
Impairment of immunity to Candida and albicans ... · 7/8/2015 · 1, , (+ + +. Science. Science +) +

,〆,
1. Introduction
Co-operation of Innate and Acquired Immunity for Controlling Tumor Cells
Hidemiγakahashl Dep目的nentof Microbiology and Immunology,
、NipponMedical School, Tokyo, Japan
τ'he body i8 compos吋 ofvほioustypes of ceおめatunite泌 harmonyto live IJロ出enaぬral臼wironment.Viral infections, toxic products,品ndenvironmenぬ1str即日es江田yaffect the cells toなutiatetransfor立国tionfor the developme設tof tumors,絞1uncontrol1able and unfavorable state of cells. Such a tumor詰tateof ce恥 i8dosely watched by偽記 intemalimmune-surveillance system設latgenerally controls or el始以at船出ernto rnaint位nth捻hぽ rnonyof the body. The internal irr盟国民付urveillancesystem is comp08ed of two distinct pa出 ; 昭 如 ¥ ate注m 聞記 sy翻 rnpredom弘 樹Iy10倒 edont抑制ぱacea玄関Sof the body, such as the s恰 lormucosal compartments, and an acquiredj adaptive総ununesystem found mainly in systemic compar加 ents,加dudingcircul出 ngblood, lymph nodes, spleen, and various org組 s(Medzhitov and Janeway, 1ヲ97a)(Fig. 1).話会 ce1l8of the加工lateimmune system伽ldand fight forei伊 bodi部 1r注目dingfro汲註leout器ide,such as viruses and bacteria, through their pat除m-reco伊 itionrecept附(PRRs)(鳩山hitov,2(07), such a器 Toll-liker酬 がorsσぽ s)(A記raet al., 2001) and C勾pelec出 receptors(Cほ s)仕latreco伊 izepathogen-associated rnole叩 larpatterns (PAMPs)倒 的蜘vand Janeway, 1997b), al泣10ロghthey do not have any specific memories of foreign bodi総Incontrast,政lea句姐redinunune system will r田 pondto forei伊 elementsonly when competent cells ofせlea叫凶redsystem recognize the総 specificallythrough仕1eirreceptors estab泌.hedvia gene叩 arrangements(palm相 dMedzhitov, 2009). A絞¥0れ.gthese ac伊悦dreceptor為 aT叩 11receptor (TCR) can specific正illyreco伊izeforeign叙 1tigensas a specific蹴 ucturalcomponent co総.posedof proteIrl-d在rivedanlIrlo acids presented partic叫arlyby担 lf酬 restricted訂¥tigen-presentingmolecul田" ter玄 関dIr¥ajorhかね∞mpatibilitycomplex小杭Qσ桃山ashi,2003)Although the precise mech仮説smsr記mainto be eluddated, s読むhspecificm慌をoriεsmediatedthr∞ghl・CRsare IrlS伽ctedby inn縦 cells,同胞larlyden制緩.ccells (DCs), which are key cells to present antig絞史的formationめou抽出eirち必iCtogether wi仕1co-stなれulatorymolecul昭 thatひ吋11promote memory formation via gene-rearrange釘陪nts訂1a亡qruredT cells. 2もerefore,inna総 DCshaveめ.eability to capture tumor-derived antigens, proむessedthe絞into an託ge凶cfragments, and prese対 theantigenic fragments in association with their 1¥心iCand c付加ulationmol削 les(AZUIr¥a et al., 1争犯;Chen et al., 1992), such as B7-1 (CD80), 昨 2(C部品), or印 '40,to田 tablishTCR-mediated an相 時specific玄関moriesin ac明住宅d勧ununi勿 (Fig.2).
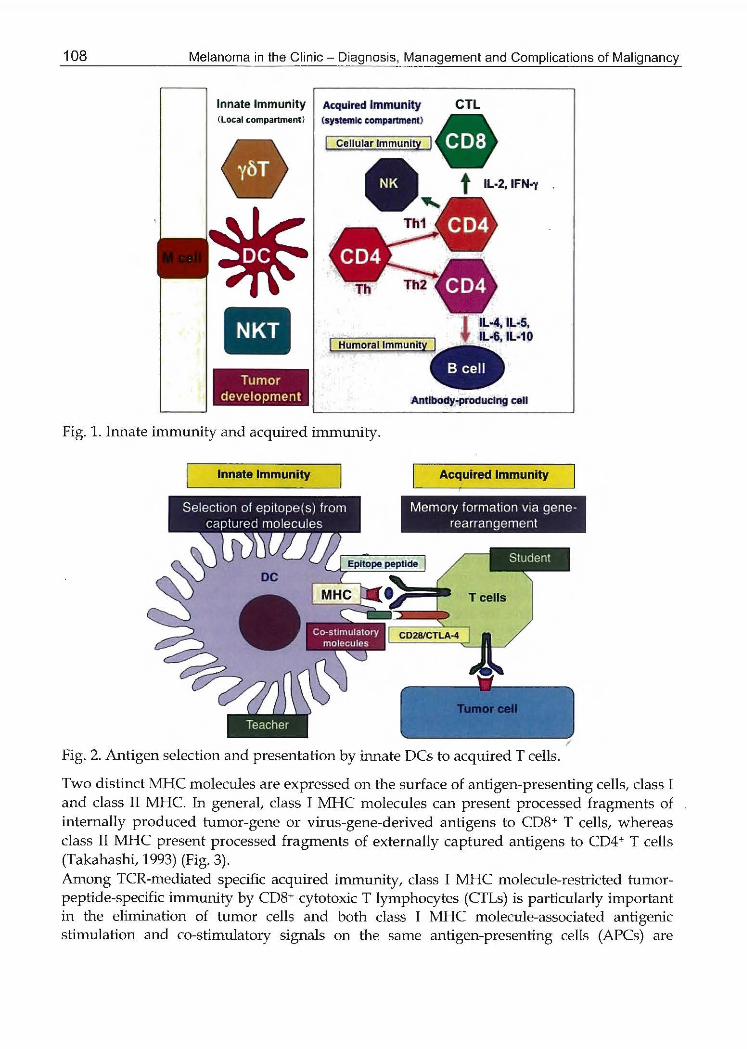
108 Melanoma in the Clinic -Diagnosis, Management and Complications of Malignancy
Innate Immunity (Local compartment)
CTL
。
EE -_.'・E・11'・'J_・~当国姐:iiN且量 Antlbody.produclngωu
Fig. 1. Innate immunity and acquired i江田mnity.
Inna旬 Immunity Acqulred Immunity ••••••••••• -e
-
門
川-e
-nHd
-
a
J
H
一
ve
-nH引uu
一-mu
即日
-
d
QU
一
mm:
聞、,,,E'
-h
a
一
yモ
ー一
O一
m-e
一
M
Fig. 2. Antigen selection and presentation by innate DCs to acquired T cells.
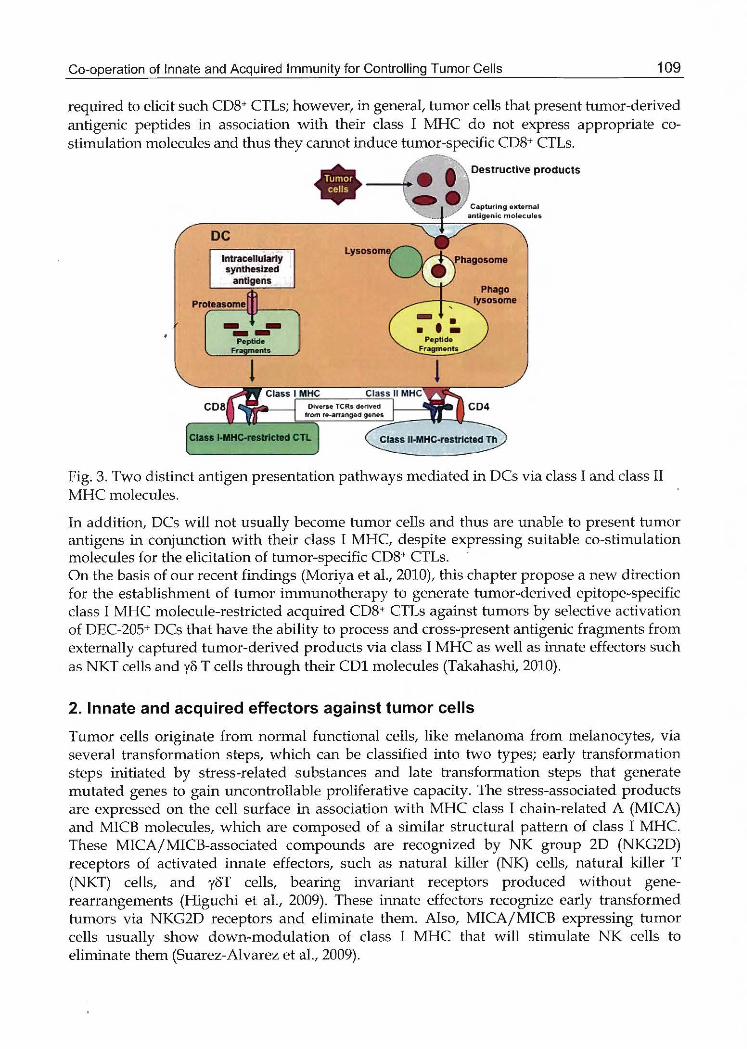
Two distinct MHC molecules are expressed on the surface of antigen-presenting cells, class 1
and class 11 MHC. In generat class 1 MHC molecules can present processed fragments of
internally produced tumor-gene or virus-gene-derived antigens to CD8+ T cells, whereas class 11 MHC present processed fragments of externally captured antigens to CD4+ T cells
(Takahashi, 1993) (Fig. 3)
Among TCR-mediated specific acquired immunity, class 1 I'v征-ICmolecule-restricted tumor-
peptide・specificimmunity by CD8+ cytotoxic T lymphocytes (CTLs) is p紅 ticularlyimportant
in the elimination of tumor cells and both cIass 1 I'v柾-IC molecule-associated田市genicstimulation and co-stim叫atorysignals on the same antigen-presenting cel1s (APCs) are
Co・operationof Innate and Acquired Immunity for Controlling Tumor Cells 109
DC
Fig. 3. Two distinct紅 ltigenpresentation pathways rnediated in DCs via class 1 and class II
MHC rnolecules.
In addition, DCs will not usually become tumor cells and thus are unable to present turnor antigens in conjunction with their class 1 MHC, despite expressing suitable co-stirnulation rnolecules for the elicitation of turnor-specific CD8+ CTLs. On the basis of ou旧rr閃ec印en川1吋tf白indi凶ngs(Mo凹r匂i
C for the e白st旬abli応shrnen叫1註toぱfturnor immunotherapy t旬og伊en即1官erat栓et加umo町r-deriv刊edep戸itωop戸e“S叩pecif凶f白仕ic class 1 MHC rnolecule-restricted acquired CD8+ CTLs against tumors by selective activation of DEC-205+ DCs that have the ability to process and cross-present antige凶cfragrnents frorn external砂capturedtumor-derived products via class 1 MHC as well as innate effectors such as NKT cells and y8 T cells through their CD1 rnolecules (Takahashi, 2010).
2. Innate and acquired effectors against tumor cells
Tumor cells originate frorn normal functional cells, like rnelanorna frorn melanocytes, via several transforrnation steps, which can be classified into two types; early transforrnation steps initiated by stress-related substances and late transformation steps that generate mutated genes to gain uncontrollable proliferative capacity. The stress-associated products are expressed on the cell surface in association with MHC class 1 chain-related A (MICA) and MICB rnolecules, which are cornposed of a similar structural pattern of class 1 MHC. These MICAjMICB-associated cornpounds are recognized by NK group 2D (NKG2D) receptors of activated innate effectors, such as natural killer (NK) cells, natural killer T (NKT) cells, and yδT cells, bearing invariant receptors produced without gene-rearrangernents (Higuchi et a1., 2009). These innate effectors recognize early transforrned tumors via NKG2D receptors and elirninate thern. Also, MICAjMICB expressing turnor cells usually show down-modulation of class 1 MHC that will stimulate NK cells to eliminate them (Suarez-Alvarez et a1., 2009).

110 Melanoma in the Clinic -Diagnosis, Management and Complications of Malignancy
In contrast, mature tumor cells with mutated genes and expressing tumor-gene-encoded antigenic peptides on their surface in association with class 1 MHC molecules c担") be specifically recognized and eliminated by acquired CD伊 CTLs.ThllS, stress-associated early transformed tumor cells appear to be regulated mainly by innate effectors, while mature tumor cells with specific s廿ucturalmutations with high proliferative capacity can be specifically controlled by acquired CD8+ CTLs in a c1ass 1 MHC molecule-restricted manner. Taken together, internally transformed tumor cells g出neduncontrolIable proliferating capacity can be recognized and regulated by both innate and acquired effectors (Fig. 4).
Progress of transformatlon
Immature 知町柚rcells
Fig. 4. Immune control of immature and mature tumor cells.
3. Antigen-presenting molecules on DCs
Induction of these innate and acquired effectors seems to be regulated by DCs via various antigen-presenting molecllles. Innate immunity is chiefly regulated via species-res仕ictedCD1 antigen-presenting molecules and acquired immunity is controlled via individually restricted MHC molecules on DCs. CD1 molecules are further divided into four classes, CD1a, CD1b, CD1c, and CDld. These CDls have been found to present lipidj glycolipid antigens to T cells bearing relatively invariant αsT -celI receptors (TCR), rnost of which are conserved among species (Barral and Brenner, 200たCohenet a1., 2009). For example, highly conserved CD1d molecules presentα-galactosyl ceramide (α-GalCer) to NKT cells of their own species (Saito et a1., 2005). Indeed, human NKT cells generall)j express unique combinations of TCRs that consist of an invariant Vα24 chain preferentially paired with a Vsll (DelIabona et aし1994),while murine α心alCer-reactiveCDld勘 restrictedNKT cells express invariant Vα14 paired with various Vs combinations (Gui et al., 2001). In contrast to species-restricted CD1 antigen-presenting molecules, both class 1 and dass II MHC molecules are highly diverse among individuals as self-restricted elements presenting internally processed peptides as antigens, which can be recognized by the same MHC molecule-bearingαsTCR-expressing T cells with antigen-specificity established by intracellular gene rearrangements. Such gene rearrangements for the establishment of T cell-mediated acquired immunity can be initiated through the combination of antigen-loaded MHC molecules plus appropriate co-stimulatory signals on DCs, although the induction of CDl-associated T celIs does not require such co-stimulatory signals. Thus, co-stimulatory

Co叩 erationof Innate and Ao号uiredImmunity for Con加lIin9Tumor Cells 111
signals app但 rto be required for the development of附{c設叫ecule-res耐 tedhi詐lyspecific ac守山ed耐m山 ty(Naka伽 kaet a1., 19担分 Generally,CD8<xs阿positiveT cells recogrせZeせを.eprocessed epiもopepeptide from internally synthesized proteなlSpresented by cl出 s1 MHC molecules, whereas CD4-positive T ce旦sr'配 ognizeepitope peptide from externally captured proteins in輔副ationwith class II減配(Dus凪 20悦 Thes加 .cturesof the two distinct groups of an均出国.presentingmolec叫es,CD1 and class 1島倒C,closeIy resemble制 chother, havi略 threeregions of a-chains (a1, a2, andα3) with non-covalently bo悶 d手2-microglobulinthat may regulate撫釦説ge恥 bindingcapacity ofめep間四出gmolecules伴侶lowsお eta1., 1空間ー However,a1though CDl-encoding genes are highly conserved and their structu総事 areshared among species with linuted polymorphism (Couedel et a1.,明書),伽 class1 MHC剛 encodinggene is highly dive四 amongindividuals. 80 far, it has be己主1reported仕18tmost innate NKT cells ~ be activated出roughCD1d and some yoT cells are activated through CD1c molβω1部 onDCs (Cohen et al., 2009)事 Mo陀 over,we have詑 cen今observedthat live bacillus Ca1me蜘 Guerin(BCG)州問総dDCs elicit innate effectoお"such拙 NKcelおi,NKT ce註s,and yST cells, 10 inhibit the growth of bladder 躍 'cinomavia IL-12 s回組問問別出合ta1., 2009). Collectively, DCs have the a私立ityto mar話pulateなmat官 会ffectorsthrough CD1s and cytokin略 asweI! as ac弓泌総deffectors via E狂!Cmolecules 10 control intemalれlmoTS.
4. DC sub諮etand “cross-presentation"
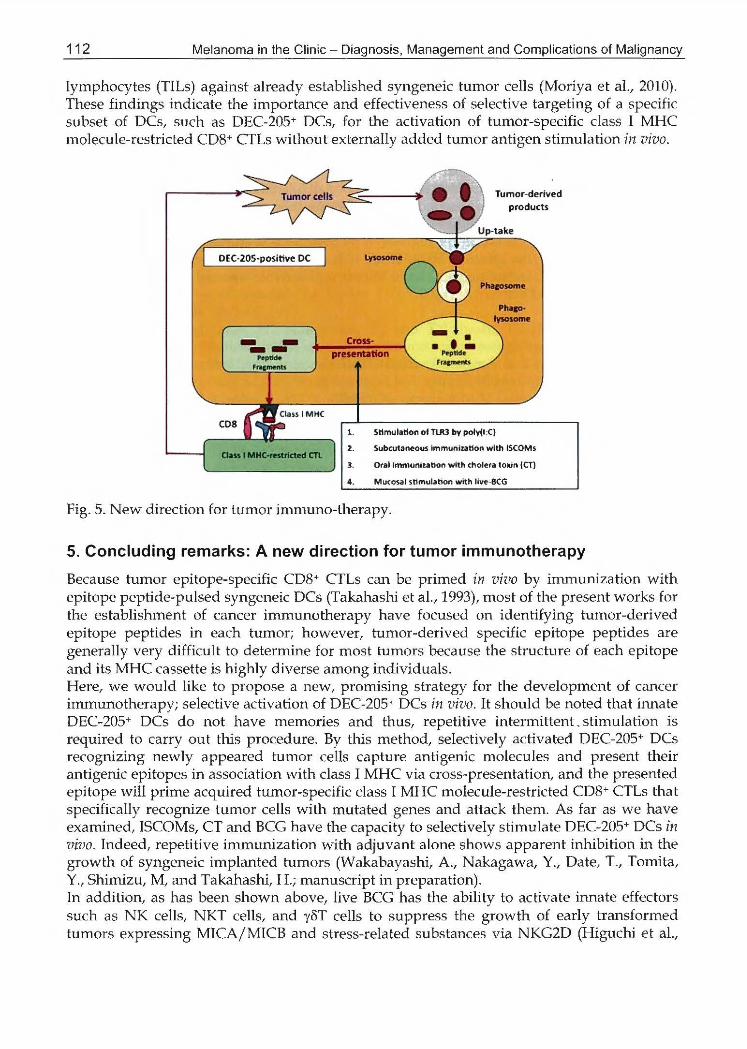
Reむently,it has been demonstrated that two non-overlapping似 bs時 ofDCs are arranged to re渉必teinternal泳ununerespons田 invivo; 33D1 (reco務uzingdenむiticcell釦hibitory校 ceptor-2(DCIR2))ゃosi伽 ea対 DEC却 5-p邸 i滋veDCs (ひなdziaket a1., 200η. It has a1so been repo対:edthat in vivo 加 getingof DECω205 wi正heither poly(I:C) (Trumpふelleret a1., 200めor知 antibodyspecific for DC倫 NKlectin gro呼 recep初予1(DNGR-l) (8剖lchoet al., 2008) induced dominant Th1加国unityor potent CTL responsξs via cross輸presenta伎の九respectiveIy. Af; indicai吋 inFig 5, it has tumed outめatthe most suitable CTL epitope peptiおswiめin航 ternaUycaptur官d初計genicproteins are器electedto present in association withcl描付加!CinDEC♂05-positiveDCs via cross-pre8entation向。riyaet al., 2010). We have sho椀 1that suchα'08Sア間四ta録。れず theshift of the antigen-pres朗 tationpaめW 句
会百mthe class II 1征!Cto class 1 1必iCprocess加groute for externally capれ.u:edar泌genicpr脚部, c四 beachieved by a bark-de市 ed叫 onin-制 ociatedaかv制~ such酪 ISCOMs(Takah描 hiet al., 1990), cholera toxin (CT) (Wakabayashi et a1., 2008),割、d話CG肝iguchi宕ta1., 2009). We have a1so demor班tratedσujirnotoet al.,お04)that TLR3-signaling of DCs, previously loaded antigenic protei抗告, by double醐 stranded RNA, polyribo註10SiniCpolyribocytidylic acid (polyび:C)), which reflects a naれぽalgenetic product from a variety of vrruses, can ge虫色ratethe cross-presentation. These results如 gg問 tthat sel,促tivestimulation of DEC回205+DCs in vivo may elicit eff,告ctiveac弓uiredCD8+ CTL 陀 sponses仕IToughTh1 dominancy by which protective i立ummityaga投獄れlffiorswill be achieved even in the absence of externally addedれlffiorantigens. We have recently established 33D1 + DC-depleted C57批 ';6mice ob悩 nedby悦 atmentwith 33D1-speciおむ monoclonalantibody (Moriya et al., 2010). As expected, 33D1+ DC-depleted 出叫 implantedwith syngeneic B16-F10 mel関 omacells into the dermis, showed apparent inhibi討onof already部 taもlishedtumor growth in vivo when subcutaneously (:双)inj伐 tedwith LPS after tumor irnpl削 tation,in which serum 1し12secretion that may be mediated by the rernaining DEC-205+ DCs was markedly enhanced up∞TLR4 si伊畠ling(Moriya et a1., 2010).じnexpectedIy,LP8-stimulated 33D1 + DC-deleted tumor糊 beariれgmice apparently produced H-2Kb-restricted epitope-spe立法c CD沙 CI'Ls among ぬmor infiltrating
112 Melanoma in the Clinic -Diagnosis, Management and Complications of Malignancy
lymphocytes (TILs) against already established syngeneic tumor cells (Moriya et aし2010)These findings indicate the importance and effectiveness of selective targeting of a specific subset of DCs, such as DEC・205+DCs, for the activation of tumor-speci白cclass 1 MHC molecule-restricted CD8+ CTLs without externally added tumor antigen stimulation in vivo.
OEω5ゃos伽 oc1 Lvsos-
Sl1mulation 01 Tほ]by poly{I:C)
Subω'.n~凶 Immunlu凶onwlth ISCOMs
Orallmmunl...l1on wlth出剖e日 toxln(CT)
Muc田 .1州 mulationwith IIv~ .8CG
Fig. 5. New direction for加 morimmuno-therapy.
5. Concluding remarks: A new direction for tumor immunotherapy
Because tumor epitope-specific CD8+ CTLs can be primed in vivo by immunization with epitope peptide-pulsed syngeneic DCs (Takahashi et a1., 1993), most of the present works for the establishment of cancer immunotherapy have focused on identifying tumor-derived epitope peptides in each tumor; however, tumor-derived specific epitope peptides are generally very difficult to determine for most tumors because the structure of each epitope and its MHC cassette is highIy diverse among individuals. Here, we would like to propose a new, promising s仕ategyfor the development of cancer immunotherapy; selective activation of DEC-205+ DCs in vivo. It should be noted that innate DEC-205+ DCs do not have memories and thus, repetitive intermittent. stimulation is required to carry out this procedure. By this method, selectively activated DEC-205+ DCs recognizing newly appeared tumor cells capture antigenic molecules and present their antigenic epitopes in association with class 1 MHC via cross-presentation, and the presented epitope wil1 prime acquired tumor・specificclass 1 ~任-IC molecule-restricted CD8+ CTLs that specifically recognize tumor cells with mutated genes and attack them. As far as we have examined, ISCOMs, CT and BCG have the capacity to selectively stimulate DEC-205+ DCs In vivo. lndeed, repetitive immunization wIth adjuvant alone shows apparent inhibition in the growth of syngeneic implanted加 mors(Wakabayashi, A., Nakagawa, Y., Date, T., Tomita, Y., Shimizu, M, and Takahashi, H.; manuscript in preparation). 1n addition, as has been shown above, live BCG has the ability to activate innate effectors such as NK cells, NKT cells, and y8T cel1s to suppress the growth of early transformed tumors expressing MICAjMICB and stress-related substances via NKG2D (Higuchi et a1.,

CCトoperationof Innate and Acqui開dImmunity for C臼ntrollingTumor Cells 113
200吟.l1lese results sなong砂suggestthat we c皿 manipulateapprop市 te泳閉山崎 tocon柱。1tumor cells by repeti位ves耐nulationof DEC-205+ひcs同校:la BCG-derived substance. Now we should recall the漁船:leof Chlsato M袋れlyama(1901-1992), a professor of 佼leDepartment of Dermatology as well as th巴pastPresident of Nippon Medica1 School in Japan, who had notic吋 thatthere were a very low number of cancer pa長.entssuffered from tuberculosis and established sp日cialsubst初 cefrom Mycobacterium tu加問losisas昌むancervaαine, n四 ed社le"Maruyama Vaccine (SSM; sp母d必subst削 .ceof Maruyama)" in 1944. τhe SSM has been widely used, part会cularlyfor various cancer patients in Japan; however, めeach除 lmecl祖対smofせleSSM has been立nknownunt延nOw.l1lus, although a number of cas時 withexcellent eff舵 tsoncanc邑r胞がessionby SSM have been reported, many dlぽ torsar告 $ 総 suspiciousof枝、eeff.日ctof SSM. Here, it is proposed that new adiuvant therapy should be considered, includ註19BCG or SSM白紙 wi1l、activatein temalおnateimmunity, 問 ticu]略。奇心205+DCs.
6.決eferences
Akira, 5., Takeda, K., and Kalsho, T. (2001). Toll動地er自ceptors:∞tical proteins lir泳inginnate and亙叫uired出 munity.Nat lmmunol, 2, 675-680.
Azuma, M., lto, D., Yagit九H., Okumura, K., Phi1lips, J.1王, Lanier, L.L., and Somoza, C (1的'3).B70 antigen is a second lig,相dforCTLA-4組dCD28. Nat:ure, 366, 76将.
Barral, D.C., and Brenner, M.B. (2007). CD1肌,ti炉 1presenta伽:how it wほお.Nat Rev Immunol, 7, 929-941.
Chen, L., Ashe, S., Brady, W.A.,偽証strom,1., Hellstro叫K.E.,L吋,be伐er,J.A., McGow;叫 れ 納 せ
L泊sley,P.S. (1労2)ーCos愉別,ulationof鉛世tumorinmlUniりrbyせleB7count閉校'epぬrfor佐官Tlyml魚崎Itemolec毛desCD28 and CTLA-4.αU, 71,潟県1102.
Cohen, N.R., Garg, 5.,四dBrenner, M.B. (2009). Antigen Pr,母総ntationby CD1 Lipids, T Cells,阻dNKT Cells in Microbial Immunity. Ad,むか問問。1,102, 1-94.
Couedel,ιPeyraもM.A., Brossay, L., Koezul五a,Y., Porcelli, S.A., Davodeau, F.,釦dBonnev泣le,M. (199め.Div告rseCD1d-res紋j胸 dreac討vitypattems of知 m釦 Tcells bearing "inv訂版nt"AV24BV11 TOえEurJ Immunol, 28, 4391-4397. 。ellabona,P., Padovan,蕊, Casora化 Gザ Brockhaus,M., and L釘邸時cchla,A.。ヲヲ4).An inv紅 iantV alpha 24-J alpha Q/V bet亘 11T cell r,記む,eptoris eχpress吋 inall individua1s by donally eχpanded CD4-8戸Tcells. J Exp Med, 180, 1171-1176.
Dudziakグ D., Iく釘nphorst,A.O., Heidka笠1p,G.F., Buchholz, V.R., Trump丘leller,c., Y鉱 nazaki,5., Cheong, c., Liミl, K., Lee, HW., Park, c.G., et a1. (200η. Differential antigen processing by dendritic cell subsets in vivo. Science, 315, 107-111.
Dus加, M.L.(200め.l1le ce蛙ularcontext of T cell signaling. Imm閥均"30,482-492. 何回oto,c., Nakagawa, Y., Ohara, K., and Takahas泌, H. (20的).Polyriboinosinic
polyrぬocy舗 ylicacid [poly(I:ql/TL防 sign泌ngallows class 1 proces伽 gof exogeno註sprotein釦 dinduction of 盟いspecificCD8十 cytotoxicT lymphocytes. 1nt lmmunol, 16, 55-63.
Gui, M., Li, J., Wen, L.J., 1如 dy,R.R., and Hayak品wa,K (2001). TC双betachain influences but doesれotsolely control autor巴activityof V alpha 14J281T cells. J 1m縦unol,167,6239舗 6246.
日ignchl,T., Shin廿zu,M., Owaki, A.,τakahashl, M., Shlnya, E., Nishl汀lura,T., and Takahashl, H. (200称Aposゑbl日mechan紘絞lof泊なavesicalBCG白erapyfor human bladder carcinoma: involvement ofなmateeffectorむ悲哀sfor the inhlbition of tumor growth. Cancer Imm詰nolImmunotheち58,1245-1255.

114 Melanoma in the ClinicωDiagnosis, Management and Complications of Malignancy
Kozlowski, 5., Takeshita, T., Boehncke, W.H, Takahashi, H., Boyd, L B齢加倍佼側rα忠紛of:必S泳尚k匂勾yあμrん川,JμA.リ, and 恥M油i匂ar菩併uli郎略, D.H. (19労矧守幻1均)巳臥χCαes鍔sb泌e!泌a2削 crogloぬb加u凶li釦npromれ¥0柑託担略g f釦日nct丘ion畠必lpept討idea邸ssociat討ionwithp滋縫r討if白iedsoluble c1ass 1乱任iCr蕊nolecul剖 .Nature, 349,74-77‘
Medzhitov, R., and Janeway, c.A., Jr. (1997a). Innate imm切註ty:impact on the adaptive immuner叫榔.e.Curr Opin lmmunol, 9,科.
Medzhitov, Rザ andJaneway, c.A., Jr. (1ヲ97b).Iぽ泌総滋rmunity:設¥evなtu田 ofanoれdona1system of recogni録的.Cell, 91, 295-298.
Medzhitov, R. (200η. Re∞伊滋onof llUcroc町ganismsand ac伽胡onof the immune responsε.1ぜature,449, 819-826.
Moriya, K., Wakabay拙 hi,A., 5hinuzu, M., T旦mura,H., Dan, K., and Taka1鴎shi,H. (2010) Induction of tumor-specific acq凶redimmむnityag.必nsta1ready establishξd tumors by selec説明 stimula悦onof 加盟teDEC-205付 dendriticce恥 αncerlmmunol Immunother, 5ヲ,1083-1095.
Nakat:引ika,K., 5ugiyama, H., Nakagawa, Y., and Taka1泊.shi,H (1ヲヲ9).Purificalioれ ofan討genicpeptide from m町'inehepatoma cells recognized by Class-I major histocOl主lpatibilitycomplex moleαlle-restricted cytotoχic T-l戸nphocytesinduced with B7-1-gene-transfected h伴 atomacells‘JHepatol, 30, 1119-1129.
Pa1m, N.W., and MedzJ:註tov,R. (2009). Pattem recogr説。nreceptors and control of adaptive 加 munity.Immunol Rev, 227,221必 3.
5aito, N., Takahashi, MリAkahata,W., Ido, E., Hidaka, c., Ibuki, K., Miura, T., Haya綜誌, Mザ
and Takahashi, H令 (200勾.Analysis of evolutio蹴 yconservation in CDld molec沿eSamo:ロgprimates. Tissue Ant炉問6ふる74-682.
5ancho, D., Mourao-Sa, D., Jof台。 O.P.,Schulz, 0., Roger吉" N.C., Pennington, D.J., Carlyle, J .R., and Reis e 50usa, C. (2:∞8). Tumor therapy in出 cevia antigen targe伽 g10 a れovel,DC-restricted C-句'pεlectin.J Clin Inves乙118,20ヲふ2110.
SuarezωA1varez, B., Lopez-Vazquez, A., Baltar, J.M., Ortega, F., and Lopez-Larrea, C. (2009). Potential role of NKG2D and its ligands in organ transpl訟.la張関:new 加 getfor immunointervention. Am J Transpl開色 9,251鵬 257.
す総ahashi,H., Takeshita, T., Morein, B., P自治ey,5., Germain, R.N., and Berzofsky, ].A. (1990). lnduction of CD8+ cytoto成主 cellsby immunization with purifi吋開制1叩.velopeproI'邑ininI配 OMs.Nature, 34長, 873-875
Tak必lllshi,H. (19ヲ'3).An託.genprocessing and presentation. Microbiol Imm問。1,37,1-9.Takahashi, H., N誌agawa,Y., Yokomuro, K.,間dBerzofsky, J.A. (1労3).InductioロofCD8十
cytω:otぬ0χ対icT lymphocytes by i 宝笠m主投1宙env刊e討,10叩p記derivedp戸ep戸討dωeや網pu叫l詰e吋ddおi誌e剖n吋d制r対i討恥cc伐恕H泌草.Int Immul101, 5,849-857.
Takahashi, H. (2003). Antigen pr記sentationin vaむcinedevelopr抗告nt.Comp Immul101 Microbiol I向井tDis,払309-328. /
Takaha話hi,H. (2010) 5pecies-specific C印刷r憾なictedIlma te 加工nunityfor the development of HIV vaccine. Vaccine, 28(Supp12), B3-7.
Tr凶 lp丘leller,c., Caskey, M., Nchinda, G., Longhi, M.P., Mizen初a,0., Hu組長;, yリSchlesinger, 5.]., Colonna, Mマa対 5teinman,R.M. (2008)刷 Themicrobial mimic poly IC induces durable and protective CD4+ T cell immunity together with a dendritic cell targeted vaccine禽 ProcNatl Acad Sci U S A, 105, 2574樽 2579.
Wakabayashi, A., Nakagawa, Y., 5hinuzu, M., Moriya, K., Nishiyarna, Y., and Takahashi, H. (2008). Suppression of初a1readyestablished tωnor growingめrou菩hactI'、ratedmucosa1に:TLSinduced by oral ad町出廿strationof tumor an説genwith cholera toxin. J Immunol, 180,必000-4010.


















