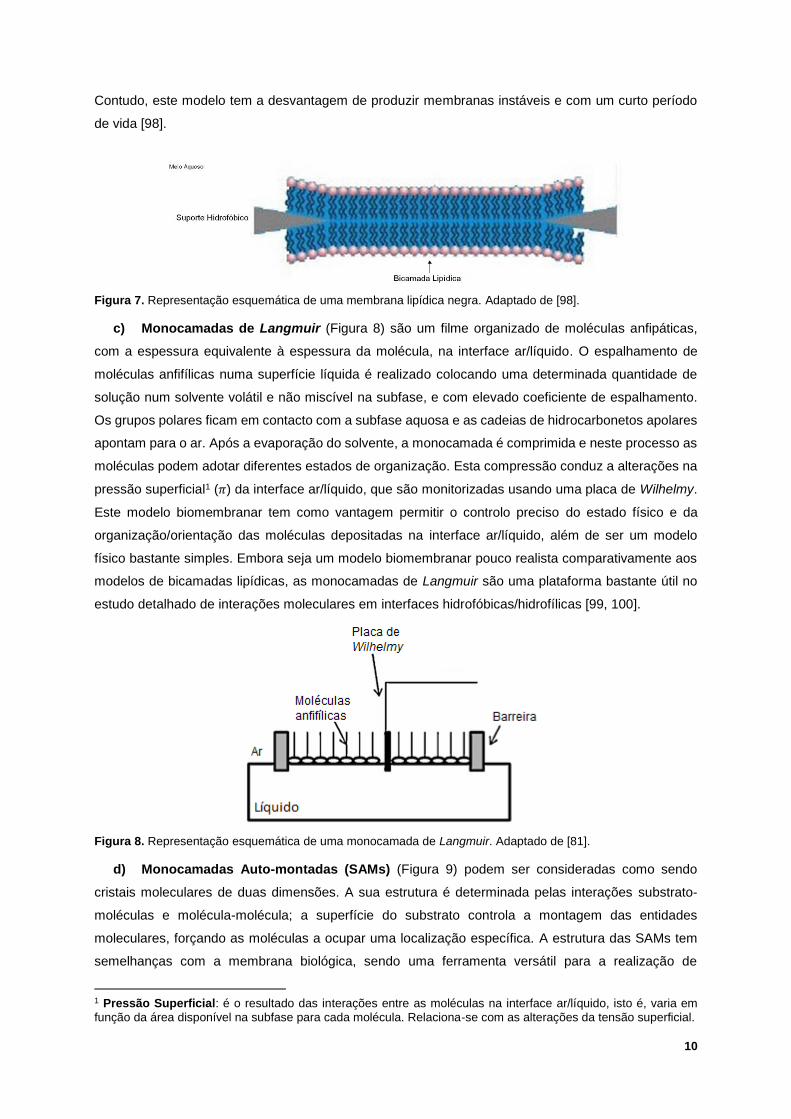
10
Contudo, este modelo tem a desvantagem de produzir membranas instáveis e com um curto período
de vida [98].
Figura 7. Representação esquemática de uma membrana lipídica negra. Adaptado de [98].
c) Monocamadas de Langmuir (Figura 8) são um filme organizado de moléculas anfipáticas,
com a espessura equivalente à espessura da molécula, na interface ar/líquido. O espalhamento de
moléculas anfifílicas numa superfície líquida é realizado colocando uma determinada quantidade de
solução num solvente volátil e não miscível na subfase, e com elevado coeficiente de espalhamento.
Os grupos polares ficam em contacto com a subfase aquosa e as cadeias de hidrocarbonetos apolares
apontam para o ar. Após a evaporação do solvente, a monocamada é comprimida e neste processo as
moléculas podem adotar diferentes estados de organização. Esta compressão conduz a alterações na
pressão superficial1 (𝜋) da interface ar/líquido, que são monitorizadas usando uma placa de Wilhelmy.
Este modelo biomembranar tem como vantagem permitir o controlo preciso do estado físico e da
organização/orientação das moléculas depositadas na interface ar/líquido, além de ser um modelo
físico bastante simples. Embora seja um modelo biomembranar pouco realista comparativamente aos
modelos de bicamadas lipídicas, as monocamadas de Langmuir são uma plataforma bastante útil no
estudo detalhado de interações moleculares em interfaces hidrofóbicas/hidrofílicas [99, 100].
Figura 8. Representação esquemática de uma monocamada de Langmuir. Adaptado de [81].
d) Monocamadas Auto-montadas (SAMs) (Figura 9) podem ser consideradas como sendo
cristais moleculares de duas dimensões. A sua estrutura é determinada pelas interações substrato-
moléculas e molécula-molécula; a superfície do substrato controla a montagem das entidades
moleculares, forçando as moléculas a ocupar uma localização específica. A estrutura das SAMs tem
semelhanças com a membrana biológica, sendo uma ferramenta versátil para a realização de
1 Pressão Superficial: é o resultado das interações entre as moléculas na interface ar/líquido, isto é, varia em
função da área disponível na subfase para cada molécula. Relaciona-se com as alterações da tensão superficial.

11
modificações superficiais; contudo, tratando-se de uma monocamada, é um modelo menos realista que
os modelos baseados em bicamadas [101-103]. As SAMs são obtidas através da modificação do
substrato com alcanotióis, de forma a obter uma superfície hidrofóbica, facilitando a formação de uma
membrana híbrida. As SAMs, comparativamente com as SLBs, são mais rígidas, devido ao
acoplamento com o substrato, normalmente de ouro; uma vez que se trata de um material relativamente
inerte, o ouro resiste à oxidação e a outras contaminações [98, 104]. As monocamadas podem ser
estudadas e analisadas por técnicas como a QCM-D; contudo, algumas técnicas rápidas e sensíveis,
como a espectroscopia NMR, não apresentam resultados tão bons para monocamadas, como
apresentam para bicamadas [98, 104, 105].
Figura 9. Representação esquemática de SAMs, com um substrato de ouro. Adaptado de [98].
e) Lipossomas, ou vesículas fosfolipídicas unilamelares ou multilamelares (Figura 10), em
suspensão ou adsorvidos/suportados num dado substrato, são um dos modelos mais adequados para
estudar a membrana lipídica das células biológicas [77, 106]. A deposição dos lipossomas é usada para
criar bicamadas homogéneas ou camadas vesiculares estáveis, dependendo do material do substrato
e do tamanho do lipossoma [91, 106, 107]. Os lipossomas são um sistema muito semelhante às células
nativas, sendo a bicamada fosfolipídica um meio de separação entre o espaço extracelular e o líquido
intracelular. [107, 108].
Figura 10. Representação esquemática de uma vesícula fosfolipídica (lipossoma). Adaptado de [107].
2.3.1. Modelos para jangadas lipídicas
São conhecidas diferentes composições lipídicas para mimetizar o comportamento de fase das
jangadas lipídicas. As bicamadas lipídicas podem ser classificadas em três fases, conforme a sua
fluidez: fase lamelar líquida-ordenada (Lo), fase lamelar líquida-desordenada (Ld) e fase sólida
ordenada (Lb) [77]. De facto, sabe-se que um local rico em esfingomielina e colesterol resulta na
formação de uma Lo e que esta coexiste com domínios ricos em glicerofosfolípidos não saturados numa
12
Ld. Assim, a separação de fase entre as fases Ld e Lo, na presença de colesterol, é essencial para a
simulação de jangadas lipídicas [109-111].
As misturas lipídicas ternárias constituídas por dois lípidos com diferentes temperaturas de fusão
e ricas em colesterol têm sido usadas na composição de membranas modelo para simular as jangadas
lipídicas [111]. Geralmente, o comportamento de fase das misturas modelo é representado como um
diagrama de fase ternário a uma temperatura constante, sendo a temperatura fisiológica (37ºC) ou a
temperatura ambiente (25ºC) as mais comuns. As misturas lipídicas ternárias apresentam uma
composição lipídica muito mais simples do que as membranas celulares mas o seu comportamento de
fase produz uma simulação satisfatória do comportamento de fase in vivo [77, 109, 112-115].
As misturas equimolares de glicerofosfolípidos/esfingolípidos/colesterol são consideradas misturas
canónicas de jangada lipídica, por apresentarem composição semelhante às DRMs extraídas das
células. As composições mais comuns destas misturas são: 1,2-dioleoil-sn-glicero-3-fosfocolina/ 1,2-
dipalmitoil-sn-glicero-3-fosfocolina/colesterol (DOPC/DPPC/Chol) e por DOPC/SM/Chol [71, 113].
Analisando as Tt, o 1-palmitoil-2-oleoil-sn-glicero-3-fosfocolina (POPC) é a melhor opção para um lípido
com baixa Tt, porque é a fosfatidil colina mais relevante na membrana celular, capaz de ser obtida por
extração a partir de fontes biológicas. Para um lípido com elevada T t, a esfingomielina deverá ser a
opção mais adequada, uma vez que permite a ligação do hidrogénio com o grupo hidroxilo do colesterol
e está presente na membrana nativa de diferentes tipos de células [77, 116].
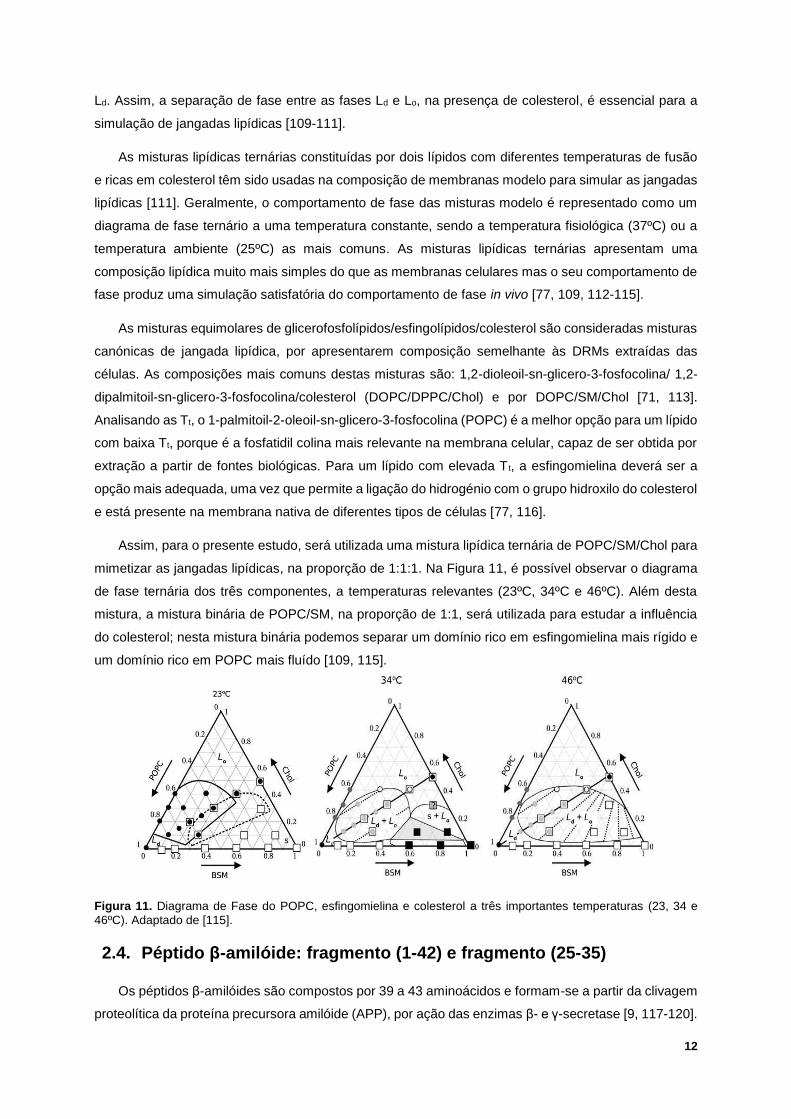
Assim, para o presente estudo, será utilizada uma mistura lipídica ternária de POPC/SM/Chol para
mimetizar as jangadas lipídicas, na proporção de 1:1:1. Na Figura 11, é possível observar o diagrama
de fase ternária dos três componentes, a temperaturas relevantes (23ºC, 34ºC e 46ºC). Além desta
mistura, a mistura binária de POPC/SM, na proporção de 1:1, será utilizada para estudar a influência
do colesterol; nesta mistura binária podemos separar um domínio rico em esfingomielina mais rígido e
um domínio rico em POPC mais fluído [109, 115].
Figura 11. Diagrama de Fase do POPC, esfingomielina e colesterol a três importantes temperaturas (23, 34 e
46ºC). Adaptado de [115].
2.4. Péptido β-amilóide: fragmento (1-42) e fragmento (25-35)
Os péptidos β-amilóides são compostos por 39 a 43 aminoácidos e formam-se a partir da clivagem
proteolítica da proteína precursora amilóide (APP), por ação das enzimas β- e γ-secretase [9, 117-120].

13
A principal função não patológica dos péptidos β-amilóides é a regulação do transporte de colesterol
[121, 121]. A parte central destes péptidos possui zonas de resíduos hidrofóbicos (17-21 e 30-40/42);
estes resíduos associam-se numa estrutura tipo gancho de cabelo (hairpin) para interagir uns com os
outros [123]. Os péptidos β-amilóides ocorrem em várias isoformas que diferem entre si pelo número
de aminoácidos no terminal C (Figura 12); péptidos Aβ1-42 é, juntamente com péptidos Aβ1-40, a forma
mais comum em espécies in vivo (Figura 12) [12, 122, 124, 125].
Segundo Vieira et. al. [126], os péptidos β-amilóides apresentam conformações hélice α em
solventes orgânicos apolares, e folha β, em solventes orgânicos polares; no seu estado solúvel, a
transição estrutural de hélice α para folha β pode conduzir à agregação molecular e à formação de
fibrilas [126, 127]. Os péptidos Aβ1-42, no estado monomérico, agregam-se em oligómeros solúveis de
baixo peso molecular. Estes oligómeros podem acumular-se, formando protofibrilas oligoméricas que,
por sua vez, podem transformar-se em fibras insolúveis que constituem as placas amilóides presentes
em doentes com doença de Alzheimer [9, 13, 127, 128].
Figura 12. Representação da sequência do péptido β-amilóide (1-42), (1-40) e (25-35), identificando os terminais
C e N. Adaptado de [129, 130].
Em particular, os péptidos Aβ1-42 possuem propriedades tensioativas e têm tendência para a
acumulação e agregação peptídica. A presença de lipossomas diminui a agregação peptídica, uma vez
que a interação entre os péptidos Aβ1-42 e os lipossomas é favorecida pela quantidade de fosfolípidos
presentes na bicamada: quanto mais lipossomas estiverem presentes, maior será a interação entre os
péptidos Aβ1-42 e os lipossomas e, consequentemente, menor a interação entre péptidos. Estes
péptidos interatuam com modelos da membrana celular, através da sua ligação a lípidos, sendo esta
interações explicadas na secção 2.5 [47, 131, 132].
O fragmento peptídico Aβ25-35 formado por 11 aminoácidos, está partcialmente localizado no
domínio hidrofóbico e apresenta as mesmas propriedades de toxicidade, alterações morfológicas e de
agregação que o péptido de comprimento total. Este fragmento peptídico (GSNKGAIIGLM) representa
o fragmento β-amilóide mais pequeno processado in vivo por proteases no cérebro [8, 133] e é
suficiente para conduzir à formação de fibrilas de péptido Aβ e à degeneração neuronal. Além disso,
os agregados do fragmento peptídico Aβ25-35 apresentam atividade neurotrófica e anfifílica, tal como o
fragmento completo [132, 134-138]. A estrutura secundária do fragmento peptídico Aβ25-35 depende de
fatores como a concentração e o pH. Loew [139] demonstrou que a estrutura secundária do fragmento
peptídico Aβ25-35 a uma concentração elevada e a pH=7 era essencialmente hélice α e folha β, enquanto
que a pH=4 a estrutura secundária incluía também espiral aleatória (random coil). Outros autores
confirmaram que a pH=7 e a uma elevada concentração peptídica este fragmento adotava a estrutura
folha β, mas a concentrações peptídicas mais baixas predominava a estrutura volta β (β turn) [140].
Sendo menos complexo do que o péptido completo Aβ1-42, mas apresentando as mesmas

14
características, o fragmento peptídico Aβ25-35 é um modelo muito atraente para estudar a agregação
dos péptidos Aβ1-42 e as interações destes péptidos, envolvidos na doença de Alzheimer, com modelos
biomembranares [137, 141].
2.4.1. Agregação Peptídica
As mudanças estruturais que ocorrem num péptido e a sua cinética são influenciadas pela
hidrofilicidade do ambiente e pela presença de lípidos em solução [142, 143]. Os péptidos β-amilóides
possuem uma elevada tendência de agregação em fibrilas amilóides, sendo estes agregados insolúveis
compostos principalmente por agregados de folha β. O processo de agregação destes péptidos passa
pela transformação de monómeros livres e solúveis em fibrilas amilóides insolúveis. De acordo com
Labbé et al. [140], esta agregação é devida à formação de estruturas ordenadas organizadas em folha
β, em resultado do estabelecimento de uma rede de ligações de hidrogénio, envolvendo os grupos CO
e NH. O processo de agregação amilóide é alvo de pesquisas biológicas e biofísicas, devido ao seu
papel em doenças neurodegenerativas, como a doença de Alzheimer [144, 145].
Os solventes fluorados, como hexafluorisopropanol (HFIP), são utilizados para estabilizar a estrutura
monomérica dos péptidos β-amilóides. O pré-tratamento das soluções com HFIP leva a um significativo
melhoramento da solubilidade do péptido β-amilóide, favorecendo o estado monomérico do péptido em
solução, devido à dissolução dos agregados peptídicos, tornando a solução mais homogénea. Deste
modo, o procedimento normal para o pré-tratamento com HFIP consiste na dissolução dos péptidos
neste álcool, conduzindo a uma solução livre de agregados e à evaporação de álcoois [125-127, 137,
146]. Este pré-tratamento tem ainda como objetivo a manutenção da estrutura hélice α do péptido β-
amilóide [125].
Existem, ainda, outras propostas para a preparação de soluções peptídicas, evitando a agregação.
O péptido β-amilóide pode ser dissolvido em dimetilsulfóxido (DMSO). Para a desagregação de um
péptido, isto é, para obter o estado monomérico deste, é possível recorrer também a uma dissolução
utilizando trifluoroacético/trifluoroetanol/hexafluorisopropanol (TFA/TFE/HFIP) [35, 127, 147-150].
2.5. Interação entre modelos biomembranares e péptido Aβ
O estudo dos efeitos dos péptidos β-amilóides em membranas lipídicas pode ser feito recorrendo a
modelos biomembranares e, ao longo dos vários anos de investigação, surgiram diversas propostas
para mecanismos de neurotoxicidade dos péptidos β-amilóide.
Segundo Terzi et al. [151], o péptido Aβ1-40 pode ligar-se à superfície da membrana celular carregada
negativamente através de ligações de atração eletrostática. Logo, a concentração de péptido Aβ1-40 à
superfície da membrana celular é maior do que em solução. Se os potenciais negativos da superfície
celular aumentarem, existe maior possibilidade de os péptidos se ligarem à superfície celular
electrostaticamente, aumentando a velocidade de agregação peptídica [15].
Ding et al. [46] estudaram as interações entre péptidos β-amilóides com bicamadas lipídicas
suportadas. Os autores concluíram que a interação entre péptidos Aβ1-40 e a bicamada lipídica inicia-

15
se com uma ligação muito forte do péptido no estado monomérico à membrana e que, posteriormente
a um período de incubação de 20 horas a 22ºC, há formação de oligómeros na membrana, verificando-
se agregação peptídica. O tamanho destes oligómeros depende diretamente da concentração de
péptido em solução.
Por seu lado, Sabaté et al. [131], utilizando lipossomas como modelo biomembranar, demonstraram
que a presença de péptidos β-amilóides pode conduzir à disrupção da membrana celular. Este efeito
de disrupção é mais forte na presença de membranas carregadas negativamente, uma vez que a
interação entre os péptidos β-amilóides e os lipossomas é favorecida pela presença de fosfolípidos
negativos na bicamada.
Ambroggio et al. [35], estudaram a estabilidade dos modelos biomembranares, representados por
vesículas unilamelares gigantes (GUVs), expostos a uma solução de péptido β-amilóide (1-42). Este
estudo revelou que o fragmento (1-42) conduz à disrupção das GUVs, compostas por POPC ou
POPC/SM/Chol (1:1:1). Demonstrou-se assim que a estabilização da membrana lipídica é diretamente
alterada quando as GUVs são expostas aos péptidos. Assim, os autores concluíram neste estudo que
diminuição da estabilização da membrana lipídica é resultado da interação entre péptidos e lípidos.
Ambroggio et al. também demonstraram, através de Microscopia de Força Atómica (AFM), que o
fragmento amilóide (1-42) forma estruturas hexaméricas em membranas celulares modelo, estando
este resultado de acordo com Lin et al. [152, 153].
Através de modelos biomembranares computacionais, Vestergaard et al. [154] investigaram a
interação dos péptidos β-amilóides com membranas lipídicas e as possíveis transformações
morfológicas resultantes. Os autores concluíram que as espécies oligoméricas do péptido Aβ1-42 se
localizam mais perto da superfície da membrana enquanto as fibrilas têm uma localização variada.
Demonstraram que a presença de péptido Aβ1-42 conduz a um aumento da área da superfície da
membrana, induzindo a transformação vesicular dos lípidos, devido aos péptidos mediarem a fusão da
membrana.
Outro estudo por simulação foi desenvolvido por Qiu et al. [155] que investigaram as interações
entre os péptidos β-amilóides e membranas enriquecidas ou não em colesterol, verificando o papel
crucial que o colesterol tem na prevenção da formação de estruturas folha β e da disrupção da
membrana.
Relativamente ao fragmento peptídico β-amilóide (25-35), também diferentes estudos foram
realizados usando modelos biomembranares. Em 1995, Terzi et al. [156], descobriram que a adição de
lipossomas de POPC/POPG a uma solução de péptido Aβ25-35 induzia uma transição na formação do
péptido de espiral aleatória para folha β.
Mason et al. [157], em 1996, afirmaram que o fragmento Aβ25-35 incorporado em lipossomas de
POPC apresenta os seus resíduos 25 a 27 localizados na parte hidrofílica da membrana, enquanto os
restantes resíduos apolares se localizam no interior do lipossoma (parte hidrofóbica).

16
Martinez-Senac et al. [158], estudaram a influência de diferentes composições de lípidos, na forma
de lipossomas, na estrutura do fragmento peptídico Aβ25-35, usando a técnica de espectroscopia de FT-
IR (espectroscopia de infravermelhos por transformada de Fourier). Os autores concluíram que os
lipossomas compostos por fosfolípidos com carga negativa, como 1,2-dimiristoil-sn-glicero-3-
fosfatidilserina (DMPS) ou 1,2-ditetradecanoil-sn-glicero-3-fosfato (DMPA), aceleram a agregação
peptídica, enquanto os lipossomas formados por fosfolípidos zwiteriónicos, como 1,2-dimiristoil-sn-
glicero-3-fosfatidilcolina (DMPC), diminuem a tendência de agregação. Através deste estudo, os
autores provaram que a interação entre estes péptidos e os fosfolípidos que formam a
membrana/modelo biomembranar tem uma natureza eletrostática.
De modo a entender o papel do colesterol, Labbé et al. [140], estudaram a interação entre o
fragmento peptídico Aβ25-35 e membranas modelo de 1,2-dimiristoil-sn-glicero-3-fosfatidilcolina /1,2-
dimiristoil-sn-glicero-3-fosfoglicerol (DMPC/DMPG), na presença e na ausência de colesterol.
Concluíram que o péptido interage com os fosfolípidos, alterando a fluidez e elasticidade da membrana,
sendo esta interação beneficiada na ausência de colesterol.
Triulzi et al. [159] modificaram o fragmento peptídico Aβ25-35 com uma cadeia alifática C18 no
terminal N do péptido de modo a estudar a agregação do péptido Aβ25-35 quando há formação de uma
monocamada de Langmuir na interface ar/líquido. Os autores concluíram que a pressões superficiais
baixas, a estrutura da monocamada apresenta uma pequena contribuição de folha β, mas que essa
contribuição aumenta significativamente quando a pressão superficial aumenta e a monocamada passa
para a fase de líquido-condensado (LC).
Lao et al. [160] estudaram a interação entre o fragmento peptídico Aβ25-35 e modelos
biomembranares de fosfolípidos aniónicos e colesterol. Os autores afirmaram que o fragmento Aβ25-35
interage preferencialmente com os grupos polares dos lípidos e que perturba pouco o núcleo de
hidrocarbonetos quando os péptidos são adicionados aos lipossomas já existentes; contudo, no caso
de os péptidos serem incluídos nos lipossomas durante a sua formação, os autores concluíram que os
péptidos penetram no interior da bicamada fosfolipídica do modelo biomembranar.
2.6. Técnicas Experimentais
2.6.1. Microbalança de Cristal de Quartzo com Dissipação (QCM-D)
A Microbalança de Cristal de Quartzo com Dissipação permite determinar de forma precisa e em
tempo real a massa e as propriedades viscoelásticas de um filme depositado sobre um sensor de
quartzo. Trata-se de uma técnica que é aplicada com sucesso para medição de processos de adsorção
em ambientes líquidos e gasosos [106, 161] e numa série de estudos relacionados com a conformação
e agregação de proteínas, por exemplo para quantificação da agregação de péptidos amilóides numa
dada interface sólido/líquido [162-165].
A QCM-D permite a medição simultânea das alterações que ocorrem na frequência de ressonância
do cristal (f) e na dissipação de energia (D) [162]. Uma possível alteração na frequência de ressonância
do cristal está relacionada com a massa adsorvida no cristal, isto é, a presença de biomoléculas na

17
solução que entra em contacto com o cristal de quartzo, leva à formação de um filme na superfície
deste, alterando a frequência de ressonância; por outro lado, a dissipação de energia está relacionada
com as propriedades viscoelásticas do filme adsorvido no cristal [108].
O princípio de funcionamento da QCM-D baseia-se essencialmente no facto dos cristais de quartzo
serem piezoelétricos, o que conduz a uma deformação mecânica quando expostos a um campo elétrico
oscilatório e a uma onda de cisalhamento que se propaga através da amostra [108, 128]. Enquanto a
variação da frequência de ressonância (Δf) é obtida antes de desligar a fonte, a variação da dissipação
de energia (ΔD) é obtida depois. Este processo é repetido a cada segundo, permitindo obter resultados
em tempo real [108, 128, 162, 166].
O parâmetro de dissipação é definido através da Equação 1:
(1)
onde 𝐸𝑑𝑖𝑠é a energia dissipada durante uma oscilação e 𝐸𝑠𝑡 é a energia armazenada pelo filme durante
a mesma oscilação [167].
Para camadas rígidas ideais, que estão acopladas à oscilação do cristal e distribuídas
homogeneamente sobre a superfície, comportando-se como um filme elástico, verificam-se alterações
na frequência e na velocidade proporcionais à massa adsorvida, sendo o fator de dissipação
desprezado. Para esta hipótese ser considerada válida, os valores de dissipação devem ser inferiores
a 1 × 10−6, para frequências na ordem dos 5 MHz [128, 167, 168]. Neste caso existe uma relação direta
entre Δf e a massa adsorvida (Δm), dada por (Equação 2):
(2)
onde 𝐶𝑓é uma constante de sensibilidade característica do sensor utilizado (para o caso dos cristais de
quartzo, a 5 MHz, 𝐶𝑓 = 17,7 𝑛𝑔. 𝐻𝑧−1. 𝑐𝑚−2) e n é o número da harmónica [107]. Esta equação designa-
se por Equação de Sauerbrey [106, 168].
Se o filme formado no cristal de quartzo não for rígido e fino, a Equação de Sauerbrey não é válida,
sendo necessário introduzir modificações nesta equação para ter em conta as propriedades
viscoelásticas do material adsorvido. Estas modificações baseadas na aplicação do Modelo de Voigt
conduzem à Equação 3 [165, 169]:
(3)
st
dis
E
ED
2
fn
Cm
f
f
f
II
f
Jn
Cfm
"
1

18
onde ω é a frequência angular, 𝜌𝐼 é a densidade do líquido, 𝜂𝐼 é a viscosidade do líquido, 𝜌𝑓 é a
densidade do filme e, por fim, 𝐽𝑓" é dado pelo quociente
𝜔.𝜂𝑓
𝜇𝑓2+𝜔2.𝜂𝑓
2 em que 𝜇𝑓 é a elasticidade do filme e
𝜂𝑓 é a viscosidade do filme.
2.6.2. Balança de Langmuir
A técnica de Langmuir é muito importante na preparação de monocamadas de moléculas biológicas,
que podem simular as membranas celulares [170-173]. A principal vantagem do método de Langmuir
é a formação de monocamadas homogéneas, compactas, de espessura e organização controladas.
Além disso, este método exige tempos de preparação curtos e permite também estudar fenómenos de
adsorção. Uma das principais características desta técnica é a possibilidade de transferir para
substratos sólidos as monocamadas formadas (técnica de Langmuir-Blodgett), permitindo o recurso a
uma vasta gama de técnicas de caracterização de superfícies que podem ser usadas para estudar a
estrutura dos filmes formados a partir da técnica de Langmuir, como AFM [171, 172, 174-177].
O método de Langmuir baseia-se na formação de um filme/monocamada na interface ar/líquido.
Nesta técnica, de modo geral, são utilizadas moléculas anfifílicas que apresentam uma porção
hidrofóbica e uma porção hidrofílica bem definidas [175]. As moléculas orgânicas, como os lípidos,
fosfolípidos ou glicolípidos, orientam-se na interface ar/líquido de modo a minimizar a energia livre e
formar uma monocamada insolúvel. A organização destas monocamadas, bem como a qualidade do
filme, depende de diversos parâmetros: pH, temperatura, velocidade, tipo de deposição e as
propriedades termodinâmicas da interface ar/líquido [172, 177].
Quando a área da monocamada é reduzida através de um sistema de compressão, usando uma ou
duas barreiras movíveis (compressão assimétrica ou simétrica, respetivamente), a distância
intermolecular diminui e a pressão superficial aumenta. A força exercida pela monocamada por unidade
de comprimento é designada por pressão superficial (𝜋) (Equação 4):
(4)
onde 𝛾0 é a tensão superficial do líquido puro e 𝛾 a tensão superficial da superfície ocupada pela
monocamada. O diagrama que representa a variação da pressão superficial em função da área
ocupada por molécula, obtido a temperatura constante, designa-se por isotérmica Pressão Superficial
vs. Área por molécula (𝜋-A) (Figura 13). A análise destas isotérmicas fornece informação sobre a
estrutura, interações, transições de fase e compressibilidade da monocamada [172, 173].
Durante a compressão das barreiras, verifica-se uma variação da pressão superficial. Esta variação
deve-se às mudanças de organização que ocorrem nas moléculas presentes na subfase. Assim, antes
de se verificar o início do aumento da pressão superficial, designado por lift-off (A0), as moléculas
encontram-se num estado gasoso (G); neste estado as moléculas não interagem entre si. Quando a
monocamada é sujeita a compressão, verifica-se um aumento da pressão superficial e
consequentemente uma diminuição da área disponível por molécula, indicando a ocorrência de uma
mudança de estado gasoso para um estado líquido-expandido (LE); neste estado as moléculas
interatuam entre si mas ainda se encontram distanciadas umas das outras, embora já se encontrem
0
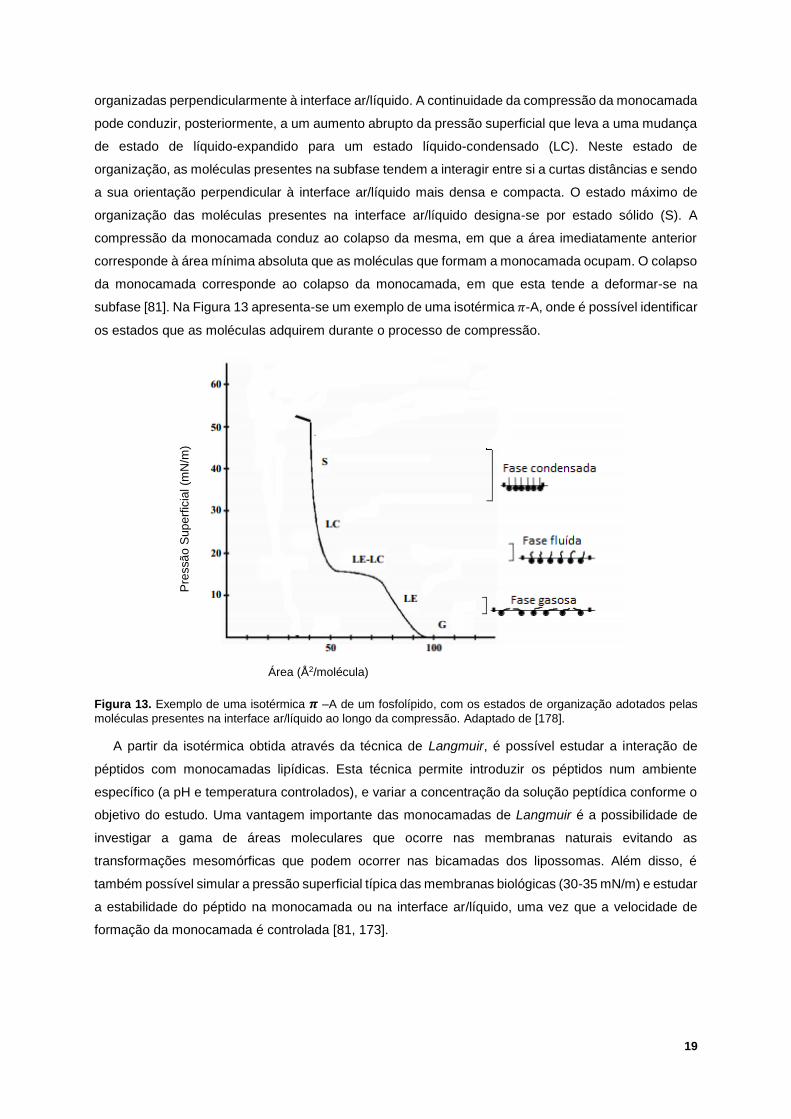
19
organizadas perpendicularmente à interface ar/líquido. A continuidade da compressão da monocamada
pode conduzir, posteriormente, a um aumento abrupto da pressão superficial que leva a uma mudança
de estado de líquido-expandido para um estado líquido-condensado (LC). Neste estado de
organização, as moléculas presentes na subfase tendem a interagir entre si a curtas distâncias e sendo
a sua orientação perpendicular à interface ar/líquido mais densa e compacta. O estado máximo de
organização das moléculas presentes na interface ar/líquido designa-se por estado sólido (S). A
compressão da monocamada conduz ao colapso da mesma, em que a área imediatamente anterior
corresponde à área mínima absoluta que as moléculas que formam a monocamada ocupam. O colapso
da monocamada corresponde ao colapso da monocamada, em que esta tende a deformar-se na
subfase [81]. Na Figura 13 apresenta-se um exemplo de uma isotérmica 𝜋-A, onde é possível identificar
os estados que as moléculas adquirem durante o processo de compressão.
Figura 13. Exemplo de uma isotérmica 𝝅 –A de um fosfolípido, com os estados de organização adotados pelas
moléculas presentes na interface ar/líquido ao longo da compressão. Adaptado de [178].
A partir da isotérmica obtida através da técnica de Langmuir, é possível estudar a interação de
péptidos com monocamadas lipídicas. Esta técnica permite introduzir os péptidos num ambiente
específico (a pH e temperatura controlados), e variar a concentração da solução peptídica conforme o
objetivo do estudo. Uma vantagem importante das monocamadas de Langmuir é a possibilidade de
investigar a gama de áreas moleculares que ocorre nas membranas naturais evitando as
transformações mesomórficas que podem ocorrer nas bicamadas dos lipossomas. Além disso, é
também possível simular a pressão superficial típica das membranas biológicas (30-35 mN/m) e estudar
a estabilidade do péptido na monocamada ou na interface ar/líquido, uma vez que a velocidade de
formação da monocamada é controlada [81, 173].
Área (Å2/molécula)
Pre
ssã
o S
up
erf
icia
l (m
N/m
)

20
2.6.3. Análise de Localização de Nanopartículas
A técnica de Análise de Localização de Nanopartículas (NTA) permite visualizar e medir, em tempo
real, o tamanho (entre 10 nm e 1 µm) de partículas presentes numa determinada solução; além disso,
esta técnica permite calcular o número de nanopartículas existentes na solução e obter informações
relativamente ao índice de refração, fluorescência e potencial zeta das nanopartículas [179, 180].
A técnica de NTA relaciona o grau de movimento Browniano com o diâmetro hidrodinâmico das
nanopartículas (consideradas esféricas), permitindo determinar a distribuição de tamanho das
nanopartículas, num curto espaço de tempo. Recorrendo a um microscópio, esta técnica determina a
posição das nanopartículas presentes na solução através da deteção da luz que estas dispersam
quando o laser incide sobre as mesmas [179, 181, 182]. Utiliza-se um feixe laser de 635, 535 ou 404
nm para iluminar as nanopartículas presentes na solução em análise. Geralmente, as nanopartículas
em solução são mais pequenas do que o comprimento de onda da luz utilizado, e a informação sobre
a morfologia das nanopartículas é pequena; assim, estas partículas atuam como pontos de dispersão
de luz, permitindo observar o seu movimento Browniano. Ajustada ao microscópio, existe uma câmara
de CCD (Electron Multiplying Charged Coupled Device), usada para realizar uma gravação da zona
com nanopartículas onde o laser incide. O movimento Browniano, por sua vez, é obtido através de um
software associado à técnica de NTA, que permite determinar o movimento do centro de cada partícula
no plano x, y. Assim, é possível observar e gravar, simultaneamente, um vídeo das partículas
visualizadas através da técnica de NTA, bem como obter um gráfico de distribuição de tamanho das
nanopartículas, recorrendo a um software de análise [180, 181, 183].
2.6.4. Calorimetria Diferencial de Varrimento (DSC)
A técnica de Calorimetria Diferencial de Varrimento (DSC) assenta na medição dos fluxos de calor
associados a transições que ocorrem nos materiais, em função da temperatura ou do tempo. O DSC
permite caracterizar os parâmetros termodinâmicos que controlam interações não covalentes e a
estabilidade de moléculas biológicas [184]. O DSC é uma técnica padrão usada para o estudo das
transições de fase dos sistemas lipídicos e das interações entre modelos de bicamada lipídica e outras
moléculas [86].
A técnica de DSC baseia-se no registo da diferença entre a capacidade calorífica, Cp, do conteúdo
presente na célula da amostra e do conteúdo da célula de referência, num dado intervalo de
temperaturas e a uma dada taxa de varrimento. A célula que contém a amostra e a célula que contém
a referência são submetidas à mesma sequência de ciclos aquecimento/arrefecimento. O principal
parâmetro que retiramos desta técnica é a temperatura de fusão das amostras analisadas; esta
temperatura corresponde à temperatura média do pico de transição de fase. Além disso, pode também
obter-se a entalpia de transição desde que seja possível traçar uma linha de base adequada entre o
início e o fim do aquecimento. O DSC permite avaliar a reversibilidade do comportamento térmico da
amostra [86, 185].
No aparelho de DSC, a amostra e a referência são analisadas ao mesmo tempo, devido ao
posicionamento do bloco de aquecimento. No decorrer de uma experiência de DSC, a amostra e a
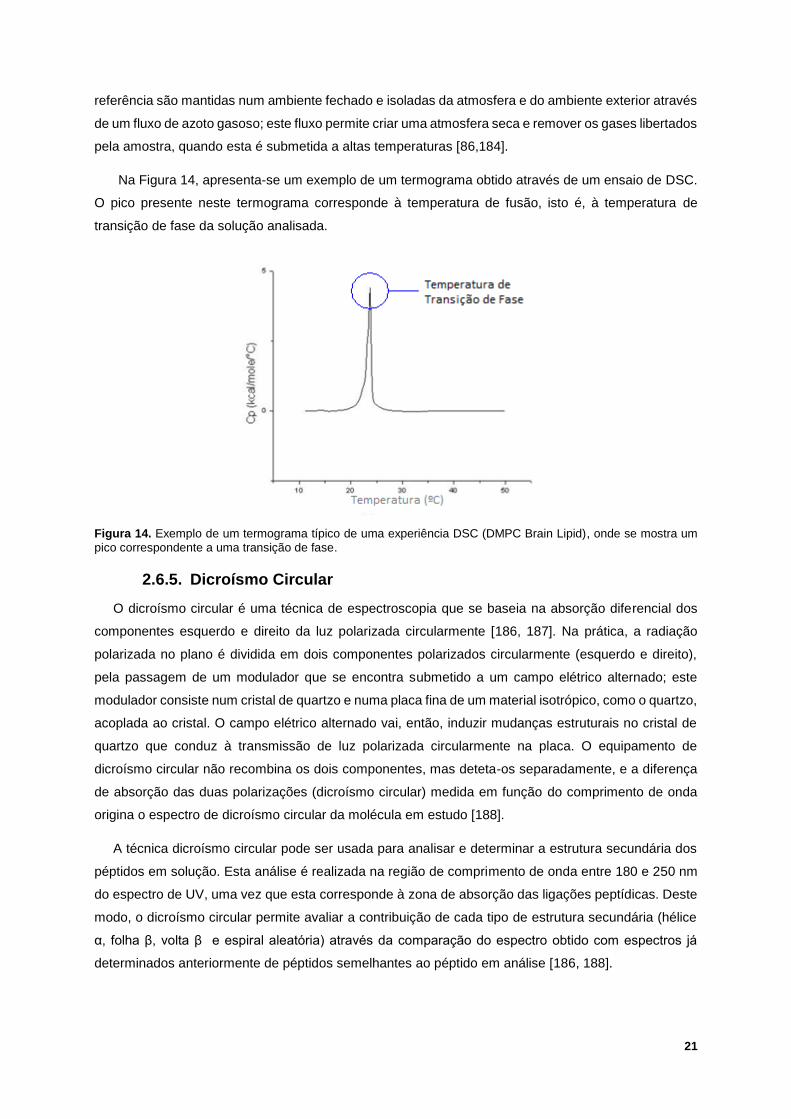
21
referência são mantidas num ambiente fechado e isoladas da atmosfera e do ambiente exterior através
de um fluxo de azoto gasoso; este fluxo permite criar uma atmosfera seca e remover os gases libertados
pela amostra, quando esta é submetida a altas temperaturas [86,184].
Na Figura 14, apresenta-se um exemplo de um termograma obtido através de um ensaio de DSC.
O pico presente neste termograma corresponde à temperatura de fusão, isto é, à temperatura de
transição de fase da solução analisada.
Figura 14. Exemplo de um termograma típico de uma experiência DSC (DMPC Brain Lipid), onde se mostra um
pico correspondente a uma transição de fase.
2.6.5. Dicroísmo Circular
O dicroísmo circular é uma técnica de espectroscopia que se baseia na absorção diferencial dos
componentes esquerdo e direito da luz polarizada circularmente [186, 187]. Na prática, a radiação
polarizada no plano é dividida em dois componentes polarizados circularmente (esquerdo e direito),
pela passagem de um modulador que se encontra submetido a um campo elétrico alternado; este
modulador consiste num cristal de quartzo e numa placa fina de um material isotrópico, como o quartzo,
acoplada ao cristal. O campo elétrico alternado vai, então, induzir mudanças estruturais no cristal de
quartzo que conduz à transmissão de luz polarizada circularmente na placa. O equipamento de
dicroísmo circular não recombina os dois componentes, mas deteta-os separadamente, e a diferença
de absorção das duas polarizações (dicroísmo circular) medida em função do comprimento de onda
origina o espectro de dicroísmo circular da molécula em estudo [188].
A técnica dicroísmo circular pode ser usada para analisar e determinar a estrutura secundária dos
péptidos em solução. Esta análise é realizada na região de comprimento de onda entre 180 e 250 nm
do espectro de UV, uma vez que esta corresponde à zona de absorção das ligações peptídicas. Deste
modo, o dicroísmo circular permite avaliar a contribuição de cada tipo de estrutura secundária (hélice
α, folha β, volta β e espiral aleatória) através da comparação do espectro obtido com espectros já
determinados anteriormente de péptidos semelhantes ao péptido em análise [186, 188].

22
Capítulo III – Materiais e Métodos Experimentais
3.1. Materiais
Os lípidos 1-palmitoíl-2-oleoíl-sn-glicero-3-fosfocolina (POPC) (referência: 850457P) e
esfingomielina (Cérebro SM, porcina) (SM) (referência: 860062P) foram fornecidos pela Avanti Polar
Lipids, Inc.; o colesterol (Chol) (referência: C8667), por sua vez, foi fornecido pela Sigma – Aldrich, Co.
O fragmento peptídico β-amilóide (25-35) (95% de pureza) (referência: SY130531_9) foi fornecido
pela Intavis. O tratamento deste péptido foi realizado usando o álcool 1,1,1,3,3,3-Hexafluoro-2-propanol
(HFIP) (referência: 105228) fornecido pela Sigma-Aldrich, Co.
Foram ainda utilizados outros compostos químicos fornecidos pela Sigma - Aldrich: ácido 4-(2-
hidroxietil)piperazina-1-etanossulfónico (HEPES) (referência: H7637), dimetilsulfóxido (DMSO)
(referência: D8779), docedil sulfato de sódio (SDS) (referência: L3771), clorofórmio (referência:
528730), acetato de sódio (referência: S8750). Além destes, utilizou-se Cloreto de Sódio (NaCl)
(referência: 12159.1211) fornecido pela Panreac, Hellmanex ® II (referência: 320.002) fornecido pela
Hellma Gmbh & Co KG, ácido acético (referência: 4001), 99% de pureza, fornecido pela Glacial
Reagent Grade.
Foram preparadas duas soluções tampão, usando água Milli-Q. A solução tampão de pH=4 foi
preparada adicionando ácido acético e acetato de sódio de modo a obter as concentrações finais de
1.2 M e 1 M, respetivamente. Para os ensaios de dicroísmo circular, o tampão pH=4 foi preparado com
concentrações finais de 19 mM para o ácido acético e de 13 mM para o acetato de sódio. Por sua vez,
a solução tampão de pH=7 foi preparada adicionando HEPES e NaCl de modo a obter concentrações
finais de 10 mM e 0.1 M, respetivamente.
As soluções lipídicas e as soluções peptídicas foram preparadas a pH=4 e pH=7 a partir das duas
soluções tampão.
Para filtrar os tampões de HEPES e de ácido acético/acetato de sódio, utilizaram-se filtros
Chromafil® Xtra PVDF-20/25 (referência: 729218), fornecidos pela Macherey Nagel, com diâmetro de
25 mm e tamanho de poro de 0.20 µm, e seringas esterilizadas (referência: CH010L), fornecidas pela
Chirana.
Para todas as soluções, foi utilizada água Milli-Q (resistividade de 18 MΩ.cm e pH=5.7.
Foram utilizados, no extrusor, filtros de policarbonato com 600, 200 e 100 nm de diâmetro fornecidos
pela Nuclepore, Whatman.
Os cristais de quartzo revestidos a ouro (AT – cut, 4.95MHz, 14mm de diâmetro) para as
experiências de QCM-D foram fornecidos pela Q-Sense AB.

23
3.2. Preparação dos Lipossomas
O protocolo fornecido pela Avanti Polar Lipids, Inc. foi utilizado como suporte de apoio à preparação
dos lipossomas [189].
A quantidade de lípidos necessária para obter uma solução lipídica com concentração final de 3.195
mM, foi dissolvida em clorofórmio e, posteriormente, colocada num balão de fundo redondo. Após a
secagem da mistura lipídica no balão através de um fluxo brando de azoto, obteve-se um filme lipídico
seco e uniforme. O filme lipídico foi mantido sob vácuo, durante toda a noite, de forma a eliminar todos
os vestígios orgânicos do solvente.
Posteriormente, o filme lipídico foi hidratado com 10mL de tampão de HEPES e mantido em banho
termostatizado a 65ºC, durante uma hora. Durante os primeiros 10 minutos, o balão que contém o filme
lipídico seco foi submetido a agitação manual; no restante tempo, este balão foi submetido a agitação
vórtex, de 10 em 10 minutos, de forma obter a dissolução completa do filme. Por sua vez, o banho
termostatizado foi mantido a 65ºC, de forma a assegurar uma temperatura superior, em pelo menos
10ºC, à temperatura de transição de fase das jangadas lipídicas. Diversos estudos afirmam que a
temperatura de extrusão dos lipossomas POPC/SM (1:1) deve variar entre 65 a 70ºC, baseado no facto
da transição de fase da esfingomielina variar dos 37 a 55ºC [109, 118]. Após este processo, obteve-se
uma suspensão de vesículas multilamelares grandes (LMV’s); esta suspensão foi, então, submetida a
cinco ciclos de aquecimento/arrefecimento, usando o banho termostatizado a 65ºC e azoto líquido,
respetivamente, de forma a ocorrer a disrupção das LMV’s.
As vesículas unilamelares grandes (LUV’s) foram obtidas a partir da extrusão de LMV’s. Para tal, a
suspensão passou através de filtros de policarbonato, com tamanho de poro de 600, 200 e 100 nm,
cerca de 20 vezes, sequencialmente: 600 nm de diâmetro - 5 vezes, 200 nm de diâmetro - 5 vezes, e
100 nm de diâmetro - 10 vezes. No final de todo o processo, obtiveram-se as LUV’s com uma
distribuição de tamanho a variar entre120 a 140 nm.
A solução de lipossomas foi diluída, com tampão de HEPES, para uma concentração final 1.12 mM
e foi, posteriormente, armazenada a 4/5ºC e usada no período máximo de sete dias após a sua
preparação, uma vez que os lípidos possuem uma cadeia de hidrocarbonetos com ligações de éster
sujeitas a uma hidrólise espontânea e o POPC, em particular, possui duas cadeias de hidrocarbonetos
com ligações de éster [190].
3.3. Preparação de Soluções Peptídicas
O filme seco de péptido β-amilóide (25-35) foi preparado com a concentração final de 0.849 mM:
10 mg de Aβ25-35 (90% de Aβ25-35 + 10% de água e contra iões, para garantir que a neutralidade
elétrica do péptido é mantida) foram dissolvidos em HFIP, com o objetivo de evitar a agregação
peptídica, através de um septo de borracha, num volume final de 10 mL. Uma vez que o frasco do
péptido não suportava o volume final pretendido de HFIP, adicionou-se apenas uma parte deste
volume;

24
Após o péptido Aβ25-35 estar em solução, o septo de borracha foi perfurado com uma agulha,
de forma a libertar o vácuo existente;
A solução peptídica de Aβ25-35 em HFIP foi incubada à temperatura ambiente, durante 30
minutos. A solução final é límpida e incolor;
Seguidamente, removeu-se o septo de borracha, cuidadosamente, de forma a não permitir que
este entre em contacto com o HFIP;
Adicionou-se o restante volume de HFIP, até perfazer 10 mL;
A solução foi sujeita a agitação vórtex e ultrassons, durante 10segundos, de forma a assegurar
a solubilização total do péptido;
A solução-mãe de Aβ25-35 foi distribuída por 40 tubos Eppendorf (0.250 mL de solução-mãe em
cada);
Os tubos Eppendorf foram deixados abertos, na hote, durante toda a noite, de forma a evaporar
o HFIP;
Transferiram-se, depois, os tubos Eppendorf para o forno de vácuo e deixou-se a solução
peptídica secar, durante 1 hora, sem aquecimento. Este passo garantiu a remoção total de qualquer
vestígio de HFIP ou humidade;
O filme peptídico seco formado em cada tubo Eppendorf foi armazenado a -20ºC.
Preparou-se, posteriormente, uma solução peptídica com uma concentração final peptídica de
80µM:
Retirou-se o tubo Eppendorf, contendo o filme peptídico seco, do congelador e deixou-se
arrefecer, à temperatura ambiente, durante 30 minutos;
Adicionou-se 400 µL de tampão ao tubo Eppendorf, para dissolver o filme peptídico;
O tubo Eppendorf, seguidamente, foi sujeito a 10 segundos de ultrassons e 20 minutos de
agitação vórtex;
A solução peptídica foi colocada um tubo de polipropileno, previamente lavado com solução
tampão;
Adicionou-se novamente 1000 µL de solução tampão ao tubo Eppendorf que é novamente
sujeito a 20 minutos de agitação vórtex e, seguidamente, a solução é colocada no tubo de
polipropileno. Este passo é realizado duas vezes.
Adiciona-se, ao tubo de polipropileno que contém já a solução peptídica, o volume de tampão
necessário para perfazer o volume final de 4.080mL.
A solução peptídica preparada com uma concentração final de 80 µM foi, posteriormente, diluída,
usando tampão pH=7 ou pH=4, de forma a obter as concentrações peptídicas utilizadas nas diferentes
técnicas.

25
3.4. Preparação de Soluções Lipídicas
Preparou-se soluções de lípidos puros (POPC, esfingomielina e colesterol) com concentração final
de 0.5 mM:
A quantidade de lípido necessária foi pesada num balão de 5 mL;
O volume final de 5 mL foi acertado com uma solução de clorofórmio e metanol (4:1), de forma
a dissolver os lípidos;
A solução-mãe de cada lípido foi, então, distribuída em frascos e estes foram armazenados
4/5ºC.
3.5. QCM-D
O equipamento utilizado no desenvolvimento do presente trabalho foi a Q-Sense E4 (Q-Sense AB),
equipada com 4 câmaras de medida ligadas a uma bomba de fluxo (Ismatec IPC-N4).
Antes de cada ensaio na QCM-D, os cristais de quartzo revestidos a ouro (Figura 15) foram
submetidos a dois ciclos de exposição a UV-Ozono, no equipamento PSD Series UVOzone Cleaning
& Sterilization da Novascan. Após cada ciclo de exposição a UV-Ozono, com a duração de 15 minutos,
os cristais de quartzo foram lavados com água Milli-Q e, posteriormente, secos usando um fluxo brando
de azoto. Este processo de limpeza teve o objetivo de remover possíveis contaminantes presentes na
superfície dos cristais e torná-los mais hidrofílicos.
Figura 15. Esquema das camadas dos cristais de quartzo a usar na QCM-D, com revestimento. 1) Ouro, 2)
Crómico, 3) Quartzo.
O comportamento da frequência e da dissipação para a frequência de ressonância fundamental e
3º, 5º, 7º e 9º harmónica foram monitorizadas usando o software QSoft 401. No início de cada ensaio,
verificou-se que a dissipação para cada cristal, em ar, era próxima do valor expectável (35 x 10-6).
Cada ensaio iniciou-se com a realização de uma linha de base; esta foi obtida com a passagem de
tampão de HEPES a uma velocidade 0.100 mL/min, durante cerca de 7 minutos, seguida de 10 minutos
de estabilização. Verificou-se que a dissipação obtida estava dentro do desvio de 20% da dissipação
nominal para o valor da frequência fundamental para um meio aquoso (350 x 10-6).
Todos os ensaios foram realizados a 37ºC e a uma velocidade de 0.100 mL/min. Em todos os
ensaios, após o período de estabilização posterior à linha de base, monitorizou-se a passagem de 3
mL de solução de lipossomas (POPC/SM ou POPC/SM/Chol), com uma concentração de 1.12 mM.

26
Posteriormente à estabilização da frequência e da dissipação, faz-se passar tampão de HEPES,
durante 10 minutos, para enxaguamento (rinsing) da camada de lipossomas, isto é, para remover os
lipossomas que se encontravam fracamente adsorvidos ao cristal de quartzo. Após a estabilização,
adicionou-se 3 mL de solução peptídica ao sistema, deixando-se, novamente, estabilizar a dissipação
e a frequência. Por fim, faz-se uma nova passagem com tampão de HEPES, durante 10 minutos,
removendo assim as moléculas de péptido fracamente adsorvidas ou algumas moléculas de lípido
provenientes da eventual disrupção dos lipossomas.
Depois de cada ensaio, a QCM-D é sujeita a limpeza, com água Milli-Q, solução de dodecil sulfato
de sódio (SDS), novamente água Milli-Q, solução de 2% de Hellmanex, e, por fim, água Milli-Q e ar,
para remover vestígios de solução. Os cristais de quartzo usados em cada ensaio são colocados no
devido suporte numa solução SDS em água Milli-Q e sujeitos a ultrassons durante 10 minutos,
seguindo-se dois ciclos de 10 minutos em ultrassons apenas com água Milli-Q. No fim, os cristais de
quartzo são lavados novamente com água Milli-Q e secos através de um fluxo brando de azoto.
Para cada um dos ensaios realizados, analisou-se a variação da frequência e da dissipação para
as harmónicas 3, 5, 7 e 9, para a mesma composição lipídica, temperatura e concentração peptídica.
Na modelação viscoelástica, os dados obtidos foram tratados através do software QTools 3, para o
modelo de Voigt. Assim, considerou-se um filme com uma única camada e utilizou-se as harmónicas 3
a 9. Adaptaram-se os valores de visocidade e densidade da água (0.001 Pa.s e 1000 Kg/m3,
respetivamente) para o fluído e a densidade do filme de 1060 Kg/m3 [171]. No Quadro 1 encontram-se
os valores apropriados para os parâmetros estimados na modelação viscoelástica.
Quadro 1. Condições limite para estimar a viscosidade do filme, o módulo de distorção e a espessura no software QTools 3.
Parâmetro Valor mínimo Valor máximo Steps
Viscosidade (Pa.s) 0.0005 0.01 50
Módulo de Elasticidade (Pa) 1000 1x109 50
Espessura (m) 1x10-10 1x10-6 50
Foi, então, escolhida a melhor modelação para cada concentração em estudo, tendo em
consideração os valores mais baixos de Qui-Quadrado, bem como os dados que fazem sentido físico
para o modelo em estudo.
3.6. Balança de Langmuir
Utilizou-se um sistema KSV 5000 (KSV Instruments) constituído por uma micro tina de
politetrafluoretileno (PTFE) com área de 8400 mm2, um volume de 50 mL e equipada com uma barreira
móvel de PTFE, foi utilizada para obter as isotérmicas Pressão Superficial vs. Área por Molécula (𝜋-A)
dos lípidos puros (POPC, SM e Chol), das misturas lipídicas (POPC/SM (1:1) e POPC/SM/Chol (1:1:1:)
e das misturas lipídicas em interação com o fragmento peptídico Aβ25-35, a diferentes concentrações.

27
Antes e depois de cada ensaio, procedeu-se à limpeza da microtina de PTFE, usando
dimetilsulfóxido (DMSO), etanol, água Milli-Q aquecida (≈50ºC) e água Milli-Q à temperatura ambiente.
Este procedimento de limpeza foi realizado cuidadosamente, de modo a garantir a total remoção de
resíduos de moléculas do ensaio ou vestígios de pó. A presença de possíveis contaminantes interferem
diretamente nos resultados obtidos, uma vez que se trata de um método experimental muito sensível,
sendo crucial a sua remoção. Posteriormente ao processo de limpeza, a micro tina foi preenchida por
tampão (HEPES para pH=7 ou ácido acético/acetato de sódio para pH=4). A temperatura foi mantida a
25ºC ± 1ºC. A pressão superficial foi medida usando uma placa de Wilhelmy. A ausência de
contaminantes na subfase foi verificada através da variação da pressão superficial do tampão,
comprimindo a subfase; esta variação não deve ser superior a 0.1 mN/m (𝜋 ≃ 0) antes do espalhamento
da solução lipídica ou peptídica.
O espalhamento das soluções lipídicas foi realizado usando uma microseringa. Este procedimento
deve ser realizado cuidadosamente, para garantir que os lípidos ficam na interface ar/líquido da subfase
aquosa. Posteriormente, fez-se 15 minutos de estabilização, onde ocorre a evaporação do solvente,
onde ocorre a interação das moléculas lipídicas. No caso das soluções peptídicas (0.25, 0.5, 1, 2 e 4
µM), estas são injectadas diretamente na subfase aquosa, usando uma microseringa. Neste caso, o
tempo de estabilização é de 75 minutos, onde o péptido Aβ25-35 difunde de forma homogénea na
subfase aquosa. Na interação lípido-péptido, primeiro foi injetado o péptido na subfase e, após 15
minutos de estabilização, foi realizado o espalhamento das soluções lipídicas na interface ar/líquido na
subfase aquosa. Neste caso, a evaporação da subfase decorre durante mais 60 minutos onde ocorre
a difusão do péptido de forma homogénea e a interação do mesmo com as moléculas lipídicas.
Posteriormente aos tempos de estabilização, procedeu-se à compressão lenta da monocamada a
uma velocidade de 10 mm/min monitorizando-se a variação da pressão superficial em função da área
ocupada por molécula.
No Quadro 2 são apresentadas as concentrações das misturas lipídicas utilizadas nos ensaios da
Balança de Langmuir.
Quadro 2. Concentrações usadas para lípidos e misturas lipídicas nas isotérmicas 𝝅-A.
POPC SM Chol POPC/SM (1:1) POPC/SM/Chol (1:1:1)
0.582 mM 0.659 mM 0.786 mM 0.615 mM 0.661 mM
Foi, então, possível estudar o comportamento das monocamadas lipídicas na presença e na
ausência do fragmento peptídico β-amilóide (25-35) a diferentes concentrações.
Foram realizadas sempre, no mínimo, três isotérmicas para cada lípido puro, mistura lipídica e
mistura peptídica, de forma a testar a sua reprodutibilidade, obtendo um desvio padrão inferior a 6%.

28
3.7. NTA
Os ensaios de Análise de Localização de Nanopartículas foram realizados usando um microscópio
e o software Capture, para visualização da solução em análise.
Foram preparados 2 mL de solução peptídica com concentrações finais de 1, 4 e 25 µM a pH=4 e a
pH=7 a 22ºC. O tampão de HEPES e o tampão de ácido acético/acetato de sódio usados para
preparação das soluções peptídicas foram sujeitos a centrifugação no equipamento Hermile Z323K,
usando filtros Centrifugal Filters Units, Amicon Ultra, durante 40 minutos a 20ºC e 8000 rpm.
Posteriormente, os tampões foram filtrados, duas vezes, com filtros da Chromafil® Xtra PVDF-20/25,
de tamanho de poro de 0.20 µm. Os tampões foram filtrados com o objetivo de evitar a presença de
poeiras em solução, uma vez que estas interferem diretamente com os resultados obtidos a partir da
técnica de NTA.
Injetou-se água Milli-Q através de uma seringa para verificar que não existia resíduos de outras
soluções na camara. Seguidamente, injetou-se, com cuidado para evitar a existência de bolhas de ar,
o volume suficiente de solução peptídica a analisar para preencher totalmente a camara. Realizaram-
se 15 medições para cada concentração de péptido a pH=4 e pH=7; entre cada medição, a amostra
em análise foi agitada com um pequeno toque na seringa para fazer passar a amostra de modo a evitar
que as 15 medições fossem realizadas sempre no mesmo local. As medições foram realizadas com
um nível de contraste de câmara de 8 e cada medição teve a duração de 90 segundos. Após cada
ensaio, lavou-se a camara com água Milli-Q até todos os resíduos de solução serem removidos da
mesma.
A partir do software Capture, obteve-se o gráfico da distribuição da concentração de fragmentos
presentes na solução peptídica em função do tamanho destas. Obteve-se, ainda, a quantidade de
péptido presente em cada concentração em estudo.
3.8. DSC
Realizaram-se 6 ciclos de varrimento, num intervalo de temperatura de 10ºC a 50ºC, três em
aquecimento e três em arrefecimento, a uma velocidade de 60ºC/hora, de modo a verificar a
reprodutibilidade dos resultados e a estabilização termodinâmica da amostra em análise. Antes de cada
ciclo, as amostras passam por um período de termostatização de 15 minutos. Estes parâmetros são
definidos através do software VP Viewer Experimental Parameter.
As medidas foram realizadas através do VP-DSC Micro-Calorimeter, da MicroCal®. Este
equipamento possui duas células: célula de referência e célula de amostra. A célula de referência é
preenchida com 0.5105 mL de tampão de HEPES 10 mM e a célula de amostra com 0.5105 mL das
diferentes amostras em análise. O enchimento destas células foi realizado usando uma seringa de
vidro. Após o seu enchimento, utilizou-se uma seringa de plástico de ajuste para retirar o excesso de
volume em ambas as células. A pressão na célula de referência e na célula de amostra é de 26 psi.

29
As amostras contendo lipossomas ou lipossomas e péptido β-amilóide foram preparadas
imediatamente antes da sua injeção diluindo o volume necessário de solução de lipossomas, usando o
tampão de HEPES, para uma concentração final de lipossomas de 1.12 mM. As amostras contendo
péptido β-amilóide foram preparadas com as seguintes concentrações peptídicas: 10, 25, 50 e 80 µM.
A amostra de tampão de HEPES 10 mM foi sujeita a desgaseificação, através do ThermoVac, da
MicroCal®, durante cerca de 10 minutos, de modo a minimizar a existência de bolhas de ar na amostra.
Quanto às amostras contendo lipossomas, estas não foram sujeitas a desgaseificação, uma vez que
os lipossomas são muito sensíveis e pode ocorrer a sua disrupção
No fim de cada ensaio, os termogramas obtidos foram analisados e modelados através do software
Origin 7.0, da OriginLab Corporation. A todos os termogramas obtidos foi subtraído o termograma
referente à linha de base do tampão de HEPES. Obteve-se, assim, a temperatura de transição de fase
para cada mistura de lipossomas e péptidos em estudo.
3.9. Dicroísmo Circular
Os espectros de dicroísmo circular do fragmento peptídico β-amilóide (25-35) em estudo foram
obtidos através do equipamento Applied Photophysics π*-180. Primeiro, registaram-se as linhas de
base com tampão de ácido acético/acetato de sódio 13mM, para o estudo a pH=4, e com tampão de
HEPES 10mM, para o estudo a pH=7. Realizaram-se 20 medições a comprimentos de onda entre 185
a 250 nm, para pH=4, e 195 e 250 nm, a pH=7, e espaçadas de 0.25 segundos. A diferença entre as
gamas de comprimento de onda utilizados nestes ensaios deve-se à concentração dos tampões usados
para cada pH, uma vez que estes interferem diretamente nos resultados dados pelo equipamento.
Estes ensaios foram efetuados a 21ºC, com uma largura de banda de 10 nm e um passo de 2.5 nm.
Os espectros de dicroísmo circular obtidos foram tratados através do software Pro-Data PiStar.
A solução peptídica em análise foi preparada com uma concentração final de 80 µM.
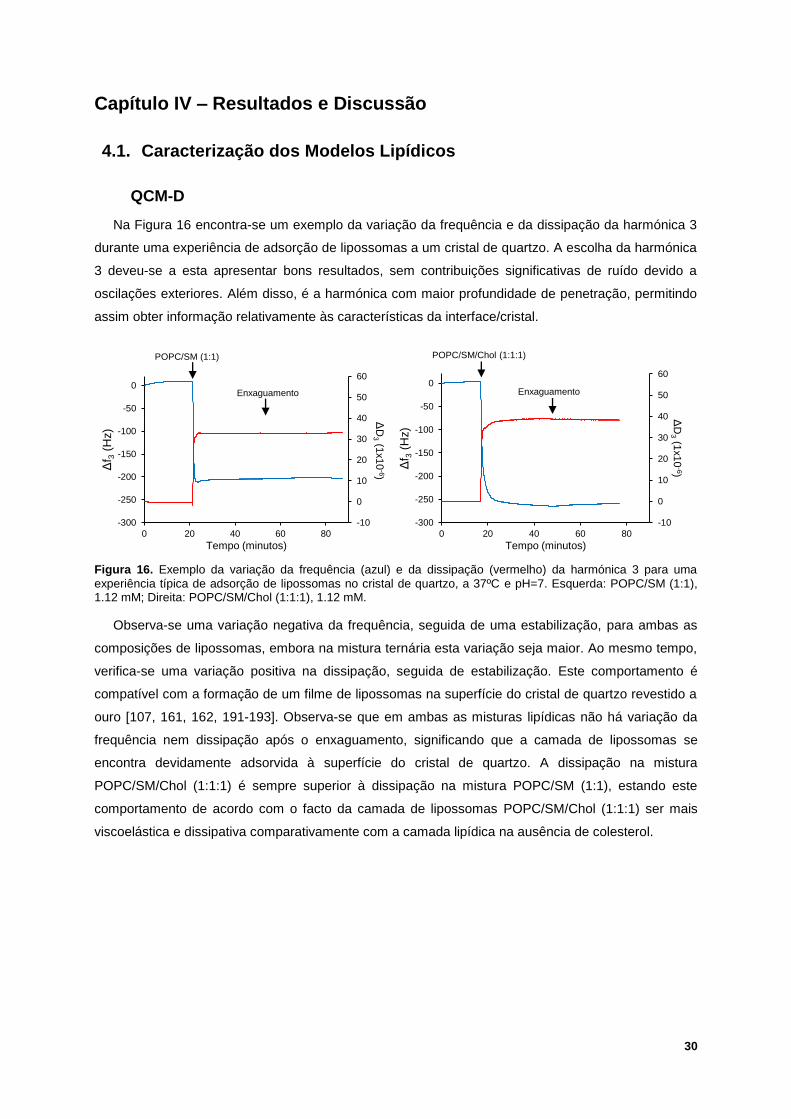
30
Capítulo IV – Resultados e Discussão
4.1. Caracterização dos Modelos Lipídicos
QCM-D
Na Figura 16 encontra-se um exemplo da variação da frequência e da dissipação da harmónica 3
durante uma experiência de adsorção de lipossomas a um cristal de quartzo. A escolha da harmónica
3 deveu-se a esta apresentar bons resultados, sem contribuições significativas de ruído devido a
oscilações exteriores. Além disso, é a harmónica com maior profundidade de penetração, permitindo
assim obter informação relativamente às características da interface/cristal.
Figura 16. Exemplo da variação da frequência (azul) e da dissipação (vermelho) da harmónica 3 para uma
experiência típica de adsorção de lipossomas no cristal de quartzo, a 37ºC e pH=7. Esquerda: POPC/SM (1:1), 1.12 mM; Direita: POPC/SM/Chol (1:1:1), 1.12 mM.
Observa-se uma variação negativa da frequência, seguida de uma estabilização, para ambas as
composições de lipossomas, embora na mistura ternária esta variação seja maior. Ao mesmo tempo,
verifica-se uma variação positiva na dissipação, seguida de estabilização. Este comportamento é
compatível com a formação de um filme de lipossomas na superfície do cristal de quartzo revestido a
ouro [107, 161, 162, 191-193]. Observa-se que em ambas as misturas lipídicas não há variação da
frequência nem dissipação após o enxaguamento, significando que a camada de lipossomas se
encontra devidamente adsorvida à superfície do cristal de quartzo. A dissipação na mistura
POPC/SM/Chol (1:1:1) é sempre superior à dissipação na mistura POPC/SM (1:1), estando este
comportamento de acordo com o facto da camada de lipossomas POPC/SM/Chol (1:1:1) ser mais
viscoelástica e dissipativa comparativamente com a camada lipídica na ausência de colesterol.
-10
0
10
20
30
40
50
60
-300
-250
-200
-150
-100
-50
0
0 20 40 60 80
ΔD
3(1
x10
-6)
Δf 3
(Hz)
Tempo (minutos)
Enxaguamento
POPC/SM (1:1)
-10
0
10
20
30
40
50
60
-300
-250
-200
-150
-100
-50
0
0 20 40 60 80
ΔD
3(1
x10
-6)Δf 3
(Hz)
Tempo (minutos)
Enxaguamento
POPC/SM/Chol (1:1:1)
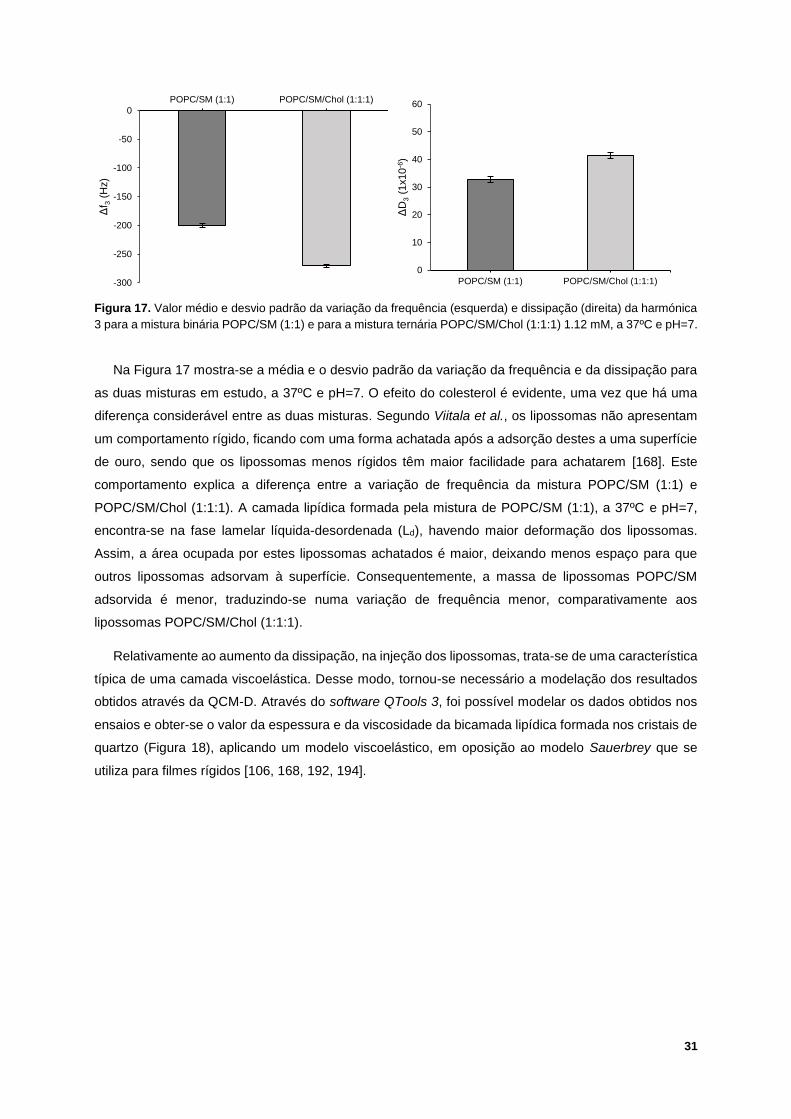
31
Figura 17. Valor médio e desvio padrão da variação da frequência (esquerda) e dissipação (direita) da harmónica
3 para a mistura binária POPC/SM (1:1) e para a mistura ternária POPC/SM/Chol (1:1:1) 1.12 mM, a 37ºC e pH=7.
Na Figura 17 mostra-se a média e o desvio padrão da variação da frequência e da dissipação para
as duas misturas em estudo, a 37ºC e pH=7. O efeito do colesterol é evidente, uma vez que há uma
diferença considerável entre as duas misturas. Segundo Viitala et al., os lipossomas não apresentam
um comportamento rígido, ficando com uma forma achatada após a adsorção destes a uma superfície
de ouro, sendo que os lipossomas menos rígidos têm maior facilidade para achatarem [168]. Este
comportamento explica a diferença entre a variação de frequência da mistura POPC/SM (1:1) e
POPC/SM/Chol (1:1:1). A camada lipídica formada pela mistura de POPC/SM (1:1), a 37ºC e pH=7,
encontra-se na fase lamelar líquida-desordenada (Ld), havendo maior deformação dos lipossomas.
Assim, a área ocupada por estes lipossomas achatados é maior, deixando menos espaço para que
outros lipossomas adsorvam à superfície. Consequentemente, a massa de lipossomas POPC/SM
adsorvida é menor, traduzindo-se numa variação de frequência menor, comparativamente aos
lipossomas POPC/SM/Chol (1:1:1).
Relativamente ao aumento da dissipação, na injeção dos lipossomas, trata-se de uma característica
típica de uma camada viscoelástica. Desse modo, tornou-se necessário a modelação dos resultados
obtidos através da QCM-D. Através do software QTools 3, foi possível modelar os dados obtidos nos
ensaios e obter-se o valor da espessura e da viscosidade da bicamada lipídica formada nos cristais de
quartzo (Figura 18), aplicando um modelo viscoelástico, em oposição ao modelo Sauerbrey que se
utiliza para filmes rígidos [106, 168, 192, 194].
-300
-250
-200
-150
-100
-50
0
POPC/SM (1:1) POPC/SM/Chol (1:1:1)Δ
f 3(H
z)
0
10
20
30
40
50
60
POPC/SM (1:1) POPC/SM/Chol (1:1:1)
ΔD
3(1
x10
-6)
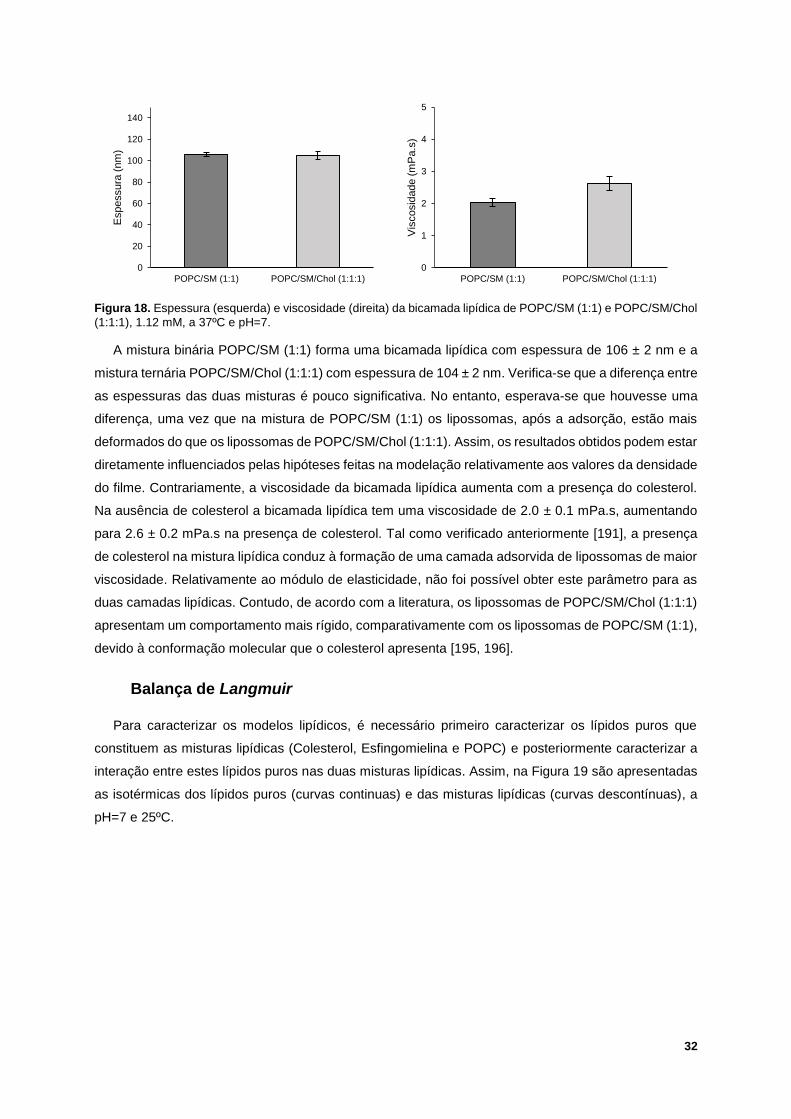
32
Figura 18. Espessura (esquerda) e viscosidade (direita) da bicamada lipídica de POPC/SM (1:1) e POPC/SM/Chol
(1:1:1), 1.12 mM, a 37ºC e pH=7.
A mistura binária POPC/SM (1:1) forma uma bicamada lipídica com espessura de 106 ± 2 nm e a
mistura ternária POPC/SM/Chol (1:1:1) com espessura de 104 ± 2 nm. Verifica-se que a diferença entre
as espessuras das duas misturas é pouco significativa. No entanto, esperava-se que houvesse uma
diferença, uma vez que na mistura de POPC/SM (1:1) os lipossomas, após a adsorção, estão mais
deformados do que os lipossomas de POPC/SM/Chol (1:1:1). Assim, os resultados obtidos podem estar
diretamente influenciados pelas hipóteses feitas na modelação relativamente aos valores da densidade
do filme. Contrariamente, a viscosidade da bicamada lipídica aumenta com a presença do colesterol.
Na ausência de colesterol a bicamada lipídica tem uma viscosidade de 2.0 ± 0.1 mPa.s, aumentando
para 2.6 ± 0.2 mPa.s na presença de colesterol. Tal como verificado anteriormente [191], a presença
de colesterol na mistura lipídica conduz à formação de uma camada adsorvida de lipossomas de maior
viscosidade. Relativamente ao módulo de elasticidade, não foi possível obter este parâmetro para as
duas camadas lipídicas. Contudo, de acordo com a literatura, os lipossomas de POPC/SM/Chol (1:1:1)
apresentam um comportamento mais rígido, comparativamente com os lipossomas de POPC/SM (1:1),
devido à conformação molecular que o colesterol apresenta [195, 196].
Balança de Langmuir
Para caracterizar os modelos lipídicos, é necessário primeiro caracterizar os lípidos puros que
constituem as misturas lipídicas (Colesterol, Esfingomielina e POPC) e posteriormente caracterizar a
interação entre estes lípidos puros nas duas misturas lipídicas. Assim, na Figura 19 são apresentadas
as isotérmicas dos lípidos puros (curvas continuas) e das misturas lipídicas (curvas descontínuas), a
pH=7 e 25ºC.
0
20
40
60
80
100
120
140
POPC/SM (1:1) POPC/SM/Chol (1:1:1)
Espessura
(nm
)
0
1
2
3
4
5
POPC/SM (1:1) POPC/SM/Chol (1:1:1)
Vis
cosid
ade (
mP
a.s
)
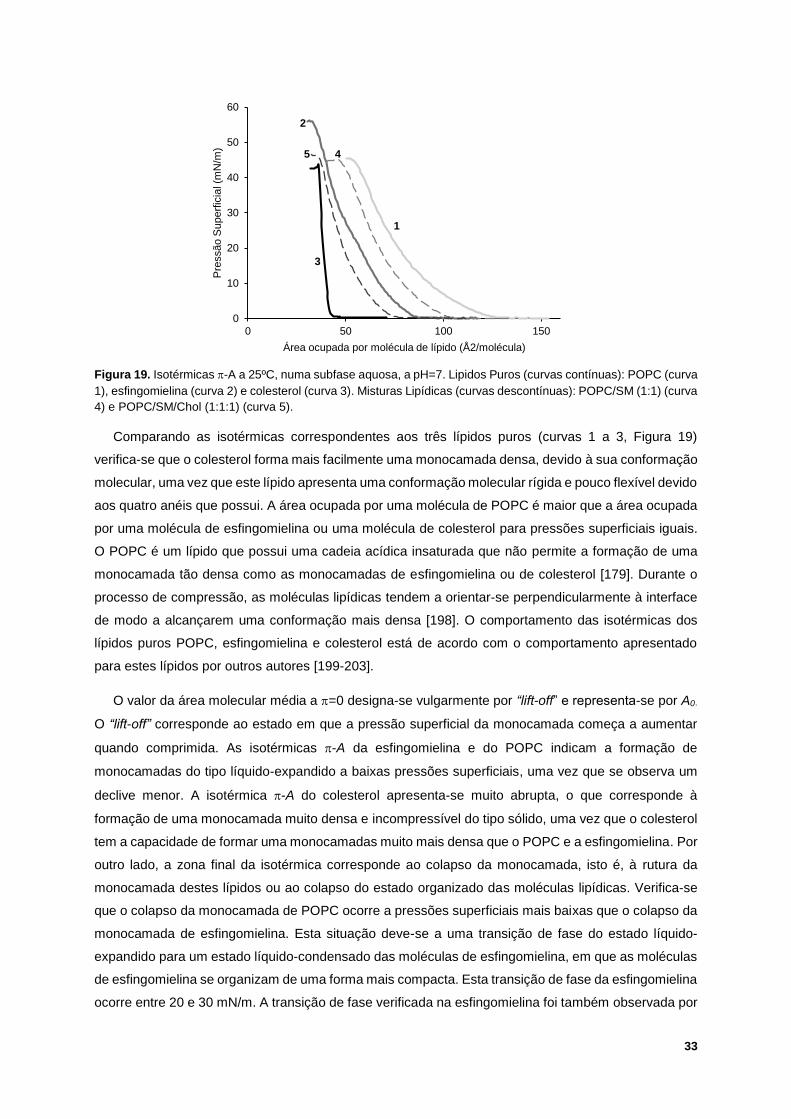
33
Figura 19. Isotérmicas -A a 25ºC, numa subfase aquosa, a pH=7. Lipidos Puros (curvas contínuas): POPC (curva
1), esfingomielina (curva 2) e colesterol (curva 3). Misturas Lipídicas (curvas descontínuas): POPC/SM (1:1) (curva
4) e POPC/SM/Chol (1:1:1) (curva 5).
Comparando as isotérmicas correspondentes aos três lípidos puros (curvas 1 a 3, Figura 19)
verifica-se que o colesterol forma mais facilmente uma monocamada densa, devido à sua conformação
molecular, uma vez que este lípido apresenta uma conformação molecular rígida e pouco flexível devido
aos quatro anéis que possui. A área ocupada por uma molécula de POPC é maior que a área ocupada
por uma molécula de esfingomielina ou uma molécula de colesterol para pressões superficiais iguais.
O POPC é um lípido que possui uma cadeia acídica insaturada que não permite a formação de uma
monocamada tão densa como as monocamadas de esfingomielina ou de colesterol [179]. Durante o
processo de compressão, as moléculas lipídicas tendem a orientar-se perpendicularmente à interface
de modo a alcançarem uma conformação mais densa [198]. O comportamento das isotérmicas dos
lípidos puros POPC, esfingomielina e colesterol está de acordo com o comportamento apresentado
para estes lípidos por outros autores [199-203].
O valor da área molecular média a =0 designa-se vulgarmente por “lift-off” e representa-se por A0.
O “lift-off” corresponde ao estado em que a pressão superficial da monocamada começa a aumentar
quando comprimida. As isotérmicas -A da esfingomielina e do POPC indicam a formação de
monocamadas do tipo líquido-expandido a baixas pressões superficiais, uma vez que se observa um
declive menor. A isotérmica -A do colesterol apresenta-se muito abrupta, o que corresponde à
formação de uma monocamada muito densa e incompressível do tipo sólido, uma vez que o colesterol
tem a capacidade de formar uma monocamadas muito mais densa que o POPC e a esfingomielina. Por
outro lado, a zona final da isotérmica corresponde ao colapso da monocamada, isto é, à rutura da
monocamada destes lípidos ou ao colapso do estado organizado das moléculas lipídicas. Verifica-se
que o colapso da monocamada de POPC ocorre a pressões superficiais mais baixas que o colapso da
monocamada de esfingomielina. Esta situação deve-se a uma transição de fase do estado líquido-
expandido para um estado líquido-condensado das moléculas de esfingomielina, em que as moléculas
de esfingomielina se organizam de uma forma mais compacta. Esta transição de fase da esfingomielina
ocorre entre 20 e 30 mN/m. A transição de fase verificada na esfingomielina foi também observada por
0
10
20
30
40
50
60
0 50 100 150
Pre
ssão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
3
2
1
45
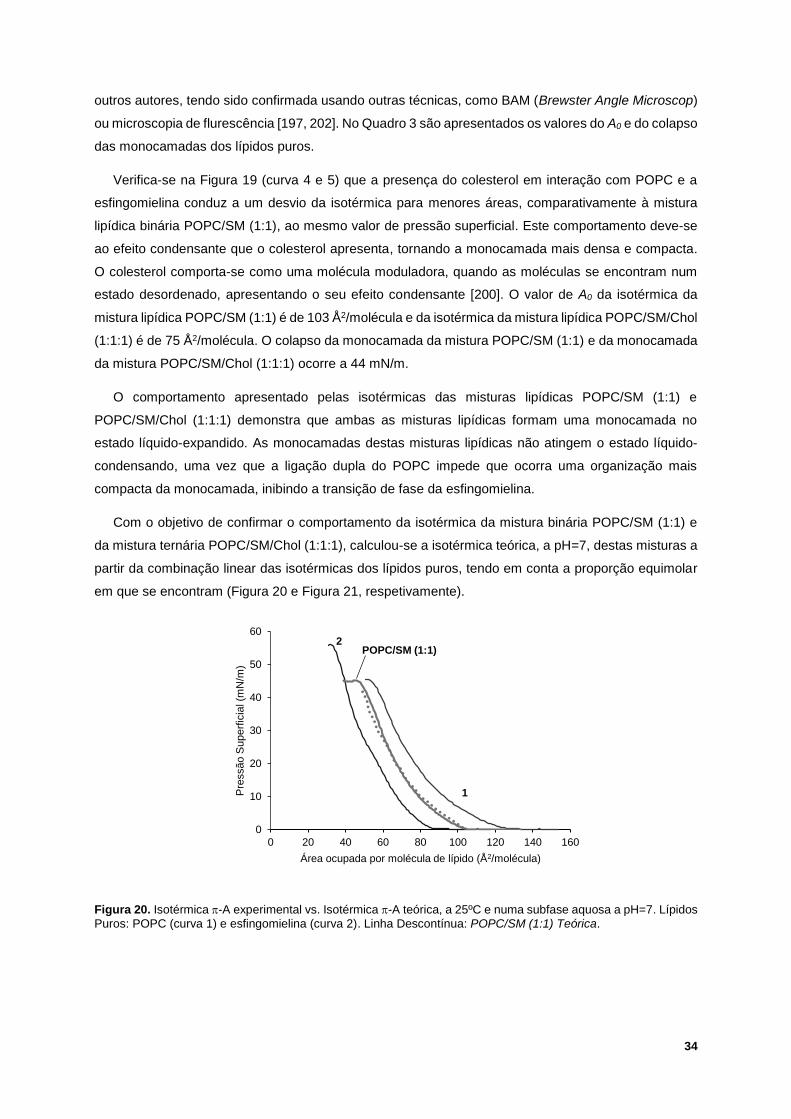
34
outros autores, tendo sido confirmada usando outras técnicas, como BAM (Brewster Angle Microscop)
ou microscopia de flurescência [197, 202]. No Quadro 3 são apresentados os valores do A0 e do colapso
das monocamadas dos lípidos puros.
Verifica-se na Figura 19 (curva 4 e 5) que a presença do colesterol em interação com POPC e a
esfingomielina conduz a um desvio da isotérmica para menores áreas, comparativamente à mistura
lipídica binária POPC/SM (1:1), ao mesmo valor de pressão superficial. Este comportamento deve-se
ao efeito condensante que o colesterol apresenta, tornando a monocamada mais densa e compacta.
O colesterol comporta-se como uma molécula moduladora, quando as moléculas se encontram num
estado desordenado, apresentando o seu efeito condensante [200]. O valor de A0 da isotérmica da
mistura lipídica POPC/SM (1:1) é de 103 Å2/molécula e da isotérmica da mistura lipídica POPC/SM/Chol
(1:1:1) é de 75 Å2/molécula. O colapso da monocamada da mistura POPC/SM (1:1) e da monocamada
da mistura POPC/SM/Chol (1:1:1) ocorre a 44 mN/m.
O comportamento apresentado pelas isotérmicas das misturas lipídicas POPC/SM (1:1) e
POPC/SM/Chol (1:1:1) demonstra que ambas as misturas lipídicas formam uma monocamada no
estado líquido-expandido. As monocamadas destas misturas lipídicas não atingem o estado líquido-
condensando, uma vez que a ligação dupla do POPC impede que ocorra uma organização mais
compacta da monocamada, inibindo a transição de fase da esfingomielina.
Com o objetivo de confirmar o comportamento da isotérmica da mistura binária POPC/SM (1:1) e
da mistura ternária POPC/SM/Chol (1:1:1), calculou-se a isotérmica teórica, a pH=7, destas misturas a
partir da combinação linear das isotérmicas dos lípidos puros, tendo em conta a proporção equimolar
em que se encontram (Figura 20 e Figura 21, respetivamente).
Figura 20. Isotérmica -A experimental vs. Isotérmica -A teórica, a 25ºC e numa subfase aquosa a pH=7. Lípidos
Puros: POPC (curva 1) e esfingomielina (curva 2). Linha Descontínua: POPC/SM (1:1) Teórica.
0
10
20
30
40
50
60
0 20 40 60 80 100 120 140 160
Pre
ssão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
2
1
POPC/SM (1:1)
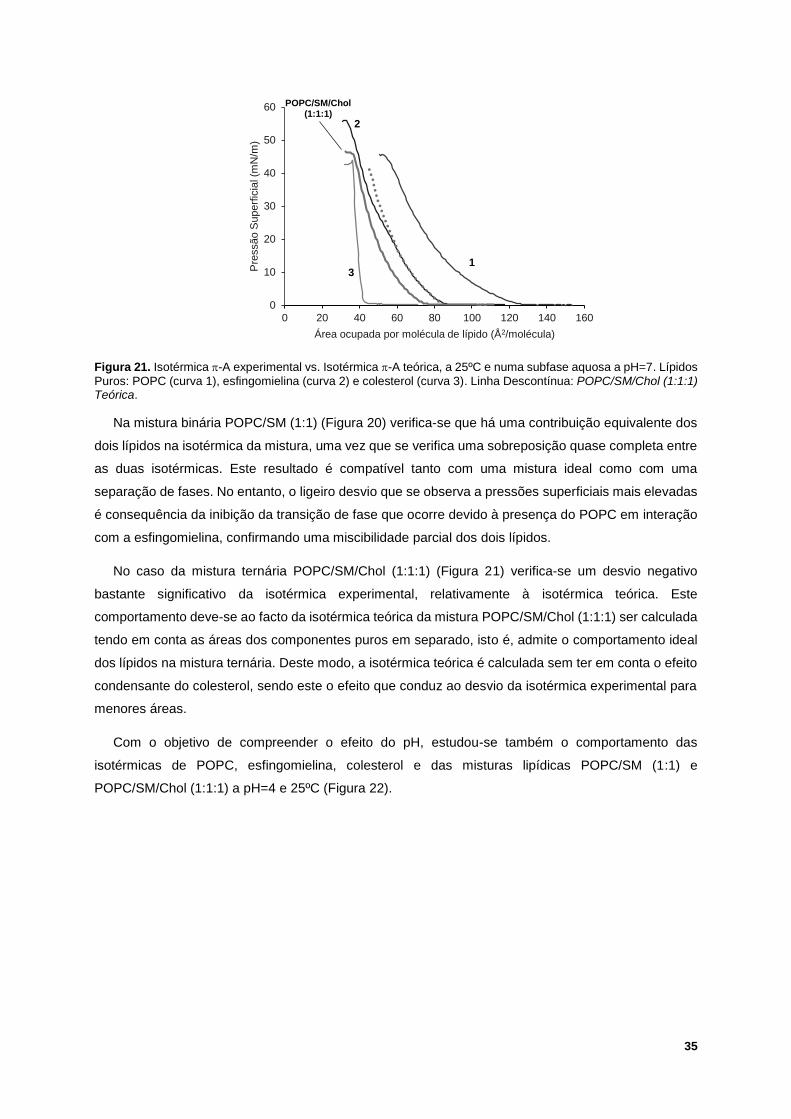
35
Figura 21. Isotérmica -A experimental vs. Isotérmica -A teórica, a 25ºC e numa subfase aquosa a pH=7. Lípidos
Puros: POPC (curva 1), esfingomielina (curva 2) e colesterol (curva 3). Linha Descontínua: POPC/SM/Chol (1:1:1) Teórica.
Na mistura binária POPC/SM (1:1) (Figura 20) verifica-se que há uma contribuição equivalente dos
dois lípidos na isotérmica da mistura, uma vez que se verifica uma sobreposição quase completa entre
as duas isotérmicas. Este resultado é compatível tanto com uma mistura ideal como com uma
separação de fases. No entanto, o ligeiro desvio que se observa a pressões superficiais mais elevadas
é consequência da inibição da transição de fase que ocorre devido à presença do POPC em interação
com a esfingomielina, confirmando uma miscibilidade parcial dos dois lípidos.
No caso da mistura ternária POPC/SM/Chol (1:1:1) (Figura 21) verifica-se um desvio negativo
bastante significativo da isotérmica experimental, relativamente à isotérmica teórica. Este
comportamento deve-se ao facto da isotérmica teórica da mistura POPC/SM/Chol (1:1:1) ser calculada
tendo em conta as áreas dos componentes puros em separado, isto é, admite o comportamento ideal
dos lípidos na mistura ternária. Deste modo, a isotérmica teórica é calculada sem ter em conta o efeito
condensante do colesterol, sendo este o efeito que conduz ao desvio da isotérmica experimental para
menores áreas.
Com o objetivo de compreender o efeito do pH, estudou-se também o comportamento das
isotérmicas de POPC, esfingomielina, colesterol e das misturas lipídicas POPC/SM (1:1) e
POPC/SM/Chol (1:1:1) a pH=4 e 25ºC (Figura 22).
0
10
20
30
40
50
60
0 20 40 60 80 100 120 140 160
Pre
ssão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
2
13
POPC/SM/Chol(1:1:1)
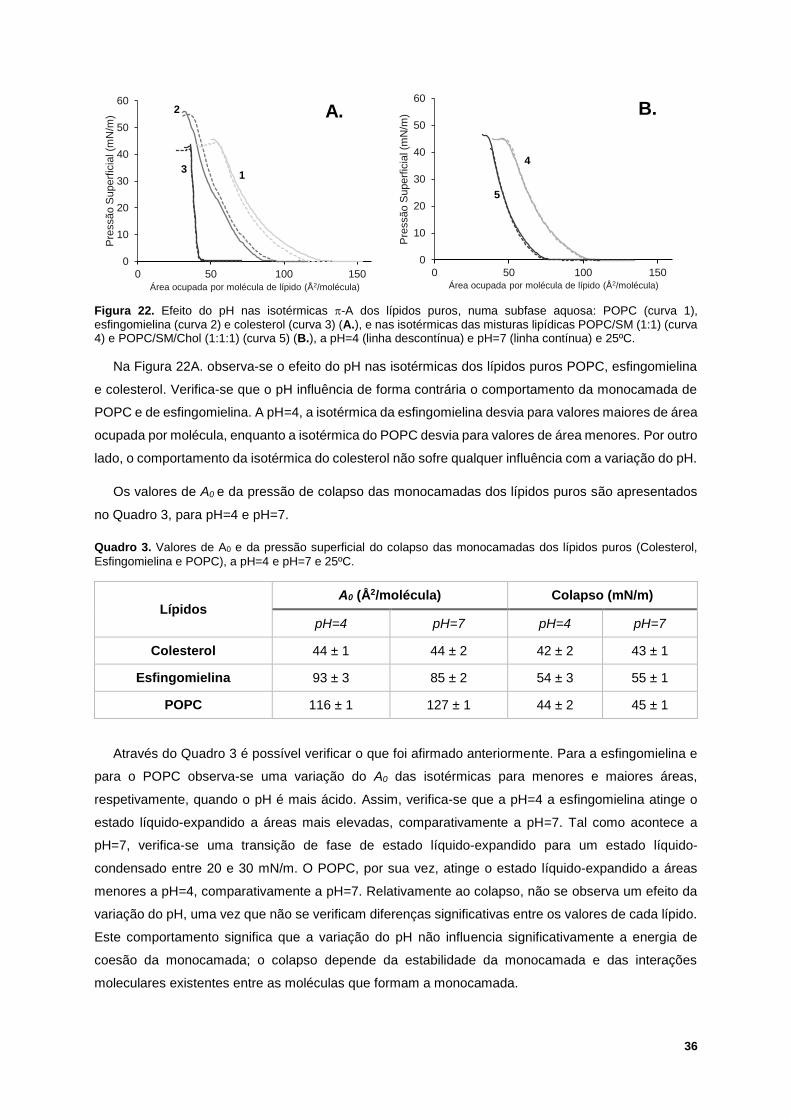
36
Figura 22. Efeito do pH nas isotérmicas -A dos lípidos puros, numa subfase aquosa: POPC (curva 1), esfingomielina (curva 2) e colesterol (curva 3) (A.), e nas isotérmicas das misturas lipídicas POPC/SM (1:1) (curva 4) e POPC/SM/Chol (1:1:1) (curva 5) (B.), a pH=4 (linha descontínua) e pH=7 (linha contínua) e 25ºC.
Na Figura 22A. observa-se o efeito do pH nas isotérmicas dos lípidos puros POPC, esfingomielina
e colesterol. Verifica-se que o pH influência de forma contrária o comportamento da monocamada de
POPC e de esfingomielina. A pH=4, a isotérmica da esfingomielina desvia para valores maiores de área
ocupada por molécula, enquanto a isotérmica do POPC desvia para valores de área menores. Por outro
lado, o comportamento da isotérmica do colesterol não sofre qualquer influência com a variação do pH.
Os valores de A0 e da pressão de colapso das monocamadas dos lípidos puros são apresentados
no Quadro 3, para pH=4 e pH=7.
Quadro 3. Valores de A0 e da pressão superficial do colapso das monocamadas dos lípidos puros (Colesterol,
Esfingomielina e POPC), a pH=4 e pH=7 e 25ºC.
Lípidos A0 (Å2/molécula) Colapso (mN/m)
pH=4 pH=7 pH=4 pH=7
Colesterol 44 ± 1 44 ± 2 42 ± 2 43 ± 1
Esfingomielina 93 ± 3 85 ± 2 54 ± 3 55 ± 1
POPC 116 ± 1 127 ± 1 44 ± 2 45 ± 1
Através do Quadro 3 é possível verificar o que foi afirmado anteriormente. Para a esfingomielina e
para o POPC observa-se uma variação do A0 das isotérmicas para menores e maiores áreas,
respetivamente, quando o pH é mais ácido. Assim, verifica-se que a pH=4 a esfingomielina atinge o
estado líquido-expandido a áreas mais elevadas, comparativamente a pH=7. Tal como acontece a
pH=7, verifica-se uma transição de fase de estado líquido-expandido para um estado líquido-
condensado entre 20 e 30 mN/m. O POPC, por sua vez, atinge o estado líquido-expandido a áreas
menores a pH=4, comparativamente a pH=7. Relativamente ao colapso, não se observa um efeito da
variação do pH, uma vez que não se verificam diferenças significativas entre os valores de cada lípido.
Este comportamento significa que a variação do pH não influencia significativamente a energia de
coesão da monocamada; o colapso depende da estabilidade da monocamada e das interações
moleculares existentes entre as moléculas que formam a monocamada.
0
10
20
30
40
50
60
0 50 100 150
Pre
ssã
o S
up
erf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
3
2
1
A.
0
10
20
30
40
50
60
0 50 100 150
Pre
ssão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
5
4
B.
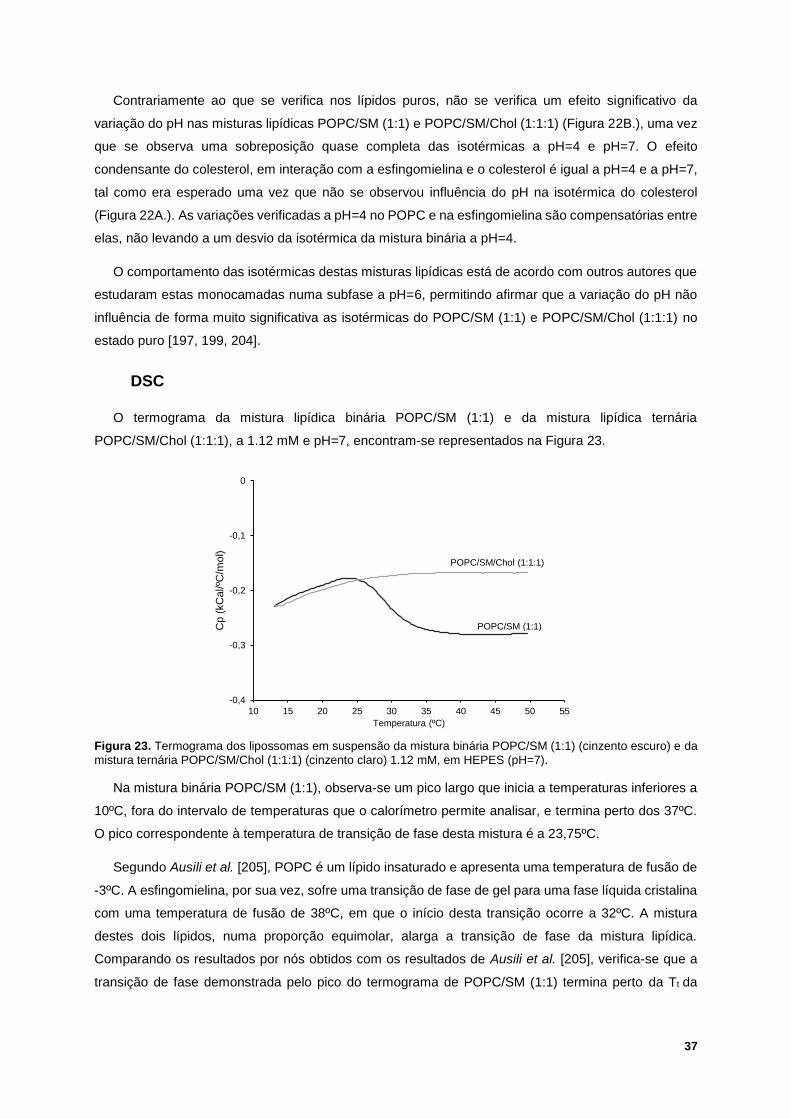
37
Contrariamente ao que se verifica nos lípidos puros, não se verifica um efeito significativo da
variação do pH nas misturas lipídicas POPC/SM (1:1) e POPC/SM/Chol (1:1:1) (Figura 22B.), uma vez
que se observa uma sobreposição quase completa das isotérmicas a pH=4 e pH=7. O efeito
condensante do colesterol, em interação com a esfingomielina e o colesterol é igual a pH=4 e a pH=7,
tal como era esperado uma vez que não se observou influência do pH na isotérmica do colesterol
(Figura 22A.). As variações verificadas a pH=4 no POPC e na esfingomielina são compensatórias entre
elas, não levando a um desvio da isotérmica da mistura binária a pH=4.
O comportamento das isotérmicas destas misturas lipídicas está de acordo com outros autores que
estudaram estas monocamadas numa subfase a pH=6, permitindo afirmar que a variação do pH não
influência de forma muito significativa as isotérmicas do POPC/SM (1:1) e POPC/SM/Chol (1:1:1) no
estado puro [197, 199, 204].
DSC
O termograma da mistura lipídica binária POPC/SM (1:1) e da mistura lipídica ternária
POPC/SM/Chol (1:1:1), a 1.12 mM e pH=7, encontram-se representados na Figura 23.
Figura 23. Termograma dos lipossomas em suspensão da mistura binária POPC/SM (1:1) (cinzento escuro) e da
mistura ternária POPC/SM/Chol (1:1:1) (cinzento claro) 1.12 mM, em HEPES (pH=7).
Na mistura binária POPC/SM (1:1), observa-se um pico largo que inicia a temperaturas inferiores a
10ºC, fora do intervalo de temperaturas que o calorímetro permite analisar, e termina perto dos 37ºC.
O pico correspondente à temperatura de transição de fase desta mistura é a 23,75ºC.
Segundo Ausili et al. [205], POPC é um lípido insaturado e apresenta uma temperatura de fusão de
-3ºC. A esfingomielina, por sua vez, sofre uma transição de fase de gel para uma fase líquida cristalina
com uma temperatura de fusão de 38ºC, em que o início desta transição ocorre a 32ºC. A mistura
destes dois lípidos, numa proporção equimolar, alarga a transição de fase da mistura lipídica.
Comparando os resultados por nós obtidos com os resultados de Ausili et al. [205], verifica-se que a
transição de fase demonstrada pelo pico do termograma de POPC/SM (1:1) termina perto da Tt da
-0,4
-0,3
-0,2
-0,1
0
10 15 20 25 30 35 40 45 50 55
Cp (
kC
al/ºC
/mol)
Temperatura (ºC)
POPC/SM/Chol (1:1:1)
POPC/SM (1:1)
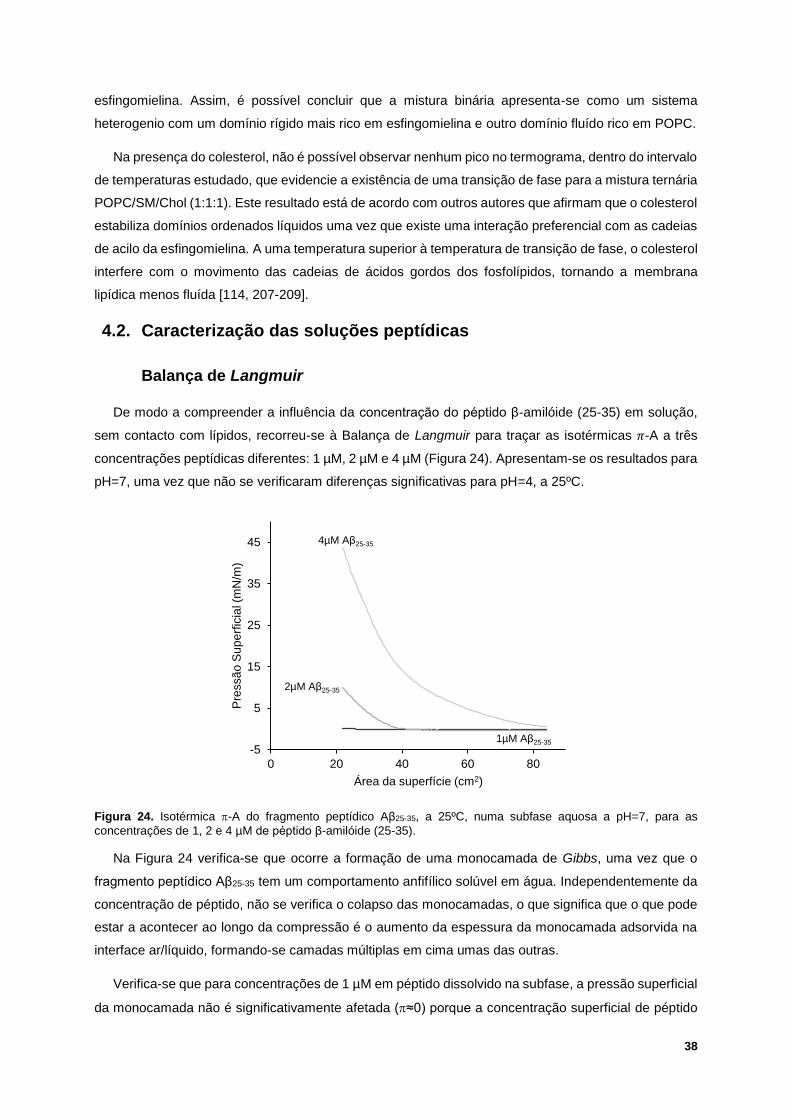
38
esfingomielina. Assim, é possível concluir que a mistura binária apresenta-se como um sistema
heterogenio com um domínio rígido mais rico em esfingomielina e outro domínio fluído rico em POPC.
Na presença do colesterol, não é possível observar nenhum pico no termograma, dentro do intervalo
de temperaturas estudado, que evidencie a existência de uma transição de fase para a mistura ternária
POPC/SM/Chol (1:1:1). Este resultado está de acordo com outros autores que afirmam que o colesterol
estabiliza domínios ordenados líquidos uma vez que existe uma interação preferencial com as cadeias
de acilo da esfingomielina. A uma temperatura superior à temperatura de transição de fase, o colesterol
interfere com o movimento das cadeias de ácidos gordos dos fosfolípidos, tornando a membrana
lipídica menos fluída [114, 207-209].
4.2. Caracterização das soluções peptídicas
Balança de Langmuir
De modo a compreender a influência da concentração do péptido β-amilóide (25-35) em solução,
sem contacto com lípidos, recorreu-se à Balança de Langmuir para traçar as isotérmicas 𝜋-A a três
concentrações peptídicas diferentes: 1 µM, 2 µM e 4 µM (Figura 24). Apresentam-se os resultados para
pH=7, uma vez que não se verificaram diferenças significativas para pH=4, a 25ºC.
Figura 24. Isotérmica -A do fragmento peptídico Aβ25-35, a 25ºC, numa subfase aquosa a pH=7, para as
concentrações de 1, 2 e 4 µM de péptido β-amilóide (25-35).
Na Figura 24 verifica-se que ocorre a formação de uma monocamada de Gibbs, uma vez que o
fragmento peptídico Aβ25-35 tem um comportamento anfifílico solúvel em água. Independentemente da
concentração de péptido, não se verifica o colapso das monocamadas, o que significa que o que pode
estar a acontecer ao longo da compressão é o aumento da espessura da monocamada adsorvida na
interface ar/líquido, formando-se camadas múltiplas em cima umas das outras.
Verifica-se que para concentrações de 1 µM em péptido dissolvido na subfase, a pressão superficial
da monocamada não é significativamente afetada (≈0) porque a concentração superficial de péptido
-5
5
15
25
35
45
0 20 40 60 80
Pre
ssã
o S
up
erf
icia
l (m
N/m
)
Área da superfície (cm2)
4µM Aβ25-35
2µM Aβ25-35
1µM Aβ25-35
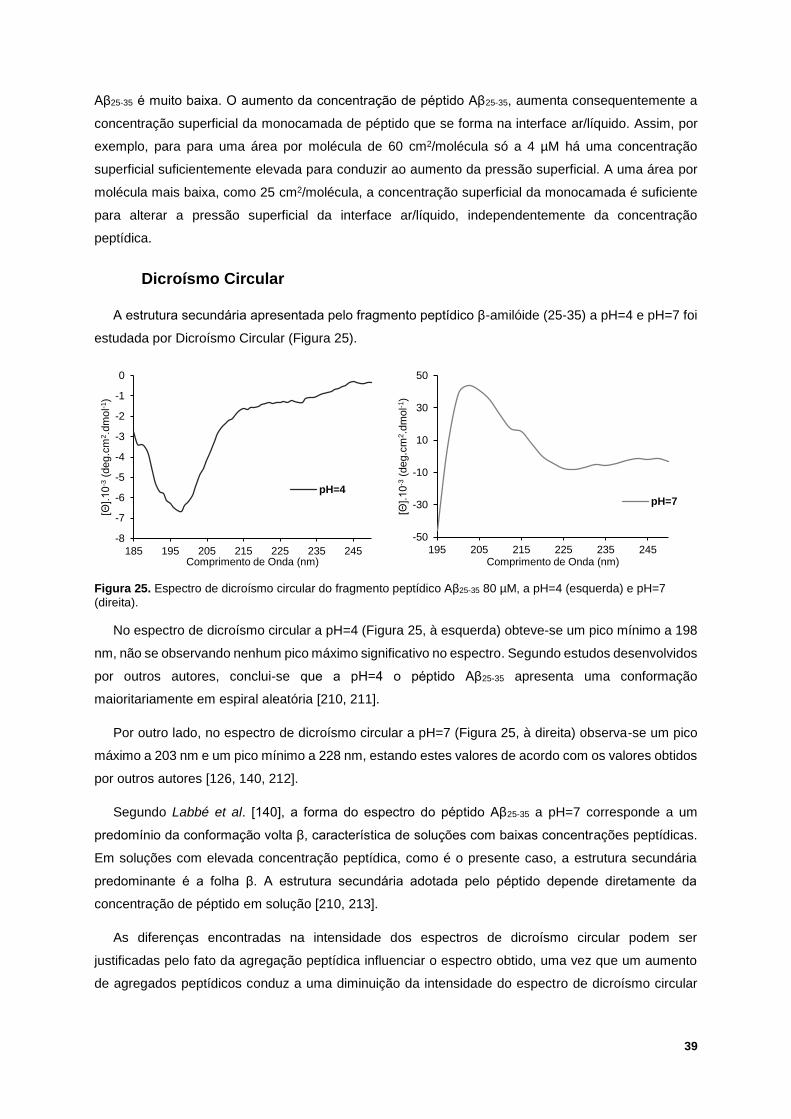
39
Aβ25-35 é muito baixa. O aumento da concentração de péptido Aβ25-35, aumenta consequentemente a
concentração superficial da monocamada de péptido que se forma na interface ar/líquido. Assim, por
exemplo, para para uma área por molécula de 60 cm2/molécula só a 4 µM há uma concentração
superficial suficientemente elevada para conduzir ao aumento da pressão superficial. A uma área por
molécula mais baixa, como 25 cm2/molécula, a concentração superficial da monocamada é suficiente
para alterar a pressão superficial da interface ar/líquido, independentemente da concentração
peptídica.
Dicroísmo Circular
A estrutura secundária apresentada pelo fragmento peptídico β-amilóide (25-35) a pH=4 e pH=7 foi
estudada por Dicroísmo Circular (Figura 25).
Figura 25. Espectro de dicroísmo circular do fragmento peptídico Aβ25-35 80 µM, a pH=4 (esquerda) e pH=7
(direita).
No espectro de dicroísmo circular a pH=4 (Figura 25, à esquerda) obteve-se um pico mínimo a 198
nm, não se observando nenhum pico máximo significativo no espectro. Segundo estudos desenvolvidos
por outros autores, conclui-se que a pH=4 o péptido Aβ25-35 apresenta uma conformação
maioritariamente em espiral aleatória [210, 211].
Por outro lado, no espectro de dicroísmo circular a pH=7 (Figura 25, à direita) observa-se um pico
máximo a 203 nm e um pico mínimo a 228 nm, estando estes valores de acordo com os valores obtidos
por outros autores [126, 140, 212].
Segundo Labbé et al. [140], a forma do espectro do péptido Aβ25-35 a pH=7 corresponde a um
predomínio da conformação volta β, característica de soluções com baixas concentrações peptídicas.
Em soluções com elevada concentração peptídica, como é o presente caso, a estrutura secundária
predominante é a folha β. A estrutura secundária adotada pelo péptido depende diretamente da
concentração de péptido em solução [210, 213].
As diferenças encontradas na intensidade dos espectros de dicroísmo circular podem ser
justificadas pelo fato da agregação peptídica influenciar o espectro obtido, uma vez que um aumento
de agregados peptídicos conduz a uma diminuição da intensidade do espectro de dicroísmo circular
-8
-7
-6
-5
-4
-3
-2
-1
0
185 195 205 215 225 235 245
[Θ].
10
-3(d
eg.c
m2.d
mo
l-1)
Comprimento de Onda (nm)
pH=4
-50
-30
-10
10
30
50
195 205 215 225 235 245
[Θ].
10
-3(d
eg.c
m2.d
mo
l-1)
Comprimento de Onda (nm)
pH=7
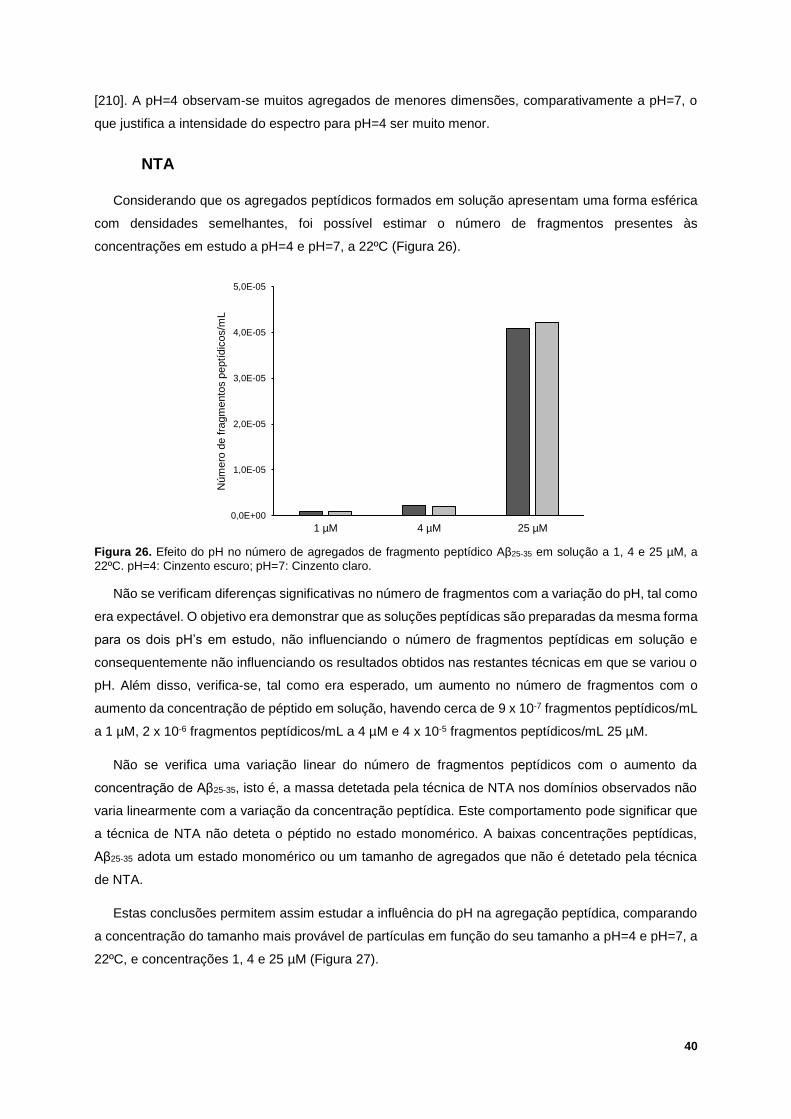
40
[210]. A pH=4 observam-se muitos agregados de menores dimensões, comparativamente a pH=7, o
que justifica a intensidade do espectro para pH=4 ser muito menor.
NTA
Considerando que os agregados peptídicos formados em solução apresentam uma forma esférica
com densidades semelhantes, foi possível estimar o número de fragmentos presentes às
concentrações em estudo a pH=4 e pH=7, a 22ºC (Figura 26).
Figura 26. Efeito do pH no número de agregados de fragmento peptídico Aβ25-35 em solução a 1, 4 e 25 µM, a
22ºC. pH=4: Cinzento escuro; pH=7: Cinzento claro.
Não se verificam diferenças significativas no número de fragmentos com a variação do pH, tal como
era expectável. O objetivo era demonstrar que as soluções peptídicas são preparadas da mesma forma
para os dois pH’s em estudo, não influenciando o número de fragmentos peptídicas em solução e
consequentemente não influenciando os resultados obtidos nas restantes técnicas em que se variou o
pH. Além disso, verifica-se, tal como era esperado, um aumento no número de fragmentos com o
aumento da concentração de péptido em solução, havendo cerca de 9 x 10-7 fragmentos peptídicos/mL
a 1 µM, 2 x 10-6 fragmentos peptídicos/mL a 4 µM e 4 x 10-5 fragmentos peptídicos/mL 25 µM.
Não se verifica uma variação linear do número de fragmentos peptídicos com o aumento da
concentração de Aβ25-35, isto é, a massa detetada pela técnica de NTA nos domínios observados não
varia linearmente com a variação da concentração peptídica. Este comportamento pode significar que
a técnica de NTA não deteta o péptido no estado monomérico. A baixas concentrações peptídicas,
Aβ25-35 adota um estado monomérico ou um tamanho de agregados que não é detetado pela técnica
de NTA.
Estas conclusões permitem assim estudar a influência do pH na agregação peptídica, comparando
a concentração do tamanho mais provável de partículas em função do seu tamanho a pH=4 e pH=7, a
22ºC, e concentrações 1, 4 e 25 µM (Figura 27).
0,0E+00
1,0E-05
2,0E-05
3,0E-05
4,0E-05
5,0E-05
1 µM 4 µM 25 µM
Núm
ero
de fra
gm
ento
s p
eptídic
os/m
L
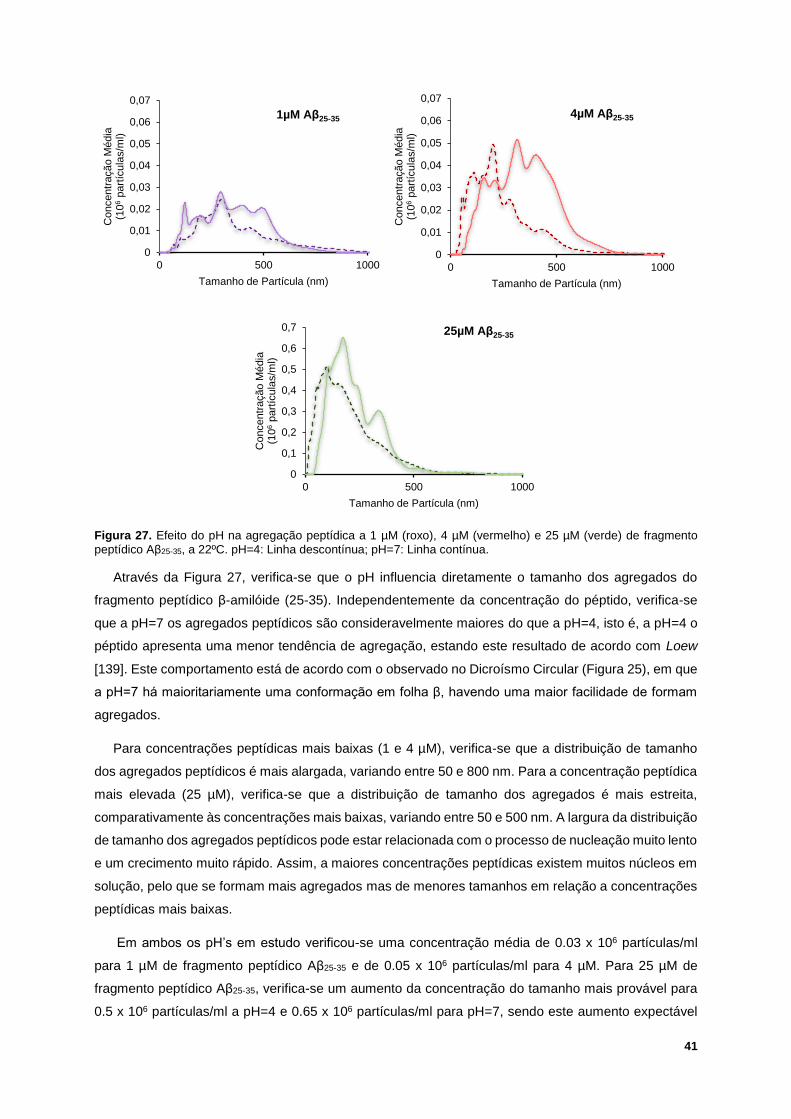
41
Figura 27. Efeito do pH na agregação peptídica a 1 µM (roxo), 4 µM (vermelho) e 25 µM (verde) de fragmento
peptídico Aβ25-35, a 22ºC. pH=4: Linha descontínua; pH=7: Linha contínua.
Através da Figura 27, verifica-se que o pH influencia diretamente o tamanho dos agregados do
fragmento peptídico β-amilóide (25-35). Independentemente da concentração do péptido, verifica-se
que a pH=7 os agregados peptídicos são consideravelmente maiores do que a pH=4, isto é, a pH=4 o
péptido apresenta uma menor tendência de agregação, estando este resultado de acordo com Loew
[139]. Este comportamento está de acordo com o observado no Dicroísmo Circular (Figura 25), em que
a pH=7 há maioritariamente uma conformação em folha β, havendo uma maior facilidade de formam
agregados.
Para concentrações peptídicas mais baixas (1 e 4 µM), verifica-se que a distribuição de tamanho
dos agregados peptídicos é mais alargada, variando entre 50 e 800 nm. Para a concentração peptídica
mais elevada (25 µM), verifica-se que a distribuição de tamanho dos agregados é mais estreita,
comparativamente às concentrações mais baixas, variando entre 50 e 500 nm. A largura da distribuição
de tamanho dos agregados peptídicos pode estar relacionada com o processo de nucleação muito lento
e um crecimento muito rápido. Assim, a maiores concentrações peptídicas existem muitos núcleos em
solução, pelo que se formam mais agregados mas de menores tamanhos em relação a concentrações
peptídicas mais baixas.
Em ambos os pH’s em estudo verificou-se uma concentração média de 0.03 x 106 partículas/ml
para 1 µM de fragmento peptídico Aβ25-35 e de 0.05 x 106 partículas/ml para 4 µM. Para 25 µM de
fragmento peptídico Aβ25-35, verifica-se um aumento da concentração do tamanho mais provável para
0.5 x 106 partículas/ml a pH=4 e 0.65 x 106 partículas/ml para pH=7, sendo este aumento expectável
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0 500 1000
Concentr
ação M
édia
(10
6part
ícula
s/m
l)
Tamanho de Partícula (nm)
1µM Aβ25-35
0
0,01
0,02
0,03
0,04
0,05
0,06
0,07
0 500 1000
Concentr
ação M
édia
(10
6part
ícula
s/m
l)
Tamanho de Partícula (nm)
4µM Aβ25-35
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0 500 1000
Concentr
ação M
édia
(10
6part
ícula
s/m
l)
Tamanho de Partícula (nm)
25µM Aβ25-35

42
em relação às outras duas concentrações em estudo. Para esta concentração existe um grande número
de partículas em solução (ver Figura 26) permitindo a formação de muitos agregados de tamanhos
distintos. Nas concentrações de 1 e 4 µM o número de partículas em solução é significativamente
menor, formando-se menos agregados em relação à concentração de 25 µM, mas de maior tamanho.
DSC
Através da Calorimetria Diferencial de Varrimento, estudou-se a temperatura de transição do
fragmento peptídico β-amilóide (25-35) a 50 µM e pH=7 (Figura 28).
Figura 28. Termograma do péptido β-amilóide (25-35) 50µM, a pH=7.
No termograma da Figura 28, não se observa nenhum pico característico de uma transição de fase
do fragmento peptídico β-amilóide (25-35). Este resultado está de acordo com o esperado a partir dos
resultados obtidos por Dicroísmo Circular, uma vez que estes demonstram que nestas condições o
péptido adota a conformação folha β, não ocorrendo uma transição de fase de hélice α para folha β.
No entanto, Chu et al. [214] afirmam que a variação da temperatura conduz a uma variação da
conformação do péptido. Através da técnica de Espectroscopia FT-IR, os autores concluíram que o
aquecimento da solução peptídica (de 25 a 120ºC) conduz à formação de estruturas em folha β. Os
autores afirmam ainda que o péptido Aβ1-40 possui propriedades térmicas irreversíveis e uma baixa
estabilidade térmica, com uma temperatura de transição de 45ºC.
4.3. Efeito do Péptido β-amilóide nas misturas lipídicas
QCM-D
Estudou-se o efeito da interação entre a mistura binária POPC/SM (1:1) e a mistura ternária
POPC/SM/Chol (1:1:1) com o fragmento peptídico β-amilóide (25-35), a pH=7 e 37ºC. A utilização
destas duas misturas lipídicas teve o objetivo de estudar também o efeito do colesterol na interação do
fragmento peptídico com os modelos biomembranares. Para tal, realizou-se a comparação da variação
da frequência e da dissipação para diferentes concentrações peptídicas, bem como a modelação dos
resultados para obter os parâmetros viscoelásticos.
0,1
0,14
0,18
0,22
0,26
10 15 20 25 30 35 40 45 50 55
Cp (
kC
al/ºC
/mol)
Temperatura (ºC)
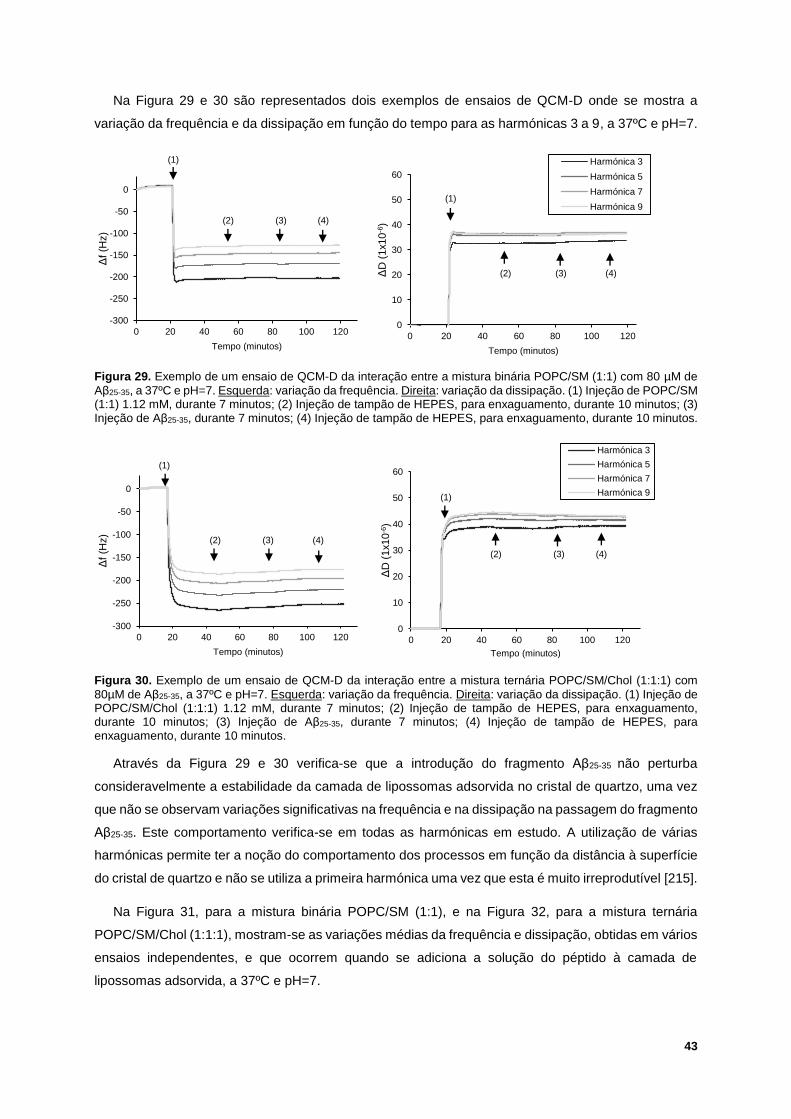
43
Na Figura 29 e 30 são representados dois exemplos de ensaios de QCM-D onde se mostra a
variação da frequência e da dissipação em função do tempo para as harmónicas 3 a 9, a 37ºC e pH=7.
Figura 29. Exemplo de um ensaio de QCM-D da interação entre a mistura binária POPC/SM (1:1) com 80 µM de
Aβ25-35, a 37ºC e pH=7. Esquerda: variação da frequência. Direita: variação da dissipação. (1) Injeção de POPC/SM (1:1) 1.12 mM, durante 7 minutos; (2) Injeção de tampão de HEPES, para enxaguamento, durante 10 minutos; (3) Injeção de Aβ25-35, durante 7 minutos; (4) Injeção de tampão de HEPES, para enxaguamento, durante 10 minutos.
Figura 30. Exemplo de um ensaio de QCM-D da interação entre a mistura ternária POPC/SM/Chol (1:1:1) com
80µM de Aβ25-35, a 37ºC e pH=7. Esquerda: variação da frequência. Direita: variação da dissipação. (1) Injeção de POPC/SM/Chol (1:1:1) 1.12 mM, durante 7 minutos; (2) Injeção de tampão de HEPES, para enxaguamento, durante 10 minutos; (3) Injeção de Aβ25-35, durante 7 minutos; (4) Injeção de tampão de HEPES, para enxaguamento, durante 10 minutos.
Através da Figura 29 e 30 verifica-se que a introdução do fragmento Aβ25-35 não perturba
consideravelmente a estabilidade da camada de lipossomas adsorvida no cristal de quartzo, uma vez
que não se observam variações significativas na frequência e na dissipação na passagem do fragmento
Aβ25-35. Este comportamento verifica-se em todas as harmónicas em estudo. A utilização de várias
harmónicas permite ter a noção do comportamento dos processos em função da distância à superfície
do cristal de quartzo e não se utiliza a primeira harmónica uma vez que esta é muito irreprodutível [215].
Na Figura 31, para a mistura binária POPC/SM (1:1), e na Figura 32, para a mistura ternária
POPC/SM/Chol (1:1:1), mostram-se as variações médias da frequência e dissipação, obtidas em vários
ensaios independentes, e que ocorrem quando se adiciona a solução do péptido à camada de
lipossomas adsorvida, a 37ºC e pH=7.
-300
-250
-200
-150
-100
-50
0
0 20 40 60 80 100 120
Δf (H
z)
Tempo (minutos)
(1)
(2) (3) (4)
0
10
20
30
40
50
60
0 20 40 60 80 100 120
ΔD
(1x10
-6)
Tempo (minutos)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9(1)
(2) (3) (4)
-300
-250
-200
-150
-100
-50
0
0 20 40 60 80 100 120
Δf (H
z)
Tempo (minutos)
(1)
(2) (3) (4)
0
10
20
30
40
50
60
0 20 40 60 80 100 120
ΔD
(1x10
-6)
Tempo (minutos)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9(1)
(2) (3) (4)
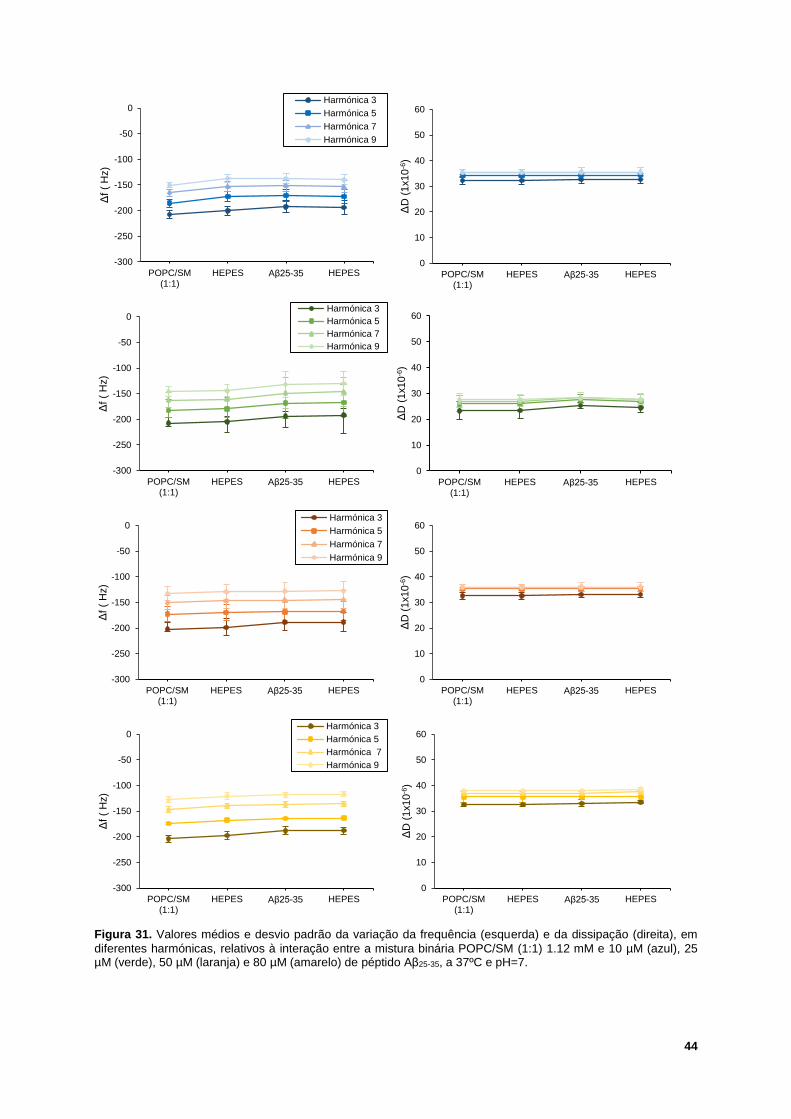
44
Figura 31. Valores médios e desvio padrão da variação da frequência (esquerda) e da dissipação (direita), em
diferentes harmónicas, relativos à interação entre a mistura binária POPC/SM (1:1) 1.12 mM e 10 µM (azul), 25 µM (verde), 50 µM (laranja) e 80 µM (amarelo) de péptido Aβ25-35, a 37ºC e pH=7.
-300
-250
-200
-150
-100
-50
0
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
-300
-250
-200
-150
-100
-50
0
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
-300
-250
-200
-150
-100
-50
0
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
-300
-250
-200
-150
-100
-50
0
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM(1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
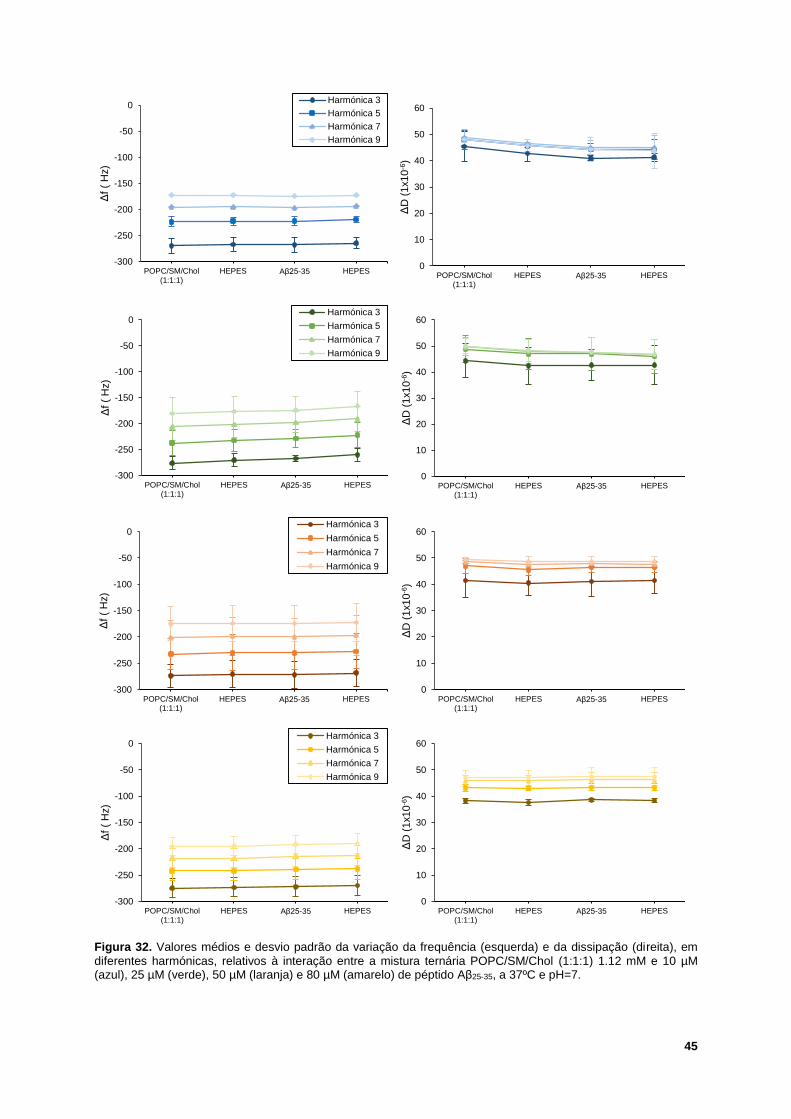
45
Figura 32. Valores médios e desvio padrão da variação da frequência (esquerda) e da dissipação (direita), em
diferentes harmónicas, relativos à interação entre a mistura ternária POPC/SM/Chol (1:1:1) 1.12 mM e 10 µM (azul), 25 µM (verde), 50 µM (laranja) e 80 µM (amarelo) de péptido Aβ25-35, a 37ºC e pH=7.
-300
-250
-200
-150
-100
-50
0
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
-300
-250
-200
-150
-100
-50
0
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
-300
-250
-200
-150
-100
-50
0
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
-300
-250
-200
-150
-100
-50
0
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
Δf (
Hz)
Harmónica 3
Harmónica 5
Harmónica 7
Harmónica 9
0
10
20
30
40
50
60
POPC/SM/Chol(1:1:1)
HEPES Aβ25-35 HEPES
ΔD
(1x10
-6)
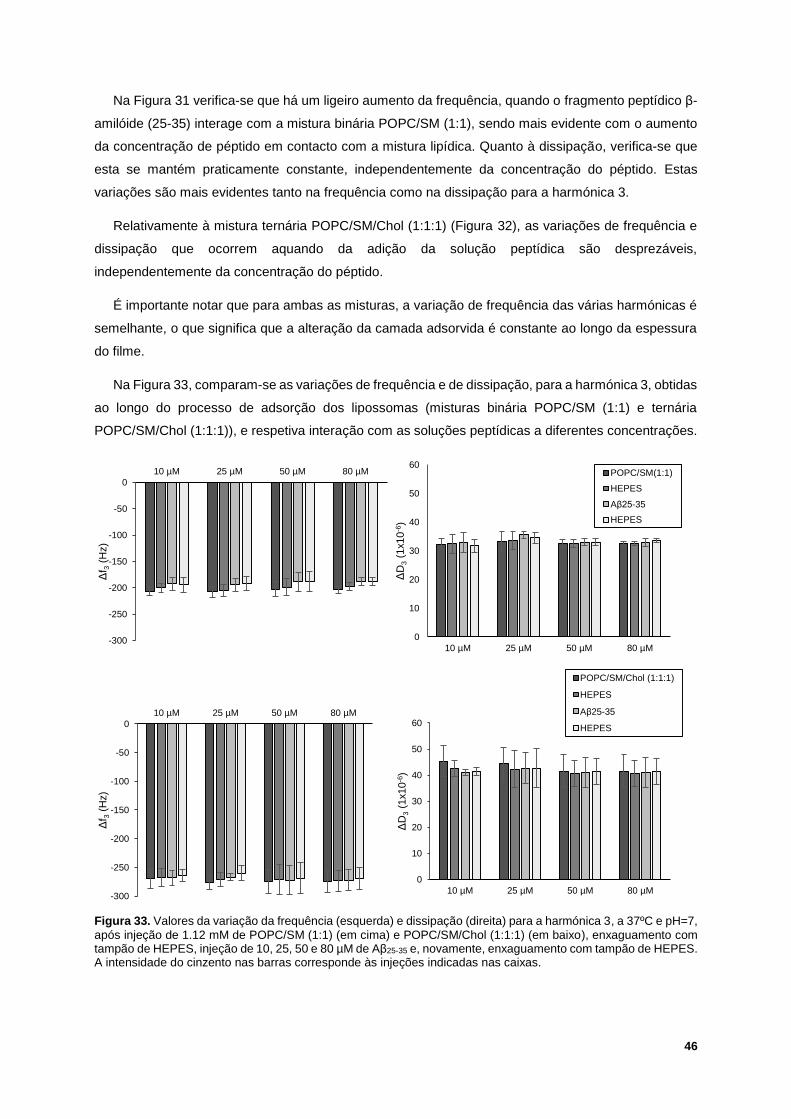
46
Na Figura 31 verifica-se que há um ligeiro aumento da frequência, quando o fragmento peptídico β-
amilóide (25-35) interage com a mistura binária POPC/SM (1:1), sendo mais evidente com o aumento
da concentração de péptido em contacto com a mistura lipídica. Quanto à dissipação, verifica-se que
esta se mantém praticamente constante, independentemente da concentração do péptido. Estas
variações são mais evidentes tanto na frequência como na dissipação para a harmónica 3.
Relativamente à mistura ternária POPC/SM/Chol (1:1:1) (Figura 32), as variações de frequência e
dissipação que ocorrem aquando da adição da solução peptídica são desprezáveis,
independentemente da concentração do péptido.
É importante notar que para ambas as misturas, a variação de frequência das várias harmónicas é
semelhante, o que significa que a alteração da camada adsorvida é constante ao longo da espessura
do filme.
Na Figura 33, comparam-se as variações de frequência e de dissipação, para a harmónica 3, obtidas
ao longo do processo de adsorção dos lipossomas (misturas binária POPC/SM (1:1) e ternária
POPC/SM/Chol (1:1:1)), e respetiva interação com as soluções peptídicas a diferentes concentrações.
Figura 33. Valores da variação da frequência (esquerda) e dissipação (direita) para a harmónica 3, a 37ºC e pH=7,
após injeção de 1.12 mM de POPC/SM (1:1) (em cima) e POPC/SM/Chol (1:1:1) (em baixo), enxaguamento com tampão de HEPES, injeção de 10, 25, 50 e 80 µM de Aβ25-35 e, novamente, enxaguamento com tampão de HEPES. A intensidade do cinzento nas barras corresponde às injeções indicadas nas caixas.
-300
-250
-200
-150
-100
-50
0
10 µM 25 µM 50 µM 80 µM
Δf 3
(Hz)
0
10
20
30
40
50
60
10 µM 25 µM 50 µM 80 µM
ΔD
3(1
x10
-6)
POPC/SM(1:1)
HEPES
Aβ25-35
HEPES
-300
-250
-200
-150
-100
-50
0
10 µM 25 µM 50 µM 80 µM
Δf 3
(Hz)
0
10
20
30
40
50
60
10 µM 25 µM 50 µM 80 µM
ΔD
3(1
x10
-6)
POPC/SM/Chol (1:1:1)
HEPES
Aβ25-35
HEPES

47
Através da Figura 33 verifica-se a existência de diferenças, embora pouco significativas, entre o
comportamento da mistura lipídica binária POPC/SM (1:1) e ternária POPC/SM/Chol (1:1:1) quando
ocorre interação com o fragmento peptídico β-amilóide (25-35). Quando o péptido Aβ25-35 interage com
os lipossomas conduz a uma ligeira diminuição da frequência em cerca de -10 Hz no caso da mistura
binária POPC/SM (1:1) + Aβ25-35. No entanto, na mistura lipídica ternária POPC/SM/Chol (1:1:1) + Aβ25-
35, a variação é praticamente nula. Verifica-se, portanto, que a interação do fragmento Aβ25-35 com a
camada de lipossomas adsorvida no cristal de quartzo revestido a ouro é mais evidente na mistura
lipídica POPC/SM (1:1). A variação positiva da frequência após a interação do péptido Aβ25-35 está de
acordo com estudos efetuados por outros autores que verificaram que, de acordo com o tipo de péptido
e a respetiva concentração, a frequência pode aumentar devido a uma rutura dos lipossomas
adsorvidos conduzindo à diminuição de massa adsorvida à superfície do cristal de quartzo revestido a
ouro ou à destabilização da camada adsorvida devido às interações dos lipossomas com os péptidos
[215-217]. Contudo, no presente caso, a diminuição de frequência é pouco significativa e não sugere
qualquer rutura de lipossomas mas sim uma ligeira fluidificação da bicamada que pode originar uma
deformação dos lipossomas adsorvidos com libertação de água intersticial.
A variação da dissipação, após interação do péptido Aβ25-35, é de cerca de 1 x 10-6 para a mistura
binária POPC/SM (1:1) e 0.5 x 10-6 para a mistura ternária POPC/SM/Chol (1:1:1), estando estas
ligeiras variações dentro do erro, podendo ser consideradas desprezáveis. Independentemente da
concentração de péptido, verifica-se que a variação da dissipação é sempre superior em valor absoluto
na mistura POPC/SM/Chol (1:1:1), mantendo as suas propriedades viscoelásticas e dissipativas
mesmo em presença de péptido Aβ25-35. Relativamente ao efeito do péptido, verifica-se um ligeiro
aumento da dissipação na interação do péptido com a mistura POPC/SM (1:1) e a mistura
POPC/SM/Chol (1:1:1), à exceção da concentração peptídica de 10 µM, onde se observa uma ligeira
diminuição da dissipação.
Os desvios de frequência e dissipação resultantes do processo de enxaguamento para ambas as
misturas lipídicas em estudo é quase nulo, significando que os lipossomas ficam na sua maioria
irreversivelmente adsorvidos à superfície do cristal de quartzo revestido a ouro [217]. A ligeira variação
que ocasionalmente se verifica após o enxaguamento é resultante da remoção de alguns lipossomas
fracamente adsorvidos, que são arrastados pela solução tampão, ou, caso seja após a adição de
péptido Aβ25-35, pode resultar na desorção de fragmentos peptídicos. A variação da frequência e da
dissipação após o enxaguamento pode também dever-se à rutura parcial dos lipossomas mais
pequenos, havendo libertação de água a partir dos lipossomas para o meio envolvente. Contudo, para
isto se verificar é necessário que ocorra uma diminuição significativa de frequência e um aumento na
dissipação, o que não se verifica neste caso.
No Quadro 4 e 5 são apresentadas as propriedades viscoelásticas para a mistura binária POPC/SM
(1:1) e mistura ternária POPC/SM/Chol (1:1:1), respetivamente, em interação com concentrações de
péptido Aβ25-35 de 10 e 80 µM, obtidas a partir da modelação dos resultados experimentais, a 37ºC e
pH=7.
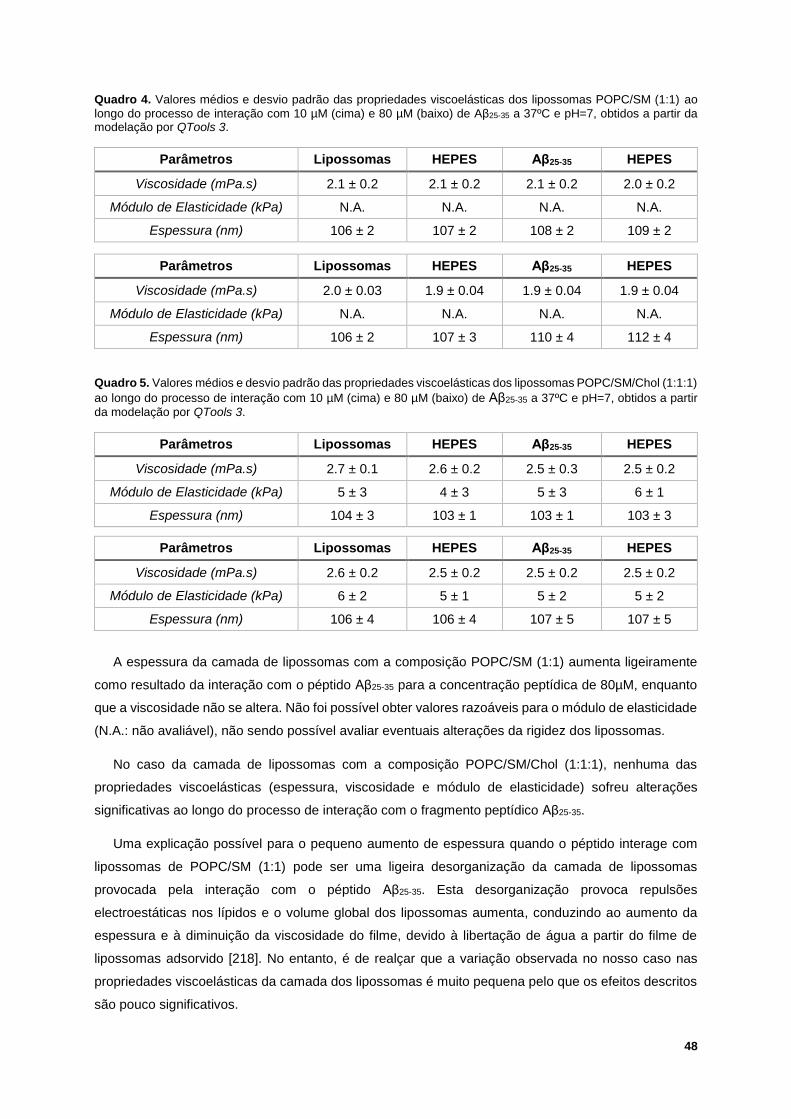
48
Quadro 4. Valores médios e desvio padrão das propriedades viscoelásticas dos lipossomas POPC/SM (1:1) ao
longo do processo de interação com 10 µM (cima) e 80 µM (baixo) de Aβ25-35 a 37ºC e pH=7, obtidos a partir da modelação por QTools 3.
Parâmetros Lipossomas HEPES Aβ25-35 HEPES
Viscosidade (mPa.s) 2.1 ± 0.2 2.1 ± 0.2 2.1 ± 0.2 2.0 ± 0.2
Módulo de Elasticidade (kPa) N.A. N.A. N.A. N.A.
Espessura (nm) 106 ± 2 107 ± 2 108 ± 2 109 ± 2
Parâmetros Lipossomas HEPES Aβ25-35 HEPES
Viscosidade (mPa.s) 2.0 ± 0.03 1.9 ± 0.04 1.9 ± 0.04 1.9 ± 0.04
Módulo de Elasticidade (kPa) N.A. N.A. N.A. N.A.
Espessura (nm) 106 ± 2 107 ± 3 110 ± 4 112 ± 4
Quadro 5. Valores médios e desvio padrão das propriedades viscoelásticas dos lipossomas POPC/SM/Chol (1:1:1)
ao longo do processo de interação com 10 µM (cima) e 80 µM (baixo) de Aβ25-35 a 37ºC e pH=7, obtidos a partir
da modelação por QTools 3.
Parâmetros Lipossomas HEPES Aβ25-35 HEPES
Viscosidade (mPa.s) 2.7 ± 0.1 2.6 ± 0.2 2.5 ± 0.3 2.5 ± 0.2
Módulo de Elasticidade (kPa) 5 ± 3 4 ± 3 5 ± 3 6 ± 1
Espessura (nm) 104 ± 3 103 ± 1 103 ± 1 103 ± 3
Parâmetros Lipossomas HEPES Aβ25-35 HEPES
Viscosidade (mPa.s) 2.6 ± 0.2 2.5 ± 0.2 2.5 ± 0.2 2.5 ± 0.2
Módulo de Elasticidade (kPa) 6 ± 2 5 ± 1 5 ± 2 5 ± 2
Espessura (nm) 106 ± 4 106 ± 4 107 ± 5 107 ± 5
A espessura da camada de lipossomas com a composição POPC/SM (1:1) aumenta ligeiramente
como resultado da interação com o péptido Aβ25-35 para a concentração peptídica de 80µM, enquanto
que a viscosidade não se altera. Não foi possível obter valores razoáveis para o módulo de elasticidade
(N.A.: não avaliável), não sendo possível avaliar eventuais alterações da rigidez dos lipossomas.
No caso da camada de lipossomas com a composição POPC/SM/Chol (1:1:1), nenhuma das
propriedades viscoelásticas (espessura, viscosidade e módulo de elasticidade) sofreu alterações
significativas ao longo do processo de interação com o fragmento peptídico Aβ25-35.
Uma explicação possível para o pequeno aumento de espessura quando o péptido interage com
lipossomas de POPC/SM (1:1) pode ser uma ligeira desorganização da camada de lipossomas
provocada pela interação com o péptido Aβ25-35. Esta desorganização provoca repulsões
electroestáticas nos lípidos e o volume global dos lipossomas aumenta, conduzindo ao aumento da
espessura e à diminuição da viscosidade do filme, devido à libertação de água a partir do filme de
lipossomas adsorvido [218]. No entanto, é de realçar que a variação observada no nosso caso nas
propriedades viscoelásticas da camada dos lipossomas é muito pequena pelo que os efeitos descritos
são pouco significativos.
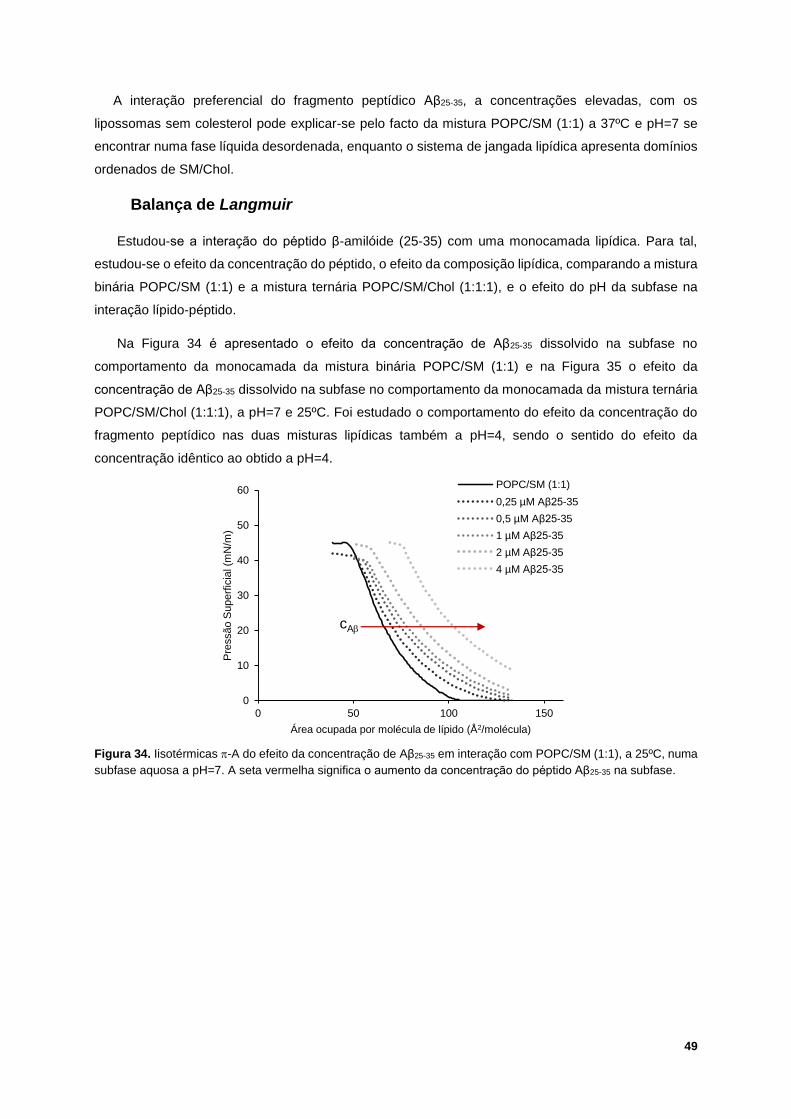
49
A interação preferencial do fragmento peptídico Aβ25-35, a concentrações elevadas, com os
lipossomas sem colesterol pode explicar-se pelo facto da mistura POPC/SM (1:1) a 37ºC e pH=7 se
encontrar numa fase líquida desordenada, enquanto o sistema de jangada lipídica apresenta domínios
ordenados de SM/Chol.
Balança de Langmuir
Estudou-se a interação do péptido β-amilóide (25-35) com uma monocamada lipídica. Para tal,
estudou-se o efeito da concentração do péptido, o efeito da composição lipídica, comparando a mistura
binária POPC/SM (1:1) e a mistura ternária POPC/SM/Chol (1:1:1), e o efeito do pH da subfase na
interação lípido-péptido.
Na Figura 34 é apresentado o efeito da concentração de Aβ25-35 dissolvido na subfase no
comportamento da monocamada da mistura binária POPC/SM (1:1) e na Figura 35 o efeito da
concentração de Aβ25-35 dissolvido na subfase no comportamento da monocamada da mistura ternária
POPC/SM/Chol (1:1:1), a pH=7 e 25ºC. Foi estudado o comportamento do efeito da concentração do
fragmento peptídico nas duas misturas lipídicas também a pH=4, sendo o sentido do efeito da
concentração idêntico ao obtido a pH=4.
Figura 34. Iisotérmicas -A do efeito da concentração de Aβ25-35 em interação com POPC/SM (1:1), a 25ºC, numa
subfase aquosa a pH=7. A seta vermelha significa o aumento da concentração do péptido Aβ25-35 na subfase.
0
10
20
30
40
50
60
0 50 100 150
Pre
ssão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
POPC/SM (1:1)
0,25 µM Aβ25-35
0,5 µM Aβ25-35
1 µM Aβ25-35
2 µM Aβ25-35
4 µM Aβ25-35
cA
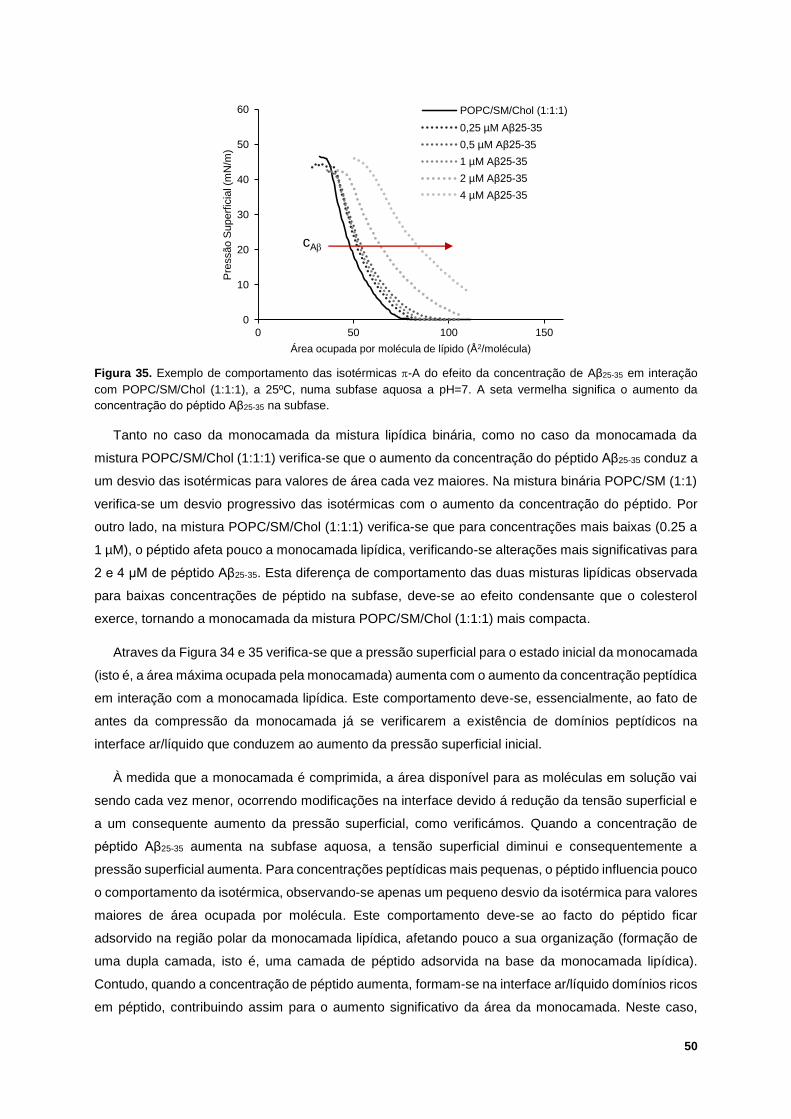
50
Figura 35. Exemplo de comportamento das isotérmicas -A do efeito da concentração de Aβ25-35 em interação
com POPC/SM/Chol (1:1:1), a 25ºC, numa subfase aquosa a pH=7. A seta vermelha significa o aumento da
concentração do péptido Aβ25-35 na subfase.
Tanto no caso da monocamada da mistura lipídica binária, como no caso da monocamada da
mistura POPC/SM/Chol (1:1:1) verifica-se que o aumento da concentração do péptido Aβ25-35 conduz a
um desvio das isotérmicas para valores de área cada vez maiores. Na mistura binária POPC/SM (1:1)
verifica-se um desvio progressivo das isotérmicas com o aumento da concentração do péptido. Por
outro lado, na mistura POPC/SM/Chol (1:1:1) verifica-se que para concentrações mais baixas (0.25 a
1 µM), o péptido afeta pouco a monocamada lipídica, verificando-se alterações mais significativas para
2 e 4 µM de péptido Aβ25-35. Esta diferença de comportamento das duas misturas lipídicas observada
para baixas concentrações de péptido na subfase, deve-se ao efeito condensante que o colesterol
exerce, tornando a monocamada da mistura POPC/SM/Chol (1:1:1) mais compacta.
Atraves da Figura 34 e 35 verifica-se que a pressão superficial para o estado inicial da monocamada
(isto é, a área máxima ocupada pela monocamada) aumenta com o aumento da concentração peptídica
em interação com a monocamada lipídica. Este comportamento deve-se, essencialmente, ao fato de
antes da compressão da monocamada já se verificarem a existência de domínios peptídicos na
interface ar/líquido que conduzem ao aumento da pressão superficial inicial.
À medida que a monocamada é comprimida, a área disponível para as moléculas em solução vai
sendo cada vez menor, ocorrendo modificações na interface devido á redução da tensão superficial e
a um consequente aumento da pressão superficial, como verificámos. Quando a concentração de
péptido Aβ25-35 aumenta na subfase aquosa, a tensão superficial diminui e consequentemente a
pressão superficial aumenta. Para concentrações peptídicas mais pequenas, o péptido influencia pouco
o comportamento da isotérmica, observando-se apenas um pequeno desvio da isotérmica para valores
maiores de área ocupada por molécula. Este comportamento deve-se ao facto do péptido ficar
adsorvido na região polar da monocamada lipídica, afetando pouco a sua organização (formação de
uma dupla camada, isto é, uma camada de péptido adsorvida na base da monocamada lipídica).
Contudo, quando a concentração de péptido aumenta, formam-se na interface ar/líquido domínios ricos
em péptido, contribuindo assim para o aumento significativo da área da monocamada. Neste caso,
0
10
20
30
40
50
60
0 50 100 150
Pre
ssão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
POPC/SM/Chol (1:1:1)
0,25 µM Aβ25-35
0,5 µM Aβ25-35
1 µM Aβ25-35
2 µM Aβ25-35
4 µM Aβ25-35
cA
51
verifica-se uma coexistência de uma monocamada de Langmuir e uma monocamada de Gibbs. O efeito
observado do péptido β-amilóide numa monocamada de lípidos está de acordo com estudos de outros
autores [20, 35].
As Figuras 36A. e Fgiura 36B. comparam as isotérmica experimentais de interação lípido-péptido
Aβ25-35, para as concentrações peptídicas de 2 µM (A) e 4 µM (B), com as curvas teóricas calculadas a
partir da soma da isotérmica da monocamada da mistura lipídica com a respectiva isotérmica de péptido
Aβ25-35 obtida na ausência da mistura lipídica.
Para a concentração de 1 µM, a curva teórica (omitida) coincide com a isotérmica da monocamada
lipídica uma vez que a contribuição independente do pépitido é aproximadamente nula (Fiurag 24).
Assim, o desvio positivo da curva experimental relativamente à previsão teórica deverá ser atribuído à
reorganização da monocamada lipídica promovida pela interação/associação lípido-péptido Aβ25-35 que
ocorre na região polar da monocamada.
Para as concentrações peptídicas de 2 µM (A) e 4 µM (B) observa-se um desvio negativo da curva
experimental relativamente à previsão teórica. Este desvio negativo pode ser devido a um maior grau
de agregação do péptido induzido pelas cadeias lipídicas que comprimem e limitam lateralmente os
domínios peptídicos incorporados na monocamada.
A razoável proximidade entre a curva teórica e curva experimental, observada para concentrações
elevadas de péptido Aβ25-35, vem confirmar a contribuição dos domínios de agregados peptídicos para
o aumento de área da monocamada.
Figura 36. Isotérmica -A experimental vs. Isotérmica -A teórica, a 25ºC e numa subfase aquosa a pH=7, da
interação entre a monocamada lipídica POPC/SM/Chol (1:1:1) com 2 μM (A.) e 4 μM (B.) de péptido Aβ25-35.
Para melhor compreender o efeito do colesterol na interação lípido-péptido Aβ25-35, apresentam-se
na Figura 36 as variações de área na monocamada lipídica, em percentagem (%), em função da
concentração de péptido Aβ25-35 na solução, observadas na mistura lipídica binária POPC/SM (1:1)
(curvas a cinzento escuro) e na mistura lipídica ternária POPC/SM/Chol (1:1:1) (curvas a cinzento
claro). A percentagem de desvio de área a pressão superficial constante é calculado a partir da equação
5:
-10
0
10
20
30
40
50
60
0 50 100 150
Pre
ssão S
uperf
icia
l (m
N/m
)
Área por molécula de lípido (Å2/molécula)
POPC/SM/Chol(1:1:1) + 2µM Aβ25-35
(Teórico)
POPC/SM/Chol(1:1:1) + 2µM Aβ25-35
(Experimental)
POPC/SM/Chol(1:1:1)
2µM Aβ25-35
A.
-10
0
10
20
30
40
50
60
0 50 100 150
Pre
ssão S
uperf
icia
l (m
N/m
)
Área por molécula de lípido (Å2/molécula)
POPC/SM/Chol(1:1:1) + 4µM Aβ25-35
(Experimental)
POPC/SM/Chol(1:1:1) + 4µM Aβ25-35
(Teórico)
4µM Aβ25-35
POPC/SM/Chol(1:1:1)
B.
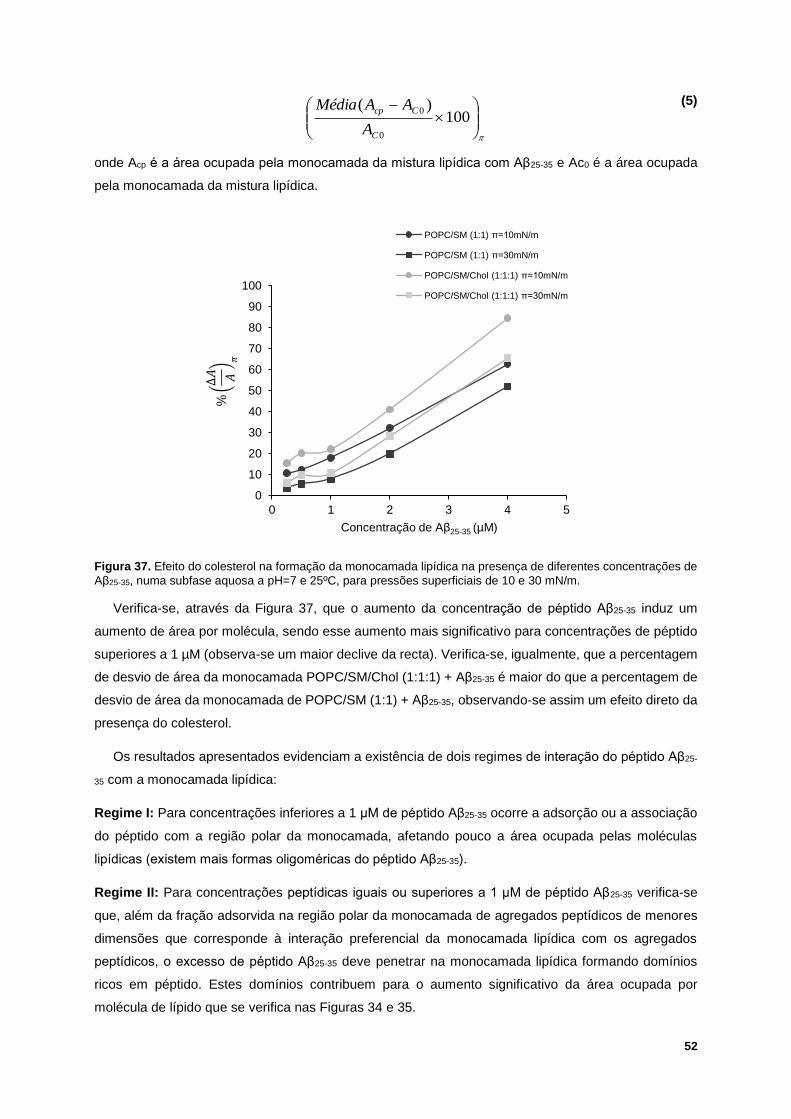
52
(5)
onde Acp é a área ocupada pela monocamada da mistura lipídica com Aβ25-35 e Ac0 é a área ocupada
pela monocamada da mistura lipídica.
Figura 37. Efeito do colesterol na formação da monocamada lipídica na presença de diferentes concentrações de
Aβ25-35, numa subfase aquosa a pH=7 e 25ºC, para pressões superficiais de 10 e 30 mN/m.
Verifica-se, através da Figura 37, que o aumento da concentração de péptido Aβ25-35 induz um
aumento de área por molécula, sendo esse aumento mais significativo para concentrações de péptido
superiores a 1 µM (observa-se um maior declive da recta). Verifica-se, igualmente, que a percentagem
de desvio de área da monocamada POPC/SM/Chol (1:1:1) + Aβ25-35 é maior do que a percentagem de
desvio de área da monocamada de POPC/SM (1:1) + Aβ25-35, observando-se assim um efeito direto da
presença do colesterol.
Os resultados apresentados evidenciam a existência de dois regimes de interação do péptido Aβ25-
35 com a monocamada lipídica:
Regime I: Para concentrações inferiores a 1 µM de péptido Aβ25-35 ocorre a adsorção ou a associação
do péptido com a região polar da monocamada, afetando pouco a área ocupada pelas moléculas
lipídicas (existem mais formas oligoméricas do péptido Aβ25-35).
Regime II: Para concentrações peptídicas iguais ou superiores a 1 µM de péptido Aβ25-35 verifica-se
que, além da fração adsorvida na região polar da monocamada de agregados peptídicos de menores
dimensões que corresponde à interação preferencial da monocamada lipídica com os agregados
peptídicos, o excesso de péptido Aβ25-35 deve penetrar na monocamada lipídica formando domínios
ricos em péptido. Estes domínios contribuem para o aumento significativo da área ocupada por
molécula de lípido que se verifica nas Figuras 34 e 35.
0
10
20
30
40
50
60
70
80
90
100
0 1 2 3 4 5
Concentração de Aβ25-35 (µM)
POPC/SM (1:1) π=10mN/m
POPC/SM (1:1) π=30mN/m
POPC/SM/Chol (1:1:1) π=10mN/m
POPC/SM/Chol (1:1:1) π=30mN/m
% ∆
𝐴 𝐴 𝜋
100
)(
0
0
C
Ccp
A
AAMédia

53
A definição destes dois regimes está de acordo com os resultados obtidos através da técnica de
NTA (Figura 26 e 27), em que se verificou-se que a baixas concentrações de péptido Aβ25-25 existem
poucos agregados peptídicos e que o número de agregados peptídicos de grandes dimensões aumenta
significativamente com o aumento da concentração de péptido.
Estes resultados mostram uma boa concordância com os modelos biomembranares computacionais
propostos Vestergaard et al. [154] para a interacção do péptido Aβ1-42 combiomembranas. Estes autores
concluíram que as espécies oligoméricas do péptido se localizam mais perto da superfície da
membrana enquanto as fibrilas têm uma localização variada. Demonstraram que a presença de péptido
Aβ1-42 conduz a um aumento da área da superfície da membrana, induzindo a transformação vesicular
dos lípidos, devido aos péptidos mediarem a fusão da membrana. Ding et al. [46] chegaram a
conclusões semelhantes ao estudar as interações do péptido Aβ1-40 com bicamadas lipídicas
suportadas. Os autores concluíram que estas interações se iniciam com uma ligação muito forte do
péptido no estado monomérico à membrana e que, posteriormente a um período de incubação de 20
horas a 22ºC ocorre a agregação peptídica com formação de oligómeros na membrana. O tamanho
destes oligómeros depende diretamente da concentração de péptido em solução.
A presença do colesterol na monocamada induz uma maior percentagem de desvio de área, uma
vez que a presença de domínios ricos em colesterol promove a interação da monocamada com o
péptido Aβ25-35. Comparativamente à monocamada da mistura lipídica POPC/SM (1:1), a monocamada
da mistura POPC/SM/Chol (1:1:1) apresenta mais domínios condensados, devido ao efeito que a
presença do colesterol induz na monocamada verificando-se assim que a interação do péptido Aβ25-35
é mais forte com domínios condensados do que com domínios expandidos. Este comportamento deve-
se ao facto dos domínios condensados apresentarem maior densidade de grupos polares disponíveis
para interagir com o péptido Aβ25-35.
Concluiu-se assim que a interação do péptido Aβ25-35 com a monocamada lipídica é promovida pela
presença do colesterol, tal como se verifica na Figura 34, 35 e 37. Este comportamento foi igualmente
verificado para pH=4.
A percentagem de desvio de área é maior à pressão superficial de 10 mN/m. Isto deve-se ao facto
das moléculas estarem na fase líquida expandida, estando mais desorganizadas, enquanto a 30 mN/m
já se encontram num estado mais organizad. Este comportamento verifica-se independentemente da
concentração de péptido em solução.
Segundo Abroggio et al., a interação do péptido β-amilóide com os lípidos conduz a uma
destabilização da monocamada de jangada lipídica [199]. Este comportamento verifica-se nos nossos
resultados obtidos com a balança de Langmuir, ao contrário dos ensaios de QCM-D onde a interação
do péptido com os lipossomas de composição POPC/SM/Chol (1:1:1) não provoca qualquer alteração
da bicamada (ver Figura 33).
Estudou-se, ainda, o efeito da variação do pH da subfase na interação das misturas lipídicas
POPC/SM (1:1) e POPC/SM/Chol (1:1:1) com 0.25, 0.5, 1, 2 e 4 µM de péptido β-amilóide (25-35). Na
Figura 38 apresentam-se as isotérmicas obtidas apenas com as concentrações peptídicas de 1 e 4 µM.
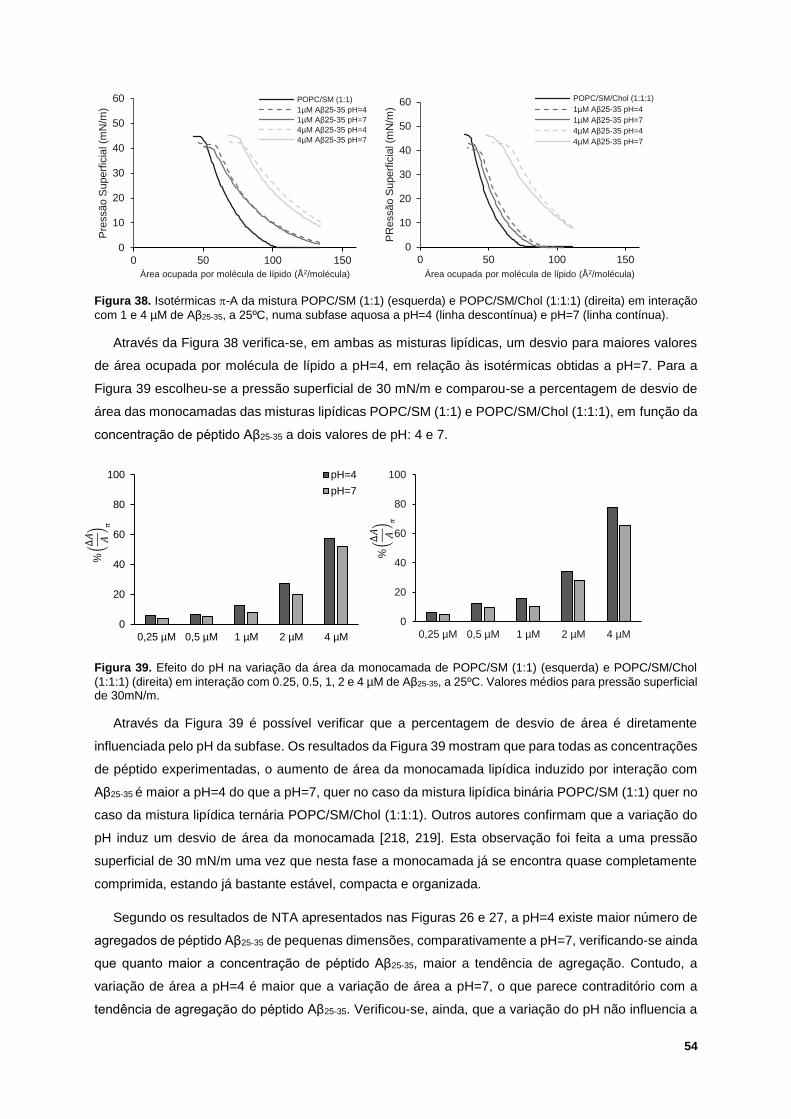
54
Figura 38. Isotérmicas -A da mistura POPC/SM (1:1) (esquerda) e POPC/SM/Chol (1:1:1) (direita) em interação
com 1 e 4 µM de Aβ25-35, a 25ºC, numa subfase aquosa a pH=4 (linha descontínua) e pH=7 (linha contínua).
Através da Figura 38 verifica-se, em ambas as misturas lipídicas, um desvio para maiores valores
de área ocupada por molécula de lípido a pH=4, em relação às isotérmicas obtidas a pH=7. Para a
Figura 39 escolheu-se a pressão superficial de 30 mN/m e comparou-se a percentagem de desvio de
área das monocamadas das misturas lipídicas POPC/SM (1:1) e POPC/SM/Chol (1:1:1), em função da
concentração de péptido Aβ25-35 a dois valores de pH: 4 e 7.
Figura 39. Efeito do pH na variação da área da monocamada de POPC/SM (1:1) (esquerda) e POPC/SM/Chol
(1:1:1) (direita) em interação com 0.25, 0.5, 1, 2 e 4 µM de Aβ25-35, a 25ºC. Valores médios para pressão superficial de 30mN/m.
Através da Figura 39 é possível verificar que a percentagem de desvio de área é diretamente
influenciada pelo pH da subfase. Os resultados da Figura 39 mostram que para todas as concentrações
de péptido experimentadas, o aumento de área da monocamada lipídica induzido por interação com
Aβ25-35 é maior a pH=4 do que a pH=7, quer no caso da mistura lipídica binária POPC/SM (1:1) quer no
caso da mistura lipídica ternária POPC/SM/Chol (1:1:1). Outros autores confirmam que a variação do
pH induz um desvio de área da monocamada [218, 219]. Esta observação foi feita a uma pressão
superficial de 30 mN/m uma vez que nesta fase a monocamada já se encontra quase completamente
comprimida, estando já bastante estável, compacta e organizada.
Segundo os resultados de NTA apresentados nas Figuras 26 e 27, a pH=4 existe maior número de
agregados de péptido Aβ25-35 de pequenas dimensões, comparativamente a pH=7, verificando-se ainda
que quanto maior a concentração de péptido Aβ25-35, maior a tendência de agregação. Contudo, a
variação de área a pH=4 é maior que a variação de área a pH=7, o que parece contraditório com a
tendência de agregação do péptido Aβ25-35. Verificou-se, ainda, que a variação do pH não influencia a
0
10
20
30
40
50
60
0 50 100 150
Pre
ssão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
POPC/SM (1:1)
1µM Aβ25-35 pH=4
1µM Aβ25-35 pH=7
4µM Aβ25-35 pH=4
4µM Aβ25-35 pH=7
0
10
20
30
40
50
60
0 50 100 150
PR
essão S
uperf
icia
l (m
N/m
)
Área ocupada por molécula de lípido (Å2/molécula)
POPC/SM/Chol (1:1:1)
1µM Aβ25-35 pH=4
1µM Aβ25-35 pH=7
4µM Aβ25-35 pH=4
4µM Aβ25-35 pH=7
0
20
40
60
80
100
0,25 µM 0,5 µM 1 µM 2 µM 4 µM
pH=4
pH=7
0
20
40
60
80
100
0,25 µM 0,5 µM 1 µM 2 µM 4 µM
% ∆
𝐴 𝐴 𝜋
% ∆
𝐴 𝐴 𝜋
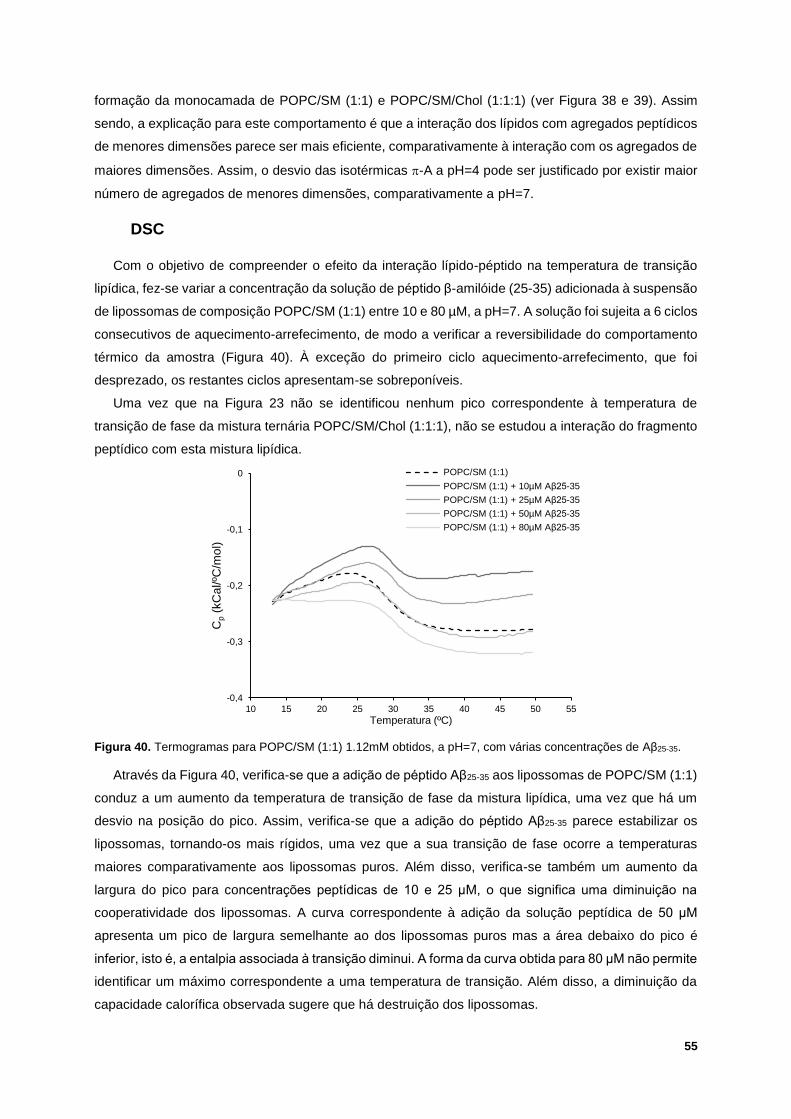
55
formação da monocamada de POPC/SM (1:1) e POPC/SM/Chol (1:1:1) (ver Figura 38 e 39). Assim
sendo, a explicação para este comportamento é que a interação dos lípidos com agregados peptídicos
de menores dimensões parece ser mais eficiente, comparativamente à interação com os agregados de
maiores dimensões. Assim, o desvio das isotérmicas -A a pH=4 pode ser justificado por existir maior
número de agregados de menores dimensões, comparativamente a pH=7.
DSC
Com o objetivo de compreender o efeito da interação lípido-péptido na temperatura de transição
lipídica, fez-se variar a concentração da solução de péptido β-amilóide (25-35) adicionada à suspensão
de lipossomas de composição POPC/SM (1:1) entre 10 e 80 µM, a pH=7. A solução foi sujeita a 6 ciclos
consecutivos de aquecimento-arrefecimento, de modo a verificar a reversibilidade do comportamento
térmico da amostra (Figura 40). À exceção do primeiro ciclo aquecimento-arrefecimento, que foi
desprezado, os restantes ciclos apresentam-se sobreponíveis.
Uma vez que na Figura 23 não se identificou nenhum pico correspondente à temperatura de
transição de fase da mistura ternária POPC/SM/Chol (1:1:1), não se estudou a interação do fragmento
peptídico com esta mistura lipídica.
Figura 40. Termogramas para POPC/SM (1:1) 1.12mM obtidos, a pH=7, com várias concentrações de Aβ25-35.
Através da Figura 40, verifica-se que a adição de péptido Aβ25-35 aos lipossomas de POPC/SM (1:1)
conduz a um aumento da temperatura de transição de fase da mistura lipídica, uma vez que há um
desvio na posição do pico. Assim, verifica-se que a adição do péptido Aβ25-35 parece estabilizar os
lipossomas, tornando-os mais rígidos, uma vez que a sua transição de fase ocorre a temperaturas
maiores comparativamente aos lipossomas puros. Além disso, verifica-se também um aumento da
largura do pico para concentrações peptídicas de 10 e 25 μM, o que significa uma diminuição na
cooperatividade dos lipossomas. A curva correspondente à adição da solução peptídica de 50 μM
apresenta um pico de largura semelhante ao dos lipossomas puros mas a área debaixo do pico é
inferior, isto é, a entalpia associada à transição diminui. A forma da curva obtida para 80 μM não permite
identificar um máximo correspondente a uma temperatura de transição. Além disso, a diminuição da
capacidade calorífica observada sugere que há destruição dos lipossomas.
-0,4
-0,3
-0,2
-0,1
0
10 15 20 25 30 35 40 45 50 55
Cp
(kC
al/ºC
/mo
l)
Temperatura (ºC)
POPC/SM (1:1)
POPC/SM (1:1) + 10µM Aβ25-35
POPC/SM (1:1) + 25µM Aβ25-35
POPC/SM (1:1) + 50µM Aβ25-35
POPC/SM (1:1) + 80µM Aβ25-35
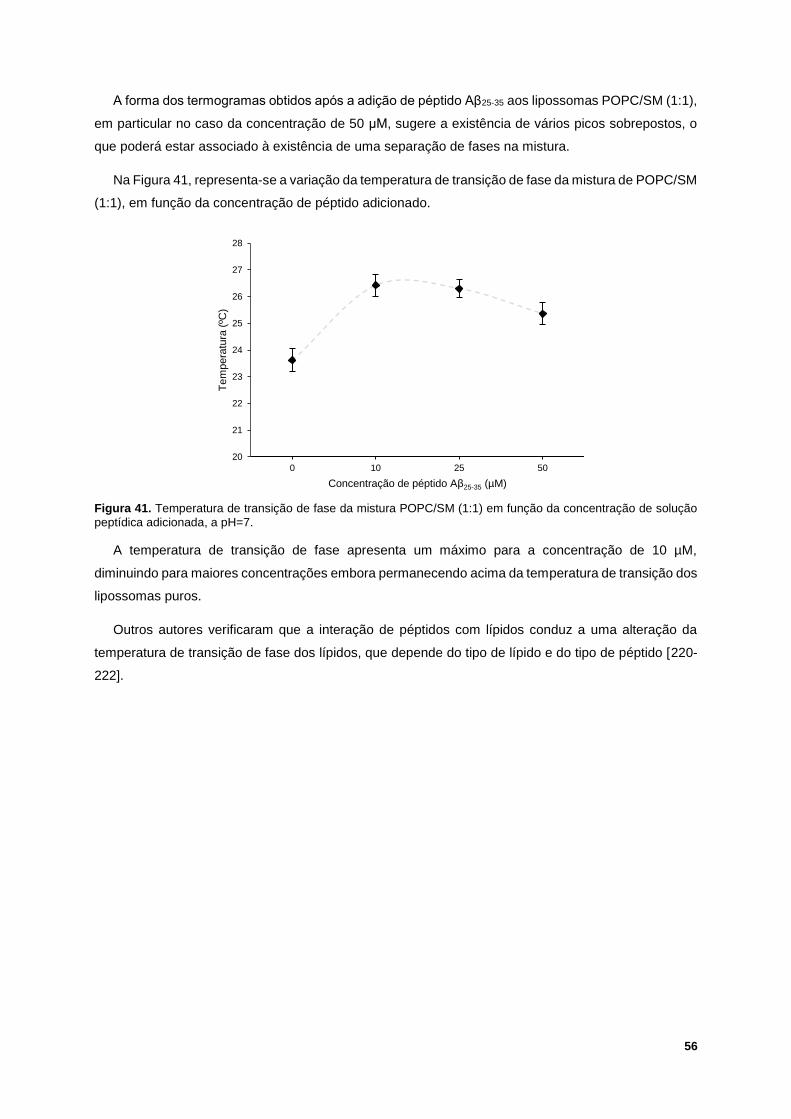
56
A forma dos termogramas obtidos após a adição de péptido Aβ25-35 aos lipossomas POPC/SM (1:1),
em particular no caso da concentração de 50 μM, sugere a existência de vários picos sobrepostos, o
que poderá estar associado à existência de uma separação de fases na mistura.
Na Figura 41, representa-se a variação da temperatura de transição de fase da mistura de POPC/SM
(1:1), em função da concentração de péptido adicionado.
Figura 41. Temperatura de transição de fase da mistura POPC/SM (1:1) em função da concentração de solução
peptídica adicionada, a pH=7.
A temperatura de transição de fase apresenta um máximo para a concentração de 10 µM,
diminuindo para maiores concentrações embora permanecendo acima da temperatura de transição dos
lipossomas puros.
Outros autores verificaram que a interação de péptidos com lípidos conduz a uma alteração da
temperatura de transição de fase dos lípidos, que depende do tipo de lípido e do tipo de péptido [220-
222].
20
21
22
23
24
25
26
27
28
0 10 25 50
Te
mpera
tura
(ºC
)
Concentração de péptido Aβ25-35 (µM)

57
Capítulo V – Conclusões e Trabalho Futuro
Uma das principais características do péptido β-amilóide é a sua elevada tendência de agregação,
pensando-se que este facto tenha uma consequência direta na doença de Alzheimer. A fim de
caracterizar o péptido Aβ25-35 nas condições do presente trabalho, investigou-se o seu estado de
agregação e a estrutura, respetivamente com as técnicas de Análise de Localização de Nanopartículas
(NTA) e Dicroísmo Circular. O péptido Aβ25-35 apresenta uma maior tendência de agregação ao pH
fisiológico. A baixas concentrações (1 e 4 µM) os agregados peptídicos têm uma distribuição de
tamanho mais alargada (50 a 800 nm), comparativamente a altas concentrações (25 µM) (50 a 500
nm). A massa de péptido detetada na forma de agregados não varia linearmente com a concentração
real da solução, existindo um número muito superior de agregados para concentrações mais elevadas.
Este facto permite concluir que a baixas concentrações existe uma fração considerável de Aβ25-35 na
forma oligomérica e/ou monomérica. Quanto à conformação do péptido Aβ25-35, verificou-se que a
variação do pH influencia diretamente a conformação adotada. Assim, a pH=4 o péptido apresenta uma
estrutura secundária maioritariamente de espiral aleatória, enquanto que a pH fisiológico o péptido
adota uma estrutura secundária de folha β. O maior nível de associação das folhas β tem levado à
associação desta estrutura com a formação dos agregados peptídicos.
A interação do péptido Aβ25-35 com os modelos biomembranares foi estudada através da balança de
Langmuir, no caso das monocamadas lipídicas e usando a Microbalança de Cristal de Quartzo com
Dissipação (QCM-D) e Calorimetria Diferencial de Varrimento (DSC), para os lipossomas suportados
ou em suspensão, respetivamente. Apesar dos modelos biomembranares escolhidos serem
estruturalmente diferentes e as condições experimentais adotadas serem diferentes, a análise e
comparação dos resultados obtidos permitiram obter algumas conclusões coincidentes e
complementares relativamente às interações existentes entre o péptido Aβ e as misturas lipídicas
POPC/SM (1:1) e POPC/SM/Chol (1:1:1).
Os resultados obtidos com as monocamadas de Langmuir revelaram-se bastante interessantes,
evidenciando uma boa concordância com os modelos biomembranares computacionais de Vestergaard
et al. [154], assim como com os resultados experimentais de Ding et. al. [46].
As misturas lipídicas usadas como modelos biomembranares, quer a mistura binária POPC/SM (1:1)
quer a mistura ternária POPC/SM/Chol (1:1:1) formam, a 25ºC, monocamadas do tipo líquido a duas
dimensões, embora o colesterol torne a monocamada de POPC/SM/Chol (1:1:1) mais densa e
compacta, devido ao efeito condensante do colesterol. A variação do pH, a 25ºC, não influencia o
comportamento das monocamadas destas misturas lipídicas. Verifica-se que o efeito do colesterol é
idêntico em monocamadas e em lipossomas adsorvidos ou em suspensão.
A interação do péptido Aβ25-35 com ambas as monocamadas lipídicas, a 25ºC, conduz a um aumento
da área ocupada por molécula que é mais significativo quanto maior for a concentração de péptido
Aβ25-35. Estes resultados permitem identificar dois regimes de interacção péptido-lípido. A baixas
concentrações, o péptido Aβ25-35, na forma de monómeros ou oligómeros, adsorve-se na região polar
da monocamada, afetando pouco a organização da monocamada lipídica – Regime I. Para

58
concentração peptídicas elevadas, existe uma concentração elevada de agregados que tendem a
penetrar na monocamada na forma de domínios ricos em Aβ25-35, contribuindo assim para um aumento
significativo da área da monocamada lipídica – Regime II. Esta interpretação é suportada pelos
resultados de NTA que indicam que a baixas concentrações existem poucos agregados e que o número
de agregados de grandes dimemsões aumenta significativamente com a concentração. Verifica-se
ainda que, para toda a gama de concentrações peptídicas estudada em monocamadas, a interação
péptido Aβ25-35 – lípido é mais forte na mistura ternária POPC/SM/Chol (1:1:1), onde, devido à presença
do colesterol, se formam jangadas lipídicas.
A pH=4 e 25ºC, o péptido Aβ25-35 conduz a um aumento de área das monocamadas lipídicas
ligeiramente superior ao observado a pH fisiológico. Este efeito é mais evidente na monocamada de
POPC/SM/Chol (1:1:1). Uma vez que a pH=4 existe uma maior quantidade de agregados de pequenas
dimensões do que a existente a pH=7 podemos concluir que a interação entre a monocamada lipídica
e os agregados de péptido Aβ25-35 de menores dimensões é mais eficiente.
Nos estudos com a QCM-D verificou-se que a interação do péptido Aβ25-35 com os lipossomas
POPC/SM (1:1) provocou um ligeiro aumento da espessura da camada que pode estar relacionado
com uma ténue rigidificação da camada dos lipossomas. No caso dos lipossomas de jangada lipídica,
a interação com o péptido Aβ25-35 não provocou qualquer alteração significativa nas propriedades
viscoelásticas da camada adsorvida. Considerando que as concentrações de péptido Aβ25-35 (10-80
µM) usadas neste estudo são elevadas, e admitindo um mecanismo de interacção semelhante ao
observado no Regime II da interacção péptido-monocamada lípidica, podemos concluir que os
agregados peptídicos de grandes dimensões não interactuam com os lipossomas suportados.
Quanto aos lipossomas em suspensão, só foi possível estudar o efeito do péptido Aβ25-35 sobre os
lipossomas de POPC/SM (1:1) uma vez que não se verificou a existência de uma transição de fase nos
lipossomas de POPC/SM/Chol (1:1:1), dentro do intervalo de temperaturas estudado (10 a 50ºC). O
efeito do péptido sobre os lipossomas de POPC/SM (1:1) depende acentuadamente da concentração
peptídica. A adição de baixas concentrações de péptido Aβ25-35 conduziu a um aumento da temperatura
da transição de fase da mistura lipídica, sugerindo que o péptido Aβ25-35 estabiliza os lipossomas em
suspensão, tornando-os mais rígidos. Esta observação confirma a ligeira rigidificação dos lipossomas
adsorvidos e está de acordo com o comportamento das monocamadas de Langmuir no regime de
baixas concentrações (Regime I) em que o péptido Aβ25-35 adsorve na região polar da monocamada
lipídica. No entanto, para concentrações peptídicas mais elevadas (80 µM), não se observou qualquer
transição de fase da mistura, evidenciando uma destruição dos lipossomas em suspensão. Este
resultado, por sua vez, compara com o observado na interacção das monocamadas para elevadas
concentrações de péptido (Regime II), onde a formação de domínios ricos em péptido Aβ25-35 levam à
disrupção da monocamada.
Os resultados obtidos com lipossomas em suspensão (DSC) para altas concentrações peptídicas
não coincidem com os obtidos com os lipossomas suportados (QCM-D), onde se verificou apenas uma
rigidificação lipossomas POPC/SM (1:1) após adição da solução peptídica de 80 µM. Assim, parece

59
poder concluir-se que os lipossomas num estado adsorvido resistem à adição de elevadas
concentrações de péptido Aβ25-35, enquanto os lipossomas em suspensão sofrem uma rutura.
Uma vez que as alterações provocadas pelo péptido Aβ25-35 nos lipossomas adsorvidos são
pequenas (QCM-D) e que não foi possível estudar o efeito deste péptido em lipossomas de jangada
lipídica em suspensão (DSC), decidimos não efetuar os estudos do efeito do pH com estas técnicas.
Alguns estudos futuros que poderiam complementar o estudo aqui apresentado seria a avaliação
da interação entre lipossomas adsorvidos e o péptido Aβ25-35 através de AFM, de modo a compreender
o efeito real do péptido na estrutura da camada de lipossomas adsorvidos. Seria também útil transferir
as monocamadas para um substrato sólido e observar as diferentes fases através da técnica de AFM.
A microscopia de ângulo de Brewster possibilitaria a observação da separação de fases nas
monocamadas lipídicas. Em termos de composição lipídica, seria interessante realizar ensaios de
QCM-D com lipossomas de POPC, de modo a compreender o efeito da ausência dos domínios lipídicos
mais rígidos na interação lipossomas-péptido Aβ25-35. Outra direção que poderia ser seguida seriam as
técnicas de espectroscopia de ressonância magnética nuclear (NMR): 31P-NMR e 1H-NMR. A técnica
de 31P-NMR seria útil para analisar o movimento do grupo polar dos fosfolípidos, permitindo
compreender o efeito do péptido β-amilóide relativamente à coexistência de diferentes fases na
membrana lipídica. A técnica de 1H-NMR, por sua vez, seria útil para determinar a organização das
cadeias de acilo dos fosfolípidos, de modo a compreender o efeito intramembranar do péptido β-
amilóide na membrana lipídica.

60
Referências bibliográficas
[1] Findeis, M. A. (2007). The role of amyloid β peptide 42 in Alzheimer ’ s disease. Pharmacology & Therapeutics, 116, 266–286.
[2] Canas, P. M., Patrícia, A., Rodrigues, R. J., & Cunha, R. A. (2014). Predominant loss of glutamatergic terminal markers in a b -amyloid peptide model of Alzheimer ’ s disease. Neuropharmacology, 76, 51–56.
[3] Lai, A. Y., & McLaurin, J. (2012). Inhibition of amyloid-beta peptide aggregation rescues the autophagic deficits in the TgCRND8 mouse model of Alzheimer disease. Biochimica et Biophysica Acta - Molecular Basis of Disease, 1822(10), 1629–1637.
[4] Fernandez-Echevarria, C., Díaz, M., Ferrer, I., Canerina-Amaro, A., & Marin, R. (2014). Aβ promotes VDAC1 channel dephosphorylation in neuronal lipd rafts. Relevance to the mechanisms of neurotoxicity in Alzheimer’s disease. Neuroscience, 278, 354–366.
[5] Vetrivel, K. S., & Thinakaran, G. (2010). Membrane rafts in Alzheimer’s disease beta-amyloid production. Biochimica et Biophysica Acta, 1801(8), 860–7.
[6] Taylor, D. R., & Hooper, N. M. (2007). Role of lipid rafts in the processing of the pathogenic prion and Alzheimer’s amyloid-β proteins. Seminars in Cell and Developmental Biology, 18(5), 638–648.
[7] McKoy, A. F., Chen, J., Schupbach, T., & Hecht, M. H. (2012). A novel inhibitor of amyloid β (Aβ) peptide aggregation: from high throughput screening to efficacy in an animal model of Alzheimer disease. The Journal of Biological Chemistry, 287(46), 38992–9000.
[8] Clementi, M. E., Marini, S., Coletta, M., Orsini, F., Giardina, B., & Misiti, F. (2005). Aβ(31-35) and Aβ(25-35) fragments of amyloid beta-protein induce cellular death through apoptotic signals: Role of the redox state of methionine-35. FEBS Letters, 579(13), 2913–2918.
[9] Jablonowska, A., Bakun, M., Kupniewska-Kozak, A., & Dadlez, M. (2004). Alzheimer’s disease Aβ peptide fragment 10-30 forms a spectrum of metastable oligomers with marked preference for N to N and C to C monomer termini proximity. Journal of Molecular Biology, 344(4), 1037–1049.
[10] Kawashima, H., Sohma, Y., Nakanishi, T., Kitamura, H., Mukai, H., Yamashita, M., Kiso, Y. (2013). A new class of aggregation inhibitor of amyloid-β peptide based on an O-acyl isopeptide. Bioorganic and Medicinal Chemistry, 21(21), 6323–6327.
[11] Sanders, H. M., Lust, R., & Teller, J. K. (2009). Amyloid-beta peptide Aβp3-42 affects early aggregation of full-length Aβ1-42. Peptides, 30(5), 849–854.
[12] Morgado, I., & Fändrich, M. (2011). Assembly of Alzheimer’s Aβ peptide into nanostructured amyloid fibrils. Current Opinion in Colloid and Interface Science, 16(6), 508–514.
[13] Kremer, J. J., Pallitto, M. M., Sklansky, D. J., & Murphy, R. M. (2000). Correlation of β-amyloid aggregate size and hydrophobicity with decreased bilayer fluidity of model membranes. Biochemistry, 39(33), 10309–10318.
[14] Mingeot-Leclercq, M. P., Lins, L., Bensliman, M., Van Bambeke, F., Van Der Smissen, P., Peuvot, J., … Brasseur, R. (2002). Membrane destabilization induced by β-amyloid peptide 29-42: Importance of the amino-terminus. Chemistry and Physics of Lipids, 120(1-2), 57–74.
[15] Watts, A., Bokvist, M., Lindstro, F., & Gro, G. (2004). Two Types of Alzheimer ’ s β-Amyloid ( 1 – 40 ) Peptide Membrane Interactions : Aggregation Preventing Transmembrane Anchoring Versus Accelerated Surface Fibril Formation. Journal of Molecular and Cellular Cardiology, 335, 1039–1049.

61
[16] Kayed, R., Sokolov, Y., Edmonds, B., McIntire, T. M., Milton, S. C., Hall, J. E., & Glabe, C. G. (2004). Permeabilization of lipid bilayers is a common conformation-dependent activity of soluble amyloid oligomers in protein misfolding diseases. The Journal of Biological Chemistry, 279(45), 46363–6.
[17] Ciccotosto, G. D., Tew, D., Curtain, C. C., Smith, D., Carrington, D., Masters, C. L., Barnham, K. J. (2004). Enhanced toxicity and cellular binding of a modified amyloid beta peptide with a methionine to valine substitution. The Journal of Biological Chemistry, 279(41), 42528–34.
[18] Tickler, A. K., Smith, D. G., Ciccotosto, G. D., Tew, D. J., Curtain, C. C., Carrington, D., … Barnham, K. J. (2005). Methylation of the imidazole side chains of the Alzheimer disease amyloid-beta peptide results in abolition of superoxide dismutase-like structures and inhibition of neurotoxicity. The Journal of Biological Chemistry, 280(14), 13355–63.
[19] Demuro, A., Mina, E., Kayed, R., Milton, S. C., Parker, I., & Glabe, C. G. (2005). Calcium dysregulation and membrane disruption as a ubiquitous neurotoxic mechanism of soluble amyloid oligomers. The Journal of Biological Chemistry, 280(17), 17294–17300.
[20] Lau, T. L., Ambroggio, E. E., Tew, D. J., Cappai, R., Masters, C. L., Fidelio, G. D., Separovic, F. (2006). Amyloid-beta peptide disruption of lipid membranes and the effect of metal ions. J Mol Biol, 356(3), 759–770.
[21] Phan, H. T. T., Hata, T., Morita, M., Yoda, T., Hamada, T., Vestergaard, M. C., & Takagi, M. (2013). The effect of oxysterols on the interaction of Alzheimer ’ s amyloid beta with model membranes. Biochimica et Biophysica Acta - BiomembranesBA - Biomembranes, 1828(11), 2487–2495.
[22] Peters, I., Igbavboa, U., Schütt, T., Haidari, S., Hartig, U., Rosello, X., Eckert, G. P. (2009). The interaction of beta-amyloid protein with cellular membranes stimulates its own production. Biochimica et Biophysica Acta, 1788, 964–972.
[23] Kremer, J. J., & Murphy, R. M. (2003). Kinetics of adsorption of β-amyloid peptide Aβ(1-40) to lipid bilayers. Journal of Biochemical and Biophysical Methods, 57(2), 159–169.
[24] Kawahara, M., Arispe, N., Kuroda, Y., & Rojas, E. (1997). Alzheimer ’ s Disease Amyloid β-Protein Forms Zn2+ -Sensitive , Cation-Selective Channels Across Excised Membrane Patches from Hypothalamic Neurons. Biophysical Chemistry, 73, 67–75.
[25] Jang, H., Zheng, J., & Nussinov, R. (2007). Models of beta-amyloid ion channels in the membrane suggest that channel formation in the bilayer is a dynamic process. Biophysical Journal, 93(6), 1938–1949.
[26] Butterfield, D., & Lauderback, C. (2002). Lipid peroxidation and protein oxidation in Alzheimer’s disease brain: potential causes and consequences involving amyloid-β-associated free radical oxidative stress. Free Radical Biology & Medicine, 32(11), 1050–1060.
[27] Butterfield, D. A., Swomley, A. M., & Sultana, R. (2013). Amyloid β-peptide (1-42)-induced oxidative stress in Alzheimer disease: importance in disease pathogenesis and progression. Antioxidants & Redox Signaling, 19(8), 823–35.
[28] Wood, W. G., Eckert, G. P., Igbavboa, U., & Mu, W. E. (2003). Amyloid beta-protein interactions with membranes and cholesterol : causes or casualties of Alzheimer ’ s disease. Biochimica et Biophysica Acta, 1610, 281–290.
[29] Demeester, N., Baier, G., Enzinger, C., Goethals, M., Vandekerckhove, J., Rosseneu, M., & Labeur, C. (2000). Apoptosis induced in neuronal cells by C-terminal amyloid β-fragments is correlated with their aggregation properties in phospholipid membranes. Molecular Membrane Biology, 17, 219–228.
[30] Hossain, S., Grande, M., Ahmadkhanov, G., & Pramanik, a. (2007). Binding of the Alzheimer amyloid beta-peptide to neuronal cell membranes by fluorescence correlation spectroscopy. Exp Mol Pathol, 82(2), 169–174.

62
[31] Buchsteiner, A., Hauß, T., Dante, S., & Dencher, N. a. (2010). Alzheimer’s disease amyloid-Β peptide analogue alters the ps-dynamics of phospholipid membranes. Biochimica et Biophysica Acta - Biomembranes, 1798(10), 1969–1976.
[32] Pham, J. D., Chim, N., Goulding, C. W., & Nowick, J. S. (2013). Structures of Oligomers of a Peptide from β-Amyloid. Journal of the American Chemical Society, 135, 12460–12467.
[33] Errico, G. D., Vitiello, G., Ortona, O., Tedeschi, A., Ramunno, A., Maria, A., & Ursi, D. (2008). Interaction between Alzheimer’s Aβ(25–35) peptide and phospholipid bilayers : The role of cholesterol. BBA - Biomembranes, 1778(12), 2710–2716.
[34] Dante, S., Hauß, Æ. T., & Dencher, Æ. N. A. (2006). Cholesterol inhibits the insertion of the Alzheimer’s peptide Aβ(25–35) in lipid bilayers. European Biophysics Journal, 35, 523–531.
[35] Ambroggio, E. E., Kim, D. H., Separovic, F., Barrow, C. J., Barnham, K. J., Bagatolli, L. A., & Fidelio, G. D. (2005). Surface Behavior and Lipid Interaction of Alzheimer β-Amyloid Peptide 1–42 : A Membrane-Disrupting Peptide. Biophysical Journal, 88(4), 2706–2713.
[36] Perluigi, M., Joshi, G., Sultana, R., Calabrese, V., Marco, C. D. E., Coccia, R., & Butterfield, D. A. (2006). In vivo protection bt the xanthate tricyclodecan-9-yl-xanthogenate against amyloid β-peptide (1-42)-induced oxidative stress. Neuroscience, 138, 1161–1170.
[37] Butterfield, D. A., Drake, J., Pocernich, C., & Castegna, A. (2001). Evidence of oxidative damage in Alzheimer ’ s disease brain : central role for amyloid β -peptide. Trends in Molecular Medicine, 7(12), 548–554.
[38] Butter, D. A., Galvan, V., Bader, M., Tang, H., Sowell, R. A., Spilman, P., Bredesen, D. E. (2010). In vivo oxidative stress in brain of Alzheimer disease transgenic mice : Requirement for methionine 35 in amyloid β -peptide of APP. Free Radical Biology & Medicine, 48, 136–144.
[39] Varadarajan, S., Kanski, J., Aksenova, M., Lauderback, C., & Butterfield, D. A. (2001). Different Mechanisms of Oxidative Stress and Neurotoxicity for Alzheimer′s Aβ(1-42) and Aβ(25-35). Journal American Chemical Society, 123, 5625–5631.
[40] Fantini, J., & Yahi, N. (2010). Molecular insights into amyloid regulation by membrane cholesterol and sphingolipids: common mechanisms in neurodegenerative diseases. Expert Reviews in Molecular Medicine, 12(September), e27.
[41] Kakio, A., Nishimoto, S. I., Kozutsumi, Y., & Matsuzaki, K. (2003). Formation of a membrane-active form of amyloid β-protein in raft-like model membranes. Biochemical and Biophysical Research Communications, 303(2), 514–518.
[42] Chakravarthy, B., Ito, S., Atkinson, T., Gaudet, C., Ménard, M., Brown, L., & Whitfield, J. (2014). Evidence that a synthetic amyloid-ß oligomer-binding peptide (ABP) targets amyloid-ß deposits in transgenic mouse brain and human Alzheimer’s disease brain. Biochemical and Biophysical Research Communications, 445(3), 656–660.
[43] Seelert, H., Dani, D. N., Dante, S., Hauß, T., Krause, F., Schäfer, E. Dencher, N. a. (2009). From protons to OXPHOS supercomplexes and Alzheimer’s disease: Structure-dynamics-function relationships of energy-transducing membranes. Biochimica et Biophysica Acta - Bioenergetics, 1787(6), 657–671.
[44] Dante, S., Hauss, T., & Dencher, N. a. (2003). Insertion of Externally Administered Amyloid β Peptide 25-35 and Perturbation of Lipid Bilayers. Biochemistry, 42(46), 13667–13672.
[45] Dong, Z., Saikumar, P., Weinberg, J. M., & Venkatachalam, M. a. (2006). Calcium in cell injury and death. Annual Review of Pathology, 1, 405–434.
[46] Ding, H., Schauerte, J. A., Steel, D. G., & Gafni, A. (2012). β-Amyloid (1–40 ) Peptide Interactions with Supported Phospholipid Membranes : A Single-Molecule Study. Biophysj, 103(7), 1500–1509.

63
[47] Eckert, G. P., Wood, W. G., & Muller, W. E. (2010). Lipid Membranes and β-Amyloid: A Harmful Connection. Current Protein & Peptide Science, 999(999), 1–7.
[48] Mirzabekov, T., Lin, M., Wei-long, Y., Marshal, P. J., Carman, M., Tomaselli, K., … Kagan, B. (1994). Channel formation in planar lipid bilayers by a neurotoxic fragment of the beta-amyloid peptide. Biochemical and Biophysical Research Communications, 202, 1142–1148.
[49] Qi, J. S., & Qiao, J. T. (2001). Amyloid β-protein fragment 31-35 forms ion channels in membrane patches excised from rat hippocampal neurons. Neuroscience, 105, 845–852.
[50] Pomorski, T. G., Nylander, T., & Cárdenas, M. (2013). Model cell membranes: Discerning lipid and protein contributions in shaping the cell. Advances in Colloid and Interface Science, 205, 207–20.
[51] Nelson, D. L., & Cox, M. M. (2004). Lehninger Princples of Biochemistry.
[52] Alekseev, A. E., Reyes, S., Selivanov, V. a., Dzeja, P. P., & Terzic, A. (2012). Compartmentation of membrane processes and nucleotide dynamics in diffusion-restricted cardiac cell microenvironment. Journal of Molecular and Cellular Cardiology, 52(2), 401–409.
[53] Fry, C. (2007). Cell physiology I. Basic Science, 25(10), 401–406.
[54] Campbell, M. K., & Farrell, S. O. (2012). Biochemistry.
[55] Russell, P. J., Hertz, P. E., & McMillian, B. (2013). Biology: The Dynamic Science.
[56] Li, X., Luu, D.-T., Maurel, C., & Lin, J. (2013). Probing plasma membrane dynamics at the single-molecule level. Trends in Plant Science, 18(11), 617–24.
[57] Dalbey, R. E., & Heijne, G. (2002). Protein Targeting, Transport and Translocation.
[58] Sembulingam, K., & Sembulingam, P. (2012). Essentials of Medical Physiology.
[59] Starr, C., Taggart, R., Evers, C., & Starr, L. (2012). Biology: Volume 1 – Cell Biology and Genetics.
[60] Frisardi, V., Panza, F., Seripa, D., Farooqui, T., & Farooqui, A. a. (2011). Glycerophospholipids and glycerophospholipid-derived lipid mediators: A complex meshwork in Alzheimer’s disease pathology. Progress in Lipid Research, 50(4), 313–330.
[61] Farooqui, A. A., & Horrocks, L. A. (2007). Glycerophospholipids in Brain: Phospholipases A2 in Neurological Disorders, Springer.
[62] McMullen, T. P. W., Lewis, R. N. a H., & McElhaney, R. N. (2004). Cholesterol-phospholipid interactions, the liquid-ordered phase and lipid rafts in model and biological membranes. Current Opinion in Colloid and Interface Science, 8(6), 459–468.
[63] Chalfant, C., & Poeta, M. Del. (2010). Advances in Experimental Medicine and Biology – Volume 688 – Sphingolipids as signaling and regulatory molecules.
[64] Harris, R. (2010). Cholesterol Binding and Cholesterol Transport Proteins – Structure and Function in Health Disease.
[65] Smith, L. L. (1981). Cholesterol Autoxidation.
[66] Patterson, G. W., & Nes, W. D. (1991). Physiology and Biochemistry of Sterols.
[67] Othman, R. a., Myrie, S. B., & Jones, P. J. H. (2013). Non-cholesterol sterols and cholesterol metabolism in sitosterolemia. Atherosclerosis, 231(2), 291–299.

64
[68] Hao, Y. H., & Chen, J. W. (2001). Influence of cholesterol on the biophysical properties of the sphingomyelin/DOPC binary system. Journal of Membrane Biology, 183(2), 85–92.
[69] Edidin, M. (2003). The State of Lipid Rafts: From Model Membranes to Cells. Annual Review of Biophysics and Biomolecular Structure, 32, 257–283.
[70] Bartels, T., Lankalapalli, R. S., Bittman, R., & Beyer, K. (2008). Raftlike Mixtures of Sphingomyelin and Cholesterol Investigated by Solid-State 2 H NMR Spectroscopy. Journal American Chemical Society, 130, 14521–14532.
[71] Holland, G. P., McIntyre, S. K., & Alam, T. M. (2006). Distinguishing individual lipid headgroup mobility and phase transitions in raft-forming lipid mixtures with 31P MAS NMR. Biophysical Journal, 90(11), 4248–4260.
[72] Lee, E. J., Yun, U. J., Koo, K. H., Sung, J. Y., Shim, J., Ye, S. K., … Kim, Y. N. (2014). Down-regulation of lipid raft-associated onco-proteins via cholesterol-dependent lipid raft internalization in docosahexaenoic acid-induced apoptosis. Biochimica et Biophysica Acta - Molecular and Cell Biology of Lipids, 1841(1), 190–203.
[73] Aliche-Djoudi, F., Podechard, N., Collin, A., Chevanne, M., Provost, E., Poul, M., Sergent, O. (2013). A role for lipid rafts in the protection afforded by docosahexaenoic acid against ethanol toxicity in primary rat hepatocytes. Food and Chemical Toxicology, 60, 286–296.
[74] Marin, R., Rojo, J. a., Fabelo, N., Fernandez, C. E., & Diaz, M. (2013). Lipid raft disarrangement as a result of neuropathological progresses: A novel strategy for early diagnosis? Neuroscience, 245, 26–39.
[75] Wang, R., Bi, J., Ampah, K. K., Ba, X., Liu, W., & Zeng, X. (2013). Lipid rafts control human melanoma cell migration by regulating focal adhesion disassembly. Biochimica et Biophysica Acta, 1833(12), 3195–205.
[76] Sebastião, A. M., Colino-Oliveira, M., Assaife-Lopes, N., Dias, R. B., & Ribeiro, J. a. (2013). Lipid rafts, synaptic transmission and plasticity: Impact in age-related neurodegenerative diseases. Neuropharmacology, 64, 97–107.
[77] Onuki, Y., & Takayama, K. (2010). Phase behavior in a ternary lipid membrane estimated using a nonlinear response surface method and Kohonen’s self-organizing map. Journal of Colloid and Interface Science, 343(2), 628–633.
[78] Mikhalyov, I., & Samsonov, A. (2011). Lipid raft detecting in membranes of live erythrocytes. Biochimica et Biophysica Acta - Biomembranes, 1808(7), 1930–1939.
[79] Hanzal-Bayer, M. F., & Hancock, J. F. (2007). Lipid rafts and membrane traffic. FEBS Lett, 581(11), 2098–2104.
[80] Simons, K., & Vaz, W. L. C. (2004). Model systems, lipid rafts, and cell membranes. Annual Review of Biophysics and Biomolecular Structure, 33, 269–95.
[81] Thakur, G., Micic, M., & Leblanc, R. M. (2009). Surface chemistry of Alzheimer ’ s disease : A Langmuir monolayer approach. Colloids and Surfaces B : Biointerfaces, 74, 436–456.
[82] Lingwood, D., Kaiser, H. J., Levental, I., & Simons, K. (2009). Lipid rafts as functional heterogeneity in cell membranes. Biochemical Society Transactions, 37(Pt 5), 955–960.
[83] Mateos, M. V., Salvador, G. a., & Giusto, N. M. (2010). Selective localization of phosphatidylcholine-derived signaling in detergent-resistant membranes from synaptic endings. Biochimica et Biophysica Acta - Biomembranes, 1798(3), 624–636.

65
[84] Tivodar, S., Paladino, S., Pillich, R., Prinetti, A., Chigorno, V., van Meer, G., … Zurzolo, C. (2006). Analysis of detergent-resistant membranes associated with apical and basolateral GPI-anchored proteins in polarized epithelial cells. FEBS Letters, 580(24), 5705–5712.
[85] Heerklotz, H. (2002). Triton Promotes Domain Formation in Lipid Raft Mixtures. Biophysical Journal, 83(5), 2693–2701.
[86] Epand, R. M. (2007). Detecting the presence of membrane domains using DSC. Biophysical Chemistry, 126(1-3), 197–200.
[87] Asanov, A., Zepeda, A., & Vaca, L. (2010). A novel form of Total Internal Reflection Fluorescence Microscopy (LG-TIRFM) reveals different and independent lipid raft domains in living cells. Biochimica et Biophysica Acta - Molecular and Cell Biology of Lipids, 1801(2), 147–155.
[88] Ishii, M., Ikushima, M., & Kurachi, Y. (2005). In vivo interaction between RGS4 and calmodulin visualized with FRET techniques: Possible involvement of lipid raft. Biochemical and Biophysical Research Communications, 338(2), 839–846.
[89] Rosolen, a., Peco, C., & Arroyo, M. (2013). An adaptive meshfree method for phase-field models of biomembranes. Part I: Approximation with maximum-entropy basis functions. Journal of Computational Physics, 249, 303–319.
[90] Makky, a., Michel, J. P., Maillard, P., & Rosilio, V. (2011). Biomimetic liposomes and planar supported bilayers for the assessment of glycodendrimeric porphyrins interaction with an immobilized lectin. Biochimica et Biophysica Acta - Biomembranes, 1808(3), 656–666.
[91] Mechler, A., Praporski, S., Piantavigna, S., Heaton, S. M., Hall, K. N., Aguilar, M.-I., & Martin, L. L. (2009). Structure and homogeneity of pseudo-physiological phospholipid bilayers and their deposition characteristics on carboxylic acid terminated self-assembled monolayers. Biomaterials, 30(4), 682–9.
[92] Kouzayha, A., Wattraint, O., & Sarazin, C. (2009). Interactions of two transmembrane peptides in supported lipid bilayers studied by a 31P and 15N MAOSS NMR strategy. Biochimie, 91(6), 774–778.
[93] Brechling, a., Pohl, M., Kleineberg, U., & Heinzmann, U. (2004). Structural organization of DMPC lipid layers on chemically micropatterned self-assembled monolayers as biomimetic systems. Journal of Biotechnology, 112(1-2), 115–125.
[94] Hamai, C., Cremer, P. S., & Musser, S. M. (2007). Single giant vesicle rupture events reveal multiple mechanisms of glass-supported bilayer formation. Biophysical Journal, 92(6), 1988–1999.
[95] El Kirat, K., Morandat, S., & Dufrêne, Y. F. (2010). Nanoscale analysis of supported lipid bilayers using atomic force microscopy. Biochimica et Biophysica Acta, 1798(4), 750–65.
[96] Alessandrini, A., Seeger, H. M., Caramaschi, T., & Facci, P. (2012). Dynamic force spectroscopy on supported lipid bilayers: Effect of temperature and sample preparation. Biophysical Journal, 103(1), 38–47.
[97] Marquês, J. T., De Almeida, R. F. M., & Viana, a. S. (2014). Lipid bilayers supported on bare and modified gold - Formation, characterization and relevance of lipid rafts. Electrochimica Acta, 126, 139–150.
[98] Castellana, E., & Cremer, P. (2006). Solid supported lipid bilayers: From biophysical studies to sensor design. Surface Science Reports, 61(10), 429–444.
[99] Giner-casares, J. J., Brezesinski, G., & Möhwald, H. (2014). Langmuir monolayers as unique physical models. Current Opinion in Colloid & Interface Science, 19(3), 176–182.
[100] Stefaniu, C., Brezesinski, G., & Möhwald, H. (2014). Langmuir monolayers as models to study processes at membrane surfaces. Advances in Colloid and Interface Science, 208, 197–213.

66
[101] Turchanin, A., & Gölzhäuser, A. (2012). Carbon nanomembranes from self-assembled monolayers: Functional surfaces without bulk. Progress in Surface Science, 87(5-8), 108–162.
[102] Qiao, Y., Shen, L., & Guo, Y. (2012). Preparation of polypyrrole films on insulating substrates by self-assembled monolayers. Materials Letters, 86, 38–41.
[103] Bhure, R., Mahapatro, A., Bonner, C., & Abdel-Fattah, T. M. (2011). Stability of phosphonic self assembled monolayers (SAMs) on cobalt chromium (Co-Cr) alloy unde oxidative conditions. Applied Surface Science, 257, 5605–5612.
[104] Smith, R. K., Lewis, P. a, & Weiss, P. S. (2004). Patterning self-assembled monolayers. Progress in Surface Science, 75(1-2), 1–68.
[105] Li, H., Anderson, M. R., Long, G. L., Morris, J. R., & Davis, R. M. (2004). Relationship Between Molecular Structure and Surface Properties of Self-Assembled Monolayers.
[106] Niri, V. H., Flatt, B. K., Fakhraai, Z., & Forrest, J. a. (2010). Simultaneous monitoring of electroformation of phospholipid vesicles by quartz crystal microbalance and optical microscopy. Chemistry and Physics of Lipids, 163(1), 36–41.
[107] Reimhult, E., Höök, F., & Kasemo, B. (2003). Intact vesicle adsorption and supported biomembrane formation from vesicles in solution: Influence of surface chemistry, vesicle size, temperature, and osmotic pressure. Langmuir, 19(5), 1681–1691.
[108] Pignataro, B., Steinem, C., Galla, H. J., Fuchs, H., & Janshoff, a. (2000). Specific adhesion of vesicles monitored by scanning force microscopy and quartz crystal microbalance. Biophysical Journal, 78(1), 487–498.
[109] Giocondi, M. C., Boichot, S., Plénat, T., & Le Grimellec, C. (2004). Structural diversity of sphingomyelin microdomains. Ultramicroscopy, 100(3-4), 135–143.
[110] Veatch, S. L., & Keller, S. L. (2005). Miscibility phase diagrams of giant vesicles containing sphingomyelin. Physical Review Letters, 94(14).
[111] Brownholland, D. P., Longo, G. S., Struts, A. V., Justice, M. J., Szleifer, I., Petrache, H. I., Thompson, D. H. (2009). Phase separation in binary mixtures of bipolar and monopolar lipid dispersions revealed by 2H NMR spectroscopy, small angle x-ray scattering, and molecular theory. Biophysical Journal, 97(10), 2700–2709.
[112] Mainali, L., Raguz, M., & Subczynski, W. K. (2011). Phase-separation and domain-formation in cholesterol-sphingomyelin mixture: pulse-EPR oxygen probing. Biophysical Journal, 101(4), 837–46.
[113] Kahya, N., Scherfeld, D., & Schwille, P. (2005). Differential lipid packing abilities and dynamics in giant unilamellar vesicles composed of short-chain saturated glycerol-phospholipids, sphingomyelin and cholesterol. Chemistry and Physics of Lipids, 135(2), 169–180.
[114] Pokorny, A., Yandek, L. E., Elegbede, A. I., Hinderliter, A., & Almeida, P. F. F. (2006). Temperature and composition dependence of the interaction of delta-lysin with ternary mixtures of sphingomyelin/cholesterol/POPC. Biophysical Journal, 91(6), 2184–2197.
[115] Nicolini, C., Kraineva, J., Khurana, M., Periasamy, N., Funari, S. S., & Winter, R. (2006). Temperature and pressure effects on structural and conformational properties of POPC / SM / cholesterol model raft mixtures — a FT-IR , SAXS , DSC , PPC and Laurdan fluorescence spectroscopy study, 1758, 248–258.
[116] De Almeida, R. F. M., Fedorov, A., & Prieto, M. (2003). Sphingomyelin/phosphatidylcholine/cholesterol phase diagram: boundaries and composition of lipid rafts. Biophysical Journal, 85(4), 2406–2416.

67
[117] Poojari, C., Kukol, A., & Strodel, B. (2013). How the amyloid- β peptide and membranes affect each other: An extensive simulation study. Biochimica et Biophysica Acta - Biomembranes, 1828(2), 327–339.
[118] Matsuzaki, S., Yasuda, Y., Kobayashi, S., Koyama, Y., Kawamoto, K., Katayama, T., & Tohyama, M. (2007). Monomeric Aβ and metals reduce their cytotoxicities to each other. Biochemical and Biophysical Research Communications, 358(2), 540–544.
[119] Giovennelli, L., Casamenti, F., Scalli, C., Bartolini, L., & Pepeu, G. (1995). Differential Effects of Amyloid Peptides β-(1-40) and β-(25-35) Injections Into The Rat Nucleus Basalis. Neuroscience, 66(4), 781–792.
[120] Maloney, B., & Lahiri, D. K. (2011). The Alzheimer’s amyloid β-peptide (Aβ) binds a specific DNA Aβ-interacting domain (A βID) in the APP, BACE1, and APOE promoters in a sequence-speci ic manner : Characterizing a new regulatory motif. Gene, 488(1-2), 1–12.
[121] Bailey, J. a., Maloney, B., Ge, Y. W., & Lahiri, D. K. (2011). Functional activity of the novel Alzheimer’s amyloid β-peptide interacting domain (AβID) in the APP and BACE1 promoter sequences and implications in activating apoptotic genes and in amyloidogenesis. Gene, 488(1-2), 13–22.
[122] Matsuzaki, K. (2007). Physicochemical interactions of amyloid beta-peptide with lipid bilayers. Biochimica et Biophysica Acta, 1768(8), 1935–1942.
[123] Chandrakesan, M., Sarkar, B., Singh, V., Abhyankar, R., Bhowmik, D., Nag, S., … Maiti, S. (2013). The basic structural motif and major biophysical properties of Amyloid-β are encoded in the fragment 18 – 35. Chemical Physics, 422, 80–87.
[124] Lin, Y. S., Bowman, G. R., Beauchamp, K. a., & Pande, V. S. (2012). Investigating how peptide length and a pathogenic mutation modify the structural ensemble of amyloid beta monomer. Biophysical Journal, 102(2), 315–324.
[125] Bartolini, M., Naldi, M., Fiori, J., Valle, F., Biscarini, F., Nicolau, D. V., & Andrisano, V. (2011). Kinetic characterization of amyloid-beta 1-42 aggregation with a multimethodological approach. Analytical Biochemistry, 414(2), 215–225.
[126] Vieira, E. P., Hermel, H., & Mo, H. (2003). Change and stabilization of the amyloid-β(1–40) secondary structure by fluorocompounds. Biochimica et Biophysica Acta, 1645, 6–14.
[127] Rocha, S., Thünemann, A. F., Pereira, M. D. C., Coelho, M., Möhwald, H., & Brezesinski, G. (2008). Influence of fluorinated and hydrogenated nanoparticles on the structure and fibrillogenesis of amyloid beta-peptide. Biophysical Chemistry, 137(1), 35–42.
[128] Zhao, J., Gao, T., Yan, Y., Chen, G., & Li, G. (2013). Probing into the interaction of β-amyloid peptides with bilayer lipid membrane by electrochemical techniques. Electrochemistry Communications, 30, 26–28.
[129] Zeng, H., Zhang, Y., Peng, L., Shao, H., Menon, N. K., Yang, J., … Zagorski, M. G. (2001). Nicotine and Amyloid Formation. Society of Biological Psychiatry, 49, 248–257.
[130] No, X., & No, R. (n.d.). Influence of Cu ( II ) and Glycodendrimers on Amyloid-beta Peptide Aggregation Oxana Klementieva Influence of Cu ( II ) and Glycodendrimers on Amyloid-beta Peptide Aggregation Oxana Klementieva.
[131] Sabaté, R., Espargaró, A., Barbosa-Barros, L., Ventura, S., & Estelrich, J. (2012). Effect of the surface charge of artificial model membranes on the aggregation of amyloid β-peptide. Biochimie, 94(8), 1730–1738.
[132] He, L., Chen, L., Lou, X., Qu, A., Zhou, Z., & Xu, T. (2002). Evaluation of β-amyloid peptide 25–35 on calcium homeostasis in cultured rat dorsal root ganglion neurons. Brain Research, 939, 65–75.

68
[133] Stix, B., & Reiser, G. (1998). β-amyloid peptide 25-35 regulates basal and hormone-stimulated Ca2+ levels in cultured rat astrocytes. Neuroscience Letters, 243(1-3), 121–124.
[134] Stepanichev, M., Zdobnova, I., Zarubenko, I., Lazareva, N., & Gulyaeva, N. V. (2006). Differential effects of tumor necrosis factor-alpha co-administered with amyloid beta-peptide (25-35) on memory function and hippocampal damage in rat. Behavioural Brain Research, 175(2), 352–61.
[135] Shanmugam, G., & Polavarapu, P. L. (2013). Site-specific structure of Aβ(25-35) peptide: isotope-assisted vibrational circular dichroism study. Biochimica et Biophysica Acta, 1834(1), 308–16.
[136] Liu, R. T., Zou, L. B., Fu, J. Y., & Lu, Q. J. (2010). Effects of liquiritigenin treatment on the learning and memory deficits induced by amyloid β-peptide (25-35) in rats. Behavioural Brain Research, 210(1), 24–31.
[137] Wei, G., & Shea, J.-E. (2006). Effects of solvent on the structure of the Alzheimer amyloid-beta(25-35) peptide. Biophysical Journal, 91(5), 1638–1647.
[138] Kellermayer, M. S. Z., Murvai, Ü., Horváth, A., Lászlóffi, E., Soós, K., & Penke, B. (2013). Epitaxial assembly dynamics of mutant amyloid β25-35-N27C fibrils explored with time-resolved scanning force microscopy. Biophysical Chemistry, 184, 54–61.
[139] Loew, C. (2011). The interaction of β -amyloid model peptides with lipid membranes.
[140] Labbé, J., Lefévre, T., Guar-Bégin, A., Auger, M. (2013). Structure and membrane interactions of the β-amyloid fragment 25–35 as viewed using spectroscopic approaches, 7228–7239.
[141] Larini, L., & Shea, J.-E. (2012). Role of β-hairpin formation in aggregation: the self-assembly of the amyloid-β(25-35) peptide. Biophysical Journal, 103(3), 576
[142] Lin, S. Y., & Chu, H. L. (2003). Fourier transform infrared spectroscopy used to evidence the prevention of β-sheet formation of amyloid β(1-40) peptide by a short amyloid fragment. International Journal of Biological Macromolecules, 32(3-5), 173–177.
[143] Anderson, V. L., Ramlall, T. F., Rospigliosi, C. C., Webb, W. W., & Eliezer, D. (2010). Identification of a helical intermediate in trifluoroethanol-induced alpha-synuclein aggregation. Proceedings of the National Academy of Sciences of the United States of America, 107(44), 18850–18855.
[144] Zhang, L., Yagnik, G., Peng, Y., Wang, J., Xu, H. H., Hao, Y., Zhou, F. (2013). Kinetic studies of inhibition of the amyloid beta (1-42) aggregation using a ferrocene-tagged β-sheet breaker peptide. Analytical Biochemistry, 434(2), 292–299.
[145] Pellarin, R., & Caflisch, A. (2006). Interpreting the Aggregation Kinetics of Amyloid Peptides. Journal of Molecular Biology, 360(4), 882–892.
[146] Rocha, S., Loureiro, J. a, Brezesinski, G., & Pereira, M. D. C. (2012). Peptide-surfactant interactions: consequences for the amyloid-beta structure. Biochemical and Biophysical Research Communications, 420(1), 136–40.
[147] Andrews, M. E., Inayathullah, N. M., Jayakumar, R., & Malar, E. J. P. (2009). Conformational polymorphism and cellular toxicity of IAPP and ??AP domains. Journal of Structural Biology, 166(2), 116–125.
[148] Boland, K., Behrens, M., Choi, D., Manias, K., & Perlmutter, D. H. (1996). The Serpin-Enzyme Complex Receptor Recognizes Soluble, Nontoxic Amyloid- Peptide but Not Aggregated, Cytotoxic Amyloid- Peptide. Journal of Biological Chemistry, 271(30), 18032–18044.
[149] Pike, C. J., & Andrea, J. (2002). Structure-Activity Analyses of β-Amyloid Peptides: Contributions of the β25–35 Region to Aggregation and Neurotoxicity. Journal of Neurochemistry, 64(1), 253–265.

69
[150] Wood, S. J., Maleeff, B., Hart, T., & Wetzel, R. (1996). Physical, morphological and functional differences between ph 5.8 and 7.4 aggregates of the Alzheimer’s amyloid peptide Abeta. Journal of Molecular Biology, 256(5), 870–877.
[151] Terzi, E., Hölzemann, G., & Seelig, A. (1995). Self-association of beta-amyloid peptide (1-40) in solution and binding to lipid-membranes. Journal Molecular Biology, 633–642.
[152] Lin, H., Zhu, Y. J., & Lal, R. (1999). Amyloid b-protein (1–40) forms calcium-permeable, Zn21-sensitive channel in reconstituted lipid vesicles. Biochemistry, 38, 11189–11196.
[153] Lin, H., Bhatia, R., & Lal, R. (2001). Amyloid beta protein forms ion channels: implications for Alzheimer’s disease pathophysiology. FASEB Journal, 15, 2433–2444.
[154] Vestergaard, M. C., Morita, M., Hamada, T., & Takagi, M. (2013). Membrane fusion and vesicular transformation induced by Alzheimer’s amyloid beta. Biochimica et Biophysica Acta, 1828(4), 1314–21.
[155] Qiu, L., Buie, C., Reay, A., Vaughn, M. W., & Cheng, K. H. (2011). Molecular dynamics simulations reveal the protective role of cholesterol in β-amyloid protein-induced membrane disruptions in neuronal membrane mimics. Journal of Physical Chemistry B, 115(32), 9795–9812.
[156] Terzi, E., Holzemann, G., & Seeling, J. (1997). Interaction of Alzheimer beta-amyloid peptide(1-40) with lipid membranes. Biochemistry, 2960(97), 14845–14852.
[157] Mason, R. P., Estermyer, J. D., Kelly, J. F., & Mason, P. E. (1996). Alzheimer ’ s Disease Amyloid β Peptide 25-35 Is Localized in the Membrane Hydrocarbon Core : X-Ray Diffraction Analysis. Biochemical and Biophysical Research Communications, 82, 78–82.
[158] Martínez-Senac, M., Villalaín, J., & Gómez-Fernández, C. (1999). Structure of the Alzheimer β-amyloid peptide ( 25 ± 35 ) and its interaction with negatively charged phospholipid vesicles. European Journal Biochemistry, 753, 744–753.
[159] Triulzi, R. C., Li, C., Naistat, D., Orbulescu, J., & Leblanc, R. M. (2007). A Two-Dimensional Approach to Study Amyloid β-Peptide Fragment (25-35). Journal of Physical Chemistry C, 111, 4661–4666.
[160] Lau, T.-L., Gehman, J. D., Wade, J. D., Perez, K., Masters, C. L., Barnham, K. J., & Separovic, F. (2007). Membrane interactions and the effect of metal ions of the amyloidogenic fragment Abeta(25-35) in comparison to Abeta(1-42). Biochimica et Biophysica Acta, 1768(10), 2400–8.
[161] Richter, R., Mukhopadhyay, A., & Brisson, A. (2003). Pathways of lipid vesicle deposition on solid surfaces: a combined QCM-D and AFM study. Biophysical Journal, 85(5), 3035–3047.
[162] Stålgren, J. J. R., Claesson, P. M., & Wärnheim, T. (2001). Adsorption of liposomes and emulsions studied with a quartz crystal microbalance. Advances in Colloid and Interface Science, 89-90, 383–394.
[163] Walters, R. H., Jacobson, K. H., Pedersen, J. a., & Murphy, R. M. (2012). Elongation kinetics of polyglutamine peptide fibrils: A quartz crystal microbalance with dissipation study. Journal of Molecular Biology, 421(2-3), 329–347.
[164] Kotarek, J. a., & Moss, M. a. (2010). Impact of phospholipid bilayer saturation on amyloid-β protein aggregation intermediate growth: A quartz crystal microbalance analysis. Analytical Biochemistry, 399(1), 30–38.
[165] Kotarek, J. a., Johnson, K. C., & Moss, M. a. (2008). Quartz crystal microbalance analysis of growth kinetics for aggregation intermediates of the amyloid-β protein. Analytical Biochemistry, 378(1), 15–24.
[166] Voinova, M. V, Rodahl, M., Jonson, M., & Kasemo, B. (n.d.). Viscoelastic Acoustic Response of Layered Polymer Films at Fluid-Solid Interfaces: Continuum Mechanics Approach, 1–22.

70
[167] Feldötö, Z., Varga, I., & Blomberg, E. (2010). Influence of salt and rinsing protocol on the structure of PAH/PSS polyelectrolyte multilayers. Langmuir : The ACS Journal of Surfaces and Colloids, 26(22), 17048–57.
[168] Viitala, T., Hautala, J. T., Vuorinen, J., & Wiedmer, S. K. (2007). Structure of Anionic Phospholipid Coatings on Silica by Dissipative Quartz Crystal Microbalance, (22), 609–618.
[169] Carapeto, a. P., Serro, a. P., Nunes, B. M. F., Martins, M. C. L., Todorovic, S., Duarte, M. T., Saramago, B. (2010). Characterization of two DLC coatings for joint prosthesis: The role of albumin on the tribological behavior. Surface and Coatings Technology, 204(21-22), 3451–3458.
[170] Wang, Z., Yu, T., Nian, P., Zhang, Q., Yao, J., Li, S., Yue, X. (2014). Fabrication of a Highly b - Oriented MFI-Type Zeolite Film by the Langmuir − Blodgett Method. Langmuir, 30, 4531–4534.
[171] Lü, Z. C., Ruan, W. D., Ji, N., Ren, L. Q., Cong, Q., & Zhao, B. (2006). Fabrication of Large-scale Nanostructure by Langmuir-Blodgett Technique. Journal of Bionic Engineering, 3(2), 59–62.
[172] Girard-Egrot, A. P., & Blum, L. J. (2007). Langmuir-Blodgett Technique for Synthesis of Biomimetic Lipid Membranes. Nanobiotechnology of Biomimetic Membranres, 11, 23–74.
[173] Crawford, N. F., & Leblanc, R. M. (2014). Serum albumin in 2D: A Langmuir monolayer approach. Advances in Colloid and Interface Science, 207(1), 131–138.
[174] Torrent-Burgués, J., Cea, P., Giner, I., & Guaus, E. (2014). Characterization of Langmuir and Langmuir-Blodgett films of an octasubstituted zinc phthalocyanine. Thin Solid Films, 556, 485–494.
[175] Guimarães, J. A., Ferraz, H. C., & Alves, T. L. M. (2014). Langmuir-Blodgett films of cholesterol oxidase and S-layer proteins onto screen-printed electrodes. Applied Surface Science, 298, 68–74.
[176] Ma, H., Perea, B., & Dai, L. L. (2010). Study of two-component colloidal particles at air / water interfaces using Langmuir – Blodgett techniques. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 372(1-3), 61–65.
[177] Pandit, P., Banerjee, M., & Gupta, A. (2014). Growth and morphological analysis of ultra thin PMMA films prepared by Langmuir–Blodgett deposition technique. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 454, 189–195.
[178] Girard-Egrot, A., Godoy, S. & Blum, L. (2005). Enzyme association with lipidic Langmuir-Bloggett films: Interests and applications in nanobioscience. Advances in Colloid and Interface Science, 116, 205-225.
[179] Gebremedhin, S., Singh, A., Koons, S., Bernt, W., Konopka, K., & Duzgunes, N. (2014). Gene delivery to carcinoma cells via novel non-viral vectors: Nanoparticle tracking analysis and suicide gene therapy. European Journal of Pharmaceutical Sciences, 60, 72–79.
[180] Wright, M. (2012). Nanoparticle Tracking Analysis for the Multiparameter Characteriation and Counting of Nanoparticle Suspensions. Methods in Molecular Biology, 906, 511–524.
[181] Zheng, Y., Campbell, E. C., Lucocq, J., Riches, A., & Powis, S. J. (2013). Monitoring the Rab27 associated exosome pathway using nanoparticle tracking analysis. Experimental Cell Research, 319(12), 1706–1713.
[182] Boyd, R. D., Pichaimuthu, S. K., & Cuenat, A. (2011). New approach to inter-technique comparisons for nanoparticle size measurements; using atomic force microscopy, nanoparticle tracking analysis and dynamic light scattering. Colloids and Surfaces A: Physicochemical and Engineering Aspects, 387(1-3), 35–42.
[183] Dragovic, R. a., Gardiner, C., Brooks, A. S., Tannetta, D. S., Ferguson, D. J. P., Hole, P., … Sargent, I. L. (2011). Sizing and phenotyping of cellular vesicles using Nanoparticle Tracking Analysis. Nanomedicine: Nanotechnology, Biology, and Medicine, 7(6), 780–788.

71
[184] Pagano, B., Randazzo, A., Fotticchia, I., Novellino, E., Petraccone, L., & Giancola, C. (2013). Differential scanning calorimetry to investigate G-quadruplexes structural stability. Methods, 64(1), 43–51.
[185] Zhang, H., Taxipalati, M., Que, F., & Feng, F. (2013). Microstructure characterization of a food-grade U-type microemulsion system by differential scanning calorimetry and electrical conductivity techniques. Food Chemistry, 141(3), 3050–3055.
[186] Reed, J., & Reed, T. A. (1997). A Set of Constructed Type Spectra for the Practical Estimation of Peptide Secondary Structure from Circular Dichroism. Analytical Biochemistry, 40(254), 36–40.
[187] Garvie, C. W. (2013). Solution-based approach to study binding to the eIF4E cap-binding site using CD spectroscopy. Analytical Biochemistry, 434(1), 166–171.
[188] Kelly, S. M., & Price, N. C. (2000). The use of circular dichroism in the investigation of protein structure and function. Current Protein & Peptide Science, 1(4), 349–384.
[189] “Avanti Polar Lipids – Preparation of Liposomes” (Online). Disponível em: http://avantilipids.com/index.php?option=com_content&view=article&id=1384&Itemid=372
[190] “Avanti Polar Lipids – How should I store my liposomes?” (Online). Disponível em: http://avantilipids.com/index.php?view=items&cid=5&id=8&option=com_quickfaq&Itemid=385
[191] Bandeiras, C., Serro, A. P., Luzyanin, K., Fernandes, A., & Saramago, B. (2013). Anesthetics interacting with lipid rafts. European Journal of Pharmaceutical Sciences, 48(1-2), 153–165.
[192] Malmstro, J., Otzen, D. E., Sutherland, D., Pe, J., & Cabre, E. J. (2009). Surfactant Protein SP-B Strongly Modifies Surface Collapse of Phospholipid Vesicles : Insights from a Quartz Crystal Microbalance with Dissipation, 97(August), 768–776.
[193] Qiu, F., Mhanna, R., Zhang, L., Ding, Y., Fujita, S., & Nelson, B. J. (2014). Artificial bacterial flagella functionalized with temperature-sensitive liposomes for controlled release. Sensors & Actuators: B. Chemical, 196, 676–681.
[194] Melzak, K. A., Bender, F., Tsortos, A., & Gizeli, E. (2008). Probing Mechanical Properties of Liposomes Using Acoustic Sensors. Langmuir, (11), 9172–9180.
[195] Khelashvili, G., Johner, N., Zhao, G., Harries, D., & Scott, H. L. (2014). Molecular origins of bending rigidity in lipids with isolated and conjugated double bonds: the effect of cholesterol. Chemistry and Physics of Lipids, 178, 18–26.
[196] Dimova, R. (2014). Recent developments in the field of bending rigidity measurements on membranes. Advances in Colloid and Interface Science, 208, 225–34.
[197] Prenner, E., Honsek, G., Dirk, H., & Lohner, K. (2007). Imaging of the domain organization in sphingomyelin and phosphatidylcholine monolayers, 145, 106–118.
[198] Reuter, S., Hofmann, A. M., Busse, K., Frey, H., & Kressler, J. (2011). Langmuir and Langmuir-Blodgett films of multifunctional, amphiphilic polyethers with cholesterol moieties. Langmuir : The ACS Journal of Surfaces and Colloids, 27(5), 1978–89.
[199] Thakur, G., Pao, C., Micic, M., Johnson, S., & Leblanc, R. M. (2011). Surface chemistry of lipid raft and amyloid Aβ(1-4 ) Langmuir monolayer. Colloids and Surfaces B: Biointerfaces, 87(2), 369–377.
[200] Guzmán, E., Liggieri, L., Santini, E., Ferrari, M., & Ravera, F. (2013). Mixed DPPC–cholesterol Langmuir monolayers in presence of hydrophilic silica nanoparticles. Colloids and Surfaces B : Biointerfaces, 105, 284–293.

72
[201] Mladenova, K., Petrova, S. D., As, G., Moskova-doumanova, V., Lalchev, Z., & Doumanov, J. A. (2014). Interaction of Bestrophin-1 with 1-palmitoyl-2-oleoyl-sn- glycero-3-phosphocholine (POPC) in surface films. Colloids and Surfaces B: Biointerfaces, 122, 432–438.
[202] Grauby-heywang, C. (2007). Comparison of the interaction of dihydrocholesterol and cholesterol with sphingolipid or phospholipid Langmuir monolayers. Colloids and Surfaces B : Biointerfaces, 59, 81–86.
[203] Gordon, D. J., Balbach, J. J., Tycko, R., & Meredith, S. C. (2004). Increasing the Amphiphilicity of an Amyloidogenic Peptide Changes the b -Sheet Structure in the Fibrils from Antiparallel to Parallel. Biophysical Journal, 86(January), 428–434.
[204] Wydro, P. (2012). Sphingomyelin/phosphatidylcholine/cholesterol monolayers--analysis of the interactions in model membranes and Brewster Angle Microscopy experiments. Colloids and Surfaces. B, Biointerfaces, 93, 174–9.
[205] Ausuili, A., Torrecillas, A., Aranda, F. J., Mollinedo, F., Gajate, C., Corbalán-García, S., Gómez-Fernández, J. C. (2008). Edelfosine is incorporated into rafts and alters their organization. Journal of Physical Chemistry B, 112(37), 11643–11654.
[206] H, K., LH, C., & GB., Z. (1983). Effects of cholesterol on acyl chain dynamics in multilamellar vesicles of various phosphatidylcholines. Biochimica et Biophysica Acta, 736(2), 137–149.
[207] Pabst, G., Chemin, C., Bourgaux, C., Jean-manuel, P., Patrick, W., Couvreur, P., & Ollivon, M. (2008). Consequences of ions and pH on the supramolecular organization of sphingomyelin and sphingomyelin / cholesterol bilayers, 153, 119–129.
[208] Galdiero, S., Falanga, A., Cantisani, M., & Vitiello, M. (2013). Peptide-Lipid Interactions : Experiments and Applications. International Journal of Molecular Sciences, 14, 18758–18789.
[209] Sonnino, S., & Prinetti, A. (2013). Membrane Domains and the “Lipid Raft” Concept. Current Medicinal Chemistry, 20(1), 4–21.
[210] Chu, H., & U, S. L. (2001). Temperature-induced conformational changes in amyloid β(1-40) peptide investigated by simultaneous FT-IR microspectroscopy with thermal system. Biophysical Chemistry, 89, 173–180.
[211] Terzi, E., Hölzemann, G., & Seelig, J. (1994). Reversible random coil-beta-sheet transition of the Alzheimer beta-amyloid fragment (25-35). Biochemistry, 33(6), 1345–1350.
[212] Chiti, F., & Dobson, C. M. (2006). Protein misfolding, functional amyloid, and human disease. Annual Review of Biochemistry, 75, 333–66.
[213] Ji, X., Naistat, D., Li, C., Orbulescu, J., & Leblanc, R. M. (2006). An alternative approach to amyloid fibrils morphology : CdSe / ZnS quantum dots labelled β-amyloid peptide fragments Aβ(31–35), Aβ(1–40) and Aβ(1–42). Colloids and Surfaces B: Biointerfaces, 50, 104–111.
[214] GD, R., LM, G., & JA., S. (1985). Turns in peptides and proteins. Advances in Protein Chemistry, 37, 1–109.
[215] Mechler, A., Praporski, S., Atmuri, K., Boland, M., Separovic, F., & Martin, L. L. (2007). Specific and selective peptide-membrane interactions revealed using quartz crystal microbalance. Biophysical Journal, 93(11), 3907–3916.
[216] Wang, K. F., Nagarajan, R., & Camesano, T. A. (2014). Antimicrobial peptide alamethicin insertion into lipid bilayer : A QCM-D exploration. Colloids and Surfaces B: Biointerfaces, 116, 472–481.

73
[217] Haw, G., & Cho, N. (2014). Rupture of zwitterionic lipid vesicles by an amphipathic , α-helical peptide : Indirect effects of sensor surface and implications for experimental analysis. Colloids and Surfaces B: Biointerfaces, 121, 340–346.
[218] Abriouel, H., Maqueda, M., Antonio, G., & Valdivia, E. (2001). Monolayer Characteristics of Bacteriocin AS-48 , pH Effect and Interactions with Dipalmitoyl Phosphatidic Acid at the Air – Water Interface. Journal of Colloid and Interface Science, 312, 306–312.
[219] Kru, P., Schalke, M., Wang, Z., Notter, R. H., & Dluhy, R. A. (1999). Effect of Hydrophobic Surfactant Peptides SP-B and SP-C on Binary Phospholipid Monolayers . I . Fluorescence and Dark-Field Microscopy. Biophysical Journal, 77(August), 903–914.
[220] Prenner, E. J., Lewis, R. N. A. H., Kondejewski, L. H., Hodges, R. S., & Y, R. N. M. (1999). Differential scanning calorimetric study of the e ¡ ect of the antimicrobial peptide gramicidin S on the thermotropic phase behavior of phosphatidylcholine , phosphatidylethanolamine and phosphatidylglycerol lipid bilayer membranes. Biochimica et Biophysica Acta, 1417, 211–223.
[221] Lohner, K., & Prenner, E. J. (1999). Differential scanning calorimetry and X-ray di ¡ raction studies of the specificity of the interaction of antimicrobial peptides with membrane-mimetic systems. Biochimica et Biophysica Acta, 1462, 141–156.
[222] Joanne, P., Galanth, C., Goasdoué, N., Nicolas, P., Sagan, S., Lavielle, S., Alves, I. D. (2009). Lipid reorganization induced by membrane-active peptides probed using differential scanning calorimetry. BBA - Biomembranes, 1788(9), 1772–1781.

74
)


![Microscopia Eletrônica de Transmissão [5] - feis.unesp.br · Imagem em campo claro: emprega o feixe transmitido para a formação de contraste, selecionado pela abertura objetiva.](https://static.fdocument.org/doc/165x107/5c0c876709d3f247038c36f8/microscopia-eletronica-de-transmissao-5-feisunespbr-imagem-em-campo.jpg)














