
Downloaded from on May 9, 2020 by guest · 131 (Sigma, St. Louis, MO) dissolved in DMSO diluted 1:5...
35
1 Epithelial PI3K signaling is required for β-catenin activation and host defense 1 against Citrobacter rodentium infection 2 3 Short title: PI3K in intestinal epithelial host defense 4 Jeffrey B. Brown 1 , Paul Cheresh 2 , Tatiana Goretsky 2 , Elizabeth Managlia 2 , Gery R. 5 Grimm 2 , Hyunji Ryu 2 , Mojgan Zadeh 2 , Ramanarao Dirisina 2 , Terrence A. Barrett 2* 6 1 Department of Pediatrics, Northwestern University Feinberg School of Medicine 7 /Children’s Memorial Hospital, 2 Department of Medicine, Northwestern University 8 Feinberg School of Medicine, Chicago, Illinois, USA 9 Correspondence address: 10 Terrence A. Barrett, MD 11 Department of Internal Medicine, Microbiology and Immunology, and Surgery 12 Northwestern University Feinberg School of Medicine 13 676 N. St. Claire Suite 1400 14 Chicago, IL 60611 15 E-mail: [email protected] 16 Tel: 312-503-0293 17 Fax: 312-503-8423 18 19 20 21 22 23 Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved. Infect. Immun. doi:10.1128/IAI.01025-10 IAI Accepts, published online ahead of print on 22 February 2011 on June 26, 2020 by guest http://iai.asm.org/ Downloaded from
Transcript of Downloaded from on May 9, 2020 by guest · 131 (Sigma, St. Louis, MO) dissolved in DMSO diluted 1:5...

1
Epithelial PI3K signaling is required for β-catenin activation and host defense 1
against Citrobacter rodentium infection 2
3
Short title: PI3K in intestinal epithelial host defense 4
Jeffrey B. Brown1, Paul Cheresh2, Tatiana Goretsky2, Elizabeth Managlia2, Gery R. 5
Grimm2, Hyunji Ryu2, Mojgan Zadeh2, Ramanarao Dirisina2, Terrence A. Barrett2* 6
1Department of Pediatrics, Northwestern University Feinberg School of Medicine 7
/Children’s Memorial Hospital, 2Department of Medicine, Northwestern University 8
Feinberg School of Medicine, Chicago, Illinois, USA 9
Correspondence address: 10
Terrence A. Barrett, MD 11
Department of Internal Medicine, Microbiology and Immunology, and Surgery 12
Northwestern University Feinberg School of Medicine 13
676 N. St. Claire Suite 1400 14
Chicago, IL 60611 15
E-mail: [email protected] 16
Tel: 312-503-0293 17
Fax: 312-503-8423 18
19
20
21
22
23
Copyright © 2011, American Society for Microbiology and/or the Listed Authors/Institutions. All Rights Reserved.Infect. Immun. doi:10.1128/IAI.01025-10 IAI Accepts, published online ahead of print on 22 February 2011
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

2
ABSTRACT 24
Citrobacter rodentium (C. rodentium) infection of mice induces cell mediated immune 25
responses associated with crypt hyperplasia and epithelial β-catenin signaling. Recent 26
data suggest that phosphatidylinositol-3-kinase (PI3K)/Akt signaling cooperates with 27
Wnt to activate β-catenin in intestinal stem and progenitor cells through phosphorylation 28
at Ser552 (P-β-catenin552). Our aim was to determine whether epithelial PI3K/Akt-29
activation is required for β-catenin signaling and host defense against C. rodentium. 30
C57/BL6 mice were infected with C. rodentium and treated with DMSO (vehicle control) 31
or with the PI3K inhibitors, LY294002 or wortmannin. Effects of infection on PI3K 32
activation and β-catenin signaling were analyzed by immunohistochemistry. Effects of 33
PI3K inhibition on host defense were analyzed by quantification of splenic and colon 34
bacterial clearance, and adaptive immune responses were measured by RT-PCR and 35
ELISA. Increased numbers of P-β-catenin552-stained epithelial cells were found 36
throughout expanded crypts in C. rodentium colitis. We show that inhibition of PI3K 37
signaling attenuates epithelial Akt activation, Ser552 phosphorylation and activation of 38
β-catenin, and epithelial proliferative responses during C. rodentium infection. PI3K 39
inhibition impairs bacterial clearance despite having no impact on mucosal cytokine 40
(IFN-γ, TNF, IL-17, IL-1β) or chemokine (CXCL1, 5, 9, 10) induction. The results 41
suggest that host defense against C. rodentium requires epithelial PI3K activation to 42
induce Akt-mediated β-catenin signaling and clearance of C. rodentium, independent of 43
adaptive immune responses. 44
45
46
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

3
Phosphatidylinositol-3-kinase (PI3K)/Akt signaling induces cell proliferative, growth, and 47
survival signals in intestinal epithelial cells (IEC)(3, 17, 61). PI3K phosphorylates PIP2 48
to generate PIP3, which functions as a ligand to recruit pleckstrin homology (PH)-49
domain-containing proteins to the inner surface of the cellular membrane. Chief among 50
these is Akt, a serine/threonine kinase that is itself phosphorylated upon activation. Akt 51
has emerged as a critical signaling node in mammalian cells and mediates the majority 52
of PI3K’s downstream effects. Amongst its many pro-proliferative functions, Akt 53
signaling enhances mRNA stability and protein synthesis (via mTOR and S6 kinase) 54
and uptake of energy substrates(36). Activated Akt opposes apoptotic signaling by 55
phosphorylating and sequestering bcl-xL/bcl-2-associated death promoter homolog 56
(BAD) and Forkhead box factors (FOXO) in the cytoplasm. Recent findings suggest 57
activated Akt also acts in the nucleus by phosphorylating substrates, such as β-catenin, 58
that increase transcription of target genes associated with proliferation (e.g. cyclinD1, c-59
Myc), tissue remodeling (e.g. MMP7), and host defense (e.g. cryptdins, Muc2, intestinal 60
trefoil factor)(15, 18, 33). 61
Wnt/β-catenin signaling is critical to the maintenance of intestinal crypt cell 62
proliferation. Wnt ligands bind intestinal stem cells (ISC) and crypt epithelial progenitor 63
cells (PC) and prevent GSK3β-dependent N-terminal phosphorylation and proteosomal 64
degradation of β-catenin(22, 29, 55, 60). When degradation is impaired by Wnt 65
signaling, β-catenin translocates to the nucleus (a hallmark of activated canonical 66
Wnt/β-catenin signaling(43)) where it initiates transcription of Wnt target genes such as 67
c-Myc and cyclinD1(23, 29, 47). Upon division, ISCs and PCs give rise to rapidly cycling 68
transit-amplifying cells(4, 6, 37). Previous studies showed that deletion of the 69
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

4
Phosphatase and Tensin homolog (PTEN), a negative regulator of PI3K, increases 70
PI3K/Akt activation and enhances nuclear β-catenin accumulation(42). More recently, 71
He et al reported that PTEN deficiency increases PI3K/Akt activation resulting in 72
excessive proliferation and crypt fissioning within small bowel (SB) polyps(24). To 73
investigate the interface between activated PI3K/Akt and β-catenin stabilization, 74
researchers developed a phospho-specific antibody that identifies β-catenin 75
phosphorylated by Akt at Ser552 (P-β-catenin552)(24). Compared to N-terminal 76
phosphorylation which targets β-catenin for degradation, C-terminal phosphorylation at 77
Ser552 is detected on nuclear β-catenin associated with enhanced transcription of Wnt/ 78
β-catenin target genes(18, 24). Staining of tissue from SB polyps found P-β-catenin552 79
within nuclei of dividing (activated) ISCs and PCs in crypt bases and at points of crypt 80
branching(24). Taken together, these results suggest PI3K/Akt activation cooperates 81
with Wnt to promote β-catenin signaling in ISCs and PCs. 82
Overactive PI3K signaling occurs in a variety of tumor types, including breast, 83
lung, ovarian, pancreatic, prostate, and colon cancers(1, 14, 20, 41, 46, 57, 62). 84
Furthermore, defects in PTEN occur in a wide spectrum of human cancers(10). Recent 85
studies by Mayer and colleagues suggest PI3K/Akt signaling is upregulated in crypt IEC 86
in chronic ulcerative colitis and active Crohn’s disease(11). We recently reported that 5-87
aminosalicylic acid treatment impairs Akt-mediated β-catenin signaling and prevents the 88
development of colitis-associated dysplasia(8). These data suggest that activation of 89
PI3K is an important step in activation of IEC β-catenin signaling in dysplasia and 90
cancer. As activating mutations of the p110 subunit of PI3K are amongst the most 91
common detected in all of human cancer(27, 45, 46), several newly developed cancer 92
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

5
therapeutics target this pathway(30, 35). Therefore, molecules that interfere with PI3K 93
signaling may be chemopreventive in inflammation-induced cancer by inhibiting β-94
catenin signaling and impairing ISC and PC activation. 95
Despite literature examining PI3K and β-catenin in crypt maintenance, cell 96
survival and malignancy, little is known about its role in intestinal inflammation. This is 97
especially important in infectious enterocolitis where maintenance of the epithelial 98
barrier is critical to survival. Citrobacter rodentium (C. rodentium) is classified as an 99
attaching and effacing (A/E) pathogen whose colonization of colon epithelia results in 100
epithelial hyperplasia and mucosal inflammation(7). C. rodentium induces crypt 101
hyperplasia, promotes PI3K recruitment to lipid rafts(51) and increases levels of nuclear 102
β-catenin in crypt IEC(49, 50). Studies here address the possibility that efficient host 103
defense responses to C. rodentium infection involve PI3K-induced β-catenin activation. 104
Our results indicate C. rodentium infection increases IEC levels of activated P-Akt and 105
nuclear P-β-catenin552. To determine the requirement for PI3K regulation of epithelial 106
β-catenin signaling in host defense, we inhibited PI3K signaling in mice inoculated with 107
C. rodentium. Loss of PI3K signaling in infected mice reduced IEC Akt and β-catenin 108
activation and attenuated IEC BrdU incorporation. Inhibition of PI3K signaling delayed 109
bacterial clearance while not affecting inflammatory responses. Taken together, these 110
results suggest PI3K-mediated Akt and β-catenin signaling play critical roles in 111
epithelial-specific host responses to enteric infection. 112
Materials and Methods 113
Animals 114
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

6
6-8 week old C57BL/6 mice (Jackson Laboratories, Bar Harbor, ME) were 115
maintained within infection-controlled containment housing in the Northwestern 116
University animal care facility. All experiments were approved by the Animal Care and 117
Usage Committee of Northwestern University. 118
Induction of Colitis and Study Design 119
C. rodentium strain DBS120(48), a kanamycin-resistant strain (a gift from David 120
Schauer, Boston, MA), were prepared by shaking at 37ºC overnight in LB broth. 121
Bacterial concentration was determined by measuring absorbance at optical density 122
600. Colony forming units (CFU’s) were confirmed through serial dilution and plating. 123
Mice were fasted for 8 hours prior to inoculation with 109 CFU’s in 200 uL sterile PBS. 124
During the study, mice were allowed free access to food and drinking water. Mice were 125
divided into 2 groups: Group 1 received 200 µL intraperitoneal (i.p.) injections of 126
dimethyl sulfoxide (DMSO) diluted 1:4 in phosphate buffered saline (PBS) (vehicle 127
control) on days 5, 8, and 13 after infection, and Group 2 received 1 mg (appr. 50 mg/kg 128
body weight) LY294002 (Sigma, St. Louis, MO) dissolved in DMSO diluted 1:4 in PBS. 129
In separate studies, a parallel set of mice in Group 2 received 0.1 mg/kg wortmannin 130
(Sigma, St. Louis, MO) dissolved in DMSO diluted 1:5 in PBS on days 4, 6, 8, and 10. 131
In studies to specifically examine IL-22 production, injections of DMSO or LY294002 132
were given on days 1, 3, and 5 after infection and mice sacrificed for analysis on days 4 133
and 7. 134
Histologic and Immunohistochemical (IHC) Analysis 135
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

7
A distal 1 cm piece of colon was removed from each mouse, fixed in 10% neutral 136
buffered formalin, routinely processed, sectioned at 5 µm, and stained with H&E for 137
microscopic examination. Crypt lengths were measured by micrometric sampling of at 138
least 15 crypts per mouse. Histologic colitis scores were determined by measuring the 139
severity of epithelial ulceration (0, none; 1, epithelial desquamation; 2, erosions with 140
gaps of 1-10 IECs; 3, ulcers >10 IECs long), the extent of inflammatory cell infiltration 141
(0, normal; 1, mild infiltration of the mucosa; 2, moderate-severe infiltration of the 142
mucosa; 3, mild infiltration of the submucosa; 4, moderate-severe infiltration of the 143
submucosa; 5, transmural infiltration into the serosa), and extent of submucosal edema 144
(0, none; 1, <200 µm; 2, 200-450 µm; 3, >450 µm). BrdU, P-Akt, and P-β-catenin552 145
IHC staining was performed as previously described(32). 146
Immunofluorescent Analysis 147
Paraffin tissue sections were subjected to antigen retrieval as described for P-β-148
catenin552 previously. Slides were incubated in rabbit anti-Myc Ab (Millipore/ Upstate, 149
Temecula, CA) diluted 1:700 in PBS for 48 hours at 4˚C. After washes with PBS, HRP-150
conjugated polymeric secondary antibody (EnVision anti-Rabbit HRP, DAKO) was 151
applied for 60 minutes at room temperature. Slides were fluorescently labeled with 152
Alexa 488 via Tyramide signal amplification, according to the manufacturer’s 153
instructions (Invitrogen), washed with PBS, stained with Hoechst 33342 (Invitrogen), 154
washed in distilled water and mounted in aqueous medium. Slides were imaged with a 155
Nikon C1 confocal microscope. 156
Colony-forming Unit Counts 157
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

8
At selected time points, colons were removed from anus to cecum and tissue and 158
stool were homogenized using a Polytron tissue homogenizer (Kinematica, 159
Switzerland). Spleens were homogenized through a 100µm mesh filter. Homogenates 160
were serially diluted, plated in duplicate on MacConkey agar (Fluka, Steinheim, 161
Germany) containing 40 µg/mL kanamycin (Fluka) (C. rodentium strain DBS120 carries 162
a kanamycin resistant plasmid), and colonies counted after 18 hours of incubation at 163
37ºC. 164
Real Time PCR Analysis 165
Tissue RNA was isolated using Trizol reagent (Invitrogen). Real-time (RT) PCR 166
analysis was performed using a qScript RT kit (Quanta, Gaithersburg, MD), QuantiTect 167
SYBR green PCR kit (Quigen), and primers on an ABI 7500 Real-time PCR System. 168
Primers were designed to span genomic DNA intron junctions for specific amplification 169
of mRNA. Samples were normalized using Gapdh. Fold increases were calculated 170
using the ddCT method. Primer sequences available upon request. 171
Colon explant culture ELISA 172
1 cm pieces of distal colonic tissue were weighed, irrigated with PBS, shaken at 173
room temperature in RPMI containing 50 µg/mL gentamicin for 30 minutes, and 174
distributed (0.05 g per well) into 24-well plates in 1 mL of RPMI supplemented with 5% 175
fetal bovine serum, 50 µg/mL gentamicin, and 1% penicillin/streptomycin/amphotericin B 176
(GIBCO, Grand Island, NY) for 20 hours at 37°C. Supernatants were collected and 177
stored at −80°C until protein quantification via ELISA kits (R&D, Minneapolis, MN). 178
Statistical Analysis 179
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

9
Data were analyzed with GraphPad Prism software by means of ANOVA 180
followed by Tukey’s range test when more than two groups were compared; otherwise, 181
the Student’s t test was used. P-values ≤ 0.05 were considered statistically significant. 182
Unless specified, values represent mean ± standard error of the mean (SEM). Unless 183
specified, data are representative of at least 2 independent experiments. 184
Results 185
C. rodentium colitis induces PI3K and β-catenin signaling in hyperplastic crypts 186
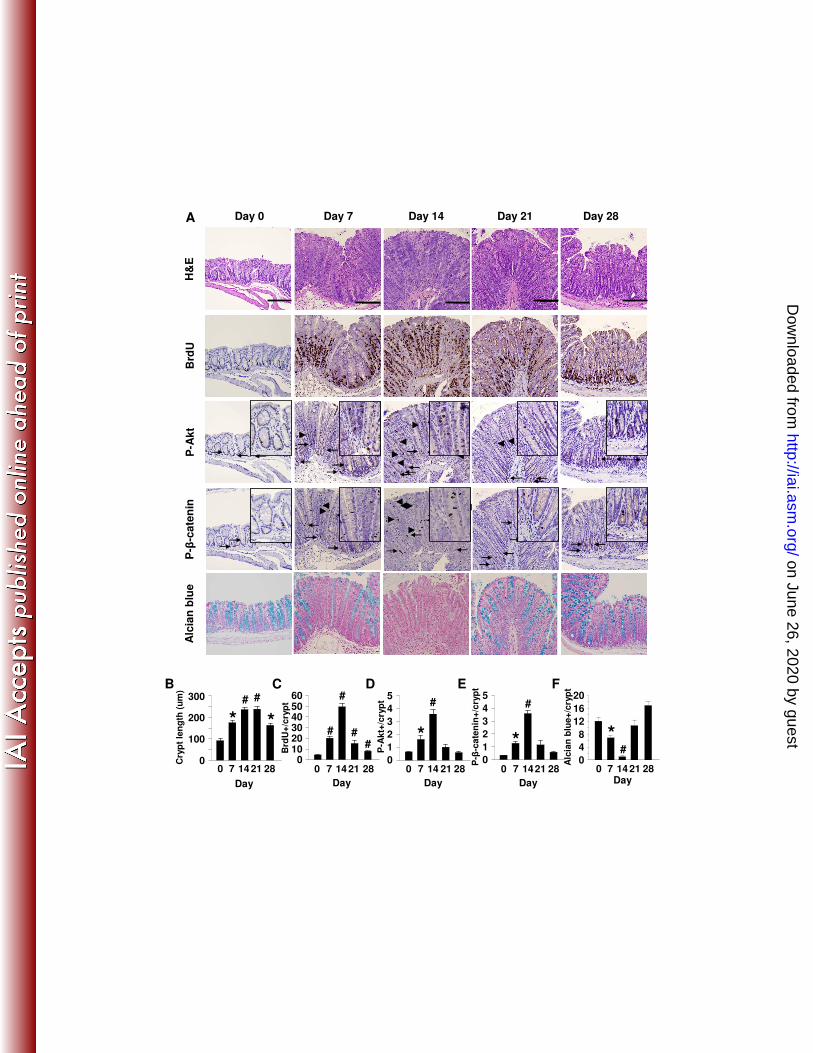
C. rodentium induces pronounced colonic crypt hyperplasia in mice. We 187
postulated that C. rodentium elicits epithelial proliferation by activating PI3K and β-188
catenin signaling. To examine this possibility, tissues were stained for BrdU 189
incorporation, P-Akt and P-β-catenin552 (Fig. 1A). Results indicate that crypt lengths 190
nearly doubled within one week of infection (baseline to day 7, mean ± SEM: 92 ± 10 to 191
176 ± 7 µm, Fig. 1B), then peaked within weeks two and three (239 ± 14 µm). 192
Coincident with crypt lengthening, numbers of cells staining for BrdU, P-Akt and P-β-193
catenin552 increased 3-5 fold within the first week, peaked by day 14 and declined 194
thereafter (Fig. 1C-E). Examination of timed sections in the first two weeks suggested 195
C. rodentium induced IEC proliferation within mid and upper crypt regions. By 196
comparison, BrdU-stained cells receded from upper crypts one week later (day 21). 197
Likewise, P-Akt and P-β-catenin552 staining reached apical crypt regions by day 14 198
before diminishing. Examination of alcian blue staining indicated that numbers of goblet 199
cells decreased during the first two weeks of infection (Fig. 1F) as previously 200
described(5, 34). To test whether the increase in BrdU and PI3K/β-catenin signaling 201
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

10
was simply an effect of increased crypt length, numbers of positive cells were calculated 202
based on average crypt lengths. Increased crypt lengths in inflamed mice reduced the 203
absolute differences in counts but data remained consistent with total crypt counts (data 204
not shown). These results raised the possibility that IEC proliferative responses to C. 205
rodentium were induced by PI3K and β-catenin signaling in mid and upper crypts. 206
PI3K signaling is required for P-β-catenin552 expression and crypt proliferation in 207
C. rodentium-induced colitis 208
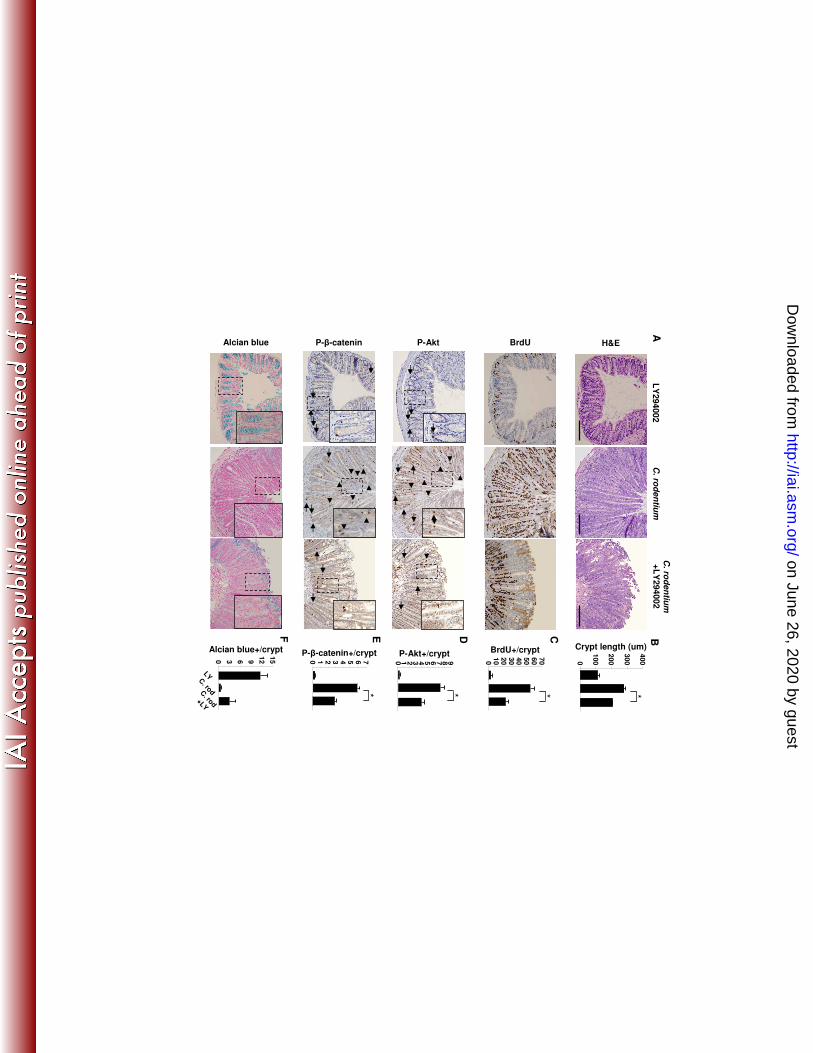
To examine the role of PI3K signaling in epithelial responses to C. rodentium 209
infection, infected mice were treated with vehicle (DMSO) or the selective PI3K inhibitor, 210
LY294002, during the first two weeks of infection (Fig. 2A). A third group of uninfected 211
mice were treated with an identical dosing schedule of LY294002 over two weeks to 212
assess the effect of PI3K inhibition on normal IEC turnover. Morphometric analysis data 213
(Fig. 2B) indicate that crypt lengths were unaffected by PI3K blockade in uninfected 214
mice. In contrast, PI3K inhibition reduced crypt elongation by 36% in infected mice. IHC 215
results show that PI3K inhibition had no effect on the distribution of staining in 216
uninfected mice, but attenuated the vertical expansion of BrdU, P-Akt and P-β-217
catenin552 staining into upper crypts of C. rodentium infected mice. Cell counting of 218
BrdU, P-Akt and P-β-catenin552 stained sections demonstrated PI3K inhibition reduced 219
numbers of BrdU+ cells by 67% (Fig. 2C) and active Akt and β-catenin signaling by 50% 220
and 42% respectively (Fig. 2D,E). Additionally, we observed that reductions in goblet 221
cell numbers in C. rodentium–infected mice were partially reversed as alcian blue 222
stained cells reappeared in the upper crypt of LY294002-treated mice (Fig. 2F). To 223
confirm that changes in PI3K signaling measured per crypt were solely due to impaired 224
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

11
proliferation (i.e. fewer numbers of IEC), BrdU, P-Akt, and P-β-catenin552 positive cells 225
were enumerated on a per IEC basis. Results were similar to crypt counts revealing 226
PI3K inhibition reduced the absolute number of proliferating epithelial cells with β-227
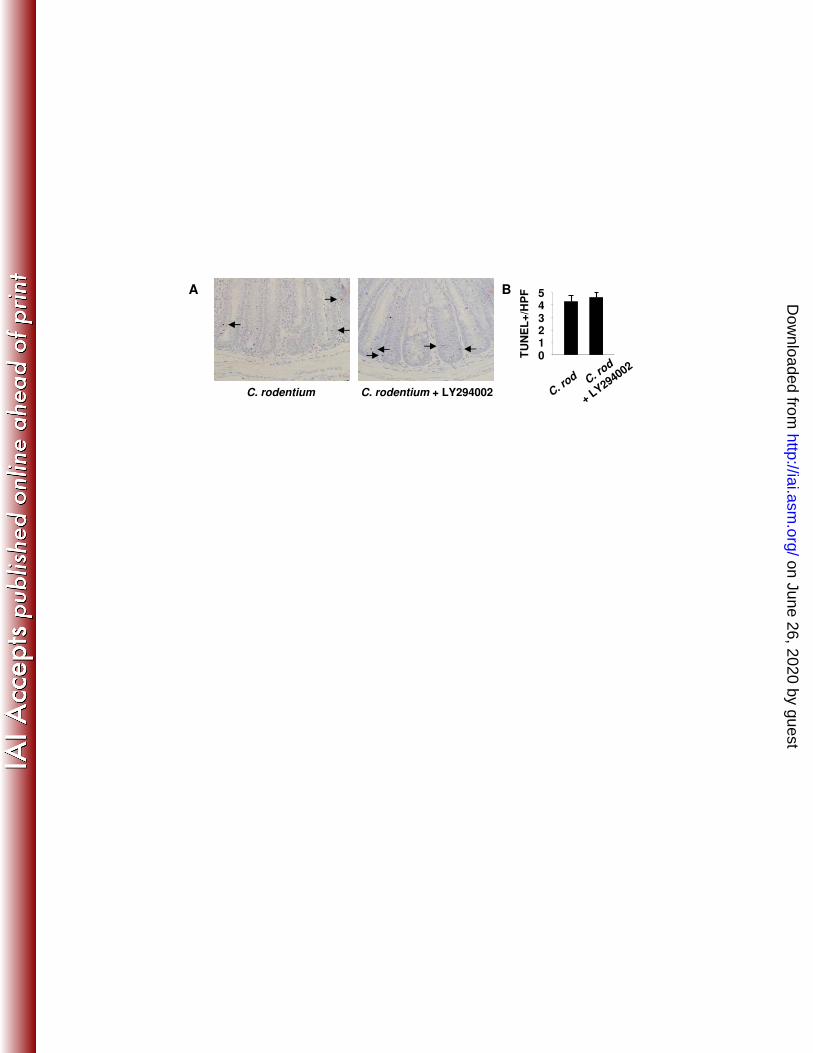
catenin signaling (data not shown). Finally, to test whether the PI3K-inhibition affected 228
levels of IEC apoptosis, sections were TUNEL stained. Data showed no difference in 229
apoptotic indices after LY294002 treatment (Figure 3). Taken together, these data 230
suggest that PI3K activation induced IEC proliferation via Akt-dependent 231
phosphorylation of β-catenin in C. rodentium colitis altering IEC cell death. The net 232
result was to increase crypt lengths. 233
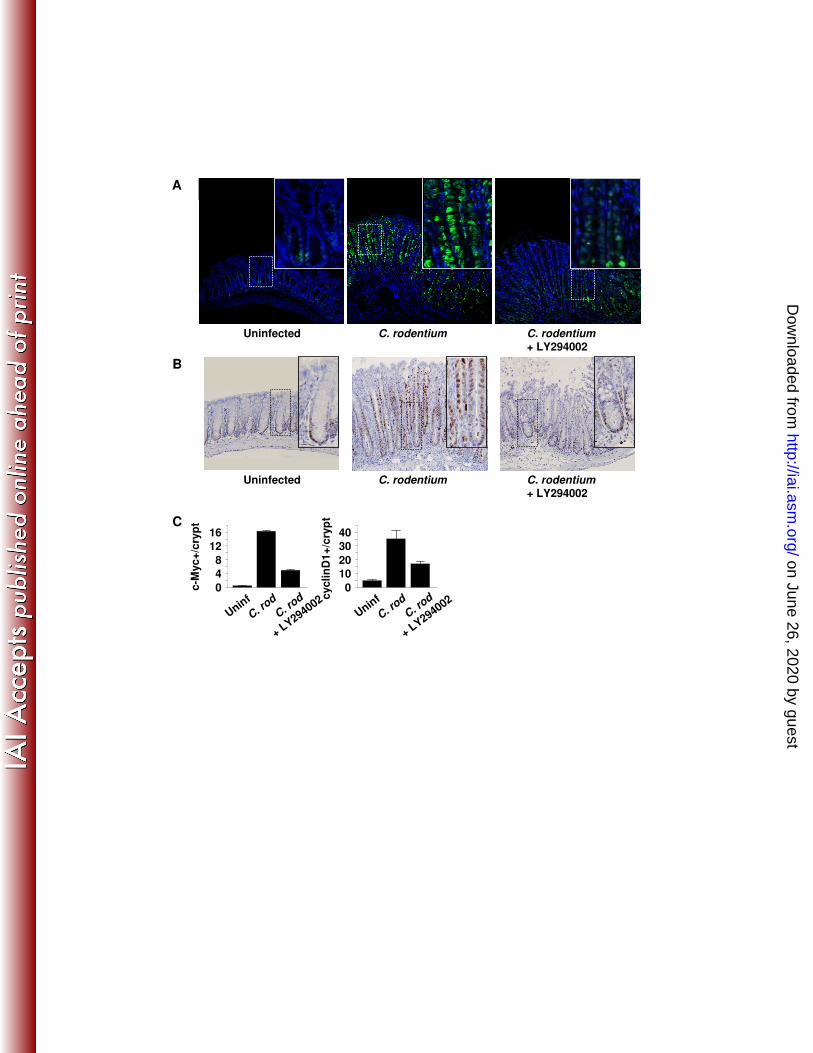
Functional activity of β-catenin is dependent on PI3K-mediated Ser552 234
phosphorylation 235
Our data in both human inflammatory bowel disease and experimental IL-10-/- 236
murine colitis(8, 32) as well as data from Fang et al(18) suggests that Ser552 237
phosphorylation of β-catenin stimulates transcriptional expression of Wnt/β-catenin 238
target genes. c-Myc and cyclinD1 are Wnt/β-catenin target genes involved in cell 239
proliferation(23, 59). In recent studies, Clevers and colleagues reported that c-Myc 240
protein expression denotes proliferative transit amplifying cells in the mid-crypt(39). To 241
assess the functional consequences of PI3K inhibition on epithelial c-Myc and cyclinD1 242
expression, colon sections were stained for c-Myc and cyclinD1 (Fig. 4). In uninfected 243
mice, c-Myc and cyclinD1 stainings localized to IECs immediately above (c-Myc) or 244
within (cyclinD1) the crypt base. In response to infection, both c-Myc and cyclinD1 245
levels increased throughout mid and upper crypts. Data also reveal that LY294002 246
treatment inhibited the expression of both proteins in mid and upper crypts. These 247
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

12
findings suggest C. rodentium infection induced expression of c-Myc and cyclinD1 in 248
transit amplifying cells through a PI3K-dependent process. Together, data suggest that 249
C. rodentium-induced IEC PI3K activation stimulated epithelial proliferation by 250
stimulating Akt and β-catenin signaling. 251
PI3K signaling is required for host defense against C. rodentium 252
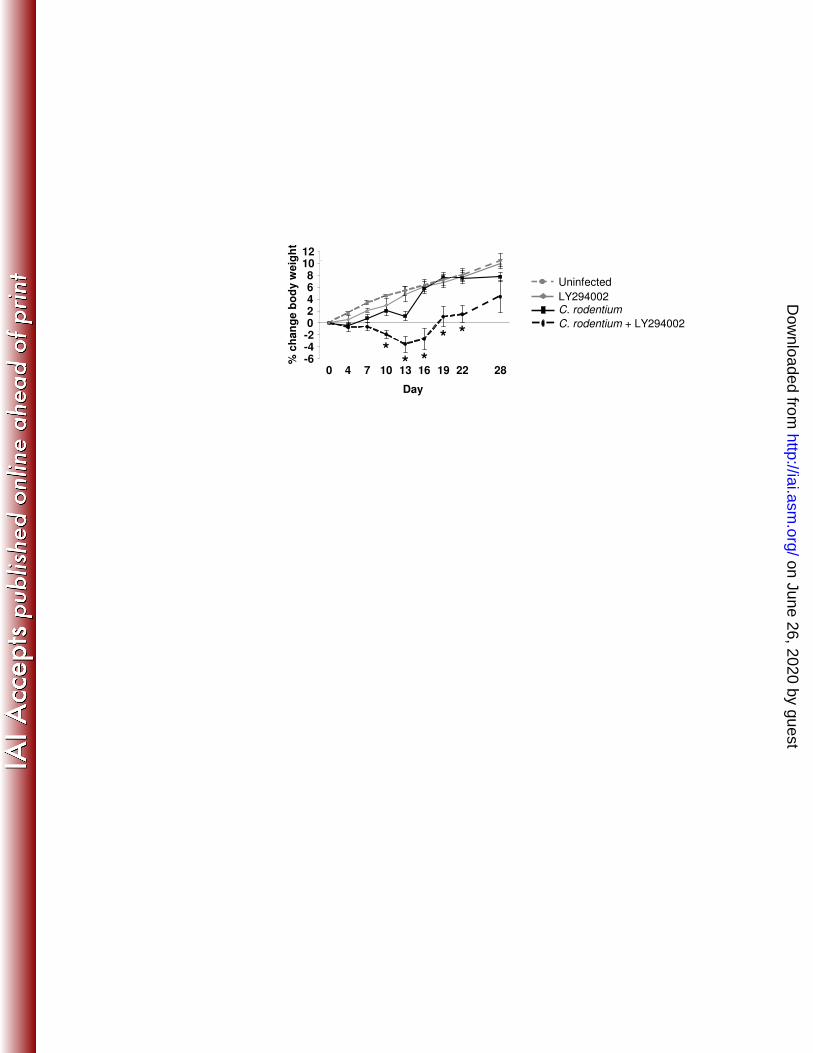
To test the role of PI3K signaling in host responses to C. rodentium, body 253
weights and C. rodentium bacterial loads in control and LY294002-treated mice were 254
measured up to 28 days after infection. Infection caused transient weight loss at day 4 255
(average body weight decreased by 0.5%) followed by weight gain (Fig. 5). Infected 256
mice treated with LY294002 failed to gain weight at day 7, followed by further weight 257
loss that continued until day 16 whereupon a gradual recovery ensued. To determine 258
whether toxicity from LY294002 itself caused weight loss, uninfected WT mice were 259
treated with 3 doses of LY294002 and weights were compared to control infected mice. 260
LY294002-treated uninfected mice gained weight in parallel with untreated mice. We 261
then considered the possibility that weight loss in LY294002-treated infected mice 262
resulted from prolonged bacterial infection. Consistent with this interpretation, we found 263
that DMSO- and LY294002-treated mice had comparable colonic and splenic bacterial 264
burdens at day 7, but LY294002 treatment delayed clearance of colonic bacteria at both 265
day 14 (4-fold more CFU’s) and day 18 (300-fold more CFU’s) (Fig. 6A), which 266
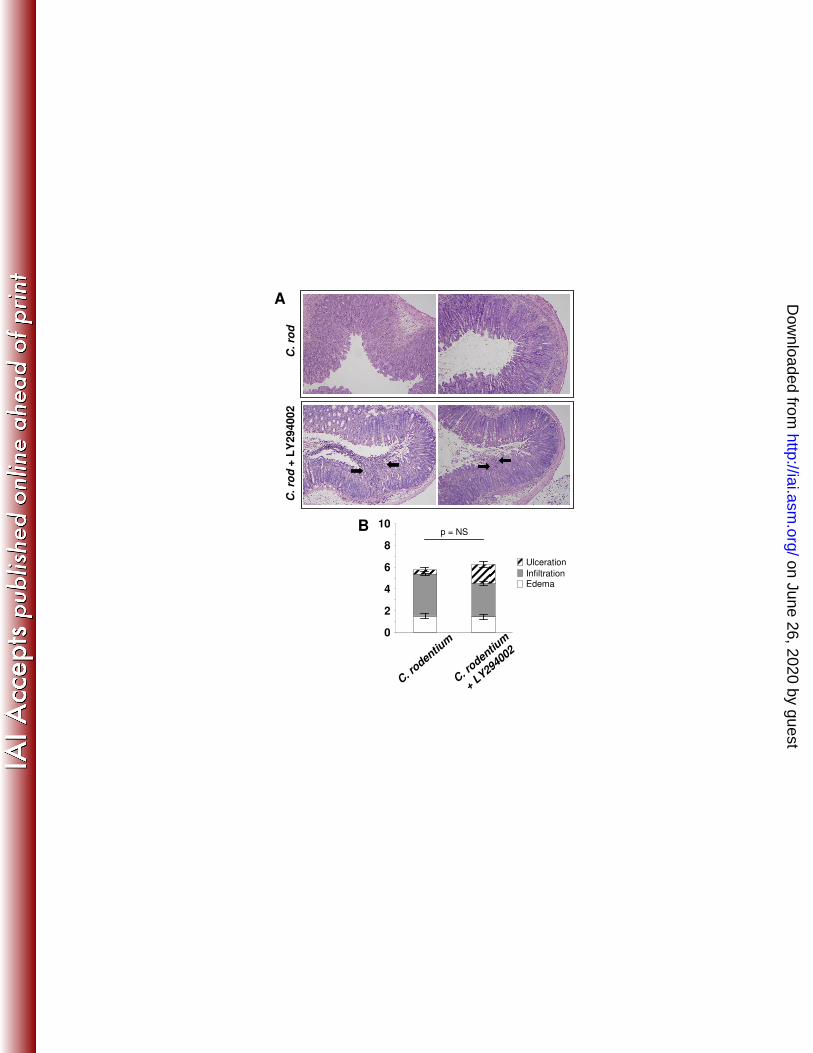
correlated with the weight divergence (day 10) between the two groups. Histologic 267
findings in LY294002-treated mice suggested that attenuated PI3K signaling impaired 268
epithelial restitution (Fig. 2) resulting in more severe, prolonged ulceration and impaired 269
barrier function (Fig. 7). We then tested whether compromised mucosa correlated with 270
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

13
enhanced bacterial translocation. Analysis of splenic C. rodentium levels revealed that 271
infected control mice cleared splenic bacteria by day 18. By comparison, splenic 272
bacterial burdens in LY294002-treated mice remained elevated at days 14 and 18 with 273
clearance at day 21 (Fig. 6B). Together, these data support the hypothesis that 274
epithelial PI3K signaling helps to prevent C. rodentium bacterial invasion. 275
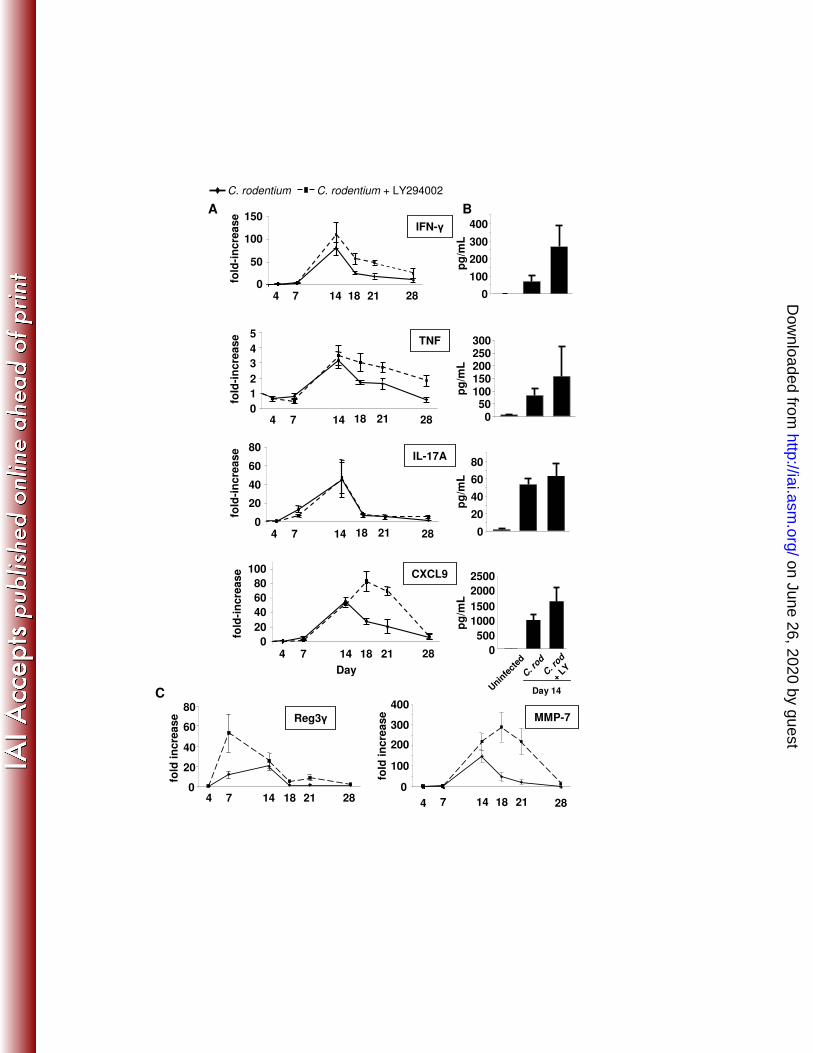
Host adaptive immune responses are not affected by treatment with LY294002 276
Previous studies indicate that cytokine(13, 21, 31, 52, 53, 56, 58) and 277
chemokine(54)-mediated adaptive immune responses promote host defense against C. 278
rodentium. To determine the effect of PI3K inhibition on adaptive mucosal host defense 279
to C. rodentium, mucosal Th1, Th17, and macrophage cytokine levels, as well as 280
chemokine responses were assessed in infected mice. Data in Figure 8A show that 281
IFN-γ, TNF, IL-17, and CXCL9 mRNA levels peaked within fourteen days of C. 282
rodentium infection. Furthermore, treatment with LY294002 during this period did not 283
affect induction of these inflammatory mediators. In fact, cytokine and chemokine 284
mRNA levels for all genes assessed increased and/or remained elevated in LY294002-285
treated mice, including CXCL1, CXCL5, CXCL10, IL1-β, and iNOS (data not shown). 286
Protein quantification by colon explant culture ELISA (Fig. 8B) at day 14 revealed that 287
C. rodentium infection also increased cytokine and chemokine protein levels 288
independent of PI3K. Interestingly, mucosal IFN-γ, TNF, CXCL9 and 10, and iNOS 289
mRNA levels increased after LY294002 treatment was stopped. Examination of tissues 290
during this period revealed that tissue remained inflamed (histologic colitis score 5.8 ± 291
0.2 vs. 6.3 ± 0.3, C. rodentium-infected vs. C. rodentium-infected with LY294002 292
treatment, p=NS, (Fig. 7B). One possibility suggested by assessments of bacterial 293
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

14
counts (Fig. 6) was that mucosal inflammation remained elevated to allow host 294
defenses to recover from PI3K inhibition. To test whether the increased cytokine and 295
chemokine levels seen in LY294002-treated infected mice was due to LY294002, colon 296
mRNA from uninfected mice that received 3 doses of LY294002 was compared to 297
control uninfected mice. Data showed no increase in cytokine/chemokine mRNA levels 298
after LY294002 treatment (data not shown). 299
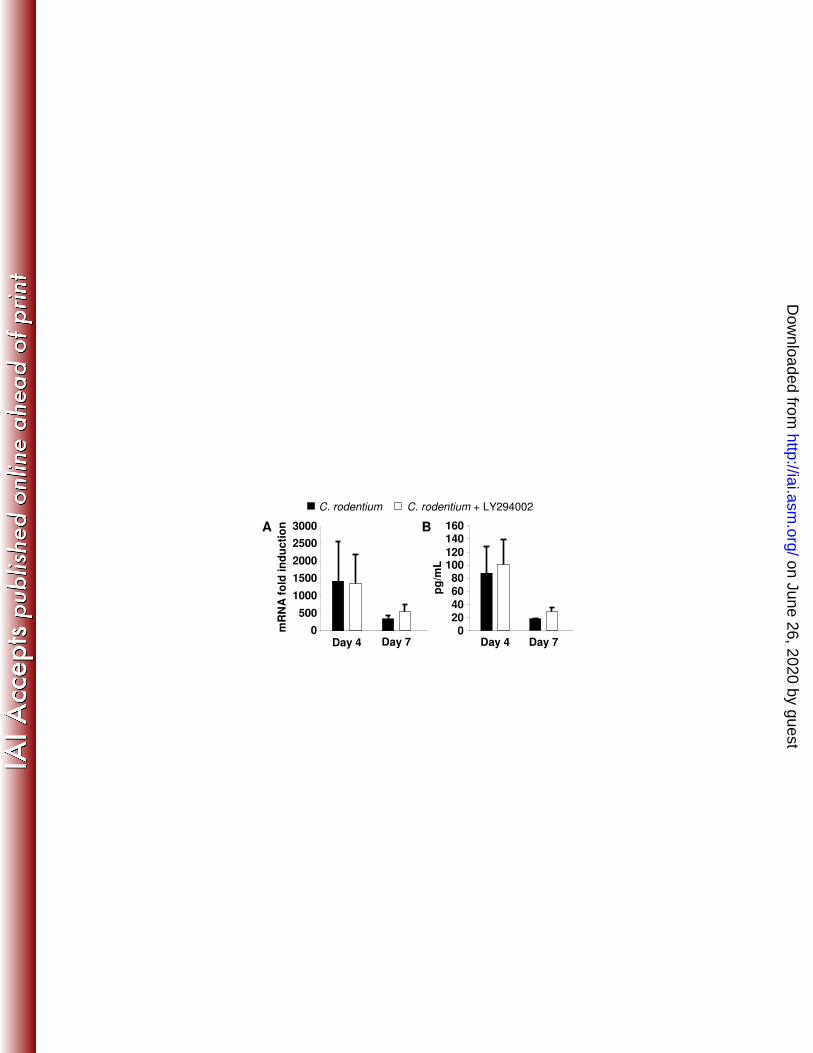
Recently, IL-22 has been implicated as a major determinant during the first week 300
of mounting host defenses to C. rodentium(63). We considered this mechanism of host 301
defense to be relevant because the IL-22 receptor is expressed on epithelial cells(2). 302
Data in Fig. 9 show that at both day 4 and 7 after infection, IL-22 induction was 303
unaffected. 304
Finally, we considered that PI3K inhibition may be affecting innate immune 305
signaling, including antimicrobial function of paneth cells and enterocytes. Data in Fig. 306
8C show that both Reg3γ (Paneth cells) and MMP-7 (enterocytes) induction was 307
unaffected. In fact, early enhanced induction of Reg3γ mRNA (day 7) and prolonged 308
induction of MMP-7 were noted. These data support the notion that PI3K signaling is 309
critical to epithelial host defenses independent of adaptive inflammatory responses. 310
Alternative PI3K inhibition with wortmannin corroborates impaired bacterial 311
clearance with LY294002 312
Although we have demonstrated that PI3K inhibition by LY294002 treatment 313
impairs mucosal host defense by inhibiting epithelial proliferative responses while 314
sparing host inflammatory responses, it remained possible that LY294002 was acting 315
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

15
through a mechanism independent of the epithelial compartment or independent of 316
PI3K. Therefore, we performed a similar set of experiments interrupting PI3K signaling 317
with the irreversible PI3K inhibitor, wortmannin. Morphometric analysis data (Fig. 10A) 318
indicate that crypt lengths were unaffected by PI3K blockade with wortmannin in 319
uninfected mice. In contrast, PI3K inhibition reduced crypt elongation by 43% in infected 320
mice, consistent with the impaired epithelial proliferative responses seen in LY294002-321
treated mice. Furthermore, wortmannin treatment resulted in similar wasting state to 322
LY294002, while having no effect on uninfected mice (Fig. 10B). As expected, colonic 323
bacterial clearance was delayed at day 18, and systemic levels of bacteria were greater 324
at day 14 and remained present ay day 18 (Fig. 10C). To measure whether wortmannin 325
spared host adaptive immune responses in a manner similar PI3K inhibition with 326
LY294002, colonic mRNA levels of IFNγ, CXCL9, and iNOS were measured at day 14 327
and 18. Data in Fig. 10D show that PI3K inhibition did not affect induction of these pro-328
inflammatory genes. This data corroborates findings in LY294002-treated mice and 329
suggests PI3K-mediated signaling plays a critical role in epithelial-specific host 330
responses to enteric infection. 331
DISCUSSION 332
The present study proposes a novel view of the role of PI3K signaling in 333
mediating epithelial responses to C. rodentium. Previous studies suggested that PI3K 334
signaling induces epithelial proliferation in response to growth factor, TNF and toll-like 335
receptor 4 ligation(16, 19, 25, 36). Data from epithelial co-culture studies indicate 336
mucosal T cells from Crohn’s disease tissue stimulate IEC PI3K signaling(11, 12). 337
Results presented here show that IEC PI3K signaling was induced during the period of 338
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

16
C. rodentium-induced inflammation associated with crypt elongation and enhanced IEC 339
proliferation (Fig. 1, 2). Reductions in crypt elongation and IEC proliferation in 340
LY294002-treated mice indicated that PI3K signaling was required for the proliferative 341
responses induced by C. rodentium. Proliferative responses may be pathogen driven to 342
facilitate increased surface area for C. rodentium colonization. However, we found that 343
blocking PI3K signaling/proliferation increased pathogen burden and prolonged mucosal 344
inflammation (Fig.’s 6-8). Thus, our findings are consistent with the notion that C. 345
rodentium-stimulated IEC PI3K signaling induces proliferation as a component of 346
mucosal host defenses needed to eliminate infection and resolve mucosal inflammation. 347
Data from our group(8, 24) and others(18) suggest that PI3K signaling serves to 348
enhance Wnt-mediated β-catenin activation. Studies in PTEN-deficient mice suggested 349
that enhanced PI3K signaling in IEC led to SB polyp formation. The detection of IEC 350
with nuclear P-β-catenin552 at points of crypt fissioning suggested that Akt-mediated 351
phosphorylation of β-catenin enhanced nuclear stabilization and activation. The anti-P-352
β-catenin552 Ab employed in these studies was used here to visualize cells with C-353
terminal phosphorylation of nuclear β-catenin in crypts of mice with C. rodentium 354
infection. Sellin et al previously reported enhanced levels of P-β-catenin552 in C. 355
rodentium colitis(49, 50). Studies here suggested that the induction of P-β-catenin552 in 356
crypts was regulated by PI3K signaling. In addition, the data were consistent with the 357
notion that PI3K mediated β-catenin activation played a key role in mounting robust 358
crypt epithelial proliferative responses during C. rodentium infection. The localization of 359
cells with nuclear P-β-catenin552 in the upper crypts of infected mice suggests that PI3K 360
signaling induced β-catenin activation in PCs within transit amplifying populations. We 361
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

17
postulate that PI3K signaling in upper crypt IEC was induced by inflammatory signals 362
received from the local environment. Of the many downstream functions of PI3K/Akt 363
signaling (protein stabilization, reduced apoptosis, etc), we suspect that Akt interacts in 364
a multi-component complex with β-catenin in the nucleus to enhance activation. In this 365
model, we suspect Akt activation did not promote nuclear localization of β-catenin, 366
which occurs during Wnt signaling. Rather, our results suggest that β-catenin 367
phosphorylation by Akt in the nucleus initiates transcription of pro-proliferative genes 368
(e.g. c-Myc, cyclinD1). This signal cooperates with effects of Akt in the cytosol (e.g. 369
mTOR activation) that promote cell cycle progression and replication. Together, the 370
data suggest that inflammatory signals derived from the surrounding milieu promote β-371
catenin signaling through a PI3K-dependent pathway. 372
The data presented show that PI3K inhibition was associated with increased 373
systemic levels of C. rodentium. Because it is not possible to confirm unperturbed 374
function of all adaptive pro-inflammatory pathways(13, 21, 52-54, 56, 58) and innate 375
host defense factors (e.g. cathelicidin, TFF)(15, 26) or unknown effects of LY294002 376
and wortmannin, it remains possible that systemic PI3K inhibition may have contributed 377
to increased systemic bacterial load. However, preservation of Th1, Th17, 378
macrophage, paneth cell, epithelial, and chemokine mRNA levels suggested that PI3K 379
inhibition did not impair mucosal inflammatory responses. Rather, we postulate that 380
PI3K inhibition reduced epithelial proliferative responses and barrier function. This 381
hypothesis is consistent with results from Podolsky and colleagues who found that 382
inhibition of IEC PI3K/Akt signaling caused disruption of tight junctions following toll-like 383
receptor 2 stimulation(9). Similarly, Resta-Lenert et al found that PI3K inhibition 384
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

18
reversed the protective effect of probiotics on barrier function in epithelial 385
monolayers(44). The findings presented here add to the potential mechanisms for the 386
role of PI3K signaling in epithelial barrier integrity. The data suggest that PI3K signaling 387
may enhance mucosal barrier function in infectious colitis by promoting epithelial 388
restitution. 389
A/E-type enteric pathogens such as Enteropathogenic and Enterohemorrhagic E. 390
coli generate substantial global morbidity (e.g. infantile diarrhea, hemolytic uremic 391
syndrome) and mortality, resulting in as many as 1 million deaths per year world 392
wide(28, 38, 40). Elucidating mechanisms required for host defense against A/E 393
pathogens may impact global health. Data here utilize C. rodentium, an A/E murine 394
pathogen, to show that PI3K-dependent activation of β-catenin and subsequent 395
epithelial proliferative responses are required for normal host-defense in C. rodentium 396
colitis. In the current study, it remains unclear whether or not the eventual clearance of 397
bacteria was due to restitution of PI3K signaling after cessation of LY294002 treatment, 398
or if enhanced inflammatory responses were enough to compensate for impaired 399
epithelial signaling. Regardless, given the global morbidity and mortality associated 400
with A/E enteric pathogens and the fact PI3K signaling may be targeted in future 401
chemotherapeutic agents(35), it is crucial to better understand its role in non-malignant, 402
inflammatory processes. 403
Acknowledgments 404
This work was supported by National Institutes of Health Grants DK54778, DKAI061701 405
(to T.A.B.) and DK066161 (to J.B.B.). 406
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

19
References 407 408 1. 2008. Comprehensive genomic characterization defines human glioblastoma genes and 409
core pathways. Nature 455:1061-8. 410 2. Aujla, S. J., and J. K. Kolls. 2009. IL-22: a critical mediator in mucosal host defense. J 411
Mol Med 87:451-4. 412 3. Bader, A. G., S. Kang, L. Zhao, and P. K. Vogt. 2005. Oncogenic PI3K deregulates 413
transcription and translation. Nat Rev Cancer 5:921-9. 414 4. Barker, N., M. van de Wetering, and H. Clevers. 2008. The intestinal stem cell. Genes 415
Dev 22:1856-64. 416 5. Bergstrom, K. S., J. A. Guttman, M. Rumi, C. Ma, S. Bouzari, M. A. Khan, D. L. 417
Gibson, A. W. Vogl, and B. A. Vallance. 2008. Modulation of intestinal goblet cell 418 function during infection by an attaching and effacing bacterial pathogen. Infect Immun 419 76:796-811. 420
6. Bjerknes, M., and H. Cheng. 1999. Clonal analysis of mouse intestinal epithelial 421 progenitors. Gastroenterology 116:7-14. 422
7. Borenshtein, D., M. E. McBee, and D. B. Schauer. 2008. Utility of the Citrobacter 423 rodentium infection model in laboratory mice. Curr Opin Gastroenterol 24:32-7. 424
8. Brown, J. B., G. Lee, E. Managlia, G. R. Grimm, R. Dirisina, T. Goretsky, P. 425 Cheresh, N. R. Blatner, K. Khazaie, G. Y. Yang, L. Li, and T. A. Barrett. 426 Mesalamine inhibits epithelial beta-catenin activation in chronic ulcerative colitis. 427 Gastroenterology 138:595-605, 605 e1-3. 428
9. Cario, E., G. Gerken, and D. K. Podolsky. 2007. Toll-like receptor 2 controls mucosal 429 inflammation by regulating epithelial barrier function. Gastroenterology 132:1359-74. 430
10. Cully, M., H. You, A. J. Levine, and T. W. Mak. 2006. Beyond PTEN mutations: the 431 PI3K pathway as an integrator of multiple inputs during tumorigenesis. Nat Rev Cancer 432 6:184-92. 433
11. Dahan, S., G. Roda, D. Pinn, F. Roth-Walter, O. Kamalu, A. P. Martin, and L. 434 Mayer. 2008. Epithelial: lamina propria lymphocyte interactions promote epithelial cell 435 differentiation. Gastroenterology 134:192-203. 436
12. Dahan, S., F. Roth-Walter, P. Arnaboldi, S. Agarwal, and L. Mayer. 2007. Epithelia: 437 lymphocyte interactions in the gut. Immunol Rev 215:243-53. 438
13. Dann, S. M., M. E. Spehlmann, D. C. Hammond, M. Iimura, K. Hase, L. J. Choi, E. 439 Hanson, and L. Eckmann. 2008. IL-6-dependent mucosal protection prevents 440 establishment of a microbial niche for attaching/effacing lesion-forming enteric bacterial 441 pathogens. J Immunol 180:6816-26. 442
14. Ding, L., G. Getz, D. A. Wheeler, E. R. Mardis, M. D. McLellan, K. Cibulskis, C. 443 Sougnez, H. Greulich, D. M. Muzny, M. B. Morgan, L. Fulton, R. S. Fulton, Q. 444 Zhang, M. C. Wendl, M. S. Lawrence, D. E. Larson, K. Chen, D. J. Dooling, A. Sabo, 445 A. C. Hawes, H. Shen, S. N. Jhangiani, L. R. Lewis, O. Hall, Y. Zhu, T. Mathew, Y. 446 Ren, J. Yao, S. E. Scherer, K. Clerc, G. A. Metcalf, B. Ng, A. Milosavljevic, M. L. 447 Gonzalez-Garay, J. R. Osborne, R. Meyer, X. Shi, Y. Tang, D. C. Koboldt, L. Lin, R. 448 Abbott, T. L. Miner, C. Pohl, G. Fewell, C. Haipek, H. Schmidt, B. H. Dunford-449 Shore, A. Kraja, S. D. Crosby, C. S. Sawyer, T. Vickery, S. Sander, J. Robinson, W. 450 Winckler, J. Baldwin, L. R. Chirieac, A. Dutt, T. Fennell, M. Hanna, B. E. Johnson, 451 R. C. Onofrio, R. K. Thomas, G. Tonon, B. A. Weir, X. Zhao, L. Ziaugra, M. C. 452
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

20
Zody, T. Giordano, M. B. Orringer, J. A. Roth, M. R. Spitz, Wistuba, II, B. 453 Ozenberger, P. J. Good, A. C. Chang, D. G. Beer, M. A. Watson, M. Ladanyi, S. 454 Broderick, A. Yoshizawa, W. D. Travis, W. Pao, M. A. Province, G. M. Weinstock, 455 H. E. Varmus, S. B. Gabriel, E. S. Lander, R. A. Gibbs, M. Meyerson, and R. K. 456 Wilson. 2008. Somatic mutations affect key pathways in lung adenocarcinoma. Nature 457 455:1069-75. 458
15. Durual, S., C. Blanchard, M. Estienne, M. F. Jacquier, J. C. Cuber, V. Perrot, and C. 459 Laboisse. 2005. Expression of human TFF3 in relation to growth of HT-29 cell 460 subpopulations: involvement of PI3-K but not STAT6. Differentiation 73:36-44. 461
16. Edelblum, K. L., J. A. Goettel, T. Koyama, S. J. McElroy, F. Yan, and D. B. Polk. 462 2008. TNFR1 promotes tumor necrosis factor-mediated mouse colon epithelial cell 463 survival through RAF activation of NF-kappaB. J Biol Chem 283:29485-94. 464
17. Engelman, J. A., J. Luo, and L. C. Cantley. 2006. The evolution of 465 phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat Rev Genet 466 7:606-19. 467
18. Fang, D., D. Hawke, Y. Zheng, Y. Xia, J. Meisenhelder, H. Nika, G. B. Mills, R. 468 Kobayashi, T. Hunter, and Z. Lu. 2007. Phosphorylation of beta-catenin by AKT 469 promotes beta-catenin transcriptional activity. J Biol Chem 282:11221-9. 470
19. Fukata, M., A. Chen, A. S. Vamadevan, J. Cohen, K. Breglio, S. Krishnareddy, D. 471 Hsu, R. Xu, N. Harpaz, A. J. Dannenberg, K. Subbaramaiah, H. S. Cooper, S. H. 472 Itzkowitz, and M. T. Abreu. 2007. Toll-like receptor-4 promotes the development of 473 colitis-associated colorectal tumors. Gastroenterology 133:1869-81. 474
20. Ghosh, P. M., S. Malik, R. Bedolla, and J. I. Kreisberg. 2003. Akt in prostate cancer: 475 possible role in androgen-independence. Curr Drug Metab 4:487-96. 476
21. Goncalves, N. S., M. Ghaem-Maghami, G. Monteleone, G. Frankel, G. Dougan, D. J. 477 Lewis, C. P. Simmons, and T. T. MacDonald. 2001. Critical role for tumor necrosis 478 factor alpha in controlling the number of lumenal pathogenic bacteria and 479 immunopathology in infectious colitis. Infect Immun 69:6651-9. 480
22. Gregorieff, A., D. Pinto, H. Begthel, O. Destree, M. Kielman, and H. Clevers. 2005. 481 Expression pattern of Wnt signaling components in the adult intestine. Gastroenterology 482 129:626-38. 483
23. He, T. C., A. B. Sparks, C. Rago, H. Hermeking, L. Zawel, L. T. da Costa, P. J. 484 Morin, B. Vogelstein, and K. W. Kinzler. 1998. Identification of c-MYC as a target of 485 the APC pathway. Science 281:1509-12. 486
24. He, X. C., T. Yin, J. C. Grindley, Q. Tian, T. Sato, W. A. Tao, R. Dirisina, K. S. 487 Porter-Westpfahl, M. Hembree, T. Johnson, L. M. Wiedemann, T. A. Barrett, L. 488 Hood, H. Wu, and L. Li. 2007. PTEN-deficient intestinal stem cells initiate intestinal 489 polyposis. Nat Genet 39:189-98. 490
25. Hynes, N. E., and G. MacDonald. 2009. ErbB receptors and signaling pathways in 491 cancer. Curr Opin Cell Biol 21:177-84. 492
26. Iimura, M., R. L. Gallo, K. Hase, Y. Miyamoto, L. Eckmann, and M. F. Kagnoff. 493 2005. Cathelicidin mediates innate intestinal defense against colonization with epithelial 494 adherent bacterial pathogens. J Immunol 174:4901-7. 495
27. Jia, S., Z. Liu, S. Zhang, P. Liu, L. Zhang, S. H. Lee, J. Zhang, S. Signoretti, M. 496 Loda, T. M. Roberts, and J. J. Zhao. 2008. Essential roles of PI(3)K-p110beta in cell 497 growth, metabolism and tumorigenesis. Nature 454:776-9. 498
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

21
28. Karch, H., P. I. Tarr, and M. Bielaszewska. 2005. Enterohaemorrhagic Escherichia 499 coli in human medicine. Int J Med Microbiol 295:405-18. 500
29. Kikuchi, A., S. Kishida, and H. Yamamoto. 2006. Regulation of Wnt signaling by 501 protein-protein interaction and post-translational modifications. Exp Mol Med 38:1-10. 502
30. Kong, D., and T. Yamori. 2008. Phosphatidylinositol 3-kinase inhibitors: promising 503 drug candidates for cancer therapy. Cancer Sci 99:1734-40. 504
31. Lebeis, S. L., K. R. Powell, D. Merlin, M. A. Sherman, and D. Kalman. 2009. 505 Interleukin-1 receptor signaling protects mice from lethal intestinal damage caused by the 506 attaching and effacing pathogen Citrobacter rodentium. Infect Immun 77:604-14. 507
32. Lee, G., T. Goretsky, E. Managlia, R. Dirisina, A. P. Singh, J. B. Brown, R. May, G. 508 Y. Yang, J. W. Ragheb, B. M. Evers, C. R. Weber, J. R. Turner, X. C. He, R. B. 509 Katzman, L. Li, and T. A. Barrett. Phosphoinositide 3-Kinase Signaling Mediates beta-510 Catenin Activation in Intestinal Epithelial Stem and Progenitor Cells in Colitis. 511 Gastroenterology 139:869-881 e9. 512
33. Lee, Y. C., H. H. Lin, C. H. Hsu, C. J. Wang, T. A. Chiang, and J. H. Chen. 513 Inhibitory effects of andrographolide on migration and invasion in human non-small cell 514 lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway. Eur J 515 Pharmacol 632:23-32. 516
34. Linden, S. K., T. H. Florin, and M. A. McGuckin. 2008. Mucin dynamics in intestinal 517 bacterial infection. PLoS One 3:e3952. 518
35. Liu, P., H. Cheng, T. M. Roberts, and J. J. Zhao. 2009. Targeting the phosphoinositide 519 3-kinase pathway in cancer. Nat Rev Drug Discov 8:627-44. 520
36. Manning, B. D., and L. C. Cantley. 2007. AKT/PKB signaling: navigating downstream. 521 Cell 129:1261-74. 522
37. Marshman, E., C. Booth, and C. S. Potten. 2002. The intestinal epithelial stem cell. 523 Bioessays 24:91-8. 524
38. Mead, P. S., L. Slutsker, V. Dietz, L. F. McCaig, J. S. Bresee, C. Shapiro, P. M. 525 Griffin, and R. V. Tauxe. 1999. Food-related illness and death in the United States. 526 Emerg Infect Dis 5:607-25. 527
39. Muncan, V., O. J. Sansom, L. Tertoolen, T. J. Phesse, H. Begthel, E. Sancho, A. M. 528 Cole, A. Gregorieff, I. M. de Alboran, H. Clevers, and A. R. Clarke. 2006. Rapid loss 529 of intestinal crypts upon conditional deletion of the Wnt/Tcf-4 target gene c-Myc. Mol 530 Cell Biol 26:8418-26. 531
40. Noel, J. M., and E. C. Boedeker. 1997. Enterohemorrhagic Escherichia coli: a family of 532 emerging pathogens. Dig Dis 15:67-91. 533
41. Parsons, D. W., S. Jones, X. Zhang, J. C. Lin, R. J. Leary, P. Angenendt, P. Mankoo, 534 H. Carter, I. M. Siu, G. L. Gallia, A. Olivi, R. McLendon, B. A. Rasheed, S. Keir, T. 535 Nikolskaya, Y. Nikolsky, D. A. Busam, H. Tekleab, L. A. Diaz, Jr., J. Hartigan, D. R. 536 Smith, R. L. Strausberg, S. K. Marie, S. M. Shinjo, H. Yan, G. J. Riggins, D. D. 537 Bigner, R. Karchin, N. Papadopoulos, G. Parmigiani, B. Vogelstein, V. E. 538 Velculescu, and K. W. Kinzler. 2008. An integrated genomic analysis of human 539 glioblastoma multiforme. Science 321:1807-12. 540
42. Persad, S., A. A. Troussard, T. R. McPhee, D. J. Mulholland, and S. Dedhar. 2001. 541 Tumor suppressor PTEN inhibits nuclear accumulation of beta-catenin and T 542 cell/lymphoid enhancer factor 1-mediated transcriptional activation. J Cell Biol 543 153:1161-74. 544
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

22
43. Pinto, D., and H. Clevers. 2005. Wnt, stem cells and cancer in the intestine. Biol Cell 545 97:185-96. 546
44. Resta-Lenert, S., and K. E. Barrett. 2006. Probiotics and commensals reverse TNF-547 alpha- and IFN-gamma-induced dysfunction in human intestinal epithelial cells. 548 Gastroenterology 130:731-46. 549
45. Samuels, Y., L. A. Diaz, Jr., O. Schmidt-Kittler, J. M. Cummins, L. Delong, I. 550 Cheong, C. Rago, D. L. Huso, C. Lengauer, K. W. Kinzler, B. Vogelstein, and V. E. 551 Velculescu. 2005. Mutant PIK3CA promotes cell growth and invasion of human cancer 552 cells. Cancer Cell 7:561-73. 553
46. Samuels, Y., Z. Wang, A. Bardelli, N. Silliman, J. Ptak, S. Szabo, H. Yan, A. Gazdar, 554 S. M. Powell, G. J. Riggins, J. K. Willson, S. Markowitz, K. W. Kinzler, B. 555 Vogelstein, and V. E. Velculescu. 2004. High frequency of mutations of the PIK3CA 556 gene in human cancers. Science 304:554. 557
47. Sancho, E., E. Batlle, and H. Clevers. 2003. Live and let die in the intestinal epithelium. 558 Curr Opin Cell Biol 15:763-70. 559
48. Schauer, D. B., and S. Falkow. 1993. The eae gene of Citrobacter freundii biotype 4280 560 is necessary for colonization in transmissible murine colonic hyperplasia. Infect Immun 561 61:4654-61. 562
49. Sellin, J. H., S. Umar, J. Xiao, and A. P. Morris. 2001. Increased beta-catenin 563 expression and nuclear translocation accompany cellular hyperproliferation in vivo. 564 Cancer Res 61:2899-906. 565
50. Sellin, J. H., Y. Wang, P. Singh, and S. Umar. 2009. beta-Catenin stabilization imparts 566 crypt progenitor phenotype to hyperproliferating colonic epithelia. Exp Cell Res 315:97-567 109. 568
51. Shen-Tu, G., D. B. Schauer, N. L. Jones, and P. M. Sherman. Detergent-resistant 569 microdomains mediate activation of host cell signaling in response to attaching-effacing 570 bacteria. Lab Invest 90:266-81. 571
52. Simmons, C. P., S. Clare, M. Ghaem-Maghami, T. K. Uren, J. Rankin, A. Huett, R. 572 Goldin, D. J. Lewis, T. T. MacDonald, R. A. Strugnell, G. Frankel, and G. Dougan. 573 2003. Central role for B lymphocytes and CD4+ T cells in immunity to infection by the 574 attaching and effacing pathogen Citrobacter rodentium. Infect Immun 71:5077-86. 575
53. Simmons, C. P., N. S. Goncalves, M. Ghaem-Maghami, M. Bajaj-Elliott, S. Clare, B. 576 Neves, G. Frankel, G. Dougan, and T. T. MacDonald. 2002. Impaired resistance and 577 enhanced pathology during infection with a noninvasive, attaching-effacing enteric 578 bacterial pathogen, Citrobacter rodentium, in mice lacking IL-12 or IFN-gamma. J 579 Immunol 168:1804-12. 580
54. Spehlmann, M. E., S. M. Dann, P. Hruz, E. Hanson, D. F. McCole, and L. Eckmann. 581 2009. CXCR2-dependent mucosal neutrophil influx protects against colitis-associated 582 diarrhea caused by an attaching/effacing lesion-forming bacterial pathogen. J Immunol 583 183:3332-43. 584
55. Staal, F. J., M. Noort Mv, G. J. Strous, and H. C. Clevers. 2002. Wnt signals are 585 transmitted through N-terminally dephosphorylated beta-catenin. EMBO Rep 3:63-8. 586
56. Symonds, E. L., C. U. Riedel, D. O'Mahony, S. Lapthorne, L. O'Mahony, and F. 587 Shanahan. 2009. Involvement of T helper type 17 and regulatory T cell activity in 588 Citrobacter rodentium invasion and inflammatory damage. Clin Exp Immunol 157:148-589 54. 590
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

23
57. Thomas, R. K., A. C. Baker, R. M. Debiasi, W. Winckler, T. Laframboise, W. M. 591 Lin, M. Wang, W. Feng, T. Zander, L. MacConaill, J. C. Lee, R. Nicoletti, C. Hatton, 592 M. Goyette, L. Girard, K. Majmudar, L. Ziaugra, K. K. Wong, S. Gabriel, R. 593 Beroukhim, M. Peyton, J. Barretina, A. Dutt, C. Emery, H. Greulich, K. Shah, H. 594 Sasaki, A. Gazdar, J. Minna, S. A. Armstrong, I. K. Mellinghoff, F. S. Hodi, G. 595 Dranoff, P. S. Mischel, T. F. Cloughesy, S. F. Nelson, L. M. Liau, K. Mertz, M. A. 596 Rubin, H. Moch, M. Loda, W. Catalona, J. Fletcher, S. Signoretti, F. Kaye, K. C. 597 Anderson, G. D. Demetri, R. Dummer, S. Wagner, M. Herlyn, W. R. Sellers, M. 598 Meyerson, and L. A. Garraway. 2007. High-throughput oncogene mutation profiling in 599 human cancer. Nat Genet 39:347-51. 600
58. Vallance, B. A., W. Deng, L. A. Knodler, and B. B. Finlay. 2002. Mice lacking T and 601 B lymphocytes develop transient colitis and crypt hyperplasia yet suffer impaired 602 bacterial clearance during Citrobacter rodentium infection. Infect Immun 70:2070-81. 603
59. van de Wetering, M., E. Sancho, C. Verweij, W. de Lau, I. Oving, A. Hurlstone, K. 604 van der Horn, E. Batlle, D. Coudreuse, A. P. Haramis, M. Tjon-Pon-Fong, P. 605 Moerer, M. van den Born, G. Soete, S. Pals, M. Eilers, R. Medema, and H. Clevers. 606 2002. The beta-catenin/TCF-4 complex imposes a crypt progenitor phenotype on 607 colorectal cancer cells. Cell 111:241-50. 608
60. van Noort, M., J. Meeldijk, R. van der Zee, O. Destree, and H. Clevers. 2002. Wnt 609 signaling controls the phosphorylation status of beta-catenin. J Biol Chem 277:17901-5. 610
61. Vivanco, I., and C. L. Sawyers. 2002. The phosphatidylinositol 3-Kinase AKT pathway 611 in human cancer. Nat Rev Cancer 2:489-501. 612
62. Wood, L. D., D. W. Parsons, S. Jones, J. Lin, T. Sjoblom, R. J. Leary, D. Shen, S. M. 613 Boca, T. Barber, J. Ptak, N. Silliman, S. Szabo, Z. Dezso, V. Ustyanksky, T. 614 Nikolskaya, Y. Nikolsky, R. Karchin, P. A. Wilson, J. S. Kaminker, Z. Zhang, R. 615 Croshaw, J. Willis, D. Dawson, M. Shipitsin, J. K. Willson, S. Sukumar, K. Polyak, 616 B. H. Park, C. L. Pethiyagoda, P. V. Pant, D. G. Ballinger, A. B. Sparks, J. Hartigan, 617 D. R. Smith, E. Suh, N. Papadopoulos, P. Buckhaults, S. D. Markowitz, G. 618 Parmigiani, K. W. Kinzler, V. E. Velculescu, and B. Vogelstein. 2007. The genomic 619 landscapes of human breast and colorectal cancers. Science 318:1108-13. 620
63. Zheng, Y., P. A. Valdez, D. M. Danilenko, Y. Hu, S. M. Sa, Q. Gong, A. R. Abbas, Z. 621 Modrusan, N. Ghilardi, F. J. de Sauvage, and W. Ouyang. 2008. Interleukin-22 622 mediates early host defense against attaching and effacing bacterial pathogens. Nat Med 623 14:282-9. 624
625 626
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

Figure 1. C. rodentium-induces epithelial hyperplasia and PI3K signaling. Colon tissue from control (Day 0) or mice inoculated with C. rodentium are shown. (A) From left to right are representative photomicrographs of tissue from infected mice at specified time points stained with H&E and for BrdU, P-Akt, P-β-catenin552, and alcian blue. Arrows indicate positively stained cells within crypt bases and arrow heads indicate staining in the mid-upper crypt regions. (B) Infection resulted in crypt lengthening as (C) proliferation (BrdU incorporation) increased 6-fold, as well as increased PI3K signaling determined by enumeration of (D) P-Akt+ epithelial cells/crypt and (E) P-β-catenin552+ IEC/crypt. (F) Enumeration of alcian blue+ cells/crypt demonstrates progressive loss of goblet cells coinciding with increased PI3K signaling to Day 14. Goblet cells re-populate crypts as PI3K signal returns to baseline. Values are mean ± SEM. Mag 200X with 400X inserts. Scale bar = 100 µm. *p ≤ 0.02, #p ≤ 0.01 vs. Day 0. Figure 2. PI3K inhibition impairs C. rodentium-induced Akt and β-catenin signaling and attenuates epithelial proliferation. Distal colon was removed from B6 mice 14 days after oral infection with C. rodentium treated with either DMSO (vehicle control, middle panels) or LY294002 (right panels). Left panels represent uninfected mice treated with LY294002. (A) Colonic tissue stained with H&E and for BrdU, P-Akt, P-β-catenin552, and alcian blue. Arrows indicate positively stained cells within crypt bases and arrow heads indicate staining in the mid-upper crypt regions. (B-F) LY294002 treatment alone had no effect relative to baseline, but impaired C. rodentium-induced crypt lengthening, PI3K signaling, proliferation, and goblet cell depletion. Values are mean ± SEM. Mag 200X with 400X inserts. Scale bar = 100 µm. *p < 0.01. Figure 3. PI3K inhibition does not affect epithelial apoptosis. (A) Distal colon sections from mice infected 14 days earlier with C. rodentium and treated with either DMSO or LY294002 were stained for apoptotic cells by the TUNEL technique (see methods). (B) Enumeration of TUNEL+ epithelial cells per 400X field. Values are mean ± SEM. Figure 4. Inhibition of PI3K signaling reduces β-catenin signaling in C. rodentium colitis. Colon sections from uninfected (day 0) and C. rodentium infected mice treated with DMSO or LY294002 (day 14) were processed and stained to assess (A) c-Myc (green) expression and (B) cyclinD1 (brown) expression. (C) Enumeration of c-Myc and cyclinD1 positive epithelial cells per crypt. Values are mean ± SEM. Mag 200X with 400X inserts. Figure 5. Inhibition of PI3K signaling increases and prolongs weight loss. C. rodentium infection caused transient weight loss at day 4 with recovery by day 7 and subsequent weight gain. Blockade of PI3K signaling worsened weight loss that did not recover until day 19 and was significantly reduced until day 28. *p < 0.01 vs. C. rodentium. (n=4-8 mice for each group). Figure 6. PI3K signaling is required for clearance of C. rodentium. In vivo colonization and clearance dynamics were monitored from day 7 after inoculation with C. rodentium to day 28. (A) Mice receiving LY294002 showed prolonged colonization within the colon and shedding of bacteria in the stool demonstrating PI3K inhibition reduces the ability to clear C. rodentium infection. (B) PI3K inhibition resulted in persistence of systemic infection within the spleen. Arrows indicate time points for LY294002 injection. *p < 0.05 vs. C. rodentium. (n=3-4 mice for each group, representative of 3 separate experiments). Figure 7. Interruption of PI3K signaling disrupts the epithelial barrier. (A) Top panels show two separate representative distal colons stained with H&E demonstrating expected crypt hyperplasia with relatively intact epithelial lining 14 days after infection with C. rodentium. Bottom panels show two separate representative distal colons in mice infected with C.
rodentium and treated with LY294002. PI3K inhibition of host epithelial responses resulted in
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

increased disruption of the epithelial barrier and frank ulceration. Arrows indicate ulcer edges. Mag 100X. (B) Histologic scores for ulceration, inflammatory cell infiltration, and edema were assessed in colonic tissue from infected mice treated with DMSO or LY294002. Figure 8. PI3K is dispensable for adaptive and innate immune responses against C. rodentium. (A) Time course of mRNA (adaptive immunity) isolated from distal colon after C. rodentium infection alone, or with LY294002 treatment. (B) Distal colon segments were removed 14 days after infection, with and without LY294002 treatment, washed, and cultured overnight for direct cytokine analysis by ELISA corresponding to 8A mRNA. (C) Time course of mRNA (innate immunity) isolated from distal colon after C. rodentium infection alone, or with LY294002 treatment. (n=3-4 mice per group). Figure 9. PI3K inhibition does not reduce IL-22 production. PI3K inhibition does not affect either (A) mRNA or (B) colon explant culture ELISA measurement of IL-22 production 4 and 7 days after infection with C. rodentium. Figure 10. PI3K inhibition with wortmannin (WM) impairs host defense to C. rodentium. Distal colon was removed from B6 mice 14 or 18 days after oral infection with C. rodentium treated with either DMSO (vehicle control) or 0.1 mg/kg WM (methods). (A) WM treatment alone had no effect relative to baseline, but impaired C. rodentium-induced crypt lengthening. (B) C.
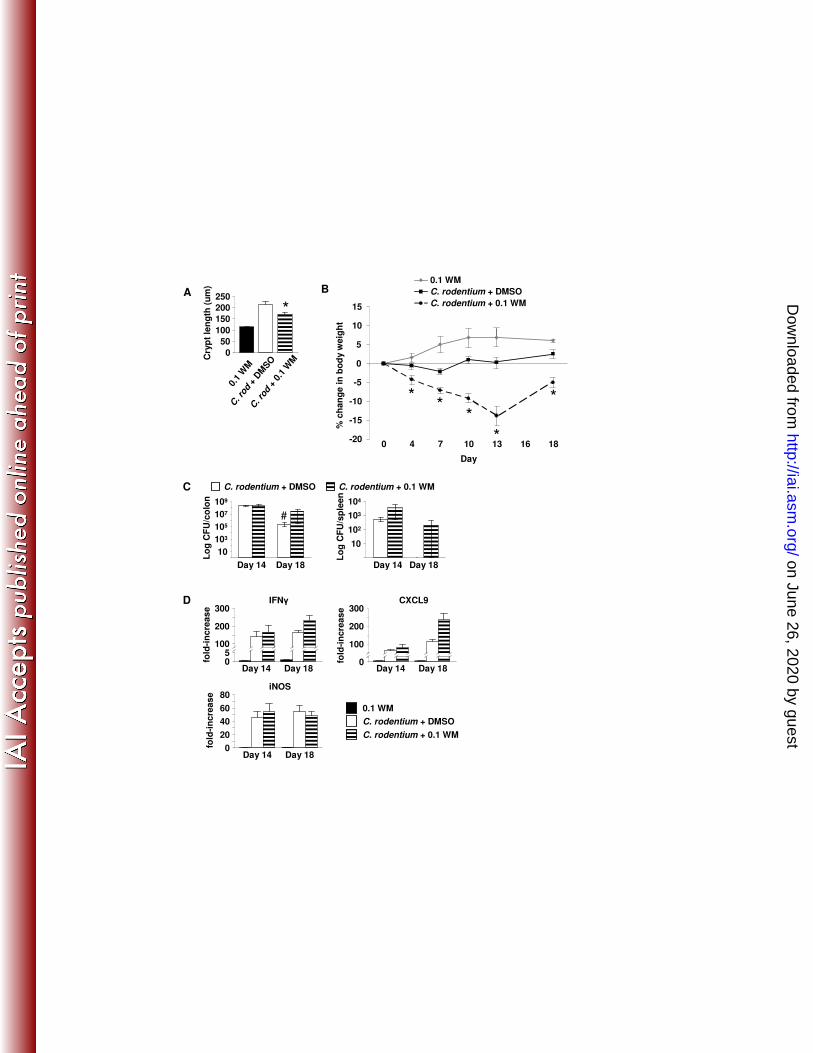
rodentium infection caused transient weight loss with recovery by day 7 and subsequent weight gain. Blockade of PI3K signaling worsened weight loss that did not begin to recover until day 16. (C) Mice receiving WM showed prolonged colonization within the colon and shedding of bacteria in the stool as well as in persistence of systemic infection within the spleen demonstrating PI3K inhibition reduces the ability to clear C. rodentium infection. (D) mRNA levels at day 14 and 18. Values are mean ± SEM. *p < 0.05 vs. C. rodentium + DMSO. #p < 0.01 vs. day 14. (n=5-8 mice per group).
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
Day 0 Day 7 Day 14 Day 21 Day 28
H&
EB
rdU
P-A
kt
P-β
-cate
nin
A
Alc
ian
blu
e
0 7 1421 28
Day
0
100
200
300
Cry
pt
len
gth
(u
m)
* *
# #B
0 7 1421 28
Day
0102030405060
Brd
U+
/cry
pt
#
#
##
C
0 7 1421 28
Day
0
1
2
3
4
5
P-β
-cate
nin
+/c
ryp
t
*
#
E
0 7 14 21 28
Day
0
1
2
3
4
5
P-A
kt+
/cry
pt
*
#
D
0 7 1421 28Day
F
Alc
ian
blu
e+
/cry
pt
0
4
8
12
16
20
*#
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
AC
. ro
den
tium
C. r
od
en
tium
+L
Y294002
BDE
C. rodC
. rod+LY
CF
BrdUP-AktP-β-cateninAlcian blue
LY
294002
Alcian blue+/crypt
0 3 6 9
12
15
LY
H&E
Crypt length (um)
0
100
200
300
400
*
010
20
30
40
50
60
70
BrdU+/crypt
*
0 1 2 3 4 5 6 7 8 9
P-Akt+/crypt
*
0 1 2 3 4 5 6 7
P-β-catenin+/crypt
*
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
C. rodentium C. rodentium + LY294002
0
1
2
3
4
5
C. r
od C. r
od
+ LY294002
TU
NE
L+
/HP
FA B
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
Uninfected C. rodentium C. rodentium
+ LY294002
Uninfected C. rodentium C. rodentium
+ LY294002
A
B
C
0
4
8
12
16
c-M
yc+
/cry
pt
0
10
20
30
40
cyclin
D1
+/c
ryp
t
Uninf
C. r
od
C. r
od
+ LY294002
Uninf
C. r
od
C. r
od
+ LY294002
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
-6-4-202468
1012
% c
han
ge b
od
y w
eig
ht
4 7 10 13 16 19 22 28
Day
0**
***
C. rodentium
C. rodentium + LY294002
Uninfected
LY294002
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from

C. rodentium
C. rodentium + LY294002
10
102
103
104
105
7 14 18 21 28
Lo
g C
FU
/sp
leen
0
Day
A
B
1
10102
103
104
105
106
107
108
Lo
g C
FU
/co
lon
1010
109
7 14 18 21 280Day
1
106
*
*
*
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
C. ro
dC
. ro
d +
LY
294002
0
2
4
6
8
10
Ulceration
InfiltrationEdema
C. r
oden
tium
C. r
oden
tium
+ LY294002
A
B p = NS
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
pg
/mL
0
100
200
300
400IFN-γ
0
50
100
150
4 7 14 18 21 28
fold
-in
cre
ase
A B
C. ro
d
+ LY
Unin
fect
ed
C. ro
d
2500
0
500
1000
1500
2000
pg
/mL
0
20
40
60
80p
g/m
L
050
100150200250300
pg
/mL
TNF
fold
-in
cre
ase
0
1
2
3
4
5
4 7 14 18 21 28
IL-17A
fold
-in
cre
ase
0
20
40
60
80
4 7 14 18 21 28
CXCL9
fold
-in
cre
ase
0
20
40
60
80
100
4 7 14 18 21 28
Day 14
Day
C. rodentium C. rodentium + LY294002
0
20
40
60
80
4 7 14 18 21 28
fold
in
cre
ase Reg3γ
0
100
200
300
400
4 7 14 18 21 28
fold
in
cre
ase MMP-7
C
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
C. rodentium C. rodentium + LY294002
02040
6080
100120
140160
Day 4 Day 7
pg
/mL
mR
NA
fo
ld in
du
cti
on
0
500
1000
1500
2000
2500
3000
Day 4 Day 7
A B
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from
0
50
100
150
200
250
Cry
pt
len
gth
(u
m)
C. ro
d +
DM
SO
C. ro
d +
0.1
WM
0.1
WM
A
-20
-15
-10
-5
0
5
10
15
0 4 7 10 13 16 18
0.1 WM
C. rodentium + DMSO
C. rodentium + 0.1 WM
Day
% c
han
ge i
n b
od
y w
eig
ht
B
10
103
105
107
109
Day 14 Day 18
Lo
g C
FU
/co
lon
10
102
103
104
Day 14 Day 18
Lo
g C
FU
/sp
leen
C. rodentium + DMSO C. rodentium + 0.1 WMC
05
100
200
300
Day 14 Day 18
fold
-in
cre
as
e
IFNγ
0.1 WM
C. rodentium + DMSO
C. rodentium + 0.1 WM
0
20
40
60
80
Day 14 Day 18
iNOS
fold
-in
cre
as
e
0
100
200
300
Day 14 Day 18
CXCL9
fold
-in
cre
as
e
D
*
#
*
*
***
on June 26, 2020 by guesthttp://iai.asm
.org/D
ownloaded from


















