Comparison of R-, S- , and RS -albuterol interaction with human β1 and β2-adrenergic receptors
9
Clinical Reviews in Allergy and Immunology @Copyright 1996 by Humana Press Inc. 1080-0549/96/37-45/$7.25 Comparison of R-, S-, and RS-Albuterol Interaction with Human 131- and 132-Adrenergic Receptors Raymond B. Penn, 7 Thomas FrielleZ John R. McCullough, 3 Gunnar Aberg, 3 and Jeffrey L. Benovic *,1 ~Department of Pharmacology, Thomas Jefferson University, Philadelphia, PA 19107; 2Department of Pharmacology, Pennsylvania State University Hershey Medical Center, Hershey, PA; and 3Sepracor Inc., Marlborough, MA Introduction Albuterol is a 0-adrenergic receptor (~AR) agonist widely used in the management of asthma. In airway smooth muscle, OAR activation leads to increased cAMP generation and activation of the cAMP- dependent protein kinase, which in turn promotes phosphorylation of intracellular proteins and subsequent airway smooth muscle relaxation (1). In all formulations, albuterol is a racemic mixture, comprised of equal amounts of the R- and S-isomers. The respective contributions of the R- and S-isomers in mediating the effects of the racemic mixture are not known. Previous studies have characterized [3ARbinding properties of racemic albuterol to guinea pig lung membranes (2) and have also measured the potency of racemic albuterol in relaxing precontracted airway smooth muscle (2,3). Although numerous studies have shown that the R-isomers of other ~-agonists are at least an order of magni- tude more potent than the corresponding S-stereoisomers (4), limited information exists examining the pharmacological properties of the R- and S-isomers of albuterol. Previous assessments of ff-agonist activity and potency have used relaxation of airway smooth muscle precontracted with muscarinic ago- nists. However, because muscarinic receptor activation of Gi~ can attenu- ate OAR-mediated signal transduction, such measurements are conditional *Author to whom all correspondence and reprint requests should be addressed. Clinical Reviews in Allergy and Immunology 37 Volume 14, 1996
-
Upload
raymond-b-penn -
Category
Documents
-
view
213 -
download
0
Transcript of Comparison of R-, S- , and RS -albuterol interaction with human β1 and β2-adrenergic receptors

Clinical Reviews in Allergy and Immunology @Copyright 1996 by Humana Press Inc. 1080-0549/96/37-45/$7.25
Comparison of R-, S-, and RS-Albuterol Interaction
with Human 131- and 132-Adrenergic Receptors
Raymond B. Penn, 7 Thomas FrielleZ John R. McCullough, 3 Gunnar Aberg, 3
and Jeffrey L. Benovic *,1
~Department of Pharmacology, Thomas Jefferson University, Philadelphia, PA 19107; 2Department of Pharmacology, Pennsylvania State University
Hershey Medical Center, Hershey, PA; and 3Sepracor Inc., Marlborough, MA
Introduction Albuterol is a 0-adrenergic receptor (~AR) agonist widely used in
the management of asthma. In airway smooth muscle, OAR activation leads to increased cAMP generation and activation of the cAMP- dependent protein kinase, which in turn promotes phosphorylation of intracellular proteins and subsequent airway smooth muscle relaxation (1). In all formulations, albuterol is a racemic mixture, comprised of equal amounts of the R- and S-isomers. The respective contributions of the R- and S-isomers in mediating the effects of the racemic mixture are not known. Previous studies have characterized [3AR binding properties of racemic albuterol to guinea pig lung membranes (2) and have also measured the potency of racemic albuterol in relaxing precontracted airway smooth muscle (2,3). Although numerous studies have shown that the R-isomers of other ~-agonists are at least an order of magni- tude more potent than the corresponding S-stereoisomers (4), limited information exists examining the pharmacological properties of the R- and S-isomers of albuterol.
Previous assessments of ff-agonist activity and potency have used relaxation of airway smooth muscle precontracted with muscarinic ago- nists. However, because muscarinic receptor activation of Gi~ can attenu- ate OAR-mediated signal transduction, such measurements are conditional
*Author to whom all correspondence and reprint requests should be addressed.
Clinical Reviews in Allergy and Immunology 3 7 Volume 14, 1996

38 Penn et al.
on the degree of precontraction (2) and may not accurately reflect ago- nist-{]AR interaction. Alternat ively, analysis of second-messenger (cAMP) generation would provide a more direct measure of agonist potency/act iv i ty , unfettered by complicating influences inherent in assays of tissue function. In this study, we examined the pharmacologi- cal properties of the R-, RS-, and S-albuterol compounds with respect to binding characteristics to recombinant human [31AR and 62AR and cAMP generation via activation of ~2AR in human prostate PC3 cells. Identification of the properties of the two isomers may improve our unders tanding of the effectiveness of albuterol inhalant treatment and establish a basis for the use of isomers as effective therapeutic agents.
Experimental Procedures Materials
Wild-type Spodoptera frugiperda (Sf9) and human prostate PC3 cells were obtained from American Type Culture Collection (Rockville, MD). Tissue-culture reagents were purchased from Gibco (Grand Island, NY) and Sigma (St. Louis, MO). The {3-agonists (-)isoproterenol and (-)norepinephrine were obtained from Sigma, and R-albuterol HC1 (98.43% purity, 99.23%ee), S-albuterol HC1 (97.61% purity, 100%ee), and RS-albuterol base were synthesized by Ultrafine Chemicals and pro- vided by Sepracor. [~25I]Iodopindolol and cAMP radioimmunoassay kits were purchased from NEN Dupont (Boston, MA). All other reagents were of the highest grade commercially available.
Cell Culture Sf9 cells were cultured in a spinner flask (80 rpm) at 27~ using
TNM-FH medium containing 10% fetal bovine serum and antibiotics (0.25 ~tg/mL fungizone, 50 ~tg/mL streptomycin, 50 ~tg/mL penicillin). Human prostate PC3 cells were cultured in MEM-Earle's medium con- taining 10% fetal bovine serum and 100 ~tg/mL gentamicin.
Preparation of Human ~,- and fi2-Adrenergic Receptors Recombinant baculoviruses containing the cDNA encoding the
human f31AR (5) or ~2AR (6) were prepared and used to infect separate flasks of Sf9 cells as previously described for the 132AR (7). Forty-eight hours postinfection, the cells were harvested by centrifugation (1000g, 20 min), washed once with ice-cold phosphate-buffered saline (PBS), and then lysed in ice-cold buffer A (50 mM Tris-HC1, pH 7.5, 5 mM EDTA, 20 ~tg/mL aprotinin, 20 ~tg/mL benzamidine) using a Brinkmann polytron (2 x 20 s). The homogenates were centrifuged at 40,000g for 20 min, the pellets were washed once with buffer A, and then resus- pended in buffer A at a concentration of -20 pmo113AR/mL.
Clinical Reviews in Allergy and Immunology Volume 14, 1996

Albuterol Interaction with flAR 39
Competitive Binding Studies The nonsubtype-se lec t ive ~-antagonis t [125I]iodopindolol was
initially determined by Scatchard analysis (8) to bind to human ~IAR and ~2AR with K~s of 27 and 20 pM, respectively. The ability of the vari- ous compounds tested in this s tudy ([-]isoproterenol, [-]norepineph- rine, ICI 118,551, and R-, RS-, and S-albuterol) to compete w i th [~25I]iodopindolol binding to human ~IAR and ~2AR was then assessed. The incubations contained Sf9 cell membranes (22 fmol of ~1AR or ~2AR), 45-85 pM [~25I]iodopindolol, and various concentrations of com- pet ing l igand (10 -H to 10-3M) in 25 mM Tris-HC1, pH 7.5, 2 mM MgC12, 0.5 mM ascorbic acid. The samples were incubated for 60 min at 22~ before filtration on GF/C glass fiber filters using a 48-place Brandel cell harvester. The filters were then washed four times with -5 mL of ice-cold 25 mM Tris-HC1, pH 7.5, 2 mM MgC12 buffer, and counted in a 7-counter.
Characterization of Human Prostate PC3 Cell flAR Initial studies were performed to characterize ~-adrenergic recep-
tors on h u m a n prosta te PC3 cells u t i l iz ing Scatchard ana lys i s of [12~I]iodopindolol binding. PC3 cells were shown to express -21,000 ~ARs/cell wi th a K, for iodopindolol b inding of 20 pM. The ~AR sub- type in PC3 cells was assessed using competitive binding assays with the ~2-specific antagonist ICI 118,551. This ligand binds to the ~2-subtype wi th -100-fold selectivity. ICI 118,551 was able to displace >95% of the iodopindolol binding with high affinity (IC50 - 4 nM) demonstrat- ing that >95% of the ~ARs on PC3 cells are of the ~2-subtype.
Measurement of cAMP Production in Human Prostate PC3 Cells
H u m a n prostate PC3 cells were harvested by trypsinization and washed twice wi th PBS. A time-course experiment of cAMP produc- tion was then performed by incubating the cells (-2 x 106 cel ls /mL) in PBS wi th either 10 ~tM (-)isoproterenol, 100 ~tM R- or RS-albuterol, or 1 mM S-albuterol at 37~ in the presence of 0.5 mM ascorbic acid and 0.2 mM isobutylmethylxanthine, a cAMP phosphodiesterase inhibitor. One hundred-microl i ter aliquots were removed at 2, 5,10, 20, 30, and 60 min and lysed by addition of 50 ~tL of 1.2M perchloric acid. Follow- ing neutralization with 100 ~tL of 7.5% sodium bicarbonate, the samples were centrifuged to remove precipitated protein and 5-10 ~tL aliquots were then analyzed for cAMP content by radio immunoassay as previ- ously described (9). All of the agents stimulated cAMP production wi th maximal levels observed after a 10-20 min incubation.
Dose-response experiments on cAMP production in the PC3 cells were also performed. In these studies, the cells were incubated wi th
Clinical Reviews in Allergy and Immunology Volume 14, 1996

40 Penn et al.
Table 1 Comparison of Ligand Affinities for the Human D~AR and ~2AR
C o m p o u n d n
Fold K, for I32AR, Ka for [3~AR, selectivity,
nM nM ~2AR:J31AR
( - ) i sopro te reno l 1 41.7 20.1 0.48 ( - )no rep ineph r ine 1 2260 102 0.045 ICI 118,551 1 0.68 62 91 R-a lbuterol 2 236 1540 6.5 RS-albuterol 2 668 2980 4.5 S-albuterol 3 33,600 111,000 3.3
various concentrations of (-)isoproterenol or R-, RS-, or S-albuterol for 10 min at 37~ as described. All of the agents produced significant increases in cAMP levels following the 10-min incubation at 37~ The samples were quenched by addition of perchloric acid as described, and cAMP levels were subsequently measured by radioimmunoassay. Maximal activation and EC~ 0 values for each of the ligands examined were calculated utilizing Sigmaplot 4.16.
Results and Discussion Studies were performed to determine the binding affinities of R-, S-,
and RS-albuterol for human [3~- and 132-adrenergic receptors. The indi- vidual receptor subtypes were prepared from Sf9 cells infected with a recombinant baculovirus containing either the human [3~AR or ~2AR cDNA. Because Sf9 cells lack endogenous [~-adrenergic receptors, this procedure enables the overexpression and preparat ion of a single, defined receptor subtype. Initial studies assessed the binding of the nonsubtype-selective [3-agonist (-)isoproterenol, the ]31AR selective ago- nist (-)norepinephrine, and the ~2AR selective antagonist ICI 118,551 to each of the 13AR subtypes. This was accomplished by determining the ability of the various compounds to compete for p25[liodopindolol bind- ing to the [3~AR or ]32AR. Each of the compounds competed for a single af f in i ty site wi th IC50 values of -140 nM, 7.6 btM, and 2.3 nM for the ~2AR, and 71,360, and 220 IG4 for the [~IAR for ( - ) isoproterenot , ( - )norepinephr ine , and ICI 118,551, respectively. The K d values were then calculated from the measured ICs0 values and measured K~ for [~25I]iodopindolol (20 pM for the [32AR and 27 pM for the ]3~AR) using the equation of Cheng and Prusoff (10). This calculation yielded KL~ val- ues of 41.7 and 20.1 nM for ( - ) i soproterenol , 2260 and 102 nM for (-)norepinephrine, and 0.68 and 62 nM for ICI 118,551 for the ~2AR and [31AR, respectively (Table 1). These studies verify previous reports (11,12) identifying the nonselectivity of (-)isoproterenol (-twofold selective
Clinical Reviews in Allergy and hnmunology Volume 14, 1996
Albuterol Interaction with flAR 41
8 . .J
g
20
120 i l I I I I
1(]0 o Q
80 �9
60
0
I -8 -7 ~5 -5 -4 -3 -2 -1
A L B U T E R O L C O N C E N T R A T I O N (log M)
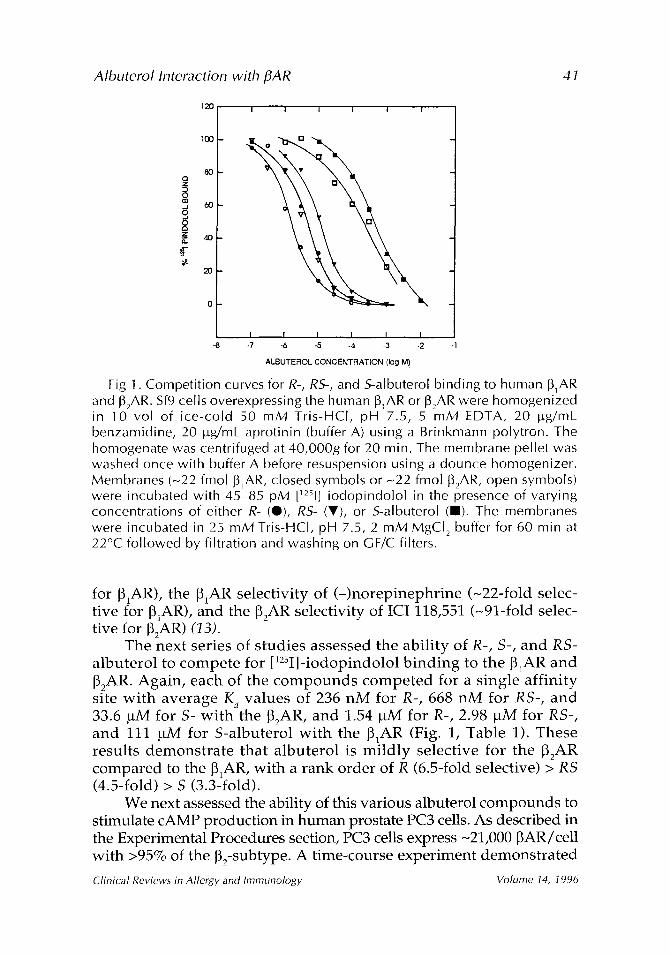
Fig 1. Competition curves for R-, RS-, and 5-albuterol binding to human J31AR and ]32AR. Sf9 cells overexpressing the human [31AR o r [32AR were homogenized in 10 vol of ice-cold 50 mM Tris-HCI, pH 7.5, 5 mM EDTA, 20 ~tg/mL benzamidine, 20 lug/mL aprotinin (buffer A) using a Brinkmann polytron. The homogenate was centrifuged at 40,000g for 20 min. The membrane pellet was washed once with buffer A before resuspension using a dounce homogenizer. Membranes (N22 fmol J31AR, closed symbols or N22 fmol IB2AR, open symbols) were incubated with 45-85 pM [~2~1] iodopindolol in the presence of varying concentrations of either R- (e), RS- (V), or S-albuterol (11). The membranes were incubated in 25 mM Tris-HCI, pH 7.5, 2 mM MgCI 2 buffer for 60 min at 22~ followed by filtration and washing on GF/C filters.
for [31AR), the J3~AR selectivity of (-)norepinephrine (-22-fold selec- tive for [~IAR), and the ~2AR selectivity of ICI 118,551 (-91-fold selec- tive for [~2AR) (13).
The next series of studies assessed the ability of R-, S-, and RS- albuterol to compete for [125I]-iodopindolol binding to the ~IAR and [32AR. Again, each of the compounds competed for a single affinity site with average K d values of 236 nM for R-, 668 nM for RS-, and 33.6 btM for S- with the ~2AR, and 1.54 btM for R-, 2.98 btM for RS-, and 111 btM for S-albuterol with the 131AR (Fig. 1, Table 1). These results demonstrate that albuterol is mildly selective for the ~2AR compared to the [3~AR, with a rank order of R (6.5-fold selective) > RS (4.5-fold) > S (3.3-fold).
We next assessed the ability of this various albuterol compounds to stimulate cAMP production in human prostate PC3 cells. As described in the Experimental Procedures section, PC3 cells express ~21,000 [~AR/cell with >95% of the J32-subtype. A time-course experiment demonstrated
Clinical Reviews in Al lergy and Immunology Volume 14, 1996
42 Penn et al.
=o -5 E o .
v
1400
1200
1000
800
600
400
200
0
I i
o / ~ ~ o ISO - / /
o
~ v �9 S
1
I I t I l I I 0 10 20 30 40 50 60 70
TIME (MIN)
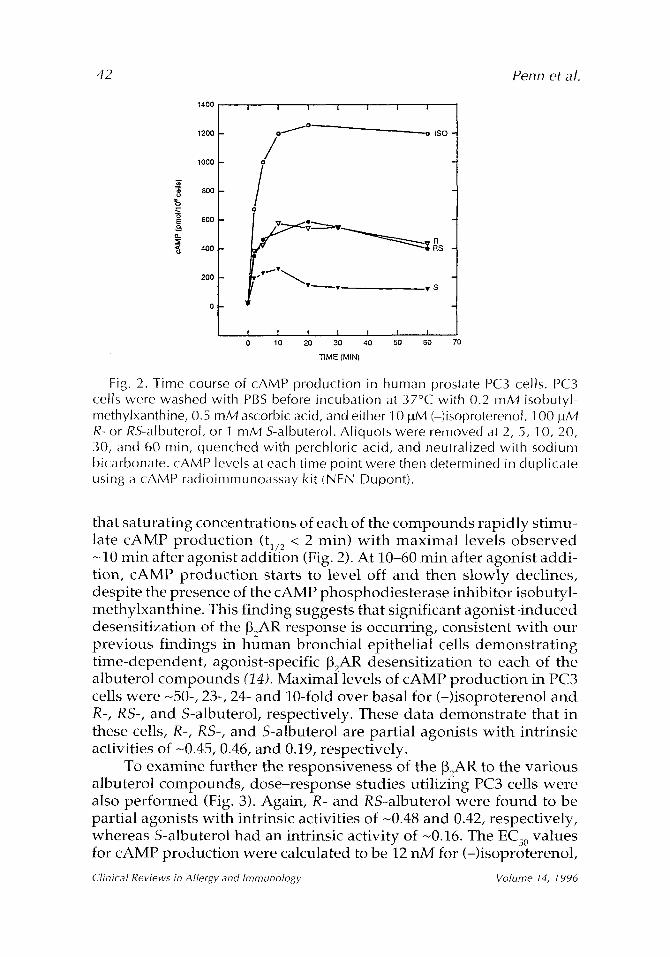
Fig. 2. Time course of cAMP production in human prostate PC3 cells. PC3 cells were washed with PBS before incubation at 37~ with 0.2 mM isobutyl- methylxanthine, 0.5 mM ascorbic acid, and either 10 ~tM (-)isoproterenol, 100 ~tM R- or RS-albuterol, or 1 mM S-albuterol. Aliquots were removed at 2, 5, 10, 20, 30, and 60 rain, quenched with perchloric acid, and neutralized with sodium bicarbonate, cAMP levels at each time point were then determined in duplicate using a cAMP radioimmunoassay kit (NEN Dupont).
that saturating concentrations of each of the compounds rapidly stimu- late cAMP product ion (tl/2 < 2 min) wi th maximal levels observed -10 min after agonist addition (Fig. 2). At 10-60 min after agonist addi- tion, cAMP product ion starts to level off and then slowly declines, despite the presence of the cAMP phosphodiesterase inhibitor isobutyl- methylxanthine. This finding suggests that significant agonist-induced desensitization of the ~2AR response is occurring, consistent with our previous findings in human bronchial epithelial cells demonstrat ing time-dependent, agonist-specific [32AR desensitization to each of the albuterol compounds (14). Maximal levels of cAMP product ion in PC3 cells were -50-, 23-, 24- and 10-fold over basal for (-)isoproterenol and R-, RS-, and S-albuterol, respectively. These data demonstrate that in these cells, R-, RS-, and S-albuterol are partial agonists with intrinsic activities of -0.45, 0.46, and 0.19, respectively.
To examine further the responsiveness of the ~32AR to the various albuterol compounds, dose-response studies utilizing PC3 cells were also performed (Fig. 3). Again, R- and RS-albuterol were found to be partial agonists with intrinsic activities of -0.48 and 0.42, respectively, whereas S-albuterol had an intrinsic activity of -0.16. The EC50 values for cAMP production were calculated to be 12 nM for (-)isoproterenol,
Clinical Reviews in Allergy and Immunology Volume 14, 1996
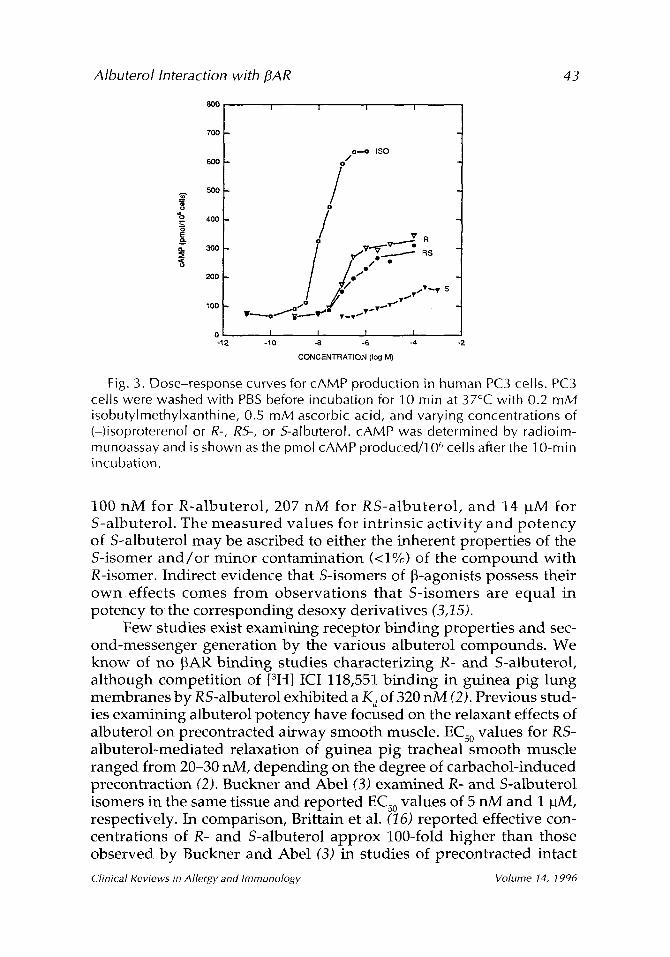
Albuterol Interaction with flAR 43
8
E
800
70O
600
500
400
300
200
'~00
0 -12
I ! I I
o---o ISO /
/ /
/ / . ."
I I 1 I -10 -8 -6 -4
CONCENTRATION (log M)
-2
Fig. 3. Dose-response curves for cAMP production in human PC3 cells. PC3 cells were washed with PBS before incubation for 10 min at 37~ with 0.2 mM isobutylmethylxanthine, 0.5 mM ascorbic acid, and varying concentrations of (-)isoproterenol or R-, RS-, or S-albuterol. cAMP was determined by radioim- munoassay and is shown as the pmol cAMP produced/106 cells after the 10-min incubation.
100 nM for R-albuterol, 207 nM for RS-albuterol , and 14 ~tM for S-albuterol. The measured values for intrinsic activity and potency of S-albuterol may be ascribed to either the inherent properties of the S-isomer and /o r minor contamination (<1%) of the compound with R-isomer. Indirect evidence that S-isomers of ~-agonists possess their own effects comes from observations that S-isomers are equal in potency to the corresponding desoxy derivatives (3,15).
Few studies exist examining receptor binding properties and sec- ond-messenger generation by the various albuterol compounds. We know of no ~AR binding studies characterizing R- and S-albuterol, although competition of [3H] ICI 118,551 binding in guinea pig lung membranes by RS-albuterol exhibited a K~ of 320 nM (2). Previous stud- ies examining albuterol potency have focused on the relaxant effects of albuterol on precontracted airway smooth muscle. ECs0 values for RS- albuterol-mediated relaxation of guinea pig tracheal smooth muscle ranged from 20-30 nM, depending on the degree of carbachoMnduced precontraction (2). Buckner and Abel (3) examined R- and S-albuterol isomers in the same tissue and reported ECs0 values of 5 nM and 1 ~VI, respectively. In comparison, Brittain et al. (16) reported effective con- centrations of R- and S-albuterol approx 100-fold higher than those observed by Buckner and Abel (3) in studies of precontracted intact
Clinical Reviews in Allergy and Immunology Volume 14, 1996

44 Penn et al.
guinea pig trachea. These disparate reports are likely explained by dif- ferences in tissue preparation and other experimental conditions. Lemoine et al. (2) examined RS-albuterol-mediated adenylyl cyclase activity in guinea pig lung membranes and determined an intrinsic activity of 61% (relative to [-]isoproterenol activity), but did not assess potency of the compound.
What can be said regarding the relative contributions to [3AR- mediated effects in the airways of the R- and S-isomers comprising a racemic mixture of albuterol? The answer would depend in part on the effective local concentration of the ligand at the principal sites of action and the activity of the compound at such a concentration. Effective local concentrations of inhaled ]3-agonists in the airways are difficult to establish empirically, and likely vary for different cell types (airway epithelium, airway smooth muscle, inflammatory cells) and along the tracheobronchial tree. Based on our findings from dose-response experiments, we would predict that concentrations of S-albuterol would have to exceed 1 ~tM before activation of the ]32AR by this compound might occur. Moreover, the respective binding affinities of the R- and S-isomers suggest that S-albuterol binding to ~2AR would be blocked effectively by equimolar amounts of the R-isomer at concentrations <10 ~tM. Although the conditions under which activity of the albuterol compounds were assessed in the present study may differ from those in situ, it appears that at concentrations of RS-albuterol likely delivered to the airway, the R-isomer is responsible for ~2AR-mediated responses of the drug.
In summary, this study examined the pharmacological properties of the R-, RS-, and S-albuterol compounds with respect to binding char- acteristics to the human ]31AR and ~2AR and cAMP generation via acti- vation of the ]32AR in human prostate PC3 cells. Competition binding studies identified a rank order of affinity for each of the ]3AR subtypes, and fold selectivity of ~2AR over {3~AR, of R > RS > S. Studies of time- dependent and concentration-dependent generation of cAMP identi- fied the albuterol compounds as partial agonists, with the R-, RS-, and S-compounds exhibiting 46, 44, and 18%, respectively, of the activity of (-)isoproterenol. Each of the compounds elicited rapid activation of ]32AR-mediated cAMP generation followed by a rapid attenuation of the response. These findings are consistent with the notion that the [3AR- mediated physiological effects of inhaled racemic mixtures of albuterol are likely affected by the R-isomer, with little if any contribution from the S-isomer.
Acknowledgment The authors acknowledge Roger Bodine for technical assistance.
Clinical Reviews in Allergy and Immunology Volume 14, 1996

Albuterol Interaction with ~AR 45
References 1. Rasmussen, H., Kelley, G., and Douglas, J. S. (1990), Am. J. Physics. 258, L279-L288. 2. Lemoine, H., Overlack, C., Kohl, A., Worth, H., and Reinhardt, D. (1992), Lung
170, 163-180. 3. Buckner, C. K. and Abel, P. (1974), J. Pharmacol. Exp. Ther. 189, 616-625. 4. Williams, L. T. and Lefkowitz, R. J. (1978), Receptor Binding Studies in Adrenergic
Receptors. Raven, New York. 5. Frielle, T. S., Collins, S., Daniel, W., Caron, M. G., Lefkowitz, R. J., and Kobilka,
B. K. (1987), Proc. Natl. Acad. Sci. USA 84, 7920-7924. 6. Kobilka, B. K., Dixon, R. A. F., Frielle, T., Dohlman, H. G., Bolanowski, M. A.,
Sigal, I. S., et al. (1987), Proc. Natl. Acad. Sci. USA 84, 46-50. 7. Kim, C. M., Dion, S. B., Onorato, J. J., and Benovic, J. L. (1993), Receptor 3, 39-55. 8. Scatchard, G. (1949), Ann. NY Acad. Sci. 51, 660-672. 9. Penn, R. B., Kelsen, S. G., and Benovic, J. L. (1992), Am. J. Respir. Cell Mol. Biol. 7,
66-72. 10. Cheng, Y. and Prusoff, W. H. (1973), Biochem. Pharmacol. 22, 3099-3108. 11. Stiles, G. L., Taylor, S., and Lefkowitz, R. J. (1983), Life Sci. 33, 467-473. 12. Suzuki, T., Nguyen, C. T., Nantel, F., Bonin, H., Valiquette, M., Frielle, T., and
Bouvier, M. (1991), Mol. Pharmacol. 41, 542-548. 13. Bilski, A., Dorries, S., Fitzgerald, J. D., Jessup, R., Tucker, H., and Wale, J. (1980),
Br. J. Pharmacol. 69, 292,293. 14. Benovic, J. L., Kong, G., Penn, R. B., and Aberg, G. (1994), Eur. Respir. J. 7, 99s. 15. Patil, P. N., LaPidus, J. B., and Tye, A. (1970), J. Pharm. Sci. 59, 1205-1234. 16. Brittain, R. T., Farmer, J. B., and Marshall, R. (1973), Br. J. Pharmacol. 48, 144-147.
Clinical Reviews in Allergy and Immunology Volume 14, 1996