
HIDROLISIS PATI PALMA MENGGUNAKAN...
70
HIDROLISIS PATI PALMA MENGGUNAKAN PULLULANASE DAN β-AMILASE SKRIPSI SITI AMINAH OKTAVIYANI F34080117 FAKULTAS TEKNOLOGI PERTANIAN INSTITUT PERTANIAN BOGOR BOGOR 2012
Transcript of HIDROLISIS PATI PALMA MENGGUNAKAN...

HIDROLISIS PATI PALMA MENGGUNAKAN PULLULANASE
DAN β-AMILASE
SKRIPSI
SITI AMINAH OKTAVIYANI
F34080117
FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
2012

HYDROLYSIS OF PALM STARCH BY USING PULLULANASE
AND β-AMYLASE
Siti Aminah Oktaviyani and Titi Candra Sunarti
Department of Agro-Industrial Technology, Faculty of Agricultural Technology,
Bogor Agricultural University, IPB Darmaga Campus, PO Box 220, Bogor, West Java, Indonesia.
Email: [email protected]
ABSTRACT
Palm trees are unexplored carbohydrate sources that wiidely spread in Indonesia. Palm
starches are still under utilized since lacked information about their pysico-chemical properties.
Hydrolysis of these starches are the right application without concerning their characteristics. This
research investigated the starches from five types of palm trees, namely Metroxylon sago (commersial
sago), Metroxylon sp. (sagu rumbia), Arenga pinnata, Arenga microcarpa (sagu baruk), and Caryota
mitis; and the hydrolysis of palm starches by using degrading enzymes, i.e. pullulanase and β-
amylase. The result showed that palm starches are normal starch which contain17-21% of amylose or
83-79% of amylopectin which differed in branch-chain lengths, and caused typical products from
each type of palm. Pullulanase as debranching enzymes, cleaveged the starch to produce DP 13.9-
20.8 of Short-Chains Amylose (SCA). The starch contained 22.2-38.8% of β-limit dextrins as the
residue of β-amylolysis. Simultaneous hydrolysis by using β-amylase and pullulanase mainly
produced maltose for all starch with DP of 1.7-2.9
Key words : palm starches, hydrolysis, pullulanase, β-amylase

Siti Aminah Oktaviyani. F34080117. Hidrolisis Pati Palma Menggunakan Pullulanase dan β-
Amilase. Di bawah bimbingan Titi Candra Sunarti. 2012.
RINGKASAN
Pati merupakan salah satu substansi yang tersebar luas di alam sebagai cadangan karbohidrat
pada tanaman. Pati dapat diperoleh dari berbagai sumber, seperti umbi-umbian, serealia, dan empulur
batang palma. Tanaman palma merupakan tanaman yang potensial sebagai sumber pati alami dan
dapat menjadi alternatif pengganti beras, jagung, maupun gandum karena jumlahnya melimpah di
Indonesia. Dari 2800 jenis palma di dunia, sekitar 460 jenis tersebar merata di daerah-daerah
Indonesia. Sagu merupakan salah satu contoh tanaman palma penghasil pati dimana 60% lahan sagu
dunia berada di Indonesia yang 90 % areanya tersebar di Papua (Matanubun 2005). Selain sagu masih
banyak tanaman palma lain yang belum termanfaatkan secara optimal, seperti aren, sagu baruk, dan
Caryota mitis.
Pengembangan dan pemanfaatan tanaman palma sebagai bahan baku industri, baik pangan
maupun non-pangan, dapat berupa hidrolisat pati yang diperoleh dari proses hidrolisis pati secara
enzimatis. Hidrolisat pati merupakan produk turunan pati hasil pemutusan rantai-rantai glukosidik
pada pati oleh enzim. Hidrolisat pati merupakan produk yang lebih mudah dan telah banyak
digunakan di industri dibandingkan pati alami karena memiliki sifat yang lebih konsisten yang
memudahkan dalam pengontrolan dan pembuatan produk dengan kualitas yang baik. Pada penelitian
ini pati palma sebagai substrat hidrolisis yang digunakan terdiri atas sagu rumbia (Metroxylon sp.),
sagu baruk (Arenga microcarpa), sagu komersial, aren (Arenga pinnata), dan Caryota mitis. Enzim
yang digunakan, yaitu β-amilase (EC 3.2.1.2) dari barley (SIGMA) yang dapat menghasilkan produk
hidrolisat pati berupa maltosa dan β-limit dekstrin serta pullulanase (A.aerogenes, Hayashibara
Biochemical Lab.) yang dapat menghasilkan produk maltooligosakarida dan Short-Chain Amylose
(SCA).
Penelitian ini bertujuan untuk mengkaji hidrolisis secara enzimatis dengan pullulanase dan β-
amilase beberapa jenis pati palma di Indonesia yang berpotensi dan belum tereksplorasi untuk bahan
baku industri dengan melihat perubahan nilai Derajat Polimerisasi (DP), Dextrose Equivalent (DE),
dan tingkat hidrolisis. Metodologi yang digunakan dalam penelitian ini terdiri atas dua tahap, yaitu
tahap persiapan dan tahap proses hidrolisis. Pada tahap persiapan dilakukan pengujian kadar amilosa
untuk masing-masing pati palma, penghilangan kandungan protein dan lemak pada pati, serta
karakterisasi enzim yang digunakan. Tahap hidrolisis dibuat larutan pati tergelatinisasi 0,4% yang
dihidrolisis oleh 0,2 U/g β-amilase dan 0,45 U/g pullulanase. Proses hidrolisis dengan kedua enzim
tersebut dilakukan selama 48 jam. Selama proses hidrolisis dilakukan sampling untuk analisa total
gula dengan metode Fenol-Sulfat dan gula pereduksi dengan modifikasi metode Park Johnson yang
digunakan untuk menghitung tingkat hidrolisis, persentase kandungan β-limit dekstrin, DE, dan DP.
Pati palma termasuk tipe pati normal dengan kandungan amilosa berkisar antara 20,1-26,7%
dengan kandungan amilosa tertinggi pada sagu rumbia. Hidrolisis pati palma dengan β-amilase terjadi
pada ikatan α-1,4-glukosidik yang menghasilkan maltosa dan residu β-limit dekstrin yang merupakan
bagian rantai percabangan pada ikatan α-1,6-glukosidik yang tidak terhidrolisis oleh β-amilase.
Adapun kandungan β-limit dekstrin yang dihasilkan dari ketujuh pati palma yang digunakan berkisar
antara 22,2–38,8% dari total pati. Hidrolisis pati palma dengan pullulanase terjadi pada ikatan
percabangan pada gugus amilopektin, yaitu pada ikatan α-1,6-glukosidik sehingga menghasilkan
produk yang memiliki rantai-rantai lurus yang cukup pendek dan memiliki bobot molekul yang lebih

rendah. Produk yang dihasilkan ini berupa Short-Chain Amylose (SCA) atau maltooligosakarida.
Berdasarkan hasil hidrolisis yang dilakukan diperoleh nilai DP rata-rata pada tingkat hidrolisis 100%
untuk masing-masing pati palma berkisar antara 13,9-20,8 dan nilai DE rata-rata berkisar antara 4,8–
7,2.
Hidrolisis dengan menggunakan kedua enzim tersebut yang ditambahkan secara suksesif
akan menghasilkan kerja enzim yang simultan dalam memecah rantai-rantai pada ikatan α-1,4-
glukosidik dan α-1,6-glukosidik sehingga dapat menghasilkan produk hidrolisat pati yang memiliki
rantai pendek-pendek berupa maltosa. Berdasarkan hasil hidrolisis masing-masing pati palma yang
dilakukan oleh β-amilase dan pullulanase ini diperoleh nilai DP rata-rata pada tingkat hidrolisis 100%
berkisar antara 1,7–2,9 dengan nilai DE rata-rata berkisar antara 35,1–57,3 yang menunjukkan produk
hidrolisat pati mengandung campuran maltosa, maltotriosa, dan glukosa.
Berdasarkan nilai DP dan DE yang dihasilkan hidrolisat pati dari tujuh pati palma yang
digunakan oleh masing-masing enzim terlihat adanya keragaman yang diakibatkan oleh keragaman
kandungan fraksi amilopektin yang menyusun masing-masing pati palma. Keragaman hasil ini dapat
dimanfaatkan oleh industri-industri hidrolisat pati untuk menghasilkan produk hidrolisat sesuai
kebutuhan dan kegunaan yang diinginkan. Dari hasil-hasil yang diperoleh dapat disimpulkan bahwa
berbagai jenis pati palma berpotensi untuk diolah menjadi hidrolisat pati yang dapat dimanfaatkan
pada berbagai bidang aplikasi terutama bidang pangan.

HIDROLISIS PATI PALMA MENGGUNAKAN PULLULANASE
DAN β-AMILASE
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar
SARJANA TEKNOLOGI PERTANIAN
Pada Departemen Teknologi Industri Pertanian
Fakultas Teknologi Pertanian
Institut Pertanian Bogor
Oleh :
SITI AMINAH OKTAVIYANI
F34080117
FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
2012

Judul Skripsi : Hidrolisis Pati Palma Menggunakan Pullulanase dan β-Amilase
Nama : Siti Aminah Oktaviyani
NIM : F34080117
Menyetujui,
Pembimbing,
Dr. Ir. Titi Candra Sunarti, M.Si.
NIP 19661219 199103 2 001
Mengetahui,
Ketua Departemen
Teknologi Industri Pertanian
Prof. Dr. Nastiti Siswi Indrasti
NIP. 19621009 198903 2 001
Tanggal lulus : 26 November 2012

PERNYATAAN MENGENAI SKRIPSI DAN SUMBER INFORMASI
Saya menyatakan dengan sebenar-benarnya bahwa skripsi dengan judul Hidrolisis Pati Palma
Menggunakan Pullulanase dan β-Amilase adalah hasil karya saya sendiri dengan arahan Dosen
Pembimbing Akademik dan belum diajukan dalam bentuk apapun pada perguruan tinggi manapun.
Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari
penulis lain disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Bogor, November 2012
Yang membuat pernyataan
Siti Aminah Oktaviyani
F34080117

© Hak cipta milik Institut Pertanian Bogor, tahun 2012
Hak cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan
sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah,
penyusunan laporan, penulisan kritik atau tinjauan suatu masalah, dan pengutipan tersebut tidak
merugikan kepentingan IPB. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh
karya tulis dalam bentuk apapun tanpa izin IPB.

BIODATA PENULIS
Siti Aminah Oktaviyani dilahirkan di Jakarta pada tanggal 17 Oktober 1990
sebagai anak ketiga dari lima bersaudara dari pasangan Hendi Setiawan dan
Etty Eliyani. Penulis menamatkan sekolah dasar di SD Negeri Pekayon 018
Jakarta pada tahun 2002 lalu melanjutkan ke SMP Negeri 91 Jakarta pada
tahun 2002-2005. Pada tahun 2008, penulis lulus dari SMA Negeri 39 Jakarta
dan melanjutkan studi di Institut Pertanian Bogor melalui jalur Seleksi
Nasional Masuk Perguruan Tinggi Negeri (SNMPTN) dengan mengambil
pilihan mayor Teknologi Industri Pertanian. Penulis menerima beasiswa
Peningkatan Prestasi Akademik (PPA) pada tahun 2009 hingga 2012. Selama
masa kuliah penulis pernah menjadi asisten praktikum Teknologi Pati, Gula, dan Sukrokimia serta
Teknologi Minyak, Lemak, dan Oleokimia. Selain menjalani aktivitas akademik, penulis pernah
terlibat dalam kegiatan RAMP 2012 sebagai tutor dan aktif mengikuti beberapa organisasi kampus,
yaitu Badan Eksekutif Mahasiswa Fakultas Teknologi Pertanian (BEM-F) sebagai bendahara
departemen Minat Bakat Mahasiswa (MBM) tahun 2010-2011, Himpunan Mahasiswa Teknologi
Industri (HIMALOGIN) sebagai bendahara departemen Public Relation tahun 2009-2010, anggota
Himpunan Mahasiswa Teknologi Industri (HIMALOGIN) tahun 2009-2011, serta anggota Unit
Kegiatan Mahasiswa (UKM) Seni Sunda Gentra Kaheman tahun 2008-2009. Penulis melaksanakan
praktik lapang di PT Tirta Marta Serang dengan judul “Mempelajari Aspek Proses Produksi dan
Pengawasan Mutu Shopping Bag Berbahan Baku Starch-Based Biodegradable Plastics”.

i
KATA PENGANTAR
Puji syukur atas kehadirat Allah SWT yang telah melimpahkan segala rahmat dan hidayah-Nya
sehingga penulis dapat menyelesaikan penulisan skripsi dengan judul Hidrolisis Pati Palma
Menggunakan Pullulanase dan β-Amilase. Skripsi ini ditulis berdasarkan hasil penelitian yang
dilaksanakan di Laboratorium Teknologi Bioindustri dan Teknik Kimia Departemen Teknologi
Industri Pertanian IPB sejak bulan Februari hingga September 2012.
Selama pelaksanaan dan penulisan skripsi ini, penulis mendapatkan banyak bantuan baik
secara moril maupun materil dari berbagai pihak. Oleh karena itu, penulis ingin mengucapkan terima
kasih kepada :
1. Dr. Ir. Titi Candra Sunarti, M.Si, selaku dosen pembimbing akademik yang telah memberikan
arahan, bantuan, serta kesabaran dalam membimbing penulis.
2. Dr. Ir. Liesbetini Haditjaroko, MS, selaku dosen penguji yang telah memberikan masukan yang
membangun dalam penyempurnaan penulisan skripsi ini.
3. Dr. Ir. Muslich, M.Si, selaku dosen penguji atas segala saran dalam penyempurnaan skripsi ini.
4. Kedua orang tua (Hendi Setiawan dan Etty Eliyani), kakak-kakak (Teh Aci dan A’Bobi), serta
adik-adik (Lia dan Iqbal) penulis yang selalu mendukung, mendoakan, dan memberikan nasihat
yang membangun.
5. Seluruh laboran Departemen Teknologi Industri Pertanian yang telah membantu, memberi
masukan, dan mengajari penulis selama penelitian berlangsung.
6. Cici, Mia, Rathi, Bunga, Fira, Niza, Cecha sebagai teman yang bersedia membantu dan memberi
semangat kepada penulis semasa kuliah dan penelitian serta Puja Dwi Sari dan Dony Noor
Romadona sebagai rekan sebimbingan yang telah membantu kelancaran selama penelitian.
7. Keluarga besar TIN 45 dan para HSL (Himpunan Solidaritas Lab), atas kebersamaan, semangat,
dan bantuannya.
8. Semua pihak yang telah membantu penulis yang tidak dapat disebutkan satu per satu.
Dengan segala kekurangan yang masih banyak terdapat di dalamnya, penulis berharap tulisan
ini dapat mendatangkan manfaat bagi siapapun yang membutuhkannya. Semoga tulisan ini menjadi
salah satu amalan baik penulis di hadapan Allah SWT. Amin.
Bogor, November 2012
Siti Aminah Oktaviyani

ii
DAFTAR ISI
Halaman
KATA PENGANTAR ........................................................................................................................ i
DAFTAR TABEL ............................................................................................................................. iii
DAFTAR GAMBAR ........................................................................................................................ iv
DAFTAR LAMPIRAN ..................................................................................................................... v
I. PENDAHULUAN .............................................................................................................................. 1
1.1 LATAR BELAKANG ........................................................................................................................ 1
1.2 TUJUAN.............................................................................................................................................. 2
II. TINJAUAN PUSTAKA .................................................................................................................... 3
2.1 PATI ............................................................................................................................................... 3
2.2 PATI PALMA ..................................................................................................................................... 4
2.3 HIDROLISIS PATI ............................................................................................................................. 8
2.4 ENZIM (β-AMILASE DAN PULLULANASE) ............................................................................... 8
III. METODOLOGI............................................................................................................................... 13
3.1 WAKTU DAN TEMPAT PENELITIAN ........................................................................................ 13
3.2 ALAT DAN BAHAN ....................................................................................................................... 13
3.2.1 Alat ................................................................................................................................... 13
3.2.2 Bahan ............................................................................................................................... 13
3.3 METODE PENELITIAN .................................................................................................................. 13
3.2.1 Persiapan Bahan ............................................................................................................... 13
3.3.2 Penentuan Aktivitas Enzim ............................................................................................. 14
3.3.3 Hidrolisis Pati Palma oleh β-amilase dan Pullulanase .................................................... 14
IV. HASIL DAN PEMBAHASAN ....................................................................................................... 16
4.1 KANDUNGAN AMILOSA PADA PATI PALMA ........................................................................ 16
4.2 HIDROLISIS ENZIMATIS PATI PALMA .................................................................................... 17
4.2.1 Hidrolisis Pati Palma dengan Pullulanase ....................................................................... 18
4.2.2 Hidrolisis Pati Palma dengan β-Amilase dan Pullulanase secara Suksesif .................... 25
4.3 APLIKASI HIDROLISAT PATI PALMA ...................................................................................... 29
V. PENUTUP ........................................................................................................................................ 34
5.1 Kesimpulan ........................................................................................................................................ 34
5.2 Saran ............................................................................................................................................. 34
DAFTAR PUSTAKA ...................................................................................................................... 35
LAMPIRAN ..................................................................................................................................... 37

iii
DAFTAR TABEL
Halaman
Tabel 1. Perbedaan amilosa dan amilopektin ................................................................................... 4
Tabel 2. Komposisi fisiko-kimia tepung sagu (per 100 g) ............................................................... 5
Tabel 3. Komponen kimia pati sagu ................................................................................................. 6
Tabel 4. Nilai gizi tepung sagu baruk basah asal Kepulauan Sangihe ............................................. 7
Tabel 5. Aplikasi produk hidrolisat pati secara umum ..................................................................... 8
Tabel 6. Produk hidrolisat enzimatis dan aplikasinya .................................................................... 12
Tabel 7. Rasio amilosa dan amilopektin pada pati palma .............................................................. 16
Tabel 8. Aktivitas dan kondisi optimum enzim .............................................................................. 18
Tabel 9. Nilai DP dan DE rata-rata pada hidrolisis 10, 40, dan 100% dengan pullulanase .......... 24
Tabel 10. Nilai β-limit dekstrin pati palma ....................................................................................... 26
Tabel 11. Nilai DP dan DE rata-rata pada hidrolisis 10, 40, dan 100% dengan β-amilase dan
pullulanase secara suksesif ............................................................................................... 29

iv
DAFTAR GAMBAR
Halaman
Gambar 1. Struktur kimia (a) amilosa (b) amilopektin ........................................................................ 3
Gambar 2. Daerah persebaran tanaman sagu di Indonesia ................................................................... 5
Gambar 3. Titik pemutusan rantai amilosa pada pati oleh β-amilase .................................................. 9
Gambar 4. Pembentukan β-limit dekstrin ............................................................................................. 9
Gambar 5. Titik pemutusan percabangan rantai amilopektin oleh pullulanase ................................. 11
Gambar 6. Produk-produk hidrolisat pati enzimatis ........................................................................... 11
Gambar 7. Pola hidrolisis pati palma dengan pullulanase .................................................................. 20
Gambar 8. Model klaster pada fraksi amilopektin ............................................................................. 20
Gambar 9. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 1 dengan pullulanase ........ 22
Gambar 10. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 2 dengan pullulanase ........ 22
Gambar 11. Perubahan nilai DP dan DE pada hidrolisis pati sagu 3 dengan pullulanase ................... 22
Gambar 12. Perubahan nilai DP dan DE pada hidrolisis pati sagu rumbia dengan pullulanase ......... 22
Gambar 13. Perubahan nilai DP dan DE pada hidrolisis pati sagu komersial dengan pullulanase ..... 23
Gambar 14. Perubahan nilai DP dan DE pada hidrolisis pati aren dengan pullulanase ...................... 23
Gambar 15. Perubahan nilai DP dan DE pada hidrolisis pati Caryota mitis dengan pullulanase ....... 23
Gambar 16. Pola hidrolisis pati palma dengan β-amilase dan pullulanase secara suksesif ................ 26
Gambar 17. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 1 dengan β-amilase dan
pullulanase secara suksesif ............................................................................................... 30
Gambar 18. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 2 dengan β-amilase dan
pullulanase secara suksesif ............................................................................................... 30
Gambar 19. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 3 dengan β-amilase dan
pullulanase secara suksesif ............................................................................................... 30
Gambar 20. Perubahan nilai DP dan DE pada hidrolisis pati sagu rumbia dengan β-amilase dan
pullulanase secara suksesif ............................................................................................... 30
Gambar 21. Perubahan nilai DP dan DE pada hidrolisis pati sagu komersial dengan β-amilase dan
pullulanase secara suksesif ............................................................................................... 31
Gambar 22. Perubahan nilai DP dan DE pada hidrolisis pati aren dengan β-amilase dan pullulanase
secara suksesif ................................................................................................................... 31
Gambar 23. Perubahan nilai DP dan DE pada hidrolisis pati Caryota mitis dengan β-amilase dan
pullulanase secara suksesif ............................................................................................... 31

v
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Jenis-jenis pati palma yang digunakan ............................................................................. 38
Lampiran 2. Prosedur analisa kadar amilosa ......................................................................................... 39
Lampiran 3. Prosedur analisa hidrolisat pati ........................................................................................ 40
Lampiran 4. Rumus perhitungan hasil analisa hidrolisat pati .............................................................. 42
Lampiran 5. Perhitungan aktivitas enzim ............................................................................................. 43
Lampiran 6. Hasil analisa nilai DE, DP, dan tingkat hidrolisis pati palma dengan pullulanase ......... 46
Lampiran 7. Hasil analisa nilai DE, DP, β-limit dekstrin dan tingkat hidrolisis pati palma dengan β-
amilase dan pullulanase secara suksesif ........................................................................... 52

1
I. PENDAHULUAN
1.1 LATAR BELAKANG
Pati merupakan salah satu substansi yang tersebar luas di alam sebagai cadangan karbohidrat
pada tanaman. Pati dapat diperoleh dari berbagai sumber, seperti umbi-umbian, serealia, dan empulur
batang palma. Tanaman palma merupakan tanaman yang potensial sebagai sumber pati alami dan
dapat menjadi alternatif pengganti beras, jagung, maupun gandum karena jumlahnya melimpah di
Indonesia. Dari 2800 jenis palma di dunia, sekitar 460 jenis tersebar merata di daerah-daerah
Indonesia. Sagu merupakan salah satu contoh tanaman palma penghasil pati dimana 60% lahan sagu
dunia berada di Indonesia yang 90 % areanya tersebar di Papua (Matanubun 2005). Selain sagu masih
banyak tanaman palma lain yang belum termanfaatkan secara optimal, seperti aren, sagu baruk, dan
Caryota mitis.
Pati hasil ekstraksi tanaman penghasilnya disebut pati alami. Pemanfaatan pati alami ini masih
sangat terbatas karena sifat fisik dan kimianya kurang sesuai untuk digunakan secara luas pada
produk-produk di industri. Oleh karena itu, pati alami akan lebih mudah diolah dalam upaya
menghasilkan produk dengan kualitas baik sehingga nilai ekonomisnya meningkat jika dimodifikasi
sifat-sifatnya melalui perlakuan fisik, kimia, atau kombinasi keduanya. Modifikasi pati bertujuan
mengubah sifat kimia atau fisik pati secara alami, yaitu dengan memotong struktur molekul,
menyusun kembali struktur molekul, oksidasi, serta substitusi gugus kimia pada molekul pati.
Modifikasi pati ini akan menghasilkan produk-produk turunan pati yang dapat diolah menjadi produk
pangan maupun non-pangan. Adapun modifikasi pati yang dilakukan pada penelitian ini adalah
hidrolisis secara enzimatis.
Pada umumnya enzim yang digunakan untuk menghasilkan hidrolisat pati di industri adalah α-
amilase. Akan tetapi, α-amilase biasanya digunakan sebagai enzim dengan hidrolisis secara sempurna
yang menghasilkan produk akhir glukosa sedangkan pada penelitian ini ingin diketahui kerja enzim
pada proses hidrolisis secara parsial serta pola hidrolisis dan perubahan komposisi gula yang terjadi
pada tingkat hidrolisis 10,40,dan 100% pada tiap pati palma yang digunakan untuk menghasilkan
beberapa produk hidrolisat pati, seperti maltosa, β-limit dekstrin, dan maltooligosakarida. Hidrolisis
parsial dapat dilakukan oleh beberapa enzim tertentu, seperti β-amilase dan pullulanase. Oleh karena
itu, pada penelitian ini digunakan kedua enzim tersebut.
Penelitian ini dilakukan untuk mengkaji hidrolisis secara enzimatis dengan pullulanase dan β-
amilase beberapa jenis pati palma di Indonesia yang berpotensi dan belum tereksplorasi untuk bahan
baku industri dengan melihat perubahan nilai Derajat Polimerisasi (DP), Dextrose Equivalent (DE),
dan tingkat hidrolisis. Pada proses hidrolisis, struktur pati yang terdiri atas fraksi amilosa maupun
amilopektin akan berubah karena rantai-rantai lurus ataupun yang bercabang akan terputus-putus
menjadi rantai yang lebih pendek yang mengindikasikan terbentuknya rantai-rantai yang mengandung
gula-gula sederhana penyusun pati. Perubahan ini perlu diamati berdasarkan peningkatan konsentrasi
gula pereduksi dan penurunan derajat polimerisasi. Produk berupa hidrolisat pati merupakan turunan
pati yang dapat dimanfaatkan menjadi berbagai produk tergantung nilai derajat polimerisasi, dextrose
equivalent, tingkat hidrolisis, dan jenis enzim yang digunakan untuk hidrolisis.
Penelitian hidrolisis enzimatis pati palma Indonesia sangat penting dilakukan untuk
mendapatkan beberapa informasi penting yang diperlukan dalam memanfaatkan lebih lanjut pati
palma tersebut. Karena produk hidrolisat pati palma diprediksikan berpotensi menjadi sumber
karbohidrat di masa depan. Oleh karena itu, masyarakat Indonesia tidak perlu takut kehabisan sumber

2
bahan pangan karena melimpahnya areal perkebunan tanaman palma di Indonesia dan berpotensinya
produk-produk turunan dari tanaman palma tersebut.
1.2 TUJUAN
Tujuan penelitian ini adalah untuk mengkaji hidrolisis beberapa jenis pati palma di Indonesia
secara enzimatis menggunakan pullulanase dan β-amilase.

3
II. TINJAUAN PUSTAKA
2.1 PATI
Pati merupakan senyawa yang memiliki berat molekul tinggi yang terdiri atas polimer glukosa
yang bercabang-cabang yang diikat dengan ikatan glukosidik. Pati termasuk salah satu jenis
polisakarida penting yang banyak terkandung pada beberapa tanaman yang tersebar di alam dan dapat
diekstrak dari sumbernya, seperti serealia (beras, gandum, jagung), umbi-umbian (ketela pohon, ubi
jalar, kentang), dan empulur batang palma (sagu, aren, sagu baruk). Pati yang terdapat pada sebagian
besar tanaman ini terdiri atas tiga fraksi penyusun, yaitu amilosa, amilopektin, dan bahan antara
seperti protein dan lemak.
Amilosa merupakan rantai lurus yang terdiri atas molekul-molekul glukosa yang berikatan
dengan α-1,4-D-glukosidik. Jumlah molekul glukosa pada rantai amilosa berkisar antara 250-350 unit.
Panjang rantai polimer akan mempengaruhi berat molekul amilosa dan panjang rantai polimer ini
sangat dipengaruhi oleh sumber pati. Derajat polimerisasi amilosa berkisar antara 500-6000 unit
glukosa tergantung sumber pati.
Struktur kimia amilopektin pada dasarnya sama seperti amilosa terdiri atas rantai pendek α-
(1,4)-D-glukosidik. Perbedaannya adalah amilopektin memiliki tingkat percabangan yang tinggi dan
memiliki bobot molekul yang lebih besar dengan adanya ikatan α-1,6-D-glukosidik dimana setiap
cabang mengandung 20-25 unit glukosa. Derajat polimerisasi amilopektin juga lebih tinggi
dibandingkan amilosa, yaitu antara 105 sampai 3x106 unit glukosa (Hustiany 2006).
Amilopektin mempunyai ukuran yang lebih besar daripada amilosa, tetapi tingkat
kekentalannya lebih rendah. Hal ini menunjukkan bahwa struktur molekul amilopektin lebih kompak
apabila terdapat dalam larutan. Begitu juga dengan kemampuan untuk membentuk kompleks lebih
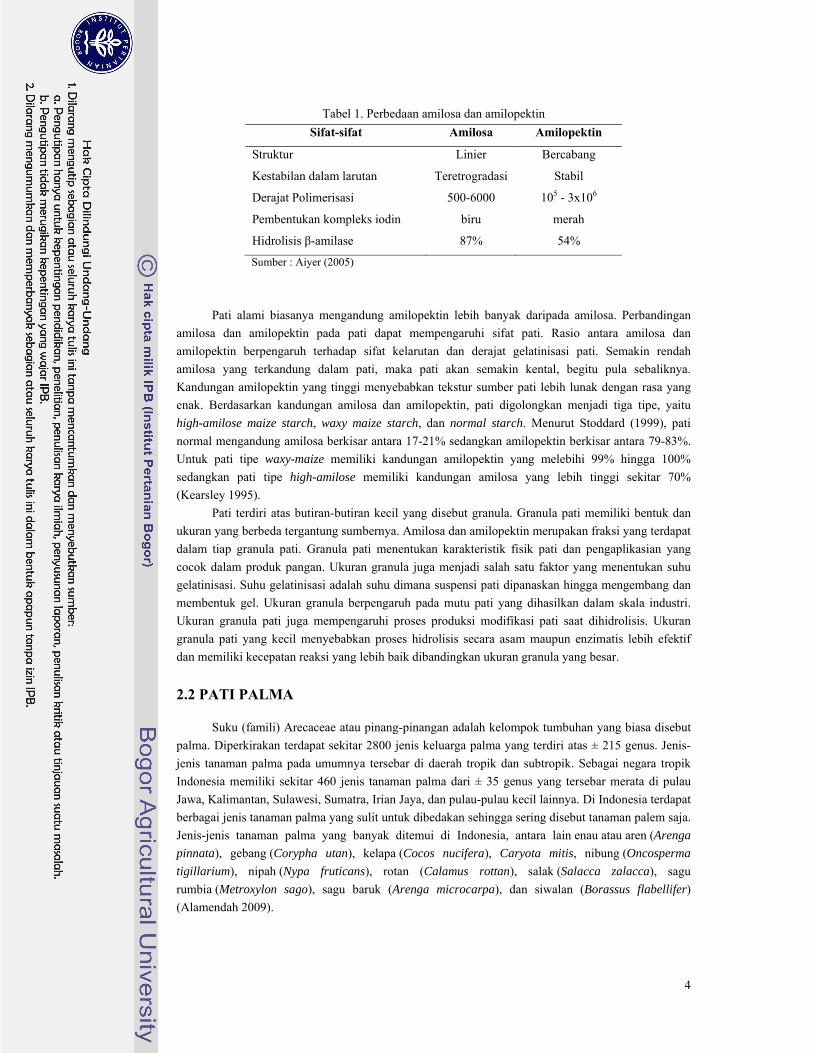
terbatas. Struktur kimia amilosa dan amilopektin dapat dilihat pada Gambar 1.
Gambar 1. Struktur kimia (a) amilosa (b) amilopektin
Fraksi amilosa dan amilopektin berada dalam granula pati. Kedua fraksi tersebut dapat
dipisahkan dalam air panas. Fraksi terlarut disebut amilosa karena banyak mengandung gugus
hidroksil dan membentuk lapisan transparan apabila dipanaskan dalam air sedangkan fraksi tidak larut
dan cenderung tidak terjadi retrogradasi disebut amilopektin. Perbedaan amilosa dan amilopektin
secara umum dapat dilihat pada Tabel 1.
a b
Ikatan α-1,6
Ikatan α-1,4
4
Tabel 1. Perbedaan amilosa dan amilopektin
Sifat-sifat Amilosa Amilopektin
Struktur Linier Bercabang
Kestabilan dalam larutan Teretrogradasi Stabil
Derajat Polimerisasi 500-6000 105 - 3x106
Pembentukan kompleks iodin biru merah
Hidrolisis β-amilase 87% 54%
Sumber : Aiyer (2005)
Pati alami biasanya mengandung amilopektin lebih banyak daripada amilosa. Perbandingan
amilosa dan amilopektin pada pati dapat mempengaruhi sifat pati. Rasio antara amilosa dan
amilopektin berpengaruh terhadap sifat kelarutan dan derajat gelatinisasi pati. Semakin rendah
amilosa yang terkandung dalam pati, maka pati akan semakin kental, begitu pula sebaliknya.
Kandungan amilopektin yang tinggi menyebabkan tekstur sumber pati lebih lunak dengan rasa yang
enak. Berdasarkan kandungan amilosa dan amilopektin, pati digolongkan menjadi tiga tipe, yaitu
high-amilose maize starch, waxy maize starch, dan normal starch. Menurut Stoddard (1999), pati
normal mengandung amilosa berkisar antara 17-21% sedangkan amilopektin berkisar antara 79-83%.
Untuk pati tipe waxy-maize memiliki kandungan amilopektin yang melebihi 99% hingga 100%
sedangkan pati tipe high-amilose memiliki kandungan amilosa yang lebih tinggi sekitar 70%
(Kearsley 1995).
Pati terdiri atas butiran-butiran kecil yang disebut granula. Granula pati memiliki bentuk dan
ukuran yang berbeda tergantung sumbernya. Amilosa dan amilopektin merupakan fraksi yang terdapat
dalam tiap granula pati. Granula pati menentukan karakteristik fisik pati dan pengaplikasian yang
cocok dalam produk pangan. Ukuran granula juga menjadi salah satu faktor yang menentukan suhu
gelatinisasi. Suhu gelatinisasi adalah suhu dimana suspensi pati dipanaskan hingga mengembang dan
membentuk gel. Ukuran granula berpengaruh pada mutu pati yang dihasilkan dalam skala industri.
Ukuran granula pati juga mempengaruhi proses produksi modifikasi pati saat dihidrolisis. Ukuran
granula pati yang kecil menyebabkan proses hidrolisis secara asam maupun enzimatis lebih efektif
dan memiliki kecepatan reaksi yang lebih baik dibandingkan ukuran granula yang besar.
2.2 PATI PALMA
Suku (famili) Arecaceae atau pinang-pinangan adalah kelompok tumbuhan yang biasa disebut
palma. Diperkirakan terdapat sekitar 2800 jenis keluarga palma yang terdiri atas ± 215 genus. Jenis-
jenis tanaman palma pada umumnya tersebar di daerah tropik dan subtropik. Sebagai negara tropik
Indonesia memiliki sekitar 460 jenis tanaman palma dari ± 35 genus yang tersebar merata di pulau
Jawa, Kalimantan, Sulawesi, Sumatra, Irian Jaya, dan pulau-pulau kecil lainnya. Di Indonesia terdapat
berbagai jenis tanaman palma yang sulit untuk dibedakan sehingga sering disebut tanaman palem saja.
Jenis-jenis tanaman palma yang banyak ditemui di Indonesia, antara lain enau atau aren (Arenga
pinnata), gebang (Corypha utan), kelapa (Cocos nucifera), Caryota mitis, nibung (Oncosperma
tigillarium), nipah (Nypa fruticans), rotan (Calamus rottan), salak (Salacca zalacca), sagu
rumbia (Metroxylon sago), sagu baruk (Arenga microcarpa), dan siwalan (Borassus flabellifer)
(Alamendah 2009).

5
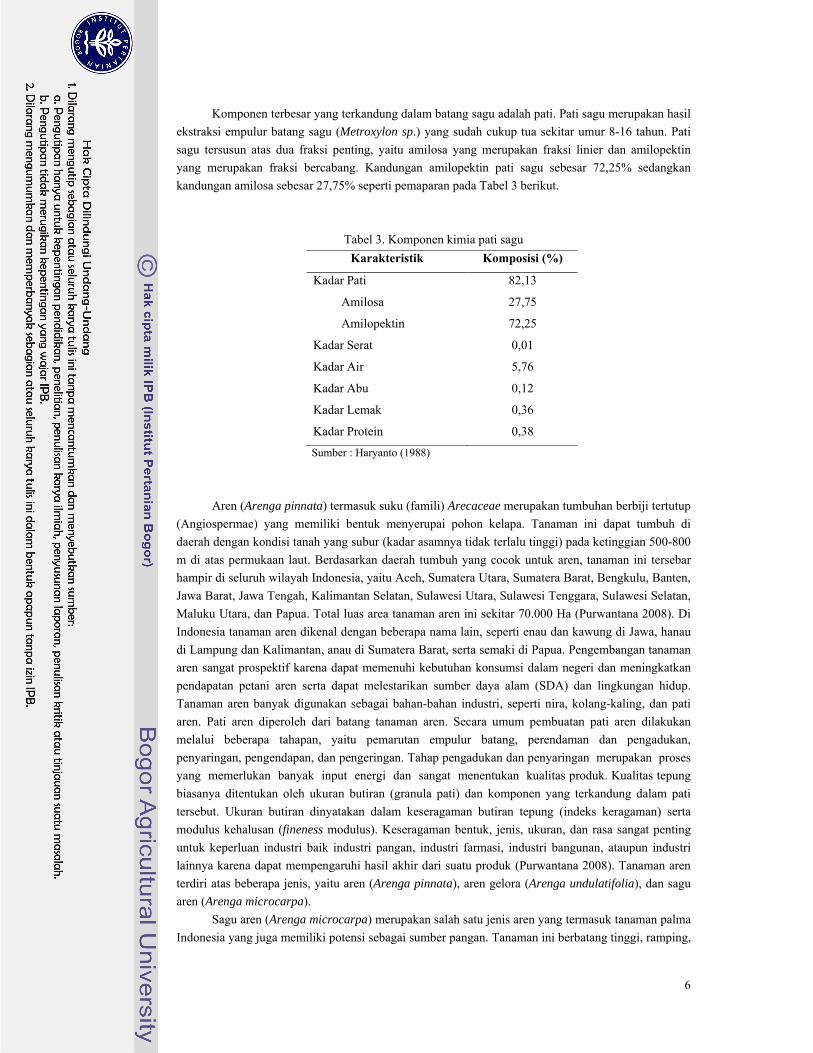
Sagu (Metroxylon sp.) banyak terdapat di Maluku dan Irian Jaya serta beberapa daerah lainnya
yang dapat dilihat pada Gambar 2. Area panen lahan sagu di Indonesia mencapai 1.250.000 Ha
dengan tingkat produktivitas 49.948 U/Ha dan produksi 5.803.447 Ton/tahun (BPS 2010). Tepung
sagu dari tanaman ini sudah sejak lama dijadikan makanan pokok oleh sebagian besar masyarakat di
Indonesia bagian Timur. Tanaman sagu di Indonesia dikenal dengan beberapa nama, seperti kirai di
Jawa Barat, bulung, kresula, bulu, atau resula di Jawa Tengah, lapia di Ambon, tumba di Gorontalo,
serta rambiam atau rabi di Aru. Tanaman ini memiliki banyak manfaat mulai dari bagian pelepah yang
dapat dipakai sebagai dinding atau pagar rumah, daunnya sebagai atap, kulit dan batangnya sebagai
kayu bakar, aci sagu, bubuk yang dihasilkan dengan mengekstraksi pati dari empulur atau batangnya
dapat diolah menjadi makanan dan pakan ternak, dan seratnya sebagai hardboard atau papan untuk
bangunan dan dapat diolah menjadi bahan bakar (bioetanol) (Haryanto 1992). Tanaman sagu termasuk
tanaman yang mampu menghasilkan pati kering sebanyak 25 ton per Ha. Satu batang pohon sagu
dapat menghasilkan 150-300 kg tepung sagu basah dengan komponen pada tepung sagu yang
tercantum pada Tabel 2.
Gambar 2. Daerah persebaran tanaman sagu di Indonesia (Matanubun 2005)
Tabel 2. Komposisi fisiko-kimia tepung sagu (per 100g)
Komponen Jumlah (%)
Air 13,69
Karbohidrat 84,89
Protein 0,46
Lemak 0,76
Serat 0,20 Abu 0,20
Sumber : Jading et al. (2011)
Sago area
6
Komponen terbesar yang terkandung dalam batang sagu adalah pati. Pati sagu merupakan hasil
ekstraksi empulur batang sagu (Metroxylon sp.) yang sudah cukup tua sekitar umur 8-16 tahun. Pati
sagu tersusun atas dua fraksi penting, yaitu amilosa yang merupakan fraksi linier dan amilopektin
yang merupakan fraksi bercabang. Kandungan amilopektin pati sagu sebesar 72,25% sedangkan
kandungan amilosa sebesar 27,75% seperti pemaparan pada Tabel 3 berikut.
Tabel 3. Komponen kimia pati sagu
Karakteristik Komposisi (%)
Kadar Pati 82,13
Amilosa 27,75
Amilopektin 72,25
Kadar Serat 0,01
Kadar Air 5,76
Kadar Abu 0,12
Kadar Lemak 0,36
Kadar Protein 0,38
Sumber : Haryanto (1988)
Aren (Arenga pinnata) termasuk suku (famili) Arecaceae merupakan tumbuhan berbiji tertutup
(Angiospermae) yang memiliki bentuk menyerupai pohon kelapa. Tanaman ini dapat tumbuh di
daerah dengan kondisi tanah yang subur (kadar asamnya tidak terlalu tinggi) pada ketinggian 500-800
m di atas permukaan laut. Berdasarkan daerah tumbuh yang cocok untuk aren, tanaman ini tersebar
hampir di seluruh wilayah Indonesia, yaitu Aceh, Sumatera Utara, Sumatera Barat, Bengkulu, Banten,
Jawa Barat, Jawa Tengah, Kalimantan Selatan, Sulawesi Utara, Sulawesi Tenggara, Sulawesi Selatan,
Maluku Utara, dan Papua. Total luas area tanaman aren ini sekitar 70.000 Ha (Purwantana 2008). Di
Indonesia tanaman aren dikenal dengan beberapa nama lain, seperti enau dan kawung di Jawa, hanau
di Lampung dan Kalimantan, anau di Sumatera Barat, serta semaki di Papua. Pengembangan tanaman
aren sangat prospektif karena dapat memenuhi kebutuhan konsumsi dalam negeri dan meningkatkan
pendapatan petani aren serta dapat melestarikan sumber daya alam (SDA) dan lingkungan hidup.
Tanaman aren banyak digunakan sebagai bahan-bahan industri, seperti nira, kolang-kaling, dan pati
aren. Pati aren diperoleh dari batang tanaman aren. Secara umum pembuatan pati aren dilakukan
melalui beberapa tahapan, yaitu pemarutan empulur batang, perendaman dan pengadukan,
penyaringan, pengendapan, dan pengeringan. Tahap pengadukan dan penyaringan merupakan proses
yang memerlukan banyak input energi dan sangat menentukan kualitas produk. Kualitas tepung
biasanya ditentukan oleh ukuran butiran (granula pati) dan komponen yang terkandung dalam pati
tersebut. Ukuran butiran dinyatakan dalam keseragaman butiran tepung (indeks keragaman) serta
modulus kehalusan (fineness modulus). Keseragaman bentuk, jenis, ukuran, dan rasa sangat penting
untuk keperluan industri baik industri pangan, industri farmasi, industri bangunan, ataupun industri
lainnya karena dapat mempengaruhi hasil akhir dari suatu produk (Purwantana 2008). Tanaman aren
terdiri atas beberapa jenis, yaitu aren (Arenga pinnata), aren gelora (Arenga undulatifolia), dan sagu
aren (Arenga microcarpa).
Sagu aren (Arenga microcarpa) merupakan salah satu jenis aren yang termasuk tanaman palma
Indonesia yang juga memiliki potensi sebagai sumber pangan. Tanaman ini berbatang tinggi, ramping,
7
dan berumpun banyak. Sagu aren tumbuh liar di hutan-hutan Maluku dan Irian Jaya. Sagu aren juga
banyak terdapat di Kepulauan Sangihe dan Taulud, Sulawesi Utara yang dikenal dengan nama sagu
baruk. Sagu baruk sudah lama dimanfaatkan oleh penduduk di wilayah tersebut sebagai makanan
pokok yang merupakan alternatif substitusi beras maupun sagu (Sunanto 1993).
Sagu baruk tergolong dalam famili Palmae dimana batang tanaman ini dapat menghasilkan
pati. Sagu baruk memiliki perbedaan dengan sagu sejati (Metroxylon sp.), yaitu hanya tumbuh di
lahan kering dengan iklim kering dan basah tidak sama dengan sagu sejati yang tumbuh di daerah
rawa. Sagu baruk memiliki struktur batang berbentuk silinder dan soliter berfungsi sebagai penyimpan
makanan cadangan dalam bentuk karbohidrat. Diameter batang beragam antara 15-20 cm tergantung
pada kondisi kesuburan tanah. Tinggi batang dapat mencapai 6-15 m. Daun berwarna hijau tua
mengkilat dan berbentuk pelepah yang tersusun dari 50-60 daun (leaflet). Bunga sagu baruk mirip
dengan bunga tanaman aren (Arenga pinnata). Bunga tanaman sagu baruk tersusun dalam satu
rangkaian bunga (inflorescensia) dimana bunga pertama muncul pada bagian pucuk (terminalis),
sedangkan bunga kedua muncul pada ketiak daun di bawah bunga pertama demikian seterusnya
sampai kurang lebih 6 rangkaian bunga. Umur berbunga antara 8–15 tahun tergantung kesuburan
tanah. Tanaman sagu baruk memperbanyak diri dengan tunas. Tanaman sagu baruk sebagai penghasil
pati (karbohidrat) mampu menghasilkan tepung sagu basah antara 25 sampai 30 kg/batang. Tepung
sagu baruk dapat dimanfaatkan untuk berbagai panganan, seperti papeda, bagea, dan aneka kue kering
lainnya (Miftahorachman 2009). Dilihat dari nilai gizi tepung sagu baruk asal Kepulauan Sangihe,
sagu baruk mengandung air, karbohidrat, pati, serat, dan sebagainya yang dipaparkan pada Tabel 4.
Tabel 4. Nilai gizi tepung sagu baruk basah asal Kepulauan Sangihe
Kandungan gizi Komposisi (%)
Air 12,54
Karbohidrat 56,11
Abu 0,32
Lemak 0,33
Serat Kasar 0,12
Kalsium 0,014
Sumber : Miftahorachman (2009)
Caryota mitis (fish-tail palm) merupakan tanaman palma yang memiliki tinggi pohon 5-12 m
dengan diameter batang 5-15 cm. Tanaman ini menyebar di beberapa daerah tropis di Indonesia,
seperti Sumatera (Aceh dan Sumatera Utara), Jawa, dan Kalimantan. Caryota mitis memiliki beberapa
nama lain di beberapa daerah di Indonesia, seperti genduru di Jawa Tengah, saray di Jawa Barat, dan
bulung talang di Kalimantan. Tanaman ini memiliki kandungan pati pada bagian batangnya dan
sukrosa pada air bunganya, yaitu sebesar 83,5%. Dengan memanfaatkan bunganya Caryota mitis
dapat dikelola sebagai tanaman perkebunan, seperti kelapa sawit, yang dapat dipanen terus-menerus
selama waktu reproduktif tanaman tersebut. Air bunga (nira) pada Caryota mitis dapat digunakan
sebagai sumber gula alternatif pengganti tebu. Proses untuk mendapatkan sukrosa murni dari air
bunga pohon tersebut dapat dilakukan melalui proses ekstraksi air bunga, pengendapan kotoran,
pemurnian air gula dan pemisahan dari kandungan senyawa lainnya, kristalisasi, dan penyimpanan
untuk selanjutnya diproses menjadi kristal gula murni.

8
2.3 HIDROLISIS PATI
Sebagian besar pati yang diperoleh langsung dari tanaman penghasilnya merupakan pati alami.
Namun sebagian besar penggunaan pati di industri berupa hidrolisat pati karena memiliki karakteristik
dan sifat yang mudah dikontrol dalam pembuatan produk-produk tertentu dengan kualitas yang baik.
Salah satu alasannya karena hidrolisat pati merupakan pati alami yang telah mengalami modifikasi
seperti penghilangan komponen-komponen minor pada pati sehingga hanya tersisa kandungan
amilosa dan amilopektin yang lebih mudah diolah menjadi beberapa produk turunan pati yang dapat
diaplikasikan di industri. Prinsip hidrolisis pati adalah pemutusan rantai polimer pati menjadi unit-unit
dekstrosa. Pada umumnya hidrolisat pati dikarakterisasi berdasarkan tingkat derajat hidrolisisnya yang
dinyatakan dengan nilai DE (dextrose equivalent) yang menunjukan persentase kandungan dekstrosa
murni dalam basis bobot kering pada produk hidrolisis (Kearsley 1995). Dextrose Equivalent (DE)
merupakan parameter kemurnian sirup glukosa atau maltosa yang didefinisikan sebagai persentase
perbandingan antar gula pereduksi dengan bobot kering sirup glukosa atau maltosa. Jika nilai DE
sebesar 100%, maka dapat diartikan seluruh bahan kering pada sirup glukosa merupakan gula
pereduksi. Gula pereduksi merupakan golongan gula yang dapat mereduksi senyawa-senyawa
penerima elektron. Pada umumnya gula pereduksi mengandung gugus aldehida atau keton bebas.
Contoh dari gula pereduksi, antara lain semua monosakarida (glukosa, fruktosa, dan galaktosa) dan
disakarida (laktosa dan maltosa). Tinggi rendahnya gula pereduksi suatu produk dipengaruhi oleh
sumber pati dan aktivitas enzim pada tiap komoditas sumbernya. Hubungan antara aktivitas enzim dan
gula pereduksi yang dihasilkan menunjukkan bahwa semakin tinggi aktivitas enzim, maka semakin
tinggi juga gula pereduksi yang dihasilkan (Lehninger 1982). Nilai DE berhubungan dengan derajat
polimerisasi (DP). Nilai DP menyatakan jumlah unit monomer dalam satu molekul. Hasil hidrolisis
pati menjadi produk hidrolisat pati dapat diketahui dari nilai DE yang dihasilkan. Adapun beberapa
contoh produk hidrolisat pati berdasarkan nilai DE yang dihasilkan beserta aplikasinya dapat dilihat
pada Tabel 5.
Tabel 5. Aplikasi produk hidrolisat pati secara umum
Produk Hidrolisat Pati DE Aplikasi
Sirup Maltosa 48-63 Pengeras permen, mencegah sifat higroskopis pada bahan, media fermentasi
Sirup Glukosa 96-98 Soft drink, media untuk fermentasi
Sirup Fruktosa - Produk susu, soft drink, industri makanan dan minuman kaleng
Sirup Campuran 42-63 Soft drink, bahan baku industri campuran
Maltodekstrin 3-20 Stabilizer, filler, lem, dan pasta
Sumber : Kennedy et al. (1995)
2.4 ENZIM (β-AMILASE DAN PULLULANASE)
Enzim adalah suatu protein yang bertindak sebagai katalisator reaksi biologis atau disebut
biokatalisator. Enzim berfungsi mengatur kecepatan dan kekhususan reaksi kimia yang berlangsung di
dalam sel. Walaupun enzim dibuat di dalam sel, tetapi untuk bertindak sebagai katalis tidak harus
berada di dalam sel. Reaksi yang dikendalikan oleh enzim, antara lain respirasi, pertumbuhan dan
perkembangan, serta fotosintesis. Enzim sebagai katalis memiliki nilai ekonomis tinggi karena sangat

9
diperlukan untuk menunjang berbagai proses industri, misalnya industri pangan. Degradasi pati
membutuhkan enzim amilase yang akan memecah atau menghidrolisis menjadi polisakarida yang
lebih pendek (dekstrin) lalu menjadi maltosa. Hidrolisis akhir maltosa menghasilkan glukosa terlarut
yang dapat ditransport masuk ke dalam sel. Amilase merupakan enzim pendegradasi pati yang dapat
dikelompokkan menjadi tiga golongan enzim, yaitu α-amilase, β-amilase, dan glukoamilase.
β-amilase (E.C 3.2.1.2) merupakan enzim golongan hidrolase yang digunakan dalam
proses sakarifikasi pati (sejenis karbohidrat). Sakarifikasi banyak berperan dalam pemecahan
makromolekul karbohidrat. Pemecahan makromolekul karbohidrat ini akan menghasilkan molekul
karbohidrat rantai pendek (sederhana). Enzim β-amilase disebut juga α-l,4-glukan maltohidrolase E.C.
3.2.1.2. karena bekerja pada ikatan α-1,4-glikosidik dengan menginversi konfigurasi posisi atom C
nomor 1 molekul glukosa dari α menjadi β. Enzim ini memutus ikatan amilosa maupun amilopektin
dari luar molekul dan menghasilkan maltosa dari ujung non-pereduksi pada rantai polisakarida. Pada
ikatan α-1,6 glikosidik aktivitas enzim ini akan berhenti (Sadikin 2002). Pola pemutusan yang
dilakukan enzim ini seperti Gambar 3.
Gambar 3. Titik pemutusan rantai amilosa pada pati oleh β-amilase (Thomas 1999)
Enzim β-amilase banyak ditemukan pada tanaman tingkat tinggi, seperti gandum, barley,
kentang, ubi, dan kacang kedelai. Di samping itu, β-amilase juga dapat ditemui pada
beberapa mikroorganisme, antara lain Pseudomonas, Bacillus, Streptococcus, dan Clostridium
thermosulfurigenes. Enzim yang berasal dari C. thermosulfurigenes umumnya lebih disukai karena
memiliki toleransi suhu dan pH yang lebih tinggi. Mekanisme kerja dari enzim β-amilase akan
memotong ikatan glikosidik pada gugus amilosa, amilopektin, dan glikogen. Amilosa merupakan
struktur rantai lurus dari pati, sedangkan amilopektin merupakan struktur percabangan dari pati. Hasil
pemotongan oleh enzim ini akan didominasi oleh molekul maltosa dan β-limit dekstrin seperti terlihat
pada Gambar 4. Dalam industri pangan, pembentukan senyawa β-limit dektrin seringkali dihindari
karena membentuk viskositas atau kekentalan yang terlalu pekat. Produk samping ini tetap dapat
digunakan sebagai bahan pengental dan tambahan pada produk-produk diet rendah kalori.
Gambar 4. Pembentukan β-limit dekstrin (Tester 2011)
β-Amilase
β-limit dekstrin
Maltosa

10
Menurut Tester (2011), β-limit dektrin merupakan produk samping yang dihasilkan dari
hidrolisis pati yang tidak sempurna oleh β-amilase yang memiliki sifat bioadesif yang cocok
digunakan sebagai perekat alami. Ukuran dan bentuk β-limit dektrin tiap pati berbeda dimana
senyawa ini dihasilkan dari 40—60% berat molekul amilopektin. Pada pati waxy maize berat molekul
β-limit dektrin mencapai 3,4 x 106 dan 3,0 x 106 pada normal waxy maize. Meskipun memiliki bobot
molekul yang cukup tinggi β-limit dektrin memiliki tekanan osmosis yang rendah, viskostas yang
tinggi, dan kemampuan dispersi kelarutan yang tinggi pada larutan. β-limit dektrin juga merupakan
produk yang free sugar sehingga cocok untuk diaplikasikan pada berbagai produk diet dan pengontrol
diabetik.
Amilosa merupakan komponen linier pada pati sehingga dapat terhidrolisis secara sempurna,
tetapi hanya 55% dari amilopektin yang merupakan percabangan yang dapat dikonversi menjadi β-
maltosa sedangkan 45% lainnya adalah limit dekstrin dengan bobot molekul yang tinggi yang
keseluruhannya termasuk bagian percabangan dari amilopektin awal. β-amilase merupakan enzim
yang memiliki pemecahan tipe ganda. Enzim tersebut memecah secara berulang pada rantai substrat
yang tersedia beberapa kali. Untuk amilosa dengan berat molekul rendah memiliki jumlah rata-rata
pemecahan sebanyak 4 sedangkan amilosa dengan berat molekul tinggi memiliki jumlah pemecahan
yang lebih banyak. Enzim ini digunakan untuk memproduksi maltosa dengan tingkat kemurnian yang
tinggi dari pati (Kainuma 1995).
Reaksi yang terjadi adalah :
Pati + n H2O n β-maltosa + β-limit dekstrin
Menurut Hii (2012), enzim perombak pullulan sebagai enzim debranching terdiri atas lima
kelompok yang dikelompokkan berdasarkan spesifikasi substrat dan produknya, yaitu :
1. Pullulan hidrolase tipe I (EC 3.2.1.135), yaitu enzim yang menyerang ikatan α-1,4 glukosidik
dalam pullulan membentuk panosa.
2. Pullulan hidrolase tipe II (EC 3.2.1.57), yaitu enzim yang menyerang ikatan α-1,4 glukosidik
dalam pullulan membentuk isopanosa.
3. Pullulanase tipe I (EC 3.2.1.41), yaitu enzim yang spesifik menghidrolisis ikatan α-1,6
glukosidik dalam pullulan membentuk maltotriosa. Enzim ini juga menyerang ikatan α-1,6
glukosidik pada amilopektin membentuk pati tinggi amilosa atau Short-Chain Amylose
(SCA).
4. Pullulanase tipe II, yaitu enzim yang menyerang ikatan α-1,6 dan α-1,4 glukosdidik pada
polisakarida. Enzim ini disebut juga amilopullulanase.
5. Glukoamilase (EC 3.2.1.3), yaitu enzim yang menghidrolisis pullulan dan secara berurutan
dari ujung gugus non-pereduksi sehingga menghasilkan glukosa.
Enzim pullulanase (EC 3.2.1.41) merupakan salah satu jenis enzim yang tergolong kelompok
enzim debranching yang memiliki aktivitas pada titik percabangan pati pada amilopektin dengan
memecah ikatan α-1,6 glikosidik dengan spesifitas substratnya. Gambar 5 menunjukkan titik
percabangan pemutusan yang dilakukan oleh enzim ini. Pullulanase (EC 3.2.1.41, pullulan 6-
glucanohydrolase) merupakan enzim yang dapat digunakan pada hidrolisis pati agar dapat stabil.
Pullulanase dapat memecahkan molekul pullulan dan memiliki aktivitas pada amilopektin dan limit
dekstrin, tetapi terdapat kesulitan untuk memecahkan glikogen. Hal ini yang membedakan pullulanase
dengan enzim debranching lainnya, seperti isoamilase. Enzim pullulanase dapat menghidrolisis pati
menjadi SCA atau maltooligosakarida pada proses hidrolisis enzimatis.
Pullulan termasuk polisakarida ekstraseluler yang diproduksi oleh Aureobasidium pullulans
atau Pullularia pullulans. Pullulanase dapat dihasilkan dari Klebsiella pneumoniae atau Aerobacter

11
aerogenes, Escherichia intermedia, dan Streptococcus mitis. Pullulanase yang berasal dari Klebsiella
pneumoniae merupakan enzim yang sangat bermanfaat untuk mempelajari struktur pati dan glikogen
yang terdiri atas ikatan α-1,4 dan α-1,6 glikosidik. Polisakarida yang dipecah oleh enzim ini akan
menghasilkan rantai-rantai lurus dan lebih pendek.
Reaksi :
Pullulan + n H2O n Maltotriosa
Pati (Glikogen) + n H2O n Maltooligosakarida
Gambar 5. Titik pemutusan percabangan rantai amilopektin oleh pullulanase (Thomas 1999).
Hidrolisis pati dapat dilakukan dengan menggunakan asam ataupun enzim. Proses ini akan
menghasilkan glukosa sebagai produk akhir. Hidrolisis secara parsial dapat dilakukan dengan
menggunakan enzim tertentu yang dapat memotong rantai karbon pada pati. Beberapa contoh produk
hidrolisat pati yang dapat dihasilkan antara lain sirup glukosa, maltosa, maltodekstrin, dan
maltooligosakarida. Enzim yang digunakan adalah enzim-enzim hidrolase yang termasuk kelompok
enzim yang sangat penting dalam pengolahan pangan. Gula yang dihasilkan melalui pemecahan oleh
enzim hidrolase ini disebut “gula pati”. Adapun beberapa contoh hidrolisat pati enzimatis dapat dilihat
pada Gambar 6 berikut.
Gambar 6. Produk-produk hidrolisat pati enzimatis (Kainuma 1995)
Pullulanase
2
4,5
1, 3
1
6
Keterangan :
1 = α-amilase
2 = β-amilase
3 = Glukosamilase
4 = Pullulanase
5 = Isoamilase
6 = Hidrogenasi
PATI
Maltosa
Glukosa
Maltodekstrin
β-Limit dekstrin
Maltooligosakarida
Maltitol

12
Dari Gambar 6 dapat diketahui bahwa tiap enzim hidrolase memiliki sifat yang spesifik dalam
menghasilkan produk hidrolisat. Adapun produk hidrolisat pati yang dihasilkan oleh beberapa enzim
serta aplikasinya di industri dapat dilihat pada Tabel 6.
Tabel 6. Produk hidrolisat enzimatis dan aplikasinya
Produk Hidrolisat Pati Enzim Aplikasi
Maltosa β-amilase Pengeras permen, mencegah sifat higroskopis pada bahan, media fermentasi, pembuat permen jelly, bahan pengental saus, pudding, dan digunakan pada industri tekstil dan kertas. a
β-Limit Dekstrin β-amilase Stabillizer, pengental rendah kalori.b
Maltooligosakarida (MO’s) Pullulanase, isoamilase, dan
amilase
Pengemulsi kosmetika, makanan berkalori rendah, agen enkapsulasi.c
Sumber : a) Said (2008) b) Poliana (2007)
c) Patel (2010)

13
III. METODOLOGI
3.1 WAKTU DAN TEMPAT PENELITIAN
Penelitian dilakukan selama delapan bulan, yaitu sejak akhir bulan Februari hingga September 2012 di Laboratorium Teknologi Bioindustri dan Laboratorium Teknik Kimia Departemen Teknologi Industri Pertanian IPB.
3.2 ALAT DAN BAHAN
3.2.1 Alat Peralatan yang digunakan dalam penelitian ini adalah alat-alat untuk pengujian, seperti
spektrofotometer Hach dan untuk analisa, seperti erlenmeyer, termometer, 3G-filter glass, penyaring
vakum, pH-meter Beckman, inkubator waterbath, neraca analitik, gelas piala, alat pengaduk,
penangas air, tabung reaksi, sentrifuse, tabung ulir, dan mikropipet.
3.2.2 Bahan Bahan-bahan utama yang digunakan dalam penelitian ini adalah tujuh pati palma Indonesia
yang berasal dari Balai Penelitian Tanaman Kelapa dan Palma Lain, Manado, meliputi pati sagu
rumbia (Metroxylon sp.), sagu komersial, Caryota mitis, Arenga pinnata, Arenga microcarpa 1 (sagu
baruk 1), Arenga microcarpa 2 (sagu baruk 2), dan Arenga microcarpa komersial (sagu baruk 3)
tersaji pada Lampiran 1. Enzim yang digunakan dalam penelitian ini adalah β-amilase dari barley
(SIGMA) dan pullulanase (A.aerogenes, Hayashibara Biochemical Lab.). Bahan-bahan kimia yang
digunakan untuk pemurnian pati (deproteinase dan defatting starch), antara lain : Dimetil Sulfoksida
(DMSO), metanol, eter, dan larutan NaOH 0,1 N. Bahan-bahan kimia lain yang digunakan adalah
untuk analisa total gula, seperti larutan fenol dan H2SO4 97% dan analisa gula pereduksi, seperti
Na2CO3, NaHCO3, KCN, potasium ferrisianida (K3Fe(CN)6), dan ferric amonium sulfat
((NH4)2Fe(SO4)2.6H2O).
3.3 METODE PENELITIAN
3.2.1 Persiapan Bahan 1.a. Penentuan Kadar Amilosa
Penentuan kadar amilosa dilakukan dengan metode IRRI yang menggunakan prinsip
pengikatan iodin (iodine binding) berdasarkan reaksi antara amilosa dengan senyawa iod yang
prosedur pengujiannya tersaji pada Lampiran 2.
1.b Penghilangan Protein pada Pati (Sunarti et al. 2001)
Deproteinisasi pati dilakukan dengan menggunakan metode denaturasi protein. Sebanyak 10 g
pati ditambahkan dengan 15-20 ml NaOH 0,1 N sampai semua bagian pati terendam. Pati yang telah
dicampur NaOH dikocok hingga terlarut homogen. Campuran yang terbentuk dipisahkan dengan alat
sentrifus. Selanjutnya pati yang terpisah dicuci dengan air lalu disentrifugasi kembali untuk
memisahkan pati dari air pencuci. Pemisahan pati dengan alat sentrifus ini dilakukan berulang
sebanyak tiga kali. Pati akhir yang diperoleh dinetralkan dengan akuades sebelum disaring dengan

14
3G-3 filter glass. Pati yang sudah ber-pH netral disaring lalu dibilas dengan metanol. Pati hasil
penyaringan dikering-anginkan hingga kering.
1.c. Penghilangan Lemak pada Pati (Sunarti et al. 2001)
Sebanyak 5 g pati yang telah dideproteinisasi dilarutkan ke dalam 100 ml DMSO (Dimethyl
sulfoxide) lalu dikocok dengan shaker pada suhu 37oC selama semalam. Larutan tersebut kemudian
dituang secara perlahan-lahan dalam 100 ml metanol lalu didiamkan semalam pada suhu 4oC.
Endapan yang terbentuk seperti butiran pati disaring dengan menggunakan 3G-3 filter glass lalu
dibilas dengan metanol dan eter di akhir. Apabila pada larutan sebelum disaring belum terdapat
endapan seperti butiran pati, ditambahkan terus metanol dan diaduk-aduk. Selanjutnya pati yang
dihasilkan dikering-anginkan.
3.3.2 Penentuan Aktivitas Enzim 2.a. Uji Aktivitas Pullulanase (Sunarti et al. 2001)
Sebanyak 3 ml larutan glutinous rice-defatted starch 0,4% ditambahkan dengan 1,5 ml buffer
fosfat 0,1 M pada pH optimal (pH 6,0) lalu diinkubasi pada suhu optimal (40oC). Kemudian larutan
ditambahkan 1,5 ml pullulanase dan dihidrolisis selama 3 jam. Selama proses dilakukan pengambilan
sampel setiap 15 menit. Tiap sampel diinaktivasi pada air mendidih selama 10 menit lalu diukur
konsentrasi gula pereduksinya. Selisih kenaikan konsentrasi gula pereduksi terbesar ditetapkan
sebagai nilai aktivitas optimum enzim.
2.b. Uji Aktivitas β-amilase (Sunarti et al. 2001)
Larutan soluble starch 2% sebanyak 5 ml ditambahkan dengan 3,75 ml buffer asetat 0,2 M
pada pH optimal enzim (pH 4,8) dan 1,25 ml akuades yang kemudian diinkubasi pada suhu optimal
enzim (40oC). Setelah itu, ditambahkan 5 ml β-amilase yang kemudian dihidrolisis selama 3 jam.
Selama proses dilakukan pengambilan sampel setiap 15 menit kemudian tiap sampel diinaktivasi pada
air mendidih selama 10 menit lalu diukur konsentrasi gula pereduksinya. Selisih kenaikan konsentrasi
gula pereduksi terbesar ditetapkan sebagai nilai aktivitas optimum enzim. Satu unit didefinisikan
sebagai jumlah enzim yang dibutuhkan untuk memproduksi 1 µmol gula pereduksi dalam bentuk
glukosa per menit pada kondisi yang telah ditentukan.
Adapun rumus untuk menghitung aktivitas masing-masing enzim sebagai berikut.
3.3.3 Hidrolisis Pati Palma oleh Pullulanase dan β-amilase 3.a. Persiapan Larutan Pati (Pati Tergelatinisasi 0,4%) (Sunarti et al. 2001)
Persiapan pati untuk tahap hidrolisis menggunakan metode gelatinisasi pati 0,4%. Pati yang
telah dihilangkan lemaknya (defatted starch) sebanyak 30 mg dilarutkan dalam 0,5 ml akuades dan
0,75 ml NaOH 1 N lalu didiamkan dengan inkubasi pada es (4oC) hingga pati tergelatinisasi. Pati yang
tergelatinisasi ditambahkan secara perlahan-lahan dengan akuades sebanyak 5,35 ml kemudian
dinetralkan oleh 0,75 ml 1 M HCl dan ditambahkan dengan 0,15 ml NaN3 3%.

15
3.b. Hidrolisis Pati Tergelatinisasi oleh Pullulanase (Sunarti et al. 2001) Sebanyak 22,5 ml pati yang telah tergelatinisasi 0,4% ditambahkan dengan 22,5 ml buffer
fosfat 0,1 M yang mengandung 0,45 U/g pullulanase kemudian diinkubasi selama 48 jam. Secara
periodik dilakukan pengambilan sampel. Setelah jam ke-48 dilakukan hidrolisis selama semalam. Tiap
sampel yang disampling tiap waktunya dianalisa total gula dengan metode Fenol-Sulfat dan gula
pereduksi dengan metode Park Johnson dengan prosedur pengujian yang tersaji pada Lampiran 3.
Setelah dianalisa masing-masing hidrolisat pati palma dihitung nilai DE, DP, dan tingkat hidrolisis
dengan rumus perhitungan yang tersaji pada Lampiran 4.
3.c. Hidrolisis Pati Tergelatinisasi oleh β-amilase dan Pullulanase secara suksesif (Sunarti et al.
2001)
Sebanyak 67,5 ml pati yang telah tergelatinisasi 0,4% ditambahkan dengan 67,5 ml buffer
asetat 0,2 M yang mengandung 2 U/g β-amilase lalu diinkubasi selama 48 jam. Secara periodik
dilakukan pengambilan sampel. Pada jam ke-24 dilakukan penambahan β-amilase berlebih dengan
konsentrasi 20 U/g kemudian hidrolisis dilanjutkan sampai jam ke-36. Pada jam ke-36 dilakukan
penambahan pullulanase 2 U/g dan dihidrolisis kembali sampai jam ke-48 kemudian sisa larutan
dihidrolisis selama semalam. Tiap sampel yang disampling tiap waktunya dianalisa total gula dengan
metode Fenol-Sulfat dan gula pereduksi dengan metode Park Johnson dengan prosedur pengujian
yang tersaji pada Lampiran 3. Setelah dianalisa masing-masing hidrolisat pati palma dihitung nilai β-
limit dekstrin, DE, DP, dan tingkat hidrolisis dengan rumus perhitungan yang tersaji pada Lampiran 4.
16
IV. HASIL DAN PEMBAHASAN
4.1 KANDUNGAN AMILOSA PADA PATI PALMA
Pati adalah karbohidrat yang merupakan polimer glukosa yang terdiri atas dua fraksi, yaitu
amilosa dan amilopektin. Selain kedua fraksi tersebut terdapat juga bahan antara yang menyusun pati,
yaitu protein dan lemak. Komponen protein dan lemak serta bahan antara lainnya ini terdapat
berkisar 5-10% (Banks et al. 1975).
Pati yang digunakan terdiri atas tujuh pati palma yang berasal dari Balai Tanaman Kelapa dan
Palma Lain, Manado, yaitu pati sagu rumbia (Metroxylon sp.), sagu komersial (Metroxylon sagoo),
Caryota mitis, aren (Arenga pinnata), serta sagu baruk 1,2, dan 3 (Arenga microcarpa). Untuk pati
sagu baruk dibedakan menjadi tiga kode karena adanya perbedaan sumber pohon sagu baruk, tekstur
dan penampakan fisik, serta penggunaannya. Untuk sagu baruk 1 memiliki tekstur agak halus dengan
warna putih bersih, sagu baruk 2 memiliki tekstur yang lebih kasar dan berwana kecoklatan,
sedangkan sagu baruk 3 merupakan pati yang sudah diperdagangkan dengan tekstur yang paling halus
yang berwarna putih. Oleh karena itu, nilai derajat polimerisasi awal untuk masing-masing pati palma
akan menghasilkan nilai yang berbeda-beda.
Pada tahap awal penelitian dilakukan penentuan kadar amilosa pati palma yang digunakan.
Ketujuh pati palma dikarakterisasi kandungan amilosa dan amilopektinnya untuk mengetahui tipe
pati. Adapun nilai amilosa dan amilopektin masing-masing pati palma disajikan pada Tabel 7.
Berdasarkan data tersebut pati palma yang digunakan memiliki kandungan amilosa berkisar antara
20,1-26,7% dengan kandungan amilopektin antara 73,3-79,9%. Hasil ini menunjukkan bahwa ketujuh
pati palma memiliki rasio amilosa dan amilopektin 15-25 : 75-85 artinya pati palma yang digunakan
termasuk tipe pati normal.
Tabel 7. Rasio amilosa dan amilopektin pada pati palma
Jenis Pati Amilosa (%) Amilopektin (%)
Sagu baruk 1 23,2 76,8
Sagu baruk 2 25,8 74,2
Sagu baruk 3 23,7 76,3
Sagu rumbia 26,7 73,3
Sagu komersial 21,8 78,2
Aren 20,1 79,9
Caryota mitis 20,6 79,4
Keragaman jumlah amilosa dan amilopektin yang menyusun tiap pati akan mempengaruhi
kerja enzim hidrolase yang digunakan. Semakin tinggi kandungan amilosanya, maka rantai lurus yang
dimiliki pati semakin banyak sehingga lebih memudahkan enzim hidrolase, seperti β-amilase dalam
menghidrolisis substrat (pati). Berdasarkan data tersebut pati sagu rumbia memiliki kandungan

17
amilosa paling tinggi, yaitu 26,67%. Oleh karena itu, pati tersebut akan lebih mudah dihidrolisis
dibandingkan pati palma lainnya. Sebaliknya, pati aren dan Caryota mitis memiliki kandungan
amilosa paling rendah, yaitu 20,04% sehingga akan lebih lambat dihidrolisis terutama oleh enzim β-
amilase. Hal ini didukung oleh pendapat Said (2008) yang mengatakan bahwa laju hidrolisis oleh
enzim amilase akan lebih cepat terjadi pada rantai lurus (amilosa) jika dibandingkan pada rantai
bercabang (amilopektin) sehingga semakin banyak rantai lurus pada suatu polisakarida, maka semakin
cepat laju hidrolisisnya.
Pada proses hidrolisis, enzim yang digunakan bersifat spesifik dan hanya memutus ikatan-
ikatan pada rantai amilosa dan amilopektin. Oleh karena itu, perlu dilakukan persiapan pati sebelum
dihidrolisis berupa proses pemurnian pati. Pemurnian pati bertujuan menghilangkan sejumlah
komponen minor pada pati, seperti lemak, protein, dan air untuk mempermudah proses hidrolisis
karena enzim dapat langsung memutus rantai glukosidik pada amilosa dan amilopektin. Pemurnian
pati terdiri atas dua tahapan, yaitu penghilangan komponen protein dan lemak.
Penghilangan komponen protein atau disebut deproteinisasi menggunakan metode denaturasi.
Protein yang terkandung dalam suatu bahan dapat terdenaturasi pada suhu ataupun pH tinggi. Oleh
karena itu, pada penelitian ini penghilangan protein pada pati palma dilakukan dengan penambahan
larutan alkali kuat, yaitu NaOH disertai pemisahan dengan metode sentrifugasi. Protein yang telah
terdenaturasi akan dipisahkan dengan proses pencucian dengan air. Untuk penghilangan komponen
lemak (defatting) menggunakan penambahan larutan kimia untuk menghilangan internal lipid yang
membentuk kompleks dengan amilosa dan ekstenal lipid yang menyelimuti granula pati.
Penghilangan internal lipid pada pati palma dilakukan dengan penambahan larutan DMSO (Dimetil
Sulfoksida) disertai pengadukan dengan shaker pada suhu 37oC. Selanjutnya penambahan metanol
dan disimpan pada suhu rendah (T=4oC) bertujuan untuk menghilangkan eksternal lipid pada pati
palma. Lemak yang telah terangkat dan sisa lemak yang belum hilang dicuci dengan metanol dan
eter. Penambahan kedua bahan kimia tersebut juga sekaligus mengikat air yang terkandung pada pati.
4.2 HIDROLISIS ENZIMATIS PATI PALMA
Hidrolisis pati dilakukan untuk memodifikasi pati alami sehingga menghasilkan produk berupa
hidrolisat pati yang memiliki karakteristik yang dapat disesuaikan dengan keperluan-keperluan
tertentu yang dibutuhkan. Karena pati yang banyak digunakan untuk berbagai kebutuhan sebagian
besar dalam bentuk hidrolisat pati bukan pati alami. Mekanisme hidrolisis yang dipilih adalah
hidrolisis secara enzimatis karena memiliki lebih banyak keunggulan dibandingkan secara asam.
Enzim merupakan katalis yang cocok untuk berbagai produk pangan maupun non-pangan. Kelebihan
hidrolisis secara enzimatis antara lain enzim bekerja secara spesifik pada substrat, lebih efisien,
produk yang dihasilkan lebih murni dan sedikit menghasilkan produk samping, kondisi proses dapat
dikontrol, serta biaya pemurnian lebih murah.
Kondisi kerja dan aktivitas optimum enzim pullulanase dan β-amilase sebagai enzim hidrolase
yang digunakan pada penelitian ini sangat penting untuk diketahui sebelum melakukan proses
hidrolisis. Hal ini dikarenakan kondisi hidrolisis harus diatur sesuai kondisi ideal enzim supaya dapat
bekerja dengan baik dan menghasilkan produk yang diharapkan. Kondisi kerja enzim, meliputi pH
dan suhu optimum yang telah diketahui berdasarkan penelitian terdahulu yang dapat dilihat pada
Tabel 8, yaitu untuk β-amilase pada suhu 40oC dengan pH 4,8 dan pullulanase pada suhu 40oC
dengan pH 6,0. Nilai pH (Power of Hydrogen) merupakan parameter penting dalam proses enzimatis
karena aktivitas optimum enzim dikondisikan pada pH tertentu dengan menggunakan buffer tertentu
yang memiliki pH tersebut. Struktur ion enzim tergantung pada pH lingkungannya karena pH
berpengaruh terhadap efektivitas bagian aktif enzim dalam membentuk kompleks enzim. Nilai pH
18
juga berkaitan dengan nilai pI (Power of Isoelectric) enzim. Titik isoelektrik adalah nilai pH dimana
protein memiliki muatan netral sehingga protein akan diam dan tidak bermigrasi kemanapun. Apabila
pH larutan penyangga (buffer) lebih besar dari titik isoelektriknya, maka molekul protein akan
bermigrasi menuju kutub positif sementara jika pH buffer lebih rendah dari titik isoelektrik, maka
protein akan bermigrasi ke kutub negatif. Apabila kondisi tersebut tidak terpenuhi, maka aktivitas
enzim dalam memutus ikatan-ikatan glukosidik pada pati tidak optimum (Lehninger 1982).
Perlakuan awal subtrat pati yang digunakan dalam menentukan aktivitas enzim dengan metode
gelatinisasi. Gelatinisasi merupakan proses yang menyebabkan granula pati mengembang karena
ikatan hidrogen melemah ketika suhu suspensi pati dinaikan. Umumnya metode gelatinisasi
dilakukan dengan pemanasan. Akan tetapi, pada penelitian ini digunakan metode gelatinisasi dengan
menggunakan basa yang dikondisikan pada suhu rendah untuk mempercepat terjadinya gelatinisasi
pati. Metode ini juga dilakukan karena dikhawatirkan proses gelatinisasi pada suhu tinggi dapat
merusak struktur pati dengan terjadinya pemutusan ikatan-ikatan antar molekul pati yang tidak
diinginkan.
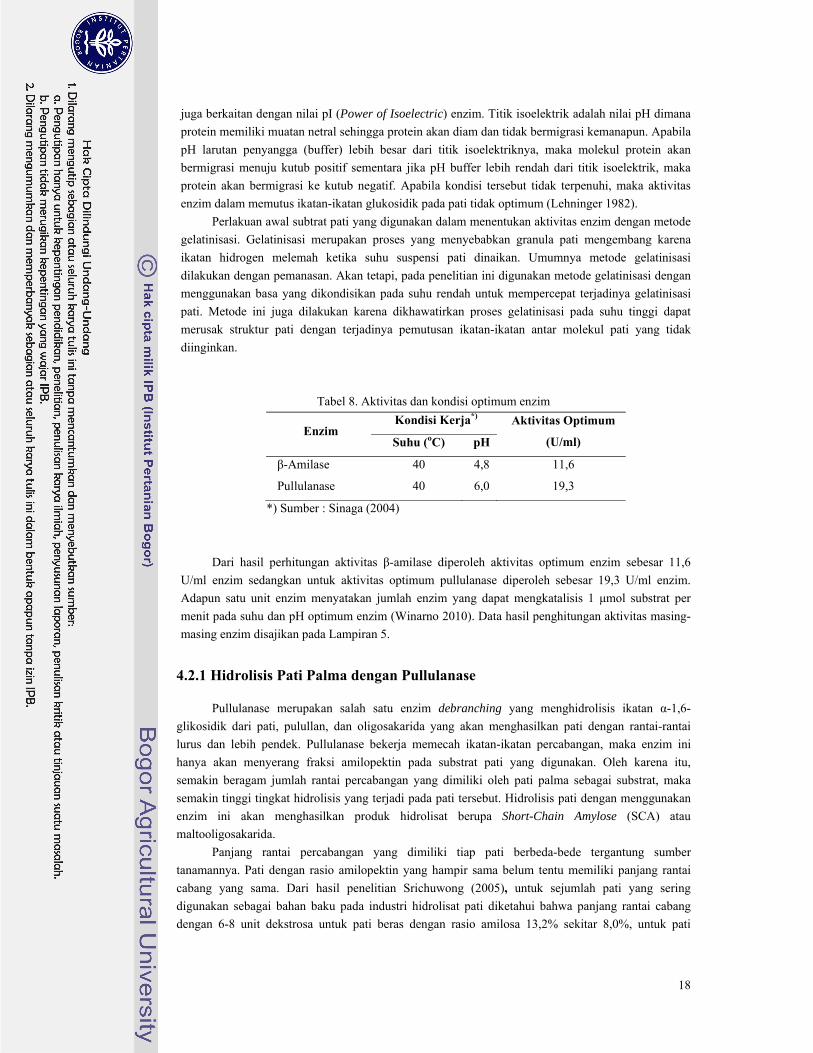
Tabel 8. Aktivitas dan kondisi optimum enzim
Enzim Kondisi Kerja*) Aktivitas Optimum
(U/ml) Suhu (oC) pH
β-Amilase 40 4,8 11,6
Pullulanase 40 6,0 19,3
*) Sumber : Sinaga (2004)
Dari hasil perhitungan aktivitas β-amilase diperoleh aktivitas optimum enzim sebesar 11,6
U/ml enzim sedangkan untuk aktivitas optimum pullulanase diperoleh sebesar 19,3 U/ml enzim.
Adapun satu unit enzim menyatakan jumlah enzim yang dapat mengkatalisis 1 μmol substrat per
menit pada suhu dan pH optimum enzim (Winarno 2010). Data hasil penghitungan aktivitas masing-
masing enzim disajikan pada Lampiran 5.
4.2.1 Hidrolisis Pati Palma dengan Pullulanase
Pullulanase merupakan salah satu enzim debranching yang menghidrolisis ikatan α-1,6-
glikosidik dari pati, pulullan, dan oligosakarida yang akan menghasilkan pati dengan rantai-rantai
lurus dan lebih pendek. Pullulanase bekerja memecah ikatan-ikatan percabangan, maka enzim ini
hanya akan menyerang fraksi amilopektin pada substrat pati yang digunakan. Oleh karena itu,
semakin beragam jumlah rantai percabangan yang dimiliki oleh pati palma sebagai substrat, maka
semakin tinggi tingkat hidrolisis yang terjadi pada pati tersebut. Hidrolisis pati dengan menggunakan
enzim ini akan menghasilkan produk hidrolisat berupa Short-Chain Amylose (SCA) atau
maltooligosakarida.
Panjang rantai percabangan yang dimiliki tiap pati berbeda-bede tergantung sumber
tanamannya. Pati dengan rasio amilopektin yang hampir sama belum tentu memiliki panjang rantai
cabang yang sama. Dari hasil penelitian Srichuwong (2005), untuk sejumlah pati yang sering
digunakan sebagai bahan baku pada industri hidrolisat pati diketahui bahwa panjang rantai cabang
dengan 6-8 unit dekstrosa untuk pati beras dengan rasio amilosa 13,2% sekitar 8,0%, untuk pati

19
tapioka dengan rasio amilosa 17,9% sekitar 9,9%, untuk pati jagung dengan rasio amilosa
23,4%sekitar 5,1% dan untuk pati sagu dengan rasio amilosa 21,9% sekitar 9,0%. Rata-rata pati
tersebut memiliki sedikit rantai cabang yang sangat pendek dan rantai cabang yang sangat panjang
dengan 25-30 unit dekstrosa penyusunnya. Namun, pada rantai percabangan yang cukup panjang
dengan jumlah unit 9-12 dekstrosa pati beras, tapioka, jagung, dan sagu memiliki jumlah panjang
rantai cabang tersebut sekitar 28,1-36,3% dimana pati tapioka memiliki panjang rantai dengan 9-12
unit dekstrosa paling banyak sedangkan pati sagu paling sedikit. Panjang rantai cabang dengan jumlah
unit 13-24 dekstrosa untuk pati beras sekitar 52,1%, pati tapioka sekitar 48,3%, pati jagung sekitar
56,7%, dan pati sagu sekitar 56,2%. Dari data tersebut terlihat bahwa tiap pati memiliki panjang rantai
percabangan dengan jumlah unit dekstrosa penyusun rantai yang berbeda-beda sehingga selain jumlah
rantai percabangan, panjang rantai percabangan yang dimiliki tiap jenis pati juga beragam yang akan
mempengaruhi penurunan derajat polimerisasi dan peningkatan nilai dextrose equivalent yang
terbentuk dari proses hidrolisis enzimatis terutama dengan enzim debranching seperti pullulanase.
Rantai cabang yang panjang akan menghasilkan penurunan nilai DP yang lebih rendah dibandingkan
pada rantai cabang yang lebih pendek dan sebaliknya akan menghasilkan nilai DE produk yang lebih
sedikit.
Pada penelitian ini, di awal hidrolisis dilakukan sampling untuk analisa total karbohidrat
sedangkan tiap waktu tertentu juga dilakukan sampling untuk analisa gula pereduksinya. Kedua
analisa dilakukan untuk menghitung nilai Derajat Polimerisasi (DP), Dextrose Equivalent (DE), dan
tingkat hidrolisis. Nilai DP, DE, dan tingkat hidrolisis pada tiap waktu hidrolisis yang dilakukan oleh
pullulanase untuk masing-masing pati palma tersaji pada Lampiran 6. Derajat Polimerisasi (DP)
menunjukkan jumlah rata-rata monomer (monosakarida) dalam suatu molekul sedangkan Dextrose
Equivalent (DE) menunjukkan jumlah gula pereduksi dalam persen dekstrosa murni dalam basis
kering. Pada proses hidrolisis semakin lama waktu hidrolisis berlangsung, maka nilai DP produk yang
dihasilkan akan semakin menurun akibat kerja enzim yang memutus ikatan-ikatan α-1,6-glikosidik
pada amilopektin sehingga menghasilkan rantai-rantai pati yang lurus dan lebih pendek yang
ditunjukkan dengan penurunan berat molekul pati. Perubahan nilai DP berbanding terbalik dengan
perubahan nilai DE selama proses hidrolisis. Sejalan waktu hidrolisis berlangsung terjadi peningkatan
nilai DE pada produk akibat adanya peningkatan pembentukan gula pereduksi yang dihasilkan dari
pemecahan rantai-rantai cabang yang panjang. Kandungan gula pereduksi yang dihasilkan dari kerja
enzim pullulanase yang memutus ikatan α-1,6-glikosidik pada amilopektin pati juga dapat mengukur
tingkat hidrolisis yang terjadi. Semakin terhidrolisis secara sempurna, maka semakin banyak gula
pereduksi yang dihasilkan.
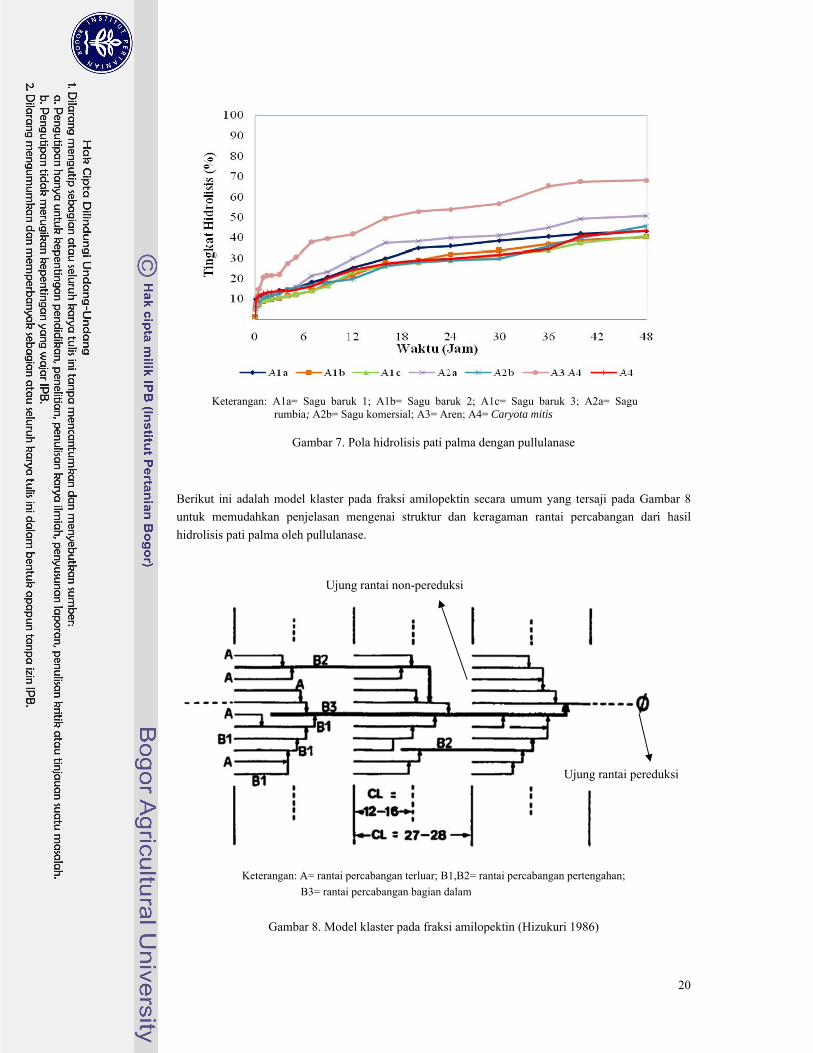
Pada Gambar 7 terlihat bahwa tingkat hidrolisis semakin meningkat secara perlahan mulai dari
jam ke-0 hingga jam ke-48. Dari ketujuh pati palma terlihat bahwa pati aren memiliki peningkatan
tingkat hidrolisis yang paling tinggi karena pati aren memiliki keragaman rantai amilopektin yang
paling tinggi. Hal ini didukung oleh data pada Lampiran 6, peningkatan hidrolisis pati aren selama 48
jam waktu inkubasi mencapai tingkat hidrolisis 68,3% jauh lebih tinggi dibandingkan keenam pati
palma lainnya. Begitu juga untuk pati sagu rumbia meskipun termasuk pati palma dengan rasio
amilopektin paling rendah, pati sagu rumbia termasuk pati yang memiliki jumlah percabangan cukup
banyak tetapi tiap rantai percabangannya pendek-pendek sehingga saat jam ke-48 hidrolisis pati sagu
rumbia mencapai tingkat hidrolisis 50,7%. Kedua pati tersebut memiliki indikasi memiliki jumlah
rantai percabangan yang cukup banyak pada bagian pertengahan klaster fraksi amilopektin. Hal ini
dikarenakan pati aren telah mencapai tingkat hidrolisis 40% pada jam ke-10 sedangkan pati sagu
rumbia terjadi pada jam ke-24. Berbeda dengan kelima pati palma lainnya yang mencapai tingkat
hidrolisis 40% pada jam ke-40 hingga jam ke-48.
20
Keterangan: A1a= Sagu baruk 1; A1b= Sagu baruk 2; A1c= Sagu baruk 3; A2a= Sagu
rumbia; A2b= Sagu komersial; A3= Aren; A4= Caryota mitis
Gambar 7. Pola hidrolisis pati palma dengan pullulanase
Berikut ini adalah model klaster pada fraksi amilopektin secara umum yang tersaji pada Gambar 8
untuk memudahkan penjelasan mengenai struktur dan keragaman rantai percabangan dari hasil
hidrolisis pati palma oleh pullulanase.
Keterangan: A= rantai percabangan terluar; B1,B2= rantai percabangan pertengahan;
B3= rantai percabangan bagian dalam
Gambar 8. Model klaster pada fraksi amilopektin (Hizukuri 1986)
Ujung rantai pereduksi
Ujung rantai non-pereduksi

21
Perubahan tingkat hidrolisis yang terjadi mengalami peningkatan yang cukup signifikan karena
enzim pullulanase bekerja dengan cepat memutus ikatan-ikatan pada rantai percabangan pati pada
gugus amilopektin, yaitu mulai dari percabangan pada rantai A lalu ke rantai B1, B2, dan terakhir
memutus ikatan percabangan pada rantai B3 hingga seluruh percabangan terputus menjadi rantai-
rantai pendek amilosa. Tingkat hidrolisis dihitung untuk mengetahui berapa lama waktu hidrolisis
dibutuhkan untuk menghasilkan produk dengan nilai DP dan DE yang diinginkan sesuai kebutuhan
sehingga untuk membuat suatu produk hidrolisat pati dapat digunakan waktu yang sesuai dengan
tingkat hidrolisis yang dibutuhkan. Nilai Derajat Polimerisasi (DP) dan Dextrose Equivalent (DE)
pada hidrolisis untuk masing-masing pati palma dengan pullulanase dapat dilihat pada Gambar 9
sampai 15.
Dari Gambar 9 sampai 15 terlihat terjadinya penurunan nilai DP terjadi karena polimer dari
pati terpecah-pecah menjadi oligosakarida dengan jumlah unit-unit yang lebih sederhana oleh enzim
sehingga menyebabkan terbentuknya senyawa-senyawa gula pereduksi yang ditandai dengan
meningkatnya nilai DE. Pada proses hidrolisis peningkatan nilai DE tiap pati terjadi cukup cepat
hingga terbentuknya hidrolisat pati yang diharapkan. Nilai DP dan DE hidrolisat pati dengan
pullulanase menghasilkan nilai yang berbeda-beda antara pati palma dengan pati lainnya. Hal ini
karena tiap pati memiliki keragaman amilosa dan amilopektin tergantung sumber patinya.
Berdasarkan hasil penelitian Sinaga (2004), pati umbi-umbian, seperti ubi jalar, ubi kayu, ganyong
memiliki nilai DP pada tingkat hidrolisis 100% berkisar antara 21,5-36,7 sedangkan nilai DP untuk
waxy potato sebesar 35 (Cai 2010). Dari nilai DP tersebut dapat diketahui bahwa pati umbi-umbian
memiliki panjang rantai percabangan yang lebih panjang dibandingkan dengan pati palma. Karena
untuk menghasilkan Short-Chains Amylose (SCA) dari pati umbi-umbian nilai DP yang dihasilkan
lebih tinggi dibandingkan hasil hidrolisat SCA oleh pullulanase pada pati palma, yaitu sebesar 21,5-
36,7. Berdasarkan hasil yang diperoleh pada penelitian ini diketahui bahwa pati palma memiliki
keragaman rantai amilopektin yang cukup tinggi dengan panjang rantai percabangan yang lebih
pendek-pendek dari pati umbi-umbian. Oleh karena itu, nilai DP hidrolisat pati palma oleh pullulanase
pada tingkat hidrolisis 100% yang dihasilkan akan lebih rendah dibandingkan pati umbi-umbian. Dari
ketujuh pati palma yang digunakan pun memiliki nilai DP dan DE yang beragam tergantung dari rasio
amilopektin yang terkandung dalam struktur patinya.
Nilai-nilai DP dan DE untuk masing-masing pati palma pada tingkat hidrolisis 10, 40, dan
100% tersaji pada Tabel 9. Dari hidrolisat pati yang dihasilkan, diperoleh nilai DP rata-rata pada
tingkat hidrolisis 10% untuk masing-masing pati palma berkisar antara 132,9-227,4 dari kisaran nilai
DP awal 273,4 untuk sagu baruk 1 hingga untuk Caryota mitis 1592,3. Hal ini menunjukkan pada
hampir semua pati palma mengandung sedikit rantai percabangan pada rantai A gugus amilopektin
sehingga enzim belum bekerja terlalu optimum dalam memecah polimer dari pati yang menyebabkan
nilai DP rata-rata tiap pati masih tinggi. Pada tingkat hidrolisis 40% nilai DP rata-rata untuk masing-
masing pati palma berkisar antara 34,9-51,5 dan nilai DE untuk masing-masing pati berkisar antara
1,9–2,6. Pada tingkat hidrolisis ini telah terbentuk rantai-rantai pendek amilosa hasil pemutusan rantai
percabangan pada amilopektin hingga bagian rantai B3 di semua pati palma. Tingkat hidrolisis 40%
ini terjadi pada jam ke-40 sampai jam ke-48 untuk hampir semua jenis pati palma yang digunakan,
kecuali pati aren dan sagu rumbia. Pati aren mengalami tingkat hidrolisis 40% pada jam ke-10 karena
memiliki rantai percabangan yang paling banyak terhidrolisis oleh pullulanase di bagian rantai A dan
B1.
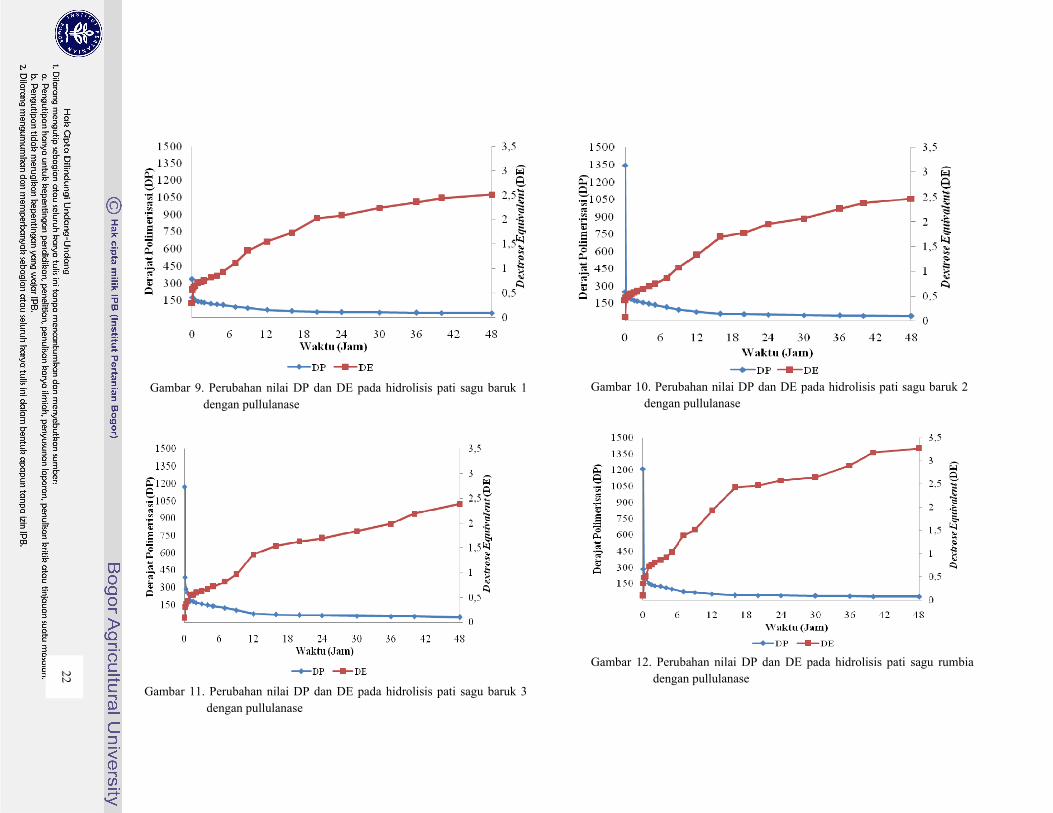
22
Gambar 11. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 3 dengan pullulanase
Gambar 9. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 1 dengan pullulanase
Gambar 10. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 2 dengan pullulanase
Gambar 12. Perubahan nilai DP dan DE pada hidrolisis pati sagu rumbia dengan pullulanase
22
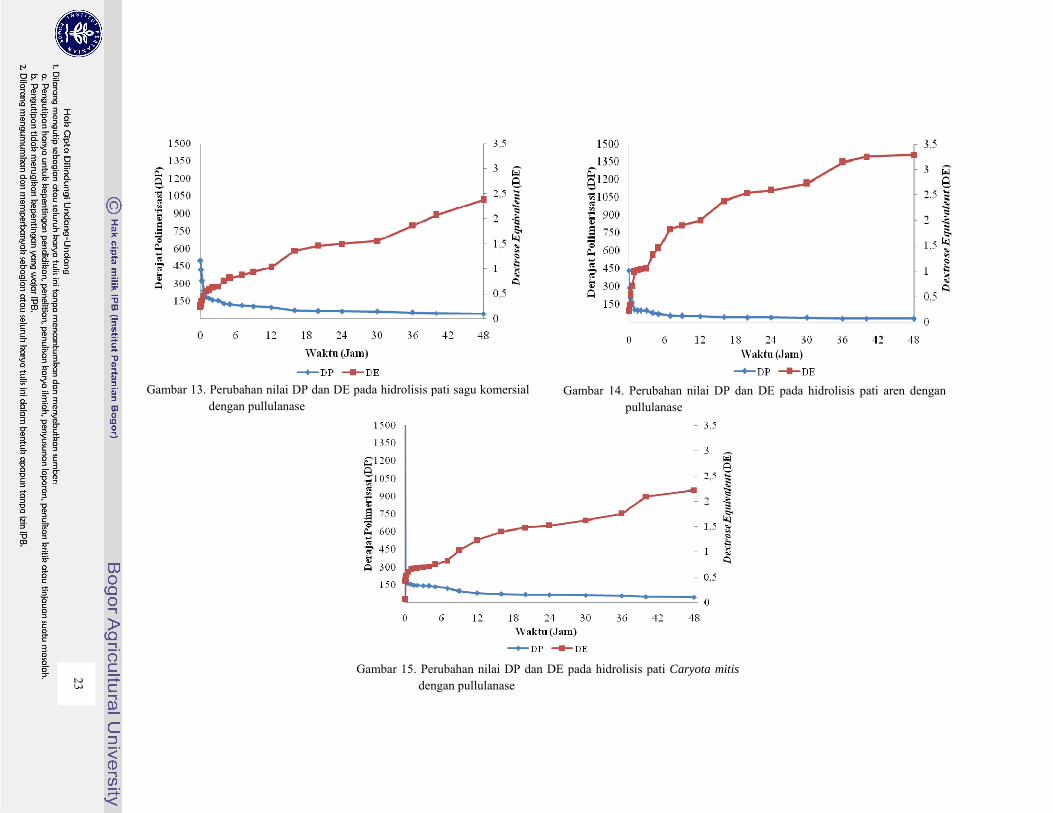
23
Gambar 13. Perubahan nilai DP dan DE pada hidrolisis pati sagu komersial dengan pullulanase
Gambar 14. Perubahan nilai DP dan DE pada hidrolisis pati aren dengan pullulanase
23
Gambar 15. Perubahan nilai DP dan DE pada hidrolisis pati Caryota mitis dengan pullulanase
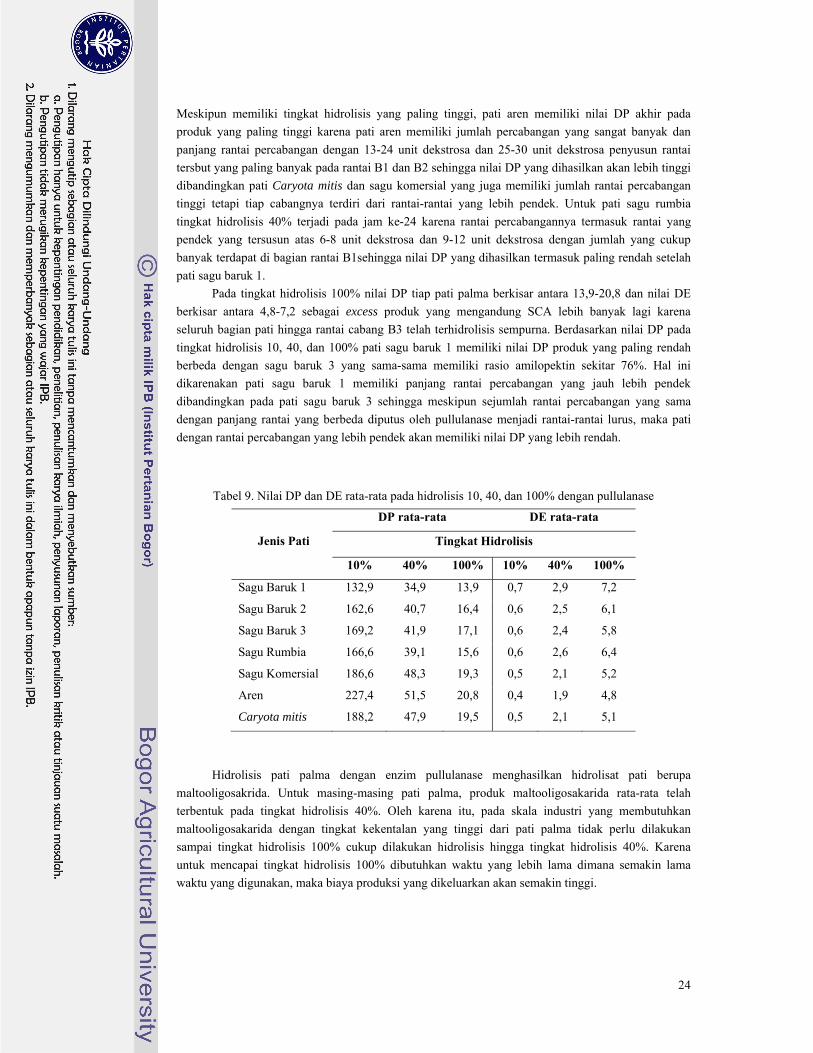
24
Meskipun memiliki tingkat hidrolisis yang paling tinggi, pati aren memiliki nilai DP akhir pada
produk yang paling tinggi karena pati aren memiliki jumlah percabangan yang sangat banyak dan
panjang rantai percabangan dengan 13-24 unit dekstrosa dan 25-30 unit dekstrosa penyusun rantai
tersbut yang paling banyak pada rantai B1 dan B2 sehingga nilai DP yang dihasilkan akan lebih tinggi
dibandingkan pati Caryota mitis dan sagu komersial yang juga memiliki jumlah rantai percabangan
tinggi tetapi tiap cabangnya terdiri dari rantai-rantai yang lebih pendek. Untuk pati sagu rumbia
tingkat hidrolisis 40% terjadi pada jam ke-24 karena rantai percabangannya termasuk rantai yang
pendek yang tersusun atas 6-8 unit dekstrosa dan 9-12 unit dekstrosa dengan jumlah yang cukup
banyak terdapat di bagian rantai B1sehingga nilai DP yang dihasilkan termasuk paling rendah setelah
pati sagu baruk 1.
Pada tingkat hidrolisis 100% nilai DP tiap pati palma berkisar antara 13,9-20,8 dan nilai DE
berkisar antara 4,8-7,2 sebagai excess produk yang mengandung SCA lebih banyak lagi karena
seluruh bagian pati hingga rantai cabang B3 telah terhidrolisis sempurna. Berdasarkan nilai DP pada
tingkat hidrolisis 10, 40, dan 100% pati sagu baruk 1 memiliki nilai DP produk yang paling rendah
berbeda dengan sagu baruk 3 yang sama-sama memiliki rasio amilopektin sekitar 76%. Hal ini
dikarenakan pati sagu baruk 1 memiliki panjang rantai percabangan yang jauh lebih pendek
dibandingkan pada pati sagu baruk 3 sehingga meskipun sejumlah rantai percabangan yang sama
dengan panjang rantai yang berbeda diputus oleh pullulanase menjadi rantai-rantai lurus, maka pati
dengan rantai percabangan yang lebih pendek akan memiliki nilai DP yang lebih rendah.
Tabel 9. Nilai DP dan DE rata-rata pada hidrolisis 10, 40, dan 100% dengan pullulanase
Jenis Pati
DP rata-rata DE rata-rata
Tingkat Hidrolisis
10% 40% 100% 10% 40% 100%
Sagu Baruk 1 132,9 34,9 13,9 0,7 2,9 7,2
Sagu Baruk 2 162,6 40,7 16,4 0,6 2,5 6,1
Sagu Baruk 3 169,2 41,9 17,1 0,6 2,4 5,8
Sagu Rumbia 166,6 39,1 15,6 0,6 2,6 6,4
Sagu Komersial 186,6 48,3 19,3 0,5 2,1 5,2
Aren 227,4 51,5 20,8 0,4 1,9 4,8
Caryota mitis 188,2 47,9 19,5 0,5 2,1 5,1
Hidrolisis pati palma dengan enzim pullulanase menghasilkan hidrolisat pati berupa
maltooligosakrida. Untuk masing-masing pati palma, produk maltooligosakarida rata-rata telah
terbentuk pada tingkat hidrolisis 40%. Oleh karena itu, pada skala industri yang membutuhkan
maltooligosakarida dengan tingkat kekentalan yang tinggi dari pati palma tidak perlu dilakukan
sampai tingkat hidrolisis 100% cukup dilakukan hidrolisis hingga tingkat hidrolisis 40%. Karena
untuk mencapai tingkat hidrolisis 100% dibutuhkan waktu yang lebih lama dimana semakin lama
waktu yang digunakan, maka biaya produksi yang dikeluarkan akan semakin tinggi.

25
4.2.2 Hidrolisis Pati Palma dengan β-Amilase dan Pullulanase secara Suksesif
β-amilase disebut juga α-l,4-glukan maltohidrolase (E.C. 3.2.1.2) merupakan tipe ekso-enzim
yang memutus ikatan α-1,4-glukosidik pada gugus amilosa dan amilopektin pati mulai dari bagian
rantai luar molekul, yaitu tiap dua molekul pada ujung rantai non-pereduksi. Mekanisme kerja enzim
ini akan mencari dan memutus seluruh ikatan α-1,4-glukosidik pada rantai di percabangan terluar
terlebih dahulu kemudian memutus ikatan α-1,4-glukosidik yang berada di rantai lurus amilosa
sehingga waktu yang dibutuhkan untuk hidrolisis dengan enzim ini sangat lama. Karena β-amilase
tidak dapat menghidrolisis ikatan α-1,6-glukosidik pada titik percabangan, maka pada hidrolisis
dengan enzim ini akan dihasilkan produk samping berupa β-limit dekstrin.
Hidrolisis dilakukan secara kontinu selama 48 jam hingga substrat (pati palma) terhidrolisis
sempurna pada tingkat hidrolisis 100%. Pada awal hidrolisis dilakukan sampling untuk analisa total
karbohidrat sedangkan tiap waktu tertentu juga dilakukan sampling untuk analisa gula pereduksinya.
Kedua analisa dilakukan untuk menghitung nilai Derajat Polimerisasi (DP), Dextrose Equivalent
(DE), tingkat hidrolisis, dan persentase kandungan β-limit dekstrin yang dihasilkan. Nilai DP, DE,
tingkat hidrolisis, dan persentase kandungan β-limit dekstrin yang dihasilkan pada tiap waktu
hidrolisis yang dilakukan oleh β-amilase dan pullulanase yang dilakukan secara suksesif untuk
masing-masing pati palma tersaji pada Lampiran 7.
Selama proses hidrolisis nilai DP mengalami penurunan yang signifikan pada menit ke-5 untuk
semua pati palma setelah ditambahkan enzim β-amilase kemudian terus menurun secara perlahan
hingga substrat (pati) mencapai tingkat hidrolisis 100% pada jam ke-48. Sebaliknya terjadi
peningkatan nilai DE secara perlahan mulai dari jam ke-0 hingga jam ke-24. Pada jam ke-24
ditambahkan β-amilase berlebih dengan konsentrasi enzim sebesar 20 U/g pati dari sisa larutan pati
yang dihidrolisis. Penambahan β-amilase berlebih ini dilakukan untuk meningkatkan kerja enzim
dalam memutus ikatan α-1,4-glukosidik pada sisa larutan pati sehingga diperoleh kandungan gula
pereduksi yang semakin meningkat. Namun, seiring dengan waktu inkubasi, β-amilase mulai berhenti
bekerja yang ditandai dengan pola grafik tingkat hidrolisis yang stasioner pada Gambar 16 dari jam
ke-28 sampai jam ke-36 sebelum penambahan enzim pullulanase karena β-amilase tidak dapat
memutus ikatan α-1,6-glukosidik pada titik percabangan. Hal ini menyebabkan terbentuknya β-limit
dekstrin dengan berat molekul yang tinggi berisi seluruh ikatan α-1,6.
Kandungan β-limit dekstrin yang terkandung pada pati kentang mencapai 54,5% dan pati
tapioka sebesar 58,3% (Bertoft 2008) sedangkan menurut Hizukuri (1981), persentase β-limit dekstrin
yang dihasilkan dalam pati kentang sebesar 68% dan pati tapioka sebesar 64%. Nilai tersebut tidak
jauh berbeda dari hasil penelitian Nilsson (2001) dimana β-limit dekstrin yang dihasilkan pada Potato
Amylopectin Starch (PAP) sebesar 54%. Dari nilai tersebut diketahui bahwa pada umbi-umbian
memiliki kandungan β-limit dekstrin yang sangat tinggi pada hidrolisis dengan β-amilase karena
rantai percabangan pati umbi-umbian cukup panjang dengan jumlah rantai percabangan yang lebih
banyak dibandingkan pada pati palma. Karena rantai cabang inilah yang menyebabkan enzim β-
amilase tidak dapat menghidrolisis pati secara sempurna sehingga dihasilkan β-limit dekstrin.
Semakin banyak dan panjang rantai percabangan pada struktur amilopektin pati, maka semakin
banyak bagian pati yang tidak dapat terhidrolisis oleh enzim β-amilase. Berbeda dengan pati umbi-
umbian, hampir semua pati palma memiliki keragaman amilopektin yang tinggi tetapi rantai cabang
yang dimiliki pendek-pendek sehingga β-limit dekstrin yang dihasilkan lebih sedikit dibandingkan
pati umbi-umbian. Untuk masing-masing pati palma yang digunakan diperoleh persentase kandungan
β-limit dekstrin yang berbeda. Persentase kandungan β-limit dekstrin tiap pati palma yang disajikan
pada Tabel 10.
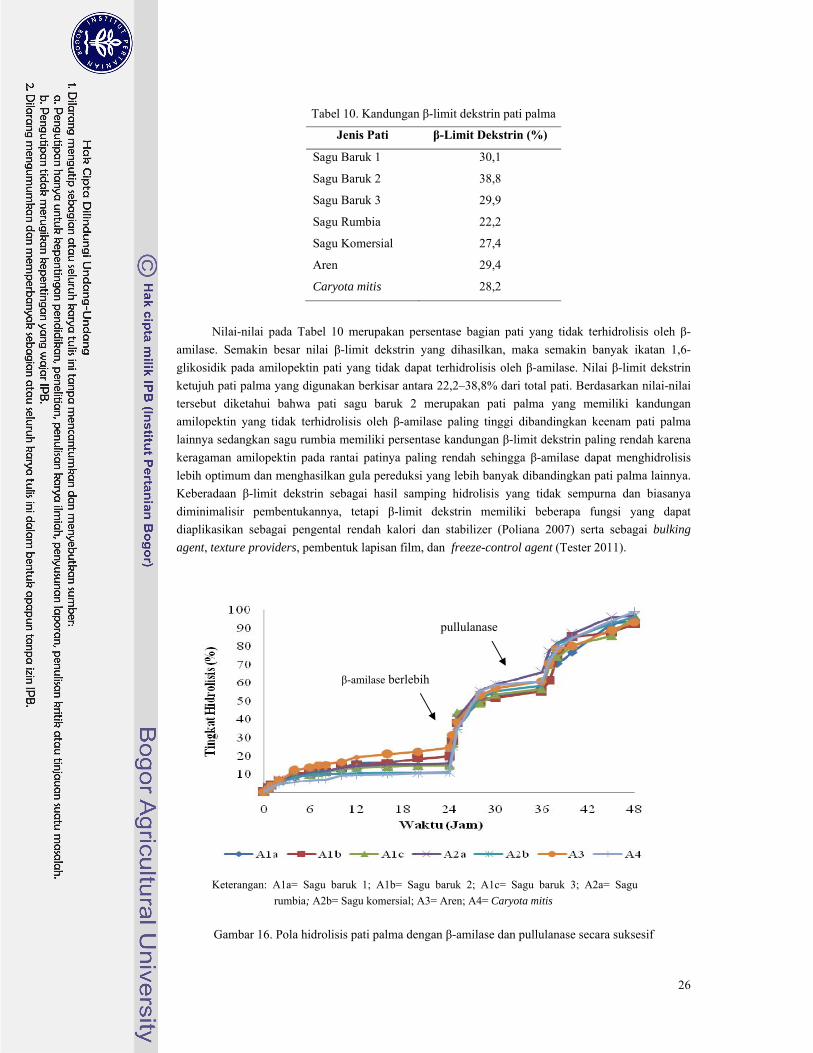
26
Tabel 10. Kandungan β-limit dekstrin pati palma
Nilai-nilai pada Tabel 10 merupakan persentase bagian pati yang tidak terhidrolisis oleh β-
amilase. Semakin besar nilai β-limit dekstrin yang dihasilkan, maka semakin banyak ikatan 1,6-
glikosidik pada amilopektin pati yang tidak dapat terhidrolisis oleh β-amilase. Nilai β-limit dekstrin
ketujuh pati palma yang digunakan berkisar antara 22,2–38,8% dari total pati. Berdasarkan nilai-nilai
tersebut diketahui bahwa pati sagu baruk 2 merupakan pati palma yang memiliki kandungan
amilopektin yang tidak terhidrolisis oleh β-amilase paling tinggi dibandingkan keenam pati palma
lainnya sedangkan sagu rumbia memiliki persentase kandungan β-limit dekstrin paling rendah karena
keragaman amilopektin pada rantai patinya paling rendah sehingga β-amilase dapat menghidrolisis
lebih optimum dan menghasilkan gula pereduksi yang lebih banyak dibandingkan pati palma lainnya.
Keberadaan β-limit dekstrin sebagai hasil samping hidrolisis yang tidak sempurna dan biasanya
diminimalisir pembentukannya, tetapi β-limit dekstrin memiliki beberapa fungsi yang dapat
diaplikasikan sebagai pengental rendah kalori dan stabilizer (Poliana 2007) serta sebagai bulking
agent, texture providers, pembentuk lapisan film, dan freeze-control agent (Tester 2011).
Keterangan: A1a= Sagu baruk 1; A1b= Sagu baruk 2; A1c= Sagu baruk 3; A2a= Sagu
rumbia; A2b= Sagu komersial; A3= Aren; A4= Caryota mitis
Gambar 16. Pola hidrolisis pati palma dengan β-amilase dan pullulanase secara suksesif
Jenis Pati β-Limit Dekstrin (%)
Sagu Baruk 1 30,1
Sagu Baruk 2 38,8
Sagu Baruk 3 29,9
Sagu Rumbia 22,2
Sagu Komersial 27,4
Aren 29,4
Caryota mitis 28,2
pullulanase
β-amilase berlebih

27
Pada Gambar 16 terlihat bahwa tingkat hidrolisis meningkat secara perlahan hingga jam ke-10
dan mulai stasioner hingga jam ke-24. Selanjutnya setelah penambahan β-amilase berlebih pada jam
ke-24 mulai mengalami peningkatan kandungan gula pereduksi yang signifikan hingga jam ke-28 lalu
kembali stasioner hingga jam ke-36. Peningkatan gula pereduksi yang dihasilkan terus terjadi setelah
penambahan pullulanase. Pada gambar tersebut terlihat pola hidrolisis dengan peningkatan yang
lambat pada awal proses hidrolisis karena kerja enzim β-amilase mencari dan memutus seluruh gugus
non-pereduksi pada rantai terluar pati terlebih dahulu lalu setelah itu memutus bagian tengah rantai
pati hingga semua bagian rantai pati terhidrolisis sehingga kerja enzim ini sangat lambat. Berdasarkan
hasil peningkatan tingkat hidrolisisnya, pada jam ke-48 ketujuh pati palma telah mengalami 92,3-99%
hidrolisis karena adanya kerja enzim β-amilase dan pullulanase yang berlangsung suksesif sehingga
hampir semua rantai baik pada gugus amilosa maupun amilopektin pati telah terputus menjadi gula-
gula pereduksi, yaitu maltosa seperti yang diharapkan. Hasil dari hidrolisis semakin lama waktu
inkubasi, maka semakin banyak gula pereduksi yang dihasilkan sehingga semakin tercapai tingkat
hidrolisis yang sempurna. Tingkat hidrolisis dihitung untuk mengetahui berapa lama waktu hidrolisis
dibutuhkan untuk menghasilkan produk dengan nilai DP dan DE yang diinginkan sesuai kebutuhan
sehingga untuk membuat suatu produk hidrolisat pati dapat digunakan waktu yang sesuai dengan
tingkat hidrolisis yang dibutuhkan.
Pada jam ke-36 ditambahkan sejumlah enzim pullulanase dengan konsentrasi yang sama
dengan jumlah β-amilase yang diberikan diawal proses hidrolisis, yaitu sebesar 2 U/g pati dari sisa
larutan pati yang dihidrolisis. Penambahan enzim ini untuk memecah titik percabangan pada pati
sehingga menghasilkan rantai-rantai lurus. Hal ini menyebabkan meningkatnya kandungan maltosa
pada hasil akhir hidrolisis. Setelah dilakukan penambahan enzim ini terlihat pada Gambar 17 sampai
23 bahwa untuk masing-masing pati palma mengalami peningkatan kandungan gula pereduksi
(maltosa) yang signifikan yang ditandai dengan peningkatan nilai DE dari jam ke-36 hingga jam ke-
38. Pullulanase yang ditambahkan bekerja memutus rantai percabangan pati menjadi rantai-rantai
lurus sehingga keberadaan rantai lurus yang dihasilkan menyebabkan β-amilase kembali bekerja
dimana pada titik inilah kerja kedua enzim berlangsung secara suksesif. Terbentuknya produk maltosa
dari masing-masing pati palma rata-rata terjadi pada jam ke-37 sampai jam ke-38, yaitu pada tingkat
hidrolisis 70-80%. Hal ini tidak jauh berbeda pada hidrolisis pati kedelai dengan β-amilase yang
menghasilkan produk maltosa pada tingkat hidrolisis 80-88% (Bird 1953).
Untuk nilai DE dan DP dari hidrolisat pati oleh β-amilase yang diperoleh tiap sumber pati
berbeda-beda tergantung pada karakteristik kandungan rantai percabangan pati. Untuk pati umbi-
umbian, seperti ubi kayu, ubi jalar, ganyong, talas, kimpul, dan suweg masing-masing pati memiliki
nilai DP berturut-turut sebesar 2,2; 2,7; 2,8; 1,7; 2,5; dan 2,2 (Sinaga 2004). Dari nilai DP tersebut
menunjukkan bahwa pada tiap jenis pati umbi-umbian memiliki keragaman rantai percabangannya
dimana pati talas memiliki jumlah rantai percabangan yang paling sedikit dibandingkan pati umbi
lainnya. Tidak jauh berbeda nilai DP yang dihasilkan oleh pati palma dari hasil proses hidrolisis
dengan β-amilase dan pullulanase secara suksesif diperoleh nilai DP dan DE rata-rata pada tingkat
hidrolisis 10, 40, dan 100% yang disajikan pada Tabel 11. Berdasarkan data tersebut diketahui nilai
DP rata-rata pada tingkat hidrolisis 10% berkisar antara 17,2-31,3 untuk masing-masing pati palma.
Nilai DP pada tingkat hidrolisis 10% menunjukkan tiap pati palma masih memiliki rantai-rantai pati
yang cukup panjang karena β-amilase baru memutus ujung-ujung gugus amilosa maupun amilopektin
pada bagian terluar rantai pati sehingga gula pereduksi yang dihasilkan pun masih sedikit. Hal ini
ditunjukkan dari nilai DE rata-rata untuk masing-masing pati pada tingkat hidrolisis 10% masih
berkisar antara 3,6-5,7 dimana pati sagu baruk 1 termasuk pati palma yang memiliki nilai DP paling
rendah dan DE paling tinggi pada tingkat hidrolisis 10%. Sebaliknya pati Caryota mitis merupakan

28
pati palma yang memiliki nilai DP paling tinggi dan nilai DE paling rendah karena pati ini termasuk
pati yang memiliki keragaman amilopektin yang tinggi selain pati aren. Untuk nilai DP rata-rata pada
tingkat hidrolisis 40% berkisar antara 4,4-7,7 dengan nilai DE berkisar antara 14,1-22,4 untuk masing-
masing pati palma. Pada tingkat hidrolisis 40% telah terjadi peningkatan gula pereduksi yang
dihasilkan yang ditandai dengan semakin tingginya nilai DE untuk tiap pati palma. Dan pada tingkat
hidrolisis 100% nilai DP rata-rata yang dihasilkan berkisar antara 1,7–2,9. Hal ini menunjukkan
bahwa seluruh rantai pada pati palma telah terhidrolisis secara sempurna dan menghasilkan produk
akhir yang sebagian besar memiliki rantai yang terdiri atas 2 unit monosakarida, yaitu glukosa.
Produk dengan 2 unit glukosa ini merupakan disakarida yang disebut maltosa. Selain itu, nilai DE
yang dihasilkan pada masing-masing pati menunjukkan produk akhir yang dihasilkan adalah maltosa
dimana nilai DE rata-rata berkisar antara 35,1–57,3. Karena menurut Kennedy (1995), nilai DE untuk
produk maltosa sekitar 48–63.
Berdasarkan Gambar 21 nilai DP pada tingkat hidrolisis 10, 40, hingga 100% pati Caryota
mitis memiliki nilai peningkatan DE yang paling rendah bahkan jika dibandingkan dengan pati aren.
Padahal dari rasio amilosa dan amilopektin kedua pati pada Tabel 7 keduanya memiliki rasio
amilopektin yang sama-sama berkisar 79%. Hal ini sama dengan hasil yang terjadi pada hidrolisis
kedua pati oleh pullulanase karena pati aren memiliki rantai percabangan yang jauh lebih panjang
dibandingkan pati Caryota mitis sehingga pati aren memiliki ikatan α-,14-glukosidik yang lebih
banyak pada gugus amilopektin pati aren dibandingkan pada pati Caryota mitis. Karena banyaknya
ikatan α-,14-glukosidik pada pati aren menyebabkan enzim β-amilase bekerja lebih optimum dalam
memecah rantai tersebut dan menghasilkan lebih banyak gula-gula pereduksi yang berdampak pada
peningkatan nila DE yang lebih tinggi pada produk hidrolisat pati aren dibandingkan pada pati
Caryota mitis.
Untuk hasil hidrolisat pada pati sagu baruk 1 pun demikian, meskipun termasuk pati yang
memiliki rasio amilopektin yang cukup tinggi, pati ini merupakan pati palma yang memiliki nilai DP
yang paling rendah dan DE yang paling tinggi dibandingkan pati sagu baruk 3 yang memiliki rasio
amilopektin sama sekitar 76%. Karena pati sagu baruk 1 memiliki rantai percabangan yang jauh lebih
pendek dibandingkan pada pati sagu baruk 3 dan pati palma lainnya sehingga setelah kedua enzim
yang digunakan, yaitu β-amilase dan pullulanase selesai memecah seluruh rantai-rantai amilosa dan
amilopektin pati menjadi beberapa unit amilosa yang pendek-pendek, maka penurunan DP yang
terjadi akan paling tinggi dan dekstrosa-dekstrosa yang terbentuk pun semakin banyak yang
menyebabkan nilai DE sagu baruk 1 paling tinggi dibandingkan pada keenam pati palma lainnya.
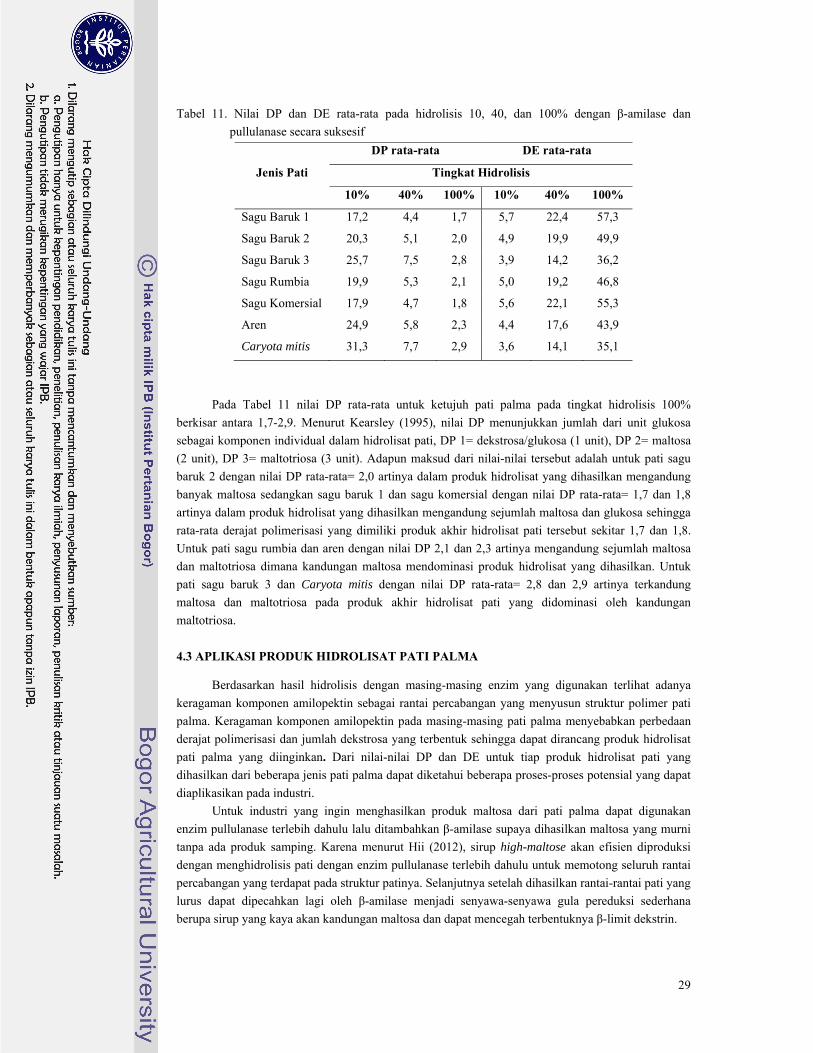
29
Tabel 11. Nilai DP dan DE rata-rata pada hidrolisis 10, 40, dan 100% dengan β-amilase dan
pullulanase secara suksesif
Jenis Pati
DP rata-rata DE rata-rata
Tingkat Hidrolisis
10% 40% 100% 10% 40% 100%
Sagu Baruk 1 17,2 4,4 1,7 5,7 22,4 57,3
Sagu Baruk 2 20,3 5,1 2,0 4,9 19,9 49,9
Sagu Baruk 3 25,7 7,5 2,8 3,9 14,2 36,2
Sagu Rumbia 19,9 5,3 2,1 5,0 19,2 46,8
Sagu Komersial 17,9 4,7 1,8 5,6 22,1 55,3
Aren 24,9 5,8 2,3 4,4 17,6 43,9
Caryota mitis 31,3 7,7 2,9 3,6 14,1 35,1
Pada Tabel 11 nilai DP rata-rata untuk ketujuh pati palma pada tingkat hidrolisis 100%
berkisar antara 1,7-2,9. Menurut Kearsley (1995), nilai DP menunjukkan jumlah dari unit glukosa
sebagai komponen individual dalam hidrolisat pati, DP 1= dekstrosa/glukosa (1 unit), DP 2= maltosa
(2 unit), DP 3= maltotriosa (3 unit). Adapun maksud dari nilai-nilai tersebut adalah untuk pati sagu
baruk 2 dengan nilai DP rata-rata= 2,0 artinya dalam produk hidrolisat yang dihasilkan mengandung
banyak maltosa sedangkan sagu baruk 1 dan sagu komersial dengan nilai DP rata-rata= 1,7 dan 1,8
artinya dalam produk hidrolisat yang dihasilkan mengandung sejumlah maltosa dan glukosa sehingga
rata-rata derajat polimerisasi yang dimiliki produk akhir hidrolisat pati tersebut sekitar 1,7 dan 1,8.
Untuk pati sagu rumbia dan aren dengan nilai DP 2,1 dan 2,3 artinya mengandung sejumlah maltosa
dan maltotriosa dimana kandungan maltosa mendominasi produk hidrolisat yang dihasilkan. Untuk
pati sagu baruk 3 dan Caryota mitis dengan nilai DP rata-rata= 2,8 dan 2,9 artinya terkandung
maltosa dan maltotriosa pada produk akhir hidrolisat pati yang didominasi oleh kandungan
maltotriosa.
4.3 APLIKASI PRODUK HIDROLISAT PATI PALMA
Berdasarkan hasil hidrolisis dengan masing-masing enzim yang digunakan terlihat adanya
keragaman komponen amilopektin sebagai rantai percabangan yang menyusun struktur polimer pati
palma. Keragaman komponen amilopektin pada masing-masing pati palma menyebabkan perbedaan
derajat polimerisasi dan jumlah dekstrosa yang terbentuk sehingga dapat dirancang produk hidrolisat
pati palma yang diinginkan. Dari nilai-nilai DP dan DE untuk tiap produk hidrolisat pati yang
dihasilkan dari beberapa jenis pati palma dapat diketahui beberapa proses-proses potensial yang dapat
diaplikasikan pada industri.
Untuk industri yang ingin menghasilkan produk maltosa dari pati palma dapat digunakan
enzim pullulanase terlebih dahulu lalu ditambahkan β-amilase supaya dihasilkan maltosa yang murni
tanpa ada produk samping. Karena menurut Hii (2012), sirup high-maltose akan efisien diproduksi
dengan menghidrolisis pati dengan enzim pullulanase terlebih dahulu untuk memotong seluruh rantai
percabangan yang terdapat pada struktur patinya. Selanjutnya setelah dihasilkan rantai-rantai pati yang
lurus dapat dipecahkan lagi oleh β-amilase menjadi senyawa-senyawa gula pereduksi sederhana
berupa sirup yang kaya akan kandungan maltosa dan dapat mencegah terbentuknya β-limit dekstrin.
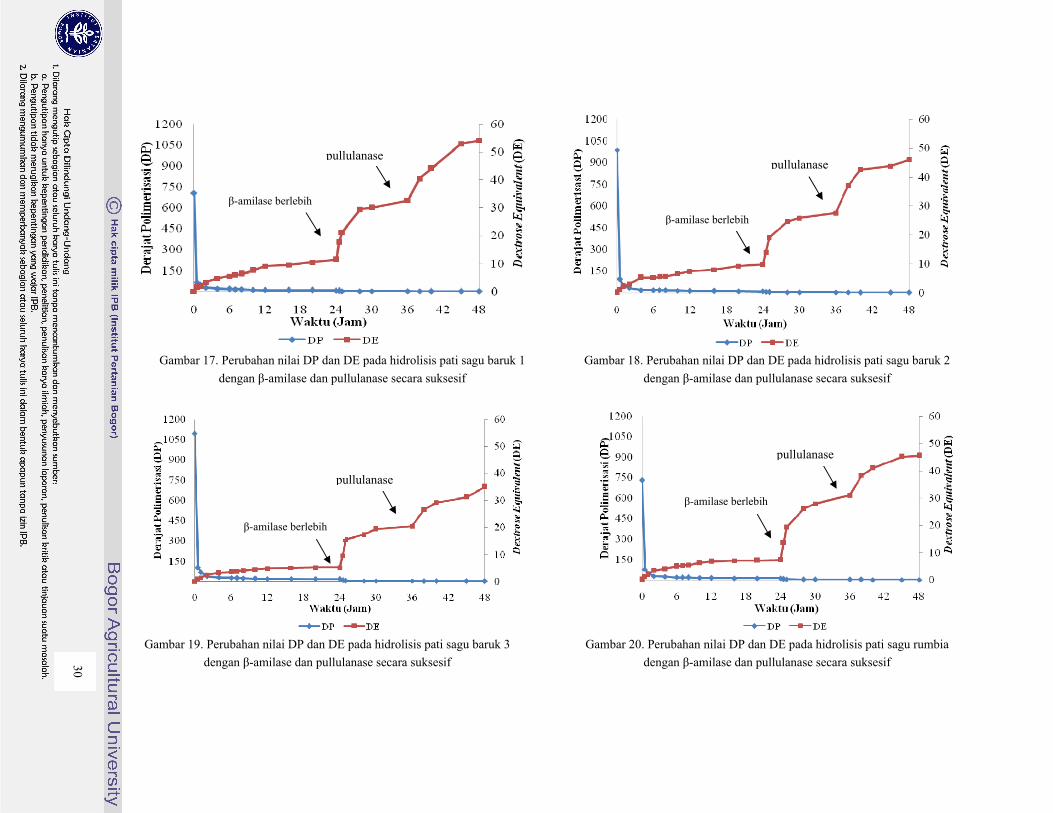
30
Gambar 17. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 1
dengan β-amilase dan pullulanase secara suksesif
pullulanase
β-amilase berlebih
pullulanase
β-amilase berlebih
Gambar 19. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 3
dengan β-amilase dan pullulanase secara suksesif
pullulanase
β-amilase berlebih
Gambar 20. Perubahan nilai DP dan DE pada hidrolisis pati sagu rumbia
dengan β-amilase dan pullulanase secara suksesif
pullulanase
β-amilase berlebih
30
Gambar 18. Perubahan nilai DP dan DE pada hidrolisis pati sagu baruk 2
dengan β-amilase dan pullulanase secara suksesif
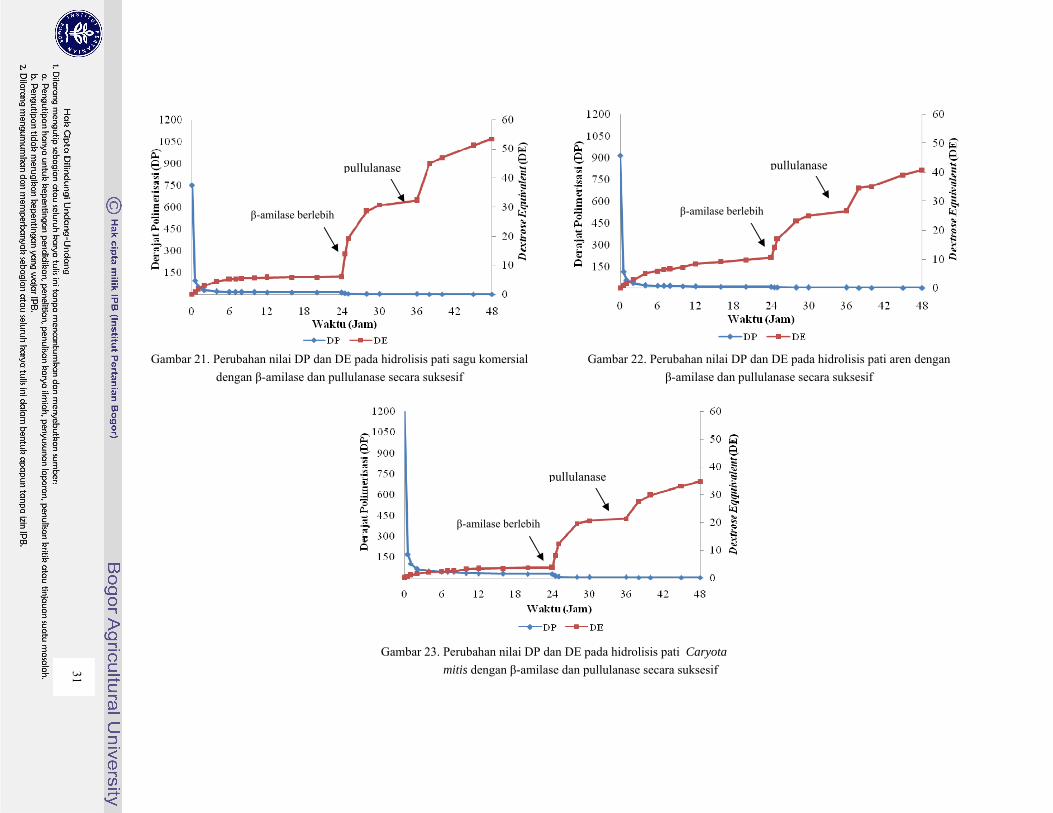
31
Gambar 23. Perubahan nilai DP dan DE pada hidrolisis pati Caryota
mitis dengan β-amilase dan pullulanase secara suksesif
pullulanase
β-amilase berlebih
Gambar 21. Perubahan nilai DP dan DE pada hidrolisis pati sagu komersial
dengan β-amilase dan pullulanase secara suksesif
pullulanase
β-amilase berlebih
Gambar 22. Perubahan nilai DP dan DE pada hidrolisis pati aren dengan
β-amilase dan pullulanase secara suksesif
31
pullulanase
β-amilase berlebih

32
Pullulanase diharapkan memutus semua rantai percabangan sehingga terbentuknya β-limit
dekstrin. Untuk menghasilkan hidrolisat pati palma berupa maltosa, pati palma yang paling potensial
digunakan sebagai substrat adalah pati sagu baruk 1, sagu baruk 2, sagu rumbia, dan sagu komersial.
Karena pada penelitian ini kedua pati tersebut memiliki nilai DP pada tingkat hidrolisis 100% yang
paling mendekati nilai DP maltosa sebesar 2. Namun, jika industri membutuhkan produk maltosa
yang tidak terlalu murni dapat dilakukan proses hidrolisis sampai tingkat hidrolisis 70-80% karena
pada tingkat hidrolisis tersebut pati-pati palma yang digunakan telah menghasilkan produk hidrolisat
berupa maltosa sehingga industri dapat menghemat biaya dengan menghemat waktu untuk mencapai
tingkat hidrolisis 70-80%.
Maltosa adalah disakarida yang terdiri atas ikatan glukosa dan glukosa. Sifat dan
pemanfaatannya hampir sama dengan sirup glukosa. Maltosa memiliki karakteristik yang khas, yaitu
tekanan osmotik dan kelarutan yang tinggi, tidak mempengaruhi flavor, dan tidak mengubah tekstur
produk. Maltosa sebagai produk hidrolisat yang dihasilkan pada penelitian ini secara alami dapat
digunakan sebagai bahan baku untuk pembuatan maltitol dan kristal maltitol serta pada industri
farmasi sebagai bahan yang digunakan untuk infus. Selain itu, dapat juga diproduksi sirup high-
maltose yang memiliki tingkat kemanisan yang ringan dengan viskositas yang rendah dalam larutan,
sifat higroskopi rendah, dan stabilitas panas yang baik.
Pada penelitian Hartanto (2005) kondisi sebenarnya yang dilakukan pada industri seperti di PT
Tainesia Jaya, Wonogiri, sirup high-maltose berbahan baku tapioka diproses melalui beberapa
tahapan, yaitu likuifikasi, sakarifikasi, filtrasi, penukaran ion, dan evaporasi. Pada tahap likuifikasi
tapioka diencerkan dalam tangki khusus yang ditambahkan enzim α-amilase pada suhu 94,5oC selama
126 menit. Selanjutnya pada tahap sakarifikasi dilakukan dalam tangki tunggal dengan alat pengaduk
dan ditambahkan enzim β-amilase dan pullulanase pada suhu 50oC. Setelah terbentuk larutan sirup
yang mengandung maltosa dilanjutkan ke tahap filtrasi dengan menggunakan vacuum filter untuk
menghilangkan partikel-partikel karbon aktif dan anorganik lainnya. Tahap selanjutnya dilakukan
penukaran ion untuk pelunakan sirup kemudian sirup maltosa murni hasil penukaran ion yang ingin
dibentuk dalam produk kristal maltosa dipekatkan di dalam alat vacuum evaporator single effect.
Tahapan proses pada PT Tainesia Jaya tidak berbeda jauh dengan sejumlah industri sirup high-
maltose di Amerika dengan metode hidrolisis secara enzimatik yang melalui beberapa tahap, yaitu
likuifikasi, filtrasi (pemisahan), dan sakarifikasi. Pada tahap likuifikasi dilakukan dengan penambahan
enzim α-amilase dalam sebuah alat pemanas seperti autoklaf pada suhu 105-110oC selama 5-7 menit
atau pada suhu 95oC selama 1-2 jam. Pada tahap ini enzim α-amilase bekerja memotong-motong
polisakarida menjadi oligosakarida sedangkan pemanasan tinggi dilakukan juga untuk memisahkan
protein dalam pati. Selanjutnya pada tahap pemisahan dilakukan pemisahan protein yang terkoagulasi
oleh panas dari larutan hidrolisat pati. Larutan hidrolisat pati yang telah mengandung sejumlah
oligosakarida masuk ke tahap sakarifikasi yang menggunakan enzim β-amilase dan pullulanase pada
suhu 40-60oC dengan pH 4-6 untuk menghasilkan sirup high-maltose dengan nilai DE sekitar 50-55
untuk substrat pati beras (Chang 2009). Sirup high-maltose sangat cocok diaplikasikan pada sejumlah
produk makanan, seperti permen dan es krim dengan kualitas yang tinggi (Hii 2012).
Untuk industri yang ingin menghasilkan produk utama dari hidrolisat pati palma berupa β-limit
dekstrin dapat menggunakan enzim β-amilase terlebih dahulu lalu pullulanase. Namun, produksi β-
limit dekstrin dengan metode tersebut juga akan menghasilkan produk samping berupa maltosa.
Berdasarkan hasil penelitian ini pati sagu baruk 1 dan sagu baruk 2 merupakan pati palma yang paling
potensial sebagai substrat karena kedua pati tersebut memiliki kandungan β-limit dekstrin yang paling
tinggi dibandingkan kelima pati palma lainnya.β-limit dekstrin umumnya dibutuhkan untuk digunakan
sebagai bahan tambahan pada berbagai produk di industri. Karena β-limit dekstrin memiliki sifat free

33
sugar, bioadesif yang baik, viskositas yang tinggi, serta laju browning yang rendah sehingga cocok
digunakan sebagai pengental rendah kalori untuk produk-produk diabetik dan bahan perekat alami
pada kertas. β-limit dekstrin juga dapat diaplikasikan sebagai bulking agent, texture providers,
pembentuk lapisan film, dan freeze-control agent (Tester 2011).
Untuk industri yang ingin menghasilkan produk maltooligosakarida sebagai produk hidrolisat
pati palma dapat digunakan enzim pullulanase saja. Maltooligosakarida umumnya digunakan sebagai
bahan pengental karena memiliki viskositas yang tinggi. Pembuatan produk hidrolisat ini disesuaikan
kembali dengan kebutuhan industri produk hilir yang menggunakan produk hidrolisat
maltooligosakarida sebagai bahan tambahannya. Untuk produk yang tidak memerlukan bahan
pengental dengan tingkat viskositas yang tinggi seperti industri produk-produk diabetik, sambel, dan
kecap dapat dibuat maltooligosakarida dengan proses hidrolisis pati palma dengan pullulanase hingga
tingkat hidrolisis 100% karena dari hasil penelitian ini pada tingkat hidrolisis 100% nilai DP rata-rata
ketujuh pati palma yang digunakan berkisar antara 13,9-20,8 sehingga karakteristik
maltooligosakarida yang dihasilkan akan memiliki daya kekentalan cukup rendah. Namun, apabila
industri hilir yang membutuhkan bahan pengental dengan daya kekentalan yang tinggi seperti industri
lem dan krim kosmetika proses hidrolisis yang digunakan untuk menghasilkan maltooligosakarida
cukup dengan tingkat hidrolisis 40% karena dari hasil penelitian ini pada tingkat hidrolisis tersebut
ketujuh pati palma yang diguanakan memiliki nilai DP yang lebih tinggi yang menyebabkan produk
hidrolisat lebih viskos (kental). Pati palma yang paling potensial digunakan untuk menghasilkan
produk hidrolisat berupa maltooligosakarida dengan daya viskositas yang paling tinggi adalah pati
aren, Caryota mitis, dan sagu komersial.

34
V. PENUTUP
5.1 Kesimpulan
Pati palma termasuk tipe pati normal dengan kandungan amilosa berkisar antara 20,1-26,7%
dengan kandungan amilosa tertinggi pada sagu rumbia. Hidrolisis pati palma dengan β-amilase terjadi
pada ikatan α-1,4-glukosidik menghasilkan maltosa dan residu β-limit dekstrin yang merupakan
bagian rantai percabangan pada ikatan α-1,6-glukosidik yang tidak terhidrolisis oleh β-amilase.
Adapun kandungan β-limit dekstrin yang dihasilkan dari ketujuh pati palma yang digunakan berkisar
antara 22,2–38,8% dari total pati. Hidrolisis pati palma dengan pullulanase terjadi pada ikatan
percabangan pada gugus amilopektin, yaitu pada ikatan α-1,6-glukosidik sehingga menghasilkan
produk yang memiliki rantai-rantai lurus yang cukup pendek dan memiliki bobot molekul yang lebih
rendah. Produk yang dihasilkan ini berupa Short-Chains Amylose (SCA) atau maltooligosakarida.
Berdasarkan hasil hidrolisis yang dilakukan diperoleh nilai DP rata-rata pada tingkat hidrolisis 100%
untuk masing-masing pati palma berkisar antara 13,9-20,8 dan nilai DE rata-rata berkisar antara 4,8–
7,2.
Hidrolisis dengan menggunakan kedua enzim tersebut yang ditambahkan secara suksesif akan
menghasilkan kerja enzim yang simultan dalam memecah rantai-rantai pada ikatan α-1,4-glukosidik
dan α-1,6-glukosidik sehingga dapat menghasilkan produk hidrolisat pati yang memiliki rantai
pendek-pendek berupa maltosa. Berdasarkan hasil hidrolisis masing-masing pati palma yang
dilakukan oleh β-amilase dan pullulanase ini diperoleh nilai DP rata-rata pada tingkat hidrolisis 100%
berkisar antara 1,7–2,9 dengan nilai DE rata-rata berkisar antara 35,1–57,3 yang menunjukkan produk
hidrolisat pati mengandung campuran maltosa, maltotriosa, dan glukosa.
Berdasarkan nilai DP dan DE yang dihasilkan hidrolisat pati dari tujuh pati palma yang
digunakan oleh masing-masing enzim terlihat adanya keragaman yang diakibatkan oleh keragaman
kandungan fraksi amilopektin yang menyusun masing-masing pati palma. Keragaman hasil ini dapat
dimanfaatkan oleh industri-industri hidrolisat pati untuk menghasilkan produk hidrolisat sesuai
kebutuhan dan kegunaan yang diinginkan. Dari hasil-hasil yang diperoleh dapat disimpulkan bahwa
berbagai jenis pati palma berpotensi sebagai sumber pati alami untuk diolah menjadi hidrolisat pati
yang dapat dimanfaatkan pada berbagai bidang aplikasi terutama bidang pangan.
5.2 Saran
Perlu dilakukan kajian lebih lanjut mengenai produksi hidrolisat enzimatis dari pati palma
supaya nilai DP dan DE yang diketahui dari hasil penelitian ini dapat diterapkan pada pengolahan di
industri-industri hidrolisat pati.

35
DAFTAR PUSTAKA
Aiyer PV. 2005. Amylases and their applications. Afr Biotechnol 4 (13) : 1525-1529.
Alamendah. 2009. Jenis-jenis Palem Arecaceae di Indonesia. http://Alamendah.wordpress.com/2009/12/12/jenis-jenis-palem-arecaceae-di-indonesia/ [29 Desember 2011].
AOAC. 1995. Official Methods of Analysis 16th ed. AOAC International , Gaithersbug, Maryland. Banks W dan Greenwood CT. 1975. Starch and Its Components. Helsted Press, John Willey and
Sons, New York. Bertoft E, Piyachomkwan K, Chatakanonda P, dan Sriroth K. 2008. Internal unit chain composition in
amylopectins. Carbohydr Polym 74 : 527-543. Bird R dan Hopkins RH. 1953. The Mechanism of β-amylase action. Biochem 56 : 140-146. [BPS] Badan Pusat Statistik. 2010. Data Luas Panen, Produktivitas, dan Produksi Tanaman Sagu
Seluruh Provinsi Indonesia. Cai L dan Shi YC. 2010. Structure and digestibility of crystalline short-chain amylose from
debranched waxy wheat, waxy maize, and waxy potato starches. Carbohydr Polym 79 : 1117-1123.
Chang SW, Chang WH, Lee MR. 2009. Simultaneous production of trehalose, bioethanol, and high-
protein product from rice by an enzymatic process. Agric Food Chem 30 : 1-7. Dubois M, Gilles KA, Hamilton JK, Rebers PA, dan Smith F. 1956. Colorimetric method for
determination of sugar and related substances. Anal Chem 28 (3) : 350-356. Hartanto, Adhi P. 2005. Proses Pembuatan Sirup Maltosa, Fruktosa, dan Glukosa di PT Tainesia Jaya
Wonogiri [skripsi]. Fakultas Teknik Universitas Negeri Surakarta, Surakarta. Haryanto B. 1988. Kualitias pati sagu asal bogor, riau, dan serawak. Majalah Insinyur Indonesia,
Jakarta. Haryanto B dan Pangloli P. 1992. Potensi dan Pemanfaatan Sagu. Kanisius, Yogyakarta.
Hii SL, Tan JS, Ling TC, dan Arif A. 2012. Pullulanase:role in starch hydrolysis and potential industrial applications. Enzyme Res 2012:1-14.
Hizukuri S, Takeda Y, Yasuda M, dan Suzuki A. 1981. Multi-branched nature of amylose and the
action of debranching enzymes. Carbohydr Res 94: 205-213. Hizukuri S. 1986. Polymodal distribution of the chain lengths of amylopectin and its significance.
Carbohydr Res 147: 342-347. Hustiany R. 2006. Modifikasi Asilasi dan Suksinilasi Pati Tapioka sebagai Bahan Enkapsulasi
Komponen Flavor [disertasi]. Departemen Teknologi Industri Pertanian, Institut Pertanian Bogor, Bogor.
Jading A, Eduard T, Paulus P, dan Sarman G. 2011. Karakteristik fisiko-kimia pati sagu hasil
pengeringan secara fluidisasi menggunakan alat pengering cross flow fluidized bed bertenaga surya dan biomassa. J Teknol Pert Universitas Negeri Papua 13 (3) : 155-164.

36
Kainuma K. 1995. The Enzymatic Process. Gordon and Breach Science Publisher, New York.
Kearsley MW dan Dziedzic SZ. 1995. Handbook Of Starch Hydrolysis Product And Their Derivatives First Edition. Great Britain by University Press, Cambridge.
Kennedy JF, Knill CJ, dan Taylor DW. 1995. Maltodextrins. Di dalam Kearsley MW dan Dziedzic
SZ. Handbook Of Starch Hydrolysis Product And Their Derivatives. Blackie Academic & Professional, London.
Lehninger AL. 1982. Dasar-Dasar Biokimia Jilid I. Erlangga, Jakarta.
Matanubun H dan Maturbongs L. 2005. Sago palm potential, biodiversity, and socio-cultural concideration for industrial sago palm plantation at south sorong papua, indonesia. The Eight International Sago Symposium in Jayapura, Indonesia.
Miftahorachman B. 2009. Potensi sagu baruk (Arenga microcarpa) sebagai sumber pangan. Warta
Penelitian dan Pengembangan 15 (3) : 14-16. Nilsson GS, Richardson S, Huber A, Torto N, Laurell T, dan Gorton L. 2001. Microdialysis clean-up
and sampling in enzyme-based methods for the characterisation of starch. Carbohydr Polym 46: 59-68.
Patel S dan Goyal A 2010. Functional oligosaccharides: production, properties, and applications.
Microbiol Biotechnol 27 (5) : 1119-1128. Poliana J dan Mac CAP. 2007 Industrial enzymes: structure, function, and applications. Dordrecht:
Springer Hal: 24. Purwantana B dan Bintoro N. 2008. Kajian kinerja mesin ekstraksi tipe ulir pada proses pembuatan
pati aren (Arenga pinnata). Makalah pada Seminar Nasional Teknik Pertanian, 18-19 November 2008, Universitas Negeri Yogyakarta, Yogyakarta.
Sadikin M. 2002. Biokimia Enzim. Widya Medika, Jakarta.
Said ZA. 2008. Produksi gula dari jagung dengan proses enzimatik secara fermentasi kultur padat. Teknologi Proses 7 (2) : 141-150.
Sinaga K. 2004. Hidrolisis Pati Umbi-umbian Asli Indonesia dengan β-amilase dan Pullulanase
[skripsi]. Departemen Teknologi Industri Pertanian, Institut Pertanian Bogor, Bogor. Srichuwong S, Sunarti TC, Mishima T, dan Hisamatsu M. 2005. Starches from diferrent botanical
sources I : contribution of amylopectin fine structure to thermal properties and enzyme digestibility. Carbohydr Polym 60 : 529-538.
Stoddard FL. 1999. Survey of Starch Particle Size Distribution on Wheat and Related Species.
Academic Press Inc, New York. Sunanto H. 1993. Aren Budidaya dan Multiguna. Kanisius Press, Yogyakarta.
Sunarti TC, Nunome T, Yoshio N, dan Hisamatsu M. 2001. Study on outer chains released from amylopectin between immobilized and free debranching enzymes. Appl Glycosci 48 (1) : 1-10.
Tester RF dan Qi X. 2011. β-limit dextrin properties and applications. Food Hydrocolloid Res 25 (8) :
1899-1903. Thomas DJ dan Atwell WA. 1999. Starches : Practical for the Food Industry. Eagan Press, USA.
Winarno. 2010. Enzim Pangan. Gramedia, Jakarta.

37
LAMPIRAN
38
Lampiran 1. Jenis-jenis pati palma yang digunakan
Kode A1a Sagu baruk 1 (Arenga microcarpa 1)
Kode A1b Sagu baruk 2 (Arenga microcarpa 2)
Kode A1c Sagu baruk 3 (Arenga microcarpa 3)
Kode A2a Sagu rumbia (Metroxylon sp.)
Kode A2b Sagu komersial
Kode A3 Aren (Arenga pinnata)
Kode A4 Caryota mitis
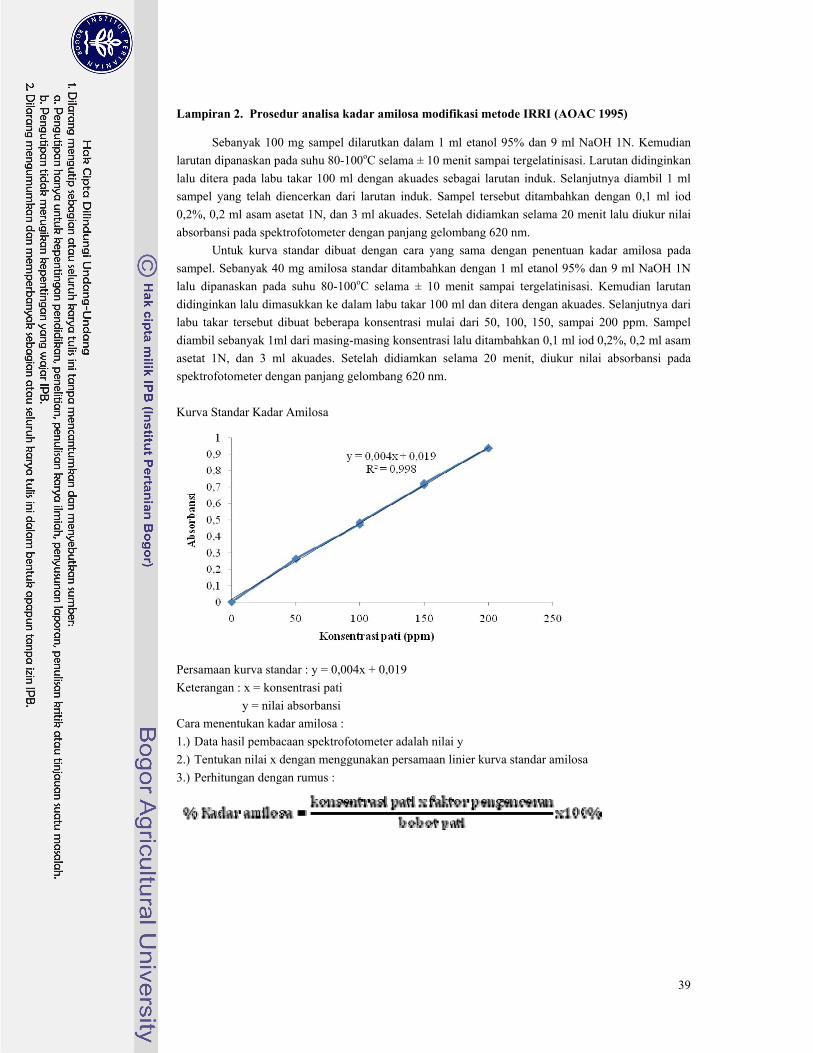
39
Lampiran 2. Prosedur analisa kadar amilosa modifikasi metode IRRI (AOAC 1995)
Sebanyak 100 mg sampel dilarutkan dalam 1 ml etanol 95% dan 9 ml NaOH 1N. Kemudian
larutan dipanaskan pada suhu 80-100oC selama ± 10 menit sampai tergelatinisasi. Larutan didinginkan
lalu ditera pada labu takar 100 ml dengan akuades sebagai larutan induk. Selanjutnya diambil 1 ml
sampel yang telah diencerkan dari larutan induk. Sampel tersebut ditambahkan dengan 0,1 ml iod
0,2%, 0,2 ml asam asetat 1N, dan 3 ml akuades. Setelah didiamkan selama 20 menit lalu diukur nilai
absorbansi pada spektrofotometer dengan panjang gelombang 620 nm.
Untuk kurva standar dibuat dengan cara yang sama dengan penentuan kadar amilosa pada
sampel. Sebanyak 40 mg amilosa standar ditambahkan dengan 1 ml etanol 95% dan 9 ml NaOH 1N
lalu dipanaskan pada suhu 80-100oC selama ± 10 menit sampai tergelatinisasi. Kemudian larutan
didinginkan lalu dimasukkan ke dalam labu takar 100 ml dan ditera dengan akuades. Selanjutnya dari
labu takar tersebut dibuat beberapa konsentrasi mulai dari 50, 100, 150, sampai 200 ppm. Sampel
diambil sebanyak 1ml dari masing-masing konsentrasi lalu ditambahkan 0,1 ml iod 0,2%, 0,2 ml asam
asetat 1N, dan 3 ml akuades. Setelah didiamkan selama 20 menit, diukur nilai absorbansi pada
spektrofotometer dengan panjang gelombang 620 nm.
Kurva Standar Kadar Amilosa
Persamaan kurva standar : y = 0,004x + 0,019
Keterangan : x = konsentrasi pati
y = nilai absorbansi
Cara menentukan kadar amilosa :
1.) Data hasil pembacaan spektrofotometer adalah nilai y
2.) Tentukan nilai x dengan menggunakan persamaan linier kurva standar amilosa
3.) Perhitungan dengan rumus :
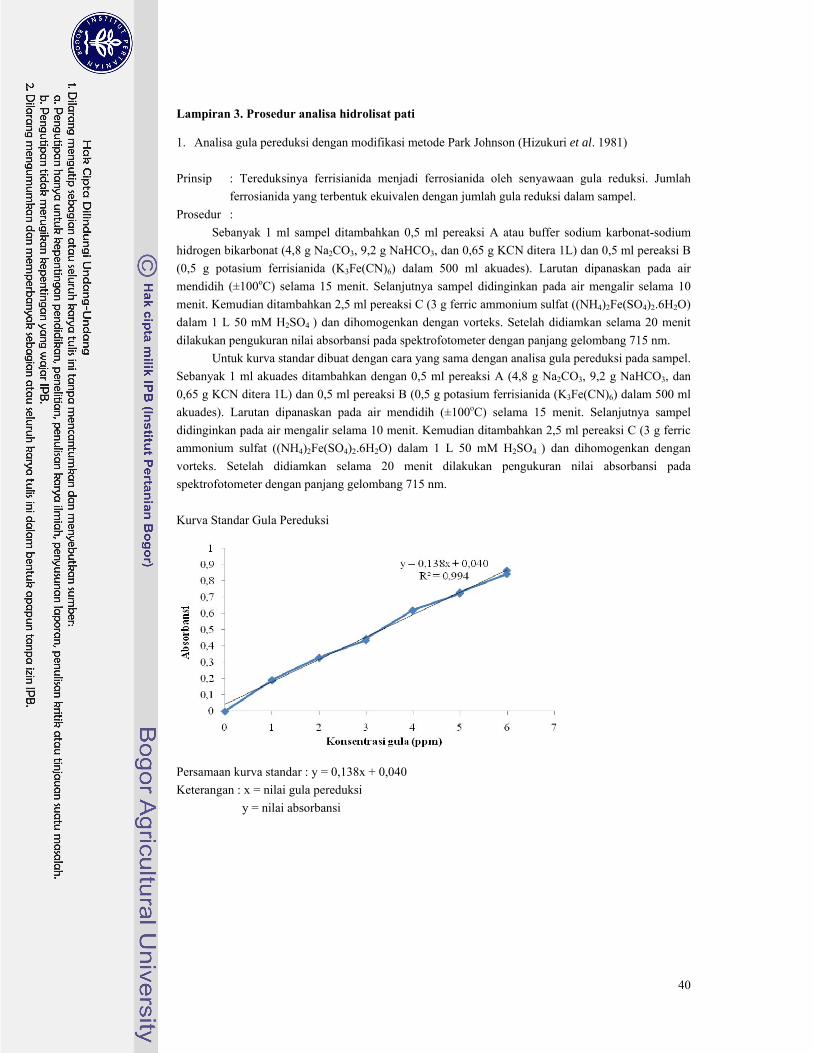
40
Lampiran 3. Prosedur analisa hidrolisat pati
1. Analisa gula pereduksi dengan modifikasi metode Park Johnson (Hizukuri et al. 1981)
Prinsip : Tereduksinya ferrisianida menjadi ferrosianida oleh senyawaan gula reduksi. Jumlah
ferrosianida yang terbentuk ekuivalen dengan jumlah gula reduksi dalam sampel.
Prosedur :
Sebanyak 1 ml sampel ditambahkan 0,5 ml pereaksi A atau buffer sodium karbonat-sodium
hidrogen bikarbonat (4,8 g Na2CO3, 9,2 g NaHCO3, dan 0,65 g KCN ditera 1L) dan 0,5 ml pereaksi B
(0,5 g potasium ferrisianida (K3Fe(CN)6) dalam 500 ml akuades). Larutan dipanaskan pada air
mendidih (±100oC) selama 15 menit. Selanjutnya sampel didinginkan pada air mengalir selama 10
menit. Kemudian ditambahkan 2,5 ml pereaksi C (3 g ferric ammonium sulfat ((NH4)2Fe(SO4)2.6H2O)
dalam 1 L 50 mM H2SO4 ) dan dihomogenkan dengan vorteks. Setelah didiamkan selama 20 menit
dilakukan pengukuran nilai absorbansi pada spektrofotometer dengan panjang gelombang 715 nm.
Untuk kurva standar dibuat dengan cara yang sama dengan analisa gula pereduksi pada sampel.
Sebanyak 1 ml akuades ditambahkan dengan 0,5 ml pereaksi A (4,8 g Na2CO3, 9,2 g NaHCO3, dan
0,65 g KCN ditera 1L) dan 0,5 ml pereaksi B (0,5 g potasium ferrisianida (K3Fe(CN)6) dalam 500 ml
akuades). Larutan dipanaskan pada air mendidih (±100oC) selama 15 menit. Selanjutnya sampel
didinginkan pada air mengalir selama 10 menit. Kemudian ditambahkan 2,5 ml pereaksi C (3 g ferric
ammonium sulfat ((NH4)2Fe(SO4)2.6H2O) dalam 1 L 50 mM H2SO4 ) dan dihomogenkan dengan
vorteks. Setelah didiamkan selama 20 menit dilakukan pengukuran nilai absorbansi pada
spektrofotometer dengan panjang gelombang 715 nm.
Kurva Standar Gula Pereduksi
Persamaan kurva standar : y = 0,138x + 0,040
Keterangan : x = nilai gula pereduksi
y = nilai absorbansi
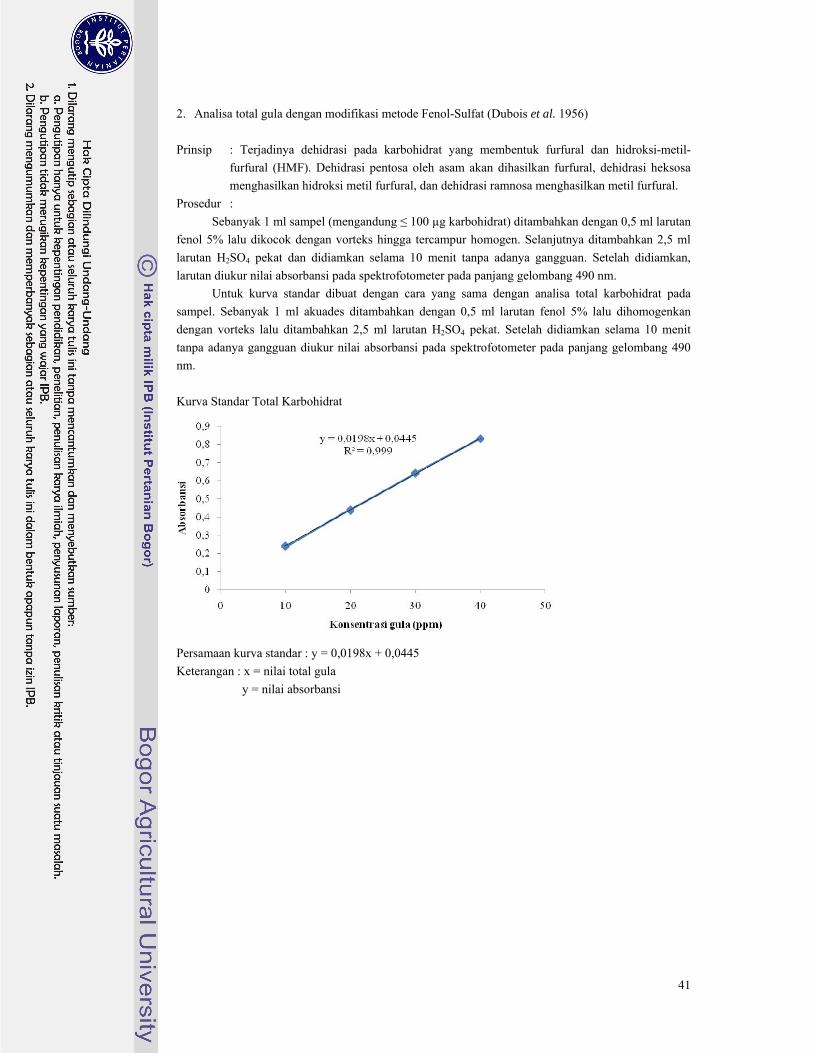
41
2. Analisa total gula dengan modifikasi metode Fenol-Sulfat (Dubois et al. 1956)
Prinsip : Terjadinya dehidrasi pada karbohidrat yang membentuk furfural dan hidroksi-metil-
furfural (HMF). Dehidrasi pentosa oleh asam akan dihasilkan furfural, dehidrasi heksosa
menghasilkan hidroksi metil furfural, dan dehidrasi ramnosa menghasilkan metil furfural.
Prosedur :
Sebanyak 1 ml sampel (mengandung ≤ 100 µg karbohidrat) ditambahkan dengan 0,5 ml larutan
fenol 5% lalu dikocok dengan vorteks hingga tercampur homogen. Selanjutnya ditambahkan 2,5 ml
larutan H2SO4 pekat dan didiamkan selama 10 menit tanpa adanya gangguan. Setelah didiamkan,
larutan diukur nilai absorbansi pada spektrofotometer pada panjang gelombang 490 nm.
Untuk kurva standar dibuat dengan cara yang sama dengan analisa total karbohidrat pada
sampel. Sebanyak 1 ml akuades ditambahkan dengan 0,5 ml larutan fenol 5% lalu dihomogenkan
dengan vorteks lalu ditambahkan 2,5 ml larutan H2SO4 pekat. Setelah didiamkan selama 10 menit
tanpa adanya gangguan diukur nilai absorbansi pada spektrofotometer pada panjang gelombang 490
nm.
Kurva Standar Total Karbohidrat
Persamaan kurva standar : y = 0,0198x + 0,0445
Keterangan : x = nilai total gula
y = nilai absorbansi

42
Lampiran 4. Rumus perhitungan hasil analisa hidrolisat pati
1. Perhitungan nilai β-limit dekstrin
% β-Limit Dekstrin =
2. Perhitungan nilai DE (Dextrose Equivalent) dan DP (Derajat Polimerisasi)
Nilai DE dihitung berdasarkan perbandingan nilai gula pereduksi terhadap total gula sampel
sedangkan DP dihitung berdasarkan perbandingan antara total gula dengan gula pereduksi sampel.
3. Perhitungan %tingkat hidrolisis
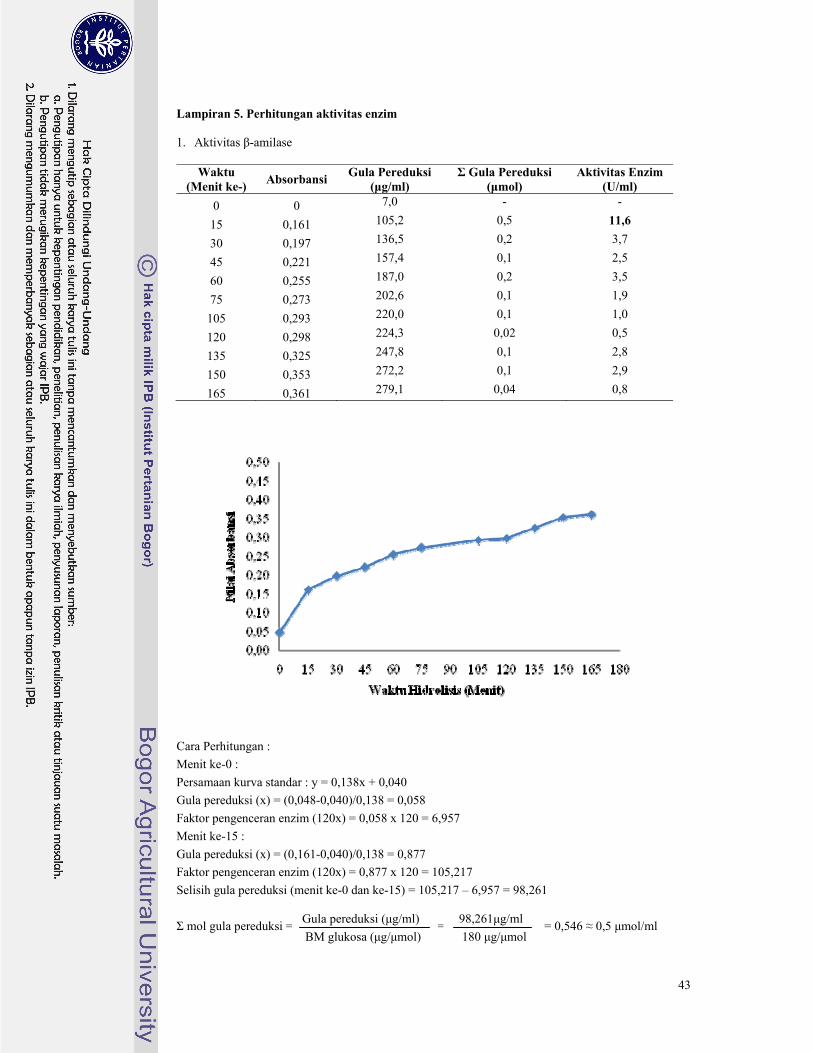
43
Lampiran 5. Perhitungan aktivitas enzim
1. Aktivitas β-amilase
Waktu (Menit ke-)
Absorbansi Gula Pereduksi
(μg/ml) Σ Gula Pereduksi
(μmol) Aktivitas Enzim
(U/ml)
0 0 7,0 - -
15 0,161 105,2 0,5 11,6
30 0,197 136,5 0,2 3,7
45 0,221 157,4 0,1 2,5
60 0,255 187,0 0,2 3,5
75 0,273 202,6 0,1 1,9
105 0,293 220,0 0,1 1,0
120 0,298 224,3 0,02 0,5
135 0,325 247,8 0,1 2,8
150 0,353 272,2 0,1 2,9
165 0,361 279,1 0,04 0,8
Cara Perhitungan :
Menit ke-0 :
Persamaan kurva standar : y = 0,138x + 0,040
Gula pereduksi (x) = (0,048-0,040)/0,138 = 0,058
Faktor pengenceran enzim (120x) = 0,058 x 120 = 6,957
Menit ke-15 :
Gula pereduksi (x) = (0,161-0,040)/0,138 = 0,877
Faktor pengenceran enzim (120x) = 0,877 x 120 = 105,217
Selisih gula pereduksi (menit ke-0 dan ke-15) = 105,217 – 6,957 = 98,261
Σ mol gula pereduksi = = = = 0,546 ≈ 0,5 μmol/ml Gula pereduksi (μg/ml)
BM glukosa (μg/μmol)
98,261μg/ml
180 μg/μmol
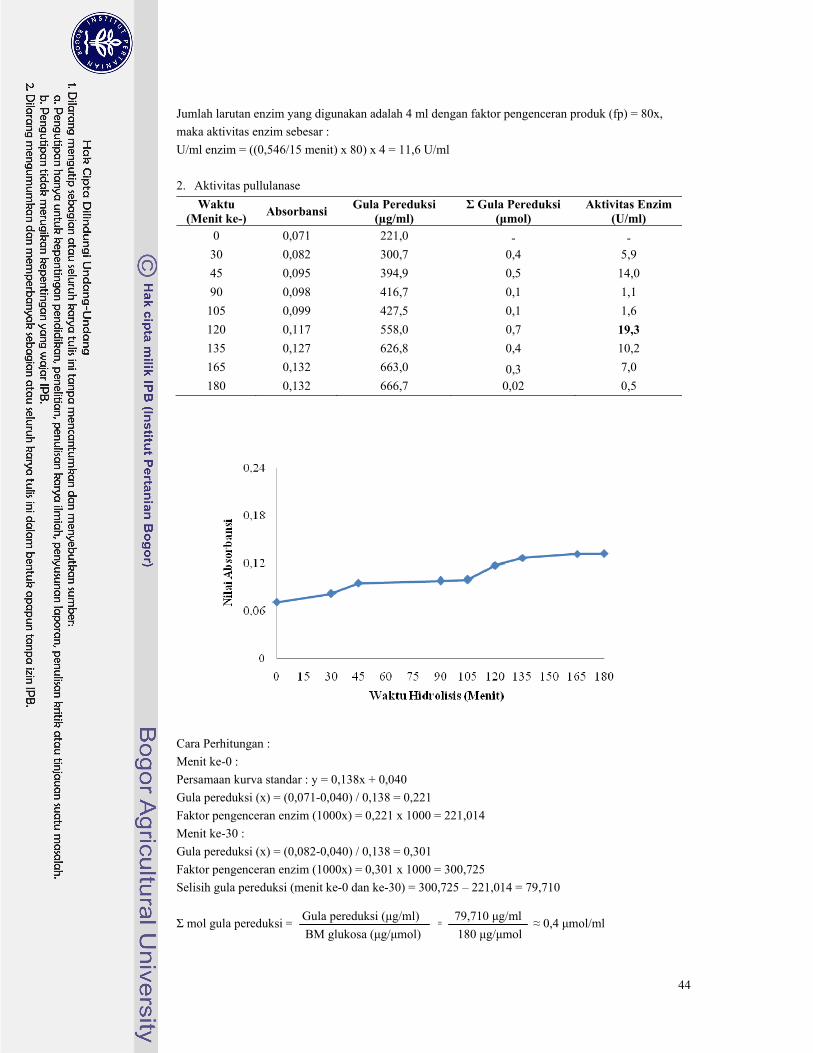
44
Jumlah larutan enzim yang digunakan adalah 4 ml dengan faktor pengenceran produk (fp) = 80x,
maka aktivitas enzim sebesar :
U/ml enzim = ((0,546/15 menit) x 80) x 4 = 11,6 U/ml
2. Aktivitas pullulanase
Waktu (Menit ke-)
Absorbansi Gula Pereduksi
(µg/ml) Σ Gula Pereduksi
(μmol) Aktivitas Enzim
(U/ml) 0 0,071 221,0 - - 30 0,082 300,7 0,4 5,9
45 0,095 394,9 0,5 14,0
90 0,098 416,7 0,1 1,1
105 0,099 427,5 0,1 1,6
120 0,117 558,0 0,7 19,3
135 0,127 626,8 0,4 10,2
165 0,132 663,0 0,3 7,0
180 0,132 666,7 0,02 0,5
Cara Perhitungan :
Menit ke-0 :
Persamaan kurva standar : y = 0,138x + 0,040
Gula pereduksi (x) = (0,071-0,040) / 0,138 = 0,221
Faktor pengenceran enzim (1000x) = 0,221 x 1000 = 221,014
Menit ke-30 :
Gula pereduksi (x) = (0,082-0,040) / 0,138 = 0,301
Faktor pengenceran enzim (1000x) = 0,301 x 1000 = 300,725
Selisih gula pereduksi (menit ke-0 dan ke-30) = 300,725 – 221,014 = 79,710
Σ mol gula pereduksi = = = = 0.546 μmol/== = 0.443 ≈ 0,4 μmol/ml Gula pereduksi (μg/ml)
BM glukosa (μg/μmol)
79,710 μg/ml
180 μg/μmol

45
Jumlah larutan enzim yang digunakan adalah 4 ml dengan faktor pengenceran produk (fp) = 100x,
maka aktivitas enzim sebesar :
U/ml enzim = ((0,443/30 menit) x 100) x 4 = 5,9 U/ml
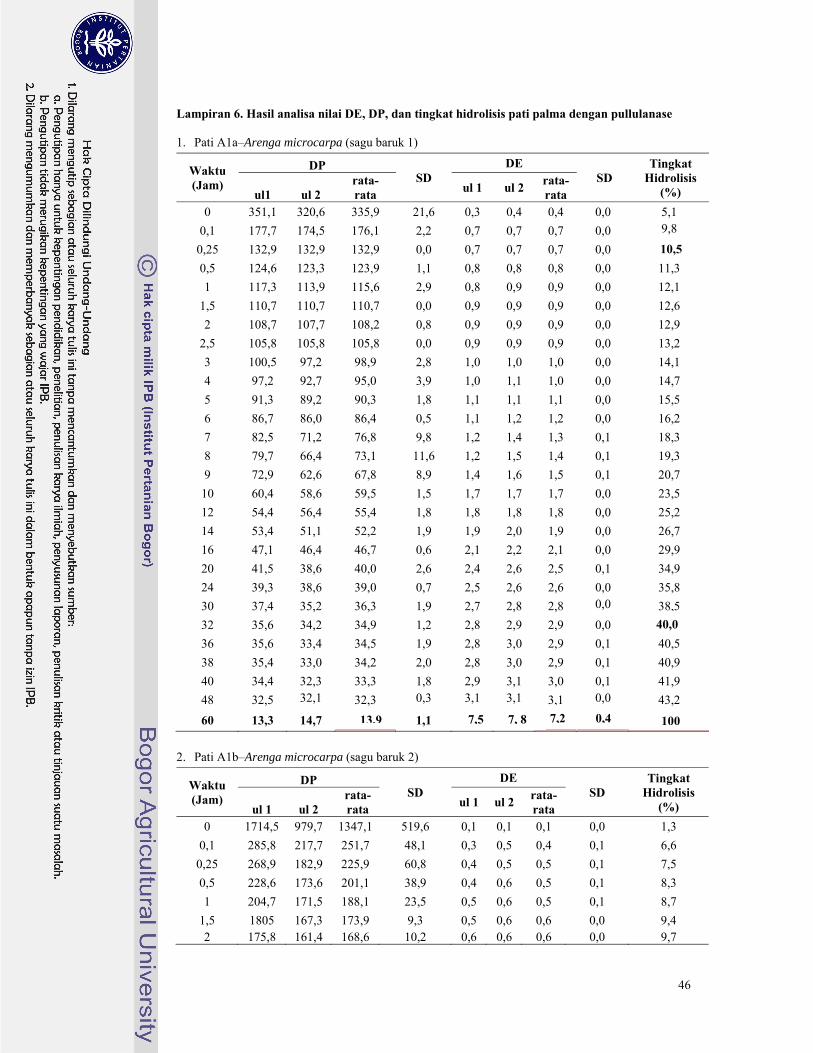
46
Lampiran 6. Hasil analisa nilai DE, DP, dan tingkat hidrolisis pati palma dengan pullulanase
1. Pati A1a–Arenga microcarpa (sagu baruk 1)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%) ul1 ul 2 rata-rata
ul 1 ul 2 rata-rata
0 351,1 320,6 335,9 21,6 0,3 0,4 0,4 0,0 5,1
0,1 177,7 174,5 176,1 2,2 0,7 0,7 0,7 0,0 9,8
0,25 132,9 132,9 132,9 0,0 0,7 0,7 0,7 0,0 10,508
0,5 124,6 123,3 123,9 1,1 0,8 0,8 0,8 0,0 11,3
1 117,3 113,9 115,6 2,9 0,8 0,9 0,9 0,0 12,1
1,5 110,7 110,7 110,7 0,0 0,9 0,9 0,9 0,0 12,6
2 108,7 107,7 108,2 0,8 0,9 0,9 0,9 0,0 12,9
2,5 105,8 105,8 105,8 0,0 0,9 0,9 0,9 0,0 13,2
3 100,5 97,2 98,9 2,8 1,0 1,0 1,0 0,0 14,1
4 97,2 92,7 95,0 3,9 1,0 1,1 1,0 0,0 14,7
5 91,3 89,2 90,3 1,8 1,1 1,1 1,1 0,0 15,5
6 86,7 86,0 86,4 0,5 1,1 1,2 1,2 0,0 16,2
7 82,5 71,2 76,8 9,8 1,2 1,4 1,3 0,1 18,3
8 79,7 66,4 73,1 11,6 1,2 1,5 1,4 0,1 19,3
9 72,9 62,6 67,8 8,9 1,4 1,6 1,5 0,1 20,7
10 60,4 58,6 59,5 1,5 1,7 1,7 1,7 0,0 23,5
12 54,4 56,4 55,4 1,8 1,8 1,8 1,8 0,0 25,2
14 53,4 51,1 52,2 1,9 1,9 2,0 1,9 0,0 26,7
16 47,1 46,4 46,7 0,6 2,1 2,2 2,1 0,0 29,9
20 41,5 38,6 40,0 2,6 2,4 2,6 2,5 0,1 34,9
24 39,3 38,6 39,0 0,7 2,5 2,6 2,6 0,0 35,8
30 37,4 35,2 36,3 1,9 2,7 2,8 2,8 0,0 38,5
32 35,6 34,2 34,9 1,2 2,8 2,9 2,9 0,0 40,047
36 35,6 33,4 34,5 1,9 2,8 3,0 2,9 0,1 40,5
38 35,4 33,0 34,2 2,0 2,8 3,0 2,9 0,1 40,9
40 34,4 32,3 33,3 1,8 2,9 3,1 3,0 0,1 41,9
48 32,5 32,1 32,3 0,3 3,1 3,1 3,1 0,0 43,2
60 13,3 14,7 13,995 1,1 7,500 6,823 7,161 0,0 100,000
2. Pati A1b–Arenga microcarpa (sagu baruk 2)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%) ul 1 ul 2 rata-rata
ul 1 ul 2 rata-rata
0 1714,5 979,7 1347,1 519,6 0,1 0,1 0,1 0,0 1,3
0,1 285,8 217,7 251,7 48,1 0,3 0,5 0,4 0,1 6,6
0,25 268,9 182,9 225,9 60,8 0,4 0,5 0,5 0,1 7,5
0,5 228,6 173,6 201,1 38,9 0,4 0,6 0,5 0,1 8,3
1 204,7 171,5 188,1 23,5 0,5 0,6 0,5 0,1 8,7
1,5 1805 167,3 173,9 9,3 0,5 0,6 0,6 0,0 9,4 2 175,8 161,4 168,6 10,2 0,6 0,6 0,6 0,0 9,7
13,9
40,0
7,2 1007,5 7, 8
10,5
0,4
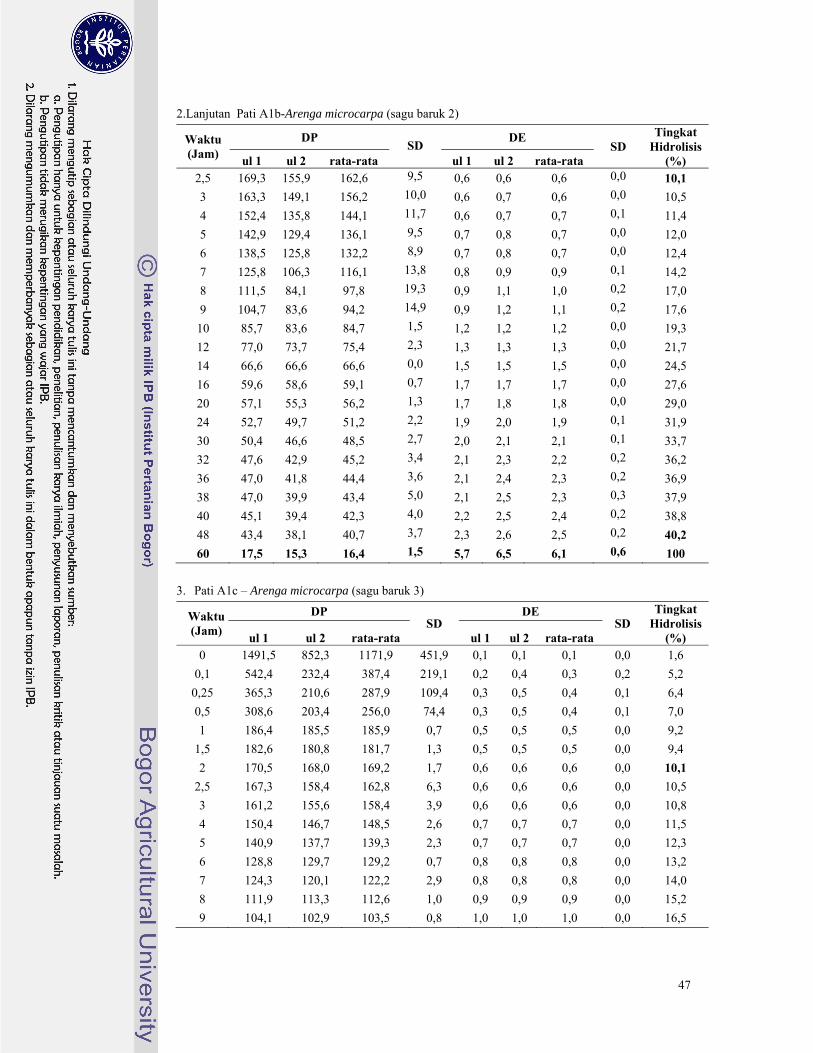
47
2.Lanjutan Pati A1b-Arenga microcarpa (sagu baruk 2)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata 2,5 169,3 155,9 162,6 9,5 0,6 0,6 0,6 0,0 10,1
3 163,3 149,1 156,2 10,0 0,6 0,7 0,6 0,0 10,5
4 152,4 135,8 144,1 11,7 0,6 0,7 0,7 0,1 11,4
5 142,9 129,4 136,1 9,5 0,7 0,8 0,7 0,0 12,0
6 138,5 125,8 132,2 8,9 0,7 0,8 0,7 0,0 12,4
7 125,8 106,3 116,1 13,8 0,8 0,9 0,9 0,1 14,2
8 111,5 84,1 97,8 19,3 0,9 1,1 1,0 0,2 17,0
9 104,7 83,6 94,2 14,9 0,9 1,2 1,1 0,2 17,6
10 85,7 83,6 84,7 1,5 1,2 1,2 1,2 0,0 19,3
12 77,0 73,7 75,4 2,3 1,3 1,3 1,3 0,0 21,7
14 66,6 66,6 66,6 0,0 1,5 1,5 1,5 0,0 24,5
16 59,6 58,6 59,1 0,7 1,7 1,7 1,7 0,0 27,6
20 57,1 55,3 56,2 1,3 1,7 1,8 1,8 0,0 29,0
24 52,7 49,7 51,2 2,2 1,9 2,0 1,9 0,1 31,9
30 50,4 46,6 48,5 2,7 2,0 2,1 2,1 0,1 33,7
32 47,6 42,9 45,2 3,4 2,1 2,3 2,2 0,2 36,2
36 47,0 41,8 44,4 3,6 2,1 2,4 2,3 0,2 36,9
38 47,0 39,9 43,4 5,0 2,1 2,5 2,3 0,3 37,9
40 45,1 39,4 42,3 4,0 2,2 2,5 2,4 0,2 38,8
48 43,4 38,1 40,7 3,7 2,3 2,6 2,5 0,2 40,2
60 17,5 15,3 16,4 1,5 5,7 6,5 6,1 0,6 100
3. Pati A1c – Arenga microcarpa (sagu baruk 3)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata 0 1491,5 852,3 1171,9 451,9 0,1 0,1 0,1 0,0 1,6
0,1 542,4 232,4 387,4 219,1 0,2 0,4 0,3 0,2 5,2
0,25 365,3 210,6 287,9 109,4 0,3 0,5 0,4 0,1 6,4
0,5 308,6 203,4 256,0 74,4 0,3 0,5 0,4 0,1 7,0
1 186,4 185,5 185,9 0,7 0,5 0,5 0,5 0,0 9,2
1,5 182,6 180,8 181,7 1,3 0,5 0,5 0,5 0,0 9,4
2 170,5 168,0 169,2 1,7 0,6 0,6 0,6 0,0 10,1
2,5 167,3 158,4 162,8 6,3 0,6 0,6 0,6 0,0 10,5
3 161,2 155,6 158,4 3,9 0,6 0,6 0,6 0,0 10,8
4 150,4 146,7 148,5 2,6 0,7 0,7 0,7 0,0 11,5
5 140,9 137,7 139,3 2,3 0,7 0,7 0,7 0,0 12,3
6 128,8 129,7 129,2 0,7 0,8 0,8 0,8 0,0 13,2
7 124,3 120,1 122,2 2,9 0,8 0,8 0,8 0,0 14,0
8 111,9 113,3 112,6 1,0 0,9 0,9 0,9 0,0 15,2
9 104,1 102,9 103,5 0,8 1,0 1,0 1,0 0,0 16,5
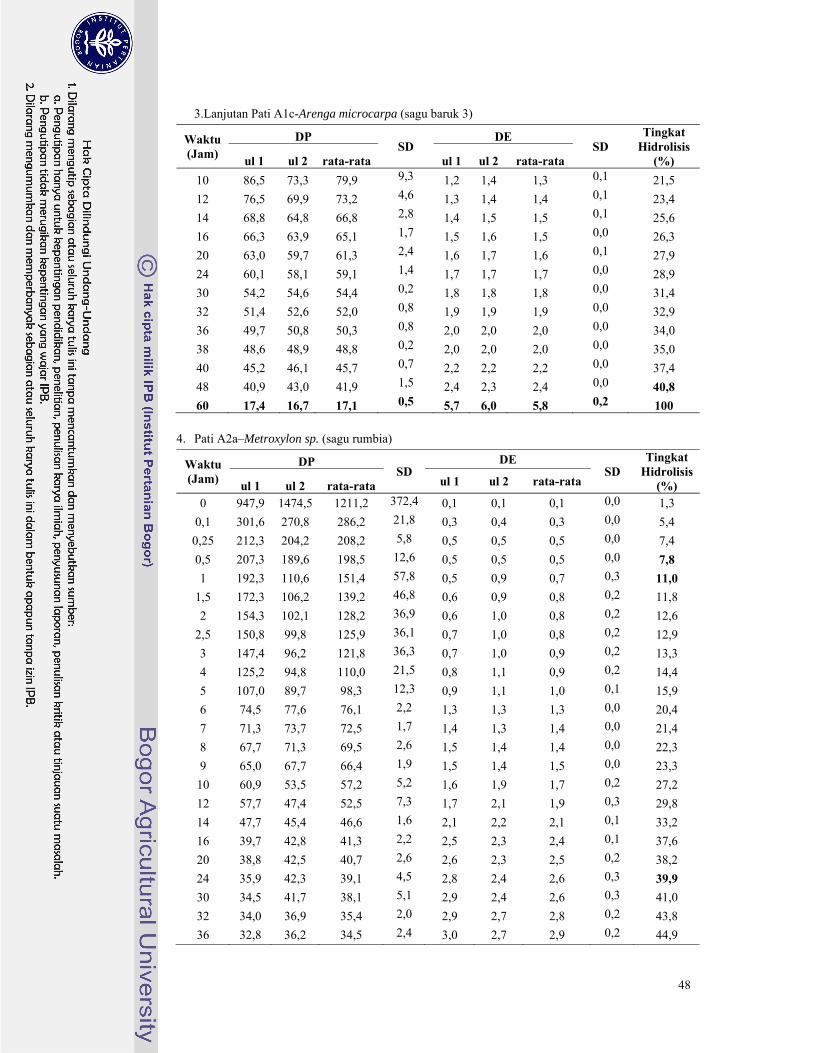
48
3.Lanjutan Pati A1c-Arenga microcarpa (sagu baruk 3)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
10 86,5 73,3 79,9 9,3 1,2 1,4 1,3 0,1 21,5
12 76,5 69,9 73,2 4,6 1,3 1,4 1,4 0,1 23,4
14 68,8 64,8 66,8 2,8 1,4 1,5 1,5 0,1 25,6
16 66,3 63,9 65,1 1,7 1,5 1,6 1,5 0,0 26,3
20 63,0 59,7 61,3 2,4 1,6 1,7 1,6 0,1 27,9
24 60,1 58,1 59,1 1,4 1,7 1,7 1,7 0,0 28,9
30 54,2 54,6 54,4 0,2 1,8 1,8 1,8 0,0 31,4
32 51,4 52,6 52,0 0,8 1,9 1,9 1,9 0,0 32,9
36 49,7 50,8 50,3 0,8 2,0 2,0 2,0 0,0 34,0
38 48,6 48,9 48,8 0,2 2,0 2,0 2,0 0,0 35,0
40 45,2 46,1 45,7 0,7 2,2 2,2 2,2 0,0 37,4
48 40,9 43,0 41,9 1,5 2,4 2,3 2,4 0,0 40,8
60 17,4 16,7 17,1 0,5 5,7 6,0 5,8 0,2 100
4. Pati A2a–Metroxylon sp. (sagu rumbia)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 947,9 1474,5 1211,2 372,4 0,1 0,1 0,1 0,0 1,3
0,1 301,6 270,8 286,2 21,8 0,3 0,4 0,3 0,0 5,4
0,25 212,3 204,2 208,2 5,8 0,5 0,5 0,5 0,0 7,4
0,5 207,3 189,6 198,5 12,6 0,5 0,5 0,5 0,0 7,8
1 192,3 110,6 151,4 57,8 0,5 0,9 0,7 0,3 11,0
1,5 172,3 106,2 139,2 46,8 0,6 0,9 0,8 0,2 11,8
2 154,3 102,1 128,2 36,9 0,6 1,0 0,8 0,2 12,6
2,5 150,8 99,8 125,9 36,1 0,7 1,0 0,8 0,2 12,9
3 147,4 96,2 121,8 36,3 0,7 1,0 0,9 0,2 13,3
4 125,2 94,8 110,0 21,5 0,8 1,1 0,9 0,2 14,4
5 107,0 89,7 98,3 12,3 0,9 1,1 1,0 0,1 15,9
6 74,5 77,6 76,1 2,2 1,3 1,3 1,3 0,0 20,4
7 71,3 73,7 72,5 1,7 1,4 1,3 1,4 0,0 21,4
8 67,7 71,3 69,5 2,6 1,5 1,4 1,4 0,0 22,3
9 65,0 67,7 66,4 1,9 1,5 1,4 1,5 0,0 23,3
10 60,9 53,5 57,2 5,2 1,6 1,9 1,7 0,2 27,2
12 57,7 47,4 52,5 7,3 1,7 2,1 1,9 0,3 29,8
14 47,7 45,4 46,6 1,6 2,1 2,2 2,1 0,1 33,2
16 39,7 42,8 41,3 2,2 2,5 2,3 2,4 0,1 37,6
20 38,8 42,5 40,7 2,6 2,6 2,3 2,5 0,2 38,2
24 35,9 42,3 39,1 4,5 2,8 2,4 2,6 0,3 39,9
30 34,5 41,7 38,1 5,1 2,9 2,4 2,6 0,3 41,0
32 34,0 36,9 35,4 2,0 2,9 2,7 2,8 0,2 43,8
36 32,8 36,2 34,5 2,4 3,0 2,7 2,9 0,2 44,9
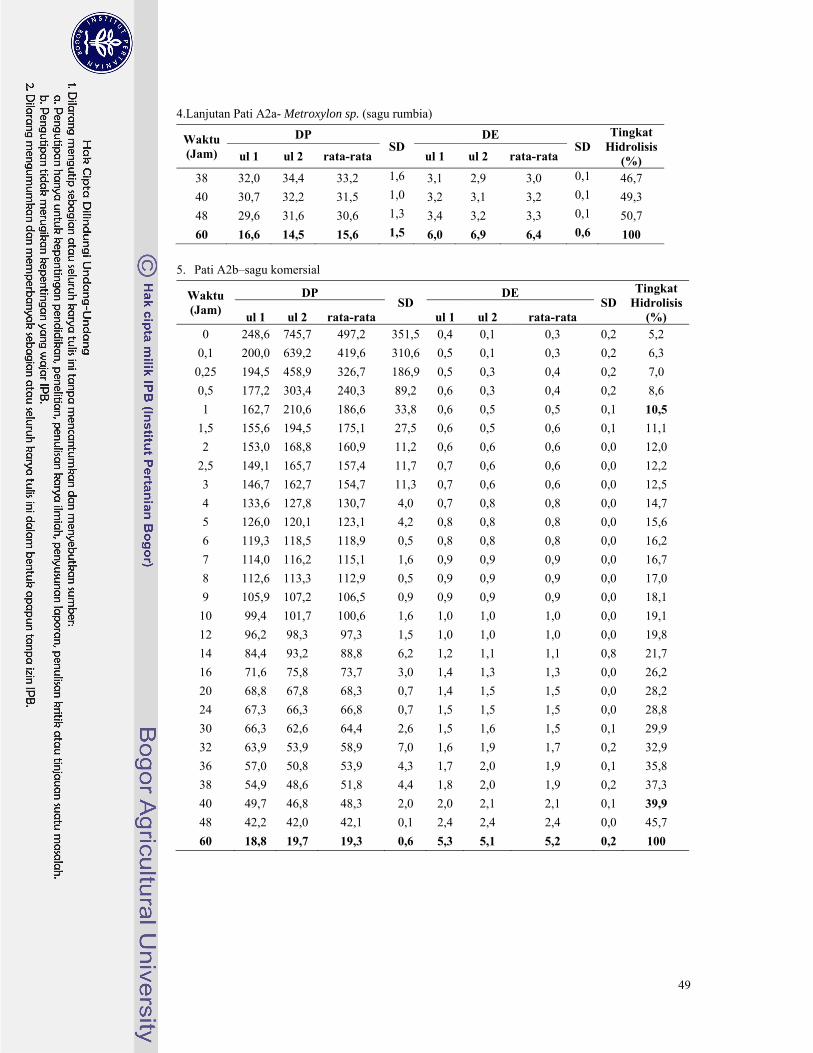
49
4.Lanjutan Pati A2a- Metroxylon sp. (sagu rumbia)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
38 32,0 34,4 33,2 1,6 3,1 2,9 3,0 0,1 46,7
40 30,7 32,2 31,5 1,0 3,2 3,1 3,2 0,1 49,3
48 29,6 31,6 30,6 1,3 3,4 3,2 3,3 0,1 50,7
60 16,6 14,5 15,6 1,5 6,0 6,9 6,4 0,6 100
5. Pati A2b–sagu komersial
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata 0 248,6 745,7 497,2 351,5 0,4 0,1 0,3 0,2 5,2
0,1 200,0 639,2 419,6 310,6 0,5 0,1 0,3 0,2 6,3
0,25 194,5 458,9 326,7 186,9 0,5 0,3 0,4 0,2 7,0
0,5 177,2 303,4 240,3 89,2 0,6 0,3 0,4 0,2 8,6
1 162,7 210,6 186,6 33,8 0,6 0,5 0,5 0,1 10,5
1,5 155,6 194,5 175,1 27,5 0,6 0,5 0,6 0,1 11,1
2 153,0 168,8 160,9 11,2 0,6 0,6 0,6 0,0 12,0
2,5 149,1 165,7 157,4 11,7 0,7 0,6 0,6 0,0 12,2
3 146,7 162,7 154,7 11,3 0,7 0,6 0,6 0,0 12,5
4 133,6 127,8 130,7 4,0 0,7 0,8 0,8 0,0 14,7
5 126,0 120,1 123,1 4,2 0,8 0,8 0,8 0,0 15,6
6 119,3 118,5 118,9 0,5 0,8 0,8 0,8 0,0 16,2
7 114,0 116,2 115,1 1,6 0,9 0,9 0,9 0,0 16,7
8 112,6 113,3 112,9 0,5 0,9 0,9 0,9 0,0 17,0
9 105,9 107,2 106,5 0,9 0,9 0,9 0,9 0,0 18,1
10 99,4 101,7 100,6 1,6 1,0 1,0 1,0 0,0 19,1
12 96,2 98,3 97,3 1,5 1,0 1,0 1,0 0,0 19,8
14 84,4 93,2 88,8 6,2 1,2 1,1 1,1 0,8 21,7
16 71,6 75,8 73,7 3,0 1,4 1,3 1,3 0,0 26,2
20 68,8 67,8 68,3 0,7 1,4 1,5 1,5 0,0 28,2
24 67,3 66,3 66,8 0,7 1,5 1,5 1,5 0,0 28,8
30 66,3 62,6 64,4 2,6 1,5 1,6 1,5 0,1 29,9
32 63,9 53,9 58,9 7,0 1,6 1,9 1,7 0,2 32,9
36 57,0 50,8 53,9 4,3 1,7 2,0 1,9 0,1 35,8
38 54,9 48,6 51,8 4,4 1,8 2,0 1,9 0,2 37,3
40 49,7 46,8 48,3 2,0 2,0 2,1 2,1 0,1 39,9
48 42,2 42,0 42,1 0,1 2,4 2,4 2,4 0,0 45,7
60 18,8 19,7 19,3 0,6 5,3 5,1 5,2 0,2 100
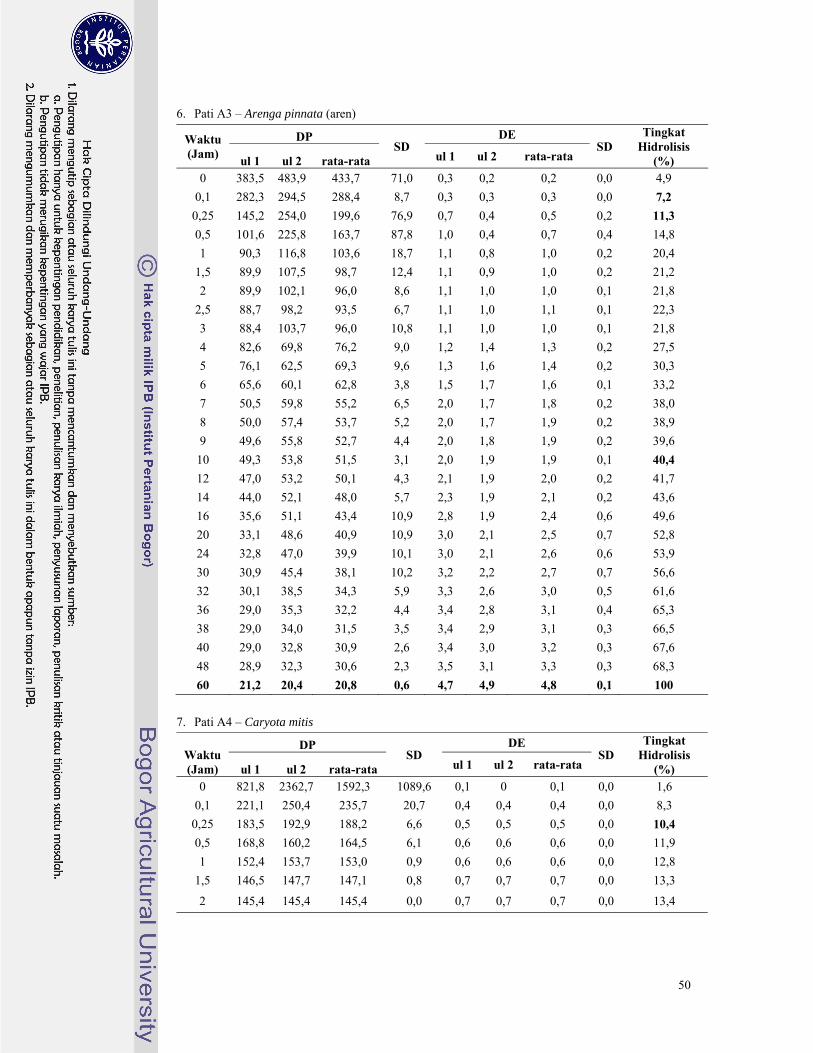
50
6. Pati A3 – Arenga pinnata (aren)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 383,5 483,9 433,7 71,0 0,3 0,2 0,2 0,0 4,9
0,1 282,3 294,5 288,4 8,7 0,3 0,3 0,3 0,0 7,2
0,25 145,2 254,0 199,6 76,9 0,7 0,4 0,5 0,2 11,3
0,5 101,6 225,8 163,7 87,8 1,0 0,4 0,7 0,4 14,8
1 90,3 116,8 103,6 18,7 1,1 0,8 1,0 0,2 20,4
1,5 89,9 107,5 98,7 12,4 1,1 0,9 1,0 0,2 21,2
2 89,9 102,1 96,0 8,6 1,1 1,0 1,0 0,1 21,8
2,5 88,7 98,2 93,5 6,7 1,1 1,0 1,1 0,1 22,3
3 88,4 103,7 96,0 10,8 1,1 1,0 1,0 0,1 21,8
4 82,6 69,8 76,2 9,0 1,2 1,4 1,3 0,2 27,5
5 76,1 62,5 69,3 9,6 1,3 1,6 1,4 0,2 30,3
6 65,6 60,1 62,8 3,8 1,5 1,7 1,6 0,1 33,2
7 50,5 59,8 55,2 6,5 2,0 1,7 1,8 0,2 38,0
8 50,0 57,4 53,7 5,2 2,0 1,7 1,9 0,2 38,9
9 49,6 55,8 52,7 4,4 2,0 1,8 1,9 0,2 39,6
10 49,3 53,8 51,5 3,1 2,0 1,9 1,9 0,1 40,4
12 47,0 53,2 50,1 4,3 2,1 1,9 2,0 0,2 41,7
14 44,0 52,1 48,0 5,7 2,3 1,9 2,1 0,2 43,6
16 35,6 51,1 43,4 10,9 2,8 1,9 2,4 0,6 49,6
20 33,1 48,6 40,9 10,9 3,0 2,1 2,5 0,7 52,8
24 32,8 47,0 39,9 10,1 3,0 2,1 2,6 0,6 53,9
30 30,9 45,4 38,1 10,2 3,2 2,2 2,7 0,7 56,6
32 30,1 38,5 34,3 5,9 3,3 2,6 3,0 0,5 61,6
36 29,0 35,3 32,2 4,4 3,4 2,8 3,1 0,4 65,3
38 29,0 34,0 31,5 3,5 3,4 2,9 3,1 0,3 66,5
40 29,0 32,8 30,9 2,6 3,4 3,0 3,2 0,3 67,6
48 28,9 32,3 30,6 2,3 3,5 3,1 3,3 0,3 68,3
60 21,2 20,4 20,8 0,6 4,7 4,9 4,8 0,1 100
7. Pati A4 – Caryota mitis
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 821,8 2362,7 1592,3 1089,6 0,1 0 0,1 0,0 1,6
0,1 221,1 250,4 235,7 20,7 0,4 0,4 0,4 0,0 8,3
0,25 183,5 192,9 188,2 6,6 0,5 0,5 0,5 0,0 10,4
0,5 168,8 160,2 164,5 6,1 0,6 0,6 0,6 0,0 11,9
1 152,4 153,7 153,0 0,9 0,6 0,6 0,6 0,0 12,8
1,5 146,5 147,7 147,1 0,8 0,7 0,7 0,7 0,0 13,3
2 145,4 145,4 145,4 0,0 0,7 0,7 0,7 0,0 13,4
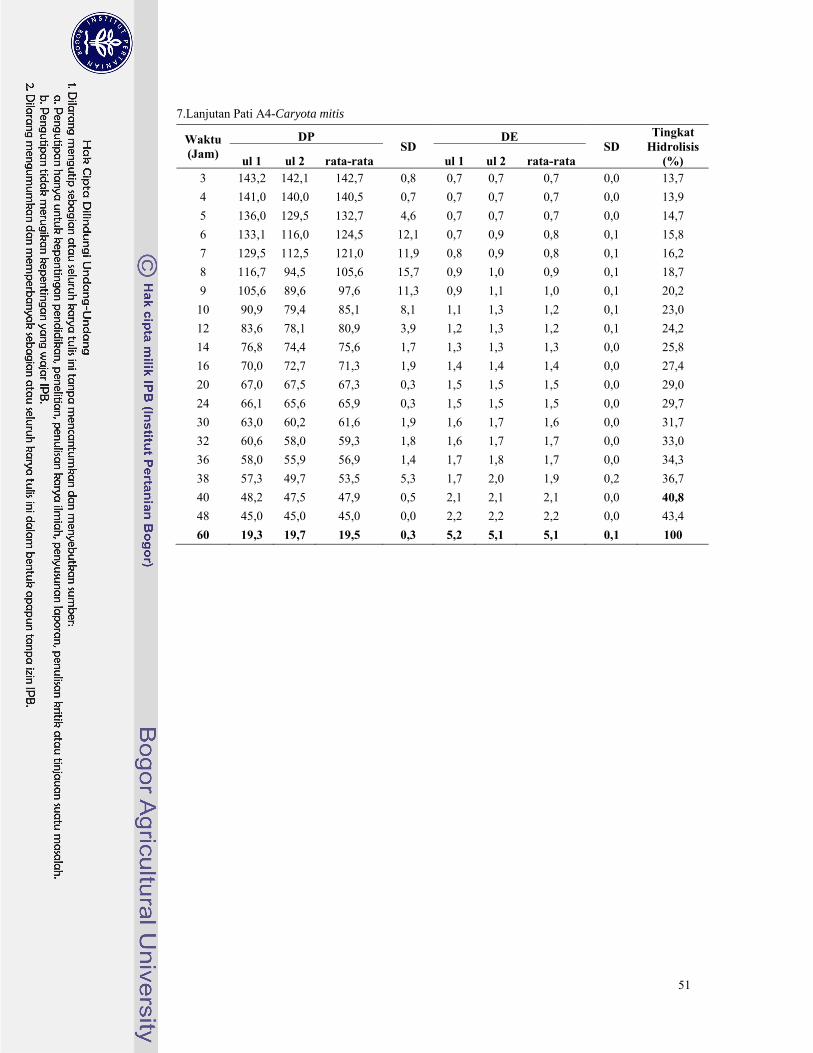
51
7.Lanjutan Pati A4-Caryota mitis
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata 3 143,2 142,1 142,7 0,8 0,7 0,7 0,7 0,0 13,7
4 141,0 140,0 140,5 0,7 0,7 0,7 0,7 0,0 13,9
5 136,0 129,5 132,7 4,6 0,7 0,7 0,7 0,0 14,7
6 133,1 116,0 124,5 12,1 0,7 0,9 0,8 0,1 15,8
7 129,5 112,5 121,0 11,9 0,8 0,9 0,8 0,1 16,2
8 116,7 94,5 105,6 15,7 0,9 1,0 0,9 0,1 18,7
9 105,6 89,6 97,6 11,3 0,9 1,1 1,0 0,1 20,2
10 90,9 79,4 85,1 8,1 1,1 1,3 1,2 0,1 23,0
12 83,6 78,1 80,9 3,9 1,2 1,3 1,2 0,1 24,2
14 76,8 74,4 75,6 1,7 1,3 1,3 1,3 0,0 25,8
16 70,0 72,7 71,3 1,9 1,4 1,4 1,4 0,0 27,4
20 67,0 67,5 67,3 0,3 1,5 1,5 1,5 0,0 29,0
24 66,1 65,6 65,9 0,3 1,5 1,5 1,5 0,0 29,7
30 63,0 60,2 61,6 1,9 1,6 1,7 1,6 0,0 31,7
32 60,6 58,0 59,3 1,8 1,6 1,7 1,7 0,0 33,0
36 58,0 55,9 56,9 1,4 1,7 1,8 1,7 0,0 34,3
38 57,3 49,7 53,5 5,3 1,7 2,0 1,9 0,2 36,7
40 48,2 47,5 47,9 0,5 2,1 2,1 2,1 0,0 40,8
48 45,0 45,0 45,0 0,0 2,2 2,2 2,2 0,0 43,4
60 19,3 19,7 19,5 0,3 5,2 5,1 5,1 0,1 100
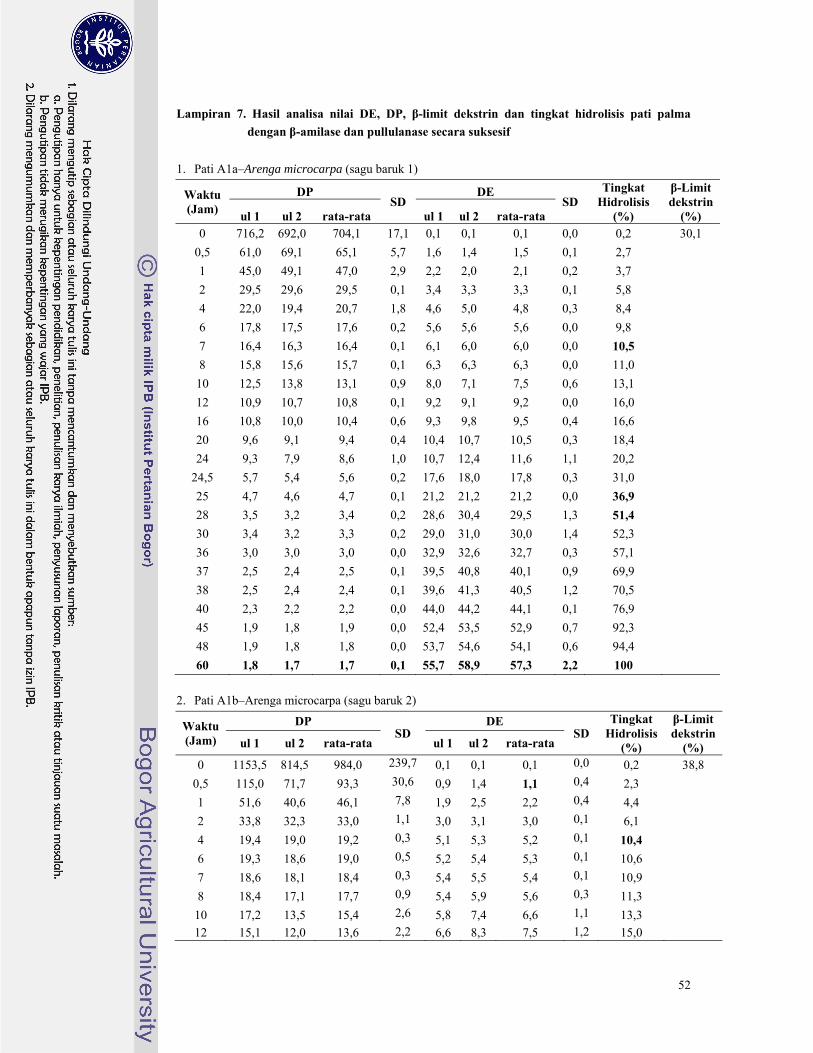
52
Lampiran 7. Hasil analisa nilai DE, DP, β-limit dekstrin dan tingkat hidrolisis pati palma
dengan β-amilase dan pullulanase secara suksesif
1. Pati A1a–Arenga microcarpa (sagu baruk 1)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-Limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata 0 716,2 692,0 704,1 17,1 0,1 0,1 0,1 0,0 0,2 30,1
0,5 61,0 69,1 65,1 5,7 1,6 1,4 1,5 0,1 2,7
1 45,0 49,1 47,0 2,9 2,2 2,0 2,1 0,2 3,7
2 29,5 29,6 29,5 0,1 3,4 3,3 3,3 0,1 5,8
4 22,0 19,4 20,7 1,8 4,6 5,0 4,8 0,3 8,4
6 17,8 17,5 17,6 0,2 5,6 5,6 5,6 0,0 9,8
7 16,4 16,3 16,4 0,1 6,1 6,0 6,0 0,0 10,5
8 15,8 15,6 15,7 0,1 6,3 6,3 6,3 0,0 11,0
10 12,5 13,8 13,1 0,9 8,0 7,1 7,5 0,6 13,1
12 10,9 10,7 10,8 0,1 9,2 9,1 9,2 0,0 16,0
16 10,8 10,0 10,4 0,6 9,3 9,8 9,5 0,4 16,6
20 9,6 9,1 9,4 0,4 10,4 10,7 10,5 0,3 18,4
24 9,3 7,9 8,6 1,0 10,7 12,4 11,6 1,1 20,2
24,5 5,7 5,4 5,6 0,2 17,6 18,0 17,8 0,3 31,0
25 4,7 4,6 4,7 0,1 21,2 21,2 21,2 0,0 36,9
28 3,5 3,2 3,4 0,2 28,6 30,4 29,5 1,3 51,4
30 3,4 3,2 3,3 0,2 29,0 31,0 30,0 1,4 52,3
36 3,0 3,0 3,0 0,0 32,9 32,6 32,7 0,3 57,1
37 2,5 2,4 2,5 0,1 39,5 40,8 40,1 0,9 69,9
38 2,5 2,4 2,4 0,1 39,6 41,3 40,5 1,2 70,5
40 2,3 2,2 2,2 0,0 44,0 44,2 44,1 0,1 76,9
45 1,9 1,8 1,9 0,0 52,4 53,5 52,9 0,7 92,3
48 1,9 1,8 1,8 0,0 53,7 54,6 54,1 0,6 94,4
60 1,8 1,7 1,7 0,1 55,7 58,9 57,3 2,2 100
2. Pati A1b–Arenga microcarpa (sagu baruk 2)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-Limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 1153,5 814,5 984,0 239,7 0,1 0,1 0,1 0,0 0,2 38,8
0,5 115,0 71,7 93,3 30,6 0,9 1,4 1,1 0,4 2,3
1 51,6 40,6 46,1 7,8 1,9 2,5 2,2 0,4 4,4
2 33,8 32,3 33,0 1,1 3,0 3,1 3,0 0,1 6,1
4 19,4 19,0 19,2 0,3 5,1 5,3 5,2 0,1 10,4
6 19,3 18,6 19,0 0,5 5,2 5,4 5,3 0,1 10,6
7 18,6 18,1 18,4 0,3 5,4 5,5 5,4 0,1 10,9
8 18,4 17,1 17,7 0,9 5,4 5,9 5,6 0,3 11,3
10 17,2 13,5 15,4 2,6 5,8 7,4 6,6 1,1 13,3
12 15,1 12,0 13,6 2,2 6,6 8,3 7,5 1,2 15,0
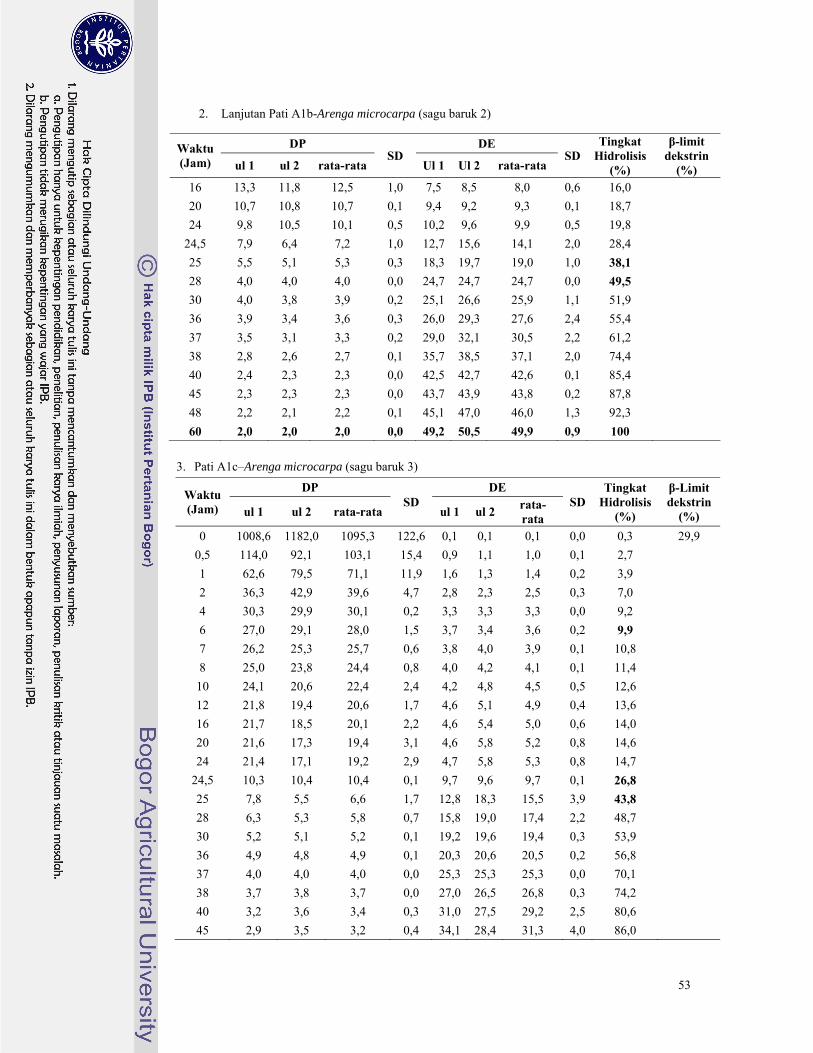
53
2. Lanjutan Pati A1b-Arenga microcarpa (sagu baruk 2)
3. Pati A1c–Arenga microcarpa (sagu baruk 3)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-Limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 1008,6 1182,0 1095,3 122,6 0,1 0,1 0,1 0,0 0,3 29,9
0,5 114,0 92,1 103,1 15,4 0,9 1,1 1,0 0,1 2,7
1 62,6 79,5 71,1 11,9 1,6 1,3 1,4 0,2 3,9
2 36,3 42,9 39,6 4,7 2,8 2,3 2,5 0,3 7,0
4 30,3 29,9 30,1 0,2 3,3 3,3 3,3 0,0 9,2
6 27,0 29,1 28,0 1,5 3,7 3,4 3,6 0,2 9,9
7 26,2 25,3 25,7 0,6 3,8 4,0 3,9 0,1 10,8
8 25,0 23,8 24,4 0,8 4,0 4,2 4,1 0,1 11,4
10 24,1 20,6 22,4 2,4 4,2 4,8 4,5 0,5 12,6
12 21,8 19,4 20,6 1,7 4,6 5,1 4,9 0,4 13,6
16 21,7 18,5 20,1 2,2 4,6 5,4 5,0 0,6 14,0
20 21,6 17,3 19,4 3,1 4,6 5,8 5,2 0,8 14,6
24 21,4 17,1 19,2 2,9 4,7 5,8 5,3 0,8 14,7
24,5 10,3 10,4 10,4 0,1 9,7 9,6 9,7 0,1 26,8
25 7,8 5,5 6,6 1,7 12,8 18,3 15,5 3,9 43,8
28 6,3 5,3 5,8 0,7 15,8 19,0 17,4 2,2 48,7
30 5,2 5,1 5,2 0,1 19,2 19,6 19,4 0,3 53,9
36 4,9 4,8 4,9 0,1 20,3 20,6 20,5 0,2 56,8
37 4,0 4,0 4,0 0,0 25,3 25,3 25,3 0,0 70,1
38 3,7 3,8 3,7 0,0 27,0 26,5 26,8 0,3 74,2
40 3,2 3,6 3,4 0,3 31,0 27,5 29,2 2,5 80,6
45 2,9 3,5 3,2 0,4 34,1 28,4 31,3 4,0 86,0
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-limit dekstrin
(%) ul 1 ul 2 rata-rata Ul 1 Ul 2 rata-rata
16 13,3 11,8 12,5 1,0 7,5 8,5 8,0 0,6 16,0
20 10,7 10,8 10,7 0,1 9,4 9,2 9,3 0,1 18,7
24 9,8 10,5 10,1 0,5 10,2 9,6 9,9 0,5 19,8
24,5 7,9 6,4 7,2 1,0 12,7 15,6 14,1 2,0 28,4
25 5,5 5,1 5,3 0,3 18,3 19,7 19,0 1,0 38,1
28 4,0 4,0 4,0 0,0 24,7 24,7 24,7 0,0 49,5
30 4,0 3,8 3,9 0,2 25,1 26,6 25,9 1,1 51,9
36 3,9 3,4 3,6 0,3 26,0 29,3 27,6 2,4 55,4
37 3,5 3,1 3,3 0,2 29,0 32,1 30,5 2,2 61,2
38 2,8 2,6 2,7 0,1 35,7 38,5 37,1 2,0 74,4
40 2,4 2,3 2,3 0,0 42,5 42,7 42,6 0,1 85,4
45 2,3 2,3 2,3 0,0 43,7 43,9 43,8 0,2 87,8
48 2,2 2,1 2,2 0,1 45,1 47,0 46,0 1,3 92,3
60 2,0 2,0 2,0 0,0 49,2 50,5 49,9 0,9 100
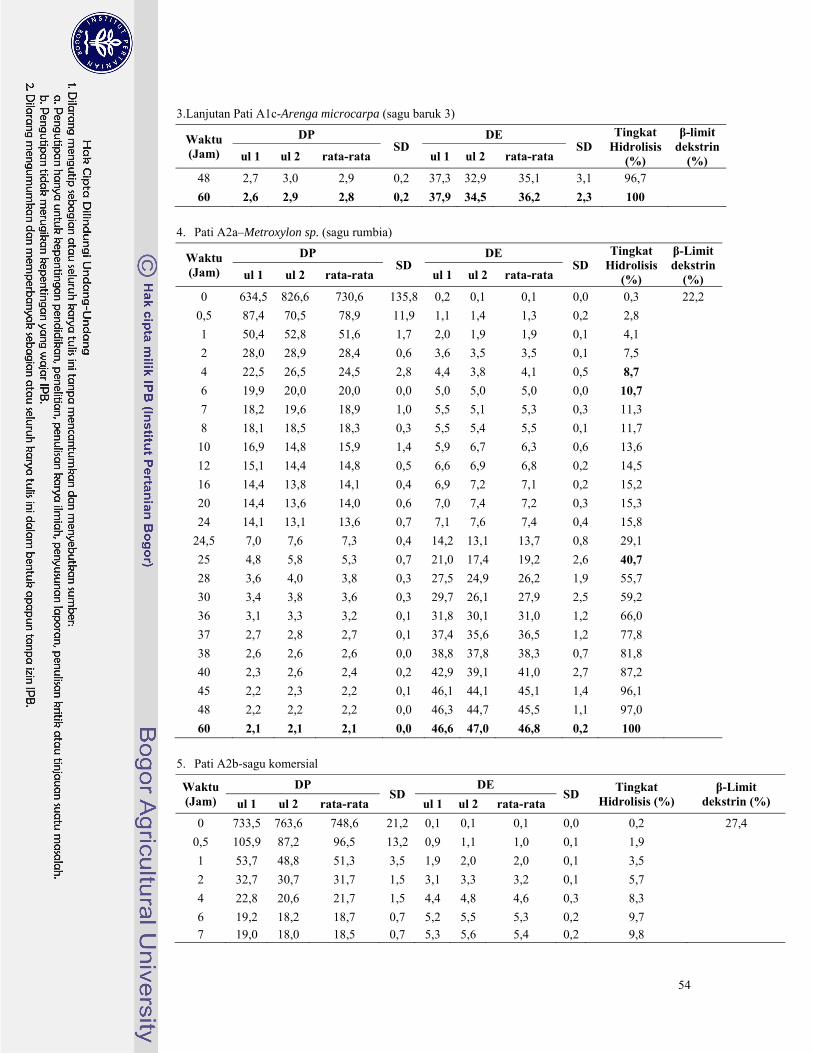
54
3.Lanjutan Pati A1c-Arenga microcarpa (sagu baruk 3)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-limit dekstrin
(%)ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
48 2,7 3,0 2,9 0,2 37,3 32,9 35,1 3,1 96,7
60 2,6 2,9 2,8 0,2 37,9 34,5 36,2 2,3 100
4. Pati A2a–Metroxylon sp. (sagu rumbia)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-Limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 634,5 826,6 730,6 135,8 0,2 0,1 0,1 0,0 0,3 22,2
0,5 87,4 70,5 78,9 11,9 1,1 1,4 1,3 0,2 2,8
1 50,4 52,8 51,6 1,7 2,0 1,9 1,9 0,1 4,1
2 28,0 28,9 28,4 0,6 3,6 3,5 3,5 0,1 7,5
4 22,5 26,5 24,5 2,8 4,4 3,8 4,1 0,5 8,7
6 19,9 20,0 20,0 0,0 5,0 5,0 5,0 0,0 10,7
7 18,2 19,6 18,9 1,0 5,5 5,1 5,3 0,3 11,3
8 18,1 18,5 18,3 0,3 5,5 5,4 5,5 0,1 11,7
10 16,9 14,8 15,9 1,4 5,9 6,7 6,3 0,6 13,6
12 15,1 14,4 14,8 0,5 6,6 6,9 6,8 0,2 14,5
16 14,4 13,8 14,1 0,4 6,9 7,2 7,1 0,2 15,2
20 14,4 13,6 14,0 0,6 7,0 7,4 7,2 0,3 15,3
24 14,1 13,1 13,6 0,7 7,1 7,6 7,4 0,4 15,8
24,5 7,0 7,6 7,3 0,4 14,2 13,1 13,7 0,8 29,1
25 4,8 5,8 5,3 0,7 21,0 17,4 19,2 2,6 40,7
28 3,6 4,0 3,8 0,3 27,5 24,9 26,2 1,9 55,7
30 3,4 3,8 3,6 0,3 29,7 26,1 27,9 2,5 59,2
36 3,1 3,3 3,2 0,1 31,8 30,1 31,0 1,2 66,0
37 2,7 2,8 2,7 0,1 37,4 35,6 36,5 1,2 77,8
38 2,6 2,6 2,6 0,0 38,8 37,8 38,3 0,7 81,8
40 2,3 2,6 2,4 0,2 42,9 39,1 41,0 2,7 87,2
45 2,2 2,3 2,2 0,1 46,1 44,1 45,1 1,4 96,1
48 2,2 2,2 2,2 0,0 46,3 44,7 45,5 1,1 97,0
60 2,1 2,1 2,1 0,0 46,6 47,0 46,8 0,2 100
5. Pati A2b-sagu komersial
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis (%)
β-Limit dekstrin (%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 733,5 763,6 748,6 21,2 0,1 0,1 0,1 0,0 0,2 27,4
0,5 105,9 87,2 96,5 13,2 0,9 1,1 1,0 0,1 1,9
1 53,7 48,8 51,3 3,5 1,9 2,0 2,0 0,1 3,5
2 32,7 30,7 31,7 1,5 3,1 3,3 3,2 0,1 5,7
4 22,8 20,6 21,7 1,5 4,4 4,8 4,6 0,3 8,3
6 19,2 18,2 18,7 0,7 5,2 5,5 5,3 0,2 9,7
7 19,0 18,0 18,5 0,7 5,3 5,6 5,4 0,2 9,8
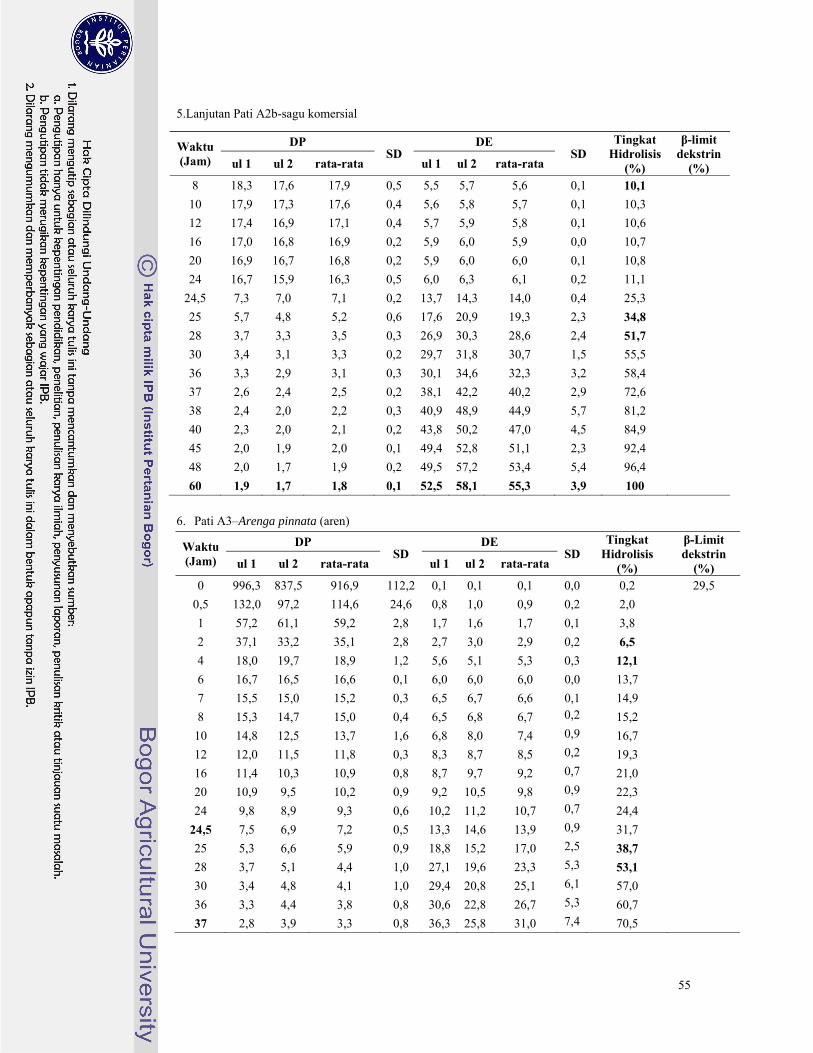
55
5.Lanjutan Pati A2b-sagu komersial
6. Pati A3–Arenga pinnata (aren)
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-Limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 996,3 837,5 916,9 112,2 0,1 0,1 0,1 0,0 0,2 29,5
0,5 132,0 97,2 114,6 24,6 0,8 1,0 0,9 0,2 2,0
1 57,2 61,1 59,2 2,8 1,7 1,6 1,7 0,1 3,8
2 37,1 33,2 35,1 2,8 2,7 3,0 2,9 0,2 6,5
4 18,0 19,7 18,9 1,2 5,6 5,1 5,3 0,3 12,1
6 16,7 16,5 16,6 0,1 6,0 6,0 6,0 0,0 13,7
7 15,5 15,0 15,2 0,3 6,5 6,7 6,6 0,1 14,9
8 15,3 14,7 15,0 0,4 6,5 6,8 6,7 0,2 15,2
10 14,8 12,5 13,7 1,6 6,8 8,0 7,4 0,9 16,7
12 12,0 11,5 11,8 0,3 8,3 8,7 8,5 0,2 19,3
16 11,4 10,3 10,9 0,8 8,7 9,7 9,2 0,7 21,0
20 10,9 9,5 10,2 0,9 9,2 10,5 9,8 0,9 22,3
24 9,8 8,9 9,3 0,6 10,2 11,2 10,7 0,7 24,4
24,5 7,5 6,9 7,2 0,5 13,3 14,6 13,9 0,9 31,7
25 5,3 6,6 5,9 0,9 18,8 15,2 17,0 2,5 38,7
28 3,7 5,1 4,4 1,0 27,1 19,6 23,3 5,3 53,1
30 3,4 4,8 4,1 1,0 29,4 20,8 25,1 6,1 57,0
36 3,3 4,4 3,8 0,8 30,6 22,8 26,7 5,3 60,7
37 2,8 3,9 3,3 0,8 36,3 25,8 31,0 7,4 70,5
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
8 18,3 17,6 17,9 0,5 5,5 5,7 5,6 0,1 10,1
10 17,9 17,3 17,6 0,4 5,6 5,8 5,7 0,1 10,3
12 17,4 16,9 17,1 0,4 5,7 5,9 5,8 0,1 10,6
16 17,0 16,8 16,9 0,2 5,9 6,0 5,9 0,0 10,7
20 16,9 16,7 16,8 0,2 5,9 6,0 6,0 0,1 10,8
24 16,7 15,9 16,3 0,5 6,0 6,3 6,1 0,2 11,1
24,5 7,3 7,0 7,1 0,2 13,7 14,3 14,0 0,4 25,3
25 5,7 4,8 5,2 0,6 17,6 20,9 19,3 2,3 34,8
28 3,7 3,3 3,5 0,3 26,9 30,3 28,6 2,4 51,7
30 3,4 3,1 3,3 0,2 29,7 31,8 30,7 1,5 55,5
36 3,3 2,9 3,1 0,3 30,1 34,6 32,3 3,2 58,4
37 2,6 2,4 2,5 0,2 38,1 42,2 40,2 2,9 72,6
38 2,4 2,0 2,2 0,3 40,9 48,9 44,9 5,7 81,2
40 2,3 2,0 2,1 0,2 43,8 50,2 47,0 4,5 84,9
45 2,0 1,9 2,0 0,1 49,4 52,8 51,1 2,3 92,4
48 2,0 1,7 1,9 0,2 49,5 57,2 53,4 5,4 96,4
60 1,9 1,7 1,8 0,1 52,5 58,1 55,3 3,9 100
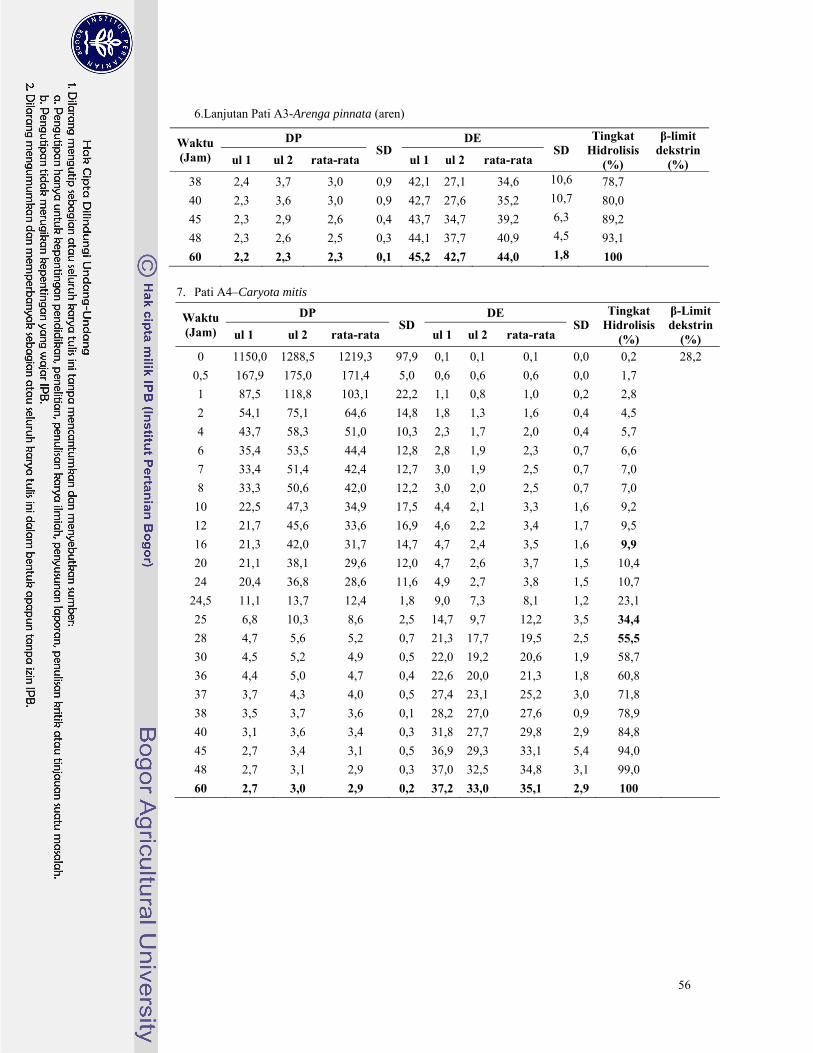
56
6.Lanjutan Pati A3-Arenga pinnata (aren)
7. Pati A4–Caryota mitis
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-Limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
0 1150,0 1288,5 1219,3 97,9 0,1 0,1 0,1 0,0 0,2 28,2
0,5 167,9 175,0 171,4 5,0 0,6 0,6 0,6 0,0 1,7
1 87,5 118,8 103,1 22,2 1,1 0,8 1,0 0,2 2,8
2 54,1 75,1 64,6 14,8 1,8 1,3 1,6 0,4 4,5
4 43,7 58,3 51,0 10,3 2,3 1,7 2,0 0,4 5,7
6 35,4 53,5 44,4 12,8 2,8 1,9 2,3 0,7 6,6
7 33,4 51,4 42,4 12,7 3,0 1,9 2,5 0,7 7,0
8 33,3 50,6 42,0 12,2 3,0 2,0 2,5 0,7 7,0
10 22,5 47,3 34,9 17,5 4,4 2,1 3,3 1,6 9,2
12 21,7 45,6 33,6 16,9 4,6 2,2 3,4 1,7 9,5
16 21,3 42,0 31,7 14,7 4,7 2,4 3,5 1,6 9,9
20 21,1 38,1 29,6 12,0 4,7 2,6 3,7 1,5 10,4
24 20,4 36,8 28,6 11,6 4,9 2,7 3,8 1,5 10,7
24,5 11,1 13,7 12,4 1,8 9,0 7,3 8,1 1,2 23,1
25 6,8 10,3 8,6 2,5 14,7 9,7 12,2 3,5 34,4
28 4,7 5,6 5,2 0,7 21,3 17,7 19,5 2,5 55,5
30 4,5 5,2 4,9 0,5 22,0 19,2 20,6 1,9 58,7
36 4,4 5,0 4,7 0,4 22,6 20,0 21,3 1,8 60,8
37 3,7 4,3 4,0 0,5 27,4 23,1 25,2 3,0 71,8
38 3,5 3,7 3,6 0,1 28,2 27,0 27,6 0,9 78,9
40 3,1 3,6 3,4 0,3 31,8 27,7 29,8 2,9 84,8
45 2,7 3,4 3,1 0,5 36,9 29,3 33,1 5,4 94,0
48 2,7 3,1 2,9 0,3 37,0 32,5 34,8 3,1 99,0
60 2,7 3,0 2,9 0,2 37,2 33,0 35,1 2,9 100
Waktu (Jam)
DP SD
DE SD
Tingkat Hidrolisis
(%)
β-limit dekstrin
(%) ul 1 ul 2 rata-rata ul 1 ul 2 rata-rata
38 2,4 3,7 3,0 0,9 42,1 27,1 34,6 10,6 78,7
40 2,3 3,6 3,0 0,9 42,7 27,6 35,2 10,7 80,0
45 2,3 2,9 2,6 0,4 43,7 34,7 39,2 6,3 89,2
48 2,3 2,6 2,5 0,3 44,1 37,7 40,9 4,5 93,1
60 2,2 2,3 2,3 0,1 45,2 42,7 44,0 1,8 100


















