Galvan et al. - COL11A1 BMC Cancer 2014
12
RESEARCH ARTICLE Open Access Validation of COL11A1/procollagen 11A1 expression in TGF-β1-activated immortalised human mesenchymal cells and in stromal cells of human colon adenocarcinoma José A Galván 1,2,6 , Jorge García-Martínez 1,2 , Fernando Vázquez-Villa 2 , Marcos García-Ocaña 2,3 , Carmen García-Pravia 2,4 , Primitiva Menéndez-Rodríguez 4 , Carmen González-del Rey 4 , Luis Barneo-Serra 1,2 and Juan R de los Toyos 2,5* Abstract Background: The human COL11A1 gene has been shown to be up-regulated in stromal cells of colorectal tumours, but, so far, the immunodetection of procollagen 11A1, the primary protein product of COL11A1, has not been studied in detail in human colon adenocarcinomas. Some cancer-associated stromal cells seem to be derived from bone marrow mesenchymal cells; the expression of the COL11A1 gene and the parallel immunodetection of procollagen 11A1 have not been evaluated in these latter cells, either. Methods: We used quantitative RT-PCR and/or immunocytochemistry to study the expression of DES/desmin, VIM/ vimentin, ACTA2/αSMA (alpha smooth muscle actin) and COL11A1/procollagen 11A1 in HCT 116 human colorectal adenocarcinoma cells, in immortalised human bone marrow mesenchymal cells and in human colon adenocarcinoma-derived cultured stromal cells. The immunodetection of procollagen 11A1 was performed with the new recently described DMTX1/1E8.33 mouse monoclonal antibody. Human colon adenocarcinomas and non-malignant colon tissues were evaluated by immunohistochemistry as well. Statistical associations were sought between anti-procollagen 11A1 immunoscoring and patient clinicopathological features. Results: Procollagen 11A1 was immunodetected in human bone marrow mesenchymal cells and in human colon adenocarcinoma-associated spindle-shaped stromal cells but not in colon epithelial or stromal cells of the normal colon. This immunodetection paralleled, in both kinds of cells, that of the other mesenchymal- related biomarkers studied: vimentin and alpha smooth muscle actin, but not desmin. Thus, procollagen 11A1 + adenocarcinoma-associated stromal cells are similar to “activated myofibroblasts”. In the series of human colon adenocarcinomas here studied, a high procollagen 11A1 expression was associated with nodal involvement (p = 0.05), the development of distant metastases (p = 0.017), and advanced Dukes stages (p = 0.047). Conclusion: The immunodetection of procollagen 11A1 in cancer-associated stromal cells could be a useful biomarker for human colon adenocarcinoma characterisation. Keywords: Procollagen 11A1, Human bone marrow mesenchymal cells, Cancer-associated stromal cells, Human colon adenocarcinoma * Correspondence: [email protected] 2 Oncology University Institute of the Principality of Asturias (IUOPA), 33006 Oviedo, Spain 5 Immunology Department, School of Medicine and Health Sciences, University of Oviedo, c/ Julián Clavería s/n, 33006 Oviedo, Spain Full list of author information is available at the end of the article © 2014 Galván et al.; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated. Galván et al. BMC Cancer 2014, 14:867 http://www.biomedcentral.com/1471-2407/14/867
-
Upload
jose-a-galvan -
Category
Documents
-
view
42 -
download
1
Transcript of Galvan et al. - COL11A1 BMC Cancer 2014
Galván et al. BMC Cancer 2014, 14:867http://www.biomedcentral.com/1471-2407/14/867
RESEARCH ARTICLE Open Access
Validation of COL11A1/procollagen 11A1expression in TGF-β1-activated immortalisedhuman mesenchymal cells and in stromal cells ofhuman colon adenocarcinomaJosé A Galván1,2,6, Jorge García-Martínez1,2, Fernando Vázquez-Villa2, Marcos García-Ocaña2,3,Carmen García-Pravia2,4, Primitiva Menéndez-Rodríguez4, Carmen González-del Rey4, Luis Barneo-Serra1,2
and Juan R de los Toyos2,5*
Abstract
Background: The human COL11A1 gene has been shown to be up-regulated in stromal cells of colorectal tumours,but, so far, the immunodetection of procollagen 11A1, the primary protein product of COL11A1, has not beenstudied in detail in human colon adenocarcinomas. Some cancer-associated stromal cells seem to be derived frombone marrow mesenchymal cells; the expression of the COL11A1 gene and the parallel immunodetection ofprocollagen 11A1 have not been evaluated in these latter cells, either.
Methods: We used quantitative RT-PCR and/or immunocytochemistry to study the expression of DES/desmin, VIM/vimentin, ACTA2/αSMA (alpha smooth muscle actin) and COL11A1/procollagen 11A1 in HCT 116 human colorectaladenocarcinoma cells, in immortalised human bone marrow mesenchymal cells and in human colonadenocarcinoma-derived cultured stromal cells. The immunodetection of procollagen 11A1 was performed with thenew recently described DMTX1/1E8.33 mouse monoclonal antibody. Human colon adenocarcinomas and non-malignantcolon tissues were evaluated by immunohistochemistry as well. Statistical associations were sought betweenanti-procollagen 11A1 immunoscoring and patient clinicopathological features.
Results: Procollagen 11A1 was immunodetected in human bone marrow mesenchymal cells and in humancolon adenocarcinoma-associated spindle-shaped stromal cells but not in colon epithelial or stromal cells ofthe normal colon. This immunodetection paralleled, in both kinds of cells, that of the other mesenchymal-related biomarkers studied: vimentin and alpha smooth muscle actin, but not desmin. Thus, procollagen 11A1+
adenocarcinoma-associated stromal cells are similar to “activated myofibroblasts”. In the series of human colonadenocarcinomas here studied, a high procollagen 11A1 expression was associated with nodal involvement(p = 0.05), the development of distant metastases (p = 0.017), and advanced Dukes stages (p = 0.047).
Conclusion: The immunodetection of procollagen 11A1 in cancer-associated stromal cells could be a usefulbiomarker for human colon adenocarcinoma characterisation.
Keywords: Procollagen 11A1, Human bone marrow mesenchymal cells, Cancer-associated stromal cells, Humancolon adenocarcinoma
* Correspondence: [email protected] University Institute of the Principality of Asturias (IUOPA), 33006Oviedo, Spain5Immunology Department, School of Medicine and Health Sciences,University of Oviedo, c/ Julián Clavería s/n, 33006 Oviedo, SpainFull list of author information is available at the end of the article
© 2014 Galván et al.; licensee BioMed CentralCommons Attribution License (http://creativecreproduction in any medium, provided the orDedication waiver (http://creativecommons.orunless otherwise stated.
Ltd. This is an Open Access article distributed under the terms of the Creativeommons.org/licenses/by/4.0), which permits unrestricted use, distribution, andiginal work is properly credited. The Creative Commons Public Domaing/publicdomain/zero/1.0/) applies to the data made available in this article,

Galván et al. BMC Cancer 2014, 14:867 Page 2 of 12http://www.biomedcentral.com/1471-2407/14/867
BackgroundA wealth of studies have reported that the COL11A1 hu-man gene, coding for the α1 chain of procollagen andmature collagen of type XI, which is an extracellularminor fibrillar collagen, is up-regulated in some humantumours and in mesenchymal-derived tumour cell lines[1-32], as well as in mesenchymal stem cells and osteo-blasts [33-35].Collagen polypeptides are synthesized as procollagens,
with the N- and C-propeptides at the ends of the proto-typical collagen triple helix. Upon secretion, the propep-tides are excised and then the mature collagen moleculesassemble in fibrils.In tumours, the expression of the COL11A1 gene is
currently associated to a fibroblast-like stromal pheno-type [12,19] but the origin and nature of the cells whichproduce procollagen and collagen 11A1 remain contro-versial to some extent [26].The so-called cancer-associated stromal cells, resulting
from the desmoplastic reaction which accompanies thedevelopment of human invasive carcinomas, comprisecells of different types, and are at least in part derivedfrom mesenchymal progenitors and local resident cells. Itis also well-established that TGF-β1 in cancer promotesthe activation of cancer-associated stromal cells [36].For the present study, we set out to verify the expression
of the COL11A1 gene, by quantitative RT-PCR in TGF-β1-exposed epithelial human colorectal HCT 116 cellsand Immortalised Human Bone Marrow MesenchymalCells (hTERT-HMCs); and the expression of procollagen11A1 by immunocytochemistry (ICC)/immunohisto-chemistry (IHC), using the DMTX1/1E8.33 monoclo-nal antibody (mAb) [37], on those cell cultures as wellas on biopsies of human colon adenocarcinomas. Concur-rently, we studied the expression of DES/desmin,VIM/vimentin and ACTA2/αSMA (alpha smooth muscle actin)as mesenchymal (myofibroblast)/stromal markers.Within the N-propeptide of human procollagen 11A1,
it is the so-called “variable region”, the most divergentamino acid sequence stretch among different procollagens.The DMTX1/1E8.33 mAb recognises an epitope in theYNYGTMESYQTEAPR amino acid stretch within thevariable region of human procollagen 11A1 [37].
MethodsCell culturesAscorbate is a well-known inducer of the synthesis of somecollagens [38,39]; thus, to favour the expression of procol-lagen 11A1, cells were habitually cultured with this supple-ment. Since TGF-β1 levels are increased in the serum ofpatients with invasive carcinomas [40], we chose to analyseits effects after continued and protracted exposure of cellcultures to this cytokine.
The human colorectal adenocarcinoma HCT 116(CCL-247) cell line, derived from a primary tumour,was obtained from the American Type Culture Collec-tion (ATCC) and cultured in DMEM, supplemented with1 mM sodium pyruvate (Biochrom), 2 mM L-glutamine(Biochrom), 1X non-essential amino acids (Biochrom),10% foetal bovine serum (Biochrom), and ascorbate 2-phosphate (37.5 μg/ml) (Wako Chemicals).Immortalised Human Bone Marrow Mesenchymal
Cells-hTERT (hTERT-HMCs) were obtained from AppliedBiological Materials (ABM) Inc., Richmond, BC, Canada(Cat. No. T0523), and grown in T25 ECM-coated flasks inPrigrow II medium (ABM, Cat. No. TM002), with theaddition of 10% foetal bovine serum, 1 μM hydrocortisone(Sigma) and ascorbate 2-phosphate (37.5 μg/ml) (WakoChemicals).For TGF-β1 induction, media were further supplemented
with 10 ng/ml of recombinant TGF-β1 (Peprotech). Themedium was replaced every 3–4 days and the cells werecultured for at least 15 days.All the cultures were carried out in a humidified atmos-
phere of 5% CO2 in air at 37°C.Culture passages and cell collections were done with
trypsin/EDTA 0.05%/0.02% (Biochrom). Three differentharvests from each cell culture type were obtained; forQ-RT-PCR, fresh cell pellets were kept at −80°C.
Colon adenocarcinoma stromal cells isolation and cultureFresh human tissue samples were procured after writteninformed consent of the patients and approval by thePrincipality of Asturias Ethics Committee of ClinicalResearch, Oviedo, Spain.Short-term cultures of colon adenocarcinoma stromal
cells were carried out, as previously described [41], fromsamples of tumoral sites, avoiding necrotic areas. A sam-ple from the operating theatre was directly transferredto a sterile tube containing DMEM culture medium (Gibco,Invitrogen), supplemented with vancomycin (40 μg/ml) andamikacin (40 μg/ml) (Normon Laboratories, Madrid,Spain), and stored for 24 hours at 4°C.After three washings with phosphate buffer saline
(PBS), the sample was cut into several small fragments.These fragments were first incubated with collagenases(Type I 2 mg/ml, Sigma) for 1.5 hours and then centri-fuged to eliminate supernatant; subsequently, the pelletwas subjected to a second incubation in trypsin/EDTAfor 30 min. After digestion, the cells were again collectedin a pellet, resuspended in DMEM culture medium, sup-plemented with 10% foetal bovine serum, L-glutamine andpenicillin/streptomycin, transferred to T-flasks and culti-vated in 5% CO2 at 37°C.Stromal cell cultures were stable up to 5–6 passages
before going into senescence. The purity of these stromal

Galván et al. BMC Cancer 2014, 14:867 Page 3 of 12http://www.biomedcentral.com/1471-2407/14/867
cell cultures was assessed by morphology and by immuno-staining for vimentin.
Q-RT-PCRFor normalisation of data, quantitative RT-PCR of DES,VIM, ACTA2 and COL11A1 mRNA, and PUM1, RPL10,and GAPDH mRNA was performed using the BioMark™HD System of the Fluidigm technology (Fluidigm, SanFrancisco, USA).Briefly, total RNA was isolated from pooled cell cul-
tures, kept at −80°C, with the RNeasy Mini kit (Qiagen).cDNA was synthesized from 100 ng of RNA from eachsample, using the AffinityScript Multiple TemperaturecDNA Synthesis kit (Agilent Technologies). A pre-amplification was carried out, applying the QIAGEN®Multiplex PCR Kit and the pool of all the 20x TaqMan®Gene Expression Assays. Real time Q-PCR reactionswere carried out with the TaqMan Universal PCR MasterMix kit (Applied Biosystems). Further details, accordingto Applied Biosystems’ recommendations, are in Table 1.Data were normalised by applying the ΔCt method,
after PCR efficiency corrections. These analyses wereperformed by Progenika Biopharma, S.A., Derio, Spain.Three independent samples (n =3) of different cell har-
vests of each cell type were studied. Data are presentedas mean and SEM. For each gene, differences betweencell culture expressions were analysed by a two-tailedunpaired t-test. A P value <0.01 was considered statisticallysignificant.
Immunohistochemistry (IHC)For immunohistochemical techniques, a cohort of 51 pa-tients with colon adenocarcinoma and 6 patients diagnosedwith incipient bowel infarction were collected from theArchive of the Pathology Department, Asturias CentralUniversity Hospital, with the Principality of AsturiasEthics Committee of Clinical Research, Oviedo, Spain,approval for guidelines on ethical procedures. The sam-ples had been fixed with 10% formaldehyde for 24 h andembedded in paraffin.
Table 1 Assays selected and PCR conditions for Q-RT-PCRof mRNA analysis
Gene Assay ID
COL11A1 Hs01097664_m1
GAPDH Hs02758991_g1
PUM1 Hs004472881_m1
RPL10 Hs00749196_s1
DES Hs00157258_m1
ACTA2 Hs00426835_g1
VIM Hs00185584_m1
PCR conditions were: 50°C – 2 min; 95°C – 10 min; and 40 amplification cycles:95°C – 15 sec and 60°C – 1 min.
Three-μm thick tissue sections were stained withHematoxylin and Eosin (H&E) for histological examin-ation. Antigen retrieval was performed by heating inPTLink (DakoCytomation, Denmark) in buffer solutionat high pH for 20 minutes. Endogenous peroxidase ac-tivity was blocked with Peroxidase Blocking Reagent(DakoCytomation, Denmark) for 5 minutes. After that,samples were first incubated at 37°C with the primaryantibodies described in Table 2. Subsequently, the EnVisionsystem (HRP Flex) (DakoCytomation) was applied for30 minutes at room temperature. Then, the samples werestained with DAB (3-3′-Diaminobenzidine) (DakoCytoma-tion, Denmark) for 10 minutes, counterstained for 10 mi-nutes with hematoxylin (DakoCytomation), dehydrated andmounted in Entellan® (Merck, Germany). Finally, thestained tissue sections were studied and photographed(40× objective) under a light microscope (Nikon -Eclipse 80i).
Immunocytochemistry (ICC)Cells were fixed in 10% formaldehyde for 10 minutes inthe chamber slide (BD Falcon™, ref. 354114). Endogen-ous peroxidase activity was blocked with PeroxidaseBlocking Reagent (DakoCytomation, Denmark) for 5 minutes.Permeabilization step was performed adding wash buffer1× (DakoCytomation, Denmark) which contains 0.05 mol/LTris/HCl, 0.15 mol/L NaCl, 0.05% Tween-20 [41]. Primaryantibodies were applied, as described in Table 2, at roomtemperature. After that, slides were incubated with theEnVision system (HRP Flex) for 10 minutes at roomtemperature. Then, the samples were visualised with DABfor 5 minutes, and counterstained with hematoxylin for5 minutes. Finally, the stained slides were dehydrated,mounted, studied and photographed as above.
Immunohistochemistry assessmentSpecimens were assessed by three observers (JAG, CGPand CGR), following these criteria: procollagen 11A1 im-munostaining was evaluated according to the cytoplasmaticsignal as the product of two parameters: extent of immuno-reactivity, which was evaluated in the most densely stainedarea (hot spot) under the 20× objective and scored on a0–3 scale, according to the proportion of positive fibro-blasts: (0) 0%; (1) <10%; (2) 10-50% and (3) >50%; andgranularity in the cytoplasm, evaluated as dispersed vs.confluent (1 and 2 points, respectively), with the 40×objective. Immunoscore values ranged from 0 to 6.Adjacent non-malignant tissue was used as a negativecontrol.
Statistical analysisThe experimental results were tested for significanceemploying the χ2 test (with Yates’ correction, when ap-propriate). The statistical analysis was carried out with
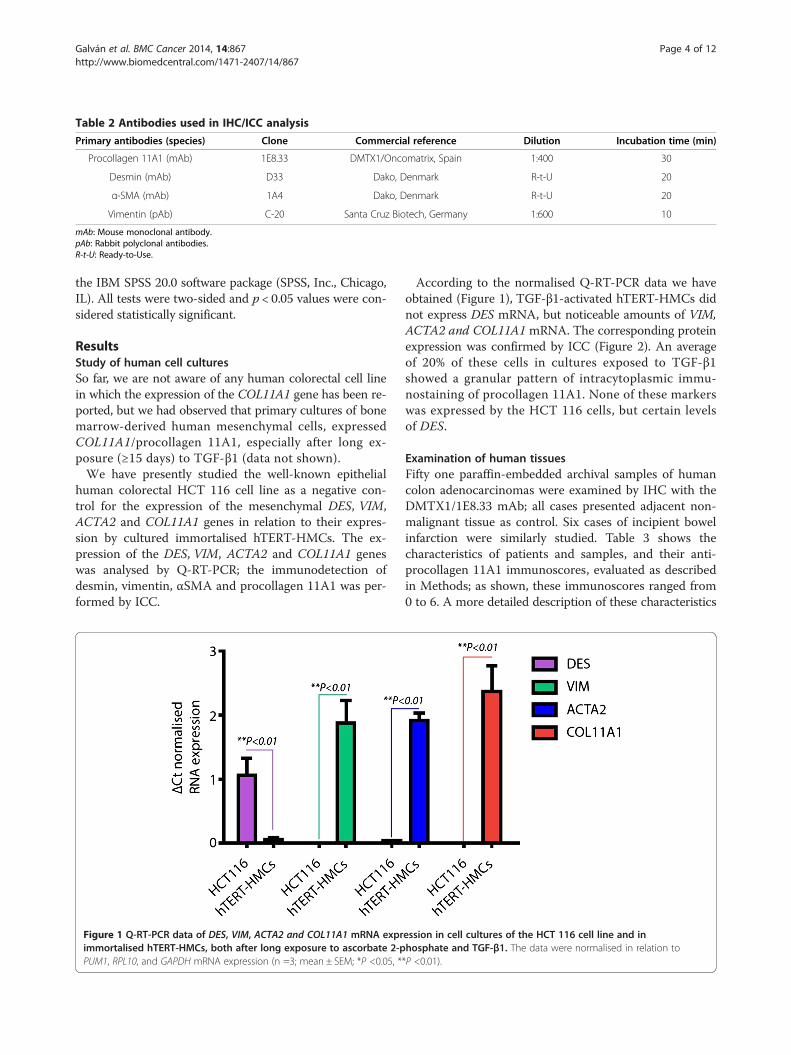
Table 2 Antibodies used in IHC/ICC analysis
Primary antibodies (species) Clone Commercial reference Dilution Incubation time (min)
Procollagen 11A1 (mAb) 1E8.33 DMTX1/Oncomatrix, Spain 1:400 30
Desmin (mAb) D33 Dako, Denmark R-t-U 20
α-SMA (mAb) 1A4 Dako, Denmark R-t-U 20
Vimentin (pAb) C-20 Santa Cruz Biotech, Germany 1:600 10
mAb: Mouse monoclonal antibody.pAb: Rabbit polyclonal antibodies.R-t-U: Ready-to-Use.
Galván et al. BMC Cancer 2014, 14:867 Page 4 of 12http://www.biomedcentral.com/1471-2407/14/867
the IBM SPSS 20.0 software package (SPSS, Inc., Chicago,IL). All tests were two-sided and p < 0.05 values were con-sidered statistically significant.
ResultsStudy of human cell culturesSo far, we are not aware of any human colorectal cell linein which the expression of the COL11A1 gene has been re-ported, but we had observed that primary cultures of bonemarrow-derived human mesenchymal cells, expressedCOL11A1/procollagen 11A1, especially after long ex-posure (≥15 days) to TGF-β1 (data not shown).We have presently studied the well-known epithelial
human colorectal HCT 116 cell line as a negative con-trol for the expression of the mesenchymal DES, VIM,ACTA2 and COL11A1 genes in relation to their expres-sion by cultured immortalised hTERT-HMCs. The ex-pression of the DES, VIM, ACTA2 and COL11A1 geneswas analysed by Q-RT-PCR; the immunodetection ofdesmin, vimentin, αSMA and procollagen 11A1 was per-formed by ICC.
Figure 1 Q-RT-PCR data of DES, VIM, ACTA2 and COL11A1 mRNA exprimmortalised hTERT-HMCs, both after long exposure to ascorbate 2-pPUM1, RPL10, and GAPDH mRNA expression (n =3; mean ± SEM; *P <0.05, *
According to the normalised Q-RT-PCR data we haveobtained (Figure 1), TGF-β1-activated hTERT-HMCs didnot express DES mRNA, but noticeable amounts of VIM,ACTA2 and COL11A1 mRNA. The corresponding proteinexpression was confirmed by ICC (Figure 2). An averageof 20% of these cells in cultures exposed to TGF-β1showed a granular pattern of intracytoplasmic immu-nostaining of procollagen 11A1. None of these markerswas expressed by the HCT 116 cells, but certain levelsof DES.
Examination of human tissuesFifty one paraffin-embedded archival samples of humancolon adenocarcinomas were examined by IHC with theDMTX1/1E8.33 mAb; all cases presented adjacent non-malignant tissue as control. Six cases of incipient bowelinfarction were similarly studied. Table 3 shows thecharacteristics of patients and samples, and their anti-procollagen 11A1 immunoscores, evaluated as describedin Methods; as shown, these immunoscores ranged from0 to 6. A more detailed description of these characteristics
ession in cell cultures of the HCT 116 cell line and inhosphate and TGF-β1. The data were normalised in relation to*P <0.01).

Figure 2 Representative immunostaining of cultured immortalised hTERT-HMCs after long exposure to ascorbate 2-phosphate andTGF-β1. A) Procollagen 11A1 B) Desmin, C) αSMA and D) Vimentin. Scale bar 50 μm (400×).
Galván et al. BMC Cancer 2014, 14:867 Page 5 of 12http://www.biomedcentral.com/1471-2407/14/867
is in Additional file 1. In three of these 51 diagnosed adeno-carcinoma cases, no procollagen 11A1 staining could bedetected, in spite of extensive re-examination. Procollagen11A1 was neither immunodetected in the adjacent non-malignant tissues nor in the infarction cases.Figure 3 shows representative procollagen 11A1 im-
munostaining patterns (panels A, B, C and D); only agranular cytoplasmic staining of peritumoral spindle-shaped fibroblast-like stromal cells was observed. Thisgranularity was either dispersed, with a few granules inthe cytoplasm of stromal cells (panel G) or frankly con-fluent (panel H). No staining was observed on specimensof bowel ischemia (panel E) or on non-malignant tissues(panel F).Figure 4 shows a representative immunostaining of an
adenocarcinoma specimen with a procollagen 11A1immunoscore of 6. As shown on panel A, only peritu-moral stromal cells were stained with the anti-procollagen11A1 mAb; besides, these cells seemed to be positive forαSMA (C) and vimentin (D), but negative for desmin (B).This immunostaining pattern was reproduced (panels E, F,G and H) in stromal cells cultured from fresh specimensof the same patient.
Association between procollagen 11A1 expression andclinicopathological featuresFor statistical purposes, some variables (age at diagnosis,tumour size, anti-procollagen 11A1 immunostaining) were
divided into 2 groups, taking the median score value as acut-off point (Table 4). Patients diagnosed with ischemiawere excluded from this analysis.9/12 patients that had developed distant metastases at
diagnosis and 15/27 patients with advanced Dukes stageswere associated with high procollagen 11A1 expression(p = 0.017 and p = 0.047, respectively). The same can beobserved for 15/26 patients with nodes affected, how-ever, only with a trend towards significance (p = 0.059).
DiscussionWe have presently shown, extending our previous obser-vations [42], that procollagen 11A1, as a protein expres-sion product of the COL11A1 gene, is immunodetectedin stromal cells of human colon adenocarcinoma. Bycontrast, and contrary to a previous report [15], we havenever observed immunodetection of procollagen 11A1in epithelial cells of normal colon tissue or colon adeno-carcinoma with the DMTX1/1E8.33 mAb. This immu-nodetection was observed in 48 of the 51 cases studied.Three cases (5.9%), which were classified under the samecriteria as the rest of the cases examined as conventionaladenocarcinomas with desmoplastic reaction, did notstain; so far, we have not identified in them any charac-teristics to which this negative immunostaining could beassociated. Except for the report of Fischer et al. [1] whodid not find either the expression of COL11A in 5 out of15 (33.3%) colonic carcinomas analysed, we are not aware
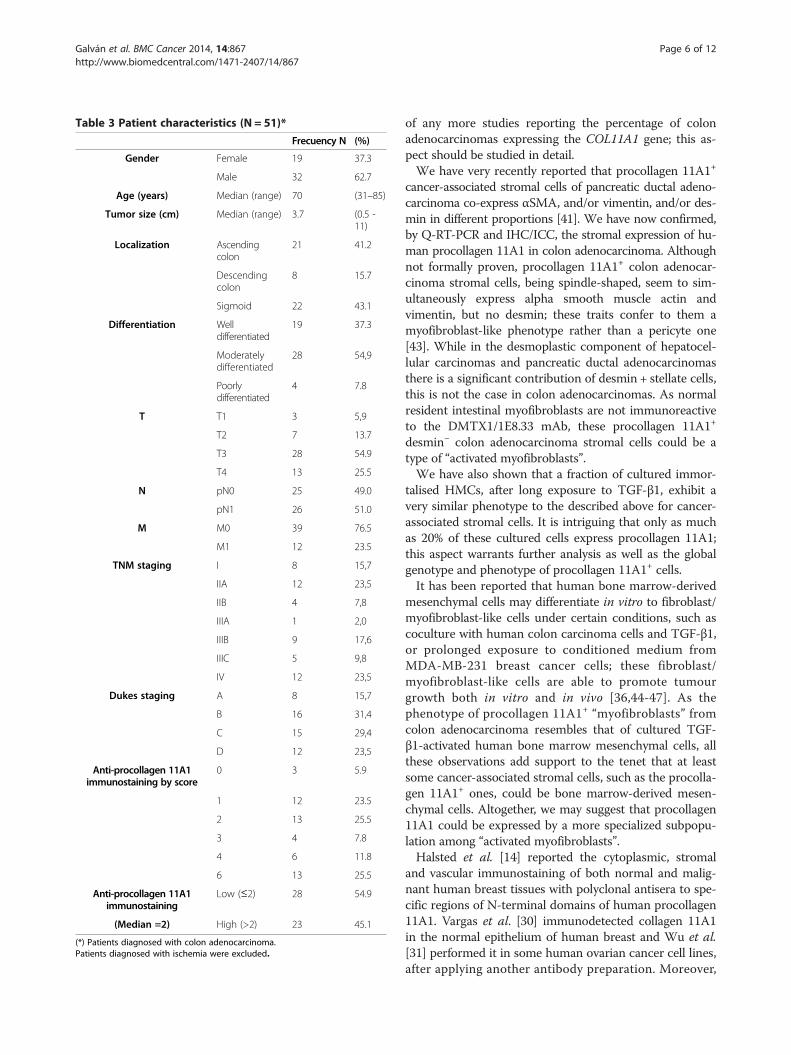
Table 3 Patient characteristics (N = 51)*
Frecuency N (%)
Gender Female 19 37.3
Male 32 62.7
Age (years) Median (range) 70 (31–85)
Tumor size (cm) Median (range) 3.7 (0.5 -11)
Localization Ascendingcolon
21 41.2
Descendingcolon
8 15.7
Sigmoid 22 43.1
Differentiation Welldifferentiated
19 37.3
Moderatelydifferentiated
28 54,9
Poorlydifferentiated
4 7.8
T T1 3 5,9
T2 7 13.7
T3 28 54.9
T4 13 25.5
N pN0 25 49.0
pN1 26 51.0
M M0 39 76.5
M1 12 23.5
TNM staging I 8 15,7
IIA 12 23,5
IIB 4 7,8
IIIA 1 2,0
IIIB 9 17,6
IIIC 5 9,8
IV 12 23,5
Dukes staging A 8 15,7
B 16 31,4
C 15 29,4
D 12 23,5
Anti-procollagen 11A1immunostaining by score
0 3 5.9
1 12 23.5
2 13 25.5
3 4 7.8
4 6 11.8
6 13 25.5
Anti-procollagen 11A1immunostaining
Low (≤2) 28 54.9
(Median =2) High (>2) 23 45.1
(*) Patients diagnosed with colon adenocarcinoma.Patients diagnosed with ischemia were excluded.
Galván et al. BMC Cancer 2014, 14:867 Page 6 of 12http://www.biomedcentral.com/1471-2407/14/867
of any more studies reporting the percentage of colonadenocarcinomas expressing the COL11A1 gene; this as-pect should be studied in detail.We have very recently reported that procollagen 11A1+
cancer-associated stromal cells of pancreatic ductal adeno-carcinoma co-express αSMA, and/or vimentin, and/or des-min in different proportions [41]. We have now confirmed,by Q-RT-PCR and IHC/ICC, the stromal expression of hu-man procollagen 11A1 in colon adenocarcinoma. Althoughnot formally proven, procollagen 11A1+ colon adenocar-cinoma stromal cells, being spindle-shaped, seem to sim-ultaneously express alpha smooth muscle actin andvimentin, but no desmin; these traits confer to them amyofibroblast-like phenotype rather than a pericyte one[43]. While in the desmoplastic component of hepatocel-lular carcinomas and pancreatic ductal adenocarcinomasthere is a significant contribution of desmin + stellate cells,this is not the case in colon adenocarcinomas. As normalresident intestinal myofibroblasts are not immunoreactiveto the DMTX1/1E8.33 mAb, these procollagen 11A1+
desmin− colon adenocarcinoma stromal cells could be atype of “activated myofibroblasts”.We have also shown that a fraction of cultured immor-
talised HMCs, after long exposure to TGF-β1, exhibit avery similar phenotype to the described above for cancer-associated stromal cells. It is intriguing that only as muchas 20% of these cultured cells express procollagen 11A1;this aspect warrants further analysis as well as the globalgenotype and phenotype of procollagen 11A1+ cells.It has been reported that human bone marrow-derived
mesenchymal cells may differentiate in vitro to fibroblast/myofibroblast-like cells under certain conditions, such ascoculture with human colon carcinoma cells and TGF-β1,or prolonged exposure to conditioned medium fromMDA-MB-231 breast cancer cells; these fibroblast/myofibroblast-like cells are able to promote tumourgrowth both in vitro and in vivo [36,44-47]. As thephenotype of procollagen 11A1+ “myofibroblasts” fromcolon adenocarcinoma resembles that of cultured TGF-β1-activated human bone marrow mesenchymal cells, allthese observations add support to the tenet that at leastsome cancer-associated stromal cells, such as the procolla-gen 11A1+ ones, could be bone marrow-derived mesen-chymal cells. Altogether, we may suggest that procollagen11A1 could be expressed by a more specialized subpopu-lation among “activated myofibroblasts”.Halsted et al. [14] reported the cytoplasmic, stromal
and vascular immunostaining of both normal and malig-nant human breast tissues with polyclonal antisera to spe-cific regions of N-terminal domains of human procollagen11A1. Vargas et al. [30] immunodetected collagen 11A1in the normal epithelium of human breast and Wu et al.[31] performed it in some human ovarian cancer cell lines,after applying another antibody preparation. Moreover,
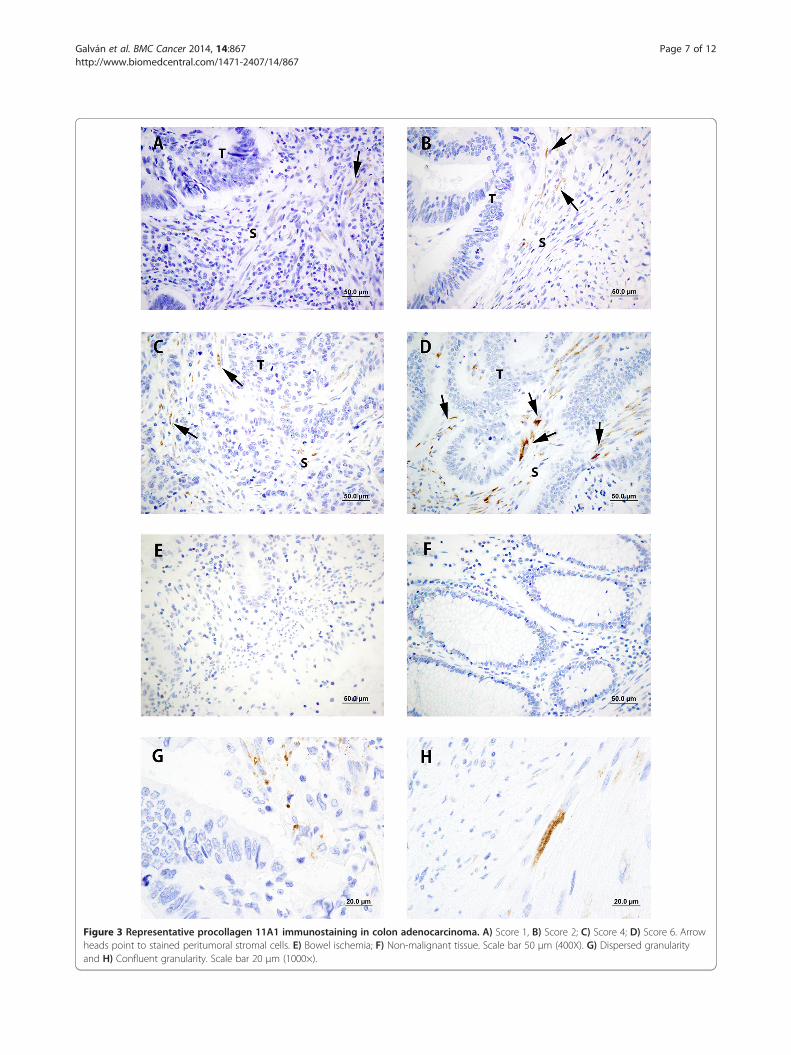
Figure 3 Representative procollagen 11A1 immunostaining in colon adenocarcinoma. A) Score 1, B) Score 2; C) Score 4; D) Score 6. Arrowheads point to stained peritumoral stromal cells. E) Bowel ischemia; F) Non-malignant tissue. Scale bar 50 μm (400X). G) Dispersed granularityand H) Confluent granularity. Scale bar 20 μm (1000×).
Galván et al. BMC Cancer 2014, 14:867 Page 7 of 12http://www.biomedcentral.com/1471-2407/14/867
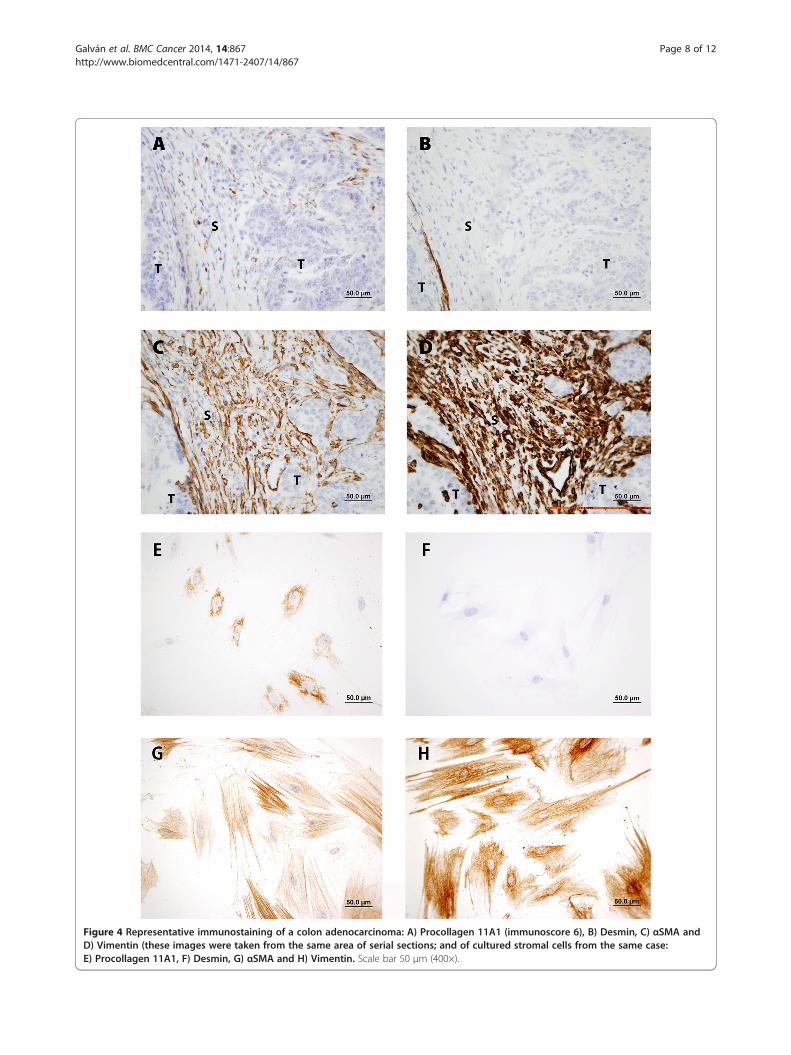
Figure 4 Representative immunostaining of a colon adenocarcinoma: A) Procollagen 11A1 (immunoscore 6), B) Desmin, C) αSMA andD) Vimentin (these images were taken from the same area of serial sections; and of cultured stromal cells from the same case:E) Procollagen 11A1, F) Desmin, G) αSMA and H) Vimentin. Scale bar 50 μm (400×).
Galván et al. BMC Cancer 2014, 14:867 Page 8 of 12http://www.biomedcentral.com/1471-2407/14/867
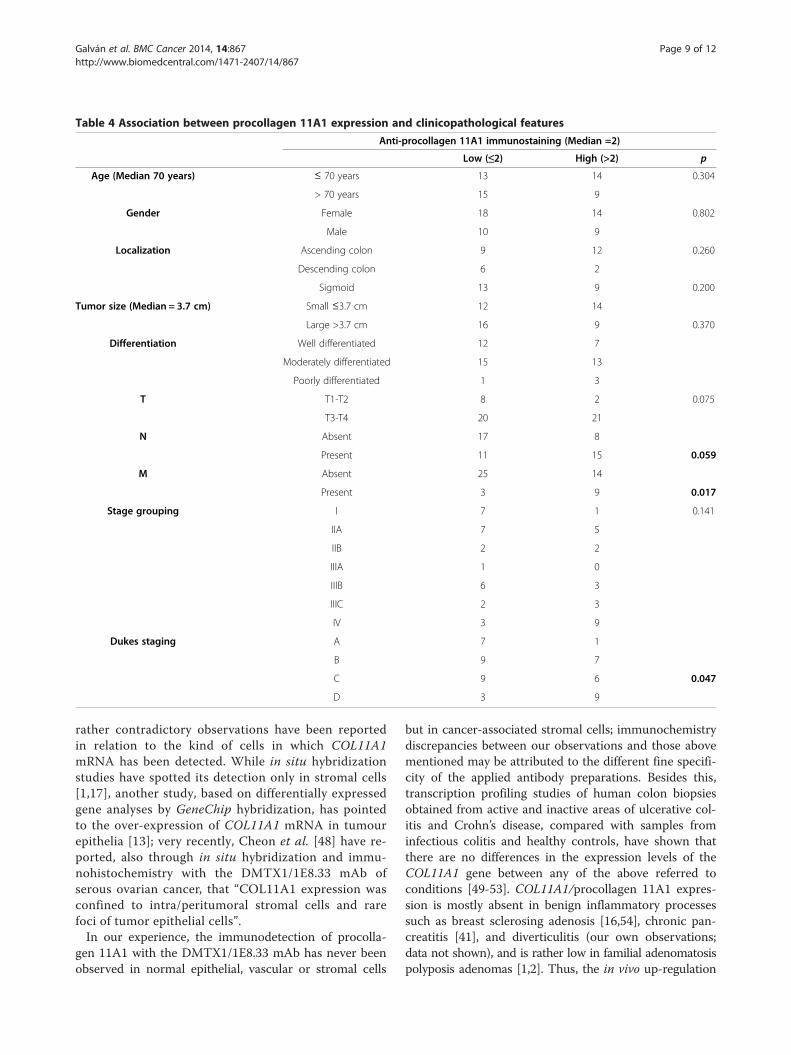
Table 4 Association between procollagen 11A1 expression and clinicopathological features
Anti-procollagen 11A1 immunostaining (Median =2)
Low (≤2) High (>2) p
Age (Median 70 years) ≤ 70 years 13 14 0.304
> 70 years 15 9
Gender Female 18 14 0.802
Male 10 9
Localization Ascending colon 9 12 0.260
Descending colon 6 2
Sigmoid 13 9 0.200
Tumor size (Median = 3.7 cm) Small ≤3.7 cm 12 14
Large >3.7 cm 16 9 0.370
Differentiation Well differentiated 12 7
Moderately differentiated 15 13
Poorly differentiated 1 3
T T1-T2 8 2 0.075
T3-T4 20 21
N Absent 17 8
Present 11 15 0.059
M Absent 25 14
Present 3 9 0.017
Stage grouping I 7 1 0.141
IIA 7 5
IIB 2 2
IIIA 1 0
IIIB 6 3
IIIC 2 3
IV 3 9
Dukes staging A 7 1
B 9 7
C 9 6 0.047
D 3 9
Galván et al. BMC Cancer 2014, 14:867 Page 9 of 12http://www.biomedcentral.com/1471-2407/14/867
rather contradictory observations have been reportedin relation to the kind of cells in which COL11A1mRNA has been detected. While in situ hybridizationstudies have spotted its detection only in stromal cells[1,17], another study, based on differentially expressedgene analyses by GeneChip hybridization, has pointedto the over-expression of COL11A1 mRNA in tumourepithelia [13]; very recently, Cheon et al. [48] have re-ported, also through in situ hybridization and immu-nohistochemistry with the DMTX1/1E8.33 mAb ofserous ovarian cancer, that “COL11A1 expression wasconfined to intra/peritumoral stromal cells and rarefoci of tumor epithelial cells”.In our experience, the immunodetection of procolla-
gen 11A1 with the DMTX1/1E8.33 mAb has never beenobserved in normal epithelial, vascular or stromal cells
but in cancer-associated stromal cells; immunochemistrydiscrepancies between our observations and those abovementioned may be attributed to the different fine specifi-city of the applied antibody preparations. Besides this,transcription profiling studies of human colon biopsiesobtained from active and inactive areas of ulcerative col-itis and Crohn’s disease, compared with samples frominfectious colitis and healthy controls, have shown thatthere are no differences in the expression levels of theCOL11A1 gene between any of the above referred toconditions [49-53]. COL11A1/procollagen 11A1 expres-sion is mostly absent in benign inflammatory processessuch as breast sclerosing adenosis [16,54], chronic pan-creatitis [41], and diverticulitis (our own observations;data not shown), and is rather low in familial adenomatosispolyposis adenomas [1,2]. Thus, the in vivo up-regulation

Galván et al. BMC Cancer 2014, 14:867 Page 10 of 12http://www.biomedcentral.com/1471-2407/14/867
of the COL11A1 gene may be considered as a biomarkerof cancer-associated stromal cells.In this study, high procollagen 11A1 immunostaining
was associated with clinicopathological variables such aslymph node involvement, advanced Dukes stages andpresence of distant metastases. These results go accordingto the role of COL11A1 in promoting carcinoma aggres-siveness and progression [11,17,20,30,31,48,55-57].
ConclusionsBased on its high specificity, our observations stress oncemore the usefulness of the DMTX1/1E8.33 mAb for can-cer research, and the clinical significance of procollagen11A1 as a very valuable biomarker to characterise cancer-associated stromal cells and to evaluate human colonadenocarcinomas.
Additional file
Additional file 1: Detailed description of patients and theirclinicopathological characteristics.
AbbreviationsHMCs: Human mesenchymal cells; hTERT-HMCs: Immortalised hTERT- humanbone marrow mesenchymal cells; ICC: Immunocytochemistry;IHC: Immunohistochemistry; mAb: Monoclonal antibody; pAb: Polyclonalantibodies; H&E: Hematoxylin and eosin; PBS: Phosphate-buffered saline;DAB: 3-3′-Diaminobenzidine.
Competing interestsThe authors declare that they have no competing interests.
Authors’ contributionsJAG carried out immunostainings, photographs and statistical analyses. Theimmunostainings were evaluated by JAG, CGP and CGR, and supervised byCGP and PMR. JGM isolated and cultured tumour stromal cells. FVV andMGO carried out other cell cultures. All authors discussed, read andapproved the final manuscript. CGP, PMR, LBS, and JRT contributed equallyto this study as senior authors.
AcknowledgementsThe authors thank Inti Zlobec for the critical reading of the manuscript andhelpful comments. The excellent technical assistance of Laura Suárez-Fernández is greatly acknowledged.This research has been co-financed by European Union ERDF Funds; by theINNPACTO-ONCOPAN IPT-010000-2010-31 Project; by the FISS-09-PS09/01911Project, Ministry of Science and Innovation, Spain; by the FC-11-PC10-23,FICYT Project, Axe 1 of the 2007–2013 ERDF Operational FrameworkProgramme of the Principality of Asturias, Spain; and by Oncomatrix, S.L.Derio, Spain.
Author details1Surgery Department, School of Medicine and Health Sciences, University ofOviedo, 33006 Oviedo, Spain. 2Oncology University Institute of thePrincipality of Asturias (IUOPA), 33006 Oviedo, Spain. 3PreparativeBiotechnology Unit, Technical-Scientific Services, University of Oviedo, 33006Oviedo, Spain. 4Pathological Anatomy Service, Asturias Central UniversityHospital (HUCA), 33006 Oviedo, Spain. 5Immunology Department, School ofMedicine and Health Sciences, University of Oviedo, c/ Julián Clavería s/n,33006 Oviedo, Spain. 6Present address: Translational Research Unit (TRU),Institute of Pathology, University of Bern, Bern, Switzerland.
Received: 29 April 2014 Accepted: 12 November 2014Published: 23 November 2014
References1. Fischer H, Stenling R, Rubio C, Lindblom A: Colorectal carcinogenesis is
associated with stromal expression of COL11A1 and COL5A2.Carcinogenesis 2001, 22:875–878. doi:10.1093/carcin/22.6.875.
2. Fischer H, Salahshor S, Stenling R, Björk J, Lindmark G, Iselius L, Rubio C,Lindblom A: COL11A1 in FAP polyps and in sporadic colorectal tumors.BMC Cancer 2001, 1:17 [http://www.biomedcentral.com/1471-2407/1/17]
3. Wang KK, Liu N, Radulovich N, Wigle DA, Johnston MR, Shepherd FA,Minden MD, Tsao MS: Novel candidate tumor marker genes for lungadenocarcinoma. Oncogene 2002, 21:7598–7604.
4. Xu SH, Qian LJ, Mou HZ, Zhu CH, Zhou XM, Liu XL, Chen Y, Bao WY:Difference of gene expression profiles between esophageal carcinomaand its pericancerous epithelium by gene chip. World J Gastroenterol2003, 9:417–422.
5. Sok JC, Kuriakose MA, Mahajan VB, Pearlman AN, DeLacure MD, Chen FA:Tissue-specific gene expression of head and neck squamous cellcarcinoma in vivo by complementary DNA microarray analysis. ArchOtolaryngol Head Neck Surg 2003, 129:760–770.
6. Schmalbach CE, Chepeha DB, Giordano TJ, Rubin MA, Teknos TN, Bradford CR,Wolf GT, Kuick R, Misek DE, Trask DK, Hanash S: Molecular profiling andthe identification of genes associated with metastatic oral cavity/pharynx squamous cell carcinoma. Arch Otolaryngol Head Neck Surg2004, 130:295–302.
7. Croner RS, Foertsch T, Brueckl WM, Guenther K, Siebenhaar R, Stremmel C,Matzel KE, Papadopoulos T, Kirchner T, Behrens J, Klein-Hitpass L, Stuerzl M,Hohenberger W, Reingruber B: Common denominator genes thatdistinguish colorectal carcinoma from normal mucosa. Int JColorectal Dis 2005, 20:353–362.
8. Barneo L, del Amo J, García-Pravia C, de los Toyos JR, Pérez-Basterrechea M,González-Pinto I, Vazquez L, Miyar A, Simón L: Identification of specificgenes by microarrays, validation and use of polyclonal antibodies inpancreatic cancer: preliminary results. In 41st Congress of the EuropeanSociety for Surgical Research-ESSR 2006. Edited by Vollmar B. Bologna, Italy:Medimond, International Proceedings; 2006:27–35.
9. del Amo-Iribarren J: Identificación de marcadores para diagnóstico diferencialy potenciales dianas terapéuticas en adenocarcinoma ductal de páncreas medianteherramientas genómicas, PhD thesis. Universidad del País Vasco, Genetics,Physical Anthropology and Animal Physiology Department; 2006.
10. Chong IW, Chang MY, Chang HC, Yu YP, Sheu CC, Tsai JR, Hung JY, ChouSH, Tsai MS, Hwang JJ, Lin SR: Great potential of a panel of multiplehMTH1, SPD, ITGA11 and COL11A1 markers for diagnosis of patientswith non-small cell lung cancer. Oncol Rep 2006, 16:981–988.
11. Vecchi M, Nuciforo P, Romagnoli S, Confalonieri S, Pellegrini C, Serio G,Quarto M, Capra M, Roviaro GC, Contessini Avesani E, Corsi C, Coggi G, DiFiore PP, Bosari S: Gene expression of early and advanced gastric cancer.Oncogene 2007, 26:4284–4294.
12. Pilarsky C, Ammerpohl O, Sipos B, Dahl E, Hartmann A, Wellmann A,Braunschweig T, Löhr M, Jesenofsky R, Friess H, Wente MN, Kristiansen G,Jahnke B, Denz A, Rückert F, Schackert HK, Klöppel G, Kalthoff H, Saeger HD,Grützmann R: Activation of Wnt signalling in stroma from pancreaticcancer identified by gene expression profiling. J Cell Mol Med 2008,12:2823–2835.
13. Badea L, Herlea V, Dima SO, Dumitrascu T, Popescu I: Combined geneexpression analysis of whole-tissue and microdissected pancreatic ductaladenocarcinoma identifies genes specifically overexpressed in tumorepithelia. Hepatogastroenterology 2008, 55:2016–2027.
14. Halsted KC, Bowen KB, Bond L, Luman SE, Jorcyk CL, Fyffe WE, Kronz JD,Oxford JT: Collagen alpha1(XI) in normal and malignant breast tissue.Mod Pathol 2008, 21:1246–1254.
15. Bowen KB, Reimers AP, Luman S, Kronz JD, Fyffe WE, Oxford JT:Immunohistochemical localization of collagen type XI alpha1 and alpha2chains in human colon tissue. J Histochem Cytochem 2008, 56:275–283.
16. Fuentes-Martínez N: Colágeno 11: Nuevo marcador en el cáncer de mama,PhD thesis. Universidad de Oviedo, Surgery and Medical Surgical SpecialitiesDepartment; 2009.
17. Zhao Y, Zhou T, Li A, Yao H, He F, Wang L, Si J: A potential role ofcollagens expression in distinguishing between premalignant andmalignant lesions in stomach. Anat Rec 2009, 292:692–700.
18. An JH, Lee SY, Jeon JY, Cho KG, Kim SU, Lee MA: Identification ofgliotropic factors that induce human stem cell migration to malignanttumor. J Proteome Res 2009, 8:2873–2881.

Galván et al. BMC Cancer 2014, 14:867 Page 11 of 12http://www.biomedcentral.com/1471-2407/14/867
19. Erkan M, Weis N, Pan Z, Schwager C, Samkharadze T, Jiang X, Wirkner U,Giese NA, Ansorge W, Debus J, Huber PE, Friess H, Abdollahi A, Kleeff J:Organ-, inflammation- and cancer specific transcriptional fingerprints ofpancreatic and hepatic stellate cells. Mol Cancer 2010, 9:88 [http://www.molecular-cancer.com/content/9/1/88]
20. Kim H, Watkinson J, Varadan V, Anastassiou D: Multi-cancer computationalanalysis reveals invasion-associated variant of desmoplastic reactioninvolving INHBA, THBS2 and COL11A1. BMC Med Genomics 2010, 3:51[http://www.biomedcentral.com/1755-8794/3/51]
21. Pavlides S, Tsirigos A, Vera I, Flomenberg N, Frank PG, Casimiro MC, Wang C,Pestell RG, Martinez-Outschoorn UE, Howell A, Sotgia F, Lisanti MP:Transcriptional evidence for the “Reverse Warburg Effect” in humanbreast cancer tumor stroma and metastasis: similarities with oxidativestress, inflammation, Alzheimer’s disease, and “Neuron-Glia MetabolicCoupling”. Aging (Albany NY) 2010, 2:185–199.
22. Chernov AV, Baranovskaya S, Golubkov VS, Wakeman DR, Snyder EY,Williams R, Strongin AY: Microarray-based transcriptional and epigeneticprofiling of matrix metalloproteinases, collagens, and related genes incancer. J Biol Chem 2010, 285:19647–19659.
23. Wilkerson MD, Yin X, Hoadley KA, Liu Y, Hayward MC, Cabanski CR, Muldrew K,Miller CR, Randell SH, Socinski MA, Parsons AM, Funkhouser WK, Lee CB,Roberts PJ, Thorne L, Bernard PS, Perou CM, Hayes DN: Lung squamous cellcarcinoma mRNA expression subtypes are reproducible, clinicallyimportant and correspond to different normal cell types. Clin Cancer Res2010, 16:4864–4875. doi:10.1158/1078-0432.CCR-10-0199.
24. Hajdu M, Singer S, Maki RG, Schwartz GK, Keohan ML, Antonescu CR: IGF2over-expression in solitary fibrous tumours is independent of anatomicallocation and is related to loss of imprinting. J Pathol 2010, 221:300–307.
25. Planche A, Bacac M, Provero P, Fusco C, Delorenzi M, Stehle JC,Stamenkovic I: Identification of prognostic molecular features in thereactive stroma of human breast and prostate cancer. PLoS One 2011,6:e18640. doi:10.1371/journal.pone.0018640.
26. Anastassiou D, Rumjantseva V, Cheng W, Huang J, Canoll PD, Yamashiro DJ,Kandel JJ: Human cancer cells express slug-based epithelial-mesenchymal transition gene expression signature obtained in vivo.BMC Cancer 2011, 11:529 [http://www.biomedcentral.com/1471-2407/11/529]
27. Navab R, Strumpf D, Bandarchi B, Zhu CQ, Pintilie M, Ramnarine VR,Ibrahimov E, Radulovich N, Leung L, Barczyk M, Panchal D, To C, Yun JJ, DerS, Shepherd FA, Jurisica I, Tsao MS: Prognostic gene-expression signatureof carcinoma-associated fibroblasts in non-small cell lung cancer. ProcNatl Acad Sci U S A 2011, 108:7160–7165. doi:10.1073/pnas.1014506108.
28. Lascorz J, Hemminki K, Försti A: Systematic enrichment analysis of geneexpression profiling studies identifies consensus pathways implicated incolorectal cancer development. J Carcinog 2011, 10:7. doi:10.4103/1477-3163.78268.
29. Seemann L, Shulman J, Gunaratne GH: A robust topology-based algorithmfor gene expression profiling. ISRN Bioinformatics 2012, Article ID 381023.doi:10.5402/2012/381023.
30. Vargas AC, McCart Reed AE, Waddell N, Lane A, Reid LE, Smart CE, Cocciardi S,da Silva L, Song S, Chenevix-Trench G, Simpson PT, Lakhani SR: Geneexpression profiling of tumour epithelial and stromal compartmentsduring breast cancer progression. Breast Cancer Res Treat 2012,135:153–165. doi:10.1007/s10549-012-2123-4.
31. Wu YH, Chang TH, Huang YF, Huang HD, Chou CY: COL11A1 promotestumor progression and predicts poor clinical outcome in ovarian cancer.Oncogene 2014, 33:3432–3440. doi:10.1038/onc.2013.307.
32. Gene expression atlas- summary for COL11A1 (Homo sapiens).[http://www.ebi.ac.uk/gxa/gene/ENSG00000060718]
33. Boshoff C, Bryson K, Clements MO, Trotter MW, Cellek S, Elliman SJ:Transcription profiling of two populations of non-hematopoieticstem cells (MSC and MAPC) isolated from human bone marrow.ArrayExpress: E-MEXP-466. [http://www.ebi.ac.uk/arrayexpress/experiments/E-MEXP-466/]
34. Grundberg E, Brändström H, Lam KC, Gurd S, Ge B, Harmsen E, Kindmark A,Ljunggren O, Mallmin H, Nilsson O, Pastinen T: Systematic assessment ofthe human osteoblast transcriptome in resting and induced primarycells. Physiol Genomics 2008, 33:301–311.
35. Kao L-P, Yu S-L, Singh S, Wang K-H, Kao A-P, Li SS: Comparative profiling ofmRNA and microRNA expression in human mesenchymal stem cellsderived from adult adipose and lipoma tissues. Open Stem Cell J 2009,1:1–9. doi:10.2174/1876893800901010001.
36. Polanska UM, Orimo A: Carcinoma-associated fibroblasts: non-neoplastictumour-promoting mesenchymal cells. J Cell Physiol 2013, 228:1651–1657.doi:10.1002/jcp.24347.
37. García-Ocaña M, Vázquez F, García-Pravia C, Fuentes-Martínez N, Menéndez-Rodríguez P, Fresno-Forcelledo F, Barneo-Serra L, Del Amo-Iribarren J,Simón-Buela L, De Los Toyos JR: Characterization of a novel mouse monoclonalantibody, clone 1E8.33, highly specific for human procollagen 11A1, atumor-associated stromal component. Int J Oncol 2012, 40:1447–1454.doi:10.3892/ijo.2012.1360.
38. Hering TM, Kollar J, Huynh TD, Varelas JB, Sandell LJ: Modulation ofextracellular matrix gene expression in bovine high-density chondrocytecultures by ascorbic acid and enzymatic resuspension. Arch BiochemBiophys 1994, 314:90–98.
39. Ronzière MC, Farjanel J, Freyria AM, Hartmann DJ, Herbage D: Analysis oftypes I, II, III, IX and XI collagens synthesized by fetal bovinechondrocytes in high-density culture. Osteoarthritis Cartilage 1997,5:205–214.
40. Elliott RL, Blobe GC: Role of transforming growth factor Beta in humancancer. J Clin Oncol 2005, 23:2078–2093.
41. García-Pravia C, Galván JA, Gutiérrez-Corral N, Solar-García L, García-Pérez E,García-Ocaña M, Del Amo-Iribarren J, Menéndez-Rodríguez P, García-GarcíaJ, de los Toyos JR, Simón-Buela L, Barneo L: Overexpression of COL11A1by cancer-associated fibroblasts: clinical relevance of a stromal marker inpancreatic cancer. PLoS One 2013, 8:e78327. doi:10.1371/journal.pone.0078327.
42. Cueva-Cayetano R, Galvan-Hernandez JÁ, Suarez-Fernandez L, Menendez-Rodriguez MP, Garcia-Pravia C, Barneo L: Preliminary analysis of collagen,type XI, alpha 1 (COL11A1), inhibin alpha (INHBA) and secreted proteinacidic and rich in cysteine (SPARC, osteonectin) as potential markers ofcolon cancer [abstract]. Brit J Surg 2013, 100(Suppl. 1):7.
43. Mifflin RC, Pinchuk IV, Saada JI, Powell DW: Intestinal myofibroblasts:targets for stem cell therapy. Am J Physiol Gastrointest Liver Physiol 2011,300:G684–G696. doi:10.1152/ajpgi.00474.2010.
44. Emura M, Ochiai A, Horino M, Arndt W, Kamino K, Hirohashi S:Development of myofibroblasts from human bone marrowmesenchymal stem cells cocultured with human colon carcinoma cellsand TGF beta 1. In Vitro Cell Dev Biol Anim 2000, 36:77–80.
45. Karnoub AE, Dash AB, Vo AP, Sullivan A, Brooks MW, Bell GW, Richardson AL,Polyak K, Tubo R, Weinberg RA: Mesenchymal stem cells within tumourstroma promote breast cancer metastasis. Nature 2007, 449:557–563.
46. ArrayExpress Experiment E-GEOD-9764: Transcription profiling ofhuman mesenchymal stem cells reveals carcinoma associatedfibroblast like differentiation. [http://www.ebi.ac.uk/arrayexpress/experiments/E-GEOD-9764]
47. Mishra PJ, Mishra PJ, Humeniuk R, Medina DJ, Alexe G, Mesirov JP, Ganesan S,Glod JW, Banerjee D: Carcinoma-associated fibroblast-like differentiation ofhuman mesenchymal stem cells. Cancer Res 2008, 68:4331–4339. doi:10.1158/0008-5472.CAN-08-0943.
48. Cheon DJ, Tong Y, Sim MS, Dering J, Berel D, Cui X, Lester J, Beach JA,Tighiouart M, Walts AE, Karlan BY, Orsulic S: A collagen-remodeling genesignature regulated by TGF-β signaling is associated with metastasis andpoor survival in serous ovarian cancer. Clin Cancer Res 2014, 20:711–723.doi:10.1158/1078-0432.CCR-13-1256.
49. ArrayExpress Experiment E-GEOD-6731: Transcription profiling of human colonbiopsies obtained from patients with ulcerative colitis, Crohn’s disease vs.normal to identify pathogenic processes underlying these disease subtypes.[http://www.ebi.ac.uk/arrayexpress/experiments/E-GEOD-6731]
50. Wu F, Dassopoulos T, Cope L, Maitra A, Brant SR, Harris ML, Bayless TM,Parmigiani G, Chakravarti S: Genome-wide gene expression differences inCrohn’s disease and ulcerative colitis from endoscopic pinch biopsies:insights into distinctive pathogenesis. Inflamm Bowel Dis 2007,13:807–821.
51. ArrayExpress Experiment E-TABM-118: Transcription profiling of biopsiesfrom the descending colon of Crohn’s disease patients. [http://www.ebi.ac.uk/arrayexpress/experiments/E-TABM-118/]
52. Csillag C, Nielsen OH, Borup R, Nielsen FC, Olsen J: Clinical phenotype andgene expression profile in Crohn’s disease. Am J Physiol Gastrointest LiverPhysiol 2007, 292:G298–G304.
53. ArrayExpress Experiment E-GEOD-1152: Transcription profiling of humanileum and colonic tissues from patients with Crohns disease or ulcerativecolitis. [http://www.ebi.ac.uk/arrayexpress/experiments/E-GEOD-1152].

Galván et al. BMC Cancer 2014, 14:867 Page 12 of 12http://www.biomedcentral.com/1471-2407/14/867
54. Fuentes-Martínez N, García-Pravia C, García-Ocaña M, Menéndez-RodríguezP, Del Amo J, Suárez-Fernández L, Galván JA, De Los Toyos JR, Barneo L:Overexpression of proCOL11A1 as a stromal marker of breast cancer.Histol Histopathol 2014, [Epub ahead of print].
55. Schuetz CS, Bonin M, Clare SE, Nieselt K, Sotlar K, Walter M, Fehm T,Solomayer E, Riess O, Wallwiener D, Kurek R, Neubauer HJ: Progression-specific genes identified by expression profiling of matched ductalcarcinomas in situ and invasive breast tumors, combining laser capturemicrodissection and oligonucleotide microarray analysis. Cancer Res 2006,66:5278–5286.
56. Lee S, Stewart S, Nagtegaal I, Luo J, Wu Y, Colditz G, Medina D, Allred DC:Differentially expressed genes regulating the progression of ductalcarcinoma in situ to invasive breast cancer. Cancer Res 2012,72:4574–4586. doi:10.1158/0008-5472.CAN-12-0636.
57. Castellana B, Escuin D, Peiró G, Garcia-Valdecasas B, Vázquez T, Pons C,Pérez-Olabarria M, Barnadas A, Lerma E: ASPN and GJB2 are implicated inthe mechanisms of invasion of ductal breast carcinomas. J Cancer Educ2012, 3:175–183. doi:10.7150/jca.4120.
doi:10.1186/1471-2407-14-867Cite this article as: Galván et al.: Validation of COL11A1/procollagen11A1 expression in TGF-β1-activated immortalised humanmesenchymal cells and in stromal cells of human colonadenocarcinoma. BMC Cancer 2014 14:867.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit