
Expressão e purificação em sistema procariota de uma...
26
1 Expressão e purificação em sistema procariota de uma proteína recombinante homóloga ao interferon beta humano para uso terapêutico Fabricio Fernandes de Souza 1,2,3 , Patrícia Cota Campos 2 , Sophie Yvette Leclercq 2 , Paula Henriques Cruz Ciscotto 1 , Roberta Márcia Marques dos Santos 3 . Resumo Atualmente o INF-β é produzido tanto em sistema eucariota quanto em procariota e uma de suas principais aplicações é como agente terapêutico no tratamento da Esclerose Múltipla. No Brasil, o INF-β ainda não é produzido como proteína recombinante, o que implica num custo muito elevado na sua importação, sendo este um dos motivos que torna de grande importância à realização desse projeto. O presente projeto tem por objetivo expressar e purificar em Escherichia coli uma proteína recombinante homóloga ao INF-β humano, utilizando códons preferenciais para essa bactéria. Para utilização de códons preferenciais, 10 oligonucleotídeos foram desenhados por meio do programa PCR Oligo Maker. O gene sintético foi construído através de duas PCRs. Após a síntese, o gene foi clonado utilizando TA Cloning Kit (vetor de fácil clonagem) seguida da subclonagem em vetor de expressão (pET-21a) e sequenciamento automático de alguns clones selecionados. Foi escolhido um clone com a sequência correta e feita a indução. Após a indução foram realizados testes de solubilidade para futura purificação. As melhores condições para construção do gene INF-β sintético foram, PCR de montagem 1pmol (oligonucleotídeos) e PCR de amplificação 65 ºC (anelamento). A melhor condição para expressão foi obtida com DO 600 igual a 1, concentração de 0,5 mM IPTG incubado à 37 ºC por 4h. A expressão do INF-β foi confirmada por meio do Western Blot com anticorpo anti-INF-β. A proteína expressa mostrou-se insolúvel. Alguns testes de solubilização foram realizados e parte da proteína insolúvel foi recuperada pelo tratamento com uréia 8M. Apesar de não concluída a purificação, realizou-se com êxito a construção, clonagem e expressão do gene INF-β sintético. Palavras chave: Interferon Beta Recombinante. Gene sintético. Códons Preferenciais. 1 Centro Universitário UNA – Belo Horizonte – MG. E-mail: [email protected] 2 Serviço de Biologia Celular e Inovação Biotecnológica – Divisão de Ciências Biomédicas - Diretoria de Pesquisa e Desenvolvimento, FUNED – Belo Horizonte –MG. 3 Serviço de Desenvolvimento Biotecnológico – Divisão de Desenvolvimento Farmacotécnico e Biotecnológico - Diretoria Industrial, FUNED – Belo Horizonte –MG.
Transcript of Expressão e purificação em sistema procariota de uma...

1
Expressão e purificação em sistema procariota de um a proteína recombinante
homóloga ao interferon beta humano para uso terapêu tico
Fabricio Fernandes de Souza1,2,3, Patrícia Cota Campos2, Sophie Yvette Leclercq2, Paula
Henriques Cruz Ciscotto1, Roberta Márcia Marques dos Santos3.
Resumo
Atualmente o INF-β é produzido tanto em sistema eucariota quanto em procariota e uma de
suas principais aplicações é como agente terapêutico no tratamento da Esclerose Múltipla.
No Brasil, o INF-β ainda não é produzido como proteína recombinante, o que implica num
custo muito elevado na sua importação, sendo este um dos motivos que torna de grande
importância à realização desse projeto. O presente projeto tem por objetivo expressar e
purificar em Escherichia coli uma proteína recombinante homóloga ao INF-β humano,
utilizando códons preferenciais para essa bactéria. Para utilização de códons preferenciais,
10 oligonucleotídeos foram desenhados por meio do programa PCR Oligo Maker. O gene
sintético foi construído através de duas PCRs. Após a síntese, o gene foi clonado utilizando
TA Cloning Kit (vetor de fácil clonagem) seguida da subclonagem em vetor de expressão
(pET-21a) e sequenciamento automático de alguns clones selecionados. Foi escolhido um
clone com a sequência correta e feita a indução. Após a indução foram realizados testes de
solubilidade para futura purificação. As melhores condições para construção do gene INF-β
sintético foram, PCR de montagem 1pmol (oligonucleotídeos) e PCR de amplificação 65 ºC
(anelamento). A melhor condição para expressão foi obtida com DO600 igual a 1,
concentração de 0,5 mM IPTG incubado à 37 ºC por 4h. A expressão do INF-β foi
confirmada por meio do Western Blot com anticorpo anti-INF-β. A proteína expressa
mostrou-se insolúvel. Alguns testes de solubilização foram realizados e parte da proteína
insolúvel foi recuperada pelo tratamento com uréia 8M. Apesar de não concluída a
purificação, realizou-se com êxito a construção, clonagem e expressão do gene INF-β
sintético.
Palavras chave: Interferon Beta Recombinante. Gene sintético. Códons Preferenciais.
1 Centro Universitário UNA – Belo Horizonte – MG. E-mail: [email protected]
2 Serviço de Biologia Celular e Inovação Biotecnológica – Divisão de Ciências Biomédicas - Diretoria de Pesquisa e Desenvolvimento, FUNED – Belo Horizonte –MG. 3 Serviço de Desenvolvimento Biotecnológico – Divisão de Desenvolvimento Farmacotécnico e Biotecnológico - Diretoria Industrial, FUNED – Belo Horizonte –MG.

2
Expression and purification in prokaryotic system o f a recombinant protein
homologous to human beta interferon for therapeutic use
Fabricio Fernandes de Souza1,2,3, Patrícia Cota Campos2, Sophie Yvette Leclercq2, Paula
Henriques Cruz Ciscotto1, Roberta Márcia Marques dos Santos3.
Abstract
Currently, IFN-β is produced both in prokaryotic and eukaryotic system, and one of its main
applications is as therapeutic agent in the treatment of Multiple Sclerosis. In Brazil, the IFN-β
is not produced as a recombinant protein, which implies a very high cost in their import, this
being one of the reasons of great importance to implementing this project. This project aims
express and purify in Escherichia coli a recombinant protein homologous to human IFN-β,
based on codon usage for this bacterium. For the codon usage, ten oligonucleotides were
designed by means of the program PCR Oligo Maker. The synthetic gene was constructed
by two PCRs, the assembly PCR, followed by amplification PCR. After the synthesis, the
gene was cloned using TA Cloning Kit (easy cloning vector) followed by subcloning in
expression vector (pET-21a) and automated sequencing of some selected clones. A clone
with the correct sequence was chosen and used to induction of expression. After induction
solubility tests were performed for further purification. The best conditions for construction of
synthetic gene IFN-β were, assembly PCR 1 pmol (oligonucleotides) and amplification PCR
65 ºC (annealing). The best expression condition was obtained with OD600 equal to 1,
concentration of 0,5 mM IPTG incubated for 37 ºC for 4h. The expression of the IFN-β was
confirmed by Western Blot with antibody anti-IFN-β. The protein was present on the insoluble
fraction. Some solubilization tests were performed and a part of the insoluble protein was
recovered by treatment with urea 8M. Although not complete the purification, was held
successfully the construction, cloning and expression of synthetic gene IFN-β.
Key words: Recombinant Beta Interferon. Synthetic Gene. Codon Usage.
1 Centro Universitário UNA – Belo Horizonte – MG. E-mail: [email protected]
2 Serviço de Biologia Celular e Inovação Biotecnológica – Divisão de Ciências Biomédicas - Diretoria de Pesquisa e Desenvolvimento, FUNED – Belo Horizonte –MG. 3 Serviço de Desenvolvimento Biotecnológico – Divisão de Desenvolvimento Farmacotécnico e Biotecnológico - Diretoria Industrial, FUNED – Belo Horizonte –MG.

3
1 INTRODUÇÃO
1.1 Interferons
Os interferons foram descritos na década de 50, pela primeira vez, por Isaacs e
Lindenmann, como fatores solúveis liberados no meio de cultura de células infectadas por
vírus que, ao interagirem com outra célula, eram capazes de induzir resistência à infecção
viral (Galvani et al., 1999).
Sabe-se que os interferons constituem uma família de diferentes proteínas classificadas em
Tipo I e Tipo II. Os interferons do Tipo I abrangem dois grupos de proteínas distintas, o INF-
α e o INF-β, enquanto o INF-γ é o único representante do Tipo II. Os INF-α e INF-β são
produzidos principalmente por fagócitos mononucleares e fibroblastos, respectivamente.
Cada um deles é codificado por um gene presente no cromossomo nove. Eles se ligam ao
mesmo receptor da superfície celular e induzem respostas biológicas semelhantes. Já o
INF-γ é uma glicoproteína, codificada por um gene no cromossomo 12, a qual é produzida
por linfócitos T auxiliares (Abbas et al.,2003; Roitt et al., 1998).
Os INF-α e INF-β exibem uma ampla variedade de atividades biológicas: ação antiviral;
antiproliferativa e citotóxica para algumas células do sistema imunológico (ex. células T, NK
e dendríticas); aumenta a expressão de antígenos de superfícies associados a tumores;
indução e/ou ativação de genes e proteínas pró-apopitóticas (ex. caspases, Bak e Bax);
repressão de genes anti-apopitóticos (ex. Bcl-2); modulação da diferenciação; atividade anti-
angiogênica. Todas essas ações fazem dos interferons um dos agentes mais promissores
para o tratamento de várias doenças (Abbas et al.,2003; Pestka, 2007).
O INF-β é uma glicoproteína constituída por 166 aminoácidos, codificado por um único gene
e não possui íntrons. A análise da sua estrutura tridimensional mostrou tratar-se de um
homodímero sendo cada monômero constituído de cinco alfa-hélices, mantidos unidos por
íons Zinco (Derynck et al., 1980; Kagawa et al., 1988; Karpusas et al., 1997; Ohno e
Taniguchi, 1981). Ainda na década de 80, o gene do INF-β foi clonado e a proteína
recombinante expressa em E. coli. O extrato bacteriano obtido foi capaz inibir a atividade
viral em ensaio in vitro (Taniguchi et al., 1980).
O seqüenciamento de aminoácidos do interferon β revelou três resíduos cisteinil nas
posições 17, 31 e 141. Após mutação gênica sítio irigida, Cys17/ Ser17, a proteína
modificada resultante, expressa em E. coli, apresentou atividade antiviral semelhante à

4
proteína nativa de fibroblasto, demonstrando que o resíduo modificado (Cys17) não era
essencial para a atividade protéica (Mark et al., 1984).
Os interferons são aprovados pelo Food and Drug Administration (FDA, EUA) para uso em
vários tratamentos em humanos. Até o momento, o INF-α é indicado para o tratamento da
leucemia de células pilosas, melanoma maligno, linfoma folicular, condilomas, sarcoma de
Kaposi relacionado com a AIDS, hepatites crônicas B e C; o INF-β para Esclerose Múltipla
(EM); e o INF-γ para a doença granulomatosa crônica e osteoporose maligna (Platanias,
2005; Pestka, 2007).
1.2 Esclerose Múltipla
A Esclerose Múltipla (EM) é uma doença auto-imune, que leva a destruição da bainha de
mielina do Sistema Nervoso Central, caracterizada por episódios repetidos de disfunção
neurológica com remissão variável (Goldman & Ausiello, 2005; Sottini A et al., 2009). A
doença pode apresentar quatro formas de evolução: remitente recorrente (EMRR), também
conhecida como surto remissão; secundariamente progressiva (EMSP); primariamente
progressiva (EMPP) e progressiva com surtos (EMPS) (Lana-Peixoto et al., 2002; Moreira et
al., 2000; Tilbery et al., 2000).
A EM acomete, predominantemente adultos jovens, e traz consigo, em adição ao sofrimento
individual e familiar, grande sobrecarga econômica à sociedade, devido à perda de força de
trabalho. Embora não apresente uma taxa de mortalidade elevada, a progressão dos déficits
neurológicos ocorre em todos os portadores da doença. Após 15 anos do seu início, cerca
de 50% dos pacientes necessitam de auxílio para caminhar e, após 25 anos, a maioria está
incapacitada para andar (Lana-Peixoto et al., 2002; Tilbery et al., 2000).
Acredita-se que a patologia se deva a uma predisposição genética que, associada a um
fator ambiental, origina uma disfunção do sistema imunológico. Essa disfunção desenvolve
uma ação auto-lesiva dirigida contra a substância branca, com perda de oligodendrócitos e
mielina. O quadro acaba levando a problemas na condução dos impulsos nervosos e
conseqüente aparecimento dos sintomas mais comuns, que são: sensitivos, disfunção do
esfíncter, neurites ópticas, ataxia e diplopia. Ainda não está totalmente esclarecido como
ocorre o ataque imunológico contra a bainha de mielina causando o dano neuronal. (Hartung
et al., 1995; Moreira et al., 2000; Siffrin et al., 2010; Tilbery et al., 2006).
Em 2005, o Departamento Científico de Neuroimunologia da Academia Brasileira de
Neurologia publicou as Diretrizes para o Tratamento da Esclerose Múltipla que trazem

5
recomendações acerca do diagnóstico, classificação e uso de imunomoduladores para
tratamento da doença. Segundo essas diretrizes, os imunomoduladores devem ser
prescritos aos pacientes que apresentem sintomas ou sinais clínicos da doença, nas formas
EMRR, EMPP e EMSP.
No Brasil há relatos de prevalência de EM nos municípios de São Paulo e Belo Horizonte,
com taxas de 15/100.000 habitantes e 18/100.000 habitantes, respectivamente. A nível
mundial, a EM afeta cerca de 2,5 milhões de pessoas (Lana-Peixoto et al., 2002; National
MS Society, 2010). Em estudo de 302 casos no país com pacientes com média de idade
igual a 37,7 anos, Moreira et al. (2000) relataram que a média de idade no início da doença
foi de 29,6 anos, sendo mais comum nas mulheres (uma proporção de 3,13 mulheres para
cada homem).
1.3 Interferon beta e a Esclerose Múltipla
Em 1993, foi publicado um estudo com INF-β 1b, em 372 pacientes com EMRR, o qual
demonstrou que o tratamento foi bem tolerado, houve significativa redução no número e
gravidade dos surtos, prolongamento do tempo de remissão entre eles e redução no número
e volume das lesões observadas nos exames de ressonância magnética nuclear e ausência
de efeitos colaterais mais sérios. Outro ponto importante foi à diminuição do número de
hospitalizações em 43% quando comparados, o grupo em tratamento com INFβ-1b e o
grupo placebo (Paty et al., 1993). O INF-β foi o primeiro tratamento efetivo para EMRR e
também o primeiro a demonstrar efeito terapêutico em pacientes com EMSP (Paty et al.,
1993; European Study Group,1998).
Os imunomoduladores atualmente empregados no tratamento da EM são: INF-β 1b
(Betaferon – Schering), INF-β 1a (Rebif – Aeres-Serono), (Avonex - Abbott). O INF-β 1a
é uma proteína recombinante glicosilada produzida em células CHO (ovário de hamster
chinês), com seqüência de aminoácidos idêntica à do INF-β humano. O INF-β 1b, também é
recombinante produzido em E. coli, apresentando na sua estrutura de aminoácidos uma
substituição da cisteína 17 por uma serina (Goldman & Ausiello, 2005).
Apesar dos esforços de centenas de laboratórios em todo o mundo, ainda não é
precisamente conhecido o mecanismo pelo qual os interferons promovem seus efeitos
(Platanias, 2005). Em um estudo com INF-β 1b recombinante in vitro, demonstrou-se sua
capacidade de induzir e aumentar a sensibilidade das células de monócitos humanos a
glicocorticóides, provavelmente devido ao aumento do número de receptores celulares para
os mesmos (Uitdehaag et al., 2001).

6
Lana-Peixoto et al. (2002) resumidamente atribuem os efeitos do INF β na EM à (1) sua
ação na diminuição da produção de INF-γ, aumento da produção de interleucina 10, e
diminuição da relação TH1/TH2; (2) diminuição da permeabilidade da barreira
hematoencefálica, por ação sobre as moléculas de adesão e metaloproteases e (3) efeito
sobre as células gliais.
1.4 Expressão de proteínas recombinantes
Dentre os avanços da engenharia genética e da biotecnologia nas últimas décadas, tornou-
se possível a manipulação do material genético permitindo a obtenção de proteínas fora do
seu organismo de origem, chamadas proteínas recombinantes. Essa possibilidade atraiu o
interesse de indústrias farmacêuticas, que ainda na década de 80, produziram a insulina em
bactérias - que antes era extraída de pâncreas suíno e bovino, requerendo grandes
quantidades de matéria-prima, tornando o processo oneroso - se tornando o primeiro
medicamento de sucesso da chamada biotecnologia moderna. Nesse contexto, o emprego
de técnicas mais eficientes, como a do DNA recombinante, abriu novas perspectivas de
produção (Lima, 2001).
Desde então, novas ferramentas vêem sendo desenvolvidas na tentativa de otimização das
técnicas de biologia molecular para tornar os processos de obtenção de proteínas
recombinantes mais eficientes. Uma alternativa para melhoria da produção de proteínas
recombinantes em E. coli, é o uso de códons preferenciais. O código genético é
degenerado, permitindo que um mesmo aminoácido seja codificado por códons distintos. Os
códons de um mesmo aminoácido não ocorrem com a mesma freqüência, em espécies
diferentes, sugerindo que existe utilização de códons preferenciais. Diferentes organismos
geralmente apresentam preferência por códons distintos e estas diferenças podem ser
fatores limitantes na expressão heteróloga. Assim, se o mRNA do gene clonado contiver
códons raros, para o organismo utilizado, é provável que haja problemas na tradução,
levando a uma redução na quantidade da proteína expressa (Jana & Deb, 2005).
A clonagem a partir do cDNA, não permite alterações na seqüência do gene de interesse,
impossibilitando a utilização do uso de códons preferenciais para o organismo que se deseja
realizar uma expressão heteróloga. Uma alternativa é a construção de um gene sintético
através da técnica de Reação em Cadeia da Polimerase (PCR), que utiliza oligonucleotídeos
para a síntese do gene ao invés de um DNA molde. De acordo com Singh et al. (1996),
quando se utiliza a técnica de PCR para construção de um gene, durante a fase de
anelamento, os oligonucleotídeos se pareiam em suas extremidades, gerando

7
sobreposições que terão a mesma função dos iniciadores, permitindo a ação da DNA
polimerase, sintetizando o gene de interesse (Fig. 1).
Figura 1. Esquema da construção do gene sintético através da técnica de PCR
Conforme exposto acima, o INF-β é uma proteína de grande interesse farmacêutico, e a
mesma pode ser obtida através da utilização de diferentes técnicas disponíveis de biologia
molecular. A Fundação Ezequiel Dias (Funed), vem investindo nos últimos anos, na
ampliação da sua capacidade para a produção de produtos biológicos, e está em fase de
implementação uma nova fábrica de soros, vacinas e biofármacos, existindo o interesse em
desenvolver novos produtos que possam ser incorporados ao seu portfólio. As técnicas de
clonagem utilizando gene sintético, mutação sitio dirigida e códons preferenciais, ainda não
haviam sido utilizadas pela equipe de Pesquisa e Desenvolvimento, se tornando uma
oportunidade para a ampliação do conhecimento e implementação de ferramentas de
biologia molecular que podem ser utilizadas no desenvolvimento de diferentes proteínas
recombinantes, além é claro, da formação de profissionais. Sendo assim, o presente estudo
teve por objetivo construir o gene sintético do INF-β através da técnica de PCR utilizando o
uso de códons preferenciais para E. coli, clonar, expressar e purificar a proteína
recombinante correspondente.

8
2 MATERIAIS E MÉTODOS
2.1 Desenho dos oligonucleotídeos e primers
A seqüência do gene INF-β humano foi obtida através do banco de dados de genes do NCBI
(número de entrada NC_000009). A partir da seqüência original foi retirada a seqüência
sinal, inserida uma mutação sítio dirigida, substituindo o aminoácido Cisteina na posição 17
por Serina. Através do programa JCat (Java Codon Adaptation Tool) foi adaptado o uso de
códons preferenciais para expressão em E. coli. Após essas alterações, 10
oligonucleotideos para a construção do gene sintético - com 501 pb - foram desenhados no
programa Assembly PCR Oligo Maker e outros 2, Sense e Anti-sense, contendo sítios de
restrição para as endonucleases Nde I e BamH I respectivamente, foram desenhados para a
amplificação e posterior clonagem (Fig. 2). As sequências dos dez oligonucleotídeos para
construção do gene foram sintetizadas pela empresa (Integrated DNA Technologies) e os
outros dois pela (Bioneer Synthesis Report).
Figura 2. Oligonucleotídeos para a construção e amplificação do gene do INF-β sintético
OLIGO SEQUÊNCIA
1 ATGACCAACAAGTGTCTCCTCCAAATTGCTCTCCTGTTGTGCTTCTCCACTACAGCTC
2 AAATTGCTGCTTCTTTGTAGGAATCCAAGCAAGTTGTAGCTCATGGAAAGAGCTGTAGTGGAGAAGCACA
3 GGATTCCTACAAAGAAGCAGCAATTTTCAGTGTCAGAAGCTCCTGTGGCAATTGAATGGGAGGCTTGAATAC
4 GCTTAATCTCCTCAGGGATGTCAAAGTTCATCCTGTCCTTGAGGCAGTATTCAAGCCTCCCATTCAATTG
5 GACATCCCTGAGGAGATTAAGCAGCTGCAGCAGTTCCAGAAGGAGGACGCCGCATTGACCATCTATGAG
6 CAGTGCTAGATGAATCTTGTCTGAAAATAGCAAAGATGTTCTGGAGCATCTCATAGATGGTCAATGCGGC
7 TTCAGACAAGATTCATCTAGCACTGGCTGGAATGAGACTATTGTTGAGAACCTCCTGGCTAATGTCTATCA
8 CTTTCTCCAGTTTTTCTTCCAGGACTGTCTTCAGATGGTTTATCTGATGATAGACATTAGCCAGGAGGTTC
9 CCTGGAAGAAAAACTGGAGAAAGAAGATTTCACCAGGGGAAAACTCATGAGCAGTCTGCACCTGAAAAG
10 ACTGTACTCCTTGGCCTTCAGGTAATGCAGAATCCTCCCATAATATCTTTTCAGGTGCAGACTGCTC
Primer*
Sense 5’ ACA CA|T ATG TCT TAC AAC CTG CTG GGT TTC CTG 3’
Primer *
Anti-sense 5’ ACA G|GA TCC GTT ACG CAG GTA ACC GGTCAG AC 3’
Nota: . *Em vermelho estão os sítios de restrição para as enzimas Nde I (primer sense) e BamH I (primer anti-
sense).

9
2.2 PCR para construção do gene INF- β sintético
O gene sintético INF-β foi construído por meio da Reação em Cadeia da Polimerase (PCR)
utilizando os 10 oligonucleotídeos desenhados ao invés de um DNA molde. Para um volume
final de 50 µl por reação continha, 1u de Deep Vent polymerase (New England Biolabs),
tampão 10 X, 200 µM do mix de dNTPs, 1,5 mM de MgCl2 e concentrações variando de 0,25
a 4 picomoles de cada oligonucleotídeo. As temperaturas e tempos durante 35 ciclos foram:
desnaturação 94 ºC, 30s; anelamento variando entre 55 e 70 ºC, 30s; e extensão 72 ºC,
60s.
A construção do gene INF-β sintético foi analisado por eletroforese em gel de agarose 1%.
2.3 PCR para amplificação do gene INF- β sintético
A amplificação do gene INF-β sintético foi realizada por meio de uma segunda PCR
utilizando o produto da primeira PCR, a PCR de construção, e os primers Sense e Anti-
sense contento os sítios para as enzimas de restrição Nde I e BamH I, respectivamente.
Para um volume final de 50 µl por reação utilizou-se, 1u de Deep Vent polymerase (New
England Biolabs), 5 µl de tampão 10 X, 200 µM do mix de dNTPs, 1,5 mM de MgCl2 e 5
picomoles de cada primer (sense e anti-sense). As temperaturas e tempos durante 35 ciclos
foram: desnaturação 94 ºC, 30s; anelamento variando entre 55 e 65 ºC, 30s; e extensão 72
ºC, 60s.
A amplificação do gene INF-β sintético foi analisada por eletroforese em gel de agarose 1%.
A partir do produto da amplificação foi feito a purificação do DNA por meio da precipitação
com isopropanol 70%.
2.4 Clonagem do gene INF- β sintético
2.4.1 Clonagem utilizando o sistema TA Cloning® Kit
Após a purificação com isopropanol 70%, o gene sintético INF-β foi clonado através do
sistema TA Cloning® Kit, que consiste em um vetor (pCR 2.1) linearizado com pontas
coesivas de base Timina (T) e possui um marcador seletivo LacZ , uma região que
corresponde ao gene da beta-galactosidase (Fig. 3).

10
Figura 3. (A) Vetor pCR 2.1; e (B) Extremidades coesivas da região LacZ do vetor
A B
Fonte: Invitrogen life technologies
A ligação entre o vetor e o inserto foi feita numa proporção aproximada de 1:4, utilizando-se
100ng do vetor para 50ng do inserto. Essa proporção foi calculada a partir da fórmula
abaixo, presente no manual do sistema TA Cloning® Kit:
X (ng) = Y (pb inserto) x Z (ng vetor) / Tamanho do vetor (pb) 1:1
A reação de ligação foi realizada de acordo com recomendações do fabricante (Invitrogen
life Technologies, 2004).
2.4.2 Preparo de células competentes da linhagem DH5α
Uma pré-cultura foi preparada contendo 10µl de E. coli da linhagem DH5α em 5ml de meio
de cultura LB, e incubada a 37 ºC por 16 horas com agitação. Uma alíquota de 200µ dessa
pré-cultura foi inoculada em 40ml de meio LB, incubada com agitação a 37 ºC por
aproximadamente 2 horas, até atingir uma densidade óptica (DO) entre 0,4 e 0,7, com
absorbância de 600nm (A600). O meio de cultura contendo as bactérias foi centrifugado a
3000rpm por 10 minutos a 4 ºC. Logo depois o sobrenadante foi descartado, o pellet
ressuspendido em 20ml de CaCl2 (50mM) gelado e incubado por 20 minutos em banho de
gelo. As bactérias foram centrifugadas como descrito anteriormente e ressuspendidas em
2ml de CaCl2 (50mM) gelado. As bactérias foram mantidas em banho de gelo por 1 hora até
serem utilizadas na transformação.

11
2.4.3 Transformação Bacteriana
A transformação bacteriana foi realizada com a adição de 5µl do produto de ligação
vetor/inserto em 100µl de células competentes e mantida em banho de gelo por 30 minutos.
O microtubo contendo as células competentes e o produto da ligação, foi colocado em
banho maria à 42 ºC por 90 segundos e logo após em banho de gelo por 2 minutos. Depois
foi acrescentado meio SOC completo e incubado com agitação a 37 ºC por 1 hora. Em
seguida 200µl da transformação foi plaqueada em meio LB ágar contendo ampicilina (AMP)
100 µg/ml e Xgal 40 µg/ml e encubado em estufa a 37 ºC por 16 horas. Foi feito um controle
negativo contendo somente células competentes.
2.4.4 PCR de colônias
As colônias brancas, indicativas de positivas para a transformação, foram submetidas a uma
PCR para a confirmação da incorporação do inserto ao vetor. Para a realização da PCR de
colônias primeiro as bactérias foram lisadas com um tampão de lise (20mM Tris-HCl pH 8,5,
2mM EDTA, 1% de Triton X-100), aquecidas por 5 minutos entre 80 e 90 ºC em banho de
óleo, centrifugadas por 5 minutos a 6000rpm. Cinco microlitros do sobrenadante foram
utilizados para fazer a PCR, utilizando o Kit Master Mix Promega. Para um volume final de
25 µl por reação utilizou-se, 12,5 µl do Máster Mix 2X, 5 pmoles de cada primer (sense e
anti-sense) e 5 µl do produto da lise bacteriana. A programação utilizada para PCR foi a
mesma descrita anteriormente para amplificação do gene INF-β sintético.
2.4.5 Sequenciamento dos clones positivos
Os clones positivos confirmados pela PCR de colônias foram repicados em uma nova placa
com meio - LB ágar e Amp - com identificação adequada, para posterior utilização. Desses
mesmos clones foram feitas culturas líquidas em LB/Amp e incubadas sob agitação à 37 ºC
por 16 horas. A partir dessas culturas contendo os clones positivos, foram feitas as
extrações de DNA plasmidial utilizando um Kit comercial (Qiagen plasmid Mini Hand Book
Kit). Em seguida foi feito um sequenciamento automático baseado no método didesoxi,
usando uma marcação fluorescente para cada ddNTP. A reação de sequenciamento foi feita
com o BigDye® Terminator v3.1 Cycle Sequencing Kit utilizando o sequenciador Abi31-30
da Applied Biosystems seguindo as recomendações do fabricante.

12
2.4.6 Subclonagem em vetor de expressão (pET-21a)
O pET-21a possui 5443pb, e como principais características, apresenta um gene que
confere resistência à ampicilina e um promotor forte, o T7 promoter de bacteriófago. Sua
região de clonagem possui sítios para várias enzimas de restrição, inclusive Nde I e BamH I,
que foram selecionadas para digestão do vetor e do inserto (Fig. 4 e 5).
Figura 4. Mapa do vetor pET-21a.
Fonte: Novagen
Figura 5. Região de clonagem e expressão do pET-21a. Em azul o promotor. Em vermelho os sítios de restrição
para as enzimas.
Fonte: Novagen

13
Foi escolhido um clone que estava com a seqüência correta do gene INF-β para ser utilizado
na subclonagem no vetor pET-21a. Esse clone foi colocado em cultura e seu plasmídeo
contendo o gene sintético INF β extraído por meio do kit comercial (Qiagen plasmid Mini
Hand Book Kit). A partir do produto da extração, foi realizada a digestão de 5µg desse
plasmídeo com as enzimas de restrição Nde I e BamH I (Fermentas Life Sciences) para
liberação do gene INF-β. Através de uma eletroforese em gel de agarose 1%, foi separada a
banda correspondente ao plasmídeo e ao gene INF-β. A banda correspondente ao gene
INF-β foi extraída do gel por meio do Kit GFX PCR DNA and Gel Band Purification (GE). O
vetor pET-21a foi digerido com as mesmas enzimas, seguindo recomendações do
fabricante.
A ligação vetor/inserto foi realizada com a T4-DNA Ligase (Promega). As quantidades de
vetor e inserto utilizadas para as reações de ligação foram obtidas de acordo com a fórmula
descrita anteriormente e as condições testadas são mostradas na tabela 1.
Tabela 1 - Reagentes e condições para reação de lig ação entre vetor/inserto
REAGENTES CONDIÇÕES
A (1:2,5) B (1:5)
pET-21a 4,0µl (280ng) 4,0µl (280ng)
Inserto 2,0µl (70ng) 4,0µl (140ng)
Tampão 10X 1,0µl 1,0µl
T4-DNA Ligase 0,5µl (1,5 und) 0,5µl (1,5 und)
H2O 2,5µl 0,5µl
Volume total 10,0µl 10,0µl
As células competentes, a transformação, a PCR de colônias e o sequenciamento, foram
realizadas como descrito anteriormente, mudando apenas a linhagem de E. coli, DH5α para
BL21. Esta linhagem é adaptada para expressão, possuindo em seu genoma o gene da
enzima T7 polimerase que reconhecerá o promotor T7 do vetor pET-21a.
2.5 Expressão da proteína recombinante INF- β
Após a transformação do plasmídeo recombinante em E. coli (linhagem BL21), foram
escolhidos aleatoriamente algumas colônias sendo submetidas a PCR para confirmação da
presença do mesmo. Confirmada a presença, esses clones foram seqüenciados e escolheu-
se o que apresentou a seqüência correta para a indução da expressão.

14
A indução foi realizada incubando-se a colônia em meio LB/AMP a 37 ºC com agitação.
Foram testadas DO entre 0,4 e 1 com absorbância de 600nm (A600). Foi retirada uma
amostra de 1 ml (T0), e ao restante da cultura foi adicionado o indutor β-D-isopropil-
tiogalactopiranosídeo (IPTG) em concentrações diferentes, variando entre 0,25 e 3 mM, e
continuada a incubação. Em intervalos de uma hora foi retirado uma amostra de 1 ml
correspondendo aos tempos T1, T2, T3, T4 e durante a noite, por aproximadamente 20h. As
amostras foram analisadas por eletroforese em gel de poliacrilamida-dodecil sulfato de sódio
(SDS-PAGE) para avaliação da expressão.
Para confirmação da expressão da proteína INF-β recombinante, uma alíquota do lisado da
cultura após 4h de indução foi submetida a SDS-PAGE e Western blot, utilizando anticorpo
anti-INF-β de coelho e como conjugado a Proteína A-peroxidase.
2.6 Purificação da proteína recombinante INF- β
2.6.1 Determinação da solubilidade da proteína recombinante INF-β
Após a indução com IPTG as células foram coletadas por centrifugação a 3000 rpm. O
sobrenadante foi descartado e o precipitado armazenado à -20 ºC por 30 minutos, e logo
após deixado a temperatura ambiente por 10 minutos para ruptura das células. Essas
células foram ressuspendidas em tampão de lise (Tris-HCl 50 mM pH 8,0; EDTA 2mM),
sonicadas 3 vezes por 30 segundos e centrifugadas a 12000 rpm a 4 ºC por 15 mim para
separação das proteínas solúveis e insolúveis (corpos de inclusão).
O precipitado de proteínas coletado na etapa anterior (corpos de inclusão) foram submetidas
a testes de solubilização com uréia. Foi adicionado ao precipitado de proteínas o tampão
uréia (Uréia 8M; 400mM de Tris-HCl pH 8,0; 2mM de EDTA pH 8,0; 100 mM de NaCl)
ficando em repouso a temperatura ambiente por 30 mim. Após esse período, o precipitado
foi ressuspendido e deixado em repouso por diferentes tempos (30 mim, 2h, 4h e 24h). Logo
após cada período de repouso, cada amostra foi centrifugada por 5 min à 14000 rpm e o
sobrenadante e o precipitado foram analisados em SDS-PAGE.

15
3 RESULTADOS E DISCUSSÃO
3.1 Construção do gene INF- β sintético
Os resultados mais expressivos da construção do gene sintético foram obtidos utilizando
concentrações de oligonucleotídeos de 0,25; 0,5; e 1 pmoles e temperaturas de anelamento
de 65, 68 e 70 ºC. À medida que aumentou-se a concentração dos oligonucleotídeos – até 1
pmol – a banda de interesse se tornou mais nítida, o que corresponde a um maior número
de amplicons (Fig. 6). Observou-se também a presença de fragmentos menores que 501 pb.
A utilização da técnica de PCR para montagem de um gene sintético, ocasiona o
aparecimento de bandas inespecíficas na primeira PCR, de construção, devido a um maior
número de oligonucleotídeos em uma única reação. Esses oligonucleotídeos podem se
anelar de forma incorreta, gerando fragmentos menores que a banda de interesse (Singh et
al., 1986).
Figura 6. Eletroforese em gel de agarose 1% da construção do gene sintético
Notas: Canaletas 1 e 8, Marcador 1 Kb Gibco. Canaletas 2, 3 e 4, concentrações dos oligos 0,25 pmol e temperaturas de anelamento 65, 68 e 70 ºC, respectivamente. Canaletas 5, 6 e 7, concentrações dos oligos 0,5 pmol e temperaturas de anelamento 65, 68 e 70 ºC, respectivamente. Canaletas 9, 10 e 11, concentração dos oligos 1pmol e temperaturas de anelamento 65, 68 e 70 ºC, respectivamente.
3.2 Amplificação do gene INF- β sintético
Na PCR para amplificação do gene sintético, foi observado que quanto maior a
concentração dos oligonucleotídeos utilizados na PCR de construção - até 1 pmol com
temperatura de anelamento igual a 55 ºC - maior o número de bandas inespecíficas e a
intensidade da banda de interesse (501pb). As melhores condições obtidas foram utilizando

16
o produto da construção do gene sintético de concentração igual a 1 pmol a uma
temperatura de anelamento de 65 ºC, que conferiu o resultado esperado, onde apenas a
banda de interesse foi amplificada (Fig. 7). Provavelmente essa especificidade se deu pelo
aumento da temperatura de anelamento, favorecendo a ligação dos primers apenas nas
regiões específicas.
Figura 7. Eletroforese em gel de agarose 1% da amplificação do gene sintético
Notas: Canaleta 1, Marcador 1 Kb Gibco. Canaletas 2, 3 e 4, amplificação com o produto da construção de 0,25 pmol com temperatura de anelamento de 55 ºC. Canaletas 5, 6 e 7, amplificação com o produto da construção de 0,5 pmol com temperatura de anelamento de 55 ºC. Canaletas 8, 9 e 10, amplificação com o produto da construção de 1pmol com temperatura de anelamento de 55 ºC. Canaletas 11, 12 e 13, amplificação com o produto da construção de 1 pmol com temperatura de anelamento de 65 ºC.
3.3 Clonagem do gene INF- β sintético
A clonagem foi feita no vetor pCR 2.1 como descrito na metodologia, obtendo dezenove
colônias positivas (brancas) identificadas pela seleção branco/azul. Selecionou-se duas
colônias rotuladas como amostras 10 e 11 e foi feita a extração plasmidial utilizando o Kit
comercial (Qiagen plasmid Mini Hand Book Kit). A confirmação de que essas colônias eram
realmente positivas, se deu através de uma PCR dos plasmídeos extraídos (Fig.8).

17
Figura 8. Eletroforese em gel de agarose 1% da PCR do vetor pCR 2.1 depois da clonagem
Notas: Canaleta 1, Marcador 1 Kb Gibco. Canaletas 2 e 3, corresponde as amostras 10 e 11 respectivamente.
O resultado do sequenciamento automático dessas amostras mostrou que a amostra 11
estava com a seqüência correta. Comparando o sequenciamento da amostra 11 com a
seqüência de nucleotídeos do gene INF-β com adaptação do uso de códons preferenciais,
foi constatado que não houve nenhuma alteração na seqüência dos nucleotídeos (Fig. 9).
Essa comparação foi importante, pois como utilizou-se o gene INF-β sintético com
adaptação do uso de códons preferenciais para E. coli, necessita-se de uma seqüência
nucleotídica sem alterações. O uso de genes sintéticos é sugerido por Kane (1995), pois
proteínas heterólogas possuem vários códons raros devido ao baixo nível de tRNA para os
mesmos em linhagens hospedeiras como a E. coli, e como conseqüência acarreta um baixo
nível da expressão da proteína de interesse.

18
Figura 9. Comparação nucleotídica no Programa Clustal
.Notas: INF-BETA corresponde à seqüência do gene com adaptação do uso preferencial de códons. TA-CLONE11 corresponde ao sequenciamento da amostra 11. O (*), indica bases iguais.
Após a confirmação que não houve alterações na seqüência nucleotídica, realizou-se uma
comparação com o banco de dados de proteínas do NCBI por meio do programa Blastx -
que através da seqüência nucleotídica busca no banco de dados a proteína correspondente
- onde se constatou que a mutação da cisteína 17 por serina também estava correta, como
mostrada na figura 10. A seqüência da proteína INF-β possui três cisteínas nas posições17,
31 e 141. As cisteínas 31 e 141 são ligadas através de ponte de sulfeto, de maneira a dar a
conformação correta para a proteína. Como os organismos procariontes possuem
deficiências em processos pós-traducionais, foi feita essa mutação para evitar a ligação
entre cisteínas em posições não desejadas. Mark et al. (1984) em seus estudos, mostrou

19
que essa mutação não alterou a atividade da proteína INF-β recombinante produzida em E.
coli quando comparada com a proteína INF-β nativa de fibroblastos.
Figura 10. Programa BlastX, NCBI.
OBS: Comparação do sequenciamento com o banco de dados de proteínas do NCBI. Em vermelho, está indicada a mutação na posição 17.
A partir da amostra 11 - vetor pCR 2.1 contendo o gene sintético INF-β - foi realizada uma
digestão como descrito na metodologia para liberação do gene sintético e posterior
clonagem em vetor de expressão, como mostra a figura 11. O vetor de expressão pET-21a
também foi digerido com as mesmas enzimas (Fig. 12).
Figura 11. Eletroforese em gel de agarose 1% da digestão do vetor pCR 2.1 contendo o gene sintético INF-β.
Nota: Canaleta 4, marcador 1Kb Ladder (Invitrogen). Canaletas 1, 2, 3, 5, 6 e 7, pCR 2.1 digerido. A seta indica a banda correspondente ao gene sintético liberado pela digestão.
20
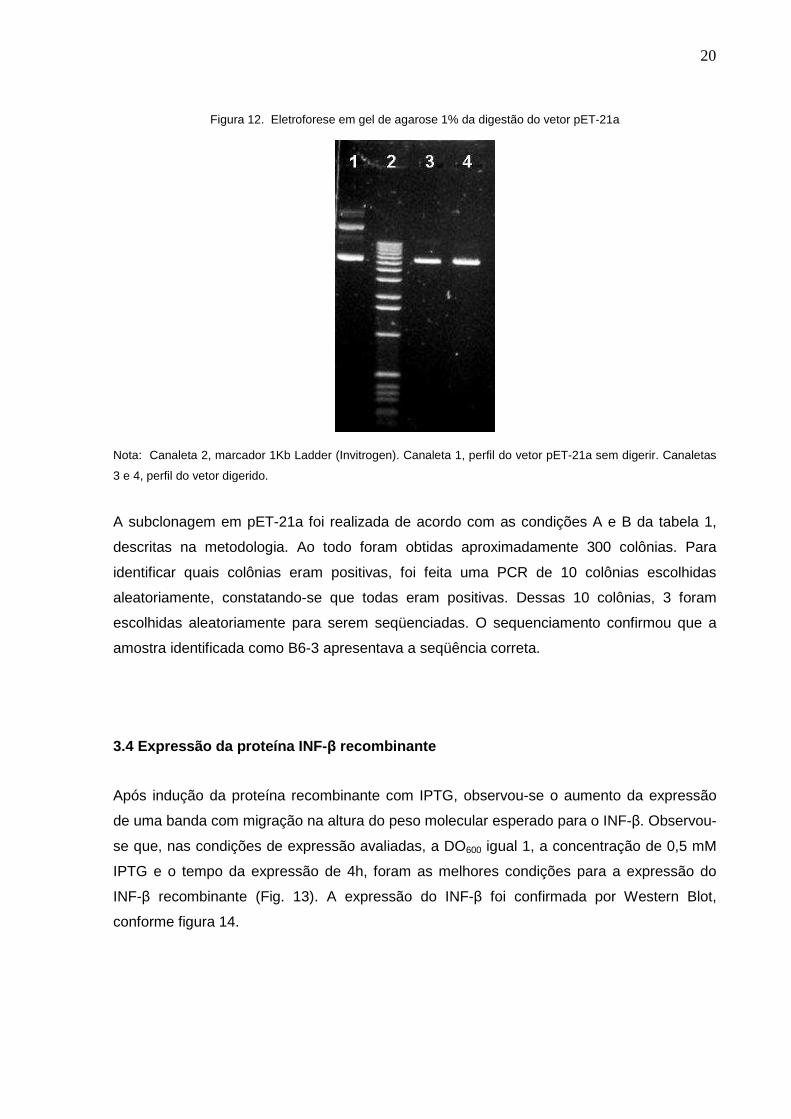
Figura 12. Eletroforese em gel de agarose 1% da digestão do vetor pET-21a
Nota: Canaleta 2, marcador 1Kb Ladder (Invitrogen). Canaleta 1, perfil do vetor pET-21a sem digerir. Canaletas
3 e 4, perfil do vetor digerido.
A subclonagem em pET-21a foi realizada de acordo com as condições A e B da tabela 1,
descritas na metodologia. Ao todo foram obtidas aproximadamente 300 colônias. Para
identificar quais colônias eram positivas, foi feita uma PCR de 10 colônias escolhidas
aleatoriamente, constatando-se que todas eram positivas. Dessas 10 colônias, 3 foram
escolhidas aleatoriamente para serem seqüenciadas. O sequenciamento confirmou que a
amostra identificada como B6-3 apresentava a seqüência correta.
3.4 Expressão da proteína INF- β recombinante
Após indução da proteína recombinante com IPTG, observou-se o aumento da expressão
de uma banda com migração na altura do peso molecular esperado para o INF-β. Observou-
se que, nas condições de expressão avaliadas, a DO600 igual 1, a concentração de 0,5 mM
IPTG e o tempo da expressão de 4h, foram as melhores condições para a expressão do
INF-β recombinante (Fig. 13). A expressão do INF-β foi confirmada por Western Blot,
conforme figura 14.

21
Figura 13. SDS-PAGE da expressão do INF-β recombinante
Nota: Canaleta 1, marcador molecular de proteínas (Fermentas). Canaletas 2, 3, T0 (sem IPTG) e T4 (4h), respectivamente. A seta indica a banda correspondente a proteína recombinante expressa.
Figura 14. Western Blot da expressão do INF-β recombinante
Notas: Canaleta 1, marcador molecular de proteínas (Fermentas). Canaleta 2, T4 (4h). A seta indica o INF-β recombinante expresso e reconhecido pelo anticorpo específico anti- INF-β.
Os baixos níveis de expressão da proteína INF-β em sistemas procariotas já foram relatados
por alguns autores. Segundo Warne & Thomes (1986) o INF-β é altamente tóxico para E.
coli e a expressão nos estágios iniciais é deletério para a célula hospedeira. No presente
trabalho observou-se este efeito deletério, pois os melhores níveis de expressão foram
alcançados com DO600 igual a 1, ou seja, a indução tardia aumentou os níveis de expressão
da proteína de interesse (INF-β).
~18kDa
~18kDa

22
A concentração de IPTG mostrou-se importante na expressão do INF-β recombinante, pois
em concentrações maiores que 1 mM não foi benéfico, levando a rendimentos inferiores
quando utilizado em concentração igual a 3 mM. Rao et al. (2009), observaram que a alta
concentração de IPTG não metabolizada presente no citoplasma, pode causar toxicidade à
E. coli.
Apesar das dificuldades encontradas na expressão do INF-β, e de se esperar maiores níveis
de expressão, quando comparado a outros trabalhos como Ghane et al. (2008), Rao et al.
(2009) e Villela et al. (2010), observou-se que os níveis de expressão obtidos no presente
estudo foram satisfatórios.
3.5 Purificação da proteína recombinante INF- β
O alto nível de expressão de proteínas eucariotas em E. coli muitas vezes resultam em
deposições citoplasmáticas insolúveis, conhecidas como corpos de inclusão. Microscopia
eletrônica mostra os corpos de inclusão como sendo agregados de proteínas sem uma
organização estrutural definida. Vários são os fatores que contribuem para a formação de
corpos de inclusão como as características intrínsecas da célula utilizada, da proteína a ser
expressa e o nível de expressão. A ocorrência de corpos de inclusão é um fato
relativamente comum na expressão de proteínas heterólogas em bactérias e pode trazer
dificuldades nas etapas seguintes para obtenção da proteína (Carrió et al., 2000).
Por serem agregados insolúveis os corpos de inclusão necessitam de condições extremas
para a sua solubilização. Dentre os sais mais utilizados estão a Uréia e Cloridrato de
Guanidina em altas concentrações. Esses permitem a solubilização dos corpos de inclusão,
no entanto, desnaturam a proteína, exigindo etapas posteriores de renaturação.
O INF-β recombinante, obtido neste trabalho, apresentou-se como uma proteína insolúvel e
o melhor resultado de solubilização foi após deixar o precipitado de proteínas em tampão
uréia 8M por 24h, como mostra a figura 15. Mesmo assim, a solubilização foi parcial e até o
momento não foram feitas tentativas de purificação em colunas cromatográficas.
Em outros estudos envolvendo a expressão do INF-β em E. coli a proteína também
apresentou-se insolúvel, necessitando de técnicas para solubilização utilizando diferentes
desnaturantes como dodecil sulfato de sódio (SDS) 10%, guanidina 6M e uréia 8M para
posterior purificação e renaturação (Ghane et al. 2008; Rao et al. 2009; Villela et al. 2010).

23
Figura 15. SDS-PAGE da solubilização do INF-β recombinante.
Notas: Canaleta 1, marcador molecular de proteínas (Fermentas). Canaletas 2, expressão do INF-β (T4). Canaleta 3, sobrenadante sem uréia (proteínas solúveis). Canaleta 4, sobrenadante depois da solubilização com uréia (proteínas solúveis). Canaleta 5, precipitado depois da solubilização com uréia (proteínas insolúveis). A seta indica a banda do INF-β recombinante.
4 CONCLUSÕES
Neste trabalho, foi realizado com êxito a construção do gene sintético utilizando códons
preferenciais, a clonagem e a expressão do gene INF-β. Ao final dessas etapas, conseguiu-
se isolar um clone eficiente com a expressão do INF-β recombinante otimizada, que será
utilizado na continuação deste trabalho até que se obtenha INF-β purificado e sua atividade
testada.
Após todas as etapas concluídas com sucesso, novos testes serão feitos em maior escala,
para que no futuro, a proteína recombinante INF-β possa ser produzida em larga escala e
incorporada ao portifólio de biofármacos da Funed, que atualmente está realizando a
transferência de tecnologia para implementação de uma unidade fabril para produção do
Interferon alfa-2a humano recombinante e eritropoetina humana recombinante.
~18kDa

24
Agradecimentos
Este trabalho foi uma iniciativa da Dra. Roberta Márcia Marques dos Santos pertencente à
Diretoria Industrial da Funed e financiado pela Fundação de Amparo a Pesquisa de Minas
Gerais (FAPEMIG). O projeto foi realizado no Laboratório de Inovação Biotecnológica da
Diretoria de Pesquisa na Funed. Agradeço a Dra. Roberta Márcia Marques dos Santos pela
orientação, a Dra. Sophie Yvette Leclercq pela co-orientação e a Msc. Patrícia Cota Campos
pela colaboração.
REFERÊNCIAS BIBLIOGRÁFICAS
Abbas AK, Pober JS, Lichtman AH. Imunologia Celular e Molecular. 4 ed. Rio de Janeiro - Livraria e editora Revinter, 2003, p. 251-260.
Carrió MM, Cubarsi R, Villaverde A. Fine architecture of bacterial inclusion bodies. FEBS Lett 2000; 471(1): 7-11.
Derynck R, Remaut E, Saman E, Stanssens P, Clercq E, Content J, Fiers W. Expression of human fibroblast interferon gene in Escherichia coli. Nature 1980; 18(287): 193-197.
European Study Group on interferon β-1b in Secundary Progressive MS. Placebo-controlled multicentre randomized trial of interferon β-1b in treatment of secondary progressive multiple sclerosis. Lancet 1998; 352(9139): 1491-1497.
Ghane M, Yakhchali B, Khodabandeh M. Over expression of biologically active interferon beta using synthetic gene in E. coli. Journal of Sciences 2008; 19(3): 203-209.
Galvani ALS, Krebs VLJ, Vaz FAC. Biochemical characteristics and properties of the humoral mediators in bacterial infections. São Paulo, Pediatria 1999; 21: 123-132.
Goldman L, Ausiello D. Cecil. Tratado de medicina interna. 22 ed. Rio de Janeiro - Editora Elsevier Ltda, 2005; 2: 2719.
Hartung H-P, Archelos JJ, Zielasek J, Gold R, Koltzenburg M, Reiners K-H, Toyka KV. Circulating adhesion molecules and inflammatory mediators in demyelination: a review. Neurology 1995; 45(6): 22-32.
Hiscott J, Pitha PM. Interferon regulatory factors: bridging antiviral defense, growth control, and immunoregulation. Journal of interferon and cytokine research 2002; 22: 3-4.
Invitrogen life technologies. TA Cloning® Kit. Manual 2004.
Jana S, Deb JK. Strategies for efficient production of heterologus proteins in Escherichia coli. Microbiology e Biotechnology 2005; 67: 289-298.

25
Kagawa Y, Takasaki S, Utsumi J, Hosoi K, Shimzu H, Kochibe N, Kobata A. Comparative study of the asparagine-linked sugar chains of natural human interferon-beta 1 and recombinant human interferon-beta 1 produced by three different mammalian cells. The Journal of Biological Chemistry 1988; 25(263): 17508-17515.
Kane, JF. Effects of rare codon clusters on high-level expression of heterologous proteins in Escherichia coli. Current opinion in biotechnology 1995; 6(5): 494-500.
Karpusas M, Nolte M, Benton CB, Meier W, Lipscomb WN, Goelz S. The crystal structure of human interferon β. A resolution Proceedings of the National Academy of Sciences 1997; 94: 11813-11818.
Lana-Peixoto MA, Callegaro D, Moreira MA, Campos GB, Marchiori PE, Gabbai AA, et al. Consenso expandido do BCTRIMS para o tratamento de esclerose múltipla. Arq Neuropsiquiatr 2002; 60(3-B): 881-886.
Lima BD. A produção de insulina humana por engenharia genética. Biotecnologia Ciência & Desenvolvimento 2001; 23: 28-31.
Mark DF, Lu SD, Creasey AA, Yamamoto R, Lin LS. Site-specific mutagenesis of the human fibroblast interferon gene. Proceedings of the National Academy of Sciences 1984; 81: 5662-5666.
Moreira MA, Felipe E, Mendes MF, Tilbery CP. Esclerose múltipla estudo descritivo de suas formas clínicas em 302 casos. Arq Neuropsiquiatr 2000; 58(2-B): 460-466.
National MS Society. How many people have MS? Disponível em: <http://www.nationalmssociety.org/about-multiple-sclerosis/what-we-know-about-ms/faqs-about-ms/index.aspx >. Acesso em: 28 out. 2010.
Novagen. pET System Manual. 11 ed. Estados Unidos da América, 2007.
Ohno S, Taniguchi T. Structure of a chromosomal gene for human interferon β. Proceedings of the National Academy of Sciences 1981; 78(9): 5305-5309.
Paty DW, Li DKB - IFNB multiple sclerosis study group. Interferon beta-1b is effective in relapsing-remitting multiple sclerosis. Clinical results of a multicenter, randomized, double-blind, placebo-controlled trial. Neurology 1993; 43: 662-667.
Platanias LC. Interferon signal: What Is Classical and What Is Nonclassical?. Journal of interferon and cytokine research 2005; 25(12): 732.
Pestka S. The interferons - 50 years after their discovery, there is much more to learn. Journal of Biological Chemistry 2007; 282(28): 20047-20051.
Rao DVK, Ramu CT, Rao JV, Narasu ML, Rao AKSB. Cloning, high expression and purification of recombinant human intereferon-β-1b in Escherichia coli. Appl Biochem Biotechnol 2009; 158: 140-154.

26
Roitt I, Brostoff J, Male D. Imunology. London, UK: Mosby International Ltda, 1998; p.222-223.
Siffrin V, Vogt J, Radbruch H, Nitsch R, Zipp F. Multiple sclerosis – candidate mechanisms underlying CNS atrophy. Trends in Neurosciences 2010; 33(4): 202-210.
Singh PK, Sarangi BK, Tuli R. A facile method for the construction of synthetic genes. Journal Bioscience 1996; 21(6):735-741.
Sottini A, Capra R, F. Serana, Chiarini M, Caimi L, Imberti L. Interferon-Beta Therapy Monitoring in Multiple Sclerosis Patients. Endocrine, Metabolic & Immune Disorders - Drug Targets 2009; 9(1): 14-28.
Taniguchi T, Guarente L, Roberts TM, Kimelman D, Douhan III J, Ptashne M. Expression of the human fibroblast interferon gene in Escherichia coli. Proceedings of the National Academy of Sciences 1980; 77(9): 5230-5233.
Tilbery CP, Mendes MF, Oliveira BES, Thomaz RB, Kelian GR. Immunomodulatory treatment in multiple sclerosis. Arq Neuropsiquiatr 2006; 64 (1): 51-54.
Tilbery CP, Moreira MA, Mendes MF, Lana-Peixoto MA. Recomendações quanto ao uso de drogas imunomoduladoras na esclerose múltipla. Arq Neuropsiquiatr 2000; 58(3-A): 769-776.
Uitdehaag BMJ, Hoekstra K, Koper JW, Polman CH, Dijkstra, CD. IFN- β1b augments glucocorticoid-induced supression of tumor necrosis factor-α production by increasing the number of glucocorticoid receptors on a human monocytic cell line. Journal of interferon and cytokine research 2001; 21: 133-135.
Villela AD, Renard G, Palma MS, Chies JM, Dalmora SL, Basso LA, Santos DS. Human interferon b1ser17: coding DNA synthesis, expression, purification and characterization of bioactive recombinant protein. Journal of Microbial & Biochemical Technology 2010; 2(5): 111-117.
Warne, S, Thomas M. Use of modified E. coli trpR gene to obtain tight regulation of high copy number expression vectors. Gene 1986; 46:103-112.

![&Z Ì ] & } u dZ /E/E'](https://static.fdocument.org/doc/165x107/629d9bc2687b3969ab0e8d75/ampz-amp-u-dz-ee.jpg)
















