COMPOSICIÓN ISOTÓPICA ( 15N y δ13C) · 2014-09-11 · composición de diferentes elementos...
98
COMPOSICIÓN ISOTÓPICA (δ 15 N y δ 13 C) DURANTE EL DESARROLLO ONTOGÉNICO DEL TIBURÓN PILOTO Carcharhinus falciformis (Müller & Henle 1839) EN LA ZONA OCEÁNICA DEL PACÍFICO ORIENTAL TROPICAL TESIS QUE PARA OBTENER EL GRADO DE MAESTRÍA EN CIENCIAS EN MANEJO DE RECURSOS MARINOS PRESENTA MARÍA ANTONIETA GALINDO ROSADO LA PAZ, B.C.S., JUNIO DE 2014 INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS
Transcript of COMPOSICIÓN ISOTÓPICA ( 15N y δ13C) · 2014-09-11 · composición de diferentes elementos...

COMPOSICIÓN ISOTÓPICA (δ15N y δ13C)
DURANTE EL DESARROLLO ONTOGÉNICO DEL
TIBURÓN PILOTO Carcharhinus falciformis (Müller
& Henle 1839) EN LA ZONA OCEÁNICA DEL
PACÍFICO ORIENTAL TROPICAL
TESIS QUE PARA OBTENER EL GRADO DE
MAESTRÍA EN CIENCIAS EN MANEJO DE RECURSOS MARINOS
PRESENTA
MARÍA ANTONIETA GALINDO ROSADO
LA PAZ, B.C.S., JUNIO DE 2014
INSTITUTO POLITECNICO NACIONAL CENTRO INTERDISCIPLINARIO DE CIENCIAS MARINAS



AGRADECIMIENTOS
Al Consejo Nacional de Ciencia y Tecnología (CONACyT) y al Programa
Institucional de Formación de Investigadores (PIFI) por el apoyo económico.
A los miembros del Comité de Tesis Dr. Felipe Galván, Dr. Fernando
Elorriaga, Dr. Alberto Sánchez, Dr. Sergio Aguíñiga y en especial al Dr. Yassir
Torres por sus conocimientos y consejos.
Al Dr. Antonio Delgado Huertas por aceptarme en su equipo de trabajo por
unos meses y recibir un excelente trato durante mi estancia.
Al Tec. Lab. Ángel Huberto Rubalcaba Díaz por su gran ayuda en el pesaje de
las muestras y evitar que fuera aburrido.
A mis compañeros del laboratorio de Ecología de Peces, en particular a
Peggy que con paciencia me explicó y ayudó con las gráficas del programa R. A
Chayo por realizar los trámites de la beca PIFI y de la reinscripción cuando estaba en
España.
A la gorda güera que se caracteriza por su gran sonrisa, ánimo y entusiasmo
(jajajaja) que siempre está para ayudarme cuando hago las cosas a último minuto
(muy pocas veces).
A MIS ALIADOS
A todos ellos, muchas gracias

DEDICATORIA
A mi familia que siempre me aconsejó y me mostró el mejor
camino a seguir. Gracias por ser mi guía. Por detenerme
cuando debías y por empujarme cuando tenía miedo de
seguir mis sueños.
A las gordas y gordos, que aunque estemos lejos saben
que en mi corazón solo puede haber agradecimiento hacia
ustedes porque son como una familia para mí.

ii
ÍNDICE
LISTA DE FIGURAS ....................................................................................................... iv
LISTA DE TABLAS ......................................................................................................... vi
GLOSARIO ................................................................................................................ viii
1. INTRODUCCIÓN .................................................................................................... 1
2. ANTECEDENTES ................................................................................................... 3
3. JUSTIFICACIÓN ..................................................................................................... 6
4. HIPÓTESIS ............................................................................................................. 7
5. OBJETIVO ............................................................................................................... 8
5.1 Objetivo General ................................................................................................ 8
5.2 Objetivos Específicos......................................................................................... 8
7.1 Trabajo de campo ........................................................................................... 10
7.2 Trabajo de laboratorio ...................................................................................... 10
7.3 Trabajo de escritorio ........................................................................................ 12
7.3.2 Composición de δ13C y δ15N ..................................................................... 13
7.3.3 Amplitud y Traslapo trófico. ....................................................................... 14
7.3.4 Estimación de la posición trófica ............................................................... 15
7.3.5 Patrones de δ13C y δ15N durante el ciclo de vida del tiburón piloto ........... 16
8. RESULTADOS ...................................................................................................... 16
8.1 Relación C: N ...................................................................................................... 17
8.2 Composición de δ13C y δ15N entre sexos. ....................................................... 19
8.3 Composición de δ13C y δ15N entre estados ontogénicos. ................................ 21
8.4 Composición isotópica entre sexos y estados ontogénicos ............................. 23
8.4 Composición de δ13C y δ15N en los años de captura. ...................................... 25
8.5 Amplitud y traslapo de nicho ............................................................................ 26
8.6 Posición Trófica ............................................................................................... 34
8.7 Composición de δ13C y δ15N durante el desarrollo ontogénico del tiburón piloto.
............................................................................................................................... 35
8.8 Perfiles ontogénicos individuales δ13C y δ15N del tiburón piloto. ..................... 38
9. DISCUSIÓN .......................................................................................................... 45

iii
9.1 Relación C: N ................................................................................................... 45
9.2 Composición de δ13C y δ15N ............................................................................... 47
9.3 Amplitud y traslapo de nicho ............................................................................ 53
9.3 Posición Trófica .............................................................................................. 57
10. CONCLUSIONES ................................................................................................ 60
11. BIBLIOGRAFÍA ................................................................................................... 61
Anexos ...................................................................................................................... 72

iv
LISTA DE FIGURAS
Figura 1. Vista lateral del tiburón piloto (Carcharhinus falciformis)……………….. 1
Figura 2. Área de operación de la flota atunera de la Comisión Interamericana del
Atún Tropical (CIAT) durante 2003 – 2005 (Los puntos representan lances donde se
capturaron tiburones pilotos de manera incidental)………………………………… 9
Figura 3. Vista de la vértebra con perforaciones, que representan las etapas o estado
ontogénico del tiburón piloto (C. falciformis), desde el foco hasta la parte más distal
(foco=0, embrión=1, neonato=2, juvenil=3 y adulto=4)…………………………….. 11
Figura 4. Relación entre C: N y la razón de δ13C en músculo de tiburón piloto (C.
falciformis) capturados del 2003 al 2005 zona del OPOT…………………………. 18
Figura 5. Relación entre C: N y la razón de δ13C en vértebra de tiburón piloto (C.
falciformis) capturados del 2003 al 2005 zona del OPOT…………………………. 19
Figura 6. Valores promedio y desviaciones estándar de δ13C y δ15N de músculo del
tiburón piloto (C. falciformis) por estado ontogénico capturados del 2003 al 2005 zona
del OPOT………………………………………………………………………………... 22
Figura 7. Valores promedio y desviaciones estándar de δ13C y δ15N de vértebra del
tiburón piloto (C. falciformis) por estado ontogénico capturado en la zona del
OPOT…………………………………………………………………………………….. 23
Figura 8. Valores promedio y desviaciones estándar de δ13C y δ15N de músculo del
tiburón piloto (C. falciformis) por sexo y estado ontogénico (Hembras Adultas = HA,
Hembras Juveniles = HJ, Hembras Neonatos = HN, Machos Adultos = MA, Machos
Juveniles= MJ, Machos Neonatos = MN) capturados del 2003 al 2005 en el
OPOT…………………………………………………………………………………….. 24
Figura 9. Diagrama de amplitud y nicho trofico entre sexos mediante los valores δ13C
y δ15N de músculo del tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la
zona del OPOT…………………………………………………………………………. 27
Figura 10. Diagrama de amplitud y nicho trofico entre sexos mediante los valores
δ13C y δ15N en vértebra del tiburón piloto (C. falciformis) capturados del 2003 al 2005
en la zona del OPOT…………………………………………………………………… 28

v
Figura 11. Diagrama de amplitud y nicho trófico entre estados ontogénicos, mediante
los valores δ13C y δ15N de músculo del tiburón piloto (C. falciformis) capturados del
2003 al 2005 zona del OPOT…………………………………………………………. ….29
Figura 12. Diagrama de amplitud y nicho trófico entre estados ontogénicos (foco,
embrión, neonato, juvenil y adulto) mediante los valores δ13C y δ15N de vértebra del
tiburón piloto (C. falciformis) capturados del 2003 al 2005 zona del OPOT……... ….31
Figura 13. Diagrama de amplitud y nicho trofico de hembras de tiburón piloto (C.
falciformis) por estado ontogénico (negro= Hembra Adulta, rojo= Hembra Juvenil y
verde= Hembra Neonato) mediante los valores δ13C y δ15N de músculo……………32
Figura 14. Diagrama de amplitud y nicho trofico de machos y sus estados
ontogénicos (negro = Macho Adulto, rojo = Machos Juvenil y verde = Macho
Neonato) mediante los valores δ13C y δ15N en músculo del tiburón piloto (C.
falciformis)……………………………………………………………………………….. ….33
Figura 15. Valores de δ13C y δ15N en vértebras de tiburón piloto (C. falciformis)
hembra capturados del 2003 al 2005 en la zona del OPOT, que muestra los distintos
estados ontogénicos dependiendo del número de perforación, desde el foco de la
vértebra hasta la parte más distal (0=foco, 1=embrión, 2=neonato, 3 en
adelante=juvenil y últimas perforaciones=adulto)…………………………………... ….36
Figura 16. Valores de δ13C y δ15N en vértebras de tiburón piloto (C. falciformis)
machos capturados del 2003 al 2005 en la zona del OPOT, que muestra los distintos
estados ontogénicos dependiendo del número de perforación, desde el foco de la
vértebra hasta la parte más distal (0=foco,1=embrión, 2=neonato, 3 en
adelante=juvenil y últimas perforaciones=adulto)…………………………………... ….37
Figura 17. Perfiles individuales por sexos de δ13C en vértebras de tiburón piloto (C.
falciformis) capturados del 2003 al 2005 en la zona del OPOT, que muestran las
etapas ontogénicas…………………………………………………………………….. ….40
Figura 18. Perfiles individuales por sexos de δ15N en vértebras de tiburón piloto (C.
falciformis) capturados del 2003 al 2005 en la zona del OPOT, que muestran las
etapas ontogénicas…………………………………………………………………….. ….43

vi
LISTA DE TABLAS
Tabla 1. Tamaño de muestra de músculo y vértebra de tiburón piloto (C. falciformis)
por sexo y estado ontogénico capturados del 2003 al 2005 en la zona del
OPOT………………….................................................................................................17
Tabla 2. Valores promedio e intervalos de δ13C y δ15N en músculo del tiburón piloto
(C.falciformis) por sexo capturados del 2003 al 2005 zona del OPOT…………….....20
Tabla 3. Valores promedio e intervalos de δ13C y δ15N en vértebras de tiburón piloto
(C.falciformis) por sexo capturados del 2003 al 2005 zona del
OPOT…………………………………………………………………………….………......20
Tabla 4. Valores promedio e intervalos de δ13C y δ15N en músculo de tiburón piloto
(C. falciformis) por estados ontogénicos (neonatos, juveniles y adultos) capturados
del 2003 al 2005 zona del OPOT………………………………………………………....21
Tabla 5. Valores promedio e intervalos de δ13C y δ15N en vértebras de tiburón piloto
(C. falciformis) por estados ontogénicos (foco, embrión, neonato, juvenil y adulto) en
la zona del OPOT tropical...........................................................................................23
Tabla 6. Valores promedio e intervalos de δ13C y δ15N en músculo de tiburón piloto
(C. falciformis) por sexos y estados ontogénicos (HN= Hembra neonato, HJ= Hembra
juvenil, HA= Hembra adulta, MN= Macho neonato, MJ= Macho juvenil y MA= Macho
adulto) capturados del 2003 al 2005 zona del OPOT……………………………......…24
Tabla 7. Valores promedio e intervalos de δ13C y δ15N en músculo de tiburón piloto
(C. falciformis) por años de captura 2003 al 2005 en la zona del OPOT……………..25
Tabla 8. Valores promedio e intervalos de δ13C y δ15N en vértebras de tiburón piloto
(C. falciformis) por años de captura 2003 al 2005………………………………………26
Tabla 9. Áreas de los elipses y polígonos por sexo en músculo de tiburón piloto (C.
falciformis) capturados del 2003 al 2005 en la zona del OPOT……………...………..26
Tabla 10. Áreas de los elipses y polígonos por sexo en vértebra de tiburón piloto
(C.falciformis) capturados del 2003 al 2005 en la zona del OPOT…………..………27
Tabla 11. Áreas de los elipses y polígonos por estado ontogénico en músculo de
tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona del OPOT..28

vii
Tabla 12. Áreas de los elipses y polígonos por estado ontogénico en vértebra de
tiburón piloto (C.falciformis) capturados del 2003 al 2005 en la zona del OPOT…30
Tabla 13. Áreas de los elipses y polígonos para hembras y sus estados ontogénicos
en músculo de tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona
del OPOT………………………………………………………………………………....….32
Tabla 14. Áreas de los elipses y polígonos de machos y sus estados ontogénicos en
músculo de tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona del
OPOT…………………………………………………………………………….…………..33
Tabla 15. Valores de la posición trófica calculados para el tiburón piloto
(C.falciformis) a partir de δ15N en la zona del OPOT…………………..………..……...34
Tabla 16. Número de tiburón (hembras) para identificar en los perfiles individuales,
con los datos de longitud, edad, fecha de captura y la zona en el OPOT……………44
Tabla 17. Número de tiburón (machos) para identificar en los perfiles individuales,
con los datos de longitud, edad, fecha de captura y la zona en el OPOT…….……...45

viii
GLOSARIO
δ13C: Diferencia expresada en partes por mil dentro de la razón de 13C/12C de una
muestra y la razón 13C/12C de un estándar (Pee Dee Belemnita en el caso del
carbono) (Rau et al. 1990).
δ15N: Diferencia expresada en partes por mil dentro de la razón de 15N/14N de una
muestra y la razón 15N/14N de un estándar (nitrógeno atmosférico en el caso del
nitrógeno) (Rau et al. 1990).
Amplitud de nicho: Selección de la dieta con respecto a un número potencial de
tipos alimenticios (Odum, 1972).
Especialista: Organismo que tiene una dieta restringida a un reducido número de
especies (Odum, 1972).
Espectrómetro de masas: Equipo que permite analizar con gran precisión la
composición de diferentes elementos químicos e isótopos atómicos, separando los
núcleos atómicos en función de su relación carga-masa (z/m). Puede utilizarse para
identificar los diferentes elementos químicos que forman un compuesto, o para
determinar el contenido isotópico de diferentes elementos en un mismo compuesto.
Con frecuencia se encuentra como detector de un cromatógrafo de gases, en una
técnica híbrida conocida por sus iniciales en inglés, GC-MS. 1
Generalista: Organismo que presenta una dieta amplia, es decir, su dieta está
conformada por una gran diversidad de presas (Odum, 1972).
Isótopo: Término que engloba a átomos de un mismo elemento químico que se
encuentran en el mismo sitio de la tabla periódica. Éstos presentan igual número

ix
atómico (número de protones en el núcleo), pero diferente número de masa (suma
del número de neutrones y de protones en el núcleo) (Glasstone 1976).
Isótopo Estable: Muchos elementos de la Tabla Periódica y entre ellos, el H, C, N,
O y S, poseen dos o más isótopos (núcleos de diferente masa pero idéntica
configuración electrónica) “estables” que no participan en ningún proceso de
desintegración nuclear, de ahí su nombre. Estos elementos (y sus isótopos) se
encuentran ampliamente distribuidos por la litosfera, hidrosfera, biosfera y atmósfera
en forma de diferentes moléculas, constituyendo unos excelentes trazadores
naturales de los procesos fisicoquímicos que ocurren en la naturaleza (Ehleringer et
al. 1989).
Nivel trófico: Posición o categoría en la que se clasifica un determinado organismo
dentro de la cadena trófica; dicha posición está dada de acuerdo a la manera en la
cual obtiene su materia y energía (Gerking, 1994).
Ontogenia: describe el desarrollo de un organismo, desde el óvulo fertilizado hasta
su senescencia, pasando por la forma adulta.
Relación carbono/ nitrógeno (C/N): La cantidad de carbono dividido por la cantidad
de nitrógeno presente en una muestra, generalmente en miligramos (Rau et al.
1990).
Tasa de recambio isotópico: Periodo en el que el valor del isótopo estable es
discriminado e incorporado en el tejido del consumidor y el cual refleja la señal
isotópica de un nuevo recurso alimenticio, después de un cambio entre dietas
isotópicamente distintas (Carlisle et al. 2012).
Traslapo Trófico: Utilización del mismo recurso alimenticio (presas) por dos o más
especies como parte de su dieta (Odum, 1972)

x
RESUMEN
El tiburón piloto (Carcharhinus falciformis) es una de las especies de tiburones más
abundantes en las capturas pesqueras en el Océano Pacífico Oriental Tropical; sin
embargo se desconoce aspectos sobre su dinámica trófica y sus posibles cambios
en las diferentes etapas ontogénicas. Se analizó la composición isotópica (δ13C y
δ15N) de músculo (n = 159) y vértebra (n = 44) durante 3 años (2003 - 2005). No se
detectaron diferencias significativas entre sexos en ambos isótopos para los dos
tejidos (p > 0.05), lo que indica una alimentación similar (especies presas y zonas de
alimentación parecidas). De acuerdo al modelo de mezcla SIBER, existe un traslapo
alto entre sexos en ambos tejidos. Sin embargo, se detectaron diferencias
significativas en músculo en particular entre juveniles vs adultos para δ13C (H = 6.78,
p < 0.05) y entre juveniles vs neonatos en δ15N (F= 6.26, p < 0.05), mientras que
para vertebras se detectaron diferencias en δ13C (F= 6.59, p= <0.05) y δ15N (F= 4.19,
p=<0.05) entre todos los estadios, en particular entre las hembras en diferentes
estados ontogénicos (H = 34.77, p < 0.05), lo que indica que: 1) consumen distintos
tipos de presas como resultado de sus requerimiento energéticos (reproducción) y 2)
diferentes procedencia. Lo anterior fue corroborado con SIBER y con el análisis de
perfiles individuales, donde las hembras presentan un traslapo trófico bajo entre sus
distintos estados ontogénicos, mientras que en machos es mayor. El tiburón piloto
presentó una posición trófica promedio de 3.7, por lo que puede ser considerado
como un depredador terciario. A partir de la información generada en el presente
estudio se pudo registrar cambios en el comportamiento trófico en función a su
estado ontogénico, lo cual es de suma importancia para el entendimiento de la
estructura de la comunidad del Océano Pacífico Oriental Tropical, así como los
posibles impactos negativos que puedan existir derivados de diferentes impactos
naturales o antropogénicos

xi
ABSTRACT
The silky shark (Carcharhinus falciformis) is one of the most abundant sharks in
capture fisheries in the Eastern Tropical Pacific Ocean; however aspects of their
trophic dynamics and potential changes in different ontogenetic stages are unknown.
Isotopic composition (δ13C and δ15N) of muscle (n= 159) and vertebrae (n = 44) for 3
years (2003-2005) were analyzed. No significant differences between sexes in both
isotopes for both tissues (p> 0.05) were detected, indicating a similar food (prey
species and similar areas). According to SIBER mixture model, there is a high overlap
between sexes in both tissues. However, significant differences in muscle particularly
among juveniles vs. adults δ13C (H= 6.78, p< 0.05) and among infants in δ15N vs.
juveniles (F= 6.26, p< 0.05) were detected, while differences were detected for
vertebrae in δ 13C (F= 6.59, p= < 0.05) and δ15N (F= 4.19, p= < 0.05) between all
stages, particularly among females in different ontogenetic states (H= 34.77 , p<
0.05), which indicates that : 1) consume different types of prey as a result of their
energy requirement (reproduction), and 2) different origin. This was corroborated with
SIBER and analysis of individual profile, where females have a low trophic overlap
between their different ontogenetic states, while in males is higher. The pilot shark
showed an average of 3.7 trophic position, so can be regarded as a tertiary predator.
From the data generated in this study was able to record changes in the feeding
behavior based on their ontogenetic state, which is critical for understanding the
community structure of Eastern Tropical Pacific Ocean, as well as possible negative
impacts that may be derived from different natural or anthropogenic impact.
.

1
1. INTRODUCCIÓN
En el Océano Pacífico Oriental Tropical (OPOT), la familia Carcharhinidae es
considerada de gran importancia pesquera, debido a su alta contribución en las
capturas artesanales, industriales y deportivas (FAO, 1998). La familia
Carcharhinidae está representada por 18 especies, de las cuales 12 pertenecen al
género Carcharhinus (Compagno et al., 1995). Dentro del género Carcharhinus se
encuentra el tiburón piloto (Carcharhinus falciformis; Fig. 1), quien representa en
promedio el 51% de las capturas de elasmobranquios en el Pacifico Mexicano y
aproximadamente el 20% de la fauna incidental de otras pesquerías como la de atún
(Litvinov, 1993; FAO, 1998, 2006; UICN, 2003; DOF, 2004).
Figura 1. Vista lateral del tiburón piloto (Carcharhinus falciformis) Tomado de biodiversityexplorer.org
A nivel internacional, C. falciformis ha sido catalogado como una especie vulnerable
y casi amenazado en el Océano Pacífico (Dulvy et al., 2008), donde ha tenido un
decremento del 50% entre los años de 1993 a 2004, donde una de las razones de
esta reducción es la captura de neonatos y juveniles en las actividades pesqueras
(Minami et al., 2007). Pratt y Casey (1990) mencionan que la sobreexplotación de un
recurso como los tiburones ocasiona alteraciones graves en las tasas de renovación
natural de dichas poblaciones.
Dado a las consecuencias que puede generar en los ecosistemas la perdida de
organismos como los tiburones, la convención sobre comercio internacional de
especies en peligro (i.e. CITES) aprobó una resolución donde se propusieran y

2
establecieran programas para recolectar la información necesaria sobre captura y
esfuerzo, así como estudios que aporten al conocimiento de los aspectos biológicos
(reproducción, edad, crecimiento, y alimentación) de C. falciformis, los cuales de
manera conjunta permitirán evaluar el estado actual que guardan las poblaciones de
tiburones del mundo y las medidas que se deben considerar para una optima
recuperación (Castro et al., 1999).
Dentro de los aspecto biológicos a evaluar está la dinámica trófica, debido a
que los organismos presentan cambios de dieta durante el desarrollo ontogénico e
incluye cambios en el hábitat, debido principalmente a los requerimientos
energéticos en las diferentes etapas de su vida (neonato, juvenil y adultos), dando
oportunidad de una posible segregación trófica por sexo o estado de madurez
(Sainsbury et al., 2000; FAO, 2003; Chapple et al., 2011). Sin embargo, son pocos
los estudios sobre la dinámica trófica de C. falciformis desconociendo el
comportamiento trófico de neonatos y juveniles.
Una manera de conocer los posibles cambios en el uso de recursos y hábitat
durante el desarrollo ontogénico de C. falciformis es mediante el análisis de la
composición isotópica de δ15N y δ13C (Post, 2002). En el caso del δ15N, ha sido
considerado como indicador de la posición trófica; mientras que los valores de δ13C
permiten conocer cuál es la fuente autotrófica en la cadena alimenticia (DeNiro y
Epstein, 1981). En el caso de los tejidos metabólicamente activos (sangre, hígado y
músculo), la información generada será de días, semanas o meses. Mientras que en
tejidos inertes (vértebras y dientes), la información generada representará escalas
semestrales o anuales (Kim y Koch, 2012). En este contexto, a partir de la composición
isotópica (δ15N y δ13C) de dos tejidos (músculo y vértebras) se determinó la dinámica
trófica de C. falciformis a diferentes escalas de tiempo, con el fin de identificar el uso
diferencial de recursos alimenticios y hábitats en diferentes etapas ontogénicas en el
OPOT.

3
2. ANTECEDENTES
Estudios previos han registrado en algunas especies de tiburones cambios en su
alimentación y por ende en su hábitat, dependiendo de su estado ontogénico
(Hernández-Aguilar, 2008; Markaida y Sosa-Nishizaki, 2010; Torres-Rojas et al.,
2013) ya que conforme aumentan de tamaño, los requerimientos energéticos y sus
capacidades para capturar sus presas son diferentes. Lowe et al. (1996) mencionan
que una alimentación diferencial entre sexos y tallas indica un comportamiento trófico
distinto, sugiriendo la existencia de una segregación trófica en diferentes áreas
geográficas, lo que podría ayudar en el establecimiento de áreas de alimentación
para la conservación de la diversidad biológica marina.
En el caso de C. falciformis, a pesar de ser una de las especies de tiburón
más abundantes de los océanos, son pocos los trabajos realizados sobre la ecología
trófica, los cuales se citan a continuación:
A nivel internacional, Estupiñan y Cedeño (2005) en Ecuador, mencionan que
las presas de mayor importancia en el espectro trófico de C. falciformis fueron peces
como: Thunnus albacares (35.47% de Importancia Relativa), Thunnus spp. (18.41%),
Thunnus obesus (6.87%) y Auxis thazard (5.84%), así como de cefalópodos:
Ancistrocheirus lesueuri (2.86%) y Dosidicus gigas (1.32%). En los machos se
identificaron dos grupos de presas: moluscos (58.95%) y peces (41.07%); mientras
que en hembras tres grupos: peces (96.98%), cefalópodos (2.67%) y reptiles
(0.35%). Los machos mostraron mayor preferencia por cefalópodos; mientras que las
hembras por peces.
En México, el estudio de la ecología trófica de C. falciformis se ha realizado
por lo general de manera conjunta con otras especies y muy pocas veces de manera
dirigida. Tal es el caso del estudio de Marín (1992), quien analizó el contenido
estomacal de 27 especies de tiburones en Tamaulipas y Veracruz, México. A partir

4
de 7 estómagos de C. falciformis, determinó que es una especie que se alimenta de
presas epipelágicas, en especial de peces óseos como Sphyraena barracuda, peces
de la familia Scombridae, así como de cefalópodos. Mientras que en el Océano
Pacifico, Galván et al. (1989) estudiaron los hábitos alimenticios de once especies de
tiburones capturados en la Isla Cerralvo, Baja California Sur, México. Entre las
especies que se capturaron con mayor frecuencia de mayo a diciembre estuvo C.
falciformis, el cual se alimentó principalmente de la langostilla Pleuroncodes planipes,
así como de peces y crustáceos bentónicos.
Entre los estudios dirigidos sobre C. falciformis destaca el de Cabrera (2000),
quien analizó los estómagos de 34 organismos capturados en el Golfo de
Tehuantepec. La presa principal fue el cangrejo Portunus xantusii con una
importancia relativa de 99.5%. Debido a que este cangrejo constituyó la mayor parte
de la dieta, se le clasificó a C. falciformis como un depredador especialista. Mientras
que Andrade (2005), analizó la dieta de C. falciformis con un total 258 estómagos
capturados en el Océano Pacifico Oriental. La estructura de tallas de las capturas
estuvo conformada por una talla mínima de 73 cm alcanzando una talla máxima de
250 cm de longitud total (LT), encontrando juveniles (180 organismos) y adultos (78
organismos). El índice de importancia relativa (%IIR) señala que el atún aleta
amarilla Thunnus albacares (66.7%) fue la especie principal encontrando una baja
amplitud trófica (0.05-0.49) y bajo traslapo trófico entre todas las categorías (machos
vs hembras, juveniles vs adultos).
Los estudios mencionados anteriormente destacan, considerando que son los
únicos que realizan un estudio dirigido para entender la dinámica trófica a partir de la
identificación taxonómica de presas extraídas de contenido estomacal de C.
falciformis. Sin embargo, una de las limitantes que se destacan del contenido
estomacal, es que los tiburones tienden a regurgitar el alimento cuando se
encuentran sometidos a estrés de captura, por lo que diferentes autores han optado
por la implementación de una herramienta complementaria como son los isótopos
estables de carbono (δ13C) y nitrógeno (δ15N).

5
Cabrera (2003) analizó la composición de la dieta de 143 organismos de C.
falciformis capturados en la costa occidental de Baja California Sur, con longitudes
totales (LT) entre los 100 y 228 cm. A partir del contenido estomacal se registraron
tres grupos tróficos principales como son cefalópodos, crustáceos y peces teleósteos
de agua oceánicas como Pleuroncodes planipes, Dosidicus gigas y Scomber
japonicus, lo cual fue corroborado a partir de isótopos estables (δ15N = 15.07 a 17.68;
δ13C = -18.51 a -17.12). De acuerdo al Índice de Levin se menciona que C.
falciformis es un depredador especialista. Mientras que el índice Morisita-Horn,
reportó un traslapo alto entre machos juveniles y adultos (0.96), lo cual indica que
consumen presas similares; sin embargo, las hembras juveniles y adultas
presentaron un traslapo bajo (0.33).
Barranco (2008) analizó 258 estómagos de C. falciformis capturados en el
Golfo de Tehuantepec, México, de los cuales 140 fueron machos y 118 hembras con
un intervalo de talla de 69 a 229 cm (LT). Por estado de madurez se registraron 152
juveniles y 106 adultos. Con base en el índice de importancia relativa (IIR) y el
análisis de importancia de especies SIMPER, las presas de mayor contribución
fueron el cangrejo pelágico Portunus xantusii, los cefalópodos del género Argonauta
spp. y el barrilete negro Euthynnus lineatus. A partir del análisis de similitud
(ANOSIM), no se detectaron diferencias significativas entre sexos, pero si entre
estados de madurez (juveniles y adultos). Según el análisis de estrategia alimenticia,
la población del tiburón piloto explota de manera diferencial los recursos dentro de su
nicho trófico y consumen preferentemente a las jaibas pelágicas y argonautas por su
alta disponibilidad, por lo que el depredador fue considerado como oportunista. A
partir de los valores de δ13C (-16.70 a -14.55‰) se determinó que C. falciformis
presenta movimientos continuos entre zonas oceánicas y costeras. La posición
trófica calculada con δ15N indico que es un consumidor terciario (PT = 4.34).

6
3. JUSTIFICACIÓN
La importancia ecológica y pesquera de los tiburones y la disminución en las
capturas a nivel global, han generado un interés común por parte de diferentes
instituciones (e.g. CIAT, CICAA, ICES) para conocer el estado actual de las
poblaciones a partir de estudios biológicos con el fin de establecer medidas de
regulación y conservación efectivas a corto, mediano y largo plazo.
El análisis de la dinámica trófica de una especie en el ecosistema forma parte
de los estudios biológicos a desarrollar en un corto plazo. Estos estudios permiten
conocer las interacciones tróficas que mantiene un depredador con sus presas, lo
que podría indicar posibles cambios en una comunidad como resultado de una
disminución de la población del depredador, así como el impacto que tendría en el
depredador la disminución de algún recurso alimenticio.
En el caso de C. falciformis, estas interacciones han sido poco estudiadas. La
información con la que se tiene en su mayoría solo corresponde a las etapas adultas
en zonas costeras, por lo que se desconoce el comportamiento trófico que mantiene
C. falciformis en otras etapas ontogénicas (neonatos vs. juveniles) en diferentes
zonas. A partir del presente estudio se podrá determinar la existencia de una posible
segregación trófica, lo cual resulta de gran relevancia para entender su papel en la
estructura y funcionamiento de la diversidad biológica.
El análisis de la composición de δ15N y δ13C a partir de diferentes tejidos
(músculo y vértebra) para el estudio de la dinámica trófica de C. falciformis en
diferentes estados ontogénicos, permitirá conocer en qué zona (costeras vs.
oceánicas o bentónicas vs. pelágicas) se alimenta el tiburón piloto a diferentes
escalas de tiempo, así como el papel funcional trófico que desempeña en el Pacífico
Oriental Tropical. En caso del músculo, al ser un tejido metabólicamente activo,
permitirá detectar cambios en la dieta en corto plazo, lo que ayudará a establecer

7
aspectos como la amplitud y traslapo trófico. Mientras que en el caso de la vértebra,
dado que es un tejido metabólicamente inerte con fijación isotópica anual, será reflejo
de un resumen anual de las actividades tróficas del depredador.
A diferencia de los estudios realizados sobre C. falciformis, el presente
estudio: 1) está dirigido a una especie en particular, 2) incluye escalas de muestreo
espacio-temporales mayores, 3) el uso de isótopos estables en diferentes tejidos y 4)
organismos en diferentes estados ontogénicos, lo cual es considerado como esencial
para una correcta interpretación de la dinámica trófica de una especie. En este
contexto, en el presente estudio se analizara la dinámica trofica de C. falciformis con
la metodología más adecuada para contar con información detallada y reciente sobre
una población clasificada como vulnerable y casi amenazada.
4. HIPÓTESIS
Dado que partir del contenido estomacal se han detectados cambios en las
interacciones tróficas durante el desarrollo ontogénicos de C. falciformis, se espera
que los valores isotópicos representen el comportamiento trófico diferencial en el uso
de presas y hábitats, donde organismos adultos presentarán valores enriquecidos de
δ15N y empobrecidos en δ13C en comparación de los neonatos y juveniles, como
resultado de la distribución preferencial en zonas oceánicas y alimentarse de presas
de alto nivel trófico.

8
5. OBJETIVO
5.1 Objetivo General
- Determinar la composición isotópica (δ15N y δ13C) en músculo y vértebra
durante el desarrollo ontogénico del tiburón piloto Carcharhinus falciformis en
el Océano Pacífico Oriental Tropical.
5.2 Objetivos Específicos
- Conocer la composición de δ13C y δ15N en el tiburón piloto (C. falciformis)
entre tejidos, sexos, estado ontogénico y año de captura.
- Determinar la amplitud trófica del tiburón piloto (C. falciformis) a partir de δ13C
y δ15N.
- Determinar el traslapo trófico sexo y estado ontogénico del tiburón piloto (C.
falciformis) a partir de δ13C y δ15N.
- Estimar la posición trófica mediante δ15N de los tiburones piloto por sexo y
estado ontogénico.
- Conocer los patrones de δ13C y δ15N en el tiburón piloto (C. falciformis) a lo
largo de su vida.
9
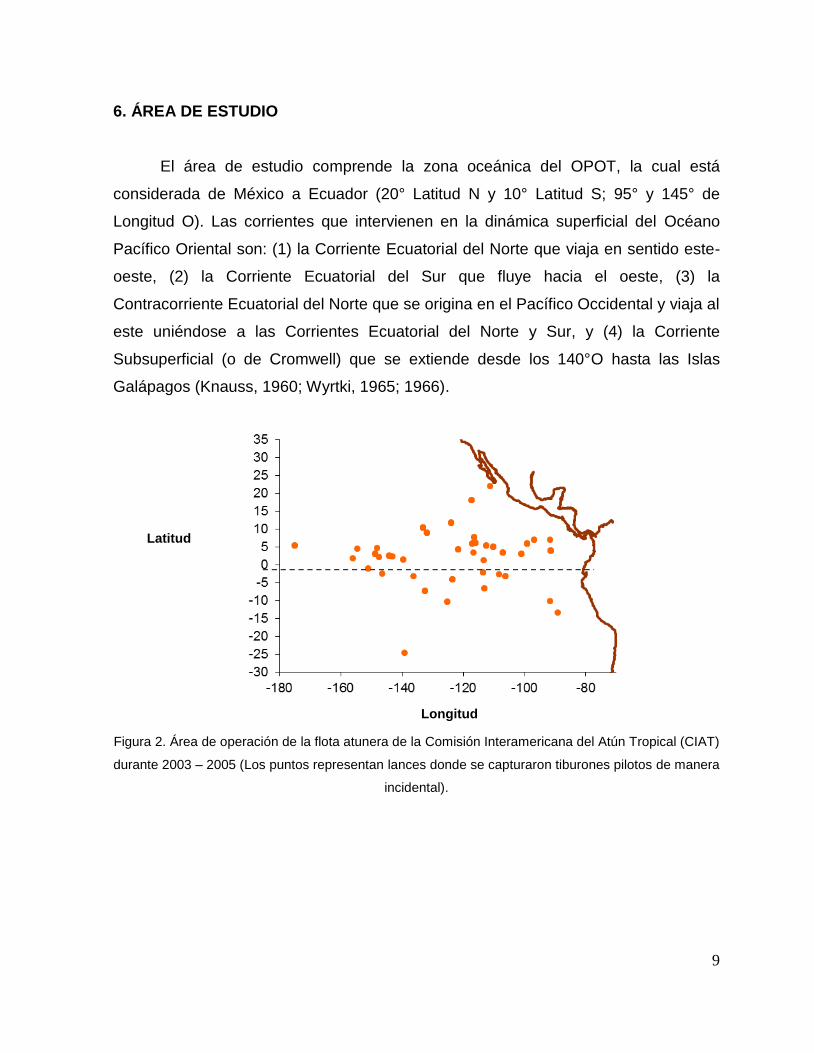
6. ÁREA DE ESTUDIO
El área de estudio comprende la zona oceánica del OPOT, la cual está
considerada de México a Ecuador (20° Latitud N y 10° Latitud S; 95° y 145° de
Longitud O). Las corrientes que intervienen en la dinámica superficial del Océano
Pacífico Oriental son: (1) la Corriente Ecuatorial del Norte que viaja en sentido este-
oeste, (2) la Corriente Ecuatorial del Sur que fluye hacia el oeste, (3) la
Contracorriente Ecuatorial del Norte que se origina en el Pacífico Occidental y viaja al
este uniéndose a las Corrientes Ecuatorial del Norte y Sur, y (4) la Corriente
Subsuperficial (o de Cromwell) que se extiende desde los 140°O hasta las Islas
Galápagos (Knauss, 1960; Wyrtki, 1965; 1966).
Figura 2. Área de operación de la flota atunera de la Comisión Interamericana del Atún Tropical (CIAT)
durante 2003 – 2005 (Los puntos representan lances donde se capturaron tiburones pilotos de manera
incidental).
Latitud
Longitud

10
7. MATERIAL Y MÉTODOS
7.1 Trabajo de campo
Las muestras fueron colectadas por técnicos de la Comisión Interamericana
de Atún Tropical (CIAT) que participaron como observadores a bordo de barcos
atuneros de México y Ecuador entre agosto de 2003 y agosto de 2005. Los tiburones
fueron identificados utilizando las claves de Compagno et al. (1995). Una vez
identificado, se registró la longitud total (“LT” cm) y el sexo. La vértebra y el músculo
fueron colectados a la altura de la primera aleta dorsal, las cuales fueron
almacenadas en bolsas de plástico, especificando el nombre de la especie y número
de organismo correspondiente, así como la fecha y localización de la captura. Las
muestras se mantuvieron congeladas hasta su traslado al Laboratorio de Ecología de
Peces del Centro Interdisciplinario de Ciencias Marinas (CICIMAR-IPN), en la ciudad
de La Paz, B.C.S., México.
7.2 Trabajo de laboratorio
Previo a la manipulación de muestras, se procedió a lavar el material (viales,
bisturí, pizas, charolas, mortero) con un limpiador detergente desinfectante, sin
fosfatos, de uso múltiple (MICRO-CHEM PLUS), diseñado para controlar el peligro
de la contaminación cruzada. Posteriormente fueron enjuagados en agua destilada y
finalmente con una solución 1:1 de cloroformo y hexano, este proceso se repitió entre
cada muestra manipulada.
En el caso del músculo: 1) se descongelaron a temperatura ambiente evitando
una calentamiento de las muestras, para posteriormente cortar aproximadamente 1 g
y colocarlo en viales estériles previamente etiquetados; 2) se extrajo la humedad en
un horno a 50º C por 24 h. hasta que las muestras se encontraron totalmente secas;
3) se maceró la muestra mediante la ayuda de un mortero de ágata hasta obtener un

11
residuo homogéneo; y 4) se utilizó una balanza analítica (OHAUS) para pesar
alrededor de 1 mg de muestra el cual fue depositado en cápsulas de estaño (8 x 5
mm).
En el caso de las vértebras, se eligieron aquellas vertebras pertenecientes a
organismos de grandes tallas y que presentaban un mayor número de anillos, ya que
éstos aportarían información de un número mayor de etapas ontogénicas (incluyendo
el foco o centro de la vértebra del cual parte el estado embrionario, neonato, juvenil y
adulto). Las muestras fueron descongeladas y limpiadas de forma manual con la
ayuda de un bisturí y agua destilada, removiendo el arco neural y el tejido conjuntivo.
Cuando las vértebras se encontraron totalmente limpias, se procedió a extraer
el tejido vertebral; para ello se utilizó un Microtaladro (Proxxon) con una broca de un
diámetro de 0.5 mm. Se hicieron perforaciones superficiales desde el foco hacia el
borde de la vértebra (Fig. 3)
Figura 3. Vista de la vértebra con perforaciones, que representan las etapas o estado ontogénico del
tiburón piloto (C. falciformis), desde el foco hasta la parte más distal (foco=0, embrión=1, neonato=2,
juvenil=3 y adulto=4)
Como se observó que la cantidad de muestra que se obtenía de las
perforaciones era reducida, se realizó una prueba con 20 muestras para conocer si la
cantidad de tejido vertebral extraído en cada perforación era suficiente, para que el

12
espectrómetro de masas pudiera obtener los valores isotópicos correctos. Ya que
dichas pruebas dieron positivas, el polvo de las perforaciones (0.6 a 0.8 mg) fue
colocado directamente en la capsula de estaño ayudando a que no se perdiera
material y así se evitó el pesaje de las mismas.
El polvo de las muestras se procesaron en el Laboratorio de Biogeoquímica de
Isótopos Estables del Instituto Andaluz de Ciencias de La Tierra (CSIC-URG) en
Granada, España, para su análisis isotópico en un espectrómetro de masas de flujo
continuo, con periféricos acoplados (Analizador Elemental Carlo Elba 1500NC). Los
valores de δ se obtuvieron mediante la siguiente fórmula (Park y Epstein, 1961):
δ X (‰) = [(R muestra/R estándar)-1] x 1000
Donde X es la proporción en unidades delta relativa a un estándar y R muestra
y R estándar son las proporciones isotópicas absolutas de la muestra y el estándar.
Al multiplicar por 1000 los valores son expresados en partes por mil (‰). El estándar
para el δ13C fue el PDB (Pee Dee Belemnita) con una precisión de 0.05 a 0.1;
mientras que para el δ15N el N2 atmosférico, con una precisión de 0.1
7.3 Trabajo de escritorio
Par el análisis de datos, se separaron los valores de δ13C y δ15N en
categorías. Para músculo se separó entre sexos, estado ontogénico y año de captura
de acuerdo a Hoyos (2003), quien menciona que organismos entre 63 y 75 cm de
longitud total (LT) corresponden a neonatos, organismos entre los 75 y 180 cm de LT
corresponden a juveniles, mientras que mayores a 180 cm de LT corresponden a
organismos adultos. También se realizó una combinación de categorías sexo/estado
ontogénico (macho neonato, macho juvenil, macho adulto, hembra neonato, hembra
juvenil y hembra adulta).

13
Para vértebra, las categorías fueron sexos; y estado ontogénico, esta última
categoría se dividió de la siguiente manera: Foco o centro de la vértebra (primera
perforación), periodo de embrión (segunda perforación, antes de la marca de
nacimiento), periodo de neonato (tercera perforación, en la marca de nacimiento),
juvenil (incluye todas las perforaciones restantes para organismos menores a 180 cm
de LT), periodo adulto (organismos de 180 a 190 cm LT), se tomo la última
perforación y para organismos mayores a 190 cm LT se consideraron las dos últimas
perforaciones) (Figura 3).
7.3.1 Relación C: N
Se graficó la relación C: N (eje Y) contra los valores de δ13C (eje x) entre estado
ontogénico. Teóricamente, se espera que los resultados obtenidos de esta razón
oscilen entre 2.9 y 3.5 (valor máximo de proteína pura). En caso de exceder este
valor, indicaría que la muestra presenta lípidos, los cuales probablemente estén
sesgando hacia valores negativos de δ13C (Mcconnaughey y McRoy, 1979; Carlson
et al., 2004).
7.3.2 Composición de δ13C y δ15N
Para ambos tejidos se aplicaron pruebas de normalidad (Shapiro - Wilks) y
homogeneidad de varianzas (Bartlett’s). Dado que en algunos casos se cumplían
ambos principios y en otros no y se aplicaron tanto pruebas paramétricas como no
paramétricas. El nivel de significancia utilizado para todas las pruebas fue de 0.05,
las pruebas estadísticas para cada categoría y tejido se especifica a continuación.
Dado que los datos isotópicos entre sexos presentaron una distribución
normal, pero no homogeneidad de varianzas. Para determinar si existen diferencias
significativas en los valores de δ13C y δ15N de músculo se utilizó la prueba de Mann-
Whitney (prueba no paramétrica “U”), mientras que entre estados ontogénicos

14
(neonato, juvenil y adulto), para δ13C se aplicó Kruskal-Wallis (prueba no paramétrica
“H”), mientras que para δ15N se aplicó un análisis de varianza de un factor (ANOVA
prueba paramétrica “F”), debido a que los datos son normales y homogéneos. Para
la categoría “sexo/estado ontogénico” se aplicó para δ13C una prueba Kruskal-Wallis
y para δ15N una ANOVA de un factor. En los casos donde se detectó diferencias
significativas se aplicaron pruebas a posteriori (prueba de Tukey para análisis
paramétricos y prueba Dunn para análisis no paramétrico). Por último, en los análisis
para detectar si existen diferencias a nivel isotópico en los años de captura se aplicó
para δ13C Kruskal-Wallis y para δ15N una ANOVA de un factor.
En las vértebras los valores entre sexos presentaron una distribución normal
pero no homogeneidad de varianzas, por lo que se realizó una prueba no
paramétrica (Mann-Whitney) para δ13C y δ15N. Mientras que entre estados
ontogénicos para δ13C y δ15N se aplicó una ANOVA de un factor y en el caso de
detectar diferencias se aplicó una prueba a posteriori de Tukey. Finalmente para los
años de captura se aplicó la prueba no paramétrica de Kruskal-Wallis para ambos
isotopos. Se realizaron tablas y gráficas donde se pueden ver representados los
valores de δ15N y δ13C, en ambos tejidos. Estadísticos y gráficos fueron realizados en
el lenguaje de programación R versión 2.15.2.
7.3.3 Amplitud y Traslapo trófico.
Se determinó la amplitud de la dieta y traslapo trófico para ambos tejidos en cada
una de sus categorías a partir del método SIBER del paquete SIAR propuesto por
Jackson et al. (2011), disponible en el programa R (R Development Core Team,
2007). El programa SIAR realiza una secuencia donde calcula probabilidades
estimando cuantitativamente la amplitud del nicho trofico la cual se refiere al área del
polígono que utiliza los valores más extremos de la muestra, mientras que las elipses
muestran el traslapo que hay entre categorías.

15
El área corregida de la elipse tiene en cuenta el tamaño de la muestra y la estructura
de los datos a partir de una matriz de covarianza seleccionada de forma aleatoria
que define la forma y área del mismo, donde valores menores a 1 corresponden a
organismos o poblaciones de hábitos especialistas, mientras que los valores
mayores a 1 corresponden a hábitos generalistas (Bearhop et al., 2004). Mientras
que para el traslapo trófico, la media de los datos determina su ubicación y el
traslapo entre las elipses, considerando que los valores cercanos a 1 o mayores
como un traslapo alto y menores a 1 un traslapo bajo (Jackson et al., 2011).
7.3.4 Estimación de la posición trófica
Con el objetivo de conocer la posición trófica de C. falciformis mediante los valores
de δ15N en músculo y sus posibles variaciones en cuanto a sexos y etapas
ontogénicas se aplicó la ecuación propuesta por Post (2002):
PT = λ + (δ15N Depredador - δ15N Base) / Δn
Donde:
λ: posición trófica de la base.
Δn: valor teórico de enriquecimiento en 15N por nivel trófico (3.1‰,Vanderklift y
Ponsard, 2003).
δ15N Depredador δ15N para el depredador.
δ15N Base: determinación de δ15N para la base.
Los valores de δ15N para la base se determinó por 12 especies dominantes de
copépodos en la zona del Pacifico Oriental Tropical del año 2003 A estas especies
de copépodos se les asignó una posición trófica (λ) de 2 y con un valor de δ15N base
de 9‰ (López, 2008).

16
7.3.5 Patrones de δ13C y δ15N durante el ciclo de vida del tiburón piloto
En el caso específico de la vértebra, para detectar posibles patrones de
comportamiento trófico a lo largo del ciclo de vida de C. falciformis entre machos y
hembras, los resultados de δ13C y δ15N de las 44 vértebras se agruparon desde la
primera perforación (corresponde al foco) hasta la última perforación (corresponde al
último anillo de crecimiento), estas perforaciones representan el estado ontogénico
de los individuos.
En el caso de las hembras para conocer si existen diferencias significativas
con respecto a las zonas de alimentación y sus presas a lo largo del ciclo de vida, los
valores de δ13C fueron comparados con un ANOVA de un factor, mientras que δ15N
con Kruskal-Wallis. Para los machos, se aplicó un ANOVA de una vía para δ13C y
δ15N respectivamente. Los gráficos y estadísticos se realizaron en el lenguaje de
programación R versión 2.15.2. Finalmente se graficaron perfiles isotópicos
individuales de δ13C y δ15N por sexos, donde se registro el número de cada individuo
que se puede identificar en una tabla la cual muestra la longitud total, la edad
determinada por las bandas de crecimiento (Cervantes, 2013), fecha y sitio de
captura. Las graficas indican los cambios isotópicos de cada tiburón a lo largo de sus
distintos estados ontogénicos.
8. RESULTADOS
Se analizaron un total de 159 muestras de músculo dorsal y 44 vértebras de tiburón
piloto, capturados en un periodo de 3 años (2003 - 2005). En el caso de las muestras
de músculo, los organismos capturados correspondían a tres etapas ontogénicas
(neonato, juvenil y adulto). En el caso de las vértebras se seleccionaron los
organismos de mayor tamaño, los cuales corresponden a 20 juveniles y 24 adultos
(Tabla 1).
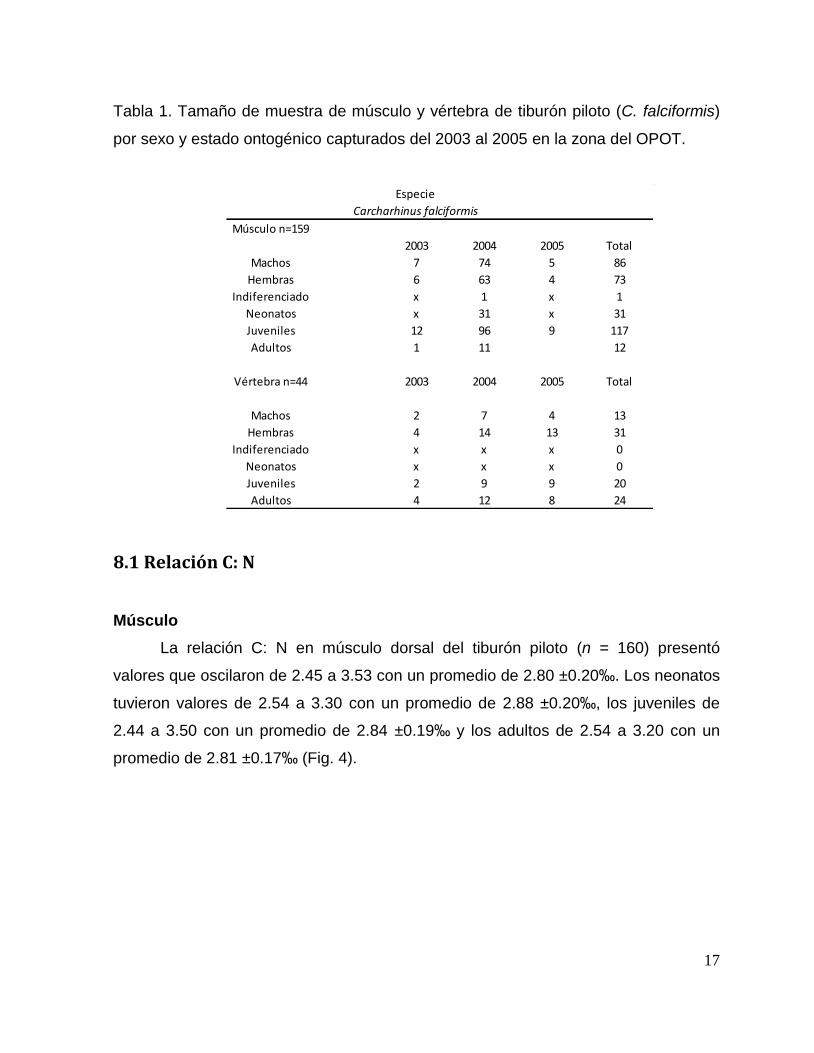
17
Tabla 1. Tamaño de muestra de músculo y vértebra de tiburón piloto (C. falciformis)
por sexo y estado ontogénico capturados del 2003 al 2005 en la zona del OPOT.
Especie
Carcharhinus falciformis
Músculo n=159
2003 2004 2005 Total
Machos 7 74 5 86
Hembras 6 63 4 73
Indiferenciado x 1 x 1
Neonatos x 31 x 31
Juveniles 12 96 9 117
Adultos 1 11 12
Vértebra n=44 2003 2004 2005 Total
Machos 2 7 4 13
Hembras 4 14 13 31
Indiferenciado x x x 0
Neonatos x x x 0
Juveniles 2 9 9 20
Adultos 4 12 8 24
8.1 Relación C: N
Músculo
La relación C: N en músculo dorsal del tiburón piloto (n = 160) presentó
valores que oscilaron de 2.45 a 3.53 con un promedio de 2.80 ±0.20‰. Los neonatos
tuvieron valores de 2.54 a 3.30 con un promedio de 2.88 ±0.20‰, los juveniles de
2.44 a 3.50 con un promedio de 2.84 ±0.19‰ y los adultos de 2.54 a 3.20 con un
promedio de 2.81 ±0.17‰ (Fig. 4).
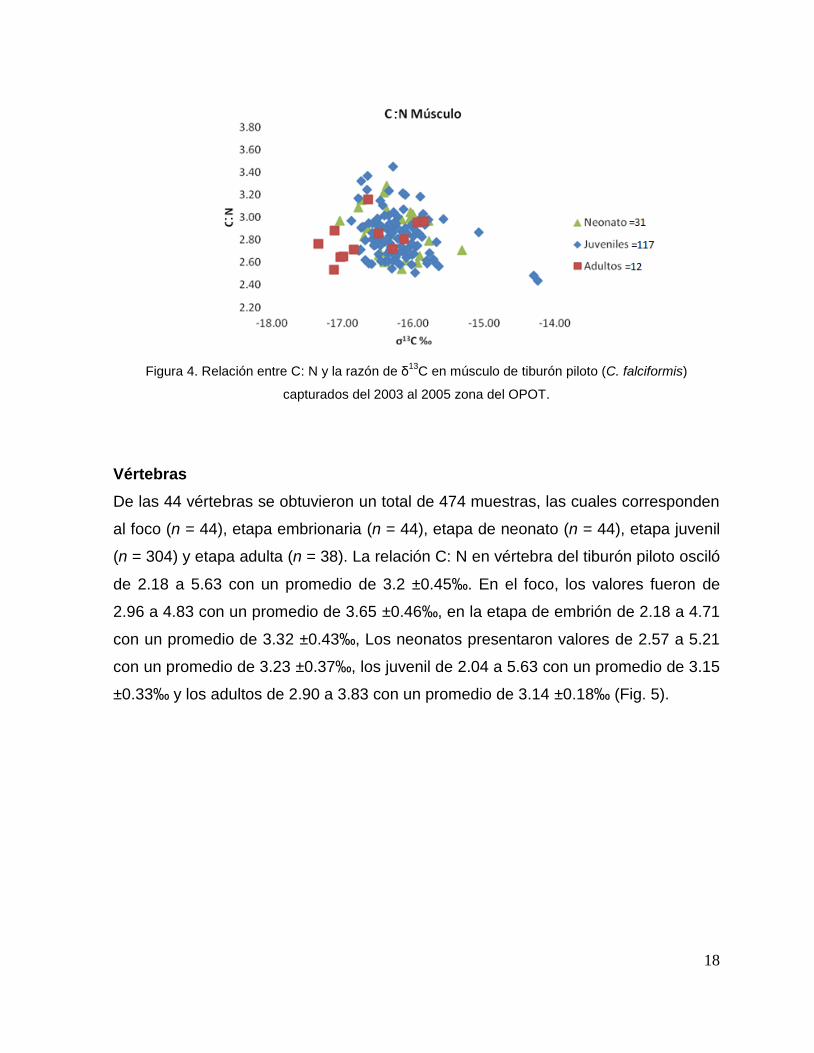
18
Figura 4. Relación entre C: N y la razón de δ13
C en músculo de tiburón piloto (C. falciformis)
capturados del 2003 al 2005 zona del OPOT.
Vértebras
De las 44 vértebras se obtuvieron un total de 474 muestras, las cuales corresponden
al foco (n = 44), etapa embrionaria (n = 44), etapa de neonato (n = 44), etapa juvenil
(n = 304) y etapa adulta (n = 38). La relación C: N en vértebra del tiburón piloto osciló
de 2.18 a 5.63 con un promedio de 3.2 ±0.45‰. En el foco, los valores fueron de
2.96 a 4.83 con un promedio de 3.65 ±0.46‰, en la etapa de embrión de 2.18 a 4.71
con un promedio de 3.32 ±0.43‰, Los neonatos presentaron valores de 2.57 a 5.21
con un promedio de 3.23 ±0.37‰, los juvenil de 2.04 a 5.63 con un promedio de 3.15
±0.33‰ y los adultos de 2.90 a 3.83 con un promedio de 3.14 ±0.18‰ (Fig. 5).
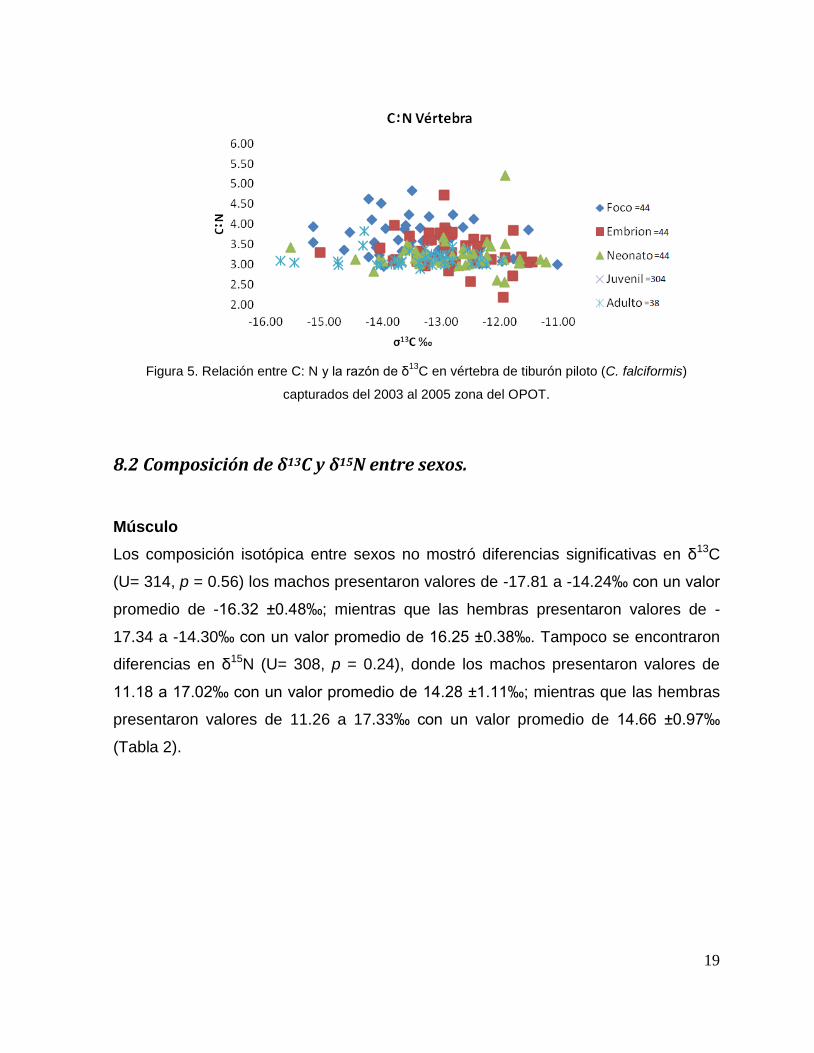
19
Figura 5. Relación entre C: N y la razón de δ13
C en vértebra de tiburón piloto (C. falciformis)
capturados del 2003 al 2005 zona del OPOT.
8.2 Composición de δ13C y δ15N entre sexos.
Músculo
Los composición isotópica entre sexos no mostró diferencias significativas en δ13C
(U= 314, p = 0.56) los machos presentaron valores de -17.81 a -14.24‰ con un valor
promedio de -16.32 ±0.48‰; mientras que las hembras presentaron valores de -
17.34 a -14.30‰ con un valor promedio de 16.25 ±0.38‰. Tampoco se encontraron
diferencias en δ15N (U= 308, p = 0.24), donde los machos presentaron valores de
11.18 a 17.02‰ con un valor promedio de 14.28 ±1.11‰; mientras que las hembras
presentaron valores de 11.26 a 17.33‰ con un valor promedio de 14.66 ±0.97‰
(Tabla 2).
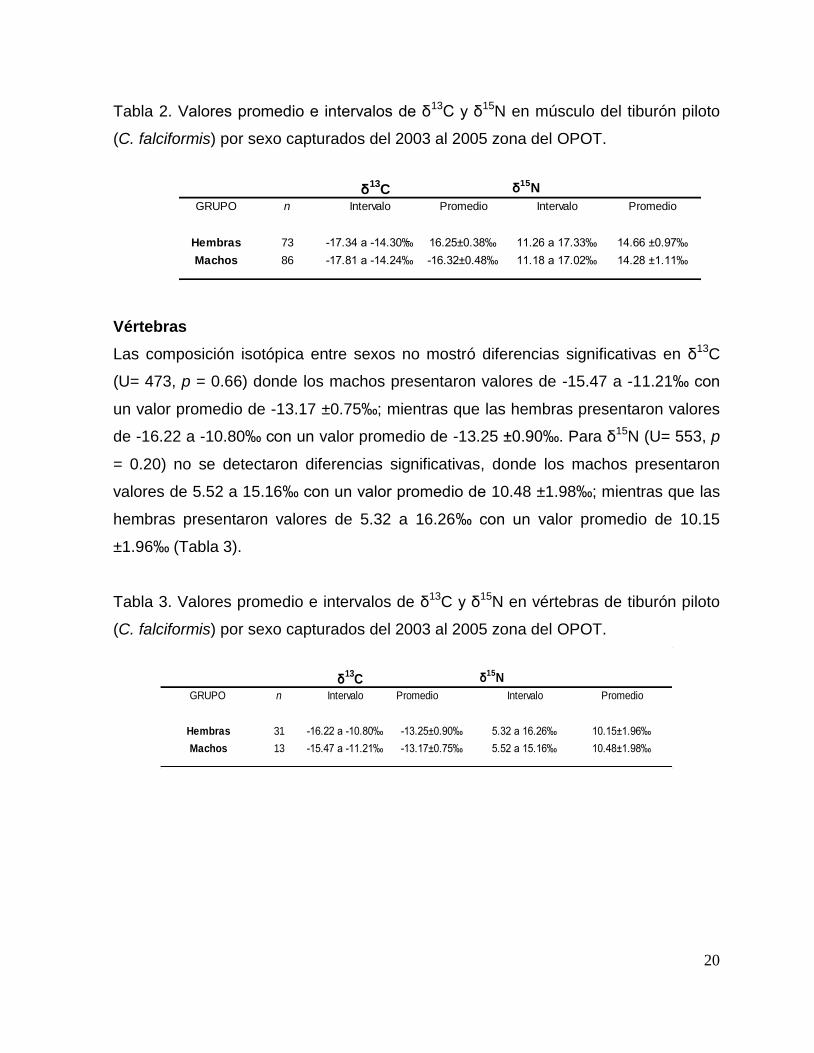
20
Tabla 2. Valores promedio e intervalos de δ13C y δ15N en músculo del tiburón piloto
(C. falciformis) por sexo capturados del 2003 al 2005 zona del OPOT.
δ15N
GRUPO n Intervalo Promedio Intervalo Promedio
Hembras 73 -17.34 a -14.30‰ 16.25±0.38‰ 11.26 a 17.33‰ 14.66 ±0.97‰
11.18 a 17.02‰ 14.28 ±1.11‰-16.32±0.48‰-17.81 a -14.24‰86Machos
δ13
C
Vértebras
Las composición isotópica entre sexos no mostró diferencias significativas en δ13C
(U= 473, p = 0.66) donde los machos presentaron valores de -15.47 a -11.21‰ con
un valor promedio de -13.17 ±0.75‰; mientras que las hembras presentaron valores
de -16.22 a -10.80‰ con un valor promedio de -13.25 ±0.90‰. Para δ15N (U= 553, p
= 0.20) no se detectaron diferencias significativas, donde los machos presentaron
valores de 5.52 a 15.16‰ con un valor promedio de 10.48 ±1.98‰; mientras que las
hembras presentaron valores de 5.32 a 16.26‰ con un valor promedio de 10.15
±1.96‰ (Tabla 3).
Tabla 3. Valores promedio e intervalos de δ13C y δ15N en vértebras de tiburón piloto
(C. falciformis) por sexo capturados del 2003 al 2005 zona del OPOT.
δ15N
GRUPO n Intervalo Promedio
Hembras 31
Intervalo Promedio
5.32 a 16.26‰ 10.15±1.96‰
5.52 a 15.16‰ 10.48±1.98‰
-16.22 a -10.80‰
-15.47 a -11.21‰
-13.25±0.90‰
-13.17±0.75‰13Machos
δ13
C
21
8.3 Composición de δ13C y δ15N entre estados ontogénicos.
Músculo
Los composición isotópica por estado ontogénico mostró diferencias significativas en
δ13C (H = 6.78, p = 0.03) en específico entre juveniles y adultos (Tukey p < 0.05). Los
adultos presentaron valores de δ13C de -17.34‰ a -15.86‰ con un promedio de -
16.65 ±0.50‰ y los juveniles de -17.63 a -15.31‰ con un promedio de -16.24
±0.69‰. Los valores de δ15N también mostraron diferencias estadísticas (F= 6.26, p=
< 0.05), en este caso los grupos que presentaron las diferencias fueron juveniles y
neonatos (Tukey p < 0.05), donde los neonatos presentaron valores de δ15N de 11.89
a 15.82‰ con un valor promedio de 13.87 ±0.50‰ y los juveniles de 11.18 a 17.33‰
con un valor promedio de 14.63 ±1.03‰ (Tabla 4 y Fig.6).
Tabla 4. Valores promedio e intervalos de δ13C y δ15N en músculo de tiburón piloto
(C. falciformis) por estados ontogénicos (neonatos, juveniles y adultos) capturados
del 2003 al 2005 zona del OPOT.
δ15N
Grupo n Intervalo Intervalo
Neonatos 31 -17.81 a -15.24‰ 11.89 a 15.82‰
Juveniles 117 -17.63 a -15.31‰ 11.18 a 17.33‰
Adultos 12 11.18 a 15.74‰
13.87 ±0.50‰
14.63±1.03‰
14.33 ±1.29‰
-16.24 ±0.69‰
-17.34 a -15.86‰ -16.65±0.50‰
δ13C
Promedio
-16.35 ±0.50‰
Promedio
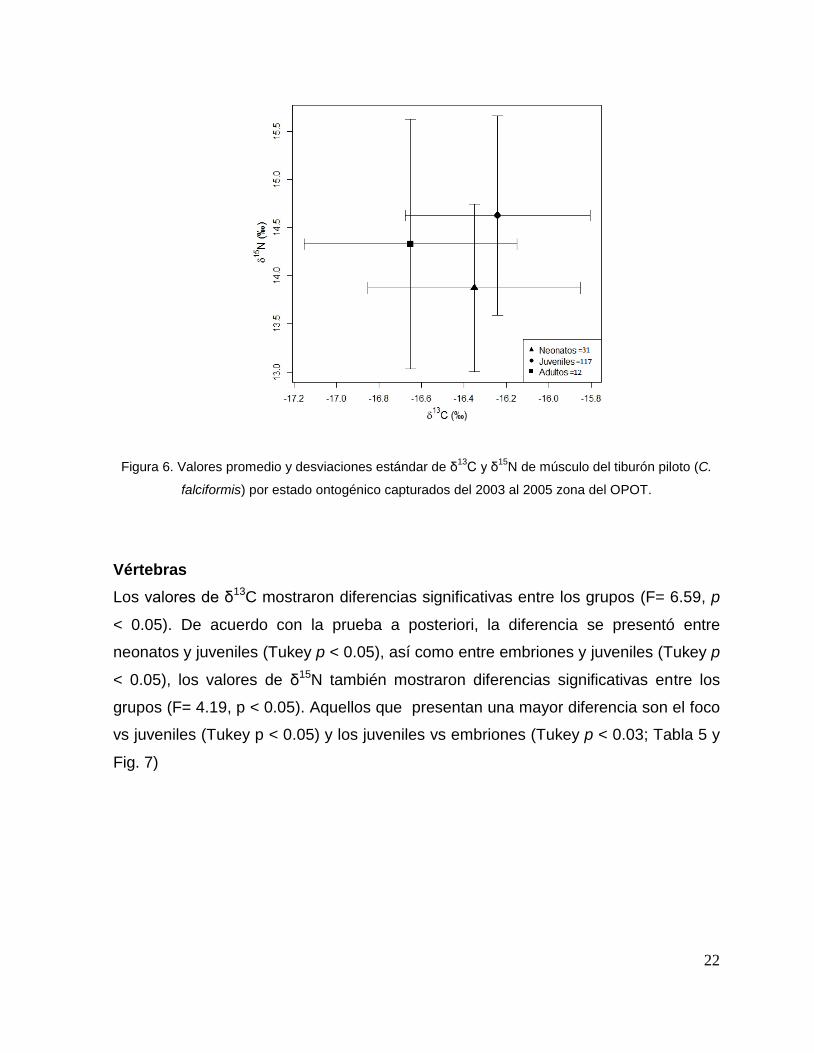
22
Figura 6. Valores promedio y desviaciones estándar de δ13
C y δ15
N de músculo del tiburón piloto (C.
falciformis) por estado ontogénico capturados del 2003 al 2005 zona del OPOT.
Vértebras
Los valores de δ13C mostraron diferencias significativas entre los grupos (F= 6.59, p
< 0.05). De acuerdo con la prueba a posteriori, la diferencia se presentó entre
neonatos y juveniles (Tukey p < 0.05), así como entre embriones y juveniles (Tukey p
< 0.05), los valores de δ15N también mostraron diferencias significativas entre los
grupos (F= 4.19, p < 0.05). Aquellos que presentan una mayor diferencia son el foco
vs juveniles (Tukey p < 0.05) y los juveniles vs embriones (Tukey p < 0.03; Tabla 5 y
Fig. 7)
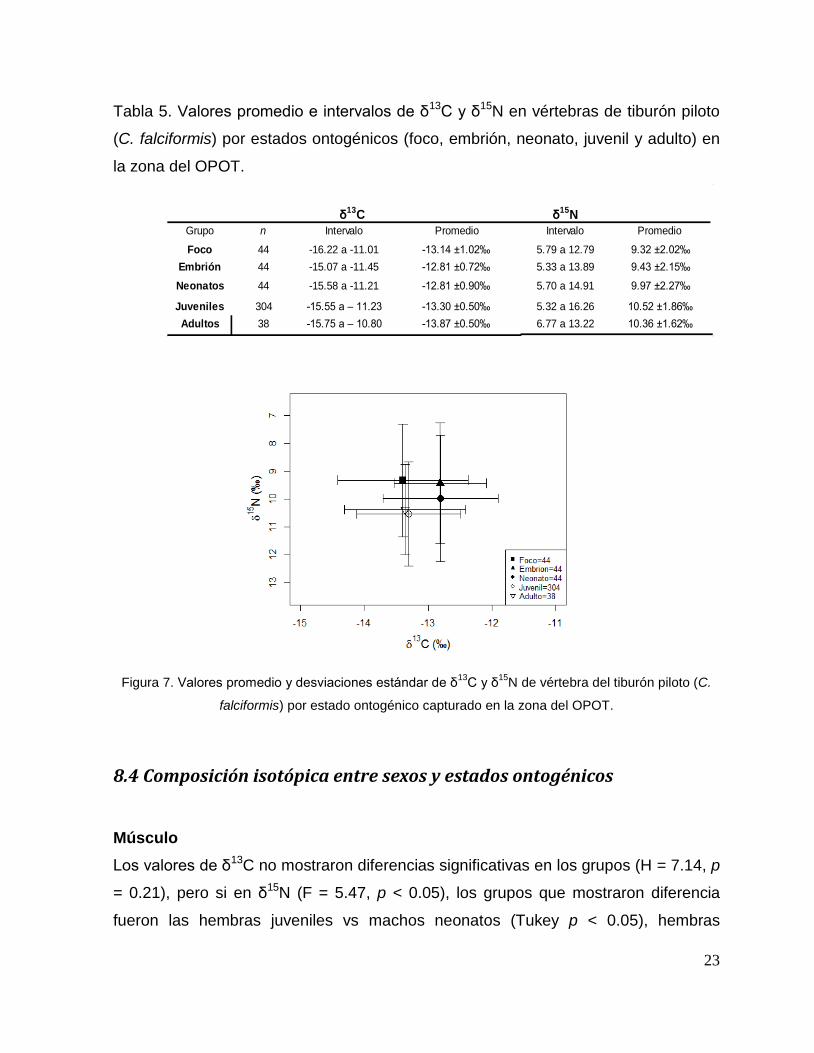
23
Tabla 5. Valores promedio e intervalos de δ13C y δ15N en vértebras de tiburón piloto
(C. falciformis) por estados ontogénicos (foco, embrión, neonato, juvenil y adulto) en
la zona del OPOT.
δ15N
Grupo n Intervalo Intervalo
Foco 44 -16.22 a -11.01 5.79 a 12.79
Embrión 44 -15.07 a -11.45 5.33 a 13.89
Neonatos 44 -15.58 a -11.21 5.70 a 14.91
Juveniles 304 -15.55 a – 11.23 5.32 a 16.26
38 6.77 a 13.22
Promedio
9.32 ±2.02‰
9.43 ±2.15‰
9.97 ±2.27‰
10.52 ±1.86‰
10.36 ±1.62‰
-12.81 ±0.72‰
-12.81 ±0.90‰
-13.30 ±0.50‰
Adultos -15.75 a – 10.80 -13.87 ±0.50‰
δ13C
Promedio
-13.14 ±1.02‰
Figura 7. Valores promedio y desviaciones estándar de δ13
C y δ15
N de vértebra del tiburón piloto (C.
falciformis) por estado ontogénico capturado en la zona del OPOT.
8.4 Composición isotópica entre sexos y estados ontogénicos
Músculo
Los valores de δ13C no mostraron diferencias significativas en los grupos (H = 7.14, p
= 0.21), pero si en δ15N (F = 5.47, p < 0.05), los grupos que mostraron diferencia
fueron las hembras juveniles vs machos neonatos (Tukey p < 0.05), hembras
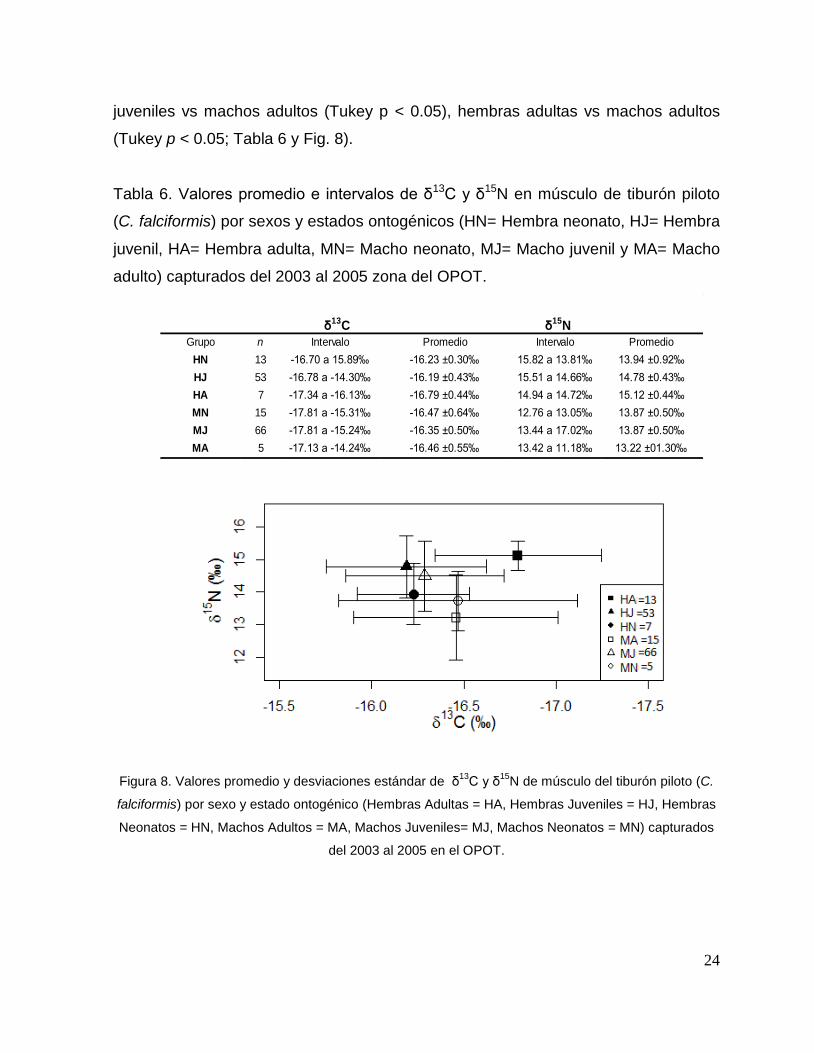
24
juveniles vs machos adultos (Tukey p < 0.05), hembras adultas vs machos adultos
(Tukey p < 0.05; Tabla 6 y Fig. 8).
Tabla 6. Valores promedio e intervalos de δ13C y δ15N en músculo de tiburón piloto
(C. falciformis) por sexos y estados ontogénicos (HN= Hembra neonato, HJ= Hembra
juvenil, HA= Hembra adulta, MN= Macho neonato, MJ= Macho juvenil y MA= Macho
adulto) capturados del 2003 al 2005 zona del OPOT.
δ15N
n Intervalo Intervalo
13 -16.70 a 15.89‰ 15.82 a 13.81‰
53 -16.78 a -14.30‰ 15.51 a 14.66‰
7 -17.34 a -16.13‰ 14.94 a 14.72‰
15 -17.81 a -15.31‰ 12.76 a 13.05‰
66 -17.81 a -15.24‰ 13.44 a 17.02‰
5 -17.13 a -14.24‰ 13.42 a 11.18‰ 13.22 ±01.30‰
Promedio
15.12 ±0.44‰
13.87 ±0.50‰
13.87 ±0.50‰
13.94 ±0.92‰
14.78 ±0.43‰
MN -16.47 ±0.64‰
MJ -16.35 ±0.50‰
MA -16.46 ±0.55‰
HN -16.23 ±0.30‰
HJ -16.19 ±0.43‰
HA -16.79 ±0.44‰
δ13C
Grupo Promedio
Figura 8. Valores promedio y desviaciones estándar de δ13
C y δ15
N de músculo del tiburón piloto (C.
falciformis) por sexo y estado ontogénico (Hembras Adultas = HA, Hembras Juveniles = HJ, Hembras
Neonatos = HN, Machos Adultos = MA, Machos Juveniles= MJ, Machos Neonatos = MN) capturados
del 2003 al 2005 en el OPOT.
25
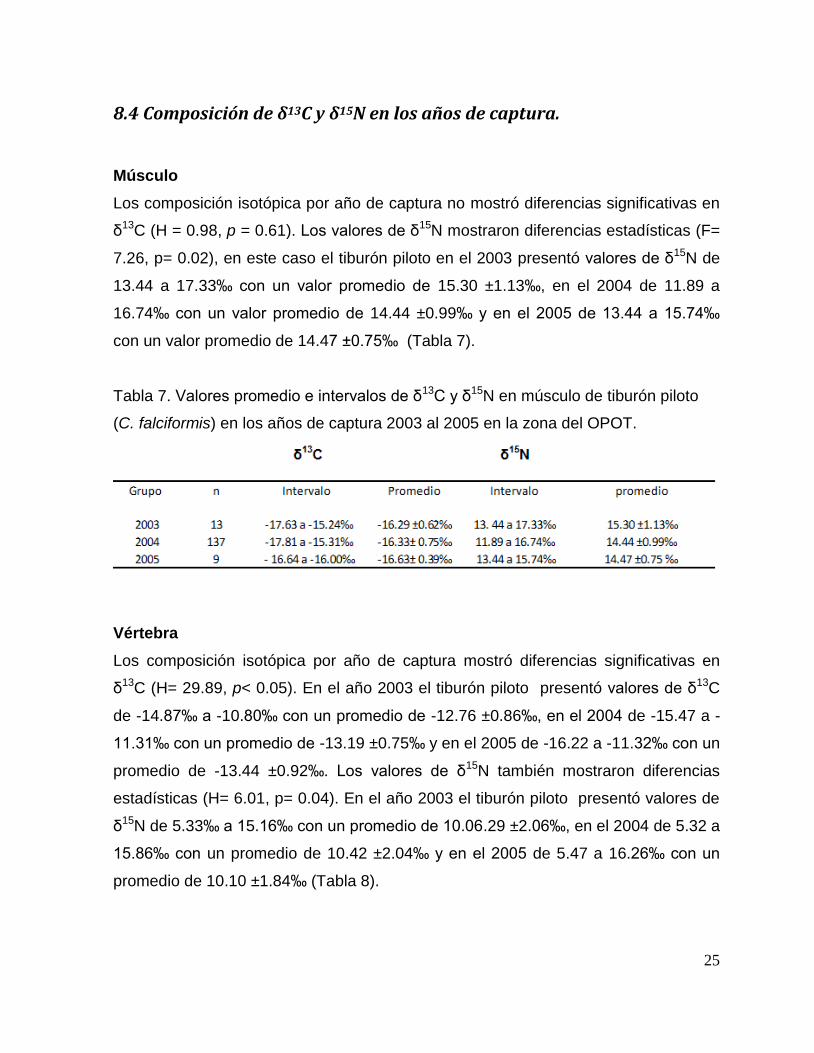
8.4 Composición de δ13C y δ15N en los años de captura.
Músculo
Los composición isotópica por año de captura no mostró diferencias significativas en
δ13C (H = 0.98, p = 0.61). Los valores de δ15N mostraron diferencias estadísticas (F=
7.26, p= 0.02), en este caso el tiburón piloto en el 2003 presentó valores de δ15N de
13.44 a 17.33‰ con un valor promedio de 15.30 ±1.13‰, en el 2004 de 11.89 a
16.74‰ con un valor promedio de 14.44 ±0.99‰ y en el 2005 de 13.44 a 15.74‰
con un valor promedio de 14.47 ±0.75‰ (Tabla 7).
Tabla 7. Valores promedio e intervalos de δ13C y δ15N en músculo de tiburón piloto
(C. falciformis) en los años de captura 2003 al 2005 en la zona del OPOT.
Vértebra
Los composición isotópica por año de captura mostró diferencias significativas en
δ13C (H= 29.89, p< 0.05). En el año 2003 el tiburón piloto presentó valores de δ13C
de -14.87‰ a -10.80‰ con un promedio de -12.76 ±0.86‰, en el 2004 de -15.47 a -
11.31‰ con un promedio de -13.19 ±0.75‰ y en el 2005 de -16.22 a -11.32‰ con un
promedio de -13.44 ±0.92‰. Los valores de δ15N también mostraron diferencias
estadísticas (H= 6.01, p= 0.04). En el año 2003 el tiburón piloto presentó valores de
δ15N de 5.33‰ a 15.16‰ con un promedio de 10.06.29 ±2.06‰, en el 2004 de 5.32 a
15.86‰ con un promedio de 10.42 ±2.04‰ y en el 2005 de 5.47 a 16.26‰ con un
promedio de 10.10 ±1.84‰ (Tabla 8).
26
Tabla 8. Valores promedio e intervalos de δ13C y δ15N en vértebra de tiburón piloto
(C. falciformis) en los años de captura 2003 al 2005 en la zona del OPOT.
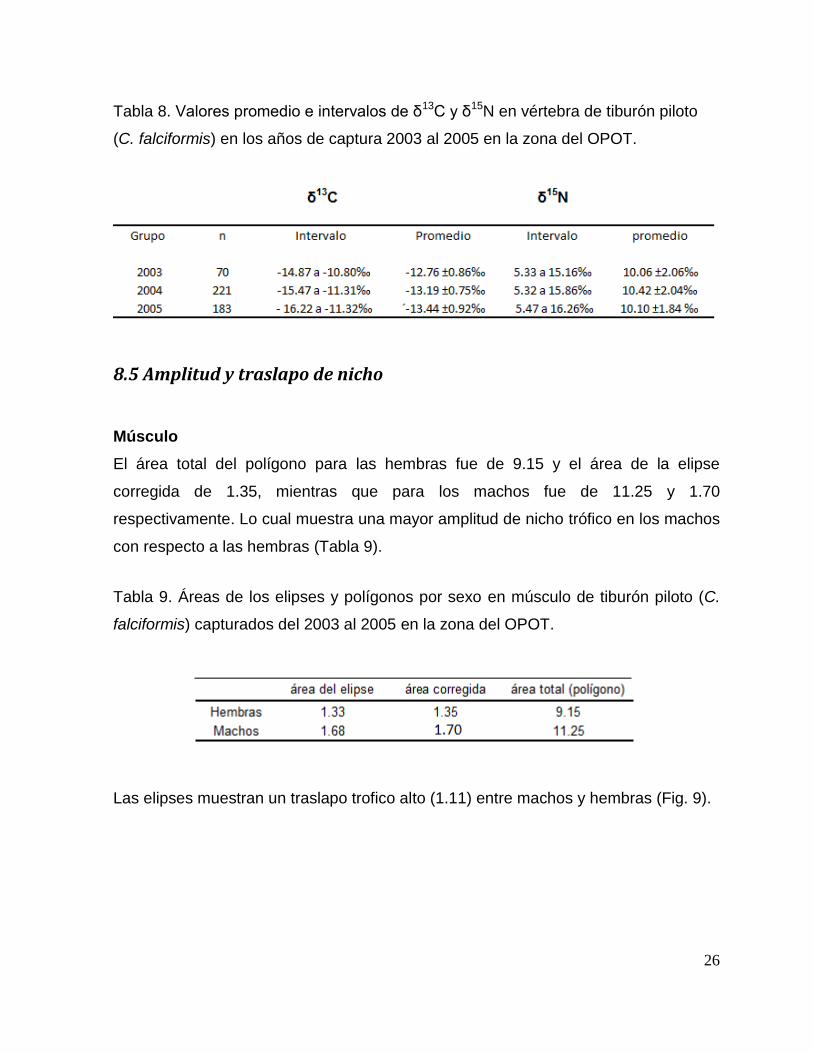
8.5 Amplitud y traslapo de nicho
Músculo
El área total del polígono para las hembras fue de 9.15 y el área de la elipse
corregida de 1.35, mientras que para los machos fue de 11.25 y 1.70
respectivamente. Lo cual muestra una mayor amplitud de nicho trófico en los machos
con respecto a las hembras (Tabla 9).
Tabla 9. Áreas de los elipses y polígonos por sexo en músculo de tiburón piloto (C.
falciformis) capturados del 2003 al 2005 en la zona del OPOT.
Las elipses muestran un traslapo trofico alto (1.11) entre machos y hembras (Fig. 9).
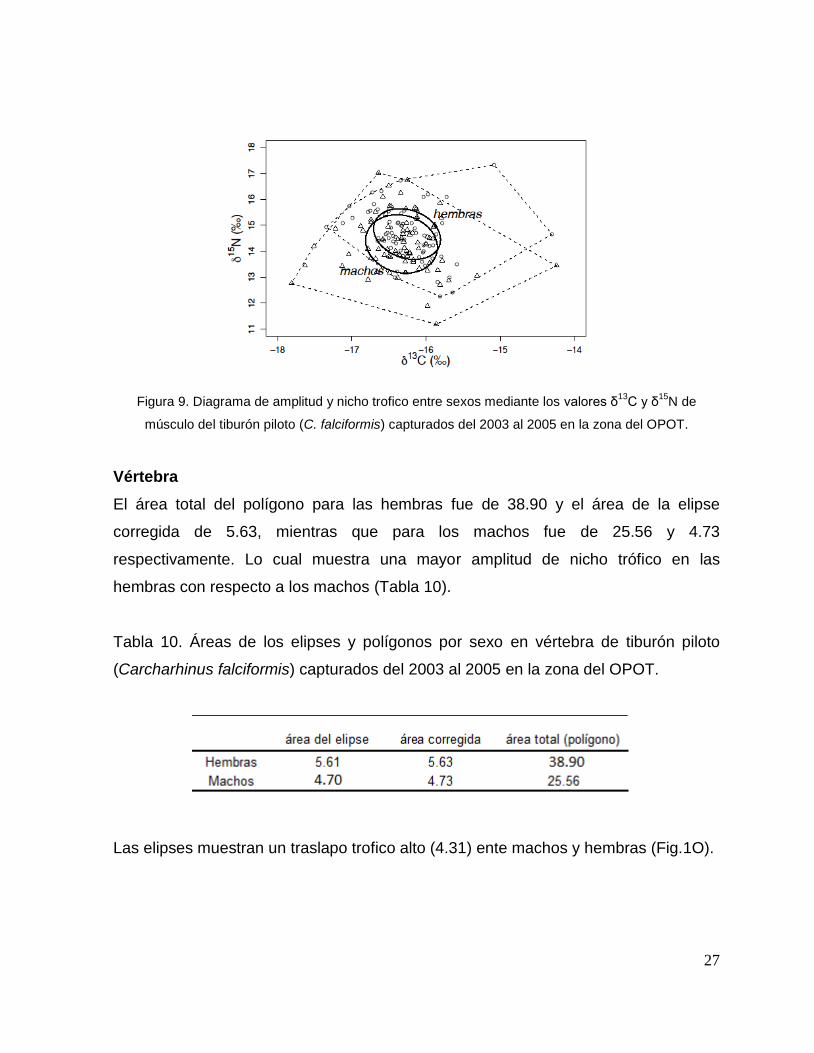
27
Figura 9. Diagrama de amplitud y nicho trofico entre sexos mediante los valores δ13
C y δ15
N de
músculo del tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona del OPOT.
Vértebra
El área total del polígono para las hembras fue de 38.90 y el área de la elipse
corregida de 5.63, mientras que para los machos fue de 25.56 y 4.73
respectivamente. Lo cual muestra una mayor amplitud de nicho trófico en las
hembras con respecto a los machos (Tabla 10).
Tabla 10. Áreas de los elipses y polígonos por sexo en vértebra de tiburón piloto
(Carcharhinus falciformis) capturados del 2003 al 2005 en la zona del OPOT.
Las elipses muestran un traslapo trofico alto (4.31) ente machos y hembras (Fig.1O).
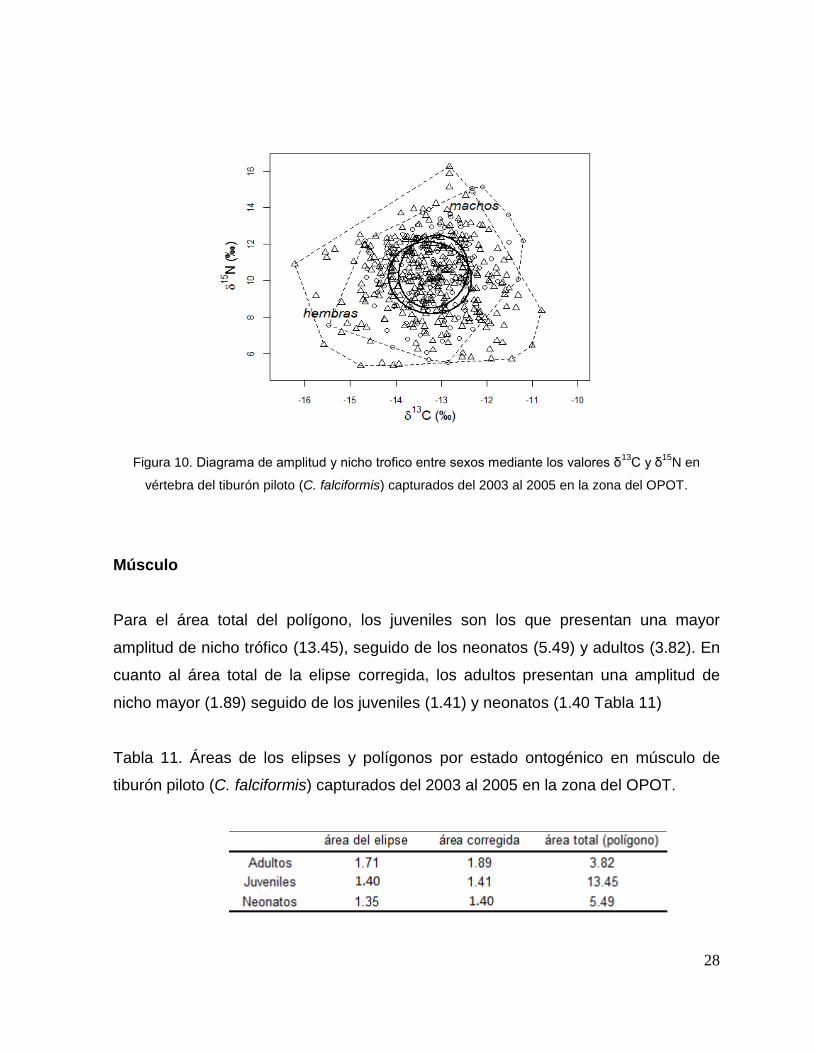
28
Figura 10. Diagrama de amplitud y nicho trofico entre sexos mediante los valores δ13
C y δ15
N en
vértebra del tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona del OPOT.
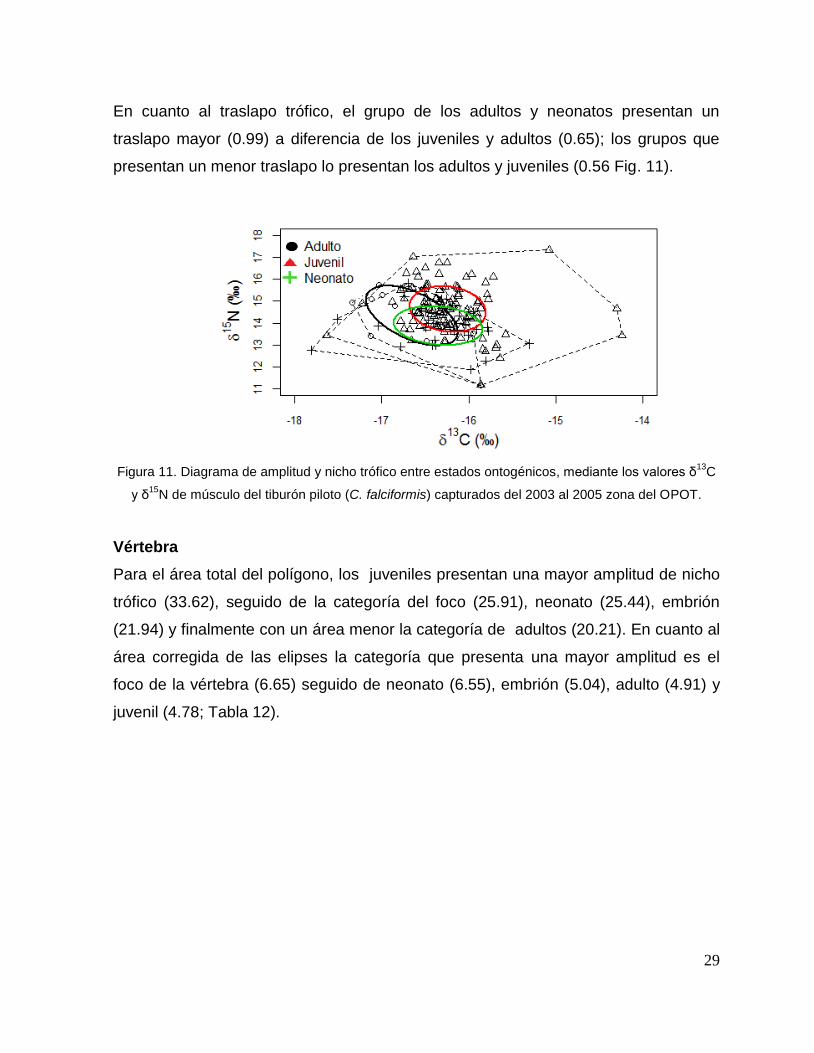
Músculo
Para el área total del polígono, los juveniles son los que presentan una mayor
amplitud de nicho trófico (13.45), seguido de los neonatos (5.49) y adultos (3.82). En
cuanto al área total de la elipse corregida, los adultos presentan una amplitud de
nicho mayor (1.89) seguido de los juveniles (1.41) y neonatos (1.40 Tabla 11)
Tabla 11. Áreas de los elipses y polígonos por estado ontogénico en músculo de
tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona del OPOT.
29
En cuanto al traslapo trófico, el grupo de los adultos y neonatos presentan un
traslapo mayor (0.99) a diferencia de los juveniles y adultos (0.65); los grupos que
presentan un menor traslapo lo presentan los adultos y juveniles (0.56 Fig. 11).
Figura 11. Diagrama de amplitud y nicho trófico entre estados ontogénicos, mediante los valores δ13
C
y δ15
N de músculo del tiburón piloto (C. falciformis) capturados del 2003 al 2005 zona del OPOT.
Vértebra
Para el área total del polígono, los juveniles presentan una mayor amplitud de nicho
trófico (33.62), seguido de la categoría del foco (25.91), neonato (25.44), embrión
(21.94) y finalmente con un área menor la categoría de adultos (20.21). En cuanto al
área corregida de las elipses la categoría que presenta una mayor amplitud es el
foco de la vértebra (6.65) seguido de neonato (6.55), embrión (5.04), adulto (4.91) y
juvenil (4.78; Tabla 12).
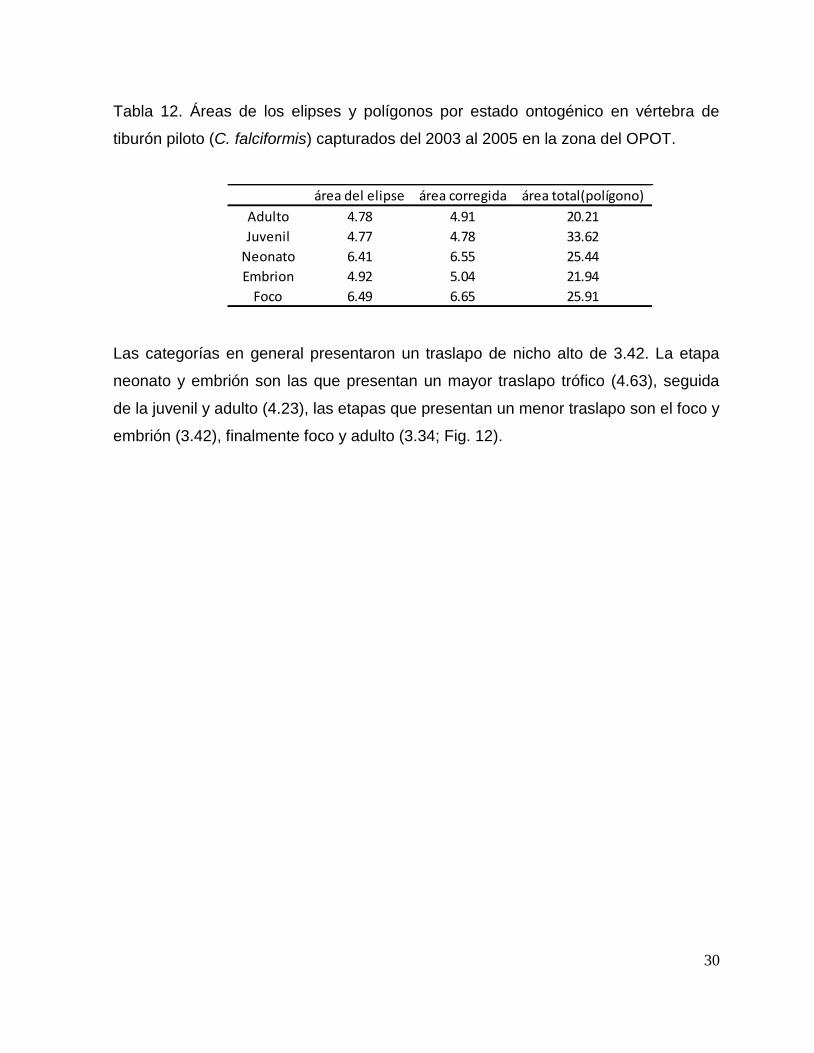
30
Tabla 12. Áreas de los elipses y polígonos por estado ontogénico en vértebra de
tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona del OPOT.
área del elipse área corregida área total(polígono)
Adulto 4.78 4.91 20.21
Juvenil 4.77 4.78 33.62
Neonato 6.41 6.55 25.44
Embrion 4.92 5.04 21.94
Foco 6.49 6.65 25.91
Las categorías en general presentaron un traslapo de nicho alto de 3.42. La etapa
neonato y embrión son las que presentan un mayor traslapo trófico (4.63), seguida
de la juvenil y adulto (4.23), las etapas que presentan un menor traslapo son el foco y
embrión (3.42), finalmente foco y adulto (3.34; Fig. 12).
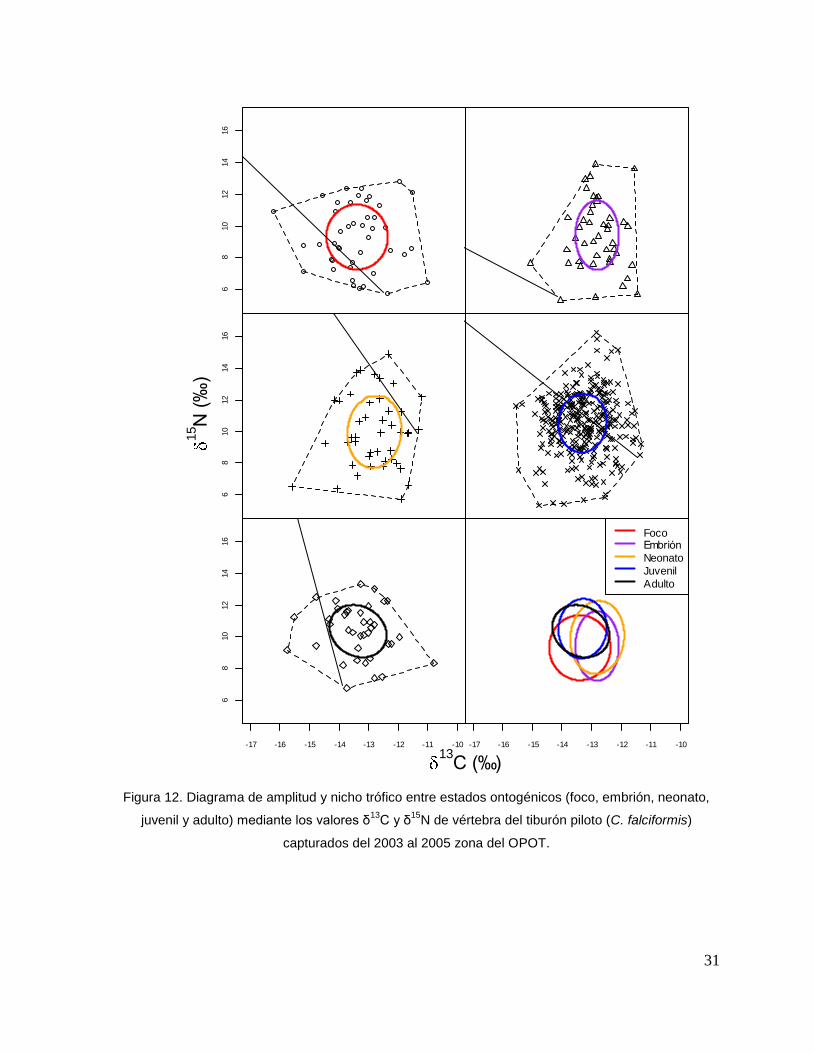
31
68
10
12
14
16
68
10
12
14
16
-17 -16 -15 -14 -13 -12 -11 -10
68
10
12
14
16
-17 -16 -15 -14 -13 -12 -11 -10
13C (‰)
15N
(‰
)
FocoEmbrión
Neonato
Juvenil
Adulto
Figura 12. Diagrama de amplitud y nicho trófico entre estados ontogénicos (foco, embrión, neonato,
juvenil y adulto) mediante los valores δ13
C y δ15
N de vértebra del tiburón piloto (C. falciformis)
capturados del 2003 al 2005 zona del OPOT.
32
Músculo
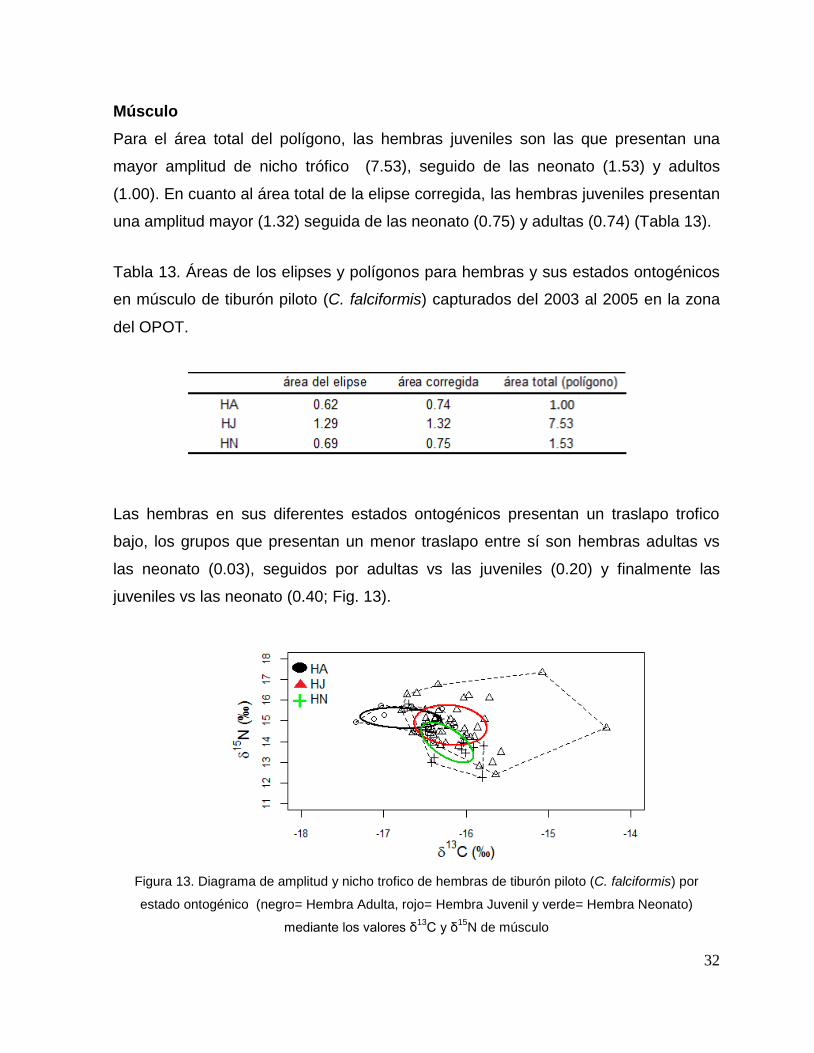
Para el área total del polígono, las hembras juveniles son las que presentan una
mayor amplitud de nicho trófico (7.53), seguido de las neonato (1.53) y adultos
(1.00). En cuanto al área total de la elipse corregida, las hembras juveniles presentan
una amplitud mayor (1.32) seguida de las neonato (0.75) y adultas (0.74) (Tabla 13).
Tabla 13. Áreas de los elipses y polígonos para hembras y sus estados ontogénicos
en músculo de tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona
del OPOT.
Las hembras en sus diferentes estados ontogénicos presentan un traslapo trofico
bajo, los grupos que presentan un menor traslapo entre sí son hembras adultas vs
las neonato (0.03), seguidos por adultas vs las juveniles (0.20) y finalmente las
juveniles vs las neonato (0.40; Fig. 13).
Figura 13. Diagrama de amplitud y nicho trofico de hembras de tiburón piloto (C. falciformis) por
estado ontogénico (negro= Hembra Adulta, rojo= Hembra Juvenil y verde= Hembra Neonato)
mediante los valores δ13
C y δ15
N de músculo
33
Músculo
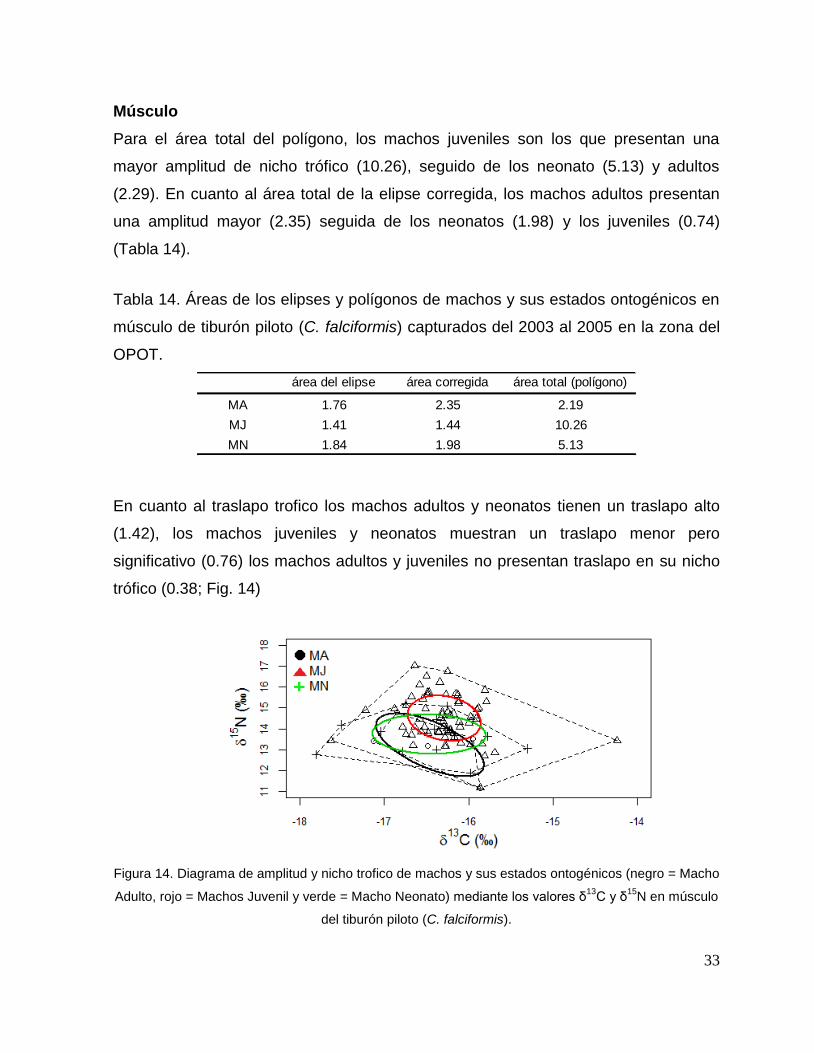
Para el área total del polígono, los machos juveniles son los que presentan una
mayor amplitud de nicho trófico (10.26), seguido de los neonato (5.13) y adultos
(2.29). En cuanto al área total de la elipse corregida, los machos adultos presentan
una amplitud mayor (2.35) seguida de los neonatos (1.98) y los juveniles (0.74)
(Tabla 14).
Tabla 14. Áreas de los elipses y polígonos de machos y sus estados ontogénicos en
músculo de tiburón piloto (C. falciformis) capturados del 2003 al 2005 en la zona del
OPOT.
área del elipse área corregida área total (polígono)
MA 1.76 2.35 2.19
MJ 1.41 1.44 10.26
MN 1.84 1.98 5.13
En cuanto al traslapo trofico los machos adultos y neonatos tienen un traslapo alto
(1.42), los machos juveniles y neonatos muestran un traslapo menor pero
significativo (0.76) los machos adultos y juveniles no presentan traslapo en su nicho
trófico (0.38; Fig. 14)
Figura 14. Diagrama de amplitud y nicho trofico de machos y sus estados ontogénicos (negro = Macho
Adulto, rojo = Machos Juvenil y verde = Macho Neonato) mediante los valores δ13
C y δ15
N en músculo
del tiburón piloto (C. falciformis).

34
8.6 Posición Trófica
Músculo
La posición trófica de C. falciformis en la zona del Océano Pacifico Oriental Tropical
presenta un valor promedio de 3.7 con mínimos de 2.5 y máximos de 4.6. Los
valores entre machos y hembras, ambos sexos presentaron valores similares (4.5 y
4.6 respectivamente). En los estados ontogénicos los juveniles son los que
obtuvieron una posición trófica mayor (PT = 3.8), mientras que los neonatos y adultos
obtuvieron valores similares (PT = 3.5). En la categoría de sexos por estado
ontogénico las hembras adultas presentan una posición trófica mayor (PT = 3.9),
seguidas de hembras juveniles, machos adultos y machos juveniles con posiciones
tróficas similares (PT = 3.7), los neonatos y hembras fueron los que presentaron
valores bajos (PT = 3.5; Tabla 15).
Tabla 15. Valores de la posición trófica calculados para el tiburón piloto (C.
falciformis) a partir de δ15N capturados del 2003 al 2005 en la zona del OPOT
.
Posición Trófica Máximos Mínimos Promedio DE
Machos 4.5 2.5 3.6 0.36
Hembras 4.6 2.9 3.8 0.36
Neonatos 4.1 2.9 3.5 0.28
Juveniles 4.6 2.5 3.8 0.33
Adultos 4.1 2.7 3.5 0.28
Machos neonatos 4 2.9 3.5 0.29
Machos juveniles 4.5 2.5 3.7 0.37
Machos Adultos 3.8 2.7 3.7 0.42
Hembras nonatos 4.1 3 3.5 0.29
Hembras juveniles 4.6 2.9 3.7 0.34
Hembras adultas 4.1 3.8 3.9 0.14
35
8.7 Composición de δ13C y δ15N durante el desarrollo ontogénico del
tiburón piloto.
Vértebra
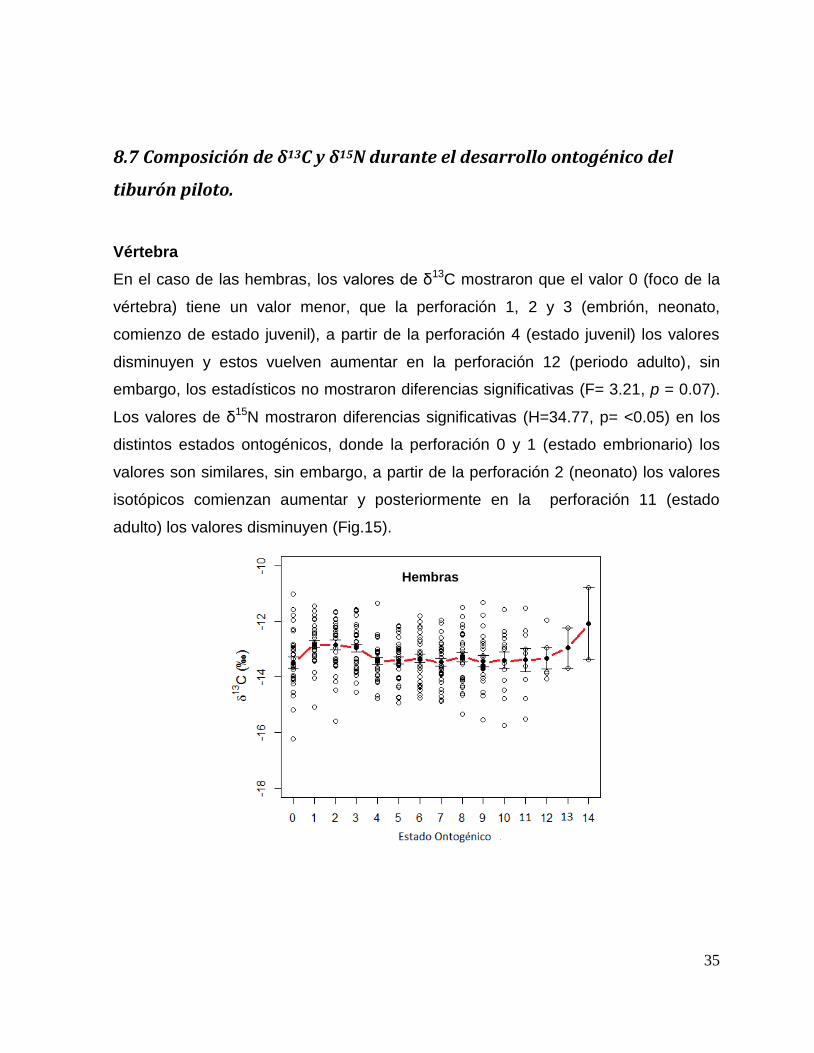
En el caso de las hembras, los valores de δ13C mostraron que el valor 0 (foco de la
vértebra) tiene un valor menor, que la perforación 1, 2 y 3 (embrión, neonato,
comienzo de estado juvenil), a partir de la perforación 4 (estado juvenil) los valores
disminuyen y estos vuelven aumentar en la perforación 12 (periodo adulto), sin
embargo, los estadísticos no mostraron diferencias significativas (F= 3.21, p = 0.07).
Los valores de δ15N mostraron diferencias significativas (H=34.77, p= <0.05) en los
distintos estados ontogénicos, donde la perforación 0 y 1 (estado embrionario) los
valores son similares, sin embargo, a partir de la perforación 2 (neonato) los valores
isotópicos comienzan aumentar y posteriormente en la perforación 11 (estado
adulto) los valores disminuyen (Fig.15).
Hembras
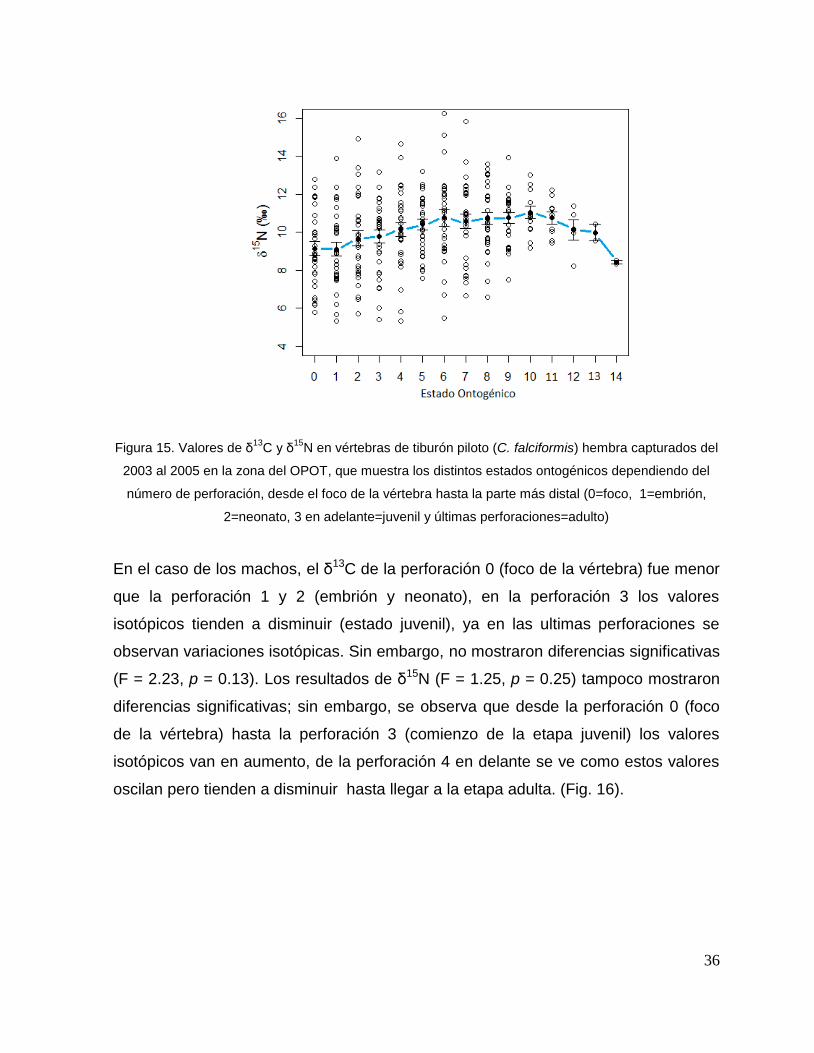
36
Figura 15. Valores de δ13
C y δ15
N en vértebras de tiburón piloto (C. falciformis) hembra capturados del
2003 al 2005 en la zona del OPOT, que muestra los distintos estados ontogénicos dependiendo del
número de perforación, desde el foco de la vértebra hasta la parte más distal (0=foco, 1=embrión,
2=neonato, 3 en adelante=juvenil y últimas perforaciones=adulto)
En el caso de los machos, el δ13C de la perforación 0 (foco de la vértebra) fue menor
que la perforación 1 y 2 (embrión y neonato), en la perforación 3 los valores
isotópicos tienden a disminuir (estado juvenil), ya en las ultimas perforaciones se
observan variaciones isotópicas. Sin embargo, no mostraron diferencias significativas
(F = 2.23, p = 0.13). Los resultados de δ15N (F = 1.25, p = 0.25) tampoco mostraron
diferencias significativas; sin embargo, se observa que desde la perforación 0 (foco
de la vértebra) hasta la perforación 3 (comienzo de la etapa juvenil) los valores
isotópicos van en aumento, de la perforación 4 en delante se ve como estos valores
oscilan pero tienden a disminuir hasta llegar a la etapa adulta. (Fig. 16).
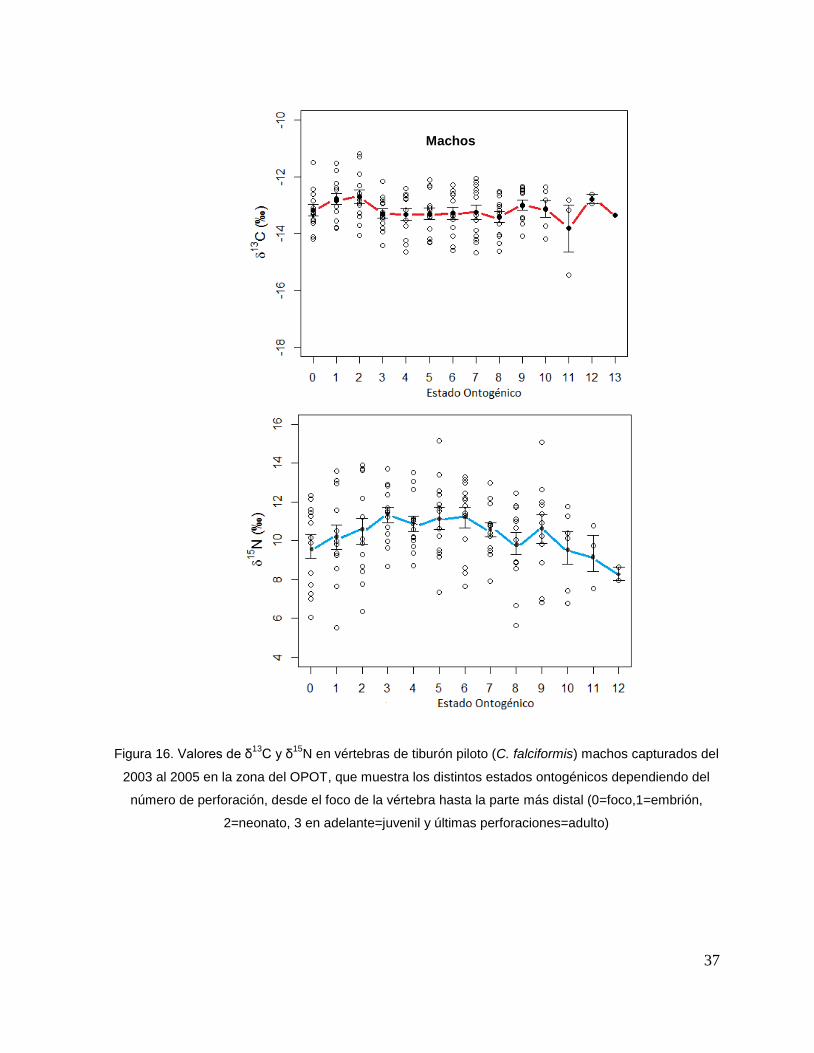
37
Figura 16. Valores de δ13
C y δ15
N en vértebras de tiburón piloto (C. falciformis) machos capturados del
2003 al 2005 en la zona del OPOT, que muestra los distintos estados ontogénicos dependiendo del
número de perforación, desde el foco de la vértebra hasta la parte más distal (0=foco,1=embrión,
2=neonato, 3 en adelante=juvenil y últimas perforaciones=adulto)
Machos

38
8.8 Perfiles ontogénicos individuales δ13C y δ15N del tiburón piloto.
Vértebra
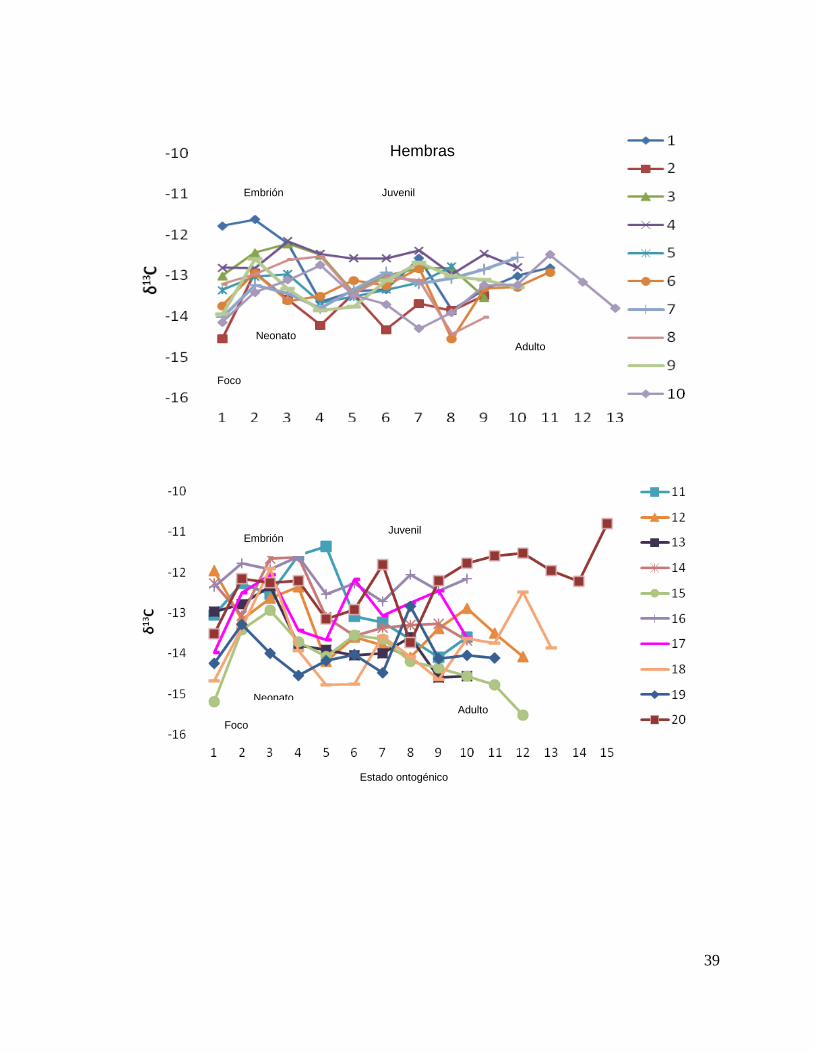
A partir del análisis de perfiles individuales de hembras se observó para δ13C que el
foco presentan valores menos enriquecidos, mientras que en la segunda perforación
(etapa de embrión), se observó un aumento (casos evidentes: tiburón 2, 15, 20, 24 y
30). En la tercera perforación (etapa de neonato), no se observó una tendencia clara
ya que en algunos casos disminuyeron (tiburón 1, 2, 19, 26 y 30) y en otros casos
aumentaron (tiburón 4, 10, 14, 17, 18 y 31). En la etapa juvenil (a partir de la cuarta
perforación) se observó que los valores oscilaron, hasta llegar a la etapa de adulto
(últimas perforaciones), donde en algunos casos se pudo observar un cambio en los
valores isotópicos (tiburón 10, 15, 18, 20, 29 y 30 (Fig. 17 y Tabla 16).
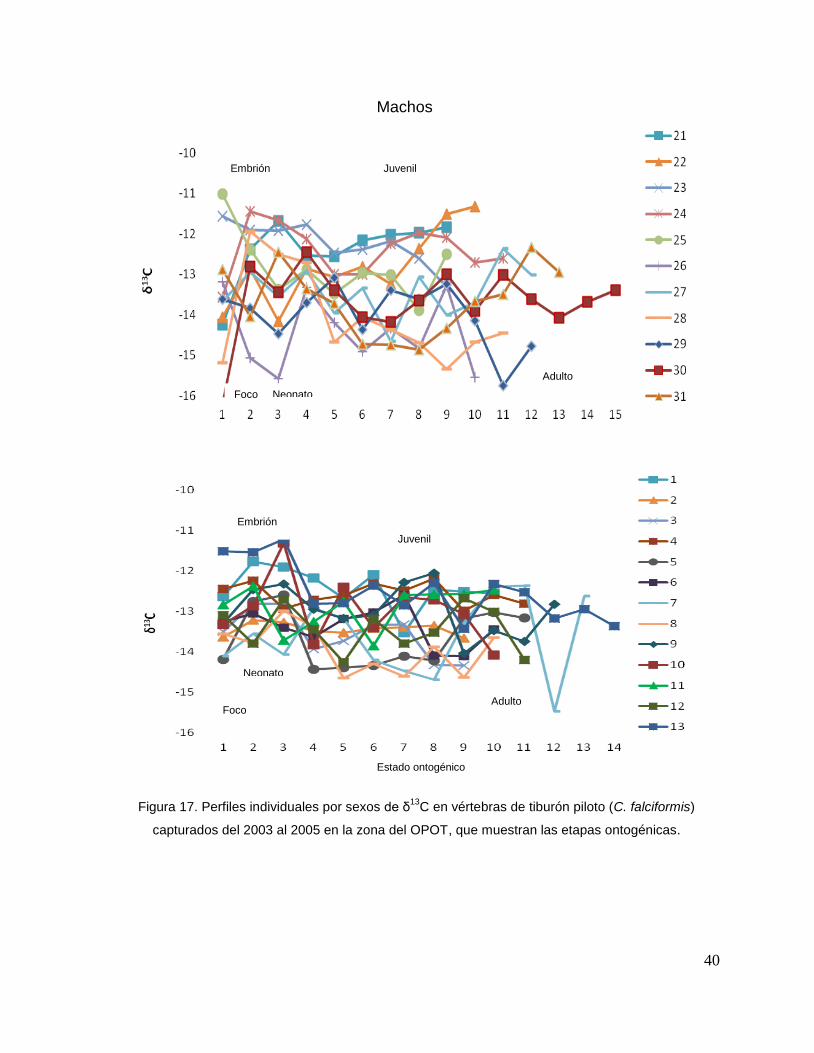
Para los machos se observó una tendencia similar, que δ13C en el foco presentan
valores menos enriquecidos, mientras que en la segunda perforación (etapa de
embrión), se observó un aumento (tiburón 1, 2, 3, 4, 5, y 10). En la tercera
perforación (etapa de neonato), se observó que en algunos casos disminuyeron
(tiburón 1, 2, 4, 6, 7, y 11) y en otros casos aumentaron los valores isotópicos
(tiburón 9, 10, 12, y 13). En la etapa juvenil (a partir de la cuarta perforación) se
observo que los valores de algunos organismos oscilaron (tiburón 1, 3, 7, 9, y 12);
mientras algunos se mantenían con poco cambio isotópico (2, 4, y 5). En la etapa de
adulto (últimas perforaciones), en algunos casos se pudo observar un cambio en los
valores isotópicos (tiburón 3, 5, 6, 7, 8, y 12) (Fig.17 y Tabla 17).
39
Neonato
Hembras
Foco
Embrión Juvenil
Adulto
Foco
Neonato
Embrión Juvenil
Adulto
Estado ontogénico
40
Figura 17. Perfiles individuales por sexos de δ13
C en vértebras de tiburón piloto (C. falciformis)
capturados del 2003 al 2005 en la zona del OPOT, que muestran las etapas ontogénicas.
Foco Neonato
Adulto
Embrión Juvenil
Machos
Neonato
Embrión
Juvenil
Foco Adulto
Estado ontogénico

41
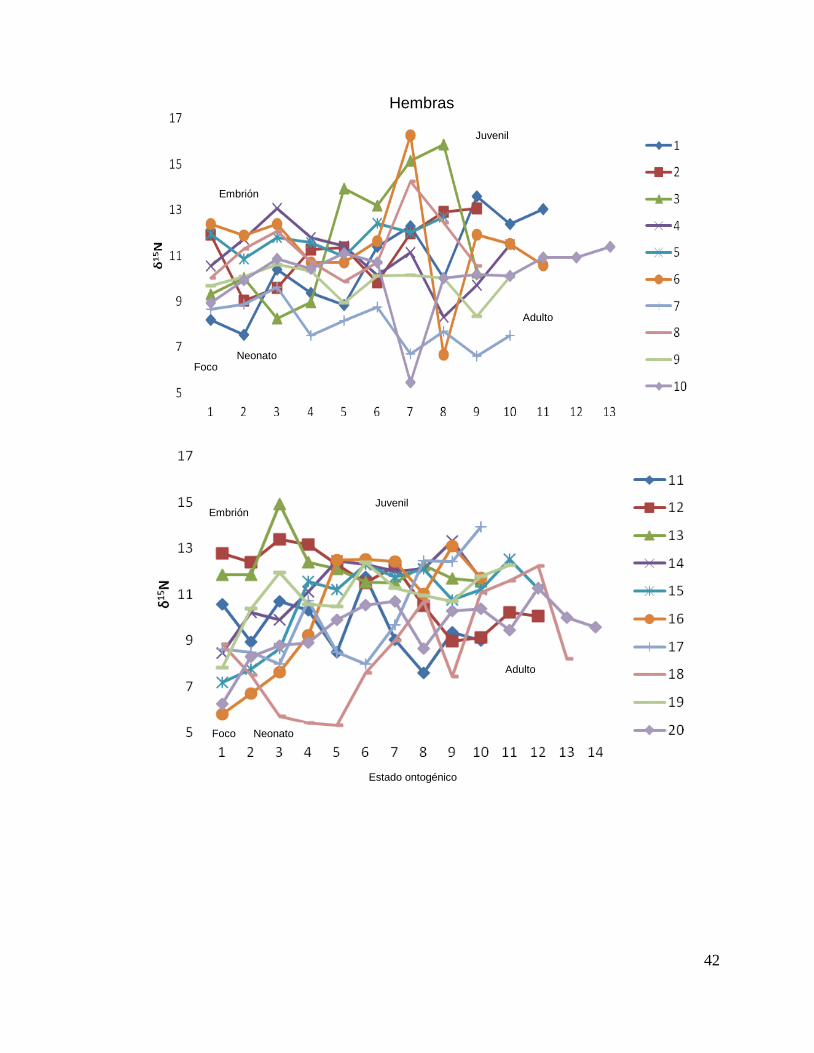
A partir del análisis de perfiles individuales de hembras se observó para δ15N que el
foco presentan valores menos enriquecidos, mientras que en la segunda perforación
(etapa de embrión), se observó en su mayoría un aumento (casos evidentes: tiburón
1, 3, 4, 8, 10, 15, 16, 19, 20, 22, 23, 25, 26 y 29). En la tercera perforación (etapa de
neonato), no se observó una tendencia clara pero en la mayoría de los casos los
valores aumentaron (tiburón 1, 2 ,4, 5, 6, 7, 8, 9, 11, 15, 16, 20, 21, 29 y 30).
En la etapa juvenil (a partir de la cuarta perforación) se observó que los
valores oscilaron en algunos casos disminuyendo (tiburón 4, 6, 7, 8, 11, 12, 13, 21, y
22) y en otros casos los valores aumentan (tiburón 3, 5, 14, 15, 16, 20, 23, 24, 29, 30
y 31), hasta llegar a la etapa de adulto (últimas perforaciones), donde en algunos
casos se pudo observar un cambio en los valores isotópicos (tiburón 3, 4, 8, 14, 15,
16, 20, 21, 25, 29, y 31 Fig.18 y Tabla16).
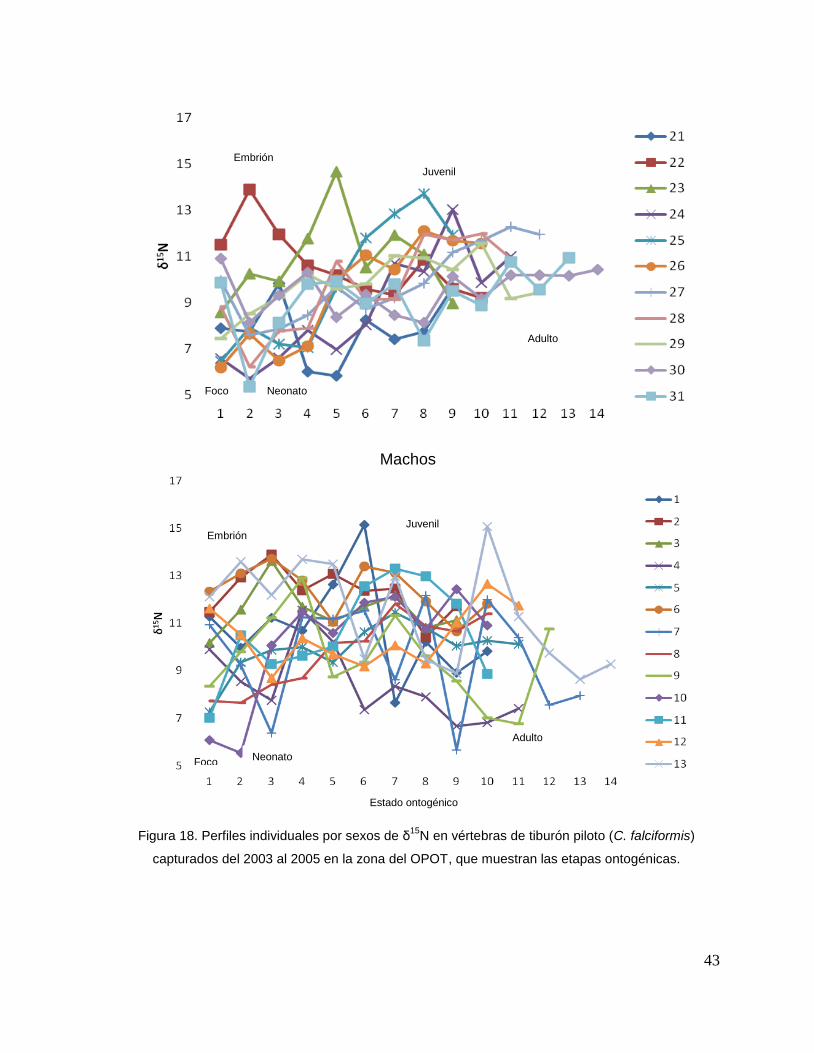
Para los machos se observó que δ15N en algunos casos los valores de foco
son menores y estos van aumentando en la etapa embrionaria (tiburón 2, 3, 6, 5, 9,
11 y 13), mientras que en otros casos sucede lo contrario (tiburón 1, 4, 7, 10, y 12).
En la tercera perforación (etapa de neonato), se observó que en algunos casos los
valores isotópicos disminuyeron (tiburón 4, 7, 12, 11,12 y 13), pero en su mayoría
aumentaron (tiburón 1, 2, 3, 5, 6, 8, 9, y 10).
En la etapa juvenil (a partir de la cuarta perforación) se observo que los
valores oscilaron ya que es una etapa con varias perforaciones, estos cambios
isotópicos en algunos organismos tiende aumentar con respecto a sus etapas
ontogénicas anteriores (tiburón 7, 8, 11 y 10) mientras que los valores isotópicos en
otros tiburones oscila mucho en esta etapa (tiburón 1, 4, 9, y 13). En la etapa de
adulto (últimas perforaciones), en algunos casos se pudo observar un cambio en los
valores isotópicos (tiburón 1, 9, 11, y 10 Fig.18 y Tabla 17).
42
Foco
Juvenil
Hembras
Hembras
Neonato
Embrión
Foco
Adulto
Embrión
Neonato
Adulto
Juvenil
Estado ontogénico
43
Figura 18. Perfiles individuales por sexos de δ15
N en vértebras de tiburón piloto (C. falciformis)
capturados del 2003 al 2005 en la zona del OPOT, que muestran las etapas ontogénicas.
Machos
Juvenil
Adulto
Neonato
Embrión
Foco
Neonato
Juvenil
Adulto
Foco
Embrión
Estado ontogénico
44
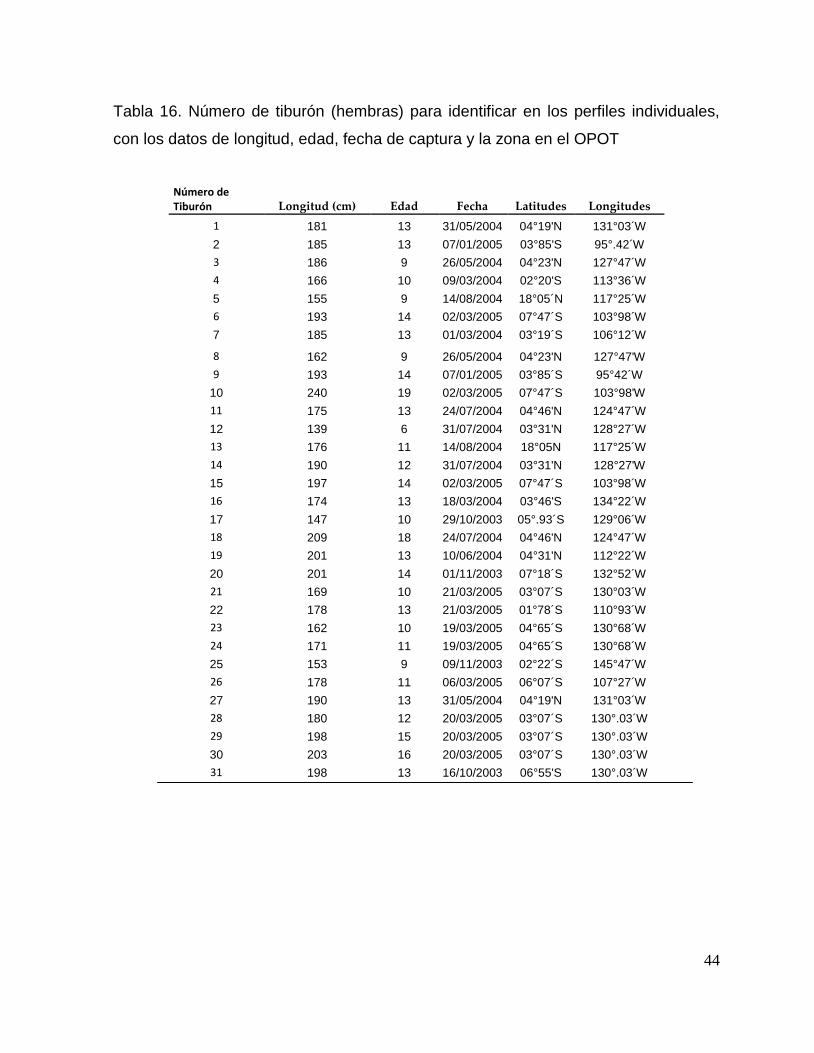
Tabla 16. Número de tiburón (hembras) para identificar en los perfiles individuales,
con los datos de longitud, edad, fecha de captura y la zona en el OPOT
Número de Tiburón Longitud (cm) Edad Fecha Latitudes Longitudes
1 181 13 31/05/2004 04°19'N 131°03´W
2 185 13 07/01/2005 03°85'S 95°.42´W
3 186 9 26/05/2004 04°23'N 127°47´W
4 166 10 09/03/2004 02°20'S 113°36´W
5 155 9 14/08/2004 18°05´N 117°25´W
6 193 14 02/03/2005 07°47´S 103°98´W
7 185 13 01/03/2004 03°19´S 106°12´W
8 162 9 26/05/2004 04°23'N 127°47'W
9 193 14 07/01/2005 03°85´S 95°42´W
10 240 19 02/03/2005 07°47´S 103°98'W
11 175 13 24/07/2004 04°46'N 124°47´W
12 139 6 31/07/2004 03°31'N 128°27´W
13 176 11 14/08/2004 18°05N 117°25´W
14 190 12 31/07/2004 03°31'N 128°27'W
15 197 14 02/03/2005 07°47´S 103°98´W
16 174 13 18/03/2004 03°46'S 134°22´W
17 147 10 29/10/2003 05°.93´S 129°06´W
18 209 18 24/07/2004 04°46'N 124°47´W
19 201 13 10/06/2004 04°31'N 112°22´W
20 201 14 01/11/2003 07°18´S 132°52´W
21 169 10 21/03/2005 03°07´S 130°03´W
22 178 13 21/03/2005 01°78´S 110°93´W
23 162 10 19/03/2005 04°65´S 130°68´W
24 171 11 19/03/2005 04°65´S 130°68´W
25 153 9 09/11/2003 02°22´S 145°47´W
26 178 11 06/03/2005 06°07´S 107°27´W
27 190 13 31/05/2004 04°19'N 131°03´W
28 180 12 20/03/2005 03°07´S 130°.03´W
29 198 15 20/03/2005 03°07´S 130°.03´W
30 203 16 20/03/2005 03°07´S 130°.03´W
31 198 13 16/10/2003 06°55'S 130°.03´W

45
Tabla 17. Número de tiburón (machos) para identificar en los perfiles individuales,
con los datos de longitud, edad, fecha de captura y la zona en el OPOT.
Número de tiburón Longitud (cm) Edad Fecha Latitudes Longitudes
1 177 11 07/11/2003 24°06´S 139°09´W
2 174 13 30/06/2005 01°78´S 110°93´W
3 196 14 31/07/2004 03°31'N 128°27´W
4 180 14 19/03/2004 03°20'S 136°10´W
5 194 14 07/01/2005 03°85´S 95°42´W
6 173 10 29/07/2004 03°85´S 124°92´W
7 167 12 13/06/2004
8 177 12 07/01/2005 03°85´S 95°42´W
9 198 15 31/05/2004 04°19'N 131°03'W
10 164 11 28/05/2004 04°93´S 128°78´W
11 176 12 20/03/2005 03°07´S 130°03´W
12 164 10 01/03/2004 03°19,S 106°12´W
13 192 12 23/10/2003 06°55'S 130°.03´W
9. DISCUSIÓN
9.1 Relación C: N
Frecuentemente los ecólogos utilizan el análisis de isótopos estables para el estudio
de la dinámica trófica de diferentes especies en el medio marino. Es por ello que
cada vez se vuelve fundamental comprender las posibles fuentes de variación que
pueden ocasionar en los valores isotópicos el tipo de muestreo biológico, las técnicas
de procesamiento y conservación de muestras, así como el tipo de tejido analizado
(Sotiropoulos et al., 2004), todos considerados como factores que pueden alterar la
interpretación de los valores isotópicos. La relación C: N se refiere a la cantidad de
carbono dividido por la cantidad de nitrógeno presente en una muestra y es utilizada
como prueba de homogeneidad bioquímica, por lo que es utilizada para detectar
posibles alteraciones en los valores isotópicos.
En el caso de los tiburones, la relación C: N sirve para evaluar si el tejido
analizado presenta altas concentraciones de urea (Sweeting et al., 2006; Kim y Koch,

46
2011) o lípidos (Post et al., 2007). La presencia de urea y lípidos en los tejidos
alteran los valores de δ15N y δ13C respectivamente, ya que la urea y el ácido úrico
tienen altas concentraciones de 14N (isótopo ligero); mientras que los lípidos están
disminuidos en 13C con relación a proteínas y carbohidratos, debido al proceso de
fraccionación isotópica que sucede durante la oxidación del piruvato a acetil
coenzima A (DeNiro y Epstein, 1977; Focken y Becker, 1998).
De manera teórica, los valores de C: N por debajo de 2.5 indicarían presencia
de urea (Sweeting et al., 2006; Kim y Koch, 2011) mientras que valores por arriba de
3.5 presencia de lípidos (Post et al., 2007). Una manera de evitar posibles efectos
negativos en los valores isotópicos por la presencia de urea o lípidos es a partir de
correcciones químicas. Sin embargo, se ha observado que dichas correcciones han
ocasionado alteraciones isotópicas en diferentes especies de peces (Gannes, 1997;
Kaehler y Pakhomov, 2001; Arrington y Winemiller, 2002). Pinnegar y Polunin (1999)
detectaron un aumento promedio de 0.78‰ en δ15N en el músculo de la trucha arco
iris juvenil una vez que le extrajeron lípidos; mientras que Sweeting et al. (2006)
recomiendan que antes de realizar alguna corrección se haga una medición de la
relación C: N.
En el presente estudio, la relación C: N de músculo (2.8) y vértebra (3.5)
estuvieron dentro del intervalo de muestras con bajas concentraciones lípidos y urea
(Post et al., 2007; Kim y Koch, 2011). Esto ha sido previamente observado en otras
especies de tiburones como Isurus oxyrinchus y Prionace glauca (Maya, 2012), por lo
que los tejidos analizados en el presente estudio de C. falciformis no presentan altas
concentraciones de lípidos y urea, siendo innecesaria hacer una corrección química y
por ende la variación de los valores obtenidos serán reflejo del comportamiento del
tiburón y no por la presencia de otros factores.

47
9.2 Composición de δ13C y δ15N
Se conoce que los tiburones presentan una amplia distribución, realizando
migraciones periódicas tanto horizontales como verticales determinadas por factores
ambientales, cambios ontogénicos, disponibilidad de alimento y por sus ciclos
reproductivos (Castro 1996), lo que en ocasiones dificulta el conocimiento del
comportamiento trófico de diferentes especies a partir de técnicas convencionales
(e.g. análisis de contenido estomacal). La composición isotópica en los tejidos de los
peces indican los efectos combinados del metabolismo, el crecimiento, ruta isotópica,
composición de proteína de tejido y la dieta (MacNeil et al., 2005).
Entre los tejidos más utilizados para la reconstrucción de dietas esta el
músculo; no obstante éste tejido no es capaz de retener la señal isotópica por un
tiempo mayor a 90 días ya que es vulnerable a cambios externos y tasas de
recambio metabólicas (MacNeil et al. 2005). Una estructura que puede ayudar a
disminuir los sesgos antes mencionados son las vértebras en específico las bandas
de crecimiento, las cuales se basan en la formación de zonas con cantidades
diferentes de calcio, cuya formación está relacionada con cambios estacionales a los
que estuvo sujeto el organismo a lo largo de su vida (Casselman, 1974; Branstetter
et al., 1987). Las diferencias en la composición química de las estructuras
calcificadas como vertebras se pueden utilizar para indicar orígenes natales,
conectividad, estructura de la población y reconstruir el historial o el movimiento
ambiental de patrones de muchos organismos marinos (Campana et al.2002).
Cabe resaltar que debido a que las bandas de crecimiento no son fácilmente
discernibles en todas las especie de tiburones en ocasiones se dificulta darle una
correspondencia a un ciclo anual (Cailliet y Goldman, 2004). Sánchez de Ita (2011) y
Cervantes (2013) mencionan que la periodicidad de formación de las marcas de
crecimiento para C. falciformis es anual, iniciando en verano-otoño y completando la
marca, en invierno primavera. En el presente trabajo el muestreo fue sistemático

48
partiendo del foco de la vértebra que es la parte central de la estructura hasta la
parte más distal, procurando tener el mayor número de muestras posible. Sin
embargo, es necesario resaltar que a diferencia de otras especies de tiburones, en
las vértebras de C falciformis fue posible detectar las diferentes bandas de
crecimiento sin la necesidad de dar tratamientos especiales (e.g. tinción de
vertebras), lo cual a su vez podría afectar los valores isotópicos.
En el presente estudio se utilizaron dos estructuras (músculo y vértebra) para
conocer los posibles patrones tróficos del tiburón piloto a lo largo de su ciclo de vida.
En el caso de δ13C, tanto la vértebra como el músculo mostraron una alta variabilidad
(Tabla 4 isotópica, lo que indica que este depredador presenta movimientos costeros
y oceánicos en las diferentes etapas de su vida. La composición isotópica en los
años de captura confirmó los posibles movimientos del tiburón piloto en distintas
escalas de tiempo, lo cual demuestra que este depredador tiene un amplio rango de
movilidad en el OPOT. Al respecto, Cabrera (2003) y Barranco (2008) a partir de
δ13C mencionan que en mares mexicanos C. falciformis presenta hábitos costeros y
oceánicos variando sus zonas de alimentación (Compagno, 1984), lo cual coincide
con lo observado en el presente estudio.
En el caso de δ15N, el valor promedio (14.40‰) fue menor a los reportados en el
Golfo de Tehuantepec (15.53‰; Barranco, 2008) y Golfo de California (alrededor de
16.00‰; Cabrera, 2003). Bonfil et al. (2005), mencionan que los tiburones son
organismos con una alta movilidad en el plano espacial y pueden realizar buceos
profundos, lo que los hace aptos para tener una fácil accesibilidad a distintos tipos de
presas, tanto oceánicos como costeros o pelágicos como bentónicos. En el caso del
tiburón piloto, esto ha sido observado a partir de análisis de contenido estomacal
(Cabrera 2000), lo que demuestra la alta movilidad al momento de alimentarse. Por
lo tanto, los valores diferenciales entre áreas demuestra que el nivel trófico de las
presas consumidas por C. falciformis varia, lo que demuestra que se está
alimentando de distintas presas dependiendo en la zona en la que este se encuentre.

49
Con el complemento de ambos tejidos, se pueden identificar individuos cuyas dietas
varían con el tiempo e individuos con dietas relativamente constantes (Bearhop et al.
2004). El cambio observado en los valores isotópicos de cada tejido demuestra la
utilidad de la técnica para detectar cambios en la dinámica trófica. Post (2002)
menciona que la comparación de los valores isotópicos entre diferentes sistemas
provee información acerca de la última fuente de carbono de cada consumidor, ya
que existe una considerable variación en las concentraciones de δ13C de la base de
la red alimenticia de la cual los organismos adquieren su carbono entre cada
ecosistema.
La composición isotópica entre sexos en ambos tejidos fue similar, lo que indica que
tanto hembras como machos se están alimentando en la misma zona de especies
presa similares. Sin embargo, Andrade (2005) reporta un bajo traslapo entre sexos.
Las diferencias podrían estar relacionadas con la técnica de análisis utilizada
(contenido estomacal vs. isotopos estables) y el periodo de tiempo que representan.
Mientras que el contenido estomacal es considerado como la última ingesta del
organismo (horas o días), los isótopos estables representan el alimento asimilado en
largos periodos de tiempo (meses). Además, unas de las limitantes más
considerables del contenido estomacal es el grado de digestión de la presas, lo cual
dificulta la identificación y tiende a subestimar o sobreestimar la importancia de
algunas especies presas.
En el caso de los isotopos estables, si bien se desconoce la resolución taxonómica
de las presas, se tiene la certeza del área y el nivel trófico de las presas, lo que
ayuda entender la dinámica trófica de la especie, por lo que la representación
isotópica sería un reflejo de un comportamiento alimenticio más constante que lo
reflejado por el contenido estomacal. Si bien Andrade (2005) reporta un bajo traslapo
entre sexos, en ambos casos la presa principal fue el atún aleta amarilla Thunnus
albacares (66.7%), mientras que las diferencias radicaron en las presas secundarias.

50
La diferencia en los resultados entre ambos estudios seria el resultado de la
asimilación de presas. Al ser T. albacares la especie principal en ambos sexos, esta
sería la que contribuye mayormente a la composición isotópica del depredador y por
lo tanto señales similares. Al respecto, diferentes autores han señalado un traslapo
alto entre sexos para C. falciformis en otras áreas, lo que demuestra que la presa
principal siempre es compartida mientras que las presas secundarias son las que
cambian (Cabrera 2000).
La composición isotópica de ambos tejidos por estado ontogénico mostro ser
diferente en δ13C, lo que indica una posible segregación trófica. En el caso del
músculo, los adultos presentaron valores de -16.65‰, lo que indica que se
encuentran en zonas oceánicas; mientras que los juveniles (-16.24‰) se mantienen
también en zonas oceánicas, pero más cercanas a la costa. Niño-Torres et al. (2006)
mencionan que valores entre los -10 a -15‰ son característicos de zonas costeras,
por lo que los valores observados en C. falciformis son característicos de zonas
oceánicas.
En el caso de las vértebras, se contó con dos perforaciones que incluyen el estado
embrionario con valores de δ13C (-16.22‰ y -15.07‰), los cuales son similares a la
de la madre o en este caso los adultos (-15.75‰). Gilbert y Schlernitzauer (1966),
mencionan que las hembras de C. falciformis proporcionan una nutrición continua
para sus crías en desarrollo a través de la placenta, donde los embriones están
protegidos dentro del cuerpo de su madre, y están adquiriendo los nutrientes de las
presas que la madre está consumiendo, obteniendo alrededor del 99% de los
nutrientes durante la gestación (Wourms, 1993). Por lo tanto, la ausencia de
diferencias entre neonatos y adultos podría estar relacionado con una distribución
espacial similar, ya que los embriones de C. falciformis están adquiriendo señales
isotópicas similares de la zona donde se alimentan las madres, sin embargo, se
presentaron diferencias entre neonatos y juveniles.

51
Diferentes autores reportan una segregación espacial en C. falciformis dependiendo
del estado ontogénico, tiburones adultos tienden a migrar hacia zonas oceánicas, ya
que ahí se alimentan de presas más grandes como los atunes; mientras que los
neonatos y juveniles se distribuyen en regiones costeras ya que funciona como una
zona de crecimiento y alimentación (Castillo-Géniz et al., 2002; Andrade, 2005). En el
presente estudio se detecto diferencias solo entre neonatos y juveniles, lo que podría
estar asociada con la etapa de transición, donde la etapa crítica es de neonato a
juvenil. Durante esta transición se ha reportado que el organismo presenta niveles de
stress alimenticio como resultado de su poca experiencia al momento de capturar
sus presas y por ende el uso de sus reservas lipídicas, esto tiene como
consecuencia cambios en las señales isotópicas del carbono fijadas en diferentes
tejidos. La falta de diferencia entre juveniles y adultos indicaría una disminución en el
stress alimenticio como consecuencia de una mayor experiencia en la captura de
presas
Un ejemplo de lo anterior son los valores de δ15N mostraron diferencias entre
juveniles y neonatos para ambos tejidos. Los embriones presentaron valores de δ15N
bajos y conforme fue aumentando tamaño los valores se incrementaron. Post (2002),
menciona que a medida que un organismo aumenta su tamaño, la señal isotópica se
incrementa como resultado del aumento en la diversidad (y posición trófica) de
especies presas en la dieta. Cabrera (2003), reporto una segregación por estado de
madurez y sexo en la población de C. falciformis de la costa occidental de Baja
California Sur, donde sugiere que las hembras adultas están separadas de los
machos, tanto juveniles como adultos, así como de las hembras juveniles. Lo anterior
demuestra que la “experiencia” adquirida conforme crecer el depredador en la
captura de especies presas podría ser considerado como factor importante para la
representación de la composición isotópica de la especie estudiada. Por lo tanto, la
diferencia isotópica observada en el presente estudio permitió detectar y corroborar
las segregaciones reportadas en estudios previos.

52
Si bien, el análisis por estado ontogénico nos permitió detectar diferencias
relevantes, el generalizar el comportamiento de la especie ocasionaría faltas graves
en el conocimiento de la dinámica poblacional. Por ejemplo, los resultados indican
que las hembras no presentan diferencias en δ13C, lo que podría indicar que a lo
largo de su ciclo de vida se están alimentando en las mismas zonas. La ausencia de
diferencias a nivel sexo/estado ontogénico en particular para hembras resalta la
necesidad de analizar detalladamente las posibles variaciones de manera
meticulosa.
La ausencia de diferencias y amplia variabilidad isotópica entre hembras en
diferentes estados ontogénicos puede estar relacionada con afinidad a zonas de
reproducción. Se ha documentado que las hembras pueden tener un alto nivel de
fidelidad a los sitios natales, regresando cada año a las mismas áreas para expulsar
a sus crías (Springer 1967; Keeney et al. 2003; Hueter et al. 2005; DiBattista et al.
2008). Estas áreas pueden ser abiertas o cerradas y son generalmente en las zonas
costeras poco profundas y de alta productividad, en donde los recién nacidos
encuentran alimento y protección de sus depredadores, incluyendo a los tiburones
adultos. Este patrón de comportamiento se observa en los resultados de las últimas
perforaciones de la vértebra, donde las hembras adultas se encuentran más
cercanas a la costa (Castro, 1993; Bonfil, 1997).
Algunos autores mencionan que se han identificado tres tipos de áreas, de acuerdo
al uso de cada una de ellas: 1) áreas de alimentación de adultos, 2) áreas de
reproducción y 3) áreas de crianza (Castro, 1993). En el litoral oaxaqueño hay
nacimientos de C. falciformis durante la temporada de lluvias, cuando se presentan
las temperaturas más elevadas del año y se encuentra una alta disponibilidad de
alimento (Pennington et al. 2006). Al respecto Branstetter (1990), Castro (1993) y
Simpfendorfer y Milward (1993), mencionan que los neonatos tienen altas tasas de
crecimiento, lo que indica un consumo elevado de alimento, por lo que tienden a
permanecer en áreas donde el alimento es abundante. Lo anterior demuestra que C.

53
falciformis presenta una alta movilidad y que su distribución estará en función de su
estado ontogénico y sexo.
Los valores de δ15N indicaron que las hembras presentaron un incremento
desde la etapa de neonato hasta la etapa juvenil para posteriormente disminuir en la
fase adulta. La similitud en los valores iniciales y finales de δ15N podría deberse: 1) al
paso de las sustancias alimenticias de la madre a embrión, y 2) la posible
especialización en un recurso alimenticio. En el primer caso, McMeans et al. (2009ª)
y Vaudo et al. (2010) reporta en el tiburón martillo (Sphyrna lewini) y el tiburón puntas
negras (Carcharhinus limbatus), los neonatos presentan valores de δ15N similares o
más enriquecidos a los de las madres. Mientras que en el segundo caso, los
requerimientos energéticos de las hembras a lo largo de su ciclo de vida son
diferentes (e.g. etapas reproductivas), por lo que se ven en la necesidad de consumir
presas diferentes. Caso contrario el de los machos, los cuales al no presentar un
gasto energético como consecuencia de la reproducción, se alimentan de especies
presa similares (valores de δ15N similares) a lo largo de su vida.
9.3 Amplitud y traslapo de nicho
El concepto de nicho ha sido definido como el lugar que ocupa un animal en
su comunidad y sus relaciones con el alimento (Elton, 1927). Pianka (1973)
menciona que los animales reparten los recursos en el ambiente en tres formas
básicas: trófica, espacial y temporalmente, por lo tanto el reparto de los recursos
disponibles en un ecosistema puede ayudar a comprender la manera en la que las
especies coexisten (Pianka, 1974).A partir de la composición isotópica, se observó
que C. falciformis presentó una amplitud de nicho similar a la de un depredador
generalista tanto entre sexos como por estado ontogénico, ya que los valores
estuvieron por arriba de 1, lo que demuestra que esta especie se alimenta de
distintas presas (Fry y Sherr, 1984; Peterson y Fry, 1987). Al respecto, Wheterbee et
al. (1990), sugieren que la disponibilidad del alimento es uno de los factores más
importantes que afectan las preferencias alimenticias de los tiburones ya que

54
observaron que cuando el alimento es abundante, los depredadores seleccionan ese
alimento en particular para maximizar su consumo y aprovechamiento de energía.
A partir del análisis de contenido estomacal se ha documentado que C.
falciformis presenta una alta diversidad de especies presas en todos los estados
ontogénicos (Castro, 1996; Cabrera, 2003), lo que podría ser interpretado como una
gran amplitud de nicho trófico. Sin embargo, se observó diferencias en los valores de
amplitud trófica entre juveniles y adultos, lo que podría estar asociado a: 1) posible
especialización conforme crece el organismo y 2) cambios en la longitud de la
cadena trófica dependiendo el área (oceánica vs costera). Cabrera (2003) menciona
que la dieta de C. falciformis puede variar dependiendo de la zona de captura y
disponibilidad de alimento, lo cuales son considerados como factores importante que
afecta las preferencias alimenticias de estos organismos, ya que se ha observado
que cuando la comida es abundante, los tiburones tienden a seleccionar un alimento
en particular, el cual maximiza su consumo y el aprovechamiento de energía
(Wetherbee et al., 1990)
Lo anterior podría ser reflejo de una posible especialización, sin embargo,
algunos autores mencionan que la especialización en un tipo de recurso tendría
como resultado una baja variabilidad isotópica (Bearhop, 2000; Newsome, 2004) lo
cual no es el caso del presente estudio, donde ambos estados (juveniles y adultos)
presentaron una amplia variabilidad. Por otra parte, la longitud de una cadena trófica
tiene una gran influencia en los valores isotópicos, ya que se ha observado que
áreas que presentan una mayor diversidad tienen como resultado una alta
variabilidad (Post 2002). Estudios previos han señalado que las zonas costeras
presentan una mayor diversidad que las zonas oceánicas y por lo tanto una mayor
amplitud y traslapo trófico (Torres-Rojas et al., 2013), lo que explicaría las diferencias
encontradas en este estudio, ya que al estar en una zona costera, los juveniles
presentarían una mayor amplitud trófica que los adultos y por lo tanto un bajo
traslapo.

55
En relación al traslapo trófico, es importante conocer el nivel de competencia
que exista entre especies o dentro de una especie (MacNaughton y Wolf 1973). Si
bien Lawlor (1980) explica que las mediciones de traslapo de nicho son reconocidas
por no ser verdaderas mediciones de competencia, estas son regularmente utilizadas
como una primera aproximación para su estudio, ya que medidas directas de
competencia son raramente disponibles para la mayoría de estudios. En el presente
estudio, los resultados mostraron que existe un traslapo alto en machos y hembras,
ya que los valores estuvieron por encima de 1. Barraco (2008) reporta un alto
traslapo entre sexos en el Golfo de Tehuantepec debido a la alta similitud en la dieta,
donde Thunnus albacares fue la especie que consumieron ambos sexos, lo que
indica nula competencia.
La competencia influye en la habilidad de un individuo para sobrevivir y
reproducirse, y puede ser modelada como cambios en el tamaño de la población con
el tiempo (MacNaughton y Wolf 1973). Darwin (1859) menciona que la competencia
será más severa entre organismos pertenecientes a un mismo género que entre
géneros, lo cual explicaría el porqué de la falta de competencia entre sexos. En
relación a los estados ontogénicos, se observó un bajo traslapo, lo que indica una
segregación trófica. Tal es el caso de las hembras, quienes mostraron bajo traslapo
en todas sus categorías, lo cual pudiera estar relacionado a que las hembras tienen
diferentes requerimientos energéticos a lo largo de su vida.
En el periodo de reproducción, las hembras buscan presas con mayor
contenido energético. Se ha reportado que las hembras grandes de Squalus
megalops tuvieron una dieta diferente y consumieron mayor cantidad de presas ricas
en calorías con respecto a las presas de los machos y las hembras pequeñas
(Braccini 2005). Mientras que en el caso de los machos, se observó un traslapo alto
entre adultos y neonatos (1.42), y más bajo entre juveniles y adultos (0.76). Andrade
(2005), reporta que el traslapo entre hembras adultas y juveniles, machos adultos y
juveniles el traslapo fue bajo (0.29) indicando que hay segregación por talla y sexo
en el alimento de C. falciformis lo cual está asociado con el área de captura.

56
El estudio de la segregación trófica en relación al estado ontogénico ha sido
observado en otras especies de tiburones. Klimley (1983) menciona que en el Golfo
de California, el tiburón martillo Sphyrna lewini en su etapa adulta es un depredador
oceánico; mientras que los juveniles prefieren zonas costeras Cortés y Gruber (1990)
observaron en la dieta del tiburón limón Negaprion brevirostris que las variaciones en
la dieta en cada intervalo de talla son debido al cambio ontogénico, lo cual permite
observar que al ir alcanzando tallas más grandes, la variedad de hábitats que ocupa
se incrementa. Así mismo, reportan que las presas más grandes fueron encontradas
en tiburones grandes y viceversa.
La segregación trófica favorece la coexistencia de depredadores al evitar una
confrontación directa de los individuos, o bien a reducir el traslapo en el uso de los
recursos (Kronfeld–Schor y Dayan, 2003). Por ejemplo, se ha observado que cuando
las especies simpátricas presentan poco traslapo en sus patrones de actividad diaria,
la competencia por otro tipo de recursos como el alimento o el espacio es menor
(Johnston y Zucker, 1983). Darwin (1859) menciona que la lucha será generalmente
más severa entre ellas si llegan a estar en competencia con el mismo género, lo cual
explica el porqué de la segregación entre hembras de diferentes estados
ontogénicos, al igual que entre machos.
Entre las causas principales de la segregación trófica destaca en general que
los tiburones cambian su dieta conforme van creciendo, ya que conforme un tiburón
aumenta de tamaño, también aumenta la eficiencia para capturar a su presa, debido
a que poseen sus sentidos totalmente desarrollados y son capaces de capturar
presas grandes y rápidas; en cambio, los tiburones pequeños no pueden presentar
este tipo de comportamiento y tienen que alimentarse de presas que no son tan
rápidas o que no presenten un alto contenido de energía, lo cual suplen comiendo las
presas a las que tengan acceso y que a su vez sean de captura fácil (Lowe et al.,
1996). Por lo tanto, la segregación trófica observada entre estados ontogénicos está
relacionada con las capacidades fisiológicas del organismo, lo que lo limita a ocupar
determinados hábitats dependiendo de su estado de madurez.

57
9.3 Posición Trófica
Las redes alimenticias o tróficas, son las relaciones alimenticias que se
establecen de forma lineal entre organismos que pertenecen a distintos niveles
tróficos (Díaz, 2006). La teoría de tramas tróficas busca comprender los patrones de
interacciones alimenticias que exhiben las comunidades en la naturaleza (Pimm et
al., 1991). A través de la alimentación los organismos acuáticos son capaces de
provocar efectos en cascada sobre niveles tróficos inferiores (Carpenter y Kitchel,
1993), que pueden afectar directa o indirectamente la mayoría de los componentes
del ecosistema, acoplarlos y promover cambios en los ciclos de nutrientes y la
dinámica energética (Motta y Uieda, 2005).
Considerando que los tiburones son organismos móviles con tiempos generacionales
y tamaños corporales relativamente grandes, suelen conectarse a través de su
alimentación en zonas litorales, bentónicas y pelágicas (Jeppesen et al., 1997;
Schindler y Scheurell, 2002; Pace et al., 2004). En el presente estudio, la posición
trófica de C. falciformis fue de 3.6 a 3.8. Sin embargo, otros estudios reportan valores
de 4.0 (Cortés 1999), 4.2 (Froese y Pauly 2007) y 4.5 (Barraco, 2008), colocando a
este tiburón como consumidor terciario.
Las diferencias entre los valores de los diferentes estudios puede depender
de: 1) la metodología usada para el cálculo de la posición trófica, o 2) cambios en la
estructura trófica del OPOT. En el primer caso, el contenido estomacal solo registra
las especies presas de mayor tamaño, lo que deriva en una posiciones tróficas altas
(estudios previos), subestimando el papel que desempeñan niveles tróficos menores;
mientras que los isótopos estables, al ser una técnica que integra tanto presas
grandes como pequeñas, la posición trófica sería un reflejo más cercano del papel
funcional que desempeña la especie (presente estudio).
Si bien lo anterior podría ser la explicación más factible, no podemos descartar
posibles cambios en el ecosistema derivado de impactos naturales (e.g. el niño) o
antropogénicos (e.g. sobrepesca) Vögler et al. (2008) mencionan que el nivel trófico

58
poblacional podría ser considerado como un indicador de la salud de las poblaciones,
debido a que resume los cambios en la dinámica poblacional. Si consideramos que
los tiburones son depredadores tope de la cadena alimenticia que juegan un papel
importante en los ecosistemas marinos al regular poblaciones de niveles tróficos
menores (Ellis et al., 1996), un aumento o disminución del nivel trófico, podría ser un
indicativo de cambios en la disponibilidad de especies presas principales en la dieta
con alto nivel trófico (e.g. atunes) y el incremento en el consumo de presas de
niveles tróficos bajos (e.g. langostilla).
En los últimos 5 años se ha registrado un descenso en las capturas de atún
(CIAT, 2012) y un aumento en la presencia de langostilla en la dieta de otros
depredadores (Torres-Rojas et al. 2014). En ambos casos se desconoce a detalle el
motivo de los cambios en las abundancias de estas especies en el OPOT, pero sirve
como indicativo de los cambios observados en el nivel trófico de C. falciformis. Lo
anterior resalta la importancia de mantener un constante monitoreo del nivel trófico
de diferentes especies como tiburones a partir de este tipo de estudios para detectar
alteraciones en el ecosistema. Sin embargo, la siguiente etapa a investigar es el
motivo por el cual de los cambios, ya sean naturales o antropogénicos.
En relación a los niveles tróficos por estado ontogénico, se conoce que la
demanda energética aumenta con el tamaño corporal (McNab, 2002) y asociado a
las características ecológicas de las especies (abundancia, tasas de crecimiento,
relaciones tróficas) que influyen en la estructura y dinámica de las tramas tróficas
(Brown et al. 2004). Por lo tanto, el tamaño corporal y por ende el estado ontogénico
tiene un importante papel en la estructura de las comunidades (Woodward et al.
2005).
En el presente estudio los machos en sus distintas etapas ontogénicas
obtuvieron valores similares de la posición trófica: neonatos (3.5), juveniles (3.7) y
adultos (3.7); por otro lado, las hembras mostraron valores más altos conforme van
aumentando de tamaño: neonatas (3.5), juveniles (3.7) y adultas (3.9). Ferry (1998)

59
menciona que los tiburones recién nacidos se alimentan por medio de la succión de
su presa o por alimentación de tipo ram, en donde la presa es tragada, debido a que
el tiburón nada con el hocico abierto. Este tipo de alimentación desaparece conforme
el organismo crece, ya que va adquiriendo experiencia depredatoria. Estos cambios
ontogénicos a partir del estudio de isótopos estables han sido previamente
reportados en otros depredadores. Abrantes y Barnett (2011) observaron en el
tiburón Notorynchus cepedianus los valores del δ15N parecen incrementarse con la
talla hasta los 250 cm de longitud total, pero los individuos mayores a esta talla
tienen valores menores que los individuos pequeños (151−250 cm LT), lo que
indicaría que los animales más grandes se alimentan en otra área caracterizada por
una línea base con valores bajos del δ15N, por lo tanto, se puede mencionar que si
bien el tiburón piloto puede ser considerado como depredador secundario-terciario, la
alta plasticidad trófica le permite cumplir diferentes roles en el ecosistema, lo cual es
considerado como esencial en el mantenimiento de un equilibrio trófico.
El potencial de un depredador de alcanzar posiciones tróficas altas depende
de sus limitaciones morfológicas para consumir las presas (Pimm, 1982). Los
depredadores generalmente son más grandes que sus presas (Cohen et al., 1993),
generando una relación positiva entre el tamaño corporal y la posición trófica
(Layman et al., 2005). Por lo tanto, el presente estudio mostro que C. falciformis
presento valores de posición trófica característicos de un consumidor terciario pero
que puede presentar cierto grado de plasticidad y que estos dependerán del estado
ontogénico en que se encuentre, por lo que el papel funcional de este depredador
puede ser considerado como clave dado que regula diferentes poblaciones de
especies presas en diferentes zonas conforme crece en el OPOT.

60
10. CONCLUSIONES
- El tejido muscular del tiburón piloto presento valores isotópicos más elevados,
que en el tejido vertebral.
- Machos y hembras no presentaron diferencias en los valores de δ13C y δ15N
en músculo y en vértebra, por lo tanto no hay una segregación por sexos.
- Los valores en músculo y vértebra de δ13C y δ15N indican que existe una
segregación por estado ontogénico del tiburón piloto a lo largo de su vida, el
cual hace uso de hábitats diferentes para alimentarse y conforme va
aumentando de tamaño sus requerimientos energéticos cambian.
- Las hembras se alimentan de distintas especies presa a lo largo de su vida, lo
cual se debe probablemente a sus requerimientos energéticos; mientras que
los machos se alimentan de presas similares en todas sus etapas
ontogénicas.
- Los valores de músculo y vértebra indican que hay un traslapo trofico entre
sexos, pero no en estados ontogénicos.
- El tiburón piloto presento valores de un consumidor terciario, debido a que no
se alimenta de presas de gran tamaño en la zona costera y las cadenas
troficas en la zona oceánica son más cortas, por lo tanto los valores de δ15N
no son muy elevados.
- En los valores de las vértebras se observo un patrón, donde las primeras
perforaciones (embrión a neonato) tienen valores menores de δ13C y δ15N, los
cuales van aumentando en la etapa juvenil y adulta.

61
11. BIBLIOGRAFÍA
Abrantes K. y A. Barnett. 2011. Intrapopulation variations in diet and habitat use in a marine
apex predator, the broadnose sevengill shark Notorynchus cepedianus. Mar. Ecol.
Prog. Ser. 442: 133–148 pp.
Andrade Z. 2005. Hábitos alimenticios del tiburón piloto Carcharhinus falciformis en el
Océano Pacífico Oriental. Tesis de Maestría. CICIMAR. La Paz, B.C.S. 121 pp.
Arrington D.A. y K.O. Winemiller. 2002. Preservation effects on stable isotope analysis of
fish muscle. Transactions of the American Fisheries Society.131: 337–342 pp.
Barranco S.L.M. 2008. Hábitos alimenticios y nivel trófico del tiburón Carcharhinus
falciformis Mûller y Henle 1841 (Elasmobranchii: Carcharhinidae) en el Golfo de
Tehuantepec, México por medio del análisis del contenido estomacal e isótopos
estables de δ13Cy δ15N. Tesis de Maestría. Universidad del Mar, Oaxaca, México. 86
pp.
Bearhop S., M. A. Teece, S. Waldron y R. W. Furness. 2000. Influence of lipid and uricacid
on δ13C and δ15N values of avian blood: Implications for trophic studies. The Auk.
117:504–507 pp.
Bearhop S., C.E. Adams, S. Waldron, R.A. Fuller y H. Macleod. 2004 Determining trophic
niche width: a novel approach using stable isotope analysis. Journal of Animal
Ecology. 73:1007-1012 pp.
Blackburn D.G. 1994. Standarized criteria for the recognition of embryonic nutritional
patterns in squamate reptils. Copeia: 925–935 pp.
Bonfil R., M. Meyer, M.C. Scholl, R. Johnson, S. O’Brien, H. Oosthuzien, S. Swanson, D.
Kotze y M. Paterson. 2005. Transoceanic migration, spatial dynamics and population
linkages of white sharks. Science. 310: 100–103 pp.
Braccini J.M., B.M. Gillander y T.I. Walker. 2005. Sources of variation in the feeding ecology
of the piked spurdog (Squalus megalops): Implications for inferring predator-prey
interactions from overall dietary composition. ICES J. Mar. Sci. 6: 1076–1094 pp.

62
Branstetter S., J.A. Musick y J.A. Colvocoresses. 1987. A comparison of the age and growth
of the tiger shark, Galeocerdo cuvieri, from off Virginia and from the Northwestern
Gulf of Mexico.Fish. Bull. 85: 26–279 pp.
Branstetter S. 1990. Early life-history implications ofselected carcharhinoid and lammoid
sharks of the northwest Atlantic. NOAA Technical Reports NMFS. 90:17-28 pp.
Brown J., J.F. Gillooly, A.P. Allen, V.M. Savage y G.B. West. 2004. Toward a metabolic
theory of ecology. Ecology. 85: 1771-1789 pp.
Cabrera Chávez-Costa A. 2000. Determinación de los hábitos alimenticios durante las
estaciones primavera y verano de Carcharhinus falciformis, Sphyrna lewini y
Nasolamia velox (Carcharhiniformes: Carcharhinidae) a partir del análisis de su
contenido estomacal en el Golfo de Tehuantepec, México. Tesis de licenciatura.
UNAM. México. 63 pp.
Cabrera Chávez-Costa A. 2003. Hábitos alimenticios del tiburón piloto Carcharhinus
falciformis (Bibron, 1839) en la costa occidental de Baja California Sur. Tesis de
Maestría. CICIMAR-IPN. 110 pp.
Cailliet G.M. y K.J. Goldman. 2004. Age determination and validation in chondrichthyan
fishes. In: Carrier J., Musick J.A., Heithaus M.R. Eds. Biology of sharks and their
relatives. CRC Press LLC, Boca Raton, Florida. 399–447 pp.
Campana, S. E., L. J. Natanson y S. Myklevoll. 2002. Bomb dating and age determination of
large pelagic sharks. Canadian Journal of Fish Aquaict Science. 59: 450-455
Carpenter S. R. y J.F. Kitchell. 1993. The trophic cascade in lakes. Cambridge University
Press, Cambridge. 19 pp.
Castillo-Géniz, J.L., S.R. Soriano-Velázquez, F. Sancho-Vazquez, C.E. Ramírez-Santiago y
A. Cid del Prado-Vera 2002. Elasmobranquios del Golfo de Tehuantepec, litoral
chiapaneco. CONABIO, México. 47 p.
Castro JI. 1993. The nursery of Bull Bay, South Carolina, with a review of the shark
nurseries of the Southeastern coast of the United States. Environmental Biology of
Fishes. 38: 37-48 pp.
Castro, J.I. 1996. The sharks of North America waters.Texas A & M University Press. Texas,
E.U.A. 180 pp.

63
Carlisle, A.B., S.L. Kim, B.X. Semmens, D.J. Madigan, S.J. Jorgensen, C.R. Perle, S.D.
Anderson, T.K. Chapple, P.W. Kanive y B.A. Block. 2012. Using stable isotope
analysis to understand the migration and trophic ecology of northeastern 57 Pacific
white sharks (Carcharodon carcharias). PloS one. 2: 1-15 pp.
Carlson, J.K., K.J. Goldman y C.G. Lowe. 2004. Metabolism, Energetic Demand, and
Endothermy. En: Carrier J.C., J.A. Musick y R.H. Michae. (Eds.). Biology of Sharks
and their relatives. CRC Press, Boca Raton. 203-246 pp.
Casselman J.M. 1974. Analysis of hard tissue of picke Esox lucius with special reference to
age and growth. En: Bagenal T.B. (Ed.).The ageing of fish. Unwin Brothers Ltd.
England. 13-27 pp.
Castro JI. 1993. The nursery of Bull Bay, South Carolina, with a review of the shark
nurseries of the Southeastern coast of the United States. Environmental Biology of
Fishes. 38: 37-48 pp.
Castro J.I., C. Woodley y R. Brudek. 1999. A preliminary evaluation of the status of shark
species. FAO Fisheries Technical paper. 380p.
Cervantes-Gutiérrez F. 2013. Edad y crecimiento de Carcharhinus falciformis (Müller &
Henle, 1839) en la zona oceánica del Pacífico oriental tropical. Tesis de Maestría.
CICIMAR, 55 pp.
Chapple T.K., S.J. Jorgensen, S.D. Anderson, P.E. Kanive y A.P. Klimley. 2011. A first
estimate of white shark, Carcharodon carcharias, abundance of Central California.
Biol. Lett. 1-4 pp.
CIAT. 2012. Tunas and billfishes in the eastern Pacific Ocean in 2011. Inter-Amer Trop Tuna
Comm Fish Status Rep 9:163 pp
Cohen J. E., S. L. Pimm, P. Yodzis y J. Saldaña. 1993. Body size of animal predator
and animal prey in food webs. Journal of Animal Ecology 62: 67-78 pp.
Compagno L. J. V. 1984. Sharks of the world. An annotated and illustrated catalogue
of shark species known to date. Part 2. Carchariniformes. FAO Fish. Synop. 4: 251-
663 pp.
Compagno L.J.V. y S.F. Cook. 1995. The exploitation and conservation of freshwater
elasmobranchs: status of taxa and prospects for the future. Journal of Aquariculture
and Aquatic Sciences. 7:62-90 pp.

64
Cortés E. y S. Gruber. 1990. Diet, feeding habits and estimated of daily ration of young
lemon sharks, Negaprion brevirostris (Poey). Copeia. 1: 204-218 pp.
Cortés E. 1999. Standardized diet compositions and trophic levels of sharks. ICES Journal
Marine Science. 56: 707-717 pp.
Darwin Ch. 1959. El origen de las especies por medio de la selección natural. Tomo I.
Estudio preliminar de Juan Comas. Universidad Nacional Autónoma de México,
México 353 pp.
DeNiro M. J. y S. Epstein. 1977. Mechanism of carbon isotope fractionation associated with
lipid synthesis. Science. 197: 261–263 pp.
DeNiro M. J. y S. Epstein. 1981. Influence of diet on the distribution of nitrogen in animals.
Geochim. Cosmochim. Acta. 45: 341-351 pp.
Diario Oficial de la Federación (DOF). 2004. Tiburón: segunda edición. Lunes 15 de Marzo
de 2004. 126 pp.
Díaz-García D. A. 2006. Construcción de un modelo trófico y análisis de la posición trófica
del grupo lutjanidae a partir de datos de la captura incidental camaronera de
Alvarado Veracruz. Tesis de licenciatura. Instituto Tecnológico de Boca del Rio
Veracruz. 78pp.
DiBattista J.D., K.A. Feldheim, X. Thibert-Plante, S.H. Gruber y A.P. Hendry. 2008. A
genetic assessment of polyandry and breeding-site fidelity in lemon sharks. Molecular
Ecology. 17: 783-795 pp.
Dulvy K. N., J.K. Baum, S. Clarke, L.J.V. Compagno, E. Cortés, A. Domingo, S. Fordham, S.
Fowler, M.P. Francis, C. Gibson, J. Martínez, J.A. Musick, A. Soldo, J.D. Stevens y S.
Valenti. 2008. You can swim but you can´t hide: the global status and conservation of
oceanic pelagic sharks and rays. Aquatic Conservation: Marine and Freshwater
Ecosystems. 459-482 pp.
Ehleringer JR y P.W. Rundel. 1989 Stable isotopes: History, units, and instrumentation. En:
P.W. Rundel, J.R. Ehleringer y K.A. Nagy (Eds.) Stable isotopes in ecological
research, Springer-Verlag. U.S.A. 1-15 pp.
Ellis J. R., M.G. Pawson y S.F. Shackley. 1996. The comparative feeding ecology of six
species of sharks and four species of ray (Elasmobranchii) in the north-east Atlantic.
Journal of the Marine Biological Association of the United Kingdom. 76: 89–106 pp.

65
Elton Ch. 1927 Animal ecology. The Macmillan Company. U.S.A. 178 pp.
Estupiñán C. y L. Cedeño. 2005. Estudio de los hábitos alimenticios mediante el análisis del
contenido estomacal del tollo mico Carcharhinus falciformis (Bibron, 1839) y los
tiburones martillo Sphyrna lewini (Griffith & Smith, 1834) y Sphyrna zygaena
(Linnaeus, 1758), en el área de Manta – Ecuador. Tesis de Licenciatura. Universidad
Laica Alfaro de Manabi. Manta, Ecuador. 133 pp.
FAO. 1998. Noticias: tiburones y aves marinas en el programa de una reunión internacional
de pesca. 22 de Octubre de 1998: http://www.fao.org/Noticias/1998/981009-s.htm
FAO. 1998. Report of the Consultation on the Management of Fishing Capacity,
Shark Fisheries and Incidental catch of Seabirds in Longline Fisheries. FAO Fish.
Rep. No. 593.
FAO. 2003. Fisheries Management-2. The Ecosystem Approach to Fisheries. FAO
Technical Guidelines for Responsible Fisheries. No. 4 Suppl. 2.FAO, Rome. At:
http://www.safmc.net/Portals/0/EMHome/MafacReportEcoMgmt.pdf. Accessed 4 April
2007
FAO. 2006. Taller sobre evaluación y Manejo de Elasmobranquios en América del Sur y
Bases Regionales para los planes de Acción (DINARA/FAO). Montevideo, Uruguay,
7-9 de Noviembre de 2005. Roma. 64 pp
Ferry-Graham. L.A. 1998. Effects of prey size and mobility on prey-capture Kinematics in
leopard sharks Triakis semifaciata. J. Exp. Biol. 201:2433-2444.
Focken, U. y K. Becker. 1998. Metabolic fractionation of stable carbon isotopes:implications
of different proximate compositions for studies of the aquatic food webs using δ13C
data. Oecologia. 115: 337–343.
Froese. R. y D. Pauly 2007. Fish Base World Wide Web. Publicación electrónica
.http://www.fishebase.org. (12/2007).
Fry B. y E.B. Sherr. 1984. 13C measurements as indicators of carbon flow in marine and
freshwater ecosystems. Contributions in Marine Science. 27:13–47 pp.
Fry B. y E.B. Sherr. 1988. d13C measurements as indicators of carbon flow in marine and
freshwater ecosystems. In Stable isotopes: Integration of biological ecological and
geochemical processes. En H. Griffiths (Ed.). Environmental plant biology series Bios
Scientific Publishers. 196–229 pp.

66
Galván-Magaña F., H. Nienhuis y P. Klimley. 1989. Seasonal abundance and feeding habits
of sharks of the lower Gulf of California, Mexico. Calif. Fish and Game. 75:74-84 pp.
Gannes L.Z., D.M. O’Brien y C. Martınez del Rio. 1997. Stable isotopes in animal ecology:
assumptions, caveats, and a call for more laboratory experiments. Ecology. 78:
1271–1276 pp.
Gerking S. D. 1994. Feeding ecology of fish. Academic Press. U.S.A. 416 pp.
Gilbert P. y D. Schlernitzauer. 1966. The placenta and gravid uterus of Carcharhinus
falciformis. Copeia. 3: 451-457 pp.
Glasstone S. 1976. Madrid, España. 1180 pp.
Graham B.S., D. Grubbs, K. Holland y B.N. Popp. 2007. A rapid ontogenetic shift in the diet
of juvenile yellowfin tuna from Hawaii. Mar. Biol. 150: 647-658 pp.
Hernández-Aguilar S. 2008. Espectro trófico del tiburón azul, Prionace glauca (Linnaeus,
1758) en la costa occidental de Baja California Sur, México. Tesis de Maestría en
Ciencias.CICIMAR. 86 pp.
Hoyos-Padilla E. M. 2003. Biología reproductiva del tiburón piloto Carcharhinus falciformis
(Bibron, 1839) de Baja California Sur. Tesis de Maestría. CICIMAR-IPN. 58 pp.
Hueter R.E., M.R. Heupel, E.J. Heist y D.B. Keeney. 2005. Evidence of philopatry in sharks
and implications for the management of shark fisheries. Journal of Northwest Atlantic
Science. 35:239–247 pp.
Jackson A., R. Inger, A. Parnell y S. Bearhop. 2011. Comparing isotopic niche widths
among and within communities: SIBER - Stable Isotope Bayesian Ellipses in R. J.
Anim. Ecol. 3: 595–602 pp.
Jeppesen E., J.P. Jensen, M. Sondergaard, T.L. Lauridsen, P. Junge y L. Jensen. 1997.
Top-down control in freshwater lakes: the role of nutrient state, submerged
macrophytes and water depth. Hydrobiologia. 343:151-164 pp.
Johnston P. G. y I. Zucker. 1983. Lability and diversity of circadian rhythms of the cotton rat
Sigmodon hispidus. American Journal of Physiology–Regulatory Integrative and
Comparative Physiology. 244: 338–346 pp.
Kaehler S. y E.A. Pakhomov. 2001. Effects of storage andpreservation on the delta C-13
and delta N-15 signatures of selected marine organisms. Mar. Ecol. Prog. Ser. 219:
299-304 pp.

67
Keeney D.B., M. Heupel, R.E. Hueter y E.J. Heist. 2003. Genetic heterogeneity among
blacktip shark, Carcharhinus limbatus, continental nurseries along the U.S. Atlantic
and Gulf of Mexico. Marine Biology. 143: 1039-1046 pp.
Kim S. y P. Koch. 2011. Methods to collect, preserve, and prepare elasmobranch tissues for
stable isotope analysis. Environ. Biol. Fish. 95: 53-63 pp.
Kim S.L y P.L. Koch. 2012 Methods to collect, preserve, and prepare elasmobranch tissues for stable
isotope analysis. Environ Biol Fishes. 1: 53-63 pp.
Klimley. 1983. Social organization of schools of the scalloped hammerhead, Sphyrna lewini
in the Gulf of California. Doctoral dissertation. University of California, San Diego, La
Jolla. 341 pp.
Knauss J. A. 1960. Measurements of the Cromwell Current. Deep Sea Research. 6: 265-
286 pp.
Kronfeld–Schor N. y T. Dayan. 2003. Partitioning of time as an ecological resource. Annual
Review of Ecology, Evolution and Systematics. 34: 153–181 pp.
Layman C. A., K.O. Winemille, D.A. Arrington y D.B. Jepsen. 2005. Body size and trophic
position in a diverse tropical food web. Ecology. 86: 2530-2535 pp.
Lawlor L. 1980. Overlap, similarity and competition coefficients. Ecology. 2 :245-25 pp.
Litvinov F.F. 1989. The structure of the epipelagic Elasmobranchii taxocene in the Atlantic
and Pacific oceans and its changes in the recent geological time. Traducción del ruso
por el autor (comunicación personal). Voprosy Ichthyologii. 6: 973–984 pp.
Lopez-Ibarra G.A. 2008. Estructura trófica de los copépodos pelagicus en el Océano
Pacífico Oriental Tropical. Tesis Doctorado. CICIMAR-IPN. 92 pp.
Lowe G.C., B.M. Wetherbee, G.L. Crow y A.L. Tester. 1996. Ontogenetic dietary shifts and
feeding behavior of the tiger shark Galeocerdo cuvier,in Hawaiian waters. Env. Biol.
Fish. 47: 203-211 pp.
MacNeil MA, G.B. Skomal y A.T. Fisk. 2005. Stable isotopes from multiple tissues reveal
diet switching in sharks. Mar. Ecol. Prog. Ser. 302: 199-206 pp.
MacNaughton S. y L. Wolf. 1973. General Ecology. 2nd Edition. Holt, Rinehart y Winston.
USA. 702 pp

68
Marín O.R. 1992. Aspectos biológicos de los tiburones capturados en las costas de
Tamaulipas y Veracruz, México. Tesis Profesional. Facultad de biología, Universidad
Veracruzana. Xalapa, Ver., México. 146 pp.
Markaida U. y O. Sosa-Nishizaki. 2010. Food and feeding habits of the blue shark Prionace
glauca caught off Ensenada, Baja California, Mexico, with a review on its feeding.
Journal of theMarine Biological Association of the United Kingdom. 90: 977-994 pp.
Maya- Meneses C.I. 2012. Traslapo trofico del tiburón azul (Prionace glauca) y tiburón mako
(Isurus oxyrinchus) capturados en la costa occidental de baja california sur. Tesis de
Licenciatura. UABCS. 65pp.
McConnaughey, T. y Mcroy C. 1979. Food-web structure and fractionation of carbon
isotopes in the Bering Sea. Marine Biology. 53: 257–262 pp.
Motta R.L. y V.S. Uieda. 2005. Food web structure in a tropical stream ecosystem. Austral
Ecology. 30: 58-73 pp.
McMeans B, J. Olin y G. Benz. 2009. Stable isotope comparisons between embryos and
mothersof a placentatrophic shark species. Journal of Fish Biology. 75: 2464-2474
pp.
McNab B. K. 2002. The physiological ecology of vertebrates. A view from energetics. Cornell
University Press,New York. 184 pp.
Minami M., C.E. Lennert-Cody y M. Román-Verdesoto. 2007. Modeling shark by catch: the
zero-inflated negative binomial regression model with smoothing. Fisheries Research.
84: 210–221 pp.
Motta R.L. y V.S. Uieda. 2005. Food web structure in a tropical stream ecosystem. Aust.
Ecol. 30: 58-73 pp.
Newsome S.D., DL. Phillips, B.J. Culleton, T.P. Guilderson y P.L. Koch. 2004. Dietary
reconstruction of an Early to Middle Holocene human population from the central CA
coast: insights from advanced stable isotope mixing models. Journal of
Archaeological Science. 3: 1101-1115 pp.
Odum E.P. 1972. Ecología. Nueva Editorial Interamericana. Tercera edición. 639 pp.
Otake T. 1990. Classification of reproductive modes in sharks with comments on female
reproductive tissue and structures. NOAA Tech. Rep. NMFS. 93: 111-130 pp.

69
Pace M. L., J.J. Cole, S.R. Carpenter, J.F. Kitchell, J.R. Hodgson, M.C. Van de Bogert, D.L.
Bade, E.S. Kritzberg y D. Bastviken. 2004. Whole-lake Carbon-13 additions reveal
terrestrial support of aquatic food webs. Nature. 427: 240-243 pp.
Park R. y S. Epstein. 1961. Metabolic fractionation of 13C and 12C in plants. Plants
Physiology. 36: 133-138 pp.
Peterson B.J. y B. Fry. 1987. Stable isotopes in ecosystem studies. Annual Review of
Ecology and Systematics. 18: 293-320 pp.
Pennington T.J., L.M. Kevin-Kuwahara, V.S. Kolber, D.D. Calientes y P.F. Chavez. 2006.
Primary production in the eastern tropical Pacific: A review. Progress in
Oceanography. 69: 287-17 pp.
Pianka E. 1973. The structure of lizard communities. Annual Review of Ecology and
Sistematics. 4: 53-74 pp.
Pianka E. R. 1974. Niche overlap and diffuse competition. Proceedings of the National
Academy of Science U.S.A. 17: 2141–2145 pp.
Pinnegar J.K. y N.V.C. Polunin. 1999. Differential fractionation of d13C and d15N among
fish tissues: implications for the study of trophic interactions. Functional Ecology. 13:
225–231 pp.
Pimm S. L. 1982. Food webs. Chapman y Hall. Londres. Reino Unido.
Pimm S.L.,Lawton J.H. y Cohen J. E. 1991. Food web patterns and their consequences.
Nature 350: 669-74 pp.
Post D.M. 2002. Using stable isotopes to estimate trophic position models, methods, and
assumptions. Ecology. 3: 703-718 pp.
Post D. M., C.A. Layman, G. Takimoto, J. Quattrochi y C.G. Montaña. 2007. Getting to the
fat of the matter: models, methods and assumptions for dealing with lipids in stable
isotope analyses. Ecologia. 152: 179-189 pp.
Pratt H.L. Jr. y J.G. Casey. 1990. Shark reproductive strategies as a limiting factor in
directed fisheries, with a review of Holden’s method of estimating growth parameters.
In: Pratt H.L.Jr., S.H. Gruber, T. Taniuchi (Eds.) Elasmobranchs as living resources.
NOAA Tech. Rep. NMFS. 90: 97–110 pp.

70
Rau G.H., J.L. Teyssie, F. Rassoulzadegan y S.W. Fowler. 1990. 12C/13C and I5N/l4N
variations among size fractionated marine particles: implications for their origin ant
trophic relationships. Mar. Ecol.Prog. Ser. 59: 33-38 pp.
Román-Reyes J. C. 2005. Análisis de contenido estomacal y la razón de isótopos estables
de carbono (13C) y nitrógeno (15N) del atún aleta amarilla (Thunnus albacares), delfín
manchado (Stenella attenuata) y delfín tornillo (Stenella longirostris) del Océano
Pacífico Oriental. Tesis Doctorado. CICIMAR-IPN. 143pp.
Sainsbury K.J., A.E. Punt, y A.D.M. Smith. 2000. Design of operational management
strategies for achieving fishery ecosystem objectives. ICES Journal of Marine
Science. 57: 731–741 pp.
Sánchez de Ita J.A., C. Quiñónez-Velázquez, F. Galván-Magaña, N. Bocanegra-Castillo y F.
Félix-Uraga. 2011. Age and growth of the silky shark Carcharhinus falciformis from
the west coast of Baja California Sur, México. Journal of Applied Ichthyology. 27: 20-
24 pp.
Schindler D.E. y M.D. Scheuerel. 2002. Habitat coupling in lake ecosystems. Oikos. 98: 177-
189 pp.
Simpfendorfer C.A. y N.E. Milward. 1993. Utilization of a tropical bay as a nursery area by
sharks of the families Carcharhinidae and Sphyrnidae. Environmental Biology of
Fishes. 37: 337-345 pp.
Sotiropoulos M.A., W.M. Tonn y L.I. Wassenaar. 2004. Effects of lipid extraction on stable
carbon and nitrogenisotope analyses of fish tissues: potential consequences for food
web studies. Ecol. Fresh. Fish. 13: 155-160 pp.
Springer S. 1967. Social organization of shark populations. En: Mathewson P.W. y D.P. Rall
(Eds). Sharks, skates and rays. John Hopkins Press, Baltimore. 149-174 pp.
Sweeting C.J., N.V.C. Polunin y S. Jennings. 2006. Effects of chemical lipid extraction and
arithmetic lipid correction on stable isotope ratios of fish tissues. Rapid Commun.
Mass Spectrom. 20: 595–601 pp.
Torres-Rojas Y.E., F.P. Osuna, A. Hernández-Herrera, F. Galván-Magaña, S.A. Gracia, H.
Villalobos-Ortiz y L. Sampson. 2014.Feeding grounds of juvenile scalloped
hammerhead sharks (Sphyrna lewini) in the south-eastern Gulf of California.
Hydrobiologia..726(1): 81-94.

71
Unión Mundial para la Naturaleza (UICN). 2003. Introducción: Biodiversidad y Hábitat.
Centro de cooperación del Mediterráneo. 6: 37pp.
Vanderklift M.A. y S. Ponsard. 2003. Sources of variation in consumer-diet delta N-15
enrichment: a meta-analysis. Ecología. 136: 169-182 pp.
Vaudo J., P. Matich y M. Heithaus. 2010 Mother–offspring isotope fractionation in two
species of placentatrophic sharks. Journal of Fish Biology. 77: 1724-1727 pp.
Vögler R., A.C. Milessi y L.O. Duarte. 2008. Changes in trophic level of Squatina
guggenheim with increasing body length: relationships with type, size and 68 trophic
level of its prey. Environ. Biol. Fish. 1: 41–52 pp.
Wetherbee B. S. Gruber y E. Cortes. 1990: Diet, feeding habits, digestion and consumption
in sharks, with special reference to thelemon shark, Negaprion brevirostris. En:
Elasmobranchs as living resources: advances in the Biology, Ecology, Systematics
and the status of the Fisheries. H. L. Pratt Jr, S. H. Gruber, T. Taniuchi (Eds), NOAA
Tech. Rep 90, Washington, U.S.A. 29–47 pp.
Winemiller K.O. 1990 Spatial and temporal variation in tropical fish trophic networks.
Ecological Monographs. 60: 331-367 pp.
Woodward G., B. Ebenman, M. Emmerson, J.M. Montoya, J.M. Olesen, A. Valido y P.H.
Warren. 2005. Body size in ecological networks. Trends in Ecology and Evolution. 20:
402-409 pp.
Wooton R.J. 1990. Ecology of teleost fishes. Champman y Hall. U.S.A. 404 pp.
Wourms J.P. 1993. Maximization of evolutionary trends for placental viviparity in the
spadenose shark, S. laticaudus. Environ. Biol. Fishes. 38: 269-294 pp.
Wyrtki K. 1965. Corrientes superficiales del Océano Pacífico Oriental Tropical. Comisión
Interamericana del Atún Tropical. 5: 295-304 pp.
Citas en línea
1- http://es.wikipedia.org/wiki/Espectr%C3%B3metro_de_masas
2- http://es.wikipedia.org/wiki/Ontogenia
3- http://www.biodiversityexplorer.org/chondrichthyes/elasmobranchii/galeomorphii/carcharh
inus_falciformis.htm
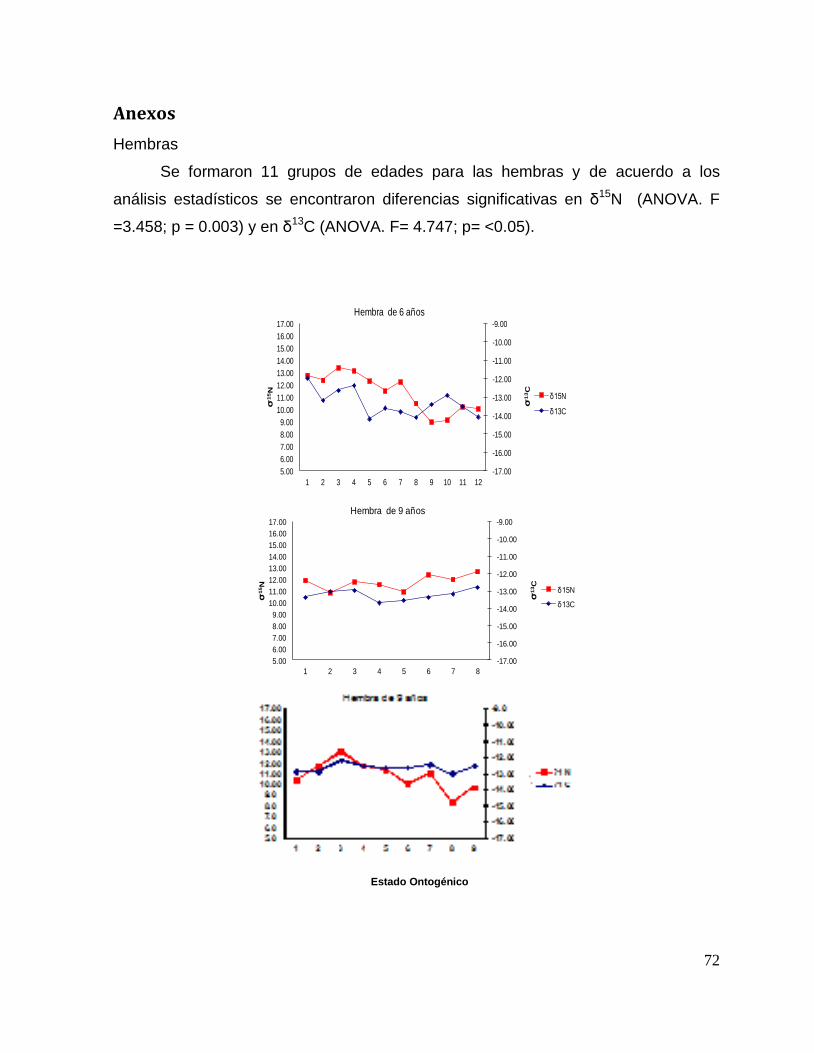
72
Anexos
Hembras
Se formaron 11 grupos de edades para las hembras y de acuerdo a los
análisis estadísticos se encontraron diferencias significativas en δ15N (ANOVA. F
=3.458; p = 0.003) y en δ13C (ANOVA. F= 4.747; p= <0.05).
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12
σ13C
σ15N
Hembra de 6 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8
σ13C
σ15N
Hembra de 9 años
δ15N
δ13C
Estado Ontogénico
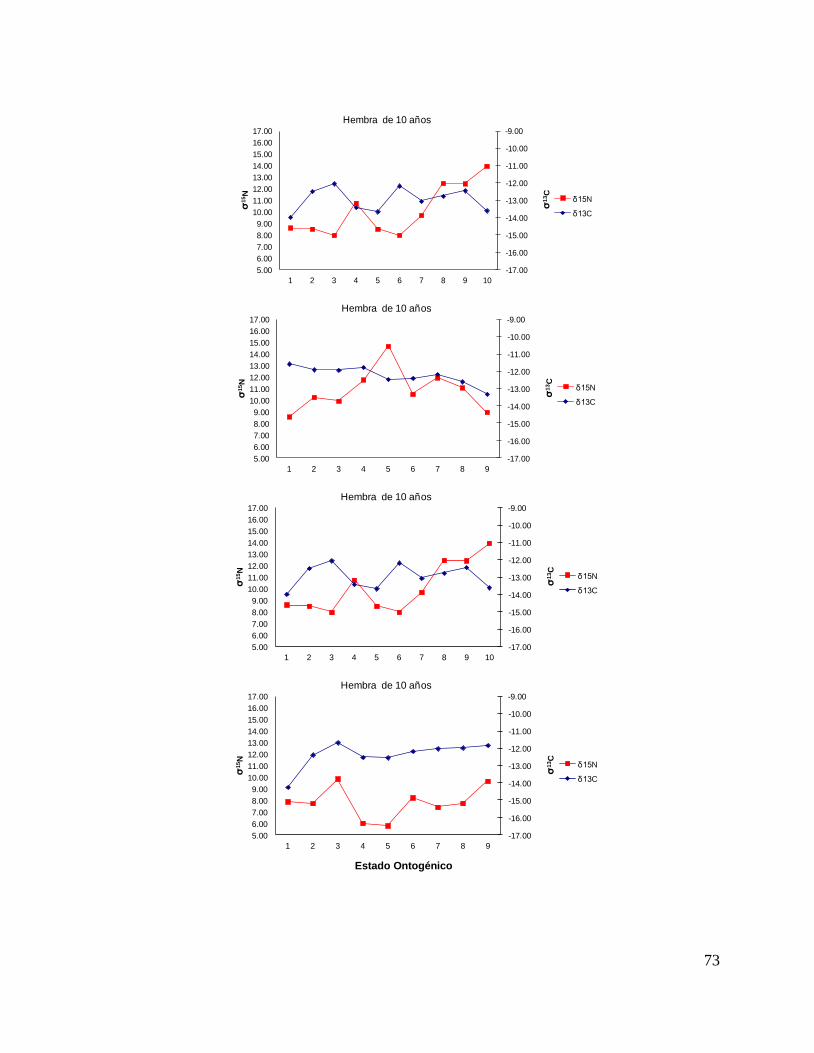
73
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 10 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9
σ13C
σ15N
Hembra de 10 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 10 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9
σ13C
σ15N
Hembra de 10 años
δ15N
δ13C
Estado Ontogénico
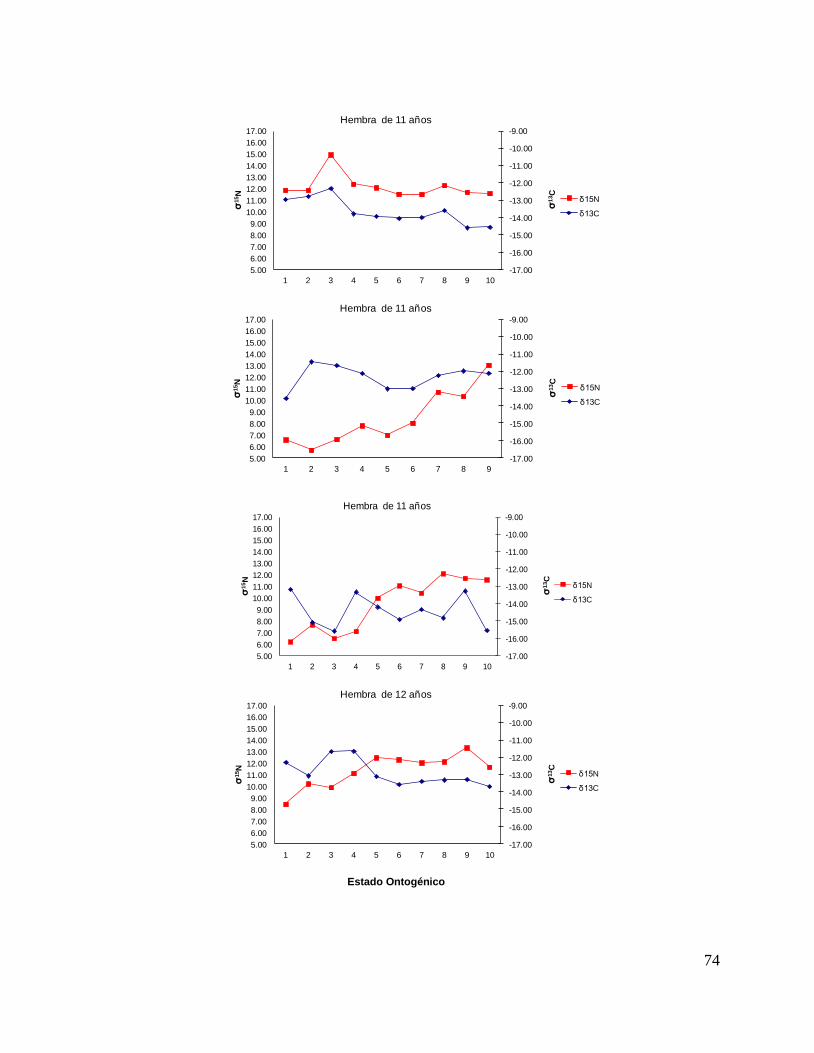
74
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 11 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9
σ13C
σ15N
Hembra de 11 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 11 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 12 años
δ15N
δ13C
Estado Ontogénico
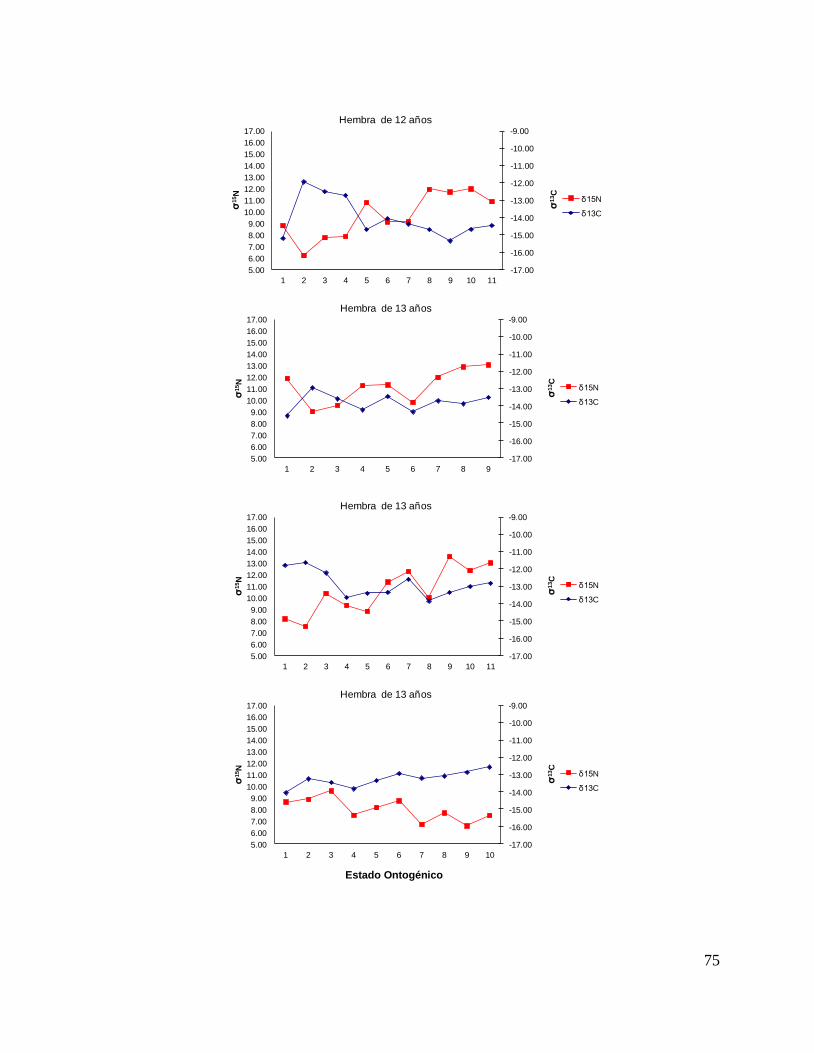
75
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11
σ13C
σ15N
Hembra de 12 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
Estado Ontogénico
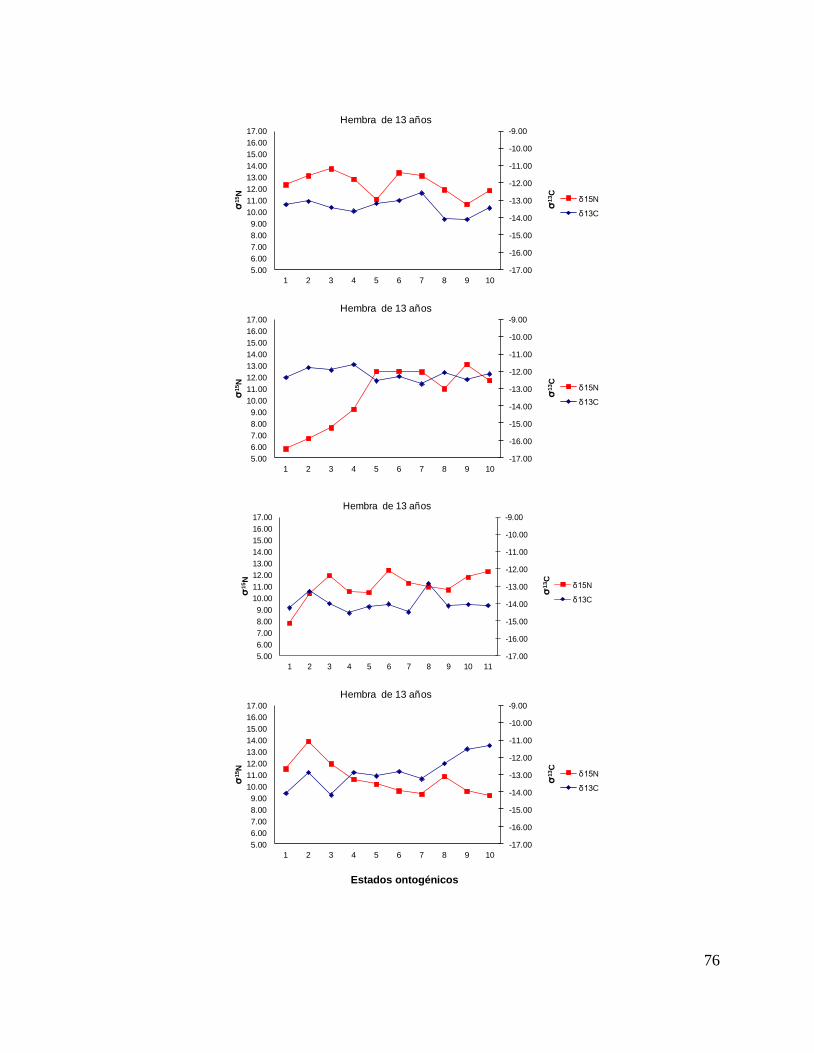
76
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
Estados ontogénicos
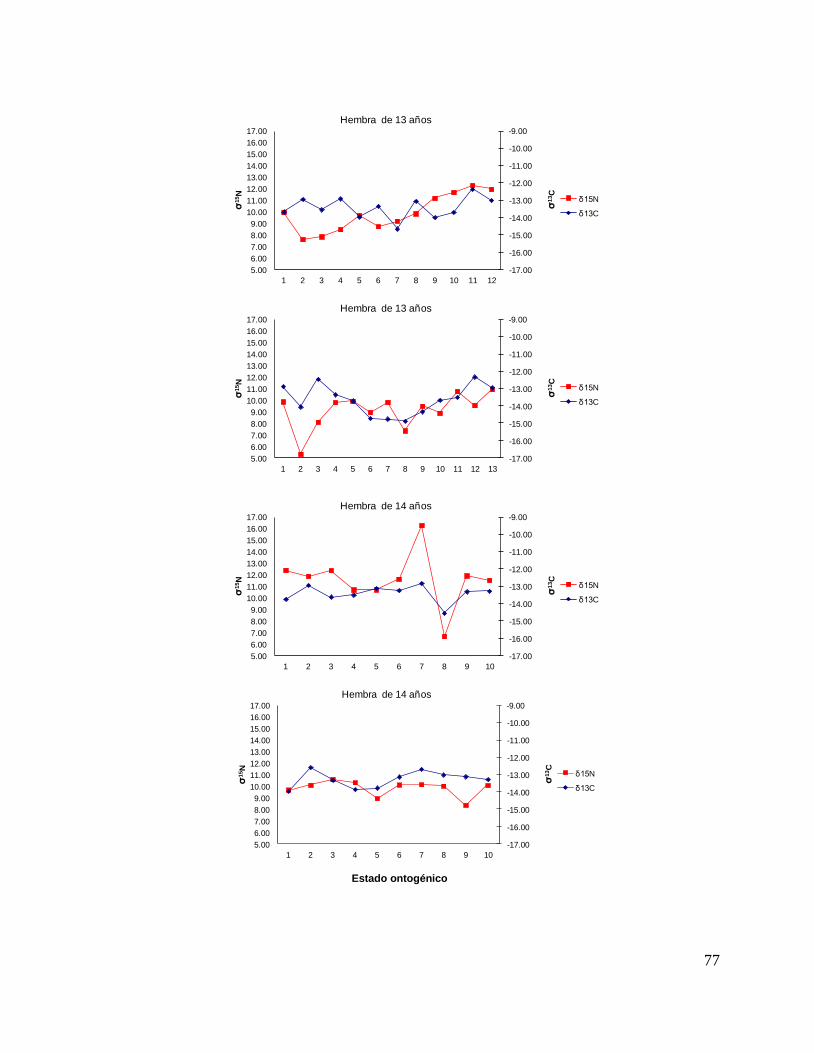
77
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12 13
σ13C
σ15N
Hembra de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 14 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Hembra de 14 años
δ15N
δ13C
Estado ontogénico
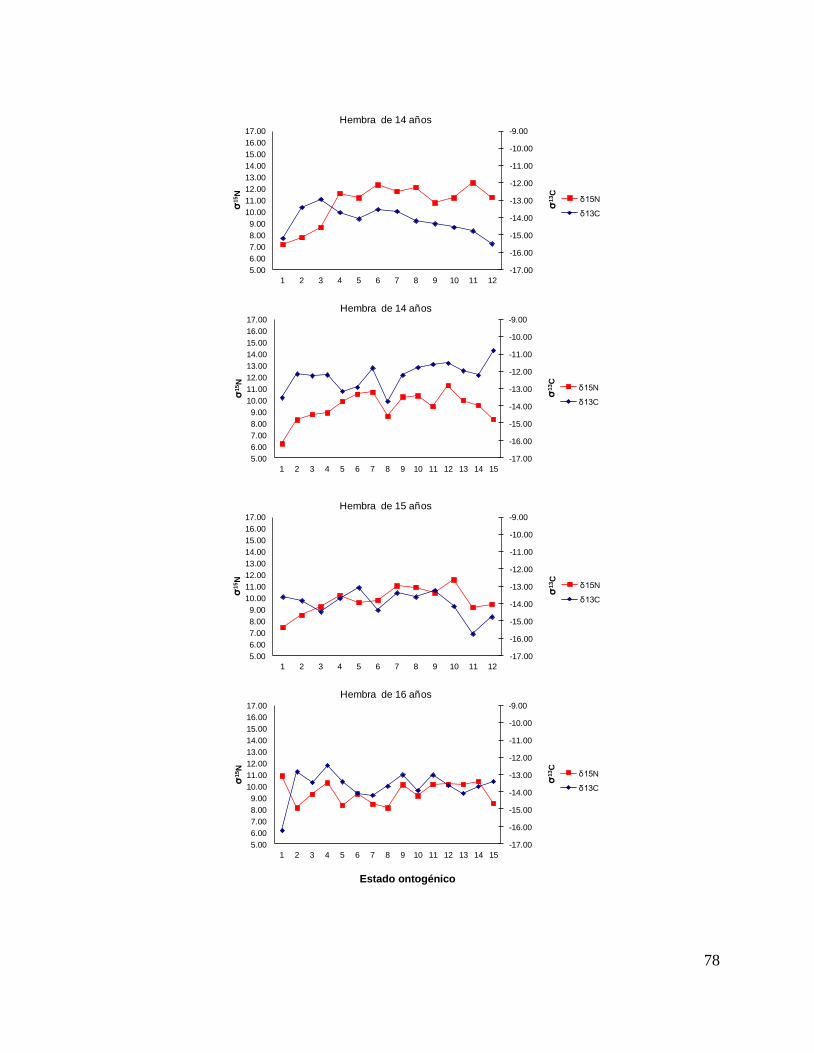
78
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12
σ13C
σ15N
Hembra de 14 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
σ13C
σ15N
Hembra de 14 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12
σ13C
σ15N
Hembra de 15 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
σ13C
σ15N
Hembra de 16 años
δ15N
δ13C
Estado ontogénico
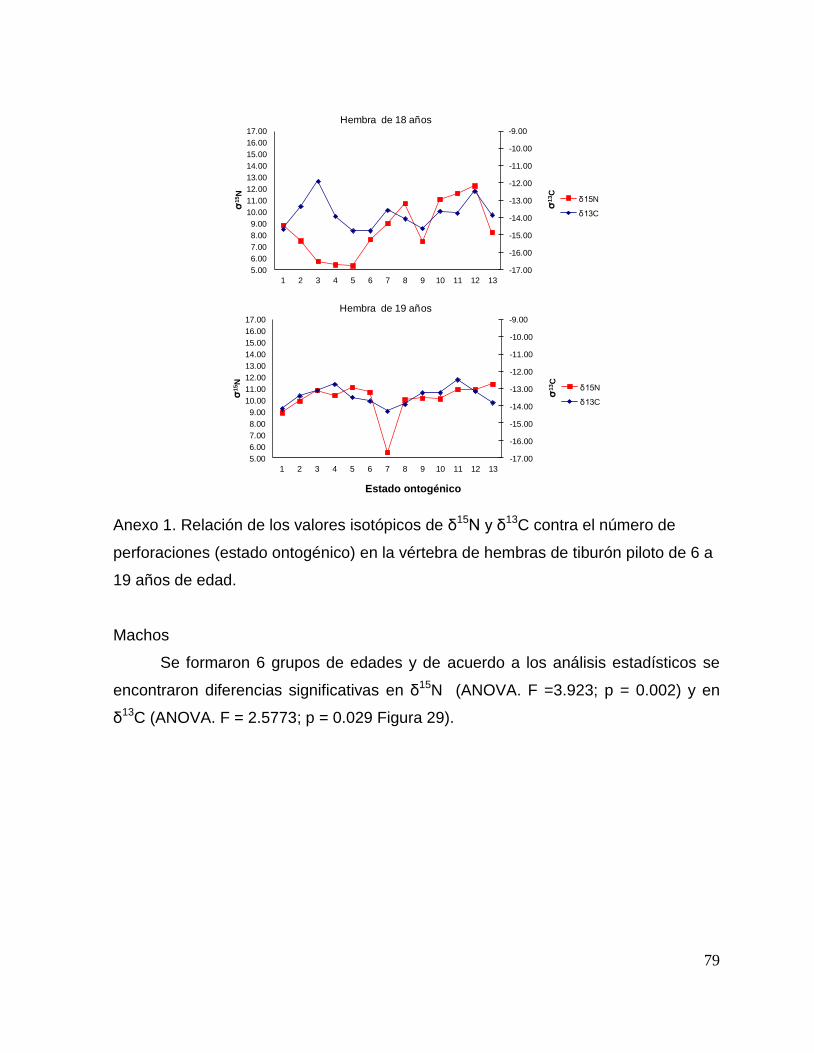
79
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12 13
σ13C
σ15N
Hembra de 18 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12 13
σ13C
σ15N
Hembra de 19 años
δ15N
δ13C
Anexo 1. Relación de los valores isotópicos de δ15N y δ13C contra el número de
perforaciones (estado ontogénico) en la vértebra de hembras de tiburón piloto de 6 a
19 años de edad.
Machos
Se formaron 6 grupos de edades y de acuerdo a los análisis estadísticos se
encontraron diferencias significativas en δ15N (ANOVA. F =3.923; p = 0.002) y en
δ13C (ANOVA. F = 2.5773; p = 0.029 Figura 29).
Estado ontogénico
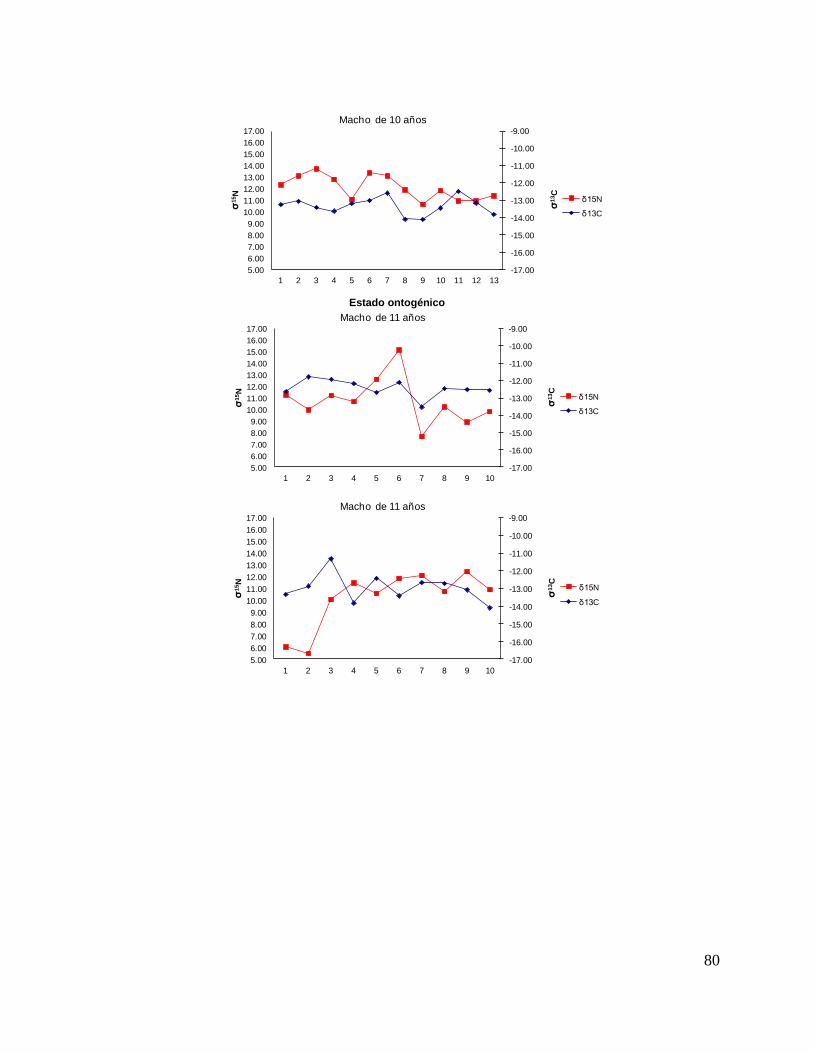
80
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12 13
σ13C
σ15N
Macho de 10 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Macho de 11 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Macho de 11 años
δ15N
δ13C
Estado ontogénico
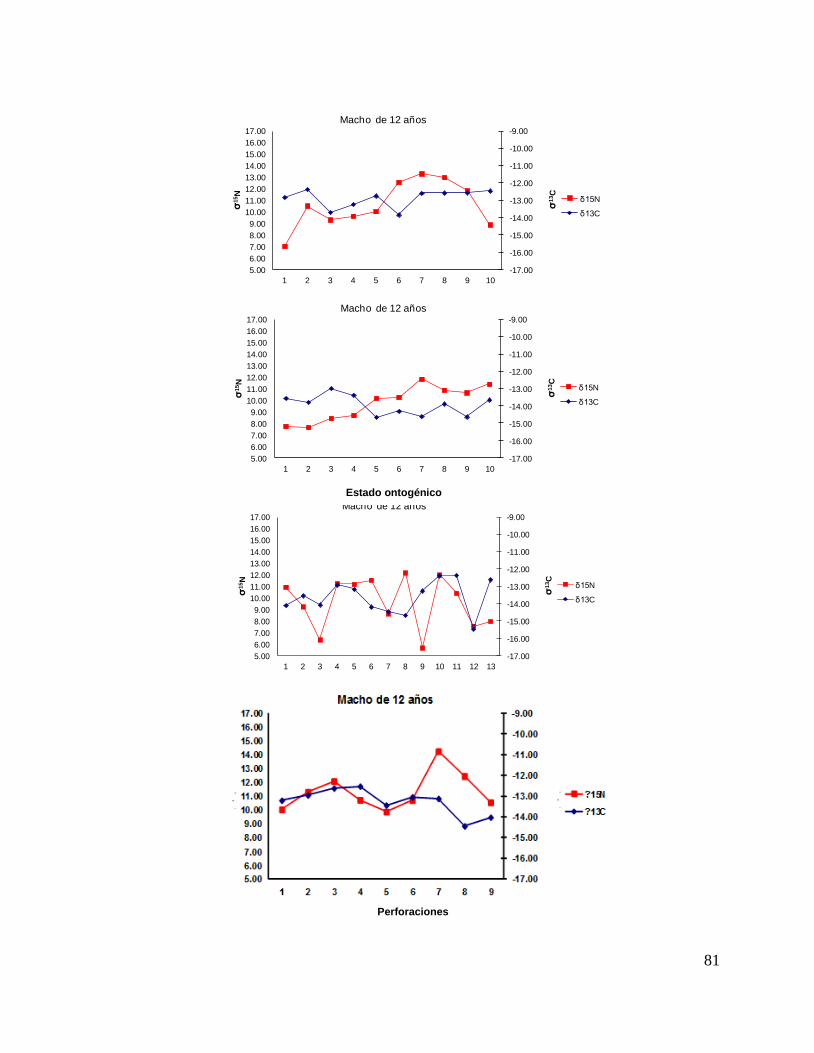
81
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Macho de 12 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10
σ13C
σ15N
Macho de 12 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12 13
σ13C
σ15N
Macho de 12 años
δ15N
δ13C
Estado ontogénico
Perforaciones
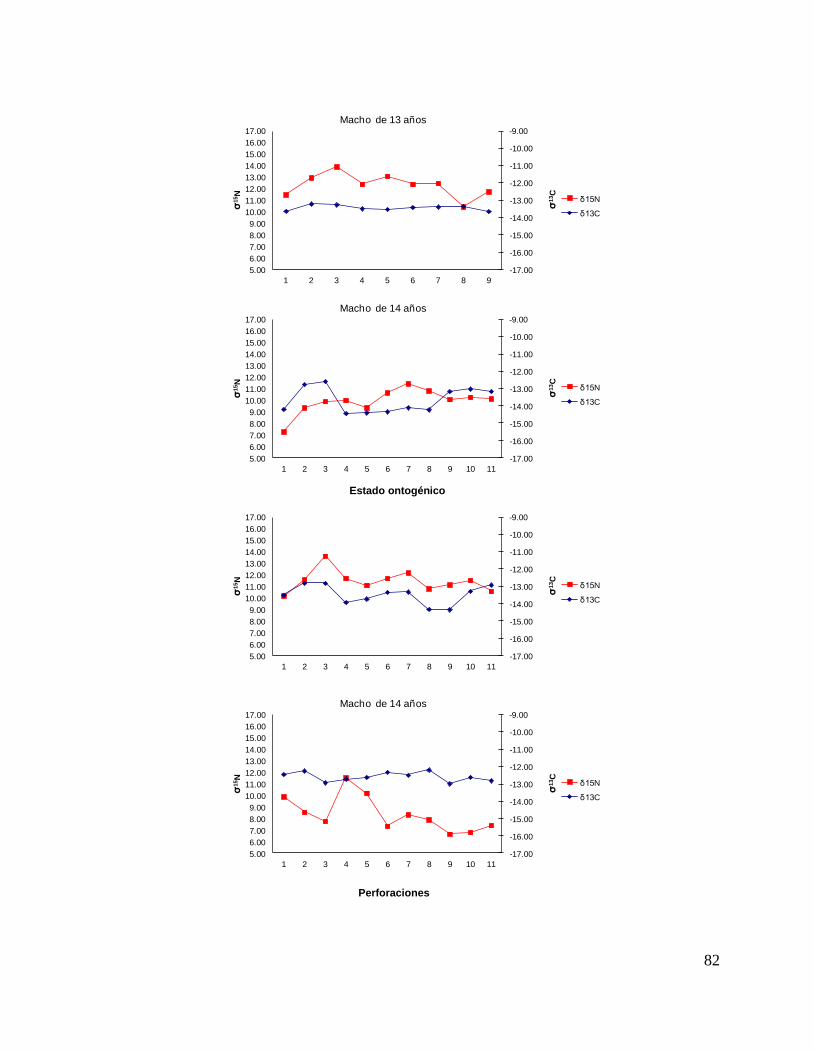
82
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9
σ13C
σ15N
Macho de 13 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11
σ13C
σ15N
Macho de 14 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11
σ13C
σ15N
Macho de 14 años
δ15N
δ13C
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11
σ13C
σ15N
Macho de 14 años
δ15N
δ13C
Perforaciones
Estado ontogénico
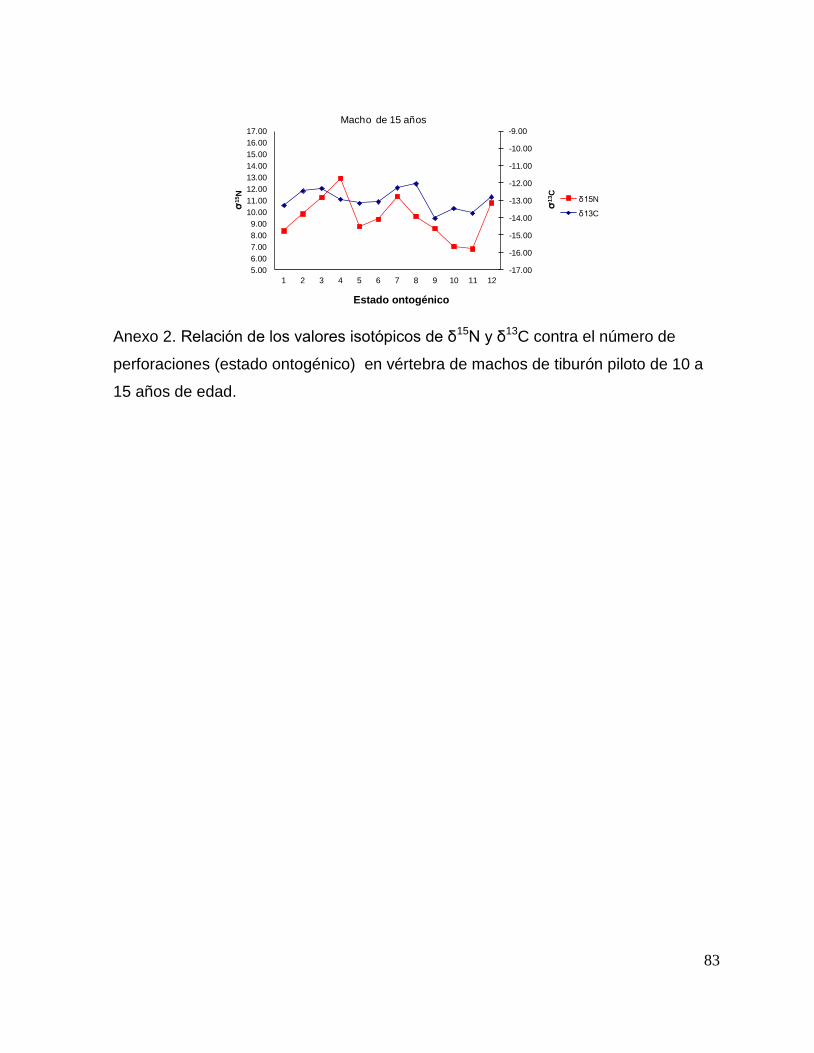
83
-17.00
-16.00
-15.00
-14.00
-13.00
-12.00
-11.00
-10.00
-9.00
5.00
6.00
7.00
8.00
9.00
10.00
11.00
12.00
13.00
14.00
15.00
16.00
17.00
1 2 3 4 5 6 7 8 9 10 11 12
σ13C
σ15N
Macho de 15 años
δ15N
δ13C
Anexo 2. Relación de los valores isotópicos de δ15N y δ13C contra el número de
perforaciones (estado ontogénico) en vértebra de machos de tiburón piloto de 10 a
15 años de edad.
Estado ontogénico