
Decreased β-adrenergic responsiveness in mouse epidermal papillomas during tumor promotion with...
5
Cancer Letters, 9 (1980) 245-249 o EIsevier/North-Holland Scientific Publishers Ltd. 245 DECREASED @DRENERGIC RESPONSIVENESS IN MOUSE EPIDERMAL PAPILLOMAS DURING TUMOR PROMOTION WITH PHORBOL MYRISTATE ACETATE SEYMOUR J. GARTE and SIDNEY BELMAN Department of Environmental Medicine, New York University, New York, NY 10016 (U.S.A.) (Received 28 December 1979) (Revised version received 18 February 1980) (Accepted 18 February 1980) SUMMARY The response to the /3-adrenergic agonist (-)-isoproterenol in mouse epidermis and in papillomas arising during tumor promotion with phorbol myristate acetate was determined by measurement of cyclic AMP levels after in vivo administration of (-)-isoproterenol. Papillomas exhibited less than half the hormonal responsiveness found in epidermis from control groups or from papilloma-bearing animals. No difference in the incorpora- tion of labelled (-)-isoproterenol into epidermal or papilloma tissue was seen. INTRODUCTION The potent skin tumor promoter phorbol myristate acetate (PMA) has been shown to alter the biological and biochemical state of the target tissue. Mouse skin becomes hyperplastic with increases in RNA, DNA, and protein synthesis [2]. Ornithine decarboxylase [14] and protease [16] levels are increased and alterations in cyclic nucleotide metabolism [4,7,13] are found after treatment with PMA. We have recently demonstrated that the loss of @-adrenergic responsiveness in mouse skin following PMA treatment [lo] is due to uncoupling of the P-adrenergic receptor from adenyl cyclase, rather than to a loss of receptor binding capacity [ 3,8]. It is still unclear which, if any, of these effects of PMA are involved specifically with the process of tumor promotion. We previously discussed Address all correspondence to: Dr. S.J. Garte, Department of Environmental Medicine, NYU Medical Center, 550 First Avenue New York, N.Y. 10016, U.S.A.
Transcript of Decreased β-adrenergic responsiveness in mouse epidermal papillomas during tumor promotion with...

Cancer Letters, 9 (1980) 245-249 o EIsevier/North-Holland Scientific Publishers Ltd.
245
DECREASED @DRENERGIC RESPONSIVENESS IN MOUSE EPIDERMAL PAPILLOMAS DURING TUMOR PROMOTION WITH PHORBOL MYRISTATE ACETATE
SEYMOUR J. GARTE and SIDNEY BELMAN
Department of Environmental Medicine, New York University, New York, NY 10016 (U.S.A.)
(Received 28 December 1979) (Revised version received 18 February 1980) (Accepted 18 February 1980)
SUMMARY
The response to the /3-adrenergic agonist (-)-isoproterenol in mouse epidermis and in papillomas arising during tumor promotion with phorbol myristate acetate was determined by measurement of cyclic AMP levels after in vivo administration of (-)-isoproterenol. Papillomas exhibited less than half the hormonal responsiveness found in epidermis from control groups or from papilloma-bearing animals. No difference in the incorpora- tion of labelled (-)-isoproterenol into epidermal or papilloma tissue was seen.
INTRODUCTION
The potent skin tumor promoter phorbol myristate acetate (PMA) has been shown to alter the biological and biochemical state of the target tissue. Mouse skin becomes hyperplastic with increases in RNA, DNA, and protein synthesis [2]. Ornithine decarboxylase [14] and protease [16] levels are increased and alterations in cyclic nucleotide metabolism [4,7,13] are found after treatment with PMA. We have recently demonstrated that the loss of @-adrenergic responsiveness in mouse skin following PMA treatment [lo] is due to uncoupling of the P-adrenergic receptor from adenyl cyclase, rather than to a loss of receptor binding capacity [ 3,8]. It is still unclear which, if any, of these effects of PMA are involved specifically with the process of tumor promotion. We previously discussed
Address all correspondence to: Dr. S.J. Garte, Department of Environmental Medicine, NYU Medical Center, 550 First Avenue New York, N.Y. 10016, U.S.A.

246
the loss of fi-adrenergic responsiveness as a significant action of PMA on cell membranes [S]. To further investigate whether this phenomenon may be related to permanent phenotypic changes in early lesions arising from PMA treatment, we have examined the response of papilloma tissue to in vivo administration of the fl-adrenergic agonist (-)-isoproterenol.
MATERIALS AND METHODS
Female Swiss Ha/ICR mice were purchased from Sprague-Dawley (Madison, WI). Mice were initiated at 8 weeks of age with a single topical dose of 25 pg dimethylbenzanthracene (DMBA) in 0.2 ml acetone to the shaved back. Promotion began 1 week later with 3 weekly topical treatments of 10 pg PMA in 0.2 ml acetone. Treatment with PMA was discontinued after 17 weeks, and mice were sacrificed after an additional 3 weeks. For determination of the P-adrenergic response, mice were injected intraperitoneally with 0.2 ml phosphate buffered saline (PBS) containing 300 nmol (-)-isoproterenol (Sigma, St. Louis, MO) 10 min before sacrifice (for maximum stimulation of adenyl cyclase [lo]), by cervical dislocation between 10-11.00 a.m. The mice were immediately frozen for 30 s in liquid nitrogen, Epidermis was removed by scraping as previously described [ 41, and papillomas were cut off at the base. One to 3 samples of papillomas (each sample contained 1 or more papillomas) and a sample of ‘uninvolved’ epidermis were harvested from each animal. Tissues were homogenized in 6% trichloroacetic acid (TCA). Cyclic AMP was deter- mined by radioimmuneassay as previously described [ 41 using antisera and 12’1-antigen purchased from Collaborative Research (Waltham, MA). DNA was assayed by a fluorometric procedure [ 61.
Incorporation of (-)-[3H]isoprotereno1 Animals with papillomas were injected with 300 nmol (-)-isoproterenol in
0.2 ml PBS containing 30 PCi (-)-[3H]isoprotereno1 (3 nmol) purchased from New England Nuclear (Boston, MA). After 10 min, the mice were sacrificed, epidermal and papilloma tissues were collected as described above and homogenized in 7 ml 6% TCA. Homogenates were centrifuged 20 min at 1000 X g. Supematants were decanted off and pellets were washed twice with TCA. Aliquots of 1 ml from supernatant and washings were counted by liquid scintillation in 10 ml Aquasol (New England Nuclear, Boston, MA). Appropriate corrections for acid quenching were used to calculate total DPM for each sample. DNA was measured in the final TCA pellet.
RESULTS AND DISCUSSION
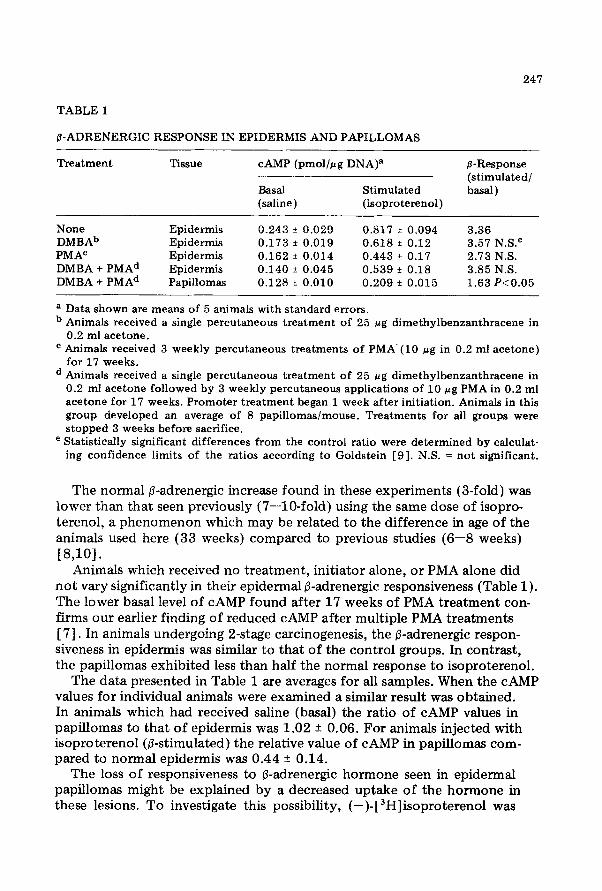
The values of CAMP in basal and isoproterenol stimulated mouse epi- dermis are given in Table 1. The p-adrenergic response is the isoproterenol stimulated increase in CAMP over the basal level.
247
TABLE 1
p-ADRENERGIC RESPONSE IN EPIDERMIS AND PAPILLOMAS
Treatment Tissue CAMP (pmol/pg DNA)=
&Sal Stimulated (saline) (isoproterenol)
@-Response (stimulated/ basal)
None Epidermis 0.243 f 0.029 0.817 F 0.094 3.36 DMBAb Epidermis 0.173 * 0.019 0.618 f 0.12 3.57 N.S? PMAe Epidermis 0.162 f 0.014 0.443 f 0.17 2.73 N.S. DMBA + PMAd Epidermis 0.140 * 0.045 0.539 + 0.18 3.85 N.S. DMBA + PMAd Papillomas 0.128 f 0.010 0.209 + 0.015 1.63 P~0.05
a Data shown are means of 5 animals with standard errors. b Animals received a single percutaneous treatment of 25 pg dimethylbenzanthracene in
0.2 ml acetone. c Animals received 3 weekly percutaneous treatments of PMA.(lO pg in 0.2 ml acetone)
for 17 weeks. d Animals received a single percutaneous treatment of 25 fig dimethylbenzanthracene in
0.2 ml acetone followed by 3 weekly percutaneous applications of 10 pg PMA in 0.2 ml acetone for 17 weeks. Promoter treatment began 1 week after initiation. Animals in this group developed an average of 8 papillomaslmouse. Treatments for all groups were stopped 3 weeks before sacrifice.
e Statistically significant differences from the control ratio were determined by calculat- ing confidence limits of the ratios according to Goldstein [9]. N.S. = not significant.
The normal p-adrenergic increase found in these experiments (3-fold) was lower than that seen previously (7-lo-fold) using the same dose of isopro- terenol, a phenomenon which may be related to the difference in age of the animals used here (33 weeks) compared to previous studies (6-8 weeks) [8,101.
Animals which received no treatment, initiator alone, or PMA alone did not vary significantly in their epidermal &adrenergic responsiveness (Table 1). The lower basal level of CAMP found after 17 weeks of PMA treatment con- firms our earlier finding of reduced CAMP after multiple PMA treatments [ 71. In animals undergoing 2-stage carcinogenesis, the P-adrenergic respon- siveness in epidermis was similar to that of the control groups. In contrast, the papillomas exhibited less than half the normal response to isoproterenol.
The data presented in Table 1 are averages for all samples. When the CAMP values for individual animals were examined a similar result was obtained. In animals which had received saline (basal) the ratio of CAMP values in papillomas to that of epidermis was 1.02 + 0.06. For animals injected with isoproterenol (P-stimulated) the relative value of CAMP in papillomas com- pared to normal epidermis was 0.44 _+ 0.14.
The loss of responsiveness to fl-adrenergic hormone seen in epidermal papillomas might be explained by a decreased uptake of the hormone in these lesions. To investigate this possibility, (-)-[ 3H]isoproterenol was

243
injected 10 min before sacrifice into animals bearing papillomas. Incorpora- tion into papillomas and normal epidermis of labelled material was 272 f 15 dpm/pg DNA and 229 f 57 dpm/pg DNA respectively.
The loss of fl-adrenergic responsiveness reported here appears to be due to a phenotypic alteration in the papilloma cells, rather than to a direct effect of the promoter. Preliminary evidence suggests that this phenomenon is present in later stage papillomas as well. In animals undergoing promotion with PMA for 31 weeks, the p-adrenergic responsiveness of normal epidermis was 1.96 compared to 0.86 for papillomas. The loss of hormonal responsive- ness in the normal epidermis of these animals (which were 41 weeks old) may be due to the increased period of promoter treatment. It is interesting, however, that the same relative decrease in responsiveness in papillomas compared to epidermis was found in the 17- and 31-week treatment groups.
Epidermal papillomas arising during tumor promotion with PMA may be considered analogous to the pre-neoplastic lesions studied in liver tumori- genesis [6]. Although the growth and progression properties of the papillo- mas have been examined [ 51, detailed biochemical characterization of these lesions has not been studied. The loss of sensitivity to /?-adrenergic hormones in papillomas may be due to a permanently uncoupled enzyme receptor complex [ 31. Decreased adenyl cyclase response to hormones has been found in submaxillary gland carcinoma of the mouse [ 121 and in rapidly growing rat hepatomas [ 11, as well as in chemically transformed epithelial cells [ 111 and leukemic lymphocytes [ 151. Further studies on biochemical pheno- typic alterations in papillomas are required to understand the process of tumorigenesis.
ACKNOWLEDGEMENTS
We thank Karen B. Fennikoh for expert technical assistance. This investi- gation was supported by Grant No. CA18536, awarded by the National Cancer Institute, DHEW, and is part of Center Programs supported by Grant No. ES00260 from the National Institute of Environmental Health Science, and Grant No. CA13343 from the National Cancer Institute, DHEW.
REFERENCES
1 Allen, D.O., Munshower, J., Morris, H.P. and Weber, G. (1971) Regulation of adenyl cyclase in hepatomas of different growth rates. Cancer Res., 31, 557-560.
2 Baird, W.M., Sedgwick, J.A. and Boutwell, R.K. (1971) Effects of phorbol and four diesters of phorbol on the incorporation of tritiated precursors into DNA, RNA and protein in mouse epidermis. Cancer Res., 31,1434-1439.
3 Belman, S. and Garte, S.J. (1980) Antagonism between phorbol myristate acetate and butyric acid on isoproterenol elevation of cyclic AMP and their effect on p-adrenergic receptors in mouse epidermis. Cancer Res., 40, 240-244.
4 Belman, S., Troll, W. and Garte, S.J. (1978) Effect of phorbol myristate acetate on cyclic nucleotide levels in mouse epidermis. Cancer Res., 38, 2978-2982.

249
5 Burns, F.J., Vanderlaan, M., Synder, E. and Albert, R.E. (1978) Induction and pro- gression, kinetics of mouse skin papillomas. In : Carcinogenesis, Vol. 2, Mechanisms of Tumor Promotion and Cocarcinogenesis, pp. 91-96. Editors: T.J. Slaga, A. Sivak and R.K. Boutwell. Raven Press, New York.
6 Farber, E. (1976) Hyperplastic areas, hyperplastic nodules and hyperbasophilic areas as putative precursor lesions. Cancer Res., 36, 2532-2533.
7 Garte, S.J. and Belman, S. (1978) Effect of multiple phorbol myristate acetate treat- ments on cyclic nucleotide levels in mouse epidermis. Biochem. Biophys. Res. Commun., 84,489-494.
8 Garte, S.J. and Belman, S. (1980) Tumor promoter uncouples p-adrenergic receptor from adenyl cyclase in mouse epidermis. Nature. In Press.
9 Goldstein, A. (1964) Biostatistics: An Introductory Text. pp. 184-187. McMillan Co., New York.
10 Grimm, W. and Marks, F. (1974) Effect of tumor promoters on the normal and iso- proterenol elevated level of adenosine 3’,5’-cyclic monophosphate in mouse epidermis in duo. Cancer Res., 34, 3128-3134.
11 Makarski, J. and Niles, R.M. (1978) Loss of adenylate cyclase hormonal sensitivity in chemically transformed epithelial cells. Exp. Cell Res., 114, 191-. 195.
12 Malamud, D. and Irwin, D. (1978) Altered cyclic adenosine 3’,5’-monophosphate metabolism and enzymatic profiles of a transplantable murine submaxillary gland carcinoma (myoepithelioma). Cancer Res., 38, 2945-2949.
13 Mufson, R.A., Simsiman, R.C. and Boutwell, R.K. (1979) Increased cyclic adenosine 3’,5’-monophosphate phosphodiesterase activity in the epidermis of phorbol ester- treated mouse skin and in papillomas. Cancer Res., 39, 2036-2040.
14 O’Brien, T.G., Simsiman, R.C. and Boutwell, R.K. (1975) Induction of the polyamine- biosynthetic enzymes in mouse epidermis by tumor promoting agents. Cancer Res., 35,1662-1670.
15 Sheppard, J.R., Gormus, R. and Moldow, C.F. (1977) Catecholamine hormone receptors are reduced on chronic lymphocytic leukaemic lymphocytes. Nature, 269, 693-695.
16 Troll, W., Klassen, A. and Janoff, A. (1970) Tumorigenesis in mouse skin: Inhibition by synthetic inhibitors of proteases. Science, 169,1211-1213.
















![HOTAIR Knockdown Decreased the Activity Wnt/β-Catenin ... · of this pathway are frequently altered in human cancer mainly by genetic and epigenetic mechanisms [25-27]. The abnormal](https://static.fdocument.org/doc/165x107/5e638e505ba2f7369635202e/hotair-knockdown-decreased-the-activity-wnt-catenin-of-this-pathway-are-frequently.jpg)
