II. TINJAUAN PUSTAKA 2.1. β-glukan -...
12
5 II. TINJAUAN PUSTAKA 2.1. β-glukan Glukan adalah turunan polisakarida alami yang terdiri dari unit D- glukopiranosil dan dapat ditemukan dalam berbagai macam sereal, tumbuhan, algae, bakteri, jamur, kapang dan khamir. Glukan terdiri dari rantai linear β-(1-3) dan terikat dengan rantai samping berupa β-(1-6) atau β-(1-4) dimana variasi rantai samping ini akan menentukan panjang dan distribusi glukan dan akan membentuk struktur tersier yang kompleks distabilkan oleh rantai ikatan hidrogen. (Zeković et al., 2005). Gambar 1. Polimer dari unit β-(1-4)-D-glikopiranosil dengan β-(1-3)-D-glikopiranosil (http://www.sigmaaldrich.com, 2018) Gambar 2. Polimer dari unit β-(1-3)-D-glikopiranosil dengan β-(1-6)-D-glikopiranosil (http://www.sigmaaldrich.com, 2018) Selulos Selulos Selulosa dan Laminarinase β(1,3)D-glukosa β(1,4)D-glukosa β(1,4)D- glukosa β(1,4)D-glukosa β(1,6)D-glukosa β(1,3)D-glukosa β(1,3)D- glukosa β(1,3)D-glukosa β-glukan Serealia β-glukan Mikroorganisme Litikase
Transcript of II. TINJAUAN PUSTAKA 2.1. β-glukan -...

5
II. TINJAUAN PUSTAKA
2.1. β-glukan
Glukan adalah turunan polisakarida alami yang terdiri dari unit D-
glukopiranosil dan dapat ditemukan dalam berbagai macam sereal, tumbuhan, algae,
bakteri, jamur, kapang dan khamir. Glukan terdiri dari rantai linear β-(1-3) dan
terikat dengan rantai samping berupa β-(1-6) atau β-(1-4) dimana variasi rantai
samping ini akan menentukan panjang dan distribusi glukan dan akan membentuk
struktur tersier yang kompleks distabilkan oleh rantai ikatan hidrogen. (Zeković et
al., 2005).
Gambar 1. Polimer dari unit β-(1-4)-D-glikopiranosil dengan β-(1-3)-D-glikopiranosil
(http://www.sigmaaldrich.com, 2018)
Gambar 2. Polimer dari unit β-(1-3)-D-glikopiranosil dengan β-(1-6)-D-glikopiranosil
(http://www.sigmaaldrich.com, 2018)
Selulos
a
Selulos
a
Selulosa dan
Laminarinase
β(1,3)D-glukosa β(1,4)D-glukosa
β(1,4)D-
glukosa
β(1,4)D-glukosa
β(1,6)D-glukosa
β(1,3)D-glukosa β(1,3)D-
glukosa β(1,3)D-glukosa
β-glukan Serealia
β-glukan Mikroorganisme
Litikase

6
Gambar 1 dan 2 menunjukkan β-glukan yang berasal dari khamir dan
mikroorganisme memiliki struktur dengan ikatan 1,3 dan 1,6 glukan, sedangkan
pada serealia mengandung ikatan β-1,3 dan 1,4 glukan. β-glukan yang diperoleh
dari serealia dapat menurunkan kolestrol dan gula darah dan glukan yang didapat
dari mikroorganisme dapat meningkatkan sistem imun dan anti tumor (Zhu et al.,
2016).
Beberapa sifat β-glukan antara lain adalah tidak beracun, tidak memiliki efek
samping yang merugikan, membantu regenerasi dan memperbaiki jaringan,
mengaktivasi dan memperkuat sistem kekebalan, serta mempertinggi keefektifan
obat antibiotik dan antiviral (Yenti, 2005). Dalam industri farmasi, β-glukan dapat
berfungsi untuk anti infeksi, mengobati luka luar, anti tumor, anti oksidan, dan
menurunkan kadar gula darah karena meningkatkan produksi insulin (Hendra,
2005).
Menurut Cheeseman dan Malcom (2000), sifat fisika dan kimia yang dimiliki
senyawa β-glukan yaitu di alam, berupa senyawa berwarna putih berupa gumpalan
besar dan tidak berbentuk kristal, tidak mempunyai rasa manis, tidak larut dalam
air netral dan dapat dipisahkan dengan mudah dalam larutan alkali, bila dicampur
dengan air maka akan membentuk larutan koloid, berbentuk gel pada suhu 54oC.
Polimer glukan ini merupakan serat yang tidak dapat dicerna, karena manusia
tidak memiliki enzim yang dapat menghidrolisis ikatan β- glikosidik. Serat-serat
yang tidak larut ini tidak dapat dimetabolisme pada saluran pencernaan sehingga
bermanfaat dalam diet yang berfungsi mengurangi kegemukan (Yenti, 2005).

7
2.2. Produksi β-glucan dari Saccharomyces cerevisiae
Khamir adalah kelompok jamur uniseluler berukuran lima hingga dua
puluh mikron yang umum dipergunakan untuk fermentasi roti dan minuman
beralkohol. Biasanya khamir berbentuk oval, namun ada yang berbentuk
memanjang dan berbentuk bola. Khamir tidak dilengkapi dengan flagelum, atau
organ penggerak lainnya (Pelchzar, 1986). Selain itu khamir bersifat anaerobik
fakultatif yang artinya mampu hidup dalam keadaan aerob ataupun anaerob. Suhu
maksimum pertumbuhan khamir antara 36-37oC, dan suhu minimumnya 9-11oC
(Judoamidjojo, 1992). Pertumbuhan maksimum biasanya terjadi sampai hari ketiga
dan mulai mengalami penurunan sampai hari ketujuh (Walker, 1995).
S. cerevisiae adalah salah satu khamir yang sering digunakan untuk
produksi β-glukan. Biasanya masyarakat mengenal S. cereviciae sebagai ragi roti
(baker’s yeast). Taksonomi dari S. cereviciae yaitu :
Kingdom : Fungi
Phylum : Ascomycota
Class : Hemiascomycota
Order : Saccharomycotales
Family : Saccharomycetaceae
Genus : Saccharomyces
Species : Saccharomyces cerevisiae
Sebuah sel khamir mampu memfermentasi glukosa dengan massa yang
sama dengan massa selnya sendiri dalam jangka waktu satu jam. Khamir dapat
bereproduksi secara aseksual dengan membentuk tunas ataupun secara seksual
dengan pembentukan ascospora. Selama proses reproduksi aseksual, sebuah tunas
baru tumbuh dari khamir dengan kondisi tertentu dan saat mencapai ukuran dewasa

8
ia akan melepaskan diri dari sel induknya. Reproduksi seksual khamir umumnya
berlangsung pada kondisi kekurangan nutrisi pertumbuhan dengan cara
pembentukan ascospora (European Bioinformatics Institute, 1996).
Gambar 3. Struktur dinding sel Saccharomyces cerevisiae
(Sumber: www.sigmaaldrich.com 2009)
Gambar 3 menunjukkan dinding sel khamir S. cereviciae terdiri dari 2 lapisan yang
dibangun oleh 4 molekul utama, yaitu manoprotein, β (1,6) glukan, β (1,3) glukan
dan kitin. Semua komponen ini tersambung oleh ikatan-ikatan kovalen. Kandungan
manoprotein dalam berat kering dinding sel menyampai 35-40%, kitin sekitar 1-2%,
pada β-1,6 glukan 5-10% dan lebih dari setengah dinding sel (50-55%) disusun
oleh β-1,3 glukan (Kapteyn,1996). Senyawa β-glukan didalam dinding sel S.
cereviciae berperan sebagai kerangka penyangga dari dinding sel dan berfungsi
memperkuat struktur dari selnya serta sebagai zat cadangan makanan. (Nguyen,
1995)
Mikroorganisme ini bersifat non patogenik dan non toksik (Lee,1992).
Saccharomyces cerevisiae ini juga menghasilkan enzim zimase dan invertase yang
berfungsi memecah polisakarida, sukrosa, dan fruktosa menjadi glukosa
(Ostergaard, 2000).
Dinding sel khamir
β-glukan
β-glukan + Kitin

9
β-glukan pada dinding sel khamir terbentuk dengan adanya metabolisme
glukosa yang diubah menjadi Glukosa-6-Fosfat, kemudian dengan adanya enzim
Phosphoglucomutase diperoleh Glukosa-1- Fosfat dan diubah lagi menjadi Uridin
difosfat glukosa (UDP-Glukosa) yang merupakan komponen penyusun dinding sel
khamir. Salah satu komponen penyusun dinding sel khamir tersebut adalah β-
glukan itu sendiri (Appeldoorn, 2002).
Menurut Cheeseman & Malcom (2000), sifat fisika dan kimia yang dimiliki
senyawa β-glukan yaitu di alam dalam bentuk senyawa berwarna putih berupa
gumpalan besar dan tidak berbentuk kristal, tidak mempunyai rasa manis, tidak
larut dalam air netral dan dapat dipisahkan dengan mudah dalam larutan alkali, bila
dicampur dengan air maka akan membentuk larutan koloid, dan berbentuk gel pada
suhu 54˚C.
β-glukan merupakan imunostimultan yang berasal dari dinding sel khamir
S. cereviciae atau dinding sel tanaman tinggi yang memiliki berat molekul tinggi
dan bercabang-cabang dan mengandung lebih dari 250.000 glukosa (Robinson,
1995). β-1,3-glukan memiliki derajat polimerisasi sebesar 1500 dengan berat
molekul 240.000 dan panjang serat sekitar 660nm. (Lipke dan Ovalle, 1998)
2.3. Produksi β-Glukan dari Aspergillus oryzae
Kapang adalah golongan organisme yang tubuh vegetatifnya (struktur
somatisnya) berupa thalus, dan tidak mempunyai klorofil. Kapang memerlukan
senyawa organik untuk nutrisinya (Pelczar dan Chan, 2005). Kapang tersusun dari
benang-benang sel panjang yang disebut hifa. (Volk dan Wheeler, 1998). Kapang
memerlukan kelembaban yang tinggi, persediaan bahan organik yang mati dan

10
mengalami pembusukan. Kapang tumbuh baik dalam lingkungan yang
mengandung banyak gula dan dengan kondisi asam yang tidak menguntungkan
bagi bakteri. Mekanisme reproduksi kapang disebut pembentukan spora. Spora
dapat bersifat seksual dan aseksual. Pada kapang, spora analog dengan biji yang
perbanyak oleh tumbuhan induk dan mampu berkembang menjadi individu baru.
Apabila spora melepaskan diri dari tumbuhan induk dalam kondisi yang
menguntungkan, benang-benang kapang baru akan tumbuh, spora bersemi dan
benang memanjang dengan pembelahan biner (Volk dan Wheeler, 1998).
Kapang genus Aspergillus sp. adalah salah satu jenis kapang yang dapat
digunakan dalam produksi β-glukan. Aspergillus sp terdapat dimana-mana sebagai
saprofit. Koloni yang mudah menghasilkan spora wamanya menjadi cokelat
kekuningan, kehijau-hijauan atau kehitam-hitaman dan miselium yang semula
bewama putih sudah tidak tampak lagi (Dwidjoseputro, 1998). Gambar 4
menunjukkan genus Aspergillus terdiri dari miselium dan konidia.
Aspergillus digunakan dalam fermentasi makanan. Kelompok Aspergillus
wenti, Aspergillus oryzae termasuk spesies yang penting dalam fermentasi
beberapa makanan tradisional dan untuk memproduksi enzim. Aspergillus oryzae
digunakan dalam fermentasi tahap pertama dalam pembuatan kecap dan tauco.
Gambar 4. Genus Aspergillus
(Sumber : Dumanov, 2006)

11
Aspergillus oryzae juga merupakan jamur yang telah diakui sebagai GRAS
(Generally Recognized As Safe). A. Oryzae tidak umum digunakan untuk produksi
β-glukan, namun spesies lain yang dipakai yaitu Aspergillus fumigatus dan
Aspergillus niger. Akira (2017) mengatakan komponen dinding sel daripada
Aspergillus fumigatus adalah polisakarida(setidaknya 90%) dan protein.
Gambar 5 menunjukkan bahwa dalam dinding sel miselium dan konidia
pada Aspergillus mengandung polisakarida yang terdiri dari rantai linear β-(1-3)-
glukan (20-35%) bercabang dengan rantai β-(1-6)-glukan (4%) ; rantai linear β-(1-
3/1-4)-glukan (10%) ; α(1-3)-glukan (45-56%); kitin dan galaktomannan (20-25%).
yang mengandung galaktomannan dan kitin. Dinding sel Aspergillus fumigatus
adalah garis pertahanan utama jamur untuk beradaptasi ke lingkungan yang tidak
mendukung, menyediakan integritas struktural dan fisik perlindungan ke dinding
sel.

12
2.4. Produksi β-Glukan dari Xanthomonas campestris
Bakteri mempunyai kemampuan untuk memproduksi bermacam jenis
polisakarida seperti kapsul dan produk ekstraselluler. Polisakarida umumnya
diproduksi sebagai respons terhadap tekanan lingkungan. Polisakarida yang berasal
dari bakteri dapat mengandung unit monosakarida yang terulang (homopolisakarida)
atau sejumlah monosakarida yang berbeda (heteropolisakarida). Sebagian besar
polisakarida ini memiliki asal bakteri dan banyak digunakan untuk makanan dan
aplikasi industri karena sifat rheological dan gel nya (1). Bakteri eksopolisakarida
asdasdasd
asdasdasd
Dinding sel Konidia
Dinding sel Miselium
β 1,3/1,4-glukan
Kitin
β 1,3-glukan
Galaktosaaminogalaktan
α 1,3 glukan
Galaktomannan Kitin oligosakarida
Membran
Plasma
β-(1,3)-glukan
sintase α-(1,3)-glukan
sintase
Kitin
Sintase
β-(1,3)-glukan
sintase
Membran
Plasma Kitin
Sintase α-(1,3)-glukan
sintase
Gambar 5. Struktur dinding sel Aspergillus Fumigatus
Sumber : Beauvais (2014)

13
(EPS) xanthan, curdlan dan gellan telah disetujui oleh Food and Drug
Administration (FDA). Menurut Zeković et al. (2005) polisakarida ini terdapat β-
(1-3)-glukan, β-(1-2)-glukan dan sellulosa. Xanthan atau Xanthan Gum merupakan
polisakarida yang memiliki nilai jual tinggi dan manfaat sebagai zat pengental
makanan serta memiliki kemampuan antioksidan yang baik (Tran, 2017) . Xanthan
Gum merupakan polisakarida yang berasal dari bakteri hasil diisolasi dari annual
bluegrass (Poa annua L.) yang berasal dari Jepang (Karen, 2014) yaitu
Xantomonas campertis (X. campertis). X. campertis merupakan bakteri gram
negatif yang diartikan memiliki sistem membran ganda dimana plasmanya
diselimuti oleh membran luar permeabel dan peptidoglikan terletak diantara
membran plasma dan membran luar (Singleton, 2006).
Gambar 6 menunjukkan X. campertis dapat menghasilkan polisakarida
berupa siklik β-glukan yang unik dan mengandung 16 residu Glukuronosil (Glc p)
15 di antaranya β-berikatan dengan C-2 dari residu berikutnya dan satu diantaranya
α-berikatan dengan C-6 dari residu berikutnya. Penelitian terhadap produksi β-(1-
2) glukan dari X. campertis masih kurang, oleh karena itu diperlukan kajian lebih
lanjut untuk X. campertis.
Gambar 6. Struktur kimia siklik β-glukan yang dihasilkan oleh X. campertis
Sumber: York (1995)

14
2.5. Produksi β-Glukan dari Bacillus natto
Bacillus natto (B.natto) merupakan bakteri hasil isolasi dari natto yang
berasal dari Jepang (Schallmey, 2004). B.natto merupakan bakteri gram positif
yang diartikan memiliki komponen peptidoglikan yang tebal pada bagian dinding
sel (Mahajan, 2010). B.natto secara umum digunakan untuk menghasilkan produk
fermentasi mikrobial, menurut Liu (2005), B.natto dapat pula dimanfaatkan untuk
menghasilkan enzim dalam lingkup industrial.
Strain Bacillus sp telah dikenal untuk membuat enzim β-glukanase, dimana
enzim ini berfungsi untuk menghidrolisis β-glukan. Berdasarkan mekanisme
kerjanya, enzim β-1,3-glukanase diklasifikasikanmenjadi 2, yaitu: enzim β-1,3-
eksoglukanase (β-1,3-glukan glukanohidrolase EC 3.2.1.58), dan enzim β-1,3-
endoglukanase (β-1,3-glukan glukanohidrolase EC 3.2.1.6 atau EC 3.2.1.39) (de la
Cruz et al.1995). Enzim β-1,3-endo-glukanase bekerja secara random
menghidrolisis rantai β-1,3-glukan menjadi 2 sampai 6 unit glukosa, sedangkan
enzim β-1,3-eksoglukanase menghidrolisis rantai β-1,3-glukan dengan melepaskan
monomer glukosa dari sisi nonreduktif (Pitson et al.1993). Ikatan yang dilepas pada
saat hidrolisis adalah ikatan β-1,3-glukosidik. Mayoritas enzim ini ialah jenis
endoglukanase (Aono et al. 1995). Enzim ini dapat berperan sebagai agen
biokontrol; digunakan untuk mengkarakterisasi β-glukan yang berperan sebagai
immunomodulator antikanker; meningkatkan nutrisi pakan ternak; berperan dalam
pembuatan bir pada proses penggilingan, fermentasi dan penyimpanan; dan
berperan dalam sintesis oligosakarida (Pitson et al. 1993). Selain daripada enzim β-
glukanase, penelitian Gummadi (2005) telah memperoleh β-glukan dari genus
Bacillus Subtilis meskipun dalam jumlah sedikit. Namun hasil metabolit

15
polisakarida dari Bacillus Natto belum diketahui saat ini. Oleh karena itu
diharapkan polisakarida yang dihasilkan oleh B. natto mengandung β-glukan.
2.6. Biosintesis β-Glukan
Lipke dan Ovalle (1998) mengatakan β-1,3-glukan yang diperoleh dari
fungi disintesis melalui reaksi enzimatis yang kompleks. β-Glukan sintetase yang
berada di membran plasma mengkatalisis sintesis β-glukan dari UDP-glukosa
menurut reaksi sebagai berikut (Shematek et al., 1980; Cabib et al., 2001) :
UDP-glukosa + (β-(1,3)-glukosa)n → UDP + (β-(1,3)-glukosa)n+1
Glukosa menjadi substrat yang sangat dimanfaatkan oleh Saccharomyces
cerevisiae. Glukosa akan memasuki sel Saccharomyces cerevisiae melalui transport
aktif. Sel kemudian akan memulai sintesis polisakarida dan menghasilkan β-glukan
(Priest, 1996).
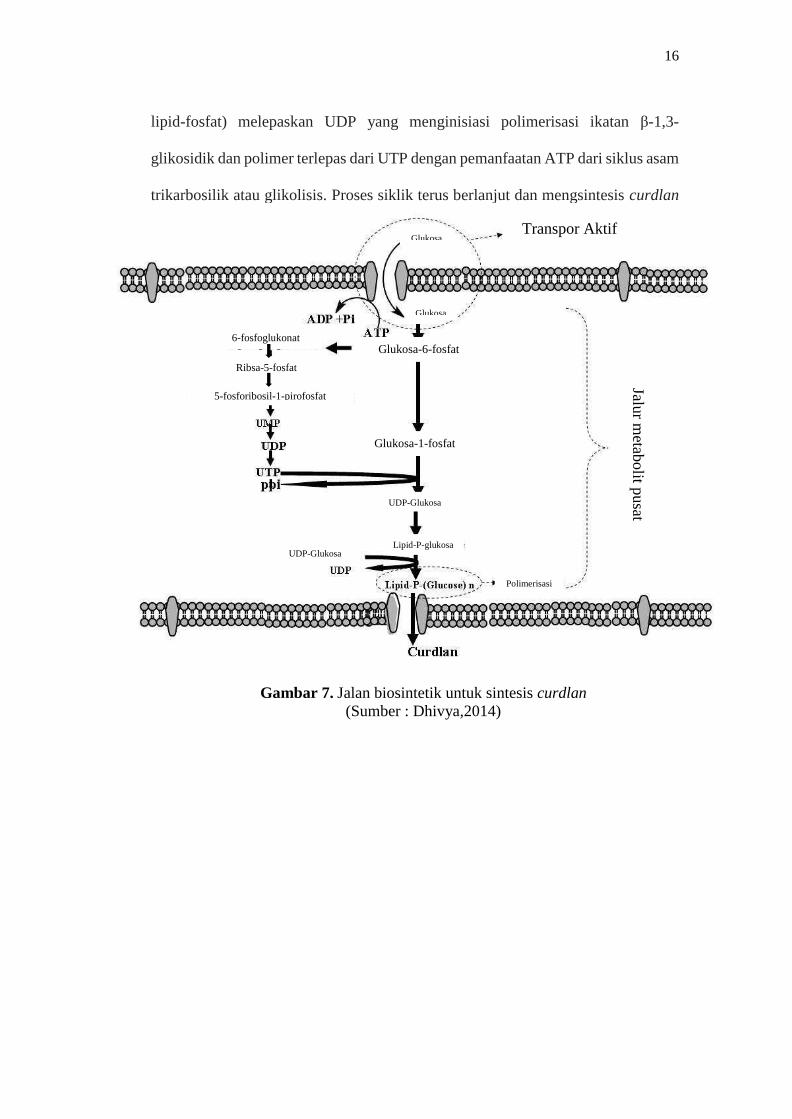
β-glukan yang dihasilkan dari bakteri Agrobacterium dikenal sebagai
curdlan. Biosintesis curdlan terdiri dari 3 tahap yaitu penyerapan substrat,
metabolisme dan polimerisasi. Substrat yang sebagian besar glukosa masuk ke
sitoplasma sel melalui transpor aktif, di tahap metabolisme susbstrat dikatabolisme
untuk membentuk metabolit primer dan prekursor untuk sintesis Extracellular
polymeric substance (EPS). Glikolisis glukosa dimulai dari fosforilisasi dengan
enzim heksokinase dalam pengeluaran ATP untuk membentuk glukosa-6-fosfat,
lalu dikonversi menjadi glukosa-1-fosfat yang terjadi di fosfoglukomutase.
Kemudian kunci prekursor (UDP-glukosa) membentuk melalui katalis UDP-
glukosa fosforilase dari Uridin Trifosfat (UTP). Glukosil-1-fosfat dari UDP-
glukosa mengandung D-glukosa yang terikat dengan prekursor lipid (isoprenoid-
16
lipid-fosfat) melepaskan UDP yang menginisiasi polimerisasi ikatan β-1,3-
glikosidik dan polimer terlepas dari UTP dengan pemanfaatan ATP dari siklus asam
trikarbosilik atau glikolisis. Proses siklik terus berlanjut dan mengsintesis curdlan
(Dhivya,2014). Biosintesis dari
curdlan dapat dilihat pada gambar 7.
Jalur m
etabolit p
usat
Transpor Aktif
Glukosa-6-fosfat
Glukosa-1-fosfat
UDP-Glukosa
Lipid-P-glukosa
Polimerisasi
Curdlan
6-fosfoglukonat
5-fosforibosil-1-pirofosfat
Ribsa-5-fosfat
Gambar 7. Jalan biosintetik untuk sintesis curdlan
(Sumber : Dhivya,2014)
Polimerisasi
Glukosa
Glukosa
UDP-Glukosa