
Gene Therapy of the β- Hemoglobinopathies by Lentiviral ...
19
Gene Therapy of the β- Hemoglobinopathies by Lentiviral Transfer of the βA(T87Q)-Globin Gene The Harvard community has made this article openly available. Please share how this access benefits you. Your story matters Citation Negre, O., A. Eggimann, Y. Beuzard, J. Ribeil, P. Bourget, S. Borwornpinyo, S. Hongeng, et al. 2016. “Gene Therapy of the β-Hemoglobinopathies by Lentiviral Transfer of the βA(T87Q)- Globin Gene.” Human Gene Therapy 27 (2): 148-165. doi:10.1089/ hum.2016.007. http://dx.doi.org/10.1089/hum.2016.007. Published Version doi:10.1089/hum.2016.007 Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:26318640 Terms of Use This article was downloaded from Harvard University’s DASH repository, and is made available under the terms and conditions applicable to Other Posted Material, as set forth at http:// nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of- use#LAA
Transcript of Gene Therapy of the β- Hemoglobinopathies by Lentiviral ...

Gene Therapy of the β-Hemoglobinopathies by Lentiviral
Transfer of the βA(T87Q)-Globin GeneThe Harvard community has made this
article openly available. Please share howthis access benefits you. Your story matters
Citation Negre, O., A. Eggimann, Y. Beuzard, J. Ribeil, P. Bourget, S.Borwornpinyo, S. Hongeng, et al. 2016. “Gene Therapy of theβ-Hemoglobinopathies by Lentiviral Transfer of the βA(T87Q)-Globin Gene.” Human Gene Therapy 27 (2): 148-165. doi:10.1089/hum.2016.007. http://dx.doi.org/10.1089/hum.2016.007.
Published Version doi:10.1089/hum.2016.007
Citable link http://nrs.harvard.edu/urn-3:HUL.InstRepos:26318640
Terms of Use This article was downloaded from Harvard University’s DASHrepository, and is made available under the terms and conditionsapplicable to Other Posted Material, as set forth at http://nrs.harvard.edu/urn-3:HUL.InstRepos:dash.current.terms-of-use#LAA

Gene Therapy of the b-Hemoglobinopathies by LentiviralTransfer of the bA(T87Q)-Globin Gene
Olivier Negre,1,2,* Anne-Virginie Eggimann,1 Yves Beuzard,2,3 Jean-Antoine Ribeil,4
Philippe Bourget,4 Suparerk Borwornpinyo,5 Suradej Hongeng,5 Salima Hacein-Bey,6
Marina Cavazzana,4 Philippe Leboulch,2,3,5,7 and Emmanuel Payen2,3,8,*1bluebird bio, Cambridge, Massachusetts; 2CEA, Institute of Emerging Disease and Innovative Therapies (iMETI), Fontenay aux Roses, France; 3UMR 007, University
of Paris 11 and CEA, CEA-iMETI, Fontenay aux Roses, France; 4Necker Hospital, Assistance Publique-Hopitaux de Paris, Paris, France; 5Mahidol University, Bangkok,
Thailand; 6Immunology Laboratory, Groupe Hospitalier Universitaire Paris-Sud, Assistance Publique–Hopitaux de Paris, Paris, France; 7Harvard Medical School and
Genetics Division, Department of Medicine, Brigham & Women’s Hospital, Boston, Massachusetts; 8INSERM, Paris, France.
b-globin gene disorders are the most prevalent inherited diseases worldwide and result from abnormal b-globin synthesis or structure. Novel therapeutic approaches are being developed in an effort to movebeyond palliative management. Gene therapy, by ex vivo lentiviral transfer of a therapeutic b-globin genederivative (bAT87Q-globin) to hematopoietic stem cells, driven by cis-regulatory elements that confer high,erythroid-specific expression, has been evaluated in human clinical trials over the past 8 years. bAT87Q-globin is used both as a strong inhibitor of HbS polymerization and as a biomarker. While long-termstudies are underway in multiple centers in Europe and in the United States, proof-of-principle of efficacyand safety has already been obtained in multiple patients with b-thalassemia and sickle cell disease.
INTRODUCTION
SICKLE CELL DISEASE (SCD) and b-thalassemia major(b-TM), the latter defined clinically as transfusion-dependent cases regardless of the underlying geno-type, are the most common monogenic disordersworldwide with approximately 400,000 affected con-ceptions or births each year.1,2 These disorders fallinto two large groups of b-globin gene mutations thatresult in either abnormal hemoglobin structure (SCD)or massively reduced/absent production of b-globinchains (b-TM). The clinical manifestations of theseinherited disorders typically appear several monthsafter birth, when gene expression switches from thefetal c-globin chain, which forms fetal hemoglobin(HbF), to the adult bA-globin chain forming hemo-globin (HbA).3 Note that adult bA-globin is also simplyreferred to as b-globin when no confusion with otherb-like globin chains is possible. In low-income coun-tries, most of the affected children succumb in earlychildhood, whereas in developed countries, neonataldiagnosis and supportive care have greatly improved
survival. However, even with modern and specializedcare, life expectancy is still reduced by several de-cades,4–6 and quality of life greatly suffers.7–9
Allogenic hematopoietic stem cell transplanta-tion (AHSCT) as a curative option is currentlyrecommended for b-TM if a human leukocyte an-tigen (HLA)-matched sibling donor is available.10,11
Disease-free survival after AHSCT is approxima-tely 88% in pediatric subjects12 and 65% in adults.13
Recent transplantation trials conducted duringthe last 15 years for young SCD patients reporteda disease free-survival rate of 85–90%.11 However,fewer than 25% of patients have a suitable in-trafamilial donor.14 In the absence of matchedsibling donors, AHSCT from HLA-matched unre-lated or haploidentical donors or minimally mis-matched cord blood products may be used as thesource of donor cells, although these approachesexhibit a lower benefit/risk ratio and thus remainexperimental.15 Although outcomes are improv-ing, AHSCT continues to carry a substantial risk
*Correspondence: Dr. Emmanuel Payen, CEA/iMETI, 18 route du Panorama, 92260 Fontenay aux Roses, France. E-mail: [email protected] or Dr. Olivier Negre,bluebird bio, 150 Second Street, Cambridge, MA 02141. E-mail: [email protected]
ª Olivier Negre et al. 2016; Published by Mary Ann Liebert, Inc. This Open Access article is distributed under the terms of the CreativeCommons Attribution Noncommercial License (http://creativecommons.org/licenses/by-nc/4.0/) which permits any noncommercial use,distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
148 j HUMAN GENE THERAPY, VOLUME 27 NUMBER 2 DOI: 10.1089/hum.2016.007Mary Ann Liebert, Inc.

of severe adverse events and mortality,16,17 bothincreasing with recipient age and disease severi-ty.18,19 Severe adverse events include graft failure,graft-versus-host disease (GVHD), early or late sideeffects from conditioning regimens that are bothmyeloablative and immunosuppressive (infections,hemorrhages, secondary malignancies), and aggra-vation of preexisting organ damage.20–22
For patients who lack a suitable HLA-matcheddonor, ex vivo gene therapy using autologous HSCsbrings hope as a potential curative treatment option.If proven safe and effective, gene therapy may thenbe extended to most b-TM and severe SCD patients,as there is no concern here for histocompatibility-related complications and the conditioning regimendoes not need to include immunosuppressive drugs.However, gene therapy shares with AHSCT therisks associated with the transplant procedureand the toxicity of the myeloablative agent. Globingene addition to HSCs by means of lentiviral vectors(LVs) is a promising approach under investigation(Fig. 1). Several clinical trials of gene therapy forb-TM and severe SCD are ongoing in France and inthe United States (Tables 1 and 2). Other recentapproaches under study for the gene therapy of theb-hemoglobinopathies include pharmacological23 orgenetic induction of c-globin production throughinterference with the BCL11A pathway24,25 or dis-ruption of the BCL11A erythroid enhancer byCRISPR/CAS9 technology as well as zinc finger ortranscription activator-like effector nuclease,26,27
or even attempts at repairing the defective bA-globin gene in HSCs by genome editing.28–30 These
approaches are at the stage of collecting evidenceof efficacy in relevant cell and animal models andscoring potentially untoward off-target events. Evenif these approaches are ultimately successful, geneaddition has the advantage of making use of a singleproduct applicable to all cases of b-TM and SCDregardless of the genotype, whereas gene repair willhave to tackle separately the hundreds of mutationsknown to cause b-thalassemia in humans.
While we recognize the important contribu-tions from other laboratories, this review willlargely focus on our own experience with the de-velopment of the HPV569 and BB305 vectors, self-inactivating (SIN) LVs bearing a human bA-globinmini-gene encoding an ‘‘antisickling’’ b-globin withamino-acid substitution (T87Q) and driven by cis-regulatory elements of the human b-globin genelocus (promoter, locus control region [LCR] ele-ments), designed in the Leboulch laboratory incollaboration with bluebird bio (formerly GenetixPharmaceuticals).
THE b-HEMOGLOBINOPATHIESThe b-thalassemias
b-TM is a microcytic hemolytic anemia that israpidly fatal in the absence of palliative life-longred blood cell (RBC) transfusions and iron chela-tion.31 The disease results from absence (b0) ormassive reduction (e.g., b+, bE/b0) in bA-globin geneexpression. While complete absence of bA-globinexpression is not compatible with RBC production,even a severe diminution in its expression also re-
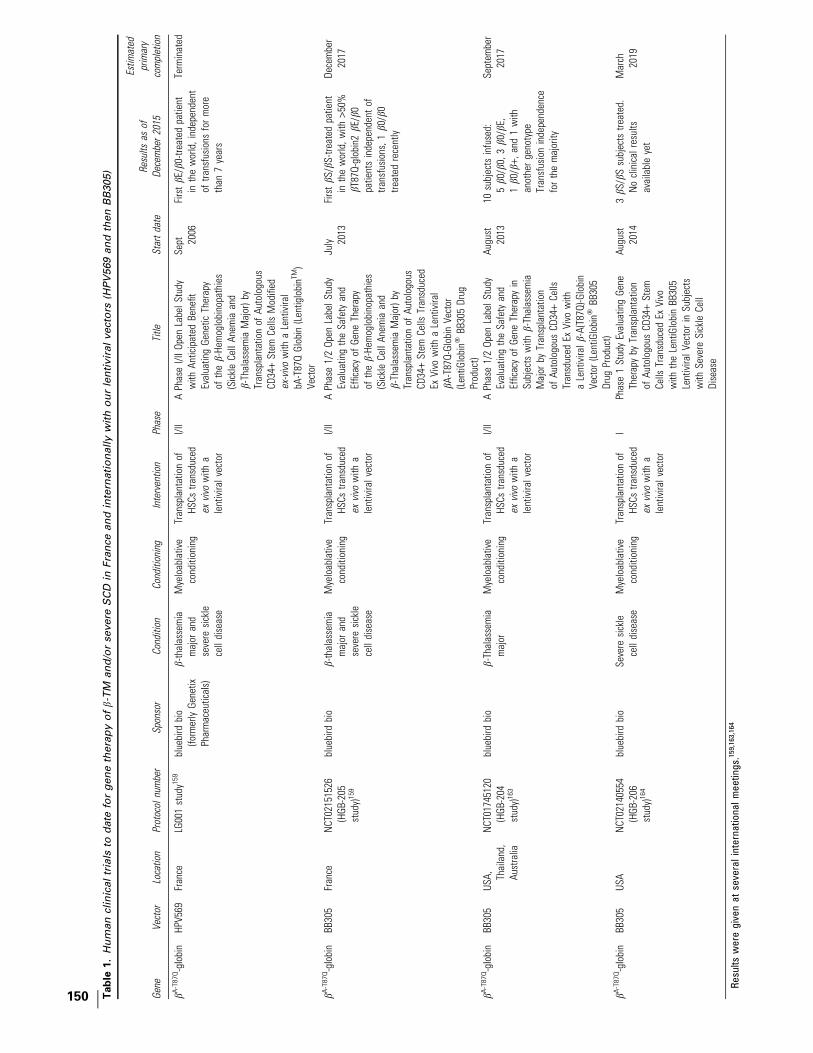
Figure 1. The milestones of ex vivo gene therapy research and development for hemoglobin disorders. LG001, HGB204, HGB205, and HGB206 clinical studiesare conducted with our lentiviral vectors (Table 1). Gene therapy trials using other lentiviral vectors are summarized in Table 2.
GENE THERAPY OF b-HEMOGLOBINOPATHIES 149
Tab
le1.
Hu
man
clin
ical
tria
lsto
date
for
gen
eth
era
py
of
b-T
Man
d/o
rse
vere
SC
Din
Fra
nce
an
din
tern
ati
on
ally
wit
ho
ur
len
tivir
al
vect
ors
(HP
V569
an
dth
en
BB
305)
Gen
eVe
ctor
Loca
tion
Prot
ocol
num
ber
Spon
sor
Cond
ition
Cond
ition
ing
Inte
rven
tion
Phas
eTi
tleSt
art
date
Resu
ltsas
ofD
ecem
ber
2015
Estim
ated
prim
ary
com
plet
ion
bA-T
87Q-g
lobi
nH
PV56
9Fr
ance
LG00
1st
udy15
9bl
uebi
rdbi
o(fo
rmer
lyG
enet
ixPh
arm
aceu
tical
s)
b-t
hala
ssem
iam
ajor
and
seve
resi
ckle
cell
dise
ase
Mye
loab
lativ
eco
nditi
onin
gTr
ansp
lant
atio
nof
HSC
str
ansd
uced
exvi
vow
itha
lent
ivira
lve
ctor
I/II
APh
ase
I/II
Ope
nLa
bel
Stud
yw
ithA
ntic
ipat
edBe
nefit
Eval
uatin
gG
enet
icTh
erap
yof
the
b-H
emog
lobi
nopa
thie
s(S
ickl
eCe
llA
nem
iaan
db-
Thal
asse
mia
Maj
or)
byTr
ansp
lant
atio
nof
Aut
olog
ous
CD34
+St
emCe
llsM
odifi
edex
-viv
ow
itha
Lent
ivira
lbA
-T87
QG
lobi
n(L
entig
lobi
nTM)
Vect
or
Sept 20
06Fi
rstb
E/b
0-tr
eate
dpa
tient
inth
ew
orld
,in
depe
nden
tof
tran
sfus
ions
for
mor
eth
an7
year
s
Term
inat
ed
bA-T
87Q-g
lobi
nBB
305
Fran
ceN
CT02
1515
26(H
GB-
205
stud
y)15
9
blue
bird
bio
b-t
hala
ssem
iam
ajor
and
seve
resi
ckle
cell
dise
ase
Mye
loab
lativ
eco
nditi
onin
gTr
ansp
lant
atio
nof
HSC
str
ansd
uced
exvi
vow
itha
lent
ivira
lve
ctor
I/II
APh
ase
1/2
Ope
nLa
bel
Stud
yEv
alua
ting
the
Safe
tyan
dEf
ficac
yof
Gen
eTh
erap
yof
the
b-H
emog
lobi
nopa
thie
s(S
ickl
eCe
llA
nem
iaan
db-
Thal
asse
mia
Maj
or)
byTr
ansp
lant
atio
nof
Aut
olog
ous
CD34
+St
emCe
llsTr
ansd
uced
ExVi
vow
itha
Lent
ivira
lbA
-T87
Q-G
lobi
nVe
ctor
(Len
tiGlo
bin�
BB30
5D
rug
Prod
uct)
July 20
13Fi
rstb
S/b
S-tr
eate
dpa
tient
inth
ew
orld
,w
ith>5
0%b
T87Q
-glo
bin2
bE/
b0pa
tient
sin
depe
nden
tof
tran
sfus
ions
,1
b0/b
0tr
eate
dre
cent
ly
Dec
embe
r20
17
bA-T
87Q-g
lobi
nBB
305
USA
,Th
aila
nd,
Aus
tral
ia
NCT
0174
5120
(HG
B-20
4st
udy)
163
blue
bird
bio
b-T
hala
ssem
iam
ajor
Mye
loab
lativ
eco
nditi
onin
gTr
ansp
lant
atio
nof
HSC
str
ansd
uced
exvi
vow
itha
lent
ivira
lve
ctor
I/II
APh
ase
1/2
Ope
nLa
bel
Stud
yEv
alua
ting
the
Safe
tyan
dEf
ficac
yof
Gen
eTh
erap
yin
Subj
ects
with
b-T
hala
ssem
iaM
ajor
byTr
ansp
lant
atio
nof
Aut
olog
ous
CD34
+Ce
llsTr
ansd
uced
ExVi
vow
itha
Lent
ivira
lb
-A(T
87Q
)-Glo
bin
Vect
or(L
entiG
lobi
n�BB
305
Dru
gPr
oduc
t)
Aug
ust
2013
10su
bjec
tsin
fuse
d:5
b0/b
0,3
b0/
bE,
1b0
/b+,
and
1w
ithan
othe
rge
noty
peTr
ansf
usio
nin
depe
nden
cefo
rth
em
ajor
ity
Sept
embe
r20
17
bA-T
87Q-g
lobi
nBB
305
USA
NCT
0214
0554
(HG
B-20
6st
udy)
164
blue
bird
bio
Seve
resi
ckle
cell
dise
ase
Mye
loab
lativ
eco
nditi
onin
gTr
ansp
lant
atio
nof
HSC
str
ansd
uced
exvi
vow
itha
lent
ivira
lve
ctor
IPh
ase
1St
udy
Eval
uatin
gG
ene
Ther
apy
byTr
ansp
lant
atio
nof
Aut
olog
ous
CD34
+St
emCe
llsTr
ansd
uced
ExVi
vow
ithth
eLe
ntiG
lobi
nBB
305
Lent
ivira
lVe
ctor
inSu
bjec
tsw
ithSe
vere
Sick
leCe
llD
isea
se
Aug
ust
2014
3b
S/bS
subj
ects
trea
ted.
No
clin
ical
resu
ltsav
aila
ble
yet
Mar
ch20
19
Res
ults
wer
egi
ven
atse
vera
lin
tern
atio
nal
mee
tings
.159,
163,
164
150 j

Tab
le2.
Hu
man
clin
ical
tria
lsfo
rg
en
eth
era
py
of
b-T
Mo
rse
vere
SC
Dw
ith
oth
er
len
tivir
al
vect
ors
Gen
eVe
ctor
Loca
tion
Prot
ocol
num
ber
Spon
sor
Cond
ition
Cond
ition
ing
Inte
rven
tion
Phas
eTi
tleSt
art
date
Resu
ltsEs
timat
edpr
imar
yco
mpl
etio
n
b-gl
obin
TNS9
.3.5
5U
SAN
CT01
6396
9016
5M
emor
ial
Sloa
nKe
tter
ing
Canc
erCe
nter
b-T
hala
ssem
iam
ajor
Part
ial
cyto
redu
ctio
n(B
u8
mg/
kg)
for
3pa
tient
s,m
yelo
abla
tive
cond
ition
ing
(Bu
14m
g/kg
)fo
r1
patie
nt
Tran
spla
ntat
ion
ofH
SCs
tran
sduc
edex
vivo
with
ale
ntiv
iral
vect
or
IA
Phas
eI
Clin
ical
Tria
lfo
rth
eTr
eatm
ent
ofb-
Thal
asse
mia
Maj
orw
ithA
utol
ogou
sCD
34+
Hem
atop
oiet
icPr
ogen
itor
Cells
Tran
sduc
edw
ithTN
S9.3
.55
aLe
ntiv
iral
Vect
orEn
codi
ngth
eN
orm
alH
uman
b-G
lobi
nG
ene
July 20
12Fo
urpa
tient
str
eate
d.Th
ree
b0/b
+an
don
eb0
/b0.
One
patie
ntha
da
sign
ifica
ntde
crea
sein
tran
sfus
ion
requ
irem
ents
.
July
2016
c-gl
obin
sGbG
USA
NCT
0218
6418
aCh
ildre
n’s
Hos
pita
lM
edic
alCe
nter
,Ci
ncin
nati
Seve
resi
ckle
cell
dise
ase
Unk
now
nTr
ansp
lant
atio
nof
HSC
str
ansd
uced
exvi
vow
itha
lent
ivira
lve
ctor
I/II
Gen
eTr
ansf
erfo
rPa
tient
sw
ithSi
ckle
Cell
Dis
ease
Usi
nga
Gam
ma
Glo
bin
Lent
iviru
sVe
ctor
:A
nO
pen
Labe
lPh
ase
I/II
Pilo
tSt
udy
July 20
14N
ore
sults
avai
labl
eye
t
July
2017
bAS3
-glo
bin
(T87
Q,
G16
D,
E22A
)
Lent
i/bA
S3-F
BU
SAN
CT02
2478
43a
Uni
vers
ityof
Calif
orni
a,Ch
ildre
n’s
Hos
pita
l,Lo
sA
ngel
es
Seve
resi
ckle
cell
dise
ase
Unk
now
nTr
ansp
lant
atio
nof
HSC
str
ansd
uced
exvi
vow
itha
lent
ivira
lve
ctor
ICl
inic
alRe
sear
chSt
udy
ofA
utol
ogou
sBo
neM
arro
wTr
ansp
lant
atio
nfo
rSi
ckle
Cell
Dis
ease
(SCD
)U
sing
Bone
Mar
row
CD34
+Ce
llsM
odifi
edw
ithth
eLe
nti/bA
S3-F
BLe
ntiv
iral
Vect
or
Aug
ust
2014
No
resu
ltsav
aila
ble
yet
Apr
il20
17
b-gl
obin
GLO
BEIta
lyN
CT02
4534
7716
6,a
IRCC
SSa
nRa
ffae
leb
-Tha
lass
emia
maj
orM
yelo
abla
tive
cond
ition
ing
Tran
spla
ntat
ion
ofH
SCs
tran
sduc
edex
vivo
with
ale
ntiv
iral
vect
or(in
trab
one
inje
ctio
n)
I/II
APh
ase
I/II
Stud
yEv
alua
ting
Safe
tyan
dEf
ficac
yof
Aut
olog
ous
Hem
atop
oiet
icSt
emCe
llsG
enet
ical
lyM
odifi
edw
ithG
LOBE
Lent
ivira
lVe
ctor
Enco
ding
for
the
Hum
anBe
taG
lobi
nG
ene
for
the
Trea
tmen
tof
Patie
nts
Aff
ecte
dby
Tran
sfus
ion
Dep
ende
ntBe
ta-T
hala
ssem
ia
May 20
15Fi
rst
patie
ntre
cent
lytr
eate
d
Aug
ust
2019
aC
linic
altr
ials
.gov
Res
ults
wer
epr
ovid
edat
inte
rnat
iona
lm
eetin
gs.16
5,16
6
j 151

sults in b-TM. This is because the protein compo-nent of adult hemoglobin comprises two a- and twob-globin chains, and massive decrease in b-globinexpression results in a relative excess of unpairedand toxic free a-chains. Free a-chains precipitate,damage the cell membrane,32 and sequester thechaperone heat shock protein 70 (HSP70), which isno longer available to protect the GATA-1 ery-throid maturation transcription factor fromcaspase-3 cleavage.33 a/b-chain imbalance resultsin cell death and ineffective erythropoiesis (dyser-ythropoiesis)34 in bone marrow as well as reducederythrocyte lifespan and hemolysis.35
Among the severe b-thalassemias, the bE/b0 ge-notype presents interesting features and is highlyfrequent. In this disease, one b-globin allele iscompletely silent (b0) while the other encodes themissense mutation (26AAG>GAG) that results bothin an amino-acid substitution (GLU26LYS) and inabnormal RNA splicing.36 Part of the transcribedbE mRNA is not translated because of abnormalsplicing, whereas the small amount of mutated(GLU26LYS) protein produced is functional butslightly unstable.37 Thus, when the bE allele iscompounded with a b0 allele, a profound decreasein b-like globin production is observed.38 Approxi-mately 50% of patients with bE/b0-thalassemiahave transfusion-dependent b-TM, depending onthe patient’s genetic makeup in modulators of dis-ease severity.39 Hemoglobin E is one of the world’smost common mutations40 and is especially preva-lent in Southeast Asia. The frequency can approach60% of the population in some parts of Thailand,Cambodia, and Laos,40 where it is estimated that100,000 new cases of bE/b0 are expected in the nextfew decades. It is also found at high frequency inother Asian nations (India, Sri Lanka, Malaysia,and southern China) and increasingly in Europeand North America through immigration.41,42
Sickle cell diseaseSCD is a multisystem disorder that results from
a single mutation in the bA-globin gene, changing aglutamic acid into a valine at the sixth position ofthe b-globin chain, known as the bS mutation.43 Inhomozygous patients (bS/bS), the most frequentgenotype, or in the compound heterozygous states(bS/bC, bS/bThalassemia) of SCD, the chronic hemo-lytic anemia is complicated by painful vaso-occlusive crises (VOC), acute chest syndrome (ACS),increased risk of infections, as well as organ vascu-lopathy and dysfunctions, particularly affectingbrain, kidney, lung, heart, bone, eye, and the skin.44
At the protein level, the single amino-acid substi-tution at codon 6 (bS chain) prompts the formation of
HbS polymers at low oxygen pressure,45 the rate ofwhich is proportional to the corpuscular concentra-tion of HbS, the extent of hemoglobin deoxygen-ation in the microcirculation,46 and the quantity ofHbF, which inhibits HbS polymerization.47 Vaso-occlusion, resulting from erythrocyte stiffness andthe adhesive interaction of abnormal erythrocytesto endothelial cells and leukocytes,48 leads to organinfarction and inflammation, which in turn en-hances adhesive interactions,49 microvascularocclusion, and ischemia. The increased number ofactivated white blood cells produces high levels ofreactive oxygen species (ROS) and other factorscontributing actively to micro-vessel clogging, in-creased local hypoxia, and increased proportionsof rigid RBCs containing HbS polymers.50 Thecycles of ischemia/reperfusion cause oxidativestress51 that contributes to the proinflammatoryphenotype,52 worsening the vicious circle. Further-more, acute and chronic intravascular hemolysisimpairs endothelial functions, causing progressivesystemic and pulmonary vasculopathy.53
CHALLENGES FOR EFFECTIVEGENE THERAPYThe therapeutic bA(T87Q)-globin gene
With regard to the gene therapy of SCD by geneaddition, wild-type human bA-globin is a relativelyweak inhibitor of HbS polymerization because itacts by mere dilution. When PO2 is relatively low,as in capillary vessels, phenylalanine 85 (bF85) andleucine 88 (bL88) form an acceptor hydrophobicpocket on one a2b
S2 tetramer that binds to the mu-
tated valine (bV6) of a close a2bS
2 complex.54 Thisphenomenon is responsible for HbS fiber polymeri-zation after further aggregation and fiber elonga-tion.46 HbA (a2b
A2) has these same hydrophobic
residues and thus does not inhibit polymer forma-tion when it is incorporated.54–56 In contrast, otherhuman ‘‘b-like’’-globin chains such as c-globin andd-globin are stronger inhibitors.57,58 This inhibitionof polymerization is best exemplified by cases ofhereditary persistence of HbF (HPFH), especiallyof the pan-cellular type (within all erythrocytes),where as little as 20% HbF is sufficient to inhibitHbS formation in vivo and to alleviate substantiallythe clinical manifestations of SCD in homozygousbS/bS patients.59,60 In these cases, the c-chainexcludes the heterotetramer a2cb
S from the HbSpolymer.61–63
However, neither c- or d-globin genes are highlyexpressed in the ‘‘adult’’ (postglobin switch) RBCenvironment. Because the bA-, c-, and d-globinchains are co-linear and comprise only a few dif-
152 NEGRE ET AL.

ferences in amino-acid residues between them, es-pecially between bA and d with only 10 differencesout of 146, biochemical studies were performed todetermine which positions were critical to inhibitHbS polymerization. Within the acceptor hydro-phobic pocket, a critical residue differs at position87 between bA (T87) and c or d (Q87), and has beendetermined to be responsible for most of the inhib-itory effect of c- or d-globins on HbS polymeriza-tion.55 A few other residues co-contribute to a lesserdegree, and in particular Ala-22.64 In an effort toexpress a strong antisickling b-globin derivative athigh levels in adult RBCs, a modified bA-globin genewas designed (Leboulch Laboratory) that comprisesa point mutation within a human b-globin mini-gene suitable for retroviral transfer, so that theexpressed therapeutic protein is bA(T87Q). The re-sulting bA(T87Q)-globin chain appears as efficientas c-globin to inhibit HbS polymerization in bio-chemical assays.65 The oxygen affinity of the mu-tated tetramer (HbAT87Q: a2b
T87Q2) is in the range
of that of HbA,65 whereas the oxygen affinity ofHBF is substantially higher than that of HbA.66
Furthermore, the b-globin T87Q is a valuablebiomarker of biological efficacy in human clinicaltrials. The bT87Q-globin chain can be distinguishedand quantified from bA- and c-globin chains byhigh-performance reverse-phase liquid chroma-tography (HPLC).67,68 In the case of b+ thalasse-mia or when RBC transfusions are still provided,there would otherwise be no possibility of quan-tifying vector-derived gene expression if wild-typebA-globin was used as the therapeutic gene.
Therapeutic levels of bA(T87Q)-globin
gene expressionThe threshold level of therapeutic bA(T87Q)-
globin expression required for effective gene ther-apy of b-TM will vary depending on residual levels,if any, of endogenous b-globin production. It willalso depend on genetic modulators that contributeto reducing the severity of the disease in each in-dividual patient. These modulators include asso-ciated a-thalassemia69,70 or the ability to producesubstantial amounts of c-globin after birth,71,72
both reducing the magnitude of free a-chains inerythroid cells and the severity of the disease.
With respect to SCD, one can surmise that theantisickling properties of bA(T87Q)-globin will becomparable to or slightly lower than that of c-globin.65 HPFH with HbF levels of 30% or evenlower are consistently associated with completeabsence of clinical and biological signs of SCD inotherwise homozygous bS/bS patients, when HbFexpression is well distributed among RBCs (pan-
cellular HPFH).73 When HbF distribution is moreheterogeneous, the antisickling effect is less pro-nounced, although SCD symptoms of homozygousbS/bS were alleviated in a few reported cases ofstable mixed chimerism as low as 11% afterAHSCT.74 It is thus likely that expression ofbA(T87Q)-globin between 20% and 30%, preferablywith moderate variegation in expression will meetthe minimum threshold to prevent most of theclinical manifestations and complications of SCDin homozygous bS/bS patients.
Cis-regulatory control elementsto be incorporated in vectors
Most of the genetic elements that control tissuespecificity and developmental switching of b-globin gene expression are located within or nearthe transcription unit in the promoter region,downstream of the polyadenylation signal, andwithin the second intron.75–78 Nevertheless, in-troducing these elements to control b-globin geneexpression in gamma-retroviral vectors (c-RV) hasresulted in very low levels of human b-globin geneexpression in erythroid cells of mice transplantedwith transduced HSCs.79,80 The discovery of chro-matin domains, referred to as DNase I hypersensi-tive sites (HS), several kilobases upstream of theb-globin gene,81,82 has shed light on the regulatoryorganization of the locus. The ability of the 15 kbLCR to confer physiological levels of human b-globin, when inserted immediately 5¢ to the b-globingene in transgenic mice,83,84 has yielded clues fordesigning effective expression systems. The mostimportant HS sites to cis-link to the human b-globingene are HS2, HS3, and HS4 that have enhanceractivity when tested individually in cell cultureand in transgenic mice.85,86 Several laboratorieshave undertaken to reduce the size of each of theHS sites while maintaining most of their en-hancer effect. However, while the so-called ‘‘coreelements’’ of 250–350 bp long show useful activitywhen combined in c-RV,87–89 expanded elementsappear necessary for near-optimal enhancer ef-fect.84,85,90,91
Gene transfer vectors suitable for the genetherapy of the b-hemoglobinopathies
Sustained gene therapy of inherited hemato-logical disorders by ex vivo gene addition requiresintegration of the vector in the chromosomes ofHSCs to obtain proper replication and segregationin the daughter cells for the lifespan of the recipi-ent. To date, only retroviral vectors have repro-ducibly shown this capability in animal models.Efforts have first made use of c-RV derived from
GENE THERAPY OF b-HEMOGLOBINOPATHIES 153

the Moloney murine leukemia virus (M-MuLV).The first M-MuLV vectors containing a human b-globin gene and its promoter were reported by theMulligan and Nienhuis laboratories.92,93 However,b-globin expression was erythroid specific but wellbelow a possible therapeutic effect.79,80,92,93
Incorporation of core LCR elements resulted inlow-titer c-RV that were also highly unstable withmultiple rearrangements of the transferred provi-ral structures.94,95 Reducing the size of the LCR tominimal elements is unsatisfactory as b-globinexpression levels are too low.96,97 Leboulch et al.88
identified untoward polyadenylation and splicingof the genomic viral RNA before packaging as themain mechanisms of both low titers and provirusrearrangements (internal splicing and poly-adenylation). LCR elements, the promoter, and theactual globin gene were placed in reverse orienta-tion to avoid splicing of the bona fide introns beforepackaging, which resulted in the unmasking ofconsensus polyadenylation and splicing signalsalong the vector genomic viral RNA. A small in-ternal deletion of sequence repeats within thesecond (minigene), together with site-directedmutagenesis eliminating those sequences, resultedin stable proviral transmission.88 Sadelain et al.89
achieved a similar result by applying the same in-ternal deletion within the second intron whilepermutating LCR elements, possibly resulting in aconformational change in the viral genomic RNAthat also inhibited untoward splicing. In spite ofthese advances, vector-encoded b-globin remainedbelow therapeutic levels, and transduction of HSCsby c-RV was suboptimal in vivo.98,99
Grants from the National Institutes of Health(NIH) were awarded in the mid-1990s to focus onmaking further progress in this area. Sadelain andLeboulch (together with R. Nagel, I. Mondon, C.Eaves, and K. Humphries) were awarded grants tofocus on the gene therapy of b-TM and SCD, re-spectively. With the discovery that the Rev re-sponsive element (RRE) system of lentiviruses,including the human immunodeficiency virus(HIV), allows for efficient nucleocytoplasmic exportand subsequent packaging of full-length, unsplicedgenomic viral RNA100 and the subsequent adventof lentiviral vectors (LVs),101,102 there appeared agreat opportunity to apply this class of vectors tothe complex genomic b-globin structures. An ad-ditional benefit is that LVs pseudotyped with theenvelope of the G protein of the vesicular stomatitisvirus (VSV-G) are amenable to concentration andare much more effective than c-RV at transducingcells arrested at the G1-S boundary of the cell cy-cle101–103 and in quiescent HSCs.104,105 Sadelain
and colleagues published the first correction ofmurine b-thalassemia by an LV containing the b-globin mini-gene described above,106,107 althoughwith substantially larger LCR elements than thoseincorporated in previous c-RV (LV referred to asTNS9). Leboulch and colleagues published the firstcorrection of transgenic mouse models of SCD,65 bymaking use of their own vector, also containing theb-globin mini-gene together with similarly largerLCR elements (644 bp for HS2, 845 bp for HS3, and1153 bp for HS4). Importantly, this LV contains theantisickling bA(T87Q)-globin gene. Human CD34+
cells transduced with a SIN version of this vectorgenerated long-term hematopoietic reconstitutionin immunodeficient mice,108 indicating effectivetransduction of human HSCs. This vector was mod-ified further in the Leboulch lab to yield HPV569109
and BB305110 vectors, which have become the basisfor our own clinical trials described in this review.We and other groups have subsequently publishedother b-globin- or c-globin-based LVs and reportedefficient transduction of mouse HSCs and high levelsof b-globin expression.111–114
REGULATORY APPROVALPROCESS IN THE UNITED STATESAND THE EUROPEAN UNION
In spite of existing regulatory frameworks forgene therapy projects,115,116 there are still manyunknowns, in part because of the lack of precedents.Innovative regulatory science can help foster asupportive ecosystem for gene therapy products andaccelerate their development in a sustainablemanner. An example of such a framework is theadaptive biomedical innovation framework.117
Frequent, science-based, transparent, and pro-active interactions with regulators can also con-tribute to the overall development strategy for genetherapy products by reducing regulatory uncer-tainty. The majority of the regulatory mechanismsthat exist to accelerate the development of medic-inal products, including gene therapy products,require (1) targeting a serious or life-threateningcondition with significant unmet medical need, (2)having the potential for outstanding efficacy withan acceptable safety profile, and (3) having a robustdevelopment plan. A clear mechanism of action andearly proof-of-concept studies are also helpful.
Typically, the first interaction with regulatoryagencies in the development of a gene therapyproduct is to discuss the nonclinical development(traditionally referred to as pharmacology andtoxicology) and the design of the planned first-in-human study. In the United States, this first in-
154 NEGRE ET AL.

teraction is a pre–Investigational New Drug (IND)meeting with the Food and Drug Administration(FDA). In the European Union (EU), this first in-teraction can be a presubmission meeting beforefiling a clinical trial application (CTA) with na-tional authorities and/or a scientific advice meetingwith the European Medicines Agency (EMA). Inaddition, ‘‘pre-pre-IND’’ informal discussions withFDA and meetings with the EMA Innovation TaskForce can be useful to discuss the early develop-ment of very innovative products, including inno-vative gene therapy products. Seeking orphandesignation for products developed for rare dis-eases (defined as fewer than 5/10,000 affected in-dividuals in the EU118 and fewer than 200,000prevalent cases in the United States),119 as well asseeking EU gene therapy,120 can also be helpfulearly on for sponsors to define their products andtheir active substance, clarify their mechanism ofaction, and outline their potential for significantclinical benefit.
Clinical trials are regulated under Directive2001/20/EC in the EU and under 21 CFR Part 312in the United States. In the EU, a new regulation121
will come into force in 2016 that will replace thecurrent directive. This regulation should stream-line the process and allow for centralized submis-sions via a new electronic portal system to onereference member state. For gene therapy prod-
ucts, seeking approval to initiate a clinical trial inthe EU currently requires filing a CTA including aninvestigational medicinal product dossier (IMPD)in each member state involved, and requires filing agenetically modified organism (‘‘GMO’’) submissionsometimes in advance of the CTA. GMO submis-sions for gene therapy product are cumbersome andoften unclear. In the future, specific applicationscustomized for gene therapy medicinal productsshould be created and these should be managed bythe same regulatory agencies receiving the CTAapplications. In the United States, filing of an INDapplication is required. INDs follow the commontechnical dossier (CTD) format of the internationalconference harmonization (ICH). IMPDs headingsare also consistent at a high level with the CTDorganization (Fig. 2).122 IMPDs and INDs mustcontain information on manufacturing and quality,on the nonclinical studies conducted, and on theplanned clinical trial, including a protocol and In-vestigator’s Brochure. For gene therapy products,filing a pediatric investigational plan (PIP) withEMA early in the development is advisable if thetargeted condition affects pediatric patients. Dur-ing this process, sponsors can receive advice ontheir preliminary long-term development plans.
Other key regulatory mechanisms that can beleveraged to accelerate the development of genetherapy products include seeking fast-track desig-
Figure 2. The common technical document, adapted from www.ich.org/products/ctd.html The common technical document is organized into five modules.Module 1 is region specific, and modules 2–5 are intended to be common for all regions.
GENE THERAPY OF b-HEMOGLOBINOPATHIES 155

nation or breakthrough therapy designation in theUnited States.123 In the EU, in addition to scientificadvice, applying for an EMA nonclinical and quali-ty/manufacturing certification procedure should beconsidered (for small and medium-size enterprises),as well as seeking the recently created prioritymedicines (PRIME) designation,91 and leveragingthe flexibility of the risk-based approach and of theadvanced therapy medicinal product (ATMP) Reg-ulation No. 1394/2007/EC, particularly for thequality/manufacturing development.124,125
InthespecificcasestudyofHPV569andBB305drugcandidates, a number of above-mentioned existingregulatory mechanisms were used to support and ac-celeratedevelopment,withanemphasisonevaluatingearly the entire life-span of the product in a creativeway, including leveraging the potential of real-worldevidence and ‘‘Safe Harbor’’ discussions with multiplestakeholders, including patients’ representatives.
Looking forward, in order to obtain marketingauthorization, a biological license application (BLA)must be submitted to the FDA in the United Statesin accordance with 21 CRF Part 601.126 In the EU, amarketing authorization application (MAA) mustbe submitted to the EMA in accordance with Direc-tive 2001/83/EC and the advanced therapy medici-nal product (ATMP) regulation 1394/2007/EC.127 Inboth regions, mechanisms exist to accelerate thereview of the BLA or MAA: priority review and ac-celerated assessment, respectively.123,128 Mechan-isms also exist to provide flexibility in the timing toobtain approval. In the United States, ‘‘acceleratedapproval’’ is the only mechanism based on the use ofa surrogate or intermediate endpoint.123 In the EU,centralized marketing authorizations that aremeant to accelerate access of medicines to patientsin need for serious conditions can be granted ‘‘underexceptional circumstances’’ or as a ‘‘conditional’’MAA.129,130
Gene therapy products provide unique opportu-nities to contribute to the evolution of regulatoryscience because of their transformative efficacypotential, their increasing complexity, and theirpending individualized nature.
HPV569 AND BB305 VECTOR DEVELOPMENTThe HPV569 lentiviral vector was assessedin the first human trial LG001
The original LVs we used to achieve long-termcorrection of SCD and b-thalassemia in mice65,112
comprised (1) HIV1’s RRE and central polypurinetract; (2) the human bA-globin gene, in reverse ori-entation, either wild-type or with the bT87Q muta-tion; (3) the human bA-globin promoter, and (4) amini-LCR composed of HS2, HS3, and HS4 of 644,845, and 1153 bp length, respectively.
To prepare for human clinical trials, safety mod-ifications were made to the original vector, as fol-lows, to yield the vector referred to as HPV569.109,112
These include mutating the GAG gene and deletionof the viral enhancer and promoter elements in theU3 region of the 3¢LTR to generate a SIN vector(Fig. 3). The SIN modification reduces the likeli-hood of propagation of replication-competent re-combinant LV in the vector producer and targetcells,105,131 decreases the risk of mobilization of thevector genome upon HIV secondary infection,132 andreduces the residual activation of cellular onco-genes by enhancer/promoter activities of integratedLTRs.133,134 Another modification has concerned theU5 region of the 3¢LTR, which was replaced by anartificial polyadenylation/termination signal de-rived from the rabbit b-globin gene.135 This lattermodification yielded higher viral titers for the SINversions of the HPV569 vector because of more effi-cient polyadenylation/termination of the viral tran-script.136 In an effort to protect transduced cells
Figure 3. Diagram of the HPV569 b-globin (bA-T87Q) lentiviral vector. The 3¢ b-globin enhancer, the 372 bp IVS2 deletion, the bA-T87Q mutation (ACA[Thr] toCAG[Gln]), and DNase I hypersensitive sites (HS) 2, HS3, and HS4 of the human b-globin locus control region (LCR) are indicated. Safety modifications,including the 2 stop codons in the w+ packaging signal, the 400 bp deletion in the U3 of the right HIV LTR, the rabbit b-globin polyA signal, and the 2 · 250 bpcHS4 chromatin insulators, are indicated. In the BB305 lentiviral vector, U3 promoter/enhancer has been replaced by cytomeglovirus (CMV) promoter/enhancerand the 2 · 250 bp cHS4 insulator elements have been removed. cPPT/flap, central polypurine tract; HIV LTR, human immunodeficiency type-1 virus longterminal repeat; ppt, polypurine tract; RRE, Rev-responsive element; bp, human b-globin promoter.
156 NEGRE ET AL.

against cis activation of adjacent genes by enhanceractivities present within the b-globin LCR,137 twocopies of the 250 bp core elements of the chicken b-globin chromatin insulator (chicken b-globin locusDNase I hypersensitive site 4: cHS4) were insertedin place of the deleted U3 region of the vector.
The HPV569 vector109 was produced at clinicalgrade68 and used to evaluate therapeutic efficacyand safety in a mouse model of b-thalassemia.138
The gene-corrected mice showed normalization oftheir phenotype. Consistent with other LV stud-ies,139,140 the HPV569 vector was shown to favortargeting of transcription units (&70%) and gene-dense regions. Importantly, no enrichment of in-tegration sites within proto-oncogenes was foundposttransplantation, and no relationship was seenbetween the proximity of integration sites to on-cogenes and site abundance in mice.138 A variety ofcomplementary efficacy and safety studies wereperformed in human CD34+ cells from patients andin several mouse models before filing a CTA dossierwith the French regulatory agency, as described inthe section Regulatory approval process in theUnited States and the European Union. The firstclinical trial with the HPV569 drug product can-didate vector (LG001) was initiated in 2006 inFrance. Three subjects with b-TM were treated andone (subject No. 1003) became transfusion inde-pendent (see details below).141
The BB305 lentiviral vectorAlthough the LG001 trial brought the proof-of-
principle of clinical efficacy in a human patient(subject No. 1003), partial clonal dominance, whichsubsequently resulted, was observed upon vectorintegration within the HMGA2 gene. This inte-gration triggered abnormal splicing of the endog-enous HMGA2 RNA using a cryptic acceptor sitewithin the cHS4 insulator core in the left LTR.Studies performed in mouse samples showed thatmost of the integrated provectors had lost one of thetwo 250 bp cHS4 insulator cores at each end.138
This phenomenon was observed in subject No. 1003as well.141 Data reported by several contributorsalso showed that inclusion of cHS4 chromatin in-sulator elements in the 3¢LTR of LVs reduced theirtiters and transduction efficacy for human CD34+
cells.142–144 Moreover, their presence had limitedefficacy on transgene expression in human hema-topoietic cells in vitro and in vivo,144 while theprotection provided was dependent on the locationof cHS4 integration in the genome.145 For all thesereasons (decreased titers and transduction effi-ciency, insulator loss, untoward splicing, and lim-ited enhancer blocking activity), removal of the
cHS4 insulator from our HPV569 vector was war-ranted.
In addition, we decided to replace the Tat-dependent U3 promoter/enhancer of the 5¢LTR bythat of the cytomegalovirus (CMV) (Fig. 3) in orderto further increase the titer of the vector.105 Over-all, the modified vector, referred to as the BB305vector,110 is identical to HPV569 after chromo-somal integration, except for the absence of thecHS4 insulator (Fig. 3). In plasmid form, however,as used in producer cells, the CMV promoter nowdrives transcription instead of the 5¢LTR.
The new-generation BB305 vector was pro-duced, purified, and tested on human CD34+ cellsas well as in a mouse model of b-thalassemia.110
Side-by-side comparison of HPV569 and BB305showed that the vector changes resulted inboth increased vector titers (3–4-fold) and in-creased transduction efficiency (2–3-fold). Com-prehensive toxicological mouse studies and vectorcomparison showed b-thalassemic phenotype cor-rection with no evidence of toxic effect related toany of the vectors, and similar integration patterns(mostly within RefSeq genes), without any sign ofclonal outgrowth or in vivo selection.110 In vitroimmortalization (IVIM) assays, developed by theBaum lab,146 were run in order to evaluate the riskof hematopoietic cell transformation upon vectorintegration. Both vectors (HPV569 and BB305)exhibited a strongly reduced risk of immortaliza-tion of murine hematopoietic cells as comparedwith control retroviral vectors, and LVs containingthe strong SFFV viral promoter.110
VECTOR AND DRUG PRODUCTMANUFACTURINGProduction of lentiviral vectors
Guidance on development and manufacturingof LVs, which are considered starting materials byboth U.S. and EU regulatory agencies when usedfor ex vivo transduction, is provided by FDA andEMA. This guidance provides recommendations forLV design, manufacturing, and characterization,including transducing activity, LV particle quan-tification, and testing for replication-competentlentiviruses (RCLs). bluebird bio has developeda GMP manufacturing process for large-scaleproduction (>40 liters) of third-generation VSV-G-pseudotyped HIV-1-based LVs. Clinical-grade ve-sicular stomatitis virus glycoprotein-pseudotypedlentiviral particles are produced by a plasmid-based co-transfection method. Purification is doneby chromatography and buffer is exchanged byultrafiltration before final filtration according to
GENE THERAPY OF b-HEMOGLOBINOPATHIES 157

published protocols.147,148 This method is used toproduce large-scale clinical-grade LV lots to sup-port our clinical trials.
Release tests performed on the manufacturedLV include potency and identity, safety, and pu-rity. The LV potency is determined using assaysthat measure both the concentration of viral par-ticles and infectious titer. The concentration ofvector particles is assessed by measuring the HIV-1p24 antigen. The titer is defined as the number offunctional transduction units per milliliter (TU/ml).149 Vector infectivity (or specific transducingactivity) is defined as the ratio between transduc-tion unit per milliliter and the concentration of p24[(TU/ml)/(ng/ml)]. This ratio gives a reliable pa-rameter to evaluate the quality of the vector prep-aration.150 High-quality vector preparations withan infectious titer >108 TU/ml and a particle/infectious ratio between 100 and 500 are obtained.151
The ability of transduced cells to produce thetherapeutic bT87Q-globin is verified after erythroiddifferentiation of transduced CD34+ hematopoieticcells. It is assessed by reverse-phase HPLC analy-sis of globin chains in differentiated erythroid cells.The therapeutic bT87Q-globin can be easily distin-guished from the normal b-chain, as well as fromthe bE and bS polypeptides.67,68
The generation of RCL is highly improbable be-cause of the multiplasmid packaging system andthe U3-deletion. Nevertheless, RCL testing is per-formed. It relies upon the permissive cell lineC8166-45, allowing the amplification and the de-tection of replicative-competent particles.152,153
Drug product (geneticallymodified CD34+ cells)
Once subjects have been screened and eligibil-ity has been determined, autologous hematopoi-etic CD34+ cells are procured either by apheresisof mobilized peripheral blood cells (for b-TM sub-jects) or by bone marrow harvest (for subjects withSCD). Multiple harvests may be undertaken ifneeded to meet the minimum cell dose required fordrug product manufacturing and untransducedbackup. Mobilization is performed with filgrastim,a recombinant form of granulocyte-colony stimu-lating factor (G-CSF), in combination with pler-ixafor. The combination of plerixafor and filgrastimmay be the most effective mobilization strategyfor subjects with b-thalassemia.154,155 Apheresisis performed on the fifth day of mobilization. Theobjective is to collect sufficient cells for bothmanufacturing and for rescue therapy. After pro-curement, the CD34+ cell population is enrichedvia purification. A portion of the product is cryo-
preserved for rescue therapy. CD34+ cells are grownin serum-free medium supplemented with recom-binant human cytokines stem cell factor (SCF),thrombopoietin (TPO), and FMS-like tyrosine ki-nase receptor-3 (Flt3-L) for approximately two days.The cells are then incubated for an additional dayfor the lentiviral transduction step. After trans-duction, a portion of the cells and supernatant areremoved for release testing. The reminder of thecells is cryopreserved. After the completion of re-lease testing and disposition, the drug product, de-fined as CD34+ hematopoietic stem cells transducedwith the BB305 LV, is infused after the patient hasundergone myeloablative conditioning.
CLINICAL TRIALS: INTERIM RESULTSFROM FRANCE
The HPV569 and BB305 vectors are designedto be used as a single product for the gene therapyof the b-hemoglobinopathies (SCD and b-TM).Vector design is intended to overcome the deficitof b-globin chains in b-thalassemia and to pro-vide anti-HbS polymerizing activity in SCD. TheHPV569 vector is the first vector to have been tes-ted worldwide in an approved human trial for thegene therapy of the b-hemoglobinopathies, withthe first patient transplanted with transduced cellsin 2006.
The French trials led the way in the testing ofthe HPV569 and BB305 drug products and are theprimary focus of this review article. The BB305program has now expanded internationally, andinitial results are summarized in Table 1 and in thesection Multicenter U.S. and international trialsfor b-TM (HGB-204) and SCD (HGB-206) with theBB305 drug product. Other ongoing trials of othergene therapies for hemoglobinopathies are sum-marized in Table 2.
Study design and pretransplant conditioningClinical studies of HPV569 and BB305 drug
products run in France are nonrandomized, open-label, single-dose, phase 1/2 studies and only enrollpatients with no sibling donor. The first trial per-formed, with regulatory approval obtained in 2006,was termed LG001 and aimed to assess the HPV569drug product, as described in the section, ‘‘TheHPV569 lentiviral vector was assessed in the firsthuman trial LG001’’ above, in 10 subjects with b-TMor severe SCD. After 3 subjects had been treated, theHPV569 vector was replaced with the improvedBB305 vector and manufacturing thereof, and a newclinical trial protocol, termed HGB-205, was im-plemented to treat the remaining 7 subjects. The
158 NEGRE ET AL.

HGB-205 study is currently ongoing in France for b-TM and SCD.
In addition to the characteristics of the clinical-grade vector (high-titer, high purity of the manu-factured lot, intrinsic therapeutic globin expres-sion properties), key parameters to success include(1) the dose of CD34+ cells infused, (2) the meanvector copy number (VCN) in CD34+ cells preinfu-sion, and (3) efficient myeloablative conditioning.Because there is little convincing evidence in ani-mal models and clinical trials that effective lenti-viral gene transfer to HSCs can be achieved with ahigh degree of sustained chimerism for vector-bearing cells in the absence of extensive myeloa-blation, when a CD34+ cell dose similar to thatapplicable to human autologous settings is used, wedecided to apply a full dose of myeloablative agentwith intravenous (IV) busulfan as pretransplantconditioning regimen. However, the additional useof immunosuppressive agent (e.g., cyclophospha-mide) is not required in this autologous setting.Monitoring of busulfan conditioning to optimizeHSC transplantation is an important componentof the success156 and is meticulously performed.When the drug product manufacture is complete,the subject receives myeloablative conditioningwith IV busulfan at a starting dose of 3.2 mg/kg/dayfor 4 days with pharmacokinetics analysis. The doseand schedule of busulfan is monitored daily andmay be adjusted based upon busulfan plasma levelsin order to maintain appropriate levels for myeloa-blation (AUC exposure of 4500–5000 [lM�min]/dayfor a daily dosing regimen, over 4 days). After bu-sulfan washout for several days after the end of IVbusulfan administration, the drug product is in-fused. The subject remains hospitalized until en-graftment occurs (absolute neutrophil count [ANC]‡0.5 · 109/liter for 3 consecutive days) and the pa-tient is medically stable. Subjects are followedmonthly for the first 6 months posttransplant, andthen every 3 months through 24 months post-transplant. Subjects are then enrolled in a long-term follow-up study for an additional 13 years.
Endpoints. The primary study objective is toassess the safety, tolerability, and success of en-graftment with autologous CD34+ HSCs transducedwith the vectors encoding the human bA-T87Q-globingene after conditioning with intravenous busulfanin subjects with b-TM and severe SCD. The primaryoutcome measures are success and kinetics of HSCengraftment, incidence of transplant-related mor-tality through 100 days posttreatment, overall sur-vival, detection of RCL, characterization of anyevents of insertional mutagenesis, and monitoring of
laboratory parameters, and frequency and severityof clinical adverse events (AEs).
Secondary endpoints include therapeutic globin(HbAT87Q) expression quantified by HPLC analy-sis, VCN levels in peripheral blood (and in bonemarrow, if collected), and RBC transfusion re-quirements posttransplant. They also include theassessment of dyserythropoiesis for b-TM subjectsand VOC and ACS frequencies for SCD subjects.
Inclusion criteria. All subjects must be between 5and 35 years old. They must have transfusion-dependent (‡100 ml/kg/year of packed RBCs forb-TM) or severe SCD, confirmed by Hb studies.Subjects must be eligible for AHSCT based on insti-tutional medical guidelines, but without a suitable,willing HLA-identical sibling donor. In addition,subjects with SCD must have failed to achieve ade-quate clinical benefit after hydroxyurea treatmentfor at least 4 months unless this treatment was notindicated or not well tolerated, and have a history of 1or more of the following poor prognostic risk factors:recurrent VOC, ACS (at least 2 episodes), significantcerebral abnormality on magnetic resonance imag-ing (MRI), stroke, antierythrocyte alloimmunization(>2 antibodies), presence of SCD cardiomyopathydocumented by Doppler echocardiography, or osteo-necrosis of 2 or more joints.
All subjects must have been treated and followedfor at least the past 2 years in a specialized centerthat maintained detailed medical records, includ-ing transfusion history. Sperm preservation ortestis or ovary biopsy is offered to subjects enrolledon the study. All subjects or their parents orguardians must provide written informed consent.
Exclusion criteria. Subjects meeting any of thefollowing criteria cannot be enrolled in the study:(1) availability of HLA-identical sibling hemato-poietic cell donor, (2) clinically significant, activebacterial, viral, parasitic, or fungal infection, (3)prior or current malignancy, myeloproliferativedisorder or immunodeficiency, (4) contraindicationto anesthesia for bone marrow harvesting, (5)white blood cell count lower than 3 · 109/liter and/or platelet count lower than 120 · 109/liter, and (6)history of major organ damage.
Trial results to dateThe LG001 and HGB-205 were the first gene
therapy studies worldwide to treat b-TM and SCDsubjects, respectively, achieving the first conver-sion to long-term transfusion independence of a b-TM subject15,141,157,158 and the first evidence ofclinical benefit in SCD.159
GENE THERAPY OF b-HEMOGLOBINOPATHIES 159

b-TM patients in the first French trial LG001 withHPV569 drug product. Four subjects with b-TMwere enrolled, 3 of whom were treated between2006 and 2011, under the LG001 protocol. Subject1002 was 29 years old with b+/b0 genotype and re-ceived a dose of 0.93 · 106 cells/kg with a mean VCNof 1.3. Subject No. 1003 was 18 years old with bE/b0
genotype and received a dose of 3.9 · 106 cells/kgwith a mean VCN of 0.6. Subject 1004 was 22 yearsold with bE/b0 genotype and received a dose of4.3 · 106 cells/kg and a VCN of 0.3. Transplantationwas uneventful in all subjects. Transduced cells didnot successfully engraft in subject 1002, althoughgene marking was detected for a few months; thissubject received backup hematopoietic cells forrescue and remains transfusion dependent. Neu-trophil engraftment was reported for subject No.1003 at day +27 and for subject 1004 at day +22postinfusion. The HPV569 drug product was welltolerated, with no nonhematologic serious AE andno drug product-related AEs reported. In addition,no subject has developed vector-derived RCL, leu-kemia, or lymphoma.
Clinical benefit of treatment with HPV569 drugproduct was observed in 1 of the 2 treated subjectswho did not receive backup cells. Subject No. 1003sustained clinical benefit as evidenced by long-term transfusion independence that was achievedapproximately 1 year posttransplant15,141 andsustained through approximately 8 years post-transplant. The hemoglobin level stabilized 1.5years posttreatment, and the mean VCN per mye-loid cell was close to 0.2. A detailed report on thissubject after 33 months of follow-up has beenpublished.15,141 After approximately 7 years oftransfusion independence, this subject received afew transfusions to address symptoms of anemia.Ongoing follow-up will determine whether or notperiodic transfusion support will again be required.His total hemoglobin and levels of HbAT87Q trans-genic hemoglobin have remained generally con-sistent from years 2 through 8 postinfusion ataround 8 g/dl total Hb of which 30% is HbAT87Q.
Integration-site analysis revealed the relativedominance of a clone bearing the vector inside in-tron 3 of the HMGA2 gene, the proportion of whichreached a maximum representation of *30% of thetransduced myeloid cells (i.e.,*4% of hematopoieticcells) 15 months posttransplantation. Molecularanalyses revealed that the integrated vector hadcaused transcriptional activation of the HMGA2promoter in erythroid cells. Furthermore, a crypticsplice acceptor site (GTAT(C)6AG), located withinthe cHS4 insulator core of the 5¢LTR, generated atruncated mRNA containing HMGA2 exons 1, 2,
and 3. Cleavage/polyadenylation occurred withinthe adjacent R region of the 5¢LTR that leads to in-creased stability of the RNA because of deletion ofthe Let7 microRNA binding sites (located in exon 5)and an excess of truncated HMGA2 mRNA. Theclone has remained under homeostatic control, witha peak at *4% of hematopoietic cells 4 years aftertransplantation, gradually decreasing to approxi-mately 1% of total nucleated blood cells at 5 yearsposttransplant.158 The clone is absent from lym-phocytes. Despite declining levels of this clone, theamount of bT87Q-globin has remained stable, indi-cating that the observed therapeutic benefit is notdependent on this specific clone. The insertion ofviral vectors in the HMGA2 gene has been observedin other gene therapy studies, with no case of leu-kemia or lymphoma related to this insertion site inany patient.158,160–162
Transduced cells successfully engrafted in sub-ject No. 1004, but this subject did not achieve sus-tained clinical benefit and remains transfusiondependent. Hemoglobin levels containing bT87Q-globin account for *5% of total hemoglobin.158
b-TM and SCD patients in French trial HGB-205with BB305 drug product. As of November 2015,four subjects with b-TM and one subject with SCDhave been treated with BB305 drug product underthe HGB-205 protocol. Initial results have beenpresented at scientific meetings but not yet pub-lished, and the trial is ongoing.157–159 All of the fourtreated b-TM subjects have so far become transfu-sion independent, and the treated SCD subjectshows early clinical benefit.
MULTICENTER U.S. AND INTERNATIONALTRIALS FOR b-TM (HGB-204) AND SCD(HGB-206) WITH THE BB305 DRUGPRODUCT
Additional subjects with b-TM and SCD arecurrently being treated under the HGB-204 andHGB-206 protocols, in two separate internationaland U.S. studies (Table 1).163,164 Early initial re-sults have been presented at scientific meetingsbut not yet published.
CONCLUSIONS
Gene therapy for hemoglobin disorders hasmade major progress, from early discovery of b-globin regulatory elements; development of LVs,including the HPV569 and BB305 vectors de-scribed here; efficient transduction of hematopoi-etic stem cells; proof-of-principle of efficacy in
160 NEGRE ET AL.

mouse models; the first conversion to multiyeartransfusion independence of a patient with b-TM;and early clinical benefit now observed in SCD.
Successfully implementing a gene therapystrategy for the b-hemoglobinopathies involves anintegrated approach of regulatory, manufacturing,and clinical trial design, and execution. Thepromise of bringing this therapeutic modality to alarge number of patients will require the successfulindustrialization and commercialization as well asregulatory product approval.
To date, no serious adverse effects have beenattributed to ex vivo LV-based HSC gene therapyfor hemoglobinopathies, but long-term data in alarger number of patients are needed to assess therisks fully. Current vectors are designed to mini-mize the possibility of genotoxicity, including on-cogenesis, but this risk cannot yet be conclusivelyquantified or excluded. Ongoing and future clinicalstudies will allow for a more complete under-standing of the benefit–risk profile of this thera-peutic approach. Implementation of gene therapyfor b-TM and SCD to a large extent will depend onthe benefit/risk/cost ratios.
ACKNOWLEDGMENTS
We thank Eliane Gluckman (Saint-Louis hospi-tal, Paris, France), the principal investigator at the
beginning of the LG001 study; Francoise Bernau-din (CHIC, Creteil, France) for contributions tostudy design and for clinical care; and OlivierHermine and Felipe Suarez (Necker Hospital,Paris), the transplanters for the HGB-205 study.We thank Christof von Kalle, Manfred Schmidt(National Center for Tumor Disease, Heidelberg,Germany) and Frederic Bushman (University ofPennsylvania, PA) for vector insertion site ana-lyses. We thank Laure Caccavelli (Necker hospital,Paris, France) and the bluebird bio France team(CEA, Fontenay aux Roses, France) for technicalassistance. We thank Philipp Gregory, SandeepSoni, Kate Lewis, Gabor Veres, Michael Paglia,John Pierciey, and Tim Douros (bluebird bio,Cambridge) for manuscript proofreading. Thework was supported by an Industrial Chair fromFrance’s Agence Nationale pour la Recherche(ANR) awarded to Philippe Leboulch. bluebird biois the sponsor of the clinical trials and providedfunds to Necker Hospital, Assistance Publique-Hopitaux de Paris.
AUTHOR DISCLOSURE
O.N. and A.-V.E. are employees of bluebird bio,Inc. Y.B., P.L., and E.P. have financial relation-ships with bluebird bio, Inc. All other authors haveno competing interests to disclose.
REFERENCES
1. Modell B, Darlison M. Global epidemiology ofhaemoglobin disorders and derived service in-dicators. Bull World Health Organ 2008;86:480–487.
2. Piel FB, Patil AP, Howes RE, et al. Global epide-miology of sickle haemoglobin in neonates: Acontemporary geostatistical model-based map andpopulation estimates. Lancet 2013;381:142–151.
3. Weatherall DJ. The genetic control of proteinsynthesis: The haemoglobin model. J Clin PatholSuppl (R Coll Pathol) 1974;8:1–11.
4. Modell B, Khan M, Darlison M, et al. Improvedsurvival of thalassaemia major in the UK andrelation to T2* cardiovascular magnetic reso-nance. J Cardiovasc Magn Reson 2008;10:42.
5. Platt OS, Brambilla DJ, Rosse WF, et al. Mor-tality in sickle cell disease. Life expectancy andrisk factors for early death. N Engl J Med1994;330:1639–1644.
6. Telfer PT, Warburton F, Christou S, et al. Im-proved survival in thalassemia major patientson switching from desferrioxamine to com-bined chelation therapy with desferrioxa-mine and deferiprone. Haematologica 2009;94:1777–1778.
7. Borgna-Pignatti C. The life of patients withthalassemia major. Haematologica 2010;95:345–348.
8. Roseff SD. Sickle cell disease: A review. Im-munohematology 2009;25:67–74.
9. Taylor LE, Stotts NA, Humphreys J, et al. A re-view of the literature on the multiple dimensionsof chronic pain in adults with sickle cell disease.J Pain Symptom Manage 2010;40:416–435.
10. Isgro A, Gaziev J, Sodani P, et al. Progress inhematopoietic stem cell transplantation as al-logeneic cellular gene therapy in thalassemia.Ann N Y Acad Sci 2010;1202:149–154.
11. Bernaudin F, Socie G, Kuentz M, et al. Long-termresults of related myeloablative stem-cell trans-plantation to cure sickle cell disease. Blood 2007;110:2749–2756.
12. Sabloff M, Chandy M, Wang Z, et al. HLA-matchedsibling bone marrow transplantation for beta-thalassemia major. Blood 2011;117:1745–1750.
13. Lucarelli G, Isgro A, Sodani P, et al. Hemato-poietic stem cell transplantation in thalassemiaand sickle cell anemia. Cold Spring Harb Per-spect Med 2012;2:a011825.
14. Pennings G, Schots R, Liebaers I. Ethical con-siderations on preimplantation genetic diagnosisfor HLA typing to match a future child as a donorof haematopoietic stem cells to a sibling. HumReprod 2002;17:534–538.
15. Payen E, Leboulch P. Advances in stem celltransplantation and gene therapy in the beta-hemoglobinopathies. Hematology 2012;2012:276–283.
16. Caocci G, Efficace F, Ciotti F, et al. Prospectiveassessment of health related quality of life inpediatric beta-thalassemia patients followinghematopoietic stem cell transplantation. BiolBlood Marrow Transplant 2011;17:861–866.
17. Luznik L, Jones RJ, Fuchs EJ. High-dose cy-clophosphamide for graft-versus-host diseaseprevention. Curr Opin Hematol 2010;17:493–499.
18. Gaziev J, Sodani P, Polchi P, et al. Bone marrowtransplantation in adults with thalassemia: Treat-ment and long-term follow-up. Ann N Y Acad Sci2005;1054:196–205.
19. Lucarelli G, Clift RA, Galimberti M, et al. Bonemarrow transplantation in adult thalassemicpatients. Blood 1999;93:1164–1167.
GENE THERAPY OF b-HEMOGLOBINOPATHIES 161

20. Faraci M, Bekassy AN, De Fazio V, et al. Non-endocrine late complications in children afterallogeneic haematopoietic SCT. Bone MarrowTransplant 2008;41 Suppl 2:S49–S57.
21. Gaziev D, Galimberti M, Lucarelli G, et al. Bonemarrow transplantation from alternative donorsfor thalassemia: HLA-phenotypically identicalrelative and HLA-nonidentical sibling or parenttransplants. Bone Marrow Transplant 2000;25:815–821.
22. Bernard F, Auquier P, Herrmann I, et al. Healthstatus of childhood leukemia survivors who re-ceived hematopoietic cell transplantation afterBU or TBI: An LEA study. Bone Marrow Trans-plant 2014;49:709–716.
23. Chou YC, Chen RL, Lai ZS, et al. PharmacologicalInduction of Human Fetal Globin Gene inHydroxyurea-Resistant Primary Adult ErythroidCells. Mol Cell Biol 2015;35:2541–2553.
24. Guda S, Brendel C, Renella R, et al. miRNA-embedded shRNAs for Lineage-specific BCL11AKnockdown and Hemoglobin F Induction. MolTher 2015;23:1465–1474.
25. Xu J, Peng C, Sankaran VG, et al. Correction ofsickle cell disease in adult mice by interferencewith fetal hemoglobin silencing. Science 2011;334:993–996.
26. Bauer DE, Orkin SH. Hemoglobin switching’s sur-prise: The versatile transcription factor BCL11A isa master repressor of fetal hemoglobin. Curr OpinGenet Dev 2015;33:62–70.
27. Vierstra J, Reik A, Chang KH, et al. Functionalfootprinting of regulatory DNA. Nat Methods2015;12:927–930.
28. Huang X, Wang Y, Yan W, et al. Production ofgene-corrected adult beta globin protein in hu-man erythrocytes differentiated from patientiPSCs after genome editing of the sickle pointmutation. Stem Cells 2015;33:1470–1479.
29. Xu P, Tong Y, Liu XZ, et al. Both TALENs andCRISPR/Cas9 directly target the HBB IVS2-654(C>T) mutation in beta-thalassemia-derivediPSCs. Sci Rep 2015;5:12065.
30. Hoban MD, Cost GJ, Mendel MC, et al. Correctionof the sickle cell disease mutation in human he-matopoietic stem/progenitor cells. Blood 2015;125:2597–2604.
31. Rachmilewitz EA, Giardina PJ. How I treat thal-assemia. Blood 2011;118:3479–3488.
32. Rouyer Fessard P, Leroy Viard K, Domenget C,et al. Mouse beta thalassemia, a model for themembrane defects of erythrocytes in the humandisease. J Biol Chem 1990;265:20247–20251.
33. Arlet JB, Ribeil JA, Guillem F, et al. HSP70 se-questration by free alpha-globin promotes inef-fective erythropoiesis in beta-thalassaemia. Nature2014;514:242–246.
34. Mathias LA, Fisher TC, Zeng L, et al. Ineffectiveerythropoiesis in beta-thalassemia major is due
to apoptosis at the polychromatophilic normo-blast stage. Exp Hematol 2000;28:1343–1353.
35. Vigi V, Volpato S, Gaburro D, et al. The corre-lation between red-cell survival and excess ofalpha-globin synthesis in beta-thalassemia. Br JHaematol 1969;16:25–30.
36. Orkin SH, Kazazian HH Jr., Antonarakis SE, et al.Abnormal RNA processing due to the exon mu-tation of beta E-globin gene. Nature 1982;300:768–769.
37. Rees DC, Clegg JB, Weatherall DJ. Is hemo-globin instability important in the interactionbetween hemoglobin E and beta thalassemia?Blood 1998;92:2141–2146.
38. Fucharoen S, Weatherall DJ. The hemoglobin Ethalassemias. Cold Spring Harb Perspect Med2012;2. pii: a011734
39. Rund D, Fucharoen S. Genetic modifiers in he-moglobinopathies. Curr Mol Med 2008;8:600–608.
40. Vichinsky E. Hemoglobin e syndromes. Hema-tology 2007:79–83.
41. Lorey F. Asian immigration and public health inCalifornia: Thalassemia in newborns in California.J Pediatr Hematol Oncol 2000;22:564–566.
42. Vichinsky EP, MacKlin EA, Waye JS, et al.Changes in the epidemiology of thalassemia inNorth America: A new minority disease. Pedia-trics 2005;116:e818–e825.
43. Marotta CA, Wilson JT, Forget BG, et al. Humanbeta-globin messenger RNA. III. Nucleotide se-quences derived from complementary DNA. JBiol Chem 1977;252:5040–5053.
44. Rees DC, Williams TN, Gladwin MT. Sickle-celldisease. Lancet 2010;376:2018–2031.
45. Brittenham GM, Schechter AN, Noguchi CT.Hemoglobin S polymerization: Primary determi-nant of the hemolytic and clinical severity of thesickling syndromes. Blood 1985;65:183–189.
46. Bunn HF. Pathogenesis and treatment of sicklecell disease. N Engl J Med 1997;337:762–769.
47. Noguchi CT, Rodgers GP, Serjeant G, et al. Levelsof fetal hemoglobin necessary for treatment ofsickle cell disease. N Engl J Med 1988;318:96–99.
48. Turhan A, Weiss LA, Mohandas N, et al. Primaryrole for adherent leukocytes in sickle cell vas-cular occlusion: A new paradigm. Proc Natl AcadSci U S A 2002;99:3047–3051.
49. Belcher JD, Bryant CJ, Nguyen J, et al. Trans-genic sickle mice have vascular inflammation.Blood 2003;101:3953–3959.
50. Amer J, Ghoti H, Rachmilewitz E, et al. Red bloodcells, platelets and polymorphonuclear neutrophilsof patients with sickle cell disease exhibit oxida-tive stress that can be ameliorated by antioxi-dants. Br J Haematol 2006;132:108–113.
51. Szocs K. Endothelial dysfunction and reactiveoxygen species production in ischemia/reperfu-
sion and nitrate tolerance. Gen Physiol Biophys2004;23:265–295.
52. Wood KC, Hebbel RP, Granger DN. Endothelialcell NADPH oxidase mediates the cerebral mi-crovascular dysfunction in sickle cell transgenicmice. FASEB J 2005;19:989–991.
53. Gladwin MT. Revisiting the hyperhemolysisparadigm. Blood 2015;126:695–696.
54. Adachi K, Reddy LR, Surrey S. Role of hydro-phobicity of phenylalanine beta 85 and leucinebeta 88 in the acceptor pocket for valine beta 6during hemoglobin S polymerization. J Biol Chem1994;269:31563–31566.
55. Adachi K, Konitzer P, Surrey S. Role of gamma87 Gln in the inhibition of hemoglobin S poly-merization by hemoglobin F. J Biol Chem1994;269:9562–9567.
56. Reddy LR, Reddy KS, Surrey S, et al. Role ofbeta87 Thr in the beta6 Val acceptor site duringdeoxy Hb S polymerization. Biochemistry 1997;36:15992–15998.
57. Goldberg MA, Husson MA, Bunn HF. Participationof hemoglobins A and F in polymerization of sicklehemoglobin. J Biol Chem 1977;252:3414–3421.
58. Poillon WN, Kim BC, Rodgers GP, et al. Sparingeffect of hemoglobin F and hemoglobin A2 onthe polymerization of hemoglobin S at physio-logic ligand saturations. Proc Natl Acad Sci U SA 1993;90:5039–5043.
59. Powars DR, Weiss JN, Chan LS, et al. Is there athreshold level of fetal hemoglobin that ame-liorates morbidity in sickle cell anemia? Blood1984;63:921–926.
60. Thomas PW, Higgs DR, Serjeant GR. Benignclinical course in homozygous sickle cell disease:A search for predictors. J Clin Epidemiol 1997;50:121–126.
61. Benesch RE, Edalji R, Benesch R, et al. Solubi-lization of hemoglobin S by other hemoglobins.Proc Natl Acad Sci U S A 1980;77:5130–5134.
62. Bookchin RM, Nagel RL, Balazs T. Role of hybridtetramer formation in gelation of haemoglobin S.Nature 1975;256:667–668.
63. Sunshine HR, Hofrichter J, Eaton WA. Gelationof sickle cell hemoglobin in mixtures with normaladult and fetal hemoglobins. J Mol Biol 1979;133:435–467.
64. Nagel RL, Bookchin RM, Johnson J, et al.Structural bases of the inhibitory effects of he-moglobin F and hemoglobin A2 on the poly-merization of hemoglobin S. Proc Natl Acad SciU S A 1979;76:670–672.
65. Pawliuk R, Westerman KA, Fabry ME, et al. Cor-rection of sickle cell disease in transgenic mousemodels by gene therapy. Science 2001;294:2368–2371.
66. Maurer HS, Behrman RE, Honig GR. Dependenceof the oxygen affinity of blood on the presence
162 NEGRE ET AL.

of foetal or adult haemoglobin. Nature 1970;227:388–390.
67. Amin A, Bourget P, Gourmel B, et al. A sensitiveand rapid HPLC assay for semi-quantitative anal-ysis of globin chain levels in blood after trans-plantation of autologous hematopoietic stem cellstransduced by a lentiviral bA-T87Q globin vector inb-thalassemia major and sickle cell disease.EBMT 2015: 41st Annual Meeting of the EuropeanSociety for Blood and Marrow Transplantation,Istanbul (Turkey).
68. Payen E, Colomb C, Negre O, et al. Lentivirusvectors in beta-thalassemia. Methods Enzymol2012;507:109–124.
69. Camaschella C, Kattamis AC, Petroni D, et al.Different hematological phenotypes caused bythe interaction of triplicated alpha-globin genesand heterozygous beta-thalassemia. Am J He-matol 1997;55:83–88.
70. Winichagoon P, Fucharoen S, Weatherall D, et al.Concomitant inheritance of alpha-thalassemia inbeta 0-thalassemia/Hb E disease. Am J Hematol1985;20:217–222.
71. Cappellini MD, Fiorelli G, Bernini LF. Interactionbetween homozygous beta (0) thalassaemia andthe Swiss type of hereditary persistence of fetalhaemoglobin. Br J Haematol 1981;48:561–572.
72. Winichagoon P, Thonglairoam V, Fucharoen S,et al. Severity differences in beta-thalassaemia/haemoglobin E syndromes: Implication of geneticfactors. Br J Haematol 1993;83:633–639.
73. Akinsheye I, Alsultan A, Solovieff N, et al. Fetalhemoglobin in sickle cell anemia. Blood 2011;118:19–27.
74. Walters MC, Patience M, Leisenring W, et al.Stable mixed hematopoietic chimerism after bonemarrow transplantation for sickle cell anemia. BiolBlood Marrow Transplant 2001;7:665–673.
75. Chada K, Magram J, Raphael K, et al. Specificexpression of a foreign beta-globin gene in ery-throid cells of transgenic mice. Nature 1985;314:377–380.
76. Kollias G, Wrighton N, Hurst J, et al. Regulatedexpression of human A gamma-, beta-, and hy-brid gamma beta-globin genes in transgenicmice: Manipulation of the developmental ex-pression patterns. Cell 1986;46:89–94.
77. Magram J, Chada K, Costantini F. Developmentalregulation of a cloned adult beta-globin gene intransgenic mice. Nature 1985;315:338–340.
78. Townes TM, Lingrel JB, Chen HY, et al. Erythroid-specific expression of human beta-globin genes intransgenic mice. EMBO J 1985;4:1715–1723.
79. Dzierzak EA, Papayannopoulou T, Mulligan RC.Lineage-specific expression of a human beta-globingene in murine bone marrow transplant recipientsreconstituted with retrovirus-transduced stem cells.Nature 1988;331:35–41.
80. Karlsson S, Bodine DM, Perry L, et al. Expressionof the human beta-globin gene following retroviral-
mediated transfer into multipotential hematopoi-etic progenitors of mice. Proc Natl Acad Sci U S A1988;85:6062–6066.
81. Forrester WC, Thompson C, Elder JT, et al. Adevelopmentally stable chromatin structure inthe human beta-globin gene cluster. Proc NatlAcad Sci U S A 1986;83:1359–1363.
82. Tuan D, Solomon W, Li Q, et al. The ‘‘beta-like-globin’’ gene domain in human erythroid cells.Proc Natl Acad Sci U S A 1985;82:6384–6388.
83. Grosveld F, van Assendelft GB, Greaves DR, et al.Position-independent, high-level expression of thehuman beta-globin gene in transgenic mice. Cell1987;51:975–985.
84. Talbot D, Collis P, Antoniou M, et al. A dominantcontrol region from the human beta-globin locusconferring integration site-independent geneexpression. Nature 1989;338:352–355.
85. Fraser P, Hurst J, Collis P, et al. DNaseI hy-persensitive sites 1, 2 and 3 of the human beta-globin dominant control region direct position-independent expression. Nucleic Acids Res1990;18:3503–3508.
86. Fraser P, Pruzina S, Antoniou M, et al. Eachhypersensitive site of the human beta-globinlocus control region confers a different devel-opmental pattern of expression on the globingenes. Genes Dev 1993;7:106–113.
87. Emery DW, Chen H, Li Q, et al. Development of acondensed locus control region cassette andtesting in retrovirus vectors for A gamma-globin.Blood Cells Mol Dis 1998;24:322–339.
88. Leboulch P, Huang GM, Humphries RK, et al.Mutagenesis of retroviral vectors transducinghuman beta-globin gene and beta-globin locuscontrol region derivatives results in stabletransmission of an active transcriptional struc-ture. EMBO J 1994;13:3065–3076.
89. Sadelain M, Wang CH, Antoniou M, et al.Generation of a high-titer retroviral vector ca-pable of expressing high levels of the humanbeta-globin gene. Proc Natl Acad Sci U S A1995;92:6728–6732.
90. Collis P, Antoniou M, Grosveld F. Definition of theminimal requirements within the human beta-globin gene and the dominant control region forhigh level expression. EMBO J 1990;9:233–240.
91. Forrester WC, Novak U, Gelinas R, et al. Mole-cular analysis of the human beta-globin locusactivation region. Proc Natl Acad Sci U S A1989;86:5439–5443.
92. Cone RD, Weber-Benarous A, Baorto D, et al.Regulated expression of a complete human beta-globin gene encoded by a transmissible retrovi-rus vector. Mol Cell Biol 1987;7:887–897.
93. Karlsson S, Papayannopoulou T, Schweiger SG,et al. Retroviral-mediated transfer of genomicglobin genes leads to regulated production ofRNA and protein. Proc Natl Acad Sci U S A1987;84:2411–2415.
94. Gelinas R, Frazier A, Harris E. A normal level ofbeta-globin expression in erythroid cells afterretroviral cells transfer. Bone Marrow Transplant1992;9 Suppl 1:154–157.
95. Novak U, Harris EA, Forrester W, et al. High-level beta-globin expression after retroviraltransfer of locus activation region-containinghuman beta-globin gene derivatives into murineerythroleukemia cells. Proc Natl Acad Sci U S A1990;87:3386–3390.
96. Chang JC, Liu D, Kan YW. A 36-base-pair coresequence of locus control region enhances ret-rovirally transferred human beta-globin geneexpression. Proc Natl Acad Sci U S A 1992;89:3107–3110.
97. Philipsen S, Talbot D, Fraser P, et al. The beta-globin dominant control region: Hypersensitivesite 2. EMBO J 1990;9:2159–2167.
98. Raftopoulos H, Ward M, Leboulch P, et al. Long-term transfer and expression of the human beta-globin gene in a mouse transplant model. Blood1997;90:3414–3422.
99. Rivella S, Sadelain M. Genetic treatment ofsevere hemoglobinopathies: The combat againsttransgene variegation and transgene silencing.Semin Hematol 1998;35:112–125.
100. Cullen BR. Retroviruses as model systems for thestudy of nuclear RNA export pathways. Virology1998;249:203–210.
101. Naldini L, Blomer U, Gage FH, et al. Efficienttransfer, integration, and sustained long-termexpression of the transgene in adult rat brainsinjected with a lentiviral vector. Proc Natl AcadSci U S A 1996;93:11382–11388.
102. Naldini L, Blomer U, Gallay P, et al. In vivo genedelivery and stable transduction of nondividingcells by a lentiviral vector. Science 1996;272:263–267.
103. Lewis P, Hensel M, Emerman M. Human immu-nodeficiency virus infection of cells arrested inthe cell cycle. Embo J 1992;11:3053–3058.
104. Case SS, Price MA, Jordan CT, et al. Stabletransduction of quiescent CD34(+)CD38(-) hu-man hematopoietic cells by HIV-1-based lenti-viral vectors. Proc Natl Acad Sci U S A 1999;96:2988–2993.
105. Miyoshi H, Smith KA, Mosier DE, et al. Trans-duction of human CD34+ cells that mediate long-term engraftment of NOD/SCID mice by HIVvectors. Science 1999;283:682–686.
106. May C, Rivella S, Callegari J, et al. Therapeutichaemoglobin synthesis in beta-thalassaemicmice expressing lentivirus-encoded human beta-globin. Nature 2000;406:82–86.
107. Rivella S, May C, Chadburn A, et al. A novelmurine model of Cooley anemia and its rescue bylentiviral-mediated human beta-globin genetransfer. Blood 2003;101:2932–2939.
108. Imren S, Fabry ME, Westerman KA, et al. High-level beta-globin expression and preferred in-
GENE THERAPY OF b-HEMOGLOBINOPATHIES 163

tragenic integration after lentiviral transductionof human cord blood stem cells. J Clin Invest2004;114:953–962.
109. Bank A, Dorazio R, Leboulch P. A phase I/IIclinical trial of beta-globin gene therapy forbeta-thalassemia. Ann N Y Acad Sci 2005;1054:308–316.
110. Negre O, Bartholomae C, Beuzard Y, et al. Pre-clinical evaluation of efficacy and safety of animproved lentiviral vector for the treatment ofbeta-thalassemia and sickle cell disease. CurrGene Ther 2015;15:64–81.
111. Hanawa H, Hargrove PW, Kepes S, et al. Ex-tended beta-globin locus control region elementspromote consistent therapeutic expression of agamma-globin lentiviral vector in murine beta-thalassemia. Blood 2004;104:2281–2290.
112. Imren S, Payen E, Westerman KA, et al. Per-manent and panerythroid correction of murinebeta thalassemia by multiple lentiviral integra-tion in hematopoietic stem cells. Proc Natl AcadSci U S A 2002;99:14380–14385.
113. Miccio A, Cesari R, Lotti F, et al. In vivo selectionof genetically modified erythroblastic progenitorsleads to long-term correction of beta-thalassemia.Proc Natl Acad Sci U S A 2008;105:10547–10552.
114. Puthenveetil G, Scholes J, Carbonell D, et al.Successful correction of the human beta-thalassemia major phenotype using a lentiviralvector. Blood 2004;3:3.
115. Maeda D, Yamaguchi T, Ishizuka T, et al. Reg-ulatory frameworks for gene and cell therapies inJapan. Adv Exp Med Biol 2015;871:147–162.
116. Martins JS, Abreu SC, Araujo ME, et al.[Strategies and results of the oral cancer pre-vention campaign among the elderly in SaoPaulo, Brazil, 2001 to 2009]. Revista Panamer-icana De Salud Publica [Pan Am J PublicHealth] 2012;31:246–252.
117. Eichler HG, Oye K, Baird LG, et al. Adaptive li-censing: Taking the next step in the evolution ofdrug approval. Clin Pharmacol Ther 2012;91:426–437.
118. European Medicines Agency, Orphan drugs andrare diseases at a glance. Doc. Ref. EMEA/290072/2007. www.ema.europa.eu/docs/en_GB/document_library/Other/2010/01/WC500069805.pdf
119. U.S. Food and Drug Administration. Developingproducts for rare disease and conditions. www.fda.gov/ForIndustry/DevelopingProductsforRareDiseasesConditions/default.htm
120. EMA. Summaries of scientific recommendationson classification of advanced-therapy medicinalproducts. www.ema.europa.eu/ema/indexjsp?curl=pages/regulation/general/general_content_000301.jsp&mid=WC0b01ac05800862c0
121. European Commission. Q&A: New rules for clinicaltrials conducted in the EU. http://europa.eu/rapid/press-release_MEMO-14–254_en.htm
122. ICH. M4: The common technical document. www.ich.org/products/ctd.html
123. FDA. Guidance for Industry—Expedited Pro-grams for Serious Conditions—Drugs and Bio-logics. www.fda.gov/downloads/drugs/guidancecomplianceregulatoryinformation/guidances/ucm358301.pdf
124. EMA. Guideline on the Risk-Based Approach Ac-cording to Annex I, Part IV of Directive 2001/83/ECApplied to Advanced Therapy Medicinal Pro-ducts. www.emaeuropa.eu/docs/en_GB/document_library/Scientific_guideline/2013/03/WC500139748.pdf
125. EC. Consultation Document—Good Manufactur-ing Practice for Advanced Therapy MedicinalProducts. http://ec.europa.eu/health/files/advtherapies/2015_pc/publ_cons_doc_2015.pdf
126. FDA. Titel 21—Food and Drugs Chapter I—Foodand drug administration department of health andhuman services subchapter F—Biologics—Part601 Licensing. www.accessdatafdagov/scripts/cdrh/cfdocs/cfCFR/CFRSearchcfm?CFRPart=601
127. EC. Regulation (EC) No. 1394/2007 of the EuropeanParliament and of the Council of 13 November2007 on Advanced Therapy Medicinal Products andAmending Directive 2001/83/EC and Regulation(EC) No 726/2004. http://ec.europa.eu/health/files/eudralex/vol-1/reg_2007_1394/reg_2007_1394_en.pdf
128. EC. Guideline on the Scientific Application andthe Practical Arrangements Necessary to Imple-ment the Procedure for Accelerated Assess-ment Pursuant to Article 14(9) of Regulation (EC)No 726/2004. http://ec.europa.eu/health/files/committee/stamp/2015–10_stamp3/stamp_3_16_3b_guideline_on_accelerated_assessment_-_public_consultation.pdf
129. EMA. Guideline on Procedures for the Grantingof a Marketing Authorization under ExceptionalCircumsances, Pursuant to Article 14 (8) of Reg-ulation (EC) No 726/2004. www.ema.europa.eu/docs/en_GB/document_library/Regulatory_and_procedural_guideline/2009/10/WC500004883.pdf
130. EMA. Guideline on the Scientific Application andthe Practical Arrangements Necessary to Imple-ment Commission Regulation (EC) No 507/2006on the Conditional Marketing Authorisation forMedicinal Products for Human use Falling withinthe Scope of Regulation (EC) No 726/2004. www.ema.europa.eu/docs/en_GB/document_library/Regulatory_and_procedural_guideline/2015/07/WC500190555.pdf
131. Zufferey R, Dull T, Mandel RJ, et al. Self-inactivating lentivirus vector for safe and efficientin vivo gene delivery. J Virol 1998;72:9873–9880.
132. Hanawa H, Persons DA, Nienhuis AW. Mobili-zation and mechanism of transcription of inte-grated self-inactivating lentiviral vectors. J Virol2005;79:8410–8421.
133. Cesana D, Ranzani M, Volpin M, et al. Un-covering and dissecting the genotoxicity of self-
inactivating lentiviral vectors in vivo. Mol Ther2014;22:774–785.
134. Zychlinski D, Schambach A, Modlich U, et al.Physiological promoters reduce the genotoxicrisk of integrating gene vectors. Mol Ther 2008;16:718–725.
135. Levitt N, Briggs D, Gil A, et al. Definition of anefficient synthetic poly(A) site. Genes Dev 1989;3:1019–1025.
136. Iwakuma T, Cui Y, Chang LJ. Self-inactivatinglentiviral vectors with U3 and U5 modifications.Virology 1999;261:120–132.
137. Hargrove PW, Kepes S, Hanawa H, et al.Globin lentiviral vector insertions can perturbthe expression of endogenous genes in beta-thalassemic hematopoietic cells. Mol Ther 2008;16:525–533.
138. Ronen K, Negre O, Roth S, et al. Distribution oflentiviral vector integration sites in mice fol-lowing therapeutic gene transfer to treat beta-thalassemia. Mol Ther 2011;19:1273–1286.
139. Schroder AR, Shinn P, Chen H, et al. HIV-1 inte-gration in the human genome favors active genesand local hotspots. Cell 2002;110:521–529.
140. Wang GP, Ciuffi A, Leipzig J, et al. HIV inte-gration site selection: Analysis by massivelyparallel pyrosequencing reveals association withepigenetic modifications. Genome Res 2007;17:1186–1194.
141. Cavazzana-Calvo M, Payen E, Negre O, et al.Transfusion independence and HMGA2 activa-tion after gene therapy of human beta-thalassaemia. Nature 2010;467:318–322.
142. Hanawa H, Yamamoto M, Zhao H, et al. Opti-mized lentiviral vector design improves titer andtransgene expression of vectors containing thechicken beta-globin locus HS4 insulator element.Mol Ther 2009;17:667–674.
143. Nielsen TT, Jakobsson J, Rosenqvist N, et al.Incorporating double copies of a chromatin in-sulator into lentiviral vectors results in less viralintegrants. BMC Biotechnol. 2009;9:13.
144. Uchida N, Washington KN, Lap CJ, et al. ChickenHS4 insulators have minimal barrier functionamong progeny of human hematopoietic cellstransduced with an HIV1-based lentiviral vector.Mol Ther 2011;19:133–139.
145. Desprat R, Bouhassira EE. Gene specificity ofsuppression of transgene-mediated insertionaltranscriptional activation by the chicken HS4insulator. PLoS One 2009;4:e5956.
146. Modlich U, Navarro S, Zychlinski D, et al. In-sertional transformation of hematopoietic cellsby self-inactivating lentiviral and gammare-troviral vectors. Mol Ther 2009;17:1919–1928.
147. Kutner RH, Puthli S, Marino MP, et al. Simplifiedproduction and concentration of HIV-1-basedlentiviral vectors using HYPERFlask vessels andanion exchange membrane chromatography.BMC Biotechnol. 2009;9:10.
164 NEGRE ET AL.

148. Kutner RH, Zhang XY, Reiser J. Production,concentration and titration of pseudotyped HIV-1-based lentiviral vectors. Nat Protoc 2009;4:495–505.
149. Skorik C, Gorman WC, Finer M, et al. Developmentof a validated method offering a potentially stan-dardized biological assay for the titration of HIV-1based lentiviral vectors. Mol Ther 2013;21:S99.
150. Follenzi A, Naldini L. Generation of HIV-1 derivedlentiviral vectors. Methods Enzymol 2002;346:454–465.
151. Gorman WC, Skorik C, Finer M, et al. Devel-opmenent of a scale-down manufacturing modelfor generation of HIV-1 based lentiviral vectors.Mol Ther 2013;21:S100.
152. Cornetta K, Yao J, Jasti A, et al. Replication-competent lentivirus analysis of clinical gradevector products. Mol Ther 2011;19:557–566.
153. Sastry L, Xu Y, Johnson T, et al. Certification as-says for HIV-1-based vectors: Frequent passage ofgag sequences without evidence of replication-competent viruses. Mol Ther 2003;8:830–839.
154. Yannaki E, Karponi G, Zervou F, et al. Hemato-poietic stem cell mobilization for gene therapy:Superior mobilization by the combination ofgranulocyte-colony stimulating factor plus pler-ixafor in patients with beta-thalassemia major.Hum Gene Ther 2013;24:852–860.
155. Karponi G, Psatha N, Lederer CW, et al.Plerixafor+G-CSF-mobilized CD34+ cells repre-sent an optimal graft source for thalassemiagene therapy. Blood 2015;126:616–619.
156. Bourget P, Falaschi L, Suarez F, et al. [A medical-pharmaceutical partnership model as a contrib-utor to the success in conditioning regimen forallogenic hematopoietic stem cell transplanta-tion in adults: A cross-reflection on our organi-zations]. Bull Cancer 2012;99:643–653.
157. Cavazzana M, Ribeil JA, Payen E, et al. StudyHgb-205: Outcomes of gene therapy for hemo-globinopathies via transplantation of autologoushematopoietic stem cells transduced ex vivowith a lentiviral bA-T87Q-globin vector (LentiGlo-bin BB305 drug product) [abstract]. Blood 2014;124:4797.
158. Cavazzana M, Ribeil JA, Payen E, et al. Out-comes of gene therapy for beta-thalassemaimajor via transplantation of autologous hema-topoietic stem cells transduced ex vivo with alentiviral beta globin vector. Haematologica2014;99:abstract S742.
159. Cavazzana M, Ribeil JA, Payen E, et al. Out-comes of gene therapy for severe sickle diseaseand beta-thalassemia major via transplantationof autologous hematopoietic stem cells trans-duced ex vivo with a lentiviral beta AT87Q-globinvector. Blood 2015;126: Abstract 202.
160. Wang GP, Berry CC, Malani N, et al. Dynamics ofgene-modified progenitor cells analyzed by track-ing retroviral integration sites in a human SCID-X1gene therapy trial. Blood 2010;115:4356–4366.
161. Adair JE, Beard BC, Trobridge GD, et al. Ex-tended survival of glioblastoma patients afterchemoprotective HSC gene therapy. Sci TranslMed 2012;4:133ra157.
162. Braun CJ, Boztug K, Paruzynski A, et al. Genetherapy for Wiskott-Aldrich syndrome—long-term efficacy and genotoxicity. Sci Transl Med2014;6:227ra233.
163. Walters M, Rasko JE, Hongeng S, et al. Up-tdate of results from the northstar study (HGB-204): A phase 1/2 study of gene therapy forb-thalassemia major via transplantation of au-tologous hematopoietic stem cells transducedex vivo with a lentiviral bA-T87Q-globin vector(Lentiglobin BB305 drug product). Blood 2015;126): Abstract 201.
164. Kanter J, Walters M, Hsieh M, et al. Initial re-sults from study HGB-206: A phase 1 studyevaluating gene therapy by transplantation ofautologous CD34+ stem cells transduced ex vivowith the Lentiglobin BB305 lentiviral vector insubjects with severe sickle cell disease. Blood2015;126: Abstract 3233.
165. Boulad F, Maggio A, Wang X, et al. Gene ther-apy for beta-thalassemia major: The MemorialSloan Kettering experience. Cooley’s AnemiaFoundation Symposium, October 2015.
166. Lidonnici MR, Aprile A, Paleari Y, et al. Updateon gene therapy clinical trial for the treat-ment of beta thalassemia major in Italy.Cooley’s Anemia Foundation Symposium, Oc-tober 2015.
Received for publication January 14, 2016;
accepted after revision January 21, 2016.
Published online: January 22, 2016.
GENE THERAPY OF b-HEMOGLOBINOPATHIES 165


















