
Hesperidin protects gentamicin-induced nephrotoxicity via...
11
Hesperidin protects gentamicin-induced nephrotoxicity via Nrf2/HO-1 signaling and inhibits inflammation mediated by NF-κB in rats Perumal Subramanian a, *, Ramaswamy Anandan a , Jaime Jacqueline Jayapalan b,c , Onn Haji Hashim b,c a Department of Biochemistry and Biotechnology, Faculty of Science,Annamalai University, Chidambaram 608 002, Tamil Nadu, India b Faculty of Medicine, University of Malaya, University of Malaya Centre for Proteomics Research (UMCPR), 50603 Kuala Lumpur, Malaysia c Department of Molecular Medicine, Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia ARTICLE INFO Article history: Received 20 July 2014 Received in revised form 14 December 2014 Accepted 19 December 2014 Available online ABSTRACT The defensive effects of hesperidin (HDN, a bioflavonoid) was investigated on gentamicin (GEN) provoked nephrotoxicity in rats.The expression patterns of nuclear factor (erythroid- derived 2)-like 2 (Nrf2), nuclear factor-κB (NF-κB), cyclooxygenase-2 (COX-2), tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), kidney injury molecule (KIM-1), osteopontin, heme oxygenase-1 (HO-1), inducible nitric oxide synthase (iNOS) and heat shock protein-70 (HSP- 70) were assessed to comprehend the mechanism of action of hesperidin. GEN treated rats showed increased expressions of KIM-1, osteopontin, COX-2, Nrf2, NF-κB, TNF-α, iNOS, IL-6 and HSP-70 and decreased expression of HO-1. HDN along with GEN decreased the expres- sions of all indices excluding HO-1 and Nrf2 and an inverse correlation of expression was seen in between Nrf2 and NF-κB. Scavenging of free radicals, suppression of inflamma- tion, upregulation of HO-1 by Nrf2 mediated antioxidant response element system (ARE) and facilitation of antioxidative system by HDN would be responsible to thwart GEN induced renal damage. © 2014 Elsevier Ltd. All rights reserved. Keywords: Gentamicin Hesperidin Nephrotoxicity Kidney injury molecule-1 Nuclear factor-κB Haem oxygenase-1 1. Introduction The aminoglycoside antibiotic gentamicin (GEN) is exten- sively used in clinical practice for the management of acute Gram-negative bacterial diseases (Valle, Imbrogno, & Fernandez, 1969). Nonetheless, a main impediment in the utilization of aminoglycoside antibiotics is their potent nephrotoxicity (Martinez-Salgado, Lopez-Hernandez, & Lopez-Novoa, 2007). Gentamicin is renowned to cause acute renal failure among * Corresponding author. Department of Biochemistry and Biotechnology, Faculty of Science,Annamalai University, Chidambaram – 608 002, Tamil Nadu, India. Tel.: +91 94432 71346. E-mail address: [email protected] (P. Subramanian). Abbreviations: ARE, antioxidant response element; CAT, catalase; COX-2, cyclooxygenase-2; ECH, echinoderm microtubule associ- ated; ERK, extracellular signal-regulated kinases; GEN, gentamicin; GST, glutathione-S-transferase; HDN, hesperidin; HO-1, haem oxygenase 1; HRP, horseradish peroxidase; HSP-70, heat shock protein-70; IL-6, interleukin-6; iNOS, inducible nitric oxide synthase; KIM-1, kidney injury molecule 1; MAPK, mitogen-activated protein kinases; NF-κB, nuclear factor-κB; NO, nitric oxide; Nrf2, nuclear factor (erythroid- derived 2)-like 2; PBS, phosphate buffered saline; ROS, reactive oxygen species; RT-PCR, reverse transcriptase-polymerase chain reaction; SOD, superoxide dismutase; TBS, Tris buffered saline; TNF-α, tumor necrosis factor-α http://dx.doi.org/10.1016/j.jff.2014.12.035 1756-4646/© 2014 Elsevier Ltd. All rights reserved. journal of functional foods 13 (2015) 89–99 Available online at www.sciencedirect.com ScienceDirect journal homepage: www.elsevier.com/locate/jff
-
Upload
nguyendung -
Category
Documents
-
view
227 -
download
4
Transcript of Hesperidin protects gentamicin-induced nephrotoxicity via...

Hesperidin protects gentamicin-inducednephrotoxicity via Nrf2/HO-1 signaling andinhibits inflammation mediated by NF-κB in rats
Perumal Subramanian a,*, Ramaswamy Anandan a,Jaime Jacqueline Jayapalan b,c, Onn Haji Hashim b,c
a Department of Biochemistry and Biotechnology, Faculty of Science, Annamalai University, Chidambaram 608002, Tamil Nadu, Indiab Faculty of Medicine, University of Malaya, University of Malaya Centre for Proteomics Research (UMCPR),50603 Kuala Lumpur, Malaysiac Department of Molecular Medicine, Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia
A R T I C L E I N F O
Article history:
Received 20 July 2014
Received in revised form 14
December 2014
Accepted 19 December 2014
Available online
A B S T R A C T
The defensive effects of hesperidin (HDN, a bioflavonoid) was investigated on gentamicin
(GEN) provoked nephrotoxicity in rats. The expression patterns of nuclear factor (erythroid-
derived 2)-like 2 (Nrf2), nuclear factor-κB (NF-κB), cyclooxygenase-2 (COX-2), tumor necrosis
factor-α (TNF-α), interleukin-6 (IL-6), kidney injury molecule (KIM-1), osteopontin, heme
oxygenase-1 (HO-1), inducible nitric oxide synthase (iNOS) and heat shock protein-70 (HSP-
70) were assessed to comprehend the mechanism of action of hesperidin. GEN treated rats
showed increased expressions of KIM-1, osteopontin, COX-2, Nrf2, NF-κB, TNF-α, iNOS, IL-6
and HSP-70 and decreased expression of HO-1. HDN along with GEN decreased the expres-
sions of all indices excluding HO-1 and Nrf2 and an inverse correlation of expression was
seen in between Nrf2 and NF-κB. Scavenging of free radicals, suppression of inflamma-
tion, upregulation of HO-1 by Nrf2 mediated antioxidant response element system (ARE)
and facilitation of antioxidative system by HDN would be responsible to thwart GEN induced
renal damage.
© 2014 Elsevier Ltd. All rights reserved.
Keywords:
Gentamicin
Hesperidin
Nephrotoxicity
Kidney injury molecule-1
Nuclear factor-κB
Haem oxygenase-1
1. Introduction
The aminoglycoside antibiotic gentamicin (GEN) is exten-sively used in clinical practice for the management of acute
Gram-negative bacterial diseases (Valle, Imbrogno, & Fernandez,1969). Nonetheless, a main impediment in the utilization ofaminoglycoside antibiotics is their potent nephrotoxicity(Martinez-Salgado, Lopez-Hernandez, & Lopez-Novoa, 2007).Gentamicin is renowned to cause acute renal failure among
* Corresponding author. Department of Biochemistry and Biotechnology, Faculty of Science, Annamalai University, Chidambaram – 608002, Tamil Nadu, India. Tel.: +91 94432 71346.
E-mail address: [email protected] (P. Subramanian).Abbreviations: ARE, antioxidant response element; CAT, catalase; COX-2, cyclooxygenase-2; ECH, echinoderm microtubule associ-
ated; ERK, extracellular signal-regulated kinases; GEN, gentamicin; GST, glutathione-S-transferase; HDN, hesperidin; HO-1, haem oxygenase1; HRP, horseradish peroxidase; HSP-70, heat shock protein-70; IL-6, interleukin-6; iNOS, inducible nitric oxide synthase; KIM-1, kidneyinjury molecule 1; MAPK, mitogen-activated protein kinases; NF-κB, nuclear factor-κB; NO, nitric oxide; Nrf2, nuclear factor (erythroid-derived 2)-like 2; PBS, phosphate buffered saline; ROS, reactive oxygen species; RT-PCR, reverse transcriptase-polymerase chain reaction;SOD, superoxide dismutase; TBS, Tris buffered saline; TNF-α, tumor necrosis factor-αhttp://dx.doi.org/10.1016/j.jff.2014.12.0351756-4646/© 2014 Elsevier Ltd. All rights reserved.
j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9
Available online at www.sciencedirect.com
ScienceDirect
journal homepage: www.elsevier.com/ locate / j ff

10–25% of the patients receiving the drug (Aronson, 2006). GENhas been documented to enhance the generation of ROS (re-active oxygen species) (Moreira et al., 2014). Elevated productionof ROS induces cellular injury and necrosis through peroxidationof membrane lipids, protein denaturation and DNA damage.The kidneys from GEN-treated rats are more susceptible to ROSdamage owing to the insufficiency of antioxidant enzymes (Comet al., 2012; Khan et al., 2009). GEN induces superoxide anion(O2
●−), hydrogen peroxide (H2O2), and hydroxyl radical (●OH)production from renal mitochondria (Moreira et al., 2014).In addition, H2O2 generation (Moreira et al., 2014), lipidperoxidation, nitrotyrosine, protein oxidation (Sener et al., 2002)and protein carbonyl (Kovacs et al., 2012) production mediatenephrotoxic effects of GEN. Furthermore, reduced glutathi-one content is diminished (Sener et al., 2002) in renal cortexof GEN treated rats. GEN was shown to induce apoptosis in renalcortical cells in rats (Hsu et al., 2014). Earlier studies exploredthat GEN could release iron from renal mitochondria and forman iron–GEN complex which enhances the production of freeradicals and ROS (Kovacs et al., 2012).
The antioxidant elevating activity, histoprotective activityof renal tissue, and normalization of kidney myeloperoxidase(MPO) and urinary enzymes (β-N-acetyl glucosaminidase (NAG),β-glucuronidase and γ-glutamyl transferase (γ-GTP)) was pre-viously reported upon hesperidin administration in GEN-treated rats (Anandan & Subramanian, 2012). Hesperidin (HDN,3, 5, 7-trihydroxyflavanone 7-rhamnoglucoside) is an abun-dant bioflavonoid present in citrus fruits, sweet oranges,tangelos and lemon (Garg, Garg, Zaneveld, & Singla, 2001; Russo,Bonaccorsi, Inferrera, Dugo, & Mondello, 2015). HDN has beenshown to exhibit a variety of pharmacological effects includ-ing antioxidant, free radical scavenging, anti-inflammatory,antiallergenic, neuroprotective, antihypertensive, antimicro-bial, hypolipidemic, anticarcinogenic, vasodilatory, analgesicand antihypercholesterolemic properties (Abuelsaad, Mohamed,Allam, & Al-Solumani, 2013; Chen et al., 2013; Kamaraj,Ramakrishnan, Anandakumar, Jagan, & Devaki, 2009; Kaur,Tirkey, & Chopra, 2006; Rong et al., 2013; Sahu, Kuncha,Sindhura, & Sistla, 2013; Tirkey, Pilkhwal, Kuhad, & Chopra,2005). HDN demonstrates antioxidative properties by diversemechanisms, including scavenging of free radicals, preven-tion of DNA damages and chelation of metal (iron and copper)ions (which play key roles in the initiation radical reactions),repression of enzymes accountable for free radical genera-tion and enhancement of endogenous antioxidative defensesystem (Cao, Sofic, & Prior, 1997; Chen et al., 2010; Oliveira et al.,2013; Sahu et al., 2013).
Despite identification of GEN nephrotoxicity and inten-sive scientific attempts directed into developing therapeuticor prophylactic agents to protect against GEN nephrotoxicity,conventional therapeutic options accessible to either treat orprevent its adverse effects, are inadequate. Although GEN-induced nephrotoxicity appears to be directly recounted to itsconcentrated accrual in the renal proximal tubular cells(Martinez-Salgado et al., 2007), the mechanisms involved in GENinduced cell injury are not completely understood. In thepresent study, the nephroprotective effects of hesperidin by as-sessing transcription factors, inflammatory mediators, renalmarkers, a cytokine and a phase II enzyme (kidney injurymolecule (KIM-1), osteopontin, haem oxygenase (HO-1),
cyclooxygenase-2 (COX-2), nuclear factor (erythroid-derived 2)-like 2 (Nrf2), nuclear factor-κB (NF-κB), tumor necrosis factor-α(TNF-α), inducible nitric oxide synthase (iNOS), interleukin-6(IL-6) and heat shock protein-70 (HSP-70)) were assessed in orderto comprehend the mechanism of action of hesperidin on GEN-induced nephrotoxicity.
KIM-1, a tubular protein, is markedly induced in responseto a number of nephrotoxins (Ichimura, Hung, Yang, Stevens,& Bonventre, 2004) and has been considered as a sensitivemarker of proximal tubule injury (Jin et al., 2013). Osteopontinis known to express in renal tubular cells under various ex-perimental or pathological conditions in vivo or in vitro (Chenet al., 2009; Jin et al., 2013), experimental cyclosporine ne-phropathy (Pichler et al., 1995) and GEN-provoked nephrotoxicity(Com et al., 2012). HO-1, a protective gene in kidney, is en-tailed in the production of anti-inflammatory, antioxidant, andantiapoptotic metabolites (He et al., 2014). Phase II genes,including HO-1, SOD, CAT, GST and NAD(P)H quinine oxidore-ductase, function in synergy to scavenge ROS or reactivenitrogen species (RNS), maintain intracellular reducing poten-tial, and detoxify xenobiotics and electrophiles (Satoh et al.,2006). The transcription factor, Nrf2 regulates ARE (antioxi-dant response element)-driven HO-1 gene expression (Zhanget al., 2013). COX-2 plays key role in prostaglandin biosynthe-sis and inflammation and is expressed constitutively in kidney(Rios, Vargas-Robles, Gamez-Mendez, & Escalante, 2012). The5′-flanking region of the COX-2 promoter contains NF-κB bindingsites and a major molecule subjected to NF-κB-driventransactivation is COX-2 (Chun & Surh, 2004).
The two redox-sensitive transcription factors, that deter-mine the role of transcriptional adaption to chemical stress,are Nrf2 and NF-κB. Nrf2, the chief regulator of the antioxi-dant response (Nguyen, Nioi, & Pickett, 2009) initiates theresponse by binding to the ARE in the promoter region of nu-merous genes, encoding for antioxidative and phase II enzymesin various types of cells (Cheng, Kalabus, Zhang, & Blanco, 2012).NF-κB (transcription factor) activation is regulated by intra-/extra-cellular ROS and/or ROS-modified target biomolecules(Gonzalez-Ramos, Defrere, & Devoto, 2012). The expression ofgenes which are involved in inflammation, cell proliferationand cytokine production is mainly regulated by NF-κB(Gonzalez-Ramos et al., 2012). Upon stimulation, NF-κB is re-leased from its inhibitory subunit (IκB) and translocates intothe nucleus, where it enhances the transcriptional activationof target genes, for instance, IL-6, iNOS, TNF-α and COX-2(Amigo, Paya, Braza-Boils, De Rosa, & Terencio, 2008;Gonzalez-Ramos et al., 2012; Ozbek et al., 2009).
TNF-α is an effective proinflammatory cytokine, producedby renal mesangial and epithelial cells which could influenceprogression of diseases and enhance the production of othercytokines interceding inflammation (Benedetti et al., 2013). GENis known to increase macrophage infiltration and elevate TNF-αlevel that led to the progression of nephritis (Sahu et al., 2013).Nitric oxide (NO) is produced from a wide variety of cell typesby nitric oxide synthase enzymes (NOS) and surfeit NO couldelevate RNS (Sunil et al., 2012). A type of NOS, iNOS (induc-ible NOS), typically entails the presence of some inducing agent(e.g., cytokines) for expression, consequently forms largeamounts of NO (Csont et al., 2005). IL-6, a pro-inflammatorycytokine is primarily involved in the regulation of immune and
90 j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9

inflammatory responses (Fielding et al., 2008). Heat shock pro-teins (HSPs) are induced in cells in response to shock,hyperthermia, and other environmental stresses (Kalmar &Greensmith, 2009). Major HSPs are known to localize in renaltubular epithelial cells and GEN specifically binds to C-terminalregion of HSP-70 (Yamamoto et al., 2010).
Our earlier study (Anandan & Subramanian, 2012) showedthat rats treated with GEN exhibit desquamation, degenera-tion, and vacuolization of proximal tubules, increased numberof hyaline casts in renal tubules, marginal localization of chro-matin in the nuclei, mononuclear cell infiltration, intertubularhemorrhage, picnosis, atrophied glomeruli, and obliterativearteriolopathy in renal tissues, including inflammatory re-sponses (Anandan & Subramanian, 2012). In rats administeredwith HDN and GEN, the renal tubules were normal with mildmononuclear cell infiltration, slight degenerative epithelialchanges in glomeruli and tubules, and a reduction in spo-radic hyaline casts in tubules (Anandan & Subramanian, 2012).We performed a mechanistic approach for the effect of HDNvia Nrf2/HO-1 signaling and scrutinized whether the inhibi-tion of inflammation is mediated by NF-κB in this study.
2. Materials and methods
2.1. Experimental animals
All the experiments were performed in male rats (Wistar strain,180–200 g), acquired from the Central Animal House, Facultyof Medicine, Annamalai University. The animals were housed(4 rats per cage) and maintained on 12:12 h day and night cyclesat 25 ± 2 °C. The animals were fed with standard pellet diets(Hindustan Unilever Ltd., Bangalore, India) and drinking waterad libitum. The experiments involved with animals were ac-complished according to the guidelines of the Committee forthe Purpose of Control and Supervision of Experiments onAnimals (CPCSEA), New Delhi, India and endorsed by the AnimalEthical Committee of Annamalai University (Approval No.160/1999 CPCSEA).
2.2. Chemicals and antibodies
Gentamicin sulfate (GEN) was purchased from Ranbaxy Labo-ratories Limited, New Delhi, India. HDN, thiobarbituric acid,phenazine methosulfate, nitroblue tetrazolium, adenosine tri-phosphate and nicotinamide adenine dinucleotide werepurchased from Sigma Chemical Company, St. Louis, MO, USA.Total RNA isolation kit and primers for PCR were obtained fromBangalore Genei, Bangalore, India. The RT-PCR kit was pro-cured from Qualigen, Carlsbad, CA, USA. All other chemicalsand biochemicals used in the study were obtained from S.D.Fine Chemicals Ltd., Mumbai, India and were of analytical grade.
The primary antibodies (polyclonal anti-rat Nrf2, NF-κB p65,TNF-α, iNOS, IL-6 and HSP-70) were acquired from Santa CruzBiotech, Dallas,TX, USA. Secondary antibodies (HRP-conjugatedanti-rat IgG for immunohistochemical analysis and alkalinephosphate-conjugated anti-rabbit IgG for Western blotting) wereobtained from Bangalore Genei.
2.3. Induction of nephrotoxicity and tissuehomogenate preparation
HDN in saline (0.9% NaCl) was administered orally at dose of200 mg/kg body weight (Anandan & Subramanian, 2012; Tirkeyet al., 2005). GEN (100 mg/kg body weight) was injected toanimals intraperitoneally (Anandan & Subramanian, 2012;Parlakpinar et al., 2005). GEN injections were carried out between09.00 and 09.30 h to maximize GEN-provoked nephrotoxicityas circadian variation in GEN-induced nephrotoxicity is wellknown (Pariat, Courtois, Cambar, Piriou, & Bouquet, 1988).
After an acclimatization period of 7 days, rats were strati-fied by body weight and allocated into following 4 groups (n = 8/group). Adequate sets of the following groups were maintainedfor RT-PCR, Western blotting and immunohistochemical studies.
Group I: control rats.Group II: rats injected with GEN (100 mg/kg b.w.) for last eight
days (15th to 22nd day).Group III: rats received HDN (200 mg/kg b.w.) orally for the
entire experimental period (22 days) and GEN (100 mg/kg bodyweight) injected from 15 to 22 days.
Group IV: rats given with HDN (200 mg/kg b.w.) orally for22 days.
At the end of experimental period, all animals were sacri-ficed by cervical dislocation. The kidney tissues were excised,sliced into pieces (250 mg) and homogenized in apposite bufferin cold condition (pH 7.0) to give 20% homogenate (w/v). Thehomogenate was centrifuged at 1000 rpm for 10 min at 4 °C.The supernatant was used for various analyses.
2.4. RT-PCR
Total RNA was isolated from the tissues using RNA isolationreagent (Trizol) (Chomczynski & Sacchi, 1987) and RT-PCR wasperformed by using the appropriate primer pairs (Table 1).Totalreaction of 25 µL was performed in 250 µL PCR vial. Amplifi-cation reaction mixture contains 2.5 µL of cDNA, 200 µM of thefour dNTPs, 100 pmol of each primer, and 2.5 U of Taq DNA poly-merase (Bangalore Genei). The samples were incubated in thethermocycler as per the following reactions: initial denatur-ation (94 °C, 2 min), denaturation (94 °C, 30 s), annealing(temperature varies depending on gene, 30 s), extension (72 °C,30 s) and final extension (72 °C, 4 min). After each step, the
Table 1 – Oligonucleotide primers used for RT-PCR.
Geneproduct
Primer sequence Fragmentsize (bp)
KIM-1 Forward-5′ACTCCTGCAGACTGGAATGG3Reverse-5′CAAAGCTCAGAGAGCCCATC3′
213
Osteopontin Forward-5′TCTAGCCCCACAGAATTCTG3′Reverse-5′GCAACTGGGATGACCTTGAT3′
416
HO-1 Forward-5′CAGAAGGGTCAGGTGTC3′Reverse-5′AGTAACTCCCACCTCGT3′
260
COX-2 Forward-5′CTGAGGGGTTACCACTTCCA3′Reverse-5′TGAGCAAGTCCGTGTTCAAG3′
209
β-actin Forward-5′AACACCCCAGCCATGTACG3′Reverse-5′ATGTCACGCACGATTTCCC3′
253
18S RNA Forward-5′TTCCTTTGGTCGCTCGCTCC3′Reverse-5′GGCTGCTGGCACCAGACTT3′
510
91j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9

contents of the vial were mixed gently and centrifuged briefly.RT-PCR was performed with 18S RNA and β-actin as internalstandards. PCR products (5 µL) were separated on a 2% agarosegel and the images were photographed using Gel Doc XR system(Bio-Rad, Hercules, CA, USA) while intensities of the bands werequantified using Quantity on 1-D analysis software and plotted(Fig. 1).
2.5. Western blotting
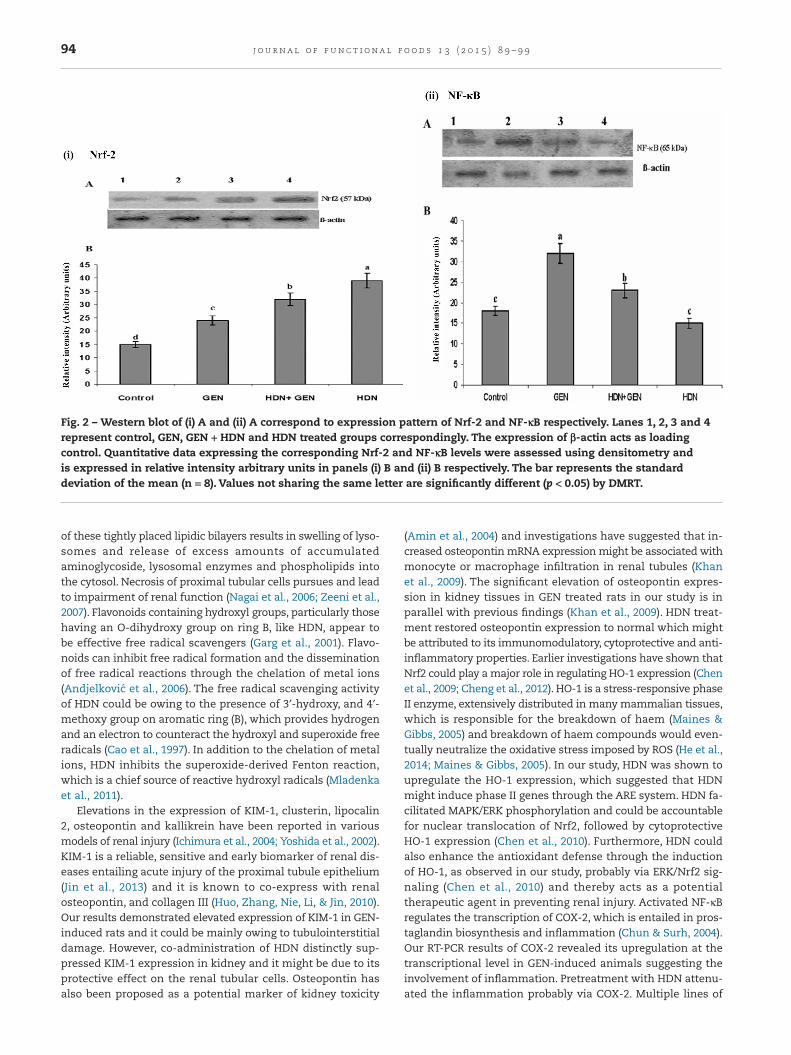
Fifty micrograms of total protein was added with 2X samplebuffer and boiled for 5 min. The sample mixture was run on12% SDS-PAGE gel and electrotransferred to a PVDF mem-brane (Millipore, Schwalbach, Germany). The membrane wasblocked in blocking buffer containing 5% skimmed milk powderfor 2 h. After 2 h, the blocked membranes were incubated withprimary antibody (rat polyclonal Nrf2 and NF-κB p65) diluted(1:1000) with blocking buffer for 6 h. Subsequent to primary an-tibody incubation, the membranes were washed three timeswith blocking buffer for 10 min each. Membranes were incu-bated with secondary antibody (anti-rat IgG alkalinephosphatase conjugate, Bangalore Genei Pvt. Ltd.). Specificbinding was identified using 5-bromo-4-chloro-3-indolylphosphate/nitroblue tetrazolium (BCIP/NBT) as substrates withβ-actin as internal control. Membrane with bands was scannedusing a scanner (HP Scan) and the intensity of bands was quan-titated with the Biorad Quantity One 1D Analysis Software(BioRad) and were plotted (Fig. 2).
2.6. Immunohistochemical analysis
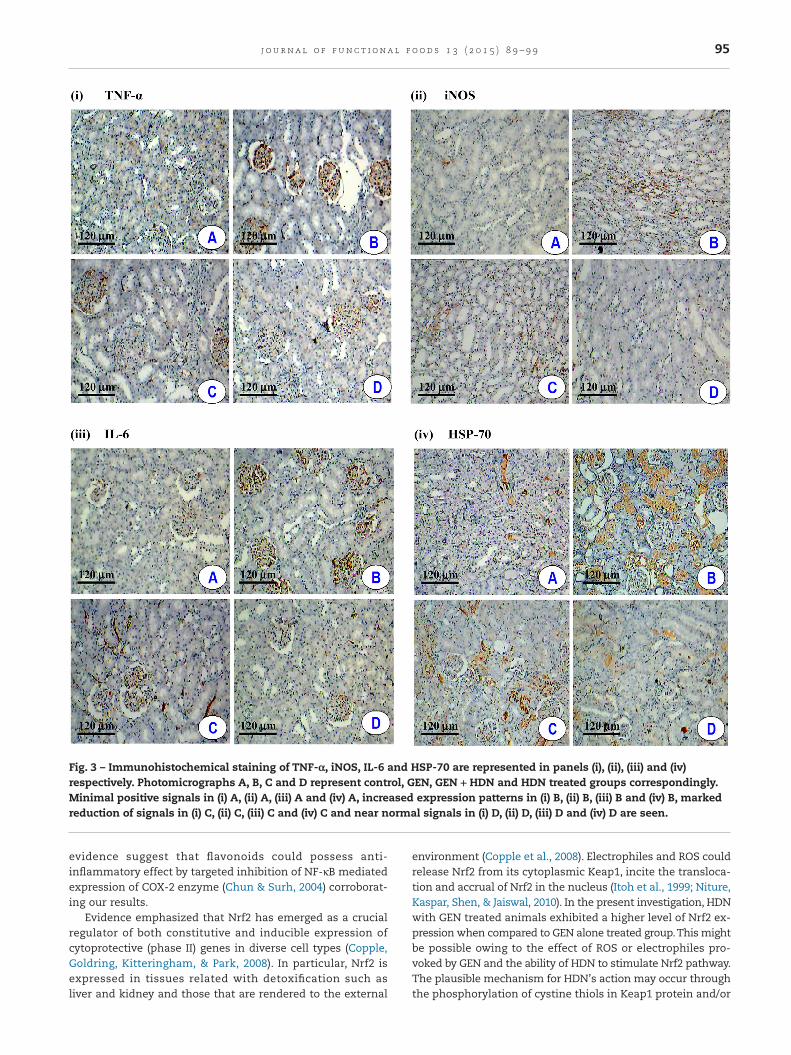
The glass section slides were dewaxed and rehydrated througha steady decrease in ethanol concentration. The slides wereincubated in sodium citrate buffer (pH 6.0) for two cycles of5 min in a microwave oven for antigen retrieval. After washingwith PBS, the slides with tissue section were treated with hy-drogen peroxide (3%) to remove any endogenous peroxidases.Blocking was carried out at 4 °C using normal goat serum ina humidified chamber for 5 h. The sections were then incu-bated 8 h at 4 °C in a humidified chamber using rat primarypolyclonal antibodies for iNOS, IL-6, HSP-70 and TNF-α respec-tively. The slides were washed with TBS and subsequentlyincubated with HRP-conjugated anti-rat IgG for 30 min eachat room temperature. The samples were washed with PBS andthe immunoprecipitate was visualized by treating with 3, 3′-diaminobenzidine. After 30 min, the slides were counterstainedwith hematoxylin and the color developed signifying the pres-ence of antigen–antibody complex was detected by lightmicroscopy. For negative controls, the primary antibody wasreplaced with TBS. Positive controls for each antibody were alsoprocessed simultaneously. The photomicrographs were takenusing Olympus BX51 microscope (Olympus Corporation, Tokyo,Japan).
2.7. Statistical analysis
The data for various indices were analyzed using analysis ofvariance (ANOVA) and the group means were compared byDuncan’s Multiple Range Test (DMRT). Values were consid-ered statistically significant when p < 0.05.
3. Results
3.1. RT-PCR analysis of KIM-1, osteopontin,HO-1 and COX-2
KIM-1 and osteopontin expressions were found to be signifi-cantly increased (p < 0.05) in the kidney of GEN induced rats(compared to controls) whereas in GEN + HDN treated rats theywere appreciably decreased (p < 0.05; Fig. 1). In HDN treated rats,there was no significant variation of expression of KIM-1 andosteopontin compared to control (Fig. 1). GEN induced ratsdisplayed a decreased expression of HO-1 when comparedto controls (Fig. 1). HDN treatment to GEN-induced group el-evated the expression level of HO-1. HDN treated groupexhibited insignificant variation compared to control group. RT-PCR analysis of COX-2 showed its upregulated expression inGEN-provoked nephrotoxic animals. COX-2 expression was con-siderably decreased in HDN pre-treated animals as comparedwith GEN-treated animals (Fig. 1).
3.2. Expression of Nrf2 and NF-κB p65 bywestern blotting
Western blot analysis showed detectable levels of Nrf2 in controlgroup (Fig. 2). GEN treatment increased the expression of Nrf2.HDN treatment to GEN-induced group resulted in further el-evation in the expression of Nrf2 (Fig. 2). Furthermore, theexpression of Nrf2 is significantly elevated (p < 0.05) in HDNtreated group (compared to control) (Fig. 2). The expression ofNF-κB p65 in the kidney is increased (p < 0.05) after GEN treat-ment (Fig. 2). In contrast, HDN treatment decreased theexpression of NF-κB p65 significantly (p < 0.05) in GEN-treatedrats.The expression in HDN treated group is insignificantly dif-ferent compared to control group (Fig. 2).
3.3. Immunohistochemical expression of TNF-α, iNOS,IL-6 and HSP-70
Control rats showed the basal level of immunohistochemicalstaining of TNF-α, iNOS, IL-6 and HSP-70 in kidney tissues(Fig. 3). The expression levels of TNF-α, iNOS, IL-6 and HSP-70are elevated in kidney tissues of GEN-treated rats. Con-versely, HDN + GEN treatment showed considerable reductionof these expressions as compared to GEN-induced rats (Fig. 3).The HDN treated rats showed similar pattern of expressionsof TNF-α, iNOS, IL-6 and HSP-70 to that of controls (Fig. 3).
4. Discussion
The specificity of GEN-provoked renal toxicity is related to itspreferential accrual in the renal convoluted tubules, lyso-somes, brush border membranes and basolateral membranesby binding with the phosphatidyl-inositide receptors (Nagai,Saito, Adachi, Yumoto, & Takano, 2006; Zeeni, Selmaoui,Beauchamp, Labrecque, & Thibault, 2007). This aminoglycosidefurther inhibits phospholipases and sphingomyelinase activ-ity causing the production of myeloid bodies.The development
92 j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9
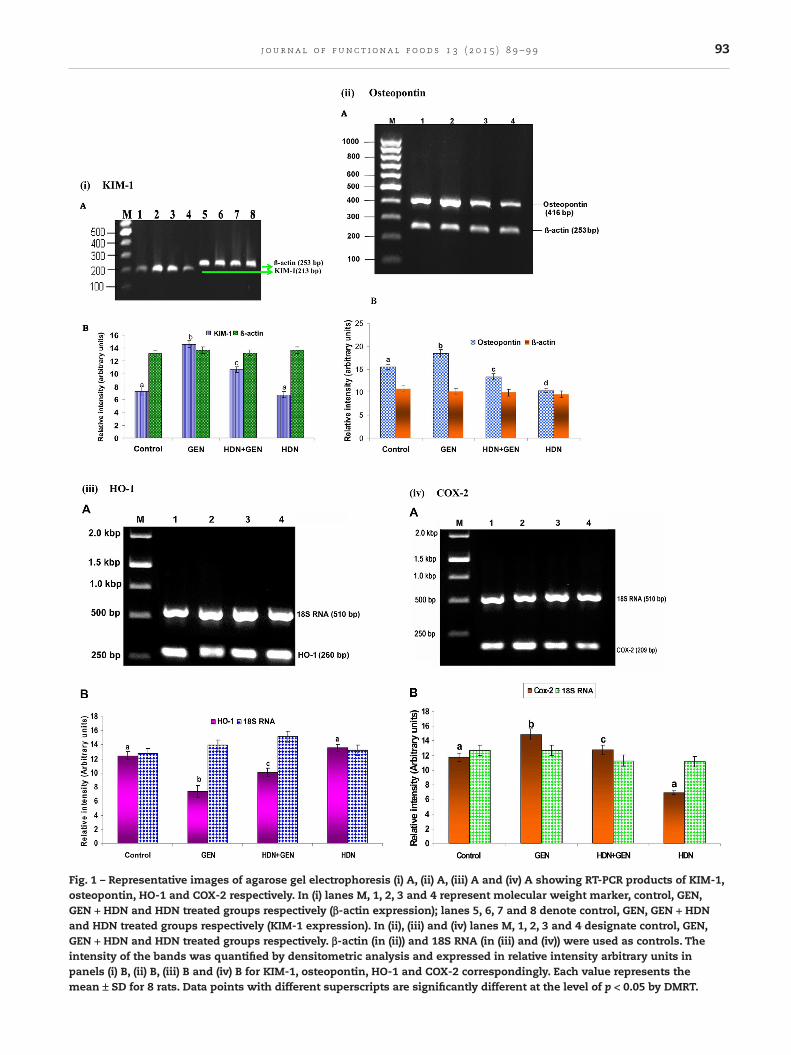
Fig. 1 – Representative images of agarose gel electrophoresis (i) A, (ii) A, (iii) A and (iv) A showing RT-PCR products of KIM-1,osteopontin, HO-1 and COX-2 respectively. In (i) lanes M, 1, 2, 3 and 4 represent molecular weight marker, control, GEN,GEN + HDN and HDN treated groups respectively (β-actin expression); lanes 5, 6, 7 and 8 denote control, GEN, GEN + HDNand HDN treated groups respectively (KIM-1 expression). In (ii), (iii) and (iv) lanes M, 1, 2, 3 and 4 designate control, GEN,GEN + HDN and HDN treated groups respectively. β-actin (in (ii)) and 18S RNA (in (iii) and (iv)) were used as controls. Theintensity of the bands was quantified by densitometric analysis and expressed in relative intensity arbitrary units inpanels (i) B, (ii) B, (iii) B and (iv) B for KIM-1, osteopontin, HO-1 and COX-2 correspondingly. Each value represents themean ± SD for 8 rats. Data points with different superscripts are significantly different at the level of p < 0.05 by DMRT.
93j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9
of these tightly placed lipidic bilayers results in swelling of lyso-somes and release of excess amounts of accumulatedaminoglycoside, lysosomal enzymes and phospholipids intothe cytosol. Necrosis of proximal tubular cells pursues and leadto impairment of renal function (Nagai et al., 2006; Zeeni et al.,2007). Flavonoids containing hydroxyl groups, particularly thosehaving an O-dihydroxy group on ring B, like HDN, appear tobe effective free radical scavengers (Garg et al., 2001). Flavo-noids can inhibit free radical formation and the disseminationof free radical reactions through the chelation of metal ions(Andjelkovic et al., 2006). The free radical scavenging activityof HDN could be owing to the presence of 3′-hydroxy, and 4′-methoxy group on aromatic ring (B), which provides hydrogenand an electron to counteract the hydroxyl and superoxide freeradicals (Cao et al., 1997). In addition to the chelation of metalions, HDN inhibits the superoxide-derived Fenton reaction,which is a chief source of reactive hydroxyl radicals (Mladenkaet al., 2011).
Elevations in the expression of KIM-1, clusterin, lipocalin2, osteopontin and kallikrein have been reported in variousmodels of renal injury (Ichimura et al., 2004; Yoshida et al., 2002).KIM-1 is a reliable, sensitive and early biomarker of renal dis-eases entailing acute injury of the proximal tubule epithelium(Jin et al., 2013) and it is known to co-express with renalosteopontin, and collagen III (Huo, Zhang, Nie, Li, & Jin, 2010).Our results demonstrated elevated expression of KIM-1 in GEN-induced rats and it could be mainly owing to tubulointerstitialdamage. However, co-administration of HDN distinctly sup-pressed KIM-1 expression in kidney and it might be due to itsprotective effect on the renal tubular cells. Osteopontin hasalso been proposed as a potential marker of kidney toxicity
(Amin et al., 2004) and investigations have suggested that in-creased osteopontin mRNA expression might be associated withmonocyte or macrophage infiltration in renal tubules (Khanet al., 2009). The significant elevation of osteopontin expres-sion in kidney tissues in GEN treated rats in our study is inparallel with previous findings (Khan et al., 2009). HDN treat-ment restored osteopontin expression to normal which mightbe attributed to its immunomodulatory, cytoprotective and anti-inflammatory properties. Earlier investigations have shown thatNrf2 could play a major role in regulating HO-1 expression (Chenet al., 2009; Cheng et al., 2012). HO-1 is a stress-responsive phaseII enzyme, extensively distributed in many mammalian tissues,which is responsible for the breakdown of haem (Maines &Gibbs, 2005) and breakdown of haem compounds would even-tually neutralize the oxidative stress imposed by ROS (He et al.,2014; Maines & Gibbs, 2005). In our study, HDN was shown toupregulate the HO-1 expression, which suggested that HDNmight induce phase II genes through the ARE system. HDN fa-cilitated MAPK/ERK phosphorylation and could be accountablefor nuclear translocation of Nrf2, followed by cytoprotectiveHO-1 expression (Chen et al., 2010). Furthermore, HDN couldalso enhance the antioxidant defense through the inductionof HO-1, as observed in our study, probably via ERK/Nrf2 sig-naling (Chen et al., 2010) and thereby acts as a potentialtherapeutic agent in preventing renal injury. Activated NF-κBregulates the transcription of COX-2, which is entailed in pros-taglandin biosynthesis and inflammation (Chun & Surh, 2004).Our RT-PCR results of COX-2 revealed its upregulation at thetranscriptional level in GEN-induced animals suggesting theinvolvement of inflammation. Pretreatment with HDN attenu-ated the inflammation probably via COX-2. Multiple lines of
Fig. 2 – Western blot of (i) A and (ii) A correspond to expression pattern of Nrf-2 and NF-κB respectively. Lanes 1, 2, 3 and 4represent control, GEN, GEN + HDN and HDN treated groups correspondingly. The expression of β-actin acts as loadingcontrol. Quantitative data expressing the corresponding Nrf-2 and NF-κB levels were assessed using densitometry andis expressed in relative intensity arbitrary units in panels (i) B and (ii) B respectively. The bar represents the standarddeviation of the mean (n = 8). Values not sharing the same letter are significantly different (p < 0.05) by DMRT.
94 j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9
evidence suggest that flavonoids could possess anti-inflammatory effect by targeted inhibition of NF-κB mediatedexpression of COX-2 enzyme (Chun & Surh, 2004) corroborat-ing our results.
Evidence emphasized that Nrf2 has emerged as a crucialregulator of both constitutive and inducible expression ofcytoprotective (phase II) genes in diverse cell types (Copple,Goldring, Kitteringham, & Park, 2008). In particular, Nrf2 isexpressed in tissues related with detoxification such asliver and kidney and those that are rendered to the external
environment (Copple et al., 2008). Electrophiles and ROS couldrelease Nrf2 from its cytoplasmic Keap1, incite the transloca-tion and accrual of Nrf2 in the nucleus (Itoh et al., 1999; Niture,Kaspar, Shen, & Jaiswal, 2010). In the present investigation, HDNwith GEN treated animals exhibited a higher level of Nrf2 ex-pression when compared to GEN alone treated group.This mightbe possible owing to the effect of ROS or electrophiles pro-voked by GEN and the ability of HDN to stimulate Nrf2 pathway.The plausible mechanism for HDN’s action may occur throughthe phosphorylation of cystine thiols in Keap1 protein and/or
Fig. 3 – Immunohistochemical staining of TNF-α, iNOS, IL-6 and HSP-70 are represented in panels (i), (ii), (iii) and (iv)respectively. Photomicrographs A, B, C and D represent control, GEN, GEN + HDN and HDN treated groups correspondingly.Minimal positive signals in (i) A, (ii) A, (iii) A and (iv) A, increased expression patterns in (i) B, (ii) B, (iii) B and (iv) B, markedreduction of signals in (i) C, (ii) C, (iii) C and (iv) C and near normal signals in (i) D, (ii) D, (iii) D and (iv) D are seen.
95j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9
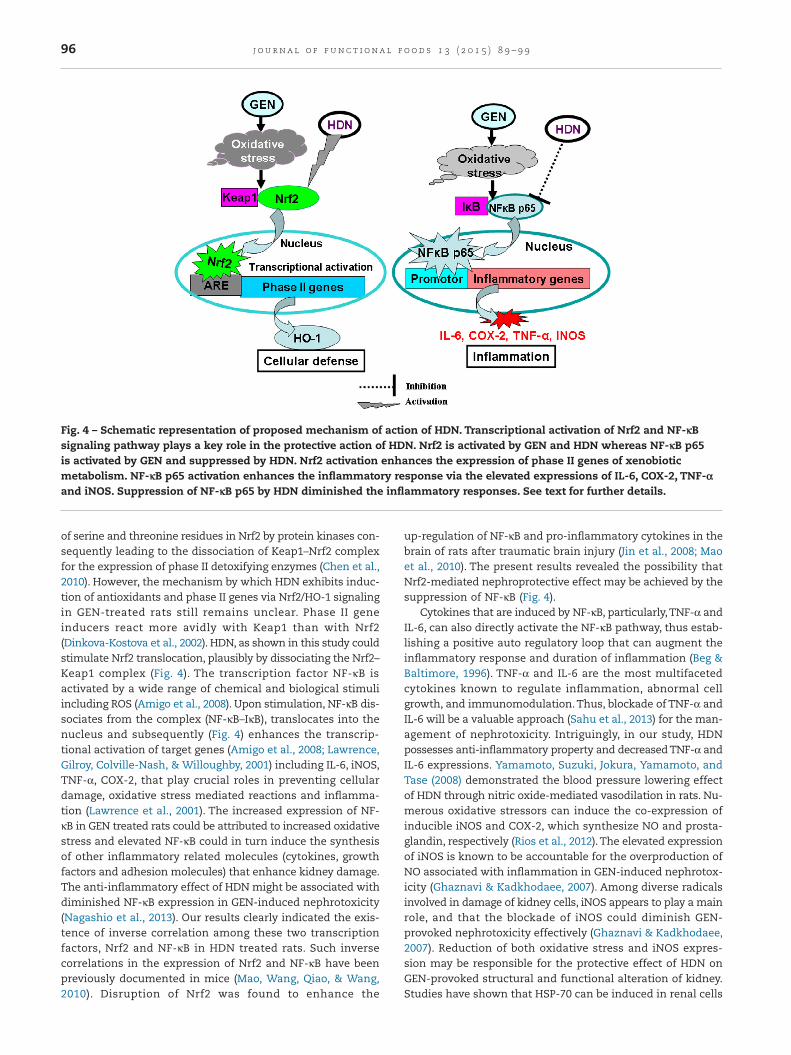
of serine and threonine residues in Nrf2 by protein kinases con-sequently leading to the dissociation of Keap1–Nrf2 complexfor the expression of phase II detoxifying enzymes (Chen et al.,2010). However, the mechanism by which HDN exhibits induc-tion of antioxidants and phase II genes via Nrf2/HO-1 signalingin GEN-treated rats still remains unclear. Phase II geneinducers react more avidly with Keap1 than with Nrf2(Dinkova-Kostova et al., 2002). HDN, as shown in this study couldstimulate Nrf2 translocation, plausibly by dissociating the Nrf2–Keap1 complex (Fig. 4). The transcription factor NF-κB isactivated by a wide range of chemical and biological stimuliincluding ROS (Amigo et al., 2008). Upon stimulation, NF-κB dis-sociates from the complex (NF-κB–IκB), translocates into thenucleus and subsequently (Fig. 4) enhances the transcrip-tional activation of target genes (Amigo et al., 2008; Lawrence,Gilroy, Colville-Nash, & Willoughby, 2001) including IL-6, iNOS,TNF-α, COX-2, that play crucial roles in preventing cellulardamage, oxidative stress mediated reactions and inflamma-tion (Lawrence et al., 2001). The increased expression of NF-κB in GEN treated rats could be attributed to increased oxidativestress and elevated NF-κB could in turn induce the synthesisof other inflammatory related molecules (cytokines, growthfactors and adhesion molecules) that enhance kidney damage.The anti-inflammatory effect of HDN might be associated withdiminished NF-κB expression in GEN-induced nephrotoxicity(Nagashio et al., 2013). Our results clearly indicated the exis-tence of inverse correlation among these two transcriptionfactors, Nrf2 and NF-κB in HDN treated rats. Such inversecorrelations in the expression of Nrf2 and NF-κB have beenpreviously documented in mice (Mao, Wang, Qiao, & Wang,2010). Disruption of Nrf2 was found to enhance the
up-regulation of NF-κB and pro-inflammatory cytokines in thebrain of rats after traumatic brain injury (Jin et al., 2008; Maoet al., 2010). The present results revealed the possibility thatNrf2-mediated nephroprotective effect may be achieved by thesuppression of NF-κB (Fig. 4).
Cytokines that are induced by NF-κB, particularly,TNF-α andIL-6, can also directly activate the NF-κB pathway, thus estab-lishing a positive auto regulatory loop that can augment theinflammatory response and duration of inflammation (Beg &Baltimore, 1996). TNF-α and IL-6 are the most multifacetedcytokines known to regulate inflammation, abnormal cellgrowth, and immunomodulation. Thus, blockade of TNF-α andIL-6 will be a valuable approach (Sahu et al., 2013) for the man-agement of nephrotoxicity. Intriguingly, in our study, HDNpossesses anti-inflammatory property and decreased TNF-α andIL-6 expressions. Yamamoto, Suzuki, Jokura, Yamamoto, andTase (2008) demonstrated the blood pressure lowering effectof HDN through nitric oxide-mediated vasodilation in rats. Nu-merous oxidative stressors can induce the co-expression ofinducible iNOS and COX-2, which synthesize NO and prosta-glandin, respectively (Rios et al., 2012). The elevated expressionof iNOS is known to be accountable for the overproduction ofNO associated with inflammation in GEN-induced nephrotox-icity (Ghaznavi & Kadkhodaee, 2007). Among diverse radicalsinvolved in damage of kidney cells, iNOS appears to play a mainrole, and that the blockade of iNOS could diminish GEN-provoked nephrotoxicity effectively (Ghaznavi & Kadkhodaee,2007). Reduction of both oxidative stress and iNOS expres-sion may be responsible for the protective effect of HDN onGEN-provoked structural and functional alteration of kidney.Studies have shown that HSP-70 can be induced in renal cells
Fig. 4 – Schematic representation of proposed mechanism of action of HDN. Transcriptional activation of Nrf2 and NF-κBsignaling pathway plays a key role in the protective action of HDN. Nrf2 is activated by GEN and HDN whereas NF-κB p65is activated by GEN and suppressed by HDN. Nrf2 activation enhances the expression of phase II genes of xenobioticmetabolism. NF-κB p65 activation enhances the inflammatory response via the elevated expressions of IL-6, COX-2, TNF-αand iNOS. Suppression of NF-κB p65 by HDN diminished the inflammatory responses. See text for further details.
96 j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9

during acute renal failure due to ischemia and nephrotoxicdrugs, and suggested their cytoprotective roles (Saito et al., 2004;Tolson, Roberts, Jortner, Pomeroy, & Barber, 2005). The presentimmunohistochemical study revealed the increased expres-sion of HSP-70 in renal tissues owing to nephrotoxicity and HDNcould effectively suppress the HSP-70 probably through its an-tioxidant and cytoprotective actions.
To summarize, HDN supplementation inhibited NF-κB me-diated expression of inflammatory genes (IL-6, iNOS, TNF-α,COX-2) and upregulated Nrf2 and Nrf2 dependant phase II an-tioxidant gene HO-1. The effects of HDN might be attributedto multiple mechanisms such as scavenging of free radicals,suppression of inflammation, chelation of metal ions, inhibi-tion of enzymes responsible for free radical generation andfacilitation of endogenous antioxidative defense system. Basedon the results, HDN could be a potential therapeutic agent inthe treatment of oxidative stress-related renal cell injury andrenal dysfunction. The potential therapeutic action of HDN toprotect or reverse renal GEN damage would have indispens-able clinical consequences.
Acknowledgements
Visiting Professorship to PS at Department of Molecular Medi-cine, University of Malaya and financial assistance to OHH(RP017-13HTM and HIR-MOHE H-20001-00-E000009) are thank-fully acknowledged.
R E F E R E N C E S
Abuelsaad, A. S., Mohamed, I., Allam, G., & Al-Solumani, A. A.(2013). Antimicrobial and immunomodulating activities ofhesperidin and ellagic acid against diarrheic Aeromonashydrophila in a murine model. Life Sciences, 93, 714–722.
Amigo, M., Paya, M., Braza-Boils, A., De Rosa, S., & Terencio, M. C.(2008). Avarol inhibits TNF-alpha generation and NF-kappaBactivation in human cells and in animal models. Life Sciences,82, 256–264.
Amin, R. P., Vickers, A. E., Sistare, F., Thompson, K. L., Roman,R. J., Lawton, M., Kramer, J., Hamadeh, H. K., Collins, J.,Grissom, S., Bennett, L., Tucker, C. J., Wild, S., Kind, C., Oreffo,V., Davis, J. W., Curtiss, S., Naciff, J. M., Cunningham, M.,Tennant, R., Stevens, J., Car, B., Bertram, T. A., & Afshari,C. A. (2004). Identification of putative gene based markersof renal toxicity. Environmental Health Perspectives, 112,465–479.
Anandan, R., & Subramanian, P. (2012). Renal protective effect ofhesperidin on gentamicin-induced acute nephrotoxicity inmale Wistar albino rats. Redox Report, 17, 219–226.
Andjelkovic, M., Van Camp, J., De Meulenaer, B., Depaemelaere,G., Socaciu, C., Verloo, M., & Verhe, R. (2006). Iron-chelationproperties of phenolic acids bearing catechol and galloylgroups. Food Chemistry, 98, 23–31.
Aronson, J. K. (2006). Meyler’s side effects of drugs (15th ed.).Amsterdam: Elsevier.
Beg, A. A., & Baltimore, D. (1996). An essential role for NF-kappaBin preventing TNF-alpha-induced cell death. Science, 274,782–784.
Benedetti, G., Fredriksson, L., Herpers, B., Meerman, J., van deWater, B., & de Graauw, M. (2013). TNF-alpha-mediated
NF-kappaB survival signaling impairment by cisplatinenhances JNK activation allowing synergistic apoptosis ofrenal proximal tubular cells. Biochemical Pharmacology, 85,274–286.
Cao, G., Sofic, E., & Prior, R. L. (1997). Antioxidant and prooxidantbehaviour of flavonoids: Structure-activity relationships. FreeRadical Biology and Medicine, 22, 749–760.
Chen, M., Gu, H., Ye, Y., Lin, B., Sun, L., Deng, W., Zhang, J., & Liu, J.(2010). Protective effects of hesperidin against oxidative stressof tert-butyl hydroperoxide in human hepatocytes. Food andChemical Toxicology, 48, 2980–2987.
Chen, S. Y., Chyau, C. C., Chu, C. C., Chen, Y. H., Chen, T. H., &Duh, P. D. (2013). Hepatoprotection using sweet orangepeel and its bioactive compound, hesperidin, forCCl4-induced liver injury in vivo. Journal of Functional Foods, 5,1591–1600.
Chen, T. H., Chang, C. F., Yu, S. C., Wang, J. C., Chen, C. H., Chan, P.,& Lee, H. M. (2009). Dipyridamole inhibits cobalt chloride-induced osteopontin expression in NRK52E cells. EuropeanJournal of Pharmacology, 613, 10–18.
Cheng, Q., Kalabus, J. L., Zhang, J., & Blanco, J. G. (2012). Aconserved antioxidant response element (ARE) in thepromoter of human carbonyl reductase 3 (CBR3) mediatesinduction by the master redox switch Nrf2. BiochemicalPharmacology, 83, 139–148.
Chomczynski, P., & Sacchi, N. (1987). Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction. Analytical Biochemistry, 162, 156–159.
Chun, K. S., & Surh, Y. J. (2004). Signal transduction pathwaysregulating cyclooxygenase-2 expression: Potential moleculartargets for chemoprevention. Biochemical Pharmacology, 68,1089–1100.
Com, E., Boitier, E., Marchandeau, J. P., Brandenburg, A.,Schroeder, S., Hoffmann, D., Mally, A., & Gautier, J. C. (2012).Integrated transcriptomic and proteomic evaluation ofgentamicin nephrotoxicity in rats. Toxicology and AppliedPharmacology, 258, 124–133.
Copple, I. M., Goldring, C. E., Kitteringham, N. R., & Park, B. K.(2008). The Nrf2-Keap1 defence pathway: Role in protectionagainst drug-induced toxicity. Toxicology, 246, 24–33.
Csont, T., Viappiani, S., Sawicka, J., Slee, S., Altarejos, J. Y.,Batinic-Haberle, I., & Schulz, R. (2005). The involvement ofsuperoxide and iNOS-derived NO in cardiac dysfunctioninduced by pro-inflammatory cytokines. Journal of Molecularand Cellular Cardiology, 39, 833–840.
Dinkova-Kostova, A. T., Holtzclaw, W. D., Cole, R. N., Itoh, K.,Wakabayashi, N., Katoh, Y., Yamamoto, M., & Talalay, P. (2002).Direct evidence that sulfhydryl groups of Keap1 are thesensors regulating induction of phase 2 enzymes that protectagainst carcinogens and oxidants. Proceedings of the NationalAcademy of Sciences of the United States of America, 99, 11908–11913.
Fielding, C. A., McLoughlin, R. M., McLeod, L., Colmont, C. S.,Najdovska, M., Grail, D., Ernst, M., Jones, S. A., Topley, N., &Jenkins, B. J. (2008). IL-6 regulates neutrophil trafficking duringacute inflammation via STAT3. Journal of Immunology, 181,2189–2195.
Garg, A., Garg, S., Zaneveld, L. J., & Singla, A. K. (2001). Chemistryand pharmacology of the citrus bioflavonoid hesperidin.Phytotherapy Research, 15, 655–669.
Ghaznavi, R., & Kadkhodaee, M. (2007). Comparative effects ofselective and non-selective nitric oxide synthase inhibition ingentamicin-induced rat nephrotoxicity. Archives of Toxicology,81, 453–457.
Gonzalez-Ramos, R., Defrere, S., & Devoto, L. (2012). Nuclearfactor-kappaB: A main regulator of inflammation and cellsurvival in endometriosis pathophysiology. Fertility andSterility, 98, 520–528.
97j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9

He, X. H., Yan, X. T., Wang, Y. L., Wang, C. Y., Zhang, Z. Z., & Zhan,J. (2014). Transduced PEP-1-haem oxygenase-1 fusion proteinprotects against intestinal ischemia/reperfusion injury.Journal of Surgical Research, 187, 77–84.
Hsu, Y. H., Chen, T. H., Wu, M. Y., Lin, Y. F., Chen, W. L., Cheng,T. H., & Chen, C. H. (2014). Protective effects of Zhibai DihuangWan on renal tubular cells affected with gentamicin-inducedapoptosis. Journal of Ethnopharmacology, 151, 635–642.
Huo, W., Zhang, K., Nie, Z., Li, Q., & Jin, F. (2010). Kidney injurymolecule-1 (KIM-1): A novel kidney-specific injury moleculeplaying potential double-edged functions in kidney injury.Transplantation Reviews (Orlando), 24, 143–146.
Ichimura, T., Hung, C. C., Yang, S. A., Stevens, J. L., & Bonventre,J. V. (2004). Kidney injury molecule-1: A tissue and urinarybiomarker for nephrotoxicant-induced renal injury. AmericanJournal of Physiology – Renal Physiology, 286, F552–F563.
Itoh, K., Wakabayashi, N., Katoh, Y., Ishii, T., Igarashi, K., Engel,J. D., & Yamamoto, M. (1999). Keap1 represses nuclearactivation of antioxidant responsive elements by Nrf2through binding to the amino-terminal Neh2 domain. Genes &Development, 13, 76–86.
Jin, W., Wang, H., Yan, W., Xu, L., Wang, X., Zhao, X., Yang, X.,Chen, G., & Ji, Y. (2008). Disruption of Nrf2 enhancesupregulation of nuclear factor-kappaB activity,proinflammatory cytokines, and intercellular adhesionmolecule-1 in the brain after traumatic brain injury. Mediatorsof Inflammation, 2008, 725174.
Jin, Z. K., Tian, P. X., Wang, X. Z., Xue, W. J., Ding, X. M., Zheng, J.,Ding, C. G., Mao, T. C., Duan, W. L., & Xi, M. (2013). Kidneyinjury molecule-1 and osteopontin: New markers forprediction of early kidney transplant rejection. MolecularImmunology, 54, 457–464.
Kalmar, B., & Greensmith, L. (2009). Induction of heat shockproteins for protection against oxidative stress. Advanced DrugDelivery Reviews, 61, 310–318.
Kamaraj, S., Ramakrishnan, G., Anandakumar, P., Jagan, S., &Devaki, T. (2009). Antioxidant and anticancer efficacy ofhesperidin in benzo(a)pyrene induced lung carcinogenesis inmice. Investigational New Drugs, 27, 214–222.
Kaur, G., Tirkey, N., & Chopra, K. (2006). Beneficial effect ofhesperidin on lipopolysaccharide-induced hepatotoxicity.Toxicology, 226, 152–160.
Khan, S. A., Priyamvada, S., Farooq, N., Khan, S., Khan, M. W., &Yusufi, A. N. (2009). Protective effect of green tea extract ongentamicin-induced nephrotoxicity and oxidative damage inrat kidney. Pharmacological Research, 59, 254–262.
Kovacs, E., Savopol, T., Iordache, M. M., Saplacan, L., Sobaru, I.,Istrate, C., Mingeot-Leclercq, M. P., & Moisescu, M. G. (2012).Interaction of gentamicin polycation with model and cellmembranes. Bioelectrochemistry (Amsterdam, Netherlands), 87,230–235.
Lawrence, T., Gilroy, D. W., Colville-Nash, P. R., & Willoughby,D. A. (2001). Possible new role for NF-kappaB in the resolutionof inflammation. Nature Medicine, 7, 1291–1297.
Maines, M. D., & Gibbs, P. E. (2005). 30 some years of haemoxygenase: From a “molecular wrecking ball” to a“mesmerizing” trigger of cellular events. Biochemical andBiophysical Research Communications, 338, 568–577.
Mao, L., Wang, H., Qiao, L., & Wang, X. (2010). Disruption of Nrf2enhances the upregulation of nuclear factor-kappaB activity,tumor necrosis factor-alpha, and matrix metalloproteinase-9after spinal cord injury in mice. Mediators of Inflammation,2010, 238321.
Martinez-Salgado, C., Lopez-Hernandez, F. J., & Lopez-Novoa, J. M.(2007). Glomerular nephrotoxicity of aminoglycosides.Toxicology and Applied Pharmacology, 223, 86–98.
Mladenka, P., Macakova, K., Filipsky, T., Zatloukalova, L., Jahodar,L., Bovicelli, P., Silvestri, I. P., Hrdina, R., & Saso, L. (2011). In
vitro analysis of iron chelating activity of flavonoids. Journal ofInorganic Biochemistry, 105, 693–701.
Moreira, M. A., Nascimento, M. A., Bozzo, T. A., Cintra, A., daSilva, S. M., Dalboni, M. A., Mouro, M. G., & Higa, E. M. (2014).Ascorbic acid reduces gentamicin-induced nephrotoxicity inrats through the control of reactive oxygen species. ClinicalNutrition, 33, 296–301.
Nagai, J., Saito, M., Adachi, Y., Yumoto, R., & Takano, M. (2006).Inhibition of gentamicin binding to rat renal brush-bordermembrane by megalin ligands and basic peptides. Journal ofControlled Release, 112, 43–50.
Nagashio, Y., Matsura, Y., Miyamoto, J., Kometani, T., Suzuki, T., &Tanabe, S. (2013). Hesperidin inhibits development of atopicdermatitis-like skin lesions in NC/Nga mice by suppressingTh17 activity. Journal of Functional Foods, 5, 1633–1641.
Nguyen, T., Nioi, P., & Pickett, C. B. (2009). The Nrf2-antioxidantresponse element signaling pathway and its activation byoxidative stress. Journal of Biological Chemistry, 284, 13291–13295.
Niture, S. K., Kaspar, J. W., Shen, J., & Jaiswal, A. K. (2010). Nrf2signaling and cell survival. Toxicology and Applied Pharmacology,244, 37–42.
Oliveira, R. M., Daniel, J. F., Aguiar, I., Silva, M. F., Fernandes, J., &Carlos, R. M. (2013). Structural effects on the hesperidinproperties obtained by chelation to magnesium complexes.Journal of Inorganic Biochemistry, 129, 35–42.
Ozbek, E., Cekmen, M., Ilbey, Y. O., Simsek, A., Polat, E. C., &Somay, A. (2009). Atorvastatin prevents gentamicin-inducedrenal damage in rats through the inhibition of p38-MAPKand NF-kappaB pathways. Renal Failure, 31, 382–392.
Pariat, C., Courtois, P., Cambar, J., Piriou, A., & Bouquet, S. (1988).Circadian variations in the renal toxicity of gentamicin inrats. Toxicology Letters, 40, 175–182.
Parlakpinar, H., Tasdemir, S., Polat, A., Bay-Karabulut, A., Vardi,N., Ucar, M., & Acet, A. (2005). Protective role of caffeic acidphenethyl ester (cape) on gentamicin-induced acute renaltoxicity in rats. Toxicology, 207, 169–177.
Pichler, R. H., Franceschini, N., Young, B. A., Hugo, C., Andoh, T. F.,Burdmann, E. A., Shankland, S. J., Alpers, C. E., Bennett, W. M.,Couser, W. G., & Johnson, R. J. (1995). Pathogenesis ofcyclosporine nephropathy: Roles of angiotensin II andosteopontin. Journal of the American Society of Nephrology, 6,1186–1196.
Rios, A., Vargas-Robles, H., Gamez-Mendez, A. M., & Escalante, B.(2012). Cyclooxygenase-2 and kidney failure. Prostaglandins &Other Lipid Mediators, 98, 86–90.
Rong, Z., Pan, R., Xu, Y., Zhang, C., Cao, Y., & Liu, D. (2013).Hesperidin pretreatment protects hypoxia-ischemic braininjury in neonatal rat. Neuroscience, 255, 292–299.
Russo, M., Bonaccorsi, I., Inferrera, V., Dugo, P., & Mondello, L.(2015). Underestimated sources of flavonoids, limnoids anddietary fiber: Availability in orange’s by-products. Journal ofFunctional Foods, 12, 150–157.
Sahu, B. D., Kuncha, M., Sindhura, G. J., & Sistla, R. (2013).Hesperidin attenuates cisplatin-induced acute renal injury bydecreasing oxidative stress, inflammation and DNA damage.Phytomedicine: International Journal of Phytotherapy andPhytopharmacology, 20, 453–460.
Saito, M., Tominaga, L., Nanba, E., Kinoshita, Y., Housi, D.,Miyagawa, I., & Satoh, K. (2004). Expression of HSP 70 and itsmRNAs during ischemia-reperfusion in the rat bladder. LifeSciences, 75, 1879–1886.
Satoh, T., Okamoto, S. I., Cui, J., Watanabe, Y., Furuta, K., Suzuki,M., Tohyama, K., & Lipton, S. A. (2006). Activation of theKeap1/Nrf2 pathway for neuroprotection by electrophilic[correction of electrophillic] phase II inducers. Proceedings ofthe National Academy of Sciences of the United States of America,103, 768–773.
98 j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9

Sener, G., Sehirli, A. O., Altunbas, H. Z., Ersoy, Y., Paskaloglu, K.,Arbak, S., & Ayanoglu-Dulger, G. (2002). Melatonin protectsagainst gentamicin-induced nephrotoxicity in rats. Journal ofPineal Research, 32, 231–236.
Sunil, V. R., Shen, J., Patel-Vayas, K., Gow, A. J., Laskin, J. D., &Laskin, D. L. (2012). Role of reactive nitrogen speciesgenerated via inducible nitric oxide synthase in vesicant-induced lung injury, inflammation and altered lungfunctioning. Toxicology and Applied Pharmacology, 261, 22–30.
Tirkey, N., Pilkhwal, S., Kuhad, A., & Chopra, K. (2005). Hesperidin,a citrus bioflavonoid, decreases the oxidative stress producedby carbon tetrachloride in rat liver and kidney. BMCPharmacology, 5, 2.
Tolson, J. K., Roberts, S. M., Jortner, B., Pomeroy, M., & Barber, D. S.(2005). Heat shock proteins and acquired resistance touranium nephrotoxicity. Toxicology, 206, 59–73.
Valle, D. A., Imbrogno, M. A., & Fernandez, E. (1969). Gentamicinin pediatric infections caused by gram-negative organisms.The Journal of Infectious Diseases, 119, 453–456.
Yamamoto, M., Suzuki, A., Jokura, H., Yamamoto, N., & Tase, T.(2008). Glycosyl hesperidin prevents endothelial dysfunctionand oxidative stress in spontaneously hypertensive rats.Nutrition (Burbank, Los Angeles County, Calif.), 24, 470–476.
Yamamoto, S., Nakano, S., Owari, K., Fuziwara, K., Ogawa, N.,Otaka, M., Tamaki, K., Watanabe, S., Komatsuda, A., Wakui, H.,Sawada, K., Kubota, H., & Itoh, H. (2010). Gentamicin inhibitsHSP70-assisted protein folding by interfering with substraterecognition. FEBS Letters, 584, 645–651.
Yoshida, T., Kurella, M., Beato, F., Min, H., Ingelfinger, J. R., Stears,R. L., Swinford, R. D., Gullans, S. R., & Tang, S. S. (2002).Monitoring changes in gene expression in renal ischemia-reperfusion in the rat. Kidney International, 61, 1646–1654.
Zeeni, N., Selmaoui, B., Beauchamp, D., Labrecque, G., & Thibault,L. (2007). Dietary protein level alters gentamicin-inducednephrotoxicity in rats. Physiology & Behaviour, 90, 760–770.
Zhang, M., An, C., Gao, Y., Leak, R. K., Chen, J., & Zhang, F. (2013).Emerging roles of Nrf2 and phase II antioxidant enzymes inneuroprotection. Progress in Neurobiology, 100, 30–47.
99j o u rna l o f f un c t i ona l f o od s 1 3 ( 2 0 1 5 ) 8 9 – 9 9





![Research Paper HO-1 induced autophagy protects against IL ... · induce apoptosis of the nucleus pulposus cells (NPCs) in the degenerative intervertebral disc [5, 6]. Autophagy is](https://static.fdocument.org/doc/165x107/5e72f110b749c078843e28fa/research-paper-ho-1-induced-autophagy-protects-against-il-induce-apoptosis-of.jpg)





![Transgenic inhibition of astroglial NF-?B protects from ... inhibition of astroglial NF-κB[1].pdf · blocked with PBS containing 0.15% Tween 20, 2% bovine serum albumin (BSA), and](https://static.fdocument.org/doc/165x107/5e0374b25abbb03275334e3a/transgenic-inhibition-of-astroglial-nf-b-protects-from-inhibition-of-astroglial.jpg)






