
3RD SEMINAR INNATE IMMUNITY: ANTIVIRAL STATE, KILLER CELLS, THE COMPLEMENT SYSTEM.

Umeå University, Medical Dissertations
New Series No. 1242-ISSN 0346-6612-ISBN 978-91-7264-721-3
Editor: The Dean of the Faculty of Medicine
From the Departments of Clinical Microbiology, Immunology and Clinical Sciences, Pediatrics
Umeå University, Umeå, Sweden
Human Intestinal Epithelial Cells in Innate Immunity
Interactions with normal microbiota and pathogenic bacteria
by
Gangwei Ou
Umeå 2009

ii
Copyright©Gangwei Ou ISBN: 978-91-7264-721-3 ISSN: 0346-6612, New Series nr: 1242 Tryck/Printed by: Arkitektkopia Umeå, Sweden 2009

Table of Contents
List of Papers…………………………………………………….…. 2 Abstract…………………………………………………………….. 3 Abbreviations……………………………………………………….. 4 1. Introduction 5 1.1 The human intestine and mucosal immunity…………………6 1.2 Hurdles for intestinal pathogens…………………………….. 8 1.3 Innate immunity...…………………………………………… 10 1.3.1 TLRs and their signaling……………………………… 10 1.3.2 TLR-independent pathogen recognition……………….12 1.3.3 TLR and NOD signaling in the human intestine………13 1.4 Mucins………………………………………………………. 14 1.5 The carcinoembryonic antigen family……………………… 15 1.6 Antimicrobial peptides and lysozyme……………………… 17 1.7 Cytokines…………………………………………………… 20 1.7.1 Interferon-γ..….………………………………………. 21 1.7.2 Tumor necrosis factor-α.…..………………………… 22 1.7.3 Interleukin-1β………………………………………… 23 1.7.4 Interleukin-6…………………………………………. 24 1.7.5 Interleukin-8…………………………………………. 25 1.8 Intestinal microflora……………………………………….. 26 1.9 Vibrio cholerae……………………………………………. 28 1.10 Celiac disease…………………………………………….. 31 2. Aims of this Thesis………………………………………………. 36 3. Results and Discussion…………………………………………... 37 3.1 Proximal small intestinal microbiota and identification of
rod-shaped bacteria associated with childhood celiac disease (Paper I)………………………………………………………...
37
3.2 Contribution of intestinal epithelial cells to innate immunity of the human gut - studies on polarized monolayers of colon carcinoma cells (Paper II)…………………………….
43
3.3 Role of V. cholerae protease, PrtV - from C. elegans to human intestine………………………………………………...
45
3.3.1 PrtV protects V. cholerae from natural predator grazing and is responsible for killing of infected C. elegans (Paper III)…………………………………………………..
45
3.3.2 V. cholera cytolysin (VCC) is the secreted proinflammatory factor and its activity is modulated by PrtV (Paper IV)…………………………………………….
45
4. Conclusions……………………………………………………… 48 5. Acknowledgements……………………………………………... 49 6. References……………………………………………………….. 51 7. Papers I – IV...………………………………………Appendix I - IV

- 2 -
List of Papers
This thesis is based on the following papers referred to in the text by their roman numbers (I-IV).
I. Ou G, Hedberg M, Hörstedt P, Baranov V, Forsberg G, Drobni M, Sandström O, Wai SN, Johansson I, Hammarström ML, Hernell O, Hammarström S. Proximal Small Intestinal Microbiota and Identification of Rod-shaped Bacteria associated with Childhood Celiac Disease. 2009. (submitted)
II. Ou G, Baranov V, Lundmark E, Hammarström S, Hammarström ML. 2009. Contribution of intestinal epithelial cells to innate immunity of the human gut - studies on polarized monolayers of colon carcinoma cells. Scand J Immunol. 69:150-61.
III. Vaitkevicius K, Lindmark B, Ou G, Song T, Toma C, Iwanaga M, Zhu J, Andersson A, Hammarström ML, Tuck S, Wai SN. 2006. A Vibrio cholerae protease needed for killing of Caenorhabditis elegans has a role in protection from natural predator grazing. Proc Natl Acad Sci U S A. 103:9280-5.
IV. Ou G, Rompikuntal PK, Bitar A, Lindmark B, Vaitkevicius K,
Bhakdi S, Wai SN, Hammarström ML. Vibrio cholerae cytolysin causes an inflammatory response in human intestinal epithelial cells that is modulated by the protease PrtV secreted by the same bacterium. 2009. (submitted)

- 3 -
Abstract Rod-shaped bacteria were previously shown to be associated with the small intestinal epithelium of children with celiac disease (CD). Using culture-dependent and independent methods, we characterized the microbiota of small intestine in children with CD and controls. The normal microbiota constitutes an unique organ-specific biofilm. Dominant bacteria are Streptococcus, Neisseria, Veillonella, Gemella, Actinomyces, Rothia and Haemophilus. Altogether 162 Genus Level Operational Taxonomic Units (GELOTU) of six different phyla were identified in a total of 63 children. In biopsies collected during 2004-2007 we did not find major differences in the microbiota between CD patients and controls. However, in biopsies collected earlier from children born during the “Swedish CD epidemic” and demonstrated to have rod-shaped bacteria by electron microscopy, we found that unclassified-Clostridales and Prevotella species were associated with CD. These anaerobic, rod-shaped bacteria showed marked affinity for the intestinal epithelium. Changes in breast-feeding practice and/or regiments for introduction of gluten containing food probably affect the composition of the bacterial flora in small intestine. We hypotesize that these bacteria contribute to contraction of CD.
An in vitro model for studies of immune mechanisms of the intestinal epithelium was established. Polarized tight monolayers of the human colon carcinoma cell lines, T84 and Caco2, were developed by culture in a two-chamber system. The two cell lines showed the features of mature- and immature columnar epithelial cells respectively. Polarized monolayers were challenged with bacteria and proinflammatory cytokines. Immune responses were estimated as quantitative changes in mRNA expression levels of a secreted mucin (MUC2), glycocalyx components (CEACAMs, MUC3), antimicrobial factors and cytokines (IFN-γ, TNF-α, IL-6 and IL-8). Tight monolayer cells were more resistant to bacterial attack than ordinary tissue culture cells and only B. megaterium induced the defensin, hBD2. Tight monolayer cells responded to cytokine challenge suggesting awareness of basolateral attack. TNF-α induced markedly increased levels of IL-8 and TNF-α itself in both cell lines suggesting recruitment and activation of immune cells. Cytokine challenge also increased levels of CEACAM1, which includes two functionally different forms, CEACAM1-L and CEACAM1-S. In T84 cells, IFN-γ was selective for CEACAM1-L while TNF-α upregulated both forms. Increased CEACAM1 expression may influence epithelial function and communication between epithelial cells and intraepithelial lymphocytes.
As a pathogenic enteric bacterium, Vibrio cholerae secretes cholera toxin that is the major factor of cholera diarrhea. However, some strains of O1 serogroup lacking the cholera toxin still cause enterocolitis and most V. cholerae vaccines candidates exhibit reactogenicity in clinical trails. An extracellular metalloprotease PrtV was characterized. It was associated with killing of bacteria predators such as the nematode Caenorhabditis elegans. Its role in human intestine was addressed by using the T84 tight monolayer in vitro model. We found that Vibrio Cholera Cytolysin (VCC), a pore-forming toxin, induces an inflammatory response in intestinal epithelial cells that includes increased epithelial permeability and induction of IL-8 and TNF-α and hence could be responsible for enterocolitis. The inflammatory response was abolished by PrtV thus VCC is indeed an autologous substrate for PrtV. In protein rich environment PrtV degradation of VCC was inhibited, suggesting that the magnitude of the inflammatory response is modulated by the milieu in the small intestine. Thus, VCC is likely to be part of the pathogenesis of cholera diarrhea and the causative agent of enteropathy in V. cholerae strains lacking the cholera toxin.

- 4 -
Abbreviations
AMP Antimicrobial peptide CD Celiac disease CEA Carcinoembryonic antigen CEACAM Carcinoembryonic antigen cell adhesion molecule CLR C-type lectin receptor CTX Vibrio cholera toxin DC Dendritic cell FDC Follicular dendritic cell HBD Human β defensin HD Human α defensin HLA Human leukocyte antigen IEC Intestinal epithelial cell IEL Intraepithelial lymphocyte IFN Interferon IL Interleukin ITIM Immunoreceptor tyrosine-based inhibition motif LP Lamina propria LPS Lipopolysaccharide LRR Leucin-rich repeat MHC Major histocompatibility complex NF-κB Nuclear factor kappa B NLR NOD-like receptor NOD Nucleotide oligomerization domain PAMP Pathogen associated molecular pattern PP Peyer´s patches PRR Pattern recognition receptor PrtV Protease of V. cholerae RLR Retinoic acid-inducible gene (RIG)-I-like receptor STAT Signal transducer and activator of transcription TLR Toll-like receptor TNF Tumor necrosis factor tTG Tissue transglutaminase VCC Vibrio cholerae cytolysin

- 5 -
1. Introduction
The intestine serves several functions, notably ingestion, digestion and absorption of food and defecation of waste products. It is also the host of the largest and most complex part of the immune system indispensable for the survival of the individual. The intestinal mucosal surface is lined by a single layer of epithelial cells. In humans it covers an area of over 100 m2 that is continuously exposed to different antigens in the form of food constituents, commensal microflora, episodic pathogens, and to noxious compounds. The epithelial cell layer constitutes the interface between the internal milieu and the external environment. It is a highly regulated, selectively permeable barrier that supports digestion and absorption of nutrients, electrolytes, and water from the lumen. It also provides an essential barrier protecting against infection and disease. This barrier is not simply physical, but is part of a complex mucosal immune system (Figure 1). At the apical side of the polarized monolayer, the intestinal epithelium directly confronts both dietary and non-dietary luminal antigens, sensing their signals and subsequently transmitting the information to the diverse populations of cells in the intraepithelial compartment and the underlying tissue, the lamina propria (LP). At the basolateral side the intestinal epithelial cells (IECs) receive signals from various immune cells, nerve cells and stromal cells that influence their barrier function, immune activity state and/or differentiation state.

- 6 -
1.1 The human intestine and mucosal immunity The intestine consists of the small intestine and the large intestine. The small intestine is divided into three anatomic sections: duodenum, jejunum and ileum measuring about 6 meters in total length. The large intestine is about 1.5 meters long and consists of the caecum, appendix, colon and rectum. In cross-section four distinct layers can be distinguished. Counting from the inside they are: mucosa, submucosa, muscularis, and serosa. The mucosa, in turn can be divided into an epithelial compartment, the LP, and muscularis mucosae. Muscularis consists of circular and longitudinal muscles. The mucosa of the small intestine has villi and shallow crypts facing the gut lumen, while the large intestine lacks villi and has deeper crypts. The intestinal villi are finger-like or leaf-like projections. The villi consist of a core of loose connective tissue covered by a single layer columnar epithelium. The core of the villus is an extension of the LP, which contains numerous fibroblasts, smooth muscle cells, lymphocytes, plasma cells, macrophages, dendritic cells (DCs), eosinophils and a network of blood capillaries as well as blind-ending lymphatic capillary, the lacteal. The intestinal crypts are composed of a single layer columnar epithelium that is continuous with the epithelium of the villi. The crypts are simple tubular structures that extend from the muscularis mucosae through the thickness of the LP, where they open to the luminal surface of the intestine at the base of the villi. Additionally, LP is enervated. The epithelial compartment contains epithelial cells and lymphocytes (Figure 2). The epithelial cells are joined firmly together by tight junctions, which create a semipermeable diffusion barrier between individual cells (Liévin-Le Moal and Servin, 2006). At least five types of epithelial cells can be distinguished: enterocytes, goblet cells, Paneth cells, enteroendocrine cells and microfold (M) cells. All epithelial cells, both in the small and large intestine, are derived from stem cells located towards the base of the crypts. In small intestine enterocytes and goblet cells migrate upward from the crypt to the villus and are shed at the tip of the villus after 5 to 6 days. In colon the cells are shed at the free luminal surface. Enterocytes are the predominant cell type of the villus epithelium and are highly specialized for nutrient absorption. At the apical surface, enterocytes are lined by densely packed microvilli (~1 µm long by 0.1 µm in diameter) termed the brush border. Large amounts of digested nutrients can be taken up because the microvilli greatly increase the apical surface area of the plasma membrane. The membrane of the microvilli is rich in protein, cholesterol as well as glycolipids and contains digestive enzymes such as disaccharidases and peptidases. Goblet cells are large mucus-secreting cells interspersed between

- 7 -
enterocytes, both in the crypts and at the villus/luminal surface. Interspersed between the enterocytes are also endocrine cells that secrete neuropeptides such as polypeptide YY and substance P. Paneth cells, only present in the small intestine, are located at the base of the crypts. They contain granules with antimicrobial peptides, antibacterial enzymes as well as antibacterial glycoproteins and most likely play a role in regulating the density and composition of the intestinal bacterial flora. A highly specialized epithelial cell is the M cell, located in the epithelium covering lymphoid follicles. The M cell has an antigen sampling function by transporting antigens from the lumen to the immune cells in the underlying follicles. The enterocytes dominate over goblet cells in small intestine and most of colon. In colon the ratio decreases, approaching 1:1 near the rectum.
Gut associated lymphoid tissue (GALT) is made up of three components: organized lymphoid aggregates [e.g. Peyer’s patches (PPs), the appendix, solitary follicles], diffuse collections of lymphocytes, macrophages, and plasma cells in the LP and intraepithelial lymphocytes (IELs) scattered within the epithelium. PPs are aggregations of lymphoid follicles present in small intestine (Cornes, 1965) while solitary follicles occur mainly in colon. The lymphoid follicles are inductive sites for the specific immune response and contain two major cell types, T and B cells and in addition macrophages, DCs and follicular dendritic cells (FDCs) (Brandtzaeg and Bjerke, 1990;

- 8 -
Yeung et al., 2000; Wittig and Zeitz, 2003). The lymphoid follicles have a germinal center structure rich in B cells with interspaced FDCs and surrounded by a T cell area containing DCs (Yeung et al., 2000). The lymphoid follicles are located beneath a single layer epithelium called the follicle-associated epithelium (FAE) containing the highly specialized M cells (Brandtzaeg and Bjerke, 1990; Kraehenbuhl and Neutra, 2000). The LP contains most cellular components of the immune system, with large numbers of B cells, plasma cells, macrophages, DCs, mucosal mast cell and T cells. The vast majority of LP T cells are αβ T cells of both the CD4 and CD8 subsets and a small subset of CD4+CD8+ cells (Melgar, et al., 2002; Wittig and Zeitz, 2003). LP T cells secrete cytokines as IL-2, IFN-γ, and TNF-α (Beagley and Elson, 1992; Forsberg et al., 2002; Melgar et al., 2003). Most plasma cells, 70-90%, produce antibodies of the IgA class (Brantzaeg et al., 1999). Recently it was shown that plasma cells also secrete β-defensins (Rahman et al., 2007). In contrast to the LP, all immune cells within the epithelium are lymphocytes. These IELs are in close contact with the epithelial cells and more frequent in the small intestine compared with large intestine (Lundqvist et al., 1995). In the human small intestine, there are 10-20 IELs per 100 villus epithelial cells. IELs are a heterogeneous mixture of T lymphocyte subsets that are chiefly of activated/memory phenotype (CD45RO+ cells). In the small intestine, the CD8+
αβT cells dominate while in the large intestine there are almost equal proportions of CD8+
αβT cells, CD4+
αβT cells and CD4/CD8 double negative αβT cells (Lundqvist et al., 1995). In both small and large intestine, about 10% of IELs are γδT cells (Lundqvist et al., 1995). The majority are CD4/CD8 double negate and preferably use Vδ1 and Vγ8 in their T cell receptor as opposed to peripheral blood γδT cells which use Vδ2 and Vγ9 (Lundqvist et al., 1995; Söderström et al., 1994). The Vδ1+
γδT cells have the ability to kill stressed cells (Groh et al., 1998) and tumor cells of epithelial origin (Maeurer et al., 1996). IELs probably play multiple functions including cytotoxic activities, induction and maintenance of oral tolerance, surveillance of the IECs, and immune protection (Wershil and Furuta, 2008). 1.2 Hurdles for intestinal pathogens Immune defense works at three levels in the gut. Secreted factors to the gut lumen, at the lumen/epithelial compartment interface and inside the mucosal tissue. (Hooper and Gordon, 2001; Guarner and Malagelada, 2003; Mowat, 2003; Magalhaes et al., 2007). To colonize mucosal surfaces and invade the

- 9 -
host, microorganisms typically must first penetrate the secreted mucus layer in order to reach the apical surface of epithelial cells or release toxins that disrupt epithelial integrity. A broad range of bacterial pathogens can produce mucus-degrading enzymes, such as glycosulfatases, sialidases and mucinases to destabilize the mucus gel and remove mucin decoy carbohydrates for adhesion (Linden et al., 2008). As mucins bind pathogens, increasing the secretion of mucins is likely to benefit the host by trapping and moving pathogens away from the epithelial surface. Underneath the secreted mucus layer, the cells have a dense forest of highly diverse glycoproteins and glycolipids, which form the glycocalyx. Membrane-anchored cell-surface mucins and Carcinoembryonic antigen (CEA)-related cell adhesion molecules (CEACAMs) are major components of glycocalyx situated at the top of the microvilli (Hammarström and Baranov, 2001). The mucus/glycocalyx compartment also contains a spectrum of antimicrobial proteins including secretory IgA with specificity for microbial antigens, defensins, lysozyme, lactoferrin etc. (Magalhaes et al., 2007). Together, these layers provide a first line of defense barrier preventing interactions between microorganisms and IECs (MacDonald, 2003; Newberry and Lorenz, 2005). Another important factor preventing access for pathogens is the commensal microbiological flora, particularly well developed in colon. Pathogens have to fight commensals, which are well adopted for the environment, for nutrients, space etc. The IECs take an active part in immune defense. This includes both production of factors of innate immunity, e.g. antimicrobial peptides and crosstalk between IECs and immune cells. The enterocytes serve as afferent sensors of danger within the luminal microenvironment by secreting chemokines and cytokines that alert and direct innate and adaptive immune responses to the infected site (Shanahan, 2005; Akira et al., 2006). M cells overlying lymphoid follicles directly take up and transport antigens from the gut lumen to underlying subadjacent DCs and other antigen-presenting cells (Miller et al., 2007). In addition, DCs are believed to sample antigen directly from the intestinal lumen by forming tight-junction-like structures with IECs and projecting dendrites through the epithelial-cell layer and into the lumen (Rescigno et al., 2001; Coombes and Powrie, 2008). The consequence of these mechanisms for antigen sensing is alerting of immune effector cells within the epithelium and LP to humoral and cell mediated defense responses, e.g. cytotoxicity and production of IgA as well as recruitment of phagocytic cells like neutrophils and macrophages.

- 10 -
1.3 Innate immunity To protect the host from invasion by microorganisms and neutralize their virulence factors, all multi-cellular organisms have developed defense mechanisms that utilize germline-encoded receptors for the detection of a wide range of ligands derived from microorganisms. These phylogenetically ancient defense mechanisms are collectively known as the innate immune system. The recognition strategy is based on the detection of conserved molecular patterns that are unique to microbes. They are called pathogen-associated molecular patterns (PAMPs) and are present in bacteria, fungi, protozoa and viruses. PAMPs include lipopolysaccharide (LPS), peptidoglycan, lipoteichoic acid (LTA), terminal mannose (a terminal sugar common in microbial glycolipids, glycoproteins and polysaccharides but rare in those of humans), bacterial flagellin, N-formylmethionine, double-stranded RNA (dsRNA) and single-stranded RNA (ssRNA), unmethylated CpG DNA and fungal glucans etc (Akira et al., 2006; Kumagai et al., 2008). Because all microbes, not just pathogenic microbes, have PAMPs, they are sometimes referred to as microbe-associated molecular patterns (MAMPs) (Mackey and McFall, 2006). The receptors on host cells recognizing PAMPs are called pattern-recognition receptors (PRRs) (Medzhitov and Janeway, 1997). There are several structurally and functionally distinct classes of PRRs: Toll-like receptors (TLRs), nucleotide oligomerization domain (NOD)-like receptors (NLRs), retinoic acid-inducible gene (RIG)-I-like receptors (RLRs) and C-type lectin receptors (CLRs). They induce partly different host defense pathways. In contrast to innate immunity, adaptive immunity is dependent on antigen receptors expressed on lymphocytes that are acquired by somatic recombination of the antigen receptor genes during B and T cell development. This system has a huge repertoire and is not limited to PAMPs. However, a major drawback with the adaptive immune response is that it requires a considerable time to become fully effective. At the first encounter of antigen it takes up to 6-7 days due to differentiation and maturation of the effector cells. During evolution adaptive immunity occurs first in primitive fishes. In man innate and adaptive immunity works in parallel and can enhance each other. Since my thesis deals with innate immunity I will not discuss adaptive immunity further. 1.3.1 TLRs and their signaling

- 11 -
The evolutionary conserved TLRs are a family of single membrane-spanning receptors. Ten members of the human- and 13 members of the mouse TLR family have been identified (Roach et al., 2005; Akira et al., 2006; Kumagai et al., 2008). TLRs are expressed as homo- or heterodimers. All TLR family members are made up of multiple leucin-rich repeat motifs (LRRs) at the N-terminal of the molecule, followed by a cysteine-rich region, a transmembrane domain, and an intracellular domain at the C-terminal, the Toll/interleukin-1 (IL-1) receptor (TIR) domain. The TLRs are either located to the cell membrane (TLR1, TLR2, TLR4, TLR5, TLR6 and TLR10) or to intracellular membranes (TLR3, TLR7, TLR8, and TLR9). The C-terminal TIR domain located in the cytosol is able to recruit TIR-containing intracellular proteins that mediate their signaling (Brikos and O'Neill, 2008; Kumagai et al., 2008). Ligand binding to the LRR, either directly or by means of accessory molecules, is believed to induce a conformational change that brings the intracellular TIR domains in the dimer together (Latz et al., 2007). Different TLRs recognize distinct PAMPs. TLR2 plays a crucial role in the recognition of peptidoglycan, LTA, and lipoproteins (Takeuchi et al., 1999). TLR2 forms a heterodimer with either TLR1 or TLR6, and these heterodimers recognize lipoproteins with different lipid moieties respectively (Takeuchi et al., 2001; 2002). TLR1-TLR2 heterodimer recognizes triacylated lipoproteins (Takeuchi et al., 2002; Jin et al., 2007), whereas the TLR2-TLR6 heterodimer recognizes diacylated lipoproteins and LTA (Takeuchi et al., 2001). LPS, a cell-wall component of Gram-negative bacteria (Poltorak et al., 1998) is recognized by TLR4 (Triantafilou and Triantafilou, 2002), which involves the accessory molecule myeloid differentiation protein 2 (MD-2) (Nagai et al., 2002; Kim et al., 2007), membrane-bound CD14 (Jiang et al., 2005), and LPS-binding protein (LBP) (Jack et al., 1997). TLR5 is a receptor for bacterial flagellin (Hayashi et al., 2001; Uematsu et al., 2006). TLR3, TLR7, TLR8 and TLR9, localized on cytoplasmic membranes of the endoplasmic reticulum or endosomes, are receptors for nucleic acids and their derivatives. TLR3 is a receptor for viral dsRNA (Alexopoulou et al., 2001). TLR7 and TLR8 recognize ssRNA from RNA viruses (Hemmi et al., 2002; Jurk et al., 2002; Diebold et al., 2004; Heil et al., 2004). TLR9 is a receptor for DNA with an unmethylated CpG-motif (CpG-DNA) (Barton GM, et al. 2006), which is abundant in bacterial DNA, whereas the frequency of CpG motifs in mammalian genome DNA is low and mostly methylated (Ishii and Akira, 2006). At ligand binding to TLR the cytoplasmic TIR domains dimerize to form a platform for the recruitment of adapter proteins and additional signaling molecules (Trinchieri and Sher, 2007). Through a series of intracellular

- 12 -
molecular reactions nuclear factor-kappa B (NF-κB) and/or interferon regulatory factor (IRFs) are activated, leading to the expression of various genes associated with host defense, including proinflammatory cytokines, chemokines and defensins. Depending on cell type, the TLR-mediated immune response may differ. Moreover, the different signaling pathways used by distinct TLRs and the presence of different PAMPs on invading microbes contributes to a complex multifunctional innate immune response. 1.3.2 TLR-independent pathogen recognition Microbial components are also recognized by the intracellular cytosolic receptor NLRs, the cytosolic RLRs and the surface expressed CLRs. The NLR family has over 30 members in humans and is characterized by the presence of a central NOD domain and a C-terminal LRR domain and a N-terminal domain responsible for the signaling, either the caspase recruit domain (CARD) or the putative protein-protein interaction (PYRIN) domain (Kanneganti et al., 2007; Franchi et al., 2008). The NLRs recognize different bacterial components notably γ-D-glutamyl-mesodiaminopimelic acid and muramyl dipeptide, respectively leading to activation of NF-κB (Franchi et al., 2008; Chamaillard et al., 2003; Girardin et al., 2003). Members of the NALP subfamily have a PYRIN domain instead of a CARD domain. Fourteen members (NALP1-14) have been described in humans (Kanneganti et al., 2007). NALP1-3 were shown to be involved in Caspase-1-mediated cleavage of proIL-1β, proIL-18 and proIL-33 into their mature forms in response to stimuli (Agostini et al., 2004; Martinon et al., 2002; Keller et al., 2008). NALP3 is activated by LPS, lipoprotein, CpG-DNA, bacterial RNA, uric acid crystals and by ATP (Kanneganti et al., 2006; Mariathasan et al., 2006; Martinon et al., 2006). The cytosolic flagellin is capable of activating caspase-1 in an NLRC4-dependent manner (Sutterwala et al., 2007). RLRs are DExD/H box RNA helicases located in the cytoplasm where they mediate recognition of virus-specific RNA and then activate IRFs inducing expression of type I interferon (IFN). Three members of RLRs have been described in mammals: retinoic acid inducible gene I protein (RIG-I), melanoma differentiation associated gene-5 protein (MDA5) and laboratory of genetics and physiology protein-2 (LGP2) (Bowie and Fitzgerald, 2007; Yoneyama and Fujita, 2007; 2008; Xiao, 2008). Interestingly, microbial ligands for members of the RLR-, NLR- and TLR-families may be the same. However, the molecular bases of their receptor-ligand interactions and the outcome of the interactions may be quite different. The cross-talk between

- 13 -
these sensing-systems probably plays an important role in immunological homeostasis. CLRs are a family of membrane-associated receptors that are able to recognize carbohydrate structures present on the pathogens. So far, more than 60 CLRs have been identified in humans. The extra-cellular portion of CLR consists of one or more C-type (Ca2+ dependent) carbohydrate recognition domains (CRDs), involved in carbohydrate recognition (Zelensky and Gready, 2005). Although microbes and vertebrate cells are able to produce similar polysaccharide chains, the density of carbohydrates on microbes is generally much higher than on vertebrate cells. At least partly due to this difference, carbohydrates present on microorganisms can be specifically recognized by the CLRs (Gijzen et al., 2006; van Vliet et al., 2008). By binding to carbohydrate moieties, CLRs mediate biological events, such as cell-cell or cell-matrix adhesion and glycoprotein turnover. Although the majority CLRs are primarily involved in antigen uptake, some CLRs such as Dectin-1 have been shown to trigger intracellular signaling cascades upon ligand binding resulting in NF-κB activation and production of cytokines (Rogers et al., 2005; Gross et al., 2006). 1.3.3 TLR and NOD signaling in the human intestine TLRs and NODs are expressed in human small and large intestine and play important roles of immune and non-immune functions in the mucosa (Melmed et al., 2003; Otte et al., 2004; Abreu et al., 2005; Sanderson and Walker, 2007). Thus, IECs and lamina propria macrophages express TLR4 and TLR2 (Hausmann et al., 2002) and jejunal smooth muscle cells express TLR4 (Rumio et al., 2006). NOD1 is ubiquitously expressed in many tissues and cells including IEC while NOD2 is constitutively or inducibly expressed in monocytes, macrophages, T and B cells, DCs, as well as IECs, including Paneth cells. (Sanderson and Walker, 2007). In order to maintain intestinal mucosal homeostasis against tissue damage, commensal microflora induced host modulatory effects seem to require functional TLRs and NODs (Rakoff-Nahoum et al., 2004; Kobayashi et al., 2005). Under normal conditions, the intestinal mucosa is constantly exposed to TLR/NOD ligands derived from the commensal microflora and a basal state of activation of downstream signaling pathways is achieved, thus ensuring rapid compensation and limited inflammatory responses (Rakoff-Nahoum et al., 2004). Deficient TLR or NOD signaling may elicit an imbalance, facilitating injury and leading to disease. There is evidence to show that commensal microflora directly assist the host to strengthen

- 14 -
intestinal epithelial barrier resistance (Hooper et al., 2001; Madsen et al., 2001; Otte et al., 2004). Altered expression of TLRs and NODs in IECs has been reported in chronic inflammation. Thus, increased expression of TLR4 has been demonstrated in inflammatory bowel disease (IBD), while the expression of TLR2 and TLR5 remained unchanged (Cario and Podolsky, 2000). Moreover, proinflammatory cytokines such as IFN-γ and tumor necrosis factor-α (TNF-α) increase expression of TLR4 and MD-2 resulting in increased LPS responsiveness in human colonic epithelial cells (Abreu et al., 2001; Suzuki et al., 2003). Polymorphisms and mutations within the NOD2 gene have been associated with the development of Crohns’ disease in a subgroup of patients ( Hugot et al., 2001; Ogura et al., 2001) and persons homozygous for a variant NOD2 may have a 20-fold higher susceptibility to Crohns’ disease (Cuthbert et al., 2002).
1.4 Mucins The intestinal mucosal surface in humans is covered by a layer of hydrated viscous mucus ranging in thickness from 50 to 700 µm. The mucus is a mixture of mucins, free protein, salts, and water. The major components of mucus are secreted gel-forming mucins produced by goblet cells (Strugala et al., 2003). This viscous, sticky layer traps particles, bacteria, and viruses, which are expelled by the peristaltic process of the gut. Mucins are family of heavily glycosylated proteins. Based on their properties, the mucins can be divided into three distinct subfamilies (Linden et al., 2008): (a) Secreted gel-forming mucins, which are the major component of mucus and confer its viscoelastic properties. They contain N- and C-terminal cysteine-rich domains that are involved in homo-oligomerization mediated by inter-molecular disulfide bonds (Gum et al., 1994; Perez-Vilar et al., 1998). (b) Cell surface mucins, also called membrane-bound mucins, which have a single membrane-spanning domain and a short cytoplasmic tail in addition to the extensive extracellular domain. (c) Secreted non-gel-forming mucins, which are relatively small glycoproteins and incapable of forming a gel (Andrianifahanana et al., 2006). Classically, mucins are known to play a central role in the protection, lubrication and hydration of the external surfaces of epithelial tissue layers lining the intricate network of ducts and lumens within the human body (Strous and Dekker, 1992; Van Klinken et al., 1995). Mucins have direct and indirect roles in defense against infection. Different mucins are expressed in varying proportions throughout the normal digestive tract and the complexity of the mucin mixture increases from esophagus to rectum (Corfield et al.,

- 15 -
2001; de Bolos et al., 2001). In the small intestine, MUC2 is predominantly expressed by the goblet cells, while MUC1, MUC3, and MUC4 are mostly found in enterocytes, and to a lesser extent in goblet cells (Audie et al., 1993; Jass and Roberton, 1994). MUC17 (Gum et al., 2002) and MUC20 (Higuchi et al., 2004) have also been detected in the small intestine. The colorectal part of the intestine exhibits a more diverse pattern of mucin expression. Here large amounts of MUC2, MUC4, and MUC11 are found in the goblet cells, whereas the membrane-bound mucins, MUC1, MUC3, MUC4, and MUC12 are detected in colonocytes (Audie et al., 1993; Jass and Roberton, 1994; Tytgat et al., 1994). MUC13 (Williams et al., 2001), MUC17 (Gum et al., 2002) and MUC20 (Higuchi et al., 2004) have also been observed in colon. Gene expression of mucins in the mucosal epithelium is both constitutive and inducible. The mucosal epithelial cells continuously produce sufficient mucins to maintain the mucus layer, whereas the inducable pathway affords a massive discharge as a response to environmental and/or (patho)physiological stimuli, including cholinergic stimuli, inflammatory cytokines, prostaglandins, LPS, bile salts and nitric oxide (Andrianifahanana et al., 2006; Linden et al., 2008). Moreover, infections can induce alterations in mucins, such as increased mucin production through goblet cell hyperplasia, increased mucin secretion and altered mucin glycosylation affecting microbial adhesion and the ability of microorganisms to degrade mucus. These alterations work in concert with other defense processes to prevent infection (Linden et al., 2008). 1.5 The Carcinoembryonic antigen family Carcinoembryonic antigen (CEA) was identified as a prominent tumor-associated antigen in human colon cancer in 1965 (Gold and Freedman, 1965). CEA and related genes make up the CEA gene family belonging to the immunoglobulin gene superfamily. The gene products are highly glycosylated glycoproteins. In human, the CEA family consists of 29 genes, 18 of which normally are expressed, including seven CEACAMs, (CEACAM1, CEACAM3-CEACAM8) and 11 pregnancy specific glycoproteins (PSG1-PSG11), and 11 PSG-related pseudogenes (Hammarström, 1999). Recently, a group of genes more distantly related to CEA than the CEACAMs discussed above was identified in the human genome (Zebhauser et al., 2005). CEACAMs are also found in other mammals and CEACAM1 appears to be the phylogenetically most ancient member of the CEACAM subfamily. According to the new nomenclature CEA should be called CEACAM5 (Beauchemin et al., 1999). However, the

- 16 -
name CEA is kept for historical and practical reason. Thus, in the extensive literature on the use of CEA/CEACAM5 as a clinical biomarker the name CEA is used. Clinical aspects of the utility of CEA as a biomarker will not be discussed here. CEACAMs are linked to the cell surface by a glycosylphosphatidylinositol (GPI) anchor (CEA, CEACAM6-8) or through a transmembrane region with a cytosolic signaling domain (CEACAM1, CEACAM3 and CEACAM4), while PSGs are secreted glycoproteins (Hammarström, 1999; Beauchemin et al., 1999). There are two major CEACAM1 isoforms, which differ with respect to the length of their cytoplasmic domain. CEACAM1-L has a long cytoplasmic domain (71-73 amino acids) and CEACAM1-S has a short cytoplasmic domain (10-12 amino acids). CEACAM1-L contains two immunoreceptor tyrosine-based inhibition motifs (ITIM) while CEACAM1-S lacks ITIM sequences (Gray-Owen and Blumberg, 2006). It has been established that four CEACAMs subgroup genes are expressed in human intestinal epithelial cells (Frängsmyr et al., 1999). In colon, CEACAM1, CEA, CEACAM6 and CEACAM7 are expressed at high level in mature and highly differentiated enterocytes at the crypt mouth and free luminal surface (Frängsmyr et al., 1995; 1999). CEA/CEACAM5 is also present in the mucus droplets of goblet cells. Immunoelectron microscopy studies of colon demonstated that all four molecules are localized to the microvilli and the glycocalyx (the fuzzy coat) at the top of the microvilli (Hammarström and Baranov, 2001). Within this compartment the 4 molecules show a different distribution. In the small intestine CEACAM1 is expressed by absorptive epithelial cells while CEA is only produced by goblet cells (Fahlgren et al., 2003). CEACAM6 and CEACAM7 appear not to be expressed in small intestine (Schölzel et al., 2000). The CEACAM family members are multifunctional molecules involved in cell adhesion, tumor suppression, regulation of signal transduction and probably also innate immunity. Several CEACAM family members have also been identified as receptors for host-specific viruses and bacteria in mice and humans, respectively, making these proteins interesting targets of pathogen (Hammarstrom, 1999; Gray-Owen and Blumberg, 2006; Kuespert et al., 2006; Griffiths et al., 2007; Hemmila et al., 2004). Deletion of CEA family members can render the host resistant to particular pathogens (Kuespert et al., 2006). The majority of the currently characterized meningococcal and gonococcal adhesions, the opacity-associated (Opa) proteins display binding specificity for human surface receptors of the CEACAMs family (OpaCEA proteins). All OpaCEA proteins bind to the non-glycosylated C′CFG-face of the immunoglobulin-domain fold of the N-terminal domain of CEACAM1, CEACAM3, CEA and CEACAM6. Binding

- 17 -
of OpaCEA proteins to CEACAM molecules is sufficient to induce the internalization of the bacteria into several cell types in vitro (Heyderman and Virji, 2007). Essentially all CEA family members, including GPI-anchored CEA and CEACAM6, function as intercellular adhesion molecules in vitro (Benchimol et al., 1989; Oikawa et al., 1989). CEACAM1 is primarily an activation-induced cell-surface molecule that functions as a co-inhibitory receptor. Homophilic ligation of CEACAM1 on T cells leads to a signaling mechanism, which results in inhibition of a broad range T-cell functions (Nagaishi et al., 2006). CEACAM1-L and CEACAM1-S have different functions. CEACAM1-L can regulate cell proliferation, invasion, cell migration, angiogenesis, lymphangiogenesis, and cytotoxicity. CEACAM1-L, containing two ITIMs, generally transmit inhibitory signals while CEACAM1-S contains sequences that can bind calmodulin (Edlund et al., 1996), tropomyosin and globular actin (Schumann et al., 2001), indicating a regulated interaction with the cytoskeleton. In the breast, CEACAM1-S mediates mammary lumen formation via an apoptotic and cytoskeletal reorganization mechanism (Kirshner et al., 2004; Chen et al., 2007). In most cell types and tissues, CEACAM1-L and CEACAM1-S are expressed together, but at characteristically different ratios (Hauck et al., 2006). The abundance of CEACAM1 and the relative ratio of CEACAM1-L to CEACAM1-S isoforms are not static, and can vary substantially according to the cell type, its phase of growth and activation state (Singer et al., 2000; Greicius et al., 2003). Moreover, in vitro studies performed using polarized epithelial cells clearly show that the CEACAM1 cytoplasmic tail governs both the distribution of the receptor between intracellular compartments and the cell membrane, and the movement of the receptor between apical and basolateral surfaces (Sundberg et al., 2004). An optimal ratio in the expression level of CEACAM1-L and CEACAM1-S isoforms were found to be essential for some of its biological functions, e.g. the suppression of cell proliferation (Obrink et al., 2002). Recent data demonstrated a high ratio of CEACAM1-S to CAECAM1-L isoforms in normal breast tissues versus breast cancer suggesting that altered splicing of CEACAM1 may play an important role in tumorogenesis (Gaur et al., 2008). 1.6 Antimicrobial peptides and lysozyme Although the alimentary tract is colonized by, and exposed to a great number of different bacteria, intestinal infection or translocation of bacterial agents is the exception, not the rule, and is largely limited to pathogenic bacteria or predisposing disease states (Liévin-Le Moal and Servin, 2006; Wehkamp et

- 18 -
al., 2007). Intestinal epithelial cells in higher organisms not only form a physical barrier but also a chemical barrier. This chemical barrier is composed of secreted anti-infectious substances, including mucus, antimicrobial peptides (AMPs), and other anti-microbial molecules, such as lysozyme, which together with resident microbiota provide the front line of defense against pathogenic microorganisms (Ganz, 2002). AMPs are small molecular weight proteins with broad-spectrum antimicrobial activity against bacteria, fungi, protozoa and some enveloped viruses. Based on common structural and functional features, the AMPs present in the human intestinal tract have been divided into two major families: defensins and cathelicidins (Otte et al., 2003). The human defensins are small cysteine containing cationic peptides (range from 28 to 50 amino acid residues) containing three intra-molecular disulfide bonds (Pazgier et al., 2007). The positions of the disulfide bonds define two subclasses: α-defensins and β-defensins (Cunliffe, 2003; Ganz, 2003). All of them have anti-microbial activity against bacteria, and some also against fungi, viruses and protozoa (Lehrer et al., 1993). Human α-defensins are 29-35 amino acid residues in length and form a triple-stranded β-sheet. Four of the α-defensins are produced by neutrophils and are named human neutrophil peptides 1-4 (HNP1-4) (Ganz et al., 1985). The primary source of two additional α-defensins, human α-defensin-5 and 6 (HD-5 and HD-6), is Paneth cells in the small intestine, where these cells are located at the crypt base (Jones and Bevins, 1992; 1993; Ganz, 1999; Porter et al., 1997). Genomic studies have revealed that there are over 40 different β-defensin genes arranged in 4 different clusters in the human genome (Liu et al., 1998). Four different β-defensins have been isolated and characterized human β-defensin 1-4 (hBD 1-4) present in the intestine. β-defensins form three β-strands arranged as an anti-parallel sheet held together by three intra-molecular disulfide bonds (Dann and Eckmann, 2007; Taylor et al., 2008). In contrast to the high and relatively stable expression level of HD-5 and HD-6 in the small intestine and low expression in other regions of the gastrointestinal tract, β-defensins expression is much more variable with respect to the individual peptides, anatomical location, and pathogenic condition. hBD1 is constantly expressed throughout the gastrointestinal tract and remains invariable even in inflammation or infections (O'Neil et al., 1999; Fahlgren et al., 2003; 2004). In normal intestine, there is little hBD-2 expression, but it is strongly upregulated in inflammatory bowel disease (Fahlgren et al., 2003; Ganz, 2002). It was reported that in the colonic IECs from patients with ulcerative colitis, the levels of mRNA expression of HD-

- 19 -
5, HD-6, hBD-2, hBD-3, hBD-4 increased significantly compared with in the IECs from control colon (Fahlgren et al., 2003; 2004). HD-5, HD-6 was produced by metaplastic Paneth cells in colon of patients with ulcerative colitis (Fahlgren et al., 2003). hBD-2, hBD-3 and hBD-4 also were produced by mature IgA and IgG producing colonic plasma cells (Rahman et al., 2007). mRNAs expression levels of HD-5 and HD-6 were also increased in proximal small intestinal IECs from children with active celiac disease (CD) but levels returned to normal in treated patients. The increased production was a consequence of Paneth cell metaplasia, which in turn correlated to the increased production of IFN-γ by IELs in CD (Forsberg et al., 2004). The increased expression of β-defensins is explained by the inducibility of these peptides by proinflammatory cytokines and bacteria probably via TLRs. The induction of hBD2, hBD3 and hBD4 is mediated by proinflammatory factors such as IL-1β, IFN-γ, mostly through NF-κB-dependent and activator protein (AP-1) -dependent pathways (O'Neil et al., 1999; Wehkamp et al., 2003; Fahlgren et al., 2004). The signaling pathways also include TLRs, especially TLR2 and TLR4, that recognize and bind PAMPs and mitogen-activated protein kinases (Froy, 2005). Complementing their role as antibacterial agents in innate immune defense, defensins also act as links to the adaptive immune system. hBD2 can bind to DCs that express the chemokine receptor CCR6; thereby recruiting DCs and memory T cells by chemotaxis to sites of infection (Yang et al., 1999). HD-5 can block the release of IL-1β from LPS-activated monocytes (Shi et al., 2007). Cathelicidins are α-helical cationic antimicrobial peptides that consist of a highly conserved N-terminal structural domain, cathelin, linked to a variable C-terminal peptide with antimicrobial activity. LL-37 is the only cathelicidin identified in humans, found predominantly in neutrophils and various epithelial cells lining the stomach, lower small intestine and throughout the colon (Gudmundsson et al., 1996; Hase et al., 2002; 2003; Schauber et al., 2003; Kai-Larsen and Agerberth, 2008). The name LL-37 derives from the first two leucines (L) residues at the N-terminus and the number of amino acids in the peptide (Gudmundsson et al., 1996). The expression is most prominent in the lower small intestine and colon with high peptide abundance in superficial epithelial cells (Schauber et al., 2003). Apart from exhibiting a broad antimicrobial spectrum, it is now evident that LL-37 possesses several additional functions, which are chemotaxi, endotoxin neutralization, angiogenesis and wound healing (Kai-Larsen and Agerberth, 2008). Lysozyme is an enzyme that acts by breaking the β1, 4 linkage between N-acetylglucosamine and N-acetylmuramic acid in the peptidoglycan layer of

- 20 -
bacterial cell walls. Paneth cells display prominent cytoplasmic granules, containing antibacterial proteins such as lysozyme, secretory phospholipase A2 type IIA, HD-5 and HD-6, which are released into the intestinal lumen in response to a range of stimuli (Keshav, 2006). Wehkamp and colleagues reported that lysozyme mRNA expression was high in the duodenum, and that the protein localized to both Brunner's glands in the lamina propria and Paneth cells (Wehkamp et al., 2006). It also has been recognized that lysozyme mRNA expression is increased in colonic intestinal epithelial cells from ulcerative colitis patients (Fahlgren et al., 2003; 2004) and in the proximal small intestinal mucosa of patients with active CD (Forsberg et al., 2004). 1.7 Cytokines Cytokines are a group of small soluble signaling proteins that are released mainly by immune cells and used extensively in cellular communication through cell-surface receptor-ligand interaction. The cytokine superfamily of proteins is an integral part of the signaling network between cells and is essential in generating and regulating the immune system. These signaling molecules can act in an autocrine, paracrine and endocrine manner that have important local and systemic effects. By binding to specific membrane receptors and acting in networks or cascades synergistically or antagonistically, cytokines signal the target cell and alter its functions, often including activation, proliferation, and expression and secretion of effectors molecules. Cytokines generally are pleiotropic and redundant molecules, which means that a particular cytokine not only acts on a number of different types of cells rather than on a single cell type but also that a number of different cytokines carry out the same function. Because of their central role in the immune system, aberrant expression levels of cytokines usually signal ongoing patho-physiological processes (Clark and Coopersmith, 2007). On the basis of their presumed function, cell of secretion, or target of action, cytokines have been classified as lymphokines, chemokines, interleukins, and interferons. Some cytokines promote inflammation and are therefore called proinflammatory cytokines, including TNF-α, IL-1β, IFN-γ and IL-18, where other cytokines suppress the activity of proinflammatory cytokines and are therefore called anti-inflammatory cytokines. IL-10 and IL-6 are pleiotropic cytokines with both pro- and anti-inflammatory effects. Chemokines are cytokines with the primary function of regulating leukocytes recruitment. IL-8 (CXCL8) is one of more than 50 chemokines identified to date. Through binding to specific cell surface receptors,

- 21 -
cytokines act on their target cells and initiate signals that are critical to a diverse spectrum of functions. Of the many cell types involved in the intestinal inflammatory response, IECs function as an integral component of the immune defence. Indeed, IECs form a critical mucosal barrier between the host’s internal milieu and the external environment. In the course of inflammation, intestinal epithelial cells serve as immuno-effector cells and are capable of up-regulating surface molecules and secreting cytokines and chemokines to facilitate antigen presentation to immune cells. The induction of inflammatory gene expression in the intestinal epithelial compartment leads to impaired barrier function, cell differentiation, and proliferation (Acheson and Luccioli, 2004). Additionally, enterocytes can act as antigen presenting cells themselves and regulate lymphocyte responses in the intestine (Hershberg and Mayer, 2000). Enterocytes are also in constant communication with IELs and T cells within the lamina propria and has been shown to exhibit increased major histocompatibility complex (MHC) class II molecules at the basolateral membrane in states of inflammation (Kaiserlian et al., 1989). The polarized expression of levels of MHC class II molecules on enterocytes suggests that antigen exposure on the luminal side as opposed to the basolateral side elicits different immunologic responses (Hershberg and Mayer, 2000). 1.7.1 Interferon-γ The interferons (IFNs), initially described as a family of antiviral proteins (Isaacs and Lindenmann, 1957). There are three types of IFNs, type I (α, β, ω and τ), type II (γ) and type III (λ), which signal through different receptors to produce distinct, but overlapping, cellular effects (Boehm et al., 1997). Type II interferon is also called interferon-γ (IFN-γ). Human IFN-γ is a homodimeric glycoprotein containing 146-amino acid subunits and is secreted by activated immune cells, primarily T and NK cells (Schroder et al., 2004). IFNγ is a pleiotropic cytokine that stimulates a wide range of cellular responses promoting cell-mediated immune responses, increased antigen presentation and production of proinflammatory cytokines. (Schroder et al., 2006; van Boxel-Dezaire and Stark, 2007). Other cellular responses include regulation of cell proliferation and stimulation of apoptosis. The biological effects of IFN-γ are elicited through activation of intracellular molecular signaling networks, the best characterized of which is the Janus kinase (JAK)/Signal Transducer and Activator of Transcription (STAT) signaling pathway providing the primary mechanism through which gene expression is induced. There are, however, differential patterns of activation of STAT1, STAT3, and STAT5 in different cells, and activation

- 22 -
of transcription factors other than STATs yielding cell type specific regulation of biological function (Darnell et al., 1994; Levy and Darnell, 2002; van Boxel-Dezaire and Stark, 2007). 1.7.2 Tumor Necrosis Factor-α Tumor necrosis factor-α (TNF-α), which was first identified in 1975 as a cytokine with anti-tumor effects in mice (Carswell et al., 1975), plays a crucial role in the immune system as it mediates inflammation, cell growth, apoptosis, and enhances the cellular immune response (Fukushima et al., 2004; Thomson and Lotze, 2003). Human TNF-α cDNA was cloned and expressed in 1985 (Pennica et al., 1985). Monocytes and macrophages produce TNF-α. However, other cell-types, including T cells and IECs have the ability to produce TNF-α in response to bacterial toxins, inflammatory products, and other stimuli (Tracey and Cerami, 1993; Bazzoni and Beutler, 1996). TNF-α is primarily synthesized as a 233-amino acid type II transmembrane precursor arranged in stable homotrimers that is subsequently cleaved proteolytically in the extracellular domain to release soluble TNF-α (17 kDa) (Beutler and Cerami, 1986; Tang et al., 1996). The biologically active soluble form of TNF-α is a non-covalently bound trimer. The cell membrane-associated form of TNF-α also possesses biological activity. The two forms of TNF-α may have distinct biological activities. A matrix metalloprotease, TNF-α-converting enzyme (TACE), not only mediates release of TNF-α from the cell membrane-bound precursor (Black et al., 1997) but is also involved in processing several cell-membrane-associated proteins, including TNF-α receptors. The released soluble TNF-α receptor forms can neutralize the actions of TNF-α (Wang et al., 2003). TNF-α exerts the biological functions via interaction with its cognate membrane receptors, comprising the TNF-α receptors (Locksley et al., 2001). Two types of TNF-α receptors, type I (TNFR1) and type II (TNFR2) are present on the plasma membrane of virtually all cell types except erythrocytes and bind both soluble and membrane-associated TNF-α. Each TNF-α trimer binds three receptor molecules to produce a hexameric (TNF-α)3-(TNFR)3 structure triggering the intracellular signaling pathway (Idriss and Naismith, 2000). TNFR1 is expressed in most tissues, and can be fully activated by both membrane-bound and soluble TNF-α, whereas TNFR2 is found only in cells of the immune system, and respond to the membrane-bound form of the TNF-α. TNFR1 and TNFR2 share structural homology in the extracellular TNF-α binding domains and exhibit similar binding affinity for TNF-α, but they show no sequence homology, and induce separate

- 23 -
cytoplasmic signaling pathways following receptor-ligand interaction (Wallach et al., 1999; Ledgerwood et al., 1999). All known responses to TNF-α are triggered by binding to one of these two receptors (Al-Lamki et al., 2001). Binding causes a conformational change in the receptor, leading to the dissociation of the inhibitory protein called “silencer of death domain” (SODD) from the intracellular death domain. This dissociation enables the adaptor protein TNFR-associated-death-domain (TRADD) protein to bind to the death domain, serving as a platform for subsequent protein binding. Following TRADD binding, these pathways can be initiated, including activation of NF-κB and mitogen-activated protein kinases (MAPK) pathway and induction of death signaling (Wajant et al., 2003; Chen and Goeddel, 2002). Based on cell culture work and studies with receptor knockout mice, both the proinflammatory and the programmed cell death pathways that are activated by TNF-α, and associated with tissue injury, are largely mediated through TNFR1. The consequences of TNFR2 signaling are less well characterized, but TNFR2 has been shown to mediate signals that promote tissue repair and angiogenesis (Bradley, 2008). 1.7.3 Interleukin-1β Interleukins are cytokines first found to be expressed by leukocytes. The IL-1 family has the following member: IL-1α, IL-1β, IL-1 receptor antagonist (IL-1RA), IL-18 and IL-33. IL-1β is the prototypic pro-inflammatory cytokine involved in immune defense against infection (Arend et al., 2008). IL-1β is produced in a variety of cells including macrophages, monocytes, neutrophils, keratinocytes and epithelial cells. In the human, IL-1β is synthesized as a 31-kDa precursor which is processed and released from cytoplasm. Pro-IL-1β is biologically inactive and must be converted to the mature 17 kDa IL-1β by caspase-1 to acquire the ability to bind to receptors and to activate cells. Both IL-1α and IL-1β bind to the same receptor and have similar if not identical biological properties. There are two types of receptors for IL-1, IL-1 receptor type I (IL-1RI) and IL-1 receptor type II (IL-1RII), and an IL-1R accessory protein (IL-1RAcP) (Eisenberg et al., 1991; Martin and Falk, 1997; May and Ghosh, 1998). IL-1RI possesses a long cytoplasmic domain of 213 amino acids and is capable of activating cells, whereas IL-1RII has only a short intracellular domain consisting of 29 amino acids and may act as a suppressor of IL-1 activity by competing for IL-1 binding (Kuno and Matsushima, 1994; Vigers et al., 2000). After IL-1 binds to IL-1RI, IL-1RAcP joins with IL-1/IL-1RI to form a complex, leading to cell activation mediated by the cytoplasmic domains of both receptor chains. This complex recruits a number of intracellular adapter

- 24 -
molecules, including MyD88, IL-1R-associated kinase (IRAK), and TNF receptor-associated factor 6 (TRAF6), to activate signal transduction pathways such as NF-κB, activator protein-1 (AP-1), JNK, and mitogen-associated protein kinase (p38 MAPK) (O'Neill and Greene, 1998; Verstrepen et al., 2008). IECs are the major source of IL-1RI in the human mucosa. Upon inflammation they respond with upregulation of IL-1RI production instead of IL-1RII (Daig et al., 2000). IL-1 can induce IECs to produce several cytokines, suggesting that IEC is important in a cytokine network at the intestinal mucosa to amplify the effects of IL-1 during an inflammatory response (McGee et al., 1996). 1.7.4 Interleukin-6 The IL-6 is a 185 amino acid multifunctional protein that acts as both a pro-inflammatory and anti-inflammatory cytokine. It is produced by many cell types including macrophages, fibroblasts, endothelial cells, keratinocytes and epithelial cells (Hirano et al., 1986; Xu et al., 1997). IL-6 expression is induced during the acute-phase response by a variety of stimuli, which include cytokines such as IL-1, TNF-α, and platelet-derived growth factor (PDGF). Bacterial and viral infection and microbial components such as LPS are also potent inducers of IL-6 (Taga and Kishimoto, 1997; Heinrich et al., 1990; Kishimoto et al., 1995; Kamimura et al., 2003; Kishimoto, 2005). IL-6 signals through the IL-6 receptor (IL-6R), which is composed of two different subunits, an alpha subunit, IL-6 receptor subunit alpha (IL-6Rα) and glycoprotein 130 (gp130), a receptor subunit for signal transduction (Paonessa et al., 1995; Hirano, 1998). IL-6Rα is the binding component specific to IL-6. In contrast, gp130 transmits signals not only of IL-6 but also of IL-6-related cytokines such as IL-11, ciliary neurotrophic factor, cardiotrophin-1, cardiotrophin-like cytokine, leukemia inhibitory factor, neuropoietin, oncostatin M, IL-27 and IL-31 (Taga and Kishimoto, 1997; Derouet et al., 2004; Dillon et al., 2004; Pflanz et al., 2004). Binding of IL-6 to IL-6Rα leads to homo- and hetero-dimerization of gp130 and these complexes bring together the intracellular regions of gp130 to initiate a signal transduction cascade through initiating cellular events including activation of gp130-associated JAK kinases and activation of ras-mediated signaling. The tyrosine-phosphorylated gp130 further transmits signals by recruiting Src homology region 2 (SH2) domain-containing signaling molecules such as the protein tyrosine phosphatase SHP-2 and STAT1 and STAT3. These factors and other transcription factors like AP-1 and SRF (serum response factor) that respond to many different signaling pathways

- 25 -
come together to regulate a variety of complex promoters and enhancers that respond to IL-6 and other signaling factors (Heinrich et al., 2003; Murray, 2007; O'Sullivan et al., 2007; Xu and Qu, 2008). Together with TNF-α and IL-1, IL-6 is also considered a major proinflammatory cytokine important in the protection from pathogens during an infection. In the intestine, the major sources of IL-6 are macrophages and epithelial cells. In human colon, IL-6 expression is induced locally in epithelial cells and macrophages in response to PAMPs (Stadnyk and Waterhouse, 1997; Kusugami et al., 1995; Reinecker et al., 1993). IL-6 deficiency causes more severe infection-associated mucosal inflammation and ulceration, indicating that this cytokine was protective against colitis caused by C. rodentium infection in mice (Dann et al., 2008). 1.7.5 Interleukin-8 IL-8 or CXCL8 is a cytokine belonging to the CXC chemokine or α-chemokine family. It has leukocyte chemotactic properties (Baggiolini et al., 1994). IL-8 is produced by macrophages, fibroblasts, endothelial cells and epithelial cells. Expression of IL-8 is regulated by different stimuli including inflammatory signals, chemical and environmental stresses, and steroid hormones (Ben-Baruch et al., 1995). IL-8 is a major mediator of immune reactions in the innate immune system. The major sources of IL-8 in the intestinal mucosa are macrophages, epithelial cells, and fibroblasts. A large number of studies have shown increased levels of IL-8 in infected mucosal (Daig et al., 1996; Rogler and Andus, 1998). In inflammation, IL-8 is secreted at high levels to attract and activate neutrophils to the diseased site (Eckmann and Kagnoff, 2005; Jung et al., 1995). Neutrophils are effector cells that kill bacteria by phagocytosis and intracellular killing mechanisms but also destroy affected tissues through the release of enzymes from granules. IL-8 also enhances the expression of adhesion molecules that promote of neutrophils recruitment into different tissue sites (Middleton et al., 1997; Kobayashi, 2008). While neutrophil granulocytes are the primary target cells for IL-8, endothelial cells, macrophages, mast cells and epithelial cells also respond to this chemokine. There are several receptors of the cell surface capable to binding IL-8. The most frequently studied are two G protein-coupled receptors, CXCR1 and CXCR2 (Holmes et al., 1991; Murphy and Tiffany, 1991). After activation of heterotrimeric G proteins, IL-8 signaling promotes activation of the primary effectors phosphatidyl-inositol-3-kinase or phospholipase C. In addition, IL-8 signaling activates members of the Rho GTPase family and a number of nonreceptor tyrosine kinases that regulate the architecture of the

- 26 -
cell cytoskeleton and its interaction with the surrounding extracellular environment (Waugh and Wilson, 2008). 1.8 Intestinal microflora The terms ‘intestinal microflora or microbiota’ are used to describe the entire population of microorganisms in the intestine regardless of type and number. The adult human intestinal microbiota is composed of an estimated 1014 commensal bacteria (Ley et al., 2006) with a total weight of approximately 1.5 kg. Somewhere between 500 and 1000 different species, of which most have not yet been properly characterized, live in the intestine based on molecular biology analysis of the collective genome, termed the microbiome. (Guarner and Malagelada, 2003; Sears, 2005; Eckburg et al., 2005; Gill et al., 2006). In human duodenum, the density of bacteria is very low, typically less then 103 colony-forming units (CFU) per gram of contents, adhering to the mucosal surface or in transit. Along the jejunum and distal ileum, the density of bacteria ranges from 104 to 107 CFU per gram of intestinal content. Further down, in the colon, the numbers increase up to around 1012 CFU per gram of luminal content (Guarner, 2006). Within each compartment the bacteria live either close to the epithelial cells or in the lumen (Schultsz et al., 1999). Both DNA and RNA viruses are present in the intestine. In adult fecal samples, the majority of the DNA viruses are phages, while the majority of the RNA viruses are plant viruses. In the feces of infants, Adenovirus, Astrovirus, Bocavirus, Enterovirus, Norovirus, Picobirnavirus, Rotavirus, Sapovirus, and Torovirus were detected by using specific PCR methods (Breitbart et al., 2003; Zhang et al., 2006; Breitbart et al., 2008). Fungi and protozoa are also part of the intestinal flora, but little is known about their numbers and distribution. Population densities of intestinal bacterial species vary widely among individuals but stay fairly constant within an individual over time (Guarner and Malagelada, 2003; Eckburg et al., 2005; Gill et al., 2006; Li et al., 2008). The vast majority (over 99%) of human intestinal bacteria belong to the phyla: Firmicutes, Actinobacteria, Bacteroides, Proteobacteria and Fusobacteria. The proportions of the different phyla differ between compartments within the same individual (Wang et al., 2005; Eckburg et al., 2005; Li et al., 2008; Andersson et al., 2008). The greatest diversity of bacteria is found in colon, where the predominant bacterial genera are: Bacteroides, Clostridium, Fusobacterium, Eubacterium, Ruminococcus, Peptococcus, Peptostreptococcus, and Bifidobacterium. Other genera such as

- 27 -
Escherichia and Lactobacillus are present to a lesser extent (Ley et al., 2006; Eckburg et al., 2005; Artis, 2008). The colonization of gastrointestinal tract starts immediately at birth. A germ-free infant is immediately exposed to the microflora of the mother and of the surrounding environment irrespective of vaginal delivery or Caesarian section. In the beginning, the composition and temporal pattern of the microbial communities varies widely from baby to baby. By the end of the first year of life, the idiosyncratic microbial ecosystem in each baby, although still distinct, has converged toward a profile characteristic of the adult gastrointestinal tract (Palmer et al., 2007). By the second year of life the fecal microflora resembles that of adults (Mackie et al., 1999). The composition of the commensal bacterial communities is strictly regulated by competition for nutrients and space. This process is dependent on complex regulatory mechanisms, which involve the microflora itself, the surrounding environment, diet, and the innate and adaptive immunity (Sanderson, 1999; McCracken and Lorenz, 2001). The intestinal commensal microflora fulfills a host of useful functions, including digestion of unutilized carbohydrates and nitrogenous substrates; production of essential nutrients and short chain fatty acids (SCFAs); stimulating and modulating development and function of the intestinal and systemic immune system; repressing the growth of harmful microorganisms; training the immune system to respond only to pathogens and preventing allergy (Guarner and Malagelada, 2003; Macpherson and Harris, 2004; Calder et al., 2005; Sears, 2005; Artis, 2008). Alterations in the acquisition or the composition of the intestinal microflora may trigger or influence not only the course of various metabolic and inflammatory disease states but also susceptibility to or progression of, immune-mediated diseases. Ley and colleagues have demonstrated that the relative proportion of commensals of Bacteroidales is reduced both in obese individuals and in inbred mouse strains that are genetically predisposed to obesity. The intestinal commensal bacteria isolated from obese mice were more effective at releasing calories from food, and these bacteria transmitted this trait to germ free mice, resulting in greater deposition of adipose tissue. (Bäckhed et al., 2004; 2007; Ley et al., 2005; 2006; 2006; Turnbaugh et al., 2006). Currently, Kalliomäki and colleagues have indicated that alterations in the gut microbiota may be linked not only to obesity in children but also to the development of allergy (Kalliomäki et al., 2008). Alterations in the intestinal microflora have also been seen both in different patient groups and murine models of airway inflammation, arthritis, inflammatory bowel disease and necrotizing enterocolitis (Swidsinski et al., 2002; 2005; 2005; Guarner and Malagelada, 2003; Kalliomäki et al., 2001; 2008; Noverr and

- 28 -
Huffnagle, 2004; Tlaskalová-Hogenová et al., 2004; de la Cochetiere et al., 2004; Ott et al., 2004; Noverr and Huffnagle, 2005; De Hertogh et al., 2006). The intestinal microflora displays a wide range of metabolic activities, which could produce potentially harmful products including toxins and carcinogens that have been implicated in such conditions as multisystem organ failure, sepsis, colon cancer, and inflammatory bowel disease. Thus, a major factor in health is the balance of bacterial numbers, species and distribution (Guarner and Malagelada, 2003; Guarner, 2006). 1.9 Vibrio cholerae Vibrio cholerae is a facultative anaerobic, motile, gram negative, curved rod-shaped bacterium with a polar flagellum, first described and named V. cholerae by the Italian doctor, Filippo Pacini, in 1854 (Kaper and Morris, 1995). There are more than 200 serogroups of V. cholerae identified on the basis of their lipopolysaccharide O side chain antigenic structures (Osawa et al., 1997). Two serogroups of V. cholerae O1 and O139 infect humans and cause epidemic and pandemic cholera. The serogroup V. cholerae O1 is subdivided into two biotypes, classical and El Tor depending on biochemical properties and phage sensitivity. Each biotype can be divided into three serotypes depending on expression of three O-antigens (A, B, and C): Ogawa (A and B), Inaba (A and C) and Hikojima (A, B and C) (Kaper et al., 1995). Cholera is a severe diarrheal disease caused by V. cholerae infection in the small bowel, generally contracted by intake of contaminated food or water. WHO estimates 3 to 5 million cases worldwide every year and that more than 120,000 people die from cholera (WHO, http://www.who.int/topics/cholera/en/). V. cholerae is widely distributed in aquatic environments and cholera outbreaks are associated with contaminated food and water supplies. The seasonality of cholera has been associated with physico-chemical and biological factors (Lipp et al., 2002). However, many factors affect the survival of V. cholerae in aquatic environments such as attachment to plankton, entering into and resuscitation from viable but non-culturable state, and predation (Cottingham et al., 2003). Factors regulating the level of viable V. cholerae O1 and O139 in aquatic environment are still being investigated (Faruque et al., 2005). The infective dose of V. cholerae to cause cholera is approximately 108 live cells (Sack et al., 2004). Therefore, the bacteria need a biological reservoir in order to grow and survive in high concentrations to infect humans. Finding of aquatic reservoirs of V. cholerae is an important factor in the epidemiology of the disease.

- 29 -
V. cholerae can secrete virulence factors in two ways either soluble reagents or carried by vesicles (Mashburn-Warren and Whiteley, 2006). V. cholerae adhere to the intestinal epithelial lining by means of toxin-coregulated pili (TCP). Cholera enterotoxin (CTX) that is produced by the adherent vibrios and secreted across the bacterial outer membrane into the extracellular environment is a major agent involved in severe diarrheal disease. It is composed of a catalytically active heterodimeric A-subunit linked with a homopentameric B-subunit. The B subunit binds to the CTX receptor ganglioside GM1 on the intestinal epithelial cell leading to conformational alterations of the toxin structure which to release the A-subunit that enters inside the epithelial cell. The intracellular glutathione reduces the disulfide bond of the A subunit that dissociates into A1 and A2 (Lencer and Saslowsky, 2005; Chinnapen et al., 2007). The A1 subunit hydrolyzes nicotinamide adenine dinucleotide to ADP-ribose and nicotinamide. The ADP-ribose activates adenylate cyclase to hydrolyze ATP to cyclic AMP resulting in a substantial increase in intracellular concentrations of cyclic AMP, followed by a protein kinase -mediated phosphorylation of the major chloride channel of intestinal epithelial cells. Osmotic movement of large quantities of water accompanies the net increase in Cl− secretion into the intestinal lumen. The subsequent loss of water and electrolytes leads to the severe diarrhea and dehydration that are characteristic of cholera (Vanden Broeck et al., 2007). However, the loss or absence of the ctx gene coding for the CTX does not eliminate all disease symptoms from EI Tor strains (Ninin et al., 2000; Saka et al., 2008). Naturally occurring CTX-negative strains both of the EI Tor biotype and of non-O1 types have been associated with enterocolitis, as well as septicemia and extraintestinal infections (Anderson et al., 2004). Additionally, studies with CTX-negative vaccine strains resulted in mild to severe diarrhea and evidence of inflammation in human volunteers (Silva et al., 1996; Tacket et al., 1997). All these findings strongly suggested the existence of additional virulence factors associated with EI Tor and non-O1 V. cholerae pathogenesis. One of the additional virulence factors is the pore-forming Vibrio cholerae cytolysin (VCC), also referred to as hemolysin A (HlyA), which is encoded by a hlyA gene located on the V. cholerae chromosome II (Heidelberg et al., 2000). VCC/HlyA was shown to be synthesized and secreted as a pro-cytolysin monomer of about 80 kDa molecular weight that is activated by proteolytical cleavage by different proteases removing a 15 kDa polypeptide at the N-terminus (Alm et al., 1988; Nagamune et al., 1996; 1997). In the presence of cholesterol and ceramide-rich membranes active VCC/HlyA forms heptameric oligomers, which insert into lipid bilayers generating membrane pores with an internal diameter of 1.2 nm (Zitzer et al.,

- 30 -
1999; Harris et al., 2002; Pantano and Montecucco, 2006). It has been reported that VCC/HlyA is capable of causing several cytotoxic effects such as cell lysis and extensive vacuolization in vitro, depending upon the cell line studied and the toxin concentration (Moschioni et al., 2002; Alm et al., 1991; Figueroa-Arredondo et al., 2001; Coelho et al., 2000; Mitra et al., 2000; Zitzer et al., 1997; Valeva et al., 2008; Saka et al., 2008). Recently, Gutierrez et al. (Gutierrez et al., 2007) described that the vacuolization phenomenon associated to VCC/HlyA was an autophagic response of the cells against this toxin and was related to a protective cell response upon VCC/HlyA intoxication. CTX-negative and TCP-negative V. cholerae strains were isolated from patients suffering cholera-like diarrhea. In addition to gastroenteritis, non-cholera toxigenic V. cholerae infections can cause extraintestinal infections such as cellulitis, meningitis, wound infections, septicemia and bacteremia (Anderson et al., 2004; Hlady and Klontz, 1996; Ou et al., 2003). No definite virulence factor(s) has yet been identified to explain their disease producing ability. It has been proposed that VCC/HlyA is implicated in the pathogenesis of these diseases. However, studies so far are contradictory regarding the real role of VCC/HlyA (Alm et al., 1991; Figueroa-Arredondo et al., 2001; Ichinose et al., 1987; Fullner et al., 2002; Levine et al., 1988; Faruque et al., 2004). The finding that purified fractions of VCC/HlyA produced fluid accumulation in ligated rabbit ileal loops supports the notion of a role for VCC/HlyA in gastroenteritis (Ichinose Y, et al. 1987). Recently Saka and colleagues (Saka et al., 2008) reported that VCC/HlyA may trigger apoptotic cell death during infection in vivo and they suggested that the apoptotic cell death pathway plays the major role in pathogenesis of a CTX negative V. cholerae non-O1, non-O139 strain. Proteases constitute important bacterial virulence factors. They may function in invasion, immune evasion, nutrition, and reproduction in a host environment (Armstrong, 2006). Several proteases produced by pathogens are able to modulate other bacterial virulence factors, for example the V. cholerae hemagglutinin/protease (Hap) can process and activate VCC/HlyA (Booth et al., 1984; Nagamune et al., 1996). One newly described V. cholerae protease, PrtV (protease of V. cholerae), was shown to confer protection against grazing by natural bacteriovorous predators (Paper III). PrtV belongs to an M6 family of metallopeptidases, with the best characterized member of the family being an immune inhibitor A (InA or InhA) from Bacillus thuringiensis, a bacterium widely used in agriculture to control insect pests (Ogierman et al., 1997). Vaitkevicius et al. recently reported that PrtV is able to degrade mammalian extracellular matrix components and plasminogen (Vaitkevicius et al., 2008).

- 31 -
1.10 Celiac disease Celiac disease (CD) is a gluten-dependent enteropathy that occurs in genetically predisposed individuals (Troncone et al., 2008). Clinical symptoms. The classical clinical manifestations of CD include chronic diarrhea, weight loss and anemia, but while CD is primarily an intestinal disorder, intestinal symptoms may also be limited or even absent. A significant proportion of CD patients present with extra-intestinal symptoms, such as skin lesions, bone pains and fractures, infertility, apthous ulceration, and polyneuropathia etc (Farrell and Kelly, 2002; Fasano, 2005). However, there are also some atypical forms with negligible symptoms (silent CD patients), but still with a positive serologic test and small intestinal villous atrophy (Gasbarrini et al., 2008). Diagnosis. Traditionally, the diagnosis of CD has required a biopsy of the mucosa taken from proximal small intestine. Histological characteristics show villus atrophy, crypt hyperplasia, and an increased number of lymphocytes in the lamina propria and within the epithelium. In 1970, the European Society of Pediatric Gastroenterology, Hepatology, and Nutrition (ESPGHAN) established a three-biopsy procedure as a standard for the diagnosis of celiac disease. The three-biopsy procedure includes one initial biopsy on suspicion of CD during active disease, a second biopsy on gluten-free diet (GDF) and a third biopsy after challenge with gluten-containing diet (Meeuwisse, 1970). Although the three-biopsy procedure has been abandoned in many centers due to the introduction of serological tests, at least one biopsy from proximal small intestine is generally required. The serologic tests for diagnosis and screening of CD include anti-gliadin antibodies (AGA), anti-endomysium antibodies (EMA) and anti-tissue transglutaminase antibodies (Carlsson et al., 2001). Prevalence. Serologic screening studies have revealed that the prevalence of CD approaches 1% in the Caucasian populations, but may be even higher due to under-diagnosis. A growing portion of CD-diagnosis is being made in asymptomatic individuals (West et al., 2003; 2007; van Heel and West, 2006). CD is one of the most common chronic childhood disorders in Sweden. The prevalence of CD in our study series was at least 1.0%, but may be as high as 2.0% (Carlsson et al., 2001). In the total childhood population (0-15 years old) the annual incidence rate of clinically diagnosed CD was 10-45 cases per 100 000 person during the period 1973 to 2003. However, among children of different age groups the annual incidence rate varied considerably (Olsson et al., 2008). In the group of children under 2 years of age the annual incidence rate varied from 65 during 1973 to 1984, to around 200 during the time period 1985 to 1995 (Ivarsson et al., 2000), and

- 32 -
then to 98 from 2001 to 2003 (Olsson et al., 2008). In the group of children over 2 years of age, the incidence rate varied from 14 to 52. The observed differences in CD incidence rates in relation to year of presentation with CD possibly indicate that some environmental factors, such as an infectious component (Ivarsson et al., 2003), infant feeding style (breast feeding versus bottle feeding), or amount and/or timing of gluten administration in infancy (Ivarsson et al., 2000; 2002), influence disease incidence. Genetics. CD has a tendency to occur more often in some families. The concordance rate of CD is about 10% in first-degree relatives, about 30% in siblings and 75% to 100% in monozygotic twins, a rate higher than in any other condition with a multi-factorial basis (Sollid and Thorsby, 1993; Houlston and Ford, 1996; Greco et al., 2002). The principal determinants of genetic susceptibility are alleles within the highly variable human MHC complex, also called human leukocyte antigen (HLA) system1. Over 97% of CD patients are HLA-DQ2 or HLA-DQ8. The reason why these genes are associated with CD is that the peptide-binding groove of HLA-DQ2 and HLA-DQ8 favors the binding of gluten peptides with negatively charged residues at key anchor positions (Sollid, 2000; Hourigan, 2006; Kagnoff, 2007). However, up to 30% of the normal population, most of them eating food containing gluten, carry the HLA-DQ2 and/or HLA-DQ8 alleles, but do not develop CD (Sollid, 2000; Hourigan, 2006). Thus, exposure to gliadin and related prolamins from wheat, barley or rye and appropriate HLA-DQ haplotypes are necessary, but not sufficient to develop the disease. Apart from HLA genes, genes involved in intestinal permeability and immunity have been proposed to be risk factors for CD (Wapenaar et al., 2008; Hunt et al., 2008). There is no doubt that additional inherited and/or environmental factors and/or epigenetic factors are involved in contraction of CD (Green and Jabri, 2006). Gluten. Almost 60 years ago, the Dutch pediatrician, Wim Dicke recognized for the first time gluten as a trigger of CD (Losowsky, 2008). Gluten is found in the seeds of cereal grains of the tribe Triticeae and is a mixture of prolamins of a group of plant storage proteins. Prolamins are characterized by a high glutamine and proline content that dissolve only in alcohol solutions. They are resistant to proteases and peptidases in the intestine. 1The functions of MHC are to act as recognition molecules and to present antigen to effector cells in the immune system. There are two classes of MHC molecules, MHC class I, which is present on almost every nucleated cell in the body and MHC class II molecules, which is present on the cell surface of antigen presenting cells, such as DCs, B-cells, macrophages, and also on IECs of small intestine. HLA-DQ is a αβ-heterodimer of the MHC class II type. The α and β chains are encoded by HLA-DQA1 and HLA-DQB1 loci respectively and contain many alleles. There are 7 HLA DQ variants (DQ2 and DQ4 through 9). Different DQ isoforms can bind to and present different antigens to T-cells.

- 33 -
Some prolamins, notably wheat gliadin, and similar proteins and are only partly degraded resulting in long peptides in barley and rye induce CD in genetically predisposed individuals (Shewry and Halford, 2002; Green and Cellier, 2007; van Heel and West, 2006). There is also evidence of a direct toxic effect of gluten peptides in several biological models. However, the failure to control the inflammatory response may be one of the factors underlying gluten intolerance in these individuals. Many different gliadin peptides have been shown to express epitopes for gliadin-specific T-cell clones and also to be active when intestinal biopsies or isolated cells from intestinal biopsies of CD patients are challenged with them in vitro (Molberg et al., 1997; van de Wal et al., 1998; Sjöström et al., 1998; Mantzaris and Jewell, 1991; de Ritis et al., 1988; Maiuri et al., 1996). Transglutaminase. Because of their high glutamine content and specific sequence patterns, prolamins are excellent substrates for deamidation by tissue transglutaminase (tTG). Tissue transglutaminase or transglutaminase 2 (TG2), a member of the vast TG enzyme family, is a ubiquitously expressed multifunctional enzyme and present in almost all cell types in the body. In the intestine, tTG is found intra-cellularly in enterocytes and myofibroblasts. Extra-cellularly, this enzyme is located in the membranes of enterocytes and in the subepithelial connective tissue of the small intestine (Biagi, et al., 2006). tTG catalyzes an ordered and specific deamidation of certain glutamine residues of food gliadins and deamidated gluten peptides bind more effectively to HLA-DQ2 and HLA-DQ8 and trigger T cell responses (van de Wal et al., 1998; Johansen et al., 1996; Godkin et al., 1997; Anderson et al., 2000). Recently, Lammers and colleagues reported that one region of α-gliadin binds to the chemokine receptor CXCR3 and leads to MyD88-dependent zonulin release and increased intestinal permeability. Disruption of tight junctions allows entry for large peptides (Lammers et al., 2008). Immuno-pathology. Gluten reactive T lymphocytes in the jejunal mucosa play a central role in the pathogenesis of CD. The inflammatory process, mediated by T cells, leads to disruption of the normal tissue architecture such as villous atrophy, crypt hyperplasia, and increased infiltration by intraepithelial lymphocytes and dysfunction of the small intestinal mucosal lining, leading to malabsorption. To delineate local T lymphocytes response to gluten, our group investigated that the gene expression of cytokines, including IL-2, IFN-γ, TNF-α, IL-4, IL-10, and TGF-β1, in jejunal T cell subsets, from pediatric CD patients with active disease (i.e. untreated- and gluten challenged CD patients) and asymptomatic CD patients and healthy controls. The results showed that the high production levels of IFN-γ and IL-10 by intraepithelial T lymphocytes in CD patients could be the reasons for

- 34 -
both recruitment of intraepithelial lymphocytes and a leaky epithelium, leading to a vicious circle with amplified immune activity and establishment of intestinal lesions (Forsberg et al., 2002). The excessive production of IFN-γ is believed to drive the formation of intestinal lesions in active CD. CD94−CD8+αβ T cells within the epithelium constitute the major sourse of IFN-γ in active disease. The production of IL-10 may be a common feature of IELs producing proinflammatory cytokines, thereby attempting to limit inflammation in an autocrine fashion. Active CD is associated with increased frequencies of αβTCR+ and γδTCR+ IEL and CD4+αβTCR+ cells in the lamina propria of the small intestinal mucosa, including increased frequencies of IEL expressing the NK-receptors CD94 and NKG2D and lamina propria lymphocytes (LPL) expressing CD25 (Halstensen and Brandtzaeg, 1993; Forsberg et al., 2007). Other groups have reported that the cytokine network involved in CD may include IL-15, IL-18, IL-21, and perhaps IL-27, which can drive the inflammatory response through STAT1 and STAT5 pathways (Benahmed et al., 2007; Garrote et al., 2008). It could be argued that the pathogenic activity of T cells in the small intestinal mucosa of CD patients is caused by an impaired local T cell development and/or failure of adequate T cell receptor (TCR)-editing upon encounter with dietary antigens, leading to an abrogated development of oral tolerance. Our group also observed two signs of extrathymic TCR-genes rearrangement in the jejunal mucosal, including recombination activating gene-1 (RAG1) and preTα-chain gene. Expression levels of the four RAG1 mRNA splice forms and preTα-chain mRNA were determined in γδTCR+ IEL, αβTCR+ IEL, and TCR−CD2+CD7+ IEL of children with CD and controls. The results show that (1) RAG1 is normally expressed in both TCR+ and TCR− IEL while preTα-chain is only expressed in the immature TCR− IEL and (2) “extrathymic” splice forms of RAG1 mRNA and preTα-chain mRNA are down-regulated in IEL of CD patients regardless of disease activity suggesting that CD patients have impaired TCR-gene rearrangement in their jejunal mucosa which could be a contributing factor for CD (Bas et al., 2003; 2009). Intestinal microbiota. It has been suggested that intestinal bacterial infection or microflora alterations might be involved in the pathogenesis of CD. An intestinal infection could cause inflammatory response and damage to the intestinal mucosa, thereby raising the activity of tTG, possibly increase the permeability of gluten peptides across the epithelial barrier, and stress the epithelial cells to express major inflammatory factors. By scanning electron microscopy, our group observed that rod-shaped bacteria were associated with the intestinal epithelium in children with CD but not in controls

- 35 -
(Forsberg et al., 2004). Furthermore, by studying short chain fatty acids in fecal samples from CD children and healthy controls, Tjellström et al. found an altered pattern of these fatty acids in both treated and untreated CD, as compared to controls (Tjellström et al., 2005; 2007). They suggested that their findings might reflect a deviant gut flora in CD patients. Moreover, by fluorescent in situ hybridization (FISH) coupled with flow cytometry, Nadal and co-workers reported that the higher incidence of Gram-negative and potentially proinflammatory bacteria in the duodenal microbiota of celiac children was linked to the symptomatic presentation of the disease and could favor the pathological process of the disorder (Nadal et al., 2007). Alterations of the intestinal microflora in fecal samples from CD children were demonstrated by FISH (Collado et al., 2007) and by PCR coupled with denaturing gradient gel electrophoresis (DGGE) analysis (Sanz et al., 2007). To understand the relationship between intestinal bacteria and CD, it is necessary to perform a detailed characterization of the intestinal microflora from both patients with CD and healthy individuals. Treatment. At present, the only effective treatment for CD is a lifelong strict gluten-free diet (Kupper, 2005). In practice, however, this generally means a supposedly harmless level of gluten rather than a complete absence (Akobeng and Thomas, 2008). The strict gluten avoidance allows the intestinal mucosal restitution and improvement of symptoms in most cases and can also eliminate the heightened risk of osteoporosis and intestinal cancer (Treem, 2004).

- 36 -
2. Aims of the Thesis
The purposes of this thesis were to study the composition of the microbiota
at the proximal small intestine in children with or without celiac disease and
also to study the immune responses of intestinal epithelial cells to different
stimuli especially Vibrio cholerae virulence factors. The specific aims were:
• To characterize the composition of microflora at proximal small
intestine in children with celiac disease and controls by culture-
independent and culture-dependent methods.
• To identify the rod-shaped bacterium/a previously detected by scanning
electron microscopy in proximal small intestine of children with celiac
disease.
• To establish an in vitro model for studies of innate defense mechanisms
of human intestinal epithelial cells.
• To study the roles of a Vibrio cholerae protease, PrtV and its putative
substrate Vibrio cholerae cytolysin, VCC as secreted pathogenic factors
for human intestine in Vibrio cholerae infection.

- 37 -
3. Results and Discussion
3.1 Proximal small intestinal microbiota and identification of rod-shaped bacteria associated with childhood celiac disease (Paper I) As discussed earlier, the fact that an individual expresses HLA-DQ2 or HLA-DQ8 on her/his immune cells and is exposed to gluten in the diet is not sufficient to develop CD. Obviously other factors, genetic, epigenetic or environmental, are also required to precipitate disease. One such factor may the composition of the intestinal microbiota. Indeed a previous investigation in our laboratory, using scanning electron microscopy (SEM), revealed that rod-shaped bacteria were associated with the epithelium of proximal small intestine of children with CD in contrast to controls (Forsberg et al., 2004). Moreover, even children with CD on a gluten-free diet and with no clinical symptoms of CD retained these bacteria indicating that it may be an inherent property to harbor rod-shaped bacteria in the small intestine. In order to identify these bacterium/a and to find possible links between specific bacterial populations and the development of CD, we have determined the composition of the bacterial flora of proximal small intestine in children with CD and controls, using culture-dependent and culture-independent methods. In total, we analyzed biopsy samples from 63 children, including 18 biopsies from clinical controls, (3 of them from healthy siblings of CD patients) and 33 biopsies from children with CD collected during 2004 to 2007. The remaining 12 biopsies were “historical” biopsies from children with CD born during the Swedish CD epidemic. These biopsies had been stored frozen at -80°C. The fresh biopsies were washed repeatedly with PBS containing dithiothreitol (DTT) to dissolve the mucus layer. Among these biopsies, 19 were from untreated CD patients, 12 from CD patients treated on a gluten-free diet and 3 from CD patients challenged with gluten containing diet. To allow different types of analysis, biopsies were divided into 4 parts, each part treated as required by the particular method utilized. The proximal small intestine is a unique ecological niche. As one of the culture independent methods we used sequencing of the bacterial 16S rRNA gene. Totally we collected 2,247 sequences of an about 440 base-pair long fragment of the bacterial 16S rRNA gene from 63 individuals. In Table 1 the data are displayed as the percentage of bacteria in proximal small intestine belonging to different phyla. Here we have pooled the results from CD and controls since, as discussed below there was no major difference between

- 38 -
CD and controls in biopsies collected during 2004-2007. Comparing with the microbiota from other sites, the composition of the bacterial biota in proximal small intestine is different from those of the other sites indicating that it is a distinct ecological niche. At the phylum level, Firmicutes, Proteobacteria and Actinobacteria dominate the bacterial community while Bacteriodetes and Fusobacteria constitute small populations. One hundred and sixty-two Genus Level Operational Taxonomic Units (GELOTUs), named jej201 to jej362, of six different phyla were identified. Thirty-five sequences were sufficiently different from all previously known bacterial sequences (i.e. ≤ 97% homology) to most probably represent previously uncharacterized phylotypes (Figure 2 and supplementary Table 2 in Paper I). The most diverse phyla were Firmicutes and Proteobacteria the least diverse phylum was TM7. Composition and properties of the normal biota of proximal small intestine. These results also show that the mucosa-associated microbiota in proximal small intestine of Swedish children with normal intestinal mucosa is dominated by two bacterial genera namely Streptococcus and Neisseria. Together with a limited number of other genera, i.e. Veillonella, Gemella, Actinomyces, Rothia and Haemophilus they constitute almost the entire bacterial community. In total 42 different GELOTUs were detected of which 30 had not been described previously in proximal small intestine, 18 of which are unknown. Our findings are in general agreement with, but substantially expand previous studies on adults using culture-based methods (Sullivan et al., 2003; Zilberstein et al., 2007) also showing a predominance of Streptococci. To ensure that bacteria identified by 16S rDNA sequencing and electron microscopy are alive and potentially able to exert biological effects at the epithelial lining, we attempted to cultivate bacteria from biopsies. Ten consecutive biopsies collected during 2007 were investigated using selective conditions for anaerobic and aerobic growth. Live bacteria were identified in all ten biopsies. Bacteria from eight of the biopsies (5 CD patients and 3 controls) were subjected to quantitative cultivation methods and characterized by standard bacteriological tests. The results (Table 3 in Paper I) demonstrated that all genera identified by 16S rDNA sequencing in controls could be cultivated except Gemella, certifying that they indeed were alive. Most of the bacteria in the “majority community” of controls were cocci or coccobacilli. The only rods regularly found were bacteria of the genus Actinomyces. The bacteria are to a significant degree associated to the mucosal surface since they were identified in biopsies washed with buffer containing DTT, a solution that will dissolve the outer mucin layer. We therefore conclude that the bacteria of the “majority community” are not just

- 39 -
transient passengers in the gut lumen but constitute an organ-specific biofilm. It is reasonable to assume that it is protective, limiting access for potential pathogens to the mucosal surface and possibly influencing the maturation of the local immune system. Apparently lactobacilli are transient visitors in proximal small intestine of children since they were not found regardless of detection method (Table 4 and Figure 2 in Paper I) while abundantly seen when the intestinal content was included (Sullivan et al., 2003; Zilberstein et al., 2007). Microbiota of children with CD, results based on biopsies collected during 2004-2007. Using bacterial 16S rDNA sequencing we compared the composition of the bacterial flora in non-CD children with that of children with CD (supplementary Table 2 in Paper I). There was no statistically significant difference between CD patients and controls in frequency of biopsies containing any of the 38 different bacterial genera discovered in the material. However, a difference between the two groups was that Haemophilus was more common and Neisseria less common in CD than in controls. Although it cannot be excluded that subtle differences exist that would have been revealed if a larger number of clones had been analyzed we conclude that the proximal small intestine microbiota is essentially the same in the two groups. Moreover, dividing the group of CD patients into those, which were on gluten-free diet and those, which presented with CD we did not find major differences in the microbiota either. Similarly only small differences were seen in the flora between young untreated- and older untreated CD patients. Almost all of these biopsies were also investigated by SEM for presence of rod-shaped bacteria. Only one biopsy in each group contained such bacteria. This finding was unexpected since the previous investigation showed that rod-shaped bacteria were present in 20-40% of the children with CD in contrast to controls, which essentially lacked such bacteria (Forsberg et al., 2004). From this study we tentatively conclude that the differences seen between CD and controls in the previous study cannot be seen in the collection of current biopsies. Microbiota of children with CD, results based on “historical” biopsies. Fortunately parts of nine CD-biopsies, which showed presence of rod-shaped bacteria by SEM (Figure 3 in Paper I) in the earlier study and which had been stored frozen at -80°C were available for analysis. When these biopsies were compared with CD biopsies lacking rod-shaped bacteria as defined by SEM, three differences stand out: bacteria similar to unclassified Clostridales, Prevotella species and Actinomyces graevenitzii were

- 40 -
significantly more common in the former biopsies than in the latter (Table 1 in Paper I). It is interesting to note that all 3 bacteria are rods. At the phylum level the two groups differed in that particularly Bacteroidetes and Fusobacteria were more common in the group of “historical” CD biopsies. It could be said that the microbiota is more colon-like. It should be pointed out that this Table displays only those genera that were different either in frequency of positive biopsies or number of clones between the two groups. Thus, bacteria similar to Streptococcus pseudopneumonie and Neisseria polysaccharea both occurred at high frequencies and in all biopsies of both groups. However, there was no statistically significant difference between the groups. Since the frozen biopsies could not be washed it could be argued that the difference between the two groups was due to lack of washing in one group and buffer/DTT washing in the other. We therefore studied the relative binding affinity of different bacterial genera to the epithelial lining. Ten DTT-washed biopsies and their corresponding wash-fluids were analyzed. The DTT-washed biopsy is expected to contain bacteria actually attached to, or inside, the mucosal tissue while the wash fluid would contain bacteria from the gut lumen or from inside the mucous layer that covers the epithelial cells. An adhesion index i.e. bacteria in biopsy/ bacteria in washings could be calculated (Table 2 in Paper I). Seven genera showed preferential binding to the epithelial lining, including jej203 (similar to unclassified Clostridiales species), jej273 (similar to Actinomyces species) and jej 308 (similar to Prevotella species). Taken together these results suggest that any or all of the bacteria of the genera Clostridium, Prevotella and Actinomyces could be the rod-shaped bacteria seen by SEM in the earlier study. On the basis of SEM morphology (Figure 3 in Paper I) it is, however, not possible to discriminate between these possibilities. Most interestingly, bacteria of the three genera mentioned above could be isolated from two current CD biopsies (Table 4 in Paper I). These biopsies were from a girl born in 1995 and a boy born in 1994, both on gluten-free diet when the biopsies were taken in 2007. Detailed analysis revealed that the Clostridium species and one of the Prevotella species were hereto-unknown species while the Actinomyces species was identified as Actinomyces graevenitzii. Rod-shaped bacteria probably expressing both a mucinase, allowing passage through the mucin layer, and forming an intimate contact with microvilli of jejunum epithelial cells was detected by transmission electron microscopy (TEM) (Figure 4 in Paper I). Four current biopsies were analyzed by TEM. In two of them (1 CD patient and 1 sibling to a CD patient), cocci and rods were found in close contact with the microvilli. Presently we do not know whether the isolated bacteria have mucinase activity and can bind to the proximal small intestine mucosa.

- 41 -
Further work with artificial tight monolayers of human small intestine as developed for colonocytes in paper II will hopefully tell us whether any of these bacteria will bind to and affect the cells biologically. Relationship to Swedish CD epidemic. Investigations have shown an epidemic pattern of childhood CD in Sweden during 1985-1996, in children younger then two years of age (Ivansson et al., 2001; Olssen et al., 2008). This epidemic coincided with not only changes in the recommendations of gluten-containing food and breast-feeding to infants (Olssen et al., 2008) but also with the frequency of CD patients with rod-shaped bacteria associated to the small intestinal mucosa (Forsberg et al., 2004; Paper I). It should be noted that the genetic background in the population before, during and after the epidemic is almost the same. These results support the notion of an infectious component in the pathogenesis of CD. The intestinal microflora is constructed during first two years after birth and the composition of commensal bacterial communities is regulated by competition for nutrients and space (Mackie et al., 1999; Palmer et al., 2007). This process is dependent on complex regulatory mechanisms, which involve the microbes themselves, diet, the surrounding environment, and innate and adaptive immunity (Sanderson, 1999; McCracken and Lorenz, 2001). The feeding-style and recommended food are capable of altering the composition of intestinal microflora. The intestinal microflora participates in stimulating and modulating the development and function of the intestinal and systemic immune system, training the immune system to respond to pathogens and preventing allergy and thus, possibly also CD (Guarner and Malagelada 2003; Macpherson and Harris, 2004; Sears, 2005; Artis, 2008). Our results suggest that changes in the composition of the proximal small intestinal microflora, especially the presence of adhering rod-shaped bacteria is one factor contributing to contraction of CD.
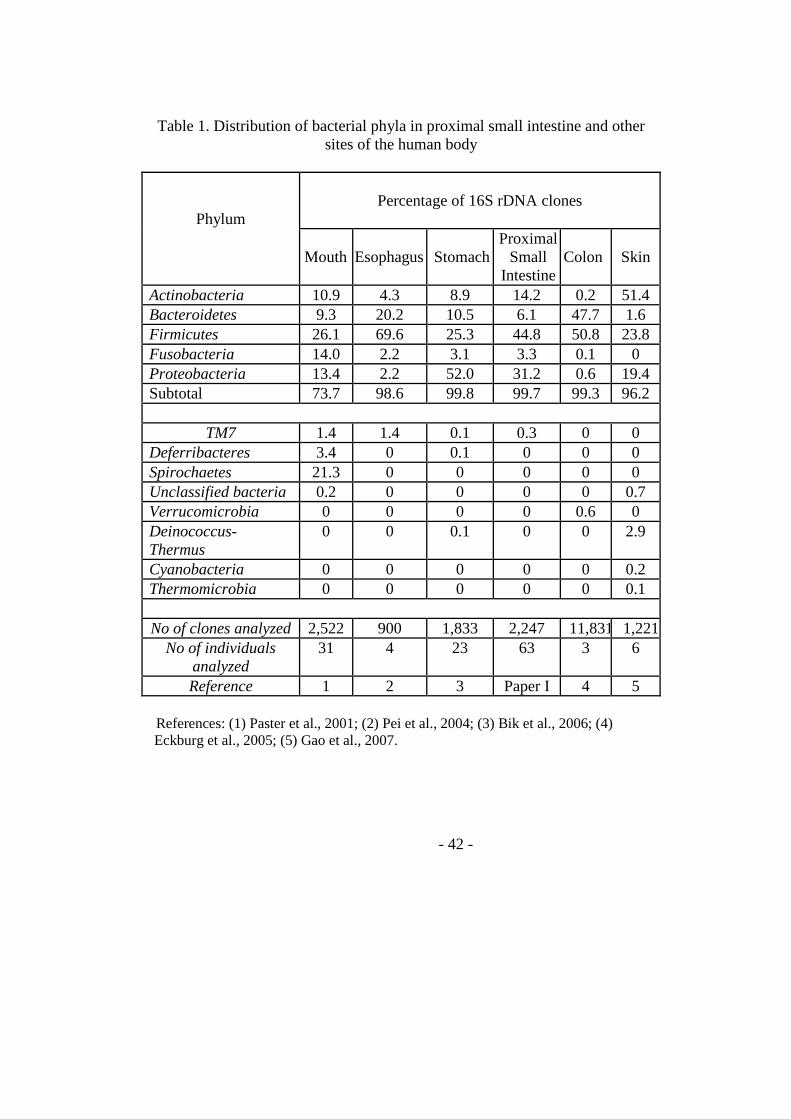
- 42 -
Table 1. Distribution of bacterial phyla in proximal small intestine and other
sites of the human body
Percentage of 16S rDNA clones
Phylum
Mouth
Esophagus
Stomach
Proximal Small
Intestine
Colon
Skin
Actinobacteria 10.9 4.3 8.9 14.2 0.2 51.4 Bacteroidetes 9.3 20.2 10.5 6.1 47.7 1.6 Firmicutes 26.1 69.6 25.3 44.8 50.8 23.8 Fusobacteria 14.0 2.2 3.1 3.3 0.1 0 Proteobacteria 13.4 2.2 52.0 31.2 0.6 19.4 Subtotal 73.7 98.6 99.8 99.7 99.3 96.2
TM7 1.4 1.4 0.1 0.3 0 0
Deferribacteres 3.4 0 0.1 0 0 0 Spirochaetes 21.3 0 0 0 0 0 Unclassified bacteria 0.2 0 0 0 0 0.7 Verrucomicrobia 0 0 0 0 0.6 0 Deinococcus-Thermus
0 0 0.1 0 0 2.9
Cyanobacteria 0 0 0 0 0 0.2 Thermomicrobia 0 0 0 0 0 0.1
No of clones analyzed 2,522 900 1,833 2,247 11,831 1,221
No of individuals analyzed
31 4 23 63 3 6
Reference 1 2 3 Paper I 4 5 References: (1) Paster et al., 2001; (2) Pei et al., 2004; (3) Bik et al., 2006; (4) Eckburg et al., 2005; (5) Gao et al., 2007.

- 43 -
3.2 Contribution of intestinal epithelial cells to innate immunity of the human gut - studies on polarized monolayers of colon carcinoma cells (Paper II) In the intestine a single layer of epithelial cells separate the bodily tisssue from the external environment. The functional intactness of this epithelium is essential for survival. Not only is the epithelium a physical barrier protecting the host from microbial invasion it also provides direct and indirect defence functions since the epithelium must be seen as a component of both innate and adaptive immunity. The purpose of this work was to establish an in vitro model for functional studies of human intestinal epithelium in innate immunity. We attempted to cultivate several colon carcinoma cell lines in a two-chamber system in which the chambers are connected by a semi-permeable membrane. Cells were grown on the semi-permeable membrane of the upper chamber and medium added both to the lower and upper chambers. As the cells grow some cell lines form a tight monolayer of polarized epithelial cells. To monitor development of a tight cell layer transepithelial resistance (TER) is measured. Two colonic carcinoma cell lines, T84 and Caco2 cells could be successfully transformed to tight monlayers. When TER reached a plateau both T84 and Caco2 cells formed a single layer of polarized epithelial cells, developed microvilli at the apical side and tight junctions between the cells (Figure 1 and Figure 2 in Paper II). For T84 cells, the monolayers exhibited several features typical for the mature colonic epithelium. Those were: 1) a distinct apical glycocalyx that expressed large amount of CEA, which is also seen on colonocytes at the free luminal surface of normal colon (Frängsmyr et al., 1999; Hammarström, 1999; Hammarström and Baranov, 2001); 2) cells with mucous granules that are characteristic for intestinal goblet cells (Figure 2 in Paper II), 3) expression levels of mRNA for the apical surface components/glycocalyx components CEACAM1, CEA, CEACAM6 and CEACAM7 similar to those expressed in normal colon IECs; 4) a normal ratio between CEACAM1-L and CEACAM1-S. As CEACAM1-L has tumor suppressor functions (Hammarström, 1999; Kunath et al., 1995; Öbrink, 1997) it is likely that malignant features of the tumor cells are repressed in the polarized T84 cells and that they indeed can be expected to give a good reflection of normal IEC functions (Table 1 in Paper II). In one respect, however, the tight monolayer was different from normal mature colon epithelium namely in the expression levels of MUC2 mRNA and number of goblet cells, which were significantly lower.

- 44 -
The Caco2 tight monolayer culture was different. It showed barely any CEA expressing microvilli/glycocalyx and the levels of mRNA for CEA, CEACAM6 and CEACAM7 were low while the CEACAM1 mRNA levels and CEACAM1-L/CEACAM1-S ratio were normal. The MUC2 mRNA level was low compared to normal IEC indicating a low frequency of fully differentiated goblet cells. Thus, Caco2 tight monolayer culture most likely is a model for immature columnar epithelial cells in the lower part of the colonic crypts. The polarized monolayers were challenged with laboratory strains of Gram+ (B. megaterium and M. luteus), Gram− (E. coli and S. typhimurium) bacteria and E. coli derived LPS from the apical side in order to mimic microbial attack from the gut lumen. They were also challenged by proinflammatory IFN-γ, TNF-α and IL-1β from the basolateral side to mimic signaling from immune cells in gut immune reactions. Immune responses were estimated as changes in mRNA expression levels for mucous component MUC2, glycocalyx components the four CEA family members and MUC3, antimicrobial factors hBD1, hBD2, hBD3, and lysozyme, chemokine IL-8, and cytokines IL-6 and TNF-α. mRNA levels for these protein components were determined by specific real-time quantitative reverse transcriptase-polymerase chain reactions with copy standards. We found that both tight monolayer cells generally were unresponsive to challenge with these bacteria except for Bacillus megaterium, which significantly increased hBD2 mRNA levels in both cell lines. Several bacterial strains and LPS also upregulated TNF-α levels in T84 tight monolayer cells. Tight monolayer cells responded to cytokine challenge suggesting awareness of basolateral attack. Thus, TNF-α induced markedly increased levels of IL-8 and of TNF-α itself in both cell lines suggesting that the epithelium participates in recruitment and activation of immune cells in the underlying mucosa in vivo. IFN-γ and TNF-α challenge most interestingly increased the level of CEACAM1-L mRNA in T84 monolayer cells while other CEACAMs or MUC3 were not affected. Although not statistically significant TNF-α also upregulated the CEACAM1-S form. Thus if the ratio CEACAM1-L/CEACAM1-S is calculated it is evident that the two cytokines have different effects, IFN-γ selectively upregulating the form with down-regulatory properties. Increased CEACAM1 expression may influence epithelial function and communication between epithelial cells and intraepithelial lymphocytes. In future studies we will use this system for biological analysis of bacteria associated to CD.

- 45 -
3.3 Role of V. cholerae protease, PrtV - from C. elegans to human intestine 3.3.1 PrtV protects V. cholerae from natural predator grazing and is responsible for killing of infected C. elegans (Paper III) By using C. elegans as a potential model system for studies of cholera diarrhea we found that C. elegans with an intestinal infection were killed by the bacterium. The killing efficiency was dependent on quorum sensing regulators. Thus, the main virulence factor of V. cholerae, cholera toxin (CTX) did not affect the C. elegans killing, while hapR mutated strains were attenuated (Figures 1 and 3 in Paper III). After testing in this C. elegans killing assay it was evident that only the depletion of a new protease, PrtV clearly attenuated the ability of V. cholerae to cause a lethal infection (Figure 3 in Paper III). Further studies on PrtV showed that it belongs to family of metallopeptidases, M6 (Vaitkevicius et al., 2008), with the best characterized member of the family being the immune inhibitor A (InA) from Bacillus thuringiensis, a bacterium widely used in agriculture to control insect pests (Ogierman et al., 1997). Moreover, PrtV conferred protection against grazing by natural bacteriovorous predators. It was recently reported that PrtV is a Zn2+ dependent protease able to degrade mammalian extracellular matrix components and plasminogen (Vaitkevicius et al., 2008). In order to study whether PrtV was also toxic to human intestine an in vitro model system consisting of polarized tight monolayers of the human colonic adenocarcinoma cell line T84 was utilized (described in Paper II). The chemokine IL-8 was used as a marker of inflammatory response.
Unexpectedly, supernatants of the ΔprtV derivates did not induce increased permeability or IL-8 expression (Figure 6 and supplementary Figure 11 in Paper III). These results suggest a role for this protease in modulating the human intestinal epithelial cell response. Considering the proteolytic activity of the PrtV protein, the observed effect could be indirect and might result from processing of other secreted V. cholerae factor(s). 3.3.2 V. cholera cytolysin (VCC) is the secreted proinflammatory factor and its activity is modulated by PrtV (Paper IV) To throw light on the contrasting effects of PrtV in human intestinal epithelial cells compared to in invertebrate intestine, we aimed at identifying secreted component(s) of V. cholerae O1 strain C6706 that induces an epithelial inflammatory response in human intestinal epithelial cells and

- 46 -
define whether the active component(s) are substrates for PrtV. Culture supernatants from V. cholerae C6706 and genetically modified strains were analyzed for capacity to induce changes in cytokine production, permeability and cell viability in the polarized tight monolayer in vitro model for human intestinal epithelium. The culture supernatants were also immunochemically characterized and monitored for capacity to lyse red blood cells. Culture supernatants from the V. cholerae ∆prtV strain when grown in Luria-Bertani broth caused permeability increase and increased expression of IL-8 and TNF-α while supernatants from wild-type bacteria did not (Paper III and IV). Experiments with supernatants from CTX deletion mutants showed that CTX was not the stimulatory component. Thus, PrtV influences the stability of one or more secreted factors other than CTX. This influence by PrtV was abolished when supernatants from bacteria cultured in brain heart infusion broth were used (Figure 2 in Paper IV), suggesting that this broth in some way inhibits PrtV degradation of the factor(s) causing the inflammatory response. VCC was found to be the sole active component causing the inflammatory response as revealed by VCC deletion mutant experiments (Figure 2 in Paper IV). The hemolytic activity of supernatants from wild-type V. cholera and its mutants showed exactly the same pattern (Figure 3 in Paper IV). Furthermore pure VCC protein was able to induce an inflammatory response of the same magnitude as the culture supernatant (Figure 5 in Paper IV). High doses of VCC were cytotoxic for epithelial cells. The inflammatory effect of VCC could be abolished by treatment with PrtV (Figure 6 in Paper IV). That PrtV is the major component causing degradation of VCC is indicated by the fact that supernatants from a derivative carrying an inframe deletion mutation in the gene for a regulator of V. cholerae proteases expression (∆hapR) caused the same changes in the inflammatory response as the ∆prtV mutant. VCC most likely is a pathogenetic factor in cholera. At high concentrations it caused hemolysis and epithelial cell death, which is in line with previous studies in a mouse model (Olivier et al., 2007). At low doses VCC induced an inflammatory response in intestinal epithelial cells that included increased epithelial permeability and induction of IL-8 and TNF-α suggesting that intestinal epithelial cells initially execute a front-line defense reaction to V. cholerae infection involving recruitment of immune cells to the infected site by IL-8 and activation of epithelial cells, intraepithelial lymphocytes, and lamina propria macrophages by TNF-α. Paracellular leakage due to decreased tight junction stability and/or formation of

- 47 -
transmembrane pores by VCC could be the mechanism behind the increased permeability in tight monolayers seen here. The reason why the bacterium produces VCC and PrtV simultaneously is unclear but the difference in VCC activity in different types of broth suggests that this might be an adaptation for different environments. In the human intestine PrtV is probably inactivated either by chelators or by high protein/peptide content allowing VCC to exert its toxic actions and yield access for the bacterium to the bodily tissues.

- 48 -
4. Conclusions
1. A unique bacterial flora inhabits the normal proximal jejunum of children. This flora is not just transient passengers in the jejunum but constitutes an organ-specific biofilm. The predominant bacterial genera are Streptococcus, Neisseria, Veillonella, Gemella, Actinomyces, Rothia and Haemophilus. 2. The rod-shaped bacteria previously found in children with CD during the CD epidemic is likely to be one or more of a previously unidentified unclassified-Clostridales species, a previously unidentified Prevotella species or a pathogenic variant of Actinomyces graevenitzii. 3. The polarized tight monolayer of both colon carcinoma cell lines T84 and Caco2 appear to be suitable in vitro models for studies of innate defense mechanisms of human intestinal epithelium. The two cell lines seem to represent intestinal epithelial cells at two different sites in the colonic mucosa, namely the luminal surface in the case of T84 and the lower crypts in the case of Caco2 cells. 4. Polarized intestinal monolayer cells seem “more resistant” to bacterial attack than ordinary tissue culture cells. 5. The inhibitory and tumor suppressive variant of CEACAM1, namely CEACAM1-L, is strongly upregulated in polarized T84 cells by stimulation with IFN-γ. This indicates that intraepithelial lymphocytes can influence epithelial cell proliferation during an immune response. 6. Vibrio cholera cytolysin (VCC) was found to be solely responsible for induction of an inflammatory response in intestinal epithelial cells that includes increased epithelial permeability and production of IL-8 and TNF-α. 7. PrtV degrades VCC thereby modulating the inflammatory and cytotoxic response.

- 49 -
5. Acknowledgements
If life is a long march, this thesis is the milestone of my journey. I have not traveled lonely during my trip through the years. There are so many people who have made this trip easier for me. I wish to express my sincere gratitude and deep appreciation to all of you who helped me along the way. First of all, I would like to thank my supervisors: Professor Sten Hammarström and Professor Marie-Louise Hammarström for providing me this opportunity to share your knowledge about mucosal immunology and favorite molecules CEACAMs. Thank you for your endless support, enthusiasm, encouragement and understanding both in academic field and daily life during these years. Professor Olle Hernell, thank you for support, enthusiasm, encouragement, discussion and the cooperation in the celiac disease project. Many thanks to Dr. Vladimir Baranov for sharing your knowledge and for all the nice electron micrographs, and for giving constructive suggestions. Professor Torgny Stigbrand for your hospitality and all the terrific foods that we had at your place. Kristina Lejon for interesting discussion. Lucia Mincheva Nilsson, thank you for interesting discussion and suggestions. Many thanks to Lena Hallström-Nylén for making everyday life much easier for me. Anne Israelsson, Marianne Sjöstedt, and Elisabeth Granström for your invaluable help on my bench work and for well-being concerns. Birgitta Sjöberg for making everyday life at the lab so much easier for all of us. I thank David, Lina, Veronika, Mia, Lotta, Aziz, Rifat, Therese, Erica and Christoffer for your friendship. I will always remember the PhD parties and Journal Club! I wish you good luck with your studies and a wonderful time. Many thanks also to the former group members and colleagues: Annas, Ali Sheikholvaezin, Arman, Silvia, Niklas, Homa, Guang-Qian, Bi-Feng and Lili for friendship and happiness. Thanks to all members at the Department of Immunology for friendship and for a series of nice dinners with excellent food and discussions.

- 50 -
Göte Forsberg, Maria Hedberg, Per Hörstedt, Mirva Drobni, Olof Sandström, Ingegerd Johansson and Elena Lundmark for cooperation and coauthorship. Sun Nyunt Wai and your group members: Karolis Vaitkevicius, Barbro Lindmark, Tianyan Song, Pramod Kumar Rompikuntal, for collaboration and coauthorship. Sucharit Bhakdi for coauthorship. Special thanks to all Chinese friends in Umeå, Teacher Gu, families of Zhang Jiexie’s, Xiong’s, Mei Yafang’s, Chen Sa’s, Gong’s, and Lao Lai ’s are acknowledged for help and friendship. I am especially grateful to Prof. Jin Taiyi for introducing me to Umeå and for never-failing encouragement and support. I would like to thank Lu Jian and Yuan Ming and Xiao Hu, Bo, Jufang, Zijiu, Qingyin and Haiyang, Daping and Yaoyi, Changchun and ShengMing, Lao Huang, Junfa, Shenyue, Yihuai, Xuhao-for all your friendship and help. I wish to thank my friends and relatives who always concern me for patience and encouragement. I also wish to thank my former colleagues in Zunyi Medical College for patience, enthusiasm, encouragement and friendship. My mom and my sisters and young brother, I love you! This thesis is dedicated to the loving memory of my father. Last but not least, Jun and Ran, without your love, it would have been impossible to show this thesis here.

- 51 -
6. References
Abreu MT, Fukata M, Arditi M. 2005. TLR signaling in the gut in health and disease. J Immunol. 174:4453-60.
Abreu MT, Vora P, Faure E, Thomas LS, Arnold ET, Arditi M. 2001. Decreased expression of Toll-like receptor-4 and MD-2 correlates with intestinal epithelial cell protection against dysregulated proinflammatory gene expression in response to bacterial lipopolysaccharide. J Immunol. 167:1609-16.
Acheson DW, Luccioli S. 2004. Microbial-gut interactions in health and disease. Mucosal immune responses. Best Pract Res Clin Gastroenterol. 18:387-404.
Agostini L, Martinon F, Burns K, McDermott MF, Hawkins PN, Tschopp J. 2004. NALP3 forms an IL-1beta-processing inflammasome with increased activity in Muckle-Wells autoinfl ammatory disorder. Immunity. 20:319-25.
Akira S, Uematsu S, Takeuchi O. 2006. Pathogen recognition and innate immunity. Cell. 124:783-801.
Akobeng AK, Thomas AG. 2008. Systematic review: tolerable amount of gluten for people with coeliac disease. Aliment Pharmacol Ther. 27:1044-52.
Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. 2001. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 413:732-8.
Al-Lamki RS, Wang J, Skepper JN, Thiru S, Pober JS, Bradley JR. 2001. Expression of tumor necrosis factor receptors in normal kidney and rejecting renal transplants. Lab Invest. 81: 1503-15.
Alm RA, Mayrhofer G, Kotlarski I, Manning PA. 1991. Amino-terminal domain of the El Tor haemolysin of Vibrio cholerae O1 is expressed in classical strains and is cytotoxic. Vaccine. 9:588-94.
Alm RA, Stroeher UH, Manning PA. 1988. Extracellular proteins of Vibrio cholerae: nucleotide sequence of the structural gene (hlyA) for the haemolysin of the haemolytic El Tor strain 017 and characterization of the hlyA mutation in the non-haemolytic classical strain 569B. Mol Microbiol. 2:481-8.
Andersson AF, Lindberg M, Jakobsson H, Bäckhed F, Nyrén P, Engstrand L. 2008. Comparative analysis of human gut microbiota by barcoded pyrosequencing. PLoS ONE. 3:e2836.
Anderson AM, Varkey JB, Petti CA, Liddle RA, Frothingham R, Woods CW. 2004. Non-O1 Vibrio cholerae septicemia: case report, discussion of literature, and relevance to bioterrorism. Diagn Microbiol Infect Dis. 49:295-7.
Anderson RP, Degano P, Godkin AJ, Jewell DP, Hill AV. 2000. In vivo antigen challenge in celiac disease identifies a single transglutaminase-modified peptide as the dominant A-gliadin T-cell epitope. Nat Med. 6:337-42.
Andrianifahanana M, Moniaux N, Batra SK. 2006. Regulation of mucin expression: mechanistic aspects and implications for cancer and inflammatory diseases. Biochim Biophys Acta. 1765:189-222.
Arend WP, Palmer G, Gabay C. 2008. IL-1, IL-18, and IL-33 families of cytokines. Immunol Rev. 223:20-38.

- 52 -
Armstrong PB. 2006. Proteases and protease inhibitors: a balance of activities in host-pathogen interaction. Immunobiology. 211:263-81.
Artis D. 2008. Epithelial-cell recognition of commensal bacteria and maintenance of immune homeostasis in the gut. Nat Rev Immunol. 8:411-20.
Audie JP, Janin A, Porchet N, Copin MC, Gosselin B, Aubert JP. 1993. Expression of human mucin genes in respiratory, digestive, and reproductive tracts ascertained by in situ hybridization. J Histochem Cytochem. 41:1479-85.
Baggiolini M, Dewald B, Moser B. 1994. Interleukin-8 and related chemotactic cytokines - CXC and CC chemokines. Adv Immunol. 55:97-179.
Bas A, Forsberg G, Sjöberg V, Hammarström S, Hernell O, Hammarström ML. 2009. Aberrant extrathymic T cell receptor gene rearrangement in the small intestinal mucosa: a risk factor for celiac disease? Gut. 58:189-95.
Bas A, Hammarström SG, Hammarström ML. 2003. Extrathymic TCR gene rearrangement in human small intestine: identification of new splice forms of recombination activating gene-1 mRNA with selective tissue expression. J Immunol. 171:3359-71.
Bazzoni F, Beutler B. 1996. The tumor necrosis factor ligand and receptor families. N Engl J Med. 334:1717-25.
Beagley KW, Elson CO. 1992. Cells and cytokines in mucosal immunity and inflammation. Gastroenterol Clin North Am. 21:347-66.
Beauchemin N, Draber P, Dveksler G, Gold P, Gray-Owen S, Grunert F, Hammarström S, Holmes KV, Karlsson A, Kuroki M, Lin SH, Lucka L, Najjar SM, Neumaier M, Obrink B, Shively JE, Skubitz KM, Stanners CP, Thomas P, Thompson JA, Virji M, von Kleist S, Wagener C, Watt S, Zimmermann W. 1999. Redefined nomenclature for members of the carcinoembryonic antigen family. Exp Cell Res. 252:243-9.
Beaugerie L, Petit JC. 2004. Microbial-gut interactions in health and disease. Antibiotic-associated diarrhoea. Best Pract Res Clin Gastroenterol. 18:337-52.
Benahmed M, Meresse B, Arnulf B, Barbe U, Mention JJ, Verkarre V, Allez M, Cellier C, Hermine O, Cerf-Bensussan N. 2007. Inhibition of TGF-beta signaling by IL-15: a new role for IL-15 in the loss of immune homeostasis in celiac disease. Gastroenterology. 132:994-1008.
Ben-Baruch A, Michiel DF, Oppenheim JJ. 1995. Signals and receptors involved in recruitment of inflammatory cells. J Biol Chem. 270:11703-6.
Benchimol S, Fuks A, Jothy S, Beauchemin N, Shirota K, Stanners CP. 1989. Carcinoembryonic antigen, a human tumor marker, functions as an intercellular adhesion molecule. Cell. 57:327-34.
Beutler B, Cerami A. 1986. Cachectin and tumour necrosis factor as two sides of the same biological coin. Nature. 320:584-8.
Biagi F, Campanella J, Laforenza U, Gastaldi G, Tritto S, Grazioli M, Villanacci V, Corazza GR. 2006. Transglutaminase 2 in the enterocytes is coeliac specific and gluten dependent. Dig Liver Dis. 38:652-8.
Bik EM, Eckburg PB, Gill SR, Nelson KE, Purdom EA, Francois F, Perez-Perez G, Blaser MJ, Relman DA. 2006. Molecular analysis of the bacterial microbiota in the human stomach. Proc Natl Acad Sci U S A. 103:732-7.
Black RA, Rauch CT, Kozlosky CJ, Peschon JJ, Slack JL, Wolfson MF, Castner BJ, Stocking KL, Reddy P, Srinivasan S, Nelson N, Boiani N, Schooley KA, Gerhart M, Davis R, Fitzner JN, Johnson RS, Paxton RJ, March CJ, Cerretti DP. 1997. A metalloproteinase disintegrin that releases tumour-necrosis factor-alpha from cells. Nature. 385:729-33.

- 53 -
Boehm U, Klamp T, Groot M, Howard JC. 1997. Cellular responses to interferon-gamma. Annu Rev Immunol. 15:749-95.
Booth BA, Boesman-Finkelstein M, Finkelstein RA. 1984. Vibrio cholerae hemagglutinin/protease nicks cholera enterotoxin. Infect Immun. 45:558-60.
Bowie AG, Fitzgerald KA. 2007. RIG-I: tri-ing to discriminate between self and non-self RNA. Trends Immunol. 28:147-50.
Bradley JR. 2008. TNF-mediated inflammatory disease. J Pathol. 214:149-60. Brandtzaeg P, Bjerke K. 1990. Immunomorphological characteristics of human Peyer's
patches. Digestion. 46:S262-73. Brandtzaeg P, Farstad IN, Johansen FE, Morton HC, Norderhaug IN, Yamanaka T. 1999.
The B-cell system of human mucosae and exocrine glands. Immunol Rev. 171:45-52. Breitbart M, Haynes M, Kelley S, Angly F, Edwards RA, Felts B, Mahaffy JM, Mueller J,
Nulton J, Rayhawk S, Rodriguez-Brito B, Salamon P, Rohwer F. 2008. Viral diversity and dynamics in an infant gut. Res Microbiol. 159:367-73.
Breitbart M, Hewson I, Felts B, Mahaffy JM, Nulton J, Salamon P, Rohwer F. 2003. Metagenomic analyses of an uncultured viral community from human feces. J Bacteriol. 185:6220-3.
Brikos C, O'Neill LA. 2008. Signalling of toll-like receptors. Handb Exp Pharmacol. 183:21-50.
Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A, Semenkovich CF, Gordon JI. 2004. The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci U S A. 101:15718-23.
Bäckhed F, Manchester JK, Semenkovich CF, Gordon JI. 2007. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc Natl Acad Sci U S A. 104:979-84.
Calder PC, Krauss-Etschmann S, de Jong EC, Dupont C, Frick JS, Frokiaer H, Heinrich J, Garn H, Koletzko S, Lack G, Mattelio G, Renz H, Sangild PT, Schrezenmeir J, Stulnig TM, Thymann T, Wold AE, Koletzko B. 2006. Early nutrition and immunity - progress and perspectives. Br J Nutr. 96:774-90.
Cario E, Podolsky DK. 2006. Toll-like receptor signaling and its relevance to intestinal inflammation. Ann N Y Acad Sci. 1072:332-8.
Carlsson AK, Axelsson IE, Borulf SK, Bredberg AC, Ivarsson SA. 2001. Serological screening for celiac disease in healthy 2.5-year-old children in Sweden. Pediatrics. 107:42-5.
Carswell EA, Old LJ, Kassel RL, Green S, Fiore N, Williamson B. 1975. An endotoxin-induced serum factor that causes necrosis of tumors. Proc Natl Acad Sci USA. 72: 3666-70.
Chamaillard M, Hashimoto M, Horie Y, Masumoto J, Qiu S, Saab L, Ogura Y, Kawasaki A, Fukase K, Kusumoto S, Valvano MA, Foster SJ, Mak TW, Nuñez G, Inohara N. 2003. An essential role for NOD1 in host recognition of bacterial peptidoglycan containing diaminopimelic acid. Nat Immunol. 4:702-7.
Chen CJ, Kirshner J, Sherman MA, Hu W, Nguyen T, Shively JE. 2007. Mutation analysis of the short cytoplasmic domain of the cell-cell adhesion molecule CEACAM1 identifies residues that orchestrate actin binding and lumen formation. J Biol Chem. 282:5749-60.
Chen G, Goeddel DV. 2002. TNF-R1 signaling: a beautiful pathway. Science. 296: 1634-5. Chinnapen DJ, Chinnapen H, Saslowsky D, Lencer WI. 2007. Rafting with cholera toxin:
endocytosis and trafficking from plasma membrane to ER. FEMS Microbiol Lett. 266:129-37.

- 54 -
Clark JA, Coopersmith CM. 2007. Intestinal crosstalk: a new paradigm for understanding the gut as the "motor" of critical illness. Shock. 28:384-93.
Coelho A, Andrade JR, Vicente AC, Dirita VJ. 2000. Cytotoxic cell vacuolating activity from Vibrio cholerae hemolysin. Infect Immun. 68:1700-5.
Collado MC, Calabuig M, Sanz Y. 2007. Differences between the fecal microbiota of coeliac infants and healthy controls. Curr Issues Intest Microbiol. 8:9-14.
Coombes JL, Powrie F. 2008. Dendritic cells in intestinal immune regulation. Nat Rev Immunol. 8:435-46.
Corfield AP, Carroll D, Myerscough N, Probert CS. 2001. Mucins in the gastrointestinal tract in health and disease. Front Biosci. 6:D1321-57.
Cornes JS. 1965. Peyer's patches in the human gut. Proc R Soc Med. 58:716. Cottingham KL, Chiavelli DA, Taylor RK. 2003. Environmental microbe and human
pathogen: the ecology and microbiology of Vibrio cholerae. Front Ecol Environ. 1:80-6.
Cunliffe RN. 2003. Alpha-defensins in the gastrointestinal tract. Mol Immunol. 40:463-7. Cuthbert AP, Fisher SA, Mirza MM, King K, Hampe J, Croucher PJ, Mascheretti S,
Sanderson J, Forbes A, Mansfield J, Schreiber S, Lewis CM, Mathew CG. 2002. The contribution of NOD2 gene mutations to the risk and site of disease in inflammatory bowel disease. Gastroenterology. 122:867-74.
Daig R, Andus T, Aschenbrenner E, Falk W, Schölmerich J, Gross V. 1996. Increased interleukin-8 expression in the colon mucosa of patients with inflammatory bowel disease. Gut. 38:216-22.
Daig R, Rogler G, Aschenbrenner E, Vogl D, Falk W, Gross V, Schölmerich J, Andus T. 2000. Human intestinal epithelial cells secrete interleukin-1 receptor antagonist and interleukin-8 but not interleukin-1 or interleukin-6. Gut. 46:350-8.
Dann SM, Eckmann L. 2007. Innate immune defenses in the intestinal tract. Curr Opin Gastroenterol. 23:115-20.
Dann SM, Spehlmann ME, Hammond DC, Iimura M, Hase K, Choi LJ, Hanson E, Eckmann L. 2008. IL-6-dependent mucosal protection prevents establishment of a microbial niche for attaching/effacing lesion-forming enteric bacterial pathogens. J Immunol. 180:6816-26.
Darnell JE Jr, Kerr IM, Stark GR. 1994. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science. 264:1415-21.
de Bolos C, Real FX, Lopez-Ferrer A. 2001. Regulation of mucin and glycoconjugate expression: from normal epithelium to gastric tumors. Front Biosci. 6:D1256-63.
De Hertogh G, Aerssens J, de Hoogt R, Peeters P, Verhasselt P, Van Eyken P, Ectors N, Vermeire S, Rutgeerts P, Coulie B, Geboes K. 2006. Validation of 16S rDNA sequencing in microdissected bowel biopsies from Crohn's disease patients to assess bacterial flora diversity. J Pathol. 209:532-9.
de la Cochetiere MF, Piloquet H, des Robert C, Darmaun D, Galmiche JP, Roze JC. 2004. Early intestinal bacterial colonization and necrotizing enterocolitis in premature infants: the putative role of Clostridium. Pediatr Res. 56:366-70.
de Ritis G, Auricchio S, Jones HW, Lew EJ, Bernardin JE, Kasarda DD. 1988. In vitro (organ culture) studies of the toxicity of specific A-gliadin peptides in celiac disease. Gastroenterology. 94:41-9.
Derouet D, Rousseau F, Alfonsi F, Froger J, Hermann J, Barbier F, Perret D, Diveu C, Guillet C, Preisser L, Dumont A, Barbado M, Morel A, deLapeyriere O, Gascan H, Chevalier S. 2004. Neuropoietin, a new IL-6-related cytokine signaling through the ciliary neurotrophic factor receptor Proc. Natl Acad Sci USA. 101:4827-32

- 55 -
Diebold SS, Kaisho T, Hemmi H, Akira S, Reis e Sousa C. 2004. Innate antiviral responses by means of TLR7-mediated recognition of single-stranded RNA. Science. 303:1529-31.
Dillon SR, Sprecher C, Hammond A, Bilsborough J, Rosenfeld-Franklin M, Presnell SR, Haugen HS, Maurer M, Harder B, Johnston J, Bort S, Mudri S, Kuijper JL, Bukowski T, Shea P, Dong DL, Dasovich M, Grant FJ, Lockwood L, Levin SD, LeCiel C, Waggie K, Day H, Topouzis S, Kramer J, Kuestner R, Chen Z, Foster D, Parrish-Novak J, Gross JA. 2004. Interleukin 31, a cytokine produced by activated T cells, induces dermatitis in mice. Nat Immunol. 5:752-60.
Eckburg PB, Bik EM, Bernstein CN, Purdom E, Dethlefsen L, Sargent M, Gill SR, Nelson KE, Relman DA. 2005. Diversity of the human intestinal microbial flora. Science. 308:1635-8.
Eckmann L, Kagnoff MF. 2005. Intestinal mucosal responses to microbial infection. Springer Semin Immunopathol. 27:181-96.
Edlund M, Blikstad I, Obrink B. 1996. Calmodulin binds to specific sequences in the cytoplasmic domain of C-CAM and down-regulates C-CAM self-association. J Biol Chem. 271:1393-9.
Eisenberg SP, Brewer MT, Verderber E, Heimdal P, Brandhuber BJ, Thompson RC. 1991. Interleukin 1 receptor antagonist is a member of the interleukin 1 gene family: evolution of a cytokine control mechanism. Proc Natl Acad Sci U S A. 88:5232-6.
Fahlgren A, Baranov V, Frängsmyr L, Zoubir F, Hammarström ML, Hammarström S. 2003. Interferon-gamma tempers the expression of carcinoembryonic antigen family molecules in human colon cells: a possible role in innate mucosal defence. Scand J Immunol. 58:628-41.
Fahlgren A, Hammarström S, Danielsson A, Hammarström ML. 2003. Increased expression of antimicrobial peptides and lysozyme in colonic epithelial cells of patients with ulcerative colitis. Clin Exp Immunol. 131:90-101.
Fahlgren A, Hammarstrom S, Danielsson A, Hammarstrom ML. 2004. beta-Defensin-3 and -4 in intestinal epithelial cells display increased mRNA expression in ulcerative colitis. Clin Exp Immunol.137:379-85.
Farrell RJ, Kelly CP. 2002. Celiac sprue. N Engl J Med. 346:180-8 Faruque SM, Chowdhury N, Kamruzzaman M, Dziejman M, Rahman MH, Sack DA, Nair
GB, Mekalanos JJ. 2004. Genetic diversity and virulence potential of environmental Vibrio cholerae population in a cholera-endemic area. Proc Natl Acad Sci U S A. 101:2123-8.
Faruque SM, Naser IB, Islam MJ, Faruque AS, Ghosh AN, Nair GB, Sack DA, Mekalanos JJ. 2005. Seasonal epidemics of cholera inversely correlate with the prevalence of environmental cholera phages. Proc Natl Acad Sci U S A. 102:1702-7.
Fasano A. 2005. Clinical presentation of celiac disease in the pediatric population. Gastroenterology. 128:S68-73.
Figueroa-Arredondo P, Heuser JE, Akopyants NS, Morisaki JH, Giono-Cerezo S, Enríquez-Rincón F, Berg DE. 2001. Cell vacuolation caused by Vibrio cholerae hemolysin. Infect Immun. 69:1613-24.
Forsberg G, Fahlgren A, Hörstedt P, Hammarström S, Hernell O, Hammarström ML. 2004. Presence of bacteria and innate immunity of intestinal epithelium in childhood celiac disease. Am J Gastroenterol. 99:894-904.
Forsberg G, Hernell O, Hammarström S, Hammarström ML. 2007. Concomitant increase of IL-10 and pro-inflammatory cytokines in intraepithelial lymphocyte subsets in celiac disease. Int Immunol. 19:993-1001.

- 56 -
Forsberg G, Hernell O, Melgar S, Israelsson A, Hammarström S, Hammarström ML. 2002. Paradoxical coexpression of proinflammatory and down-regulatory cytokines in intestinal T cells in childhood celiac disease. Gastroenterology. 123:667-78.
Franchi L, Park JH, Shaw MH, Marina-Garcia N, Chen G, Kim YG, Núñez G. 2008. Intracellular NOD-like receptors in innate immunity, infection and disease. Cell Microbiol. 10:1-8.
Froy O. 2005. Regulation of mammalian defensin expression by Toll-like receptor-dependent and independent signalling pathways. Cell Microbiol. 7:1387-97.
Frängsmyr L, Baranov V, Hammarström S. 1999. Four carcinoembryonic antigen subfamily members, CEA, NCA, BGP and CGM2, selectively expressed in the normal human colonic epithelium, are integral components of the fuzzy coat. Tumour Biol. 20:277-92.
Frängsmyr L, Baranov V, Prall F, Yeung MM, Wagener C, Hammarström S. 1995. Cell- and region-specific expression of biliary glycoprotein and its messenger RNA in normal human colonic mucosa. Cancer Res. 55:2963-7.
Fukushima K, Ishiyama C, Yamashita K. 2004. Recognition by TNF-alpha of the GPI-anchor glycan induces apoptosis of U937 cells. Arch Biochem Biophys. 426:298-305.
Fullner KJ, Boucher JC, Hanes MA, Haines GK 3rd, Meehan BM, Walchle C, Sansonetti PJ, Mekalanos JJ. 2002. The contribution of accessory toxins of Vibrio cholerae O1 El Tor to the proinflammatory response in a murine pulmonary cholera model. J Exp Med. 195:1455-62.
Ganz T. 1999. Defensins and host defense. Science. 286:420-1. Ganz T. 2002. Epithelia: not just physical barriers. Proc Natl Acad Sci U S A. 99:3357-8. Ganz T. 2003. Defensins: antimicrobial peptides of innate immunity. Nat. Rev. Immunol.
3:710-20. Ganz T, Selsted ME, Szklarek D, Harwig SS, Daher K, Bainton DF, Lehrer RI. 1985.
Defensins. Natural peptide antibiotics of human neutrophils. J Clin Invest. 76:1427-35. Gao Z, Tseng CH, Pei Z, Blaser MJ. 2007. Molecular analysis of human forearm superficial
skin bacterial biota. Proc Natl Acad Sci U S A. 104:2927-32. Garrote JA, Gómez-González E, Bernardo D, Arranz E, Chirdo F. 2008. Celiac disease
pathogenesis: the proinflammatory cytokine network. J Pediatr Gastroenterol Nutr. 47:S27-32.
Gasbarrini G, Malandrino N, Giorgio V, Fundarò C, Cammarota G, Merra G, Roccarina D, Gasbarrini A, Capristo E. 2008. Celiac disease: what's new about it? Dig Dis. 26:121-7.
Gaur S, Shively JE, Yen Y, Gaur RK. 2008. Altered splicing of CEACAM1 in breast cancer: identification of regulatory sequences that control splicing of CEACAM1 into long or short cytoplasmic domain isoforms. Mol Cancer. 7:46.
Gijzen K, Cambi A, Torensma R, Figdor CG. 2006. C-type lectins on dendritic cells and their interaction with pathogen-derived and endogenous glycoconjugates. Curr Protein Pept Sci. 7:283-94.
Gill SR, Pop M, Deboy RT, Eckburg PB, Turnbaugh PJ, Samuel BS, Gordon JI, Relman DA, Fraser-Liggett CM, Nelson KE. 2006. Metagenomic analysis of the human distal gut microbiome. Science. 312:1355-9.
Girardin SE, Boneca IG, Carneiro LA, Antignac A, Jéhanno M, Viala J, Tedin K, Taha MK, Labigne A, Zähringer U, Coyle AJ, DiStefano PS, Bertin J, Sansonetti PJ, Philpott DJ. 2003. Nod1 detects a unique muropeptide from gram-negative bacterial peptidoglycan. Science. 300:1584-7.
Godkin A, Friede T, Davenport M, Stevanovic S, Willis A, Jewell D, Hill A, Rammensee HG. 1997. Use of eluted peptide sequence data to identify the binding characteristics of

- 57 -
peptides to the insulin-dependent diabetes susceptibility allele HLA-DQ8 (DQ 3.2). Int Immunol. 9:905-11.
Gold P, Freedman SO. 1965. Specific carcinoembryonic antigens of the human digestive system. J Exp Med. 122:467-81.
Gray-Owen SD, Blumberg RS. 2006. CEACAM1: contact-dependent control of immunity. Nat Rev Immunol. 6:433-46.
Greco L, Romino R, Coto I, Di Cosmo N, Percopo S, Maglio M, Paparo F, Gasperi V, Limongelli MG, Cotichini R, D'Agate C, Tinto N, Sacchetti L, Tosi R, Stazi MA. 2002. The first large population based twin study of coeliac disease. Gut. 50:624-8.
Green PH, Cellier C. 2007. Celiac disease. N Engl J Med. 357:1731-43. Green PH, Jabri B. 2006. Celiac disease. Annu Rev Med. 57:207-21. Greicius G, Severinson E, Beauchemin N, Obrink B, Singer BB. 2003. EACAM1 is a potent
regulator of B cell receptor complex-induced activation. J Leukoc Biol. 74:126-34. Griffiths NJ, Bradley CJ, Heyderman RS, Virji M. 2007. IFN-gamma amplifies NFkappaB-
dependent Neisseria meningitidis invasion of epithelial cells via specific upregulation of CEA-related cell adhesion molecule 1. Cell Microbiol. 9:2968-83.
Groh V, Steinle A, Bauer S, Spies T. 1998. Recognition of stress-induced MHC molecules by intestinal epithelial gammadelta T cells. Science. 279:1737-40.
Gross O, Gewies A, Finger K, Schäfer M, Sparwasser T, Peschel C, Förster I, Ruland J. 2006. Card9 controls a non-TLR signalling pathway for innate anti-fungal immunity. Nature. 442:651-6.
Guarner F. 2006. Enteric flora in health and disease. Digestion. 73 S1:5-12. Guarner F, Malagelada JR. 2003. Gut flora in health and disease. Lancet. 361:512-9. Gudmundsson GH, Agerberth B, Odeberg J, Bergman T, Olsson B, Salcedo R. 1996. The
human gene FALL39 and processing of the cathelin precursor to the antibacterial peptide LL-37 in granulocytes. Eur J Biochem. 238:325-32.
Gum JR Jr, Crawley SC, Hicks JW, Szymkowski DE, Kim YS. 2002. MUC17, a novel membrane-tethered mucin. Biochem Biophys Res Commun. 291:466-75.
Gum JR Jr, Hicks JW, Toribara NW, Siddiki B, Kim YS. 1994. Molecular cloning of human intestinal mucin (MUC2) cDNA. Identification of the amino terminus and overall sequence similarity to prepro-von Willebrand factor. J Biol Chem. 269:2440-6.
Gutierrez MG, Saka HA, Chinen I, Zoppino FC, Yoshimori T, Bocco JL, Colombo MI. 2007. Protective role of autophagy against Vibrio cholerae cytolysin, a pore-forming toxin from V. cholerae. Proc Natl Acad Sci U S A. 104:1829-34.
Halstensen TS, Brandtzaeg P. 1993. Activated T lymphocytes in the celiac lesion: non-proliferative activation (CD25) of CD4+ alpha/beta cells in the lamina propria but proliferation (Ki-67) of alpha/beta and gamma/delta cells in the epithelium. Eur J Immunol. 23:505-10.
Hammarström S. 1999. The carcinoembryonic antigen (CEA) family: structures, suggested functions and expression in normal and malignant tissues. Semin Cancer Biol. 9: 67-81.
Hammarström S, Baranov V. 2001. Is there a role for CEA in innate immunity in the colon? Trends Microbiol. 9:119-25.
Harris JR, Bhakdi S, Meissner U, Scheffler D, Bittman R, Li G, Zitzer A, Palmer M. 2002. Interaction of the Vibrio cholerae cytolysin (VCC) with cholesterol, some cholesterol esters, and cholesterol derivatives: a TEM study. J Struct Biol. 139:122-35.
Hase K, Eckmann L, Leopard JD, Varki N, Kagnoff MF. 2002. Cell differentiation is a key determinant of cathelicidin LL-37/human cationic antimicrobial protein 18 expression by human colon epithelium. Infect Immun. 70:953-63.

- 58 -
Hase K, Murakami M, Iimura M, Cole SP, Horibe Y, Ohtake T, Obonyo M, Gallo RL, Eckmann L, Kagnoff MF. 2003. Expression of LL-37 by human gastric epithelial cells as a potential host defense mechanism against Helicobacter pylori. Gastroenterology. 125:1613-25.
Hauck CR, Agerer F, Muenzner P, Schmitter T. 2006. Cellular adhesion molecules as targets for bacterial infection. Eur J Cell Biol. 85:235-42.
Hausmann M, Kiessling S, Mestermann S, Webb G, Spöttl T, Andus T, Schölmerich J, Herfarth H, Ray K, Falk W, Rogler G. 2002. Toll-like receptors 2 and 4 are up-regulated during intestinal inflammation. Gastroenterology. 122:1987-2000.
Hayashi F, Smith KD, Ozinsky A, Hawn TR, Yi EC, Goodlett DR, Eng JK, Akira S, Underhill DM, Aderem A. 2001. The innate immune response to bacterial flagellin is mediated by Toll-like receptor 5. Nature. 410:1099-103.
Heidelberg JF, Eisen JA, Nelson WC, Clayton RA, Gwinn ML, Dodson RJ, Haft DH, Hickey EK, Peterson JD, Umayam L, Gill SR, Nelson KE, Read TD, Tettelin H, Richardson D, Ermolaeva MD, Vamathevan J, Bass S, Qin H, Dragoi I, Sellers P, McDonald L, Utterback T, Fleishmann RD, Nierman WC, White O, Salzberg SL, Smith HO, Colwell RR, Mekalanos JJ, Venter JC, Fraser CM. 2000. DNA sequence of both chromosomes of the cholera pathogen Vibrio cholerae. Nature. 406:477-83.
Heil F, Hemmi H, Hochrein H, Ampenberger F, Kirschning C, Akira S, Lipford G, Wagner H, Bauer S. 2004. Species-specific recognition of single-stranded RNA via toll-like receptor 7 and 8. Science. 303:1526-9.
Heinrich PC, Behrmann I, Haan S, Hermanns HM, Müller-Newen G, Schaper F. 2003. Principles of interleukin (IL)-6-type cytokine signalling and its regulation. Biochem J. 374:1-20.
Heinrich PC, Castell JV, Andus T. 1990. Interleukin-6 and the acute phase response. Biochem J. 265:621-36.
Hemmi H, Kaisho T, Takeuchi O, Sato S, Sanjo H, Hoshino K, Horiuchi T, Tomizawa H, Takeda K, Akira S. 2002. Small anti-viral compounds activate immune cells via the TLR7 MyD88-dependent signaling pathway. Nat Immunol. 3:196-200.
Hemmila E, Turbide C, Olson M, Jothy S, Holmes KV, Beauchemin N. 2004. Ceacam1a-/- mice are completely resistant to infection by murine coronavirus mouse hepatitis virus A59. J Virol. 78:10156-65.
Hershberg RM, Mayer LF. 2000. Antigen processing and presentation by intestinal epithelial cells - polarity and complexity. Immunol Today. 21:123-8.
Heyderman RS, Virji M. 2007. IFN-gamma amplifies NFkappaB-dependent Neisseria meningitidis invasion of epithelial cells via specific upregulation of CEA-related cell adhesion molecule 1. Cell Microbiol. 9:2968-83.
Higuchi T, Orita T, Katsuya K, Yamasaki Y, Akiyama K, Li H, Yamamoto T, Saito Y, Nakamura M. 2004. MUC20 suppresses the hepatocyte growth factor-induced Grb2-Ras pathway by binding to a multifunctional docking site of met. Mol Cell Biol. 24:7456-68.
Hirano T. 1998. Interleukin 6 and its receptor: ten years later. Int Rev Immunol. 16:249-84. Hirano T, Yasukawa K, Harada H, Taga T, Watanabe Y, Matsuda T, Kashiwamura S,
Nakajima K, Koyama K, Iwamatsu A, Tsunasawa S, Sakiyama F, Matsui H, Takahara Y, Taniguchi T, Kishimoto T. 1986. Complementary DNA for a novel human interleukin (BSF-2) that induces B lymphocytes to produce immunoglobulin. Nature. 324:73-6.
Hlady WG, Klontz KC. 1996. The epidemiology of Vibrio infections in Florida, 1981-1993. J Infect Dis. 173:1176-83.

- 59 -
Holmes WE, Lee J, Kuang WJ, Rice GC, Wood WI. 1991. Structure and functional expression of a human interleukin-8 receptor. Science. 253:1278-80.
Hooper LV, Gordon JI. 2001. Commensal host-bacterial relationships in the gut. Science. 292:1115-8.
Hooper LV, Wong MH, Thelin A, Hansson L, Falk PG, Gordon JI. 2001. Molecular analysis of commensal host-microbial relationships in the intestine. Science. 291:881-4.
Houlston RS, Ford D. 1996. Genetics of coeliac disease. QJM. 89:737-43. Hourigan CS. 2006. The molecular basis of coeliac disease. Clin Exp Med. 6:53-9. Hugot JP, Chamaillard M, Zouali H, Lesage S, Cezard JP, Belaiche J, Almer S, Tysk C,
O'Morain CA, Gassull M, Binder V, Finkel Y, Cortot A, Modigliani R, Laurent-Puig P, Gower-Rousseau C, Macry J, Colombel JF, Sahbatou M, Thomas G. 2001. Association of NOD2 leucine-rich repeat variants with susceptibility to Crohn's disease. Nature. 411:599-603.
Hunt KA, Zhernakova A, Turner G, Heap GA, Franke L, Bruinenberg M, Romanos J, Dinesen LC, Ryan AW, Panesar D, Gwilliam R, Takeuchi F, McLaren WM, Holmes GK, Howdle PD, Walters JR, Sanders DS, Playford RJ, Trynka G, Mulder CJ, Mearin ML, Verbeek WH, Trimble V, Stevens FM, O'Morain C, Kennedy NP, Kelleher D, Pennington DJ, Strachan DP, McArdle WL, Mein CA, Wapenaar MC, Deloukas P, McGinnis R, McManus R, Wijmenga C, van Heel DA. 2008. Newly identified genetic risk variants for celiac disease related to the immune response. Nat Genet. 40:395-402.
Ichinose Y, Yamamoto K, Nakasone N, Tanabe MJ, Takeda T, Miwatani T, Iwanaga M. 1987. Enterotoxicity of El Tor-like hemolysin of non-O1 Vibrio cholerae. Infect Immun. 55:1090-3.
Idriss HT, Naismith JH. 2000. TNF alpha and the TNF receptor superfamily: structure-function relationship(s). Microsc Res Tech. 50:184-95.
Isaacs A, Lindenmann J. 1957. Virus interference I. The interferon. Proc R Soc Lond B: Biol Sci. 147:258-67.
Ishii KJ, Akira S. 2006. Innate immune recognition of, and regulation by, DNA. Trends Immunol. 27:525-32.
Ivarsson A, Hernell O, Nyström L, Persson LA. 2003. Children born in the summer have increased risk for coeliac disease. J Epidemiol Community Health. 57:36-9.
Ivarsson A, Hernell O, Stenlund H, Persson LA. 2002. Breast-feeding protects against celiac disease. Am J Clin Nutr. 75:914-21.
Ivarsson A, Persson LA, Nyström L, Ascher H, Cavell B, Danielsson L, Dannaeus A, Lindberg T, Lindquist B, Stenhammar L, Hernell O. 2000. Epidemic of coeliac disease in Swedish children. Acta Paediatr. 89:165-71.
Jack RS, Fan X, Bernheiden M, Rune G, Ehlers M, Weber A, Kirsch G, Mentel R, Fürll B, Freudenberg M, Schmitz G, Stelter F, Schütt C. 1997. Lipopolysaccharide-binding protein is required to combat a murine gram-negative bacterial infection. Nature. 389:742-5.
Jass JR, Roberton AM. 1994. Colorectal mucin histochemistry in health and disease: a critical review. Pathol Int. 44:487-504.
Jiang Z, Georgel P, Du X, Shamel L, Sovath S, Mudd S, Huber M, Kalis C, Keck S, Galanos C, Freudenberg M, Beutler B. 2005. CD14 is required for MyD88-independent LPS signaling. Nat Immunol. 6:565-70.
Jin MS, Kim SE, Heo JY, Lee ME, Kim HM, Paik SG, Lee H, Lee JO. 2007. Crystal structure of the TLR1-TLR2 heterodimer induced by binding of a tri-acylated lipopeptide. Cell. 130:1071-82.

- 60 -
Johansen BH, Gjertsen HA, Vartdal F, Buus S, Thorsby E, Lundin KE, Sollid LM. 1996. Binding of peptides from the N-terminal region of alpha-gliadin to the celiac disease-associated HLA-DQ2 molecule assessed in biochemical and T cell assays. Clin Immunol Immunopathol. 79:288-93.
Jones DE, Bevins CL. 1992. Paneth cells of the human small intestine express an antimicrobial peptide gene. J Biol Chem. 267:23216-25.
Jones DE, Bevins CL. 1993. Defensin-6 mRNA in human Paneth cells: implications for antimicrobial peptides in host defense of the human bowel. FEBS Lett. 315:187-92.
Jung HC, Eckmann L, Yang SK, Panja A, Fierer J, Morzycka-Wroblewska E, Kagnoff MF. 1995. A distinct array of proinflammatory cytokines is expressed in human colon epithelial cells in response to bacterial invasion. J Clin Invest. 95:55-65.
Jurk M, Heil F, Vollmer J, Schetter C, Krieg AM, Wagner H, Lipford G, Bauer S. 2002. Human TLR7 or TLR8 independently confer responsiveness to the antiviral compound R-848. Nat Immunol. 3:499.
Kagnoff MF. 2007. Celiac disease: pathogenesis of a model immunogenetic disease. J Clin Invest. 117:41-9.
Kai-Larsen Y, Agerberth B. 2008. The role of the multifunctional peptide LL-37 in host defense. Front Biosci. 13:3760-7.
Kaiserlian D, Vidal K, Revillard JP. 1989. Murine enterocytes can present soluble antigen to specific class II-restricted CD4+ T cells. Eur J Immunol. 19:1513-6.
Kalliomäki M, Collado MC, Salminen S, Isolauri E. 2008. Early differences in fecal microbiota composition in children may predict overweight. Am J Clin Nutr. 87:534-8.
Kalliomäki M, Kirjavainen P, Eerola E, Kero P, Salminen S, Isolauri E. 2001. Distinct patterns of neonatal gut microflora in infants in whom atopy was and was not developing. J Allergy Clin Immunol. 107:129-34.
Kamimura D, Ishihara K, Hirano T. 2003. IL-6 signal transduction and its physiological roles: the signal orchestration model. Rev Physiol Biochem Pharmacol. 149:1-38.
Kanneganti TD, Lamkanfi M, Núñez G. 2007. Intracellular NOD-like receptors in host defense and disease. Immunity. 27:549-59.
Kanneganti TD, Ozören N, Body-Malapel M, Amer A, Park JH, Franchi L, Whitfield J, Barchet W, Colonna M, Vandenabeele P, Bertin J, Coyle A, Grant EP, Akira S, Núñez G. 2006. Bacterial RNA and small antiviral compounds activate caspase-1 through cryopyrin/Nalp3. Nature. 440:233-6.
Kaper JB, Morris JG Jr, Levine MM. 1995. Cholera. Clin Microbiol Rev. 8:48-86. Keller M, Rüegg A, Werner S, Beer HD. 2008. Active caspase-1 is a regulator of
unconventional protein secretion. Cell. 132:818-31. Keshav S. 2006. Paneth cells: leukocyte-like mediators of innate immunity in the intestine. J
Leukoc Biol. 80:500-8. Kim HM, Park BS, Kim JI, Kim SE, Lee J, Oh SC, Enkhbayar P, Matsushima N, Lee H,
Yoo OJ, Lee JO. 2007. Crystal structure of the TLR4-MD-2 complex with bound endotoxin antagonist Eritoran. Cell. 130:906-17.
Kirshner J, Hardy J, Wilczynski S, Shively JE. 2004. Cell-cell adhesion molecule CEACAM1 is expressed in normal breast and milk and associates with beta1 integrin in a 3D model of morphogenesis. J Mol Histol. 35:287-99.
Kishimoto T. 2005. Interleukin-6: from basic science to medicine: 40 years in immunology. Annu Rev Immunol. 23:1-21.
Kishimoto T, Akira S, Narazaki M, Taga T. 1995. Interleukin-6 family of cytokines and gp130. Blood. 86:1243-54.

- 61 -
Kobayashi KS, Chamaillard M, Ogura Y, Henegariu O, Inohara N, Nuñez G, Flavell RA. 2005. Nod2-dependent regulation of innate and adaptive immunity in the intestinal tract. Science. 307:731-4.
Kobayashi Y. 2008. The role of chemokines in neutrophil biology. Front Biosci. 13:2400-7. Kraehenbuhl JP, Neutra MR. 2000. Epithelial M cells: differentiation and function. Annu
Rev Cell Dev Biol. 16:301-32. Kuespert K, Pils S, Hauck CR. 2006. CEACAMs: their role in physiology and
pathophysiology. Curr Opin Cell Biol. 18: 565-71. Kumagai Y, Takeuchi O, Akira S. 2008. Pathogen recognition by innate receptors. J Infect
Chemother. 14:86-92. Kunath T, Ordonez-Garcia C, Turbide C, Beauchemin N. 1995. Inhibition of colonic tumor
cell growth by biliary glycoprotein. Oncogene. 11:2375-82. Kuno K, Matsushima K. 1994. The IL-1 receptor signaling pathway. J Leukoc Biol. 56:542-
7. Kupper C. 2005. Dietary guidelines and implementation for celiac disease.
Gastroenterology. 128:S121-7. Kusugami K, Fukatsu A, Tanimoto M, Shinoda M, Haruta J, Kuroiwa A, Ina K, Kanayama
K, Ando T, Matsuura T, Yamaguchi T, Morise K, Jeda M, Iokawa H, Ishihara A, Sarai S. 1995. Elevation of interleukin-6 in inflammatory bowel disease is macrophage- and epithelial cell-dependent. Dig Dis Sci. 40:949-59.
Lammers KM, Lu R, Brownley J, Lu B, Gerard C, Thomas K, Rallabhandi P, Shea-Donohue T, Tamiz A, Alkan S, Netzel-Arnett S, Antalis T, Vogel SN, Fasano A. 2008. Gliadin induces an increase in intestinal permeability and zonulin release by binding to the chemokine receptor CXCR3. Gastroenterology. 135:194-204.
Latz E, Verma A, Visintin A, Gong M, Sirois CM, Klein DC, Monks BG, McKnight CJ, Lamphier MS, Duprex WP, Espevik T, Golenbock DT. 2007. Ligand induced conformational changes allosterically activate Toll-like receptor 9. Nat Immunol. 8:772-9.
Ledgerwood EC, Pober JS, Bradley JR. 1999. Recent advances in the molecular basis of TNF signal transduction. Lab Invest. 79:1041-50.
Lehrer RI, Lichtenstein AK, Ganz T. 1993. Defensins: antimicrobial and cytotoxic peptides of mammalian cells. Annu Rev Immunol. 11:105-28.
Lencer WI, Saslowsky D. 2005. Raft trafficking of AB5 subunit bacterial toxins. Biochim Biophys Acta. 1746:314-21.
Levine MM, Kaper JB, Herrington D, Losonsky G, Morris JG, Clements ML, Black RE, Tall B, Hall R. 1988. Volunteer studies of deletion mutants of Vibrio cholerae O1 prepared by recombinant techniques. Infect Immun. 56:161-7.
Levy DE, Darnell JE Jr. 2002. Stats: transcriptional control and biological impact. Nat Rev Mol Cell Biol. 3:651-62.
Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI. 2005. Obesity alters gut microbial ecology. Proc Natl Acad Sci U S A. 102:11070-5.
Ley RE, Peterson DA, Gordon JI. 2006. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell. 124:837-48.
Ley RE, Turnbaugh PJ, Klein S, Gordon JI. 2006. Microbial ecology: human gut microbes associated with obesity. Nature. 444:1022-3.
Li M, Wang B, Zhang M, Rantalainen M, Wang S, Zhou H, Zhang Y, Shen J, Pang X, Zhang M, Wei H, Chen Y, Lu H, Zuo J, Su M, Qiu Y, Jia W, Xiao C, Smith LM, Yang S, Holmes E, Tang H, Zhao G, Nicholson JK, Li L, Zhao L. 2008. Symbiotic gut

- 62 -
microbes modulate human metabolic phenotypes. Proc Natl Acad Sci U S A. 105:2117-22.
Linden SK, Sutton P, Karlsson NG, Korolik V, McGuckin MA. 2008. Mucins in the mucosal barrier to infection. Mucosal Immunol. 1:183-97.
Lipp EK, Huq A, Colwell RR. 2002. Effects of global climate on infectious disease: the cholera model. Clin Microbiol Rev. 15:757-70.
Liu L, Wang L, Jia HP, Zhao C, Heng HH, Schutte BC, McCray PB Jr, Ganz T. 1998. Structure and mapping of the human beta-defensin HBD-2 gene and its expression at sites of inflammation. Gene. 222:237-44.
Liévin-Le Moal V, Servin AL. 2006. The front line of enteric host defense against unwelcome intrusion of harmful microorganisms: mucins, antimicrobial peptides, and microbiota. Clin Microbiol Rev. 19:315-37.
Locksley RM, Killeen N, Lenardo MJ. 2001. The TNF and TNF receptor superfamilies: integrating mammalian biology. Cell. 104:487-501.
Losowsky MS. 2008. A history of coeliac disease. Dig Dis. 26:112-20. Lundqvist C, Baranov V, Hammarström S, Athlin L, Hammarström ML. 1995. Intra-
epithelial lymphocytes. Evidence for regional specialization and extrathymic T cell maturation in the human gut epithelium. Int Immunol. 7:1473-87.
MacDonald TT. 2003. The mucosal immune system. Parasite Immunol. 25:235-46. Mackey D, McFall AJ. 2006. MAMPs and MIMPs: proposed classifications for inducers of
innate immunity. Mol Microbiol. 61:1365-71. Mackie RI, Sghir A, Gaskins HR. 1999. Developmental microbial ecology of the neonatal
gastrointestinal tract. Am J Clin Nutr. 69:1035S-45S. Macpherson AJ, Harris NL. 2004. Interactions between commensal intestinal bacteria and
the immune system. Nat Rev Immunol. 4:478-85. Madsen K, Cornish A, Soper P, McKaigney C, Jijon H, Yachimec C, Doyle J, Jewell L, De
Simone C. 2001. Probiotic bacteria enhance murine and human intestinal epithelial barrier function. Gastroenterology. 121:580-91.
Maeurer MJ, Martin D, Walter W, Liu K, Zitvogel L, Halusczcak K, Rabinowich H, Duquesnoy R, Storkus W, Lotze MT. 1996. Human intestinal Vdelta1+ lymphocytes recognize tumor cells of epithelial origin. J Exp Med. 183:1681-96.
Magalhaes JG, Tattoli I, Girardin SE. 2007. The intestinal epithelial barrier: how to distinguish between the microbial flora and pathogens. Semin Immunol. 19:106-15.
Maiuri L, Troncone R, Mayer M, Coletta S, Picarelli A, De Vincenzi M, Pavone V, Auricchio S. 1996. In vitro activities of A-gliadin-related synthetic peptides: damaging effect on the atrophic coeliac mucosa and activation of mucosal immune response in the treated coeliac mucosa. Scand J Gastroenterol. 31:247-53.
Mantzaris G, Jewell DP. 1991. In vivo toxicity of a synthetic dodecapeptide from A gliadin in patients with coeliac disease. Scand J Gastroenterol. 26:392-8.
Mariathasan S, Weiss DS, Newton K, McBride J, O'Rourke K, Roose-Girma M, Lee WP, Weinrauch Y, Monack DM, Dixit VM. 2006. Cryopyrin activates the inflammasome in response to toxins and ATP. Nature. 440:228-32.
Martin MU, Falk W. 1997. The interleukin-1 receptor complex and interleukin-1 signal transduction. Eur Cytokine Netw. 8:5-17.
Martinon F, Burns K, Tschopp J. 2002. The inflammasome: a molecular platform triggering activation of infl ammatory caspases and processing of proIL-beta. Mol Cell. 10:417-26.
Martinon F, Pétrilli V, Mayor A, Tardivel A, Tschopp J. 2006. Gout-associated uric acid crystals activate the NALP3 inflammasome. Nature. 440:237-41.

- 63 -
Mashburn-Warren LM, Whiteley M. 2006. Special delivery: vesicle trafficking in prokaryotes. Mol Microbiol. 61:839-46.
May MJ, Ghosh S. 1998. Signal transduction through NF-kappa B. Immunol Today. 19:80-8.
McCracken VJ, Lorenz RG. 2001. The gastrointestinal ecosystem: a precarious alliance among epithelium, immunity and microbiota. Cell Microbiol. 3:1-11.
McGee DW, Vitkus SJ, Lee P. 1996. The effect of cytokine stimulation on IL-1 receptor mRNA expression by intestinal epithelial cells. Cell Immunol. 168:276-80.
Medzhitov R, Janeway CA Jr. 1997. Innate immunity: the virtues of a nonclonal system of recognition. Cell. 91:295-8.
Meeuwisse GW. 1970. Diagnostic criteria in coeliac disease. Acta Paediatr Scand, 59:461-3. Melgar S, Bas A, Hammarström S, Hammarström ML. 2002. Human small intestinal
mucosa harbours a small population of cytolytically active CD8+ alphabeta T lymphocytes. Immunology. 106:476-85.
Melgar S, Yeung MM, Bas A, Forsberg G, Suhr O, Oberg A, Hammarstrom S, Danielsson A, Hammarstrom ML. 2003. Over-expression of interleukin 10 in mucosal T cells of patients with active ulcerative colitis. Clin Exp Immunol. 134:127-37.
Melmed G, Thomas LS, Lee N, Tesfay SY, Lukasek K, Michelsen KS, Zhou Y, Hu B, Arditi M, Abreu MT. 2003. Human intestinal epithelial cells are broadly unresponsive to Toll-like receptor 2-dependent bacterial ligands: implications for host-microbial interactions in the gut. J Immunol. 170:1406-15.
Middleton J, Neil S, Wintle J, Clark-Lewis I, Moore H, Lam C, Auer M, Hub E, Rot A. 1997. Transcytosis and surface presentation of IL-8 by venular endothelial cells. Cell. 91:385-95.
Miller H, Zhang J, Kuolee R, Patel GB, Chen W. 2007. Intestinal M cells: the fallible sentinels? World J Gastroenterol. 13:1477-86.
Mitra R, Figueroa P, Mukhopadhyay AK, Shimada T, Takeda Y, Berg DE, Nair GB. 2000. Cell vacuolation, a manifestation of the El tor hemolysin of Vibrio cholerae. Infect Immun. 68:1928-33.
Molberg O, Kett K, Scott H, Thorsby E, Sollid LM, Lundin KE.1997. Gliadin specific, HLA DQ2-restricted T cells are commonly found in small intestinal biopsies from coeliac disease patients, but not from controls. Scand J Immunol. 46:103-9.
Moschioni M, Tombola F, de Bernard M, Coelho A, Zitzer A, Zoratti M, Montecucco C. 2002. The Vibrio cholerae haemolysin anion channel is required for cell vacuolation and death. Cell Microbiol. 4:397-409.
Mowat AM. 2003. Anatomical basis of tolerance and immunity to intestinal antigens. Nat Rev Immunol. 3:331-41.
Murphy PM, Tiffany HL. 1991. Cloning of a complimentary DNA encoding a functional human interleukin-8 receptor. Science. 253:1280-3.
Murray PJ. 2007. The JAK-STAT signaling pathway: input and output integration. J Immunol. 178:2623-9.
Nadal I, Donat E, Ribes-Koninckx C, Calabuig M, Sanz Y. 2007. Imbalance in the composition of the duodenal microbiota of children with coeliac disease. J Med Microbiol. 56:1669-74.
Nagai Y, Akashi S, Nagafuku M, Ogata M, Iwakura Y, Akira S, Kitamura T, Kosugi A, Kimoto M, Miyake K. 2002. Essential role of MD-2 in LPS responsiveness and TLR4 distribution. Nat Immunol. 3:667-72.

- 64 -
Nagaishi T, Iijima H, Nakajima A, Chen D, Blumberg RS. 2006. Role of CEACAM1 as a regulator of T cells. Ann N Y Acad Sci. 1072:155-75.
Nagamune K, Yamamoto K, Honda T. 1997. Intramolecular chaperone activity of the pro-region of Vibrio cholerae El Tor cytolysin. J Biol Chem. 272:1338-43.
Nagamune K, Yamamoto K, Naka A, Matsuyama J, Miwatani T, Honda T. 1996. In vitro proteolytic processing and activation of the recombinant precursor of El Tor cytolysin/hemolysin (pro-HlyA) of Vibrio cholerae by soluble hemagglutinin/protease of V. cholerae, trypsin, and other proteases. Infect Immun. 64:4655-8.
Newberry RD, Lorenz RG. 2005. Organizing a mucosal defense. Immunol Rev. 206:6-21. Ninin E, Caroff N, El Kouri D, Espaze E, Richet H, Quilici ML, Fournier JM. 2000.
Nontoxigenic vibrio Cholerae O1 bacteremia: case report and review. Eur J Clin Microbiol Infect Dis. 19:489-91.
Noverr MC, Huffnagle GB. 2004. Does the microbiota regulate immune responses outside the gut? Trends Microbiol. 12:562-8.
Noverr MC, Huffnagle GB. 2005. The 'microflora hypothesis' of allergic diseases. Clin Exp Allergy. 35:1511-20.
Obrink B, Sawa H, Scheffrahn I, Singer BB, Sigmundsson K, Sundberg U, Heymann R, Beauchemin N, Weng G, Ram P, Iyengar R. 2002. Computational analysis of isoform-specific signal regulation by CEACAM1-A cell adhesion molecule expressed in PC12 cells. Ann N Y Acad Sci. 971:597-607.
Ogierman MA, Fallarino A, Riess T, Williams SG, Attridge SR, Manning PA. 1997. Characterization of the Vibrio cholerae El Tor lipase operon lipAB and a protease gene downstream of the hly region. J Bacteriol. 179:7072-80.
Ogura Y, Bonen DK, Inohara N, Nicolae DL, Chen FF, Ramos R, Britton H, Moran T, Karaliuskas R, Duerr RH, Achkar JP, Brant SR, Bayless TM, Kirschner BS, Hanauer SB, Nunez G, Cho JH. 2001. A frameshift mutation in NOD2 associated with susceptibility to Crohn's disease. Nature. 411:603-6.
Oikawa S, Inuzuka C, Kuroki M, Matsuoka Y, Kosaki G, Nakazato H. 1989. Cell adhesion activity of non-specific cross-reacting antigen (NCA) and carcinoembryonic antigen (CEA) expressed on CHO cell surface: homophilic and heterophilic adhesion. Biochem Biophys Res Commun. 164:39-45.
Olivier V, Haines III GK, Tan Y, Fullner Satchell KJ. 2007. Hemolysin and the multifunctional autoprocessing RTX toxin are virulence factors during intestinal infection of mice with Vibrio cholerae El Tor O1 strains. Infect Immun. 75:5035-42.
Olsson C, Hernell O, Hörnell A, Lönnberg G, Ivarsson A. 2008. Difference in celiac disease risk between Swedish birth cohorts suggests an opportunity for primary prevention. Pediatrics. 122:528-34
O'Neil DA, Porter EM, Elewaut D, Anderson GM, Eckmann L, Ganz T, Kagnoff MF. 1999. Expression and regulation of the human beta-defensins hBD-1 and hBD-2 in intestinal epithelium. J Immunol. 163:6718-24.
O'Neill LA, Greene C. 1998. Signal transduction pathways activated by the IL-1 receptor family: ancient signaling machinery in mammals, insects and plants. J Leukoc Biol. 63:650-7.
Osawa R, Okitsu T, Sata S, and Yamai S. 1997. Rapid screening method for identification of cholera toxin-producing Vibrio cholerae O1 and O139. J Clin Microbiol. 35:951-3.
O'Sullivan LA, Liongue C, Lewis RS, Stephenson SE, Ward AC. 2007. Cytokine receptor signaling through the Jak-Stat-Socs pathway in disease. Mol Immunol. 44:2497-506.

- 65 -
Ott SJ, Musfeldt M, Wenderoth DF, Hampe J, Brant O, Fölsch UR, Timmis KN, Schreiber S. 2004. Reduction in diversity of the colonic mucosa associated bacterial microflora in patients with active inflammatory bowel disease. Gut. 53:685-93.
Otte JM, Cario E, Podolsky DK. 2004. Mechanisms of cross hyporesponsiveness to Toll-like receptor bacterial ligands in intestinal epithelial cells. Gastroenterology. 126:1054-70.
Otte JM, Kiehne K, Herzig KH. 2003. Antimicrobial peptides in innate immunity of the human intestine. J Gastroenterol. 38:717-26.
Otte JM, Podolsky DK. 2004. Functional modulation of enterocytes by gram-positive and gram-negative microorganisms. Am J Physiol Gastrointest Liver Physiol. 286:G613-26.
Ou TY, Liu JW, Leu HS. 2003. Independent prognostic factors for fatality in patients with invasive vibrio cholerae non-O1 infections. J Microbiol Immunol Infect. 36:117-22.
Palmer C, Bik EM, Digiulio DB, Relman DA, Brown PO. 2007. Development of the Human Infant Intestinal Microbiota. PLoS Biol. 5:e177.
Pantano S, Montecucco C. 2006. A molecular model of the Vibrio cholerae cytolysin transmembrane pore. Toxicon. 47:35-40.
Paonessa G, Graziani R, De Serio A, Savino R, Ciapponi L, Lahm A, Salvati AL, Toniatti C, Ciliberto G. 1995. Two distinct and independent sites on IL-6 trigger gp 130 dimer formation and signalling. EMBO J. 14:1942-51.
Paster BJ, Boches SK, Galvin JL, Ericson RE, Lau CN, Levanos VA, Sahasrabudhe A, Dewhirst FE. 2001. Bacterial diversity in human subgingival plaque. J Bacteriol. 183:3770-83.
Pazgier M, Li X, Lu W, Lubkowski J. 2007. Human defensins: synthesis and structural properties. Curr Pharm Des. 13:3096-118.
Pei Z, Bini EJ, Yang L, Zhou M, Francois F, Blaser MJ. 2004. Bacterial biota in the human distal esophagus. Proc Natl Acad Sci U S A. 101:4250-5.
Pennica D, Hayflick JS, Bringman TS, Palladino MA, Goeddel DV. 1985. Cloning and expression in Escherichia coli of the cDNA for murine tumor necrosis factor. Proc Natl Acad Sci USA. 82: 6060-4.
Perez-Vilar J, Eckhardt AE, DeLuca A, Hill RL. 1998. Porcine submaxillary mucin forms disulfide-linked multimers through its amino-terminal D-domains. J Biol Chem. 273:14442-9.
Pflanz S, Hibbert L, Mattson J, Rosales R, Vaisberg E, Bazan JF, Phillips JH, McClanahan TK, de Waal Malefyt R, Kastelein RA. 2004. WSX-1 and glycoprotein 130 constitute a signal-transducing receptor for IL-27. J Immunol. 172:2225-31.
Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C, Du X, Birdwell D, Alejos E, Silva M, Galanos C, Freudenberg M, Ricciardi-Castagnoli P, Layton B, Beutler B. 1998. Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science. 282:2085-8.
Porter EM, Liu L, Oren A, Anton PA, Ganz T. 1997. Localization of human intestinal defensin 5 in Paneth cell granules. Infect Immun. 65:2389-95.
Rahman A, Fahlgren A, Sitohy B, Baranov V, Zirakzadeh A, Hammarström S, Danielsson A, Hammarström ML. 2007. Beta-defensin production by human colonic plasma cells: a new look at plasma cells in ulcerative colitis. Inflamm Bowel Dis. 13:847-55.
Rakoff-Nahoum S, Paglino J, Eslami-Varzaneh F, Edberg S, Medzhitov R. 2004. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell. 118:229-41.

- 66 -
Reinecker HC, Steffen M, Witthoeft T, Pflueger I, Schreiber S, MacDermott RP, Raedler A. 1993. Enhanced secretion of tumour necrosis factor-alpha, IL-6, and IL-1 beta by isolated lamina propria mononuclear cells from patients with ulcerative colitis and Crohn's disease. Clin Exp Immunol. 94:174-81.
Rescigno M, Urbano M, Valzasina B, Francolini M, Rotta G, Bonasio R, Granucci F, Kraehenbuhl JP, Ricciardi-Castagnoli P. 2001. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat Immunol. 2:361-7.
Roach JC, Glusman G, Rowen L, Kaur A, Purcell MK, Smith KD, Hood LE, Aderem A. 2005. The evolution of vertebrate Toll-like receptors. Proc Natl Acad Sci U S A. 102:9577-82.
Rogers NC, Slack EC, Edwards AD, Nolte MA, Schulz O, Schweighoffer E, Williams DL, Gordon S, Tybulewicz VL, Brown GD, Reis e Sousa C. 2005. Syk-dependent cytokine induction by Dectin-1 reveals a novel pattern recognition pathway for C type lectins. Immunity. 22:507-17.
Rogler G, Andus T. 1998. Cytokines in inflammatory bowel disease. World J Surg. 22:382-9.
Rumio C, Besusso D, Arnaboldi F, Palazzo M, Selleri S, Gariboldi S, Akira S, Uematsu S, Bignami P, Ceriani V, Ménard S, Balsari A. 2006. Activation of smooth muscle and myenteric plexus cells of jejunum via Toll-like receptor 4. J Cell Physiol. 208:47-54.
Sack DA, Sack RB, Nair GB, Siddique AK. 2004. Cholera. Lancet. 363:223-33. Saka HA, Bidinost C, Sola C, Carranza P, Collino C, Ortiz S, Echenique JR, Bocco JL.
2008. Vibrio cholerae cytolysin is essential for high enterotoxicity and apoptosis induction produced by a cholera toxin gene-negative V. cholerae non-O1, non-O139 strain. Microb Pathog. 44:118-28.
Sanderson IR. 1999. The physicochemical environment of the neonatal intestine. Am J Clin Nutr. 69:1028S-34S.
Sanderson IR, Walker WA. 2007. TLRs in the Gut I. The role of TLRs/Nods in intestinal development and homeostasis. Am J Physiol Gastrointest Liver Physiol. 292:G6-10.
Sanz Y, Sánchez E, Marzotto M, Calabuig M, Torriani S, Dellaglio F. 2007. Differences in faecal bacterial communities in coeliac and healthy children as detected by PCR and denaturing gradient gel electrophoresis. FEMS Immunol Med Microbiol. 51:562-8.
Schauber J, Svanholm C, Termén S, Iffland K, Menzel T, Scheppach W, Melcher R, Agerberth B, Lührs H, Gudmundsson GH. 2003. Expression of the cathelicidin LL-37 is modulated by short chain fatty acids in colonocytes: relevance of signalling pathways. Gut. 52:735-41.
Schroder K, Hertzog PJ, Ravasi T, Hume DA. 2004. Interferon-gamma: an overview of signals, mechanisms and functions. J Leukoc Biol. 75:163-89.
Schroder K, Sweet MJ, Hume DA. 2006. Signal integration between IFNgamma and TLR signalling pathways in macrophages. Immunobiology. 211:511-24.
Schultsz C, Van Den Berg FM, Ten Kate FW, Tytgat GN, Dankert J. 1999. The intestinal mucus layer from patients with inflammatory bowel disease harbors high numbers of bacteria compared with controls. Gastroenterology. 117:1089-97.
Schumann D, Chen CJ, Kaplan B, Shively JE. 2001. Carcinoembryonic antigen cell adhesion molecule 1 directly associates with cytoskeleton proteins actin and tropomyosin.J Biol Chem. 276:47421-33.
Schölzel S, Zimmermann W, Schwarzkopf G, Grunert F, Rogaczewski B, Thompson J. 2000. Carcinoembryonic antigen family members CEACAM6 and CEACAM7 are

- 67 -
differentially expressed in normal tissues and oppositely deregulated in hyperplastic colorectal polyps and early adenomas. Am J Pathol. 156:595-605.
Sears CL. 2005. A dynamic partnership: celebrating our gut flora. Anaerobe. 11:247-51. Shanahan F. 2005. Physiological basis for novel drug therapies used to treat the
inflammatory bowel diseases. I. Pathophysiological basis and prospects for probiotic therapy in inflammatory bowel disease. Am J Physiol Gastrointest Liver Physiol. 288: G417-21.
Shewry PR, Halford NG. 2002. Cereal seed storage proteins: structures, properties and role in grain utilization. J Exp Bot. 53:947-58.
Shi J, Aono S, Lu W, Ouellette AJ, Hu X, Ji Y, Wang L, Lenz S, van Ginkel FW, Liles M, Dykstra C, Morrison EE, Elson CO. 2007. A novel role for defensins in intestinal homeostasis: regulation of IL-1beta secretion. J Immunol. 179:1245-53.
Silva TM, Schleupner MA, Tacket CO, Steiner TS, Kaper JB, Edelman R, Guerrant R. 1996. New evidence for an inflammatory component in diarrhea caused by selected new, live attenuated cholera vaccines and by El Tor and Q139 Vibrio cholerae. Infect Immun. 64:2362-4.
Singer BB, Scheffrahn I, Obrink B. 2000. The tumor growth-inhibiting cell adhesion molecule CEACAM1 (C-CAM) is differently expressed in proliferating and quiescent epithelial cells and regulates cell proliferation. Cancer Res. 60:1236-44.
Sjöström H, Lundin KE, Molberg O, Körner R, McAdam SN, Anthonsen D, Quarsten H, Norén O, Roepstorff P, Thorsby E, Sollid LM. 1998. Identification of a gliadin T-cell epitope in coeliac disease: general importance of gliadin deamidation for intestinal T-cell recognition. Scand J Immunol. 48:111-5.
Sollid LM. 2000. Molecular basis of celiac disease. Annu Rev Immunol. 18:53-81. Sollid LM, Thorsby E. 1993. HLA susceptibility genes in celiac disease: genetic mapping
and role in pathogenesis. Gastroenterology. 105:910-22. Stadnyk AW, Waterhouse CC. 1997. Epithelial cytokines in intestinal inflammation and
mucosal immunity. Curr Opin Gastroenterol. 13:510-7. Strous GJ, Dekker J. 1992. Mucin-type glycoproteins. Crit Rev Biochem Mol Biol. 27:57-
92. Strugala V, Allen A, Dettmar PW, Pearson JP. 2003. Colonic mucin: methods of measuring
mucus thickness. Proc Nutr Soc. 62:237-43. Sundberg U, Beauchemin N, Obrink B. 2004. The cytoplasmic domain of CEACAM1-L
controls its lateral localization and the organization of desmosomes in polarized epithelial cells. J Cell Sci. 117:1091-104.
Sutterwala FS, Mijares LA, Li L, Ogura Y, Kazmierczak BI, Flavell RA. 2007. Immune recognition of Pseudomonas aeruginosa mediated by the IPAF/NLRC4 inflammasome. J Exp Med. 204:3235-45.
Suzuki M, Hisamatsu T, Podolsky DK. 2003. Gamma interferon augments the intracellular pathway for lipopolysaccharide (LPS) recognition in human intestinal epithelial cells through coordinated up-regulation of LPS uptake and expression of the intracellular Toll-like receptor 4-MD-2 complex. Infect Immun. 71:3503-11.
Swidsinski A, Ladhoff A, Pernthaler A, Swidsinski S, Loening-Baucke V, Ortner M, Weber J, Hoffmann U, Schreiber S, Dietel M, Lochs H. 2002. Mucosal flora in inflammatory bowel disease. Gastroenterology. 122:44-54.
Swidsinski A, Loening-Baucke V, Lochs H, Hale LP. 2005. Spatial organization of bacterial flora in normal and inflamed intestine: a fluorescence in situ hybridization study in mice. World J Gastroenterol. 11:1131-40.

- 68 -
Swidsinski A, Weber J, Loening-Baucke V, Hale LP, Lochs H. 2005. Spatial organization and composition of the mucosal flora in patients with inflammatory bowel disease. J Clin Microbiol. 43:3380-9.
Söderström K, Bucht A, Halapi E, Lundqvist C, Gronberg A, Nilsson E, Orsini DL, van de Wal Y, Koning F, Hammarström ML, Kiessling R. 1994. High expression of V gamma 8 is a shared feature of human gamma delta T cells in the epithelium of the gut and in the inflamed synovial tissue. J Immunol. 152:6017-27.
Tacket CO, Kotloff KL, Losonsky G, Nataro JP, Michalski J, Kaper JB, Edelman R, Levine MM. 1997. Volunteer studies investigating the safety and efficacy of live oral El Tor Vibrio cholerae O1 vaccine strain CVD 111. Am J Trop Med Hyg. 56:533-7.
Taga T, Kishimoto T. 1997. Gp130 and the interleukin-6 family of cytokines. Annu Rev Immunol. 15:797-819.
Takeuchi O, Hoshino K, Kawai T, Sanjo H, Takada H, Ogawa T, Takeda K, Akira S. 1999. Differential roles of TLR2 and TLR4 in recognition of gram-negative and gram-positive bacterial cell wall components. Immunity. 11:443-51.
Takeuchi O, Kawai T, Mühlradt PF, Morr M, Radolf JD, Zychlinsky A, Takeda K, Akira S. 2001. Discrimination of bacterial lipoproteins by Toll-like receptor 6. Int Immunol. 13:933-40.
Takeuchi O, Sato S, Horiuchi T, Hoshino K, Takeda K, Dong Z, Modlin RL, Akira S. 2002. Cutting edge: role of Toll-like receptor 1 in mediating immune response to microbial lipoproteins. J Immunol. 169:10-4.
Tang P, Hung MC, Klostergaard J. 1996. Human pro-tumor necrosis factor is a homotrimer. Biochemistry. 35:8216-25.
Taylor K, Barran PE, Dorin JR. 2008. Structure-activity relationships in beta-defensin peptides. Biopolymers. 90:1-7.
Thomson AW and Lotze MT. Editors, The Cytokine Handbook, Academic Press (2003), pp. 837-60.
Tjellström B, Stenhammar L, Högberg L, Fälth-Magnusson K, Magnusson KE, Midtvedt T, Sundqvist T, Houlston R, Popat S, Norin E. 2007. Gut microflora associated characteristics in first-degree relatives of children with celiac disease. Scand J Gastroenterol. 42:1204-8.
Tjellström B, Stenhammar L, Högberg L, Fälth-Magnusson K, Magnusson KE, Midtvedt T, Sundqvist T, Norin E. 2005. Gut microflora associated characteristics in children with celiac disease. Am J Gastroenterol. 100:2784-8.
Tlaskalová-Hogenová H, Stepánková R, Hudcovic T, Tucková L, Cukrowska B, Lodinová-Zádníková R, Kozáková H, Rossmann P, Bártová J, Sokol D, Funda DP, Borovská D, Reháková Z, Sinkora J, Hofman J, Drastich P, Kokesová A. 2004. Commensal bacteria (normal microflora), mucosal immunity and chronic inflammatory and autoimmune diseases. Immunol Lett. 93:97-108.
Tracey KJ, Cerami A. Tumor necrosis factor, other cytokines and disease. 1993. Annu Rev Cell Biol. 9:317-43.
Treem WR. 2004. Emerging concepts in celiac disease. Curr Opin Pediatr. 16:552-9. Triantafilou M, Triantafilou K. 2002. Lipopolysaccharide recognition: CD14, TLRs and the
LPS-activation cluster. Trends Immunol. 23:301-4. Trinchieri G, Sher A. 2007. Cooperation of Toll-like receptor signals in innate immune
defence. Nat Rev Immunol. 7:179-90. Troncone R, Ivarsson A, Szajewska H, Mearin ML; Members of European Multistakeholder
Platform on CD (CDEUSSA). 2008. Review article: future research on coeliac disease -

- 69 -
a position report from the European multistakeholder platform on coeliac disease (CDEUSSA). Aliment Pharmacol Ther. 27:1030-43.
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER, Gordon JI. 2006. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 444:1027-31.
Tytgat KM, Büller HA, Opdam FJ, Kim YS, Einerhand AW, Dekker J. 1994. Biosynthesis of human colonic mucin: Muc2 is the prominent secretory mucin. Gastroenterology. 107:1352-63.
Uematsu S, Jang MH, Chevrier N, Guo Z, Kumagai Y, Yamamoto M, Kato H, Sougawa N, Matsui H, Kuwata H, Hemmi H, Coban C, Kawai T, Ishii KJ, Takeuchi O, Miyasaka M, Takeda K, Akira S. 2006. Detection of pathogenic intestinal bacteria by Toll-like receptor 5 on intestinal CD11c+ lamina propria cells. Nat Immunol. 7:868-74.
Vaitkevicius K, Rompikuntal PK, Lindmark B, Vaitkevicius R, Song T, Wai SN. 2008. The metalloprotease PrtV from Vibrio cholerae. FEBS J. 275:3167-77.
Valeva A, Walev I, Weis S, Boukhallouk F, Wassenaar TM, Bhakdi S. 2008. Pro-inflammatory feedback activation cycle evoked by attack of Vibrio cholerae cytolysin on human neutrophil granulocytes. Med Microbiol Immunol. 197:285-93.
van Boxel-Dezaire AH, Stark GR. 2007. Cell type-specific signaling in response to interferon-gamma. Curr Top Microbiol Immunol. 316:119-54.
van de Wal Y, Kooy Y, van Veelen P, Peña S, Mearin L, Papadopoulos G, Koning F. 1998. Selective deamidation by tissue transglutaminase strongly enhances gliadin-specific T cell reactivity. J Immunol. 161:1585-8.
van Heel DA, West J. 2006. Recent advances in coeliac disease. Gut. 55:1037-46. Van Klinken BJ, Dekker J, Büller HA, Einerhand AW. 1995. Mucin gene structure and
expression: protection vs. adhesion. Am J Physiol. 269:G613-27. van Vliet SJ, García-Vallejo JJ, van Kooyk Y. 2008. Dendritic cells and C-type lectin
receptors: coupling innate to adaptive immune responses. Immunol Cell Biol. 86:580-7. Vanden Broeck D, Horvath C, De Wolf MJ. 2007. Vibrio cholerae: cholera toxin. Int J
Biochem Cell Biol. 39: 1771-5. Verstrepen L, Bekaert T, Chau TL, Tavernier J, Chariot A, Beyaert R. 2008. TLR-4, IL-1R
and TNF-R signaling to NF-kappaB: variations on a common theme. Cell Mol Life Sci. 65:2964-78.
Vigers GP, Dripps DJ, Edwards CK 3rd, Brandhuber BJ. 2000. X-ray crystal structure of a small antagonist peptide bound to interleukin-1 receptor type 1. J Biol Chem. 275:36927-33.
Wajant H, Pfizenmaier K, Scheurich P. 2003. Tumor necrosis factor signaling. Cell Death Differ. 10:45-65.
Wallach D, Varfolomeev EE, Malinin NL, Goltsev YV, Kovalenko AV, Boldin MP. 1999. Tumor necrosis factor receptor and Fas signaling mechanisms. Annu Rev Immunol. 17:331-67.
Wang J, Al-Lamki RS, Zhang H, Kirkiles-Smith N, Gaeta ML, Thiru S, Pober JS, Bradley JR. 2003. Histamine antagonizes tumor necrosis factor (TNF) signaling by stimulating TNF receptor shedding from the cell surface and Golgi storage pool. J Biol Chem. 278:21751-60.
Wang M, Ahrné S, Jeppsson B, Molin G. 2005. Comparison of bacterial diversity along the human intestinal tract by direct cloning and sequencing of 16S rRNA genes. FEMS Microbiol Ecol. 54:219-31.
Wapenaar MC, Monsuur AJ, van Bodegraven AA, Weersma RK, Bevova MR, Linskens RK, Howdle P, Holmes G, Mulder CJ, Dijkstra G, van Heel DA, Wijmenga C. 2008.

- 70 -
Associations with tight junction genes PARD3 and MAGI2 in Dutch patients point to a common barrier defect for coeliac disease and ulcerative colitis. Gut. 57:463-7.
Waugh DJ, Wilson C. 2008. The interleukin-8 pathway in cancer. Clin Cancer Res. 14:6735-41.
Wehkamp J, Chu H, Shen B, Feathers RW, Kays RJ, Lee SK, Bevins CL. 2006. Paneth cell antimicrobial peptides: topographical distribution and quantification in human gastrointestinal tissues. FEBS Lett. 580:5344-50.
Wehkamp J, Harder J, Weichenthal M, Mueller O, Herrlinger KR, Fellermann K, Schroeder JM, Stange EF. 2003. Inducible and constitutive beta-defensins are differentially expressed in Crohn's disease and ulcerative colitis. Inflamm Bowel Dis. 9:215-23.
Wehkamp J, Schauber J, Stange EF. 2007. Defensins and cathelicidins in gastrointestinal infections. Curr Opin Gastroenterol. 23:32-8.
Wershil BK, Furuta GT. 2008. 4. Gastrointestinal mucosal immunity. J Allergy Clin Immunol. 121:S380-3.
West J, Logan RF, Hill PG, Khaw KT. 2007. The iceberg of celiac disease: what is below the waterline? Clin Gastroenterol Hepatol. 5:59-62.
West J, Logan RF, Hill PG, Lloyd A, Lewis S, Hubbard R, Reader R, Holmes GK, Khaw KT. 2003. Seroprevalence, correlates, and characteristics of undetected coeliac disease in England. Gut. 52:960-5.
Williams SJ, Wreschner DH, Tran M, Eyre HJ, Sutherland GR, McGuckin MA. 2001. Muc13, a novel human cell surface mucin expressed by epithelial and hemopoietic cells. J Biol Chem. 276:18327-36.
Wittig BM, Zeitz M. 2003. The gut as an organ of immunology. Int J Colorectal Dis. 18: 181-7.
World Health Organization, Cholera Fact Sheet No. 107, Revised September 2007, Geneva, Switzerland: World Health Organization
Xiao T. 2008. Innate immune recognition of nucleic acids. Immunol Res. Sep 23. [Epub ahead of print]
Xu D, Qu CK. 2008. Protein tyrosine phosphatases in the JAK/STAT pathway. Front Biosci. 13, 4925-32.
Xu GY, Yu HA, Hong J, Stahl M, McDonagh T, Kay LE, Cumming DA. 1997. Solution structure of recombinant human interleukin-6. J Mol Biol. 268:468-81.
Yang D, Chertov O, Bykovskaia SN, Chen Q, Buffo MJ, Shogan J, Anderson M, Schröder JM, Wang JM, Howard OM, Oppenheim JJ. 1999. Beta-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science. 286:525-8.
Yeung MM, Melgar S, Baranov V, Öberg Å, Danielsson Å, Hammarström S, Hammarström ML. 2000. Characterisation of mucosal lymphoid aggregates in ulcerative colitis: immune cell phenotype and TcR-gammadelta expression. Gut. 47:215-27.
Yoneyama M, Fujita T. 2007. Function of RIG-I-like receptors in antiviral innate immunity. 2007. J Biol Chem. 282:15315-8.
Yoneyama M, Fujita T. 2008. Structural mechanism of RNA recognition by the RIG-I-like receptors. Immunity. 29:178-81.
Zebhauser R, Kammerer R, Eisenried A, McLellan A, Moore T, Zimmermann W. 2005. Identification of a novel group of evolutionarily conserved members within the rapidly diverging murine CEA family.Genomics. 86:566-80.
Zelensky AN, Gready JE. 2005. The C-type lectin-like domain superfamily. FEBS J. 272: 6179-217

- 71 -
Zhang T, Breitbart M, Lee WH, Run JQ, Wei CL, Soh SW, Hibberd ML, Liu ET, Rohwer F, Ruan Y. 2006. RNA viral community in human feces: prevalence of plant pathogenic viruses. PLoS Biol. 4:e3.
Zitzer A, Wassenaar TM, Walev I, Bhakdi S. 1997. Potent membrane-permeabilizing and cytocidal action of Vibrio cholerae cytolysin on human intestinal cells. Infect Immun. 65:1293-8.
Zitzer A, Zitzer O, Bhakdi S, Palmer M. 1999. Oligomerization of Vibrio cholerae cytolysin yields a pentameric pore and has a dual specificity for cholesterol and sphingolipids in the target membrane. J Biol Chem. 274:1375-80.
Öbrink B. 1997. CEA adhesion molecules: multifunctional proteins with signal-regulatory properties. Curr Opin Cell Biol. 9:616-26.


















