
INSTITUTO OSWALDO CRUZ PATRÍCIA MELLO FERRÃO · Obrigada por compartilhar comigo seu alto-astral...
157
I Pós-Gra PATRÍCIA MELLO F Estudo da modulação de de TGF-β: uma abordage Orientadoras: Dr a . Leila Dr a . Mari INSTITUTO OSWALDO CRUZ aduação em Biologia Celular e Molecu FERRÃO e proteínas de Trypanosoma cruzi em re em proteômica. Dissertação apresentada ao Cruz como parte dos requisito título de Mestre em Biologia Ce a de Mendonça Lima iana Caldas Waghabi RIO DE JANEIRO 2009 ular esposta ao estímulo Instituto Oswaldo os para obtenção do elular e Molecular
Transcript of INSTITUTO OSWALDO CRUZ PATRÍCIA MELLO FERRÃO · Obrigada por compartilhar comigo seu alto-astral...

INSTITUTO OSWALDO CRUZ
Pós-Graduação em Biologia Celular e Molecular
PATRÍCIA MELLO FERRÃO
Estudo da modulação de proteínas de
de TGF-β: uma abordagem prote
Orientadoras: Dra. Leila de Mendonça Lima
Dra. Mariana Caldas Waghabi
INSTITUTO OSWALDO CRUZ
Graduação em Biologia Celular e Molecular
PATRÍCIA MELLO FERRÃO
Estudo da modulação de proteínas de Trypanosoma cruzi em resposta ao estímulo
: uma abordagem proteômica.
Dissertação apresentada ao Instituto Oswaldo
Cruz como parte dos requisitos para obtenção do
título de Mestre em Biologia Celular e Molecular
. Leila de Mendonça Lima
. Mariana Caldas Waghabi
RIO DE JANEIRO
2009
Graduação em Biologia Celular e Molecular
em resposta ao estímulo
Dissertação apresentada ao Instituto Oswaldo
Cruz como parte dos requisitos para obtenção do
título de Mestre em Biologia Celular e Molecular

Ficha catalográfica elaborada pela
Biblioteca de Ciências Biomédicas/ ICICT / FIOCRUZ - RJ
F395
Ferrão, Patrícia Mello.
Estudo da modulação de proteínas de Trypanosoma cruzi em resposta ao estímulo de TGF-β: uma abordagem proteômica / Patrícia Mello Ferrão. – Rio de Janeiro, 2009.
xx, 136f. : il. ; 30cm
Dissertação (mestrado) – Instituto Oswaldo Cruz, Pós-
Graduação em Biologia Celular e Molecular, 2009. Bibliografia: f. 112-136
1. TGF-β. 2. Trypanosoma cruzi 3. Fosforilação de proteínas. 4.
Fosfoproteômica. I. Título.
CDD 616 9363

INSTITUTO OSWALDO CRUZ
Pós-Graduação em
PATRÍCIA MELLO FERRÃO
Estudo da modulação de proteínas de
estímulo de TGF
ORIENTADORAS: Dr a. Leila de Mendonça Lima
Dra. Mariana Caldas Waghabi
Aprovada em: 06/11/2009
EXAMINADORES:
Drª. Tânia Cremonini de Araújo
Dr. Adeílton Alves Brandão
Dr. Gilberto Barbosa Domont
Drª. Claudia Masini d`Avila Levy
Dr. Richard Hemmi Valente (IOC/FIOCRUZ)
ii
INSTITUTO OSWALDO CRUZ
Graduação em Biologia Celular e Molecular
PATRÍCIA MELLO FERRÃO
Estudo da modulação de proteínas de Trypanosoma cruzi
estímulo de TGF -β: uma abordagem prote ômica.
. Leila de Mendonça Lima
. Mariana Caldas Waghabi
06/11/2009
Tânia Cremonini de Araújo-Jorge (IOC/FIOCRUZ) – Presidente
Adeílton Alves Brandão (IOC/FIOCRUZ) – Titular
Gilberto Barbosa Domont (IQ/UFRJ) – Titular
d`Avila Levy (IOC/FIOCRUZ) – Revisora e Suplente
Richard Hemmi Valente (IOC/FIOCRUZ) - Suplente
Biologia Celular e Molecular
Trypanosoma cruzi em resposta ao
ômica.
Presidente
Revisora e Suplente

Estudo da modulação de proteínas de
estímulo de TGF
Estudos recentes mostraram que TGFaguda e crônica. O aumento de seus níveis plasmáticos e a ativação da sua via de sinalização celular são aspectos peculiares da doença chagásica crônica. Além do seu relevante papel na patologia chagásica, também se observou que esta citocina está intimamente associada ao seu ciclo de vida. Trabalhos anteriores demonstraram que TGF-β latente, utilizandoT. cruzi se ligam e internalizam TGFrelacionado à capacidade de proliferação de amastigotas e sua diferenciação em tripomastigotas no final do ciclo intracelular. ao questionamento de quais moléculas de processos de proliferação e diferenciação celular frente ao estímulo por TGFNeste sentido, o presente projeto tem por principal objetmoléculas responsivas ao estímulo de TGFfosfoproteômica. Para tal, extratos de proteínas totais de epimastigotas de (cepa Y), incubadas ou não com TGFexponencial de crescimento do parasito. Evidenciamos que a dose ótima de TGFpara maior indução de fosforilação seria de 5 ng/ml. Em seguida, o tempo ótimo de indução com TGF- β (1, 5, 15, 30 e 60 minutos) foi testado e concludiferenças entre os padrõunidimensionais, nos fazendo optar pela análise exclusiva dos perfis em géis bidimensionais de 7 cm com faixa de pH 3bidimensionais demonstrou diferenças nos padrões de fosfestudados, nos levando a manter um estudo de cinética de tempo. As imagens dos géis foram analisadas e algumas das proteínas consideradas responsivas a TGFforam identificadas por espectrometria de massas. Observamos que as protechoque térmico, tubulinas, desidrogenases, enolases, ciclofilina A, GrpE, cruzipaína, fator de elongamento 1-fosforilação e/ou expressão moduladas em resposta a TGFcorrelacionar a função já descrita na literatura para cada proteína com seu possível papel na sinalização intracelular disparada por TGFcomportamento de fosforilação e/ou expressão apresentado em nossas análises. Por último, foi avaliado se efeito sobre a proliferação dos parasitos. Verificamos que a adição de TGFpromoveu um aumento de até 73% no crescimento dos parasitos nas primeiras 24 horas de estudo. O conjunto de dados obtidosmecanismos moleculares relacionados à sinalização de TGFfonte para detecção de novos alvos terapêuticos para a doença de Chagas.
iii
Estudo da modulação de proteínas de Trypanosoma cruzi
estímulo de TGF -β: uma abordagem prote ômica.
RESUMO
Estudos recentes mostraram que TGF-β está envolvido na cardiopatia chagásica aguda e crônica. O aumento de seus níveis plasmáticos e a ativação da sua via de sinalização celular são aspectos peculiares da doença chagásica crônica. Além do
el na patologia chagásica, também se observou que esta citocina está intimamente associada ao T. cruzi como um regulador de diferentes etapas de seu ciclo de vida. Trabalhos anteriores demonstraram que T. cruzi
latente, utilizando-o na invasão às células hospedeiras e que amastigotas de se ligam e internalizam TGF-β recombinante, estando este evento
relacionado à capacidade de proliferação de amastigotas e sua diferenciação em tripomastigotas no final do ciclo intracelular. Este conjunto de informações nos levou ao questionamento de quais moléculas de T. cruzi poderiam estar envolvidas nos processos de proliferação e diferenciação celular frente ao estímulo por TGFNeste sentido, o presente projeto tem por principal objetivo a caracterização de moléculas responsivas ao estímulo de TGF-β através de uma abordagem fosfoproteômica. Para tal, extratos de proteínas totais de epimastigotas de (cepa Y), incubadas ou não com TGF-β, foram preparados durante a fase
al de crescimento do parasito. Evidenciamos que a dose ótima de TGFpara maior indução de fosforilação seria de 5 ng/ml. Em seguida, o tempo ótimo de
β (1, 5, 15, 30 e 60 minutos) foi testado e concludiferenças entre os padrões de fosforilação são muitos sutis em géis unidimensionais, nos fazendo optar pela análise exclusiva dos perfis em géis bidimensionais de 7 cm com faixa de pH 3-10 não-linear. A avaliação dos perfis bidimensionais demonstrou diferenças nos padrões de fosforilação entre os tempos estudados, nos levando a manter um estudo de cinética de tempo. As imagens dos géis foram analisadas e algumas das proteínas consideradas responsivas a TGFforam identificadas por espectrometria de massas. Observamos que as protechoque térmico, tubulinas, desidrogenases, enolases, ciclofilina A, GrpE, cruzipaína,
-α, fator de iniciação eucariótica 5a, entre outras, têm sua fosforilação e/ou expressão moduladas em resposta a TGF
ar a função já descrita na literatura para cada proteína com seu possível papel na sinalização intracelular disparada por TGF-β, em concordcomportamento de fosforilação e/ou expressão apresentado em nossas análises. Por último, foi avaliado se a adição de TGF-β a culturas de epimastigotas teria algum efeito sobre a proliferação dos parasitos. Verificamos que a adição de TGFpromoveu um aumento de até 73% no crescimento dos parasitos nas primeiras 24 horas de estudo. O conjunto de dados obtidos contribui para a elucidação dos mecanismos moleculares relacionados à sinalização de TGF-β, proporcionando uma fonte para detecção de novos alvos terapêuticos para a doença de Chagas.
Trypanosoma cruzi em resposta ao
ômica.
á envolvido na cardiopatia chagásica aguda e crônica. O aumento de seus níveis plasmáticos e a ativação da sua via de sinalização celular são aspectos peculiares da doença chagásica crônica. Além do
el na patologia chagásica, também se observou que esta citocina como um regulador de diferentes etapas de
T. cruzi é capaz de ativar na invasão às células hospedeiras e que amastigotas de
recombinante, estando este evento relacionado à capacidade de proliferação de amastigotas e sua diferenciação em
Este conjunto de informações nos levou poderiam estar envolvidas nos
processos de proliferação e diferenciação celular frente ao estímulo por TGF-β. ivo a caracterização de
és de uma abordagem fosfoproteômica. Para tal, extratos de proteínas totais de epimastigotas de T. cruzi
, foram preparados durante a fase al de crescimento do parasito. Evidenciamos que a dose ótima de TGF-β
para maior indução de fosforilação seria de 5 ng/ml. Em seguida, o tempo ótimo de (1, 5, 15, 30 e 60 minutos) foi testado e concluímos que as
es de fosforilação são muitos sutis em géis unidimensionais, nos fazendo optar pela análise exclusiva dos perfis em géis
linear. A avaliação dos perfis orilação entre os tempos
estudados, nos levando a manter um estudo de cinética de tempo. As imagens dos géis foram analisadas e algumas das proteínas consideradas responsivas a TGF-β foram identificadas por espectrometria de massas. Observamos que as proteínas de choque térmico, tubulinas, desidrogenases, enolases, ciclofilina A, GrpE, cruzipaína,
ção eucariótica 5a, entre outras, têm sua fosforilação e/ou expressão moduladas em resposta a TGF-β. Buscamos
ar a função já descrita na literatura para cada proteína com seu possível , em concordância com o
comportamento de fosforilação e/ou expressão apresentado em nossas análises. a culturas de epimastigotas teria algum
efeito sobre a proliferação dos parasitos. Verificamos que a adição de TGF-β promoveu um aumento de até 73% no crescimento dos parasitos nas primeiras 24
contribui para a elucidação dos β, proporcionando uma
fonte para detecção de novos alvos terapêuticos para a doença de Chagas.

Study of protein modulation in
Recent studies show that TGFcardiopathy. High levels of TGFshown to be peculiar aspects of patients with chronic Chagas disease. Besides its relevant role in the pathology of Chagas disbe intimately associated with the parasite´s life cycle. Previous works have shown that latent TGF-β, using it in the process of hforms are able to bind and internalize recombinant TGFtheir capacity to proliferate and differentiate into trypomastigote forms at the end of the intracellular cycle. Taken togetheto which T. cruzi molecules are involved in the processe of cellular proliferation and differentiation stimulated by TGFresponsive molecules through a phoprotein extracts from T. cruziwere prepared from parasites in the exponential growth phase. We determined that 5 ng/ml of TGF-β was the dose that induceevents. Next, the optimal induction time (1, 5, 15, 30 and 60 minutes) was evaluated and we concluded that the phosphorylation patterns from the studied times showed only subtle differences using 1exclusively through 2-DE gels (7cm pH 3showed differences in phosphorylation patterns during the timewhich led us to maintain a timepossible TGF-β responsive proteins were identified by mass spectrometry. We observed that heat shock proteins, tubulins, dehydrogenases, enolases, cyclophilin A, GrpE, cruzipain, elongation factortheir phosphorylation and/or expression levels modulated by TGFcorrelate the function already described in the literature for each protein with their possible role in intracellular signaling triggered by TGFphosphorylation and/or expression behavior shown in our analysis. Finally, we assessed whether the addition of TGFeffect on parasite proliferation. We found that TGFto 73% in parasite growth in the first 24 hours of culture. The data presented here contributes to the elucidation of the molecular mechanisms related to TGFsignaling in T. cruzi, providing a source of new potential therapeutic targets against Chagas disease.
iv
Study of protein modulation in Trypanosoma cruzi in response to TGF
stimuli: a proteomic approach.
ABSTRACT
Recent studies show that TGF-β is involved in the acute and chronic chagasic cardiopathy. High levels of TGF-β and the activation of its signaling pathway were shown to be peculiar aspects of patients with chronic Chagas disease. Besides its relevant role in the pathology of Chagas disease, this cytokine was also observed to be intimately associated with Trypanosoma cruzi as a regulator of different stages of the parasite´s life cycle. Previous works have shown that T. cruzi
, using it in the process of host cell invasion. In addition, amastigote forms are able to bind and internalize recombinant TGF-β, and this event is related to their capacity to proliferate and differentiate into trypomastigote forms at the end of the intracellular cycle. Taken together, this set of information raises the question as
molecules are involved in the processe of cellular proliferation and differentiation stimulated by TGF-β. Therefore, this work aims to characterize TGFresponsive molecules through a phosphoproteomic approach. For this purpose, total
T. cruzi epimastigotes (Y strain), incubated or not with TGFwere prepared from parasites in the exponential growth phase. We determined that 5
was the dose that induced the highest number of phosphorylation events. Next, the optimal induction time (1, 5, 15, 30 and 60 minutes) was evaluated and we concluded that the phosphorylation patterns from the studied times showed only subtle differences using 1-DE analysis, leading us to evaluate these profiles
DE gels (7cm pH 3-10 non-linear). The profiles obtained showed differences in phosphorylation patterns during the time-which led us to maintain a time-kinetics study. Gel images wer
responsive proteins were identified by mass spectrometry. We observed that heat shock proteins, tubulins, dehydrogenases, enolases, cyclophilin A, GrpE, cruzipain, elongation factor-1α, eukaryotic initiation factortheir phosphorylation and/or expression levels modulated by TGFcorrelate the function already described in the literature for each protein with their possible role in intracellular signaling triggered by TGF-β, in agreement with theiphosphorylation and/or expression behavior shown in our analysis. Finally, we assessed whether the addition of TGF-β to epimastigotes cultures would have some effect on parasite proliferation. We found that TGF-β addition led to an increase of up
in parasite growth in the first 24 hours of culture. The data presented here contributes to the elucidation of the molecular mechanisms related to TGF
, providing a source of new potential therapeutic targets against
in response to TGF -β
is involved in the acute and chronic chagasic and the activation of its signaling pathway were
shown to be peculiar aspects of patients with chronic Chagas disease. Besides its ease, this cytokine was also observed to
as a regulator of different stages of T. cruzi is able to activate
ost cell invasion. In addition, amastigote , and this event is related to
their capacity to proliferate and differentiate into trypomastigote forms at the end of r, this set of information raises the question as
molecules are involved in the processe of cellular proliferation and . Therefore, this work aims to characterize TGF-β
sphoproteomic approach. For this purpose, total epimastigotes (Y strain), incubated or not with TGF-β,
were prepared from parasites in the exponential growth phase. We determined that 5 d the highest number of phosphorylation
events. Next, the optimal induction time (1, 5, 15, 30 and 60 minutes) was evaluated and we concluded that the phosphorylation patterns from the studied times showed
ng us to evaluate these profiles linear). The profiles obtained
-course under study, kinetics study. Gel images were analyzed and
responsive proteins were identified by mass spectrometry. We observed that heat shock proteins, tubulins, dehydrogenases, enolases, cyclophilin
, eukaryotic initiation factor-5a and others had their phosphorylation and/or expression levels modulated by TGF-β. We tried to correlate the function already described in the literature for each protein with their
, in agreement with their phosphorylation and/or expression behavior shown in our analysis. Finally, we
to epimastigotes cultures would have some addition led to an increase of up
in parasite growth in the first 24 hours of culture. The data presented here contributes to the elucidation of the molecular mechanisms related to TGF-β
, providing a source of new potential therapeutic targets against

v
Trabalho realizado no Laboratório de
Gênomica Funcional e Bioinformática
(LAGFB/IOC), sob a orientação das
doutoras Leila de Mendonça Lima e
Mariana Caldas Waghabi

vi
Aos meus avós, pelo carinho,
confiança e apoio e a minha mãe,
pela dedicação e ternura.

vii
“Só existem dois dias no ano em que nada pode ser feito. Um se chama ontem e o
outro se chama amanhã. Portanto, hoje é o dia certo para amar, acreditar, fazer e
principalmente viver”.
DALAI LAMA
"Assim como a semente traça a forma e o destino da árvore, os teus próprios
desejos é que te configuram a vida".
EMMANUEL
“A mente que se abre a uma nova idéia jamais volta ao seu tamanho original”.
ALBERT EINSTEIN
"Um pouco de ciência nos afasta de Deus. Muito, nos aproxima”.
LOUIS PASTEUR

viii
Agradecimentos
À Deus, por sua sabedoria infinita, que ensina com a paciência e o amor que jamais
poderemos alcançar. A Ele que nos dá saúde e forças para que diariamente
possamos ter a oportunidade de errarmos e nos reerguemos, bastando apenas ser
esta a nossa vontade.
À minha mãe Marília e irmã Marina, pela convivência não mais de todos os dias,
mas ainda assim, sempre juntas comigo no meu coração. A minha tia Sandra, pelo
exemplo de pessoa boa, honesta e generosa que sempre foi. Ao meu pai Paulo, que
apesar da distância, por tantas vezes auxiliou na expansão dos meus pensamentos
e idéias. Aos meus avós, Laura e Francisco, por todo amor, carinho e confiança
incomensuráveis depositados em mim durante todo a minha vida. Obrigada por
sempre acreditarem em mim!
Às minhas queridas Suzi e Lola, companheiras de todas as horas, que com alegria e
simplicidade me lembram a todo o momento a alegria de viver. Ao Zoé, meu
pequeno, que ainda vive, no meu coração.
Às minhas velhas amigas Thaís Vilela e Gabriela Vasquez, que me acompanham há
tanto tempo e com quem compartilho as alegrias, as tristezas e as últimas
novidades. Vocês são minhas jóias raras!
À minha queridíssima amiga Tatiana Gutierrez, aquisição mais sólida e valiosa dos
tempos de fundão, com quem vibro a cada nova conquista pessoal e profissional.
Obrigada por compartilhar comigo seu alto-astral e pensamentos positivos. Ah, e
curte bem essa vida de casada!
Ao Diego, a maior e melhor surpresa da minha vida. Quem poderia imaginar que
daquela conversa informal num domingo de 2x2 no maraca poderia começar uma
história de amor tão bonita e tão forte, que até hoje me enche de orgulho e
felicidade. Ainda me pego por aí, sonhando acordada e suspirando quando meus
pensamentos voam até você. Você ainda mexe muito comigo... Te amo, amo, amo
demais!

ix
À minha orientadora Mariana Waghabi, pilar incontestável da idealização e
construção desta dissertação. Sem você, nada disso teria acontecido. Apesar dos
momentos turbulentos, gostaria que soubesse que tenho grande admiração por
você, pela sua paixão e envolvimento com a pesquisa, por você ser uma
pesquisadora talentosa e dedicada. Muito obrigada por tudo que me ensinou, pela
paciência que teve comigo e por ter me confiado à realização deste trabalho.
À minha orientadora Leila de Mendonça Lima, por ter me dado a oportunidade de
estar aqui hoje, tornando possível todo o conhecimento e experiência que adquiri ao
longo desses últimos anos. Pela contribuição intelectual à dissertação, através de
seu raciocínio lógico e capacidade notável de enxergar os resultados além.
Às meninas do lab 101:
À Palominha, minha doce menina, futura gêninha da pesquisa. Trabalhar com você
é fácil e prazeroso. Confio cegamente em tudo o que você faz. Te adoro
incondicionalmente, como se fosse minha irmã!
À Fabi, com seus trejeitos engraçados, sempre de bom-humor e disposta a ajudar
pessoas perfeccionistas e minuciosas (ou como ela diria, pessoas chatas e lerdas)
como eu. Muito obrigada pela ajuda valiosa com a cultura das epis e outros
experimentos da dissertação!
À Melissa, aluna sênior do laboratório, que, apesar dos momentos ranzinzas, é um
amor de pessoa e ótima companhia, ouvindo, dando bons conselhos e incentivando
a todas. Além, é claro, de ser uma ótima consultora de modas (chique demais essa
menina!).
À Talita, menina doidinha, sempre perdida num monte de afazeres dentro e fora do
laboratório. Obrigada por tudo que me ensinou nesses últimos 5 anos. Torço muito
para que você alcance suas aspirações profissionais e, acima de tudo, que seu
coração esteja tranqüilo e apaixonado como está hoje!

x
À Cris, a novata do grupo, pelas conversas maduras e pelo exemplo de virar a mesa
e dar a volta por cima.
Às meninas do lab 106:
À Márcia, por ser uma pessoa que naturalmente esbanja simpatia e acessibilidade.
Você é acima de tudo uma pessoa sincera, que não tem medo de expor seus
sentimentos e emoções. O coração imenso de mãe que você guarda no peito é sem
dúvida seu maior tesouro. Continue sempre a pessoa amiga, que cativa a todos
onde passa.
À Luciana, pela animação diária e histórias divertidas, que tornam o dia-a-dia leve e
agradável. Além, é claro, de sua prestatibilidade ímpar, estando sempre disposta a
ajudar (e como ajudou)!
À galera do lab 105:
Ao Marcos, Vanessa e Renata pelo alto-astral e energia que passam pra mim, por
serem sempre atenciosos e prestativos nos momentos que precisei de ajuda.
Marcelo e Teca, obrigada pelas idéias e sugestões construtivas dadas ao trabalho
durante as apresentações nos seminários de laboratório.
Aos amigos, Leonardo, Marcos, Aline e Andressa pelas conversas, conselhos e
amizade.
À dona Neiva, cuja ajuda silenciosa é de extrema importância para a realização do
trabalho de todos do laboratório.
Aos meninos do lab 201, pelos momentos descontraídos nas festinhas do
laboratório.
Às pesquisadoras Cristina Pessolani e Ana Gisele Neves-Ferreira, pela contribuição
intelectual dada ao desenvolvimento desta dissertação.

xi
À Claudia Levy, pela revisão desta dissertação, por aceitar participar da banca
avaliadora e por, assim como na minha monografia, enriquecer com idéias e
sugestões os trabalhos que realizo.
Um agradecimento super, hiper, ultra, mega especial a todos (Paloma, Fabiane,
Melissa, Luciana, Talita, Leonardo e Márcia) que bravamente participaram da
maratona do processamento de 200 “spots” em uma semana! Serei eternamente
grata pela ajuda (e deverei para sempre favores em troca)!
Agradeço ao Instituto Oswaldo Cruz pelo apoio financeiro, através do qual recebi
minha bolsa de mestrado e pela oportunidade de realizar este trabalho, que tanto
acrescentou no meu amadurecimento profissional.
Às plataformas de Eletroforese 2D (RTP02C) e de Espectrometria de massas -
PDTIS/Fiocruz, que viabilizaram a realização deste trabalho.

xii
LISTA DE SIGLAS E ABREVIATURAS
°C - grau Celsius
µm - micrômetro
1,3-BPGA - 1,3-bifosfo-D-ácido glicérico
1DE - eletroforese unidimensional
2DE - eletroforese bidimensional
ACN - acetonitrila
ADP- adenosina difosfato
AHADH - desidrogenase L-alfa-hidróxi-ácida aromática
AKR- aldo-ceto redutase
ALK1 – “activin receptor-like kinase 1”
ALK2 – “activin receptor-like kinase 2”
ATP - adenosina trifosfato
BMP – proteínas morfogênicas do osso
BPG- bifosfoglicerato
BSA – albumina de soro bovino
CBB – “coomassie brilliant blue”
CDC – “Centers for Disease Control and Prevention”
Cdc42- “cell division cycle 42”
CHAPS – 3-[(3-cholamidopropil)dimetilamonio]-1-propanosulfonato
CK2- “casein kinase 2”
Clp – “caseinolytic protease”
cm - centímetro
CRM1 – “chromosome region maintenance 1”
Da - dalton
DNA – ácido desoxirribonucléico
DTT – ditiotreitol
EF-1α – “elongation factor 1α” (fator de elongamento-1α)
EIF-5A – “eukaryotic initiation factor 5a” (fator eucariótico de iniciação da tradução –
5a)
ERK- “extracellular signal-regulated kinase”
Fos- “fructo oligosaccharide”
FtsH – “filamentation temperature-sensitive”
g – aceleração da gravidade

xiii
GAPDH - gliceraldeído-3-fosfato desidrogenase
GDF – fatores de crescimento e diferenciação
GDP- guanosina difosfato
gp63s- glicoproteína 63
GTP- guanosina trifosfato
h – hora
HIV – “human imunodeficiency vírus” (vírus da imunodeficiência humana)
Hsp – “heat shock protein” (proteína de choque térmico)
IEF – “isoelectric focusing” (focalização isoelétrica)
IgE – imunoglobulina E
IPG – gradiente imobilizado de pH
JNK- Jun N-terminal kinase
kb - quilobase
kDa – quilodalton
LAP – “latency associated peptide” (peptídeo associado à latência)
M – molar
mA - miliamper
MALDI – “matrix assisted laser desorption ionization” (desorção/Ionização a Laser
Assistido por Matriz)
MAPKs- “mitogen activated protein kinase”
MH2- “MAD homolog 2”
mL – mililitro
mm - milímetro
mM – milimolar
mRNP – “messenger ribonucleoprotein”
MS – espectrometria de massas
MW– massa molecular
MyoD- “Myogenic diferentiation”
NAC - complexo associado ao polipeptídeo nascente
NAD(P)- nicotinamida adenina dinucleotídeo fosfato
NTA – ácido nitrilotriacético
OD – “optical density” (densidade óptica)
OMS - Organização Mundial de Saúde
OPAS – Organização Pan-Americana de Saúde
p/v – peso/volume

xiv
PAGE – eletroforese em gel de poliacrilamida
pb (s) – par(es) de base
PBS – “phophate buffered saline” (salina tamponada com fosfato)
PDI - proteína dissulfeto isomerase
PDTIS – Programa de Desenvolvimento Tecnológico em Insumos para Saúde
PGFS- prostaglandina F sintase
pH - potencial hidrogeniônico
pI - ponto isoelétrico
PI3K- fosfatidilinositol 3 quinase
PMF – “peptide mass fingerprint”
PP2A- proteína fosfatase 2A
PPIase - peptidil-prolil isomerase
PPM – parte por milhão
PTM- “post-translational modifications” (modificações pós-traducionais)
qsp – quantidade suficiente para
RNA – ácido ribonucléico
rpm – rotações por minuto
SDS – dodecil sulfato de sódio
Smad – união dos nomes “Mothers against decapentaplegic” (MAD) e SMA de
Caenorhabditis elegans
SnoN- “Ski-related novel protein N”
T - tempo
TAT - tirosina aminotransferase
TBP – tributilfosfina
TCA – ácido tricloro acético
TcMPX - triparedoxina peroxidase mitocondrial
TCTP - proteína tumoral traducionalmente controlada
TEA - trietanolamina
TEMED – N,N,N’,N’ – tetrametiletilenodiamina
TFA – ácido trifluoro acético
TGF-β – “transforming growth factor beta” (fator transformador de crescimento beta)
TOF – “time of flight” (tempo de vôo)
TR- tripanotiona redutase
TRIS - tris-hidroxi-metil-amino-metano
TriX - triparedoxina

xv
TβRI - receptor para TGF-β tipo I
TβRII - receptor para TGF-β tipo II
TβRIII - receptor para TGF-β tipo III
v – volts
v/v – volume/volume
WB - western-blot
WHO – “World Health Organization” (Organização Mundial da Saúde)
µg – micrograma
µL – microlitro

xvi
LISTA DE TABELAS MATERIAIS E MÉTODOS
3.1 Solução Inorgânica do meio de cultura LIT 25
3.2 Solução Orgânica do meio de cultura LIT 25
3.3 Meio de cultura LIT 25
3.4 Tampão de amostras de proteína em gel SDS-PAGE (1X) 27
3.5 Géis 12% SDS-PAGE para “strips” de 7cm 27
3.6 Tampão de corrida Laemmli 27
3.7 Parâmetros para focalização isoelétrica 28
3.8 Tampão de Equilíbrio I 29
3.9 Tampão de Equilíbrio II 29
3.10 Comprimentos de onda ideais para emissão e excitação da fluorescência dos corantes ProQ Diamond e Sypro Ruby
31
3.11 Parâmetros utilizados nas buscas do Mascot 35
RESULTADOS
4.1 Identificação das proteínas moduladas por TGF-β 47
4.2 Valores dos “spots” responsivos a TGF-β nos tempos estudados 55
DISCUSSÃO
5.1 Comportamento das proteínas em resposta a TGF-β 95

xvii
LISTA DE FIGURAS INTRODUÇÃO
1.1 Mapa da distribuição geográfica da doença de Chagas 01
1.2 Manifestações clínicas da doença de Chagas 03
1.3 Formas evolutivas de T. cruzi 04
1.4 Ciclo Biológico de T. cruzi 05
1.5 Superfamília TGF-β 07
1.6 Via clássica de sinalização por TGF-β 10
1.7 Vias alternativas de sinalização por TGF-β 12
RESULTADOS
4.1 Curva de crescimento das formas epimastigotas de Trypanosoma cruzi 37
4.2 Padronização da dose ótima de incubação com TGF-β 39
4.3 Padronização do tempo ótimo de incubação com TGF-β 41
4.4 Perfis bidimensionais dos extratos protéicos de epimastigotas incubadas ou não com TGF-β nos tempos de estudo
43
4.5 Quantidade total de “spots” protéicos em cada condição estudada 45
4.6 Mapa protéico do extrato das epimastigotas tratadas com TGF-β por 1 minuto
46
4.7 Comportamento das proteínas de choque térmico 61
4.8 Comportamento da proteína da DnaK 62
4.9 Comportamento da proteína Hsc70 63
4.10 Comportamento da proteína GrpE 64
4.11 Comportamento da proteína Ciclofilina A 65
4.12 Comportamento da proteína β-Tubulina 67
4.13 Comportamento da proteína Prostaglandina F2α sintase 69
4.14 Comportamento da proteína Aldo-ceto redutase 70
4.15 Comportamento da proteína Enolase 71
4.16 Comportamento das proteínas GAPDH e Malato desidrogenase 73
4.17 Comportamento das proteínas dissulfeto isomerase e desidrogenase aromática L-α hidroxiácida
74
4.18 Comportamento da proteína Tirosina aminotransferase 75
4.19 Comportamento das proteínas Hipotéticas 77
4.20 Comportamento da proteína hipotética (“spot” 17) 78
4.21 Comportamento do complexo associado ao polipeptídeo nascente 80

xviii
4.22 Comportamento do fator de iniciação eucariótica – 5a 81
4.23 Comportamento do fator de elongamento – 1α 82
4.24 Comportamento da proteína Cruzipaína 83
4.25 Comportamento da subunidade proteolítica do complexo hslvu 84
4.26 Comportamento da proteína Triparedoxina peroxidase 85
4.27 Comportamento do fator de liberação de histamina dependente de IgE 87
4.28 Proliferação das epimastigotas após adição de TGF-β ao meio de cultura
88

xix
ÍNDICE
Conteúdo Página
RESUMO iii
ABSTRACT iv
LISTA DE SIGLAS E ABREVIATURAS xii
LISTA DE TABELAS xvi
LISTA DE FIGURAS xvii
1. INTRODUÇÃO 01
1.1 Doença de Chagas. 01
1.2 Trypanosoma cruzi. 03
1.3 Fator Transformador de Crescimento beta (TGF-β). 06
1.3.1 A superfamília TGF-β. 06
1.3.2 Receptores de TGF-β. 08
1.3.3 Via de sinalização clássica por TGF-β. 09
1.3.4 Vias alternativas de sinalização por TGF-β. 11
1.3.5 Genes e fatores de transcrição responsivos a TGF-β 13
1.3.6 Papel do TGF-β na doença de Chagas. 15
1.3.7 Papel de TGF-β na biologia do T.cruzi. 16
1.4 Modificações pós-traducionais. 18
1.4.1 Fosforilação de Proteínas: Aspectos Gerais. 19
1.5 Fosforilação em Trypanosoma cruzi. 20
1.6 Fosfoproteômica. 21
1.7 Proteoma de T.cruzi. 22
2. OBJETIVOS 24
2.1 Objetivos Gerais 24
2.2 Objetivos Específicos 24
3. MATERIAL E MÉTODOS 25
3.1 Cultivo dos parasitos. 25
3.2 Preparo das amostras, Lise e Extração das Proteínas. 26
3.3 Precipitação e Dosagem das Proteínas. 26
3.4 Eletroforese Unidimensional. 27

xx
3.5 Eletroforese Bidimensional. 28
3.5.1 Focalização Isoelétrica (IEF, 1ª Dimensão). 28
3.5.2 Eletroforese em SDS/PAGE (2ª Dimensão) 29
3.6 Coloração dos Géis e Obtenção das Imagens. 30
3.7 Análise de Imagens. 31
3.8 Retirada e Processamento dos “spots”. 32
3.9 Identificação de proteínas por Espectrometria de Massas. 34
3.10 Identificação dos peptídeos. 35
3.11 Proliferação de epimastigotas em resposta a adição de TGF-β. 36
4. RESULTADOS 37
4.1 Curva de crescimento das formas epimastigotas de Trypanosoma cruzi.
37
4.2 Padronização da dose ótima de TGF-β. 38
4.3 Padronização do tempo ótimo de incubação com TGF-β. 40
4.4 Perfis bidimensionais de fosforilação e expressão de proteínas de formas epimastigotas de T. cruzi em resposta a TGF-β.
41
4.5 Análises dos perfis bidimensionais diferenciais de formas epimastigotas de T. cruzi em resposta ao TGF-β.
44
4.5.1 Quantidade total de “spots” protéicos em cada condição estudada 44
4.5.2 Identificação dos “spots” presentes nos mapas proteômicos de extratos de epimastigotas de T. cruzi.
45
4.5.3 Proteínas moduladas após a adição de TGF-β. 53
4.6 Proliferação das epimastigotas após adição de TGF-β. 88
5. DISCUSSÃO 89
6. CONCLUSÕES 111
7. REFERÊNCIAS BIBLIOGRÁFICAS 112

Introdução
1
1. INTRODUÇÃO
1.1. Doença de Chagas
A Doença de Chagas, identificada pelo brasileiro Carlos Chagas em 1909, há
exatos 100 anos, é causada pelo protozoário flagelado Trypanosoma cruzi e é
apontada como a maior causa de miocardite aguda e cardiomiopatia crônica
progressiva em regiões endêmicas da America Latina (Moncayo & Silveira, 2009).
Estimativas epidemiológicas recentes indicam que cerca de 12 milhões de pessoas
estão infectadas com T. cruzi e que haja de 100.000 a 200.000 novos casos por ano,
com 20 a 40 mil mortes ao ano (Fapesp, 2008).
A distribuição geográfica da doença (Figura 1.1), incluindo seus reservatórios
e vetores, se estende do sul dos Estados Unidos ao sul da Argentina e Chile. Dessa
forma, a infecção cobre praticamente toda a América, onde 90 milhões de pessoas
encontram-se expostas a infecção (Coura & Dias, 2009).
Os mecanismos de transmissão da doença de Chagas podem ser divididos
em dois grupos: (1) mecanismos principais, por via vetorial (triatomíneos), transfusão
sanguínea, alimentos contaminados e transmissão placentária; (2) mecanismos
secundários, por acidentes laboratoriais, transplantes de órgãos, transmissão sexual
e compartilhamento de seringas por usuários de drogas (Prata, 2001; Coura, 2007;
Coura & Dias, 2009). Devido ao movimento de migração populacional, um número
cada vez maior de casos de importação da doença de Chagas tem sido detectado
em áreas não endêmicas, como América do Norte e várias partes da Europa, Ásia e
Oceania (Schmunis, 2007).
___________________________________________________________________
Figura 1.1: mapa da distribuição geográfica destacando (em vermelho) os países onde a doença de Chagas é endêmica. Fonte: Organização Mundial de Saúde (1996).

Introdução
2
A doença de Chagas possui duas fases sucessivas: aguda e crônica. A fase
aguda inicia-se após um período de incubação de 7 a 10 dias e tem como principais
características a curta duração (1 a 3 meses) e a presença de parasitos no sangue
circulante do indivíduo infectado. Em alguns casos, são encontrados chagomas de
inoculação e/ou o sinal de romaña (figura 1.2), que são lesões na porta de entrada
do parasita, geradas principalmente em resposta à ruptura das células hospedeiras
pelo T. cruzi. Apesar disso, a sintomatologia pode passar despercebida, podendo
ser confundida com uma gripe comum. Os sintomas mais comuns são febre, astenia,
cefaléia, dores no corpo e anorexia, mas também pode ser observada a presença de
miocardite aguda em aproximadamente 80 a 90% dos casos agudos. A resposta
imune forte e específica associada à resposta imune inata controla a infecção
aguda, que evolui para a fase crônica. A cardiomiopatia chagásica é ainda uma
doença incurável, sendo segundo dados da Organização Pan-Americana de Saúde
(OPAS) uma das principais causas de doenças do miocárdio na América Latina
(Zeledon, 1997).
A fase crônica pode ser dividida em duas formas clínicas: (i) a indeterminada
(assintomática, mas com sorologia positiva), e a (ii) determinada, na qual sinais e
sintomas clínicos (figura 1.2) são observados em cerca de 30% dos indivíduos
infectados. As manifestações clínicas da doença de Chagas crônica estão
relacionadas com patologias envolvendo o coração, esôfago e intestino (Coura,
2007). Alterações cardíacas são as manifestações mais sérias e frequentes da
doença de Chagas crônica e afetam cerca de 20 a 40% dos indivíduos, em geral,
anos ou décadas após a infecção inicial (Rassi et al., 2009). A doença cardíaca
chagásica é também a causa mais comum de cardiomiopatia na América Latina e,
em áreas endêmicas, é a causa principal de morte cardiovascular entre pacientes de
30 a 50 anos (Rassi et al., 2000). A cardiomiopatia chagásica está geralmente
associada ao aumento do coração através de uma fibrose exacerbada e disfunções
que incluem arritmias, distúrbios de condução elétrica e falência congestiva do
coração (Higuchi et al., 1999).

Introdução
3
___________________________________________________________________
Figura 1.2: Manifestações clínicas características das fases aguda (a) e crônica (b e c) da doença de Chagas. a. Sinal de romanã, caracterizado como um edema unilateral bipalpebral (imagem retirada do endereço eletrônico http://www.jyi.org/features/ft.php?id=185). b. Chapa radiográfica e c. coração dissecado, revelando a cardiomegalia característica de pacientes com cardiomiopatia chagásica crônica (as imagens b e c foram retiradas de Gilles, 2000). 1.2. Trypanosoma cruzi
O T. cruzi é o agente etiológico da doença de Chagas e pertence à família
Trypanosomatidae, que compreende um grande grupo de parasitos flagelados
heteroxênico capazes de causar doenças em humanos e outros animais (De Souza,
2002). Este parasito possui um ciclo de vida complexo, que se caracteriza pela
presença de vários estágios de desenvolvimento observados em hospedeiros
vertebrados e invertebrados (Kirchhoff, 1993; De Souza, 2002). Durante seu ciclo de
vida, três diferentes formas evolutivas de T. cruzi podem ser facilmente identificadas
(figura 1.3) por microscopia ótica em preparações coradas com Giemsa ou por
microscopia de fase, sendo elas: (a) Amastigotas, que também são conhecidas
como esferomastigotas, micromastigotas ou forma leishmanial. São formas
replicativas, que apresentam morfologia arredondada com flagelo curto e
cinetoplasto anterior ao núcleo. Estudos com amastigotas, obtidas de fontes
variadas, mostraram que as mesmas também são infectivas para células de
vertebrados (de Carvalho & de Souza, 1986; Andrews, 1990); (b) Epimastigotas, que
possuem formato fusiforme com 20 a 40 µm de comprimento, são formas flageladas
replicativas e apresentam o cinetoplasto em forma de disco côncavo, localizado
anteriormente ao núcleo. Estas formas de T. cruzi são observadas principalmente no
intestino de hospedeiros invertebrados. Epimastigotas são também encontradas
dentro de células de vertebrados durante o processo de diferenciação amastigota-
tripomastigota, sendo denominadas formas “epimastigotas- like” (Almeida-de-Faria et
al., 1999); (c) Tripomastigotas são as formas infectivas clássicas e apresentam
morfologia alongada com comprimento aproximado de 25 µm e um diâmetro de 2
µm. Estas formas apresentam cinetoplasto redondo e flagelo longo localizados na
região posterior e anterior ao núcleo, respectivamente. Elas são observadas em (i)
a b cc

células tissulares e no san
nas fezes, e na urina do hospedeiro invertebrado, (iii) na fase estacionária de
crescimento de culturas axênicas, e (iv) na fase líquida de culturas celulares. Essa
forma é incapaz de se dividir.
___________________________________________________________________
Figura 1.3: Formas evolutivas de Procopio et al., 1999), http://www.fiocruz.br/~ccs/arquivosite/estetica/chagas.htm) e http://www.stanford.edu/class/humbio103/ParaSites2004/Trypanosomiasis/morphology.htm).
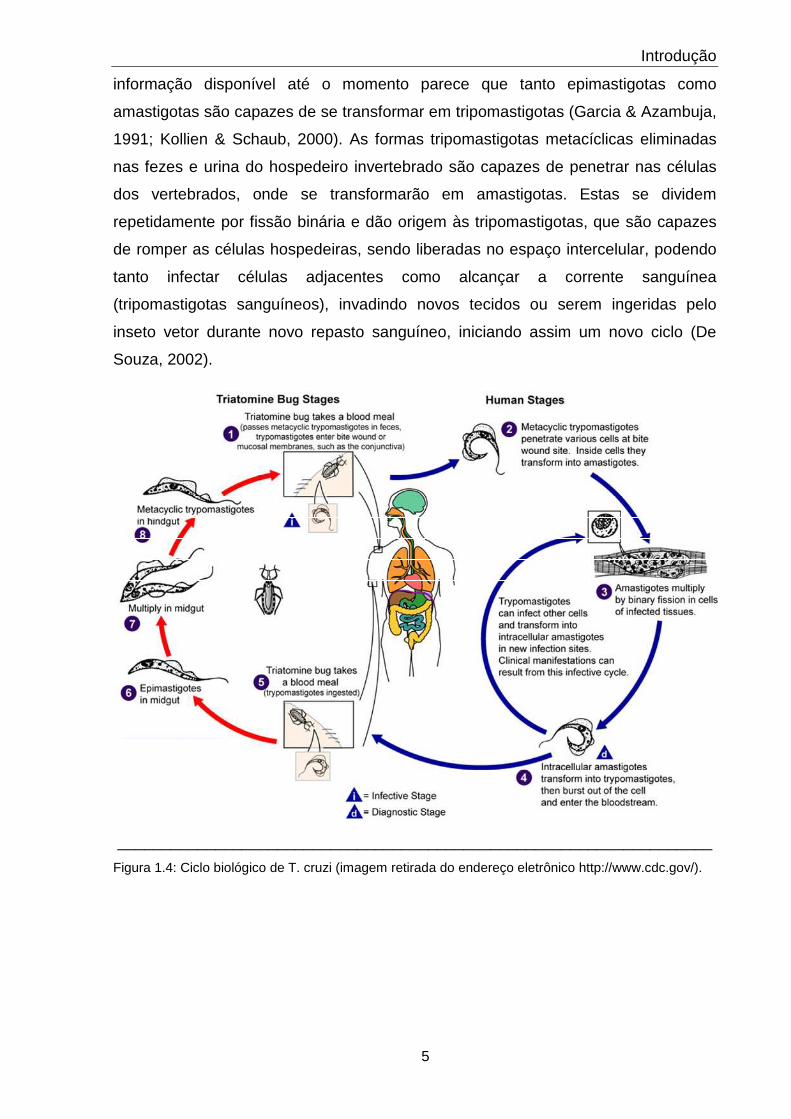
O ciclo biológico (figura 1.4) começa quando o hospedeiro invertebrado se
alimenta do sangue do hospedeiro vertebrado. Os hospedeiros invertebrados são
membros das famílias Hemiptera e Reduvidae, como
infestans, Panstrongylus megistus
formas tripomastigotas presentes
pelo inseto. Sabe-se que é
tripomastigotas sanguíneas se transforma em epimastigotas e em formas
arredondadas. No intestino, as epimastigotas se dividem repetidam
processo denominado fissão binária. Essas formas podem se prender às células
intestinais através de hemidesmossomas, e no reto, uma certa parte das
epimastigotas se transforma em tripomastigotas metacíclicas, que são eliminadas
com as fezes e urina sendo capazes de infectar os hospedeiros vertebrados
& Azambuja, 1991; Zeledon, 1997; Kollien & Schaub, 2000)
Tem sido demonstrado que formas tripomastigotas podem se transformar em
formas arredondadas que possuem um flagelo livre. Esta forma, que aparece no
estômago do vetor invertebrado, é capaz de se transformar tanto em pequenas
epimastigotas que iniciam um pr
longas epimastigotas que se movem para a região mais posterior do trato digestivo
do inseto. Aparentemente essas formas longas de epimastigotas são incapazes de
se dividirem. Alguns dias após a ingestão do sang
transformarem em tripomastigotas são encontradas no reto. Com base na
4
células tissulares e no sangue de hospedeiros vertebrados, (ii) no intestino posterior,
nas fezes, e na urina do hospedeiro invertebrado, (iii) na fase estacionária de
crescimento de culturas axênicas, e (iv) na fase líquida de culturas celulares. Essa
forma é incapaz de se dividir.
___________________________________________________________________
Figura 1.3: Formas evolutivas de T. cruzi. a. tripomastigotas sanguíneas (imagem retirada de , b. epimastigotas (imagem retirada do endereço eletrônico
z.br/~ccs/arquivosite/estetica/chagas.htm) e c. amastigotas (imagem retirada de http://www.stanford.edu/class/humbio103/ParaSites2004/Trypanosomiasis/morphology.htm).
O ciclo biológico (figura 1.4) começa quando o hospedeiro invertebrado se
angue do hospedeiro vertebrado. Os hospedeiros invertebrados são
membros das famílias Hemiptera e Reduvidae, como Rhodinus prolixus
Panstrongylus megistus, entre outros. Durante o repasto sanguíneo, as
formas tripomastigotas presentes no sangue do hospedeiro vertebrado são ingeridas
se que é no estômago do inseto onde a maior parte das
tripomastigotas sanguíneas se transforma em epimastigotas e em formas
arredondadas. No intestino, as epimastigotas se dividem repetidam
processo denominado fissão binária. Essas formas podem se prender às células
intestinais através de hemidesmossomas, e no reto, uma certa parte das
epimastigotas se transforma em tripomastigotas metacíclicas, que são eliminadas
rina sendo capazes de infectar os hospedeiros vertebrados
& Azambuja, 1991; Zeledon, 1997; Kollien & Schaub, 2000).
Tem sido demonstrado que formas tripomastigotas podem se transformar em
formas arredondadas que possuem um flagelo livre. Esta forma, que aparece no
estômago do vetor invertebrado, é capaz de se transformar tanto em pequenas
epimastigotas que iniciam um processo de multiplicação no intestino, como em
longas epimastigotas que se movem para a região mais posterior do trato digestivo
do inseto. Aparentemente essas formas longas de epimastigotas são incapazes de
se dividirem. Alguns dias após a ingestão do sangue, as amastigotas capazes de se
transformarem em tripomastigotas são encontradas no reto. Com base na
Introdução
gue de hospedeiros vertebrados, (ii) no intestino posterior,
nas fezes, e na urina do hospedeiro invertebrado, (iii) na fase estacionária de
crescimento de culturas axênicas, e (iv) na fase líquida de culturas celulares. Essa
___________________________________________________________________
tripomastigotas sanguíneas (imagem retirada de epimastigotas (imagem retirada do endereço eletrônico
amastigotas (imagem retirada de http://www.stanford.edu/class/humbio103/ParaSites2004/Trypanosomiasis/morphology.htm).
O ciclo biológico (figura 1.4) começa quando o hospedeiro invertebrado se
angue do hospedeiro vertebrado. Os hospedeiros invertebrados são
Rhodinus prolixus, Triatoma
, entre outros. Durante o repasto sanguíneo, as
no sangue do hospedeiro vertebrado são ingeridas
inseto onde a maior parte das
tripomastigotas sanguíneas se transforma em epimastigotas e em formas
arredondadas. No intestino, as epimastigotas se dividem repetidamente por um
processo denominado fissão binária. Essas formas podem se prender às células
intestinais através de hemidesmossomas, e no reto, uma certa parte das
epimastigotas se transforma em tripomastigotas metacíclicas, que são eliminadas
rina sendo capazes de infectar os hospedeiros vertebrados (Garcia
Tem sido demonstrado que formas tripomastigotas podem se transformar em
formas arredondadas que possuem um flagelo livre. Esta forma, que aparece no
estômago do vetor invertebrado, é capaz de se transformar tanto em pequenas
ocesso de multiplicação no intestino, como em
longas epimastigotas que se movem para a região mais posterior do trato digestivo
do inseto. Aparentemente essas formas longas de epimastigotas são incapazes de
ue, as amastigotas capazes de se
transformarem em tripomastigotas são encontradas no reto. Com base na
Introdução
5
informação disponível até o momento parece que tanto epimastigotas como
amastigotas são capazes de se transformar em tripomastigotas (Garcia & Azambuja,
1991; Kollien & Schaub, 2000). As formas tripomastigotas metacíclicas eliminadas
nas fezes e urina do hospedeiro invertebrado são capazes de penetrar nas células
dos vertebrados, onde se transformarão em amastigotas. Estas se dividem
repetidamente por fissão binária e dão origem às tripomastigotas, que são capazes
de romper as células hospedeiras, sendo liberadas no espaço intercelular, podendo
tanto infectar células adjacentes como alcançar a corrente sanguínea
(tripomastigotas sanguíneos), invadindo novos tecidos ou serem ingeridas pelo
inseto vetor durante novo repasto sanguíneo, iniciando assim um novo ciclo (De
Souza, 2002).
___________________________________________________________________
Figura 1.4: Ciclo biológico de T. cruzi (imagem retirada do endereço eletrônico http://www.cdc.gov/).

Introdução
6
1.3. Fator Transformador de Crescimento beta (TGF- β)
1.3.1. A superfamília TGF- β
O Fator Transformador de Crescimento beta (“Transforming growth factor
beta” - TGF-β) pertence a uma superfamília que compreende um grande número de
fatores de crescimento polipeptídicos estruturalmente relacionados, capazes de
regular uma vasta série de processos celulares, como proliferação celular,
determinação de linhagem, diferenciação, motilidade, adesão e morte (Lawrence,
1995; Massague, 1998).
A superfamília TGF-β (figura 1.5) engloba uma ampla variedade de proteínas
sinalizadoras, incluindo isoformas do TGF-β, proteínas morfogênicas do osso
(BMPs), fatores de crescimento e diferenciação (GDFs), activinas, inibinas e
substância inibidora Mülleriana (Roberts & Sporn, 1993; Piek et al., 1999; Moustakas
& Heldin, 2005). Os ligantes TGF-β apresentam moléculas relacionadas estrutural e
funcionalmente, codificadas por diferentes genes, e suas isoformas são designadas
TGF-β, TGF-β2 e TGF-β3 (Burt & Law, 1994).

Introdução
7
Figura 1.5: Superfamília TGF-β (imagem retirada do endereço eletrônico http://www.rndsystems.com/dam_public/5895.pdf).
A atividade biológica do TGF-β depende de sua interação com receptores
específicos que transduzem o sinal bioquímico, resultando em um efeito biológico.
Somente a molécula ativa é capaz de se ligar ao seu receptor, e este mecanismo
representa um sistema típico de interação ligante-receptor, seguido de transdução
de sinal e culminando em um efeito final.

Introdução
8
O TGF-β é sintetizado sob uma forma latente contendo domínios C- e N-
terminais, que são clivados intracelularmente antes de serem secretados pelas
células, mas mantêm-se associados por interações eletrostáticas como um
complexo não-covalente (Murphy-Ullrich & Poczatek, 2000). No entanto, para
exercer seu efeito biológico, a forma latente precisa sofrer uma alteração na sua
conformação passando a um estado ativado. A forma latente é produzida por
diferentes tipos celulares, entre eles, macrófagos, plaquetas, fibroblastos e células
do baço. A síntese de TGF-β tem caráter heterogêneo: células T e monócitos
sintetizam principalmente TGF-β, e células do sistema nervoso central sintetizam
altos níveis de TGF-β3 (Moustakas et al., 2002). A maioria das células em cultura
produz TGF-β latente. No entanto, tanto células T ativadas quanto células
fagocíticas mononucleares ativadas podem secretar a forma ativa de TGF-β (Chen
et al., 2001).
Como os outros membros desta superfamília, a forma latente é uma proteína
homodimérica, cujas cadeias de aproximadamente 12,5 kDa são mantidas por
pontes dissulfeto. A latência é conferida pelo peptídeo de 75 kDa, denominado LAP
(“latency associated peptide”), localizado na região N-terminal da sequência do
precursor e que permanece ligado não-covalentemente ao TGF-β após sua
secreção (Massague, 1990; Lyons & Moses, 1990; Taipale et al., 1996).
A forma latente pode ser ativada por diferentes fatores incluindo proteases,
endoglicosidases e mudanças de pH ou temperatura (Hall et al., 1992). É possível
que, em condições fisiológicas (in vivo), um mecanismo em particular de ativação de
TGF-β latente seja dominante em relação aos outros.
1.3.2. Receptores de TGF- β
Uma vez ativado, o TGF-β pode então ser reconhecido pelos seus receptores
específicos de superfície, desencadeando o processo de sinalização intracelular.
TGF-β apresenta três receptores de superfície, sendo eles denominados: receptor
para TGF-β tipo I (TβRI), receptor para TGF-β tipo II (TβRII) e receptor para TGF-β
tipo III (TβRIII), que existem em virtualmente todos os tipos celulares estudados. Os
receptores do tipo I (53 kDa) e do tipo II (75 kDa) são serina/treonina quinase
transmembranares. Após a ligação ao TGF-β, TβRI é fosforilado pelo TβRII, que
detém as funções de sinalização, sendo mediador da maioria dos efeitos biológicos

Introdução
9
de TGF-β (Massague, 1998; Lijnen et al., 2000). TβRII é constitutivamente
fosforilado. Em mamíferos, cinco receptores do tipo II e sete do tipo I foram
identificados como ligantes da superfamília de TGF-β (Lutz & Knaus, 2002). Todos
são receptores transmembranares que contém um domínio intracelular
serina/treonina quinase. Além do TβRI, já foram descritos outros receptores de tipo I,
como ALK1 e ALK2 que transmitem a sinalização gerada pela ligação ao TGF-β
(Massague, 1998).
1.3.3. Via de sinalização clássica por TGF- β
A ligação inicial de TGF-β ativo ao TβRII, é seguida pelo recrutamento de
TβRI, formando um complexo heterotetramérico (Massague, 1998; Lutz & Knaus,
2002). TβRI é transfosforilado na sua região justamembranar, rica em glicina e
serina, denominada GS-Box. Esta fosforilação é mediada por TβRII, o que leva a
ativação de TβRI. Esta ativação não é devida a um aumento da atividade quinase,
mas é baseada na criação de um sítio de ligação para as proteínas Smad, que
representam o substrato para TβRI (Lutz & Knaus, 2002).
A ligação do TGF-β aos seus receptores de superfície não resulta apenas na
sinalização, mas também na endocitose do ligante associado ao receptor, que são
direcionados intracelularmente para a via de degradação nos compartimentos
tardios da via endocítica (Massague, 1998; Lutz & Knaus, 2002). O mecanismo
preciso da endocitose do receptor de TGF-β ainda é controverso. Alguns grupos
sugerem que esta internalização ocorre por endocitose mediada por clatrina (Anders
et al., 1997), outros por endocitose mediada por receptor independente de clatrina
ou ainda pela via das cavéolas (Zwaagstra et al., 2001). O receptor pode ainda,
dependendo do estímulo externo, retornar à superfície celular e participar de uma
nova cascata de sinalização intracelular (Mitchell et al., 2004).
Na via de sinalização clássica de TGF-β (figura 1.6) participam as proteínas
Smad, que compõem uma família de fatores de transcrição, sendo os substratos
para o receptor de tipo I (TβRI). Essas proteínas apresentam peso molecular entre
42 e 60 kDa e podem ser divididas em três sub-famílias: (i) Smads ativadas pelo
receptor (R- Smad) incluindo Smads ativadas por BMP (Smad 1, 5 e 8) e por TGF-β
(Smad 2 e 3); (ii) o mediador comum Smad4 (Co- Smad), e (iii) as Smad inibitórias,
Smad 6 e 7 (I-Smad) (Massague, 1998). TβRI causa a fosforilação das R-Smads no

Introdução
10
motivo SSXS da porção C-terminal que é conservado entre todas as R-Smads,
causando a dissociação do receptor e a formação de um sítio âncora para Smad 4,
formando então um complexo heteromérico. O complexo de Smads é translocado
para o núcleo, onde se associa a co-fatores e co-moduladores que se ligam ao DNA
para ativar a transcrição de determinados genes responsivos a TGF-β. A escolha
dos genes alvo é determinada pela composição do complexo de transcrição formado
(Massague & Chen, 2000; Lutz & Knaus, 2002).
Esta via clássica das Smads é conservada evolutivamente e é regulada por
diversos eventos de fosforilação, transporte núcleo-citoplasmático, degradação
proteassomal mediada por ubiquitina e, finalmente, por Smads inibitórias ((I)-Smads)
(Shi & Massague, 2003). As proteínas Smad são os únicos substratos conhecidos
para TβRI capazes de promover funções de sinalização celular em resposta a TGF-
β. Embora outras proteínas também sejam capazes de interagir com o receptor e
mediar o processo de sinalização, a característica de transmissão de sinal
diretamente do receptor para a maquinaria transcricional é única das Smads (Piek et
al., 1999). Por isso, a via clássica de Smads foi por muito tempo considerada como a
única via responsiva a TGF-β.
As proteínas Smad inibitórias são induzidas pela sinalização por Smad,
acumuladas no núcleo e podem ser então exportadas do núcleo após estímulo das
células por TGF-β ou BMP. Em seguida, elas se ligam aos receptores do tipo I e
exercem feedback negativa, através do bloqueio da fosforilação de proteínas R-
Smads e da formação do complexo R-Smad–Smad 4, estimulando a defosforilação
do receptor pelo recrutamento de fosfatases e promovendo a ubiquitinação e
degradação lisossomal do receptor (Moustakas & Heldin, 2005).
Figura 1.6: via clássica de sinalização por TGF-β (imagem retirada de Derynck & Zhang, 2003).

Introdução
11
1.3.4. Vias alternativas de sinalização por TGF- β
A via de Smads representada acima é de extrema importância para a
execução precisa de programas tissulares durante o desenvolvimento animal. Tem
sido firmemente estabelecido que as vias das Smads são mediadoras centrais da
transdução de sinais dos receptores dos membros da superfamília de TGF-β ao
núcleo (Barolo et al., 2002; Waite & Eng, 2003). Contudo, novas evidências
bioquímicas e de desenvolvimento apóiam a idéia de que, alternativamente, vias
não-dependentes de Smads também participam na sinalização por TGF-β
(Moustakas & Heldin, 2005).
A sinalização por vias independentes de Smads (figura 1.7) possui três
mecanismos gerais que contribuem para as respostas fisiológicas em resposta a
TGF-β: (1) vias de sinalização independentes de Smads modificam diretamente (ex.:
fosforilam) as Smads, e assim, modulam a atividade de efetores centrais; (2) as
proteínas Smad interagem e modulam diretamente a atividade de outras proteínas
sinalizadoras (ex.: quinases), deste modo, transmitindo sinais a outras vias; (3) os
receptores de TGF-β interagem diretamente com proteínas não-Smad ou fosforilam
as mesmas, iniciando uma sinalização paralela que colabora com a via das Smads
na produção de respostas fisiológicas.
Assim, transdutores de sinal não-Smad sob o controle de TGF-β fornecem
regulação quantitativa da via de sinalização e servem como pontos de interação
cruzada com outras importantes vias de sinalização, como as da tirosina quinase,
receptores acoplados a proteína G ou outros receptores de citocinas. A seguir,
daremos um breve relato sobre as vias alternativas já descritas na literatura.
Algumas evidências científicas relatam a associação de TGF-β a outras vias
de sinalização, incluindo as vias MAPKs, ERK, JNK, p38, PI3K, fosfatases PP2A e
ainda membros da família Rho. Algumas dessas vias regulam a ativação de Smads,
como descrito anteriormente, mas outras podem induzir respostas não relacionadas
à transcrição (Derynck & Zhang, 2003). A ativação dessas vias com cinética lenta
pode ser resultado de respostas geradas a partir da transcrição induzida pelas
Smads, entretanto, a rápida ativação (5 a 15 minutos), sugere independência da
transcrição por essas proteínas (Massague & Wotton, 2000).
Estudos científicos usando células deficientes em Smad 4 apóiam a
possibilidade da ativação da via MAPK que é independente de Smads (Engel et al.,

Introdução
12
1999). Além disso, receptores de TGF-β do tipo I mutados, deficientes na ativação
de Smads, ativam a sinalização por p38MAPK em resposta a TGF-β (Yu et al.,
2000). Os mecanismos de ativação de ERK, JNK ou p38MAPK por TGF-β e suas
consequências biológicas ainda estão pouco caracterizadas. A ativação induzida por
TGF-β das vias de ERK e JNK pode resultar na fosforilação de Smads e regular a
ativação das mesmas (de Caestecker et al., 1998); (Engel et al., 1999); (Kretzschmar
et al., 1999); (Funaba et al., 2002). Já a ativação da sinalização de Ras/Erk MAPK
por TGF-β pode induzir a expressão de TGF-β1, amplificando assim a resposta e
induzindo respostas secundárias à TGF-β (Yue & Mulder, 2000). A ativação das vias
MAPK por TGF-β pode também afetar respostas de transcrição através de efeitos
diretos sobre os fatores de transcrição que interagem com as proteínas Smad
(Massague & Chen, 2000; Itoh et al., 2000; Moustakas et al., 2001).
Dependendo da linhagem celular, outras vias de sinalização podem ser
induzidas. TGF-β pode rapidamente ativar GTPases semelhantes a Rho (“Rho-like
GTPases”), incluindo RhoA, Rac e Cdc42, apesar da ativação atrasada de RhoA e
Cdc42 (devido à síntese de novo de proteínas) ter sido também observada
(Bhowmick et al., 2001; Bakin et al., 2002; Edlund et al., 2002).
Figura 1.7: vias alternativas de sinalização por TGF-β (imagem retirada de Derynck & Zhang, 2003).

Introdução
13
1.3.5. Genes e fatores de transcrição responsivos a TGF-β
TGF-β ativa a transcrição por meio de interações físicas e da ajuda funcional
de Smads ligadoras de DNA com sequências específicas de fatores de transcrição e
com os co-ativadores CBP e p300 (Derynck & Zhang, 2003). Uma vez que o
complexo de Smad se liga ao DNA, ele pode controlar a transcrição de genes alvo
alterando a estrutura do nucleossoma, e assim, remodelando o molde de cromatina.
Através de seu domínio MH2, as Smads podem se ligar aos co-ativadores
p300/CBP, que possuem atividade histona acetil transferase, e também aos co-
repressores TGIF, c-ski e SnoN, que recrutam histonas desacetilases (Derynck et
al., 1998; Massague & Wotton, 2000). A atividade transcricional do domínio MH2 da
Smad se manifesta em fusões com o domínio de ligação ao DNA Gal4p, e exige a
presença de uma co-Smad. Smads e co-Smads podem recrutar o conjunto
necessário de co-ativadores ou co-repressores para orquestrar uma resposta
transcricional. (Massague & Chen, 2000).
O número de fatores de transcrição que se ligam ao DNA, com os quais as
Smads podem interagir funcionalmente é impressionante. Muitos genes são ativados
em resposta ao TGF-β, enquanto outros são transcricionalmente reprimidos. Assim,
dependendo do fator de transcrição com o qual interagem, do promotor e do
contexto intracelular, mecanismos distintos levam a sinalização mediada por Smad e
determinam se essas proteínas irão ativar ou reprimir a transcrição.
TGF-β inibe a progressão do ciclo celular através da regulação da transcrição
dos reguladores do ciclo celular. Entre eles, c-Myc e membros da família Id são
regulados negativamente por TGF-β. Em células onde a expressão do c-Myc é
regulada negativamente por TGF-β, a Smad3 reprime a transcrição de c-Myc em
associação com os fatores de transcrição E2F4 e E2F5, e o co-repressor p107. Este
complexo é pré-montado no citoplasma e, em resposta ao TGF-β, é translocado
para o núcleo, onde, em associação com a Smad4, se liga a um sítio de ligação
E2F-Smad no promotor c-Myc e reprime a expressão de c-Myc. Na regulação
negativa de Id1, a Smad3 ativada por TGF-β induz diretamente a expressão de
ATF3, e então, a Smad3 junto a ATF3 formam um complexo que reprime o promotor
Id1. Da mesma forma, SIP1 induzida por TGF-β, regula negativamente a expressão
de E-caderina. TGF-β também inibe a diferenciação de mioblastos, osteoblastos e
adipócitos através da repressão funcional de fatores de transcrição-chave que
conduzem essas vias de diferenciação. Smad3 reprime a transcrição por

Introdução
14
Runx2/CBFA1 na diferenciação dos osteoblastos; MyoD e outros fatores de
transcrição miogênicos básicos hélice-alça-hélice nos mioblastos e CCAAT/proteínas
estimuladoras de ligação na diferenciação dos adipócitos. A repressão ou ativação
da transcrição pelas Smads depende também do tipo de célula e da sequência
promotora a qual se ligam (Derynck & Zhang, 2003).
TGF-β pode também causar a rápida inibição da transcrição de genes, como
o da fosfatase ativadora de CdK Cdc25A (Iavarone & Massague, 1997), mediando
efeitos anti-proliferativos. A ativação transcricional por TGF-β da PAI-1 (Chang &
Goldberg, 1995), receptores de ácido retinóico (Chen et al., 1996), α2 de colágeno(I)
e outros genes (Chang & Goldberg, 1995) parece precisar da atividade da AP-1.
Além disso, um repressor contendo Fos (“Fos-containing repressor”) foi relacionado
à regulação negativa da protease secretora transina/estromalisina por TGF-β (Kerr
et al., 1990). Não é certo que as Smads participem em todas, ou até mesmo na
maioria das respostas gênicas por TGF-β (Massague, 1998).
Vários genes responsivos a TGF-β têm sido descritos, mas apenas uma
fração desses tem as características de uma resposta transcricional imediata.
p15Ink4b e p21Cip1 são inibidores quinase dependentes de ciclina, cuja rápida indução
em resposta a TGF-β medeia a parada do ciclo celular. “Clusters” de sítios do tipo
Sp1 próximos ao sítio de início da transcrição de p15Ink4b e p21Cip1 contam como
regiões responsivas a TGF-β em ensaios de gene repórter.
A expressão estimulada por TGF-β de colágenos intersticiais e outras
proteínas da matriz extracelular revelam um papel importante do TGF-β nos
processos de desenvolvimento e regenerativos (Massague, 1990; Roberts et al.,
1990; Roberts & Sporn, 1993). As regiões responsivas a TGF-β dos genes que
codificam as proteínas da matriz extracelular, como α1 do colágeno (I) (Ritzenthaler
et al., 1993), α2 de colágeno (I) (Rossi et al., 1988; Inagaki et al., 1994), inibidor do
ativador de plasminogênio do tipo 1 (PAI-1) (Keeton et al., 1991; Riccio et al., 1992),
elastina (Marigo et al., 1994), e perlecan (Iozzo et al., 1997) se assemelham aos
sítios Sp1 ou aos sítios CTF/NF-I.

Introdução
15
1.3.6. Papel do TGF- β na Doença de Chagas
O TGF-β foi inicialmente descrito como fator transformador de crescimento e
proliferação de células tumorais (Todaro et al., 1980). No entanto, estudos
posteriores indicaram que ele apresentava uma função pleiotrópica, podendo
desempenhar atividades proliferativas e anti-proliferativas, sendo capaz tanto de
inibir como estimular o crescimento de um mesmo tipo celular, dependendo das
condições da cultura e do grau de confluência das células (Massague, 1998).
Além da ação na proliferação celular, o TGF-β é uma das principais citocinas
envolvidas na regulação da formação e degradação de matriz extracelular
(Massague, 1998), induzindo a expressão de vários componentes de matriz, como
fibronectina, laminina, colágeno, vitronectina e trombospondina. A inibição da
degradação de matriz é devida à diminuição da atividade proteolítica pericelular
(Taipale et al., 1996).
A produção exacerbada de TGF-β na infecção pelo T. cruzi foi inicialmente
descrita por Silva e colaboradores em 1991. Este estudo demonstrou que células do
baço de camundongo apresentam no oitavo dia de cultura in vitro um aumento na
produção de TGF-β, variando aproximadamente de 2 ng/ml na cultura não infectada
para 5 ng/ml na cultura infectada por T. cruzi. Além disso, o TGF-β se mostrou como
um potente inibidor de INF-γ, sendo responsável pelo aumento da replicação de
T.cruzi dentro dos macrófagos.
Estudos demonstraram o envolvimento do TGF-β na cardiopatia chagásica
aguda e crônica, com exacerbação de seus níveis plasmáticos e da ativação da sua
via de sinalização celular como aspectos peculiares de pacientes nos estágios mais
avançados da doença (Araujo-Jorge et al., 2002; Waghabi et al., 2002).
A cardiopatia chagásica está geralmente associada ao aumento do coração
através de uma fibrose exacerbada e disfunções incluindo arritmias, distúrbios de
condução elétrica e falência congestiva do coração (Higuchi et al., 1999). A fibrose é
uma das manifestações mais significativas da cardiopatia chagásica crônica (CCC) e
encontra-se associada com infiltrados inflamatórios e cardiomiócitos em
degeneração (Andrade et al., 1989; Rossi & Bestetti, 1995; Rossi, 2001). Além de
sua participação na produção de proteínas da matriz extracelular e conseqüente
fibrose, TGF-β pode modular outras manifestações cardíacas, incluindo a
proliferação e morte dos cardiomiócitos. A sinalização por TGF-β está relacionada a

Introdução
16
resposta de hipertrofia dos cardiomiócitos à estimulação pelo receptor β1-
adrenérgico (Rosenkranz, 2004), a qual pode amplificar o crescimento hipertrófico
após o infarto do miocárdio e causar a transição entre uma hipertrofia adaptativa e
uma hipertrofia descompensada.
1.3.7. Papel de TGF-β na biologia do T. cruzi
Diversos estudos foram desenvolvidos demonstrando claramente que TGF-β
desempenha várias funções não somente na progressão da doença de Chagas,
como também no controle em diferentes etapas do ciclo de vida e infectividade de T.
cruzi (Waghabi et al., 2002; Waghabi et al., 2005a; Waghabi et al., 2005b; Waghabi
et al., 2007; Araujo-Jorge et al., 2008).
Sabe-se que o T. cruzi é capaz de ativar diretamente TGF-β latente (Waghabi
et al., 2005b), e com isto, dispara sua via de sinalização dependente de Smad,
sendo a ativação desta via necessária durante os eventos de invasão à célula
hospedeira (Ming et al., 1995; Hall & Pereira, 2000). Estudos com cardiomiócitos
infectados in vitro por T. cruzi demonstram que formas amastigotas de T. cruzi são
capazes de captar e internalizar TGF-β do hospedeiro, estocando-o, o que sugere
um novo papel de TGF-β no ciclo celular do parasito: seu uso no momento de
sinalizar a parada da proliferação e iniciar a diferenciação para formas
tripomastigotas (Waghabi et al., 2005a), podendo refletir uma dependência do
parasito por uma molécula da célula hospedeira que pode ser usada para regular
seu próprio ciclo intracelular.
Esses resultados também sugerem a presença de receptores para TGF-β na
superfície celular de T. cruzi, assim como a existência de uma via de sinalização
intracelular capaz de transduzir os efeitos de TGF-β. Genes ortólogos para os
receptores serina/treonina quinase canônicos (TβRI e TβRII) e proteínas Smads já
foram identificadas no parasito helminto Schistosoma mansoni, mas não foram
encontradas após análises “in silico” do genoma de T. cruzi (Waghabi et al., 2005a).
Sendo assim, espera-se que neste microorganismo deva ocorrer uma via alternativa
de sinalização por TGF-β e que seus possíveis receptores celulares sejam distintos
dos já descritos para mamíferos.

Introdução
17
Como descrito acima, muitas vias de sinalização Smad-independentes são
conhecidas por serem ativadas ou moduladas por TGF-β em células eucarióticas.
Entre estas estão incluídas Jun-kinase, p38MAP-kinase, Ras/MEK/ERK, Rho-A/
p160ROCK e PP2A/S6kinase. Alguns estudos mostraram a presença de homólogos
de Ras (Razani et al., 2001), Rho (Dong et al., 2000) e ERK (Sasaki et al., 2001;
Tang et al., 2003) em T. cruzi e T. brucei, sugerindo que pelo menos algumas
dessas vias alternativas de TGF-β possam ser funcionais nesses parasitos, embora
os mecanismos envolvidos nesta ativação, assim como suas consequências
biológicas, ainda não estejam elucidados (Derynck & Zhang, 2003).
O novo papel do TGF-β da célula hospedeira descrito nesses trabalhos
acrescenta complexidade para a biologia do T. cruzi, com implicações adicionais
para esta citocina na Doença de Chagas. De forma geral, o TGF-β parece: (1) ser
ativado na superfície da célula hospedeira a partir de TGF-β latente, mecanismo
este mediado pelo parasito, (2) induzir sinalização via receptores de TGF-β da célula
hospedeira, favorecendo assim a invasão celular, (3) ser captado e estocado
intracelularmente pelos parasitos, participando do processo de diferenciação de
formas amastigotas para tripomastigotas, e (4) desencadear a fibrose na
cardiomiopatia chagásica.
Outro estudo recente mostra que o bloqueio da via de sinalização intracelular
de TGF-β nas células hospedeiras, pela ação de um inibidor do receptor de TGF-β
tipo I (TbRI), SB-431542, é capaz de inibir a invasão de T. cruzi e induzir um retardo
na proliferação de formas amastigotas e na diferenciação destas formas em
tripomastigotas ao final do ciclo, mostrando mais uma vez o envolvimento de TGF-β
no ciclo de vida de T. cruzi (Waghabi et al., 2007). A reversão da capacidade do
TGF-β em desenvolver fibrose no miocárdio infectado pelo T. cruzi foi testada in vivo
utilizando-se o modelo murino através do uso de SB-431542. Foi observada uma
redução dos níveis de parasitemia, das taxas de mortalidade, dos infiltrados
inflamatórios e carga parasitária no coração dos animais infectados tratados na fase
aguda da doença (Waghabi et al., 2009).
A atividade final de TGF-β sempre parece ser dependente de eventos de
sinalização celular, ativação e degradação de genes responsivos e, em todos estes
aspectos, a fosforilação de proteínas ocorre como processo-chave. Daremos a
seguir um breve relato sobre fosforilação de proteínas.

Introdução
18
1.4. Modificações pós-traducionais
Após serem sintetizadas, as proteínas podem sofrer alterações, denominadas
modificações pós-traducionais (“post translational modifications” - PTM). As PTMs
alteram as ligações covalentes, modificando as propriedades de uma proteína,
através de clivagem proteolítica. As PTMs de uma proteína podem determinar seu
estado de atividade, localização, reciclagem (“turnover”) e interação com outras
proteínas, além de alterarem as estruturas terciárias e quaternárias da proteína
modificada (Mann & Jensen, 2003).
Muitas PTMs foram descobertas durante estudos de proteínas individuais com
a ajuda de técnicas moleculares clássicas, como a deleção de aminoácidos
carreadores da modificação. Para a análise dessas modificações em uma única
proteína é interessante primeiro obtê-la pura, em grandes quantidades, o que
normalmente é feito pelo uso dos métodos de purificação cromatográfica,
precipitação com anticorpos específicos ou ambas as técnicas combinadas. A
análise de modificações é, em geral, feita pela comparação da sequência de
aminoácidos obtida experimentalmente para a proteína possivelmente modificada
com a sequência de aminoácidos previamente conhecida da proteína intacta. Desta
forma, o primeiro passo é a identificação da proteína a ser estudada, o que pode ser
feito com alta sensibilidade pelo reconhecimento de anticorpos (western blot) ou por
técnicas de espectrometria de massas (MS) (Mann & Jensen, 2003).
A espectrometria de massas tem continuamente aumentado a sensibilidade
de detecção das PTMs. A MS mede a razão massa/carga, fornecendo o peso
molecular e o padrão de fragmentação de peptídeos derivados de proteínas. Dessa
forma, a MS constitui um método geral para a identificação de modificações que
alteram a massa molecular de uma proteína. Por essa razão, tem sido vastamente
utilizada para mapear a estrutura primária completa de proteínas individuais (Carr et
al., 1989; Ling et al., 1991; Roepstorff, 1997).
O novo campo da Proteômica é também promissor na identificação de PTMs.
A técnica tem sido bastante valiosa na resolução e identificação de grandes
conjuntos de proteínas presentes em organelas ou em todo um organismo em
estudo, a partir de um único experimento. A eletroforese bidimensional separa as
populações de proteínas com base em suas cargas e massas moleculares. A

Introdução
19
técnica oferece resolução suficiente para separar diretamente os diferentes estados
modificados de cada proteína presente na amostra (Mann & Jensen, 2003).
A intensidade da coloração dos “spots” protéicos presentes nos géis
bidimensionais reflete fortemente a quantidade da proteína contida no gel,
especialmente para proteínas similares. Dessa forma, o método é capaz de gerar
informações sobre a proporção relativa dos vários estados modificados de uma
proteína. Entre algumas das principais modificações pós-traducionais temos:
acetilação, metilação, glicosilação, acilação, deamidação, ubiquitinação e
fosforilação (Mann & Jensen, 2003).
1.4.1. Fosforilação de Proteínas: Aspectos Gerais
Desde o isolamento da fosfoserina em 1932, descrita primeiramente como
ácido fosfórico serina, a fosforilação de proteínas tornou-se uma das modificações
pós-traducionais mais estudadas e biologicamente relevantes. A fosforilação é uma
modificação reversível, que afeta o enovelamento e, consequentemente, a atividade
de proteínas, regulando funções essenciais como divisão celular, transdução de
sinal, atividade enzimática, entre outras (Paradela & Albar, 2008).
A presença ou ausência de grupamentos fosfato nas cadeias laterais de
aminoácidos, tais como serina, treonina e tirosina, é usada para modular a atividade
biológica da proteína e propagar sinais em vias e redes de sinalização celular
(Cohen, 1992; Cohen, 2000). Sendo a fosforilação um processo dinâmico, os sítios
de fosforilação não podem ser preditos a partir da análise da sequência genômica de
um organismo, requerendo medições analíticas para a verificação experimental
desses sítios de modificação. A análise de sítios de fosforilação pelo uso de
espectrometria de massas já é desafiadora quando apenas uma proteína é
analisada, e torna-se uma tarefa ainda mais difícil quando se propõe executar
medições em escala proteômica (fosfoproteômica) (Goshe, 2006).
A fosforilação e a defosforilação nos resíduos S (serina), T (treonina) e Y
(tirosina) são as modificações mais conhecidas envolvidas na ativação e inativação
reversível de atividades enzimáticas e modulação de interações moleculares em vias
de sinalização nos eucariotos (Pawson, 2002). Contudo, a fração de proteínas
fosforiladas é geralmente muito pequena in vivo (<0.1%) e a reversão desta

Introdução
20
modificação ocorre de maneira rápida. Assim, para detecção específica de
peptídeos fosforilados é indispensável o uso de inibidores de fosfatases (Seo & Lee,
2004).
1.5. Fosforilação em Trypanosoma cruzi
Com o desenvolvimento do projeto genoma de T. cruzi, estima-se que
aproximadamente 2% de seu genoma codifica para proteínas quinases, sugerindo
um importante papel regulatório para estas proteínas no controle das funções e
desenvolvimento deste parasito (Parsons et al., 2005).
Recentemente, um estudo comparativo dos quinomas de tripanossomatídeos
mostrou que T. brucei, T. cruzi e Leishmania major possuem 176, 190 e 199 genes
preditos como codificantes para proteínas quinases, respectivamente (Parsons et al.,
2005; Naula et al., 2005). Dessas quinases, aproximadamente 12% são exclusivas
dos tripanossomatídeos (Parsons et al., 2005; Naula et al., 2005), não tendo sido
ainda identificadas no genoma de outros organismos. Entre as proteínas fosfatases,
T. brucei, T. cruzi e L. major possuem 78, 86 e 88 genes, respectivamente.
Aproximadamente 40% dos genes codificantes para fosfatases são atípicos, sem
nenhum ortólogo claro identificado no genoma de outros eucariotos (Brenchley et al.,
2007). As diferenças significativas identificadas entre as quinases de T. cruzi e da
célula hospedeira sugerem que a inibição específica de enzimas do parasito pode
ser alcançada, podendo representar uma nova abordagem terapêutica no controle
da doença de Chagas.
Apesar da importância da fosforilação de proteínas em muitos processos
celulares, poucos estudos identificaram sítios de fosforilação em proteínas dos
tripanossomatídeos (Nakayasu et al., 2009). Recentemente, a análise proteômica de
formas amastigotas e promastigotas de L. donovani permitiu a identificação de 73
fosfoproteínas com diversas funções biológicas. Contudo, os sítios específicos de
fosforilação dessas proteínas ainda não foram identificados (Morales et al., 2008).
Outro estudo proteômico em L. donovani identificou 18 sítios de fosforilação
provenientes de 16 fosfopeptídeos distintos (Rosenzweig et al., 2008).

Introdução
21
1.6. Fosfoproteômica
Em geral, a análise das modificações pós-traducionais pode ser feita por um
método combinando eletroforese bidimensional e espectrometria de massas. No
caso de fosfoproteínas, o poder único de resolução da eletroforese bidimensional
permite a separação de diferentes formas modificadas de uma mesma proteína em
“spots” distintos no gel (Gianazza et al., 1989). Fosforilações heterogêneas de
proteínas geram uma série de “spots” com o mesmo peso molecular, mas com
diferentes valores de ponto isoelétrico (pI). Estes “spots” provenientes de uma
mesma proteína são visualizados como uma espécie de linha horizontal pontilhada
no gel.
As análises de modificações pós-traducionais são mais difíceis do que a
identificação direta de proteínas, pelas seguintes razões: (a) métodos altamente
sensíveis são requeridos para detecção, devido à baixa estequiometria. Por
exemplo, já que apenas 5 a 10% do substrato da proteína quinase é fosforilado, são
necessários métodos para detectar a proteína modificada em níveis muito baixos
(<5-10 fmol); (b) já que a ligação covalente entre a modificação pós-traducional e a
cadeia lateral de aminoácidos é tipicamente instável, é muitas vezes difícil manter o
peptídeo em seu estado modificado durante a preparação da amostra e
subseqüente ionização durante a espectrometria de massas, e (c) modificações pós-
traducionais são frequentemente transientes na homeostase dinâmica da natureza
(Seo & Lee, 2004).
Corantes fluorescentes foram recentemente desenvolvidos para detectar
proteínas, incluindo proteínas modificadas pós-traducionalmente, com alta
sensibilidade e linearidade. Um desses corantes se chama ProQ Diamond
(Molecular Probes) e é usado para detecção de fosfoproteínas. O ProQ Diamond
liga-se direta e especificamente a(s) molécula(s) de fosfato das fosfoproteínas em
níveis tão baixos como 1 ng e é totalmente compatível com outros corantes e com a
espectrometria de massas (Steinberg et al., 2003; Schulenberg et al., 2004; Agrawal,
2009).
Portanto, a combinação de eletroforese bidimensional com ProQ Diamond
permite a análise quantitativa de fosfoproteínas em uma escala global, sendo este
um grande avanço no campo da fosfoproteômica.

Introdução
22
1.7. Proteoma de T. cruzi
Durante o processo de diferenciação das formas evolutivas de T. cruzi
ocorrem mudanças no seu padrão de expressão protéica, como já descrito na
literatura na diferenciação das promastigotas para amastigotas em espécies de
Leishmania (Bente et al., 2003). Algumas proteínas estágio-específicas
desempenham papéis importantes na infectividade e sobrevivência, podendo ser
considerados como alvos racionais para o desenho de drogas. Com o intuito de
entender as bases moleculares da diferenciação do parasito, trabalhos anteriores se
interessaram pela identificação de proteínas específicas, expressas nas diferentes
formas do parasito (Parodi-Talice et al., 2004; Paba et al., 2004).
A abordagem proteômica é uma ferramenta valiosa para o estudo de padrões
globais de expressão gênica (Hancock et al., 1999; Tyers & Mann, 2003). Esta
metodologia tem sido empregada no estudo dos tripanossomatídeos (Paba et al.,
2004; Parodi-Talice et al., 2004; Atwood et al., 2005; Magalhaes et al., 2008; Ferella
et al., 2008; Sodre et al., 2009; Ayub et al., 2009) e sua aplicação tem se mostrado
particularmente importante, já que nestes organismos a regulação da expressão
gênica ocorre principalmente ao nível pós-transcricional (Clayton & Shapira, 2007).
Todos os genes codificantes para proteínas em tripanossomatídeos estão
organizados em grandes unidades policistrônicas de transcrição que geram RNAs
precursores policistrônicos, onde estes são então processados a RNAs
monocistrônicos pelo mecanismo de “trans-splicing”. Supõe-se que nesses
microorganismos a regulação gênica ocorra apenas através do controle da
estabilidade e/ou tradução de RNAs específicos (Graham & Barry, 1995; Vanhamme
& Pays, 1995).
Além disso, modificações pós-traducionais desempenham um papel
importante na modulação da função de proteínas nesses parasitos. Juntas, essas
considerações apóiam as análises proteômicas como uma abordagem ideal para
avaliar os níveis de expressão protéica em diferentes estágios ou sob tratamentos
específicos nos tripanossomatídeos. Além disso, uma abordagem proteômica para
T. cruzi se torna ainda mais significante dado o fato de que o projeto genoma para
este parasito já foi concluído (El-Sayed et al., 2005; Degrave et al., 2001; Luchtan et
al., 2004).

Introdução
23
Análises proteômicas em larga escala ou “high-throughput” são ainda
incompletas, uma vez que as metodologias disponíveis não possuem faixa dinâmica
suficiente para identificar e quantificar todas as proteínas expressas por um
organismo. Nos estudos realizados até o momento, estima-se que aproximadamente
50% de todo espectro possível de proteínas pode ser mapeado dentro de 67 grupos
de proteínas mais abundantes (Atwood et al., 2005). Um número maior de proteínas
de baixa abundância pode ser revelado após a depleção dessas proteínas altamente
abundantes, antes da realização da análise proteômica. Análises do proteoma de T.
cruzi revelam a operação de várias vias estágio-específicas, previamente não
documentadas e que podem ser alvos apropriados para o desenvolvimento de novas
drogas contra a Doença de Chagas. Entre os alvos mais interessantes estão as vias
propostas para geração de energia em amastigotas e epimastigotas.
Adicionalmente, a identificação das proteínas expressas em abundância em formas
tripomastigotas e amastigotas de T. cruzi forneceram uma nova fonte substancial de
candidatos para o desenvolvimento de vacinas (Atwood et al., 2005).
Possíveis novos candidatos poderiam surgir do mapeamento de vias de
transdução de sinais essenciais para sobrevivência e desenvolvimento desses
parasitos, porém ausentes em seus hospedeiros vertebrados. A fosforilação de
proteínas desempenha um papel importante em diversas vias de sinalização de T.
cruzi, e seu estudo pode constituir fonte para a detecção de novos alvos
terapêuticos contra a doença de Chagas.

Objetivos
24
2. OBJETIVOS
2.1. Objetivos Gerais
Nosso trabalho possui como principal objetivo a caracterização dos padrões
de fosforilação de proteínas de epimastigotas de T. cruzi em resposta a adição de
TGF-β ao meio de cultura, através de uma abordagem proteômica; e a identificação
de moléculas envolvidas na via de sinalização disparada por TGF-β nesse
microorganismo.
2.2. Objetivos Específicos
Para o alcance dos principais objetivos do nosso trabalho, temos como metas
específicas:
(1) Padronizar a dose ótima de indução de fosforilação nas proteínas de
epimastigotas de T. cruzi com TGF-β;
(2) Caracterizar o tempo ótimo de indução de fosforilação nas proteínas de
epimastigotas de T. cruzi com TGF-β;
(3) Analisar as diferenças entre os perfis unidimensionais dos extratos protéicos das
epimastigotas incubadas ou não nos tempos de 1, 5, 15, 30 e 60 minutos com TGF-
β a 4°C;
(4) Analisar as diferenças entre os perfis bidimensionais das epimastigotas
incubadas ou não nos tempos de 1, 5, 15, 30 e 60 minutos com TGF-β a 4°C;
(5) Identificar os “spots” referentes às proteínas que em resposta a TGF-β
apresentam alterações nos padrões de fosforilação e expressão;
(6) Observar o efeito na proliferação de epimastigotas após adição de 5 ng/ml de
TGF-β à cultura de T. cruzi.
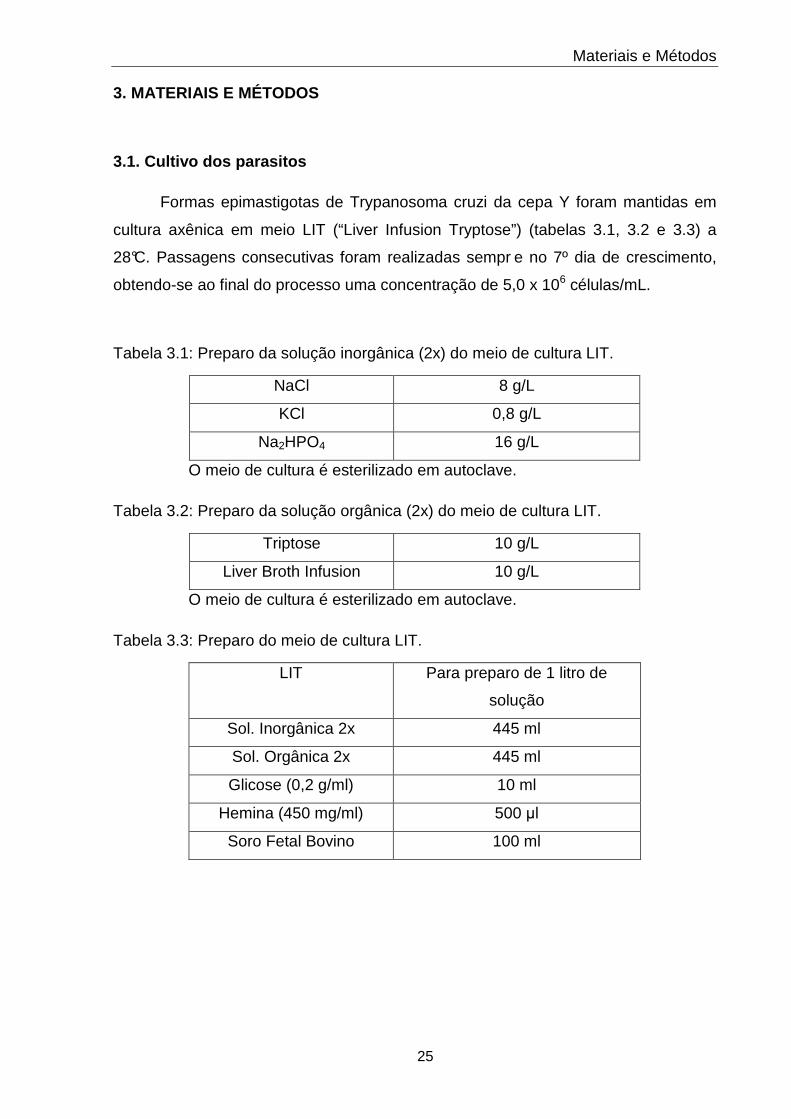
Materiais e Métodos
25
3. MATERIAIS E MÉTODOS
3.1. Cultivo dos parasitos
Formas epimastigotas de Trypanosoma cruzi da cepa Y foram mantidas em
cultura axênica em meio LIT (“Liver Infusion Tryptose”) (tabelas 3.1, 3.2 e 3.3) a
28°C. Passagens consecutivas foram realizadas sempr e no 7º dia de crescimento,
obtendo-se ao final do processo uma concentração de 5,0 x 106 células/mL.
Tabela 3.1: Preparo da solução inorgânica (2x) do meio de cultura LIT.
NaCl 8 g/L
KCl 0,8 g/L
Na2HPO4 16 g/L
O meio de cultura é esterilizado em autoclave.
Tabela 3.2: Preparo da solução orgânica (2x) do meio de cultura LIT.
Triptose 10 g/L
Liver Broth Infusion 10 g/L
O meio de cultura é esterilizado em autoclave.
Tabela 3.3: Preparo do meio de cultura LIT.
LIT Para preparo de 1 litro de
solução
Sol. Inorgânica 2x 445 ml
Sol. Orgânica 2x 445 ml
Glicose (0,2 g/ml) 10 ml
Hemina (450 mg/ml) 500 µl
Soro Fetal Bovino 100 ml

Materiais e Métodos
26
3.2. Preparo das Amostras, Lise e Extração das Prot eínas
Para o preparo de extratos de proteínas totais, os parasitos foram coletados
da cultura no quinto dia após a passagem, quando as epimastigotas encontram-se
no final da fase logarítimica de crescimento. Epimastigotas na concentração de
5x108 parasitos/ml passaram por dois ciclos de lavagem em PBS/BSA 0,1% seguido
por centrifugação a 720xg por 15 minutos, sendo finalmente ressuspensos nesta
mesma solução. Os parasitos foram então transferidos para microtubos. Para os
ensaios da dose ótima de TGF-β indutora de fosforilação de proteínas, foi
adicionado ou não TGF-β recombinante humano (R&D) nas concentrações de 1,25 /
2,5 / 5 e 10 ng/ml durante 15 minutos a 4ºC. Para avaliarmos o tempo ótimo de
indução de fosforilação, os parasitos foram incubados ou não com TGF-β (5 ng/ml)
nos tempos de 1, 5, 15, 30 ou 60 minutos a 4ºC. Após os tempos de incubação
acima determinados, os tubos foram centrifugados a 720xg por 5 minutos e lavados
duas vezes com PBS. O processo de lise dos parasitos se inicia após a retirada do
sobrenadante, quando então o sedimento é ressuspenso em 500 µl de tampão de
lise (PBS diluído 10x/coquetel de inibidores de protease 1:100 (Sigma)/coquetel de
inibidores de fosfatase 1:100 (Sigma). Em seguida, as células são completamente
lisadas com quatro ciclos de congelamento/descongelamento e armazenadas a uma
temperatura de – 70ºC.
3.3. Precipitação e Dosagem das Proteínas
As proteínas foram precipitadas com ácido tricloroacético a uma concentração
final de 17% por 10 minutos a -20ºC seguido de centrifugação por 10 minutos a uma
velocidade de 16.100xg. Após centrifugação, o sedimento foi lavado com solução
gelada de acetona/trietanolamina 1%. Em seguida, as amostras foram centrifugadas,
o sobrenadante descartado e o sedimento ressuspenso em tampão de focagem
isoelétrica (tampão IEF – 8M Uréia/2% CHAPS) contendo inibidores de fosfatase
(1:100; Sigma). A dosagem de proteínas foi realizada pelo Kit de dosagem RCDC -
BioRad (modificado de Lowry et al., 1951), segundo instruções do fabricante,
empregando-se BSA como padrão.

Materiais e Métodos
27
3.4. Eletroforese Unidimensional
Uma quantidade de 20 µg de amostra de extratos totais de epimastigotas de
T. cruzi foi misturada a tampão de amostra de proteína 1x (tabela 3.4) para um
volume final de 20 µl. A mistura foi em seguida aquecida a 100ºC por 5 minutos.
Posteriormente, as amostras foram submetidas à eletroforese em géis 12% SDS-
PAGE (tabela 3.5) de 7 centímetros em tampão de corrida Laemmli (Laemmli, 1970)
a 200 volts (tabela 3.6).
Tabela 3.4: Tampão para aplicação de amostras de proteína em gel SDS-PAGE (1X).
Tris-HCl (pH6,8) 62,5mM
SDS 2%
β-mercaptoetanol 5% (v/v)
Azul de bromofenol 0,002%
Glicerol 10% (v/v)
Tabela 3.5: Géis 12% SDS-PAGE para “strips” de 7cm.
Tris-HCl 1,5M (pH 8,8) 1,5mL
Acrilamida 30%/ Bis Acrilamida 0,8% 2,4mL
SDS 10% 60µL
APS 10% 60µL
TEMED 6µL
H2O Milli-Q 1,98mL
Tabela 3.6: Tampão de corrida Laemmli.
Tris-base 24,6 mM
Glicina 192 mM
SDS 3,48 mM

Materiais e Métodos
28
3.5. Eletroforese Bidimensional
3.5.1. Focalização Isoelétrica (IEF, 1ª dimensão)
Para a focalização isoelétrica foram utilizadas 100 µg (gel de 7 centímetros)
ou 500 µg (gel de 17 centímetros) de proteínas dos extratos de epimastigotas
obtidos como descrito acima, já solubilizadas na presença de agentes caotrópicos.
As amostras foram descongeladas a 4°C e a cada uma delas foram adicionados: o
agente redutor TBP para uma concentração final de 2 mM, 1% de anfólitos (p/v),
traços de azul de bromofenol e o volume final foi ajustado para 125 µl (géis de 7 cm)
ou 300 µl (géis de 17 cm) com tampão IEF. A mistura foi homogeneizada e mantida
à temperatura ambiente por uma hora, período suficiente para interação entre
amostra e reagentes. Ao término do período, as amostras foram colocadas em
contato por uma hora com tiras de gel (“strips” - ReadyStrip IPG Strip - BIO-RAD)
com gradiente de pH imobilizado na faixa de pH de 3 a 10 não-linear. As tiras de gel
foram colocadas cuidadosamente na cuba de focagem isoelétrica (BioRad). Após
uma hora, as tiras IPG são cobertas com 1 ml (7 cm) ou 2 ml (17 cm) de óleo mineral
e a cuba é colocada na célula de focalização isoelétrica (Protein IEF Cell - BioRad).
A reidratação ativa é realizada por 11 horas a 50 volts e após seu término, inicia-se
o programa de focalização isoelétrica apropriado, conforme instruções do fabricante,
descrito na tabela 3.7.
Tabela 3.7: Parâmetros para focalização isoelétrica.
Géis de 17 centímetros
Passo 1 : 250V – Corrente constante – 20 minutos
Passo 2 : 10000V – Corrente constante – 2,5 horas
Passo 3 : 10000V – 80000V - Rampagem Rápida
Passo 4 : 500V – Rampagem Rápida – 5 horas
Géis de 7 centímetros
Passo 1 : 250V – Corrente constante – 20 minutos
Passo 2 : 4000V – Corrente constante – 2 horas
Passo 3 : 4000V – 12500V - Rampagem Rápida
Passo 4 : 500V – Rampagem Rápida – 5 horas

Materiais e Métodos
29
3.5.2. Eletroforese em SDS/PAGE (2 a dimensão)
Uma etapa fundamental é aplicada às proteínas separadas por focalização
isoelétrica antes da corrida de segunda dimensão. O processo denominado
equilíbrio dos “strips” é responsável por: (1) manter a redução das pontes dissulfeto
e (2) alquilação dos grupos sulfidrilas resultantes dos resíduos de cisteína.
Dessa forma, ao final da focalização isoelétrica as tiras de IPG são incubadas
por 15min, sob leve agitação à temperatura ambiente no tampão de equilíbrio I
(tabela 3.8). Em seguida, o tampão de equilíbrio I é devidamente descartado e as
tiras IPG são incubadas com o tampão de equilíbrio II (tabela 3.9) por 15min à
temperatura ambiente, sob leve agitação.
Tabela 3.8: Tampão de Equilíbrio I.
Uréia 6 M
SDS 10% (p/v) 2%
Tris-HCl 1,5M pH 8,8 0,375 M
Glicerol 100% 20%
DTT 130 mM
Tabela 3.9: Tampão de Equilíbrio II.
Uréia 6 M
SDS 10% (p/v) 2%
Tris-HCl 1,5M pH 8,8 0,375 M
Glicerol 100% 20%
Iodoacetamida 135 mM
As tiras IPG são então lavadas em tampão Laemmli e aplicadas
cuidadosamente na superfície de géis 12% SDS-PAGE, onde as proteínas serão
separadas, perpendicularmente à primeira dimensão, de acordo com sua massa

Materiais e Métodos
30
molecular. Para os géis de 7cm foi utilizada uma voltagem constante de 200 volts;
para os géis de 17 cm utilizamos uma corrente de 20 mA durante 1h, e 40 mA até o
final da corrida, conforme instruções do fabricante.
3.6. Coloração dos Géis e Obtenção das Imagens
Após os ensaios de eletroforese uni- e bi-dimensionais, os géis foram corados
com corante fluorescente Pro-Q Diamond (Molecular Probes), que permite a
visualização das proteínas fosforiladas. O corante permite a detecção direta no gel
de grupamentos fosfatos ligados a resíduos de tirosina, serina ou treonina, sem a
necessidade de anticorpos ou radioisótopos. A coloração por ProQ Diamond permite
a detecção das fosfoproteínas em quantidades tão pequenas como 1 a 16 ng por
banda de géis unidimensionais. Para fosfoproteínas individuais (“spots” nos géis
bidimensionais), a força do sinal se correlaciona com o número de grupamentos
fosfatos, sendo linear por mais de três ordens de magnitude. Na seqüência, os géis
foram corados com o corante fluorescente Sypro Ruby (Molecular Probes), que se
liga covalentemente a aminoácidos básicos e ao esqueleto da cadeia polipeptídica,
permitindo a coloração da maioria das classes de proteínas, incluindo glicoproteínas,
fosfoproteínas, lipoproteínas, proteínas ligadoras de cálcio, proteínas fibrilares e
outras proteínas de difícil coloração. O corante possui taxa de quantificação linear de
mais de três ordens de magnitude e um limite mínimo de detecção variando entre
0,25 a 1 ng de proteína. Como a visualização das proteínas marcadas por estes
corantes só é possível através de aparelhos específicos, coramos por fim os géis
com Coomassie coloidal, que nos permite visualizar as proteínas (bandas e “spots”)
à olho nu, condição fundamental para a posterior retirada dos “spots” protéicos dos
géis, a fim de serem identificados por espectrometria de massas. O Coomassie
coloidal G-250 possui uma sensibilidade de detecção de até 10ng e melhores
contrastes podem ser obtidos lavando o gel em água após a coloração. As
colorações por ProQ, Sypro e Coomassie coloidal são completamente compatíveis
para análise dos peptídeos por espectrometria de massas. A captura das imagens
dos géis corados com ProQ Diamond e Sypro Ruby foi feita com o scanner Typhoon
Trio (Amersham Biosciences). Para os géis corados com Coomassie coloidal foi
utilizado o scanner GS-800 Calibrated Densitometer (BioRad). Os comprimentos de
onda considerados ideais de acordo com o fabricante para a excitação e emissão da
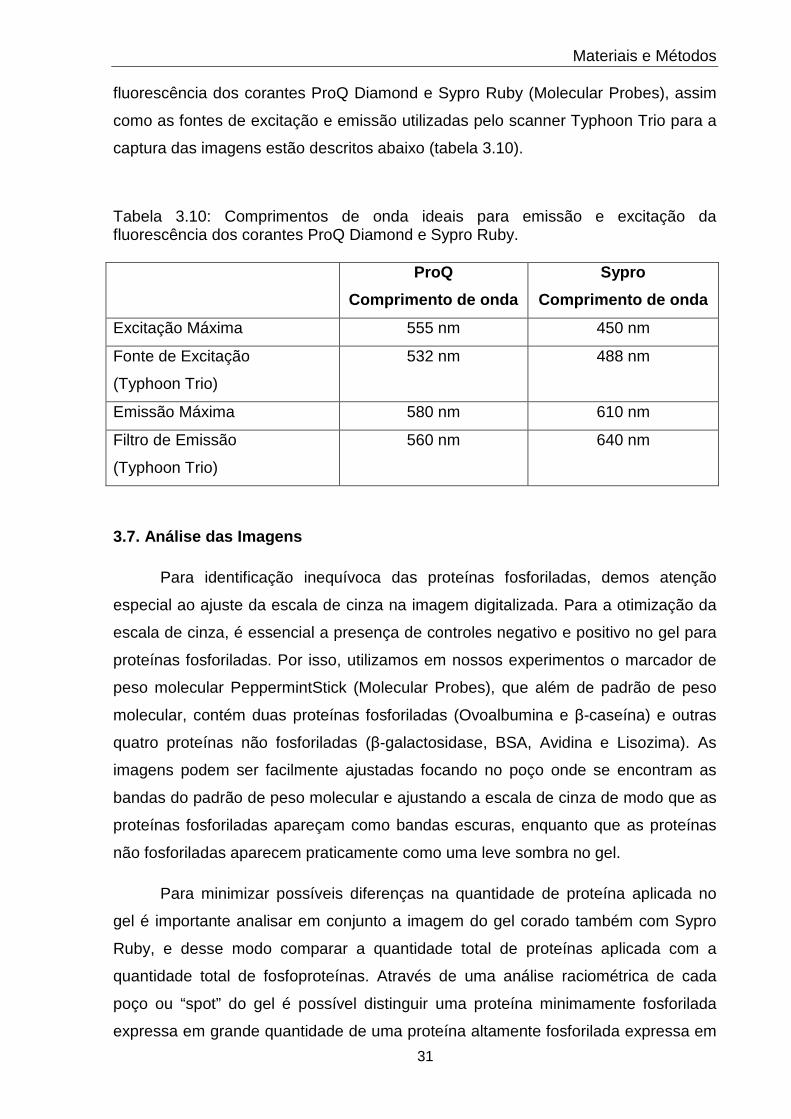
Materiais e Métodos
31
fluorescência dos corantes ProQ Diamond e Sypro Ruby (Molecular Probes), assim
como as fontes de excitação e emissão utilizadas pelo scanner Typhoon Trio para a
captura das imagens estão descritos abaixo (tabela 3.10).
Tabela 3.10: Comprimentos de onda ideais para emissão e excitação da fluorescência dos corantes ProQ Diamond e Sypro Ruby.
ProQ
Comprimento de onda
Sypro
Comprimento de onda
Excitação Máxima 555 nm 450 nm
Fonte de Excitação
(Typhoon Trio)
532 nm 488 nm
Emissão Máxima 580 nm 610 nm
Filtro de Emissão
(Typhoon Trio)
560 nm 640 nm
3.7. Análise das Imagens
Para identificação inequívoca das proteínas fosforiladas, demos atenção
especial ao ajuste da escala de cinza na imagem digitalizada. Para a otimização da
escala de cinza, é essencial a presença de controles negativo e positivo no gel para
proteínas fosforiladas. Por isso, utilizamos em nossos experimentos o marcador de
peso molecular PeppermintStick (Molecular Probes), que além de padrão de peso
molecular, contém duas proteínas fosforiladas (Ovoalbumina e β-caseína) e outras
quatro proteínas não fosforiladas (β-galactosidase, BSA, Avidina e Lisozima). As
imagens podem ser facilmente ajustadas focando no poço onde se encontram as
bandas do padrão de peso molecular e ajustando a escala de cinza de modo que as
proteínas fosforiladas apareçam como bandas escuras, enquanto que as proteínas
não fosforiladas aparecem praticamente como uma leve sombra no gel.
Para minimizar possíveis diferenças na quantidade de proteína aplicada no
gel é importante analisar em conjunto a imagem do gel corado também com Sypro
Ruby, e desse modo comparar a quantidade total de proteínas aplicada com a
quantidade total de fosfoproteínas. Através de uma análise raciométrica de cada
poço ou “spot” do gel é possível distinguir uma proteína minimamente fosforilada
expressa em grande quantidade de uma proteína altamente fosforilada expressa em

Materiais e Métodos
32
baixa quantidade. Para esse tipo de análise, medimos a intensidade de
fluorescência do sinal gerado por ProQ Diamond (P) e por Sypro Ruby (S), e
calculamos a razão P/S de cada spot ou poço do gel.
As imagens bidimensionais foram analisadas utilizando-se o programa
PDQuest (BioRad). A análise dos géis bidimensionais englobou as seguintes etapas:
normalização das imagens; detecção e quantificação dos spots; escolha de spots
referência (spots que estejam presentes em todos os géis a serem analisados);
alinhamento dos géis a partir da definição dos spots de referência; pareamento
automático entre os spots dos diferentes géis e a identificação dos spots
diferencialmente expressos.
Em nossas análises, foi utilizado o método de normalização denominado
“Total Density in Gel Image”, onde o valor de intensidade de pixel de cada “spot”,
medido em partes por milhão (ppm), é dividido pelo valor de intensidade total de
todos os pixels presentes na imagem. Este modelo de normalização assume que a
densidade total de uma imagem (incluindo densidade do background e densidade do
“spot”) será relativamente consistente de gel para gel, favorecendo a validação das
análises.
Neste trabalho, foram realizadas análises provenientes de comparações de
três experimentos independentes (triplicatas biológicas), entre as diferentes
condições estudadas (amostras tratadas ou não com TGF-β nos tempos de 1, 5, 15,
30 ou 60 minutos), observando a expressão qualitativa (presença ou não de
determinado “spot”) ou quantitativa em cada condição estudada.
3.8. Retirada e Processamento dos spots
Os “spots” protéicos de interesse foram excisados manualmente do gel, com
auxílio de ponteiras de 1 ml, cujas pontas foram cortadas com tesouras devidamente
limpas. Os spots de interesse foram transferidos para tubos novos previamente
lavados seqüencialmente em metanol/água milli-Q/metanol.
O processamento iniciou-se pela descoloração dos spots, adicionando-se a
cada tubo, contendo um spot coletado, 200 µL de uma solução de acetonitrila (ACN)
50% (p/v)/ bicarbonato de amônio 25 mM, pH 8.0. Esta etapa é feita por 16 h, com o

Materiais e Métodos
33
intuito de descorar completamente os “spots” e retirar o detergente SDS presente
nos géis de SDS-PAGE. Essa lavagem é repetida por mais duas vezes com o
mesmo volume e pelo tempo de 15 minutos cada, em agitação contínua. O
descorante é então removido e é adicionado a cada tubo um volume de 200µL da
solução de ACN 100% (solução de desidratação) por 5 minutos, sob agitação
contínua. Ao final desta etapa, os pedaços de géis tornam-se opacos. Em seguida,
os “spots” são colocados no evaporador a vácuo (Speed Vac) por 15 minutos, para
evaporação de toda a solução de acetonitrila. Posteriormente, foi realizada a etapa
de digestão tríptica, adicionando-se 10 µL da solução a 20 ng/µL de tripsina (Tripsin
Gold, Mass Spectrometry Grade – PROMEGA), de forma a cobrir completamente os
pedaços de gel. Os tubos são mantidos no gelo durante este procedimento de
digestão enzimática, garantindo que a mesma não sofra degradação. Após o
período de 1 h, todo o excesso de tripsina é removido e adiciona-se 20 µL de
solução de bicarbonato de amônio 50mM, disparando-se a digestão. Os tubos são
incubados por 16 h, a 37°C.
Aos tubos contendo as amostras é aplicado ultrasom por 10 minutos e em
seguida são agitados no vortex. Os 20 µL da solução acima são removidos para um
tubo limpo de 500 µL e o material restante re-extraído com 30 µL de solução de
ácido fórmico 5% (v/v) / ACN 50% (v/v). Os tubos são agitados em vortex por 20
segundos e a solução permanece no tubo durante 15 minutos a temperatura
ambiente. Após esse período, é novamente aplicado ultrasom por 2 minutos,
seguido por agitação no vortex. A solução contendo os peptídeos é removida e
unida aos 20 µL de solução reservada anteriormente em tubo de 500 µL. A extração
é repetida mais uma vez e todos os extratos são, ao final do processo, combinados
em um mesmo tubo, com um volume final de aproximadamente 60 µL. A amostra é
então concentrada no evaporador a vácuo até atingir o volume aproximado de 5-10
µL. Os peptídeos foram armazenados a -20ºC para posterior dessalinização.
Para retirar os possíveis contaminantes e sais que interferem na identificação
das proteínas por espectrometria de massas, além de concentrar a amostra protéica
para melhorar a sua identificação, utilizamos ponteiras contendo uma resina de fase
reversa (Zip Tip C18 - Millipore).
O ZipTip deve ser preparado previamente à sua utilização. Cada ponteira foi
umedecida em ACN 100% e equilibrada em uma solução de 0,1% TFA (ácido

Materiais e Métodos
34
trifluoroacético). Este procedimento é feito com sucessivas aspirações, de forma a
reter na resina os peptídeos e deixar no tubo os possíveis interferentes. Os
peptídeos são lavados com 0,1% TFA e finalmente eluídos em um novo tubo
utilizando uma solução de 0,1% TFA / ACN 50 %.
Depois de concentradas em Ziptip (Millipore), as amostras são co-
cristalizadas com uma matriz de ácido α-ciano-4-hidroxâmico em uma placa de aço
inox especialmente utilizada para a análise no espectrômetro de massas MALDI-
TOF-TOF. Foram misturadas quantidades iguais de amostra e matriz (0,3 µL)
estando a matriz à uma concentração de 1,0 mg/ ml diluída em solução 50% ACN
/0,3% TFA em água milliQ, sendo em seguida, armazenados para posterior análise
por espectrometria de massas.
3.9. Identificação de proteínas por Espectrometria de Massas
As amostras foram analisadas em espectrômetro de massas do tipo MALDI-
TOF-TOF 4700 Proteomics Analyser (Applied Biosystems) da Plataforma de
Espectrometria de Massas/PDTIS (Laboratório de Toxinologia, IOC-FIOCRUZ).
Inicialmente foi feita uma análise (MS) para determinar as massas dos peptídeos
trípticos referentes a cada “spot”. Em seguida, o equipamento faz uma nova
fragmentação (MS/MS) dos peptídeos com consequente geração de íons que, em
geral, podem ser íons y e íons b, ambos informativos e complementares entre si.
Assim, para cada peptídeo inicial (cuja massa já foi determinada) são gerados vários
valores de massa como produtos da fragmentação MS/MS. As diferenças entre
esses valores correspondem à massa dos aminoácidos que compõem o peptídeo,
permitindo a determinação de sua sequência. Após a conclusão da determinação
das massas dos diferentes peptídeos, os resultados foram capturados através do
software integrado ao equipamento (4000 Explorer Remote Client), sendo retirados
os picos de contaminantes. Os mesmos foram transferidos para um programa de
busca, visando à identificação das possíveis proteínas, como descrito abaixo.

Materiais e Métodos
35
3.10. Identificação dos peptídeos
Os resultados obtidos pelo espectrômetro de massas MALDI-TOF-TOF foram
analisados no programa Mascot Online (endereço eletrônico
www.matrixscience.com/cgi/search_form.pl?FORMVER=2&SEARCH=MIS). Para
identificação dos peptídeos foram utilizados os parâmetros descritos na tabela
abaixo (tabela 3.11):
Tabela 3.11: Parâmetros utilizados nas buscas do Mascot.
Database: NCBInr
Taxonomy : All entries
Enzyme: Trypsin
Allow up to: 1 missed cleavages
Fixed modifications : none
Variable modifications : Acetyl (protein N-term), Carbamidomethyl (C), Deamidated (NQ), Gln
→Pyro-Glu (N-termQ), Glu →Pyro-Glu (N-termE) ,Oxidation (HW), Oxidation (M), Phospho (ST),
Phospho (Y).
Peptide tol. : ± 1.2 Da
MS/MS tol.: ± 0.6 Da
Peptide charge: +1 Da
Monoisotopic
Instrument : Maldi TOF-TOF
Overview : Report top 50 hits
Consideramos uma identificação encontrada no Mascot como positiva após
verificarmos os dados obtidos para alguns dos parâmetros do programa. Foram eles:
o valor do “score” obtido para a identificação (valores superiores a 42 foram
considerados), se a proteína encontrada era proveniente de um microorganismo da
mesma espécie em estudo ou, pelo menos, da família dos tripanossomatídeos, se
havia equivalência entre os pontos isoelétricos e massas moleculares teóricas e
experimentais e a porcentagem de cobertura das proteínas.

Materiais e Métodos
36
3.11. Proliferação de epimastigotas em resposta a a dição de TGF- β
Um total de 2,5x106 parasitos foi obtido a partir de uma cultura inicial de
epimastigotas na concentração de 5x106 parasitos/mL. Esta massa foi ressuspensa
em 500 µl de meio LIT e a solução final distribuída em placas de 24 poços mantidas
a 28ºC. Para testar o efeito de TGF-β sobre a proliferação das epimastigotas de T.
cruzi, 5 ng/mL de TGF-β foram ou não adicionados diariamente às culturas de
epimastigotas. A cada 24 horas (24, 48 e 72 horas) os parasitos foram quantificados
em câmara de Neubauer com diluição de 1:100 em PBS. As amostras (incubada ou
não com TGF-β) foram preparadas em duplicata nos poços da placa. Apresentamos
nos resultados o gráfico obtido da média e desvio padrão de três experimentos
independentes realizados.
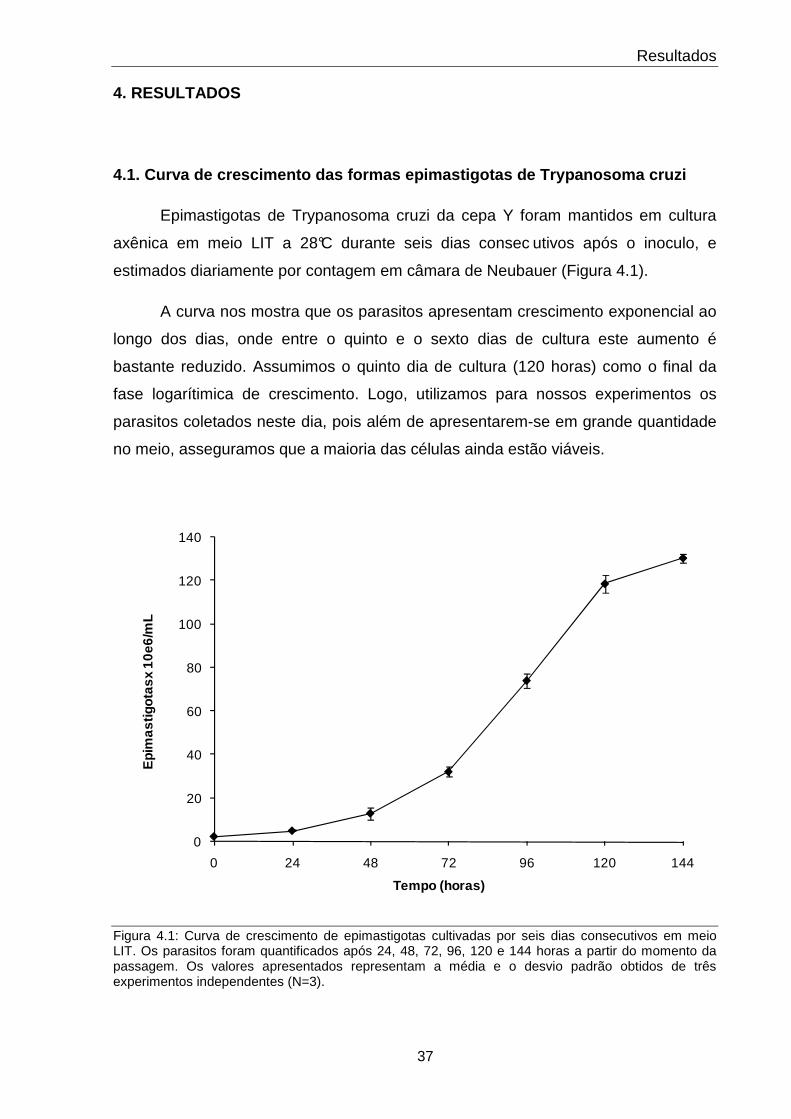
Resultados
37
4. RESULTADOS
4.1. Curva de crescimento das formas epimastigotas de Trypanosoma cruzi
Epimastigotas de Trypanosoma cruzi da cepa Y foram mantidos em cultura
axênica em meio LIT a 28°C durante seis dias consec utivos após o inoculo, e
estimados diariamente por contagem em câmara de Neubauer (Figura 4.1).
A curva nos mostra que os parasitos apresentam crescimento exponencial ao
longo dos dias, onde entre o quinto e o sexto dias de cultura este aumento é
bastante reduzido. Assumimos o quinto dia de cultura (120 horas) como o final da
fase logarítimica de crescimento. Logo, utilizamos para nossos experimentos os
parasitos coletados neste dia, pois além de apresentarem-se em grande quantidade
no meio, asseguramos que a maioria das células ainda estão viáveis.
Figura 4.1: Curva de crescimento de epimastigotas cultivadas por seis dias consecutivos em meio LIT. Os parasitos foram quantificados após 24, 48, 72, 96, 120 e 144 horas a partir do momento da passagem. Os valores apresentados representam a média e o desvio padrão obtidos de três experimentos independentes (N=3).
0
20
40
60
80
100
120
140
0 24 48 72 96 120 144
Epi
mas
tigot
asx
10e6
/mL
Tempo (horas)

Resultados
38
4.2. Padronização da dose ótima de TGF- β
Com o intuito de avaliar qual seria a dose ótima de TGF-β capaz de induzir a
maior fosforilação de proteínas em formas epimastigotas de T. cruzi, iniciamos
nossos estudos com um ensaio utilizando diferentes doses desta citocina. Para tal,
massas de 5x108 epimastigotas foram incubadas por 15 minutos com doses de 1,25
/ 2,5 / 5 ou 10 ng/ml de TGF-β, e em seguida, foram preparados os extratos de
proteínas totais das epimastigotas. Para avaliarmos qual seria a dose ótima de
indução de fosforilação de proteínas em resposta a TGF-β, delimitamos o volume de
cada canal (“lane”) do gel, utilizando ferramentas do programa Quantity One
(BioRad), tanto na imagem do gel corado com ProQ, como na imagem do gel corado
com Sypro. Calculamos então a razão (P/S) entre o valor da intensidade de pixels
totais do poço corado com ProQ (P) e o valor da intensidade de pixels totais do poço
corado com Sypro (S), com a intenção de normalizar os valores obtidos.
Os resultados obtidos (Figura 4.2) nos permitiram inferir que a dose ótima de
incubação com TGF-β, ou seja, aquela capaz de induzir o maior número de eventos
de fosforilação de proteínas nas formas epimastigotas de T. cruzi, foi a de 5 ng/ml.
Sendo assim, adotamos 5 ng/ml como a dose ótima de indução de
fosforilação em todos os nossos experimentos.

Resultados
39
Figura 4.2: A. Géis SDS-PAGE12% de 7cm corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagens do gel unidimensional contendo extratos de proteínas totais de epimastigotas, que foram previamente incubados com as doses de 0 (controle), 1,25, 2,5 5 e 10 ng/ml de TGF-β. Marcadores de massa molecular (MM) indicados na figura, em kDa. B. Gráfico representativo dos valores de intensidade relativa de pixels da razão ProQ/Sypro de um único experimento (N=1).
ProQ Sypro
M.M. C 1,25 2,5 5 10
116,2
66,2
45
23,6
18
14,4
45
23,6
M.M. C 1,25 2,5 5 10
0,9
0,95
1
1,05
1,1
1,15
1,2
C 1,25 2,5 5 10
Inte
nsi
dade
Rel
ativ
a TG
F-β
/CT
R
Dose de TGF-β adicionada
A
B

Resultados
40
4.3. Padronização do tempo ótimo de incubação com T GF-β
A partir da avaliação da dose ótima de TGF-β, realizamos um ensaio para
avaliar qual seria o tempo ideal de indução com esta citocina, ou seja, o tempo de
incubação com TGF-β capaz de induzir o maior número de eventos de fosforilação
de proteínas de formas epimastigotas de T. cruzi. Para tal, incubamos ou não
massas de 5x108 epimastigotas por 1, 5, 15, 30 ou 60 minutos com 5 ng/ml da
citocina em estudo, a 4ºC. O procedimento é feito em baixa temperatura com o
objetivo de retardar os eventos iniciais de fosforilação, que costumam ocorrer muito
rapidamente na situação fisiológica natural (in vivo). Contudo, incubamos ou não
massas de 5x108 epimastigotas por 1 ou 5 minutos com 5 ng/ml de TGF-β também a
28ºC, temperatura normal de cultivo de formas epimastigotas de T. cruzi.
Verificamos que o perfil unidimensional das amostras mantidas a 28ºC é bastante
semelhante àqueles das amostras incubadas a 4ºC (dados não mostrados).
O mesmo procedimento de normalização utilizado para a análise da dose
ótima foi aplicado na verificação do tempo ótimo de incubação com TGF-β. Nossas
análises, utilizando géis unidimensionais (Figura 4.3), indicaram que não há
diferenças significativas entre as amostras estimuladas e não estimuladas (controle)
de cada tempo estudado. As diferenças entre as amostras estimuladas com TGF-β
nos diferentes tempos estudados também parecem ser muito sutis, através da
análise unidimensional. Assim, optamos por concentrar nossos esforços na análise
do tempo ótimo de indução com TGF-β exclusivamente através de géis
bidimensionais, que permitem uma maior resolução das amostras em estudo, além
da visualização de diferenças pontuais encontradas em cada proteína estudada ao
longo do tempo.
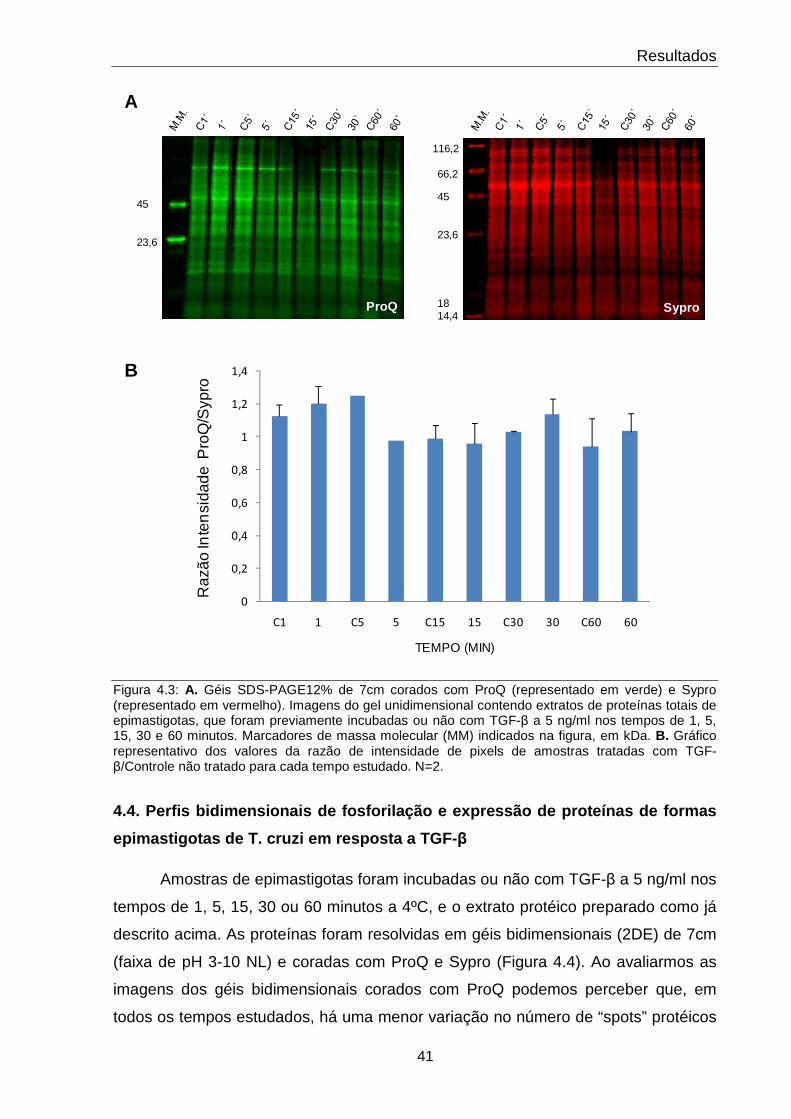
Resultados
41
Figura 4.3: A. Géis SDS-PAGE12% de 7cm corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagens do gel unidimensional contendo extratos de proteínas totais de epimastigotas, que foram previamente incubadas ou não com TGF-β a 5 ng/ml nos tempos de 1, 5, 15, 30 e 60 minutos. Marcadores de massa molecular (MM) indicados na figura, em kDa. B. Gráfico representativo dos valores da razão de intensidade de pixels de amostras tratadas com TGF-β/Controle não tratado para cada tempo estudado. N=2.
4.4. Perfis bidimensionais de fosforilação e expres são de proteínas de formas
epimastigotas de T. cruzi em resposta a TGF-β
Amostras de epimastigotas foram incubadas ou não com TGF-β a 5 ng/ml nos
tempos de 1, 5, 15, 30 ou 60 minutos a 4ºC, e o extrato protéico preparado como já
descrito acima. As proteínas foram resolvidas em géis bidimensionais (2DE) de 7cm
(faixa de pH 3-10 NL) e coradas com ProQ e Sypro (Figura 4.4). Ao avaliarmos as
imagens dos géis bidimensionais corados com ProQ podemos perceber que, em
todos os tempos estudados, há uma menor variação no número de “spots” protéicos
0
0,2
0,4
0,6
0,8
1
1,2
1,4
C1 1 C5 5 C15 15 C30 30 C60 60
TEMPO (MIN)
45
23,6
116,2
66,2
45
23,6
1814,4
Raz
ão In
ten
sida
de P
roQ
/Syp
ro
ProQ Sypro
A
B

Resultados
42
fosforilados nas porções mais ácidas (região de pH 3 a 5) e mais alcalinas (região de
pH 10) dos géis.
Na região de pH 5 a 8 há uma maior variação no perfil de fosforilação entre os
tempos estudados. Ao comparar as imagens, vemos que de 1 a 15 minutos há uma
concentração mais forte de “spots” no centro dos géis tratados com TGF-β, contra
uma fosforilação mais fraca nesta mesma região nos géis da amostra controle. Nos
tempos de 30 e 60 minutos parece haver defosforilação parcial de alguns “spots”
nesta região central em relação aos tempos mais curtos de incubação. Nestes
tempos mais longos de indução observamos uma fosforilação menos acentuada das
proteínas de ambas as amostras (tratadas ou não com TGF-β) e vemos também que
os perfis dessas amostras parecem se igualar.
Os perfis de expressão das proteínas nas diferentes condições estudadas
foram evidenciados através da coloração das proteínas totais por Sypro.
Observamos que, em todos os tempos estudados, os “spots” protéicos encontram-se
dispersos nos géis, sendo encontrados praticamente em todas as faixas de pH
estudadas, principalmente na região de pH 4 a 10. Contudo, a maior concentração
de proteínas encontra-se na faixa de pH 4 a 7.
Assim como nas análises unidimensionais, também preparamos géis
bidimensionais com extratos de epimastigotas incubados ou não com TGF-β a 5
ng/ml por 1 minuto a 28ºC. Verificamos que o perfil bidimensional das amostras
incubadas ou não com TGF-β por 1min a 28ºC se apresenta com um perfil
intermediário aos perfis das amostras incubadas ou não com TGF-β por 5 e 15
minutos a 4ºC (dados não mostrados). É importante ressaltar que nossos resultados
são baseados na análise de triplicatas biológicas de cada uma das amostras
estudadas. Consideramos apenas como dados dignos de avaliação em nosso
trabalho àqueles que foram reprodutíveis em todas as replicatas ou em pelo menos
duas das replicatas biológicas obtidas para cada amostra.
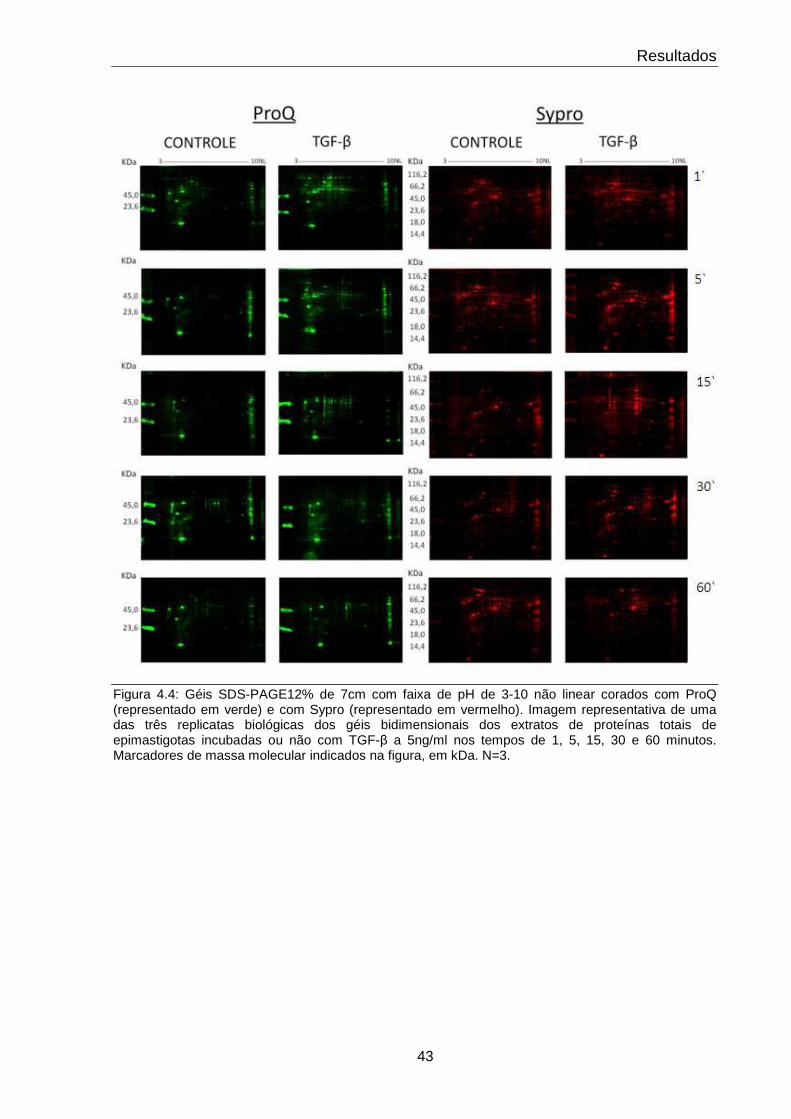
Resultados
43
Figura 4.4: Géis SDS-PAGE12% de 7cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas biológicas dos géis bidimensionais dos extratos de proteínas totais de epimastigotas incubadas ou não com TGF-β a 5ng/ml nos tempos de 1, 5, 15, 30 e 60 minutos. Marcadores de massa molecular indicados na figura, em kDa. N=3.

Resultados
44
4.5. Análises dos perfis bidimensionais diferenciai s de formas epimastigotas
de T. cruzi em resposta ao TGF-β
4.5.1. Quantidade total de “spots” protéicos em cad a condição estudada
As imagens dos géis bidimensionais (Fig. 4.4) foram analisadas através do
programa PDQuest (BioRad). A quantidade total de “spots” presente em cada
condição foi obtida através de uma das ferramentas do programa, denominada
“Experiment Summary”. Recolhemos os valores obtidos das replicatas de cada
condição estudada e calculamos a média e o desvio padrão destes valores,
representados graficamente na figura 4.5.
Em todos os tempos estudados observamos um maior número de eventos de
fosforilação nas amostras tratadas com TGF-β quando comparadas a seus
respectivos controles. Podemos também observar que no tempo de 1 minuto
ocorrem mais eventos de fosforilação do que nos outros tempos estudados. Os
tempos de 30 e 60 minutos apresentam quantidades semelhantes de eventos de
fosforilação, sendo estas inferiores as dos tempos menores de incubação nas
amostras tratadas com TGF-β. Após 30 minutos observamos uma tendência do
número de “spots” fosforilados em se igualar entre as amostras controle e tratadas
com TGF-β.
Com base nestes dados, calculamos a média do percentual de aumento de
fosforilação pela adição de TGF-β na cultura nos diferentes tempos. Observamos
que nos tempos de 5 e 15 minutos ocorrem os maiores aumentos no índice de
fosforilação quando os parasitos são tratados com TGF-β (58% e 41%,
respectivamente), seguido do tempo de 1 minuto (13%) e dos tempos de 30 e 60
minutos (9% e 5%, respectivamente).
Por termos observado diferenças nos perfis de fosforilação entre os tempos
estudados, resolvemos não mais selecionar um único tempo para análise, e sim
realizar um estudo de cinética de tempo.
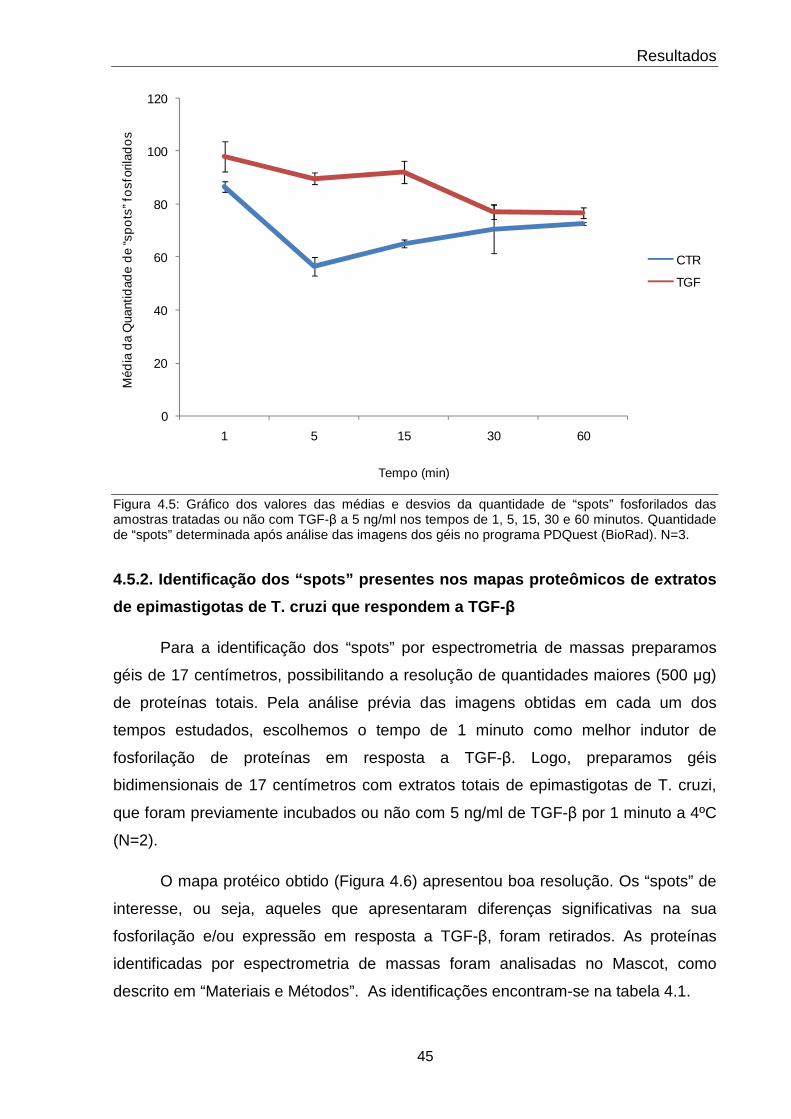
Resultados
45
Figura 4.5: Gráfico dos valores das médias e desvios da quantidade de “spots” fosforilados das amostras tratadas ou não com TGF-β a 5 ng/ml nos tempos de 1, 5, 15, 30 e 60 minutos. Quantidade de “spots” determinada após análise das imagens dos géis no programa PDQuest (BioRad). N=3.
4.5.2. Identificação dos “spots” presentes nos mapa s proteômicos de extratos
de epimastigotas de T. cruzi que respondem a TGF- β
Para a identificação dos “spots” por espectrometria de massas preparamos
géis de 17 centímetros, possibilitando a resolução de quantidades maiores (500 µg)
de proteínas totais. Pela análise prévia das imagens obtidas em cada um dos
tempos estudados, escolhemos o tempo de 1 minuto como melhor indutor de
fosforilação de proteínas em resposta a TGF-β. Logo, preparamos géis
bidimensionais de 17 centímetros com extratos totais de epimastigotas de T. cruzi,
que foram previamente incubados ou não com 5 ng/ml de TGF-β por 1 minuto a 4ºC
(N=2).
O mapa protéico obtido (Figura 4.6) apresentou boa resolução. Os “spots” de
interesse, ou seja, aqueles que apresentaram diferenças significativas na sua
fosforilação e/ou expressão em resposta a TGF-β, foram retirados. As proteínas
identificadas por espectrometria de massas foram analisadas no Mascot, como
descrito em “Materiais e Métodos”. As identificações encontram-se na tabela 4.1.
0
20
40
60
80
100
120
1 5 15 30 60
CTR
TGF
Méd
ia d
a Q
uant
idad
e d
e “s
po
ts” f
osf
orila
do
s
Tempo (min)

Resultados
46
Figura 4.6: Gel SDS-PAGE12% de 17cm com faixa de pH de 3-10 não linear corado por Coomassie coloidal. Gel bidimensional do extrato proteínas totais de epimastigotas incubada com TGF-β no tempo de 1 minuto. O mapa protéico representado acima apresenta circulados os “spots” identificados em nosso trabalho, cujos valores de fosforilação e/ou expressão variam significativamente nos diferentes tempos estudados. Marcadores de massa molecular indicados na figura, em kDa. N=2.
116,25
66,2
45,0
23,6
18,0
3 10 NL
55
52
53
73
6059
38
39
51
48
42
40
8
23
9
89
10
102 87 3
45
6 7 1516
17
19 20
27
25 26 28
29
31
3232a
2
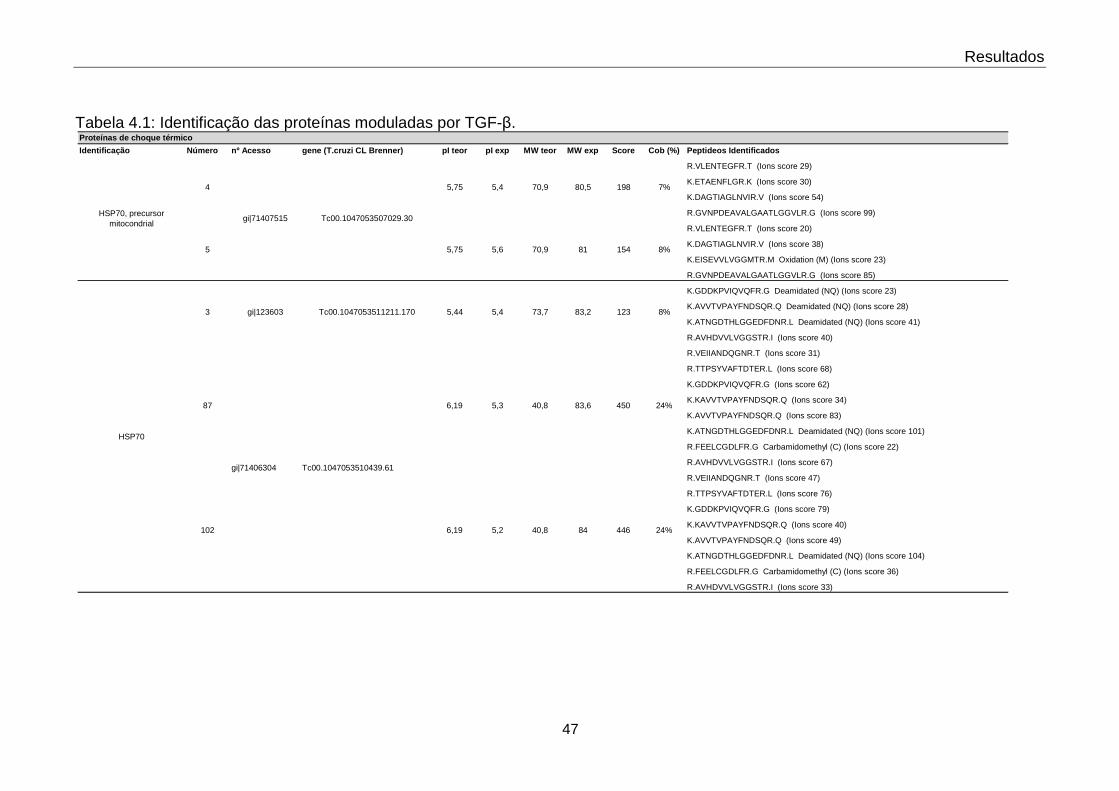
Resultados
47
Tabela 4.1: Identificação das proteínas moduladas por TGF-β.
Identificação Número nº Acesso gene (T.cruzi CL Brenne r) pI teor pI exp MW teor MW exp Score Cob (%) Peptideos I dentificados
R.VLENTEGFR.T (Ions score 29)
K.ETAENFLGR.K (Ions score 30)
K.DAGTIAGLNVIR.V (Ions score 54)
R.GVNPDEAVALGAATLGGVLR.G (Ions score 99)
R.VLENTEGFR.T (Ions score 20)
K.DAGTIAGLNVIR.V (Ions score 38)
K.EISEVVLVGGMTR.M Oxidation (M) (Ions score 23)
R.GVNPDEAVALGAATLGGVLR.G (Ions score 85)
K.GDDKPVIQVQFR.G Deamidated (NQ) (Ions score 23)
K.AVVTVPAYFNDSQR.Q Deamidated (NQ) (Ions score 28)
K.ATNGDTHLGGEDFDNR.L Deamidated (NQ) (Ions score 41)
R.AVHDVVLVGGSTR.I (Ions score 40)
R.VEIIANDQGNR.T (Ions score 31)
R.TTPSYVAFTDTER.L (Ions score 68)
K.GDDKPVIQVQFR.G (Ions score 62)
K.KAVVTVPAYFNDSQR.Q (Ions score 34)
K.AVVTVPAYFNDSQR.Q (Ions score 83)
K.ATNGDTHLGGEDFDNR.L Deamidated (NQ) (Ions score 101)
R.FEELCGDLFR.G Carbamidomethyl (C) (Ions score 22)
R.AVHDVVLVGGSTR.I (Ions score 67)
R.VEIIANDQGNR.T (Ions score 47)
R.TTPSYVAFTDTER.L (Ions score 76)
K.GDDKPVIQVQFR.G (Ions score 79)
K.KAVVTVPAYFNDSQR.Q (Ions score 40)
K.AVVTVPAYFNDSQR.Q (Ions score 49)
K.ATNGDTHLGGEDFDNR.L Deamidated (NQ) (Ions score 104)
R.FEELCGDLFR.G Carbamidomethyl (C) (Ions score 36)
R.AVHDVVLVGGSTR.I (Ions score 33)
Proteínas de choque térmico
123 8%5,44 5,4 73,7 83,2
Tc00.1047053510439.61
5,4 70,9 80,5 198 7%5,75
5,75 5,6 70,9 81 154 8%
24%
6,19 5,3 40,8 83,6 450 24%
6,19 5,2 40,8 84 446
gi|71406304
HSP70, precursor mitocondrial
Tc00.1047053507029.30gi|71407515
4
5
Tc00.1047053511211.170
87
102
gi|1236033
HSP70

Resultados
48
Identificação Número nº Acesso gene (T.cruzi CL Brenne r) pI teor pI exp MW teor MW exp Score Cob (%) Peptideos I dentificados
R.AVGVILQSVAEQSR.K (Ions score 37)
R.GLIDGETSDYNR.E (Ions score 34)
R.AAVQEGIVPGGGVALLR.A (Ions score 79)
K.ALDSLLGDSSLTADQR.T (Ions score 41)
K.VLENNDVTVGYDAQR.D (Ions score 85)
R.NVIIEQSYGAPK.I (Ions score 47)
R.AVGVILQSVAEQSR.K (Ions score 64)
R.GYISPYFVTDAK.A (Ions score 21)
R.GLIDGETSDYNR.E (Ions score 49)
R.AAVQEGIVPGGGVALLR.A (Ions score 27)
K.ALDSLLGDSSLTADQR.T (Ions score 63)
K.VLENNDVTVGYDAQR.D (Ions score 99)
K.SQTFSTNADNQR.N Deamidated (NQ) (Ions score 60)
R.ATLSEPDVEAGITLEDR.Q (Ions score 32)
R.VLSGVELEAVNR.V (Ions score 60)
K.AIELMGQALR.I Oxidation (M) (Ions score 28)
K.AMYWAQR.A Oxidation (M) (Ions score 23)
R.ALDLNPENVR.A (Ions score 52)
R.SYGISSFGK.D (Ions score 34)
K.TPASSEFPSGHISIVLK.V (Ions score 55)
M.SYKPHHATVPTNPK.V Acetyl (Protein N-term) (Ions score 44)
K.VFFDVSIGGQSAGR.V (Ions score 70)
K.NFGYAGSGFHR.I (Ions score 94)
K.FADESFAGK.A (Ions score 10)
R.INVYFDEATGGR.Y (Ions score 44)
R.YLTASALFR.G (Ions score 21)
R.INVYFDEATGGR.Y (Ions score 80)
R.AVLIDLEPGTMDSVR.A Oxidation (M) (Ions score 62)
R.FPGQLNSDLR.K (Ions score 66)
K.LAVNLVPFPR.L (Ions score 61)
Proteínas de citoesqueleto
89 11%6,2 24,3 19,8
8,9 18,7
4,88 4,7 30,1 83,5 87 10%
58,8 159 10%4,91 4.2 41,5
17%
HSP60
59,3 74,6 258 13%6 5,73 5
gi|71665064gi|71665068
Tc00.1047053507641.280
7 5,73 5,1 59,3 74,5 345
59GRPE gi|71401098 Tc00.1047053509045.20 8,49
gi|122054742
beta-tubulina Tc00.1047053506563.40
gi|185681399
gi|71659715 14 2098,44
4,69 4,7
27%
4,74 4,4 49,6 66,4 60 4%
38,5 266 10%49,4
Proteínas de choque térmico (Cont.)
Tc00.1047053511257.10Tc00.1047053509543.50
gi|71649573gi|71408185
2DNAK
Tc00.1047053508409.210Tc00.1047053510143.24
gi|71663660gi|71420437
10HSC70
73Ciclofilina A
(Peptidil-prolil cis-trans-isomerase)
Tc00.1047053506925.300
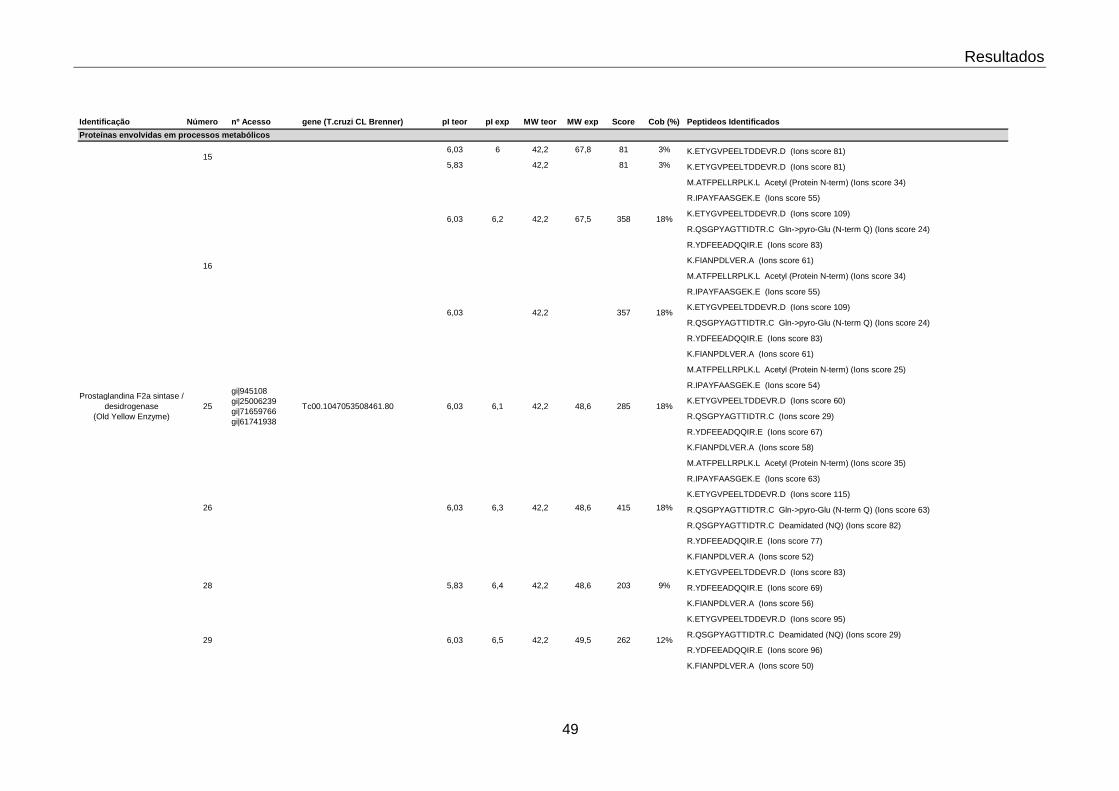
Resultados
49
Identificação Número nº Acesso gene (T.cruzi CL Brenne r) pI teor pI exp MW teor MW exp Score Cob (%) Peptideos I dentificados
6,03 6 42,2 67,8 81 3% K.ETYGVPEELTDDEVR.D (Ions score 81)
5,83 42,2 81 3% K.ETYGVPEELTDDEVR.D (Ions score 81)
M.ATFPELLRPLK.L Acetyl (Protein N-term) (Ions score 34)
R.IPAYFAASGEK.E (Ions score 55)
K.ETYGVPEELTDDEVR.D (Ions score 109)
R.QSGPYAGTTIDTR.C Gln->pyro-Glu (N-term Q) (Ions score 24)
R.YDFEEADQQIR.E (Ions score 83)
K.FIANPDLVER.A (Ions score 61)
M.ATFPELLRPLK.L Acetyl (Protein N-term) (Ions score 34)
R.IPAYFAASGEK.E (Ions score 55)
K.ETYGVPEELTDDEVR.D (Ions score 109)
R.QSGPYAGTTIDTR.C Gln->pyro-Glu (N-term Q) (Ions score 24)
R.YDFEEADQQIR.E (Ions score 83)
K.FIANPDLVER.A (Ions score 61)
M.ATFPELLRPLK.L Acetyl (Protein N-term) (Ions score 25)
R.IPAYFAASGEK.E (Ions score 54)
K.ETYGVPEELTDDEVR.D (Ions score 60)
R.QSGPYAGTTIDTR.C (Ions score 29)
R.YDFEEADQQIR.E (Ions score 67)
K.FIANPDLVER.A (Ions score 58)
M.ATFPELLRPLK.L Acetyl (Protein N-term) (Ions score 35)
R.IPAYFAASGEK.E (Ions score 63)
K.ETYGVPEELTDDEVR.D (Ions score 115)
R.QSGPYAGTTIDTR.C Gln->pyro-Glu (N-term Q) (Ions score 63)
R.QSGPYAGTTIDTR.C Deamidated (NQ) (Ions score 82)
R.YDFEEADQQIR.E (Ions score 77)
K.FIANPDLVER.A (Ions score 52)
K.ETYGVPEELTDDEVR.D (Ions score 83)
R.YDFEEADQQIR.E (Ions score 69)
K.FIANPDLVER.A (Ions score 56)
K.ETYGVPEELTDDEVR.D (Ions score 95)
R.QSGPYAGTTIDTR.C Deamidated (NQ) (Ions score 29)
R.YDFEEADQQIR.E (Ions score 96)
K.FIANPDLVER.A (Ions score 50)
Proteínas envolvidas em processos metabólicos
gi|945108gi|25006239gi|71659766gi|61741938
Tc00.1047053508461.80Prostaglandina F2a sintase /
desidrogenase (Old Yellow Enzyme)
6,03
15
25
18%6,03 6,3
18%
6,2 42,2 67,5 358 18%
42,2
5,83 6,4 42,2 48,6 203 9%
48,6 285 18%6,03 6,1
6,03
6,03
42,2 357
6,5 42,2 49,5 262
42,2 48,6 415
29
28
26
16
12%

Resultados
50
Identificação Número nº Acesso gene (T.cruzi CL Brenne r) pI teor pI exp MW teor MW exp Score Cob (%) Peptideos I dentificados
R.WAIEAGYR.H (Ions score 41)
R.HIDTAYFYNNEK.G (Ions score 77)
K.NHVLGEIAK.K (Ions score 24)
R.QIDELNEDKR.F Gln->pyro-Glu (N-term Q) (Ions score 42)
R.QIDELNEDKR.F (Ions score 35)
R.MGAEVYHSLK.S Oxidation (M) (Ions score 23)
K.QYNLTFK.S (Ions score 25)
K.AQVVGDDLTVTNVSR.I (Ions score 77)
R.SAVPSGASTGIHEACELR.D Carbamidomethyl (C) (Ions score 24)
K.AGSFNEALR.M (Ions score 37)
K.AQVVGDDLTVTNVSR.I Deamidated (NQ) (Ions score 84)
K.INQIGTITEAIEASK.F (Ions score 40)
20a 5,59 6,9 37,6 58,6 34 4% K.AQVVGDDLTVTNVSR.I (Ions score 36)
GAPDH 39 gi|120679 Tc00.1047053506943.50 8,87 9,5 39 45,1 79 4%R.VPTPDVSVVDLTFTAAR.D (Ions score 79)
8,96 9,6 34,1 37,4 42 3% R.RLPIGPITTVEK.E (Ions score 42)
8,82 34 42 3% R.RLPIGPITTVEK.E (Ions score 42)
7,79 7,1 29,5 46,3 51 5% K.AAADLTGVEAVQYPR.I (Ions score 53)
7,79 29,5 76 5% K.AAADLTGVEAVQYPR.I (Ions score 78)
K.NAAIFSEHGR.L (Ions score 57)
R.EAIKDDAYLDGEFMTTVQQR.G Oxidation (M) (Ions score 15)
R.EAVATWWR.N (Ions score 18)
R.KHVEDIVR.L (Ions score 18)
K.GKDPNATFTSVADFETTVPR.V (Ions score 56)
K.NLVVPGWR.L (Ions score 37)
13%6,6 46,4 58 173
5,59
5,92
46,1 51 122 10%
177
37,6 118 9%
67 9%gi|7109725 6,82 7 33,7 41,5
gi|71665461gi|5566209
Tc00.1047053504105.140
gi|71659493 Tc00.1047053510187.3027Tirosina aminotransferase
gi|14279174gi|189396135
13%48Aldo-ceto redutase 7,18 7,9 32,4 31,1
Malato desidrogenasegi|2961256gi|71423452
31Proteína dissulfeto
isomerasegi|71410849
5,82 6,3
Tc00.1047053511287.49
Enolase
32Desidrogenase aromática L-
alfa hidroxi-ácida
20
19
51 Tc00.1047053511293.69
Tc00.1047053509505.10
Proteínas envolvidas em processos metabólicos (Cont .)

Resultados
51
Identificação Número nº Acesso gene (T.cruzi CL Brenne r) pI teor pI exp MW teor MW exp Score Cob (%) Peptideos I dentificados
R.LILNYDVDPAR.I (Ions score 37)
R.IGFLGLGTESSTDNSAGSIIVR.G (Ions score 21)
R.ADKDFSSLLSR.K (Ions score 24)
K.EFVVAHIGSR.N (Ions score 22)
K.SYPLNTPSQR.L (Ions score 23)
R.YEYGLEAR.Y (Ions score 37)
R.LEYEEVLR.M (Ions score 45)
R.YEYGLEAR.Y (Ions score 26)
R.AVSGVVPETESR.V (Ions score 55)
R.VEEVPTTAETPETPVPAEK.T (Ions score 113)
K.VSIVATDIFTGNK.M (Ions score 96)
K.MEDQAPTTHNVDVPFVK.T Deamidated (NQ); Oxidation (M) (Ions score 75)
K.TSTYSVLDIQEDR.T (Ions score 131)
R.DNLDMPPNAELAAQIK.E Oxidation (M) (Ions score 94)
K.SVNFAQER.Y Deamidated (NQ) (Ions score 36)
K.IGGIGTVPVGR.V (Ions score 73)
R.FIPISGWQGDNMIDK.S Oxidation (M) (Ions score 26)
K.IGGIGTVPVGR.V (Ions score 19)
R.VETGTMKPGDVVTFAPANVTTEVK.S Oxidation (M) (Ions score 23)
K.SIEMHHEQLAEATPGDNVGFNVK.N Oxidation (M) (Ions score 65)
K.NSWTTQWGEDGYIR.I (Ions score 108)
K.NSWTTQWGEDGYIR.I Oxidation (HW) (Ions score 71)
K.NSWTTQWGEDGYIR.I Deamidated (NQ); 2 Oxidation (HW) (Ions score 38)
R.KGDTVVLIGDR.Q (Ions score 56)
R.LEASLIVCGR.E Carbamidomethyl (C) (Ions score 25)
R.ALIDVDGYDAEK.I (Ions score 77)
Proteínas hipotéticas
Proteínas associadas ao processo de tradução
Proteases/peptidades
106 2%
54,7 67,6 99 10%
93,6 40,5 69
15,7 394 35%4,82
167 17%
4,86 3,9
6,46,17
4,85 3,9
93 43,6 54 2%
1%
4,66 4,1 19,5 30,6
4,2 18
Sub-unidade proteolítica do complexo Hslvu
Fator de elongação 1 alfa
Tc00.1047053509429.320gi|11363088 5,63 3,9 49,6 53,3
5,2 22,8 22,8 157 15%6,77
105 4%
61 123 18%7,55 9,6 43,5
7,55 6,6 43,5 41,9Fator de elongação 1 alfa
Tc00.1047053510119.20gi|1929445gi|52424046gi|704459
38
Tc00.1047053506925.120
40Complexo associado a
peptídeo nascentegi|71419111
Tc00.1047053510579.70Tc00.1047053510241.60
23
Proteína hipotética
gi|71405542 Tc00.1047053507569.10
Tc00.1047053511903.40
89 gi|71409780 Tc00.1047053504423.30
gi|71416273 Tc00.1047053506275.20
17Proteína hipotética gi|71661816
53Fator de iniciação
eucariótico 5agi|71659667
32a
Cruzipaína
gi|704459
55

Resultados
52
Identificação Número nº Acesso gene (T.cruzi CL Brenne r) pI teor pI exp MW teor MW exp Score Cob (%) Peptideos I dentificados
R.GGLGQMNIPILADK.T Oxidation (M) (Ions score 52)
K.SYGVLKEEDGVAYR.G (Ions score 81)
K.EEDGVAYR.G (Ions score 18)
R.QITVNDLPVGR.D Gln->pyro-Glu (N-term Q) (Ions score 52
R.QITVNDLPVGR.D (Ions score 64)
K.GSYIEVGGEDYGIAANVDEDAGEGAK.G (Ions score 121)
R.VVDVVHNNR.Y (Ions score 45)
K.NSYMAHIR.G Oxidation (M) (Ions score 20)
Proteínas envolvidas na regulação do estresse oxida tivo
Proteínas envolvidas na transdução de sinal
5,71 22,7 209 19%
52Fator de liberação de
histamina dependente de IgE
gi|71407834Tc00.1047053506207.50Tc00.1047053510065.30
25%4,52 1844.0 19,5 18,7
Tc00.1047053507259.10Tc00.1047053487507.10
gi|17224953gi|71396508gi|58176947
60Tryparedoxina peroxidase

Resultados
53
4.5.3. Proteínas moduladas após a adição de TGF- β
A partir de nossas análises foi possível observar que a adição de TGF-β às
culturas de epimastigotas de T. cruzi induziu tanto a fosforilação de determinadas
proteínas, principalmente nos primeiros minutos após a introdução da citocina,
quanto o aumento ou redução na expressão relativa de proteínas do extrato total de
T. cruzi. Para esses “spots” que sofreram algum tipo de modulação (seja no perfil de
fosforilação seja no perfil de expressão) obtivemos, através do programa PDQuest
(BioRad), os seus valores de intensidade de pixel quando corados com ProQ e com
Sypro, respectivamente. Dessa forma, para cada “spot” obtivemos dois valores de
média de intensidade: um referente ao seu nível de fosforilação (ProQ) e outro
referente ao seu nível de expressão (Sypro).
Tanto para as análises de fosforilação (ProQ), como para as de expressão
(Sypro) calculamos a razão (C/I) entre o valor da amostra controle (C) e da amostra
estimulada com TGF-β (I) e vice-versa (razão I/C) para cada “spot” de interesse.
Consideramos como expressão ou fosforilação significativas quando encontrado um
aumento ou uma redução superior a duas vezes o valor da intensidade de pixel do
“spot” tratado com TGF-β em relação a seu controle.
Nas análises de fosforilação calculamos, além da razão descrita acima, a
razão (P/S) da intensidade de pixel do “spot” corado por ProQ (P) pela intensidade
de pixel do “spot” corado por Sypro (S), de modo a normalizar a quantidade total de
fosforilação pela quantidade total de proteína presente no gel.
Estas análises demonstram que alguns “spots” apresentam diferenças quanto
ao seu nível de fosforilação, mantendo seus níveis de expressão constantes ao
longo do tempo estudado; outros - fosforilados ou não – apresentaram diferenças
apenas quanto aos seus perfis de expressão ao longo do tempo estudado; um
terceiro grupo apresentou alterações significativas tanto no nível de fosforilação
quanto de expressão. Estas proteínas foram classificadas em grupos funcionais e o
comportamento de cada uma delas ao longo do tempo encontra-se representado
graficamente nas figuras a seguir.
A cada “spot” do gel foi atribuído um número que permite a sua localização no
mapa proteômico (Figura 4.6). Algumas proteínas são identificadas em diferentes

Resultados
54
“spots” no gel e observamos que o comportamento de fosforilação e expressão
destes podem apresentar diferenças entre si.
Em nossas análises, quando encontramos diferentes “spots” (de mesma massa
molecular, que apresentavam apenas pequenas variações de pI), identificados como
uma proteína com seqüência idêntica de aminoácidos, relacionamos esses “spots”,
somando seus valores e os representando também unificados em gráficos. A nossa
intenção é mostrar o comportamento geral da proteína, já que no gel bidimensional
uma mesma proteína pode ser separada em vários “spots” horizontalmente
dispostos, devido a modificações pós-traducionais (como fosforilação) que essas
proteínas podem sofrer durante diversos processos celulares.
Os valores da média da intensidade de pixel (em ppm) dos “spots” em cada
tempo de estudo estão representados na tabela a seguir. Na tabela, as proteínas
estão organizadas em grupamentos funcionais (tabela 4.2).

Resultados
55
Tabela 4.2: Valores das médias da intensidade de pixels dos “spots” responsivos a TGF-β ao longo dos tempos estudados.
Identificação Número 1 min 5 min 15 min 30 min 60 min
Fosforilação 8,39 -1,39 0 1,76 -2,33
Expressão 1,02 1,01 -4,65 -1,75 -1,75
Fosforilação 3,22 0 0 4,23 2,43
Expressão 1,50 1,58 -1,91 4,23 2,43
Fosforilação 0 0 0 0 0
Expressão 1,04 6,15 -2,72 -3,35 -2,65
Fosforilação 1,95 1,09 1,11 2,05 1,48
Expressão 0 1,55 0,82 -3,78 -19,20
Fosforilação 0 0 0 0 -2,16
Expressão 0 0 0 0 5,72
Fosforilação 3,65 0 0 1,11 1,92
Expressão 1,62 0,85 -6,00 -3,27 -2,10
Fosforilação 0 0 0 0 0
Expressão 1,09 1,28 -3,20 -2,33 -3,55
DNAK 2 Expressão 3,90 3,13 -2,33 -3,18 0
Fosforilação 1,90 1,33 0,95 1,35 -1,88
Expressão 1,08 1,55 -1,39 1,14 1,22
GrpE 59 Expressão -1,65 0,93 5,34 2,83 2,24
Ciclofilina A 73 Expressão -1,2 1,11 -3,50 4,13 0,63
Proteínas de citoesqueleto
Fosforilação 1,06 2,88 1,08 2,13 -2,32
Expressão 2,23 3,39 1,71 -4,39 -2,38
Fosforilação 3,13 1,00 -2,86 1,33 -2,24
Expressão -1,35 1,27 -2,64 -1,41 -2,45
Valores positivos representam a média de ppm da razão TGF-beta/ Controle
Valores negativos representam a média de ppm da razão Controle/ TGF-beta
Proteínas de choque térmico
HSP70 precursor
mitocondrial
HSP70
HSP60
Proteina de interação a
Hsc70
beta Tubulina
9
42
7
10
87
102
6
4
5
3

Resultados
56
Identificação Número 1 min 5 min 15 min 30 min 60 min
Proteínas de metabolismo
Fosforilação 6,36 0 0 0 3,91
Expressão 1,18 1,70 1,22 1,16 -2,2
Fosforilação 5,10 4,48 0 -1,21 3,72
Expressão -2,59 -1,19 1,64 0,95 -5
25 Expressão 1,51 1,00 -5 -5,16 -1,37
Fosforilação -2,26 -9,80 0 3,34 -2,67
Expressão 1,37 4,14 -2,99 -4,07 1,67
Fosforilação 0 0 0 0 1,55
Expressão 2,58 2,95 1,15 -4,0 1,18
29 Expressão -1,72 3,52 0 6,20 -1,98
Aldo-Ceto Redutase 48 Expressão -1,39 1,29 1,32 -2,70 -1,34
Fosforilação 3,26 -12,44 0 -3,59 1,61
Expressão -1,96 13,06 -3,80 1,48 1,32
Fosforilação 0 0 0 2,06 2,30
Expressão -1,34 -1,22 -2,78 -6,92 1,92
Fosforilação 3,42 -5,51 0,96 1,13 2,52
Expressão -2,55 2,34 1,12 0,95 -1,84
Fosforilação -3,09 2,12 17,79 1,66 24,07
Expressão 1,06 3,37 2,91 1,37 -5,86
Valores positivos representam a média de ppm da razão TGF-beta/ Controle
Valores negativos representam a média de ppm da razão Controle/ TGF-beta
Gliceraldeido 3 Fosfato
Desidrogenase
malato Desidrogenase
glicossomal
Prostaglandina F2 alfa
sintase/ Desidrogenase
(Old yellow enzyme)
39
51
19
20
Enolase
15
16
26
28
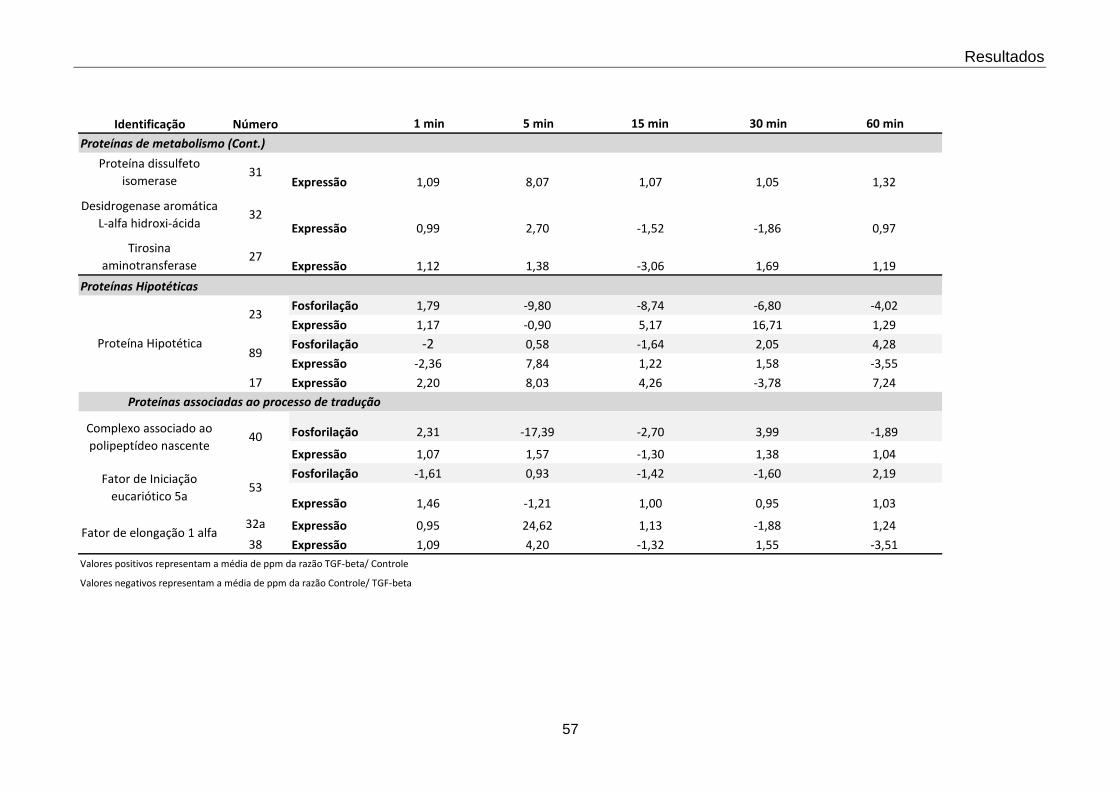
Resultados
57
Identificação Número 1 min 5 min 15 min 30 min 60 min
Proteínas de metabolismo (Cont.)
Proteína dissulfeto
isomerase31
Expressão 1,09 8,07 1,07 1,05 1,32
Desidrogenase aromática
L-alfa hidroxi-ácida32
Expressão 0,99 2,70 -1,52 -1,86 0,97
Tirosina
aminotransferase27
Expressão 1,12 1,38 -3,06 1,69 1,19
Proteínas Hipotéticas
Fosforilação 1,79 -9,80 -8,74 -6,80 -4,02
Expressão 1,17 -0,90 5,17 16,71 1,29
Fosforilação -2 0,58 -1,64 2,05 4,28
Expressão -2,36 7,84 1,22 1,58 -3,55
17 Expressão 2,20 8,03 4,26 -3,78 7,24
Proteínas associadas ao processo de tradução
Fosforilação 2,31 -17,39 -2,70 3,99 -1,89
Expressão 1,07 1,57 -1,30 1,38 1,04
Fosforilação -1,61 0,93 -1,42 -1,60 2,19
Expressão 1,46 -1,21 1,00 0,95 1,03
32a Expressão 0,95 24,62 1,13 -1,88 1,24
38 Expressão 1,09 4,20 -1,32 1,55 -3,51
Valores positivos representam a média de ppm da razão TGF-beta/ Controle
Valores negativos representam a média de ppm da razão Controle/ TGF-beta
Proteína Hipotética
Complexo associado ao
polipeptídeo nascente
Fator de Iniciação
eucariótico 5a
Fator de elongação 1 alfa
53
23
89
40

Resultados
58
Identificação Número 1 min 5 min 15 min 30 min 60 min
Proteases/ Peptidases
Cruzipaina 8 Expressão -1,24 38,07 -4,00 -2,43 1,49
Sub-unidade proteolítica
do complexo Hslvu
55
Expressão 0,94 4,00 2,02 -1,38 -1,9
Proteínas de regulação de estresse oxidativo
Triparedoxina peroxidase60
Expressão -1,6 1,38 1,15 -1,39 11,82
Proteínas de Transdução de sinal
Fosforilação 1,90 -1,35 -7,70 4,58 18,58
Expressão 0,80 -1,23 1,00 1,82 -2,90
Valores positivos representam a média de ppm da razão TGF-beta/ Controle
Valores negativos representam a média de ppm da razão Controle/ TGF-beta
Fator de liberação de
Histamina dependente
de IgE
52

Resultados
59
• Proteínas de choque térmico: HSPs (“heat shock prot eins”)
No mapa proteômico encontramos 5 “spots” identificados como HSP70 (“heat
shock protein 70 kDa”), dentre estes, dois “spots” são diferencialmente fosforilados
(“spots” 4 e 5) e dois “spots” diferencialmente expressos (“spots” 3 e 4). A união dos
“spots” 4 e 5 gera um gráfico onde se observa um aumento na fosforilação em 1
minuto e expressão reduzida em 15 minutos. A união dos “spots” 3, 87 e 102 gera
um gráfico onde se observa um aumento na fosforilação em 1 e 30 minutos. A
expressão é aumentada em 1 minuto e reduzida em 15, 30 e 60 minutos (Figura
4.7).
No mapa proteômico encontramos 2 “spots” identificados como HSP60 (“heat
shock protein 60 kDa”); observamos que um “spot” é diferencialmente fosforilado
(“spot” 6) e dois “spots” diferencialmente expressos (“spots” 6 e 7) (Figura 4.7).
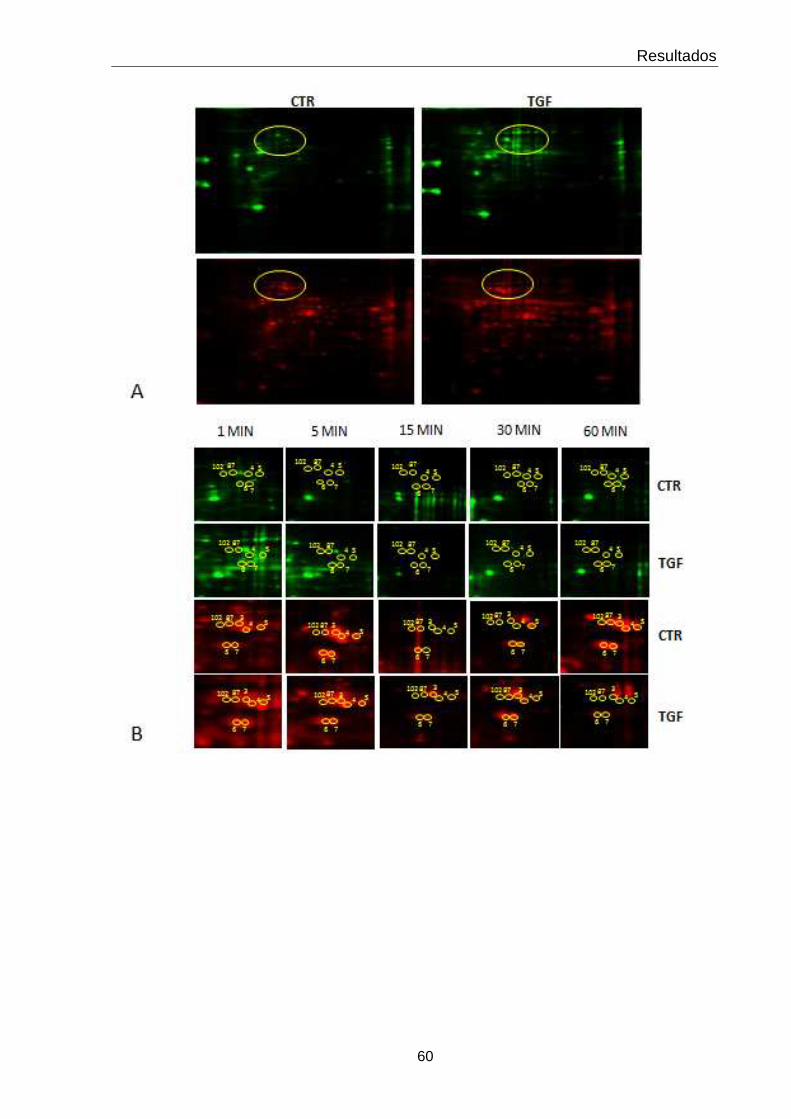
Resultados
60
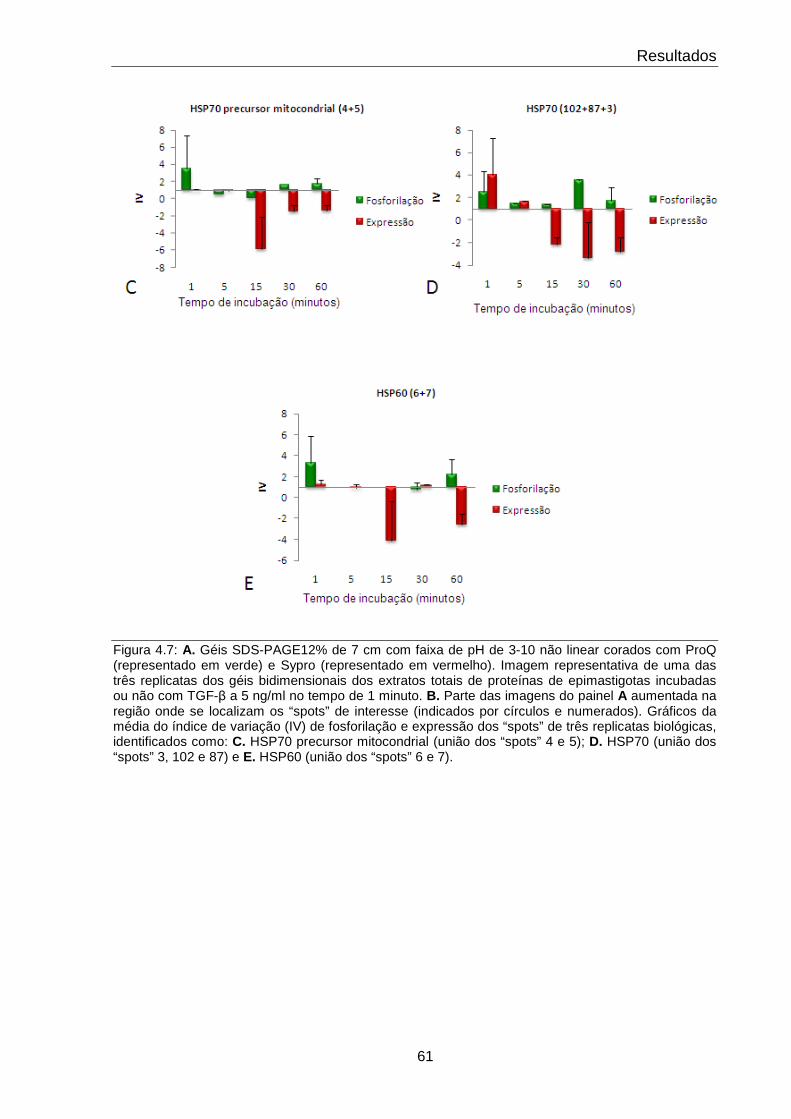
Resultados
61
Figura 4.7: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localizam os “spots” de interesse (indicados por círculos e numerados). Gráficos da média do índice de variação (IV) de fosforilação e expressão dos “spots” de três replicatas biológicas, identificados como: C. HSP70 precursor mitocondrial (união dos “spots” 4 e 5); D. HSP70 (união dos “spots” 3, 102 e 87) e E. HSP60 (união dos “spots” 6 e 7).
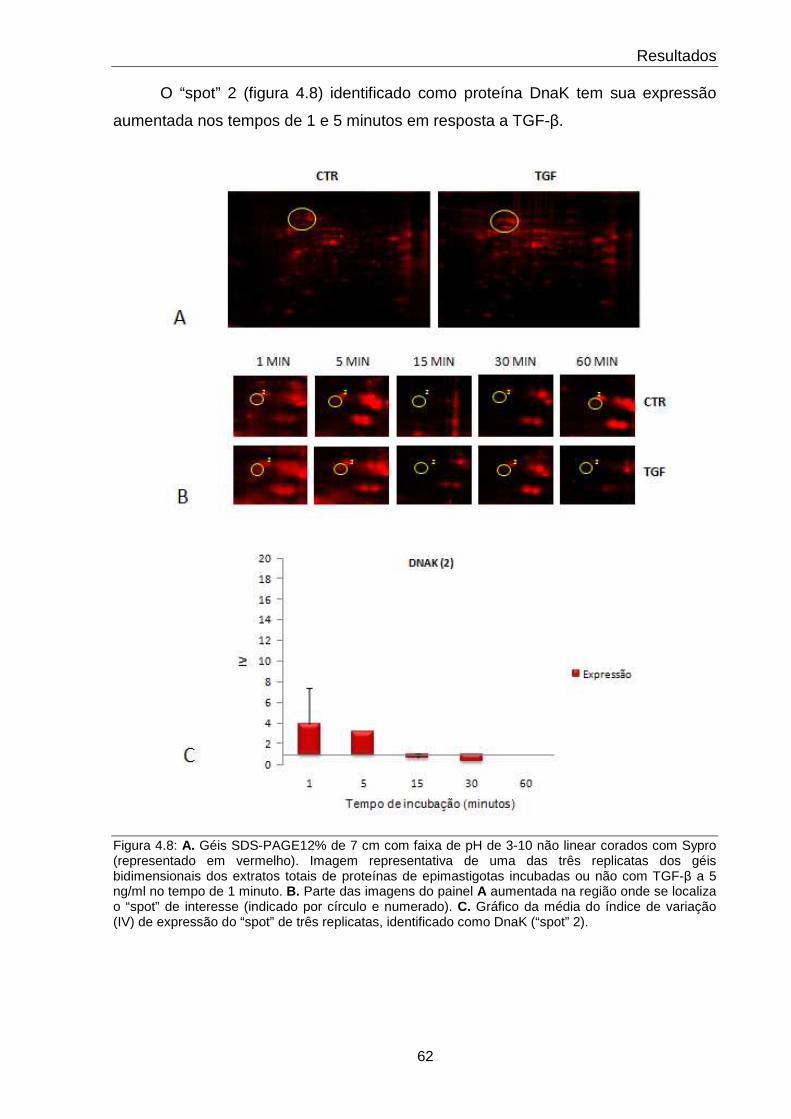
Resultados
62
O “spot” 2 (figura 4.8) identificado como proteína DnaK tem sua expressão
aumentada nos tempos de 1 e 5 minutos em resposta a TGF-β.
Figura 4.8: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como DnaK (“spot” 2).
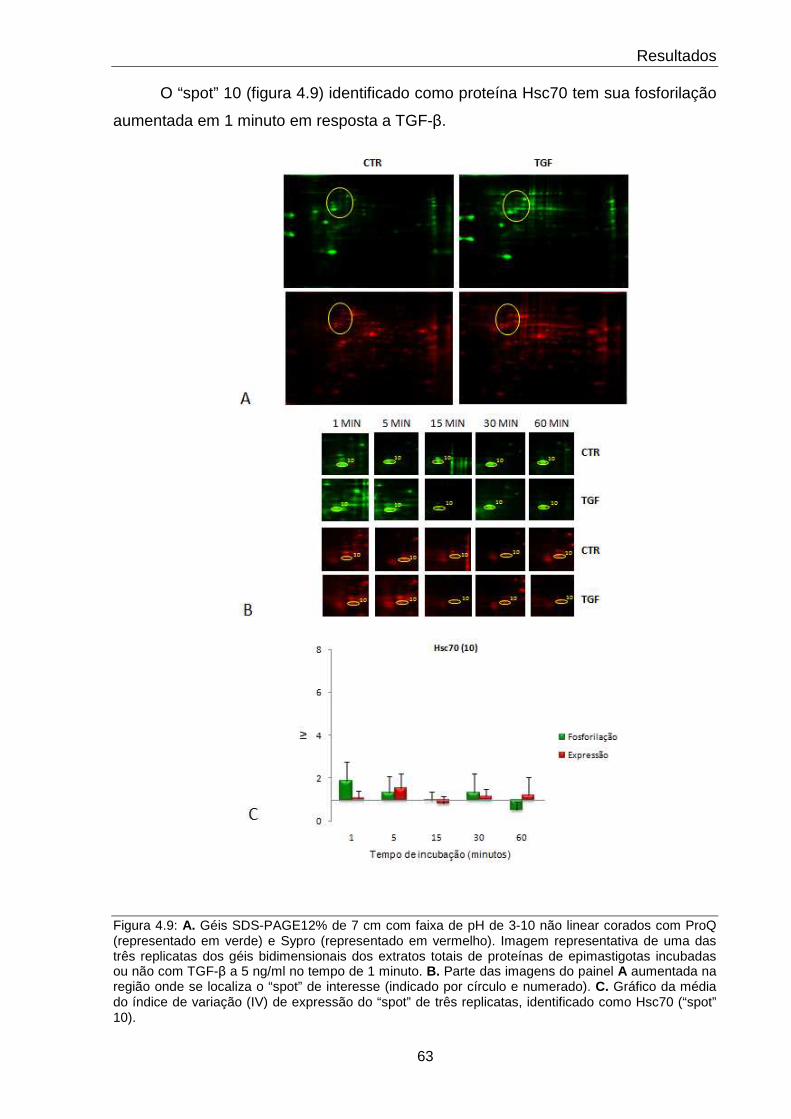
Resultados
63
O “spot” 10 (figura 4.9) identificado como proteína Hsc70 tem sua fosforilação
aumentada em 1 minuto em resposta a TGF-β.
Figura 4.9: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Hsc70 (“spot” 10).
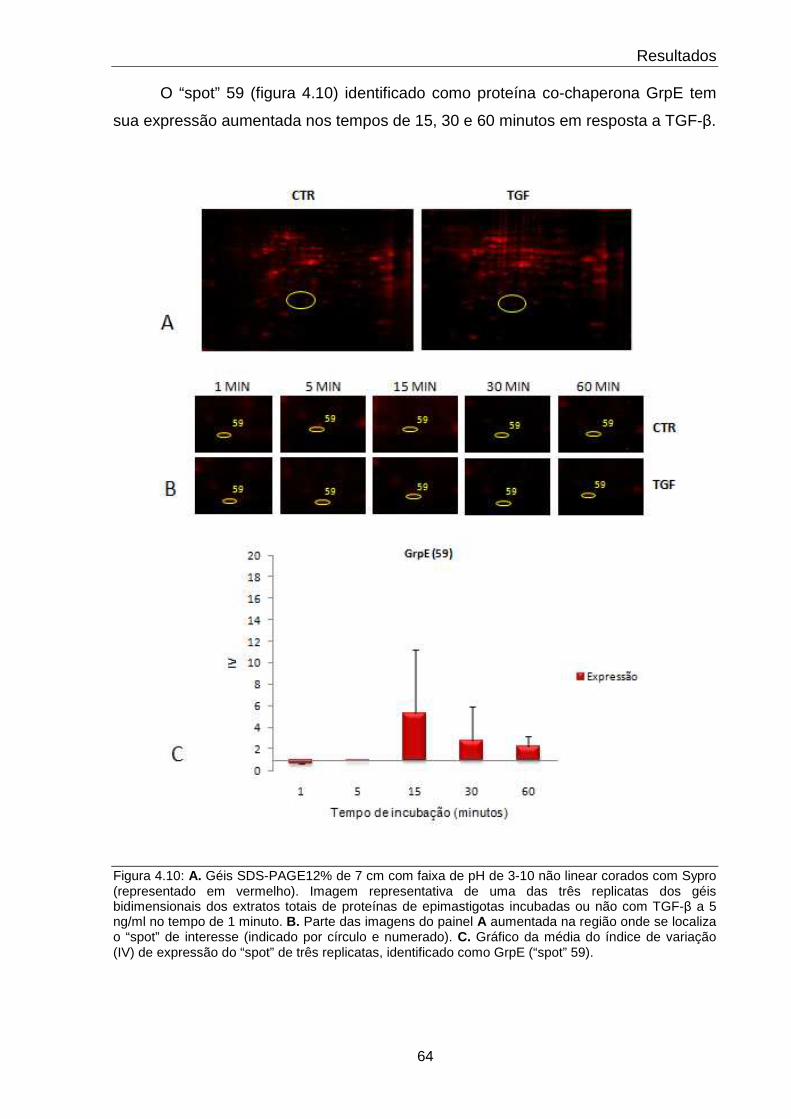
Resultados
64
O “spot” 59 (figura 4.10) identificado como proteína co-chaperona GrpE tem
sua expressão aumentada nos tempos de 15, 30 e 60 minutos em resposta a TGF-β.
Figura 4.10: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como GrpE (“spot” 59).
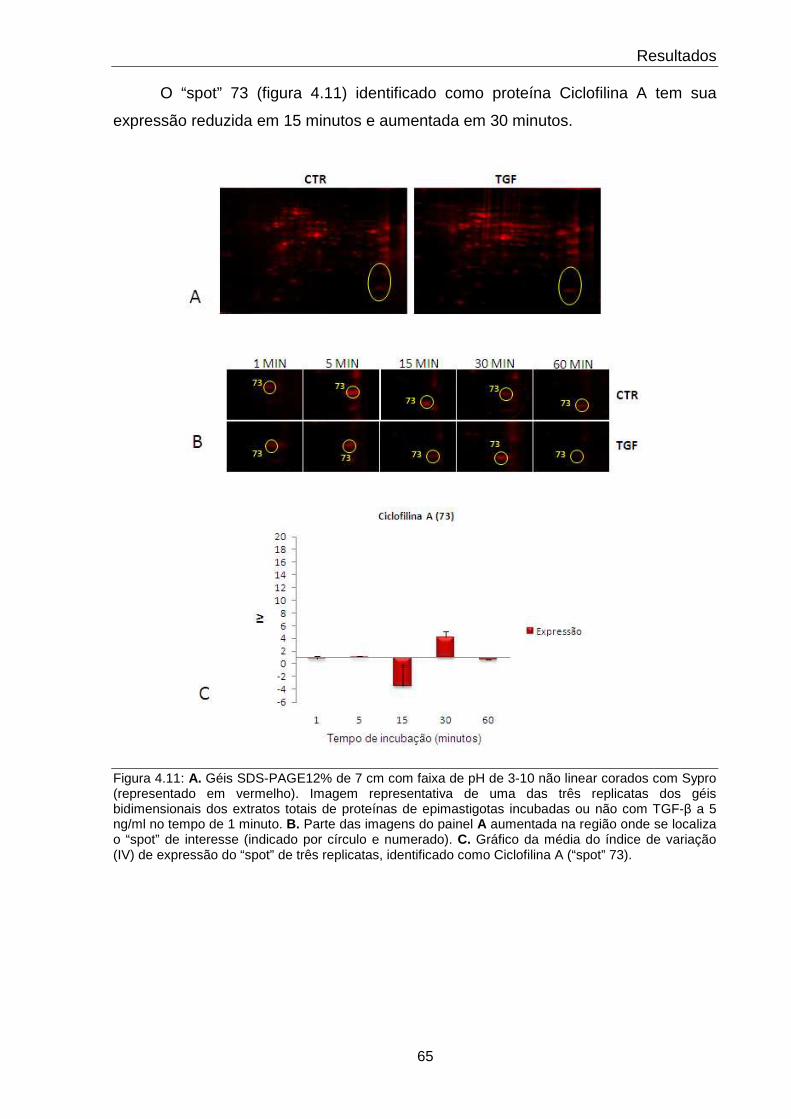
Resultados
65
O “spot” 73 (figura 4.11) identificado como proteína Ciclofilina A tem sua
expressão reduzida em 15 minutos e aumentada em 30 minutos.
Figura 4.11: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Ciclofilina A (“spot” 73).

Resultados
66
• Proteínas de citoesqueleto ( β-tubulina).
No mapa proteômico observamos dois “spots” identificados como β-tubulina,
ambos diferencialmente fosforilados e expressos (“spots” 9 e 42).
O “spot” 9 (figura 4.12) apresenta um aumento na fosforilação em 5 e 30 minutos
e sofre defosforilação em 60 minutos. A sua expressão é aumentada em 1 e 5
minutos e reduzida em 30 e 60 minutos.
O “spot” 42 (figura 4.12) tem fosforilação aumentada em 1 minuto e é
defosforilado em 15 minutos. Sua expressão é reduzida em 15 minutos e 60
minutos.

Resultados
67
Figura 4.12: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). Gráficos da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como: C. β-Tubulina (“spot” 9) e D. β-Tubulina (“spot” 42).

Resultados
68
• Proteínas envolvidas em processos metabólicos
No mapa proteômico, foram observados nove “spots” identificados como
diferentes tipos de desidrogenases; dentre estes, cinco são diferencialmente
fosforilados (“spots” 15, 16, 26, 39 e 51) e seis diferencialmente expressos (“spots”
25, 26, 28 32, 39 e 51) (figura 4.13).
Os “spots” 15, 16, 25, 26, 28 e 29 foram identificados como prostaglandina F2α
sintase. A partir da união dos “spots” 15 e 16, observa-se que ocorre fosforilação
aumentada nos tempos de 1, 5 e 60 minutos e defosforilação em 15 e 30 minutos. A
expressão destas proteínas é rapidamente reduzida em 1 minuto, mas aumentada
em 5 minutos, tornando a reduzir em 15 e 60 minutos de indução com TGF-β(figura
4.13). A união dos “spots” 25, 26, 28 e 29 gera um gráfico (figura 4.13) onde se
observa defosforilação em 5 minutos e aumento na fosforilação em 30 minutos. A
sua expressão é aumentada em 5 minutos e reduzida em 15 e 30 minutos.
O “spot” 48 (figura 4.14) identificado como proteína aldo-ceto redutase tem sua
expressão reduzida apenas em 30 minutos, não sofrendo regulação nos demais
tempos de estudo.
No mapa proteômico, foram observados dois “spots” identificados como enolase,
onde ambos são diferencialmente fosforilados e expressos (“spots” 19 e 20). A união
dos “spots” 19 e 20 gera um gráfico (figura 4.15) onde se observa uma
defosforilação em 5 minutos e um aumento na fosforilação em 30 e 60 minutos. A
sua expressão é reduzida em 1, 15 e 30 minutos e aumentada em 5 minutos.
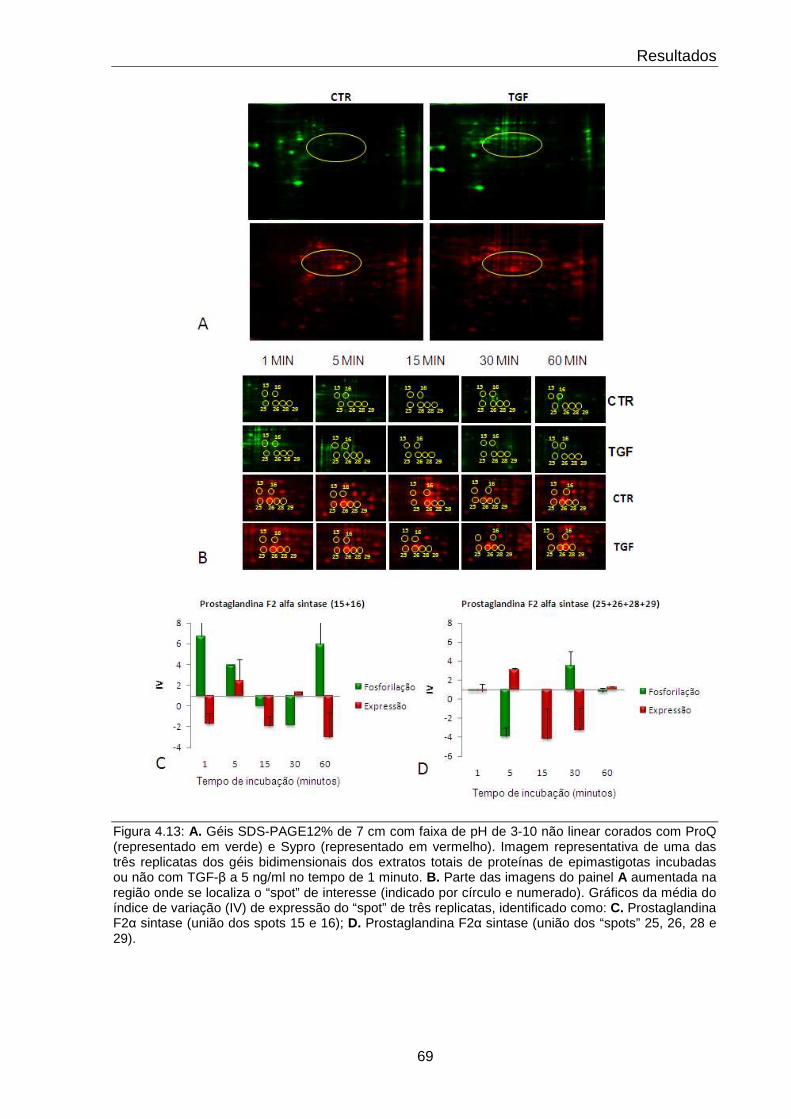
Resultados
69
Figura 4.13: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). Gráficos da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como: C. Prostaglandina F2α sintase (união dos spots 15 e 16); D. Prostaglandina F2α sintase (união dos “spots” 25, 26, 28 e 29).
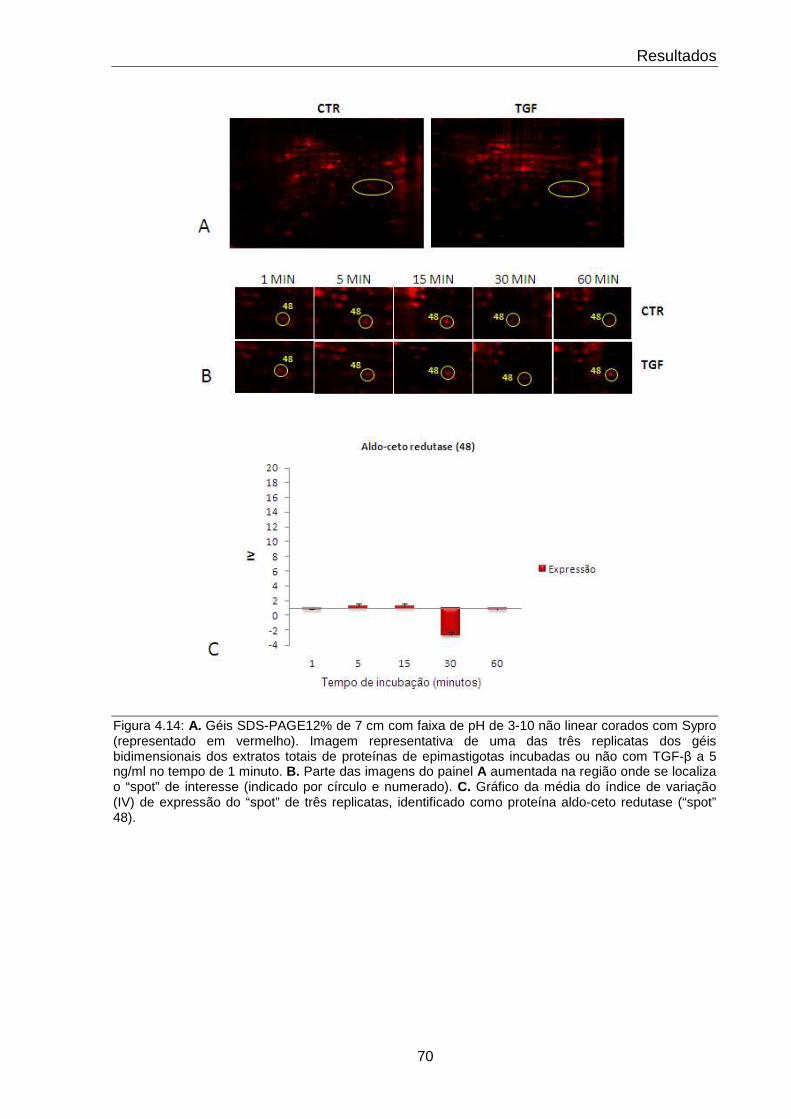
Resultados
70
Figura 4.14: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como proteína aldo-ceto redutase (“spot” 48).
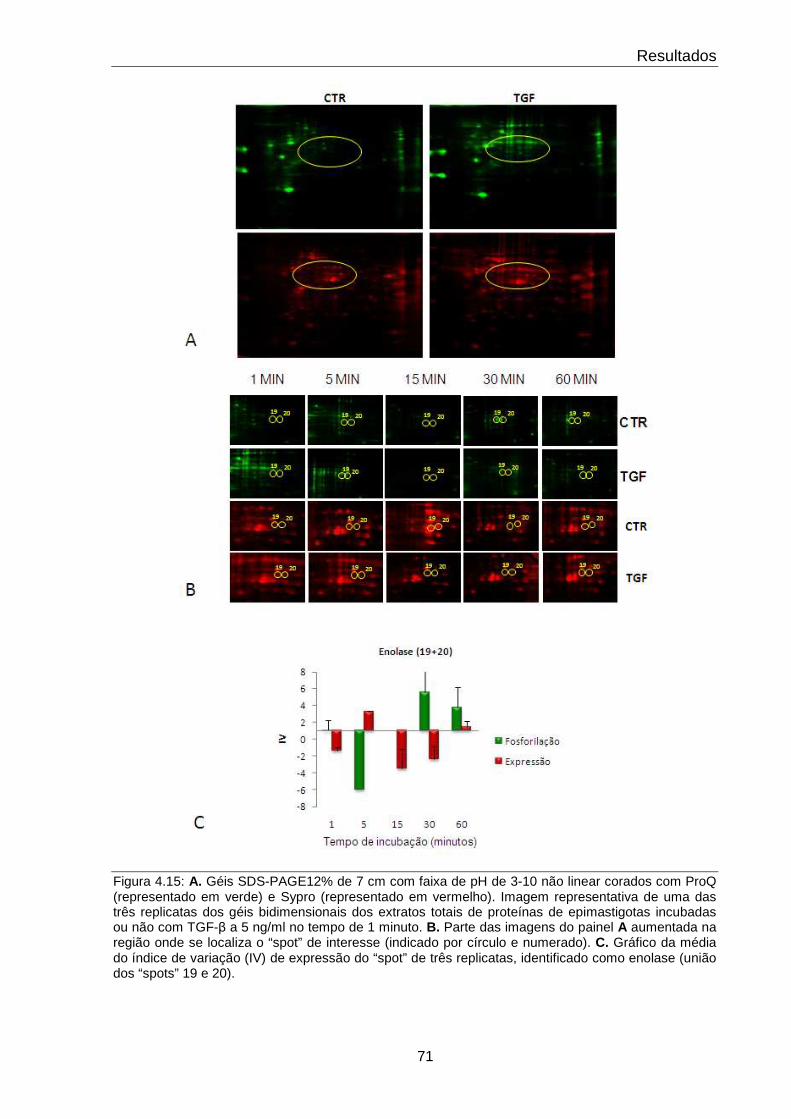
Resultados
71
Figura 4.15: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como enolase (união dos “spots” 19 e 20).

Resultados
72
O “spot” 39 (figura 4.16), identificado como gliceraldeído-3-fosfato desidrogenase
(GAPDH), tem sua fosforilação aumentada em 1 e 60 minutos e sofre defosforilação
em 5 minutos. Sua expressão é reduzida em 1 minuto e aumentada em 5 minutos. O
“spot” 51 (figura 4.16) identificado como malato desidrogenase glicossomal é
defosforilado em 1 minuto, mas tem sua fosforilação aumentada em resposta a TGF-
β nos tempos de 5, 15 e 60 minutos. Sua expressão é aumentada em 5 e 15 minutos
e reduzida em 60 minutos.
Na figura 4.17 identificamos o “spot” 31 como dissulfeto isomerase e o “spot” 32
como desidrogenase aromática L-alfa hidroxiácida. Ambos apresentam expressão
aumentada em 5 minutos e apenas o “spot” 32 apresenta expressão reduzida em 30
minutos (figura 4.17).
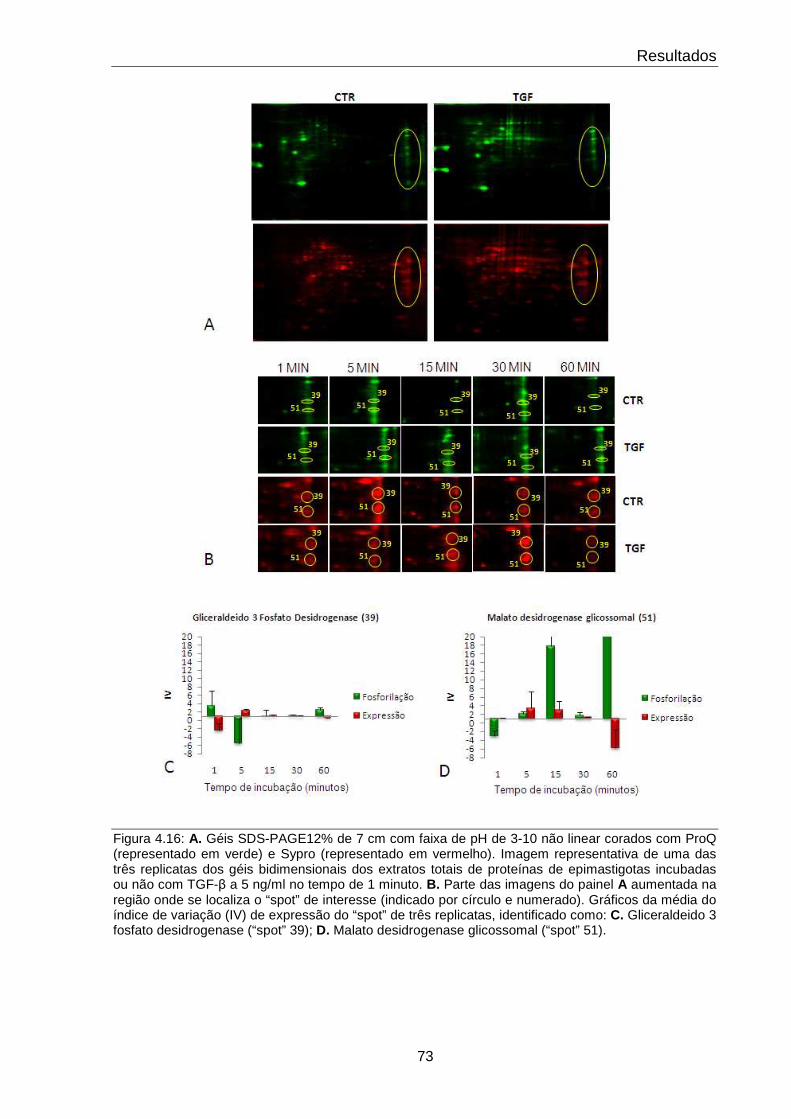
Resultados
73
Figura 4.16: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). Gráficos da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como: C. Gliceraldeido 3 fosfato desidrogenase (“spot” 39); D. Malato desidrogenase glicossomal (“spot” 51).
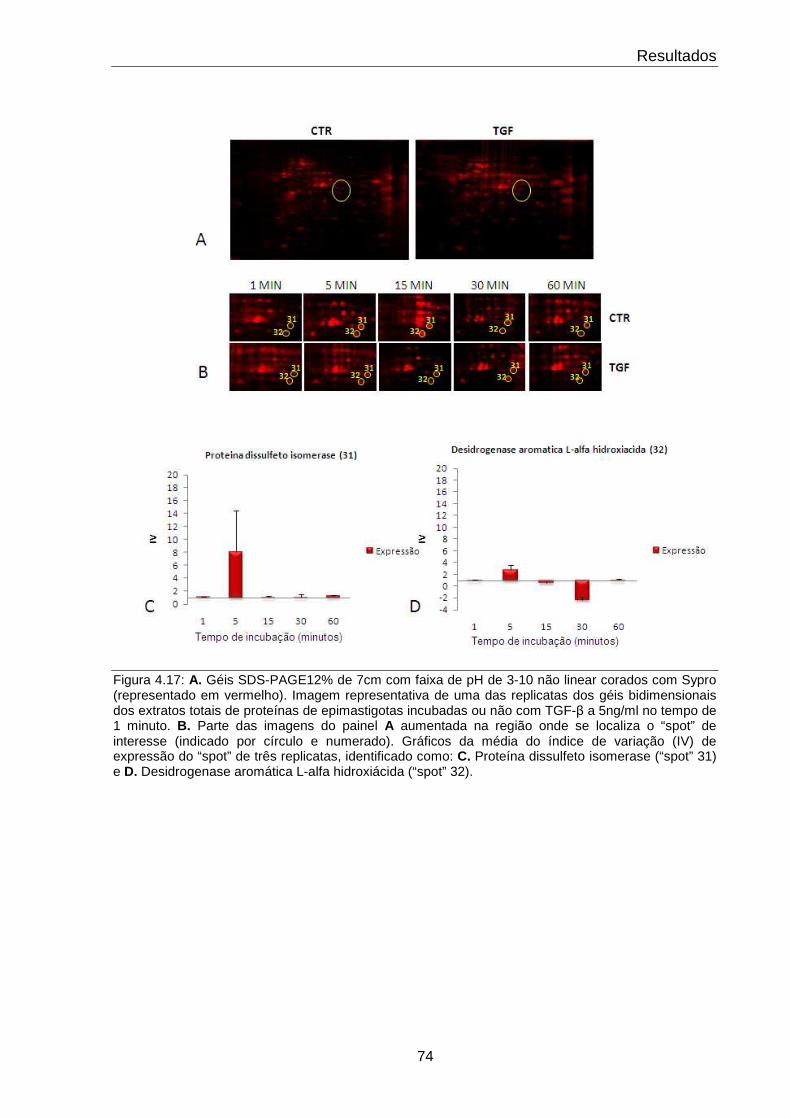
Resultados
74
Figura 4.17: A. Géis SDS-PAGE12% de 7cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). Gráficos da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como: C. Proteína dissulfeto isomerase (“spot” 31) e D. Desidrogenase aromática L-alfa hidroxiácida (“spot” 32).
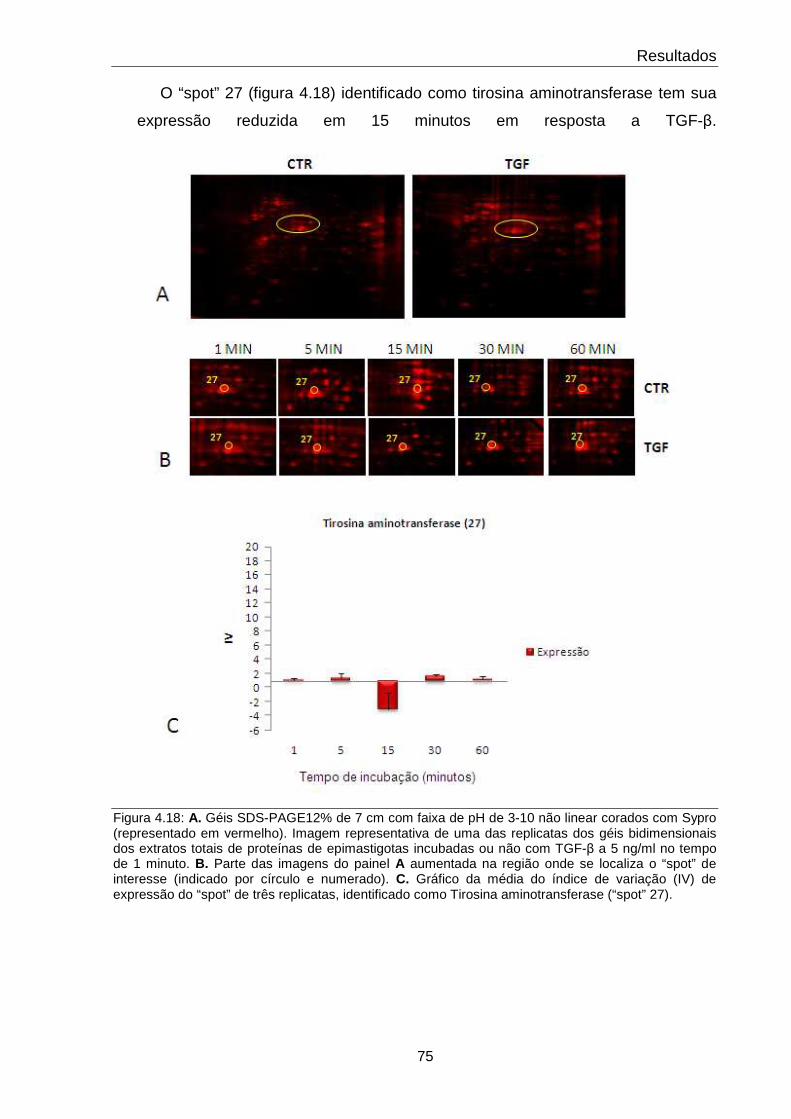
Resultados
75
O “spot” 27 (figura 4.18) identificado como tirosina aminotransferase tem sua
expressão reduzida em 15 minutos em resposta a TGF-β.
Figura 4.18: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Tirosina aminotransferase (“spot” 27).

Resultados
76
• Grupo das proteínas hipotéticas.
No mapa proteômico, foram observados três “spots” identificados como proteínas
hipotéticas (figura 4.19); dois destes “spots” protéicos foram observados como
diferencialmente fosforilados (“spots” 23 e 89) e dois diferencialmente expressos
(“spots” 17 e 23).
O “spot” 17 (figura 4.20) tem sua expressão aumentada nos tempos de 1, 5, 15 e
60 minutos e reduzida em 30 minutos.

Resultados
77
Figura 4.19: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Proteínas Hipotéticas (união dos “spots” 23 e 89).
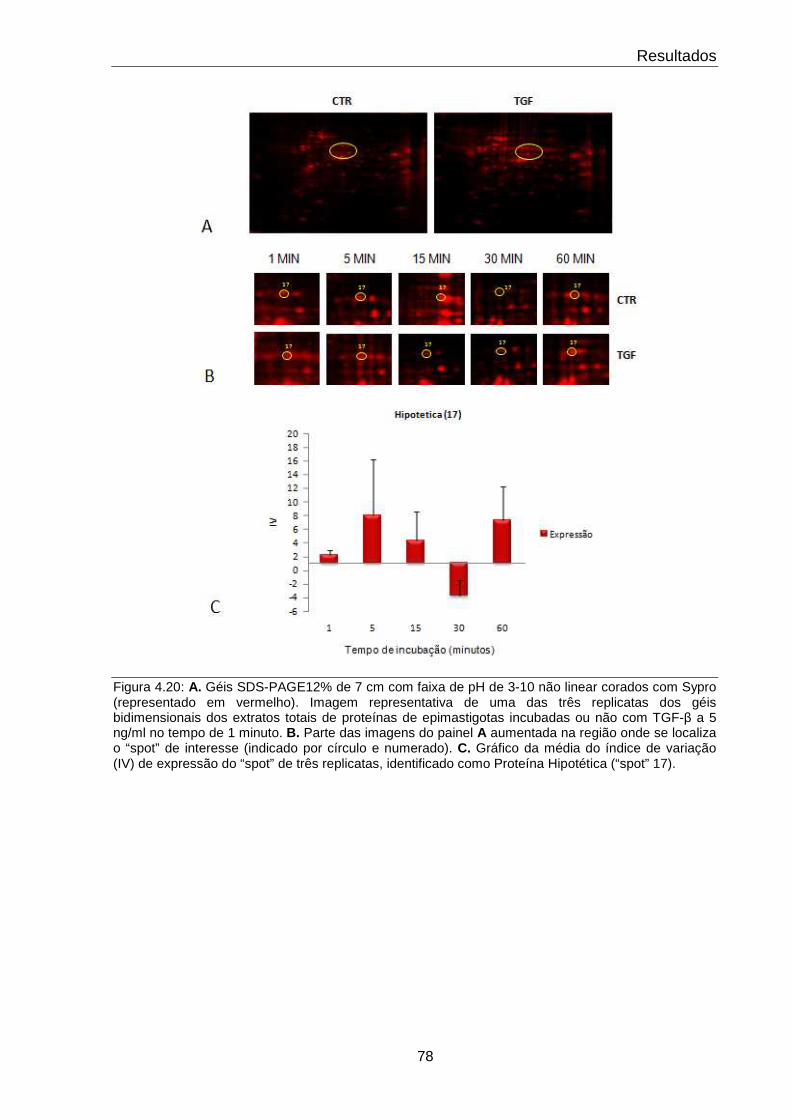
Resultados
78
Figura 4.20: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Proteína Hipotética (“spot” 17).

Resultados
79
• Proteínas associadas ao processo de tradução (fator de elongamento 1
alfa, fator de iniciação eucariótico 5a e complexo associado ao polipeptídeo
nascente).
O “spot” 40 (figura 4.21) identificado como subunidade do complexo associado ao
polipeptídeo nascente tem sua fosforilação aumentada em 1 minuto e sofre
defosforilação em resposta a TGF-β nos tempos de 5 e 15 minutos, sendo mais
marcante em 5 minutos. Já o “spot” 53 (figura 4.22) identificado como fator de
iniciação eucariótica 5a tem sua fosforilação aumentada apenas no tempo de 60
minutos em resposta a TGF-β, sem nenhuma alteração em sua expressão ao longo
do tempo estudado.
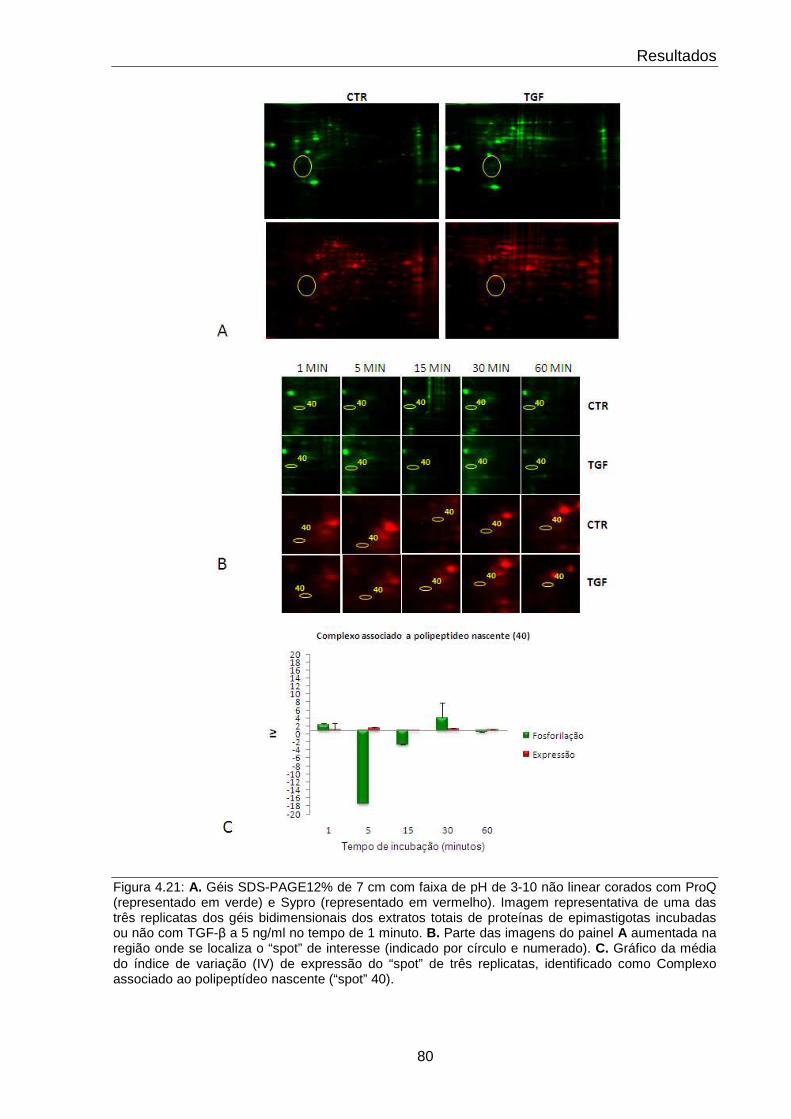
Resultados
80
Figura 4.21: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Complexo associado ao polipeptídeo nascente (“spot” 40).
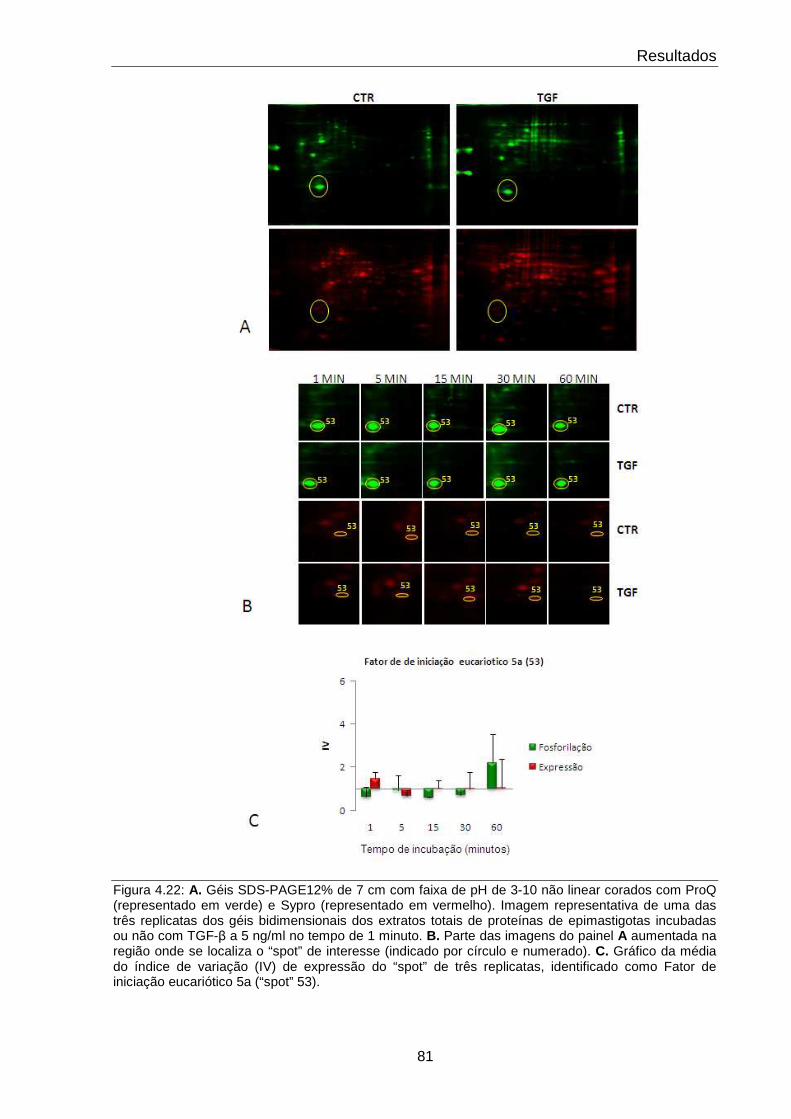
Resultados
81
Figura 4.22: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Fator de iniciação eucariótico 5a (“spot” 53).

Resultados
82
No mapa proteômico observamos dois “spots” identificados como fator de
elongamento-1α, localizados em regiões distintas do gel 2DE (regiões 1 e 2
marcadas na figura 4.23), e os resultados demonstram que ambos são
diferencialmente expressos (“spots” 32a e 38), com expressão aumentada em 5
minutos. Além disto, o “spot” 38 também apresenta modulação no tempo de 60
minutos, no qual se observa uma redução de expressão.
Figura 4.23: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). Gráficos da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como: C. Fator de elongamento-1α (“spot” 32a) e D. Fator de elongamento-1α (“spot” 38).

Resultados
83
• Proteases e Peptidases (cruzipaína, e subunidade pr oteolítica do
complexo Hslvu).
O “spot” 8 (figura 4.24) identificado como cruzipaína tem sua expressão
aumentada em 5 minutos e reduzida em 30 minutos.
O “spot” 55 (figura 4.25) identificado como subunidade proteolítica do
complexo Hslvu tem sua expressão aumentada em 5 e 15 minutos.
Figura 4.24: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Cruzipaína (“spot” 8).
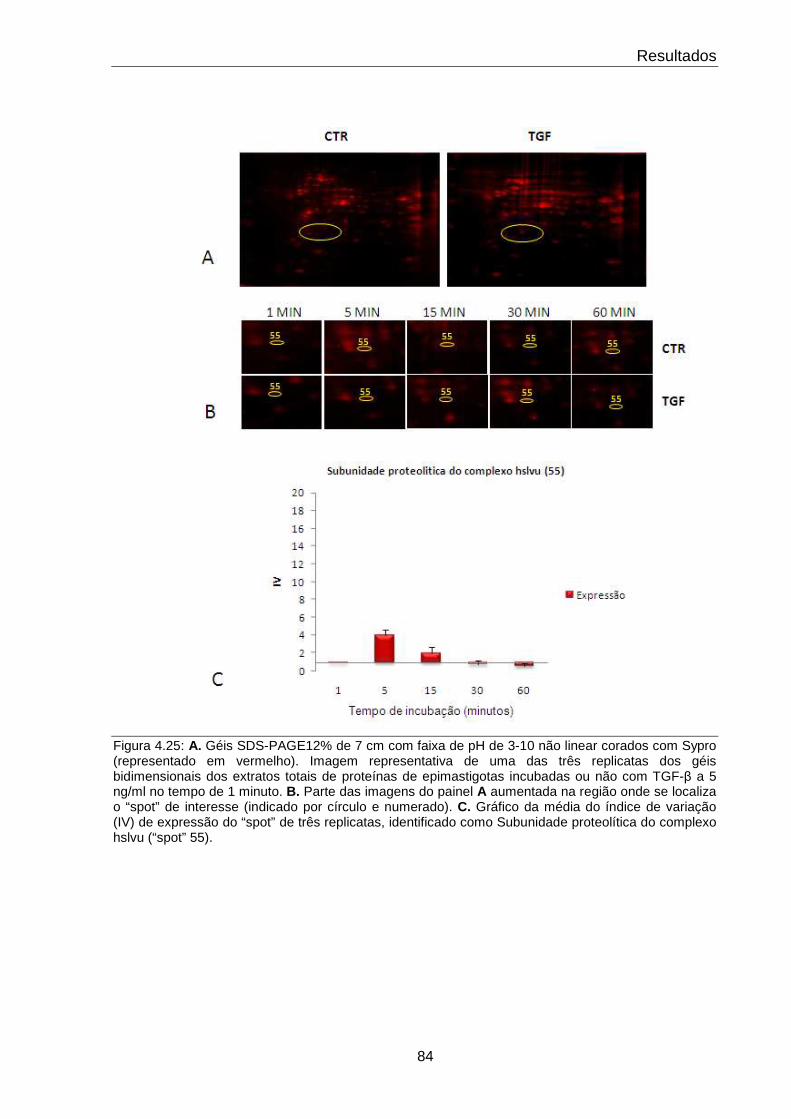
Resultados
84
Figura 4.25: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Subunidade proteolítica do complexo hslvu (“spot” 55).
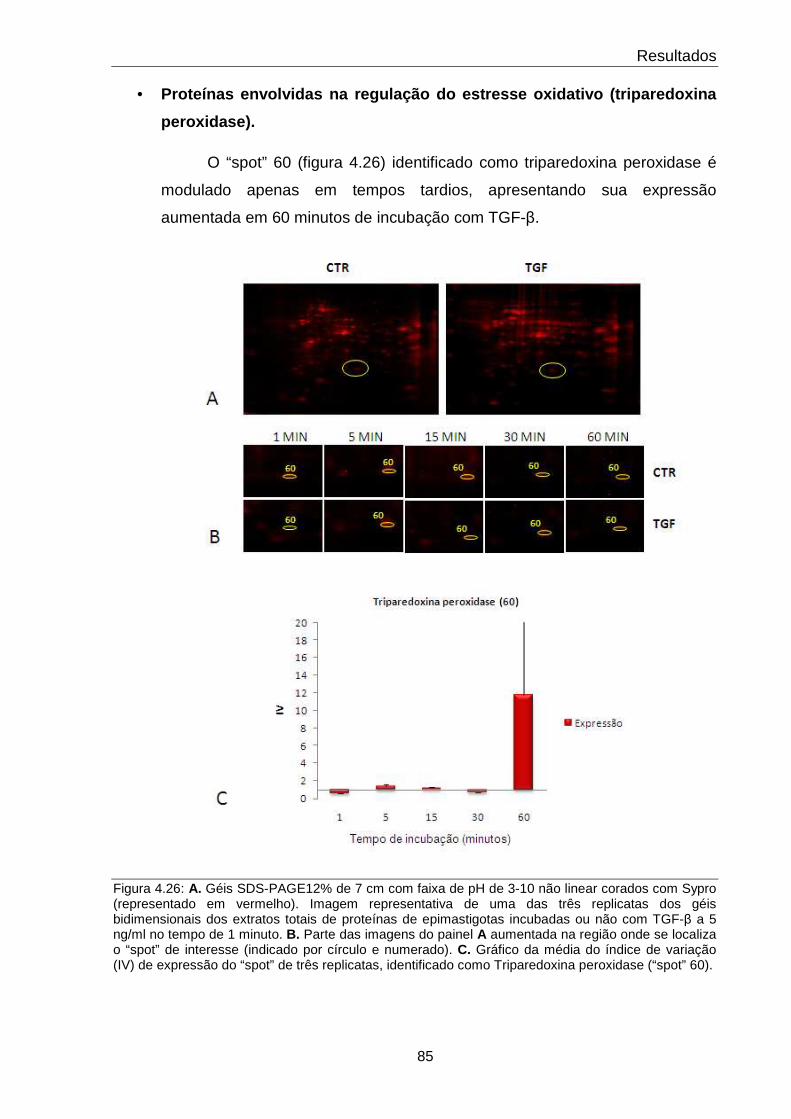
Resultados
85
• Proteínas envolvidas na regulação do estresse oxida tivo (triparedoxina
peroxidase).
O “spot” 60 (figura 4.26) identificado como triparedoxina peroxidase é
modulado apenas em tempos tardios, apresentando sua expressão
aumentada em 60 minutos de incubação com TGF-β.
Figura 4.26: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Triparedoxina peroxidase (“spot” 60).

Resultados
86
• Proteínas envolvidas na transdução de sinal (fator de liberação de
histamina dependente de IgE).
O “spot” 52 (figura 4.27) identificado como fator de liberação de histamina
dependente de IgE sofre defosforilação em 15 minutos, mas volta a ter sua
fosforilação aumentada nos tempos de 30 e 60 minutos. Sua expressão é reduzida
em 60 minutos.
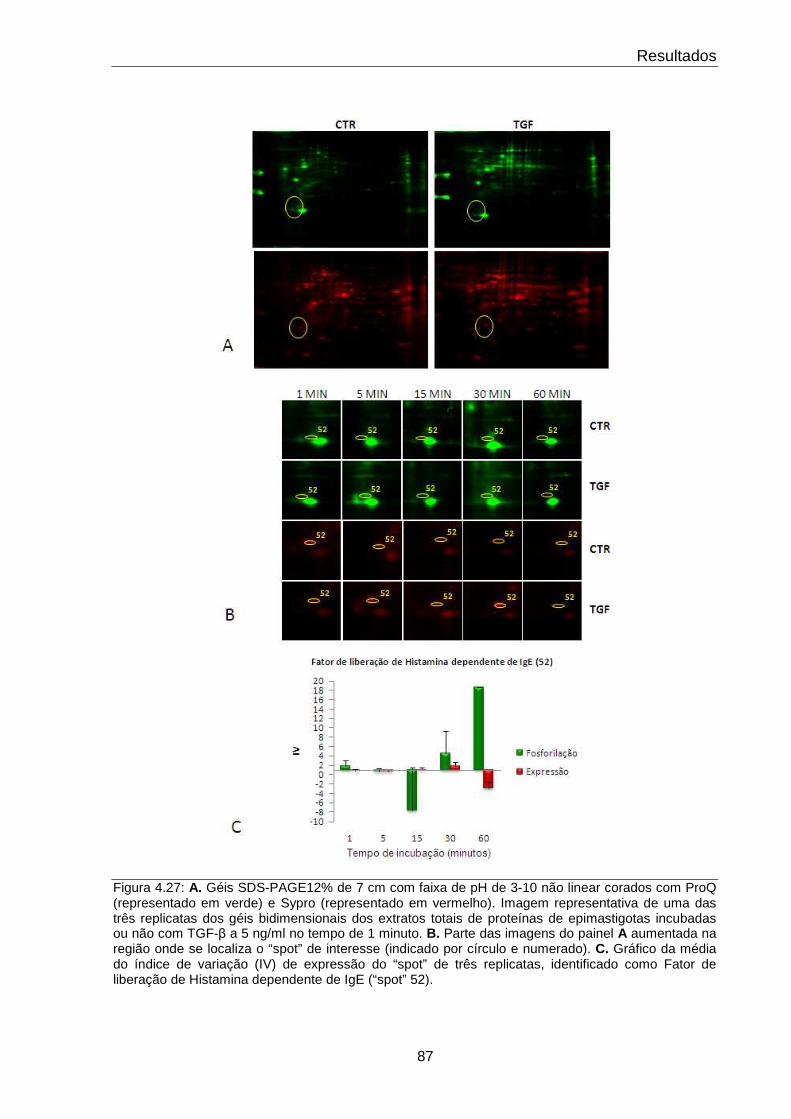
Resultados
87
Figura 4.27: A. Géis SDS-PAGE12% de 7 cm com faixa de pH de 3-10 não linear corados com ProQ (representado em verde) e Sypro (representado em vermelho). Imagem representativa de uma das três replicatas dos géis bidimensionais dos extratos totais de proteínas de epimastigotas incubadas ou não com TGF-β a 5 ng/ml no tempo de 1 minuto. B. Parte das imagens do painel A aumentada na região onde se localiza o “spot” de interesse (indicado por círculo e numerado). C. Gráfico da média do índice de variação (IV) de expressão do “spot” de três replicatas, identificado como Fator de liberação de Histamina dependente de IgE (“spot” 52).
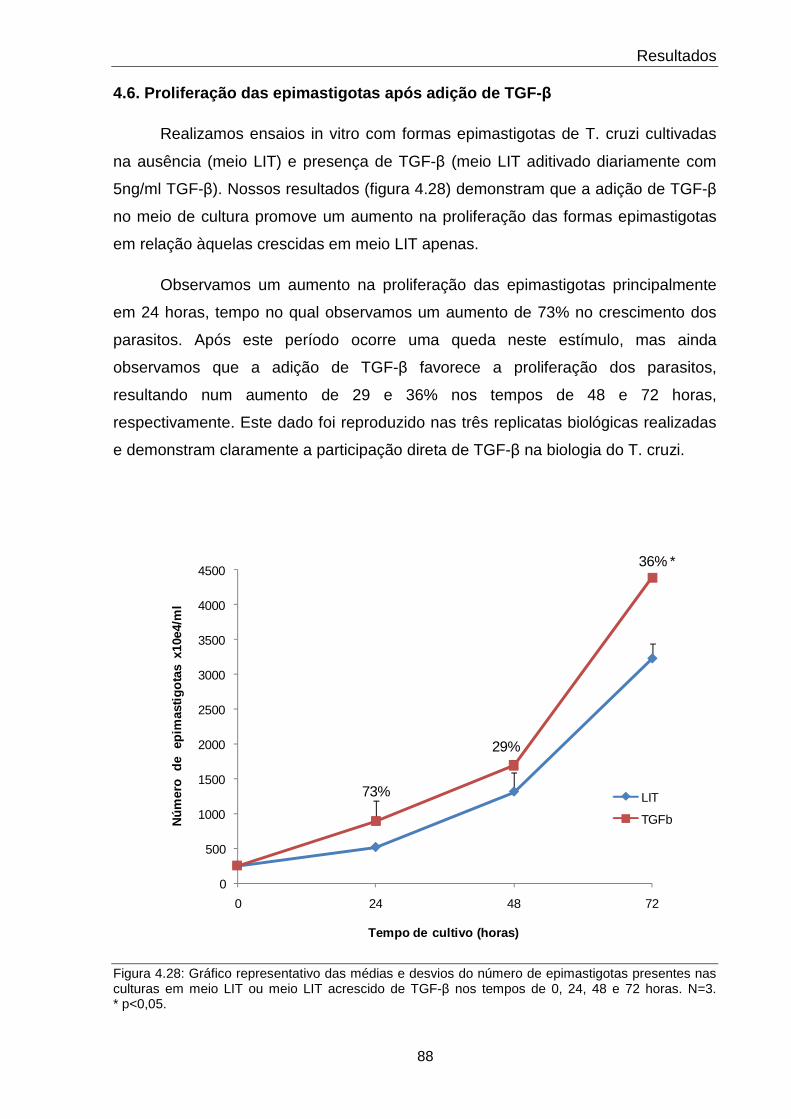
Resultados
88
4.6. Proliferação das epimastigotas após adição de TGF-β
Realizamos ensaios in vitro com formas epimastigotas de T. cruzi cultivadas
na ausência (meio LIT) e presença de TGF-β (meio LIT aditivado diariamente com
5ng/ml TGF-β). Nossos resultados (figura 4.28) demonstram que a adição de TGF-β
no meio de cultura promove um aumento na proliferação das formas epimastigotas
em relação àquelas crescidas em meio LIT apenas.
Observamos um aumento na proliferação das epimastigotas principalmente
em 24 horas, tempo no qual observamos um aumento de 73% no crescimento dos
parasitos. Após este período ocorre uma queda neste estímulo, mas ainda
observamos que a adição de TGF-β favorece a proliferação dos parasitos,
resultando num aumento de 29 e 36% nos tempos de 48 e 72 horas,
respectivamente. Este dado foi reproduzido nas três replicatas biológicas realizadas
e demonstram claramente a participação direta de TGF-β na biologia do T. cruzi.
Figura 4.28: Gráfico representativo das médias e desvios do número de epimastigotas presentes nas culturas em meio LIT ou meio LIT acrescido de TGF-β nos tempos de 0, 24, 48 e 72 horas. N=3. * p<0,05.
0
500
1000
1500
2000
2500
3000
3500
4000
4500
0 24 48 72
Núm
ero
de
epi
mas
tigot
as x
10e4
/ml
Tempo de cultivo (horas)
LIT
TGFb
73%
29%
36% *

Discussão
89
5. DISCUSSÃO
A citocina TGF-β desempenha importantes funções, tanto na doença de
Chagas, como no ciclo de vida de T. cruzi. De forma sucinta, os dados atuais da
literatura indicam que TGF-β participa em pelo menos quatro diferentes processos
que influenciam o desenvolvimento da miocardiopatia da doença de Chagas: (a)
invasão de fibroblastos e miócitos cardíacos, (b) ciclo intracelular do parasita, (c)
regulação da inflamação e da resposta imunológica, (d), fibrose e remodelamento
cardíaco durante a doença aguda e crônica (Araujo-Jorge et al., 2008).
Com relação ao seu papel no ciclo intracelular de T. cruzi, TGF-β latente é
ativado pelo parasito na superfície da célula hospedeira (Waghabi et al., 2005b),
favorecendo sua invasão celular através da sinalização via receptores de TGF-β de
superfície (Ming et al., 1995; Waghabi et al., 2005b). Amastigotas de T. cruzi são
capazes de captar e estocar TGF-β da célula hospedeira e usar esta molécula para
regular a transição entre parada da proliferação e início da diferenciação para
tripomastigotas (Waghabi et al., 2005a). Apesar da importância desta citocina na
biologia do T. cruzi, ainda não são conhecidos os mecanismos moleculares exatos
de transdução dos sinais gerados por TGF-β, que o permitem exercer seus efeitos
finais no microorganismo.
Por essa razão, em nosso estudo visamos caracterizar os padrões de
fosforilação e expressão de proteínas de epimastigotas de T. cruzi em resposta a
adição de TGF-β e identificar moléculas do parasito que estejam relacionadas à via
de sinalização disparada por esta citocina.
Para isso, utilizamos uma abordagem fosfoproteômica, que permite analisar,
em um único experimento, um conjunto de proteínas possivelmente envolvidas na
via de sinalização de interesse. A fosforilação reversível de proteínas é um
mecanismo essencial para a regulação de diversos processos biológicos, incluindo,
enovelamento, transdução de sinal, atividade enzimática, proliferação, divisão e
diferenciação celular, entre outros (Paradela & Albar, 2008). A sinalização por TGF-β
envolve a fosforilação de uma série de moléculas, incluindo seus receptores e
proteínas transdutoras de sinal, que permite a ativação de sua via, e logo, a
efetivação de seu efeito final na célula. Assim, a caracterização do fosfoproteoma de
T. cruzi nas nossas condições de estudo se torna objeto de grande utilidade para a
identificação de uma série de proteínas responsivas a TGF-β, que podem participar

Discussão
90
de sua via intracelular de transdução de sinal. O uso do corante fluorescente ProQ
Diamond (Molecular Probes) viabilizou a detecção das fosfoproteínas nos géis
bidimensionais com alta sensibilidade e linearidade. A coloração seguinte dos géis
com o corante fluorescente Sypro Ruby (Molecular Probes) permitiu a avaliação do
grau de expressão das proteínas presentes no gel, estivessem elas fosforiladas ou
não.
Em nosso trabalho foi utilizada a forma evolutiva epimastigota de T. cruzi
(cepa Y). O uso desta forma é ideal para o desenvolvimento da tecnologia
proteômica, pois qualquer tipo de contaminante proveniente de outros tipos celulares
presente na amostra pode interferir seriamente na interpretação dos resultados. As
formas amastigotas e tripomastigotas são obtidas a partir de culturas de células de
mamíferos, sendo praticamente impossível obter total separação entre os
componentes celulares dos parasitos daqueles dos mamíferos. Em contrapartida, as
epimastigotas são mantidas em culturas axênicas, garantindo que todo material
avaliado pertença exclusivamente ao parasito e possibilitando um maior controle e
reprodutibilidade dos experimentos. Além disso, trabalhos recentes do grupo
demonstraram por diferentes tipos de ensaios biológicos que as formas
epimastigotas de T. cruzi também são capazes de responder ao estímulo de TGF-β
presente nas culturas (Waghabi, M.C., comunicação pessoal). A coleta dos parasitos
foi feita sempre no mesmo dia de cultivo e ainda na fase de crescimento
exponencial, assegurando que os parasitos estejam em plena atividade biológica,
condição fundamental para a realização de nossos ensaios experimentais.
Inicialmente verificamos que a dose ideal para indução de fosforilação das
proteínas de epimastigotas de T. cruzi é de 5 ng/ml. A escolha da dose foi feita a
partir da análise do perfil unidimensional dos extratos protéicos de T. cruzi, e por
esta abordagem, observamos apenas diferenças sutis no padrão diferencial das
bandas. Talvez a aplicação da metodologia bidimensional gerasse resultados mais
precisos do que aqueles obtidos, mas este processo seria extenso demais para o
período de realização do nosso trabalho. Dessa forma, resolvemos nos basear nos
resultados das imagens unidimensionais e optamos pela dose que mostrou uma
fosforilação um pouco mais acentuada, que é a de 5 ng/ml. Esta dose também
coincide com aquela usada pelo grupo em outros ensaios experimentais com as
diferentes formas de T.cruzi (Waghabi, M.C., comunicação pessoal).

Discussão
91
Determinamos em seguida o tempo ótimo de incubação das epimastigotas a
4ºC com TGF-β (5 ng/ml), capaz de induzir o maior número de eventos de
fosforilação nas proteínas do parasito. As análises em géis unidimensionais não
foram capazes de discriminar diferenças significativas entre os tempos estudados.
Sendo assim, optamos por realizar os experimentos em géis bidimensionais,
empregando todos os tempos acima.
As imagens dos géis bidimensionais mostraram que os perfis protéicos entre
os tempos estudados apresentavam diferenças na fosforilação e expressão em
função da adição de TGF-β. Esta informação indicou que cada tempo de estudo
poderia estar mostrando uma etapa da cascata de eventos disparada pela adição de
TGF-β no meio, e sendo assim, optamos por manter um estudo de cinética de
tempo.
O uso de “strips” com faixa de pH 3 a 10 é bastante abrangente, o que
permitiu a visualização da maioria das proteínas em um único gel. Como muitas
proteínas são focalizadas na região central da faixa de pH 3 a 10, usamos
gradientes não-lineares (NL), pois os mesmos permitem uma melhor resolução das
proteínas na faixa central de pH.
Cada experimento bidimensional foi realizado em triplicatas biológicas, para
que somente os dados reprodutíveis fossem selecionados. Notamos que no tempo
de 1 minuto houve a indução do maior número de “spots” fosforilados, tanto na
amostra não tratada (controle), quanto na tratada com TGF-β. A realização dos
experimentos a 4ºC retarda o desencadeamento de eventos da cascata de
sinalização disparada por TGF-β, e tal fato, permitiu a visualização dos efeitos
marcantes sobre a fosforilação de determinados grupos de proteínas em 1 minuto de
incubação. O fato de que em 1 minuto também se observa a maior indução de
fosforilação nas proteínas de epimastigotas não tratadas com TGF-β (controle) pode
ser explicada como uma resposta do parasito para se adaptar ao novo ambiente,
com temperatura mais baixa. A fosforilação ainda mais abundante nas proteínas de
epimastigotas tratadas com TGF-β por 1 minuto a 4ºC pode ser dada pela união de
dois novos fatores introduzidos: resposta rápida à baixa na temperatura e adição de
TGF-β no meio. Nos tempos mais longos de incubação (30 e 60 minutos) observa-se
que a fosforilação das proteínas de ambas as amostras (tratadas ou não com TGF-
β) é menos acentuada e que seus perfis tendem a se igualar. Este comportamento

Discussão
92
pode estar refletindo o término dos eventos moleculares relacionados à adição de
TGF-β ao meio. A resposta de adaptação ao choque térmico, neste momento, pode
ou não estar encerrada ou em vias de finalização.
Um total de 463 “spots” foram retirados de géis de 7 ou 17 cm corados com
Coomassie coloidal. Destes, 156 “spots” obtiveram identificação positiva, dos quais
38 foram destacados por apresentarem fosforilação e/ou expressão mais de duas
vezes aumentada(s) ou diminuída(s) em resposta a TGF-β. Entre os analisados em
nosso trabalho, 11 “spots” foram identificados como proteínas de choque térmico, 2
como proteínas de citoesqueleto, 14 como proteínas envolvidas em vias
metabólicas, 4 como proteínas associadas ao processo de tradução, 2 como
proteases/peptidases, 1 como proteína de regulação de estresse oxidativo, 1 como
uma proteína envolvida na transdução de sinal e 3 como proteínas hipotéticas.
É válido ressaltar que para a maioria dos “spots” responsivos a TGF-β, a
confirmação da identificação foi feita com pelo menos mais uma replicata biológica.
Além disso, as identificações obtidas são confirmadas por dados de outros mapas
proteômicos de T. cruzi obtidos em nosso laboratório (dados não publicados), e
também por outros mapas proteômicos de T. cruzi presentes na literatura (Paba et
al., 2004; Parodi-Talice et al., 2004; Sodre et al., 2009).
O projeto de sequenciamento do genoma de T. cruzi foi feito com base na
cepa CL Brener, que é um híbrido de dois genótipos possuindo, assim, alelos
distintos para a maioria dos genes. Mais de 50% do seu genoma consiste em
sequências repetidas, como retrotransposons e genes para grandes famílias de
moléculas de superfície, que incluem as trans-sialidases, mucinas, gp63s e uma
nova grande família (mais de 1300 cópias) de proteínas de superfície associadas à
mucina (El-Sayed et al., 2005).
As análises dos genomas de T. cruzi, T. brucei e Leishmania major (Tritryp)
mostram que esses organismos diferem de outros eucariotos no que diz respeito ao
seu sistema de reparo de DNA e ao início da replicação, reflexo do seu DNA
mitocondrial incomum. Embora o Tritryp não tenha identificado várias classes de
moléculas de sinalização, seus quinomas contêm um grande e diversificado conjunto
de proteínas quinases e fosfatases, cujo número e diversidade implicam em
interações previamente desconhecidas e em processos regulatórios, que podem ser
alvos de intervenção farmacológica (Atwood et al., 2005). Recentemente um grupo

Discussão
93
de pesquisadores (Nakayasu et al., 2009) identificou, por espectrometria de massas,
237 fosfopeptídeos provenientes de 119 proteínas distintas de epimastigotas de T.
cruzi. Os resultados indicam que a propagação de cascatas de sinalização celular
por proteínas quinases e fosfatases desempenha um papel importante nos
processos fisiológicos de T. cruzi, incluindo a motilidade celular, metabolismo,
transporte de íons, diferenciação e sobrevivência.
É interessante lembrar que o tratamento da doença de Chagas é atualmente
limitado a dois medicamentos, o nifurtimox e o benznidazol, que têm eficácia limitada
e causam efeitos colaterais graves (Urbina & Docampo, 2003; Wilkinson et al.,
2008). Além disso, não existe até o momento uma vacina contra T. cruzi para uso
em humanos (Garg & Bhatia, 2005; Dumonteil, 2007). Portanto, há uma necessidade
crítica de desenvolver novas estratégias terapêuticas para prevenir ou tratar a
doença de Chagas. Em consonância com o interesse de desenvolver novos
fármacos, a verificação de que T. cruzi possui uma série de quinases que participam
de vias metabólicas não encontradas nos hospedeiros vertebrados, tem tornado
essas proteínas alvos moleculares interessantes para o tratamento da doença de
Chagas (Doerig, 2004; Nakayasu et al., 2009).
A análise das imagens dos géis bidimensionais pelo programa PDQuest
(BioRad) permitiu a atribuição de um valor numérico para o grau de fosforilação e
expressão de cada um dos “spots”. Apesar dos resultados gerarem valores precisos,
sabemos que a técnica apresenta limitações por ser baseada na quantificação de
proteínas através da avaliação de imagens. Este método de quantificação pode não
ser preciso o suficiente para gerar afirmações numéricas exatas. Deste modo, com o
intuito de tornar nossas análises menos suscetíveis a erros, nos concentramos
apenas na indicação de aumento ou redução da fosforilação e/ou expressão das
proteínas em estudo.
Nosso estudo identificou uma série de proteínas cuja fosforilação e/ou
expressão parece estar alterada após a incubação de formas epimastigotas, por
diferentes tempos, com TGF-β a 5 ng/ml. Essas proteínas são representadas nos
géis bidimensionais por um ou mais “spots”. Nos casos onde diferentes “spots” (de
mesma massa molecular, mas com pI diferentes) foram identificados para uma
mesma proteína com idênticas sequências de aminoácidos, seus valores foram
somados, na tentativa de avaliar o comportamento geral da proteína.

Discussão
94
Os padrões de fosforilação e expressão das proteínas de epimastigotas de T.
cruzi em resposta a adição de TGF-β foram determinados nos tempos de 1, 5, 15,
30 e 60 minutos de incubação a 4ºC. Tendo em vista os tempos de estudo podem
representar etapas sequenciais da resposta induzida por TGF-β, é possível associar
as proteínas que apresentaram fosforilação e/ou expressão alteradas em um tempo
de estudo com determinado passo da via, e ainda, propor interações e/ou
regulações das proteínas alteradas em tempos tardios pelas proteínas moduladas
nos tempos mais curtos de incubação. As proteínas cuja fosforilação e/ou expressão
foram moduladas em um ou mais tempo de estudo estão representadas na tabela
5.1.
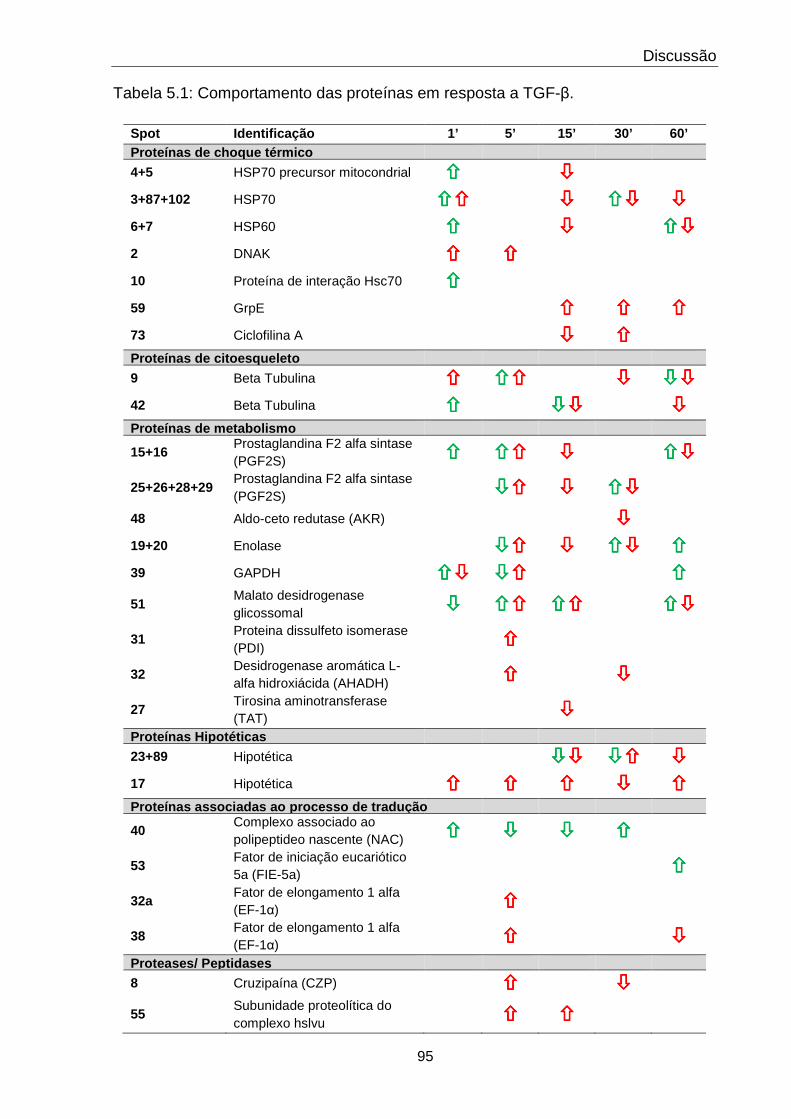
Discussão
95
Tabela 5.1: Comportamento das proteínas em resposta a TGF-β.
Spot Identificação 1’ 5’ 15’ 30’ 60’ Proteínas de choque térmico 4+5 HSP70 precursor mitocondrial ���� ����
3+87+102 HSP70 �������� ���� �������� ����
6+7 HSP60 ���� ���� ��������
2 DNAK ���� ����
10 Proteína de interação Hsc70 ����
59 GrpE ���� ���� ����
73 Ciclofilina A ���� ����
Proteínas de citoesqueleto 9 Beta Tubulina ���� �������� ���� ��������
42 Beta Tubulina ���� �������� ����
Proteínas de metabolismo 15+16
Prostaglandina F2 alfa sintase (PGF2S) ���� �������� ���� ��������
25+26+28+29 Prostaglandina F2 alfa sintase (PGF2S)
�������� ���� ��������
48 Aldo-ceto redutase (AKR) ����
19+20 Enolase �������� ���� �������� ����
39 GAPDH �������� �������� ����
51 Malato desidrogenase glicossomal ���� �������� �������� ��������
31 Proteina dissulfeto isomerase (PDI)
����
32 Desidrogenase aromática L-alfa hidroxiácida (AHADH)
���� ����
27 Tirosina aminotransferase (TAT)
����
Proteínas Hipotéticas 23+89 Hipotética �������� �������� ����
17 Hipotética ���� ���� ���� ���� ����
Proteínas associadas ao processo de tradução
40 Complexo associado ao polipeptideo nascente (NAC) ���� ���� ���� ����
53 Fator de iniciação eucariótico 5a (FIE-5a)
����
32a Fator de elongamento 1 alfa (EF-1α)
����
38 Fator de elongamento 1 alfa (EF-1α)
���� ����
Proteases/ Peptidases
8 Cruzipaína (CZP) ���� ����
55 Subunidade proteolítica do complexo hslvu
���� ����

Discussão
96
Proteínas de regulação de estresse oxidativo 60
Triparedoxina peroxidase (TriX)
����
Proteínas de transdução de sinal 52
Fator de liberação de histamina dependente de IgE (FLHdIgE)
���� ���� ���� ��������
Tabela 5.1: tabela mostrando o comportamento nos padrões de fosforilação (setas verdes) e/ou expressão (setas vermelhas) de cada proteína ao longo dos tempos de estudo, em função da adição de TGF-β. O(s) número(s) do(s) “spot(s)” que representa(m) cada proteína está(ão) indicado(s) na coluna à esquerda. As proteínas estão ordenadas de acordo com as categorias funcionais indicadas nos resultados.
As proteínas de choque térmico foram inicialmente caracterizadas como
proteínas responsivas a alterações bruscas na temperatura ambiente e a situações
de estresse celular, mas também desempenham funções vitais no funcionamento
celular, como enovelamento, montagem, localização intracelular, secreção,
regulação e endereçamento de proteínas instáveis para a via de degradação
proteolítica (Bukau & Horwich, 1998; Nollen & Morimoto, 2002; Young et al., 2004;
Mayer & Bukau, 2005). As HSP70 são constitutivamente expressas e estão
localizadas em uma variedade de compartimentos celulares, como citoplasma,
membrana, no retículo endoplasmático, nas mitocôndrias e nos cloroplastos. O
precursor mitocondrial HSP70 (MPHSP70) exerce a função molecular de chaperona
junto a proteínas precursoras mitocondriais. Esses precursores são devidamente
enovelados pela MPHSP70 no citoplasma, o que permite sua translocação à matriz
mitocondrial, onde exercerão suas funções. A Hsc70 interage com a HSP70,
participando no processo de enovelamento protéico. Uma nova função putativa foi
sugerida para estas proteínas, mostrando que as HSP70 e Hsc70 de mamíferos são
capazes de se ligar seletivamente a certas sequências de RNA através do seu
domínio N-terminal de ligação ATP. A ligação a molécula de RNA parece ser
influenciada pela atividade da co-chaperonas HSP40 e pelo co-fator Bag (Zimmer et
al., 2001).
Trabalhos recentes demonstraram que TGF-β e chaperonas podem estar
diretamente relacionados. Bahn e colaboradores (2007) adicionaram TGF-β a
explantes de células epiteliais de ratos previamente aquecidos a 45ºC. Eles
verificaram que as chaperonas desempenham papel protetor na transição epitélio-
mesênquima induzida por TGF-β e estimulam a sobrevivência celular. Xin e
colaboradores (2005) evidenciaram um novo mecanismo para regulação da
sensibilidade de TGF-β dependente de CHIP (região carboxi-terminal da proteína de

Discussão
97
interação Hsc70). CHIP regula os níveis basais de Smad 3, através da degradação
ubiquitina-dependente desta proteína. Assim, CHIP é capaz de controlar a amplitude
da resposta celular induzida por TGF-β.
O rápido aumento na fosforilação das proteínas de choque térmico (HSP60,
HSP70 e HSP70 precursor mitocondrial e da proteína de interação Hsc70) pode ser
explicado como uma resposta combinada do parasito, onde o mesmo busca se
adaptar à variação de temperatura do meio (mudança de 28 para 4°C), e também
responder rapidamente ao estímulo com TGF-β. As proteínas de choque térmico
podem estar atuando como reguladores de outras moléculas, possivelmente
alterando suas conformações, o que pode resultar na translocação dessas
moléculas do citoplasma para outras organelas, mudança de conformação para um
estado ativo ou endereçamento para vias de degradação. A ligação às moléculas de
RNA pode, por sua vez, estar regulando positiva ou negativamente a expressão de
outras proteínas (através da estabilização ou desestabilização de mRNAs).
Nos procariotos, a proteína equivalente a HSP70 é denominada DnaK, e esta
também exerce a função de chaperona e de ligação ao RNA. O aumento rápido na
expressão dessas proteínas está provavelmente influenciando o aumento nas
atividades já descritas para as proteínas de choque térmico. A GrpE é o co-fator da
DnaK, auxiliando a proteína de choque térmico no enovelamento correto de
proteínas. A DnaK parece ser capaz de se ligar a certas sequências de RNA, em um
processo influenciado pela atividade da GrpE (Zimmer et al., 2001).
O aumento em tempos mais tardios na expressão da GrpE indica que ela
pode estar sendo regulada positivamente por outras moléculas responsivas ou
ativadas por TGF-β. Sua atividade deve estar relacionada tanto ao correto
enovelamento de proteínas, como também a um possível papel na estabilização de
moléculas de RNA, regulando a taxa de transcrição, e consequentemente, a
expressão de determinadas proteínas.
As ciclofilinas (ou peptidil-prolil isomerases - PPIases) pertencem a um grupo
de proteínas com atividade peptidil cis-trans isomerase, conhecidas coletivamente
como imunofilinas. As PPIases são enzimas que aceleram o enovelamento de
proteínas, catalisando a isomerização das ligações peptídicas anteriores aos
resíduos de prolina. As ciclofilinas são encontradas em todas as células de todos os
organismos procariotos e eucariotos estudados, entre eles, mamíferos, plantas,

Discussão
98
insetos, fungos e bactérias (Marks, 1996). As ciclofilinas são proteínas de 18 kDa,
estruturalmente conservadas, que compartilham um domínio comum de
aproximadamente 109 aminoácidos, cercado por domínios exclusivos de cada
membro da família, associados a compartimentalização subcelular e a
especialização funcional (Arevalo-Rodriguez et al., 2004). Estudos recentes têm
indicado uma série de funções celulares adicionais para as ciclofilinas, incluindo
papéis na sinalização celular e como chaperonas (Wang & Heitman, 2005).
A redução na sua expressão em 15 minutos de incubação com TGF-β pode
indicar que esta proteína esteja sendo negativamente regulada por outra molécula
direta ou indiretamente ativada por TGF-β em algum dos tempos anteriores de
estudo. A redução na sua atividade pode acarretar no retardamento do
enovelamento protéico, e assim, atrasar o correto endereçamento e ativação de
proteínas por ela reguladas. Pela sua função de chaperona, a redução na sua
expressão pode levar a diminuição na marcação de proteínas para vias de
degradação, gerando um aumento na abundância de certas proteínas. Contudo em
30 minutos, a sua expressão é aumentada sugerindo retorno da sua atividade
fisiológica.
Com relação às proteínas de citoesqueleto, temos que as α e β-tubulinas são
os principais componentes dos microtúbulos. As tubulinas delta são encontradas em
diversas localidades celulares, mas ao contrário das tubulinas α e β, elas não são
igualmente distribuídas entre os eucariotos. As delta tubulinas já foram localizadas
nos centríolos e desempenham uma função na formação do fuso mitótico durante a
mitose (Dutcher, 2001). Essas proteínas sofrem uma série de modificações pós-
traducionais, incluindo acetilação, tirosinação, fosforilação, poliglutaminação e
poliglicilação (Chapin & Bulinski, 1991). Os microtúbulos são os principais
componentes do citoesqueleto dos tripanossomatídeos e desempenham um papel
importante nas mudanças morfológicas associada do seu ciclo de vida (Kohl & Gull,
1998). O citoesqueleto dos tripanossomas é composto por um arranjo complexo de
microtúbulos, que incluem a rede subpelicular, o axonema, o corpo basal, o eixo
para-flagelar, a zona de aderência ao flagelo e os filamentos responsáveis pela
fixação do parasita às células hospedeiras (Robinson et al., 1991).
Alterações na expressão das tubulinas ocorrem durante o ciclo de vida dos
tripanosomatídeos, e são responsáveis por uma série de adaptações bioquímicas e

Discussão
99
morfológicas sofridas pelos parasitos. Estudos mostraram que a quantidade de
transcritos de α e β-tubulinas parece ser de 3 a 6 vezes maior em epimastigotas do
que em tripomastigotas e amastigotas, apesar de suas taxas de transcrição serem
bastante similares em epimastigotas e amastigotas (Bartholomeu et al., 2002). Um
estudo sobre a expressão dos genes de tubulina durante o crescimento de formas
epimastigotas do T. cruzi por Northern blot mostrou que há uma diminuição nos
níveis de mRNAs de α e β-tubulinas quando as epimastigotas passam da fase
logarítmica para a fase estacionária de crescimento (Gonzalez-Pino et al., 1999). As
alterações foram associadas com uma diminuição semelhante nas taxas de
transcrição para ambos os genes. Este estudo observou ainda que a expressão de
β-tubulina parece ser controlada em nível pós-transcricional.
A proteína β-tubulina é representada em nosso mapa proteômico pelos
“spots” 9 e 42, que apresentam comportamentos diferentes em seus padrões de
fosforilação e expressão ao longo de nosso tempo de estudo. Considerando que a β-
tubulina é uma proteína encontrada em diversas estruturas do parasito, e que os
dois “spots” que a representam em nosso mapa possuem pesos moleculares e
pontos isoelétricos bem distintos, é possível que estejamos analisando β-tubulinas
de diferentes localizações celulares, que exercem diferentes funções na estrutura
celular do parasito. O aumento rápido na expressão da β-tubulina representada pelo
“spot” 9 pode indicar que a mesma já existia pronta em vesículas ou organelas de
reserva. Um aumento rápido na taxa de transcrição de seu RNA também é possível,
apesar de parecer pouco provável pelo curto período de tempo que o parasito teria
para re-organizar sua maquinaria replicativa. Outra explicação seria a da redução na
taxa de degradação dos mRNAs, o que levaria a um aumento na quantidade de
proteínas traduzidas pela célula. A redução mais tardia na expressão de ambos os
“spots” pode ser por causa da regulação negativa das β-tubulinas por parte de
outras moléculas envolvidas direta ou indiretamente na via de sinalização por TGF-
β. Em resposta a TGF-β, as β-tubilinas podem estar alterando a organização dos
microtúbulos, permitindo a ocorrência de novas interações entre as moléculas das
epimastigotas.
Entre as proteínas envolvidas no metabolismo celular, a prostaglandina F2alfa
sintase (PGFS) é uma aldo-ceto redutase que pertence à família oxidorredutase
flavina-dependente (Kubata et al., 2000). Esta enzima localiza-se no citoplasma das
epimastigotas (Blehert et al., 1999) e, além da produção de prostaglandinas, tem a

Discussão
100
capacidade de reduzir tanto agentes tripanocidas (nifurtimox e outros) como
hidroperóxidos. Tem sido sugerido que a Prostaglandina F2 alfa sintase de T. cruzi
(TcPGFS) seja responsável pela maior parte da atividade antioxidante neste parasito
(Kubata et al., 2000). A PGFS catalisa a redução dependente de NADPH da 9,11-
endoperóxido a prostaglandina F2 alfa (Okano et al., 2002). A PGFS é uma proteína
com estrutura do tipo TIM barrel, e logo, possui um motivo conservado de ligação ao
fosfato. O fosfato vem do substrato ao qual a enzima se liga, ou de co-fatores, como
a riboflavina-5'-fosfato (FMN).
O aumento na sua fosforilação sugere maior atividade da PGFS, o que pode
estar levando ao aumento na síntese de prostaglandinas, que são compostos
lipídicos derivados de ácidos graxos, que se ligam a receptores de membranas,
como os receptores de membrana acoplados a proteína G. O aumento na síntese
das prostaglandinas pode estar associado a regulação de cálcio e ao controle do
crescimento e a proliferação celular.
As aldo-ceto redutases (AKR) são oxidoredutases monoméricas, amplamente
distribuídas em mamíferos, anfíbios, plantas, fungos, bactérias e protozoários
(Mathe et al., 1977; Samuelsson, 1979; Kubata et al., 2000; Ellis, 2002). As AKRs
formam uma superfamília de NAD(P)(H) oxidoredutases solúveis, cujo principal
objetivo é o de reduzir aldeídos e cetonas a álcoois primários e secundários. As
AKRs exibem ampla especificidade de substrato e catalisam a oxidação e/ou
redução de substratos, como as prostaglandinas, hormônios esteróides,
monossacarídeos, isoflavonóides, aldeídos alifáticos e aromáticos e hidrocarbonetos
aromáticos policíclicos (McCracken et al., 1972; Dubois et al., 1998; Hayaishi, 2000;
Okano et al., 2002).
A redução na expressão da AKR indica que a mesma é negativamente
regulada por moléculas envolvidas ou ativadas na sinalização por TGF-β. A
identificação genérica da proteína como uma aldo-ceto redutase diminui nossas
chances de relacionar sua regulação negativa a moléculas ou vias específicas, já
que este grupo é formado por uma série de enzimas com funções diversificadas.
A enzima enolase (2-fosfo-D-glicerato hidrolase), também conhecida como
fosfopiruvato desidratase, é uma metaloenzima responsável por catalisar a
desidratação reversível da D-2-fosfoglicerato (PGA) a fosfoenolpiruvato (PEP), tanto
na glicólise como na gliconeogênese (Verlinde et al., 2001). A enolase é uma

Discussão
101
proteína com dobradamento do tipo TIM “barrel”, possuindo um motivo conservado
de ligação ao fosfato. O grupamento fosfato é proveniente tanto de seu substrato
como de co-fatores, como a riboflavina-5'-fosfato (FMN). A enolase também já foi
descrita como um componente estrutural do degradosoma de RNA de procariotos,
promovendo a degradação de moléculas de RNA nestes microorganismos (Kuhnel &
Luisi, 2001; Carpousis, 2002). Dessa forma, além de atuar como uma enzima
metabólica, a enolase parece também ser induzida por proteínas de choque térmico
(entre elas, a DnaK), como parte da resposta a situações de estresse celular.
A defosforilação da enolase pode estar relacionada à redução na sua
atividade, acarretando na perda de produção de energia e na regulação positiva ou
negativa da degradação de moléculas de RNA.
A enzima glicosomal gliceraldeído-3-fosfato desidrogenase (gGAPDH)
desempenha várias funções importantes no processo glicolítico. A GAPDH catalisa a
oxidação e a fosforilação da D-gliceraldeído-3-fosfato (GAP) a 1,3-bifosfo-D-ácido
glicérico (1,3-BPGA) na presença de NAD+ e de fosfato inorgânico (Tisdale &
Artalejo, 2007). Além da sua função no metabolismo da glicose, a GAPDH exerce
múltiplas atividades intracelulares, incluindo a participação na ativação da
transcrição (Zheng et al., 2003) no transporte vesicular (Tisdale, 2001), na fusão
membrana-membrana, na modulação do citoesqueleto, no reparo e replicação do
DNA, na apoptose (Tarze et al., 2007) e na exportação do tRNA (Tisdale & Artalejo,
2007).
Sua rápida ativação pode estar associada a um maior requerimento de
energia para a viabilização da resposta celular induzida por TGF-β. A GAPDH pode
também estar ativando ou ampliando a transcrição de genes, gerando ou
aumentando a expressão de proteínas que atuarão na via de sinalização de TGF-β.
Além disso, a GAPDH pode estar associada ao transporte de moléculas, permitindo
o surgimento de novas interações celulares.
A malato desidrogenase é uma proteína que se liga a NAD, sendo seus sítios
ativos altamente conservados, compartilhando um mecanismo catalisador comum
(Golding & Dean, 1998). A malato desidrogenase citosólica e a malato
desidrogenase mitocondrial trabalham juntas na maioria dos organismos
eucarióticos, permitindo a re-oxidação mitocondrial do NADH glicolítico produzido no
citosol. Contudo, os tripanossomatídeos são uma exceção, porque a maioria das

Discussão
102
enzimas glicolíticas, incluindo a GAPDH, estão presentes no glicosoma (Cannata &
Cazzulo, 1984). O sistema de trocas envolve principalmente as malato
desidrogenases glicosomal e mitocondrial (Cannata & Cazzulo, 1984; Cazzulo,
1994), e logo, uma malato desidrogenase citosólica se torna inútil para o que pode
ser considerado o papel mais importante desta enzima em outros organismos.
A malato desidrogenase encontrada no nosso estudo é glicosomal, e assim,
desempenha papel importante no metabolismo do T. cruzi. O decréscimo na sua
fosforilação sugere a redução na atividade desta proteína, o que deve estar afetando
a re-oxidação do NADH e, portanto, o transporte de elétrons e a produção de
energia pelo parasito.
As proteínas dissulfeto isomerases (PDIs) são tiol-dissulfeto oxidoredutases
que catalisam a formação, redução e isomerização de ligações dissulfeto,
dependendo do ambiente redox, e também podem funcionar como chaperonas
moleculares, auxiliando no enovelamento de proteínas, embora este mecanismo não
esteja completamente elucidado (Noiva & Lennarz, 1992; Ferrari & Soling, 1999;
Rubotham et al., 2005).
O aumento na sua expressão pode estar relacionado à sua maior ativação, o
que reflete na geração de uma série de reações envolvidas na formação e ruptura
de pontes dissulfeto, as quais desempenham papel importante no enovelamento e
estabilidade de proteínas.
No T. cruzi, a desidrogenase aromática L-alfa-hidróxi-ácida (AHADH) está
envolvida no catabolismo de aminoácidos aromáticos. Esta é uma enzima citosólica
muito ativa ligada a NAD (Nowicki et al., 1992), que reduz os alfa-oxiácidos
aromáticos para L-alfa-hidróxi-ácidos (Cazzulo Franke et al., 1999).
Embora a AHADH seja completamente incapaz de usar oxaloacetato ou
piruvato como substrato, esta enzima apresenta semelhanças na sua sequência de
aminoácidos com as enzimas malato desidrogenase e lactato desidrogenase, onde a
maior similaridade foi observada com a malato desidrogenase citosólica
(Montemartini et al., 1994). As epimastigotas, durante sua vida no intestino do
inseto, são quase que exclusivamente dependentes de aminoácidos e proteínas
como fontes de energia, porque a glicose presente no repasto sanguíneo é
rapidamente esgotada. A expressão diferencial da AHADH nas várias formas do

Discussão
103
parasito, com os níveis mais elevados da enzima nas formas do inseto (epimastigota
e tripomastigotas metacíclicos), está de acordo com a possibilidade de um
catabolismo de aminoácidos mais ativo realizado por essas formas. Isto significa que
elas devem ter suas vias catabólicas conectadas (pelo menos parcialmente) a um
ciclo de Krebs e a uma cadeia respiratória funcionais, permitindo a utilização de
aminoácidos como combustível (Nowicki & Cazzulo, 2008).
O aumento na expressão da AHADH deve estar relacionado ao aumento na
degradação de proteínas para geração de energia e de aminoácidos livres, processo
essencial, por exemplo, para aceleração no seu desenvolvimento e proliferação.
As epimastigotas de T. cruzi têm um conjunto intracelular considerável de
aminoácidos livres, que são utilizados na síntese de proteínas e na produção de
energia (Rohloff et al., 2003). As principais enzimas envolvidas na troca e eliminação
do nitrogênio do grupo amino são as aminotransferases, muitas das quais
transferem o grupo amino a 2- oxoglutarato para gerar L-glutamato. A tirosina
aminotransferase (TAT) de T. cruzi é muito abundante, representando 3% do total de
proteínas solúveis; o genoma de T. cruzi possui aproximadamente 70 cópias do
gene tat (Bontempi et al., 1993). Enquanto a TAT dos mamíferos prefere tirosina e
alfa-cetoglutarato como substratos (Iwasaki et al., 1973), a enzima de T. cruzi aceita
três alfa-cetoácidos (piruvato, a-cetoglutarato e oxaloacetato), todos os três
aminoácidos aromáticos e alanina como substratos (Montemartini et al., 1993). Esta
enzima tem uma ampla especificidade de substrato, utilizando de forma eficiente a
leucina, metionina, tirosina, fenilalanina, triptofano e alanina como doadores de
aminoácidos (Montemartini et al., 1993; Nowicki et al., 2001; Blankenfeldt et al.,
1999).
A redução na sua expressão sugere que a TAT esteja sendo negativamente
regulada por outra molécula direta ou indiretamente ativada por TGF-β em algum
dos tempos anteriores de estudo e indica uma diminuição na síntese de algumas
proteínas e menor produção de energia pela via da TAT.
Entre as proteínas envolvidas no processo de tradução, temos o complexo
associado ao polipeptídeo nascente (NAC), que se associa com ribossomos e
cadeias nascentes em uma estequiometria aparente de 1:1 (Wiedmann et al., 1994;
Funfschilling & Rospert, 1999; Rospert et al., 2002; Olesya, 2009). A NAC é um
complexo heterodimérico altamente conservado, presente em arqueobactérias,

Discussão
104
leveduras e células de mamíferos. O conhecimento sobre sua função in vivo ainda
apresenta várias lacunas a serem preenchidas. Alguns estudos especulam que a
NAC desempenha um papel no enovelamento de proteínas recém-sintetizadas,
protegendo-as da interação com fatores citosólicos inadequados. Estudos mostram
que a NAC pode contribuir para a fidelidade nos processos co-traducionais tais
como marcação de alvos e enovelamento (Wang et al., 1995). Há também
evidências de que a NAC interaja diretamente com a sequência de reconhecimento
de sinal e está envolvida na translocação correta de proteínas para o retículo
endoplasmático, através da regulação da acessibilidade do poro de translocação e
evitando a marcação errada de proteínas não secretórias (Lauring et al., 1995;
Moller et al., 1998). Além disso, um papel regulador da NAC na importação de
proteínas para a mitocôndria foi proposto (George et al., 1998; Funfschilling &
Rospert, 1999), no entanto, ainda não há evidência direta para apoiar esta hipótese.
A ativação da NAC em reposta a TGF-� pode estar relacionada à síntese de
proteínas corretamente enoveladas e ao endereçamento dessas moléculas ao
retículo endoplasmático e ao Golgi, onde provavelmente sofrerão modificações pós-
traducionais que permitirão o encaminhamento ao local onde exercerão suas
atividades celulares.
O fator eucariótico de iniciação da tradução 5A (EIF-5a) é uma proteína de 18
kDa, altamente conservada desde leveduras a células de mamíferos (Schnier et al.,
1991; Park et al., 1997; Caraglia et al., 2000). A EIF-5a promove a formação da
primeira ligação peptídica durante a fase inicial da síntese de proteínas (Hershey,
1991). Entretanto, a verdadeira função “in vivo” da EIF-5a, é apenas parcialmente
conhecida. Uma série de observações sugere que a EIF-5a desempenha um papel
essencial no crescimento e diferenciação celular. A EIF-5a pode ser uma proteína
bimodular, interagindo tanto com RNA quanto com proteínas, e presume-se assim,
que ela possua um papel importante na maquinaria de tradução (Park, 2006). Outras
funções foram propostas para a EIF-5a, incluindo a de co-fator celular da proteína
REV de HIV-1 (Ruhl et al., 1993), de um fator envolvido na exportação nuclear
(Lipowsky et al., 2000) e no “turnover” de mRNA (Zuk & Jacobson, 1998; Valentini et
al., 2002). A EIF-5a é encontrada no citoplasma em duas situações: uma é livre e a
outra é vinculada ao retículo endoplasmático (provavelmente o seu local de ação
apropriado) (Shi et al., 1996). No entanto, foi também recentemente relatado que a
EIF-5a pode se acumular nos filamentos intranucleares associados ao poro nuclear

Discussão
105
em células de mamíferos (Rosorius et al., 1999). Além disso, o fator interage com o
receptor nuclear geral de exportação CRM1 e é transportado do núcleo para o
citoplasma (Rosorius et al., 1999). Estes dados indicam que a EIF-5A pode também
funcionar como uma proteína de translocação núcleo-citoplasma de mRNAs
eventualmente correlacionados com a proliferação celular.
O aumento na fosforilação da EIF-5a indica uma resposta tardia desta
proteína à sinalização disparada por TGF-β. Sua ativação deve estar relacionada à
regulação da tradução de proteínas, e logo, a produção de novas proteínas, que
podem ser os efetores da resposta celular a TGF-β.
O fator de elongamento-1α (EF-1α) desempenha papel essencial na
biossíntese de proteínas eucarióticas (Billaut-Mulot et al., 1996). A EF-1α se liga a
aminoacil-tRNAs no sítio aceptor da subunidade ribossomal 80S através da hidrólise
de GTP (Moazed & Noller, 1989). A EF-1α possui uma série de outras funções além
da biossíntese de proteínas. Esta proteína foi encontrada em associação com a valil-
tRNA sintetase de mamíferos, complexadas a partículas mRNP, ligada a actina do
citoesqueleto da célula, ligada ao retículo endoplasmático ou ao aparato mitótico,
estando envolvida na degradação de proteínas ou em associação ao ribossomo
(Merrick, 1992). A EF-lα também tem sido relacionada a uma variedade de
processos, como a embriogênese, a divisão celular, a proliferação celular, o
envelhecimento e a transformação oncogênica (Janssen et al., 1994). Além disso,
uma quantidade considerável de EF-1α foi localizada no núcleo de diversos tipos de
células (Collings et al., 1994; Barbarese et al., 1995). Existe ainda a possibilidade da
EF-lα desempenhar um papel na regulação da transcrição (Blumenthal et al., 1972).
Grandes aumentos nos níveis de mRNA de EFlα são observadas em células
de cultura de rápida proliferação (Sanders et al., 1992), nos embriões (Krieg et al.,
1989) e em células tumorais humanas de proliferação rápida (Grant et al., 1992).
Estes dados indicam que o nível de expressão de EFlα se correlaciona com a taxa
de crescimento e proliferação celular (Condeelis, 1995). Ensaios de
imunofluorescência utilizando anticorpos para a EF-1α de T. cruzi (TCEF-1α),
visando estudar a distribuição celular da (TCEF-1α) durante o crescimento in vitro de
epimastigotas, revelaram que esta proteína é acumulada no núcleo de parasitos em
processo de envelhecimento mostrando níveis elevados de células binucleadas,
indicativo de parada na citocinese, e aqueles em processo de morte por apoptose,

Discussão
106
sugerindo, portanto, que a TCEF-1α é capaz de fazer a translocação entre
citoplasma e núcleo celular (Billaut-Mulot et al., 1996; Ouaissi, 2003).
No nosso estudo, a EF-1α tem sua expressão aumentada em resposta ao
TGF-β, podendo, portanto, estar participando da regulação da síntese de proteínas,
influenciando positivamente na proliferação das epimastigotas.
Entre as proteases/peptidases, temos a cruzipaína (CZP), uma cisteína
protease do tipo papaína, que parece ser maturada no complexo de Golgi (Engel et
al., 1998) e é altamente acumulada (Soares et al., 1992) e ativa (Cunha-e-Silva et
al., 2002) nos reservossomas, que contêm proteínas usadas na diferenciação para
tripomastigotas metacíclicos (Soares et al., 1992). A via que endereça a cruzipaína
aos reservossomas não é totalmente compreendida, porém trabalhos anteriores
sugerem que esta enzima seja transportada do complexo de Golgi através de
vesículas secretórias (Soares et al., 1992) em uma via independente de manose 6-
fosfato (Cazzulo et al., 1990). A CZP é expressa em diferentes níveis nas diferentes
formas evolutivas do parasito (Cazzulo et al., 1990; Martinez et al., 1991). Esta
enzima é a principal protease encontrada na forma epimastigota, sendo ativa pelo
menos dez vezes mais nesta forma do que nas amastigotas e tripomastigotas
(Campetella et al., 1990; Sant'Anna et al., 2008).
A endocitose e a reciclagem de proteínas são vias metabólicas essenciais
para o desenvolvimento do ciclo de vida do T. cruzi (De Souza, 2002). A via
endocítica está sob controle rigoroso de desenvolvimento nas diferentes fases do
ciclo de vida do parasito. As formas epimastigotas são altamente ativas e
armazenam macromoléculas obtidas do meio em estruturas denominadas
reservossomas (Soares & de Souza, 1991). Esta estrutura não foi observada em
formas tripomastigotas ou amastigotas (Soares & de Souza, 1991). Durante o
processo de diferenciação de formas epimastigotas em tripomastigotas metacíclicos,
a conversão da forma proliferativa para a forma infectante é realizada com consumo
de energia e é dependente da disponibilidade de aminoácidos, sendo necessário o
uso dos substratos resultantes da degradação de moléculas pela via endocítica
(Soares et al., 1989). Utilizando uma abordagem imunocitoquímica, a cruzipaína
também foi localizada na membrana plasmática e na bolsa flagelar do parasito
(Souto-Padron et al., 1990). Portanto, é possível que pelo menos uma fração da
CZP seja primeiramente endereçada para a superfície do parasito, e em seguida,

Discussão
107
internalizada como parte da via endocítica e encaminhada aos reservossomas. Já foi
descrita na literatura uma relação entre TGF-β e cruzipaína. A adição de cruzipaína
à macrófagos de murinos induziu um aumento na secreção de TGF-β (Stempin et
al., 2002). A cruzipaína foi sugerida ainda como a provável molécula ativadora de
TGF-β em Trypanosoma cruzi (Waghabi et al., 2005b).
A Cruzipaína também está envolvida na virulência do T. cruzi. Embora esteja
menos presente em amastigotas e tripomastigotas, a cruzipaína é um antígeno
imunodominante reconhecido nos soros de pacientes humanos com a doença de
Chagas crônica (Martinez et al., 1991; Cazzulo, 2002).
O aumento na expressão da cruzipaína pode estar associado ao aumento na
atividade desta proteína, que em resposta a TGF-β deve regular a degradação e
reciclagem de proteínas nos reservosomas de T. cruzi.
O complexo HslVU consiste em dois componentes: o componente HslU, que
atua como chaperona e ATPase (Rohrwild et al., 1996; Neuwald et al., 1999; Seong
et al., 2000), e o componente HslV, a treonina protease amino-terminal (Chuang et
al., 2005), que confere a atividade peptidase (Azim et al., 2005). A protease HslVU
está envolvida na proteólise de proteínas de vida curta, como a proteína inibidora da
divisão celular SulA (Cordell et al., 2003), o fator de choque térmico 32 (Kanemori et
al., 1997), e o ativador de transcrição RcsA (Torres-Cabassa & Gottesman, 1987). A
protease HslVU é considerada como um análogo de bactérias do proteassoma 26S
eucariótico (Rohrwild et al., 1996; Gille et al., 2003). Essas proteases desempenham
um papel essencial no controle dos níveis das principais proteínas reguladoras e na
eliminação de polipeptídeos anormais (Park et al., 2005).
O aumento na expressão da subunidade proteolítica do complexo Hsvlu deve
se relacionar ao aumento de sua atividade, e assim, esta molécula pode estar
desempenhando função semelhante a da cruzipaína, regulando a degradação de
proteínas do parasito.
A triparedoxina peroxidase (TriX) 1 é uma tiol-dissulfeto oxidorredutase
encontrada em parasitos tripanosomatídeos pertencentes à ordem Kinetoplastida. A
principal função biológica da TriX é a de regular o estresse oxidativo como um
componente da via da tripanotiona peroxidase (Alphey et al., 2003). Em Leishmania
e outros tripanosomatídeos, as enzimas da família triparedoxina peroxidase são

Discussão
108
consideradas as principais responsáveis pela detoxicação de peróxidos, protegendo
as células de danos causados pelo estresse oxidativo (Flohe et al., 2003). Estas
enzimas obtêm seus equivalentes redutores da tripanotiona, através da proteína
triparedoxina ditiol (Wyllie et al., 2008). Várias enzimas antioxidantes, incluindo a
triparedoxina peroxidase mitocondrial (TcMPX) são reguladas positivamente durante
a transformação da forma não infectiva do inseto (epimastigota) para a forma
infectiva tripomastigota metacíclica (Atwood et al., 2005; Piacenza et al., 2008).
O aumento tardio na expressão da triparredoxina peroxidase indica a
necessidade prévia da ativação de outras moléculas pela via de TGF-β. TGF-β está
envolvido na proliferação das epimastigotas e este evento é fruto de uma série de
interações moleculares. O aumento na proliferação das epimastigotas pode estar
associado aos primeiros sinais para a diferenciação das epimastigotas em
tripomatigotas que, como descrito na literatura, regula positivamente a triparredoxina
peroxidase. A ativação da triparredoxina peroxidase deve ocorrer em etapas
anteriores à diferenciação, ainda na fase de proliferação das epimastigotas.
O fator de liberação de histamina dependente de IgE está envolvido
primordialmente na resposta imunológica produzida por células do sistema de
defesa em eucariotos superiores, sendo também conhecida como proteína tumoral
traducionalmente controlada (TCTP). A TCTP é expressa em tecidos mitoticamente
ativos (Thiele et al., 2000). Em inúmeras situações experimentais e sistemas
biológicos, foi estabelecido que os níveis de TCTP são altamente regulados em
resposta a uma ampla gama de sinais extracelulares e às condições celulares.
Vários relatos destacaram a importância da TCTP para a progressão do ciclo celular
e a transformação maligna (Bommer & Thiele, 2004). Normalmente, os sinais de
crescimento (Bommer et al., 2002) e citocinas (Nielsen et al., 1998; Teshima et al.,
1998) foram relatados por rapidamente induzir a síntese desta proteína. Além disso,
a TCTP exerce uma função extracelular como fator de liberação de histamina e tem
atividade anti-apoptótica (MacDonald et al., 1995). A TCTP já foi caracterizada em
vários organismos parasitos, tais como subespécies de Plasmodium (Bhisutthibhan
et al., 1999; Walker et al., 2000) e vários vermes parasitos (Gnanasekar et al., 2002;
Rao et al., 2002). Em todos estes casos, a proteína do parasito é secretada no
organismo hospedeiro e é capaz de causar infiltrados inflamatórios de eosinófilos
(Gnanasekar et al., 2002; Rao et al., 2002) e / ou liberação de histamina de basófilos
(Rao et al., 2002; MacDonald et al., 2001).

Discussão
109
Várias condições de estresse, como privação de nutrientes (Bommer et al.,
2002; Bonnet et al., 2000), choque térmico, metais pesados, estresse de cálcio (Xu
et al., 1999) ou sinais pró-apoptóticos ou citotóxicos (Sinha et al., 2000) resultam
tanto na regulação positiva como negativa dos níveis de TCTP (Bommer et al., 2002;
Gachet et al., 1999). A adaptação rápida dos níveis da proteína TCTP a alterações
nas condições celulares nos mostra que tanto sua síntese como a degradação são
altamente reguladas, existindo várias evidências mostrando que a síntese de TCTP
é regulada, tanto no nível transcricional como traducional (Bommer et al., 2002).
TCTP possui uma região de ligação a tubulina e foi mostrado que em células
de mamíferos, parte desta proteína se liga aos microtúbulos durante a maior parte
do ciclo celular (Gachet et al., 1999). Os níveis de TCTP são regulados
positivamente durante a entrada das células no ciclo celular, e logo, acredita-se que
esta proteína seja importante para o crescimento e divisão celular. A maioria das
publicações descreve a TCTP como uma proteína citoplasmática, mas sua
localização nuclear também já foi relatada (Li et al., 2001).
A ativação desta proteína em formas epimastigotas de T.cruzi em resposta ao
TGF-β pode estar associada a uma série de processos celulares. Assim como as
proteínas de choque térmico, a TCTP pode estar sendo duplamente ativada: como
resposta à baixa na temperatura do ambiente e em resposta a adição de TGF-β ao
meio. Com relação à última, a TCTP pode estar envolvida na proliferação das
epimastigotas. Uma hipótese seria a da ligação desta proteína às tubulinas, de modo
a provocar re-estruturações no parasito e permitir que novas interações celulares
ocorram.
A fosforilação de proteínas está fortemente associada aos processos de
sinalização celular e de desenvolvimento nos eucariotos. Apesar do amplo espectro
de informação gerado pela fosfoproteômica, é extremamente desafiador conectar as
quinases em estudo com seus devidos alvos moleculares. O desafio de associar as
quinases a seus substratos reflete a complexidade das funções desempenhadas por
essas proteínas. As quinases tendem a exercer seus efeitos biológicos através da
fosforilação simultânea de múltiplos resíduos, muitas vezes em componentes de
complexos protéicos múltiplos. Embora estes eventos de fosforilação sejam
biologicamente significantes, a identificação das quinases responsáveis por eles se
torna extremamente difícil. Por isso, apesar de termos identificado uma série de

Discussão
110
proteínas responsivas a TGF-β, constitui tarefa difícil relacionar o papel de cada uma
delas na sinalização disparada por TGF-β.
Por último, fizemos um ensaio de proliferação das epimastigotas na presença
ou não de TGF-β nas culturas. A adição da citocina no meio promoveu um aumento
de até 73% no crescimento dos parasitos nas primeiras 24 horas de estudo. Com 48
e 72 horas ainda é observado um aumento na proliferação devido à adição de TGF-
β, porém este já não é tão acentuado. Esse dado acrescenta importância ao papel
de TGF-β sobre a forma epimastigota de T. cruzi e reforça a relevância do nosso
estudo, mostrando que esta forma evolutiva também é influenciada positivamente
pela citocina em questão.
Considerações Finais
Este estudo integra um amplo conjunto de trabalhos elaborados em nosso
laboratório, que visam compreender a relação de TGF-β com o T. cruzi. Aprofundar
nosso conhecimento acerca de quais papéis TGF-β desempenha e de como os
mesmos são executados no parasito é uma oportunidade para entender melhor
como T. cruzi regula as diferentes etapas de seu ciclo de vida, e consequentemente,
como ele consegue se desenvolver nos seus hospedeiros vertebrados e
invertebrados. A compreensão dos mecanismos moleculares relacionados à
sinalização por TGF-β em T. cruzi pode ser de grande relevância para a busca de
novos alvos que visem o tratamento ou até mesmo a cura da doença de Chagas.
Nosso estudo mostrou que algumas proteínas de epimastigotas de T. cruzi
respondem aos estímulos de TGF-β, mas o papel exato que elas desempenham na
sua via de sinalização continua sendo objeto de investigação em nosso laboratório.
De uma forma geral, podemos dizer que as epimastigotas respondem ao estímulo
de TGF-β, que por sua vez é capaz de interagir direta e indiretamente com proteínas
do parasito, alterando seus padrões de fosforilação e modulando seus níveis de
expressão. TGF-β deve, então, ativar ou desativar via(s) específica(s) do parasito,
podendo causar um aumento na sua proliferação, e possivelmente, regulando outras
funções celulares neste microorganismo.

Conclusões
111
7. CONCLUSÕES
Após a realização de nosso estudo, podemos concluir que:
� A dose de 5 ng/ml de TGF-β induziu o maior número de eventos de
fosforilação nas proteínas das epimastigotas de T. cruzi;
� A metodologia fosfoproteômica se mostrou eficaz na caracterização das
proteínas de epimastigotas de T. cruzi diferencialmente fosforiladas e/ou
expressas em resposta a TGF-β;
� No estudo de cinética de tempo, os perfis protéicos bidimensionais das
epimastigotas tratadas com 5 ng/ml de TGF-β por 1, 5, 15, 30 ou 60 minutos
apresentaram diferenças nos padrões de fosforilação e/ou expressão de
algumas proteínas;
� As proteínas responsivas à TGF-β estão relacionadas a diversas funções
celulares, entre elas, enovelamento protéico, resposta a choque térmico,
formação de citoesqueleto, síntese de proteínas, transdução de sinal, sendo
as mais abundantes envolvidas no metabolismo celular;
� TGF-β modulou positivamente o processo de proliferação das epimastigotas
de T. cruzi.

Referências
112
8. REFERÊNCIAS
Agrawal, GK, Thelen, J. A high-resolution two dimensional gel- and Pro-Q DPS-
based proteomics workflow for phosphoprotein identification and quantitative
profiling. In: Graauw, Md, editor. Phospho-Proteomics, Methods and Protocols.
New York: Humana Press; 2009. p. 3-19.
Almeida-de-Faria, M, Freymuller, E, Colli, W, Alves, MJ. Trypanosoma cruzi:
characterization of an intracellular epimastigote-like form. Exp Parasitol. 1999
Aug;92(4):263-74.
Alphey, MS, Gabrielsen, M, Micossi, E, Leonard, GA, McSweeney, SM, Ravelli, RB,
et al. Tryparedoxins from Crithidia fasciculata and Trypanosoma brucei:
photoreduction of the redox disulfide using synchrotron radiation and evidence
for a conformational switch implicated in function. J Biol Chem. 2003 Jul
11;278(28):25919-25.
Anders, RA, Arline, SL, Dore, JJ, Leof, EB. Distinct endocytic responses of
heteromeric and homomeric transforming growth factor beta receptors. Mol Biol
Cell. 1997 Nov;8(11):2133-43.
Andrade, SG, Grimaud, JA, Stocker-Guerret, S. Sequential changes of the
connective matrix components of the myocardium (fibronectin and laminin) and
evolution of cardiac fibrosis in mice infected with Trypanosoma cruzi. Am J Trop
Med Hyg. 1989 Mar;40(3):252-60.
Andrews, NW. The acid-active hemolysin of Trypanosoma cruzi. Exp Parasitol. 1990
Aug;71(2):241-4.
Araujo-Jorge, TC, Waghabi, MC, Hasslocher-Moreno, AM, Xavier, SS, Higuchi Mde,
L, Keramidas, M, et al. Implication of transforming growth factor-beta1 in
Chagas disease myocardiopathy. J Infect Dis. 2002 Dec 15;186(12):1823-8.
Araujo-Jorge, TC, Waghabi, MC, Soeiro Mde, N, Keramidas, M, Bailly, S, Feige, JJ.
Pivotal role for TGF-beta in infectious heart disease: The case of Trypanosoma
cruzi infection and consequent Chagasic myocardiopathy. Cytokine Growth
Factor Rev. 2008 Oct-Dec;19(5-6):405-13.

Referências
113
Arevalo-Rodriguez, M, Wu, X, Hanes, SD, Heitman, J. Prolyl isomerases in yeast.
Front Biosci. 2004 Sep 1;9:2420-46.
Atwood, JA, 3rd, Weatherly, DB, Minning, TA, Bundy, B, Cavola, C, Opperdoes, FR,
et al. The Trypanosoma cruzi proteome. Science. 2005 Jul 15;309(5733):473-6.
Ayub, MJ, Atwood, J, Nuccio, A, Tarleton, R, Levin, MJ. Proteomic analysis of the
Trypanosoma cruzi ribosomal proteins. Biochem Biophys Res Commun. 2009
Apr 24;382(1):30-4.
Azim, MK, Goehring, W, Song, HK, Ramachandran, R, Bochtler, M, Goettig, P.
Characterization of the HslU chaperone affinity for HslV protease. Protein Sci.
2005 May;14(5):1357-62.
Bakin, AV, Rinehart, C, Tomlinson, AK, Arteaga, CL. p38 mitogen-activated protein
kinase is required for TGFbeta-mediated fibroblastic transdifferentiation and cell
migration. J Cell Sci. 2002 Aug 1;115(Pt 15):3193-206.
Banh, A, Deschamps, PA, Vijayan, MM, Sivak, JG, West-Mays, JA. The role of
Hsp70 and Hsp90 in TGF-beta-induced epithelial-to-mesenchymal transition in
rat lens epithelial explants. Mol Vis. 2007;13:2248-62.
Barbarese, E, Koppel, DE, Deutscher, MP, Smith, CL, Ainger, K, Morgan, F, et al.
Protein translation components are colocalized in granules in oligodendrocytes.
J Cell Sci. 1995 Aug;108 ( Pt 8):2781-90.
Barolo, S, Stone, T, Bang, AG, Posakony, JW. Default repression and Notch
signaling: Hairless acts as an adaptor to recruit the corepressors Groucho and
dCtBP to Suppressor of Hairless. Genes Dev. 2002 Aug 1;16(15):1964-76.
Bartholomeu, DC, Silva, RA, Galvao, LM, el-Sayed, NM, Donelson, JE, Teixeira, SM.
Trypanosoma cruzi: RNA structure and post-transcriptional control of tubulin
gene expression. Exp Parasitol. 2002 Nov-Dec;102(3-4):123-33.
Bente, M, Harder, S, Wiesgigl, M, Heukeshoven, J, Gelhaus, C, Krause, E, et al.
Developmentally induced changes of the proteome in the protozoan parasite
Leishmania donovani. Proteomics. 2003 Sep;3(9):1811-29.

Referências
114
Bhisutthibhan, J, Philbert, MA, Fujioka, H, Aikawa, M, Meshnick, SR. The
Plasmodium falciparum translationally controlled tumor protein: subcellular
localization and calcium binding. Eur J Cell Biol. 1999 Sep;78(9):665-70.
Bhowmick, NA, Ghiassi, M, Bakin, A, Aakre, M, Lundquist, CA, Engel, ME, et al.
Transforming growth factor-beta1 mediates epithelial to mesenchymal
transdifferentiation through a RhoA-dependent mechanism. Mol Biol Cell. 2001
Jan;12(1):27-36.
Billaut-Mulot, O, Fernandez-Gomez, R, Loyens, M, Ouaissi, A. Trypanosoma cruzi
elongation factor 1-alpha: nuclear localization in parasites undergoing
apoptosis. Gene. 1996 Sep 26;174(1):19-26.
Blankenfeldt, W, Nowicki, C, Montemartini-Kalisz, M, Kalisz, HM, Hecht, HJ. Crystal
structure of Trypanosoma cruzi tyrosine aminotransferase: substrate specificity
is influenced by cofactor binding mode. Protein Sci. 1999 Nov;8(11):2406-17.
Blehert, DS, Fox, BG, Chambliss, GH. Cloning and sequence analysis of two
Pseudomonas flavoprotein xenobiotic reductases. J Bacteriol. 1999
Oct;181(20):6254-63.
Blumenthal, T, Landers, TA, Weber, K. Bacteriophage Q replicase contains the
protein biosynthesis elongation factors EF Tu and EF Ts. Proc Natl Acad Sci U
S A. 1972 May;69(5):1313-7.
Bommer, UA, Borovjagin, AV, Greagg, MA, Jeffrey, IW, Russell, P, Laing, KG, et al.
The mRNA of the translationally controlled tumor protein P23/TCTP is a highly
structured RNA, which activates the dsRNA-dependent protein kinase PKR.
RNA. 2002 Apr;8(4):478-96.
Bommer, UA & Thiele, BJ. The translationally controlled tumour protein (TCTP). Int J
Biochem Cell Biol. 2004 Mar;36(3):379-85.
Bonnet, C, Perret, E, Dumont, X, Picard, A, Caput, D, Lenaers, G. Identification and
transcription control of fission yeast genes repressed by an ammonium
starvation growth arrest. Yeast. 2000 Jan 15;16(1):23-33.
Bontempi, EJ, Bua, J, Aslund, L, Porcel, B, Segura, EL, Henriksson, J, et al. Isolation
and characterization of a gene from Trypanosoma cruzi encoding a 46-

Referências
115
kilodalton protein with homology to human and rat tyrosine aminotransferase.
Mol Biochem Parasitol. 1993 Jun;59(2):253-62.
Brenchley, R, Tariq, H, McElhinney, H, Szoor, B, Huxley-Jones, J, Stevens, R, et al.
The TriTryp phosphatome: analysis of the protein phosphatase catalytic
domains. BMC Genomics. 2007;8:434.
Bukau, B & Horwich, AL. The Hsp70 and Hsp60 chaperone machines. Cell. 1998
Feb 6;92(3):351-66.
Burt, DW & Law, AS. Evolution of the transforming growth factor-beta superfamily.
Prog Growth Factor Res. 1994;5(1):99-118.
Campetella, O, Martinez, J, Cazzulo, JJ. A major cysteine proteinase is
developmentally regulated in Trypanosoma cruzi. FEMS Microbiol Lett. 1990
Jan 15;55(1-2):145-9.
Cannata, JJ & Cazzulo, JJ. Glycosomal and mitochondrial malate dehydrogenases in
epimastigotes of Trypanosoma cruzi. Mol Biochem Parasitol. 1984 Apr;11:37-
49.
Caraglia, M, Budillon, A, Vitale, G, Lupoli, G, Tagliaferri, P, Abbruzzese, A.
Modulation of molecular mechanisms involved in protein synthesis machinery
as a new tool for the control of cell proliferation. Eur J Biochem. 2000
Jul;267(13):3919-36.
Carpousis, AJ. The Escherichia coli RNA degradosome: structure, function and
relationship in other ribonucleolytic multienzyme complexes. Biochem Soc
Trans. 2002 Apr;30(2):150-5.
Carr, SA, Hemling, ME, Folena-Wasserman, G, Sweet, RW, Anumula, K, Barr, JR, et
al. Protein and carbohydrate structural analysis of a recombinant soluble CD4
receptor by mass spectrometry. J Biol Chem. 1989 Dec 15;264(35):21286-95.
Cazzulo Franke, MC, Vernal, J, Cazzulo, JJ, Nowicki, C. The NAD-linked aromatic
alpha-hydroxy acid dehydrogenase from Trypanosoma cruzi. A new member of
the cytosolic malate dehydrogenases group without malate dehydrogenase
activity. Eur J Biochem. 1999 Dec;266(3):903-10.

Referências
116
Cazzulo, JJ. Intermediate metabolism in Trypanosoma cruzi. J Bioenerg Biomembr.
1994 Apr;26(2):157-65.
Cazzulo, JJ. Proteinases of Trypanosoma cruzi: patential targets for the
chemotherapy of Changas desease. Curr Top Med Chem. 2002
Nov;2(11):1261-71.
Cazzulo, JJ, Cazzulo Franke, MC, Martinez, J, Franke de Cazzulo, BM. Some kinetic
properties of a cysteine proteinase (cruzipain) from Trypanosoma cruzi. Biochim
Biophys Acta. 1990 Feb 9;1037(2):186-91.
Chang, E & Goldberg, H. Requirements for transforming growth factor-beta
regulation of the pro-alpha 2(I) collagen and plasminogen activator inhibitor-1
promoters. J Biol Chem. 1995 Mar 3;270(9):4473-7.
Chapin, SJ & Bulinski, JC. Preparation and functional assay of pure populations of
tyrosinated and detyrosinated tubulin. Methods Enzymol. 1991;196:254-64.
Chen, W, Jin, W, Tian, H, Sicurello, P, Frank, M, Orenstein, JM, et al. Requirement
for transforming growth factor beta1 in controlling T cell apoptosis. J Exp Med.
2001 Aug 20;194(4):439-53.
Chen, Y, Takeshita, A, Ozaki, K, Kitano, S, Hanazawa, S. Transcriptional regulation
by transforming growth factor beta of the expression of retinoic acid and retinoid
X receptor genes in osteoblastic cells is mediated through AP-1. J Biol Chem.
1996 Dec 6;271(49):31602-6.
Chuang, DM, Hough, C, Senatorov, VV. Glyceraldehyde-3-phosphate
dehydrogenase, apoptosis, and neurodegenerative diseases. Annu Rev
Pharmacol Toxicol. 2005;45:269-90.
Clayton, C & Shapira, M. Post-transcriptional regulation of gene expression in
trypanosomes and leishmanias. Mol Biochem Parasitol. 2007 Dec;156(2):93-
101.
Cohen, P. Signal integration at the level of protein kinases, protein phosphatases and
their substrates. Trends Biochem Sci. 1992 Oct;17(10):408-13.
Cohen, P. The regulation of protein function by multisite phosphorylation--a 25 year
update. Trends Biochem Sci. 2000 Dec;25(12):596-601.

Referências
117
Collings, DA, Wasteneys, GO, Miyazaki, M, Williamson, RE. Elongation factor 1
alpha is a component of the subcortical actin bundles of characean algae. Cell
Biol Int. 1994 Nov;18(11):1019-24.
Condeelis, J. Elongation factor 1 alpha, translation and the cytoskeleton. Trends
Biochem Sci. 1995 May;20(5):169-70.
Cordell, SC, Robinson, EJ, Lowe, J. Crystal structure of the SOS cell division inhibitor
SulA and in complex with FtsZ. Proc Natl Acad Sci U S A. 2003 Jun
24;100(13):7889-94.
Coura, JR. Chagas disease: what is known and what is needed--a background
article. Mem Inst Oswaldo Cruz. 2007 Oct 30;102 Suppl 1:113-22.
Coura, JR & Dias, JC. Epidemiology, control and surveillance of Chagas disease:
100 years after its discovery. Mem Inst Oswaldo Cruz. 2009 Jul;104 Suppl 1:31-
40.
Cunha-e-Silva, NL, Atella, GC, Porto-Carreiro, IA, Morgado-Diaz, JA, Pereira, MG,
De Souza, W. Isolation and characterization of a reservosome fraction from
Trypanosoma cruzi. FEMS Microbiol Lett. 2002 Aug 27;214(1):7-12.
de Caestecker, MP, Parks, WT, Frank, CJ, Castagnino, P, Bottaro, DP, Roberts, AB,
et al. Smad2 transduces common signals from receptor serine-threonine and
tyrosine kinases. Genes Dev. 1998 Jun 1;12(11):1587-92.
de Carvalho, TU & de Souza, W. Infectivity of amastigotes of Trypanosoma cruzi.
Rev Inst Med Trop Sao Paulo. 1986 Jul-Aug;28(4):205-12.
De Souza, W. Basic cell biology of Trypanosoma cruzi. Curr Pharm Des.
2002;8(4):269-85.
Degrave, WM, Melville, S, Ivens, A, Aslett, M. Parasite genome initiatives. Int J
Parasitol. 2001 May 1;31(5-6):532-6.
Derynck, R, Zhang, Y, Feng, XH. Smads: transcriptional activators of TGF-beta
responses. Cell. 1998 Dec 11;95(6):737-40.
Derynck, R & Zhang, YE. Smad-dependent and Smad-independent pathways in
TGF-beta family signalling. Nature. 2003 Oct 9;425(6958):577-84.

Referências
118
Doerig, C. Protein kinases as targets for anti-parasitic chemotherapy. Biochim
Biophys Acta. 2004 Mar 11;1697(1-2):155-68.
Dong, C, Li, Z, Alvarez, R, Jr., Feng, XH, Goldschmidt-Clermont, PJ. Microtubule
binding to Smads may regulate TGF beta activity. Mol Cell. 2000 Jan;5(1):27-
34.
Dubois, RN, Abramson, SB, Crofford, L, Gupta, RA, Simon, LS, Van De Putte, LB, et
al. Cyclooxygenase in biology and disease. FASEB J. 1998 Sep;12(12):1063-
73.
Dumonteil, E. DNA Vaccines against Protozoan Parasites: Advances and
Challenges. J Biomed Biotechnol. 2007;2007(6):90520.
Dutcher, SK. The tubulin fraternity: alpha to eta. Curr Opin Cell Biol. 2001 Feb:
13(1):49-54.
Edlund, S, Landstrom, M, Heldin, CH, Aspenstrom, P. Transforming growth factor-
beta-induced mobilization of actin cytoskeleton requires signaling by small
GTPases Cdc42 and RhoA. Mol Biol Cell. 2002 Mar;13(3):902-14.
El-Sayed, NM, Myler, PJ, Bartholomeu, DC, Nilsson, D, Aggarwal, G, Tran, AN, et al.
The genome sequence of Trypanosoma cruzi, etiologic agent of Chagas
disease. Science. 2005 Jul 15;309(5733):409-15.
Ellis, EM. Microbial aldo-keto reductases. FEMS Microbiol Lett. 2002 Nov
5;216(2):123-31.
Engel, ME, Datta, PK, Moses, HL. Signal transduction by transforming growth factor-
beta: a cooperative paradigm with extensive negative regulation. J Cell
Biochem Suppl. 1998;30-31:111-22.
Engel, ME, McDonnell, MA, Law, BK, Moses, HL. Interdependent SMAD and JNK
signaling in transforming growth factor-beta-mediated transcription. J Biol
Chem. 1999 Dec 24;274(52):37413-20.
Fapesp. Transmissão silenciosa. 2008. Boletim de Comunicação, 2008 atualizado
em 21/02/2008. Disponível no endereço eletrônico:
http://www.agencia.fapesp.br/materia/8445/entrevistas.

Referências
119
Ferella, M, Nilsson, D, Darban, H, Rodrigues, C, Bontempi, EJ, Docampo, R, et al.
Proteomics in Trypanosoma cruzi--localization of novel proteins to various
organelles. Proteomics. 2008 Jul;8(13):2735-49.
Ferrari, DM & Soling, HD. The protein disulphide-isomerase family: unravelling a
string of folds. Biochem J. 1999 Apr 1;339 ( Pt 1):1-10.
Flohe, L, Budde, H, Hofmann, B. Peroxiredoxins in antioxidant defense and redox
regulation. Biofactors. 2003;19(1-2):3-10.
Funaba, M, Zimmerman, CM, Mathews, LS. Modulation of Smad2-mediated signaling
by extracellular signal-regulated kinase. J Biol Chem. 2002 Nov
1;277(44):41361-8.
Funfschilling, U & Rospert, S. Nascent polypeptide-associated complex stimulates
protein import into yeast mitochondria. Mol Biol Cell. 1999 Oct;10(10):3289-99.
Gachet, Y, Tournier, S, Lee, M, Lazaris-Karatzas, A, Poulton, T, Bommer, UA. The
growth-related, translationally controlled protein P23 has properties of a tubulin
binding protein and associates transiently with microtubules during the cell
cycle. J Cell Sci. 1999 Apr;112 ( Pt 8):1257-71.
Garcia, ES & Azambuja, P. Development and interactions of Trypanosoma cruzi
within the insect vector. Parasitol Today. 1991 Sep;7(9):240-4.
Garg, N & Bhatia, V. Current status and future prospects for a vaccine against
American trypanosomiasis. Expert Rev Vaccines. 2005 Dec;4(6):867-80.
George, R, Beddoe, T, Landl, K, Lithgow, T. The yeast nascent polypeptide-
associated complex initiates protein targeting to mitochondria in vivo. Proc Natl
Acad Sci U S A. 1998 Mar 3;95(5):2296-301.
Gianazza, E, Osnaghi, A, Bontempi, L, Righetti, PG, Celentano, F. Ion retention by
immobilized pH matrices. Appl Theor Electrophor. 1989;1(3):155-9.
Gille, C, Goede, A, Schloetelburg, C, Preissner, R, Kloetzel, PM, Gobel, UB, et al. A
comprehensive view on proteasomal sequences: implications for the evolution
of the proteasome. J Mol Biol. 2003 Mar 7;326(5):1437-48.

Referências
120
Gilles, H. Parasitic disease affecting the heart in childhood. Images Paediatr Cardiol.
2000;5:28-39.
Gnanasekar, M, Rao, KV, Chen, L, Narayanan, RB, Geetha, M, Scott, AL, et al.
Molecular characterization of a calcium binding translationally controlled tumor
protein homologue from the filarial parasites Brugia malayi and Wuchereria
bancrofti. Mol Biochem Parasitol. 2002 Apr 30;121(1):107-18.
Golding, GB & Dean, AM. The structural basis of molecular adaptation. Mol Biol Evol.
1998 Apr;15(4):355-69.
Gonzalez-Pino, MJ, Rangel-Aldao, R, Slezynger, TC. Expression of alpha- and beta-
tubulin genes during growth of Trypanosoma cruzi epimastigotes. DNA Cell
Biol. 1999 Jun;18(6):449-55.
Goshe, MB. Characterizing phosphoproteins and phosphoproteomes using mass
spectrometry. Brief Funct Genomic Proteomic. 2006 Feb;4(4):363-76.
Graham, SV & Barry, JD. Transcriptional regulation of metacyclic variant surface
glycoprotein gene expression during the life cycle of Trypanosoma brucei. Mol
Cell Biol. 1995 Nov;15(11):5945-56.
Grant, AG, Flomen, RM, Tizard, ML, Grant, DA. Differential screening of a human
pancreatic adenocarcinoma lambda gt11 expression library has identified
increased transcription of elongation factor EF-1 alpha in tumour cells. Int J
Cancer. 1992 Mar 12;50(5):740-5.
Hall, BS & Pereira, MA. Dual role for transforming growth factor beta-dependent
signaling in Trypanosoma cruzi infection of mammalian cells. Infect Immun.
2000 Apr;68(4):2077-81.
Hall, SW, LaMarre, J, Marshall, LB, Hayes, MA, Gonias, SL. Binding of transforming
growth factor-beta 1 to methylamine-modified alpha 2-macroglobulin and to
binary and ternary alpha 2-macroglobulin-proteinase complexes. Biochem J.
1992 Jan 15;281 (Pt 2):569-75.
Hancock, W, Apffel, A, Chakel, J, Hahnenberger, K, Choudhary, G, Traina, JA, et al.
Integrated genomic/proteomic analysis. Anal Chem. 1999 Nov 1;71(21):742A-
8A.

Referências
121
Hayaishi, O. Molecular mechanisms of sleep-wake regulation: a role of prostaglandin
D2. Philos Trans R Soc Lond B Biol Sci. 2000 Feb 29;355(1394):275-80.
Hershey, JW. Translational control in mammalian cells. Annu Rev Biochem.
1991;60:717-55.
Higuchi, ML, Fukasawa, S, De Brito, T, Parzianello, LC, Bellotti, G, Ramires, JA.
Different microcirculatory and interstitial matrix patterns in idiopathic dilated
cardiomyopathy and Chagas' disease: a three dimensional confocal microscopy
study. Heart. 1999 Sep;82(3):279-85.
Iavarone, A & Massague, J. Repression of the CDK activator Cdc25A and cell-cycle
arrest by cytokine TGF-beta in cells lacking the CDK inhibitor p15. Nature. 1997
May 22;387(6631):417-22.
Inagaki, Y, Truter, S, Ramirez, F. Transforming growth factor-beta stimulates alpha
2(I) collagen gene expression through a cis-acting element that contains an
Sp1-binding site. J Biol Chem. 1994 May 20;269(20):14828-34.
Iozzo, RV, Pillarisetti, J, Sharma, B, Murdoch, AD, Danielson, KG, Uitto, J, et al.
Structural and functional characterization of the human perlecan gene promoter.
Transcriptional activation by transforming growth factor-beta via a nuclear factor
1-binding element. J Biol Chem. 1997 Feb 21;272(8):5219-28.
Itoh, S, Itoh, F, Goumans, MJ, Ten Dijke, P. Signaling of transforming growth factor-
beta family members through Smad proteins. Eur J Biochem. 2000
Dec;267(24):6954-67.
Iwasaki, Y, Lamar, C, Danenberg, K, Pitot, HC. Studies on the induction and
repression of enzymes in rat liver. Characterization and metabolic regulation of
multiple forms of tyrosine aminotransferase. Eur J Biochem. 1973
Apr;34(2):347-57.
Janssen, GM, van Damme, HT, Kriek, J, Amons, R, Moller, W. The subunit structure
of elongation factor 1 from Artemia. Why two alpha-chains in this complex? J
Biol Chem. 1994 Dec 16;269(50):31410-7.

Referências
122
Kanemori, M, Nishihara, K, Yanagi, H, Yura, T. Synergistic roles of HslVU and other
ATP-dependent proteases in controlling in vivo turnover of sigma32 and
abnormal proteins in Escherichia coli. J Bacteriol. 1997 Dec;179(23):7219-25.
Keeton, MR, Curriden, SA, van Zonneveld, AJ, Loskutoff, DJ. Identification of
regulatory sequences in the type 1 plasminogen activator inhibitor gene
responsive to transforming growth factor beta. J Biol Chem. 1991 Dec
5;266(34):23048-52.
Kerr, LD, Miller, DB, Matrisian, LM. TGF-beta 1 inhibition of transin/stromelysin gene
expression is mediated through a Fos binding sequence. Cell. 1990 Apr
20;61(2):267-78.
Kirchhoff, LV. Chagas disease. American trypanosomiasis. Infect Dis Clin North Am.
1993 Sep;7(3):487-502.
Kohl, L & Gull, K. Molecular architecture of the trypanosome cytoskeleton. Mol
Biochem Parasitol. 1998 May 15;93(1):1-9.
Kollien, AH & Schaub, GA. The development of Trypanosoma cruzi in triatominae.
Parasitol Today. 2000 Sep;16(9):381-7.
Kretzschmar, M, Doody, J, Timokhina, I, Massague, J. A mechanism of repression of
TGFbeta/ Smad signaling by oncogenic Ras. Genes Dev. 1999 Apr
1;13(7):804-16.
Krieg, PA, Varnum, SM, Wormington, WM, Melton, DA. The mRNA encoding
elongation factor 1-alpha (EF-1 alpha) is a major transcript at the midblastula
transition in Xenopus. Dev Biol. 1989 May;133(1):93-100.
Kubata, BK, Duszenko, M, Kabututu, Z, Rawer, M, Szallies, A, Fujimori, K, et al.
Identification of a novel prostaglandin f(2alpha) synthase in Trypanosoma
brucei. J Exp Med. 2000 Nov 6;192(9):1327-38.
Kuhnel, K & Luisi, BF. Crystal structure of the Escherichia coli RNA degradosome
component enolase. J Mol Biol. 2001 Oct 26;313(3):583-92.
Laemmli, UK. Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature. 1970 Aug 15;227(5259):680-5.

Referências
123
Lauring, B, Sakai, H, Kreibich, G, Wiedmann, M. Nascent polypeptide-associated
complex protein prevents mistargeting of nascent chains to the endoplasmic
reticulum. Proc Natl Acad Sci U S A. 1995 Jun 6;92(12):5411-5.
Lawrence, DA. Transforming growth factor-beta: an overview. Kidney Int Suppl. 1995
Jun;49:S19-23.
Li, F, Zhang, D, Fujise, K. Characterization of fortilin, a novel antiapoptotic protein. J
Biol Chem. 2001 Dec 14;276(50):47542-9.
Lijnen, PJ, Petrov, VV, Fagard, RH. Induction of cardiac fibrosis by transforming
growth factor-beta(1). Mol Genet Metab. 2000 Sep-Oct;71(1-2):418-35.
Ling, V, Guzzetta, AW, Canova-Davis, E, Stults, JT, Hancock, WS, Covey, TR, et al.
Characterization of the tryptic map of recombinant DNA derived tissue
plasminogen activator by high-performance liquid chromatography-electrospray
ionization mass spectrometry. Anal Chem. 1991 Dec 15;63(24):2909-15.
Lipowsky, G, Bischoff, FR, Schwarzmaier, P, Kraft, R, Kostka, S, Hartmann, E, et al.
Exportin 4: a mediator of a novel nuclear export pathway in higher eukaryotes.
EMBO J. 2000 Aug 15;19(16):4362-71.
Lowry, OH, Rosebrough, NJ, Farr, AL, Randall, RJ. Protein measurement with the
Folin phenol reagent. J Biol Chem. 1951 Nov;193(1):265-75.
Luchtan, M, Warade, C, Weatherly, DB, Degrave, WM, Tarleton, RL, Kissinger, JC.
TcruziDB: an integrated Trypanosoma cruzi genome resource. Nucleic Acids
Res. 2004 Jan 1;32(Database issue):D344-6.
Lutz, M & Knaus, P. Integration of the TGF-beta pathway into the cellular signalling
network. Cell Signal. 2002 Dec;14(12):977-88.
Lyons, RM & Moses, HL. Transforming growth factors and the regulation of cell
proliferation. Eur J Biochem. 1990 Feb 14;187(3):467-73.
MacDonald, SM, Bhisutthibhan, J, Shapiro, TA, Rogerson, SJ, Taylor, TE, Tembo, M,
et al. Immune mimicry in malaria: Plasmodium falciparum secretes a functional
histamine-releasing factor homolog in vitro and in vivo. Proc Natl Acad Sci U S
A. 2001 Sep 11;98(19):10829-32.

Referências
124
MacDonald, SM, Rafnar, T, Langdon, J, Lichtenstein, LM. Molecular identification of
an IgE-dependent histamine-releasing factor. Science. 1995 Aug
4;269(5224):688-90.
Magalhaes, AD, Charneau, S, Paba, J, Guercio, RA, Teixeira, AR, Santana, JM, et
al. Trypanosoma cruzi alkaline 2-DE: Optimization and application to
comparative proteome analysis of flagellate life stages. Proteome Sci.
2008;6:24.
Mann, M & Jensen, ON. Proteomic analysis of post-translational modifications. Nat
Biotechnol. 2003 Mar;21(3):255-61.
Marigo, V, Volpin, D, Vitale, G, Bressan, GM. Identification of a TGF-beta responsive
element in the human elastin promoter. Biochem Biophys Res Commun. 1994
Mar 15;199(2):1049-56.
Marks, AR. Cellular functions of immunophilins. Physiol Rev. 1996 Jul;76(3):631-49.
Martinez, J, Campetella, O, Frasch, AC, Cazzulo, JJ. The major cysteine proteinase
(cruzipain) from Trypanosoma cruzi is antigenic in human infections. Infect
Immun. 1991 Nov;59(11):4275-7.
Massague, J. The transforming growth factor-beta family. Annu Rev Cell Biol.
1990;6:597-641.
Massague, J. TGF-beta signal transduction. Annu Rev Biochem. 1998;67:753-91.
Massague, J & Chen, YG. Controlling TGF-beta signaling. Genes Dev. 2000 Mar
15;14(6):627-44.
Massague, J & Wotton, D. Transcriptional control by the TGF-beta/Smad signaling
system. EMBO J. 2000 Apr 17;19(8):1745-54.
Mathe, AA, Hedqvist, P, Strandberg, K, Leslie, CA. Aspects of prostaglandin function
in the lung (second of two parts). N Engl J Med. 1977 Apr 21;296(16):910-4.
Mayer, MP & Bukau, B. Hsp70 chaperones: cellular functions and molecular
mechanism. Cell Mol Life Sci. 2005 Mar;62(6):670-84.

Referências
125
McCracken, JA, Carlson, JC, Glew, ME, Goding, JR, Baird, DT, Green, K, et al.
Prostaglandin F 2 identified as a luteolytic hormone in sheep. Nat New Biol.
1972 Aug 2;238(83):129-34.
Merrick, WC. Mechanism and regulation of eukaryotic protein synthesis. Microbiol
Rev. 1992 Jun;56(2):291-315.
Ming, M, Ewen, ME, Pereira, ME. Trypanosome invasion of mammalian cells
requires activation of the TGF beta signaling pathway. Cell. 1995 Jul
28;82(2):287-96.
Mitchell, H, Choudhury, A, Pagano, RE, Leof, EB. Ligand-dependent and -
independent transforming growth factor-beta receptor recycling regulated by
clathrin-mediated endocytosis and Rab11. Mol Biol Cell. 2004 Sep;15(9):4166-
78.
Moazed, D & Noller, HF. Intermediate states in the movement of transfer RNA in the
ribosome. Nature. 1989 Nov 9;342(6246):142-8.
Moller, I, Jung, M, Beatrix, B, Levy, R, Kreibich, G, Zimmermann, R, et al. A general
mechanism for regulation of access to the translocon: competition for a
membrane attachment site on ribosomes. Proc Natl Acad Sci U S A. 1998 Nov
10;95(23):13425-30.
Moncayo, A & Silveira, AC. Current epidemiological trends for Chagas disease in
Latin America and future challenges in epidemiology, surveillance and health
policy. Mem Inst Oswaldo Cruz. 2009 Jul;104 Suppl 1:17-30.
Montemartini, M, Santome, JA, Cazzulo, JJ, Nowicki, C. Purification and partial
structural and kinetic characterization of tyrosine aminotransferase from
epimastigotes of Trypanosoma cruzi. Biochem J. 1993 Jun 15;292 ( Pt 3):901-6.
Montemartini, M, Santome, JA, Cazzulo, JJ, Nowicki, C. Purification and partial
structural and kinetic characterization of an aromatic L-alpha-hydroxy acid
dehydrogenase from epimastigotes of Trypanosoma cruzi. Mol Biochem
Parasitol. 1994 Nov;68(1):15-23.

Referências
126
Morales, MA, Watanabe, R, Laurent, C, Lenormand, P, Rousselle, JC, Namane, A, et
al. Phosphoproteomic analysis of Leishmania donovani pro- and amastigote
stages. Proteomics. 2008 Jan;8(2):350-63.
Moustakas, A & Heldin, CH. Non-Smad TGF-beta signals. J Cell Sci. 2005 Aug
15;118(Pt 16):3573-84.
Moustakas, A, Pardali, K, Gaal, A, Heldin, CH. Mechanisms of TGF-beta signaling in
regulation of cell growth and differentiation. Immunol Lett. 2002 Jun 3;82(1-
2):85-91.
Moustakas, A, Souchelnytskyi, S, Heldin, CH. Smad regulation in TGF-beta signal
transduction. J Cell Sci. 2001 Dec;114(Pt 24):4359-69.
Murphy-Ullrich, JE & Poczatek, M. Activation of latent TGF-beta by thrombospondin-
1: mechanisms and physiology. Cytokine Growth Factor Rev. 2000 Mar-
Jun;11(1-2):59-69.
Nakayasu, ES, Gaynor, MR, Sobreira, TJ, Ross, JA, Almeida, IC. Phosphoproteomic
analysis of the human pathogen Trypanosoma cruzi at the epimastigote stage.
Proteomics. 2009 Jul;9(13):3489-506.
Naula, C, Parsons, M, Mottram, JC. Protein kinases as drug targets in trypanosomes
and Leishmania. Biochim Biophys Acta. 2005 Dec 30;1754(1-2):151-9.
Neuwald, AF, Aravind, L, Spouge, JL, Koonin, EV. AAA+: A class of chaperone-like
ATPases associated with the assembly, operation, and disassembly of protein
complexes. Genome Res. 1999 Jan;9(1):27-43.
Nielsen, HV, Johnsen, AH, Sanchez, JC, Hochstrasser, DF, Schiotz, PO.
Identification of a basophil leukocyte interleukin-3-regulated protein that is
identical to IgE-dependent histamine-releasing factor. Allergy. 1998
Jul;53(7):642-52.
Noiva, R & Lennarz, WJ. Protein disulfide isomerase. A multifunctional protein
resident in the lumen of the endoplasmic reticulum. J Biol Chem. 1992 Feb
25;267(6):3553-6.

Referências
127
Nollen, EA & Morimoto, RI. Chaperoning signaling pathways: molecular chaperones
as stress-sensing 'heat shock' proteins. J Cell Sci. 2002 Jul 15;115(Pt 14):2809-
16.
Nowicki, C & Cazzulo, JJ. Aromatic amino acid catabolism in trypanosomatids. Comp
Biochem Physiol A Mol Integr Physiol. 2008 Nov;151(3):381-90.
Nowicki, C, Hunter, GR, Montemartini-Kalisz, M, Blankenfeldt, W, Hecht, H, Kalisz,
HM. Recombinant tyrosine aminotransferase from Trypanosoma cruzi:
structural characterization and site directed mutagenesis of a broad substrate
specificity enzyme. Biochim Biophys Acta. 2001 Apr 7;1546(2):268-81.
Nowicki, C, Montemartini, M, Duschak, V, Santome, JA, Cazzulo, JJ. Presence and
subcellular localization of tyrosine aminotransferase and p-hydroxyphenyllactate
dehydrogenase in epimastigotes of Trypanosoma cruzi. FEMS Microbiol Lett.
1992 Apr 15;71(2):119-24.
Okano, Y, Inoue, T, Kubata, BK, Kabututu, Z, Urade, Y, Matsumura, H, et al.
Crystallization and preliminary X-ray crystallographic studies of Trypanosoma
brucei prostaglandin F(2 alpha) synthase. J Biochem. 2002 Dec;132(6):859-61.
Ouaissi, A. Apoptosis-like death in trypanosomatids: search for putative pathways
and genes involved. Kinetoplastid Biol Dis. 2003 Jun 25;2(1):5.
Paba, J, Santana, JM, Teixeira, AR, Fontes, W, Sousa, MV, Ricart, CA. Proteomic
analysis of the human pathogen Trypanosoma cruzi. Proteomics. 2004
Apr;4(4):1052-9.
Paradela, A & Albar, JP. Advances in the analysis of protein phosphorylation. J
Proteome Res. 2008 May;7(5):1809-18.
Park, E, Rho, YM, Koh, OJ, Ahn, SW, Seong, IS, Song, JJ, et al. Role of the GYVG
pore motif of HslU ATPase in protein unfolding and translocation for
degradation by HslV peptidase. J Biol Chem. 2005 Jun 17;280(24):22892-8.
Park, MH. The post-translational synthesis of a polyamine-derived amino acid,
hypusine, in the eukaryotic translation initiation factor 5A (eIF5A). J Biochem.
2006 Feb;139(2):161-9.

Referências
128
Park, MH, Lee, YB, Joe, YA. Hypusine is essential for eukaryotic cell proliferation.
Biol Signals. 1997 May-Jun;6(3):115-23.
Parodi-Talice, A, Duran, R, Arrambide, N, Prieto, V, Pineyro, MD, Pritsch, O, et al.
Proteome analysis of the causative agent of Chagas disease: Trypanosoma
cruzi. Int J Parasitol. 2004 Jul;34(8):881-6.
Parsons, M, Worthey, EA, Ward, PN, Mottram, JC. Comparative analysis of the
kinomes of three pathogenic trypanosomatids: Leishmania major, Trypanosoma
brucei and Trypanosoma cruzi. BMC Genomics. 2005;6:127.
Pawson, T. Regulation and targets of receptor tyrosine kinases. Eur J Cancer. 2002
Sep;38 Suppl 5:S3-10.
Piacenza, L, Peluffo, G, Alvarez, MN, Kelly, JM, Wilkinson, SR, Radi, R.
Peroxiredoxins play a major role in protecting Trypanosoma cruzi against
macrophage- and endogenously-derived peroxynitrite. Biochem J. 2008 Mar
1;410(2):359-68.
Piek, E, Heldin, CH, Ten Dijke, P. Specificity, diversity, and regulation in TGF-beta
superfamily signaling. FASEB J. 1999 Dec;13(15):2105-24.
Prata, A. Clinical and epidemiological aspects of Chagas disease. Lancet Infect Dis.
2001 Sep;1(2):92-100.
Procopio, DO, Barros, HC, Mortara, RA. Actin-rich structures formed during the
invasion of cultured cells by infective forms of Trypanosoma cruzi. Eur J Cell
Biol. 1999 Dec;78(12):911-24.
Rao, KV, Chen, L, Gnanasekar, M, Ramaswamy, K. Cloning and characterization of
a calcium-binding, histamine-releasing protein from Schistosoma mansoni. J
Biol Chem. 2002 Aug 23;277(34):31207-13.
Rassi, A, Jr., Rassi, A, Little, WC. Chagas' heart disease. Clin Cardiol. 2000
Dec;23(12):883-9.
Rassi, A, Jr., Rassi, A, Marin-Neto, JA. Chagas heart disease: pathophysiologic
mechanisms, prognostic factors and risk stratification. Mem Inst Oswaldo Cruz.
2009 Jul;104 Suppl 1:152-8.

Referências
129
Razani, B, Zhang, XL, Bitzer, M, von Gersdorff, G, Bottinger, EP, Lisanti, MP.
Caveolin-1 regulates transforming growth factor (TGF)-beta/SMAD signaling
through an interaction with the TGF-beta type I receptor. J Biol Chem. 2001 Mar
2;276(9):6727-38.
Riccio, A, Pedone, PV, Lund, LR, Olesen, T, Olsen, HS, Andreasen, PA.
Transforming growth factor beta 1-responsive element: closely associated
binding sites for USF and CCAAT-binding transcription factor-nuclear factor I in
the type 1 plasminogen activator inhibitor gene. Mol Cell Biol. 1992
Apr;12(4):1846-55.
Ritzenthaler, JD, Goldstein, RH, Fine, A, Smith, BD. Regulation of the alpha 1(I)
collagen promoter via a transforming growth factor-beta activation element. J
Biol Chem. 1993 Jun 25;268(18):13625-31.
Roberts, AB, Flanders, KC, Heine, UI, Jakowlew, S, Kondaiah, P, Kim, SJ, et al.
Transforming growth factor-beta: multifunctional regulator of differentiation and
development. Philos Trans R Soc Lond B Biol Sci. 1990 Mar 12;327(1239):145-
54.
Roberts, AB & Sporn, MB. Physiological actions and clinical applications of
transforming growth factor-beta (TGF-beta). Growth Factors. 1993;8(1):1-9.
Robinson, D, Beattie, P, Sherwin, T, Gull, K. Microtubules, tubulin, and microtubule-
associated proteins of trypanosomes. Methods Enzymol. 1991;196:285-99.
Roepstorff, P. Mass spectrometry in protein studies from genome to function. Curr
Opin Biotechnol. 1997 Feb;8(1):6-13.
Rohloff, P, Rodrigues, CO, Docampo, R. Regulatory volume decrease in
Trypanosoma cruzi involves amino acid efflux and changes in intracellular
calcium. Mol Biochem Parasitol. 2003 Feb;126(2):219-30.
Rohrwild, M, Coux, O, Huang, HC, Moerschell, RP, Yoo, SJ, Seol, JH, et al. HslV-
HslU: A novel ATP-dependent protease complex in Escherichia coli related to
the eukaryotic proteasome. Proc Natl Acad Sci U S A. 1996 Jun
11;93(12):5808-13.

Referências
130
Rosenkranz, S. TGF-beta1 and angiotensin networking in cardiac remodeling.
Cardiovasc Res. 2004 Aug 15;63(3):423-32.
Rosenzweig, D, Smith, D, Myler, PJ, Olafson, RW, Zilberstein, D. Post-translational
modification of cellular proteins during Leishmania donovani differentiation.
Proteomics. 2008 May;8(9):1843-50.
Rosorius, O, Reichart, B, Kratzer, F, Heger, P, Dabauvalle, MC, Hauber, J. Nuclear
pore localization and nucleocytoplasmic transport of eIF-5A: evidence for direct
interaction with the export receptor CRM1. J Cell Sci. 1999 Jul;112 ( Pt
14):2369-80.
Rospert, S, Dubaquie, Y, Gautschi, M. Nascent-polypeptide-associated complex. Cell
Mol Life Sci. 2002 Oct;59(10):1632-9.
Rossi, MA. Connective tissue skeleton in the normal left ventricle and in hypertensive
left ventricular hypertrophy and chronic chagasic myocarditis. Med Sci Monit.
2001 Jul-Aug;7(4):820-32.
Rossi, MA & Bestetti, RB. The challenge of chagasic cardiomyopathy. The pathologic
roles of autonomic abnormalities, autoimmune mechanisms and microvascular
changes, and therapeutic implications. Cardiology. 1995;86(1):1-7.
Rossi, P, Karsenty, G, Roberts, AB, Roche, NS, Sporn, MB, de Crombrugghe, B. A
nuclear factor 1 binding site mediates the transcriptional activation of a type I
collagen promoter by transforming growth factor-beta. Cell. 1988 Feb
12;52(3):405-14.
Rubotham, J, Woods, K, Garcia-Salcedo, JA, Pays, E, Nolan, DP. Characterization of
two protein disulfide isomerases from the endocytic pathway of bloodstream
forms of Trypanosoma brucei. J Biol Chem. 2005 Mar 18;280(11):10410-8.
Ruhl, M, Himmelspach, M, Bahr, GM, Hammerschmid, F, Jaksche, H, Wolff, B, et al.
Eukaryotic initiation factor 5A is a cellular target of the human immunodeficiency
virus type 1 Rev activation domain mediating trans-activation. J Cell Biol. 1993
Dec;123(6 Pt 1):1309-20.
Samuelsson, B. Prostaglandins, thromboxanes, and leukotrienes: formation and
biological roles. Harvey Lect. 1979;75:1-40.

Referências
131
Sanders, J, Maassen, JA, Moller, W. Elongation factor-1 messenger-RNA levels in
cultured cells are high compared to tissue and are not drastically affected
further by oncogenic transformation. Nucleic Acids Res. 1992 Nov
25;20(22):5907-10.
Sant'Anna, C, Parussini, F, Lourenco, D, de Souza, W, Cazzulo, JJ, Cunha-e-Silva,
NL. All Trypanosoma cruzi developmental forms present lysosome-related
organelles. Histochem Cell Biol. 2008 Dec;130(6):1187-98.
Sasaki, A, Masuda, Y, Ohta, Y, Ikeda, K, Watanabe, K. Filamin associates with
Smads and regulates transforming growth factor-beta signaling. J Biol Chem.
2001 May 25;276(21):17871-7.
Schmunis, GA. Epidemiology of Chagas disease in non-endemic countries: the role
of international migration. Mem Inst Oswaldo Cruz. 2007 Oct 30;102 Suppl
1:75-85.
Schnier, J, Schwelberger, HG, Smit-McBride, Z, Kang, HA, Hershey, JW. Translation
initiation factor 5A and its hypusine modification are essential for cell viability in
the yeast Saccharomyces cerevisiae. Mol Cell Biol. 1991 Jun;11(6):3105-14.
Schulenberg, B, Goodman, TN, Aggeler, R, Capaldi, RA, Patton, WF.
Characterization of dynamic and steady-state protein phosphorylation using a
fluorescent phosphoprotein gel stain and mass spectrometry. Electrophoresis.
2004 Aug;25(15):2526-32.
Seo, J & Lee, KJ. Post-translational modifications and their biological functions:
proteomic analysis and systematic approaches. J Biochem Mol Biol. 2004 Jan
31;37(1):35-44.
Seong, IS, Oh, JY, Lee, JW, Tanaka, K, Chung, CH. The HslU ATPase acts as a
molecular chaperone in prevention of aggregation of SulA, an inhibitor of cell
division in Escherichia coli. FEBS Lett. 2000 Jul 21;477(3):224-9.
Shi, XP, Yin, KC, Zimolo, ZA, Stern, AM, Waxman, L. The subcellular distribution of
eukaryotic translation initiation factor, eIF-5A, in cultured cells. Exp Cell Res.
1996 Jun 15;225(2):348-56.

Referências
132
Shi, Y & Massague, J. Mechanisms of TGF-beta signaling from cell membrane to the
nucleus. Cell. 2003 Jun 13;113(6):685-700.
Silva, JS, Twardzik, DR, Reed, SG. Regulation of Trypanosoma cruzi infections in
vitro and in vivo by transforming growth factor beta (TGF-beta). J Exp Med.
1991 Sep 1;174(3):539-45.
Sinha, P, Kohl, S, Fischer, J, Hutter, G, Kern, M, Kottgen, E, et al. Identification of
novel proteins associated with the development of chemoresistance in
malignant melanoma using two-dimensional electrophoresis. Electrophoresis.
2000 Aug;21(14):3048-57.
Soares, MJ & de Souza, W. Endocytosis of gold-labeled proteins and LDL by
Trypanosoma cruzi. Parasitol Res. 1991;77(6):461-8.
Soares, MJ, Souto-Padron, T, Bonaldo, MC, Goldenberg, S, de Souza, W. A
stereological study of the differentiation process in Trypanosoma cruzi. Parasitol
Res. 1989;75(7):522-7.
Soares, MJ, Souto-Padron, T, De Souza, W. Identification of a large pre-lysosomal
compartment in the pathogenic protozoon Trypanosoma cruzi. J Cell Sci. 1992
May;102 ( Pt 1):157-67.
Sodre, CL, Chapeaurouge, AD, Kalume, DE, de Mendonca Lima, L, Perales, J,
Fernandes, O. Proteomic map of Trypanosoma cruzi CL Brener: the reference
strain of the genome project. Arch Microbiol. 2009 Feb;191(2):177-84.
Souto-Padron, T, Campetella, OE, Cazzulo, JJ, de Souza, W. Cysteine proteinase in
Trypanosoma cruzi: immunocytochemical localization and involvement in
parasite-host cell interaction. J Cell Sci. 1990 Jul;96 ( Pt 3):485-90.
Steinberg, TH, Agnew, BJ, Gee, KR, Leung, WY, Goodman, T, Schulenberg, B, et al.
Global quantitative phosphoprotein analysis using Multiplexed Proteomics
technology. Proteomics. 2003 Jul;3(7):1128-44.
Stempin, C, Giordanengo, L, Gea, S, Cerban, F. Alternative activation and increase
of Trypanosoma cruzi survival in murine macrophages stimulated by cruzipain,
a parasite antigen. J Leukoc Biol. 2002 Oct;72(4):727-34.

Referências
133
Taipale, J, Saharinen, J, Hedman, K, Keski-Oja, J. Latent transforming growth factor-
beta 1 and its binding protein are components of extracellular matrix microfibrils.
J Histochem Cytochem. 1996 Aug;44(8):875-89.
Tang, Y, Katuri, V, Dillner, A, Mishra, B, Deng, CX, Mishra, L. Disruption of
transforming growth factor-beta signaling in ELF beta-spectrin-deficient mice.
Science. 2003 Jan 24;299(5606):574-7.
Tarze, A, Deniaud, A, Le Bras, M, Maillier, E, Molle, D, Larochette, N, et al. GAPDH,
a novel regulator of the pro-apoptotic mitochondrial membrane
permeabilization. Oncogene. 2007 Apr 19;26(18):2606-20.
Teshima, S, Rokutan, K, Nikawa, T, Kishi, K. Macrophage colony-stimulating factor
stimulates synthesis and secretion of a mouse homolog of a human IgE-
dependent histamine-releasing factor by macrophages in vitro and in vivo. J
Immunol. 1998 Dec 1;161(11):6356-66.
Thiele, H, Berger, M, Skalweit, A, Thiele, BJ. Expression of the gene and processed
pseudogenes encoding the human and rabbit translationally controlled tumour
protein (TCTP). Eur J Biochem. 2000 Sep;267(17):5473-81.
Tisdale, EJ. Glyceraldehyde-3-phosphate dehydrogenase is required for vesicular
transport in the early secretory pathway. J Biol Chem. 2001 Jan
26;276(4):2480-6.
Tisdale, EJ & Artalejo, CR. A GAPDH mutant defective in Src-dependent tyrosine
phosphorylation impedes Rab2-mediated events. Traffic. 2007 Jun;8(6):733-41.
Todaro, GJ, Fryling, C, De Larco, JE. Transforming growth factors produced by
certain human tumor cells: polypeptides that interact with epidermal growth
factor receptors. Proc. Natl. Acad. Sci. USA. 1980 Sep;77(9): 5258-5262.
Torres-Cabassa, AS & Gottesman, S. Capsule synthesis in Escherichia coli K-12 is
regulated by proteolysis. J Bacteriol. 1987 Mar;169(3):981-9.
Tyers, M & Mann, M. From genomics to proteomics. Nature. 2003 Mar
13;422(6928):193-7.
Urbina, JA & Docampo, R. Specific chemotherapy of Chagas disease: controversies
and advances. Trends Parasitol. 2003 Nov;19(11):495-501.

Referências
134
Valentini, SR, Casolari, JM, Oliveira, CC, Silver, PA, McBride, AE. Genetic
interactions of yeast eukaryotic translation initiation factor 5A (eIF5A) reveal
connections to poly(A)-binding protein and protein kinase C signaling. Genetics.
2002 Feb;160(2):393-405.
Vanhamme, L & Pays, E. Control of gene expression in trypanosomes. Microbiol
Rev. 1995 Jun;59(2):223-40.
Verlinde, CL, Hannaert, V, Blonski, C, Willson, M, Perie, JJ, Fothergill-Gilmore, LA, et
al. Glycolysis as a target for the design of new anti-trypanosome drugs. Drug
Resist Updat. 2001 Feb;4(1):50-65.
Waghabi, MC, Coutinho, CM, Soeiro, MN, Pereira, MC, Feige, JJ, Keramidas, M, et
al. Increased Trypanosoma cruzi invasion and heart fibrosis associated with
high transforming growth factor beta levels in mice deficient in alpha(2)-
macroglobulin. Infect Immun. 2002 Sep;70(9):5115-23.
Waghabi, MC, de Souza, EM, de Oliveira, GM, Keramidas, M, Feige, JJ, Araujo-
Jorge, TC, et al. Pharmacological Inhibition of Transforming Growth Factor Beta
Signalling Decreases Infection and Prevents Heart Damage in Acute Chagas
Disease. Antimicrob Agents Chemother. 2009 Sep 8.
Waghabi, MC, Keramidas, M, Bailly, S, Degrave, W, Mendonca-Lima, L, Soeiro Mde,
N, et al. Uptake of host cell transforming growth factor-beta by Trypanosoma
cruzi amastigotes in cardiomyocytes: potential role in parasite cycle completion.
Am J Pathol. 2005a Oct;167(4):993-1003.
Waghabi, MC, Keramidas, M, Calvet, CM, Meuser, M, de Nazare, CSM, Mendonca-
Lima, L, et al. SB-431542, a transforming growth factor beta inhibitor, impairs
Trypanosoma cruzi infection in cardiomyocytes and parasite cycle completion.
Antimicrob Agents Chemother. 2007 Aug;51(8):2905-10.
Waghabi, MC, Keramidas, M, Feige, JJ, Araujo-Jorge, TC, Bailly, S. Activation of
transforming growth factor beta by Trypanosoma cruzi. Cell Microbiol. 2005b
Apr;7(4):511-7.
Waite, KA & Eng, C. From developmental disorder to heritable cancer: it's all in the
BMP/TGF-beta family. Nat Rev Genet. 2003 Oct;4(10):763-73.

Referências
135
Walker, DJ, Pitsch, JL, Peng, MM, Robinson, BL, Peters, W, Bhisutthibhan, J, et al.
Mechanisms of artemisinin resistance in the rodent malaria pathogen
Plasmodium yoelii. Antimicrob Agents Chemother. 2000 Feb;44(2):344-7.
Wang, P & Heitman, J. The cyclophilins. Genome Biol. 2005;6(7):226.
Wang, S, Sakai, H, Wiedmann, M. NAC covers ribosome-associated nascent chains
thereby forming a protective environment for regions of nascent chains just
emerging from the peptidyl transferase center. J Cell Biol. 1995
Aug;130(3):519-28.
Wiedmann, B, Sakai, H, Davis, TA, Wiedmann, M. A protein complex required for
signal-sequence-specific sorting and translocation. Nature. 1994 Aug
11;370(6489):434-40.
Wilkinson, SR, Taylor, MC, Horn, D, Kelly, JM, Cheeseman, I. A mechanism for
cross-resistance to nifurtimox and benznidazole in trypanosomes. Proc Natl
Acad Sci U S A. 2008 Apr 1;105(13):5022-7.
Wyllie, S, Vickers, TJ, Fairlamb, AH. Roles of trypanothione S-transferase and
tryparedoxin peroxidase in resistance to antimonials. Antimicrob Agents
Chemother. 2008 Apr;52(4):1359-65.
Xin, H, Xu, X, Li, L, Ning, H, Rong, Y, Shang, Y, et al. CHIP controls the sensitivity of
transforming growth factor-beta signaling by modulating the basal level of
Smad3 through ubiquitin-mediated degradation. J Biol Chem. 2005 May
27;280(21):20842-50.
Xu, A, Bellamy, AR, Taylor, JA. Expression of translationally controlled tumour
protein is regulated by calcium at both the transcriptional and post-
transcriptional level. Biochem J. 1999 Sep 15;342 Pt 3:683-9.
Young, JC, Agashe, VR, Siegers, K, Hartl, FU. Pathways of chaperone-mediated
protein folding in the cytosol. Nat Rev Mol Cell Biol. 2004 Oct;5(10):781-91.
Yu, R, Chen, C, Mo, YY, Hebbar, V, Owuor, ED, Tan, TH, et al. Activation of
mitogen-activated protein kinase pathways induces antioxidant response
element-mediated gene expression via a Nrf2-dependent mechanism. J Biol
Chem. 2000 Dec 22;275(51):39907-13.

Referências
136
Yue, J & Mulder, KM. Activation of the mitogen-activated protein kinase pathway by
transforming growth factor-beta. Methods Mol Biol. 2000;142:125-31.
Zeledon, R, Alvarenga, N. J, Schosinsky, K. . Chagas Disease: Pan American Health
Organization1997 Contract No.: 347.
Zheng, L, Roeder, RG, Luo, Y. S phase activation of the histone H2B promoter by
OCA-S, a coactivator complex that contains GAPDH as a key component. Cell.
2003 Jul 25;114(2):255-66.
Zimmer, C, von Gabain, A, Henics, T. Analysis of sequence-specific binding of RNA
to Hsp70 and its various homologs indicates the involvement of N- and C-
terminal interactions. RNA. 2001 Nov;7(11):1628-37.
Zuk, D & Jacobson, A. A single amino acid substitution in yeast eIF-5A results in
mRNA stabilization. EMBO J. 1998 May 15;17(10):2914-25.
Zwaagstra, JC, El-Alfy, M, O'Connor-McCourt, MD. Transforming growth factor
(TGF)-beta 1 internalization: modulation by ligand interaction with TGF-beta
receptors types I and II and a mechanism that is distinct from clathrin-mediated
endocytosis. J Biol Chem. 2001 Jul 20;276(29):27237-45.


