
ChemComm SI wo track. Comm. 2 AFM tip functionalization. Lysenin- and θ-functionalized AFM tips...
10
Chem. Comm. 1 Materials and methods Materials. 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC), brain and egg sphingomyelin (SM) and egg phosphatidylcholine (PC) were purchased from Avanti Polar Lipids. Cholesterol (Chol) was purchased from Sigma Aldrich. Two buffer solutions were freshly prepared and used as follows i) a fusion Buffer A containing 10 mM Tris, 150 mM, NaCl 3 mM and CaCl 2 and ii) an imaging Buffer B of 10 mM Tris, 150 mM NaCl at pH 7.4. Buffer solutions were filtered before use with a 0.2 μm pore size inorganic membrane filter. Preparation of supported lipid bilayers. DOPC, SM and Chol were dissolved in chloroform to give a final lipid concentration of 10 mM. Aliquots of DOPC and SM or Chol solutions were mixed in different DOPC:SM and DOPC:Chol molar ratios (either 70:30 or 50:50), poured into a glass vial and evaporated to dryness under a nitrogen flow. Multilamellar vesicles were obtained by hydration with the Buffer A solution to give a final lipid concentration of 1 mM and then subjecting the vial to 3 x 2.5 min cycles of tip sonication to obtain unilamellar vesicles. Circular mica surfaces were used as substrates for AFM experiments. Prior to use, mica surfaces were glued onto Teflon discs with epoxy-based mounting glue. Phospholipid supported bilayers were prepared by the deposition of the small unilamellar vesicles suspension on freshly cleaved mica followed by incubation at 59°C. The samples were then slowly cooled to room temperature and thoroughly rinsed with Buffer B solution. Toxin production, purification and validation. The SM-specific toxin fragment, non-toxic (NT) lysenin, was expressed in E. coli BL21 (DE3) as a fusion protein with a 6xHis-tag followed by the fluorescent protein mCherry (total MW ~45 kDa) at N-terminal, as purified and validated previously 1 . The chol-specific toxin fragment, i.e. the fourth domain of perfringolysine (theta D4), was cloned in pET28 containing 6xHis- and LPETGG-tags in N- and C-terminal, respectively (Fig S1a). It was generated from pET28/His-mCherry-theta 2 by removing the mCherry sequence and adding the LPETGG-tag. The resulting plasmid was transformed in E. coli BL21 (DE3) and the protein expressed in LB medium for 72h at 16°C after addition of 0.4 mM isopropyl-β-D-thiogalactoside. Bacterial extracts and protein purification were prepared as previously described 2 . Analysis of the purified protein by western blot revealed recombinant theta at the expected size (~16 kDa; Fig. S1b). The most enriched fractions were pooled, concentrated using Vivaspin turbo 15 columns (Sartorius), and the imidazole was removed by desalting on NAP-5 columns (GE Healthcare). Purified protein was finally kept in aliquots in 10 mM HEPES (pH 7.2) 10 mM NaCl and stored at -80°C until use. Protein concentration in the purified fraction was estimated by measuring A 280 and assuming a molar absorptivity of 44500 M - 1. cm -1 . From 1 l of culture, we obtained 10 g of protein. The toxin binding specificity was verified on MLVs containing increasing amounts of Chol (Fig. S1b) and prepared as previously described 1 . Electronic Supplementary Material (ESI) for ChemComm. This journal is © The Royal Society of Chemistry 2018
Transcript of ChemComm SI wo track. Comm. 2 AFM tip functionalization. Lysenin- and θ-functionalized AFM tips...

Chem.Comm. 1
Materialsandmethods
Materials.1,2-dioleoyl-sn-glycero-3-phosphocholine(DOPC),brainandeggsphingomyelin(SM)and
egg phosphatidylcholine (PC) were purchased from Avanti Polar Lipids. Cholesterol (Chol) was
purchasedfromSigmaAldrich.Twobuffersolutionswerefreshlypreparedandusedas follows i)a
fusionBufferAcontaining10mMTris,150mM,NaCl3mMandCaCl2andii)animagingBufferBof
10mMTris,150mMNaClatpH7.4.Buffersolutionswerefilteredbeforeusewitha0.2μmporesize
inorganicmembranefilter.
Preparationofsupportedlipidbilayers.DOPC,SMandCholweredissolvedinchloroformtogivea
final lipid concentration of 10 mM. Aliquots of DOPC and SM or Chol solutions were mixed in
differentDOPC:SMandDOPC:Cholmolarratios(either70:30or50:50),pouredintoaglassvialand
evaporatedtodrynessunderanitrogenflow.Multilamellarvesicleswereobtainedbyhydrationwith
theBufferAsolutiontogiveafinallipidconcentrationof1mMandthensubjectingthevialto3x2.5
mincyclesoftipsonicationtoobtainunilamellarvesicles.
CircularmicasurfaceswereusedassubstratesforAFMexperiments.Priortouse,micasurfaceswere
glued onto Teflon discs with epoxy-based mounting glue. Phospholipid supported bilayers were
prepared by the deposition of the small unilamellar vesicles suspension on freshly cleaved mica
followed by incubation at 59°C. The samples were then slowly cooled to room temperature and
thoroughlyrinsedwithBufferBsolution.
Toxin production, purification and validation. The SM-specific toxin fragment, non-toxic (NT)
lysenin, was expressed in E. coli BL21 (DE3) as a fusion protein with a 6xHis-tag followed by the
fluorescentproteinmCherry(totalMW~45kDa)atN-terminal,aspurifiedandvalidatedpreviously1.
Thechol-specifictoxinfragment, i.e.thefourthdomainofperfringolysine(thetaD4),wasclonedin
pET28 containing 6xHis- and LPETGG-tags in N- and C-terminal, respectively (Fig S1a). It was
generated from pET28/His-mCherry-theta2 by removing the mCherry sequence and adding the
LPETGG-tag.TheresultingplasmidwastransformedinE.coliBL21(DE3)andtheproteinexpressedin
LBmediumfor72hat16°Cafteradditionof0.4mMisopropyl-β-D-thiogalactoside.Bacterialextracts
andproteinpurificationwerepreparedaspreviouslydescribed2.Analysisofthepurifiedproteinby
westernblotrevealedrecombinantthetaattheexpectedsize(~16kDa;Fig.S1b).Themostenriched
fractionswerepooled,concentratedusingVivaspinturbo15columns(Sartorius),andtheimidazole
was removed by desalting onNAP-5 columns (GE Healthcare). Purified proteinwas finally kept in
aliquotsin10mMHEPES(pH7.2)10mMNaClandstoredat-80°Cuntiluse.Proteinconcentrationin
thepurifiedfractionwasestimatedbymeasuringA280andassumingamolarabsorptivityof44500M-
1.cm-1.From1lofculture,weobtained10gofprotein.Thetoxinbindingspecificitywasverifiedon
MLVscontainingincreasingamountsofChol(Fig.S1b)andpreparedaspreviouslydescribed1.
Electronic Supplementary Material (ESI) for ChemComm.This journal is © The Royal Society of Chemistry 2018

Chem.Comm. 2
AFMtip functionalization.Lysenin-andθ-functionalizedAFMtipswereobtainedusingNHS-PEG27-
acetal linkers3.ToobtainNHS-PEG27-acetaltips,AFMcantileverswerefirstcleanedwithchloroform
for10min,rinsedwithethanol,N2driedandthencleanedfor15mininanultravioletradiationand
ozonecleaner(UV-O,Jetlight,CA,USA).Thecantileverswereimmersedinanethanolaminesolution
(3.3gethanolaminein6.6mlDMSO)overnightandthenrinsedinDMSO(3x1min)andethanol(3x
1min), followedbyN2drying.To attach the linker to theAFMtip,1mgofNHS-PEG27-acetalwas
dilutedin0.5mlchloroformwith30µltriethylamineandcantileverswereimmersedinthissolution
for 2 h. The cantileverswere then cleaned 3 x 10min in chlorofom and driedwith N2. Next, the
cantileverswereimmersedina1%citricacidsolutionfor10minutesandrinsedwithpurewater(3x
5min),followedbydryingwithN2.
To obtain Lysenin-tips, 100 µl of a 100 µM tris-nitrilotriacetic amine trifluoroacetate (tris-NTA)
solutionwaspipettedontothecantileversand2μlofafreshlyprepared1MNaCNBH3solutionwas
then added and gently mixed. The cantilevers were incubated for 1 h, then 5 μl of a 1 M
ethanolaminesolutionpH8.0wereaddedfor10minutestoquenchthereaction.Cantileverswere
washed in Tris buffer (3 x 5 min) and then incubated for 1.5 h with 100 μl of a 10 μM Lysenin
solution.Lysenin-tipswererinsedwithTrisbufferandstored in individualwellsofamultiwelldish
untilusedinAFMexperiments(typicallywithin48h).
θ-functionalizedAFMtipswereobtainedbyincubatingNHS-PEG27-acetalcantileversin100µlofa1
mMGGGGGGGGGGKpeptide(Gly10Lys,Genscript,USA)solutiontowhich2μlofafreshlyprepared1
MNaCNBH3 solutionwasadded.After1h, then5μlofa1Methanolamine solutionpH8.0were
addedfor10minutestoquenchthereaction.Cantileverswerethenincubatedwith20µlofa10µM
θ-toxinsolutionand20µlofa10µMSortaseAsolutionfor1hat37⁰C.θ-tipswererinsedwithTris
bufferandstoredinindividualwellsofamultiwelldishuntilusedinAFMexperimentsthesameday.
FD-based AFM on supported lipid bilayers. AFM experiments were performed with a Bioscope
Resolve AFM (Bruker) operated in “PeakForce Tapping QNMmode” in imaging Buffer B at room
temperature (≈24⁰C). Rectangular Si3N4 cantilevers (AC40, Bruker) with a sharpened tetrahedral
silicontip,nominalspringconstantsof0.09N/mandresonancefrequencyinliquidof≈25kHzwere
used.The springconstantof thecantileverswascalibratedusing the thermalnoisemethodat the
endofeachexperiment4andwasfoundtobeof0.08±0.01N/m.
InFD-basedAFMmeasurements,theAFMcantileverisoscillatedwellbelowitsresonancefrequency
inasinusoidalmanner,whilethesamplesurfaceiscontouredpixel-by-pixel.Aforce-distancecurveis
recorded for each approach and retraction of the oscillating cantilever. FD-based AFM height,
Young’smodulusandadhesionmapsare thenobtainedbydoingapixel-by-pixel reconstructionof
theacquireddata.FD-basedmultiparametricmapswereacquiredusingaforcesetpointof100-200

Chem.Comm. 3
pN.TheAFMcantileverwasoscillatedverticallyat0.25kHzwithpeak-to-peakoscillationamplitudes
of100nm.Imageswererecordedusingascanrateof0.2Hzand256x256pixels.
Data analysis.Raw FD curveswere processed offline using theNanoScopeAnalysis 1.80 Software
(Bruker).ToreconstructYoung’smodulusmaps,weanalyzedtheapproachpartoftheforce-distance
curves from PeakForce QNMmaps. The best quality of the fit was obtained when by fitting the
contactpartofthecurvewiththeHertzmodel5,6:
𝐹!/! = !!
!(!!!!)
𝑅!/!
𝛿 (1)
whereEistheYoung’smodulus,δistheindentationdepth,νisthePoissonratio,andRisthecontact
radius.We used a Poisson’s ratio value of 0.3. Young’smoduluswas calculated from the slope of
Equation1.
HeightimageswereprocessedusingtheGwyddionfreeSPMsoftware.Afirstorsecondorderplane
fitwasperformed.Imagesdidnotundergofurtherprocessing.

Chem.Comm. 4
Fig.S1.Toxinfragmentproduction(a)andvalidationofthethetatoxinfragment(b).(a)MappingofpET28-expressing lysenin or theta toxin. (b) Theta toxin fragment (16 kDa) binding specificityvalidationonmultilamellarvesicles(MLVs)containingincreasingCholcontents.Thetawasincubatedwith MLVs made of SM and PC and containing (4 last wells) or not (2 first wells) Chol. Aftercentrifugation,pellets(P)containingMLVsandsupernatants(SN)wereanalysedbywesternblottingusinganti-Hisantibodies.The6xHis-taggedtoxinonlybindsChol-containingMLVs(P).

Chem.Comm. 5
Fig. S2. Validating the specificity of the interactions detected by toxin-derivatized AFM tips. Thelysenin-tipwasusedtoprobeaSM-free lipidbilayerofDOPC:Chol (70:30). (a,b,c)Height,adhesionandYoung’smodulus imagesoftheDOPC:Chol(70:30).NospecificunbindingeventwereobservedbetweentheLysenin-tipandtheDOPC:Cholsample. (d,e,f)Aθ-functionalizedAFMtipwasusedtomap a Chol-free lipid bilayer made of DOPC:SM (70:30) (d) High-resolution height image of theDOPC:SM(70:30)bilayerand(e)correspondingadhesionshowingnointeractionbetweenthetipandthesampleand(f)Young’smodulusmap

Chem.Comm. 6
Fig. S3. Mapping SM-enriched domains with bare AFM tips on DOPC:SM 50:50 and 70:30 lipidbilayers. (a,b) Height images of the bilayers and corresponding (c,d) adhesion and (e,f) Young’smodulusmaps.
Chem.Comm. 7
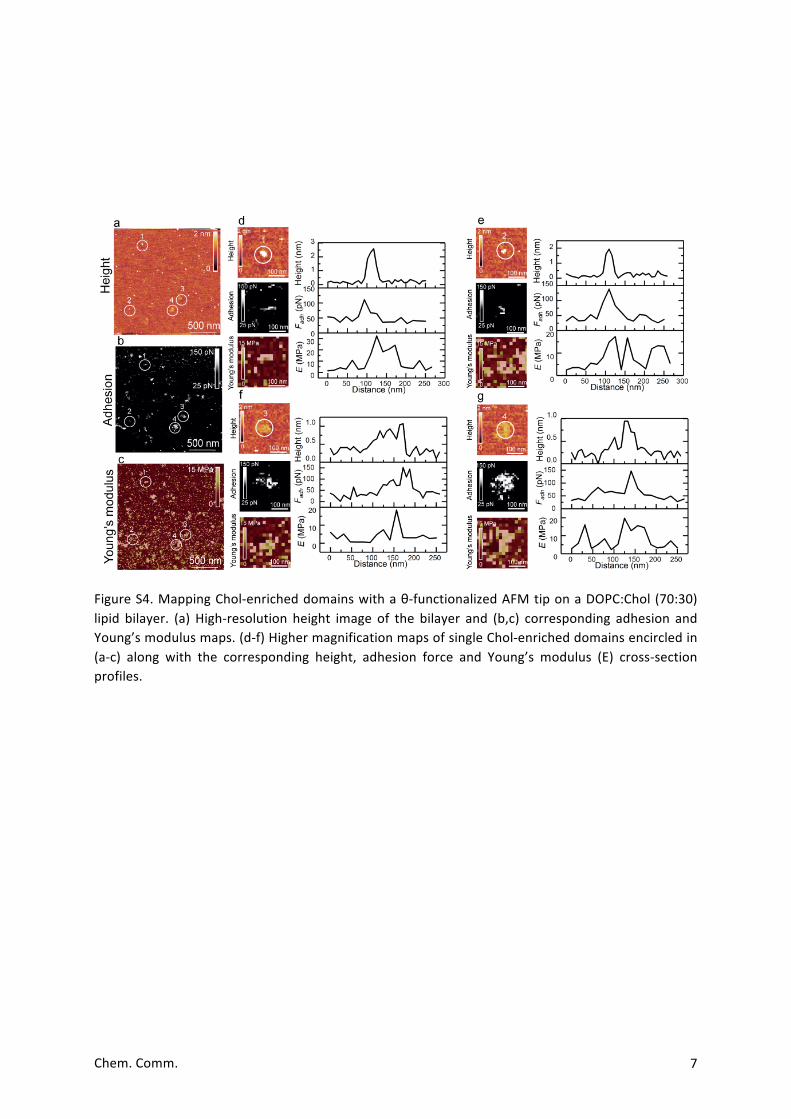
FigureS4.MappingChol-enricheddomainswithaθ-functionalizedAFMtiponaDOPC:Chol(70:30)lipid bilayer. (a) High-resolution height image of the bilayer and (b,c) corresponding adhesion andYoung’smodulusmaps.(d-f)HighermagnificationmapsofsingleChol-enricheddomainsencircledin(a-c) along with the corresponding height, adhesion force and Young’s modulus (E) cross-sectionprofiles.

Chem.Comm. 8
FigS5.MappingChol-enricheddomainswithaθ-functionalizedAFMtiponaDOPC:Chol(70:30)lipidbilayer.(a)High-resolutionheightimageofthebilayerand(b,c)correspondingadhesionandYoung’smodulusmaps.
Chem.Comm. 9
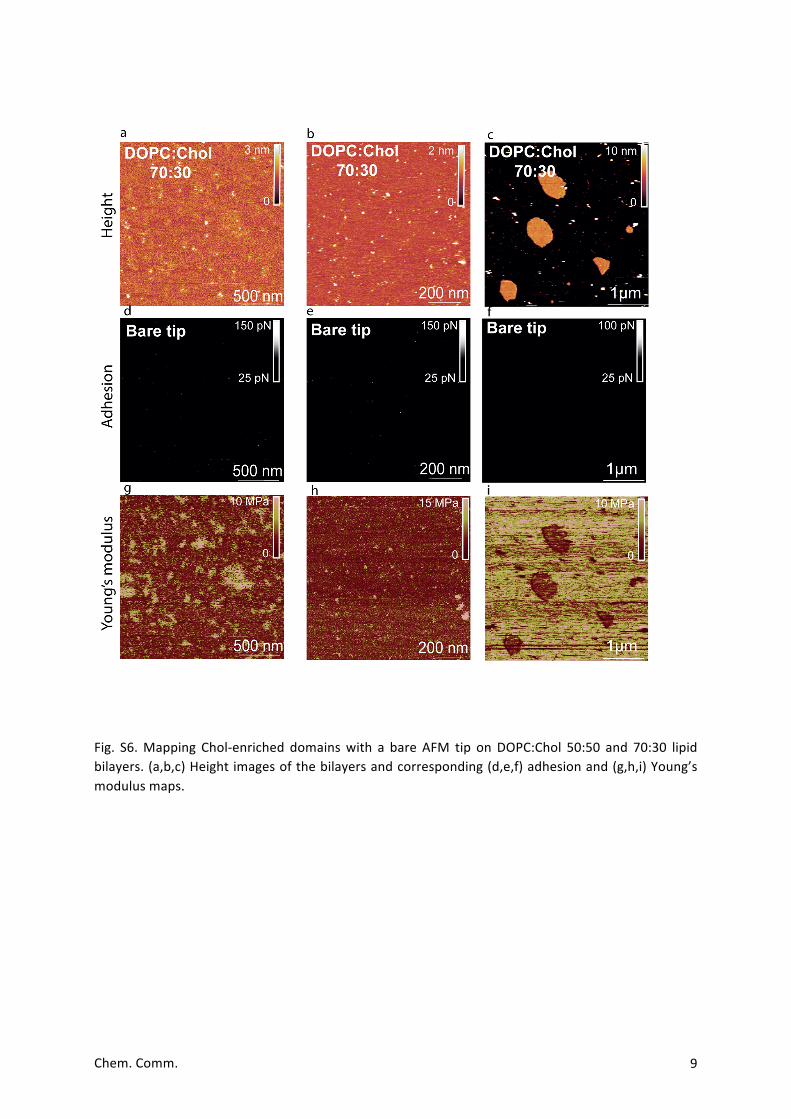
Fig. S6.Mapping Chol-enriched domainswith a bare AFM tip onDOPC:Chol 50:50 and 70:30 lipidbilayers.(a,b,c)Heightimagesofthebilayersandcorresponding(d,e,f)adhesionand(g,h,i)Young’smodulusmaps.

Chem.Comm. 10
References
1. M.Carquin,H.Pollet,M.Veiga-da-Cunha,A.Cominelli,P.VanDerSmissen,F.N’kuli,H.Emonard,P.Henriet,H.Mizuno,P.J.CourtoyandD.Tyteca,JournalofLipidResearch,2014,55,1331-1342.
2. M.Carquin,L.Conrard,H.Pollet,P.VanDerSmissen,A.Cominelli,M.Veiga-da-Cunha,P.J.CourtoyandD.Tyteca,CellularandMolecularLifeSciences,2015,72,4633-4651.
3. L.Wildling,B.Unterauer,R.Zhu,A.Rupprecht,T.Haselgrübler,C.Rankl,A.Ebner,D.Vater,P.Pollheimer,E.E.Pohl,P.HinterdorferandH.J.Gruber,BioconjugateChemistry,2011,22,1239-1248.
4. J.L.HutterandJ.Bechhoefer,ReviewofScientificInstruments,1993,64,1868-1873.5. H.Schillers,C.Rianna,J.Schäpe,T.Luque,H.Doschke,M.Wälte,J.J.Uriarte,N.Campillo,G.
P.A.Michanetzis,J.Bobrowska,A.Dumitru,E.T.Herruzo,S.Bovio,P.Parot,M.Galluzzi,A.Podestà,L.Puricelli,S.Scheuring,Y.Missirlis,R.Garcia,M.Odorico,J.-M.Teulon,F.Lafont,M.Lekka,F.Rico,A.Rigato,J.-L.Pellequer,H.Oberleithner,D.NavajasandM.Radmacher,ScientificReports,2017,7,5117.
6. H.Hertz,JournalfürdiereineundangewandteMathematik,1882,92,156-171.



![Index [] · The Power of Functional Resins in Organic Synthesis. ... α-chymotrypsin 603 Aβ (β-amyloid (1-42)) synthesis 504, 507, 508 Accurel MP 1000 373 acetal-protected carbonyls](https://static.fdocument.org/doc/165x107/5f6421717515ab779846508d/index-the-power-of-functional-resins-in-organic-synthesis-chymotrypsin.jpg)














