Dendritic cells: controllers of adaptive...
1
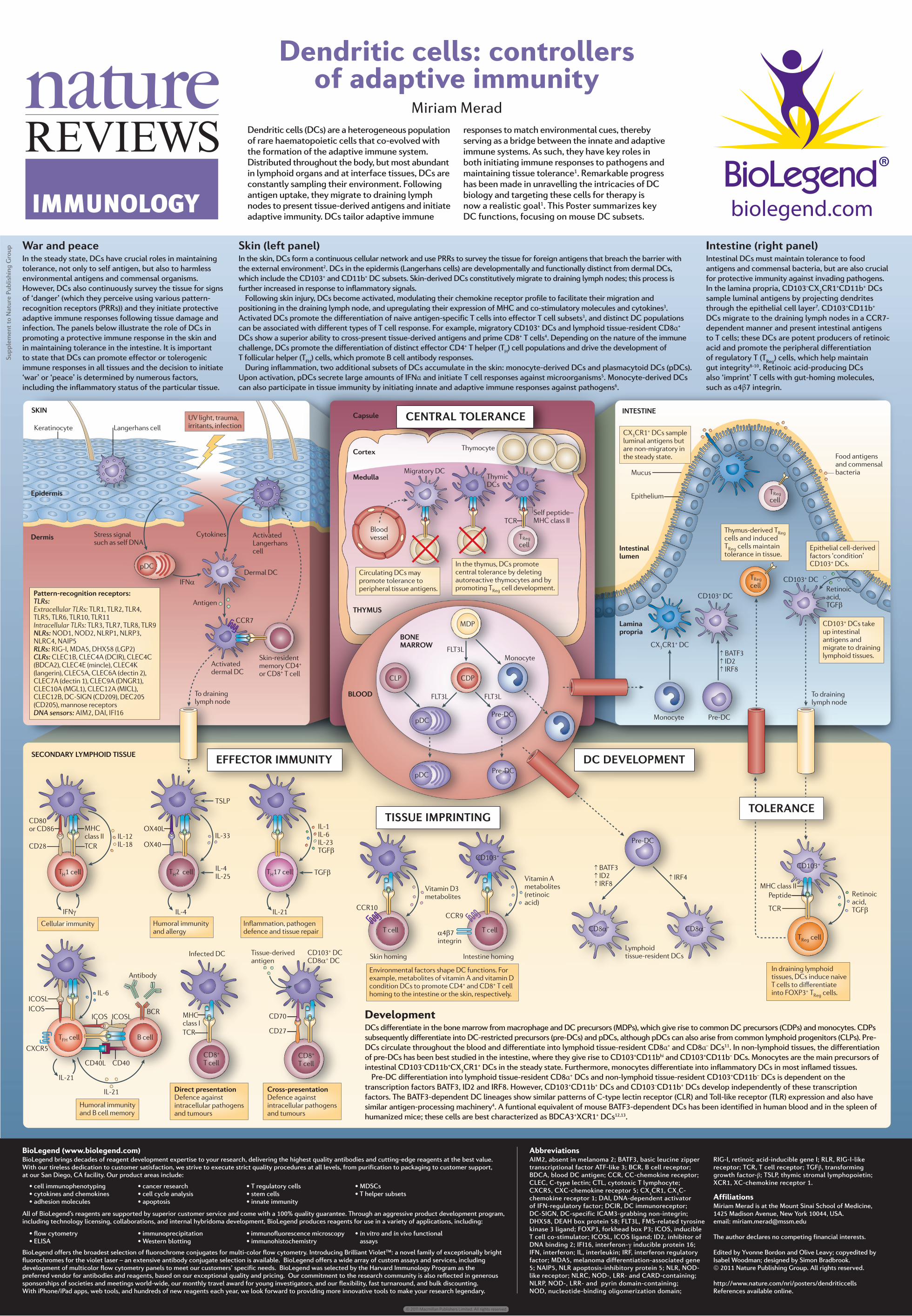
biolegend.com .CPIGTJCPU EGNN #EVKXCVGF .CPIGTJCPU EGNN -GTCVKPQE[VG /KITCVQT[ &% 5GNH RGRVKFGs /*% ENCUU ++ /*% ENCUU ++ 6%4 6%4 6J[OKE &%U 6J[OQE[VG %& &% %: %4 &% %& %& &% 2TG&% 6Q FTCKPKPI N[ORJ PQFG ↑ $#6( ↑ +& ↑ +4( ↑ $#6( ↑ +& ↑ +4( ↑ +4( /&2 6 4GI EGNN 6 4GI EGNN /QPQE[VG /QPQE[VG %&2 %.2 (.6. (.6. (.6. R&% R&% $.11& $10' /#4419 %CRUWNG %QTVGZ /GFWNNC .[ORJQKF VKUUWGTGUKFGPV &%U 2TG&% 2TG&% 2TG&% %& QT %& %& 6%4 /*% ENCUU ++ +. +. +. +. +. 6 * EGNN +(0γ 5'%10 .;/2*1+& 6+557' $NQQF XGUUGN 2CVVGTPTGEQIPKVKQP TGEGRVQTU 6.4U 'ZVTCEGNNWNCT 6.4U 6.4 6.4 6.4 6.4 6.4 6.4 6.4 +PVTCEGNNWNCT 6.4U 6.4 6.4 6.4 6.4 0.4U 01& 01& 0.42 0.42 0.4% 0#+2 4.4U 4+)+ /&# &*: .)2 %.4U %.'%$ %.'%# &%+4 %.'%% $&%# %.'%' OKPENG %.'%- NCPIGTKP %.'%# %.'%# FGEVKP %.'%# FGEVKP %.'%# &0)4 %.'%# /). %.'%# /+%. %.'%$ &%5+)0 %& &'% %& OCPPQUG TGEGRVQTU &0# UGPUQTU #+/ &#+ +(+ >OCN &% #EVKXCVGF FGTOCN &% %%4 5VTGUU UKIPCN UWEJ CU UGNH &0# R&% 5MKPTGUKFGPV OGOQT[ %& QT %& 6 EGNN 6Q FTCKPKPI N[ORJ PQFG #PVKIGP +(0α +P FTCKPKPI N[ORJQKF VKUUWGU &%U KPFWEG PCKXG 6 EGNNU VQ FKȭGTGPVKCVG KPVQ (1:2 6 4GI EGNNU %GNNWNCT KOOWPKV[ 4GVKPQKE CEKF 6)(β 4GVKPQKE CEKF 6)(β 'RKFGTOKU >OKU %[VQMKPGU (QQF CPVKIGPU CPF EQOOGPUCN DCEVGTKC /WEWU 'RKVJGNKWO .COKPC RTQRTKC +06'56+0' 6*;/75 5-+0 +PVGUVKPCN NWOGP 6J[OWUFGTKXGF 6 4GI EGNNU CPF KPFWEGF 6 4GI EGNNU OCKPVCKP VQNGTCPEG KP VKUUWG %: %4 &%U UCORNG NWOKPCN CPVKIGPU DWV CTG PQPOKITCVQT[ KP VJG UVGCF[ UVCVG +P VJG VJ[OWU &%U RTQOQVG EGPVTCN VQNGTCPEG D[ FGNGVKPI CWVQTGCEVKXG VJ[OQE[VGU CPF D[ RTQOQVKPI 6 4GI EGNN FGXGNQROGPV %KTEWNCVKPI &%U OC[ RTQOQVG VQNGTCPEG VQ RGTKRJGTCN VKUUWG CPVKIGPU 'RKVJGNKCN EGNNFGTKXGF HCEVQTU nEQPFKVKQPo %& &%U 6 4GI EGNN 1: 6 * EGNN +. *WOQTCN KOOWPKV[ CPF CNNGTI[ +. +. +. 6)(β 6)(β 6 * EGNN +. +PȯCOOCVKQP RCVJQIGP FGHGPEG CPF VKUUWG TGRCKT %& %&. +. 6%4 $%4 #PVKDQF[ +PHGEVGF &% /*% ENCUU + +. +%15 %:%4 +%15 +%15. +%15. %& %& 6 (* EGNN $ EGNN %& 6 EGNN %& 6 EGNN *WOQTCN KOOWPKV[ CPF $ EGNN OGOQT[ &KTGEV RTGUGPVCVKQP &GHGPEG CICKPUV KPVTCEGNNWNCT RCVJQIGPU CPF VWOQWTU %TQUURTGUGPVCVKQP &GHGPEG CICKPUV KPVTCEGNNWNCT RCVJQIGPU CPF VWOQWTU %& &% %&α &% 6KUUWGFGTKXGF CPVKIGP %&α %&α s 78 NKIJV VTCWOC KTTKVCPVU KPHGEVKQP %& &%U VCMG WR KPVGUVKPCN CPVKIGPU CPF OKITCVG VQ FTCKPKPI N[ORJQKF VKUUWGU 1:. +. 'PXKTQPOGPVCN HCEVQTU UJCRG &% HWPEVKQPU (QT GZCORNG OGVCDQNKVGU QH XKVCOKP # CPF XKVCOKP & EQPFKVKQP &%U VQ RTQOQVG %& CPF %& 6 EGNN JQOKPI VQ VJG KPVGUVKPG QT VJG UMKP TGURGEVKXGN[ 6 EGNN 6 EGNN 5MKP JQOKPI +PVGUVKPG JQOKPI %%4 8KVCOKP & OGVCDQNKVGU 8KVCOKP # OGVCDQNKVGU TGVKPQKE CEKF %%4 αβ KPVGITKP 6 4GI EGNN 2GRVKFG %& 65.2 %'064#. 61.'4#0%' '(('%614 +//70+6; &% &'8'.12/'06 6+557' +/24+06+0) 61.'4#0%' Supplement to Nature Publishing Group BioLegend (www.biolegend.com) BioLegend brings decades of reagent development expertise to your research, delivering the highest quality antibodies and cutting-edge reagents at the best value. With our tireless dedication to customer satisfaction, we strive to execute strict quality procedures at all levels, from purification to packaging to customer support, at our San Diego, CA facility. Our product areas include: All of BioLegend’s reagents are supported by superior customer service and come with a 100% quality guarantee. Through an aggressive product development program, including technology licensing, collaborations, and internal hybridoma development, BioLegend produces reagents for use in a variety of applications, including: BioLegend offers the broadest selection of fluorochrome conjugates for multi-color flow cytometry. Introducing Brilliant Violet™: a novel family of exceptionally bright fluorochromes for the violet laser – an extensive antibody conjugate selection is available. BioLegend offers a wide array of custom assays and services, including development of multicolor flow cytometry panels to meet our customers’ specific needs. BioLegend was selected by the Harvard Immunology Program as the preferred vendor for antibodies and reagents, based on our exceptional quality and pricing. Our commitment to the research community is also reflected in generous sponsorships of societies and meetings world-wide, our monthly travel award for young investigators, and our flexibility, fast turnaround, and bulk discounting. With iPhone/iPad apps, web tools, and hundreds of new reagents each year, we look forward to providing more innovative tools to make your research legendary. • cell immunophenotyping • cytokines and chemokines • adhesion molecules • cancer research • cell cycle analysis • apoptosis • T regulatory cells • stem cells • innate immunity • MDSCs • T helper subsets • flow cytometry • ELISA • immunoprecipitation • Western blotting • immunofluorescence microscopy • immunohistochemistry • in vitro and in vivo functional • assays Development DCs differentiate in the bone marrow from macrophage and DC precursors (MDPs), which give rise to common DC precursors (CDPs) and monocytes. CDPs subsequently differentiate into DC-restricted precursors (pre-DCs) and pDCs, although pDCs can also arise from common lymphoid progenitors (CLPs). Pre- DCs circulate throughout the blood and differentiate into lymphoid tissue-resident CD8α + and CD8α – DCs 11 . In non-lymphoid tissues, the differentiation of pre-DCs has been best studied in the intestine, where they give rise to CD103 + CD11b hi and CD103 + CD11b – DCs. Monocytes are the main precursors of intestinal CD103 – CD11b + CX 3 CR1 + DCs in the steady state. Furthermore, monocytes differentiate into inflammatory DCs in most inflamed tissues. Pre-DC differentiation into lymphoid tissue-resident CD8α + DCs and non-lymphoid tissue-resident CD103 + CD11b – DCs is dependent on the transcription factors BATF3, ID2 and IRF8. However, CD103 + CD11b + DCs and CD103 – CD11b + DCs develop independently of these transcription factors. The BATF3-dependent DC lineages show similar patterns of C-type lectin receptor (CLR) and Toll-like receptor (TLR) expression and also have similar antigen-processing machinery 4 . A funtional equivalent of mouse BATF3-dependent DCs has been identified in human blood and in the spleen of humanized mice; these cells are best characterized as BDCA3 + XCR1 + DCs 12,13 . War and peace In the steady state, DCs have crucial roles in maintaining tolerance, not only to self antigen, but also to harmless environmental antigens and commensal organisms. However, DCs also continuously survey the tissue for signs of ‘danger’ (which they perceive using various pattern- recognition receptors (PRRs)) and they initiate protective adaptive immune responses following tissue damage and infection. The panels below illustrate the role of DCs in promoting a protective immune response in the skin and in maintaining tolerance in the intestine. It is important to state that DCs can promote effector or tolerogenic immune responses in all tissues and the decision to initiate ‘war’ or ‘peace’ is determined by numerous factors, including the inflammatory status of the particular tissue. Skin (left panel) In the skin, DCs form a continuous cellular network and use PRRs to survey the tissue for foreign antigens that breach the barrier with the external environment 2 . DCs in the epidermis (Langerhans cells) are developmentally and functionally distinct from dermal DCs, which include the CD103 + and CD11b + DC subsets. Skin-derived DCs constitutively migrate to draining lymph nodes; this process is further increased in response to inflammatory signals. Following skin injury, DCs become activated, modulating their chemokine receptor profile to facilitate their migration and positioning in the draining lymph node, and upregulating their expression of MHC and co-stimulatory molecules and cytokines 3 . Activated DCs promote the differentiation of naive antigen-specific T cells into effector T cell subsets 3 , and distinct DC populations can be associated with different types of T cell response. For example, migratory CD103 + DCs and lymphoid tissue-resident CD8α + DCs show a superior ability to cross-present tissue-derived antigens and prime CD8 + T cells 4 . Depending on the nature of the immune challenge, DCs promote the differentiation of distinct effector CD4 + T helper (T H ) cell populations and drive the development of T follicular helper (T FH ) cells, which promote B cell antibody responses. During inflammation, two additional subsets of DCs accumulate in the skin: monocyte-derived DCs and plasmacytoid DCs (pDCs). Upon activation, pDCs secrete large amounts of IFNα and initiate T cell responses against microorganisms 5 . Monocyte-derived DCs can also participate in tissue immunity by initiating innate and adaptive immune responses against pathogens 6 . Intestine (right panel) Intestinal DCs must maintain tolerance to food antigens and commensal bacteria, but are also crucial for protective immunity against invading pathogens. In the lamina propria, CD103 – CX 3 CR1 + CD11b + DCs sample luminal antigens by projecting dendrites through the epithelial cell layer 7 . CD103 + CD11b + DCs migrate to the draining lymph nodes in a CCR7- dependent manner and present intestinal antigens to T cells; these DCs are potent producers of retinoic acid and promote the peripheral differentiation of regulatory T (T Reg ) cells, which help maintain gut integrity 8-10 . Retinoic acid-producing DCs also ‘imprint’ T cells with gut-homing molecules, such as α4β7 integrin. Dendritic cells: controllers of adaptive immunity Miriam Merad Dendritic cells (DCs) are a heterogeneous population of rare haematopoietic cells that co-evolved with the formation of the adaptive immune system. Distributed throughout the body, but most abundant in lymphoid organs and at interface tissues, DCs are constantly sampling their environment. Following antigen uptake, they migrate to draining lymph nodes to present tissue-derived antigens and initiate adaptive immunity. DCs tailor adaptive immune responses to match environmental cues, thereby serving as a bridge between the innate and adaptive immune systems. As such, they have key roles in both initiating immune responses to pathogens and maintaining tissue tolerance 1 . Remarkable progress has been made in unravelling the intricacies of DC biology and targeting these cells for therapy is now a realistic goal 1 . This Poster summarizes key DC functions, focusing on mouse DC subsets. Abbreviations AIM2, absent in melanoma 2; BATF3, basic leucine zipper transcriptional factor ATF-like 3; BCR, B cell receptor; BDCA, blood DC antigen; CCR, CC-chemokine receptor; CLEC, C-type lectin; CTL, cytotoxic T lymphocyte; CXCR5, CXC-chemokine receptor 5; CX 3 CR1, CX 3 C- chemokine receptor 1; DAI, DNA-dependent activator of IFN-regulatory factor; DCIR, DC immunoreceptor; DC-SIGN, DC-specific ICAM3-grabbing non-integrin; DHX58, DEAH box protein 58; FLT3L, FMS-related tyrosine kinase 3 ligand; FOXP3, forkhead box P3; ICOS, inducible T cell co-stimulator; ICOSL, ICOS ligand; ID2, inhibitor of DNA binding 2; IFI16, interferon-γ inducible protein 16; IFN, interferon; IL, interleukin; IRF, interferon regulatory factor; MDA5, melanoma differentiation-associated gene 5; NAIP5, NLR apoptosis-inhibitory protein 5; NLR, NOD- like receptor; NLRC, NOD-, LRR- and CARD-containing; NLRP, NOD-, LRR- and pyrin domain-containing; NOD, nucleotide-binding oligomerization domain; RIG-I, retinoic acid-inducible gene I; RLR, RIG-I-like receptor; TCR, T cell receptor; TGFβ, transforming growth factor-β; TSLP, thymic stromal lymphopoietin; XCR1, XC-chemokine receptor 1. Affiliations Miriam Merad is at the Mount Sinai School of Medicine, 1425 Madison Avenue, New York 10044, USA. email: [email protected] The author declares no competing financial interests. Edited by Yvonne Bordon and Olive Leavy; copyedited by Isabel Woodman; designed by Simon Bradbrook. © 2011 Nature Publishing Group. All rights reserved. http://www.nature.com/nri/posters/dendriticcells References available online. IMMUNOLOGY © 2011 Macmillan Publishers Limited. All rights reserved
Transcript of Dendritic cells: controllers of adaptive...
biolegend.com
↑↑↑
↑↑↑
↑
γ
α
β
β
β
β
α
α αα β
Supp
lem
ent
to N
atur
e Pu
blis
hing
Gro
up
BioLegend (www.biolegend.com)BioLegend brings decades of reagent development expertise to your research, delivering the highest quality antibodies and cutting-edge reagents at the best value. With our tireless dedication to customer satisfaction, we strive to execute strict quality procedures at all levels, from purification to packaging to customer support, at our San Diego, CA facility. Our product areas include:
All of BioLegend’s reagents are supported by superior customer service and come with a 100% quality guarantee. Through an aggressive product development program, including technology licensing, collaborations, and internal hybridoma development, BioLegend produces reagents for use in a variety of applications, including:
BioLegend offers the broadest selection of fluorochrome conjugates for multi-color flow cytometry. Introducing Brilliant Violet™: a novel family of exceptionally bright fluorochromes for the violet laser – an extensive antibody conjugate selection is available. BioLegend offers a wide array of custom assays and services, including development of multicolor flow cytometry panels to meet our customers’ specific needs. BioLegend was selected by the Harvard Immunology Program as the preferred vendor for antibodies and reagents, based on our exceptional quality and pricing. Our commitment to the research community is also reflected in generous sponsorships of societies and meetings world-wide, our monthly travel award for young investigators, and our flexibility, fast turnaround, and bulk discounting. With iPhone/iPad apps, web tools, and hundreds of new reagents each year, we look forward to providing more innovative tools to make your research legendary.
• cell immunophenotyping• cytokines and chemokines• adhesion molecules
• cancer research• cell cycle analysis• apoptosis
• T regulatory cells• stem cells• innate immunity
• MDSCs• T helper subsets
• flow cytometry• ELISA
• immunoprecipitation• Western blotting
• immunofluorescence microscopy• immunohistochemistry
• in vitro and in vivo functional • assays
DevelopmentDCs differentiate in the bone marrow from macrophage and DC precursors (MDPs), which give rise to common DC precursors (CDPs) and monocytes. CDPs subsequently differentiate into DC-restricted precursors (pre-DCs) and pDCs, although pDCs can also arise from common lymphoid progenitors (CLPs). Pre-DCs circulate throughout the blood and differentiate into lymphoid tissue-resident CD8α+ and CD8α– DCs11. In non-lymphoid tissues, the differentiation of pre-DCs has been best studied in the intestine, where they give rise to CD103+CD11bhi and CD103+CD11b– DCs. Monocytes are the main precursors of intestinal CD103–CD11b+CX3CR1+ DCs in the steady state. Furthermore, monocytes differentiate into inflammatory DCs in most inflamed tissues.
Pre-DC differentiation into lymphoid tissue-resident CD8α+ DCs and non-lymphoid tissue-resident CD103+CD11b– DCs is dependent on the transcription factors BATF3, ID2 and IRF8. However, CD103+CD11b+ DCs and CD103–CD11b+ DCs develop independently of these transcription factors. The BATF3-dependent DC lineages show similar patterns of C-type lectin receptor (CLR) and Toll-like receptor (TLR) expression and also have similar antigen-processing machinery4. A funtional equivalent of mouse BATF3-dependent DCs has been identified in human blood and in the spleen of humanized mice; these cells are best characterized as BDCA3+XCR1+ DCs12,13.
War and peace In the steady state, DCs have crucial roles in maintaining tolerance, not only to self antigen, but also to harmless environmental antigens and commensal organisms. However, DCs also continuously survey the tissue for signs of ‘danger’ (which they perceive using various pattern-recognition receptors (PRRs)) and they initiate protective adaptive immune responses following tissue damage and infection. The panels below illustrate the role of DCs in promoting a protective immune response in the skin and in maintaining tolerance in the intestine. It is important to state that DCs can promote effector or tolerogenic immune responses in all tissues and the decision to initiate ‘war’ or ‘peace’ is determined by numerous factors, including the inflammatory status of the particular tissue.
Skin (left panel)In the skin, DCs form a continuous cellular network and use PRRs to survey the tissue for foreign antigens that breach the barrier with the external environment2. DCs in the epidermis (Langerhans cells) are developmentally and functionally distinct from dermal DCs, which include the CD103+ and CD11b+ DC subsets. Skin-derived DCs constitutively migrate to draining lymph nodes; this process is further increased in response to inflammatory signals.
Following skin injury, DCs become activated, modulating their chemokine receptor profile to facilitate their migration and positioning in the draining lymph node, and upregulating their expression of MHC and co-stimulatory molecules and cytokines3. Activated DCs promote the differentiation of naive antigen-specific T cells into effector T cell subsets3, and distinct DC populations can be associated with different types of T cell response. For example, migratory CD103+ DCs and lymphoid tissue-resident CD8α+ DCs show a superior ability to cross-present tissue-derived antigens and prime CD8+ T cells4. Depending on the nature of the immune challenge, DCs promote the differentiation of distinct effector CD4+ T helper (TH) cell populations and drive the development of T follicular helper (TFH) cells, which promote B cell antibody responses.
During inflammation, two additional subsets of DCs accumulate in the skin: monocyte-derived DCs and plasmacytoid DCs (pDCs). Upon activation, pDCs secrete large amounts of IFNα and initiate T cell responses against microorganisms5. Monocyte-derived DCs can also participate in tissue immunity by initiating innate and adaptive immune responses against pathogens6.
Intestine (right panel)Intestinal DCs must maintain tolerance to food antigens and commensal bacteria, but are also crucial for protective immunity against invading pathogens. In the lamina propria, CD103–CX3CR1+CD11b+ DCs sample luminal antigens by projecting dendrites through the epithelial cell layer7. CD103+CD11b+ DCs migrate to the draining lymph nodes in a CCR7-dependent manner and present intestinal antigens to T cells; these DCs are potent producers of retinoic acid and promote the peripheral differentiation of regulatory T (TReg) cells, which help maintain gut integrity8-10. Retinoic acid-producing DCs also ‘imprint’ T cells with gut-homing molecules, such as α4β7 integrin.
Dendritic cells: controllers of adaptive immunity
Miriam MeradDendritic cells (DCs) are a heterogeneous population of rare haematopoietic cells that co-evolved with the formation of the adaptive immune system. Distributed throughout the body, but most abundant in lymphoid organs and at interface tissues, DCs are constantly sampling their environment. Following antigen uptake, they migrate to draining lymph nodes to present tissue-derived antigens and initiate adaptive immunity. DCs tailor adaptive immune
responses to match environmental cues, thereby serving as a bridge between the innate and adaptive immune systems. As such, they have key roles in both initiating immune responses to pathogens and maintaining tissue tolerance1. Remarkable progress has been made in unravelling the intricacies of DC biology and targeting these cells for therapy is now a realistic goal1. This Poster summarizes key DC functions, focusing on mouse DC subsets.
Abbreviations AIM2, absent in melanoma 2; BATF3, basic leucine zipper transcriptional factor ATF-like 3; BCR, B cell receptor; BDCA, blood DC antigen; CCR, CC-chemokine receptor; CLEC, C-type lectin; CTL, cytotoxic T lymphocyte; CXCR5, CXC-chemokine receptor 5; CX3CR1, CX3C-chemokine receptor 1; DAI, DNA-dependent activator of IFN-regulatory factor; DCIR, DC immunoreceptor; DC-SIGN, DC-specific ICAM3-grabbing non-integrin; DHX58, DEAH box protein 58; FLT3L, FMS-related tyrosine kinase 3 ligand; FOXP3, forkhead box P3; ICOS, inducible T cell co-stimulator; ICOSL, ICOS ligand; ID2, inhibitor of DNA binding 2; IFI16, interferon-γ inducible protein 16; IFN, interferon; IL, interleukin; IRF, interferon regulatory factor; MDA5, melanoma differentiation-associated gene 5; NAIP5, NLR apoptosis-inhibitory protein 5; NLR, NOD-like receptor; NLRC, NOD-, LRR- and CARD-containing; NLRP, NOD-, LRR- and pyrin domain-containing; NOD, nucleotide-binding oligomerization domain;
RIG-I, retinoic acid-inducible gene I; RLR, RIG-I-like receptor; TCR, T cell receptor; TGFβ, transforming growth factor-β; TSLP, thymic stromal lymphopoietin; XCR1, XC-chemokine receptor 1.
AffiliationsMiriam Merad is at the Mount Sinai School of Medicine, 1425 Madison Avenue, New York 10044, USA. email: [email protected]
The author declares no competing financial interests.
Edited by Yvonne Bordon and Olive Leavy; copyedited by Isabel Woodman; designed by Simon Bradbrook.© 2011 Nature Publishing Group. All rights reserved.
http://www.nature.com/nri/posters/dendriticcellsReferences available online.
IMMUNOLOGY
© 2011 Macmillan Publishers Limited. All rights reserved
![The Garvagh Madonna Illuminating the Photochemistry of ...posters/Examples...Rafaello Santi (Raphael): The Garvagh Madonna Madder Lake [Alizarin + Puprurin] Alizarin Purpurin Madder](https://static.fdocument.org/doc/165x107/60e7340affeae4294037f4d9/the-garvagh-madonna-illuminating-the-photochemistry-of-postersexamples-rafaello.jpg)