
Human NK Cells Express Endothelial Nitric Oxide Synthase ... · This article cites 55 articles, 27...
9
of November 13, 2017. This information is current as α Regulating Expression of TNF- Them from Activation-Induced Cell Death by Oxide Synthase, and Nitric Oxide Protects Human NK Cells Express Endothelial Nitric Eda T. Bloom Horvath-Arcidiacono, Kotaro Hori, Howard Mostowski and Keizo Furuke, Parris R. Burd, Judith A. http://www.jimmunol.org/content/163/3/1473 1999; 163:1473-1480; ; J Immunol average * 4 weeks from acceptance to publication Speedy Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • ? The JI Why References http://www.jimmunol.org/content/163/3/1473.full#ref-list-1 , 27 of which you can access for free at: cites 55 articles This article Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 1999 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on November 13, 2017 http://www.jimmunol.org/ Downloaded from by guest on November 13, 2017 http://www.jimmunol.org/ Downloaded from
Transcript of Human NK Cells Express Endothelial Nitric Oxide Synthase ... · This article cites 55 articles, 27...

of November 13, 2017.This information is current as
αRegulating Expression of TNF-Them from Activation-Induced Cell Death byOxide Synthase, and Nitric Oxide Protects Human NK Cells Express Endothelial Nitric
Eda T. BloomHorvath-Arcidiacono, Kotaro Hori, Howard Mostowski and Keizo Furuke, Parris R. Burd, Judith A.
http://www.jimmunol.org/content/163/3/14731999; 163:1473-1480; ;J Immunol
average*
4 weeks from acceptance to publicationSpeedy Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
?The JIWhy
Referenceshttp://www.jimmunol.org/content/163/3/1473.full#ref-list-1
, 27 of which you can access for free at: cites 55 articlesThis article
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 13, 2017
http://ww
w.jim
munol.org/
Dow
nloaded from

Human NK Cells Express Endothelial Nitric Oxide Synthase,and Nitric Oxide Protects Them from Activation-Induced CellDeath by Regulating Expression of TNF-a
Keizo Furuke,* Parris R. Burd,* Judith A. Horvath-Arcidiacono,* Kotaro Hori, †
Howard Mostowski,* and Eda T. Bloom1*
Although NO appears important in rodent immune responses, its involvement in the human immune system is unclear. We reportthat human NK cells express constitutive endothelial NO synthase mRNA and protein, but not detectable levels of inducible NOsynthase. They produce NO following activation by coculture with target cells or cross-linking with anti-CD16 mAb, and pro-duction is increased in the presence of IL-2.N-monomethyl-L-arginine (L-NMA), a NOS inhibitor, partially inhibited NK cell lysisof four different target cells (<40% inhibition at 500 mM L-NMA), but not granule release following coculture with target cells,or Fas ligand induction following cross-linking with anti-CD16 mAb. However, L-NMA augmented apoptosis of NK cells inducedby activation through CD16 ligation or coculture with K562. An NO donor, S-nitroso-N-acetylpenicillamine (SNAP), suppressedapoptosis of NK cells induced by CD16 cross-linking or coculture with target cells, suggesting that endogenous NO production isinvolved in protection of NK cells from activation-induced apoptosis, thereby maintaining NK activity. SNAP also suppressed, andL-NMA enhanced, expression of TNF-a, reported to be involved in activation-induced NK cell death, in response to CD16cross-linking. Suppression of anti-CD16-induced apoptosis by SNAP was reversed by the addition of rTNF-a. DNA-bindingactivity of the transcription factor, NF-AT, which is involved in TNF- a induction upon ligation of CD16, was inhibited by SNAPand enhanced byL-NMA. Our results suggest that down-regulation of TNF-a expression, possibly due to suppression of NF-ATactivation, is a mechanism by which endogenous NO protects NK cells from activation-induced apoptosis, and maintains lyticcapacity. The Journal of Immunology,1999, 163: 1473–1480.
N itric oxide has been implicated in a number of physio-logical functions, including smooth muscle relaxation,platelet inhibition, neurotransmission, and macrophage-
mediated cytotoxicity (1, 2). The wide variety of effects isachieved from its interactions with target molecules via a rich ox-idation-reduction (redox) potential (3). Synthesis of NO fromL-arginine is catalyzed by the enzyme NO synthase (NOS),2 whichincludes two constitutive isoforms, endothelial NOS (eNOS) andneuronal NOS, and an inducible isoform, termed iNOS (4). iNOSis induced by stimuli such as bacterial endotoxins or cytokines and,in mice, has been shown to participate in immune defense againstbacterial, fungal, viral, or parasitic pathogens through productionof large amounts of NO by macrophages (5). However, althoughhuman iNOS is also reported to be induced in monocytes and/ormacrophages by LPS, IFN-g, or HIV infection, the levels of NOproduction appear relatively low compared with that in rodent sys-
tems (6–8). Thus, a physiological role for NO in the human im-mune system has not been identified. Activity of constitutive NOSsis Ca21 dependent, transient, and responsible for NO production atvery low levels in comparison with that of iNOS (4, 9). The lowlevels of NO produced by the constitutive isoforms are believed tofunction in signaling for regulation of vascular tonus or neuro-transmission (4, 9).
NK cells play a crucial role in immune surveillance by recog-nizing and eliminating virus-infected or neoplastic target cells(10–12). We previously reported that NK cell functions are regu-lated by the intracellular redox status (13, 14). Thiol-reducingcompounds, such as cysteine and glutathione, are required for op-timal proliferation and killing activity of NK cells in response toIL-2. Conversely, a variety of oxidative stresses is known to me-diate apoptosis in many systems (15, 16) and facilitates Fas-in-duced apoptosis in NK cells (17). NO-releasing agents have beenreported to inhibit human NK cell activity (18, 19). Production ofNO by murine NK cells has been reported (20), whereas NO pro-duction by human NK cells has not been confirmed. In the presentstudy, we showed that human NK cells express eNOS mRNA andprotein and identified a potential physiological role for NO pro-duction and mechanism of action in human NK cells.
Materials and MethodsReagents, Abs, and media
Human rIL-2 and rIL-12 were provided by Amgen (Thousand Oaks, CA)and Genetics Institute (Cambridge, MA), respectively. Human rTNF-a waspurchased from Genzyme (Cambridge, MA).N-monomethyl-L-arginine,monoacetate salt (L-NMA) was purchased from Calbiochem-Novabiochem(La Jolla, CA).S-nitroso-N-acetylpenicillamine (SNAP) was obtained fromSigma (St. Louis, MO). Metalloproteinase inhibitor KB8301 was pur-chased from PharMingen (San Diego, CA). Anti-CD5 mAb was purified
*Laboratory of Cellular Immunology, Division of Cellular and Gene Therapies, and†Division of Cytokine Biology, Center for Biologics Evaluation and Research, Foodand Drug Administration, Bethesda, MD 20892
Received for publication November 25, 1998. Accepted for publication May21, 1999.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 Address correspondence and reprint requests to Dr. Eda T. Bloom, Chief, Labora-tory of Cellular Immunology (HFM-518), Division of Cellular and Gene Therapies,Center for Biologics Evaluation Research, Food and Drug Administration, 8800Rockville Pike, Bethesda, MD 20892. E-mail address: [email protected] Abbreviations used in this paper: NOS, NO synthase; AO, acridine orange; BLT,benzyloxycarbonyl-L-lysine thiobenzyl ester; EB, ethidium bromide; FasL, naturalcellular ligand for Fas/CD95/APO-1; GaM, goat anti-mouse IgG; eNOS, endothelialNOS; iNOS, inducible NOS; L-NMA, N-monomethyl-L-arginine; SNAP,S-nitroso-N-acetylpenicillamine.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from

from ascites, as described previously (14). Anti-CD22 mAb was purchasedfrom PharMingen. Anti-CD16 (clone 3G8), anti-CD36, anti-CD16 PE, andanti-CD56 PE mAbs were purchased from Immunotech (Westbrook, ME).Anti-CD16 FITC mAb and anti-CD56 mAb as an isotype control for 3G8were purchased from Becton Dickinson (Mountain View, CA). Goat anti-mouse IgG (GaM) mAb was purchased from Sigma.
RPMI 1640 media with or without phenol red or without arginine werepurchased from Life Technologies (Grand Island, NY). Standard RPMI1640 medium was prepared with 10% FCS (HyClone Laboratories, Logan,UT), 50 mg/ml gentamicin sulfate, and 2 mM glutamine.
Cells and purification of human NK cells
Human NK cell lines, YT and NK3.3, were kindly provided by Dr. J.Yodoi (Kyoto University, Kyoto, Japan) and Dr. J. Kornbluth (Universityof Arkansas, Little Rock, AR), respectively.
Human NK cells were purified from buffy coats from healthy donors(Blood Bank, National Institutes of Health, Bethesda, MD), as described(17). Briefly, mononuclear cells were obtained by centrifugation over Lym-phocyte Separation Medium (LSM; Organon Teknika, Durham, NC) andtreated with 100 mg/108 cells of sterilized carbonyl iron (Sigma) in stan-dard RPMI 1640 medium at 37°C for 30 min, followed by exposure to amagnetic field to remove monocytes. Cell suspensions were centrifugedover 47.5% Percoll to obtain low density large granular cells. NK-enrichedmononuclear cells were further treated with anti-CD5, anti-CD22, and anti-CD36 mAbs at 4°C for 2–4 h, incubated with GaM-coated magnet beads(PerSeptive Biosystems, Framingham, MA) at 4°C for 30 min, and ex-posed to a magnetic field to eliminate residual T cells, B cells, and mono-cytes. The purity of NK cells (CD161 and/or CD561) in each experimentwas always 85–95%, as assessed by flow cytometry (FACScan; BectonDickinson). Of the population defined as CD561 and/or CD161 cells,;10% expressed CD16 alone, 30% expressed CD56 alone, and greaterthan 50% expressed both markers.
A human glioblastoma cell line, A172, stimulated with IFN-g and LPSwas used as a positive control for human iNOS mRNA and protein (21).
RT-PCR analysis of RNA
Human iNOS, eNOS, and TNF-a mRNAs were identified by semiquanti-tative RT-PCR, as described previously (22). Total RNA was isolated fromcells using TRIzol (Life Technologies), according to the manufacturer’sinstructions. The sequences of the primer pairs in this experiment were asfollows: humanb-actin as an internal control (59-ATC TGG CAC CACACC TTC TAC A-39, 59-GTT TCG TGG ATG CCA CAG GAC T-39),human iNOS (59-TGT GCC ACC TCC AGT CCA GTG ACA-39, 59-GCTCAT CTC CCG TCA GTT GGT AGG-39), human eNOS (59-GCA TCCCTA CTC CCA CCA GCG CCA-39, 59-TCC CGG GCA TCG AAC ACCTGC AGC-39), and human TNF-a (59-TCA GCT TGA GGG TTT GCTACA A-39, 59-TCT GGC CCA GGC AGT CAG ATC-39). Amplificationswere performed in a thermocycler (GeneAmp PCR system 9600; Perkin-Elmer, Norwalk, CT), as follows: 94°C, 3 min, followed by 21 (b-actin),40 (NOSs), or 30 (TNF-a) cycles (94°C, 1 min; 57°C, 1.5 min; 72°C, 2min). Aliquots of each amplification were analyzed by electrophoresis in6% (b-actin and NOSs) or 8% (TNF-a) acrylamide Tris-borate gels. Thelanes were normalized tob-actin mRNA levels.
Immunoblot analysis
iNOS and eNOS protein levels were assayed by immunoblot. A total of5 3 106 cells was lysed with lysis buffer (50 mM Tris-HCl (pH 7.5), 150mM NaCl, 1% Nonidet P-40, 0.5% sodium deoxycholate, 0.1% SDS, 1mM EGTA, 50mg/ml leupeptin, 50mg/ml aprotinin, 50mM p-nitrophenylp9-guanidino-benzoate). Equivalent amounts of total cellular protein (50mg/lane) were separated on 7.5% SDS-polyacrylamide gels and blotted topolyvinyl difluoride membranes (Millipore, Bedford, MA). After blockingwith 1.25% BSA, 1.25% chicken egg albumin, 25 mM Tris-HCl (pH 7.5),150 mM NaCl, and 3 mM KCl, the membrane was incubated with rabbitanti-eNOS or iNOS Abs (Calbiochem-Novabiochem) and then with don-key HRP-conjugated anti-rabbit IgG Ab (Amersham, Arlington Heights,IL) at room temperature for 1 h. Protein bands were visualized using en-hanced chemoluminescence detection reagents (Amersham) and exposureto XAR film (Eastman Kodak, Rochester, NY) for 10–30 s.
Measurement of NO oxidation products, nitrite plus nitrate(nitrite/nitrate) by Griess reaction using nitrate reductase
Because NO in oxygen-containing solutions is chemically unstable andundergoes rapid oxidation to nitrite, and cellular components catalyze itsfurther oxidation to nitrate, production of NO was determined by measur-ing the formation of the stable oxidation products of NO, nitrite, and ni-
trate, as described (23). Cells were treated as described inResultsat 107/ml.Nitrate in cell culture supernatants was reduced to nitrite by incubation ofthe samples for 30 min with nitrate reductase (0.1 U/ml; Sigma) in thepresence of 100mM nicotinamide adenine dinucleotide phosphate(NADPH) and 10mM flavin adenine dinucleotide. Remaining NADPHwas oxidized with lactate dehydrogenase (10 U/ml) in the presence of 10mM sodium pyruvate. Total nitrite concentration was then determined byusing a procedure based on the Griess reaction (24). The baseline level ofthis assay was 0.1 nmol/106 cells according to the levels detected in mediaalone.
Cytotoxicity assay
The 51Cr release assay was performed as described (14) using four E:Tratios in quadrupling dilutions beginning at 5:1 of viable cells (determinedby trypan blue exclusion). K562, Molt-4, U937, and UCLA-SO-M14(M14) cells were used as target cells. Data were expressed as lytic units in107 cells, in which 1 lytic unit is the number of effector cells required tolyse 20% of the target cells, obtained by fitting the titration curves to scalefamilies of curves, as described previously (25).
Measurements of apoptosis
The percentage of apoptotic cells was measured by fluorescence micros-copy, after staining with acridine orange (AO) and ethidium bromide (EB),as described (26).
The JAM test of Matzinger (27) was used to quantify target-inducedDNA fragmentation in [3H]TdR-labeled NK cells, with some modificationsas described (28), except that NK cells were pulsed with 0.2mCi of[3H]TdR per 53 105 cells/ml in standard medium at 37°C for 12 h, andlabeled cells were cocultured with or without K562 cells at E:T (NK:K562)ratios of 3:1, with the indicated treatment at 37°C for 4 h. The percentageof DNA fragmentation was calculated as: percentage of DNA fragmenta-tion 5 (S2 R)/S3 100;S5 cpm retained in the absence of effector cells,R 5 cpm retained in the presence of effector cells.
Granule exocytosis assay
Granule exocytosis by NK cells upon coculture with target cells was mea-sured by BLT-esterase assay, as described, with modifications (29). Inbrief, 50ml of 2 3 106 cells/ml IL-2-activated NK cells were cultured with50 ml of 2 3 105 cells/ml K562 cells in the absence or presence of theindicated concentrations ofL-NMA for 4 h at37°C. Esterase was measuredin supernatants, and total esterase content was obtained in supernatantsfollowing incubation of cells with 1% Nonidet P-40 for 4 h at37°C. Per-centage of secretion of esterase was calculated as: percentage of secre-tion 5 1003 (E 2 B)/(T 2 B); E 5 mean absorbance of the supernatantsfrom NK cells incubated with K562 cells,B 5 mean absorbance of thesupernatants from NK cells incubated without K562 cells, andT 5 totalesterase content.
Flow-cytometric analyses for expression of FasL and NOS
Cell surface FasL expression was quantitated by flow-cytometric analysis,as described previously (30). Briefly, cells were treated with a metallopro-tease inhibitor, KB8301, which blocks FasL cleavage resulting in highlevels of cell surface FasL expression (31), for 6 h, washed by FACS buffer(PBS with 1% FCS and 0.1% NaN3), and incubated with biotin-conjugatedanti-human FasL mAb (PharMingen) or biotin-conjugated mouse IgG1(PharMingen) for 20 min on ice. After washing with FACS buffer, theywere incubated with streptavidin PE (Becton Dickinson), and fluorescenceintensity was measured by FACScan.
NOS expression was examined in NK cells by double staining with Absto surface and intracytoplasmic Ags. Cells were stained with anti-CD16 PEand anti-CD56 PE or a control mouse IgG PE (Sigma), followed by fixationwith 4% paraformaldehyde in FACS buffer (1% FCS and 0.1% sodiumazide in PBS) for 20 min on ice. Cells were washed with FACS buffer,permeabilized with saponin (0.1% saponin in FACS buffer), vortexedgently, incubated with rat IgG (Sigma) for 20 min on ice to block nonspe-cific binding and then with anti-eNOS Ab, anti-iNOS Ab, or a controlrabbit IgG (Sigma) for 30 min on ice, and finally washed with saponinsolution. Next they were stained with FITC-conjugated anti-rabbit Ab (Sig-ma) in saponin solution for 30 min on ice, washed twice with saponinsolution. Fluorescence was analyzed by FACScan.
Electrophoretic mobility shift assay
Nuclear extracts were prepared from cells cultured with the indicated treat-ment, as described by Schreiber et al. (32).
We used the sequence for human IL-2 distal NF-AT site (GGA GGAAAA ACT GTT TCA TAC AGA AGG (33)) as a probe to measure NF-AT
1474 ENDOTHELIAL NOS IN HUMAN NK CELLS
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from
activity. dsDNA probe was synthesized in the CBER Core Facility Services(Center for Biologics Evaluation and Research, Food and Drug Adminis-tration, Bethesda, MD). The dsDNA probe was end labeled with[g-32P]ATP (3000 Ci/mmol; Amersham) and T4 polynucleotide kinase(Amersham Pharmacia Biotech, Piscataway, NJ), according to the manu-facturer’s instructions.
EMSA was performed as described (34) with some modifications. Inbrief, nuclear extracts (5mg per reaction volume) were incubated for 20min at room temperature with 1mg poly(dI-dC) (Amersham PharmaciaBiotech) and 0.5 ng32P-labeled dsDNA probe in 20ml of binding buffer(10 mM Tris-HCl, pH 7.5), 50 mM NaCl, 30 mM KCl, 5 mM MgCl2, 10%glycerol, and 0.05% Nonidet P-40. The reaction was stopped with 2ml of0.1% bromophenol blue, followed by electrophoresis on 5% polyacryl-amide gels in 0.253 Tris-buffered EDTA (pH 8.4) under nondenaturingconditions. Five nanograms of unlabeled cold probe were added at thebeginning of the reaction as a competitor. For supershift, antisera againstmurine NF-AT1 (Upstate Biotechnology, Lake Placid, NY), which cross-reacts with human NF-AT1, were added to nuclear extracts and incubatedon ice for 30 min before the binding reaction.
Statistical analysis
The one-tailed Student’st test was used for data analysis.
ResultsHuman NK cells expressed eNOS, but not iNOS, mRNA andprotein
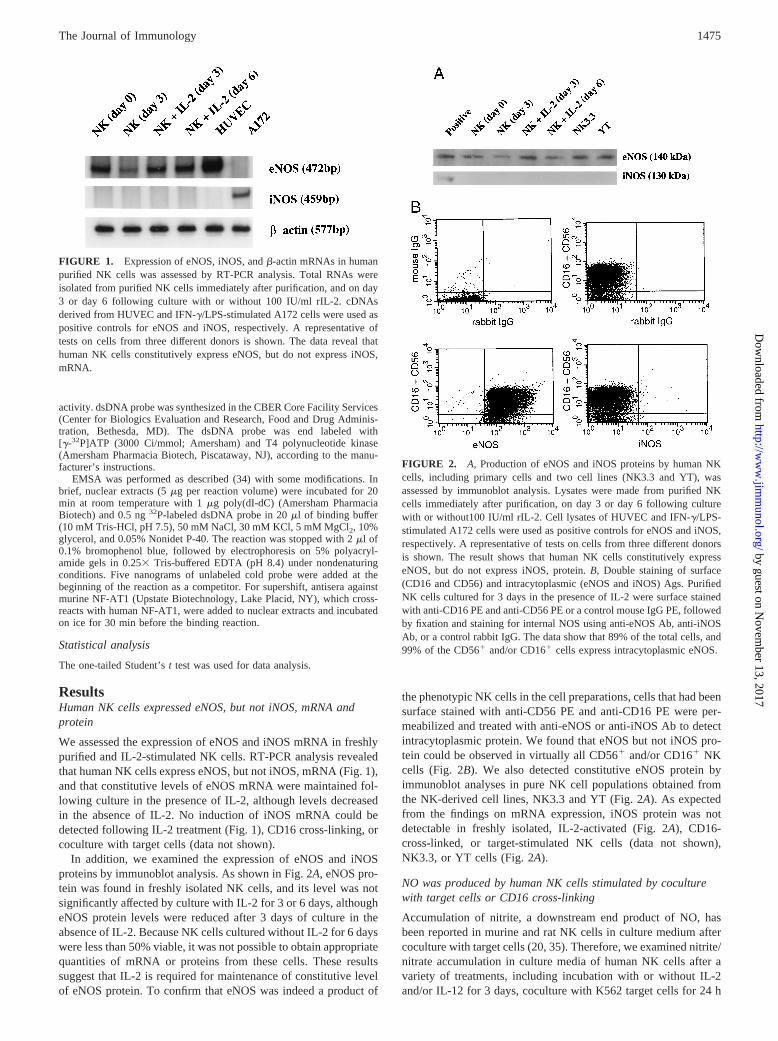
We assessed the expression of eNOS and iNOS mRNA in freshlypurified and IL-2-stimulated NK cells. RT-PCR analysis revealedthat human NK cells express eNOS, but not iNOS, mRNA (Fig. 1),and that constitutive levels of eNOS mRNA were maintained fol-lowing culture in the presence of IL-2, although levels decreasedin the absence of IL-2. No induction of iNOS mRNA could bedetected following IL-2 treatment (Fig. 1), CD16 cross-linking, orcoculture with target cells (data not shown).
In addition, we examined the expression of eNOS and iNOSproteins by immunoblot analysis. As shown in Fig. 2A, eNOS pro-tein was found in freshly isolated NK cells, and its level was notsignificantly affected by culture with IL-2 for 3 or 6 days, althougheNOS protein levels were reduced after 3 days of culture in theabsence of IL-2. Because NK cells cultured without IL-2 for 6 dayswere less than 50% viable, it was not possible to obtain appropriatequantities of mRNA or proteins from these cells. These resultssuggest that IL-2 is required for maintenance of constitutive levelof eNOS protein. To confirm that eNOS was indeed a product of
the phenotypic NK cells in the cell preparations, cells that had beensurface stained with anti-CD56 PE and anti-CD16 PE were per-meabilized and treated with anti-eNOS or anti-iNOS Ab to detectintracytoplasmic protein. We found that eNOS but not iNOS pro-tein could be observed in virtually all CD561 and/or CD161 NKcells (Fig. 2B). We also detected constitutive eNOS protein byimmunoblot analyses in pure NK cell populations obtained fromthe NK-derived cell lines, NK3.3 and YT (Fig. 2A). As expectedfrom the findings on mRNA expression, iNOS protein was notdetectable in freshly isolated, IL-2-activated (Fig. 2A), CD16-cross-linked, or target-stimulated NK cells (data not shown),NK3.3, or YT cells (Fig. 2A).
NO was produced by human NK cells stimulated by coculturewith target cells or CD16 cross-linking
Accumulation of nitrite, a downstream end product of NO, hasbeen reported in murine and rat NK cells in culture medium aftercoculture with target cells (20, 35). Therefore, we examined nitrite/nitrate accumulation in culture media of human NK cells after avariety of treatments, including incubation with or without IL-2and/or IL-12 for 3 days, coculture with K562 target cells for 24 h
FIGURE 1. Expression of eNOS, iNOS, andb-actin mRNAs in humanpurified NK cells was assessed by RT-PCR analysis. Total RNAs wereisolated from purified NK cells immediately after purification, and on day3 or day 6 following culture with or without 100 IU/ml rIL-2. cDNAsderived from HUVEC and IFN-g/LPS-stimulated A172 cells were used aspositive controls for eNOS and iNOS, respectively. A representative oftests on cells from three different donors is shown. The data reveal thathuman NK cells constitutively express eNOS, but do not express iNOS,mRNA.
FIGURE 2. A, Production of eNOS and iNOS proteins by human NKcells, including primary cells and two cell lines (NK3.3 and YT), wasassessed by immunoblot analysis. Lysates were made from purified NKcells immediately after purification, on day 3 or day 6 following culturewith or without100 IU/ml rIL-2. Cell lysates of HUVEC and IFN-g/LPS-stimulated A172 cells were used as positive controls for eNOS and iNOS,respectively. A representative of tests on cells from three different donorsis shown. The result shows that human NK cells constitutively expresseNOS, but do not express iNOS, protein.B, Double staining of surface(CD16 and CD56) and intracytoplasmic (eNOS and iNOS) Ags. PurifiedNK cells cultured for 3 days in the presence of IL-2 were surface stainedwith anti-CD16 PE and anti-CD56 PE or a control mouse IgG PE, followedby fixation and staining for internal NOS using anti-eNOS Ab, anti-iNOSAb, or a control rabbit IgG. The data show that 89% of the total cells, and99% of the CD561 and/or CD161 cells express intracytoplasmic eNOS.
1475The Journal of Immunology
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from
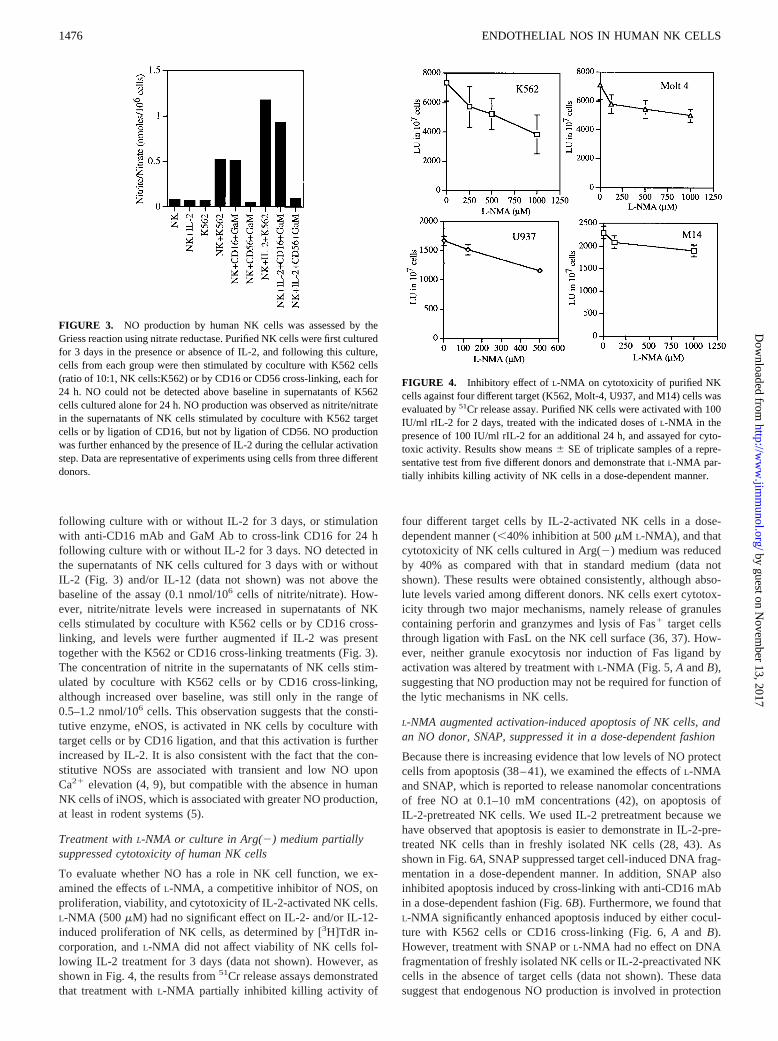
following culture with or without IL-2 for 3 days, or stimulationwith anti-CD16 mAb and GaM Ab to cross-link CD16 for 24 hfollowing culture with or without IL-2 for 3 days. NO detected inthe supernatants of NK cells cultured for 3 days with or withoutIL-2 (Fig. 3) and/or IL-12 (data not shown) was not above thebaseline of the assay (0.1 nmol/106 cells of nitrite/nitrate). How-ever, nitrite/nitrate levels were increased in supernatants of NKcells stimulated by coculture with K562 cells or by CD16 cross-linking, and levels were further augmented if IL-2 was presenttogether with the K562 or CD16 cross-linking treatments (Fig. 3).The concentration of nitrite in the supernatants of NK cells stim-ulated by coculture with K562 cells or by CD16 cross-linking,although increased over baseline, was still only in the range of0.5–1.2 nmol/106 cells. This observation suggests that the consti-tutive enzyme, eNOS, is activated in NK cells by coculture withtarget cells or by CD16 ligation, and that this activation is furtherincreased by IL-2. It is also consistent with the fact that the con-stitutive NOSs are associated with transient and low NO uponCa21 elevation (4, 9), but compatible with the absence in humanNK cells of iNOS, which is associated with greater NO production,at least in rodent systems (5).
Treatment withL-NMA or culture in Arg(2) medium partiallysuppressed cytotoxicity of human NK cells
To evaluate whether NO has a role in NK cell function, we ex-amined the effects ofL-NMA, a competitive inhibitor of NOS, onproliferation, viability, and cytotoxicity of IL-2-activated NK cells.L-NMA (500 mM) had no significant effect on IL-2- and/or IL-12-induced proliferation of NK cells, as determined by [3H]TdR in-corporation, andL-NMA did not affect viability of NK cells fol-lowing IL-2 treatment for 3 days (data not shown). However, asshown in Fig. 4, the results from51Cr release assays demonstratedthat treatment withL-NMA partially inhibited killing activity of
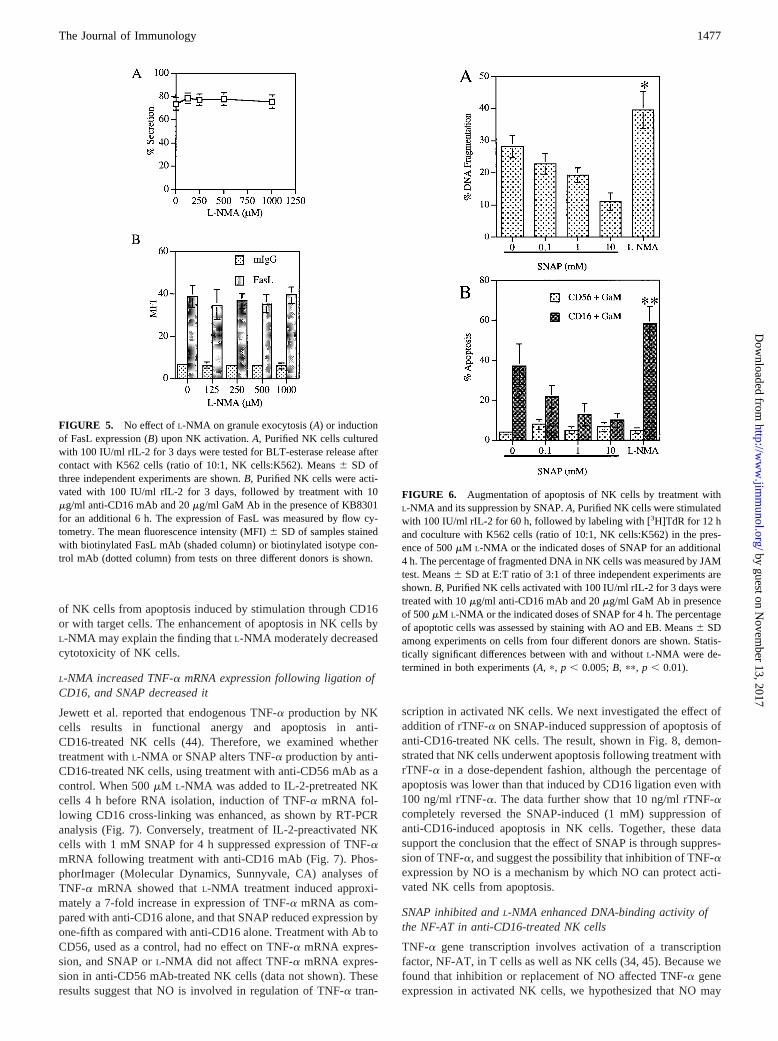
four different target cells by IL-2-activated NK cells in a dose-dependent manner (,40% inhibition at 500mM L-NMA), and thatcytotoxicity of NK cells cultured in Arg(2) medium was reducedby 40% as compared with that in standard medium (data notshown). These results were obtained consistently, although abso-lute levels varied among different donors. NK cells exert cytotox-icity through two major mechanisms, namely release of granulescontaining perforin and granzymes and lysis of Fas1 target cellsthrough ligation with FasL on the NK cell surface (36, 37). How-ever, neither granule exocytosis nor induction of Fas ligand byactivation was altered by treatment withL-NMA (Fig. 5, A andB),suggesting that NO production may not be required for function ofthe lytic mechanisms in NK cells.
L-NMA augmented activation-induced apoptosis of NK cells, andan NO donor, SNAP, suppressed it in a dose-dependent fashion
Because there is increasing evidence that low levels of NO protectcells from apoptosis (38–41), we examined the effects ofL-NMAand SNAP, which is reported to release nanomolar concentrationsof free NO at 0.1–10 mM concentrations (42), on apoptosis ofIL-2-pretreated NK cells. We used IL-2 pretreatment because wehave observed that apoptosis is easier to demonstrate in IL-2-pre-treated NK cells than in freshly isolated NK cells (28, 43). Asshown in Fig. 6A, SNAP suppressed target cell-induced DNA frag-mentation in a dose-dependent manner. In addition, SNAP alsoinhibited apoptosis induced by cross-linking with anti-CD16 mAbin a dose-dependent fashion (Fig. 6B). Furthermore, we found thatL-NMA significantly enhanced apoptosis induced by either cocul-ture with K562 cells or CD16 cross-linking (Fig. 6,A and B).However, treatment with SNAP orL-NMA had no effect on DNAfragmentation of freshly isolated NK cells or IL-2-preactivated NKcells in the absence of target cells (data not shown). These datasuggest that endogenous NO production is involved in protection
FIGURE 3. NO production by human NK cells was assessed by theGriess reaction using nitrate reductase. Purified NK cells were first culturedfor 3 days in the presence or absence of IL-2, and following this culture,cells from each group were then stimulated by coculture with K562 cells(ratio of 10:1, NK cells:K562) or by CD16 or CD56 cross-linking, each for24 h. NO could not be detected above baseline in supernatants of K562cells cultured alone for 24 h. NO production was observed as nitrite/nitratein the supernatants of NK cells stimulated by coculture with K562 targetcells or by ligation of CD16, but not by ligation of CD56. NO productionwas further enhanced by the presence of IL-2 during the cellular activationstep. Data are representative of experiments using cells from three differentdonors.
FIGURE 4. Inhibitory effect ofL-NMA on cytotoxicity of purified NKcells against four different target (K562, Molt-4, U937, and M14) cells wasevaluated by51Cr release assay. Purified NK cells were activated with 100IU/ml rIL-2 for 2 days, treated with the indicated doses ofL-NMA in thepresence of 100 IU/ml rIL-2 for an additional 24 h, and assayed for cyto-toxic activity. Results show means6 SE of triplicate samples of a repre-sentative test from five different donors and demonstrate thatL-NMA par-tially inhibits killing activity of NK cells in a dose-dependent manner.
1476 ENDOTHELIAL NOS IN HUMAN NK CELLS
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from
of NK cells from apoptosis induced by stimulation through CD16or with target cells. The enhancement of apoptosis in NK cells byL-NMA may explain the finding thatL-NMA moderately decreasedcytotoxicity of NK cells.
L-NMA increased TNF-a mRNA expression following ligation ofCD16, and SNAP decreased it
Jewett et al. reported that endogenous TNF-a production by NKcells results in functional anergy and apoptosis in anti-CD16-treated NK cells (44). Therefore, we examined whethertreatment withL-NMA or SNAP alters TNF-a production by anti-CD16-treated NK cells, using treatment with anti-CD56 mAb as acontrol. When 500mM L-NMA was added to IL-2-pretreated NKcells 4 h before RNA isolation, induction of TNF-a mRNA fol-lowing CD16 cross-linking was enhanced, as shown by RT-PCRanalysis (Fig. 7). Conversely, treatment of IL-2-preactivated NKcells with 1 mM SNAP for 4 h suppressed expression of TNF-amRNA following treatment with anti-CD16 mAb (Fig. 7). Phos-phorImager (Molecular Dynamics, Sunnyvale, CA) analyses ofTNF-a mRNA showed thatL-NMA treatment induced approxi-mately a 7-fold increase in expression of TNF-a mRNA as com-pared with anti-CD16 alone, and that SNAP reduced expression byone-fifth as compared with anti-CD16 alone. Treatment with Ab toCD56, used as a control, had no effect on TNF-a mRNA expres-sion, and SNAP orL-NMA did not affect TNF-a mRNA expres-sion in anti-CD56 mAb-treated NK cells (data not shown). Theseresults suggest that NO is involved in regulation of TNF-a tran-
scription in activated NK cells. We next investigated the effect ofaddition of rTNF-a on SNAP-induced suppression of apoptosis ofanti-CD16-treated NK cells. The result, shown in Fig. 8, demon-strated that NK cells underwent apoptosis following treatment withrTNF-a in a dose-dependent fashion, although the percentage ofapoptosis was lower than that induced by CD16 ligation even with100 ng/ml rTNF-a. The data further show that 10 ng/ml rTNF-acompletely reversed the SNAP-induced (1 mM) suppression ofanti-CD16-induced apoptosis in NK cells. Together, these datasupport the conclusion that the effect of SNAP is through suppres-sion of TNF-a, and suggest the possibility that inhibition of TNF-aexpression by NO is a mechanism by which NO can protect acti-vated NK cells from apoptosis.
SNAP inhibited andL-NMA enhanced DNA-binding activity ofthe NF-AT in anti-CD16-treated NK cells
TNF-a gene transcription involves activation of a transcriptionfactor, NF-AT, in T cells as well as NK cells (34, 45). Because wefound that inhibition or replacement of NO affected TNF-a geneexpression in activated NK cells, we hypothesized that NO may
FIGURE 5. No effect ofL-NMA on granule exocytosis (A) or inductionof FasL expression (B) upon NK activation.A, Purified NK cells culturedwith 100 IU/ml rIL-2 for 3 days were tested for BLT-esterase release aftercontact with K562 cells (ratio of 10:1, NK cells:K562). Means6 SD ofthree independent experiments are shown.B, Purified NK cells were acti-vated with 100 IU/ml rIL-2 for 3 days, followed by treatment with 10mg/ml anti-CD16 mAb and 20mg/ml GaM Ab in the presence of KB8301for an additional 6 h. The expression of FasL was measured by flow cy-tometry. The mean fluorescence intensity (MFI)6 SD of samples stainedwith biotinylated FasL mAb (shaded column) or biotinylated isotype con-trol mAb (dotted column) from tests on three different donors is shown.
FIGURE 6. Augmentation of apoptosis of NK cells by treatment withL-NMA and its suppression by SNAP.A, Purified NK cells were stimulatedwith 100 IU/ml rIL-2 for 60 h, followed by labeling with [3H]TdR for 12 hand coculture with K562 cells (ratio of 10:1, NK cells:K562) in the pres-ence of 500mM L-NMA or the indicated doses of SNAP for an additional4 h. The percentage of fragmented DNA in NK cells was measured by JAMtest. Means6 SD at E:T ratio of 3:1 of three independent experiments areshown.B, Purified NK cells activated with 100 IU/ml rIL-2 for 3 days weretreated with 10mg/ml anti-CD16 mAb and 20mg/ml GaM Ab in presenceof 500mM L-NMA or the indicated doses of SNAP for 4 h. The percentageof apoptotic cells was assessed by staining with AO and EB. Means6 SDamong experiments on cells from four different donors are shown. Statis-tically significant differences between with and withoutL-NMA were de-termined in both experiments (A, p, p , 0.005;B, pp, p , 0.01).
1477The Journal of Immunology
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from
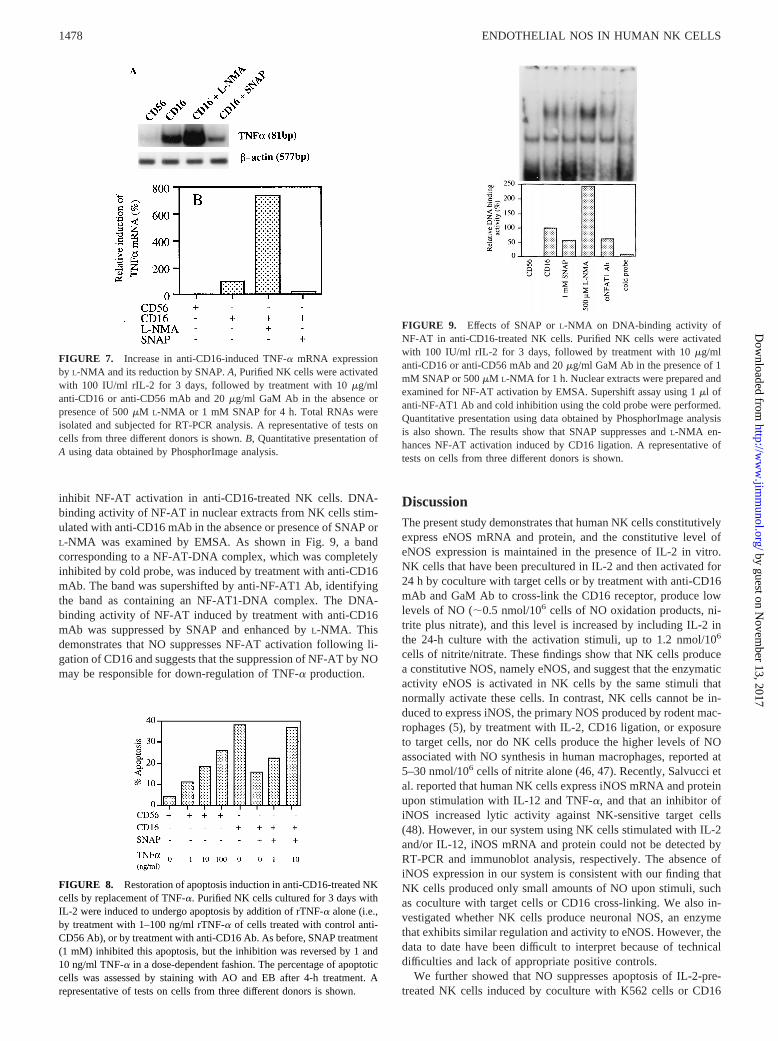
inhibit NF-AT activation in anti-CD16-treated NK cells. DNA-binding activity of NF-AT in nuclear extracts from NK cells stim-ulated with anti-CD16 mAb in the absence or presence of SNAP orL-NMA was examined by EMSA. As shown in Fig. 9, a bandcorresponding to a NF-AT-DNA complex, which was completelyinhibited by cold probe, was induced by treatment with anti-CD16mAb. The band was supershifted by anti-NF-AT1 Ab, identifyingthe band as containing an NF-AT1-DNA complex. The DNA-binding activity of NF-AT induced by treatment with anti-CD16mAb was suppressed by SNAP and enhanced byL-NMA. Thisdemonstrates that NO suppresses NF-AT activation following li-gation of CD16 and suggests that the suppression of NF-AT by NOmay be responsible for down-regulation of TNF-a production.
DiscussionThe present study demonstrates that human NK cells constitutivelyexpress eNOS mRNA and protein, and the constitutive level ofeNOS expression is maintained in the presence of IL-2 in vitro.NK cells that have been precultured in IL-2 and then activated for24 h by coculture with target cells or by treatment with anti-CD16mAb and GaM Ab to cross-link the CD16 receptor, produce lowlevels of NO (;0.5 nmol/106 cells of NO oxidation products, ni-trite plus nitrate), and this level is increased by including IL-2 inthe 24-h culture with the activation stimuli, up to 1.2 nmol/106
cells of nitrite/nitrate. These findings show that NK cells producea constitutive NOS, namely eNOS, and suggest that the enzymaticactivity eNOS is activated in NK cells by the same stimuli thatnormally activate these cells. In contrast, NK cells cannot be in-duced to express iNOS, the primary NOS produced by rodent mac-rophages (5), by treatment with IL-2, CD16 ligation, or exposureto target cells, nor do NK cells produce the higher levels of NOassociated with NO synthesis in human macrophages, reported at5–30 nmol/106 cells of nitrite alone (46, 47). Recently, Salvucci etal. reported that human NK cells express iNOS mRNA and proteinupon stimulation with IL-12 and TNF-a, and that an inhibitor ofiNOS increased lytic activity against NK-sensitive target cells(48). However, in our system using NK cells stimulated with IL-2and/or IL-12, iNOS mRNA and protein could not be detected byRT-PCR and immunoblot analysis, respectively. The absence ofiNOS expression in our system is consistent with our finding thatNK cells produced only small amounts of NO upon stimuli, suchas coculture with target cells or CD16 cross-linking. We also in-vestigated whether NK cells produce neuronal NOS, an enzymethat exhibits similar regulation and activity to eNOS. However, thedata to date have been difficult to interpret because of technicaldifficulties and lack of appropriate positive controls.
We further showed that NO suppresses apoptosis of IL-2-pre-treated NK cells induced by coculture with K562 cells or CD16
FIGURE 7. Increase in anti-CD16-induced TNF-a mRNA expressionby L-NMA and its reduction by SNAP.A, Purified NK cells were activatedwith 100 IU/ml rIL-2 for 3 days, followed by treatment with 10mg/mlanti-CD16 or anti-CD56 mAb and 20mg/ml GaM Ab in the absence orpresence of 500mM L-NMA or 1 mM SNAP for 4 h. Total RNAs wereisolated and subjected for RT-PCR analysis. A representative of tests oncells from three different donors is shown.B, Quantitative presentation ofA using data obtained by PhosphorImage analysis.
FIGURE 8. Restoration of apoptosis induction in anti-CD16-treated NKcells by replacement of TNF-a. Purified NK cells cultured for 3 days withIL-2 were induced to undergo apoptosis by addition of rTNF-a alone (i.e.,by treatment with 1–100 ng/ml rTNF-a of cells treated with control anti-CD56 Ab), or by treatment with anti-CD16 Ab. As before, SNAP treatment(1 mM) inhibited this apoptosis, but the inhibition was reversed by 1 and10 ng/ml TNF-a in a dose-dependent fashion. The percentage of apoptoticcells was assessed by staining with AO and EB after 4-h treatment. Arepresentative of tests on cells from three different donors is shown.
FIGURE 9. Effects of SNAP orL-NMA on DNA-binding activity ofNF-AT in anti-CD16-treated NK cells. Purified NK cells were activatedwith 100 IU/ml rIL-2 for 3 days, followed by treatment with 10mg/mlanti-CD16 or anti-CD56 mAb and 20mg/ml GaM Ab in the presence of 1mM SNAP or 500mM L-NMA for 1 h. Nuclear extracts were prepared andexamined for NF-AT activation by EMSA. Supershift assay using 1ml ofanti-NF-AT1 Ab and cold inhibition using the cold probe were performed.Quantitative presentation using data obtained by PhosphorImage analysisis also shown. The results show that SNAP suppresses andL-NMA en-hances NF-AT activation induced by CD16 ligation. A representative oftests on cells from three different donors is shown.
1478 ENDOTHELIAL NOS IN HUMAN NK CELLS
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from

cross-linking. This effect on apoptosis is associated with alterationin lytic activity, such that suppression of NO production partiallyinhibits NK lytic activity without affecting known lytic mecha-nisms, namely granule exocytosis and FasL expression. Stimula-tion of NK cells with either K562 or anti-CD16 mAb has beenshown to elicit intracellular Ca21 elevation (49). ConstitutiveeNOS is known to be activated for short periods of time by Ca21-elevating stimuli and to be responsible for the low levels of NOthat are involved in homeostatic processes, such as in blood vessels(4, 9). Moreover, addition of exogenous NO appears to regulateactivity of Janus kinase 3 immunoprecipitated from an NK cell line(50). Our findings, showing that an inhibitor of NOS,L-NMA,enhanced apoptosis and inhibited cytotoxicity by human NK cells,suggest that NO produced through eNOS activation is involved insuppression of activation-induced cell death and maintenance ofcytotoxicity mediated by NK cells. Therefore, our data suggest thateNOS may play a homeostatic role in the maintenance of a min-imum level of activity by activated NK cells through inhibitingtheir apoptosis.
Although NO has been reported to inhibit apoptosis in variouscells, including human EBV-transformed B cells (38), human Tcells (39), rat hepatocytes (40), sheep endothelial cells (41), andhuman eosinophils (51), the underlying mechanism remains un-clear. It is reported that endogenous TNF-a is involved in andrequired for apoptosis in NK cells activated with anti-CD16 mAb(44) or cytokines including IL-2 and IL-15 (52). We hypothesizedthat inhibition of transcription of TNF-a may be a mechanism ofsuppression of apoptosis in activated NK cells by NO. Our datashowed that inhibition of NOS activity augmented TNF-a produc-tion, and SNAP, an NO donor, decreased it, suggesting that NOindeed suppresses TNF-a mRNA expression in anti-CD16-treatedNK cells. This hypothesis was supported by the demonstration thataddition of rTNF-a reversed suppression by NO of apoptosis inanti-CD16 mAb-treated NK cells. A nuclear factor, NF-AT, isknown to be involved in TNF-a induction following CD16 cross-linking in NK cells (34). Our study revealed that SNAP treatmentsuppressed DNA-binding activity of NF-AT in NK cells activatedby anti-CD16 mAb, suggesting further that NO inhibits TNF-ainduction in activated NK cells by suppression of transcriptionthrough NF-AT activation. The nuclear translocation of NF-AT isknown to be regulated by the serine/threonine phosphatase, cal-cineurin, which contains Fe in its active site (53, 54). Because NOis highly reactive with transition metals, including Fe (3), it ispossible that NO may modulate NF-AT activity through an effecton calcineruin.
Although NF-AT has been recently reported to be involved inFasL expression in activated human T cells (55), our data did notshow any alteration byL-NMA in FasL expression in anti-CD16-activated NK cells. Because apoptosis of NK cells can also occurin part through Fas/FasL interaction (28, 56), NO may be involvedin modulation of this pathway. In fact, Fas-induced apoptosis ofJurkat T cells is reported to be blocked by treatment with SNAP(39), and a disruption by NO of Fas receptor signaling in humaneosinophils has been proposed (51), and NOS activity has beenreported to inhibit Fas-induced apoptosis in a number of humanleukocytic cell lines (57). However, our data suggest that NO maynot play a major role in the Fas-FasL-mediated pathway of NKcell-mediated cytotoxicity because FasL expression was not al-tered byL-NMA treatment. In addition, we observed thatL-NMAtreatment had no effect on apoptosis of purified NK cells inducedby agonist anti-Fas Ab (data not shown).
A number of reports demonstrate that NO inhibits the activationof NF-kB, which regulates transcription of a number of immuno-logically crucial genes (42, 58). Therefore, we investigated the
effect of SNAP on DNA-binding activity of NF-kB in anti-CD16mAb-treated NK cells. Activation of NF-kB in NK cells in re-sponse to anti-CD16 mAb occurred only after 4 h of stimulationwith anti-CD16 mAb, much more slowly than in response to otherstimuli, such as TNF-a and PMA, and was inhibited by SNAP(data not shown). This delayed kinetics of NF-kB activation maybe explained by the hypothesis that NF-kB may be activated afterCD16 ligation through synthesis of endogenous TNF-a followingNF-AT activation.
This is a first report that human NK cells express eNOS and thatNO modulates NF-AT activation. In addition, our results suggest aphysiological role for NO production, i.e., protection of NK cellsfrom apoptosis to maintain a level of cytotoxic activity. Finally,our data suggest a possible mechanism by which this protectionoccurs, namely inhibition of TNF-a transcription by inactivationof NF-AT. We hypothesize that endogenous NO production maybe important in vivo to maintain a basal level of activated NK cellactivity by protecting such cells from apoptosis (Fig. 10).
AcknowledgmentsWe thank Amgen for supplying human rIL-2, Dr. J. Yodoi for the kind giftof the YT cell line, and Dr. J. Kornbluth for providing the NK3.3 cell line.We thank Drs. Giovanna Tosato and Mark Hayes for critical review of themanuscript.
References1. Schmidt, H. H. H. W., and U. Walter. 1994. NO at work.Cell 78:919.2. Nathan, C. Z. 1992. Nitric oxide as a secretory product of mammalian cells.
FASEB J. 6:3051.3. Stamler, J. S. 1994. Redox signaling: nitrosylation and related target interactions
of nitric oxide.Cell 78:931.4. Nathan, C., and Q.-w. Xie. 1994. Nitric oxide synthases: roles, tolls, and controls.
Cell 78:915.5. MacMicking, J., Q.-w. Xie, and C. Nathan. 1997. Nitric oxide and macrophage
function.Annu. Rev. Immunol. 15:323.6. Reiling, N., A. J. Ulmer, M. Duchrow, M. Ernst, H.-D. Flad, and S. Hauschildt.
1994. Nitric oxide synthase: mRNA expression of different isoforms in humanmonocytes/macrophages.Eur. J. Immunol. 24:1941.
7. Weinberg, J. B., M. A. Misukonis, P. J. Shami, S. N. Mason, D. L. Sauls,W. A. Dittman, E. R. Wood, G. K. Smith, B. McDonald, K. E. Bachus, et al.1995. Human mononuclear phagocyte inducible nitric oxide synthase (iNOS):analysis of iNOS mRNA, iNOS protein, biopterin, and nitric oxide production byblood monocytes and peritoneal macrophages.Blood 86:1184.
FIGURE 10. Diagrammatic representation of our hypothesis that in NKcells activated through CD16 ligation or coculture with target cells, en-dogenous NO, produced in response to activation of eNOS, may suppressactivation-induced apoptosis in human NK cells through inhibition ofNF-AT activation and subsequent TNF-a production. We further speculatethat this mechanism may lead to survival of some level of activated NKcells, thereby maintaining a basal level of activity.
1479The Journal of Immunology
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from

8. Bukrinsky, M. I., H. S. L. M. Nottet, H. Schmidtmayerova, L. Dubrovsky,C. R. Flanagan, M. E. Mullins, S. A. Lipton, and H. E. Gendelman. 1995. Reg-ulation of nitric oxide synthase activity in human immunodeficiency virus type 1(HIV-1)-infected monocytes: implications for HIV-associated neurological dis-ease.J. Exp. Med. 181:735.
9. Marletta, M. A. 1994. Nitric oxide synthase: aspects concerning structure andcatalysis.Cell 78:927.
10. Lotzova, E. 1989. Analysis of effector mechanisms in cancer.Curr. Opin. Im-munol. 1:904.
11. Garrido, F., F. Ruiz-Cabello, T. Cabrera, J. J. Perez-Villar, M. Lopez-Botet,M. Duggan-Keen, and P. L. Stern. 1997. Implications for immunosurveillance ofaltered HLA class I phenotypes in human tumors.Immunol. Today 18:89.
12. Biron, C. A. 1997. Activation and function of natural killer cell responses duringviral infections.Curr. Opin. Immunol. 9:24.
13. Yamauchi, A., and E. T. Bloom. 1997. Control of cell cycle progression humannatural killer cells through redox regulation of expression and phosphorylation ofretinoblastoma gene product protein.Blood 89:4092.
14. Yamauchi, A., and E. T. Bloom. 1993. Requirement of thiol compounds as re-ducing agents for IL-2-mediated induction of LAK activity and proliferation ofhuman NK cells.J. Immunol. 151:5535.
15. Buttke, T. M., and P. A. Sandstrom. 1994. Oxidative stress as a mediator ofapoptosis.Immunol. Today 15:7.
16. Powis, G., J. R. Gasdaska, and A. Baker. 1997. Redox signaling and the controlof cell growth and death.Adv. Pharmacol. 38:329.
17. Furuke, K., and E. T. Bloom. 1998. Redox-sensitive events in Fas-induced ap-optosis in human natural killer cells include ceramide generation and proteintyrosine dephosphorylation.Int. Immunol. 10:1261.
18. Ito, M., M. Watanabe, H. Kamiya, and M. Sakurai. 1996. Inhibition of naturalkiller (NK) cell activity against varicella-zoster virus (VZV)-infected fibroblastsand lymphocyte activation in response to VZV antigen by nitric oxide-releasingagents.Clin. Exp. Immunol. 106:40.
19. Ito, M., M. Watanabe, H. Kamiya, and M. Sakurai. 1996. Inhibition of naturalkiller cell activity against cytomegalovirus-infected fibroblasts by nitric oxide-releasing agents.Cell. Immunol. 174:13.
20. Filep, J. G., C. Baron, S. Lahance, C. Perreault, and J. S. D. Chan. 1996. In-volvement of nitric oxide in target-cell lysis and DNA fragmentation induced bymurine natural killer cells.Blood 87:5136.
21. Hori, K., P. R. Burd, K. Furuke, J. Kutza, K. A. Weih, and K. A. Clouse. 1999.Human immunodeficiency virus-1-infected macrophages induce inducible nitricoxide synthase and nitric oxide (NO) production in astrocytes: astrocytic NO asa possible mediator of neural damage in acquired immunodeficiency syndrome.Blood 93:1843.
22. Burd, P. R., W. C. Thompson, E. E. Max, and F. C. Mills. 1995. Activated mastcells secrete interleukin-13.J. Exp. Med. 181:1373.
23. Ding, M., B. A. St. Pierre, J. F. Parkinson, P. Medberry, J. L. Wong,N. E. Rogers, L. J. Ignarro, and J. E. Merrill. 1997. Inducible nitric-oxide syn-thase and nitric oxide production in human fetal astrocytes and microglia.J. Biol.Chem. 272:11327.
24. Green, L. C., D. A. Wagner, J. Glogowski, P. L. Skipper, J. S. Wishnock, andS. R. Tannenbaum. 1982. Analysis of nitrate, nitrite, and (15N) nitrate in biolog-ical fluids.Anal. Biochem. 126:131.
25. Bloom, E. T., and E. L. Korn. 1983. Quantification of natural cytotoxicity byhuman lymphocyte subpopulations isolated by density: heterogeneity of the ef-fector cells.J. Immunol. Methods 58:323.
26. Duke, R. C., and J. J. Cohen. 1992. Morphological and biological assays ofapoptosis. InCurrent Protocols in Immunology. J. E. Coligan, A. M. Kruisbeek,D. H. Margulies, E. M. Shevach, and W. Strober, eds. Greene/Wiley-Interscience,New York, p. 3.17.1.
27. Matzinger, P. 1991. The JAM test: a simple assay for DNA fragmentation andcell death.J. Immunol. Methods 145:185.
28. Yamauchi, A., K. Taga, H. S. Mostowski, and E. T. Bloom. 1996. Target cell-induced apoptosis of interleukin-2-activated human natural killer cells: roles ofcell surface molecules and intracellular events.Blood 87:5127.
29. Taffs, R., and M. Sitkovsky. 1991. Granule enzyme exocytosis assay for cyto-toxic T lymphocyte activation. In Current Protocols in Immunology.J. E. Coligan, A. M. Kruisbeek, D. H. Margulies, E. M. Shevach, and W. Strober,eds. Greene/Wiley-Interscience, New York, p. 3.16.1.
30. Furuke, K., M. Shiraishi, H. S. Mostowski, and E. T. Bloom. 1999. Fas ligandinduction in human NK cells is regulated by redox through a calcineurin-nuclearfactors of activate T cell-dependent pathway.J. Immunol. 162:1988.
31. Kayagaki, N., A. Kawasaki, T. Ebata, H. Ohmoto, S. Ikeda, S. Inoue, K. Yoshino,K. Okumura, and H. Yagita. 1995. Metalloproteinase-mediated release of humanFas ligand.J. Exp. Med. 182:1777.
32. Schreiber, E., P. Matthias, M. M. Muller, and W. Schaffner. 1989. Rapid detec-tion of octamer binding proteins with ‘mini-extracts,’ prepared from a smallnumber of cells.Nucleic Acids Res. 17:6419.
33. Jain, J., Z. Miner, and A. Rao. 1993. Analysis of the preexisting and nuclearforms of nuclear factor of activated T cells.J. Immunol. 151:837.
34. Aramburu, J., L. Azzoni, A. Rao, and B. Perussia. 1995. Activation and expres-sion of the nuclear factors of activated T cells, NFATp and NFATc, in humannatural killer cells: regulation upon CD16 ligand binding.J. Exp. Med. 182:801.
35. Cifone, M. G., C. Festuccia, L. Cironi, G. Cavallo, M. A. Chessa, V. Pensa,E. Tubaro, and A. Santoni. 1994. Induction of the nitric oxide-synthesizing path-way in fresh and interleukin 2-cultured rat natural killer cells.Cell. Immunol.157:181.
36. Montel, A. H., M. R. Bochan, W. S. Goebel, and Z. Brahmi. 1995. Fas-mediatedcytotoxicity remains intact in perforin and granzyme B antisense transfectants ofhuman NK-like cell line.Cell. Immunol. 165:312.
37. Nakazawa, T., K. Agematsu, and A. Yabuhara. 1997. Later development of Fasligand-mediated cytotoxicity as compared with granule-mediated cytotoxicityduring the maturation of natural killer cells.Immunology 92:180.
38. Mannick, J. B., K. Asano, K. Izumi, E. Kieff, and J. S. Stamler. 1994. Nitric oxideproduced by human B lymphocytes inhibits apoptosis and Epstein-Barr virusreactivation.Cell 79:1137.
39. Melino, G., F. Bernassola, R. A. Knight, M. T. Corasaniti, G. Nistico, and A.Finazzi-Agro. 1997.S-nitrosylation regulates apoptosis.Nature 388:432.
40. Kim, Y. M., M. E. de Vera, S. C. Watkins, and T. R. Billiar. 1997. Nitric oxideprotects cultured rat hepatocytes from tumor necrosis factor-a-induced apoptosisby inducing heat shock protein 70 expression.J. Biol. Chem. 272:1402.
41. Tzeng, E., Y. M. Kim, B. R. Pitt, A. Lizonova, I. Kovesdi, and T. R. Billiar. 1997.Adenoviral transfer of the inducible nitric oxide synthase gene blocks endothelialcell apoptosis.Surgery 122:255.
42. Matthews, J. R., C. H. Botting, M. Panico, H. R. Morris, and R. T. Hay. 1996.Inhibition of NF-kB DNA binding by nitric oxide.Nucleic Acids Res. 24:2236.
43. Taga, K., A. Yamauchi, E. T. Bloom, and G. Tosato. 1996. Target-induced deathby apoptosis in human lymphokine-activated natural killer cells.Blood 87:2411.
44. Jewett, A., M. Cavalcanti, and B. Bonavida. 1997. Pivotal role of endogenousTNF-a in the induction of functional inactivation and apoptosis in NK cells.J. Immunol. 159:4815.
45. McCaffrey, P. G., A. E. Goldfeld, and A. Rao. 1994. The role of NFATp incyclosporin A-sensitive tumor necrosis factor-a gene transcription.J. Biol. Chem.269:30445.
46. Martin, J. H. J., and S. W. Edwards. 1993. Changes in mechanisms of monocyte/macrophage-mediated cytotoxicity during culture.J. Immunol. 150:3478.
47. De Maria, R., M. G. Cifone, R. Trotta, M. R. Rippo, C. Festuccia, A. Santoni, andR. Testi. 1994. Triggering of human monocyte activation through CD69, a mem-ber of the natural killer cell gene complex family of signal transducing receptors.J. Exp. Med. 180:1999.
48. Salvucci, O., J. P. Kolb, B. Dugas, N. Dugas, and S. Chouaib. 1998. The induc-tion of nitric oxide by interleukin-12 and tumor necrosis factor-a in human nat-ural killer cells: relationship with the regulation of lytic activity.Blood 92:2093.
49. Leibson, P. J. 1997. Signal transduction during natural killer cell activation: in-side the mind of a killer.Immunity 6:655.
50. Duhe, R. J., G. A. Evans, R. A. Erwin, R. A. Kirken, G. W. Cox, andW. L. Farrar. 1998. Nitric oxide and thiol redox regulation of Janus kinase ac-tivity. Proc. Natl. Acad. Sci. USA 95:126.
51. Hebestreit, H., B. Dibbert, I. Balatti, D. Braun, A. Schapowal, K. Blaser, andH.-U. Simon. 1998. Disruption of Fas receptor signaling by nitric oxide in eo-sinophils.J. Exp. Med. 187:415.
52. Ross, M. E., and M. A. Caligiuri. 1997. Cytokine-induced apoptosis of humannatural killer cells identifies a novel mechanism to regulate the innate immuneresponse.Blood 89:910.
53. Kissinger, C. R., H. E. Parge, D. R. Knighton, C. T. Lewis, L. A. Pelletier,A. Tempczyk, V. J. Kalish, K. D. Tucker, R. E. Showalter, E. W. Moomaw, etal. 1995. Crystal structures of human calcineurin and the human FKBP12-FK506-calcineurin complex.Nature 378:641.
54. Wang, X., V. C. Culotta, and C. B. Klee. 1996. Superoxide dismutase protectscalcineurin from inactivation.Nature 383:434.
55. Latinis, K. M., L. A. Norian, S. L. Eliason, and G. A. Koretzky. 1997. Two NFATtranscription factor binding sites participate in the regulation of CD95 (Fas) li-gand expression in activated human T cells.J. Biol. Chem. 272:31427.
56. Eischen, C. M., J. D. Schilling, D. H. Lynch, P. H. Krammer, and P. J. Leibson.1996. Fc receptor-induced expression of Fas ligand on activated NK cells facil-itates cell-mediated cytotoxicity and subsequent autocrine NK cell apoptosis.J. Immunol. 156:2693.
57. Mannick, J. B., X. Q. Miao, and J. S. Stamler. 1997. Nitric oxide inhibits Fas-induced apoptosis.J. Biol. Chem. 272:24125.
58. Baldwin, A. S., Jr. 1996. The NF-kB and IkB proteins: new discoveries andinsights.Annu. Rev. Immunol. 14:649.
1480 ENDOTHELIAL NOS IN HUMAN NK CELLS
by guest on Novem
ber 13, 2017http://w
ww
.jimm
unol.org/D
ownloaded from


















