Human Pol ζ purified with accessory subunits is active in ...Human Pol ζ purified with accessory...
6
Human Pol ζ purified with accessory subunits is active in translesion DNA synthesis and complements Pol η in cisplatin bypass Young-Sam Lee 1 , Mark T. Gregory, and Wei Yang 2 Laboratory of Molecular Biology, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD 20892 Contributed by Wei Yang, December 27, 2013 (sent for review December 5, 2013) DNA polymerase ζ (Pol ζ) is a eukaryotic B-family DNA polymerase that specializes in translesion synthesis and is essential for normal embryogenesis. At a minimum, Pol ζ consists of a catalytic subunit Rev3 and an accessory subunit Rev7. Mammalian Rev3 contains >3,000 residues and is twice as large as the yeast homolog. To date, no vertebrate Pol ζ has been purified for biochemical char- acterization. Here we report purification of a series of human Rev3 deletion constructs expressed in HEK293 cells and identification of a minimally catalytically active human Pol ζ variant. With a tagged form of an active Pol ζ variant, we isolated two additional acces- sory subunits of human Pol ζ, PolD2 and PolD3. The purified four- subunit Pol ζ4 (Rev3–Rev7–PolD2–PolD3) is much more efficient and more processive at bypassing a 1,2-intrastrand d(GpG)-cisplatin cross-link than the two-subunit Pol ζ2 (Rev3–Rev7). We show that complete bypass of cisplatin lesions requires Pol η to insert dCTP opposite the 3′ guanine and Pol ζ4 to extend the primers. TLS | REV3L | MAD2L2 | processivity | two-polymerase lesion bypass D NA polymerase ζ (Pol ζ), composed of the catalytic Rev3 and accessary Rev7 subunits, is an error-prone DNA translesion polymerase that causes both spontaneous and DNA damage-induced mutagenesis (1, 2). More than two-thirds of the 1,504 residues in yeast Rev3 share sequence homology with all B-family DNA polymerases, including Pols α, δ, and e, which are responsible for the bulk of high-fidelity genomic replication in eukaryotes (3). Unlike the typical B-family polymerases, Pol ζ lacks an intrinsic 3′–5′ exonuclease activity and thus has no proofreading function (2). Human homologs of REV3 (REV3L) and REV7 (MAD2L2; hereafter referred to as REV7) genes were identified shortly after yeast Pol ζ was characterized. Human Rev3 contains 3,130 residues and is twice as large as the yeast counterpart (4). Human and yeast Rev7 are homologous (5) and bear sequence similarity to the mitotic checkpoint proteins Mad2 (6). Unlike Saccharomyces cerevisiae REV3 and REV7 genes, which are nonessential and whose knockout leads only to a de- creased rate of damage-induced mutagenesis (7, 8), Rev3l knock- out in mice is embryonic-lethal (9), and mouse Rev3l −/− embryonic stem cells are not viable (10, 11). Human and mouse cell cultures obtained from conditional Rev3l knockout show genome instability and growth defects without an external challenge of DNA damage (12–14). DNA pol ζ is apparently essential for normal cell pro- liferation and embryogenesis in mammals. Translesion synthesis (TLS) and DNA-damage-induced mu- tagenesis are the best-characterized functions of Pol ζ. Absence of the yeast REV3 gene leads to sensitivity to UV light and intrastrand and interstrand cross-linking agents (2, 15). DNA Pol ζ has been shown to induce multiple base substitutions as well as more complex mutations in yeast (7, 16, 17) and may contribute to hypermutation in Ig genes in mammals (18, 19). The TLS function of DNA Pol ζ has been implicated in its role of medi- ating resistance to platinum-based chemotherapies (20–22). Owing to the conservation of B-family DNA polymerases, a distorted DNA template base is unlikely to be accommodated in the active site of DNA Pol ζ. In fact, yeast DNA Pol ζ is unable to insert a nucleotide opposite either a cis–syn thymine or a 6-4 photo- product (23). Genetic data indicate that a complete lesion bypass event may require two TLS DNA polymerases (24)—one for nucleotide incorporation opposite a lesion (insertion step) and the other for the subsequent primer extension (extension step). The insertion step of TLS is often accomplished by a Y-family poly- merase, whose active site is uncommonly large, solvent-exposed, and flexible (25). Studies of another B-family TLS DNA polymerase from Escherichia coli (Pol II) show that it efficiently extends a DNA primer after a lesion by looping out the damaged DNA template strand, leading to frameshift and mixed-type mutations (26). In budding yeast, REV3 has been shown to be epistatic with POL32, a subunit of DNA Pol δ. Inactivating either REV3 or POL32 leads to reduced spontaneous mutagenesis (27–29). As with all eukaryotic B-family DNA polymerases, Rev3 contains a Cys-rich C-terminal domain (CTD) (30–33), which forms a zinc-finger domain followed by a [4Fe–4S] cluster (34). In Pol α, δ, and e, each CTD interacts with its specific accessary subunits (32, 35). Recently, three groups have independently shown that the [4Fe–4S] cluster of yeast Rev3 interacts with Pol31 and Pol32 subunit (36), thus forming an stoichiometric four-subunit Pol ζ (Pol ζ4; Rev3–Rev7–Pol31–Pol32) (23, 37, 38). Baranovskiy et al. further showed that the CTDs of human Pol ζ and δ share the same accessary subunits p50 and p66, homologs of yeast Pol31 and Pol32, respectively (37). The interaction between yeast Rev3 and Pol31 is reported to be direct, and Pol32 is essential to stabilize Pol31 and, furthermore, via its interactions with pro- liferating cell nuclear antigen (PCNA), recruits and activates Pol ζ to carry out TLS (38). The catalytic activity of yeast Pol ζ is im- proved by the presence of Pol31 and Pol32 (23, 38). Significance Although human DNA polymerase ζ (Pol ζ) is essential for DNA replication and cell proliferation, difficulties purifying active Pol ζ have hindered its biochemical characterization. We report here the first purification of an active form of human Pol ζ holoenzyme composed of Rev3, Rev7, PolD2, and PolD3, which opens up the possibility for detailed biochemical and structural studies of this essential enzyme. Based on genetic data, it has been postulated that two specialized DNA polymerases are needed for successful translesion synthesis. We show here that human Pol η inserts a nucleotide opposite the lesion, followed by Pol ζ extending the DNA primer; thus, the two complement each other to fully bypass the cisplatin cross-link. Author contributions: Y.-S.L. and W.Y. designed research; Y.-S.L. and M.T.G. performed re- search; Y.-S.L., M.T.G., and W.Y. analyzed data; and Y.-S.L. and W.Y. wrote the paper. The authors declare no conflict of interest. See Commentary on page 2864. 1 Present address: Well-Aging Research Center, Samsung Advanced Institute of Technol- ogy, Samsung Electronics, Yongin, Gyeonggido, Korea. E-mail: [email protected]. 2 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1324001111/-/DCSupplemental. 2954–2959 | PNAS | February 25, 2014 | vol. 111 | no. 8 www.pnas.org/cgi/doi/10.1073/pnas.1324001111 Downloaded by guest on March 29, 2020
Transcript of Human Pol ζ purified with accessory subunits is active in ...Human Pol ζ purified with accessory...

Human Pol ζ purified with accessory subunits is activein translesion DNA synthesis and complements Pol η incisplatin bypassYoung-Sam Lee1, Mark T. Gregory, and Wei Yang2
Laboratory of Molecular Biology, National Institute of Diabetes and Digestive and Kidney Diseases, National Institutes of Health, Bethesda, MD 20892
Contributed by Wei Yang, December 27, 2013 (sent for review December 5, 2013)
DNA polymerase ζ (Pol ζ) is a eukaryotic B-family DNA polymerasethat specializes in translesion synthesis and is essential for normalembryogenesis. At a minimum, Pol ζ consists of a catalytic subunitRev3 and an accessory subunit Rev7. Mammalian Rev3 contains>3,000 residues and is twice as large as the yeast homolog. Todate, no vertebrate Pol ζ has been purified for biochemical char-acterization. Here we report purification of a series of human Rev3deletion constructs expressed in HEK293 cells and identification ofa minimally catalytically active human Pol ζ variant. With a taggedform of an active Pol ζ variant, we isolated two additional acces-sory subunits of human Pol ζ, PolD2 and PolD3. The purified four-subunit Pol ζ4 (Rev3–Rev7–PolD2–PolD3) is much more efficientand more processive at bypassing a 1,2-intrastrand d(GpG)-cisplatincross-link than the two-subunit Pol ζ2 (Rev3–Rev7). We show thatcomplete bypass of cisplatin lesions requires Pol η to insert dCTPopposite the 3′ guanine and Pol ζ4 to extend the primers.
TLS | REV3L | MAD2L2 | processivity | two-polymerase lesion bypass
DNA polymerase ζ (Pol ζ), composed of the catalytic Rev3and accessary Rev7 subunits, is an error-prone DNA
translesion polymerase that causes both spontaneous and DNAdamage-induced mutagenesis (1, 2). More than two-thirds of the1,504 residues in yeast Rev3 share sequence homology with allB-family DNA polymerases, including Pols α, δ, and e, which areresponsible for the bulk of high-fidelity genomic replication ineukaryotes (3). Unlike the typical B-family polymerases, Pol ζlacks an intrinsic 3′–5′ exonuclease activity and thus has noproofreading function (2). Human homologs of REV3 (REV3L)and REV7 (MAD2L2; hereafter referred to as REV7) genes wereidentified shortly after yeast Pol ζ was characterized. HumanRev3 contains 3,130 residues and is twice as large as the yeastcounterpart (4). Human and yeast Rev7 are homologous (5) andbear sequence similarity to the mitotic checkpoint proteins Mad2(6). Unlike Saccharomyces cerevisiae REV3 and REV7 genes,which are nonessential and whose knockout leads only to a de-creased rate of damage-induced mutagenesis (7, 8), Rev3l knock-out in mice is embryonic-lethal (9), and mouse Rev3l−/− embryonicstem cells are not viable (10, 11). Human and mouse cell culturesobtained from conditional Rev3l knockout show genome instabilityand growth defects without an external challenge of DNA damage(12–14). DNA pol ζ is apparently essential for normal cell pro-liferation and embryogenesis in mammals.Translesion synthesis (TLS) and DNA-damage-induced mu-
tagenesis are the best-characterized functions of Pol ζ. Absenceof the yeast REV3 gene leads to sensitivity to UV light andintrastrand and interstrand cross-linking agents (2, 15). DNA Polζ has been shown to induce multiple base substitutions as well asmore complex mutations in yeast (7, 16, 17) and may contributeto hypermutation in Ig genes in mammals (18, 19). The TLSfunction of DNA Pol ζ has been implicated in its role of medi-ating resistance to platinum-based chemotherapies (20–22). Owingto the conservation of B-family DNA polymerases, a distortedDNA template base is unlikely to be accommodated in the activesite of DNA Pol ζ. In fact, yeast DNA Pol ζ is unable to insert
a nucleotide opposite either a cis–syn thymine or a 6-4 photo-product (23). Genetic data indicate that a complete lesion bypassevent may require two TLS DNA polymerases (24)—one fornucleotide incorporation opposite a lesion (insertion step) and theother for the subsequent primer extension (extension step). Theinsertion step of TLS is often accomplished by a Y-family poly-merase, whose active site is uncommonly large, solvent-exposed,and flexible (25). Studies of another B-family TLS DNA polymerasefrom Escherichia coli (Pol II) show that it efficiently extends a DNAprimer after a lesion by looping out the damaged DNA templatestrand, leading to frameshift and mixed-type mutations (26).In budding yeast, REV3 has been shown to be epistatic with
POL32, a subunit of DNA Pol δ. Inactivating either REV3 orPOL32 leads to reduced spontaneous mutagenesis (27–29). Aswith all eukaryotic B-family DNA polymerases, Rev3 containsa Cys-rich C-terminal domain (CTD) (30–33), which forms azinc-finger domain followed by a [4Fe–4S] cluster (34). In Pol α,δ, and e, each CTD interacts with its specific accessary subunits(32, 35). Recently, three groups have independently shown thatthe [4Fe–4S] cluster of yeast Rev3 interacts with Pol31 and Pol32subunit (36), thus forming an stoichiometric four-subunit Pol ζ(Pol ζ4; Rev3–Rev7–Pol31–Pol32) (23, 37, 38). Baranovskiy et al.further showed that the CTDs of human Pol ζ and δ share the sameaccessary subunits p50 and p66, homologs of yeast Pol31 andPol32, respectively (37). The interaction between yeast Rev3and Pol31 is reported to be direct, and Pol32 is essential tostabilize Pol31 and, furthermore, via its interactions with pro-liferating cell nuclear antigen (PCNA), recruits and activates Pol ζto carry out TLS (38). The catalytic activity of yeast Pol ζ is im-proved by the presence of Pol31 and Pol32 (23, 38).
Significance
Although human DNA polymerase ζ (Pol ζ) is essential for DNAreplication and cell proliferation, difficulties purifying activePol ζ have hindered its biochemical characterization. We reporthere the first purification of an active form of human Pol ζholoenzyme composed of Rev3, Rev7, PolD2, and PolD3, whichopens up the possibility for detailed biochemical and structuralstudies of this essential enzyme. Based on genetic data, it hasbeen postulated that two specialized DNA polymerases areneeded for successful translesion synthesis. We show here thathuman Pol η inserts a nucleotide opposite the lesion, followedby Pol ζ extending the DNA primer; thus, the two complementeach other to fully bypass the cisplatin cross-link.
Author contributions: Y.-S.L. and W.Y. designed research; Y.-S.L. and M.T.G. performed re-search; Y.-S.L., M.T.G., and W.Y. analyzed data; and Y.-S.L. and W.Y. wrote the paper.
The authors declare no conflict of interest.
See Commentary on page 2864.1Present address: Well-Aging Research Center, Samsung Advanced Institute of Technol-ogy, Samsung Electronics, Yongin, Gyeonggido, Korea. E-mail: [email protected].
2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1324001111/-/DCSupplemental.
2954–2959 | PNAS | February 25, 2014 | vol. 111 | no. 8 www.pnas.org/cgi/doi/10.1073/pnas.1324001111
Dow
nloa
ded
by g
uest
on
Mar
ch 2
9, 2
020
Purification and characterization of Pol ζ has so far beenlimited to the yeast protein. Perhaps because of its large size,mammalian Pol ζ has not been purified for biochemical char-acterization. To overcome this roadblock, we coexpressed hu-man REV3L and REV7 in mammalian cells in culture. Initially,very low expression level and heterogeneity was encountered, butthese problems were solved by targeted deletion of various in-ternal segments of human REV3L. We succeeded in purifying anactive two-subunit form of human Pol ζ (Pol ζ2). By differentialpull-down experiments using Pol ζ2 variants with and without theCTD of Rev3, we isolated two CTD-dependent Pol ζ accessarysubunits, PolD2 and PolD3. We report here purification of anactive form of human four-subunit Pol ζ4 and the collaborationof two TLS polymerases, Pol η and Pol ζ, in lesion bypass.
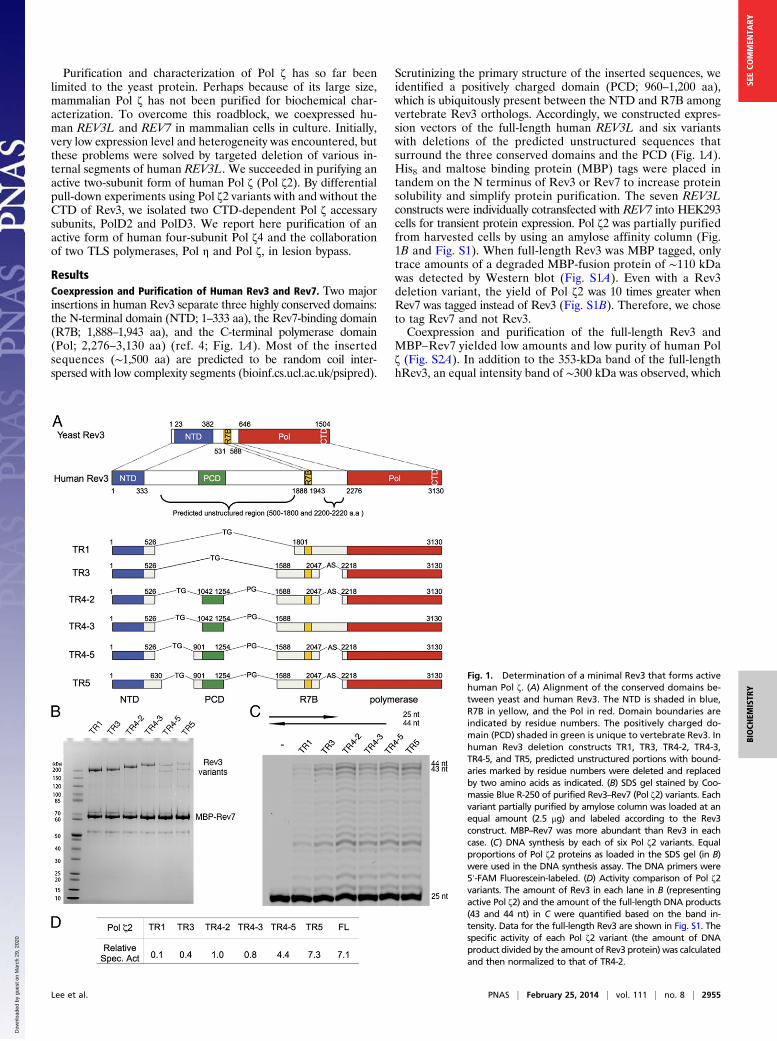
ResultsCoexpression and Purification of Human Rev3 and Rev7. Two majorinsertions in human Rev3 separate three highly conserved domains:the N-terminal domain (NTD; 1–333 aa), the Rev7-binding domain(R7B; 1,888–1,943 aa), and the C-terminal polymerase domain(Pol; 2,276–3,130 aa) (ref. 4; Fig. 1A). Most of the insertedsequences (∼1,500 aa) are predicted to be random coil inter-spersed with low complexity segments (bioinf.cs.ucl.ac.uk/psipred).
Scrutinizing the primary structure of the inserted sequences, weidentified a positively charged domain (PCD; 960–1,200 aa),which is ubiquitously present between the NTD and R7B amongvertebrate Rev3 orthologs. Accordingly, we constructed expres-sion vectors of the full-length human REV3L and six variantswith deletions of the predicted unstructured sequences thatsurround the three conserved domains and the PCD (Fig. 1A).His8 and maltose binding protein (MBP) tags were placed intandem on the N terminus of Rev3 or Rev7 to increase proteinsolubility and simplify protein purification. The seven REV3Lconstructs were individually cotransfected with REV7 into HEK293cells for transient protein expression. Pol ζ2 was partially purifiedfrom harvested cells by using an amylose affinity column (Fig.1B and Fig. S1). When full-length Rev3 was MBP tagged, onlytrace amounts of a degraded MBP-fusion protein of ∼110 kDawas detected by Western blot (Fig. S1A). Even with a Rev3deletion variant, the yield of Pol ζ2 was 10 times greater whenRev7 was tagged instead of Rev3 (Fig. S1B). Therefore, we choseto tag Rev7 and not Rev3.Coexpression and purification of the full-length Rev3 and
MBP–Rev7 yielded low amounts and low purity of human Polζ (Fig. S2A). In addition to the 353-kDa band of the full-lengthhRev3, an equal intensity band of ∼300 kDa was observed, which
Fig. 1. Determination of a minimal Rev3 that forms activehuman Pol ζ. (A) Alignment of the conserved domains be-tween yeast and human Rev3. The NTD is shaded in blue,R7B in yellow, and the Pol in red. Domain boundaries areindicated by residue numbers. The positively charged do-main (PCD) shaded in green is unique to vertebrate Rev3. Inhuman Rev3 deletion constructs TR1, TR3, TR4-2, TR4-3,TR4-5, and TR5, predicted unstructured portions with bound-aries marked by residue numbers were deleted and replacedby two amino acids as indicated. (B) SDS gel stained by Coo-massie Blue R-250 of purified Rev3–Rev7 (Pol ζ2) variants. Eachvariant partially purified by amylose column was loaded at anequal amount (2.5 μg) and labeled according to the Rev3construct. MBP–Rev7 was more abundant than Rev3 in eachcase. (C) DNA synthesis by each of six Pol ζ2 variants. Equalproportions of Pol ζ2 proteins as loaded in the SDS gel (in B)were used in the DNA synthesis assay. The DNA primers were5′-FAM Fluorescein-labeled. (D) Activity comparison of Pol ζ2variants. The amount of Rev3 in each lane in B (representingactive Pol ζ2) and the amount of the full-length DNA products(43 and 44 nt) in C were quantified based on the band in-tensity. Data for the full-length Rev3 are shown in Fig. S1. Thespecific activity of each Pol ζ2 variant (the amount of DNAproduct divided by the amount of Rev3 protein) was calculatedand then normalized to that of TR4-2.
Lee et al. PNAS | February 25, 2014 | vol. 111 | no. 8 | 2955
BIOCH
EMISTR
YSE
ECO
MMEN
TARY
Dow
nloa
ded
by g
uest
on
Mar
ch 2
9, 2
020

likely corresponds to a large fragment of Rev3 (see below).With a single internal deletion of residues 527–1,800 in Rev3(TR1), the yield of Pol ζ increased by ∼10-fold (Fig. 1 A andB). However, the specific polymerase activity of TR1 ona normal DNA template–primer pair was ∼2% that of full-length Pol ζ (Fig. 1 and Fig. S2). Inclusion of the positivelycharged patch (1,042–1,254 aa) and alteration of boundariessurrounding the R7B domain (TR3, TR4-2, and TR4-3)resulted in an approximately eightfold specific-activity increaseof TR4-2 relative to TR1 without reducing the protein yield(Fig. 1). Comparison among TR1, TR3, TR4-2, and TR4-3constructs suggested that inclusion of amino acids 1,588–1,800and deletion of amino acids 2,048–2,217 enhanced the proteinyield and specific activity. The most significant increase in thePol ζ-specific activity was achieved by the addition of residues900–1,041 (TR4-5 and TR5) (Fig. 1), which restored thespecific catalytic activity of TR5 (a total of 2,358 aa) to that ofthe full-length Rev3 (3,130 aa). However, this activity increasewas accompanied by a significant decrease in protein yield andthe appearance of a proteolyzed Rev3 band (Fig. 1B). The sizedifference between the expected Rev3—whether full-length,TR4-5, or TR5—and the proteolyzed bands on SDS gelsappeared to be constant. We therefore estimate that the cleav-age site is around residue 500 and suspect that the presence ofresidues 900–1,041 somehow alters the Rev3 structure to exposethis protease cleavage site. For the remaining Pol ζ studiesreported here, we focus on the TR4-2 variant (2,113 of 3,130 aatotal), which produces a single Rev3 band on SDS gels and anactive Pol ζ2 that has the same nucleotide preference and gen-erates the same DNA products as the full-length Rev3 (Fig. S2).
Identification of Human DNA Pol ζ Accessory Subunits. The con-served Cys-rich CTDs of B-family DNA polymerases α, δ, and eeach interact with unique accessory subunits. To determinewhether the CTD of human Rev3 (3,016–3,130 aa) also interactswith its own accessory proteins, we constructed a CTD-deletionclone of TR4-2 (TR4-2ΔCTD) and cotransfected expressionvectors of TR4-2 with or without CTD and His8–MBP–Rev7 intoHEK293 cells (Fig. 2A). To ensure that the amount of Rev3 andRev7 did not greatly exceed endogenous Pol ζ cofactors, wereduced the DNA amounts used for transfection. Without as-suming which proteins might interact with the CTD of Pol ζ, wefractionated the total HEK293 cell lysate with an amylose pull-down followed by a cation exchange column. The eluents weretreated with PreScission protease to separate Rev7 from theHis8–MBP tag and analyzed on a SDS gel with Coomassie bluestain (Fig. 2B). The presence of the Rev3 CTD resulted in twoadditional pull-down protein bands. Band 1 (∼65 kDa) wascompletely absent in the lane without the Rev3 CTD, andband 2 (∼50 kDa) ran very closely to a protein band (band x)present in the lane without the Rev3 CTD (Fig. 2B). With >55%of each protein band sequenced by mass spectrometry, bands 1and 2 were identified as human PolD3 (p66) and PolD2 (p50),homologs of yeast Pol32 and Pol31, respectively. Band x wasidentified to be α and β tubulins. The other protein band (band y)pulled down with or without the Rev3 CTD was ∼75 kDa andis too small to be Rev1, which is known to associate weaklywith Rev7 (39–42). Based on our prior experience with proteinexpression in HEK293 cells, band y is likely a protein chap-erone. We conclude that, like yeast Pol ζ, human Pol ζ is afour-subunit DNA polymerase and shares two accessory subunitswith DNA Pol δ.
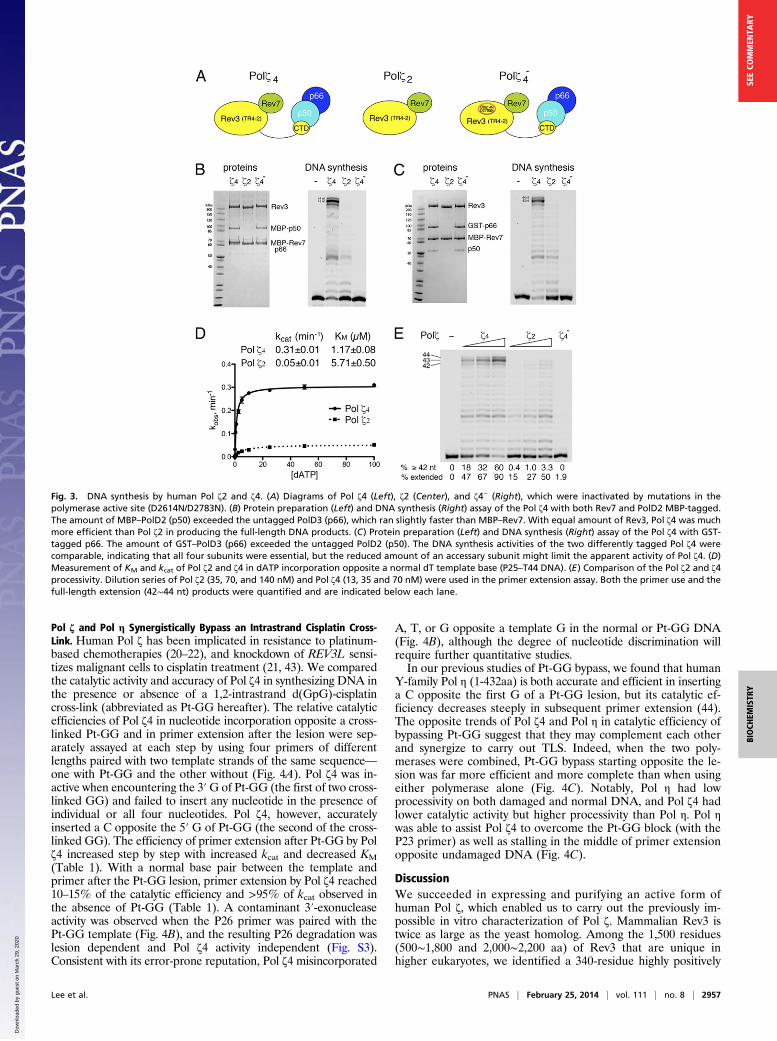
PolD2 and PolD3 Enhance the Catalytic Efficiency and Processivity ofPol ζ. To investigate how PolD2 and PolD3 influence the stabilityand catalytic activity of human Pol ζ, we prepared Pol ζ2 and Polζ4 by cotransfection of two or four relevant expression vectors ofTR4-2, His8–MBP–Rev7, PolD2 (p50 with or without an MBP
tag), and PolD3 (p66 with or without a GST tag) (Fig. 3 A–C)into HEK293 cells. To ensure that no endogenous PolD2 andPolD3 copurified with Pol ζ2, a TR4-2ΔCTD clone was used togenerate Pol ζ2. For the negative control in enzymatic assays, wegenerated and purified inactive Pol ζ4− protein by mutating twocatalytically essential aspartates in Rev3 to asparagines (D2614Nand D2783N). Differently tagged Pol ζ4 proteins (MBP-taggedp50 or GST-tagged p66) were purified by amylose or amyloseand glutathione affinity columns, followed by cation-exchangechromatography. Pol ζ2, ζ4, and ζ4− were equally well expressedand equally stable (Fig. 3). Interestingly, PolD2 (p50) or PolD3(p66) that was untagged—and thus might suffer loss during theaffinity purification—was present in less than stoichiometricamounts in the purified four-subunit complex Pol ζ4 (Fig. 3 Band C). The varying ratios of PolD2 and PolD3 in the human Polζ4 preparations when different subunits were tagged suggest thatPolD3 may interact with human Rev3 or Rev7 independent ofthe PolD2–CTD interaction (23, 37, 38).To compare the Pol ζ catalytic activities, equimolar concen-
trations of Pol ζ2, ζ4, and ζ4− proteins were tested in normalDNA synthesis assays (Fig. 3 B–D). Although negligible, residualDNA synthesis activity was detected with the Pol ζ4− protein(Fig. 3 B–E), which could be due to a trace contaminant withactive DNA polymerases—for example, Pol δ or Rev1 copur-ifying via interaction with PolD2 or Rev7. Even with PolD2 orPolD3 in substoichiometric amounts relative to Rev3 (TR4-2),Pol ζ4 exhibited higher kcat and lower KM than Pol ζ2 (Fig. 3D)and was nearly 30 times more efficient than Pol ζ2 in catalyzingDNA synthesis in the absence of a lesion.The presence of PolD2 and PolD3 accessory subunits also
increased the processivity of human Pol ζ4 on DNA substrates.Full-length DNA primer-extension products (42- to 44-nt bands)were prominent with Pol ζ4 but very weak with Pol ζ2 (Fig. 3 Band C). To semiquantitatively assess the processivity of Pol ζ4and Pol ζ2, the primer extension assay was performed witha range of concentrations of the two protein complexes, andprimer use and product distributions were then compared (Fig.3E). At the lowest concentration of Pol ζ4 and highest concen-tration of Pol ζ2, for which nearly 50% of DNA primers wereextended in each case, Pol ζ4 produced at least five times morefull-length DNA products than Pol ζ2. The increased processivitywas likely due to enhanced DNA binding by human Pol ζ4, whichmay also explain the reduced KM.
Fig. 2. Identification of human Pol ζ accessary subunits. (A) A diagram ofPol ζ and potential accessary subunits. (B) Pull-down of Rev3 accessary sub-units from HEK293 cells. The TR4-2 variants of Rev3 with or without CTDwere constructed and coexpressed with MBP–Rev7 as baits to pull down Polζ2-interacting proteins. Bands 1 and 2 were purified only in the presence ofthe CTD. Band 2 overlapped with band x, which was copurified with Rev3with or without the CTD. (C) Mass spectrometry analysis identified bands 1and 2 as PolD3 (p66) and PolD2 (p50), respectively, and band x as tubulins.
2956 | www.pnas.org/cgi/doi/10.1073/pnas.1324001111 Lee et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 2
9, 2
020
Pol ζ and Pol η Synergistically Bypass an Intrastrand Cisplatin Cross-Link. Human Pol ζ has been implicated in resistance to platinum-based chemotherapies (20–22), and knockdown of REV3L sensi-tizes malignant cells to cisplatin treatment (21, 43). We comparedthe catalytic activity and accuracy of Pol ζ4 in synthesizing DNA inthe presence or absence of a 1,2-intrastrand d(GpG)-cisplatincross-link (abbreviated as Pt-GG hereafter). The relative catalyticefficiencies of Pol ζ4 in nucleotide incorporation opposite a cross-linked Pt-GG and in primer extension after the lesion were sep-arately assayed at each step by using four primers of differentlengths paired with two template strands of the same sequence—one with Pt-GG and the other without (Fig. 4A). Pol ζ4 was in-active when encountering the 3′G of Pt-GG (the first of two cross-linked GG) and failed to insert any nucleotide in the presence ofindividual or all four nucleotides. Pol ζ4, however, accuratelyinserted a C opposite the 5′ G of Pt-GG (the second of the cross-linked GG). The efficiency of primer extension after Pt-GG by Polζ4 increased step by step with increased kcat and decreased KM(Table 1). With a normal base pair between the template andprimer after the Pt-GG lesion, primer extension by Pol ζ4 reached10–15% of the catalytic efficiency and >95% of kcat observed inthe absence of Pt-GG (Table 1). A contaminant 3′-exonucleaseactivity was observed when the P26 primer was paired with thePt-GG template (Fig. 4B), and the resulting P26 degradation waslesion dependent and Pol ζ4 activity independent (Fig. S3).Consistent with its error-prone reputation, Pol ζ4 misincorporated
A, T, or G opposite a template G in the normal or Pt-GG DNA(Fig. 4B), although the degree of nucleotide discrimination willrequire further quantitative studies.In our previous studies of Pt-GG bypass, we found that human
Y-family Pol η (1-432aa) is both accurate and efficient in insertinga C opposite the first G of a Pt-GG lesion, but its catalytic ef-ficiency decreases steeply in subsequent primer extension (44).The opposite trends of Pol ζ4 and Pol η in catalytic efficiency ofbypassing Pt-GG suggest that they may complement each otherand synergize to carry out TLS. Indeed, when the two poly-merases were combined, Pt-GG bypass starting opposite the le-sion was far more efficient and more complete than when usingeither polymerase alone (Fig. 4C). Notably, Pol η had lowprocessivity on both damaged and normal DNA, and Pol ζ4 hadlower catalytic activity but higher processivity than Pol η. Pol ηwas able to assist Pol ζ4 to overcome the Pt-GG block (with theP23 primer) as well as stalling in the middle of primer extensionopposite undamaged DNA (Fig. 4C).
DiscussionWe succeeded in expressing and purifying an active form ofhuman Pol ζ, which enabled us to carry out the previously im-possible in vitro characterization of Pol ζ. Mammalian Rev3 istwice as large as the yeast homolog. Among the 1,500 residues(500∼1,800 and 2,000∼2,200 aa) of Rev3 that are unique inhigher eukaryotes, we identified a 340-residue highly positively
Fig. 3. DNA synthesis by human Pol ζ2 and ζ4. (A) Diagrams of Pol ζ4 (Left), ζ2 (Center), and ζ4− (Right), which were inactivated by mutations in thepolymerase active site (D2614N/D2783N). (B) Protein preparation (Left) and DNA synthesis (Right) assay of the Pol ζ4 with both Rev7 and PolD2 MBP-tagged.The amount of MBP–PolD2 (p50) exceeded the untagged PolD3 (p66), which ran slightly faster than MBP–Rev7. With equal amount of Rev3, Pol ζ4 was muchmore efficient than Pol ζ2 in producing the full-length DNA products. (C) Protein preparation (Left) and DNA synthesis (Right) assay of the Pol ζ4 with GST-tagged p66. The amount of GST–PolD3 (p66) exceeded the untagged PolD2 (p50). The DNA synthesis activities of the two differently tagged Pol ζ4 werecomparable, indicating that all four subunits were essential, but the reduced amount of an accessary subunit might limit the apparent activity of Pol ζ4. (D)Measurement of KM and kcat of Pol ζ2 and ζ4 in dATP incorporation opposite a normal dT template base (P25–T44 DNA). (E) Comparison of the Pol ζ2 and ζ4processivity. Dilution series of Pol ζ2 (35, 70, and 140 nM) and Pol ζ4 (13, 35 and 70 nM) were used in the primer extension assay. Both the primer use and thefull-length extension (42∼44 nt) products were quantified and are indicated below each lane.
Lee et al. PNAS | February 25, 2014 | vol. 111 | no. 8 | 2957
BIOCH
EMISTR
YSE
ECO
MMEN
TARY
Dow
nloa
ded
by g
uest
on
Mar
ch 2
9, 2
020

charged domain (residues 960–1,200), which contains low-com-plexity sequence dominated by Lys (17.8%), Ser (10%), and Arg(8.3%). Its inclusion enhances the specific activity of DNA syn-thesis by Pol ζ. The exact amino acid sequence of PCD is nothighly conserved, but the Lys- and Arg-rich character is con-served from invertebrates to mammals, suggesting that charge–charge interactions may lead to increased DNA binding by Pol ζ.Our deletion analysis also revealed that the region between R7B(Rev7 binding) and the polymerase catalytic domain (residues2,048–2,217) negatively impacts the polymerase activity of Pol ζ2(Fig. 1). The remaining 770 residues, which can be removed fromhuman Rev3 without reducing the polymerase activity of Pol ζ2as shown by our TR-5 construct, may play a role in interactionswith other factors to regulate the activity of Pol ζ and its re-cruitment to a replication fork.Based on studies of yeast Pol ζ and the CTD of human Rev3, it
is not surprising that human PolD2 and PolD3, the equivalents ofyeast Pol31 and Pol32, are accessory subunits of human Pol ζ.Our pull-down experiments using whole-cell lysates of HEK293cells, however, were conducted without the assumption ofPolD2 and PolD3 involvement. Only PolD2 and PolD3 co-purified with Rev3 (TR4-2) and Rev7 after stringent washingwith a 1.0 M NaCl solution, and Rev1, a known cofactor of Polζ via interactions with Rev7 (41, 42, 45), was not detected(Fig. 2B). PolD2 and PolD3 must have stronger interactionswith Rev3 and Rev7 than Rev1. In yeast, Pol31 directlyinteracts with the CTD of Pol ζ and Pol δ, and Pol32 interacts
only with Pol31 (38). However, the recent EM study of yeastPol ζ4 placed Pol32 proximal to Rev7 (46). Potential inter-actions between human PolD3 (yeast Pol32 homolog) andRev7 may explain our observation that PolD3 remains a com-ponent of Pol ζ4, even when PolD3 is in molar excess of PolD2(Fig. 3C). Unless PolD2 and PolD3 are limiting in vivo, bothPol δ and ζ may exist as a four-subunit complex without com-peting for the accessory units.In human Pol ζ4, the presence of PolD2 and PolD3, even at
submolar ratios to Rev3 (TR4-2), dramatically increased theefficiency (kcat/KM) and processivity of DNA synthesis (Fig. 3D).The effect of decreased KM and increased processivity suggeststhat PolD2 and PolD3 significantly enhance association of Polζ with DNA substrates. The cocrystal structure of humanPolD2 and the NTD of PolD3 has been determined (35) andshows a primarily negatively charged molecular surface with-out obvious features for DNA binding. It is possible that theunstructured, but positively charged, region in PolD3 that isnot included in the crystal structure may play a crucial role inenhancing the Pol ζ catalytic activity and processivity. Incontrast, inclusion of two accessory subunits in yeast Pol ζ4primarily increases the kcat by twofold to threefold without anappreciable change of KM or processivity (23). This differencebetween yeast and human Pol ζ4 may result from low similaritybetween human PolD3 and yeast Pol32 accessory subunits.Pol ζ and Pol η are synergistic in improving the efficiency and
processivity of DNA synthesis both in the absence and presence
Fig. 4. Pol ζ4 and Pol η carry out TLS synergistically bypassing a Pt-GG lesion. (A) Diagrams of DNA primer and template used in the assay. The four primersallow separate analyses of nucleotide insertions opposite the Pt-GG dimer and primer extension 1 or 2 nt beyond the lesion. The template strands containeither cisplatin cross-linked Pt-GG (in red) or undamaged DNA of the same sequence. (B) Primer extension of undamaged and Pt-GG DNA template by Pol ζ4in the presence of each (A, T, G, or C) or all four nucleotides (labeled as 4). Percentage of each primer extended in the presence of a correct dNTP or mixedfour nucleotides was quantified and is indicated. (Upper) Insertion step. (Lower) Extension step. (C) Pol ζ4 and Pol η were synergistic in DNA synthesis.(Upper) With normal DNA template, Pol η was efficient in nucleotide incorporation but had poor processivity. (Lower) In the presence of a Pt-GG lesion,Pol ζ4 failed to insert opposite the 3′ G of the Pt-GG, whereas Pol η failed to extend after the Pt-GG lesion. Together, Pol ζ4 and Pol η resulted in 10% offull-length product.
Table 1. Catalytic efficiency of Pol ζ4 in normal DNA synthesis and Pt-GG bypass
Primer TemplateIncomingnucleotide kcat, min−1 KM, μM kcat/KM, M
−1·min−1Relative efficiency,
% of kcat/Km
Relative rate,% of kcat
P24 (second insertion) Undamaged dCTP 0.22 ± 0.01 1.50 ± 0.12 144.7 x 103 100 100Pt-GG 0.09 ± 0.01 55.8 ± 10.4 1.7 x 103 1.2 43.3
P25 (first extension) Undamaged dATP 0.31 ± 0.01 1.17 ± 0.08 261.5 x 103 100 100Pt-GG 0.15 ± 0.01 35.6 ± 4.2 4.1 x 103 1.6 47.7
P26 (second extension) Undamaged dGTP 0.22 ± 0.01 0.82 ± 0.09 268.3 x 103 100 100Pt-GG 0.21 ± 0.01 8.80 ± 0.97 24.1 x 103 9.0 96.4
2958 | www.pnas.org/cgi/doi/10.1073/pnas.1324001111 Lee et al.
Dow
nloa
ded
by g
uest
on
Mar
ch 2
9, 2
020

of a DNA lesion (Fig. 4C). In the absence of direct interactionsbetween Pol ζ and Pol η, the synergy most likely takes place bypassing DNA substrate between the two polymerases. Pol η andPol ζ appear to complement each other in catalytic efficiencyand processivity in both TLS, as demonstrated here and in main-tenance of common fragile sites in human genome as reported(14, 47). However, without necessary regulation of their access toDNA replication forks, the substantial efficiency and processivityof these two low-fidelity DNA polymerases could lead to a highfrequency of mutagenesis. We expect that stringent down-regulationof Pol ζ may keep this essential polymerase engaged only whennecessary for lesion bypass and cell proliferation.
Materials and MethodsHuman Pol ζ2 and ζ4 with His8–MBP-labeled Rev7 were expressed in tran-siently transfected HEK293 cells and purified by using affinity pull-down,ion-exchange, and size-exclusion chromatography. The catalytically inactivePol ζ4− mutant was generated by using QuikChange (Stratagene). Catalyticactivities of Pol ζ variants were measured with Fluorescein (6-FAM)-labeledprimers as described (26). A full description of materials and methods can befound in SI Materials and Methods.
ACKNOWLEDGMENTS. We thank Dr. Christopher W. Lawrence for humanREV3L andMAD2L2 cDNAs and Drs. R. Craigie and D. J. Leahy for editing thismanuscript. This work is supported by National Institutes of Health Intra-mural Research Grant DK036146-07.
1. Nelson JR, Lawrence CW, Hinkle DC (1996) Thymine-thymine dimer bypass by yeastDNA polymerase zeta. Science 272(5268):1646–1649.
2. Lawrence CW (2004) Cellular functions of DNA polymerase zeta and Rev1 protein.Adv Protein Chem 69:167–203.
3. Kunkel TA, Burgers PM (2008) Dividing the workload at a eukaryotic replication fork.Trends Cell Biol 18(11):521–527.
4. Gibbs PE, McGregor WG, Maher VM, Nisson P, Lawrence CW (1998) A human ho-molog of the Saccharomyces cerevisiae REV3 gene, which encodes the catalytic sub-unit of DNA polymerase zeta. Proc Natl Acad Sci USA 95(12):6876–6880.
5. Murakumo Y, et al. (2000) A human REV7 homolog that interacts with the poly-merase zeta catalytic subunit hREV3 and the spindle assembly checkpoint proteinhMAD2. J Biol Chem 275(6):4391–4397.
6. Aravind L, Koonin EV (1998) The HORMA domain: A common structural denominatorin mitotic checkpoints, chromosome synapsis and DNA repair. Trends Biochem Sci23(8):284–286.
7. Lemontt JF (1971) Mutants of yeast defective in mutation induced by ultraviolet light.Genetics 68(1):21–33.
8. Lawrence CW, Das G, Christensen RB (1985) REV7, a new gene concerned with UVmutagenesis in yeast. Mol Gen Genet 200(1):80–85.
9. Van Sloun PP, Romeijn RJ, Eeken JC (1999) Molecular cloning, expression and chro-mosomal localisation of the mouse Rev3l gene, encoding the catalytic subunit ofpolymerase zeta. Mutat Res 433(2):109–116.
10. Esposito G, et al. (2000) Disruption of the Rev3l-encoded catalytic subunit of poly-merase zeta in mice results in early embryonic lethality. Curr Biol 10(19):1221–1224.
11. Gan GN, Wittschieben JP, Wittschieben BO, Wood RD (2008) DNA polymerase zeta(pol zeta) in higher eukaryotes. Cell Res 18(1):174–183.
12. Lange SS, et al. (2013) Dual role for mammalian DNA polymerase ζ in maintaininggenome stability and proliferative responses. Proc Natl Acad Sci USA 110(8):E687–E696.
13. Lange SS, Wittschieben JP, Wood RD (2012) DNA polymerase zeta is required forproliferation of normal mammalian cells. Nucleic Acids Res 40(10):4473–4482.
14. Bhat A, Andersen PL, Qin Z, Xiao W (2013) Rev3, the catalytic subunit of Polζ, is re-quired for maintaining fragile site stability in human cells. Nucleic Acids Res 41(4):2328–2339.
15. Ho TV, Guainazzi A, Derkunt SB, Enoiu M, Schärer OD (2011) Structure-dependentbypass of DNA interstrand crosslinks by translesion synthesis polymerases. NucleicAcids Res 39(17):7455–7464.
16. Harfe BD, Jinks-Robertson S (2000) DNA polymerase zeta introduces multiple muta-tions when bypassing spontaneous DNA damage in Saccharomyces cerevisiae. MolCell 6(6):1491–1499.
17. Stone JE, Lujan SA, Kunkel TA, Kunkel TA (2012) DNA polymerase zeta generatesclustered mutations during bypass of endogenous DNA lesions in Saccharomycescerevisiae. Environ Mol Mutagen 53(9):777–786.
18. Saribasak H, et al. (2012) DNA polymerase ζ generates tandem mutations in immu-noglobulin variable regions. J Exp Med 209(6):1075–1081.
19. Daly J, et al. (2012) Altered Ig hypermutation pattern and frequency in comple-mentary mouse models of DNA polymerase ζ activity. J Immunol 188(11):5528–5537.
20. Wu F, Lin X, Okuda T, Howell SB (2004) DNA polymerase zeta regulates cisplatin cy-totoxicity, mutagenicity, and the rate of development of cisplatin resistance. CancerRes 64(21):8029–8035.
21. Doles J, et al. (2010) Suppression of Rev3, the catalytic subunit of Polzeta, sensitizesdrug-resistant lung tumors to chemotherapy. Proc Natl Acad Sci USA 107(48):20786–20791.
22. Sharma S, Shah NA, Joiner AM, Roberts KH, Canman CE (2012) DNA polymerase ζ isa major determinant of resistance to platinum-based chemotherapeutic agents. MolPharmacol 81(6):778–787.
23. Johnson RE, Prakash L, Prakash S (2012) Pol31 and Pol32 subunits of yeast DNApolymerase δ are also essential subunits of DNA polymerase ζ. Proc Natl Acad Sci USA109(31):12455–12460.
24. Livneh Z, Ziv O, Shachar S (2010) Multiple two-polymerase mechanisms in mammaliantranslesion DNA synthesis. Cell Cycle 9(4):729–735.
25. Yang W, Woodgate R (2007) What a difference a decade makes: Insights intotranslesion DNA synthesis. Proc Natl Acad Sci USA 104(40):15591–15598.
26. Wang F, Yang W (2009) Structural insight into translesion synthesis by DNA Pol II. Cell139(7):1279–1289.
27. Huang ME, Rio AG, Galibert MD, Galibert F (2002) Pol32, a subunit of Saccharomycescerevisiae DNA polymerase delta, suppresses genomic deletions and is involved in themutagenic bypass pathway. Genetics 160(4):1409–1422.
28. Gibbs PE, McDonald J, Woodgate R, Lawrence CW (2005) The relative roles in vivo ofSaccharomyces cerevisiae Pol eta, Pol zeta, Rev1 protein and Pol32 in the bypass andmutation induction of an abasic site, T-T (6-4) photoadduct and T-T cis-syn cyclo-butane dimer. Genetics 169(2):575–582.
29. Hanna M, Ball LG, Tong AH, Boone C, Xiao W (2007) Pol32 is required for Pol zeta-dependent translesion synthesis and prevents double-strand breaks at the replicationfork. Mutat Res 625(1-2):164–176.
30. Dua R, Levy DL, Campbell JL (1998) Role of the putative zinc finger domain of Sac-charomyces cerevisiae DNA polymerase epsilon in DNA replication and the S/Mcheckpoint pathway. J Biol Chem 273(45):30046–30055.
31. Sanchez Garcia J, Ciufo LF, Yang X, Kearsey SE, MacNeill SA (2004) The C-terminal zincfinger of the catalytic subunit of DNA polymerase delta is responsible for direct in-teraction with the B-subunit. Nucleic Acids Res 32(10):3005–3016.
32. Klinge S, Núñez-Ramírez R, Llorca O, Pellegrini L (2009) 3D architecture of DNA Polalpha reveals the functional core of multi-subunit replicative polymerases. EMBO J28(13):1978–1987.
33. Tahirov TH, Makarova KS, Rogozin IB, Pavlov YI, Koonin EV (2009) Evolution of DNApolymerases: an inactivated polymerase-exonuclease module in Pol epsilon anda chimeric origin of eukaryotic polymerases from two classes of archaeal ancestors.Biol Direct 4:11.
34. Netz DJ, et al. (2012) Eukaryotic DNA polymerases require an iron-sulfur cluster forthe formation of active complexes. Nat Chem Biol 8(1):125–132.
35. Baranovskiy AG, et al. (2008) X-ray structure of the complex of regulatory subunits ofhuman DNA polymerase delta. Cell Cycle 7(19):3026–3036.
36. Johansson E, Majka J, Burgers PM (2001) Structure of DNA polymerase delta fromSaccharomyces cerevisiae. J Biol Chem 276(47):43824–43828.
37. Baranovskiy AG, et al. (2012) DNA polymerase δ and ζ switch by sharing accessorysubunits of DNA polymerase δ. J Biol Chem 287(21):17281–17287.
38. Makarova AV, Stodola JL, Burgers PM (2012) A four-subunit DNA polymerase ζcomplex containing Pol δ accessory subunits is essential for PCNA-mediated muta-genesis. Nucleic Acids Res 40(22):11618–11626.
39. Acharya N, Johnson RE, Prakash S, Prakash L (2006) Complex formation with Rev1enhances the proficiency of Saccharomyces cerevisiae DNA polymerase zeta for mis-match extension and for extension opposite from DNA lesions. Mol Cell Biol 26(24):9555–9563.
40. Kikuchi S, Hara K, Shimizu T, Sato M, Hashimoto H (2012) Structural basis of re-cruitment of DNA polymerase ζ by interaction between REV1 and REV7 proteins.J Biol Chem 287(40):33847–33852.
41. Wojtaszek J, et al. (2012) Structural basis of Rev1-mediated assembly of a quaternaryvertebrate translesion polymerase complex consisting of Rev1, heterodimeric poly-merase (Pol) ζ, and Pol κ. J Biol Chem 287(40):33836–33846.
42. Xie W, Yang X, Xu M, Jiang T (2012) Structural insights into the assembly of humantranslesion polymerase complexes. Protein Cell 3(11):864–874.
43. Adachi M, et al. (2008) Hypersensitivity to cisplatin after hRev3 mRNA knockdown inhead and neck squamous cell carcinoma cells. Mol Med Rep 1(5):695–698.
44. Zhao Y, et al. (2012) Structural basis of human DNA polymerase η-mediated chemo-resistance to cisplatin. Proc Natl Acad Sci USA 109(19):7269–7274.
45. Hashimoto K, et al. (2012) The vital role of polymerase ζ and REV1 in mutagenic, butnot correct, DNA synthesis across benzo[a]pyrene-dG and recruitment of polymerase ζby REV1 to replication-stalled site. J Biol Chem 287(12):9613–9622.
46. Gomez-Llorente Y, et al. (2013) The architecture of yeast DNA polymerase zeta. CellRep 5(1):79–86.
47. Bergoglio V, et al. (2013) DNA synthesis by Pol η promotes fragile site stability bypreventing under-replicated DNA in mitosis. J Cell Biol 201(3):395–408.
Lee et al. PNAS | February 25, 2014 | vol. 111 | no. 8 | 2959
BIOCH
EMISTR
YSE
ECO
MMEN
TARY
Dow
nloa
ded
by g
uest
on
Mar
ch 2
9, 2
020