Capítulo 8. homeostasis de la glucosa
11
Capítulo 8. Homeostasis de la glucosa L. N. Noé González Gallegos
-
Upload
centro-universitario-del-norte-universidad-de-guadalajara -
Category
Health & Medicine
-
view
497 -
download
2
Transcript of Capítulo 8. homeostasis de la glucosa

Capítulo 8. Homeostasis de la
glucosaL. N. Noé González Gallegos

Fig. 8.1 Los tres estados de la
homeostasis de la glucosa.

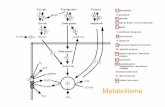
Fig. 8.3 Resumen del metabolismo de los combustibles en la situación posprandial
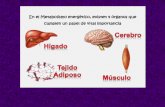
1. La elevación de la glucemia provoca la liberación de insulina por parte de las células β del páncreas. La disponibilidad del sustrato y el aumento de la insulina estimulan la síntesis de glucógeno, triacilgliceroles (triglicéridos) y proteínas en los tejidos; este es un estado anabólico.
2. La glucosa es el único combustible para el cerebro; su captación por este órgano es independiente de la insulina.
3. El músculo y el tejido adiposo también utilizan glucosa pero su captación es dependiente de la insulina.

Fig. 8.4 Resumen del metabolismo de los combustibles en ayunas, de 4 a 12 h después de una comida. La elevación del cociente glucagón:insulina activa la degradación del glucógeno hepático, que proporciona glucosa al cerebro. El descenso de la concentración de insulina y el aumento de noradrenalina promueven la hidrólisis de los triacilgliceroles almacenados, liberando ácidos grasos que pueden utilizarse como combustible por parte del hígado y el músculo. El músculo utiliza su propio glucógeno como combustible.1. La degradación del glucógeno
hepático proporciona glucosa para que sea oxidada por el cerebro. Este depósito solo dura entre 12 y 24 h.
2. La hidrólisis de triacilgliceroles de los depósitos libera ácidos grasos, usados preferencialmente por músculo e hígado como combustibles.
3. Los músculos también pueden utilizar su propio glucógeno como combustible.

Fig. 8.5 Resumen del metabolismo de los combustibles en la inanición inicial. La noradrenalina y el cortisol activan la degradación de proteínas musculares para liberar aminoácidos, especialmente alanina y glutamina. La noradrenalina también activa la hidrólisis de los triacilgliceroles para liberar glicerol. El glicerol, la alanina y la glutamina se transportan al hígado, donde se incorporan a la gluconeogenia y son oxidados para formar glucosa. La glucosa es utilizada fundamentalmente por el cerebro. Los ácidos grasos liberados por la hidrólisis de los triacilgliceroles pueden trasladarse al hígado y usarse para generar cuerpos cetónicos, usados por el cerebro y otros tejidos (los números hacen referencia al texto a continuación).
1. El glucagón y después el cortisol, activan la degradación de proteínas musculares, liberándose aminoácidos (especialmente alanina y glutamina).
2. La hidrólisis de los depósitos de triacilgliceroles (tejido adiposo) libera glicerol. El hígado emplea los aminoácidos y el glicerol para la gluconeogenia.
3. El cerebro utiliza la glucosa producida.4. Los ácidos grasos liberados de los
triacilgliceroles también son usados por el hígado para producir cuerpos cetónicos, que pueden emplearse como combustible alternativo en los tejidos periféricos, así como en el cerebro.

Fig. 8.7 El ciclo de Cori distribuye la carga metabólica entre el hígado y el músculo. El lactato, que se acumula en el músculo durante la actividad intensa, se transporta al hígado para volver a convertirse en glucosa gracias a la gluconeogenia. Esto repone combustible para el músculo y previene la acidosis láctica.

Fig. 8.8 Vía de señales de la insulina. La insulina se une a su receptor, lo que resulta en una autofosforilación. Entonces, esto fosforila la proteína Cbl, que forma un complejo con la proteína adaptadora CAP. El complejo Cbl/CAP interacciona a continuación con la proteína adaptadora Crk, que está asociada estructuralmente con C3G, una proteína de intercambio GTP/GDP. La C3G activa las TC10, que promueven la translocación del GLUT4 a la membrana plasmática. La autofosforilación del receptor de insulina también fosforila el sustrato 1 del receptor de insulina (IRS-1). El IRS-1 atrae a p85, que se une a p110, que entonces activa la fosfatidilinositol-3-cinasa (PI-3-cinasa). La PI-3-cinasa activa la proteína cinasa B (PKB; Akt), que actúa sobre vías posteriores, afectando al metabolismo de la glucosa, los lípidos y las proteínas, y a la expresión de genes específicos. La Akt también promueve la translocación del GLUT-4 a la membrana plasmática, lo que resulta en un aumento de la captación de glucosa.

Fig. 8.9 Efecto del aumento del cociente glucagón:insulina en la diabetes.

Fig. 8.12 Hiperglucemia y cetoacidosis diabética. En ausencia de insulina, la hiperglucemia causa diuresis osmótica. La pérdida de líquido y electrólitos resulta en deshidratación. El aumento de la cetogenia provoca acidosis metabólica. La compensación respiratoria da lugar a hiperventilación. Hay que corregir paralelamente la deshidratación y la hiperglucemia mediante el tratamiento con insulina.

Fig. 8.15 Complicaciones a largo plazo de la diabetes.

Bibliografía Metabolismo de los hidratos de carbono. En: Horton-Szar D, editor. Lo esencial en metabolismo y nutrición. 4 ed. España: Elsevier; 2013.