Biosynthèse du lactosucrose à partir du lactose et du saccharose ...
219
Biosynthèse du lactosucrose à partir du lactose et du saccharose par approche homogène en utilisant la β-galactosidase libre et par approche hétérogène en utilisant la β-galactosidase immobilisée sur différents matériaux siliceux mésoporeux Mémoire Zeineb Ben Rejeb Maîtrise en génie agroalimentaire Maître ès sciences (M. Sc.) Québec, Canada © Zeineb Ben Rejeb, 2014
Transcript of Biosynthèse du lactosucrose à partir du lactose et du saccharose ...

Biosynthèse du lactosucrose à partir du lactose et du saccharose par approche homogène en utilisant la
β-galactosidase libre et par approche hétérogène en utilisant la β-galactosidase immobilisée sur différents
matériaux siliceux mésoporeux
Mémoire
Zeineb Ben Rejeb
Maîtrise en génie agroalimentaire Maître ès sciences (M. Sc.)
Québec, Canada
© Zeineb Ben Rejeb, 2014


III
Résumé court
Le lactosucrose est un trisaccharide synthétisé par la bioconversion du lactose et du
saccharose en utilisant la β-galactosidase à partir d’Aspergillus oryzae.
La production industrielle du lactosucrose est onéreuse à cause du coût élevé des
enzymes, d’où le recours à immobiliser ces enzymes. Dans cette optique, sept différents
matériaux siliceux mesostructurés ont été utilisés pour immobiliser la β-galactosidase, en
utilisant le glutaraldhéyde. La majorité des matériaux présente une bonne activité.
Cependant, l’enzyme immobilisée sur la silice en forme de mousse montre un rendement
maximal, dépassant celui de l’enzyme libre. En plus, un rendement équivalent à 75% de
celui obtenu à la première utilisation du matériau est maintenu pour la plupart des
biocatalyseurs, après un 5ème recyclage.
Une concentration équimolaire de 60% entre les substrats, un pH de 5 et une température
de 40°C se sont révélés optimaux pour la production du lactosucrose qui a atteint une
concentration de 76,27 g/L.


V
Résumé long
Les oligosaccharides sont des glucides qui modulent la flore colique et améliorent
l’équilibre de la santé humaine. Ils sont largement utilisés comme prébiotiques. Le
lactosucrose est un oligosaccharide qui attire de plus en plus le milieu scientifique et
industriel. Il est un trisaccharide (Glucose-Fructose-Galactose), non digestible avec un
pouvoir sucrant faible. Ce trisaccharide peut être synthétisé par voie enzymatique, via la
bioconversion du lactose et du saccharose, et ceci en utilisant la levansucrose, la
fructofuranosidase ou la β-galactosidase.
Durant ce travail, la β-galactosidase extraite à partir d’Aspergillus oryzae a été utilisée
pour synthétiser le lactosucrose. Dans un premier temps, et afin d’optimiser le rendement
en lactosucrose, nous avons étudié différents paramètres avec l’enzyme libre. Une
concentration équimolaire de 60% entre le lactose et le saccharose, un pH de 5 et une
température de 40°C se sont révélés optimaux pour la production du lactosucrose.
L’optimum est obtenu avec un rendement de 25% (76,27 g/L) en lactosucrose. Un sous-
produit a été isolé, il s’agit d’un autre oligosaccharide (tetrahexose) avec une
concentration de 14,52 g/L. La production actuelle du lactosucrose et d’autres
oligosaccharides à l’échelle industrielle est onéreuse due au coût élevé des enzymes, d’où
le recours à immobiliser celles-ci pour pouvoir les recycler et les réutiliser et par
conséquent, minimiser les coûts de production. Dans cette deuxième partie, nous avons
utilisé sept différents matériaux siliceux pour immobiliser l’enzyme (β-galactosidase
d’Aspergillus oryzae) en utilisant le glutaraldéhyde (GA) comme agent de ‘cross-linking’, et
effectuer la réaction de transglycosylation par la suite dans une approche hétérogène et
comparer le rendement de la réaction avec celui de l’enzyme libre (approche homogène).
Les matériaux synthétisés ont différentes tailles de pores qui varient de 5 à 24 nm et aussi
différentes formes poreuses (hexagonale, cubique, sphérique, forme d’oignon, etc.). Cette
différence de taille et de forme a affecté énormément la rétention enzymatique par ces
matériaux ainsi que le rendement de la réaction en phase hétérogène.
La majorité des matériaux synthétisés présentent une bonne activité. Cependant,
l’enzyme immobilisée sur la silice en forme de mousse (MCF) montre un rendement
maximal, qui dépasse celui obtenu par l’enzyme libre, ce qui confirme que l’immobilisation
d’enzyme sur des supports siliceux mesostructurés peut conférer à l’enzyme des

VI
conditions réactionnelles améliorées afin d’augmenter sa stabilité et par conséquent sa
performance.
Le recyclage de ces biocatalyseurs a été effectué 5 fois successives. Le rendement
obtenu à la 5ème régénération est équivalant à plus que 80% du rendement initial pour la
majorité des matériaux testés.

VII
Abstract
Oligosaccharides are non-digestible carbohydrates that modulate the colonic flora, and
improve the human health balance. They are widely used as prebiotics. Lactosucrose is an
oligosaccharide increasingly attracting scientists and industrials. It is a non-digestible
trisaccharide (Glucose-Fructose-Galactose) with a low sweetening power. This
trisaccharide can be synthesized enzymatically through bioconversion of lactose and
sucrose, using the levansucrose, the fructofuranosidase and the β-galactosidase.
During this work, Aspergillus oryzae β-galactosidase was used to synthetize the
lactosucrose. First, and for lactosucrose yield optimization, we investigated different
parameters with the free enzyme in solution. Equimolar concentrations of 60% lactose and
sucrose, a pH of 5, and a temperature of 40°C, have been found optimal for lactosucrose
production. Optimum lactosucrose yield of 25% (76.27 g/L) was obtained under these
conditions. Another oligosaccharide (tetrahexose) was also isolated as a by-product with a
concentration of 14.52 g/L, so that the biosynthesis produced a total oligosaccharides yield
of 30%.
Currently, the production of lactosucrose and other oligosaccharides at the industrial scale
is still highly costly due to the high cost of enzymes. Thus, immobilizing them is greatly
beneficial being able to be recycled and reused, and therefore minimizes the production
process costs. In this second part, we used seven different highly structuredsiliceous
materials for the enzyme immobilization (Aspergillus oryzae β -galactosidase) using
glutaraldehyde (GA) as a cross-linking agent, and then performing the transglycosylation
reaction in a heterogeneous approach, as well as comparing the reaction yield with that
obtained with free enzyme in solution (homogeneous approach). The materials have pore
sizes ranging from 5 to 24 nm, with different porous shapes (hexagonal, cubic, spherical,
rod-like, vesicle-like, etc.). These differences in size and shape affected greatly the
enzyme immobilization on these materials as well as the heterogeneous phase reaction
yield.
Most materials exhibit good activity. However, the enzyme immobilized on mesostructured
cellular silica foam (MCF) shows a maximum yield, which exceeded that obtained with free
enzyme, confirming that immobilization of enzyme on mesostructured silica supports may

VIII
improve the enzyme performance under the investigated conditions by increasing its
stability and therefore its catalytic activity and selectivity.
Theses biocatalysts were regenerated and recycled, and then they were involved in five
successive cycles. The yield obtained on the 5th cycle represented more than 80% of the
initial yield for 86% of the tested materials.

IX
Table des matières
RÉSUMÉ COURT .............................................................................................................................. III
RÉSUMÉ LONG .................................................................................................................................. V
ABSTRACT ....................................................................................................................................... VII
TABLE DES MATIERES .................................................................................................................... IX
LISTE DES TABLEAUX ................................................................................................................... XIII
LISTE DES FIGURES ....................................................................................................................... XV
LISTE DES ABRÉVIATIONS ET DES SIGLES ............................................................................... XXI
DÉDICACE .....................................................................................................................................XXIII
REMERCIEMENTS ........................................................................................................................ XXV
AVANT-PROPOS ......................................................................................................................... XXVII
CHAPITRE 1. INTRODUCTION GENERALE ................................................................................ 1
CHAPITRE 2. REVUE DE LITTERATURE .................................................................................... 3
2.1. LACTOSÉRUM ........................................................................................................................ 3
2.1.1. Définition et composition ............................................................................................. 3
2.1.2. Utilisation et valorisation du lactosérum ...................................................................... 5
2.2. LACTOSE ............................................................................................................................... 7
2.2.1. Production et composition ........................................................................................... 7
2.2.2. Applications du lactose .............................................................................................. 10
2.3. OLIGOSACCHARIDES ET ALIMENTS FONCTIONNELS ................................................................. 12
2.3.1. Oligosaccharide : Définition et propriété ................................................................... 12
2.3.2. Aliments fonctionnels: définition et propriétés ........................................................... 14
2.4. LE LACTOSUCROSE .............................................................................................................. 15
2.4.1. Structure .................................................................................................................... 15
2.4.2. Caractéristiques et production mondiale ................................................................... 15
2.4.3. Synthèse du lactosucrose ......................................................................................... 18
2.5. Β–GALACTOSIDASES: STRUCTURE, PRODUCTION, MECANISME DE REACTION ET APPLICATION .. 22
2.5.1. Structure .................................................................................................................... 22
2.5.2. Production ................................................................................................................. 23
2.5.3. Mécanisme de la réaction et applications ................................................................. 25
2.5.4. β-Galactosidase de Aspergillus oryzae et son activité de transgalactosylation........ 27
2.6. IMMOBILISATION DES ENZYMES ............................................................................................. 28

X
2.6.1. Avantage d’immobilisation des enzymes .................................................................. 29
2.6.2. Immobilisation des enzymes : supports .................................................................... 30
2.6.3. Méthode d’immobilisation des enzymes .................................................................... 34
2.7. CONCLUSION ....................................................................................................................... 42
CHAPITRE 3. HYPOTHESES ET OBJECTIFS DE RECHERCHE ICI ........................................ 43
3.1. HYPOTHÈSES DU TRAVAIL ..................................................................................................... 43
3.2. OBJECTIFS DE TRAVAIL ......................................................................................................... 43
3.2.1. Objectif principal ........................................................................................................ 43
3.2.2. Objectifs spécifiques .................................................................................................. 43
CHAPITRE 4. MATERIELS ET METHODES ............................................................................... 45
4.1. MATERIELS BIOLOGIQUES ..................................................................................................... 45
4.2. PRODUITS CHIMIQUES .......................................................................................................... 45
4.3. ÉQUIPEMENTS ..................................................................................................................... 46
4.4. METHODOLOGIE ................................................................................................................... 46
4.4.1. Réaction enzymatique avec l’enzyme libre (approche homogène) ........................... 46
4.4.2. Les analyses HPLC ................................................................................................... 48
4.4.3. Calculs des rendements ............................................................................................ 48
4.4.4. Purification et caractérisation des produits formés .................................................... 48
4.4.4.1. UPLC-MS ............................................................................................................................ 48
4.4.4.2. Analyse du lactosucrose par résonance magnétique nucléaire .......................................... 49
4.4.5. La synthèse des supports .......................................................................................... 49
4.4.5.1. Synthèse du support mésoporeux SBA-15 ......................................................................... 49
4.4.5.2. Synthèse de nanomateriaux en forme de barre (Rod-like) ‘’S2.9’’ et en forme de vésicule
(Vesicle-like) ‘’S34.8’’ ........................................................................................................................... 50
4.4.5.3. Synthèse des MCF (Mesocellular Siliceous Foams) ........................................................... 50
4.4.5.4. Synthèse de LP-FDU-12 ..................................................................................................... 51
4.4.6. Caractérisations des supports ................................................................................... 51
4.4.6.1. Analyses BET ..................................................................................................................... 51
4.4.6.2. Diffraction des rayons X (DRX) ........................................................................................... 52
4.4.6.3. Analyses de Microscopie Électronique à Transmission (MET) ........................................... 52
4.4.7. Immobilisation de β-Galactosidase ........................................................................... 52
4.4.7.1. Immobilisation ‘Cross-linking’ .............................................................................................. 52
4.4.7.2. Rétention d’enzymes par les supports ................................................................................ 53
4.4.8. Réaction enzymatique avec l’enzyme immobilisée (approche hétérogène) ............. 54
4.4.9. Analyses statistiques ................................................................................................. 55
CHAPITRE 5. RESULTATS ET DISCUSSIONS .......................................................................... 57
5.1. RÉACTION ENZYMATIQUE ...................................................................................................... 57

XI
5.1.1. Identification des produits par HPLC, spectrométrie de masse et par la RMN du
proton 57
5.2. RÉACTION ENZYMATIQUE EN PHASE HOMOGÈNE .................................................................... 67
5.2.1. La cinétique de la réaction enzymatique ................................................................... 67
5.2.2. Effet de la concentration enzymatique ...................................................................... 68
5.2.3. Effet de concentration des substrats ......................................................................... 74
5.2.4. Effet du pH................................................................................................................. 80
5.2.5. Effet de la température .............................................................................................. 82
5.2.6. Conclusion ................................................................................................................. 85
5.3. REACTION ENZYMATIQUE EN PHASE HETEROGENE ................................................................. 87
5.3.1. Caractérisation des matériaux................................................................................... 87
5.3.1.1. SBA-15 ............................................................................................................................... 87
5.3.1.1.1. Analyse de la surface BET ........................................................................................... 87
5.3.1.1.2. Analyse par diffraction des rayons X (DRX) ................................................................. 88
5.3.1.1.3. Microscopie Électronique à Transmission (MET) ......................................................... 89
5.3.1.2. Mousse mésocellulaire (MCF) ............................................................................................ 90
5.3.1.2.1. Analyse de la surface BET ........................................................................................... 90
5.3.1.2.2. Analyse par diffraction des rayons X (DRX) ................................................................. 91
5.3.1.2.3. Microscopie Électronique à Transmission (MET) ......................................................... 92
5.3.1.3. Nanomatériaux sous forme de barres (rod-like) S-2,9 et sous forme de vésicules (vesicle-
like) S-34,8 ........................................................................................................................................... 94
5.3.1.3.1. Analyse de la surface BET ........................................................................................... 94
5.3.1.3.2. Analyse par diffraction des rayons X (DRX) ................................................................. 95
5.3.1.3.3. Microscopie Électronique à Transmission (MET) ......................................................... 97
5.3.1.4. Les LP-FDU-12 ................................................................................................................... 99
5.3.1.4.1. Analyse de la surface BET ........................................................................................... 99
5.3.1.4.2. Analyse par diffraction des rayons X (DRX) ............................................................... 101
5.3.1.4.3. Microscopie Électronique à Transmission (MET) ....................................................... 103
5.3.2. Immobilisation de β-galactosidase .......................................................................... 105
5.3.2.1. Effet de pH sur la rétention des enzymes ......................................................................... 106
5.3.2.2. Effet de la concentration d’enzyme sur la quantité immobilisée lors de la rétention ......... 107
5.3.2.3. Effet de la concentration de Glutaraldéhyde (GA) sur la rétention des enzymes
immobilisées ...................................................................................................................................... 113
5.3.3. Réaction de transglycosylation avec l’enzyme immobilisée ................................... 115
5.3.3.1. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée sur la MCF .............. 115
5.3.3.2. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée sur la SBA-15 ......... 118
5.3.3.3. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-2.9 ............. 120
5.3.3.4. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-34.8 ........... 123
5.3.3.5. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-10-100 ....... 125
5.3.3.6. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-15-100 ....... 128

XII
5.3.3.7. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-T-100 ......... 130
5.3.3.8. Comparaison entre la réactivité des matériaux ................................................................. 134
5.3.4. Activité des biocatalyseurs synthétisé après recyclage .......................................... 136
CONCLUSION GÉNÉRALE ............................................................................................................ 143
RÉFÉRENCES BIBLIOGRAPHIQUES ........................................................................................... 145
ANNEXES........................................................................................................................................ 153

XIII
Liste des tableaux
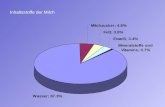
TABLEAU 1. COMPOSITION DU LAIT BOVIN ET DU LACTOSERUM (SMITHERS ET AL., 1996) ..................................................... 4
TABLEAU 2. COMPOSITION MOYENNE DU LAIT SELON L’ESPECE (G/L) ................................................................................ 8
TABLEAU 3. PRODUCTION DES OLIGOSACCHARIDES DE QUALITE ALIMENTAIRE ESTIMEE EN 1995 .......................................... 17
TABLEAU 4. CONDITIONS ET RENDEMENT EN LACTOSUCROSE PAR LES DIFFÉRENTES ESPÈCES ENZYMATIQUES............................ 21
TABLEAU 5. SOURCE DE Β GALACTOSIDASE ................................................................................................................. 24
TABLEAU 6. LES PRINCIPAUX PRODUITS FABRIQUÉS À L'ÉCHELLE INDUSTRIELLE EN UTILISANT DES ENZYMES IMMOBILISÉES
(BORNSCHEUER, 2008) ................................................................................................................................. 30
TABLEAU 7. PROPRIÉTÉS CARACTÉRISTIQUES DES DIFFÉRENTS MATÉRIAUX DE SILICE MÉSOPOREUSES UTILISÉS POUR ENCAPSULER
DES ENZYMES ............................................................................................................................................... 34
TABLEAU 8. DIFFERENTES METHODES ET DIFFERENTS SUPPORTS D’IMMOBILISATION DE Β-GALACTOSIDASE A PARTIR DE
ASPERGILLUS ORYZAE ..................................................................................................................................... 40
TABLEAU 9. LES CONDITIONS OPERATOIRES DE LA BIOSYNTHESE DE LACTOSUCROSE A PARTIR DE LACTOSE ET DE SACCHAROSE ..... 47
TABLEAU 10. PREPARATION DES STANDARDS A CONCENTRATION PROTEIQUE CONNUE ........................................................ 53
TABLEAU 11. RECAPITULATIF DES DEPLACEMENTS CHIMIQUES DU GOS I ANALYSE PAR RESONANCE MAGNETIQUE NUCLEAIRE (1H-
RMN) ET IDENTIFIE COMME DU LACTOSUCROSE. ................................................................................................. 64
TABLEAU 12. LES DIFFERENTES CONCENTRATIONS DE SACCHAROSE ET DE LACTOSE ETUDIEES POUR LA BIOSYNTHESE DE
LACTOSUCROSE ............................................................................................................................................. 74
TABLEAU 13. LES PROPRIETES TEXTURALES DES SUPPORTS SYNTHETISES. ........................................................................ 105
TABLEAU 14. RESULTATS DU TEST DU TUKEY POUR LA MCF ET LA CONCENTRATION D’ENZYME ........................................... 110
TABLEAU 15. RESULTATS DU TEST DE TUKEY POUR LA S 2,9 ET LA CONCENTRATION D’ENZYME ........................................... 111
TABLEAU 16. RESULTATS DU TEST DE TUKEY POUR LA S 34,8 ET LA CONCENTRATION D’ENZYME ......................................... 111
TABLEAU 17. RESULTATS DU TEST DE TUKEY POUR LA S-T-100 ET LA CONCENTRATION D’ENZYME ...................................... 112
TABLEAU 18. RESULTATS DU TEST DE TUKEY POUR LA S-15-100ET LA CONCENTRATION D’ENZYME ..................................... 112
TABLEAU 19. RESULTATS DU TEST DE TUKEY POUR LA S-10-100 ET LA CONCENTRATION D’ENZYME .................................... 112
TABLEAU 20. RESULTATS DU TEST DE TUKEY POUR LA S 34,8 ET LES CONCENTRATIONS DE GA ........................................... 114
TABLEAU 21. RESULTATS RECAPITULATIFS DES RENDEMENTS EN GALACTO-OLIGOSACCHARIDES (LACTOSUCROSE ET TETRAHEXOSE)
OBTENUS AVEC L’ENZYME IMMOBILISEE SUR LES DIFFERENTS MATERIAUX ............................................................... 133


XV
Liste des figures
FIGURE 1. CAILLES (COAGULATION DE LA CASEINE) ET LACTOSERUM .................................................................................. 4
FIGURE 2. PRINCIPAUX PRODUCTEURS DE FROMAGE DANS LE MONDE EN 2005 (EXPRIMES EN MILLIERS DE TONNES) .................. 5
FIGURE 3. L'UTILISATION COMMERCIALE DE LACTOSERUM DE LAIT ..................................................................................... 7
FIGURE 4. STRUCTURE CHIMIQUE DU LACTOSE............................................................................................................... 9
FIGURE 5. STRUCTURE DES MOLECULES DE LACTOSE EN CONFIGURATION Α ET Β (GÄNZLE ET AL., 2008) ............................. 9
FIGURE 6. VOIES DE TRANSFORMATION DE LACTOSE: LES DERIVES DE LACTOSE (SEKI AND SAITO, 2012) .......................... 11
FIGURE 7. REPRESENTATION SCHEMATIQUE DU PROCESSUS DE PRODUCTION D'OLIGOSACCHARIDES (SAKO ET AL., 1999) ..... 13
FIGURE 8. STRUCTURE DE LACTOSUCROSE .................................................................................................................. 15
FIGURE 9. BIOSYNTHESE DE LACTOSUCROSE PAR A: TRANSFRUCTOSYLATION ET B: TRANSGALACTOSYLATION (DAUDE ET AL., 2012,
LI ET AL., 2009) ........................................................................................................................................... 19
FIGURE 10. REACTIONS D’HYDROLYSE DE SACCHAROSE ET DE LACTOSUCROSE (FUJITA ET AL., 1990) ................................ 21
FIGURE 11. STRUCTURE 3-D DE Β-D-GALACTOSIDASE (A PARTIR DE PROTEIN DATA BANK). ................................................. 23
FIGURE 12. MECANISME DE REACTION POUR LA HYDROLYSE ET LA TRANSGLYCOSYLATION DU LACTOSE PAR KLUYVEROMYCES LACTIS
Β-GALACTOSIDASE (ZHOU AND CHEN, 2001) ............................................................................................... 26
FIGURE 13. LA BIOSYNTHESE DE LACTOSUCROSE ET SES ANALOGUES PAR Β-GALACTOSIDASE DE BACILLUS CIRCULANS A PARTIR DU
LACTOSE ET DU SACCHAROSE COMME SUBSTRATS (LI ET AL., 2009, MU ET AL., 2013) ...................................... 26
FIGURE 14. COMPARAISON DE LA TAILLE DES PORES DE CERTAINS MATERIAUX NANOPOREUX ET LA TAILLE MOYENNE MOLECULAIRE
DE CERTAINES ENZYMES COURAMMENT UTILISES (HUMPHREY AND WRIGHT, 2004) .................................................. 32
FIGURE 15. SCHEMA REPRESENTATIF DE HMMS (KIM ET AL., 2007) .............................................................................. 33
FIGURE 16. IMMOBILISATION COVALENTE DE L'ENZYME SUR UN SUPPORT: (A) DES RESIDUS D'ACIDE AMINE ACTIF; (B)
FONCTIONNALITE DE LIAISON DU SUPPORT; (C) SUPPORT; (D) ESPACE (CAO, 2006) .................................................. 36
FIGURE 17. REPRESENTATION DE L’IMMOBILISATION DES ENZYMES PAR ENCAPSULATION ..................................................... 36
FIGURE 18. REPRESENTATION DE L’IMMOBILISATION PAR RETICULATION «CROSS-LINKING» (KIM ET AL., 2008) .................. 38
FIGURE 19. CHROMATOGRAMME DES SOLUTIONS STANDARDS DES SUCRES (0,5G/L CHAQUE STANDARD). AVEC : A) SACCHAROSE,
B) LACTOSE, C) GLUCOSE, D) GALACTOSE, E) FRUCTOSE. ........................................................................................ 58
FIGURE 20. CHROMATOGRAMME TYPIQUE DES SUBSTRATS ET DES PRODUITS FORMES PAR TRANSGALACTOSILATION ET HYDROLYSE
................................................................................................................................................................. 59
FIGURE 21. MECANISME PROPOSE POUR LA BIOSYNTHESE DE LACTOSUCROSE AVEC LA Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE
................................................................................................................................................................. 60
FIGURE 22. SPECTRE MS DU GOS I .......................................................................................................................... 61
FIGURE 23. SPECTRE 13C-RMN DU GOS I ................................................................................................................ 62
FIGURE 24. SPECTRE 1H- RMN DU GOS I ................................................................................................................. 63
FIGURE 25. SPECTRE MS DU GOS II ......................................................................................................................... 66
FIGURE 26. CINETIQUE DE LA REACTION DE CONVERSION DES SUBSTRATS ET LA FORMATION DES PRODUITS ............................. 68

XVI
FIGURE 27. EFFET DE LA CONCENTRATION D'ENZYME SUR LA PRODUCTION DU LACTOSUCROSE .............................................. 69
FIGURE 28. EFFET DE LA CONCENTRATION D'ENZYME SUR LA PRODUCTION DU TETRAHEXOSE ................................................ 70
FIGURE 29. EFFET DE LA CONCENTRATION D'ENZYME SUR LA BIOCONVERSION DU LACTOSE .................................................. 71
FIGURE 30. LA FORMATION DE GALACTOSE (PRODUIT D'HYDROLYSE DE LACTOSE) AVEC DIFFÉRENTES CONCENTRATIONS D'ENZYME
.................................................................................................................................................................. 73
FIGURE 31. LA FORMATION DE GLUCOSE (PRODUIT D'HYDROLYSE DE LACTOSE AVEC DIFFÉRENTES CONCENTRATIONS D'ENZYME .. 73
FIGURE 32. EFFET DE LA CONCENTRATION INITIALE DE LACTOSE AVEC LE RAPPORT MOLAIRE DES SUBSTRATS 1:1 SUR LA
PRODUCTION DES GOS TOTAUX ....................................................................................................................... 75
FIGURE 33. EFFET DE LA CONCENTRATION INITIALE DE SACCHAROSE AVEC LE RAPPORT MOLAIRE DES SUBSTRATS 2:1 SUR LA
PRODUCTION DES GOS TOTAUX ....................................................................................................................... 75
FIGURE 34. EFFET DE LA CONCENTRATION INITIALE DE LACTOSE AVEC LE RAPPORT MOLAIRE DES SUBSTRATS 1:1 SUR LA
PRODUCTION DES GOS TOTAUX ....................................................................................................................... 76
FIGURE 35. EFFET DE LA CONCENTRATION INITIALE DE LACTOSE AVEC LE RAPPORT MOLAIRE DES SUBSTRATS 2:1 SUR LA
PRODUCTION DES GOS TOTAUX ....................................................................................................................... 76
FIGURE 36. EFFET DE LA CONCENTRATION DES REACTIFS INITIAUX SUR LE RENDEMENT DU LACTOSUCROSE ............................... 78
FIGURE 37. EFFET DE LA CONCENTRATION DES REACTIFS INITIAUX SUR LE RENDEMENT DU TETRAHEXOSE ................................. 78
FIGURE 38. EFFET DE LA CONCENTRATION ET LES RAPPORTS MOLAIRES SUR LA FORMATION DU GALACTOSE ............................. 79
FIGURE 39. EFFET DE LA CONCENTRATION ET LES RAPPORTS MOLAIRES SUR LA FORMATION DU GLUCOSE ................................ 80
FIGURE 40. EFFET DU PH SUR LA CONVERSION DE LACTOSE ............................................................................................ 81
FIGURE 41. EFFET DU PH SUR LE RENDEMENT EN GOS TOTAUX ...................................................................................... 82
FIGURE 42. EFFET DE LA TEMPERATURE SUR LA CONVERSION DU LACTOSE ......................................................................... 83
FIGURE 43. EFFET DE LA TEMPERATURE SUR LA FORMATION DES GOS ............................................................................. 84
FIGURE 44. EFFET DE LA TEMPERATURE SUR LE RENDEMENT EN LACTOSUCROSE ................................................................. 84
FIGURE 45. EFFET DE LA TEMPERATURE SUR LE RENDEMENT EN TETRAHEXOSE ................................................................... 85
FIGURE 46. LES ISOTHERMES D'ADSORPTION/DESORPTION D'AZOTE POUR LA SBA-15. LA FIGURE INSEREE REPRESENTE LA
DISTRIBUTION DE LA TAILLE DES PORES. .............................................................................................................. 88
FIGURE 47. LE DIFFRACTOGRAMME (DRX) OBTENU AUX PETITS ANGLES OBTENUS POUR LA SBA-15 ..................................... 89
FIGURE 48. MICROGRAPHES MET DE LA SBA-15 OBSERVE DE DIFFERENTS ANGLES............................................................ 90
FIGURE 49. LES ISOTHERMES D'ADSORPTION/DESORPTION D'AZOTE POUR LA MCF. LA FIGURE INSEREE REPRESENTE LA
DISTRIBUTION DE LA TAILLE DES PORES ............................................................................................................... 91
FIGURE 50. LE DIFFRACTOGRAMME (DRX) OBTENU AUX PETITS ANGLES OBTENUS POUR LA MCF ......................................... 92
FIGURE 51. MICROGRAPHE D’ELECTRON DE TRANSMISSION DE LA MCF (20 NM) .............................................................. 93
FIGURE 52. LES ISOTHERMES D'ADSORPTION/DESORPTION D'AZOTE POUR LA S-2.9. LA FIGURE INSEREE REPRESENTE LA
DISTRIBUTION DE LA TAILLE DES PORES ............................................................................................................... 94

XVII
FIGURE 53. LES ISOTHERMES D'ADSORPTION/DESORPTION D'AZOTE POUR LA S-34,8. LA FIGURE INSEREE REPRESENTE LA
DISTRIBUTION DE LA TAILLE DES PORES ............................................................................................................... 95
FIGURE 54. LE DIFFRACTOGRAMME (DRX) OBTENU AUX PETITS ANGLES POUR S-2.9 .......................................................... 96
FIGURE 55. LE DIFFRACTOGRAMME (DRX) OBTENU AUX PETITS ANGLES POUR S-34.8 ........................................................ 97
FIGURE 56. MICROGRAPHE MET DES SILICES SOUS FORME DE BARRES (S2.9 7.7 NM) ....................................................... 98
FIGURE 57. MICROGRAPHE MET DES SILICES SOUS FORME DES VESICULES (S-34.8 16.7 NM) ............................................. 98
FIGURE 58. LES ISOTHERMES D'ADSORPTION/DESORPTION D'AZOTE POUR LA S-10-100 ..................................................... 99
FIGURE 59. LES ISOTHERMES D'ADSORPTION/DESORPTION D'AZOTE POUR LA S-15-100 ................................................... 100
FIGURE 60. LES ISOTHERMES D'ADSORPTION/DESORPTION D'AZOTE POUR LA S-T-100 ..................................................... 100
FIGURE 61. LE DIFFRACTOGRAMME (DRX) OBTENU AUX PETITS ANGLES OBTENUS POUR LA S-10-100 ................................ 101
FIGURE 62. LE DIFFRACTOGRAMME (DRX) OBTENU AUX PETITS ANGLES OBTENUS POUR LA S-15-100 ................................ 102
FIGURE 63. LE DIFFRACTOGRAMME (DRX) OBTENU AUX PETITS ANGLES OBTENUS POUR LA S-T-100 .................................. 103
FIGURE 64. MICROGRAPHE MET DE LA S-10-100(24 NM) ........................................................................................ 104
FIGURE 65. MICROGRAPHE MET DE LA S-15-100(19-20 NM) ................................................................................... 104
FIGURE 66. MICROGRAPHE MET DE LA S-T-100(7.5 NM) ......................................................................................... 104
FIGURE 67. EFFET DU CHANGEMENT DE PH SUR LA RETENTION DES MATERIAUX ............................................................... 106
FIGURE 68. DISTRIBUTION DE RETENTION PRESENTEE PAR LA PROCEDURE GLM DE SAS ..................................................... 107
FIGURE 69. EFFET DE LA QUANTITE D’ENZYME SUR L’IMMOBILISATION ET LA RETENTION DES MATERIAUX .............................. 109
FIGURE 70. EFFET DE LA CONCENTRATION DE GA SUR L’IMMOBILISATION ET LA RETENTION DES ENZYMES PAR LES DIFFERENTS
MATERIAUX ................................................................................................................................................ 114
FIGURE 71. CINETIQUE DE LA REACTION DE FORMATION DES GOS CATALYSEE PAR Β-GALACTOSIDASE IMMOBILISEE SUR LA MCF
COMME SUPPORT A UNE CONCENTRATION ENZYMATIQUE DE 100% ...................................................................... 117
FIGURE 72. COMPARAISON ENTRE LES PRODUITS DE LA REACTION AVEC ENZYMES IMMOBILISEES SUR LA MCF ET LES PRODUITS DE
LA REACTION AVEC DES ENZYMES EN SOLUTION A UNE CONCENTRATION ENZYMATIQUE DE 60% ................................. 118
FIGURE 73. CINETIQUE DE LA REACTION DE FORMATION DES GOS CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA SBA-15 A
UNE CONCENTRATION D’ENZYME DE 100% ...................................................................................................... 119
FIGURE 74. CINETIQUE DES PRODUITS FORMES AU COURS DE LA REACTION CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA
SBA-15 A UNE CONCENTRATION D’ENZYME DE 100% ....................................................................................... 119
FIGURE 75. COMPARAISON ENTRE LES PRODUITS DE LA REACTION AVEC ENZYMES IMMOBILISEES SUR LA SBA-15 ET LES PRODUITS
DE LA REACTION AVEC DES ENZYMES EN SOLUTION A UNE CONCENTRATION ENZYMATIQUE DE 60% ............................. 120
FIGURE 76. CINETIQUE DE LA REACTION DE FORMATION DES GOS CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA ‘S 2,9’ A
UNE CONCENTRATION D’ENZYME DE 100% ...................................................................................................... 121
FIGURE 77. CINETIQUE DES PRODUITS FORMES AU COURS DE LA REACTION CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA
‘S 2,9’ A UNE CONCENTRATION D’ENZYME DE 100% ......................................................................................... 122

XVIII
FIGURE 78. COMPARAISON ENTRE LES PRODUITS DE LA REACTION AVEC ENZYMES IMMOBILISEES SUR LA ‘S 2,9’ ET LES PRODUITS
DE LA REACTION AVEC DES ENZYMES EN SOLUTION A UNE CONCENTRATION ENZYMATIQUE DE 60% ............................. 122
FIGURE 79. CINETIQUE DE LA REACTION DE FORMATION DES GOS CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA ‘S 34,8’ A
UNE CONCENTRATION D’ENZYME DE 100% ...................................................................................................... 124
FIGURE 80. CINETIQUE DES PRODUITS FORMES AU COURS DE LA REACTION CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA
‘S 34,8’ A UNE CONCENTRATION D’ENZYME DE 100% ....................................................................................... 124
FIGURE 81. COMPARAISON ENTRE LES PRODUITS DE LA REACTION AVEC ENZYMES IMMOBILISEES SUR LA ‘S 34,8’ ET LES PRODUITS
DE LA REACTION AVEC DES ENZYMES EN SOLUTION A UNE CONCENTRATION ENZYMATIQUE DE 60% ............................. 125
FIGURE 82. CINETIQUE DE LA REACTION DE FORMATION DES GOS CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA ‘S-10-
100’ A UNE CONCENTRATION D’ENZYME DE 100% ............................................................................................ 126
FIGURE 83. CINETIQUE DES PRODUITS FORMES AU COURS DE LA REACTION CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA
‘S-10-100’ A UNE CONCENTRATION D’ENZYME DE 100% ................................................................................... 127
FIGURE 84. COMPARAISON ENTRE LES PRODUITS DE LA REACTION AVEC ENZYMES IMMOBILISEES SUR LA ‘S-10-100’ ET LES
PRODUITS DE LA REACTION AVEC DES ENZYMES EN SOLUTION A UNE CONCENTRATION ENZYMATIQUE DE 60% ............... 127
FIGURE 85. CINETIQUE DE LA REACTION DE FORMATION DES GOS CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA ‘S-15-
100’ A UNE CONCENTRATION D’ENZYME DE 100% ............................................................................................ 128
FIGURE 86. CINETIQUE DES PRODUITS FORMES AU COURS DE LA REACTION CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA
‘S-15-100’ A UNE CONCENTRATION D’ENZYME DE 100% ................................................................................... 129
FIGURE 87. COMPARAISON ENTRE LES PRODUITS DE LA REACTION AVEC ENZYMES IMMOBILISEES SUR LA ‘S-15-100’ ET LES
PRODUITS DE LA REACTION AVEC DES ENZYMES EN SOLUTION A UNE CONCENTRATION ENZYMATIQUE DE 60% ............... 129
FIGURE 88. CINETIQUE DE LA REACTION DE FORMATION DES GOS CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA ‘S-T-
100’ A UNE CONCENTRATION D’ENZYME DE 100% ............................................................................................ 131
FIGURE 89. CINETIQUE DES PRODUITS FORMES AU COURS DE LA REACTION CATALYSEE PAR Β-GALACTOSIDASE SUPPORTEE SUR LA
‘S-T-100’ A UNE CONCENTRATION D’ENZYME DE 100% ..................................................................................... 131
FIGURE 90. COMPARAISON ENTRE LES PRODUITS DE LA REACTION AVEC ENZYMES IMMOBILISEES SUR LA ‘S-T-100’ ET LES PRODUITS
DE LA REACTION AVEC DES ENZYMES EN SOLUTION A UNE CONCENTRATION ENZYMATIQUE DE 60% ............................. 132
FIGURE 91. CONVERSION DE LACTOSE EN FONCTION DU RENDEMENT EN LACTOSUCROSE DE DIFFERENTS MATERIAUX ............. 135
FIGURE 92. CONVERSION DE LACTOSE EN FONCTION DE RENDEMENT EN TETRAHEXOSE DE DIFFERENTS MATERIAUX ................. 136
FIGURE 93. LA PRODUCTION EN LACTOSUCROSE ET EN TETRAHEXOSE PAR Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE IMMOBILISEE
SUR LA SBA-15 POUR 5 CYCLES SUCCESSIFS ...................................................................................................... 138
FIGURE 94. LA PRODUCTION EN LACTOSUCROSE ET EN TETRAHEXOSE PAR Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE IMMOBILISEE
SUR LA MCF POUR 5 CYCLES SUCCESSIFS .......................................................................................................... 138
FIGURE 95. LA PRODUCTION EN LACTOSUCROSE ET EN TETRAHEXOSE PAR Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE IMMOBILISEE
SUR LA S-34.8 POUR 5 CYCLES SUCCESSIFS ....................................................................................................... 139

XIX
FIGURE 96. LA PRODUCTION EN LACTOSUCROSE ET EN TETRAHEXOSE PAR Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE IMMOBILISEE
SUR LA S-10-100 POUR 5 CYCLES SUCCESSIFS ................................................................................................... 139
FIGURE 97. LA PRODUCTION EN LACTOSUCROSE ET EN TETRAHEXOSE PAR Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE IMMOBILISEE
SUR LA S-15-100 POUR 5 CYCLES SUCCESSIFS ................................................................................................... 140
FIGURE 98. LA PRODUCTION EN LACTOSUCROSE ET EN TETRAHEXOSE PAR Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE IMMOBILISEE
SUR LA S-2.9 POUR 5 CYCLES SUCCESSIFS ......................................................................................................... 140
FIGURE 99. LA PRODUCTION EN LACTOSUCROSE ET EN TETRAHEXOSE PAR Β-GALACTOSIDASE D’ASPERGILLUS ORYZAE IMMOBILISEE
SUR LA S-T-100 POUR 5 CYCLES SUCCESSIFS ..................................................................................................... 141


XXI
Liste des abréviations et des sigles
A.oryzae : Aspergillus oryzae.
BCA: Acide bicinchoninique.
BSA : Sérum albumine bovine.
DRX : Diffraction des rayons X.
Ez : Enzyme.
FOS : Fructooligosaccharide.
FOSHU : Food for Specified Health Uses.
Fru : Fructose.
GA: Glutaraldéhyde.
Gal : Galactose.
Glc : Glucose.
GOS : Galactooligosaccharide.
HCl : Acide chlorhydrique.
HMMS : Hierarchically-ordered mesocellular mesoporous silica
HPLC : Chromatographie phase liquide à haute performance.
I : Intensité.
KCl : Chlorure de potassium.
m/v (%) : Pourcentage massique par rapport au volume.
MET : Microscopie électronique à transmission.
mL : Millilitre.

XXII
MPS : Mesoporous silica « Silice mésopoureuse ».
MS : Spectrométrie de masse.
ONPG : Orthonitrophényl-β-galactoside.
P123 : Acide pluronique (agent structurant, EO20PO70EO20).
rpm : « Rotation per minute » tour par minute.
T : Température.
t : Tonnes.
TEOS : Tétraéthylorthosilicate (précurseur de silice).
TIPB : 1,3,5-triisopropylbenzène.
TMB : 1,3,5-triméthylbenzène.
U : Unité d’enzyme.
u.a. : Unité arbitraire.
UV : Ultra- violet.

XXIII
Dédicace
A mes très chers parents, qui ont toujours été les
meilleurs au monde
A mon époux qui ne cesse de m’encourager
A mon frère et mes deux petites sœurs, qui sont les
plus adorables au monde
A tous mes vrais amis …
A toutes les mains qui m’ont été tendues


XXV
Remerciements
Ce travail a été réalisé à l’Université Laval au laboratoire du Professeur Khaled
Belkacemi, mon directeur de recherche, auprès de qui j'ai énormément appris, à qui je
tiens particulièrement à exprimer toute ma reconnaissance et mes remerciements pour
le soutien et la confiance qu’il m’a accordés durant cette période, et pour m'avoir
accueilli au sein de son laboratoire. Ses précieux conseils m’ont permis de mener à
terme ce projet. Je le remercie énormément pour le temps et l’intérêt qu’il m’a
consacrés.
Je tiens à exprimer ma profonde gratitude au Professeure Safia Hamoudi pour ses
conseils judicieux, sa bonne humeur, son aide indispensable et appréciable.
Je remercie profondément mes collègues de laboratoire Abdelnasser Abidli et Cherif
Larabi pour leurs sympathies, pour l’esprit de travailler en équipe qu’ils m’ont fait
partager, pour leurs disponibilités exceptionnelles et leurs précieux conseils. Ils ont su
faire créer une bonne ambiance durant cette période et m’ont soutenu sans conditions et
n’ont pas hésité à m’aider dans mon travail.
Je voudrais remercier chaleureusement toutes les nombreuses personnes que j’ai
rencontrées durant ces deux ans et qui m’ont aidé lors de la réalisation de ce travail. Je
mentionne spécialement Ahmed Amara et Atef Sahli pour leur disponibilité et leurs
encouragements, Pascal Dubè pour son aide indispensable.
Je remercie tous mes amis qui étaient là et qui m’entouraient dans toutes les situations.
Merci chers amis pour votre amitié si précieuse.
Pour terminer, je tiens plus que tout à adresser mon affection et ma gratitude à ma chère
maman Souad, mon cher papa Habib, mon grand frère Mohamed, mes deux anges

XXVI
Asma & Mariem, et mon cher époux Koutheir pour leur amour et leur soutien et
encouragements tout au long de mes études de maîtrise. Sans leur amour, je ne serais
pas en mesure de faire face aux problèmes même minimes rencontrés au cours de mes
études.

XXVII
Avant-propos
Le présent travail a été réalisé dans le cadre de mon projet de maîtrise. Ce projet portait
sur la synthèse d’un nouveau biocatalyseur à base de β-galactosidase à partir
d’Aspergillus oryzae immobilisée sur différents matériaux siliceux mésoporeux, dans le
but de synthétiser le lactosucrose obtenu par la bioconversion du lactose et du
saccharose. L’immobilisation est effectuée après un processus d’optimisation de
différentes conditions opératoires de la réaction de transglycosylation. Cette
immobilisation permet l’amélioration de la stabilité et la performance de l’enzyme. Ce
projet s’inscrit dans le cadre d’une thématique de recherche d’actualité et d’une pratique
de la chimie verte, visant l’utilisation de l’approche hétérogène pour la biosynthèse des
galacto-oligosaccharides bénéfiques en excluant l’usage des agents chimiques.
Le présent document comporte cinq chapitres. Le chapitre 1, l'introduction, présente la
problématique générale du mémoire. Au chapitre 2, la revue de littérature résume l’état
actuel des connaissances dans la production des oligosaccharides à intérêts bénéfiques
pour la santé humaine, plus précisément le lactosucrose, ainsi qu’une section décrivant
l’enzyme utilisée (β-galactosidase à partir d’Aspergillus oryze) et ses applications ainsi
que les différentes voies d’immobilisation, tout en abordant les principaux aspects
théoriques et pratiques de cette technique. La revue de littérature est suivie d’un 3e
chapitre qui présente respectivement et brièvement, l’hypothèse et les objectifs du
travail. Le chapitre 4 porte sur la présentation du matériel et des méthodes
expérimentales utilisés ainsi que les techniques de caractérisation et d’identification et
les analyses statistiques des données. Les principaux résultats et la discussion sont
présentés au 5 e chapitre.
Les travaux réalisés et les résultats obtenus dans ce chapitre (partie homogène) portant
sur la synthèse du lactosucrose utilisant la β-Galactosidase libre ont fait l’objet d’un
article rédigé. Ce premier article sera soumis à la revue Food Chemistry, sous le titre:
’’Optimization of lactose and sucrose bioconversion using Aspergillus oryzae β-
Galactosidase for the production of lactosucrose’’. Zeineb Ben Rejeb est premier auteur
de cet article.
Un deuxième article détaillant la catalyse hétérogène sera soumis pour publication à la
revue Bioresource Technology, sous le titre : ‘’Biosynthesis of lactosucrose using a novel

XXVIII
heterogeneous biocatalyst based on Aspergillus oryzae β-Galactosidase immobilized on
different silica-based materials for lactose and sucrose conversion’’. Zeineb Ben Rejeb
sera comme premier auteur de cet article.
Ces résultats ont fait également l'objet d'une participation à une séance de posters dans
le cadre de la 5e conférence annuelle du Centre en Chimie Verte et Catalyse CCVC qui
s'est tenue à l’Université McGill, Montréal le 19 mai 2014.
La conclusion et les perspectives sont présentées à la fin en résumant les principaux
résultats obtenus dans le cadre de ce projet. Afin d’atteindre les objectifs visés par cette
étude, j’ai réalisé une revue de littérature et suggéré des protocoles expérimentaux. J’ai
également réalisé l’ensemble des manipulations en laboratoire, traité les données
expérimentales (caractérisation et analyse statistique des données), interprété les
résultats et rédigé ce présent mémoire. Le Professeur Khaled Belkacemi, mon directeur
de recherche, était responsable de la supervision des travaux. Il a contribué à la
planification des expériences et à la discussion des résultats de même que la révision du
manuscrit final.

1
Chapitre 1. Introduction générale
Le lactosérum est un sous-produit de l’industrie fromagère, produit en grande quantité
dans le monde entier et cette production ne cesse d’augmenter. Ce sous-produit
contient environ 20% des protéines du lait et la majorité des sucres du lait (Lactose)
(Smithers et al., 1996). Malgré l’existence d’approches utilisées pour bénéficier d’une
valeur ajoutée au lactosérum, une grande quantité du ce sous-produit est toujours
rejetée (Panesar et Kennedy, 2012). Ceci explique le grand intérêt pour les procédés
innovants, afin de transformer le lactose et explorer ses applications dans les industries
pharmaceutiques et alimentaires. En effet, les oligosaccharides comme le lactulose, le
lactitol, l'acide lactobionique, le lactosucrose, et le galacto-oligosaccharide (GOS)
dérivent du lactose et sont des produits à haute valeur ajoutée, bénéfiques pour la santé
humaine (Gänzle et al., 2008).
Les oligosaccharides et leurs dérivés sont des biomolécules importantes, ayant une
large gamme de fonctions dans les systèmes biologiques. Ils ont des propriétés
prébiotiques qui aident à : (i) l’amélioration de l’équilibre de la flore intestinale ; (ii)
l’amélioration de l'absorption du calcium alimentaire ; (iii) la réduction du taux de
cholestérol sérique et (vi) la synthèse de vitamines B (Aider et De Halleux, 2007, Honda
K1, 1999, Sako et al., 1999).
Les oligosaccharides sont bénéfiques pour la santé, mais leur synthèse par voie
chimique est fastidieuse. Cette synthèse nuit à l’environnement et à la santé humaine à
cause de résidus des solvants persistant. Leur synthèse passe inexorablement par
lusieurs étapes de réactions chimiques et aussi de purification qui sont souvent
onéreuses. Ainsi, d’autres alternatives biologiques sont recherchées, telles que
l’utilisation des enzymes (Emaga et al., 2012, Zhang Ning et al., 2001). En effet, les
oligosaccharides peuvent être synthétisés par voie enzymatique à partir des
disaccharides. La source de l'enzyme et les conditions de la réaction contrôlent le type
et la quantité d'oligosaccharides produits (Crittenden et Playne, 1996). L'utilisation de
l’enzyme β-galactosidase en tant que catalyseur est intéressante dans la réalisation de
telles réactions. où une multitude de produits à haute valeur sont synthétisés, tels que
les galactooligosaccharides (GOS) (Albayrak et Yang, 2002, Gaur et al., 2006, Urrutia et
al., 2013), le lactulose (Vera et al., 2011) et le lactosucrose (Li et al., 2009).

2
Généralement, les β–galactosidases sont utilisées pour l’hydrolyse du lactose et
peuvent également catalyser la formation des galacto-oligosaccharides (GOS) en
présence de lactose comme substrat. La littérature (Li et al., 2009) montre que la
présence de saccharose avec le lactose comme substrats offre la possibilité de
synthèse de lactosucrose, un prébiotique important (Li et al., 2009).
Plusieurs sources microbiennes de la β-galactosidase ont été étudiées pour des
applications alimentaires telles que Aspergillus Niger, Aspergillus oryzae,
Kluyveromyces lactis et Kluyveromyces fragilis (Richmond et al., 1981). La β-
galactosidase à partir de A. oryzae a été appliquée à la biosynthèse de différents
produits tels que les GOS (Albayrak et Yang, 2002), le lactulose (Guerrero et al., 2011),
et les galactosyl-polyhydroxyalcohols (Irazoqui et al., 2009). Elle était utilisée sous la
forme libre ainsi qu’immobilisée. La β-galactosidase à partir de A. oryzae a été
immobilisée par différentes stratégies : (i) encapsulation dans l'alginate; (ii) fixation
covalente sur différents supports, ou (iii) adsorption ionique combinée à la réticulation
(Urrutia et al., 2013).
L’immobilisation est une technique largement utilisée dans les applications industrielles
des enzymes pour améliorer la stabilité, la sélectivité en vers les produits désirés,
l’isolation des produits dans un système biphasique, et la réutilisation des catalyseurs
(régénération et recyclage).
Dans ce travail, il est question de synthétiser le lactosucrose par voie enzymatique,
selon deux approches; homogène et hétérogène. L’immobilisation de β-galactosidase (à
partir de A. oryzae) s’effectuera sur différents matériaux siliceux mésoporeux pour
étudier et comparer la rétention et le rendement de la biosynthèse du lactosucrose afin
de choisir la combinaison matériau/enzyme la plus efficace.

3
Chapitre 2. Revue de littérature
2.1. Lactosérum
2.1.1. Définition et composition
Le Lactosérum est le liquide jaune-vert qui reste après la précipitation et l'élimination de
la caséine du lait au cours de la fabrication du fromage (Figure 1). Il est considéré
comme un sous-produit de l’industrie laitière (Smithers et al., 1996).
Il existe principalement deux variétés de lactosérum, selon l’approche utilisée pour la
précipitation des caséines:
a. Le lactosérum acide (pH<5) : résultant de la production de fromages frais ou
doux (comme le fromage crémeux et le fromage cottage) suite à l’action des
bactéries lactiques.
b. Le lactosérum doux (pH 6-7) : résultant de la production de fromages à pâte dure
suite à la précipitation de la caséine par la présure (fromages à pâte pressée,
semi-pressée, ou molle).
La composition des différents types de lactosérum est variable (Guimarães et al., 2010,
Mawson, 1994).

4
Figure 1. Caillés (coagulation de la caséine) et lactosérum.
Tableau 1. Composition du lait bovin et du lactosérum (Smithers et al., 1996)
Composition Concentration
Lait Lactosérum
(%, wt/vol)
Caséine 2,8 0,0
Protéines solubles 0,7 0,7
Lipides 3,7 0,05
Cendre 0,7 0,7
Lactose 4,9 4,9
Total 12,8 6,35
Le lactosérum représente environ 85-95% du volume du lait et conserve 55% des
nutriments du lait. Ce sous-produit contient essentiellement 100% de glucides totaux
(lactose), et environ 20% des protéines totales du lait (Tableau 1). Le lactosérum
contient également des quantités appréciables d'autres composants tels que les lipides
(0,4-0,5% m/v), les sels minéraux (8-10% d’extrait sec), l’acide lactique (0,05% m/v),
l’acide citrique, ainsi que des composés azotés non protéiques (Guimarães et al., 2010,
Mawson, 1994).

5
Le lactosérum est produit de plus en plus vu l’augmentation mondiale de production du
fromage. En général, pour faire 1 kg de fromage, 9 L de lactosérum sont générés
(Kosikowski, 1979). En effet, plus de 20 millions de tonnes de fromages sont produits
chaque année, soit près de 655 kg/s avec un taux d’augmentation de 2% chaque année.
Comme l’illustre la Figure 2, les États-Unis sont les plus grands producteurs de
fromage, avec 24% de la production mondiale totale en 2005, alors que le Canada est
situé en neuvième position (McDonald, Novembre 2006). La production du lactosérum
continue de croître à travers le monde et dépasse actuellement les 80 x l09 L/J
(Smithers et al., 1996).
De nos jours, trouver des voies de valorisation prometteuses pour le lactosérum est
devenu un ‘leitmotiv’ au niveau mondial.
.
Figure 2. Principaux producteurs de fromage dans le monde en 2005 (exprimés en milliers de tonnes).
2.1.2. Utilisation et valorisation du lactosérum
Les différents modes d'élimination et d’utilisation du lactosérum ont présenté un défi
pour l'homme depuis qu’il a commencé à fabriquer du fromage. Ces modes de
valorisation et d’élimination ont été décrits dans plusieurs publications (Mawson, 1994,
Siso, 1996, Smithers, 2008), et peuvent être illustrés dans trois catégories:
a. Utilisation directe : le lactosérum est utilisé avec peu ou sans traitement ultérieur.
Cela comprend l'utilisation traditionnelle du lactosérum comme ingrédient

6
alimentaire pour les animaux d’élevage et comme engrais pour l'agriculture.
L'inconvénient avec cette utilisation est relié à la trop forte demande en oxygène
du lactosérum (Mawson, 1994).
b. Stabilisation du lactosérum: le lactosérum est traité par des moyens physiques
et/ou chimiques afin de le rendre beaucoup plus stable et résistant à la
dégradation microbienne. Parmi les techniques utilisée, on trouve: (1) la
récupération des protéines par ultrafiltration, et (2) la dénaturation des protéines
par la chaleur, et (3) la concentration par osmose inverse et/ou évaporation
suivie d’une cristallisation du lactose, et (4) la transformation du lactosérum en
poudre. Dans certains de ces procédés, le lactose et les sels du lait retenus dans
le produit sont également manipulés pour étendre les possibilités d’exploitation
de ces produits (Mawson, 1994).. Différentes poudres condensées sont
préparées, telles que (1) la poudre du lactosérum complet, et (2) la poudre du
lactosérum déminéralisé, et (3) la poudre du lactosérum délactosé ou déprotéiné,
et enfin (4) le lactose en poudre (Siso, 1996).
c. Processus de conversion: Le lactose est converti en un autre composé par les
activités de micro-organismes (bioconversion) ou par une réaction chimique. Des
exemples de ces derniers incluent la production du lactulose, du lactitol, du
bioéthanol ainsi que du méthane et la production de levure par fermentation
(Mawson, 1994). Ces procédés de bioconversion sont actuellement exploités à
l'échelle commerciale (Figure 3).

7
Figure 3. L'utilisation commerciale de lactosérum de lait.
2.2. Lactose
2.2.1. Production et composition
Le lactose, ou le sucre de lait, est le glucide majeur (97% des glucides du lait) présent
dans le lait de tous les mammifères, avec quelques exceptions mineures. Sa
concentration approximative est comprise entre 2% et 10%. À titre d’exemple, la teneur
en lactose du lait de vache varie entre 4,4% et 5,2%, avec une moyenne de 4,8%,
comparée à une teneur de 7% dans le lait maternel (Gänzle et al., 2008).
D’autre part, le lactose représente le glucide majoritaire du lait avec la teneur la plus
élevée comparée aux autres constituants du lait maternel et de vache standard (lipides,
protéines et minéraux). Ceci explique son importance en nutrition (Seki et Saito, 2012).

8
Le Tableau 2 illustre la composition moyenne du lait maternel et du lait de vache (Vilain,
2010).
Tableau 2. Composition moyenne du lait selon l’espèce (g/L)
Eau Lipides Protéines
Totales
Glucides
(Lactose)
Matières
minérales
Lait
maternel 905 35 12 – 14 65 - 70 3
Lait de
vache 900 35 – 40 30 -35 45 - 50 8 – 10
Le lactose (β-D-galactopyrannosyl(1→4)β-D-glucopyrannose, C12H22O11, 342,3 g mol−1)
est un disaccharide composé d'une molécule de β-D-galactose (Gal) et d'une molécule
de α/β-D-glucose (Glc) reliées entre elles par une liaison osidique β(1→4) (Gänzle et al.,
2008).
La structure de la molécule de lactose est représentée par la Figure 4. Structure
chimique du lactose. Dans une solution aqueuse, le lactose est présent sous deux
formes (Figure 5) avec des proportions à l’équilibre de 62,7 % comme β-lactose et 37,3
% comme α-lactose. Cet d'équilibre est légèrement affecté par des différences de
température, mais pas par les différences de pH (Gänzle et al., 2008).

9
Figure 4. Structure chimique du lactose.
Figure 5. Structure des molécules de lactose en configuration α et β (Gänzle et al., 2008).

10
La majeure partie de production du lactose, fabriquée à l’échelle industrielle, est
produite à partir de lactosérum (qui est le sous-produit de la production fromagère) suite
à des procédés de purification et de cristallisation (Seki et Saito, 2012).
La production mondiale de lactosérum a été estimée à 137,9 millions de tonnes en 1998
représentant près de 5,5 millions de tonnes de lactose par an. Au Canada, la production
du fromage en 2004 a été estimée à 0,34 million de tonnes, ce qui implique que plus de
0,25 million de tonnes de lactosérum ont été produites cette année (Aider et De Halleux,
2007). Ceci confirme l’abondance de la matière première pour la production de lactose
et de ses dérivés qui prédominent l’intérêt industriel vu leurs qualités et bienfaits
demandés, en plus de la disponibilité de la matière première à bas prix.
2.2.2. Applications du lactose
Les applications du lactose sont de plus en plus nombreuses et variées dans le domaine
alimentaire et pharmaceutique. Le lactose est utilisé largement dans les confiseries, le
pain, les aliments pour animaux et les aliments pour bébé. Il est encore utilisé comme le
glucide le plus adapté à la préparation de nombreux milieux de fermentation. Les
industries pharmaceutiques utilisent le lactose depuis des années comme agent vecteur
de médicament. En outre, le lactose est utilisé comme matière première pour la
fabrication de plusieurs produits (les dérivés de lactose), tels que les oligosaccharides,
les sucres rares et les alcools (Seki et Saito, 2012). Les principales voies de
transformation de lactose sont illustrées dans la Figure 6.
La transformation de lactose suit différentes voies telles que : l’hydrolyse, la
déshydratation, l’isomérisation, la polymérisation, l’hydrogénation, l’oxydation, la
réduction, la fermentation et la voie enzymatique. En effet, le tagatose, un sucre rare, est
un monosaccharide préparé par isomérisation de galactose qui est lui-même produit par
l'hydrolyse du lactose. On le trouve en petites quantités dans les produits laitiers. Bien
que le degré de douceur de tagatose soit d'environ 90% de celui du saccharose, son
pouvoir calorifique est faible. Le lactulose est un disaccharide comprenant le galactose
et le fructose. Il est le produit de la réaction d’isomérisation de lactose dans un milieu
alcalin comme par exemple l’hydroxyde de calcium. Le lactulose a été ajouté aux
préparations pour nourrissons comme un facteur bifidus depuis de nombreuses années.
Le lactosucrose qui est un trisaccharide comprenant le galactose, le glucose et le
fructose est synthétisé par voie enzymatique. L’epilactose est un disaccharide

11
comprenant le galactose et le mannose est préparé par épimérisation du lactose (Aider
et De Halleux, 2007, Seki et Saito, 2012).
Figure 6. Voies de transformation de lactose: les dérivés de lactose (Seki et Saito, 2012).
L'acide lactobionique qui est un acide aldonique comportant du galactose et de l'acide
gluconique. Il est de plus en plus utilisé comme facteur bifidus. Il est généralement
fabriqué par oxydation du groupe aldéhyde du fragment glucose du lactose, mais ces
dernières années une nouvelle méthode de synthèse a été décrite à l'aide d'une enzyme
à partir des cellules Burkholderia cepacia. D’autre part, le lactose peut être une matière
première pour les alcools de sucre tel que le lactitol préparé par hydrogénation
catalytique dans des conditions de haute température et haute pression (Seki et Saito,
2012).
Lactose
Hydrolyse
Fermentation
Polymérisation enzymatique (avec saccharose)
Polymérisation
enzymatique
Isomérisation
Epimérisation
Oxydation
Réduction
Fermentation
Glucose+Galacto
se
Isomérisation Tagatose
Lactulose
Galacto-oligosaccharide
Lactosucrose
Epilactose
Acide Lactobionique
Lactitol
Éthanol
Acide
lactique
Déshydratation
Lactide
Polymérisation
Résine
polylactique

12
2.3. Oligosaccharides et aliments fonctionnels
2.3.1. Oligosaccharide : Définition et propriété
Les oligosaccharides, définis comme étant des oligomères de différents degrés de
polymérisation, peuvent êtres synthétisés aussi bien par voie enzymatique que par voie
chimique. Ils sont solubles dans l’eau et ont un pouvoir sucrant relativement faible (de
l’ordre de 0,3-0,6 fois celui du sucrose), qui dépend de leur structure chimique et de leur
masse moléculaire (Sako et al., 1999). Pour ces raisons, ils sont communément utilisés
comme des substituants au sucre à faible cariogènes dans la confiserie, la confiture, la
pâtisserie, le chewing-gum, le yaourt, les boissons ainsi que dans le régime alimentaire
faible en calories et les aliments pour diabétiques. Outre le secteur alimentaire, d'autres
domaines, tels que l’industrie pharmaceutique et cosmétique, peuvent également
exploiter les propriétés physico-chimiques et physiologiques des oligosaccharides
(Torres et al., 2010). Parmi ces oligosaccharides, on trouve le lactulose, le reffinose, les
maltooligosaccharides, l’inulin, les fructooligosaccharides (FOS) et les
galactooligosaccharides (GOS). Les FOS et les GOS sont généralement produits par
transglycosylation enzymatique, en présence de fructanotransferase et β-galactosidase,
respectivement (Figure 7) (Sako et al., 1999).
« Galactooligosaccharide » est le terme générique pour les oligosaccharides produits à
partir du lactose avec β-galactosidase. Au Japon, les GOS ont été approuvés comme
ingrédients alimentaires FOSHU (Food for Specified Health Uses) et ils sont également
utilisés comme prébiotiques ainsi que dans la préparation des produits pharmaceutiques
(Seki et Saito, 2012). Les GOS peuvent sont considérés comme des oligosaccharides
non digestibles. Ils sont fermentescibles par les bactéries dans le gros intestin (Champ
et al., 2003). Ceci est dû à la spécificité du substrat des enzymes digestives gastro-
intestinales humaines qui sont pour la plupart spécifiques aux liaisons α-glycosidique,
alors que les liaisons glycosidiques des GOS ont une configuration β. Certaines β-
galactosidases, localisées dans l’intestin grêle, sont capables de digérer les GOS, mais
leur activité reste faible dans la plupart du temps (Ito et Kimura, 1993, Qiang et al.,
2009).

13
Les galactooligosaccharides sont particulièrement adaptés dans des produits
alimentaires spécifiques à des catégories bien particulières, tels que la nutrition infantile,
la nutrition clinique et les aliments pour les personnes âgées. Certaine frange de la
population peut tirer un avantage supplémentaire de la consommation de
galactooligosaccharides. Les nourrissons par exemple, peuvent bénéficier de l'effet
bifidogène des GOS car leur microflore contient une faible quantité de bifidobactéries
(Sako et al., 1999). La consommation des GOS peut également être bénéfique pour les
personnes âgées, car ils ont un nombre inferieur de bifidobactéries dans leur microflore
comparé à celui des jeunes et des adultes. Ils souffrent souvent de la constipation et leur
capacité à absorber le calcium est inférieure à celle des jeunes adultes (Zhang Ning et
al., 2001).
Bettrave
Lait de vache
Amidon
Saccharose
Lactose
Amidon soluble
Lactosérum de soya
Xylan
Chitin
Raffinose
Fructo-oligosaccharides
Lactosucrose
Lactulose
Galacto-oligosaccharides
Glycosylsucrose
Oligosaccharides de soya
Malto-oligosaccharides
Isomalto-oligosaccharides
Soya
Xylo-oligosaccharides
Chitin oligosaccharides
Extraction
Transglycosylation
Isomérisation
Hydrolyse
Hydrolyse transglycosylation
Figure 7. Représentation schématique du processus de production d'oligosaccharides (Sako et al., 1999)

14
En général, les GOS peuvent être utilisés comme des :
additifs alimentaires dans les boissons, les aliments congelés, les biscuits, etc.
compléments alimentaires : dans les préparations pour nourrisson, par exemple.
édulcorants: ceci est particulièrement utile pour les diabétiques.
2.3.2. Aliments fonctionnels: définition et propriétés
Un aliment fonctionnel, de façon générale, est tout aliment ou ingrédient qui, en plus de
fournir des effets bénéfiques nutritionnels, peut contribuer à des bénéfices de santé tel
que réduire les risques chroniques des maladies (Marriott, 2000).
Le concept des aliments fonctionnels a été développé au Japon au milieu des années
1980 et a suscité un grand intérêt chez les grandes entreprises alimentaires dans les
quatre coins du monde. Le marché mondial des aliments fonctionnels a augmenté de 10
milliards de dollars américain en 1995, pour atteindre 33 milliards de dollars en l’an 2000
Ce marché a dépassé les 50 milliards en 2010 (Menrad, 2003). Une étude européenne a
estimé que le marché de l’alimentation fonctionnelle valait à peu près 1 milliard d’euros
en 1997 et se base essentiellement sur les produits laitiers.
Traditionnellement, les fruits et les légumes sont considérés comme la source principale
des aliments fonctionnels, mais de récentes recherches ont établi que des aliments
d’origine animale tels que le lait et les produits laitiers peuvent aussi être d’excellentes
sources pour ces composants (Bauman et al., 2006). Prenons le cas du lait fermenté, il
contient des oligosaccharides ainsi que des probiotiques qui améliorent la qualité de la
microflore intestinale humaine. De ce fait, les propriétés fonctionnelles du lait fermenté
sont liées à la fois à ses propriétés probiotiques et à la présence des oligosaccharides
(Martínez-Villaluenga et al., 2008).

15
2.4. Le lactosucrose
2.4.1. Structure
Le lactosucrose, connu comme 4G-β-D-galactosylsucrose, est un trisaccharide composé
d’une molécule de galactose, d’une molécule de glucose et d’une molécule de fructose.
Son nom scientifique est β-D-fructofuranosyl-4-O-β-D-galactopyranosyl-α-D-
glucopyranoside et peut être symbolisé par Gal-Glu-Fru. Sa formule chimique est
C18H32O16 et sa masse molaire est de 504,4 g mol-1 (Mu et al., 2013, Seki et Saito, 2012)
(Figure 8).
Figure 8. Structure de lactosucrose.
2.4.2. Caractéristiques et production mondiale
Le pouvoir sucrant du lactosucrose est de 0,3 à 0,6 par rapport au saccharose, avec un
pouvoir cariogène faible et un apport calorique faible(Gänzle, 2012).
Le Japon est le premier pays producteur du lactosucrose commercialisé, ainsi que de
nombreux autres types d'oligosaccharides. Le lactosucrose est utilisé, non seulement
dans beaucoup d’aliments japonais, mais aussi aux États-Unis comme supplément
diététique et fonctionnel (Sharma, 2011, Gänzle, 2012). Il est considéré comme FOSHU
par les japonais depuis 1991 (Crittenden et Playne, 1996). Il possède un effet
probiotique (bifidogène) démontré dans certains essais in-vitro, appliqués sur les

16
animaux et quelques sujets humains (Hussein et al., 1999, Mizote et al., 2009, Teramoto
et al., 2006).
Le lactosucrose est classé parmi les oligosaccharides non digestibles avec d’autres
dérivés de lactose. Il est rapporté comme étant bénéfique pour la santé humaine
(Gänzle, 2012). En effet, le lactosucrose peut stimuler sélectivement la croissance de
Bifidobacterium et il a un effet bénéfique sur les patients atteints de maladies
inflammatoires chroniques intestinales dû à son effet bifidogénique (Fusako et al., 1996,
Ohkusa, 1995, Teramoto et al., 1996). Selon des expériences effectuées sur des rats et
des chiens, le lactosucrose a montré des effets de protection contre l’indométacine
d’ulcère gastrique (Honda, 1999, Terada et al., 1992), ainsi qu’une diminution de la
valeur du pH du son contenu intestinal. Ce dernier changement, accompagné d'autres
facteurs, peuvent augmenter l'absorption du calcium par l'intestin. Ce résultat a été
prouvé chez des femmes jeunes en bonne santé ayant moins d'apport en calcium pour
lesquelles la supplémentation des aliments en lactosucrose à long terme a favorisé
l'absorption intestinale du calcium et a diminué la résorption osseuse (Teramoto et al.,
2006).
Grace aux bienfaits du lactosucrose cités précédemment, sa commercialisation et sa
production mondiale ainsi que celle des autres oligosaccharides non digestibles dérivés
du lactose augmentent de plus en plus dans le monde comme illustré dans le
Tableau 3 (Crittenden et Playne, 1996).

17
Tableau 3. Production des oligosaccharides de qualité alimentaire éstimée en 1995
Classe des
oligosaccharides
Production
estimée en
1995 (t)
principaux fabricants noms
commerciaux
GOS 15000 Yakult Honsha (Japan) Oligomate
Nissin Sugar Manufacturing Company (Japan) Cup-Oligo
Snow Brand Milk Products (Japan) P7L and others
Borculo Whey Products (The Netherlands) TOS-Syrup
Lactulose 20000 Morinaga Milk Industry Co. (Japan) MLS/P/C
Solvay (Germany)
Milei GmbH (Germany)
Canlac Corporation (Canada)
Laevosun (Austria)
lnalco SPA (Italy)
Lactosucrose 1600 Ensuiko Sugar Refining Co. (Japan) Newka-Oligo
Hayashibara Shoji Inc. (Japan) Newka-Oligo
FOS 12000 Meiji Seika Kaisha (Japan) Meioligo
Beghin-Meiji Industries (France) Actilight
Golden Technologies (USA) NutraFlora
Cheil Foods and Chemicals (Korea) Oligo-Sugar
ORAFTI (Belgium) Rafilose and
Raftiline
Cosucra (Belgium) Fibruline
Isomaltulose
oligosaccharides
5000 Mitsui Sugar Co. (Japan) ICP/O
Glucosyl sucrose 4000 Hayashibara Shoji Inc. (Japan) Coupling Sugar
Maltooligosaccharides 10000 Nihon Shokuhin Kako (Japan) Fuji-Oligo
Hayashibara Shoji Inc. (Japan) Tetrup
Isomaltooligosaccharid
es
11000 Showa Sangyo (Japan) lsomalto-900
Hayashibara Shoji Inc. (Japan) Panorup
Nihon Shokuhin Kako (Japan) Biotose and
Panorich
Cyclodextrine 4000 Nihon Shokuhin Kako (Japan) Celdex
Ensuiko Sugar Refining Co. (Japan) Dexy Pearl
Asahi Kasei Kagyo Co. (Japan)
Gentiooligosaccharides 400 Nihon Shokuhin Kako (Japan) Gentiose
Soybean
oligosaccharides
Xylo-oligosaccharides
2000 The Calpis Food Industry Co. (Japan)
Suntory Ltd (Japan)
Soya-oligo
Xylo-oligo

18
2.4.3. Synthèse du lactosucrose
Le lactosucrose est un trisaccharide obtenu suite à une réaction de transfert réversible à
partir du lactose et du saccharose, à l'aide de l'activité des enzymes de
transfructosylation ou transgalactosylation (Schéma 7) (Daude et al., 2012, Kawase et
al., 2001). Dans la première réaction, la fraction fructosyle du saccharose est transférée
au lactose, ainsi le lactosucrose est formé (Gänzle, 2011, Mussatto et Mancilha, 2007). Il
peut être aussi obtenu par la transgalactosylation du saccharose avec la ß-
galactosidase en utilisant le lactose comme donneur de galactosyle et du saccharose
comme accepteur de galactosyle (Gänzle, 2011).

19
Figure 9. Biosynthèse de lactosucrose par A: transfructosylation et B: transgalactosylation (Daude et al., 2012, Li et al., 2009).

20
La synthèse de lactosucrose est obtenue lorsque les réactifs initiaux (lactose et
saccharose) sont présents dans la réaction avec des concentrations équimolaires pour
optimiser le rendement, ainsi que la présence de l’enzyme libre en solution (Mussatto
and Mancilha, 2007).
Le lactosucrose comme les autres oligosaccharides peut être synthétisé par voie
enzymatique ou chimique, mais la méthode enzymatique reste toujours plus
avantageuse; en éliminant et en réduisant l’utilisation des produits toxiques qui peuvent
altérer le produit avec la présence des résidus néfastes pour la santé. La voie
enzymatique nous épargne les traitements et les purifications, en utilisant des solvants
nuisibles à l’environnement. L'utilisation d'enzymes pour la synthèse de sucres
complexes offre plusieurs avantages par rapport à la méthode chimique; les glucides
sont difficiles à manipuler en raison de la présence de nombreux groupements
hydroxyles réactifs. La biosynthèse enzymatique est donc une catalyse très efficace, qui
peut être effectuée sans protection des groupements hydroxyles, avec un large éventail
possible de réactions régiospécifiques et stéréospécifiques. En plus, ces réactions sont
réalisées dans des conditions douces (Hamed et al., 2013).
Il y a trois grandes familles d’enzymes utilisées pour la synthèse de lactosucrose
(Tableau 4) :
a. Les galactosidases et plus précisément les β-galactosidases qui sont
synthétisées par différents microorganismes tels qu’E.Coli, Bacillus circulans,
Aspergillus niger, Bacillus stearothermophilus, Pyrococcus woesei, Arthrobacter
psychrolactophilus (Gänzle, 2011, Li et al., 2009, Oliveira et al., 2011, Sheik-
Asraf et Gunasekaran, 2010).
b. La levansucrases qui est l’enzyme la plus spécifique pour cette réaction dont
ses substrats sont le saccharose et le lactose. La levansucrase est synthétisée
aussi par plusieurs microorganismes (Choi et al., 2004, Gänzle et al., 2008,
Han, 2009).
c. Les β–Fructofuranosidases qui catalysent l'hydrolyse du saccharose pour
donner un mélange de glucose et de fructose, ainsi que la formation des
oligosaccharides comme le lactosucrose dans notre cas (Kawase et al., 2001,

21
Terada et al., 1992). Il est possible aussi d’utiliser les microorganismes
directement pour la production de l’enzyme (Choi et al., 2004, Gänzle, 2011).
Tableau 4. Conditions et rendement en lactosucrose par les différentes espèces
enzymatiques
β-galactosidase β-fructofurannosidase Levansucrase
pH 5 6 7
Température (°C) 40 54 23
Concentration initiale de
substrats (Lactose :
saccharose)
30 : 30 48 : 48 18 :18
Concentration
enzymatique 6 U* 5 U 1 U
Rendement en
lactosucrose (%) 21 4 28,5
Temps de réaction 12h 30 min 12h
*U : Une unité d’enzyme permet d’hydrolyser 1,0 µmole de lactose par minute.
En effet, l'enzyme non seulement catalyse la réaction de transfert mais aussi l'hydrolyse
du saccharose et du lactosucrose ainsi que la formation des produits secondaires
(Figure 10) (Fujita et al., 1990, Mussatto et Mancilha, 2007).
Sucrose + H2O Fructose + Glucose
Lactosucrose + H2O Fructose + Lactose
Figure 10. Réactions d’hydrolyse de saccharose et de lactosucrose (Fujita et al., 1990).

22
Pour cette raison, le rendement maximal de lactosucrose peut être atteint avec un
processus discontinu et une concentration équimolaire de saccharose et de lactose
(Mussatto et Mancilha, 2007).
2.5. β–Galactosidases: Structure, production,
mécanisme de réaction et application
La β-galactosidase, connue aussi comme la lactase (β-D-galactohydrolase, EC 3.2.1.23)
est une enzyme qui hydrolyse des résidus D-galactosyle à partir des oligosaccharides
ou des métabolites secondaires (Husain, 2010). De façon classique, son application
principale est dans l'hydrolyse du lactose et du lactosérum (Oliveira et al., 2011). En plus
de l'hydrolyse, la β-D-galactosidase est capable de catalyser la réaction de
transgalactosylation, conduisant à la formation d'une nouvelle liaison glycosidique entre
deux unités de galactose (Panesar et al., 2010). Les réactions de transgalactosylation
consécutives, conduisent à la formation d'une chaîne de plusieurs unités de galactose,
avec l'unité de glucose terminale (Panesar et al., 2010). Cette enzyme a été classée
parmi les glycosides hydrolases 2 (GH 2) des enzymes actives des glucides (Panesar et
al., 2006).
2.5.1. Structure
A partir de la structure tridimensionnelle (Figure 11) de la β-D-galactosidase
(Kluyveromyces lactis), proposée par Pereira-Rodriguez et al.(Pereira-Rodríguez et al.,
2012), la β-galactosidase est formée d’un tétramère composé de quatre sous-unités
identiques (A B C D), avec 1024 acides aminés.
La taille moléculaire de β-galactosidase dépend de sa source. La plus grande taille
revient à la β-galactosidase obtenue à partir de E. coli lactase (520 à 850 KDa). La β-
galactosidase est aussi obtenue à partir de S. fragilis et A. oryzae avec des poids
moléculaires de 201 et 110 KDa, respectivement (Zhang Ning et al., 2001).

23
Figure 11. Structure 3-D de β-D-galactosidase (à partir de Protein Data Bank).
2.5.2. Production
La β-D-galactosidase est obtenue à partir d'une grande variété de sources telles que les
microorganismes (bactéries, champignons, levures), les animaux et les plantes, en
particulier les amandes, pêches, abricots, pommes (Tableau 5). En fonction de son
origine, les paramètres du procédé enzymatique comme le pH optimal, la température
optimale de réaction, la vitesse de conversion de lactose ainsi que le rendement en
GOS diffèrent nettement (Husain, 2010, Panesar et al., 2006, Richmond et al., 1981).

24
Tableau 5. Source de β galactosidase
Plantes Organes
animaux
Levure Bactéries Champignons
Pêche
Abricot
Amande
Les grains de Kéfir
Roses sauvages
Graines de
Luzerne
Les grains de Café
intestin
cerveau
Les tissus
de la
peau
le foie de
bovin
Kluyveromyces
lactis ya
Kluyveromyces
fragilis
Candida
pseudotropicalis
Escherichia coli
Bacillus megaterium
Thermus acquaticus
Streptococcus lactis
S. thermophilus
L.. bulgaricus
L. helareticus
Neurospora crassa
Aspergillus foetidus
Aspergillus niger
Aspergillus flavus
Aspergillus oryzae
A. phoenicis
Mucor pucillus
Mucor meuhei
En effet, les β galactosidases de sources bactériennes ont été largement utilisées pour
l'hydrolyse du lactose en raison de la facilité pour la fermentation, de l’activité
enzymatique élevée ainsi qu’une bonne stabilité. Traditionnellement, les β-
galactosidases les plus largement utilisées en industrie ont été obtenues à partir
d'Aspergillus spp. et Kluyveromyces spp. (Husain, 2010, Oliveira et al., 2011)
Les β galactosidases d’origine fongique ont généralement un pH optimal acide entre 2,5
et 5,4. Ainsi, elles sont plus efficaces pour l'hydrolyse du lactose présent dans le
lactosérum. Elles sont thermostables, mais bien plus sensibles à l'inhibition des produits
d’hydrolyse. Le galactose est souvent un inhibiteur compétitif. En effet, les enzymes
extraites de Aspergillus Niger sont fortement inhibées par le galactose, tandis que celles
extraites de Aspergillus oryzae sont moins exposées à cette inhibition (Husain, 2010,
Panesar et al., 2006).
La levure Kluyveromyces lactis est une importante source commerciale de β-
galactosidase. La production de la β-galactosidase par cette levure connait plus d'intérêt
depuis son utilisation par l'industrie alimentaire pour la production de lait pauvre en

25
lactose (Husain, 2010, Martínez-Villaluenga et al., 2008). En outre, au cours de cette
hydrolyse il y a production des galactooligosaccharides par transgalactosylation qui ont
suscité beaucoup d'intérêt au cours de ces dernières années (Martínez-Villaluenga et
al., 2008).
2.5.3. Mécanisme de la réaction et applications
Le mécanisme de la réaction d’hydrolyse de lactose par les β-galactosidases est
présenté dans la Figure 12, qui montre que la réaction se fait en deux étapes :
Étape (a) : La molécule de lactose forme le complexe enzyme-galactosyl dans le site
actif d’enzyme, avec libération simultanée du glucose.
Étape (b) : Le complexe enzyme-galactosyle est transféré à l’accepteur nucléophile
contenant un groupe hydroxyle. Le transfert à l’eau produit du galactose (réaction
d’hydrolyse, Schéma 9b avec R : hydrogène). Le complexe enzyme-galactose, formé
par hydrolyse du lactose peut réagir avec les molécules de glucides (avec R : mono ou
disaccharides), conduisant à la formation des di-, tri- et supérieur- galactosyl-
saccharides, appelés galactooligosaccharides (GOS) (Zhou et Chen, 2001).
La production de lactosucrose par β-galactosidase est due au transfert du résidu de
galactosyle du lactose (donneur) au saccharose (accepteur). Contrairement à la
transgalactosylation, la production du lactosucrose par levansucrases et Arthrobacter sp.
K-1 β-fructofuranosidase (transfructosylation), utilise le lactose comme accepteur de
glycosyle et le saccharose comme donneur (Figure 13)(Mu et al., 2013).

26
Figure 12. Mécanisme de réaction pour la hydrolyse et la transglycosylation du lactose par Kluyveromyces lactis β-galactosidase (Zhou et Chen, 2001).
Figure 13. La biosynthèse de lactosucrose et ses analogues par β-galactosidase de Bacillus circulans à partir du lactose et du saccharose comme substrats (Li et al., 2009,
Mu et al., 2013).
Glc
Glc Glc
Glc
Glc
Fru Gal
Gal Gal
Gal
Fru
Fru
Glc Fru Gal
Glc Gal Gal
α-1,2
α-1,2
α-1,2
β-1,4
α-1,2
β-1,4
β-1,4
β-1,4 β-1,4
β-1,4
β-1,3 Lactose Glucose
B. circulans β-galactosidase

27
Le mécanisme de la réaction de la β-galactosidase inclut l’hydrolyse du lactose et une
réaction de transglycosylation. Le processus utilisé pour l’hydrolyse par β-D-
galactosidase du lactose, du lactosérum et d’autres produits laitiers dépend énormément
de la substance hydrolysée. Différents processus sont utilisés suivant la nature du
substrat et les caractéristiques de l'enzyme. La première caractéristique, qui détermine
le choix et l'application d'une enzyme donnée, est la gamme de pH. Les enzymes
tolérant un pH acide sont plus adaptées pour le traitement du lactosérum, tandis que les
enzymes tolérant un pH neutre sont adaptées pour la transformation du lait et de
lactosérum doux. La β-galactosidase est également utilisée pour améliorer la production
et le vieillissement du fromage Cheddar par l’hydrolyse préliminaire du lactose en
glucose (Husain, 2010, Luckarift -Heather et Spain- Jim, 2008, Panesar et al., 2006).
Il y a un marché considérable pour le lait et les produits laitiers sans lactose, qui sont
obtenus par hydrolyse enzymatique à l'aide de β-galactosidases. Ces produits sont
appréciés et consommés par les gens ayant une intolérance au lactose, qui constitue un
problème commun. Il est estimé que plus de 70% de la population adulte dans le monde
a des problèmes de digestion du lactose (Oliveira et al., 2011).
La β-galactosidase est très appréciée pour son action hydrolytique sur le lactose. De
plus, l’activité de transférase de la β-galactosidase produit une série d'oligosaccharides.
Ces oligosaccharides ont un effet bénéfique sur la croissance de la microflore intestinale
souhaitable. Ceci attire de plus en plus l'attention des chercheurs afin d’étudier l’activité
de transférase de la β-galactosidase. En outre, l’action de transférase est utilisée dans
diverses applications pour la production d'ingrédients alimentaires, de produits
pharmaceutiques et d'autres substances biologiquement actives (Panesar et al., 2006).
2.5.4. β-Galactosidase de Aspergillus oryzae et son activité de
transgalactosylation
L'activité de transgalactosylation augmente avec la température dans l’intervalle de 40 à
55°C, ce qui se traduit par l'augmentation de rendement en GOS (Vera et al., 2011). En
effet, la température optimale pour l'hydrolyse du lactose et ONPG (Orthonitrophényl-β-
galactoside) par l'enzyme purifiée est de 50°C, tandis que l’enzyme brute est plus
résistante à la chaleur et son maximum d’activité hydrolytique pour ces mêmes substrats
est atteint à 55-60°C (Park et al., 1979). Cette gamme de température où l’activité

28
enzymatique est maximale, est maintenue aussi bien pour la β-galactosidase libre que
celle immobilisée (Freitas et al., 2011).
L’étude de la stabilité et de l’activité de β-Galactosidase effectuée par plusieurs
chercheurs (Freitas et al., 2011, Park et al., 1979, Vera et al., 2011) montrent que
l’hydrolyse maximale de lactose et ONPG par l'enzyme purifiée libre et immobilisée se
déroule à un pH de 4,.5 et 5,0, respectivement. Mais l’enzyme ne perd pas son activité
lorsqu'elle est maintenue à un pH compris entre 3,6 et 8,0 pendant une nuit à
température ambiante (Park et al., 1979).
Concernant l'influence du pH et de la température sur la synthèse des GOS, il a été
montré que, même si les deux paramètres affectent les vitesses de réaction, ils
n'affectent pas la concentration des produits. Dans la synthèse des GOS cinétiquement
contrôlée, l'équilibre entre la transgalactosylation et l’activité hydrolytique n'est pas
altérée par la température (Vera et al., 2011). Cependant, il a été prouvé que
l'augmentation de la concentration du lactose améliore le rendement maximal. Pour
cela, il faut bien prendre en considération la concentration initiale en lactose. Selon la
littérature, avec l’utilisation β-galactosidase de Aspergillus oryzae à une concentration
initiale en lactose de 500 g/L, le rendement en GOS obtenu est de 27% (p/p), dont
environ 70% sont des trisaccharides et le reste est distribué entre les tetrasaccharides et
les pentasaccharides avec des proportions différentes (Urrutia et al., 2013). D’autre part,
avec une solution de lactose à 200 g/L, l’Aspergillus oryzae β-galactosidase libre forme
seulement des trisaccharides, avec un rendement maximal de 22,6% (p/p) (Gaur et al.,
2006). En effet, la β-galactosidase de Aspergillus oryzae serait préférablement utilisée
lorsque la cible est un produit avec une teneur importante en di-et trisaccharides (Urrutia
et al., 2013).
2.6. Immobilisation des enzymes
Les enzymes, généralement, utilisées dans les réactions sont des enzymes libres, d’où
la difficulté de les récupérer (Luckarift -Heather et Spain- Jim, 2008, Oliveira et al.,
2011). De plus, ces enzymes sont sensibles aux conditions environnementales, et
subissent généralement une dénaturation ou une désactivation par des réactifs
chimiques, la température, le pH, les solvants, etc. (Falahati et al., 2012).

29
Afin de surmonter ces obstacles, et par conséquent diminuer le coût de la réaction,
l’immobilisation des espèces biologiquement actives sur des matériaux inorganiques et
organiques est souvent utilisée dans une tentative de stabilisation de l'activité
enzymatique et afin de permettre la réutilisation du biocatalyseur. Cette stratégie
combine la haute sélectivité des réactions enzymatiques et les propriétés mécaniques
du support (Falahati et al., 2012, Husain, 2010).
2.6.1. Avantage d’immobilisation des enzymes
L’immobilisation des enzymes présente plusieurs avantages tels que :
Utilisation efficace de l’enzyme en garantissant sa stabilité vis-à-vis du pH, de
la température, et des solvants (Husain, 2010, Kim et al., 2007);
Utilisation répétitive de l’enzyme (biocatalyseur) pour la séparation et
récupération du catalyseur à partir du flux de produits, et par conséquent
diminution de coût des réactions enzymatiques (Humphrey et Wright, 2004);
Augmentation de la résistance vis-à-vis le cisaillement, et par conséquent,
une prolongation de la durée de vie de l’enzyme (End et Schöning, 2004);
Amélioration de l’activité de enzymes immobilisée grâce à une meilleure
disponibilité des centres catalytiques actifs (End et Schöning, 2004);
Facilité de développer des procédés continus (Humphrey et Wright, 2004) ;
Amélioration de la résistance mécanique, la densité, la stabilité thermique;
Facilité de manipulation permettant une synthèse à débits élevés dans les
réacteurs continus, et facilité de la régénération des biocatalyseurs (enzymes
supportées) (Husain, 2010);
Amélioration de la vitesse de réaction suite à une cinétique favorablement
améliorée (pour certains types de réacteurs) (End et Schöning, 2004).
Ce domaine est en progression continue vu son intérêt scientifique et ses applications
multiples qui attirent les industriels (Falahati et al., 2012).

30
Le Tableau 6 illustre la majorité des produits fabriqués industriellement suite à des
biocatalyseurs immobilisés. Les principaux domaines industriels liés à ce procédé sont;
la production de sucres, des acides aminés ainsi que des produits chimiques et
pharmaceutiques (Bornscheuer, 2008). En effet, l’immobilisation des enzymes dépend
de deux paramètres : le support et la méthode utilisée pour l’immobilisation.
Tableau 6. Les principaux produits fabriqués à l'échelle industrielle en utilisant des enzymes immobilisées (Bornscheuer, 2008)
Produit Enzyme Capacité (t/an)
Sirop de maïs à haute teneur en
fructose
glucose isomérase (xylose
isomérase)
> 7, 000,000
Acrylamide nitrile hydratase > 30,000
Acide 6-Aminopenicillanique pénicilline acylase 2,000
Acides L-amino Aminoacylase > 300
2.6.2. Immobilisation des enzymes : supports
Étant donné que les exigences spécifiques sont différentes pour chaque enzyme ainsi
que pour ses applications, il est impossible de trouver un support universel pour
l'immobilisation de toutes les enzymes. En plus, il faut prendre en considération les
caractéristiques chimiques et physiques du matériau pour permettre la sélection du
support le plus approprié. Les principales caractéristiques de la composition chimique
sont les produits chimiques de départ pour les synthèses, les groupes fonctionnels
disponibles pour les interactions avec l'enzyme, et la stabilité chimique dans les
conditions réactionnelles employées (Yiu et Wright, 2005). Parmi les caractéristiques
physiques, on cite :
La forme et la taille des pores;
La surface appropriée;
La stabilité thermique;
Le caractère hydrophile;

31
L’insolubilité dans le milieu réactionnel;
Le volume accessible de support pour les enzymes.
En fonction de leurs compositions chimiques, et en se basant sur la littérature, on
distingue deux types de supports pour le procédé d’immobilisation des enzymes: les
supports organiques et inorganiques (End et Schöning, 2004).
En effet, de nombreuses matrices inorganiques ont été testées comme supports pour
l'immobilisation d'enzymes tels que le gel de silice, le verre, l'alumine, des minéraux
argileux, Les zéolites, et les oxydes de métaux de transition (par exemple; NiO et TiO2).
Une partie de ces matériaux sont synthétiques et d'autres sont d'origine naturelle. Bien
qu'ils soient disponibles et non coûteux, les matériaux inorganiques naturels ne sont pas
couramment utilisés pour l’immobilisation des enzymes, puisque leurs caractéristiques
chimiques et physiques sont moins compatibles avec les applications enzymatiques
(End et Schöning, 2004). Tandis que les propriétés des matériaux synthétiques telles
que la surface, la composition chimique, la porosité, la distribution des pores, etc. sont
plus modulables, d’où l’utilisation avantageuse de ces matériaux en biocatalyse.
Les matériaux organiques sont les principaux supports utilisés pour des applications à
grande échelle. Cette utilisation des supports organiques pour l'immobilisation
d'enzymes est due à la variété de groupes fonctionnels organiques qui peuvent disposer
de nombreuses interactions avec l’espèce enzymatique, en particulier par des liaisons
covalentes. Mais ces matrices présentent quelques inconvénients dus à l’instabilité
thermique par rapport aux matrices inorganiques ainsi qu’une instabilité vis-à-vis des
solvants et les attaques microbiennes (End et Schöning, 2004). D’où l’intérêt accordé
aux supports inorganiques ces dernières années, et exceptionnellement aux supports
mésoporeux à base de silice. Ils présentent toute une famille de matériaux
nanostructurés très ordonnés qui permettent l’encapsulation des molécules et des
enzymes de dimensions différentes (Takahashi et Miyazaki-Imamura, 2008) (Figure 14).
À guise d’exemple, la silice mésoporeuse ordonnée offre d'excellentes possibilités pour
l'immobilisation d'enzymes, surtout par liaison covalente, en raison de la disponibilité de
groupes silanols bien définis. Ces groupes fournissent des sites réactifs pour la
fonctionnalisation et offrent des propriétés de surface appropriées, permettant de
contrôler la position et la densité du biocatalyseur immobilisé avec précision (Lee et al.,
2009).

32
En effet, cette attention attribuée à ces matériaux est due au fait qu’ils possèdent:
Une large surface active (surface externe et à l’intérieur des pores);
Une uniformité de structure bien ordonnée;
Une taille de pores contrôlables (2 nm < Ø < 50 nm) (Kim et al., 2006).
Figure 14. Comparaison de la taille des pores de certains matériaux nanoporeux et la taille moyenne moléculaire de certaines enzymes couramment utilisés (Humphrey et
Wright, 2004).
La forme et la taille de ces matériaux aident beaucoup l’immobilisation des enzymes. Or,
les MCM-41, les FSM-16 et la SBA-15 sont parmi les premiers matériaux utilisés comme
supports d’enzymes depuis les années 90. Cependant, de nos jours, il y a une multitude
de supports utilisés pour l’immobilisation des enzymes tels que : la SBA-15 (avec sa
forme hexagonale qui est utilisée comme un support pour différentes enzymes comme la
lipase et la trypsine) (Humphrey et Wright, 2004), les HMMS qui sont utilisés
efficacement comme supports pour les α-chymotrypsin et les lipases grâce à leur forme
particulière. Les HMMS sont des mésoporeux avec des pores allant jusqu’à 37 nm reliés
par des canaux de 13 nm (Kim et al., 2007) (Figure 15). Les MCF sont aussi utilisés
5 10 15 20 30 40 50 60 70 80 90 100 150 200
250 300 500 1000
Cyctochrome c
Trypsin
Lipase
Penicillin Acylase
Chloroperoxidase
Horseradish peroxidase
Diamètre de pore / Å
Zeolites Matériaux
macroporeux
MCF
FDU-5 MCM-48
SBA-1 MCM-41 SBA-15
Région mésoporeuse
(Diamètre de pore = 20 Å – 200 Å

33
largement pour l’immobilisation des enzymes, grâce à la forme sphérique et la taille de
leurs pores atteignant 22 nm (Schmidt-Winkel et al., 1999).
Le Tableau 7 résume les propriétés caractéristiques de différents matériaux
mésoporeux à base de silice utilisés pour encapsuler des enzymes (Lee et al., 2009).
Figure 15. Schéma représentatif de HMMS (Kim et al., 2007).
En effet, le choix approprié d’un support est critique. Les propriétés chimiques et
physiques du support (la structure chimique du support, sa porosité, la taille de ses
pores et sa morphologie), ainsi que la taille de l’enzyme et ses caractéristiques, doivent
tous être considérés pour faire un bon choix. Lors du choix du matériau, il faut aussi
prendre en considération la méthode d’immobilisation utilisée et vice-versa.

34
Tableau 7. Propriétés caractéristiques des différents matériaux de silice mésoporeuses utilisés pour encapsuler des enzymes
Matériaux
mésoporeux
Source de silice Description Diamètre des
pores (nm)
MCM - 41 TEOS, sodium
silicate
2D canaux hexagonaux 2-10
MCM-48 TEOS, sodium
silicate
Cubique 3D 2-4
FSM-16 Polysilicate kanemite 2D canaux hexagonaux ~ 4
SBA-1 TEOS Mésostructure cubique
3D
2-3
SBA-15 TEOS, sodium
silicate
2D canaux hexagonaux 5-30
SBA-16 TEOS, TMOS Cages sphériques 5-30
MCF TEOS Mousse cellulaire 10-50
HMS TEOS Mésostructure
désordonnée
2-0
MSU-X TEOS, TMOS Mésostructure
désordonnée
2-15
IBN-X TEOS Nanoparticules 5-20
PMOs (RO)3Si-R-Si(OR)3 2D ou 3D hexagonal 2-20
2.6.3. Méthode d’immobilisation des enzymes
Plusieurs méthodes d'immobilisation d'enzymes ont été utilisées depuis les années
1980. Chaque méthode a ses avantages et ses inconvénients, et le choix dépend de la

35
nature de l'enzyme cible et des conditions de la réaction. Toutefois, aucune méthode
d’immobilisation ne peut être généralisée pour toutes les enzymes et leurs applications.
En effet, il y a plusieurs méthodes d’immobilisation d’enzyme dont les plus utilisées sont
les suivants :
Adsorption : L'immobilisation d'enzymes par adsorption a été parmi les
premières méthodes d'immobilisation d'enzymes. Elle représente la méthode la
plus simple et la plus naturelle (sans produits chimiques) pour immobiliser des
enzymes sur des supports mésoporeux (Husain, 2010). Cette méthode consiste
à un attachement physique entre l’enzyme et le support par des forces de Van
Der Waals, des liaisons hydrogène, ou l'interaction hydrophile. Cet attachement
est obtenu suite au procédé suivant : le support mésoporeux est mis en
suspension dans une solution d'enzyme pour une certaine durée, habituellement
pour une nuit, ensuite on sépare le support de la solution par centrifugation. En
général, les enzymes immobilisées préparées par adsorption ont tendance à se
desorber du support, en raison de l'interaction relativement faible (Humphrey et
Wright, 2004).
Liaison covalente : L’immobilisation des enzymes par liaison covalente est la
technique la plus fréquemment associée au terme « enzyme immobilisée »
(Bowers et Carr, 1980). En effet, une liaison covalente d'une enzyme à un
support est basée sur une réaction chimique entre les résidus d'acides aminés
actifs situés sur la surface de l'enzyme et la partie fonctionnalisée du support, ou
inversement (Cao, 2006), comme illustré dans la Figure 16. Cette méthode
d'immobilisation d'enzymes, fournit habituellement de plus forts liens entre
l’enzyme et le support, comparée à d’autres méthodes d’immobilisation
d’enzymes telles que l’immobilisation par adsorption. Ainsi, le détachement de
l'enzyme de la matrice utilisée est souvent minimisé (Cao, 2006). Cependant,
les méthodes covalentes sont relativement coûteuses et compliquées. Les
rendements d'activité peuvent être faible en raison de l'exposition de l'enzyme à
des conditions sévères et parfois, le site actif des enzymes peut être modifié par
l'intermédiaire des réactions chimiques utilisées pour créer la liaison covalente
(Husain, 2010).

36
Figure 16. Immobilisation covalente de l'enzyme sur un support: (A) des résidus d'acide aminé actif; (B) fonctionnalité de liaison du support; (C) support; (D) espace (Cao, 2006).
Encapsulation : Par définition, l’encapsulation d'une enzyme est la formation
d'une barrière similaire à une membrane physique semi-perméable autour
d’une préparation enzymatique (Cao, 2006). Par exemple, l'encapsulation
des enzymes dans des sol-gel est basée sur la condensation des
précurseurs de silice autour des molécules enzymatiques, de sorte qu'un
réseau poreux est formé dans lequel la protéine est piégée au hasard (Figure
17) (Urrego et al., 2010). L’encapsulation de l'enzyme exige des conditions
extrêmement bien contrôlées (Husain, 2010).
Figure 17. Représentation de l’immobilisation des enzymes par encapsulation.
Réticulation «cross-linking» (Figure 18): la réticulation est une méthode simple
et efficace pour immobiliser des enzymes sur des surfaces mésoporeuses
avec une grande stabilité, car elle peut empêcher le détachement des
enzymes lors de la lixiviation, et par conséquent améliorer la stabilité

37
enzymatique. La réticulation est basée sur la formation de liaisons covalentes
entre les molécules d'enzyme, au moyen de réactifs bi-ou multifonctionnels,
conduisant à des agrégats réticulés en tridimensionnels. En raison de la
réticulation, très peu de désorption se produit et par conséquent un meilleur
attachement de l'enzyme est observé. Un avantage supplémentaire de
préparations d'enzymes réticulées, est qu'il n'y a pas d'exigence pour le
support pour l'immobilisation d’enzymes (Husain, 2010).
La réticulation est mieux utilisée en conjonction avec l'une des autres
méthodes. Elle est principalement utilisée comme un moyen de stabiliser les
enzymes adsorbées et empêcher leur détachement. Cependant, la
réticulation peut provoquer des changements considérables dans le site actif
des enzymes, et également des limitations de diffusion accentués, qui
peuvent entraîner une perte importante de l'activité. La perte de l'activité
enzymatique se produit également lors de la réticulation de l'enzyme en
utilisant des réactifs bifonctionnels ou multifonctionnels. Cette méthode
présente un contrôle difficile de la taille des agrégats, une accessibilité
difficile des substrats aux noyaux d'agrégats, et un manque de résistance
mécanique de l'enzyme réticulée (Zhao, 2006). Le réactif le plus couramment
utilisé pour la réticulation comprend du glutaraldéhyde (GA). On peut citer
d'autres réactifs utilisés pour la réticulation, notamment le bis-diazobenzidine-
2,2-disulfonique, diazobenzidine, l'hexaméthylène diisocyanate (López-
Gallego et al., 2005).
La réticulation implique généralement un processus en deux étapes :
Premièrement, des molécules enzymatiques sont adsorbées à l'intérieur des
nanocanaux de la silice mésopoureuse (MPS). Deuxièmement, les enzymes
adsorbées sont réticulées par addition d'un agent de réticulation bifonctionnel
comme le glutaraldéhyde, afin de produire une charge enzymatique très
stable, et active. En effet, le glutaraldéhyde représente l'agent de réticulation
de choix car il est peu coûteux et facilement disponible en quantités
commerciales.

38
Figure 18. Représentation de l’immobilisation par réticulation «cross-linking» (Kim et al., 2008).

39
2.6.4. Immobilisation de β-galactosidase à partir de Aspergillus
oryzae
La majorité des systèmes d’immobilisation de β-galactosidase ont été développés dans
le but de réalisé l'hydrolyse du lactose; il y a eu quelques travaux concernant la
production de GOS. Il est important de noter que les méthodes d’immobilisation
favorables pour l'hydrolyse du lactose peuvent être différentes de celles de la production
des GOS.
La β-galactosidase à partir de Aspergillus oryzae a été immobilisée par plusieurs
procédés et sur une variété de matrices, y compris l’encapsulation, la réticulation,
l'adsorption, les liaisons covalentes ou la combinaison de ces méthodes (Tableau 8).
Étant donné que chaque méthode a ses propres avantages et inconvénients, le choix de
la méthode appropriée dépend de l'enzyme, de la matrice, de la réaction, des conditions
expérimentales et du réacteur.

40
Tableau 8. Différentes méthodes et différents supports d’immobilisation de β-galactosidase à partir de Aspergillus oryzae
Méthodes d’immobilisation Activité
enzymatique
* (%)
Références
Réticulation avec du glutaraldéhyde
sur des fibres composées de gélatine
et d'alginate de
56
Tanriseven & Dogan
(Zhang et al., 2001)
couplage avec carbodiimide à des
billes d'alginate 76
Dominguez et al.
(Domínguez et al., 1988)
Encapsulation dans l’alcool spongeux
(polyvinyl cryogel) --
Rossi et al. (Parhi et al.,
2008)
encapsulation dans des billes
d'alginate de cobalt réticulée avec du
glutaraldéhyde
83
Ates & Mehmetoglu (Lee
et al., 2005)
micro-encapsulation dans des billes
d'alginate 64
Dashevsky (Dashevsky,
1998)
encapsulation dans de la gélatine et
réticulée avec la transglutaminase 8–46
Fuchsbauer et
al.(Fuchsbauer et al., 1996)
adsorption sur du chlorure de
polyvinyle (PVC)
adsorption sur membrane de gel de
silice
--
Bakken & Hill Jr (Bakken
et al., 1990)
adsorption sur la célite
liaison covalente de chitosane
agrégation réticulé par le
2
18,4
13.5
Gaur et al. (Gaur et al.,
2006)

41
glutaraldéhyde
liaison covalente dans les mousses de
polyuréthane --
Hu et al. (Hu et al., 1993)
la liaison covalente à la toile de coton 55
Albayrak et al. (Albayrak
et Yang, 2002)
Liaison covalente sur des supports
siliceux macroporeux 48
Bernal.C et al.(Bernal et
al., 2014)
Liaison covalente sur des
microsphères de chlorure de vinyle
amino-Fonctionné avec le couplage de
glutaraldéhyde
80
Mohy Eldin et al.(Mohy
Eldin et al., 2014)
encapsulation dans des nanoparticules
de silice agrégées --
Wu. et al. (Wu et al., 2013)
Liaison covalente sur des méso-
macroporeux silices greffées par des
groupements glyoxyl
--
Bernal.c et al.(Bernal et al.,
2013)
Liaison covalente sur la surface des
nanoparticules d'argent fonctionnalisé
via le glutaraldéhyde
--
Ansari. et al. (Ansari et al.,
2012)
Adsorption sur la résine Duolite A568 --
Gürdaş. et al. (Güleç et al.,
2010)
Adsorption sur des nanoparticules
d'oxyde de zinc 73
Husain. et al. (Husain et
al., 2011)
*Activité enzymatique : c’est le pourcentage de formation de produit par rapport au
substrat initial.

42
2.7. Conclusion
Les oligosaccharides prennent de plus en plus une place dans notre alimentation grâce
à leurs effets prouvés bénéfiques pour la santé humaine. En effet, ils existent
naturellement dans les fruits et les légumes mais ils peuvent aussi être synthétisés soit
par voie chimique ou bien par voie enzymatique. La voie enzymatique reste toujours un
meilleur choix comparé à la voie chimique pleine d’inconvénients. Le lactosucrose est un
trisaccharide non digestible favorable au développement de la flore intestinale,
synthétisé par le biais de levansucrase, de fructofuranosidase ou de β-galactosidase en
présence de lactose et de sucrose. La principale limite de la méthode enzymatique est
son coût élevé. Pour diminuer ce coût, on a recours à l'immobilisation des enzymes sur
des supports pour les réutiliser ultérieurement. La méthode d’immobilisation ainsi que la
nature de support affectent considérablement la réaction catalytique et son rendement.

43
Chapitre 3. Hypothèses et objectifs de
recherche
3.1. Hypothèses du travail
D'après la revue de littérature, nous émettons les hypothèses suivantes pour
synthétiser le lactosucrose par catalyse hétérogéne.
1) La β-galactosidase pourrait être immobilisée sur différents supports siliceux
mesoporeux sans perdre son activité et par conséquent le lactosucrose peut être
synthétisé avec l’enzyme immobilisée par catalyse hétérogène.
2) La taille et la forme des pores des supports pourraient avoir des effets sur
l’immobilisation et l’activité des enzymes ainsi que la cinétique de la réaction.
3.2. Objectifs de travail
3.2.1. Objectif principal
L’objectif principal de ce travail est de produire du lactosucrose avec une approche
biotechnologique verte basée sur la catalyse hétérogène en utilisant l’enzyme β–
galactosidase supportée sur des matériaux inorganiques à base de silice.
3.2.2. Objectifs spécifiques
L’objectif général de ce sujet de recherche engendre plusieurs autres objectifs
spécifiques. Ces objectifs peuvent être représentés comme suit :
1) Synthétiser et caractériser les différents matériaux mésoporeux;
2) Optimiser les conditions d’immobilisation de β–galactosidase sur des matériaux
inorganiques à base de silice;
3) Synthétiser le lactosucrose avec la β-galactosidase immobilisée;
4) Réutiliser des biocatalyseurs plusieurs fois tout en gardant l’activité et la stabilité
d’enzyme.


45
Chapitre 4. Matériels et méthodes
4.1. Matériels biologiques
β-Galactosidase synthétisée à partir d’Aspergillus oryzae (Sigma-Aldrich).
Elle hydrolyse le lactose en ses composants monosaccharides ; glucose et
galactose. Elle catalyse également la transglycosylation de glucose. Une
unité de β-Galactosidase hydrolyse 1,0 µmole de lactose par minute à pH de
4,5 à 30°C. Chaque milligramme d’enzyme contient 8 unités de β-
Galactosidase.
Albumine de sérum bovine (BSA) : pureté ≥98%, Sigma-Aldrich.
4.2. Produits chimiques
D-(+)-lactose monohydrate, Sigma-Aldrich.
Saccharose, Sigma-Aldrich.
Fructose: (pureté > 98%), Sigma-Aldrich.
D-(+)-Glucose, Sigma-Aldrich.
D-(+)-Galactose, Sigma-Aldrich.
Tétraéthylorthosilicate, précurseur de silice (TEOS; pureté > 98%), Sigma-
Aldrich.
Pluronic P123 (acide) : Surfactant (agent structurant) (EO20PO70EO20),
Sigma-Aldrich.
1,3,5-triisopropylbenzène (TIPB) : pureté > 95%, Sigma-Aldrich.
1,3,5-triméthylbenzène (TMB) : pureté > 96%, Sigma-Aldrich.
Acide chlorhydrique (HCl, 36-38%), Fisher Scientific.
Chlorure de potassium (KCl, 99-100%), BDH Inc.
Hydrogénophosphate de potassium (K2HPO4, ≥98.0 %). Fisher Scientific.
Phosphate de potassium monobasique (KH2PO4, ≥99.0%), Sigma-Aldrich.
Glutaraldehyde (GA, Grade I, 25% dans H2O), Sigma-Aldrich.
Acide bicinchoninique (BCA) : pureté ≥98%), Sigma-Aldrich.
Carbonate de sodium (Na2CO3, ≥99.5%), Sigma-Aldrich.
Bicarbonate de sodium (NaHCO3, ≥99.7%), Fisher Scientific.

46
Tartrate de sodium (C4H4Na2O6, ≥99.0%), Fisher Scientific.
Soude en pastille : (NaOH, pureté ≥ 97%), Laboratoire Mat.
Sulfate de cuivre pentahydraté (CuO4S·5H2O, ≥98.0%), Fisher Scientific.
Chlorure de sodium (NaCl, ≥99.0%), Sigma-Aldrich.
4.3. Équipements
Verreries de laboratoire.
Balances (maximum de 210 g, déviation = 0,1 mg).
Agitateurs magnétiques (Corning Inc., Fisher Scientific)
pH-mètre (Accumet, Fisher Scientific)
Étuves (Maximum de 275°C), Fisher Scientific.
Four à calcination, programmable, Vulcan 3-550 PD NEY USA.
Diffractomètre de rayon X (Ultima III, Rigaku, Japan)
Spectrophotomètre d’absorption UV visible, Genesys 20.
Équipement complet de physisorption Autosorb 1 (Quantachrome, Florida,
USA).
Système HPLC Waters (Millipore Corp., Milford, Mass. U.S.A).
Spectromètre de masse TQD (Waters).
Un microscope électronique à transmission JEM-3010 (JEOL, Japon).
4.4. Méthodologie
4.4.1. Réaction enzymatique avec l’enzyme libre (approche
homogène)
La synthèse du lactosucrose a été réalisée à partir du lactose et du saccharose en tant
que substrats, en présence de β-galactosidase en tant que catalyseur.
Le lactose et le saccharose avec différentes proportions, ont été dissous dans 50 mL de
solution aqueuse. Ensuite, une concentration bien déterminée de β-galactosidase a été
ajoutée à cette solution. Le mélange enzyme/substrats est placé à l’étuve à différentes
températures pendant des intervalles de temps variés allant jusqu’à 60 heures.

47
Le but de l’expérience est d’étudier la stabilité et l’influence des conditions opératoires
sur le rendement enzymatique et la cinétique de réaction. La synthèse a été effectuée
avec différentes concentrations de substrats, selon deux rapports molaires différents de
donneur/accepteur (lactose/saccharose). Les combinaisons sont résumées dans le
Tableau 9.
L’optimisation des paramètres expérimentaux (concentration d’enzyme, pH, et T°) a été
effectuée en utilisant le rapport des substrats (1 :1) à une concentration de 30% (m/v) de
chacun de ces derniers (lactose et saccharose), en variant les paramètres
expérimentaux comme suit:
Quatre concentrations différentes d’enzyme (1, 4, 6 et 10 U/mL).
Trois pH différents (7 ; 5 ; 3,5).
Trois températures différentes (30°C, 40°C, 50°C).
Tableau 9. Les conditions opératoires de la biosynthèse de lactosucrose à partir de lactose et de saccharose
Réaction
Rapport molaire 1 :1
Réaction
Rapport molaire 2 :1
Lactose
(g/100ml)
Saccharose
(g/100ml)
Lactose
(g/100ml)
Saccharose
(g/100ml)
A 15 15 D 50 25
B 30 30 E 30 15
C 50 50 F 15 7,5
Au cours de l'incubation, 100 µL des échantillons ont été prélevés à des intervalles de
temps bien définis. Ces quantités prélevées ont été diluées 100 fois, et chauffées à
100°C pendant 10 minutes pour désactiver l'enzyme. Afin d’identifier et de quantifier les
produits, tous les échantillons ont été analysés par Chromatographie en phase liquide à
haute performance (HPLC).

48
4.4.2. Les analyses HPLC
La teneur en saccharides des échantillons a été déterminée en utilisant un appareil
HPLC Waters (Millipore Corp., Milford, Mass. U.S.A) comprenant un détecteur à indice
de réfraction (modèle 2142, LKB, Bromma). La colonne (Waters Sugar-Pack-I, Waters,
Millipore Corp.) était maintenue à une température de 90ºC. La phase mobile
isocratique, constituée d’une solution d’EDTA (50mg/L), est assurée avec un débit de
0.5 mL/min. L’identification ainsi que la quantification ont été faites en comparaison
avec des solutions standards à différentes concentrations connues. L’appareil est équipé
d’un injecteur automatique, fixé à 50µL par injection.
4.4.3. Calculs des rendements
Durant ce travail, l’évolution des produits formés et des substrats consommés est définis
comme:
La conversion du lactose (%) :
(%))(.
)()(.(%).
int
int
gLactose
glactoseglactoseLacCon
ial
tial (1)
La conversion du saccharose (%) :
(%))(.
)()(.(%).
int
int
gSaccharose
gsaccharosegsaccharoseSaccCon
ial
tial (2)
Le rendement en GOS (%) :
(%))(.
)((%).Rendement
int gLactose
gGOSGOS
ial
(3)
4.4.4. Purification et caractérisation des produits formés
4.4.4.1. UPLC-MS
Pour l'isolement de GOS inconnus produits au cours de la synthèse, la réaction a été
arrêtée après 20 heures. Le biocatalyseur a été désactivé par l'ébullition de la solution
pendant 10 min. Les échantillons n’ont subi aucune dilution au préalable, pour avoir une
concentration très élevée lors de séparation des pics par UPLC. Par la suite, les pics
séparés sont analysés par spectrométrie de masse (MS). L'analyse MS a été réalisée

49
sur un spectromètre de masse TQD (Waters) équipé d'une interface electrospray Z-
spray. L'acquisition des données a été réalisée avec le logiciel MassLynx 4.1 (Waters).
4.4.4.2. Analyse du lactosucrose par résonance magnétique nucléaire
Les structures chimiques des galactooligosaccharides biosynthétisés ont été identifiées
par résonance magnétique nucléaire unidimensionnelle (1D, 1H-RMN). Les spectres
RMN ont été enregistrés à température ambiante (25°C) sur un spectromètre Bruker
Avance 500 MHz spectromètre III (Bruker Ltd, Rheinstetten, Allemagne), équipé d'une
tête de sonde Bruker 5 mm TXI avec gradient de l'axe Z, en opérant à 500 MHz. Les
échantillons de galactooligosaccharides purifiés ont été dissous dans de l'eau deutérée
(D2O). Les spectres 1H-RMN sont obtenus avec l'accumulation de 128 balayages par
une largeur d'impulsion de 45°, temps d'acquisition de 2,0 s et un retard de recyclage de
1 s. Les spectres 1H -RMN ont été acquis à travers une fenêtre spectrale de 9 ppm. Les
déplacements chimiques 1H ont été référencés au tétraméthylsilane TMS (δ 0,00 ppm)
comme étalon externe. Les valeurs des déplacements chimiques (δ) sont données en
partie par million (ppm) et les constantes de couplage (J) sont données en hertz (Hz).
Les pics ont été signalés comme singulet (s), doublet (d), triplet (t), doublet dédoublet
(dd) ou multiplet (m), afin de faciliter la lecture et la compréhension. Le traitement des
données acquises a été réalisée en utilisant les logiciels: Topspin 2.1 (Bruker Ltd,
Rheinstetten, Allemagne), et aussi Mestre-C Lite et MestReNova (Mestrelab Research,
Escondido, Californie, USA).
4.4.5. La synthèse des supports
Dans le but d’étudier l’effet de la taille et la forme des pores des supports, ainsi leur
surfaces spécifiques sur l’immobilisation et l’activité des enzymes, nous avons choisi
sept différents supports ayant des tailles de pores et des surfaces spécifiques
différentes.
4.4.5.1. Synthèse du support mésoporeux SBA-15
La méthodologie de synthèse de la SBA-15 a été mise au point par Zhao et ses
collègues (Zhao et al., 1998). La SBA-15 de forme hexagonale a été synthétisée en
milieu acide en utilisant comme agent structurant (tensioactif) le poly (éthylène glycol)-
poly (propylène glycol)-poly (éthylène glycol), EO20PO70EO20, qui est un copolymère
(Pluronic 123).

50
Dans une synthèse typique, et afin de produire environ 10 g de la SBA-15 calciné, 20 g
de Pluronic 123 a été dissous dans 600 mL d'une solution de HCl (2 N) à température
ambiante, sous agitation magnétique, pendant 3 h. Une fois le tensioactif est solubilisé,
la solution obtenue est chauffée lentement jusqu'à atteindre 40°C, puis 37 g de
tetraethoxysilane, TEOS (précurseur de silice) sont ajoutés goutte à goutte à la solution.
Le mélange réactionnel résultant reste sous agitation à 40°C pendant 24 h. Par la suite,
le mélange est versé dans une bouteille L-Pyrex et est transféré vers une chambre de
four sans agitation pendant 24 h à 100°C pour le vieillissement de la synthèse de la
SBA-15. Le produit final a été récupéré par filtration, lavé plusieurs fois avec l'eau et
l'éthanol, et séché à température ambiante. Finalement, la poudre obtenue est calcinée
dans un four programmable à 540°C pendant 5 heures avec un pas de 5°C/min
(Belkacemi et al., 2006).
4.4.5.2. Synthèse de nanomateriaux en forme de barre (Rod-like) ‘’S2.9’’ et
en forme de vésicule (Vesicle-like) ‘’S34.8’’
Le mode de préparation est similaire à celui de la SBA-15, mais avec quelques
modifications. Tout d'abord, 1 g de P123 a été dissous dans un mélange de 8,5 mL de
H2O et 30 g de solution aqueuse de HCl (2 M). La solution résultante a été agitée à
température ambiante jusqu'à ce qu’elle devienne limpide. Ensuite, des quantités
spécifiées de TIPB (1,3,5-triisopropylbenzène) ont été ajoutées à cette solution, avec
des rapports molaires (TIPB: P123) de 2,9: 1 (pour la ‘’S2.9’’) et 34,8: 1 (pour la
‘’S34.8’’). Le mélange a été agité à température ambiante pendant 18h. Puis, 2,1 g de
TEOS ont été ajoutés à ces solutions sous agitation pendant 8 min, et le tout a été
maintenu sous des conditions statiques à 35°C pendant 24 h. Par la suite, ces deux
solutions ont été transférées dans un autoclave revêtu de téflon, maintenu à 130°C
pendant 24h. Enfin, des précipités blancs ont été filtrés et lavés avec de l'eau, séchés à
température ambiante, puis calcinés à 500°C pendant 6 h à une vitesse de chauffage de
1°C/min dans un four tubulaire afin d’éliminer les résidus organiques (Zhou et al., 2007).
4.4.5.3. Synthèse des MCF (Mesocellular Siliceous Foams)
Dans une synthèse typique, 2 g de P123 ont été dissous dans une solution aqueuse de
75 mL de HCl (1,6 M) à température ambiante. Par la suite, 2 g de TMB (1,3,5-
triméthylbenzène) ont été ajoutés. Après au moins 45 minutes à une température de 35-
40°C, 4,4 g de TEOS ont été ajoutés. Le mélange est maintenu à cette température (35-

51
40°C) sous agitation pendant 20 h. Puis il a été transféré vers une étuve à 100°C
pendant 24 h pour le vieillissement. Le solide obtenu est filtré puis lavé et séché à
température ambiante pendant 24 h. Finalement, il est calciné à 500°C pendant 8 h à
une vitesse de chauffe de 5°C/min (Schmidt-Winkel et al., 1998).
4.4.5.4. Synthèse de LP-FDU-12
La synthèse de ce type de matériaux a été effectuée selon le protocole suivant : 0,5 g de
P123, 0,6g de TMB et 2,5g de KCl ont été dissout dans 30 mL de HCl (2M). Après 2 h
d’agitation, 2,08 g de TEOS ont été ajoutés à la solution. Ensuite, la solution a été mise
sous agitation à une basse température (10°C, 15°C et température ambiante) pendant
24h. Lors de vieillissement, le mélange a été transféré vers une température de 100°C
pendant 24 h. Finalement, les matériaux ont été filtrés, lavés et séchés à température
ambiante avant la calcination à 550°C/5h à une vitesse de chauffe de 2°/min (Fan et al.,
2005, Kruk et Hui, 2008).
4.4.6. Caractérisations des supports
4.4.6.1. Analyses BET
Les isothermes d'adsorption / désorption des échantillons calcinés (supports
synthétisés) ont été obtenues en utilisant un analyseur d'adsorption volumétrique
(modèle Autosorb-1, Quantachrome Instruments, Boyton Beach, FL) à 77 K (-196°C).
Avant l'analyse d'adsorption, les échantillons ont été dégazés pendant 22 h à 300°C.
Le volume total des pores a été estimé à partir de la quantité adsorbée à 0,99 de
pressions relatives P/P0. Les distributions des tailles des pores du matériau ont été
calculées en utilisant la branche de désorption des isothermes d'adsorption/désorption
d’azote (N2), en appliquant la méthode Barrett-Joyner-Halenda, BJH (pour la SBA-15,
‘’S.9’’, ‘’S34.8’’ et les FDU-12) (Zhao et al., 1998, Zhou et al., 2009). Par contre la
méthode BJH n’est pas calibrée pour le calcul des tailles de pores sphériques larges
(FDU-12) qui conduit à une sous-estimation de diamètre des pores, (Huang et Kruk,
2012). D’autre part, la méthode Broekhoff-de Boer–Frenkel-Halsey-Hill, BdB–FHH a été
utilisée dans le cas MCF (Desch et al., 2014).

52
4.4.6.2. Diffraction des rayons X (DRX)
Les structures ordonnées des supports sont déterminées par diffractométrie de rayons X
(DRX). Les diffractogrammes des poudres des supports synthétisés ont été obtenus en
utilisant un appareil DRX Ultima III de Rigaku, Japon, avec le rayonnement Cu Ka (λ =
1.5406 Å). Les diffractogrammes de poudre aux petits angles ont été acquis à une
vitesse de 0,5°/min dans la plage 2θ de 0,6 à 3° de balayage.
4.4.6.3. Analyses de Microscopie Électronique à Transmission (MET)
Les analyses de microscopie électronique à transmission (MET) ont été réalisées avec
un appareil JEM-3010 (JEOL, Japon) fonctionnant à une tension d'accélération de 200
kV. Les échantillons des supports ont été suspendus dans le méthanol et traités par
ultrasons pendant 10 min. Ensuite, la suspension (5 µL) a été disposée uniformément,
puis elle a été séchée sur une grille de nickel. Cette analyse donne une idée sur la taille
et la forme des pores du matériau, ainsi que sa structure.
4.4.7. Immobilisation de β-Galactosidase
4.4.7.1. Immobilisation ‘Cross-linking’
La méthode ‘cross-linking’ utilisée pour l’immobilisation de β-Galactosidase suit le
protocole suivant : 10 mg du support siliceux ont été mélangés avec 1,5 mL de β-
galactosidase libre dans une solution tampon (tampon de phosphate de sodium 10 mM)
de concentration de 4 g/L. Le mélange a été agité au vortex pendant 30 secondes,
soumis à une sonication pendant 5 secondes, et puis incubé à température ambiante
sous agitation (250 rpm). Après une incubation de 20 min (phase d’adsorption), les
échantillons ont été brièvement lavés par un tampon de phosphate de sodium 100 mM,
puis incubés dans le même tampon contenant 0,1% de glutaraldéhyde (GA) à 200 rpm
pendant 30 min. Après le traitement avec le GA, les échantillons ont été lavés par un
excès de tampon de phosphate à 100 mM et stockés à 4°C (Kim et al., 2008).
Afin d’étudier la capacité de rétention de différents matériaux, l’immobilisation a été
effectuée avec 4 différentes concentrations de β-Galactosidase par rapport à la quantité
des supports (25, 60, 75 et 100% massique), 3 pH différents (3,5 ; 5 et 7) ainsi que 3
concentrations de glutaraldhyéde (0,1%, 1% et 2% v/v).

53
4.4.7.2. Rétention d’enzymes par les supports
Les enzymes non retenues sur les supports après chaque lavage ont été mesurées par
la méthode BCA (Smith et al., 1985) (méthode de dosage colorimétrique des protéines).
La concentration totale en protéines se manifeste par un changement de couleur de la
solution d'échantillon du vert au pourpre. Cette couleur pourpre absorbe fortement la
lumière à une longueur d'onde de 562 nm.
La solution de BCA de couleur verte, est composée de 50 volumes de réactif A et 1
volume de réactif B. Cette solution est stable pour une durée bien déterminée, environ
8h.
Le réactif A est composé de plusieurs produits à des concentrations bien
précises : 1% d’acide bicinchoninique, 2% de carbonate de sodium, 0,16% de
tartrate de sodium, 0,4% de soude, et 0,95% de bicarbonate de sodium.
Cette solution est fortement alcaline (pH = 11,25).
Le réactif B est composé de 4% de sulfate de cuivre pentahydraté.
La quantité de protéine présente dans une solution peut être quantifiée par une
spectrométrie d’absorption UV, calibrée avec des solutions protéiques de concentration
connue au préalable (Tableau 10).
Tableau 10. Préparation des standards à concentration protéique connue
Blanc 1 2 3 4 5
Étalon sérum albumine bovine à
1 mg/mL en NaCl 0,15 mol/L
0 µL 20 µL 40 µL 60 µL 80 µL 100 µL
Eau 100 µL 80 µL 60 µL 40 µL 20 µL 0 µL
Réactif Cu2+
/BCA du jour 2 mL
Quantité de protéines par tube
réactionnel
0 µL 20 µg 40 µg 60 µg 80 µg 100 µg
Les standards ainsi que les solutions à doser sont incubés pendant 2 h à température
ambiante ou 30 min à 37°C afin de favoriser la réaction, et par conséquent, mettre en
relief le changement de couleur. Ensuite, les échantillons sont placés dans des cuvettes
en plastique translucide de 1 cm de largeur, sur 3,5 cm de longueur compatible avec le
spectrophotomètre d’absorption UV visible, Genesys 20. A l’aide ce dernier, des

54
mesures d’absorbance, de transmittance (%), et de concentration (ppm, g/L, mg/ml)
peuvent être effectuées, à une longueur d’onde située entre 325 nm et 1100 nm. La
concentration en ppm des échantillons a été déterminée pour une longueur d’onde
optimale de 562 nm.
En se basant sur les concentrations des protéines dosées, la charge d’enzyme finale
immobilisée dans la silice a été calculée à partir de la différence entre la quantité initiale
de β-galactosidase et la quantité d’enzymes non retenues sur les supports, dosées dans
les solutions de lavage.
4.4.8. Réaction enzymatique avec l’enzyme immobilisée
(approche hétérogène)
Une fois que la préparation enzymatique est immobilisée sur les différents matériaux,
elle est placée dans une solution aqueuse de 50 mL à des concentrations équimolaires
de saccharose et de galactose. Cette réaction a été incubée, pendant 28h, à une
température de 40°C et un pH=5.
Durant ce temps, des prélèvements de 100 µL de chaque échantillon ont été prélevés et
dilués 100 fois. Ensuite, pour d’arrêter la réaction, les prélèvements ont été chauffés à
100°C pendant 10min. La même procédure est appliquée avec l’approche homogène.
La régénération et le recyclage des biocatalyseurs (β-galactosidase immobilisée sur
différents matériaux siliceux) ont été effectués en récupérant la matrice enzymatique par
filtration à la fin de chaque cycle et en la réintroduisant dans une nouvelle réaction avec
les mêmes conditions.
Tous les échantillons ont été analysés par chromatographie phase liquide à haute
performance, HPLC (voir la section 4.4.2).

55
4.4.9. Analyses statistiques
En utilisant la procédure ANOVA du logiciel SAS (1987), une étude statistique est
effectuée afin de déterminer s'il existe une différence significative entre les différents
paramètres étudiés, et si possible, quel est le paramètre le plus efficace dans cette
étude. Dans la plupart du temps, le plan factoriel en utilisant le test de Tukey est effectué
(Joiner et Rosenblatt, 1971).


57
Chapitre 5. Résultats et discussions
5.1. Réaction enzymatique
La réaction enzymatique a été étudiée selon deux approches: homogène (enzyme libre)
et hétérogène (enzyme immobilisée). La première approche consiste à étudier l’effet de la
concentration de l’enzyme libre, l’effet des différentes concentrations de substrats
(lactose et saccharose), ainsi que l’effet du pH et de la température sur la production du
lactosucrose. Par la suite, les conditions optimisées en phase homogène sont appliquées
à la réaction en phase hétérogène après optimisation des conditions d’immobilisation.
5.1.1. Identification des produits par HPLC, spectrométrie de
masse et par la RMN du proton
Suite à une réaction enzymatique utilisant le saccharose et le lactose en présence de la
β-galactosidase libre, un échantillon du milieu réactionnel a été prélevé puis a subit des
analyses par HPLC, spectrométrie de masse et par la RMN du proton pour pouvoir
procéder à l’identification des produits obtenus.
Comme il est anticipé la formation du glucose, du galactose, du fructose et des
oligosaccharides (GOS) comme le lactosucrose et la présence aussi d’une partie du
saccharose et du lactose qui n’a pas réagi dans le milieu réactionnel, il tout à fait naturel
de vérifier leur temps de rétention par HPLC en utilisant des standards purs.
Cependant, comme les standards du lactosucrose et d’autres GOS ne sont pas
commercialement disponibles, des méthodes puissantes d’identification de structure
chimique comme la spectrométrie de masse et la RMN du proton (1H-RMN) ont été
utilisées pour cette fin après une étape préalable de purification.
La Figure 19 présente le chromatogramme des standards disponibles utilisés comme
référence pour des fins d’identification avec leur temps de rétention des sucres simples et
les disaccharides comme le lactose et le saccharose.
Les temps de rétentions des standards sont présentés en minutes, comme suit :
saccharose (7,838 min), lactose (8,169 min), glucose (9,770 min), galactose (10,677 min) et

58
fructose (11,316 min). Quatre différentes concentrations de chaque solution ont été
préparées afin de tracer leurs courbes d’étalonnage. Les concentrations croissantes
utilisées sont les suivantes 0,05g/L ; 0,125g/L ; 0,5g/l et 1g/L.
Figure 19. Chromatogramme des solutions standards des sucres (0,5g/L chaque standard). Avec : a) saccharose, b) lactose, c) glucose, d) galactose, e) fructose.

59
Figure 20. Chromatogramme typique des substrats et des produits formés par transgalactosilation et hydrolyse.

60
Glc Fru Glc Gal
LactoseSaccharose
B-Galactosidase
d'Aspergillus oryzae
Glc Fru Glc Gal
Lactose
+
Glc FruGal ++ +
FruGlc Gal
Lactosucrose Tetrahexose
+
GOS I (produit désiré) GOS II (produits secondaire)
B-Galactosidase
d'Aspergillus oryzae
Etape d'hydrolyse
Etape de transglycosylation
Saccharose
Figure 21. Mécanisme proposé pour la biosynthèse de lactosucrose avec la β-Galactosidase d’Aspergillus oryzae.

61
Dans l’espoir d’identifier les pics inconnus attribués à la présence des GOS (I et II). Les
produits de la réaction ont été fractionnés par UPLC et les produits dus aux pics GOS I et
GOS II ont été récupérés puis concentrés par évaporation avec un évaporateur rotatif et
ont été sujets aux analyses de spectrométrie de masse et par 1H-RMN et 13C-RMN.
En effet, Le spectre de masse du GOS I tel que montré par la Figure 22 montre la
fragmentation majeure [M-2H] 2- du lactosucrose déprotoné à m/z=502 (504-2) qui
correspond à la masse moléculaire exacte du la molécule ciblée du lactosucroce avec la
distribution isotopique. Ces résultats ont été confirmés par les analyses RMN du proton et
du carbon de l’échantillon qui révèlent la structure désirée du lactosucrose avec une très
haute pureté.
Figure 22. Spectre MS du GOS I.
m/z
150
200
250
300
350
400
450
500
550
600
650
700
750
800
850
900
950
1000
% 2
ZANE
IB_M
AI14
_PIC
2_2
1 (0
.002
)Sc
an E
S-
5.43
e650
2.7
178.
6
211.
734
0.9
245.
638
2.2
443.
056
6.2
601.
1

62
La structure du composé GOS I a été élucidée par l’analyse 1H-RMN et 13C-RMN.
L’analyse 13C-RMN révèle 18 pics correspondant à des carbones incluant des carbones
de sucres anomériques vers les déplacements chimiques de 105.80, 92.81 et 106.80
ppm et 4 carbones methylene oxylés vers les déplacements chimiques de 62.70, 61.39.,
61.76 et 64.39 ppm, indiquant la présence de trois hexoses dans la structure du composé
(Figure 23).
Figure 23. Spectre 13C-RMN du GOS I.
L’analyse 1H-RMN confirme que les 3 hexoses consistent en des galactopyranose,
glucopyranose et un fructopyranose (Figure 24). Les résultats de ces analyses
suggèrent que le composé en question est un O--galactopyranosyl-(1→4)-O--D-
glucopyranosyl-(1→2)--D-fructofuranoside, communément appelé lactusucrose. Les
assignements entiers des spectres 1H-RMN sont montrés dans le Tableau 11.

63
Figure 24. Spectre 1H- RMN du GOS I.
O O
O
O
O
OH HO
OH
OH
OH
OHHO
HO
HO
HO
OH

64
Tableau 11. Récapitulatif des déplacements chimiques du GOS I analysé par résonance magnétique nucléaire (1H-RMN) et identifié comme du lactosucrose.
O O
O
O
O
OH HO
OH
OH
OH
OHHO
HO
HO
HO
OH
1
23
4
6
51'2'
3'4'
5'
6'
1''
2''3''
4''
5''
6''
lactosucrose
C18H32O16
Mol. Wt.: 504,44
1H RMN (500 MHz, D2O, 25°C):
1H-RMN
Fructose
Position des
protons
(ppm) Nature du pic J (Hz) Nombre de protons
associés au pic
1 5.26 T 3.1 1H
2 4.06-4.03 Dd 8.5, 3.5 1H
3 5.06 D 3.5 1H
4 - - - -
5 4.16 S 2H
6 4.18 S 2H

65
Glucose
1’ 5.40 D 4.1 1H
2’ 4.33-4.24 M 1H
3’ 3.54-3.48 M 1H
4’ 4.52-4.46 M 1H
5’ 4.44 D 6.5 1H
6’ 4.35 D 7.4 2H
Galactose
1’’ 5.24 D 3.9 1H
2’’ 4.02-3.84 M 1H
3’’ 3.42-3.36 M 1H
4’’ 3.74-3.57 M 1H
5’’ 3.29 T 9.1 1H
6’’ 3.76 D 3.0 2H
Donc toutes ces analyses confirment que le composé GOS I correspond à la présence du
lactosucrose.
Le composé GOS II a été analysé par spectrométrie de masse. Son Spectre MS est
montré dans la Figure 25.

66
Figure 25. Spectre MS du GOS II.
m/z
150
200
250
300
350
400
450
500
550
600
650
700
750
800
850
900
950
1000
%
3
ZA
NE
IB_M
AI1
4_P
IC1_2 1
(0.0
02)
Scan E
S-
3.7
8e6
664.5
340.7
200.1
173.4
227.1
322.2
503.7
428.3
367.4
588.2
732.2
687.2
800.7
754.5
890.9
843.8
816.6
869.6
965.2
905.7

67
Cette analyse révèle que le GOS II s’identifie à un tetrahexose dont m/z = 664.5
5.2. Réaction enzymatique en phase homogène
5.2.1. La cinétique de la réaction enzymatique
La Figure 26 montre une cinétique typique de la réaction enzymatique, qui représente la
conversion du lactose et du saccharose conduisant à la formation des oligosaccharides
sous l’effet biocatalytique de β-Galactosidase d’Aspergillus oryzae. Dans les premières
24 heures, le taux de formation des GOS a augmenté, avec une diminution rapide de la
concentration du lactose et une diminution moyenne de la concentration du saccharose.
En effet, cette différence de diminution est due à la contribution du lactose dans la
formation des GOS et dans la formation des monosaccharides après hydrolyse. Par
contre, le saccharose contribue majoritairement à la formation de tetrahexose . Il est clair
que la transgalactosylation domine au début de la réaction, favorisant ainsi la production
des GOS avec un rendement élevé. Cependant, l'hydrolyse des GOS formés prend forme
au fur et à mesure de l’avancement de la réaction. On remarque que la concentration en
GOS diminue après 24 heures de réaction, alors que les concentrations du glucose et du
galactose continuent à augmenter avec une conversion en glucose supérieure à celle en
galactose. En même temps, on observe que la quantité des disaccharides continue à
diminuer, ce qui s’explique par l’hydrolyse de ces disaccharides en monosaccharides
(glucose, galactose et fructose). Quand on a laissé la réaction se poursuivre, seuls les
produits d'hydrolyse (glucose et galactose) et le saccharose étaient retrouvés dans la
solution du produit final (Annexe1). Cette diminution des oligosaccharides formés peut
être attribuée à une inhibition compétitive de l’enzyme par le glucose et le galactose issus
de l’hydrolyse. Albayrak et Yang (Albayrak et Yang, 2002) ont confirmé que les
monosaccharides formés par l’hydrolyse de lactose (glucose et galactose) ne sont pas
avantageux pour la formation des GOS, en particulier du lactosucrose. Il faut donc
essayer d’enlever ces monosaccharides du milieu réactionnel à fur et à mesure pour
déplacer l’équilibre vers la synthèse des oligosaccharides. En effet, l’effet inhibiteur de
galactose sur la β-Galactosidase d’Aspergillus oryzae est souvent démontré majoritaire
beaucoup plus que celui du glucose.(Boon et al., 2000, Park et al., 1979)

68
Figure 26. Cinétique de la réaction de conversion des substrats et la formation des produits.
5.2.2. Effet de la concentration enzymatique
L’étude de l’effet de concentration enzymatique sur la synthèse des GOS catalysée par
une préparation commerciale de β-galactosidase à partir de A. oryzae (Sigma-Aldrich) a
été réalisée avec un mélange réactionnel de disaccharides à des concentrations
identiques (300 g/L de lactose, et 300 g/L de saccharose),(Li et al., 2009) avec des
concentrations enzymatiques différentes (1U / ml de solution, 4U / ml de solution, 6U / ml
de solution et 10U / ml de solution) pendant 60 heures.
0
50
100
150
200
250
300
350
0 10 20 30 40 50 60 70
Co
ncen
trati
on
(g
/L)
Temps (h)
saccharose (g/l)
lactose (g/l)
glucose (g/l)
galactose (g/l)
fructose (g/l)
GOS I
GOS II
Total GOS

69
Figure 27. Effet de la concentration d'enzyme sur la production du lactosucrose.
0
10
20
30
40
50
60
70
80
90
0 10 20 30 40 50 60 70
Co
ncen
trati
on
(g
/l)
Temps (h)
Lactosucrose1U/ml
Lactosucrose4U/ml
Lactosucrose6U/ml
Lactosucrose10U/ml

70
Figure 28. Effet de la concentration d'enzyme sur la production du tetrahexose.
0
2
4
6
8
10
12
14
16
18
0 10 20 30 40 50 60 70
Co
nc
en
tra
tio
n (
g/L
)
Temps (h)
Tetrahexose 1U/ml
Tetrahexose 4U/ml
Tetrahexose 6U/ml
Tetrahexose 10U/ml

71
Figure 29. Effet de la concentration d'enzyme sur la bioconversion du lactose.
Les Figure 27Figure 28 et Figure 29Erreur ! Source du renvoi introuvable. montrent,
que l’augmentation de la concentration d’enzyme de 1 à 6 U/mL entraine une
augmentation dans la production des oligosaccharides et dans la conversion de lactose
ainsi qu’une augmentation dans la vitesse de la réaction (Li et al., 2009). Quand la
concentration en enzymes est augmentée à 10U/mL, on observe une diminution de la
production des GOS qui serait due à une inhibition de la réaction de transgalactosylation
pour la synthèse des GOS par la concentration trop élevée de l’enzyme ou par la
formation des produits eux-mêmes.
Les courbes de formation du lactosucrose, à part celles avec une concentration
enzymatique de 1U/mL, présentent des maximas de production après 24 h de temps de
réaction. Au-delà de ce temps la production en se produit diminue. La courbe
représentative de la concentration 1U/mL montre une augmentation quasi-linéaire avec
le temps de réaction qui fait penser que la cinétique de transgalactosylation pour ce
produit n’est pas inhibée à ce niveau de concentration de l’enzyme.
Avec une concentration de 4U/mL, le temps nécessaire pour atteindre le maximum de
concentration du lactosucrose (64,4 g/l) était de 48h et de 36h dans le cas du tetrahexose
(12,8 g/l). Cependant, ce temps est réduit jusqu’à 24h pour la synthèse du lactosucrose
0
50
100
150
200
250
300
350
0 10 20 30 40 50 60 70
Co
nc
en
tra
tio
n (
g/L
)
Temps (h)
lactose 1U/ml
lactose 4U/ml
lactose 6U/ml
lactose 10U/ml

72
avec les concentrations 6U/mL et 10U/mL dont les quantités maximales produites sont
respectivement 76,3 g/l et 63,7 g/l. La concentration enzymatique de 6U/mL est la
concentration optimale de synthèse des GOS avec la meilleure vitesse de réaction et le
meilleur rendement des GOS estimé à 30,6% (avec une composition de 84% du
lactosucrose et 16% du tetrahexose).
-Selon les courbes de transformation du lactose, la conversion du lactose évolue
l’augmentation de la concentration d’enzyme. Ceci prouve que la réaction de conversion
du lactose est bien favorisée par la concentration enzymatique de 10U/mL et elle est
totale (100% de conversion de lactose) à partir de 60 heures. Néanmoins, les résultats
présentés ci-dessus montrent que la concentration de 10U/mL vient en deuxième position
après les 6U/mL dans la synthèse des GOS. A partir de ces observations, nous pouvons
conclure que la concentration de 10U/mL contribue plus que les autres concentrations
enzymatiques dans la réaction d’hydrolyse de lactose en déplaçant l'équilibre de la
réaction de β-galactosidase à l'hydrolyse plutôt qu’à la transglycosylation. La
concentration de 10U/mL peut avoir donc, un effet inhibiteur dans la formation des GOS
de par son activité d’hydrolyse qui serait favorisée à ce niveau de concentration. Cette
activité élevée d’hydrolyse peut être attribuée à la disponibilité d’un nombre très élevé de
sites enzymatiques libres à 10U/mL, qui favorisent et accélèrent la formation des GOS, et
par la suite leur hydrolyse rapide ainsi que l’hydrolyse du lactose (Figure 30 et Figure
31). Alors, qu’avec les concentrations faibles de 1U/mL, la vitesse de production des
GOS est faible où aucune formation du tetrahexose n’est observée avant 24 h de
reaction. L’hydrolyse pour l’obtention des monosaccharides étant nécessaire pour la
formation du tetrahexose, d’où l’apparition tardive de ce dernier avec la concentration
enzymatique 1U/mL.
Ces résultats confirment que la concentration en enzyme 6U/mL et le temps de réaction
de 24 heures sont les conditions optimales pour la réaction de transgalactosylation,
favorisant la synthèse des GOS et limitant la réaction d’hydrolyse de lactose. Dans ces
conditions optimales, la conversion du lactose et de saccharose étaient respectivement :
75% et 29%.

73
Figure 30. La formation de galactose (produit d'hydrolyse de lactose) avec différentes concentrations d'enzyme.
Figure 31. La formation de glucose (produit d'hydrolyse de lactose avec différentes concentrations d'enzyme.
0
20
40
60
80
100
120
140
0 10 20 30 40 50 60 70
Co
nce
ntr
atio
n (
g/L)
Temps (h)
galactose (g/l) 1U
galactose (g/l) 4U
galactose (g/l) 6U
galactose (g/l) 10U
0
20
40
60
80
100
120
140
160
0 20 40 60
Co
nce
ne
trat
ion
(g/
L)
Temps (h)
glucose (g/l) 1U
glucose (g/l) 4U
glucose (g/l) 6U
glucose (g/l) 10U

74
5.2.3. Effet de concentration des substrats
Afin de déterminer le rapport molaire optimal du donneur / accepteur de galactosyls, ainsi
que la concentration favorable des substrats pour la production du lactosucrose et du
tetrahexose, on a utilisé différentes concentration de saccharose (50% ; 30% ; 25% ; 15%
et 7,5%) et de lactose (50% ; 30% ; 15%) avec deux ratios molaires de lactose /
saccharose; 1:1 et 2:1 (Tableau 12). Les réactions sont effectuées à 30°C, à pH=5 et
avec la concentration d’enzyme optimisée de 6U/mL.
Tableau 12. Les différentes concentrations de saccharose et de lactose étudiées pour la biosynthèse de lactosucrose
Réaction
Rapport molaire 1 :1
Réaction
Rapport molaire 2 :1
Lactose
(g/100mL)
Saccharose
(g/100mL)
Lactose
(g/100mL)
Saccharose
(g/100mL)
A 15 15 D 50 25
B 30 30 E 30 15
C 50 50 F 15 7,5
Les concentrations initiales du saccharose (Figure 32 et Figure 33) et du lactose (Figure
34 et Figure 35) affectent clairement le rendement en GOS totaux. En balayant les
différentes concentrations des deux sucres, on remarque que le meilleur rendement est
celui à 30% pour les deux rapports molaires. La concentration de 50% du lactose avec
les deux ratios molaires occupe la deuxième position en termes de rendement des GOS.
Cela s’explique par la réaction d’hydrolyse et la saturation des sites enzymatique, vu
l’accumulation des substrats et leur disponibilité pour l’hydrolyse. La concentration de
15%, avec les deux rapports molaires, s’est révélée insuffisante et faible par rapport à la
concentration de l’enzyme. Ce rendement faible observé en GOS s’explique par la
formation des GOS sur des sites enzymatiques et l’hydrolyse rapide des autres GOS déjà
formés par d’autres sites enzymatiques.

75
Figure 32. Effet de la concentration initiale de lactose avec le rapport molaire des substrats 1:1 sur la production des GOS totaux.
Figure 33. Effet de la concentration initiale de saccharose avec le rapport molaire des substrats 2:1 sur la production des GOS totaux.
0
5
10
15
20
25
30
35
0 10 20 30 40 50
Re
nd
em
en
t d
e G
OS
% de conversion de saccharose
50%
30%
15%
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
Re
nd
em
en
t d
e G
OS
% de conversion de saccharose
25%
15%
7,50%

76
Figure 34. Effet de la concentration initiale de lactose avec le rapport molaire des substrats 1:1 sur la production des GOS totaux.
Figure 35. Effet de la concentration initiale de lactose avec le rapport molaire des substrats 2:1 sur la production des GOS totaux.
0
5
10
15
20
25
30
35
0 20 40 60 80 100
Re
nd
em
en
t d
e G
OS
%conversion de lactose
50%
30%
15%
0
5
10
15
20
25
30
0 20 40 60 80 100
Ren
de
me
nt
GO
S
%conversion de lactose
50%
30%
15%

77
En effet, le rendement maximal du lactosucrose (24% correspondant à une production de
73 g/l) a été obtenu au rapport molaire de 1:1 au bout de 24 heures (Figure 36). Cette
production présente 1,3 fois (4/3) la quantité obtenue avec les rapports molaires 2:1. Ces
résultats sont en concordance avec les résultats de (Li et al., 2009). et (Mussatto et
Mancilha, 2007), qui confirment que l’optimisation du rendement en lactosucrose
nécessite la présence de réactifs initiaux (lactose et saccharose) en concentrations
équimolaires. Cette relation entre l’équimolarité des substrats et le rendement en
lactosucrose peut être justifiée par une faible activité d'hydrolyse par rapport à une
activité élevée de transgalactosylation dans ces conditions. Concernant le rendement en
tetrahexose (Figure 37), la concentration en lactose à 30% donne un meilleur rendement
mais avec un rapport molaire de 2:1 cette fois. Le rendement maximal du tetrahexose
(5,38%) est 1,25 (5/4) fois plus élevé que celui obtenu avec un rapport molaire de 1:1. Le
rapport molaire affecte la cinétique de la réaction et de formation du tetrahexose qui est
produit plus rapidement (on passe de 24h pour un rapport molaire de 1:1 à 16 heures
pour un rapport de 2:1). Ces résultats peuvent s’expliquer par la formation des
monosaccharides (glucose, galactose et fructose) lors de la réaction d’hydrolyse,
favorisant la production du tetrahexose. Cependant, lorsque la concentration de lactose
dépasse les 30%, la formation de ces produits secondaires (les monosaccharides)
devient plus élevée (Figure 38 et Figure 39), et donc plus compétitive et inhibitrice à la
production des GOS (particulièrement du lactosucrose) (Albayrak et Yang, 2002, Boon et
al., 2000, Li et al., 2009).

78
Figure 36. Effet de la concentration des réactifs initiaux sur le rendement du lactosucrose.
Figure 37. Effet de la concentration des réactifs initiaux sur le rendement du tetrahexose.
0
5
10
15
20
25
30
35
0 5 10 15 20 25 30
Re
nd
em
en
t la
cto
su
cro
se
(%
)
Temps (h)
Rendement GOS I %(50% - 1:1)
Rendement GOS I %(30% - 1:1)
Rendement GOS I %(15% - 1:1)
Rendement GOS I %(50% - 2:1)
Rendement GOS I %(30% - 2:1)
Rendement GOS I %(15% - 2:1)
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
Ren
de
men
t t
etr
ah
exo
se (
%)
Temps (h)
Rendement GOS II % (50% -1:1)
Rendement GOS II % (30% -1:1)
Rendement GOS II % (15% -1:1)
Rendement GOS II % (50% -2:1)
Rendement GOS II % (30% -2:1)
Rendement GOS II % (15% -2:1)

79
Figure 38. Effet de la concentration et les rapports molaires sur la formation du galactose.
0
10
20
30
40
50
60
70
0 5 10 15 20 25 30
Gal
acto
se g
/L
Temps (h)
galactose (g/l)(50%-1:1)
galactose (g/l)(30%-1:1)
galactose (g/l)(15%-1:1)
galactose (g/l)(50%-2:1)
galactose (g/l)(30%-2:1)
galactose (g/l)(15%- 2:1)

80
Figure 39. Effet de la concentration et les rapports molaires sur la formation du glucose.
5.2.4. Effet du pH
La Figure 40 montre l’effet de pH, qui semble minime, sur la conversion de lactose ainsi
que la production totale des GOS (Figure 41). Le changement de pH entre 7 ; 5 et 3,5
affecte la vitesse de la réaction de conversion de lactose plus que la formation de GOS,
ce qui est en accord avec la littérature (Vera et al., 2011).
En effet, le pH 5 présente le meilleur taux de conversion de lactose et de rendement en
lactosucrose et tetrahexose (Figure 40 et Figure 41), ensuite on trouve les pH 7 et 3,5
avec presque les mêmes rendements de transformation.
Ces résultats s’expliquent par le point isoélectrique de l’enzyme qui constitue le point
pour lequel l’enzyme est électriquement neutre. A l’approche de celui-ci, l’activité de
l’enzyme diminue considérablement, ce qui est le cas pour la β–galactosidase. Le pH=3,6
est indiqué par (Bernal et al., 2014) comme le point isoélectrique de la β-galactosidase
isolée à partir de l’Aspergillus oryzae. D’où, le rendement le plus faible au pH =3,5 en
GOS. Le rendement à pH=7 est plus faible que celui à pH=5 car il s’agissait d’un pH
neutre et la β-galactosidase a une activité favorisée avec des pH un peu plus acides
0
20
40
60
80
100
120
0 5 10 15 20 25 30
Glu
cose
(g/
L)
Temps (h)
glucose (g/l)(50%-1:1)
glucose (g/l)(30%-1:1)
glucose (g/l)(15%-1:1)
glucose (g/l)(50%-2:1)
glucose (g/l)(30%-2:1)
glucose (g/l)(15%- 2:1)

81
mais aussi un peu loin du point isoélectrique. Ces résultats sont en accords avec les
résultats de (Freitas et al., 2011), qui annoncent que le pH 4,8 fournit la meilleure activité
et stabilité enzymatique avec les β–galactosidases (Aspergillus oryzae).
Figure 40. Effet du pH sur la conversion de lactose.
0
50
100
150
200
250
300
350
0 5 10 15 20 25 30
co
nc
en
tra
tio
n g
/l
temps (h)
lactose (g/l) pH 7
lactose (g/l) pH 5
lactose (g/l)pH 3,5

82
Figure 41. Effet du pH sur le rendement en GOS totaux.
5.2.5. Effet de la température
Afin de déterminer l’effet de la température sur la conversion du lactose et la production
de GOS, la biosynthèse a été effectuée à trois températures différentes (30°C ; 40°C et
50°C). En utilisant les paramètres déjà optimisés; avec une concentration de 60% de
lactose et saccharose ayant un rapport équimolaire, pH=5 et une concentration
enzymatique de 6 U/mL.
Comme le montre la Figure 42, la vitesse de conversion du lactose dépend étroitement
de la température, incluant la transformation par hydrolyse ainsi que la production des
GOS. Un excellent rendement est atteint à 40°C et ce rapidement, (au bout de 12
heures), mais le rendement optimal est observé à 30°C après 24 heures (Figure 43). La
réaction à 40°C est très efficace pour former les GOS en 12 heures seulement.
Cependant, à cette température l’enzyme est très active pour l’hydrolyse aussi, ce qui
conduit à l’hydrolyse des produits formés par la suite.
0
10
20
30
40
50
60
70
80
90
100
0 5 10 15 20 25 30
co
nc
en
tra
tio
n g
/l
temps (h)
Total GOS pH 7
Total GOS pH 5
Total GOS pH 3,5

83
A une température de 30°C, l’hydrolyse est minimale, ce qui conduit à une production
optimale en GOS. La formation du lactosucrose est optimale à 30°C (Figure 44), et est
accompagnée par une activité d’hydrolyse minimale. Cependant l’hydrolyse est maximale
à 40°C, ce qui par contre favorise la production du tetrahexose (Figure 45), suite à la
formation des monosaccharides nécessaires à son synthèse.
Il est très important de mentionner que la synthèse du lactosucrose n’est pas beaucoup
affectée par l’augmentation de la température (même à 50 °C), ce qui est en
concordance avec les résultats cités par Albayrak et Yang(Albayrak et Yang, 2002) et par
Vera et al(Vera et al., 2011). Cependant, le rendement du tetrahexose subi une chute
significative à 50°C. Ceci peut être attribué à l’inhibition totale de l’activité d’hydrolyse
enzymatique, et donc l’absence des monosaccharides nécessaires à la formation du
tetrahexose. Toutefois, la température de réaction est critique pour la synthèse
enzymatique. Des résultats similaires ont été rapportés par (Li et al., 2009)concernant
l’effet de la température sur les produits de β-galactosidase.
Figure 42. Effet de la température sur la conversion du lactose.
0
50
100
150
200
250
300
350
0 5 10 15 20 25 30
La
cto
se
(g
/L)
Temps (h)
lactose (g/l) 30°C
lactose (g/l) 40°C
lactose (g/l) 50°C

84
Figure 43. Effet de la température sur la formation des GOS.
Figure 44. Effet de la température sur le rendement en lactosucrose.
0
10
20
30
40
50
60
70
80
90
100
0 5 10 15 20 25 30
Re
nd
em
en
t G
OS
%
Temps (h)
Total GOS 30°C
Total GOS 40°C
Total GOS 50°C
0
5
10
15
20
25
30
0 20 40 60 80 100
Ren
de
me
nt
lac
tos
uc
ros
e %
% Conversion lactose
30°C
40°C
50°C

85
Figure 45. Effet de la température sur le rendement en tetrahexose.
5.2.6. Conclusion
Ces résultats indiquent que la formation des différents oligosaccharides par la même
enzyme ne dépend pas seulement de l’enzyme, mais aussi des conditions de la réaction
qui peuvent favoriser la synthèse d’un ou de plusieurs produits.
Après optimisation des paramètres réactionnels (concentration d’enzyme, concentration
et ratios molaires des substrats, pH et température), nous avons effectué la biosynthèse
de lactosucrose à partir du lactose et du saccharose avec l’action enzymatique de β-
Galactosidase d’Aspergillus oryzae, dans les conditions optimisées (6U/mL, une
concentration équimolaire (1 :1) de saccharose (30%) et de lactose (30%), pH= 5 et
T°= 40°C). Sous ces conditions optimales, nous avons obtenu un rendement de 27,47%
des GOS avec une composition d’environ 84% du lactosucrose (68,2 g/L) et 16% du
tetrahexose (13,3 g/L), c’est-à-dire un rendement de 23% en lactosucrose. Ce rendement
obtenu est supérieur à celui atteint par (Li et al., 2009), et confirment la formation de
lactosucrose à une concentration de 50.4 g/L (rendement de 17%), après 4h de réaction
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
0 20 40 60 80 100
Re
nd
em
en
t te
tra
he
xo
se
%
% Conversion lactose
30°C
40°C
50°C

86
en utilisant la β-D-Galactosidase de Bacillus circulans. L’activité observée dans le présent
travail avec la β-Galactosidase d’Aspergillus oryzae s’est révélée plus sélective en
trisaccharides, par rapport à celle déjà obtenue précédemment dans la littérature pour la
bioconversion de lactose (Albayrak et Yang, 2002, Gaur et al., 2006, Güleç et al., 2010,
Huerta et al., 2011, Neri et al., 2011).

87
5.3. Réaction enzymatique en phase hétérogène
Dans cette section, on discute, séparément, la synthèse et la caractérisation des
matériaux et l’étude de la rétention d’enzyme (β-galactosidase d’A. oryzae) sur les
différents matériaux préparés. Ensuite, on étudie l’activité catalytique de chaque
biocatalyseur sur la réaction de transglycosylation, ainsi que leurs performances après
recyclage.
5.3.1. Caractérisation des matériaux
5.3.1.1. SBA-15
5.3.1.1.1. Analyse de la surface BET
Les isothermes d'adsorption/désorption d'azote et la distribution correspondante des
tailles des pores obtenus sont présentées dans la Figure 46. Le support siliceux (SBA-
15) présente une surface specifique BET de ≈ 901.9 m2/g, un volume total des pores de
1.31cm3/g, ainsi qu’un diamètre de pore moyen d’environ 6.8 nm. Ces résultats sont
comparables aux résultats rapportés dans la littérature pour ce type de matériaux
(Belkacemi et al., 2006, Zhao et al., 1998). L’isotherme de ces matériaux est de type IV
avec une hystérèse de type H1, qui caractérise les matériaux mésoporeux (Lynch, 2001).

88
Figure 46. Les isothermes d'adsorption/désorption d'azote pour la SBA-15. La figure insérée représente la distribution de la taille des pores.
5.3.1.1.2. Analyse par diffraction des rayons X (DRX)
Le diffractogramme (DRX) obtenu aux petits angles pour la SBA-15 synthétisée (Figure
47), montre un pic majeur de diffraction (100) à l’angle 2θ = 1,0°, confirmant une
structure ordonnée de ce matériau. On observe également la présence de deux petits
pics supplémentaires à 2θ = 1,6° et 2θ = 1,8°, caractéristiques des diffractions (110) et
(200), respectivement, affirmant une structure mésoporeuse hexagonale (Belkacemi et
al., 2006, Zhao et al., 1998).

89
1 2 3
2 Thêta (degrés)
I (u. a.)
SBA-15100
110 200
Figure 47. Le diffractogramme (DRX) obtenu aux petits angles obtenus pour la SBA-15.
5.3.1.1.3. Microscopie Électronique à Transmission (MET)
Les images MET de la SBA-15 (Figure 48) sont en concordance avec les résultats DRX
et BET. Ils confirment la forme hexagonale et la mésostructure ordonnée du matériau.
Les images obtenues montrent des mailles répétitives avec une taille des pores d’environ
7nm, ainsi que des canaux assemblés en grappes parallèles visibles lorsqu’on change
l’angle du vue. Ces résultats sont satisfaisants et sont comparables avec les travaux de
(Zhao et al., 1998) et aussi avec ceux rapporté par (Belkacemi et al., 2006).

90
Figure 48. Micrographes MET de la SBA-15 observé de différents angles.
5.3.1.2. Mousse mésocellulaire (MCF)
5.3.1.2.1. Analyse de la surface BET
Les isothermes d'adsorption / désorption d'azote et la distribution correspondante de la
taille des pores obtenus sont présentés dans la Figure 49. Les calculs ont été réalisés à
l’aide du model de BdB-FHH (Méthode d'analyse de la porosité de Broekhoff de Boer,
Frenkel Halsey Hill) (Desch et al., 2014). L’aire de la surface BET calculée est de ≈ 487.2
m2/g, le volume total des pores est considérablement large environ 0.52 cm3/g, et le
diamètre de pore large est d’environ 20.0 nm. L’isotherme de la MCF est de type IV avec
une boucle hystérèse large de type H2. Ces résultats montrent la présence des pores
sphériques.

91
Figure 49. Les isothermes d'adsorption/désorption d'azote pour la MCF. La figure insérée représente la distribution de la taille des pores.
5.3.1.2.2. Analyse par diffraction des rayons X (DRX)
Le diffractogramme des rayons X de la MCF aux petits angles montre des pics bien
résolus (Figure 50). Ces derniers donnent l’information sur la forme structurale et
sphérique de matériau. Le diffractogramme montre deux pics majeurs à 2θ = 0,7° et deux
petits pics à 2θ = 1,0° et à 2θ = 1,2°.Ces résultats sont en concordance avec ceux
obtenus par (Schmidt-Winkel et al., 1999) Cette structure est révélée sphérique
monodispersée, ce qui a été prouvé avec des simulations de ces diffractogrammes
(Feigin et al., 1987).

92
1 2 3 4
I (u. a.)
2 Thêta (degrés)
MCF
Figure 50. Le diffractogramme (DRX) obtenu aux petits angles obtenus pour la MCF.
5.3.1.2.3. Microscopie Électronique à Transmission (MET)
Les images MET (Figure 51) affirment la forme sphérique obtenue par les analyses DRX
et la structure ordonnée du matériau. On distingue bien des sphères creuses de
diamètres ≈ 20 nm. en concordance avec la morphologie observée par (Schmidt-Winkel
et al., 1999)

93
Figure 51. Micrographe d’électron de transmission de la MCF (20 nm).

94
5.3.1.3. Nanomatériaux sous forme de barres (rod-like) S-2,9 et sous forme
de vésicules (vesicle-like) S-34,8
5.3.1.3.1. Analyse de la surface BET
Les isothermes d'adsorption / désorption d'azote et la distribution correspondante des
tailles de pores obtenues sont présentées dans la Figure 52 (S-2,9) et la Figure 53
(S-34,8). Les calculs ont été réalisés à l’aide du model de Barrett-Joyner-Halenda (BJH)
(Zhou et al., 2009) pour la S-2,9 et pour la S-34,8. Le model BJH pour ce matériau donne
une taille moyenne des pores de 3,8 nm largement au-deçà de la taille réelle observée
par le TEM. L’aire de la surface BET résultante est de 376 ,7 m2/g pour la S-2,9 et de
458,1 m2/g pour la S-34.8, le volume total des pores est de 0.60 cm3/g pour la S-2,9 et
0,62 cm3/g pour la S-34,8. Le diamètre des pores est de 7,7 nm pour la S-2,9 et
d’environ 17 nm pour la S-34,8. L’isotherme de la S-2,9 est de type IV avec une boucle
d’hystérèse de type H1, qui est similaire à celle de la SBA-15. Par contre, la S-34,8
présente une large boucle d’hystérèse.
Figure 52. Les isothermes d'adsorption/désorption d'azote pour la S-2.9. La figure insérée représente la distribution de la taille des pores.

95
Figure 53. Les isothermes d'adsorption/désorption d'azote pour la S-34,8. La figure insérée représente la distribution de la taille des pores.
5.3.1.3.2. Analyse par diffraction des rayons X (DRX)
Pour la synthèse de ce matériau deux concentrations de TIPB sont utilisées: une petite
quantité dont le rapport molaire TIPB:P123=2,9:1 (noté S-2,9) et une quantité importante
avec un rapport molaire de TIPB:P123=34,8:1 (noté S-34,8).
Les diffractogrammes des deux matériaux ne sont pas très différents. Le matériau ayant
une faible concentration en TIPB, le S-2,9 (Figure 54) présente un diffractogramme avec
quatre pics caractéristiques à 2θ ≈ 0,7° (100), à 2θ ≈ 1,3° (110), à 2θ ≈ 1,5° (200), et à 2θ
≈ 2,0° (210).Ces pics reflètent la structure mésoporeuse ordonnée hexagonale du
matériau synthétisé dans le cadre de ce travail.
La grande quantité de TIPB introduite dans le deuxième matériau a conduit à une
modification au niveau de la structure du matériau (Figure 55). Un pic de diffraction bien
résolu est observé à 2θ ≈ 0,5 (001) et un autre petit pic est observé à 2θ = 0,9 (002)
après l’analyse DRX du matériau S-34,8.
Tous ces résultats sont en concordance avec les résultats obtenus par Zhou et ses
collègues (Zhou et al., 2007, Zhou et al., 2009)

96
1 2 3
I (u. a.)
2 Thêta (degrés)
TIPB:P123 / 2,9:1 (S-2,9)100
110
200
210
Figure 54. Le diffractogramme (DRX) obtenu aux petits angles pour S-2,9.

97
0 1 2 3
2 Thêta (degrés)
I (u. a.)
TIPB:P123 / 34,8:1 (S-34,8)
001
002
Figure 55. Le diffractogramme (DRX) obtenu aux petits angles pour S-34,8.
5.3.1.3.3. Microscopie Électronique à Transmission (MET)
Les images MET des deux matériaux montrent clairement la morphologie ainsi que la
structure de ces deux matériaux. On distingue, à partir de la Figure 56 la structure
hexagonale ordonnée du premier matériau ainsi que la forme de barres d’où la
nomination silice mésoporeuse en forme de barres (Rod-like). Selon les images MET la
taille de diamètre des pores est d’environ 7-8nm. Cependant, l’ajout excessif de TIPB a
mené à un changement radical aussi bien au niveau de la forme qu’au niveau de la
structure de la silice. La forme de barres et de mailles hexagonales se transforme en
forme de vésicules en forme ‘ de rondelles d’oignon’ (Figure 57), avec des couches de
silices régulièrement espacées (entre 16-17nm). Ces résultats sont très satisfaisants, ils
sont similaires à ceux présentés par Zhou et son groupe de recherche (Zhou et al., 2007,
Zhou et al., 2009).

98
Figure 56. Micrographe MET des silices sous forme de barres (S2,9 – 7,7 nm).
Figure 57. Micrographe MET des silices sous forme des vésicules (S-34,8 – 16,7 nm).

99
5.3.1.4. Les LP-FDU-12
5.3.1.4.1. Analyse de la surface BET
Les isothermes d'adsorption / désorption d'azote et la distribution correspondante des
tailles de pores obtenues sont présentées dans les Figure 58, Figure 59 et Figure 60,
respectivement, la S-10-100, la S-15-100 et la S-T-100. Une tentative de calculs a été
faite à l’aide du model de BdB-FHH. Cependant, ce modèle ne converge pas pour ces
matériaux. La méthode BJH ne permet pas non plus d’estimer la distribution de la taille
des pores. Les aires de surface BET obtenues sont, respectivement, de ≈ 690 m2/g, de ≈
378 m2/g et de ≈ 632 m2/g pour la S-10-100, la S-15-100 et la S-T-100. Le volume total
des pores est de 0,72 cm3/g, 0,49 cm3/g et 0,61 cm3/g pour la S-10-100, la S-15-100 et
la S-T-100, respectivement. Aussi un diamètre de pore large d’environ 24 nm, 20 nm et
un plus modeste de 7,5 nm pour la S-10-100, la S-15-100 et la S-T-100, respectivement
a été obtenu. Les isothermes obtenues sont de type IV avec des boucles hystérèses de
type H2. Ces résultats montrent des propriétés texturales très intéressantes.
Figure 58. Les isothermes d'adsorption/désorption d'azote pour la S-10-100.
0
50
100
150
200
250
300
350
400
450
500
0,0 0,2 0,4 0,6 0,8 1,0
Volu
me a
dsorb
é (
cm
3/g
)
P/P0
Adsorption
Désorption

100
Figure 59. Les isothermes d'adsorption/désorption d'azote pour la S-15-100.
Figure 60. Les isothermes d'adsorption/désorption d'azote pour la S-T-100.
0
50
100
150
200
250
300
350
0,0 0,2 0,4 0,6 0,8 1,0
Vo
lum
e a
dso
rbé
(cm
3/g
)
P/P0
Adsorption
Desorption
0
50
100
150
200
250
300
350
400
450
0,0 0,2 0,4 0,6 0,8 1,0
Vo
lum
e a
dso
rbé
(cm
3/g
)
P/P0
Adsorption
Désorption

101
5.3.1.4.2. Analyse par diffraction des rayons X (DRX)
Le diffractogramme (DRX) obtenu aux petits angles pour la S-10-100 (Figure 61)
montrent des pics à 2θ ≈ 0,8°; 0,9°; 1,1°; 1,2° et à 2θ ≈ 1.6° qui caractérisent les
diffractions (311), (331), (331), (442), et (644), respectivement. Le diffractogramme
représentant la S-15-100 (Figure 62), montre un pic majeur de diffraction (111) à l’angle
2θ = 0,3°, ainsi qu’un pic similaire est observé avec la S-T-100 (Figure 63) à environ 2θ ≈
0,5°, on observe aussi un petit pic caractérisant la diffraction (331) à 2θ ≈ 0.6° et à 2θ ≈
0.9° pour la S-15-100 et la S-T-100, respectivement, confirmant une structure
mésoporeuse cubique ordonnée de ces matériaux (Fan et al., 2005).
1 2 3
I (u. a.)
2 Thêta (degrés)
S-10-100311
331 333 442
644
Figure 61. Le diffractogramme (DRX) obtenu aux petits angles obtenus pour la S-10-100.

102
0 1 2 3
2 Thêta (degrés)
I (u. a.)
S-15-100111
331
Figure 62. Le diffractogramme (DRX) obtenu aux petits angles obtenus pour la S-15-100.

103
1 2 3
2 Thêta (degrés)
I (u. a.)
S-T-100111
331
Figure 63. Le diffractogramme (DRX) obtenu aux petits angles obtenus pour la S-T-100.
5.3.1.4.3. Microscopie Électronique à Transmission (MET)
Les images MET montrent une excellente structure cubique de ces matériaux, qui sont en
concordance avec les résultats obtenus par les analyses DRX. Ces images (Figure 64,
Figure 65 et Figure 66) montrent des unités cellulaires larges qui sont 24 nm pour les
silices mésoporeuses S-10-100 et de 20 nm pour les silices mésoporeuses S-15-100.
Mais des unités plus étroites ont été observées pour les silices mésoporeuses S-T-100
de l’ordre de 8 nm. Ces résultats sont en concordance avec les dimensions estimées à
l’aide des isothermes d’absorption/désorption de l’azote.

104
Figure 64. Micrographe MET de la S-10-100 (24 nm).
Figure 65. Micrographe MET de la S-15-100 (19-20 nm).
Figure 66. Micrographe MET de la S-T-100 (7,5 nm).

105
Le tableau suivant résume les résultats obtenus par analyse BET des différents
matériaux synthétisés.
Tableau 13. Les propriétés texturales des supports synthétisés.
Matériaux La surface BET
(m2/g)
Volume des pores
VP (cm3 g-1)
Taille des pores D
(nm)
SBA-15 901,9 1,31 6,8
MCF 487,2 0,52 20,0
S-2.9 376,7 0,60 7,7
S-34.8 458,1 0,62 16,9
S-10-100 689,9 0,72 24,0
S-15-100 378,4 0,49 19,5
S-T-100 632,4 0,61 7,5
5.3.2. Immobilisation de β-galactosidase
Afin d’étudier la rétention de β-galactosidase sur les matériaux on a utilisé la méthode
BCA (Smith et al., 1985) décrite en détail dans la partie 4.4.6.2 pour déterminer la
quantité d’enzyme immobilisée.
Dans cette partie on a essayé d’immobiliser l’enzyme sur les sept matériaux synthétisés
avec trois pH différents : neutre (pH=7) et acide (pH=3,5 et pH=5). Ensuite, on a utilisé
différentes concentrations d’enzyme pour voir la capacité de rétention maximale de
chaque matériau : 250µg d’enzyme/mg de matériaux (25%) ; 600 µg d’enzyme/mg de
matériaux (60%) ; 750 µg d’enzyme/mg de matériaux (75%) et 1000 µg d’enzyme/mg de
matériaux (100%) ainsi que l’effet de la concentration de glutaraldéhyde utilisé (0,1%; 1%
et 2%).

106
5.3.2.1. Effet de pH sur la rétention des enzymes
La représentation graphique de l’effet du pH sur l’immobilisation de β-galactosidase ainsi
que le test d’ANNOVA avec la procédure glm de SAS (Figure 67 et Figure 68) montrent
une différence significative (F<0,0001) entre le pH 7 et les autres pH (Annexe2). Le pH 5
offre une rétention légèrement meilleure que le pH 3,5 pour la majorité des matériaux
(Figure 67). Cependant, cette différence reste statistiquement non significative. De plus,
il est observé que travailler à pH=5 défavorise l’hydrolyse des sucres due à une forte
acidité (pH=3,5) (Parhi et al., 2008). Ce qui est en surcroît un avantage pour la formation
des GOS.
Figure 67. Effet du changement de pH sur la rétention des matériaux.
Ces résultats s’expliquent par le point isoélectrique du matériau et de l’enzyme.
Effectivement, les pH=3,6 et pH=4,6 sont rapportés dans la littérature comme étant des
valeurs de références de pI de la β-galactosidase à partir de l’A.oryzae (Ansari et Husain,
2010, Bernal et al., 2014). En effet, l’enzyme est chargée positivement, à un pH < pI et
chargée négativement à un pH > pI. Souvent, le pI de la surface du support siliceux est
supposé être au voisinage de 2 (Vinu et al., 2004), sauf qu’il existe des matériaux siliceux
ayant des pI supérieurs à 2, comme le montre les résultats obtenus par Hudson et ses
50
55
60
65
70
75
80
85
90
95
100
% d
e r
éte
nti
on
pH=7
pH=3,5
PH=5

107
collègues (Hudson et al., 2005) qui rapportent que le pI de la SBA-15 est d'environ 3,7 ±
0,3.
Figure 68. Distribution de rétention présentée par la procédure glm de SAS.
De plus, plusieurs facteurs contrôlent l’immobilisation de l’enzyme. En effet, des forces
impliquées dans la physisorption de la β-galactosidase sur la silice mésoporeuse comme
les laissons de van der Waals, les interactions par liaisons hydrogène, les interactions
électrostatiques entre les résidus d'acides aminés à la surface de la β-galactosidase et
les groupes silanol sur la surface des matériaux mésoporeux peuvent avoir un rôle
prépondérant (Zhou et al., 2009).
Ces dernières ont un impact très important sur l’immobilisation : lorsque le pH de la
solution est supérieur au pI de l’enzyme, la surface de la protéine devient chargée
négativement et donc une répulsion électrostatique s’exerce entre la β-galactosidase
chargée négativement et le support de silice mésoporeuse (groupes silanol également
chargés négativement), ce qui réduit la quantité de β-galactosidase adsorbée sur le
matériau. A un pH proche du point isoélectrique, la charge nette de l’enzyme est de zéro
(neutre) et donc l'interaction électrostatique entre les molécules protéiques est la plus

108
basse possible, et entraine par conséquent une diminution de la répulsion électrostatique
entre les β-galactosidases (Yiu et Wright, 2005). Les supports maximale d’enzyme
lorsque la valeur de pH de la solution est proche du pI de la protéine, ce qui est en
accord avec les résultats rapportés dans la littérature (Salis et al., 2005, Yiu et Wright,
2005).
En se basant sur ces résultats, tout au long du reste de travail, on va utiliser le pH 5
optimisé pour l’immobilisation de β-galactosidase et la réaction de synthèse des GOS.
5.3.2.2. Effet de la concentration d’enzyme sur la quantité immobilisée lors
de la rétention
Pour étudier l’effet de la concentration d’enzyme sur la rétention avec chaque matériau,
on a utilisé :
Quatre quantités d’enzymes : 250, 600, 750 et 1000 µg d’enzyme/mg de
matériaux
pH optimisé de 5
Concentration initiale de 0,1% de GA.
Selon la Figure 69, les réponses de rétention observées diffèrent d’un matériau à un
autre. Cette différence s’explique par la présence de différentes formes de matériaux
siliceux, ainsi que des diamètres et volumes poreux distincts. D’après les résultats du test
d’ANNOVA appliqué pour les différentes quantités de β-galactosidases, on remarque une
différence significative des interactions entre les matériaux et l’enzyme à des
concentrations différentes. Ceci empêche de faire directement des comparaisons entre
les différentes quantités d’enzymes. Par conséquent, on a eu besoin d’étudier l’effet de la
concentration d’enzyme pour chacun des matériaux, à part (Annexe 3).
On remarque que la concentration d’enzyme à 25% présente le taux de rétention le plus
faible pour la plupart des matériaux, sauf dans le cas des MCF. Pour ce dernier, la
différence de rétention en présence de différentes concentrations d’enzyme est non
significative, comme le montrent les analyses d’ANNOVA en utilisant le test de Tukey
(Tableau 14). Ces résultats affirment que la MCF est capable de retenir la même quantité
d’enzyme (entre 93% et 94%) indépendamment de sa concentration. Pour le reste des
matériaux, une différence significative entre les concentrations d’enzyme est mise en
évidence par le test de Tukey.

109
La MCF retient 94,2% d’enzyme avec la plus faible concentration d’enzyme (25%) par
rapport à une rétention de 84,5% pour la S 2,9 et de 88,0% pour la S 34,8 et la S-15-100.
La rétention optimale de la MCF à cette faible concentration est due à l’adsorption des
enzymes à l’intérieur des pores du matériau (MCF). Ces pores ont une habilité supérieure
à retenir ces protéines grâce à leur forme de mousse. En plus, la forme et la taille des
pores favorisent la formation des agrégats qui les protègent contre les conditions de
lavage (lavage avec HCl ou lavage avec l’eau). Ceci n’est pas le cas avec les autres
matériaux ayant des pores cylindriques uniformes.
Figure 69. Effet de la quantité d’enzyme sur l’immobilisation et la rétention des matériaux.
Même si la concentration d’enzyme à 25% de matériaux est la plus faible en termes de
rétention pour 86% des matériaux (6 matériaux sur 7), elle présente toujours un taux de
rétention satisfaisant (de 84% à 91% de rétention) en comparaison avec des résultats
rapportés dans la littérature (Kim et al., 2007, Kim et al., 2008, Lee et al., 2005).
Dans le cas de la SBA-15, la meilleure rétention est observée avec la concentration
d’enzyme de 75% (Figure 69). Mais en quantité d’enzyme immobilisée, la SBA-15 est
capable de retenir plus de 910 µg d’enzyme/mg, ce qui représente ~ 92 % de la quantité
d’enzyme ajoutée. Cette capacité de rétention est due, essentiellement, à la grande
75
80
85
90
95
100
MCF SBA-15 S 2,9 S 34,8 S-15-100 S-10-100 S-T-100
% R
éte
nti
on
25%
60%
75%
100%

110
surface spécifique de la SBA-15 (901,9 m2/g), mais aussi à la rétention au niveau des
pores relativement étroits (Taille des pores=6,8 nm) par rapport à la taille de l’enzyme (β-
galactosidase=110 KDa).
Tableau 14. Résultats du test du Tukey pour la MCF et la concentration d’enzyme
1
Groupement de Tukey
Moyenne rep Ez
A 94,16 3 25
A 94,06 3 75
A 93,50 3 60
A 93,16 3 100
LA S 2,9 (Tableau 15), la S 34,8 (Tableau 16) et la S-T-100 (Tableau 17) présentent la
même réponse que la SBA-15, avec un meilleur taux de rétention à 75% et une
concentration maximale d’enzyme en µg/mg de matériau de 947,6 ; 957 et 946,
respectivement. Dans le cas de la S 2,9, ces résultats sont attribués, en grande partie, à
sa surface spécifique. Ceci est dû à la taille de ses pores qui ne dépasse pas 7,7 nm.
Dans le cas de la S-T-100, qui présente une rétention similaire à celle de la S 2,9, les
résultats sont plutôt attribués à la forme et la taille de ses pores (7,5 nm), vu que ce
matériau dispose d’une surface spécifique plus petite que la S 2,9. En ce qui concerne la
S 34,8, la rétention est plus importante, essentiellement grâce à sa structure
mésoporeuse particulière mais aussi pour la forme et la taille de ses pores (16,7 nm) qui
semblent très efficaces pour l’immobilisation d’enzyme. En effet, les courbures des méso-
canaux de la silice et la flexibilité inter-coque résultant de la grande taille des pores
empêchent l’enzyme de s’échapper (Zhou et al., 2009).

111
Tableau 15. Résultats du test de Tukey pour la S 2,9 et la concentration d’enzyme
Les moyennes avec la même lettre ne sont pas significativement différentes
Groupement de Tukey
Moyenne rep GA
A 94,86 3 75
A 94,76 3 100
B A 90,50 3 60
B 84,43 3 25
Tableau 16. Résultats du test de Tukey pour la S 34,8 et la concentration d’enzyme
Les moyennes avec la même lettre ne sont pas significativement différentes
Groupement de Tukey
Moyenne rep Ez
A 96,80 3 75
A 95,73 3 100
A 94,46 3 60
B 88,90 3 25

112
Tableau 17. Résultats du test de Tukey pour la S-T-100 et la concentration d’enzyme
Les moyennes avec la même lettre ne sont pas significativement différentes
Groupement de Tukey
Moyenne rep Ez
A 96,46 3 75
A 94,63 3 100
A 94,60 3 60
B 87,40 3 25
Les S-10-100 et S-15-100 ont presque la même forme, la même taille des pores et la
même surface. Ceci explique pourquoi ils se comportent de la même façon et montrent
un meilleur pourcentage de rétention à 60% et une rétention de 940 µg d’enzyme/mg de
matériau (Tableau 18 et Tableau 19). Cette rétention est due particulièrement à la taille
des pores de ces deux matériaux : 24 nm et 19,5 nm, respectivement.
Tableau 18. Résultats du test de Tukey pour la S-15-100et la concentration d’enzyme
Les moyennes avec la même lettre ne sont pas significativement différentes
Groupement de Tukey
Moyenne rep Ez
A 96,53 3 60
B 94,46 3 100
B 94,40 3 75
C 88,73 3 25
Tableau 19. Résultats du test de Tukey pour la S-10-100 et la concentration d’enzyme
Les moyennes avec la même lettre ne sont pas significativement différentes

113
Groupement de Tukey
Moyenne rep GA
A 95,56 3 60
B A 94,46 3 100
B 94,20 3 75
C 91,73 3 25
Pour conclure, la majorité des silices synthétisées au cours de notre étude montre une
capacité de rétention remarquable qui atteint 95%. Les matériaux sont capables de
retenir presque leur poids en enzyme. Ces résultats sont très encourageants pour tester
l’activité de chaque biocatalyseur (enzyme supportée).
5.3.2.3. Effet de la concentration de Glutaraldéhyde (GA) sur la rétention des
enzymes immobilisées
Dans cette partie, on a utilisé trois concentrations de GA : 0,1%, 1% et 2% de la solution
tampon ajoutée, afin d’étudier l’effet de la concentration de GA sur la rétention des sept
matériaux. Ces concentrations sont utilisées à un pH=5 et une concentration d’enzyme
de 75% avec les sept matériaux examinés dans cette étude.
Les analyses d’ANNOVA avec la procédure glm de SAS montrent qu’il y a une interaction
significative entre les matériaux et les concentrations de GA. Donc, on ne peut pas
conclure directement, et il faut passer par l’étude de chaque matériau séparément. Mais il
est clair selon la Figure 70 que la totalité des matériaux présente une meilleure rétention
avec la concentration 0,1% (Annexe4).
Les analyses statistiques avec le test Tukey de chaque matériau confirment que le
pourcentage 0,1% de GA est la meilleure concentration pour la rétention de β-
galactosidase, par rapport aux autres concentrations. Dans le cas de la S 34,8, il n’y a
pas de différence significative entre les trois traitements. Ceci est probablement dû à son
unique forme de vésicule (Tableau 20).
Trevisan et ses collègues (Trevisan et al., 1997) montrent que l’augmentation de la
concentration de GA entraîne une diminution de l’activité et même une désactivation de

114
l’enzyme, mais elle assure une meilleure stabilité. Donc, il s’agit d’un compromis entre
l'effet sur l'activité enzymatique dégénérative et sur le renforcement de la structure
protéique qui affecte la rétention de l’enzyme.
Figure 70. Effet de la concentration de GA sur l’immobilisation et la rétention des enzymes par les différents matériaux.
Tableau 20. Résultats du test de Tukey pour la S 34,8 et les concentrations de GA
Les moyennes avec la même lettre ne sont pas significativement différentes
Groupement de Tukey
Moyenne rep GA
A 81,86 3 1
A 81,80 3 0.1
A 78,80 3 2
70
75
80
85
90
95
100
MCF SBA-15 S 2,9 S 34,8 S-15-100 S-10-100 S-T-100
% R
éte
nti
on
0,1%
1%
2%

115
5.3.3. Réaction de transglycosylation avec l’enzyme immobilisée
Dans cette partie, on a étudié la réponse de chaque matériau vis-à-vis de la réaction de
transglycosylation pour la formation du lactosucrose et du tetrahexose, tout en la
comparant d’une part à la réaction en phase homogène, et d’autre part aux différents
matériaux entre eux. La réaction a été effectuée dans les conditions déjà optimisées.
Dans un premier lieu on fixe un pH de 5 et une température de 40°C ainsi qu’une
concentration d’enzyme de 100% qui a été optimisée durant le processus de rétention.
Dans un second lieu, on va utiliser une concentration enzymatique de 60% pour pouvoir
comparer les différentes approches : hétérogène (enzyme immobilisée) et homogène
(enzyme libre).
5.3.3.1. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée
sur la MCF
La MCF est un support siliceux ayant des pores sphériques interconnectés, de tailles
uniformes atteignant 20 nm avec une surface spécifique de 487.2 m2/g. Ce matériau a
montré une rétention d’enzyme de 93,2% à pH acide (pH=5) avec une concentration
de 100% en enzyme.
La Figure 71 montre une formation du lactosucrose dépassant les 75 g/L (25%) (Avec
une concentration de 100% en enzyme) ainsi que la présence du glucose avec une
quantité de 94,8 g/l, ensuite on trouve la formation du fructose qui ne cesse
d’augmenter en fonction du temps, ainsi qu’une formation faible du tetrahexose (11,6
g/l après 24h de réaction) et du galactose (9,2 g/l après 24h de réaction). En effet,
l’enzyme révèle une réactivité très importante après son immobilisation sur la MCF.
Cette réactivité peut être due au taux très important d’enzymes immobilisées et leur
disponibilité pour effectuer la réaction de transglycosylation, ainsi que les phénomènes
de transfert de matière qui se trouveraient favorisés par des pores larges de la MCF.
D’un autre côté, la forme mousseuse exceptionnelle ainsi que la surface spécifique
importante de la MCF favorisent considérablement les interactions entre les matrices
protéiques de l’enzyme conduisant à la formation des agrégats responsables de
l’activité catalytique, ainsi que leur rôle dans l’amélioration de la rétention de ce
matériau.

116
Avec une concentration enzymatique de 60%, la MCF montre une rétention de 93,5%.
La formation maximale du lactosucrose sous ces conditions réactionnelles est de 69,5
g/L (23%), ce qui représente une concentration supérieure à celle obtenue avec
l’enzyme en solution à pH 5 (un maximum de 62,8 g/L (21%)) comme le montre la
Figure 72. Cependant, il est bien clair selon la Figure 72 que la vitesse de réaction de
formation du lactosucrose est plus élevée dans le cas de la réaction en phase
homogène et cela peut être expliqué par l’accessibilité améliorée des substrats aux
sites actifs de l’enzyme en solution par rapport à celle qui est immobilisée. On
remarque aussi la grande différence entre l’approche hétérogène et homogène
concernant la formation du glucose et du galactose. Dans le cas de la phase
homogène, il y avait une production de 105,2 g/L de glucose et de 69,6 g/L de
galactose contre une production correspondante de 74,4 g/L de glucose et 8,7 g/L de
galactose en phase hétérogène. Cette différence peut être attribuée à la réaction
d’hydrolyse du lactose qui a été majoritaire dans la phase homogène d’où la
production intense des monosaccharides galactose et glucose. Cette réaction
indésirable est limitée dans le cas de l’enzyme immobilisée. Ces résultats montrent
une sélectivité importante de ce biocatalyseur (MCF) qui permet d’augmenter la
synthèse du lactosucrose, tout en limitant la réaction d’hydrolyse du lactose.

117
Figure 71. Cinétique de la réaction de formation des GOS catalysée par β-galactosidase immobilisée sur la MCF comme support à une concentration enzymatique de 100%.
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Co
nce
ntr
atio
n (
g/L)
Temps (h)
Saccharose (g/L)
Lactose (g/L)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)

118
Figure 72. Comparaison entre les produits de la réaction avec enzymes immobilisées sur la MCF et les produits de la réaction avec des enzymes en solution à une concentration
enzymatique de 60%.
5.3.3.2. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée
sur la SBA-15
La SBA-15 est un support mésoporeux siliceux avec une forme hexagonale ayant une
taille de pore de 6,8 nm, et une surface très importante de 901.9 m2/g. Cette surface
exceptionnellement large assure une rétention de 91,9% à un pH=5 avec une
concentration enzymatique de 100%.
Au cours de la réaction enzymatique utilisant la β-Galactosidase immobilisée sur la
SBA-15, la formation des produits ne reflète pas cette rétention enzymatique (91,9%
avec une concentration d’enzyme de 100%), surtout avec des pores de très petite
taille, comparée à la taille de la β-galactosidase. Une telle rétention s’explique par la
surface externe importante de la SBA-15. En effet, le maximum de production de
lactosucrose par la SBA-15 à une concentration d’enzyme de 100% est de 17,1 g/l
(5,8 %) (Figure 73 et Figure 74 ). Pandya et ses collègues (Pandya et al., 2005)
confirment que l’enzyme est immobilisée uniquement sur la surface externe dans le
0
20
40
60
80
100
120
0 5 10 15 20 25 30
Co
nce
ntr
atio
n (
g/l)
Time (h)
glu-MCF
gal-MCF
fru-MCF
GOS I-MCF
GOS II-MCF
glu-E libre
gal-E libre
fru-E libre
GOS I-E libre
GOS II-E libre

119
cas de la SBA-15. Ceci conduit à une formation faible des agrégats enzymatiques
responsables de l’activité catalytique, ce qui implique une activité faible.
Figure 73. Cinétique de la réaction de formation des GOS catalysée par β-galactosidase supportée sur la SBA-15 à une concentration d’enzyme de 100%.
Figure 74. Cinétique des produits formés au cours de la réaction catalysée par β-galactosidase supportée sur la SBA-15 à une concentration d’enzyme de 100%.
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Saccharose (g/L)
Lactose (g/L)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)
0
5
10
15
20
25
30
35
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)

120
Avec une concentration enzymatique de 60% (Figure 75), la SBA-15 offre un
rendement plus faible (18% en lactosucrose) comparé à celui obtenu avec l’approche
homogène. Cependant, il est important de mentionner que, tout au long de la réaction
avec l’enzyme immobilisée, ni le galactose ni le tetrahexose ne se forment. En effet, le
galactose est un produit d’hydrolyse de lactose et son absence montre que la réaction
de transglycosylation domine dans ces conditions expérimentales. L’absence du
tetrahexose confirme que la SBA-15 montre une très haute sélectivité vis-à-vis la
production du lactosucrose.
Figure 75. Comparaison entre les produits de la réaction avec enzymes immobilisées sur la SBA-15 et les produits de la réaction avec des enzymes en solution à une
concentration enzymatique de 60%.
5.3.3.3. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée
sur la S-2.9
Le support siliceux S 2,9 présente une surface BET de 376.7 m2/g et une taille des
pores de 7,7 nm. Ces caractéristiques permettent à la ‘S 2,9’ de retenir 94,76% de
l’enzyme, mais entraine cependant une activité catalytique très limitée (Figure 76 et
Figure 77). Un rendement faible en lactosucrose qui ne dépasse pas 3,5% (production
maximal de 10,5 g/L du lactosucrose) et qui est attribué à l’immobilisation d’enzyme
restreinte à la surface externe du matériau (Pandya et al., 2005).
0
20
40
60
80
100
120
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/l
Temps (h)
glu-SBA-15
gal-SBA-15
fru-SBA-15
GOS I-SBA-15
GOS II-SBA-15
glu-E libre
gal-E libre
fru-E libre
GOS I-E libre
GOS II-E libre

121
Cependant, une sélectivité très intéressante est observée, une absence de formation
de galactose qui montre que la transglycosylation emporte la totalité de la réaction,
afin de favoriser la synthèse de lactosucrose.
La Figure 78 montre une large différence entre le rendement de β-galactosidase en
solution à 60% et celle immobilisée sur la ‘S 2,9’. En effet, le rendement en
lactosucrose formé avec l’enzyme immobilisée est de 7,1% par rapport à celle produite
avec l’enzyme libre.
Figure 76. Cinétique de la réaction de formation des GOS catalysée par β-galactosidase supportée sur la ‘S 2,9’ à une concentration d’enzyme de 100%.
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Saccharose (g/L)
Lactose (g/L)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)

122
Figure 77. Cinétique des produits formés au cours de la réaction catalysée par β-galactosidase supportée sur la ‘S 2,9’ à une concentration d’enzyme de 100%.
Figure 78. Comparaison entre les produits de la réaction avec enzymes immobilisées sur la ‘S 2,9’ et les produits de la réaction avec des enzymes en solution à une concentration
enzymatique de 60%.
0
5
10
15
20
25
30
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)
0
20
40
60
80
100
120
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/l
Temps h
glu-S 2,9
gal-S 2,9
fru-S 2,9
GOS I-S 2,9
GOS II-S 2,9
glu-E libre
gal-E libre
fru-E libre
GOS I-E libre
GOS II-E libre

123
5.3.3.4. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée
sur la S-34.8
La silice avec la structure en forme d’oignons (S 34,8) montre une capacité très
particulière pour la rétention des enzymes qui peut atteindre jusqu’à 95%, ainsi qu’une
activité importante pour la réaction dans l’approche hétérogène. Cela est dû
principalement à sa forme qui piège l’enzyme à l’intérieur des pores, et à la taille élevée
de ses pores (16.9 nm) ainsi qu’une surface de BET considérable de 458.1 m2/g.
La Figure 79 ainsi que la Figure 80 (avec une concentration de 100% en enzyme)
montrent une formation importante du lactosucrose qui dépasse les 54 g/L (18%), ainsi
que la présence du tetrahexose avec une quantité considérable de 11 g/L (4%). En effet,
l’enzyme révèle une réactivité importante suite à son immobilisation sur la ‘S 34,8’. Cette
réactivité peut être attribuée au taux très important d’enzymes immobilisées et leur
disponibilité pour effectuer la réaction de transglycosylation, De plus, la forme en vésicule
et la taille des pores ainsi que la surface spécifique importante du support, contribuent
considérablement à l’amélioration du transfert de matière et par conséquent l’accessibilité
des substrats aux sites catalytiques actifs sur la matrice enzymatique.
La vitesse de formation des produits avec l’enzyme immobilisée à 60% de concentration
enzymatique est plus faible que celle avec l’enzyme libre (Figure 81). En effet, la réaction
en approche hétérogène atteint son maximum de production de lactosucrose (45,9 g/L du
lactosucrose, 15,5%) après 28 heures de réaction. Tandis que ce maximum de
production (62,8 g/L du lactosucrose, 21,2 %) est atteint après seulement 12 heures de
réaction en approche homogène. Ceci est dû à une meilleure accessibilité des substrats
aux sites actifs de l’enzyme libre, ce qui améliore la cinétique de transglycosylation.
La Figure 81 montre une formation faible de glucose et de galactose dans le cas de
l’approche hétérogène par rapport à celle produite avec l’approche homogène. Cette
formation est probablement due à la réaction d’hydrolyse du lactose. Cette réaction
secondaire est limitée dans le cas des enzymes immobilisées sur la S 34,8.

124
Figure 79. Cinétique de la réaction de formation des GOS catalysée par β-galactosidase supportée sur la ‘S 34,8’ à une concentration d’enzyme de 100%.
Figure 80. Cinétique des produits formés au cours de la réaction catalysée par β-galactosidase supportée sur la ‘S 34,8’ à une concentration d’enzyme de 100%.
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Saccharose (g/L)
Lactose (g/L)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)
0
10
20
30
40
50
60
70
80
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)

125
Figure 81. Comparaison entre les produits de la réaction avec enzymes immobilisées sur la ‘S 34,8’ et les produits de la réaction avec des enzymes en solution à une
concentration enzymatique de 60%.
5.3.3.5. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée
sur la S-10-100
Le support siliceux S-10-100 est un support à base de la SBA -15 gonflée avec l’ajout de
TMB. Suite à ce gonflement, on obtient un matériau avec une taille des pores de 24 nm
et une surface de BET très importante de 689.9 m2/g. Des caractéristiques très
favorables conduisant à une rétention de 94,46% de β-galactosidase à pH= 5 et une
concentration d’enzyme de 100%.
Dans ces conditions réactionnelles, 51 g/L (17 %) du lactosucrose et 10 g/L (4 %) du
tetrahexose sont obtenus après 28 heures de réaction ainsi que la formation du
galactose, du glucose et deu fructose suite à la réaction d’hydrolyse (Figure 82 et Figure
83). Ce rendement considérable des galacto-oligosaccharides est favorisé principalement
par la taille considérable des pores de la S-10-100 ainsi que sa surface spécifique
importante.
La Figure 84 présente une comparaison entre les produits formés lors des deux
approches homogène et hétérogène de la réaction à 60% de concentration d’enzyme.
-20
0
20
40
60
80
100
120
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/l
Temps (h)
glu-S 34,8
gal-S 34,8
fru-S 34,8
GOS I-S 34,8
GOS II-S 34,8
glu-E libre
gal-E libre
fru-E libre
GOS I-E libre
GOS II-E libre

126
Cette figure montre aussi une vitesse de réaction très élevée et une sélectivité faible dans
le cas de l’utilisation de l’enzyme libre.
Figure 82. Cinétique de la réaction de formation des GOS catalysée par β-galactosidase supportée sur la ‘S-10-100’ à une concentration d’enzyme de 100%.
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Co
nce
ntr
atio
n (
g/L)
Temps (h)
Saccharose (g/L)
Lactose (g/L)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)

127
Figure 83. Cinétique des produits formés au cours de la réaction catalysée par β-galactosidase supportée sur la ‘S-10-100’ à une concentration d’enzyme de 100%
Figure 84. Comparaison entre les produits de la réaction avec enzymes immobilisées sur la ‘S-10-100’ et les produits de la réaction avec des enzymes en solution à une
concentration enzymatique de 60%.
0
10
20
30
40
50
60
70
80
0 5 10 15 20 25 30
Co
nce
ntr
atio
n (
g/L)
Temps (h)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)
0
20
40
60
80
100
120
0 10 20 30
Co
nce
ntr
atio
n g
/l
Temps (h)
glu-S-10-100
gal-S-10-100
fru-S-10-100
GOS I-S-10-100
GOS II-S-10-100
glu-E libre
gal-E libre
fru-E libre

128
5.3.3.6. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée
sur la S-15-100
Le support siliceux S-15-100 est une SBA-15 gonflée par l’ajout de TMB afin d’obtenir
des pores plus large (19,5 nm) et des formes irrégulières (hexagonales déformées). La S-
15-100 a réussi à retenir 94,44% de l’enzyme à 100% de concentration.
Les Figure 85 et Figure 86 montrent une concentration considérable du lactosucrose (43
g/L, 14,5 %) dans le cas de la réaction hétérogène. Elles montrent aussi une sélectivité
importante, surtout au niveau de la formation minime du galactose. Cette sélectivité est
illustrée dans la Figure 87, où 14 g/L du galactose sont formés par la voie hétérogène à
une concentration de 60% d’enzyme, contre 69,5 g/L de galactose formés par l’enzyme
en solution. Ces résultats peuvent être expliqués, d’une part, par la taille des pores de la
silice, et d’autre part, par l’amélioration de la stabilité de la matrice enzymatique vis-à-vis
des conditions de la réaction de transgalactosylation, grâce à l’immobilisation des
enzymes à l’intérieur et à la surface du support siliceux.
Figure 85. Cinétique de la réaction de formation des GOS catalysée par β-galactosidase supportée sur la ‘S-15-100’ à une concentration d’enzyme de 100%.
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Saccharose (g/L)
Lactose (g/L)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)

129
Figure 86. Cinétique des produits formés au cours de la réaction catalysée par β-galactosidase supportée sur la ‘S-15-100’ à une concentration d’enzyme de 100%.
Figure 87. Comparaison entre les produits de la réaction avec enzymes immobilisées sur la ‘S-15-100’ et les produits de la réaction avec des enzymes en solution à une
concentration enzymatique de 60%.
0
10
20
30
40
50
60
70
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)
0
20
40
60
80
100
120
0 5 10 15 20 25 30
Co
ncn
trat
ion
g/l
Temps (h)
glu-S-15-100
gal-S-15-100
fru-S-15-100
GOS I-S-15-100
GOS II-S-15-100
glu-E libre
gal-E libre
fru-E libre
GOS I-E libre
GOS II-E libre

130
5.3.3.7. Réactivité de la β-Galactosidase d’Aspergillus oryzae immobilisée
sur la S-T-100
La S-T-100 est un autre support à base de la SBA -15 gonflée avec l’ajout de TMB. Ce
matériau présente une surface de BET très importante de 632,4 m2/g, avec une taille des
pores de 7,5 nm. Ces caractéristiques et surtout l’aire spécifique élevée, sont très
favorables, pour avoir une rétention importante de 96,46 % de β-galactosidase à pH= 5 et
une concentration d’enzyme de 100%.
Cette rétention (avec 100% d’enzyme) très élevée a donné un rendement faible de 3,4
g/L (1,2 %) en lactosucrose, ainsi que 1,8g/L (0.59 %) du tetrahexose. Cette production
est comparable avec celle obtenue par la SBA-15 et la S-2.9, mais reste néanmoins plus
faible qu’elles, à cause de leurs pores étroits. On observe aussi une production minimale
du galactose (Figure 88 et Figure 89) suite à une réaction d’hydrolyse minimale dans
ces conditions opératoires.
La Figure 90 montre une grande différence entre la production des GOS en comparant
les deux phases catalytiques homogène (enzyme libre) et hétérogène (enzyme
immobilisée). Une production plus élevée est observée avec l’enzyme libre avec une
cinétique rapide. On remarque aussi une formation très élevée du glucose et du
galactose dans le cas de l’approche homogène par rapport à celle produite dans
l’approche hétérogène, ce qui peut être attribué à une limitation de la réaction d’hydrolyse
du lactose après immobilisation.

131
Figure 88. Cinétique de la réaction de formation des GOS catalysée par β-galactosidase supportée sur la ‘S-T-100’ à une concentration d’enzyme de 100%.
Figure 89. Cinétique des produits formés au cours de la réaction catalysée par β-galactosidase supportée sur la ‘S-T-100’ à une concentration d’enzyme de 100%.
0
50
100
150
200
250
300
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Saaccharose (g/L)
Lactose (g/L)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)
0
2
4
6
8
10
12
14
0 5 10 15 20 25 30
Co
nce
ntr
atio
n g
/L
Temps (h)
Glucose (g/L)
Galactose (g/L)
Fructose (g/L)
Lactosucrose (g/L)
Tetrahexose (g/L)

132
Figure 90. Comparaison entre les produits de la réaction avec enzymes immobilisées sur la ‘S-T-100’ et les produits de la réaction avec des enzymes en solution à une
concentration enzymatique de 60%.
Les résultats obtenus avec l’enzyme supportée sur les différents matériaux siliceux, ainsi
que ceux obtenus avec l’approche homogène utilisant l’enzyme libre, sont rassemblés
dans le Tableau 21, qui présente les différentes proportions obtenues en termes du
lactosucroseI et du tetrahexose ainsi que les GOS totaux.
0
20
40
60
80
100
120
0 10 20 30
Co
nce
ntr
atio
n g
/l
Temps (h)
glu-S-T-100
gal-S-T-100
fru-S-T-100
GOS I-S-T-100
GOS II-S-T-100
glu-E libre
gal-E libre
fru-E libre
GOS I-E libre
GOS II-E libre

133
Tableau 21. Résultats récapitulatifs des rendements en galacto-oligosaccharides (lactosucrose et tetrahexose) obtenus avec l’enzyme immobilisée sur les différents matériaux
Concentration d’enzyme 100% Concentration d’enzyme 60%
Lactosucrose g/L
(%)
Tetrahexose g/L
(%)
Total GOS
(%)
Lactosucrose g/L
(%)
Tetrahexose g/L
(%)
Total GOS
(%)
E/MCF 75,2
(25,33%)
11,0
(3,68%)
86,1
(29,02%)
69,5
(23,41%)
11,5
(3,87%)
80,9
(27,28%)
E/SBA-15 17,1
(5,78%)
1,0
(0,33%)
18,1
(6,11%)
11,5
(3,87%)
00,0 11,5
(3,87%)
E/S 2,9 10,5
(3,52%)
3,9
(1,33%)
14,4
(4,86%)
4,5
(1,50%)
1,5
(0,5%)
5,9
(2%)
E/S 34,8 54,6
(18,40%)
11,2
(3,78%)
65,8
(22,18%)
45,9
(15,46%)
10,8
(3,62%)
56,6
(19,09%)
E/S-10-100 51,9
(17,48%)
9,7
(3,61%)
61,5
(20,75%)
40,5
(13,66%)
9,9
(3,32%)
50,4
(16,99%)
E/S-15-100 43,0
(14,48%)
10,7
(3,61%)
53,7
(18,09%)
37,9
(12,78%)
10,6
(3,58%)
48,6
(16,37%)
E/S-T-100 3,4
(1,15%)
1,8
(0,59%)
5,2
(1,75%)
3,1
(1,03%)
1,4
(0,47%)
4,5
(1,5%)
Approche
Homogène
63,7
(21,47%)
10,6
(3,58%)
74,3
(25,06%)
62,8
(21,16%)
13,4
(4,53%)
76,2
(25,69%)

134
5.3.3.8. Comparaison entre la réactivité des matériaux
Le Tableau 21 illustre la production de β-Galactosidase d’Aspergillus oryzae immobilisée
sur chaque matériau en lactosucrose et en tetrahexose pour les deux concentrations
d’enzyme (100% et 60%). Tous les matériaux présentent une production en GOS plus
élevée avec une concentration enzymatique de 100% par rapport à 60%. Ce constat
s’explique par la rétention plus élevée des enzymes à la concentration de 100%, ce qui
implique leur disponibilité pour la réaction, en augmentant le nombre de sites actifs.
Les Figure 91 et 92 (avec une concentration enzymatique de 100%) montrent que la
MCF présente le meilleur taux de conversion de lactose (75% de conversion), ainsi que le
meilleur biocatalyseur pour la production du lactosucrose et du tetrahexose. En effet, les
larges pores de la MCF et sa forme mousseuse font d’elle un support plus approprié pour
accueillir confortablement l'enzyme dans ses pores, en plus de faciliter l’accès des
substrats aux sites actifs de l’enzyme et faciliter un transfert de matière favorable.
En deuxième position, on trouve la S 34,8 suivie de la S-10-100 et de la S-15-100 avec
des taux de conversion du lactose très proches et une allure similaire comme illustrée
dans les Figure 91 etFigure 92. Dans la littérature (Chouyyok et al., 2009, Kannan et
Jasra, 2009), l'activité de l'enzyme immobilisée sur différents supports siliceux dépend
étroitement de la taille des pores, vu qu’un diamètre large permet un meilleur transfert de
matière. De plus, la structure des matériaux et leurs surfaces affectent le transfert de
matière et donc l’activité de l’enzyme. Chouyyok et ses collègues (Chouyyok et al., 2009)
confirment que la structure de la MCF en cellules sphériques est plus appropriée pour un
bon transfert de matière et pour l’activité enzymatique, par rapport aux structures de la
SBA-15 et la MCM-41 cylindriques longues.

135
Figure 91. Conversion de lactose en fonction du rendement en lactosucrose de différents matériaux.
0
5
10
15
20
25
30
0 10 20 30 40 50 60 70
Re
nd
em
en
t la
cto
sucr
ose
(%
)
Coversion de lactose %
MCF
SBA-15
S 2,9
S 34,8
S-15-100
S-10-100
S-T-100

136
Figure 92. Conversion de lactose en fonction de rendement en tetrahexose de différents matériaux.
5.3.4. Activité des biocatalyseurs synthétisé après recyclage
Dans cette section, on étudie un paramètre important en catalyse hétérogène : la stabilité
des biocatalyseurs. Il s’agit de mesurer l’activité de chaque biocatalyseurs (β-
Galactosidase d’Aspergillus oryzae immobilisée sur la silice) après son recyclage. Pour
avoir une idée plus large sur la performance de ces biocatalyseurs, on a décidé
d’effectuer la réaction de transglycosylation en présence de biocatalyseurs régénérés. On
a donc déterminé les rendements en GOS, jusqu'au 5ème cycle.
La Figure 93 montre une désactivation minimale du biocatalyseur (β-Galactosidase
d’Aspergillus oryzae immobilisée sur la SBA-15). On rappelle que la SBA-15 présente
une activité modeste (18,1%) à cause de ses pores étroits, donc l’enzyme active est
immobilisée à l’extérieur. Donc à cause de cette immobilisation faible, on observe une
baisse d’activité qui passe à 14,7% (en GOS totaux) après un 5ème recyclage du
biocatalyseur.
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 10 20 30 40 50 60 70
Re
nd
em
en
t Te
trah
exo
se (
%)
Coversion de lactose %
MCF
SBA-15
S 2,9
S 34,8
S-15-100
S-10-100
S-T-100

137
Pour la MCF (Figure 94), une activité très importante a été observée atteignant 86,08 g/l
de GOS totaux. Cette activité a été pratiquement maintenue après plusieurs recyclages
du biocatalyseur. On a observé seulement environ 8,1% (en GOS totaux) de perte en
activité avec ce matériau, accompagnée de 86,1% de rendement en GOS. Ceci montre
encore une fois que c’est un excellent choix de support pour immobiliser la β-
Galactosidase d’Aspergillus oryzae, grâce à ses propriétés texturales uniques. Elles
permettent une accessibilité favorable à la formation d’agrégats qui sont résistants, d’une
part, aux conditions opératoires de lavage lors du processus de recyclage de ce
biocatalyseurs, et d’autre part, à l’effet de l’environnement acide du milieu réactionnel.
Les autres biocatalyseurs à base des matériaux similaires, ayant des diamètres de pores
entre 17 et 20 nm, maintiennent leurs activités catalytiques avec des pertes légères de
15,4% pour la S-34,8 (Figure 95), de 13,15% pour la S-10-100 (Figure 96) et de 13,80%
pour la S-15-100 (Figure 97), accompagnées des rendements de 65,8, de 61,6 et de
53,7 g/L, respectivement.
Une perte considérable de 33,19% (en GOS totaux) en activité illustrée dans la Figure
98, s’ajoute à un rendement faible en production des GOS 14,42%, et font du S-2,9 un
support inadéquat pour immobiliser l’enzyme de taille encombrante (β-Galactosidase
d’Aspergillus oryzae). Cette perte est principalement attribuée au faible diamètre des
pores de ce matériau, ce qui limite l’accessibilité à l’enzyme, et par conséquent
défavorise la formation des agrégats protéiques responsables de l’activité catalytique. Ce
comportement est analogue à la S-T-100 (Figure 99), qui présente aussi une grande
perte près de plus que la moitié (54,70%) et qui présentait déjà une production faible de
5,20 g/L en GOS.
.

138
Figure 93. La production en lactosucrose et en tetrahexose par β-Galactosidase d’Aspergillus oryzae immobilisée sur la SBA-15 pour 5 cycles successifs.
Figure 94. La production en lactosucrose et en tetrahexose par β-Galactosidase d’Aspergillus oryzae immobilisée sur la MCF pour 5 cycles successifs.
0
2
4
6
8
10
12
14
16
18
20
22
Lactosucrose Tetrahexose GOS totaux
Co
nce
ntr
atio
n (
g/L)
Cycle 1
Cycle 2
Cycle 3
Cycle 4
Cycle 5
0
10
20
30
40
50
60
70
80
90
100
Lactosucrose tetrahexose GOS Totaux
Co
nce
ntr
atio
n (
g/L)
Cycle 1
Cycle 2
Cycle 3
Cycle 4
Cycle 5

139
Figure 95. La production en lactosucrose et en tetrahexose par β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-34.8 pour 5 cycles successifs.
Figure 96. La production en lactosucrose et en tetrahexose par β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-10-100 pour 5 cycles successifs.
0
10
20
30
40
50
60
70
80
Lactosucrose Tetrahexose GOS Totaux
Co
nce
ntr
atio
n (
g/L)
Cycle 1
Cycle 2
Cycle 3
Cycle 4
Cycle 5
0
10
20
30
40
50
60
70
Lactosucrose Tetrahexose GOS Totaux
Co
ncn
trat
ion
(g/
L) Cycle 1
Cycle 2
Cycle 3
Cycle 4
Cycle 5

140
Figure 97. La production en lactosucrose et en tetrahexose par β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-15-100 pour 5 cycles successifs.
Figure 98. La production en lactosucrose et en tetrahexose par β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-2.9 pour 5 cycles successifs.
0
10
20
30
40
50
60
Lactosucrose Tetrahexose GOS Totaux
Co
nce
ntr
atio
n (
g/L)
Cycle 1
Cycle 2
Cycle 3
Cycle 4
Cycle 5
0
2
4
6
8
10
12
14
16
18
Lactosucrose Tetrahexose GOS Totaux
Co
nce
ntr
atio
n (
g/L)
Cycle 1
Cycle 2
Cycle 3
Cycle 4
Cycle 5

141
Figure 99. La production en lactosucrose et en tetrahexose par β-Galactosidase d’Aspergillus oryzae immobilisée sur la S-T-100 pour 5 cycles successifs.
0
1
2
3
4
5
6
7
Lactosucrose Tetrahexose GOS Totaux
Co
nce
ntr
atio
n (
g/L)
Cycle 1
Cycle 2
Cycle 3
Cycle 4
Cycle 5


143
Conclusion générale
Lors de cette étude, on a réussi à synthétiser deux galacto-oligosaccharides, dont le
plus important est le lactosucrose. La synthèse de ce dernier était catalysée par la β-
galactosidase à partir d’Aspergillus oryzae par bioconversion de lactose et de
saccharose. Deux approches ont été utilisées pour la biosynthèse de lactosucrose, une
approche homogène et une approche hétérogène.
Dans l’approche homogène, l’effets du pH, de la température, de la concentration
d’enzyme ainsi que des substrats a été étudiées et ensuite optimisées pour obtenir un
rendement de 25% en lactosucrose avec une concentration équimolaire de 60% entre
les substrats, un pH de 5, une température de 40°C, et 6U/mL d’enzymes.
Dans l’approche hétérogène, sept différents matériaux siliceux ont été synthétisés. Ces
matériaux ont servi à immobiliser la β-galactosidase en utilisant le glutaraldhéyde (GA)
comme agent de réticulation. Malgré les différences en diamètres des pores (variant de
6,8 à 24,0 nm), en formes de pores, ainsi qu’en formes de supports (hexagonale,
sphérique, forme de barre, forme d’oignon, etc.), les matériaux montrent une très bonne
rétention de l’enzyme utilisée dépassant les 90% avec des résultats très proches pour
les différents matériaux. Une telle rétention est due principalement à la taille et la forme
des pores, à la surface spécifique de chaque matériau et au positionnement des
enzymes lors de l’adsorption (à l’intérieur des pores ou à la surface).
L’effet de la taille et de la forme des pores, ainsi que la structure des matériaux a été
mise en relief lors de la réaction enzymatique. En effet, l'activité de l'enzyme immobilisée
sur les différents supports de silice est résumée dans l’ordre suivant :
S-T-100 < S 2,9 < SBA-15 < S-15-100 < S-10-100 < S 34,8 < MCF
La MCF montre une très bonne activité enzymatique, grâce à sa forme particulière ainsi
que la taille de ses pores. Cette activité dépasse celle obtenue par l’enzyme libre. Par
contre, la SBA-15 montre une activité modeste, mais avec une excellente sélectivité
envers le lactosucrose.
L’approche hétérogène montre une sélectivité très importante par rapport à l’approche
homogène. D’une part, l’approche hétérogène favorise la réaction de

144
transgalactosylation, donc la formation des GOS, et d’autre part, elle limite la réaction
d’hydrolyse des substrats. Ces deux propriétés sont ainsi très intéressantes et donnent
plus d’efficacité aux biocatalyseurs ainsi synthétisés.
Le recyclage et la réutilisation des différents biocatalyseurs ont été effectués en 5 cycles
successifs, et révèlent une performance très élevée de ces biocatalyseurs avec des
performances légèrement plus faibles pour certains matériaux.
Suite à cette étude, d’autres méthodes d’immobilisation sont envisagées comme
perspectives. Principalement, l’immobilisation d’enzymes par des liaisons covalentes en
modifiant des fonctionnalités des matériaux afin d’améliorer plusieurs paramètres, tels
que la rétention, la formation des agrégats enzymatiques ainsi que l’amélioration de leur
résistances vis-à-vis des conditions opératoires de synthèse et de recyclage.
Il est à noter aussi d’étudier d’autres matériaux qui présentent des fonctionnalités
différentes de celle de la silice.

145
Références bibliographiques
Aider, M., De Halleux, D., 2007. Isomerization of lactose and lactulose production: review. Trends in Food Science & Technology 18 (7), 356-364.
Albayrak, N., Yang, S.-T., 2002. Production of galacto-oligosaccharides from lactose by Aspergillus oryzae β-galactosidase immobilized on cotton cloth. Biotechnology and Bioengineering 77 (1), 8-19.
Ansari, S.A., Husain, Q., 2010. Lactose hydrolysis by β galactosidase immobilized on concanavalin A-cellulose in batch and continuous mode. Journal of Molecular Catalysis B: Enzymatic 63 (1–2), 68-74.
Ansari, S.A., Satar, R., Alam, F., Alqahtani, M.H., Chaudhary, A.G., Naseer, M.I., Karim, S., Sheikh, I.A., 2012. Cost effective surface functionalization of silver nanoparticles for high yield immobilization of Aspergillus oryzae β-galactosidase and its application in lactose hydrolysis. Process Biochemistry 47 (12), 2427-2433.
Ashish K Sharma, V.A., Rajesh Kumar2, Himanshu Chaurasia1, Deepti Chaurasia1, Peeyush Bhardwaj, 2011. Prebiotics: A Review of Therapeutic Potential. INTERNATIONAL JOURNAL OF PHARMACEUTICAL INNOVATIONS 1 (3), 28-40.
Bakken, A.P., Hill, C.G., Amundson, C.H., 1990. Use of novel immobilized β-galactosidase reactor to hydrolyze the lactose constituent of skim milk. Biotechnology and Bioengineering 36 (3), 293-309.
Bauman, D.E., Mather, I.H., Wall, R.J., Lock, A.L., 2006. Major Advances Associated with the Biosynthesis of Milk. Journal of Dairy Science 89 (4), 1235-1243.
Belkacemi, K., Boulmerka, A., Arul, J., Hamoudi, S., 2006. Hydrogenation of Vegetable Oils with Minimum trans and Saturated Fatty Acid Formation Over a New Generation of Pd-catalyst. Topics in Catalysis 37 (2-4), 113-120.
Bernal, C., Sierra, L., Mesa, M., 2014. Design of β-galactosidase/silica biocatalysts: Impact of the enzyme properties and immobilization pathways on their catalytic performance. Engineering in Life Sciences 14 (1), 85-94.
Bernal, C., Urrutia, P., Illanes, A., Wilson, L., 2013. Hierarchical meso-macroporous silica grafted with glyoxyl groups: opportunities for covalent immobilization of enzymes. New Biotechnology 30 (5), 500-506.
Boon, M.A., Janssen, A.E.M., Van 't Riet, K., 2000. Effect of temperature and enzyme origin on the enzymatic synthesis of oligosaccharides. Enzyme and Microbial Technology 26 (2-4), 271-281.
Bornscheuer, U.T., 2008. Industrial Biotransformations. In, Biotechnology Set. Wiley-VCH Verlag GmbH, pp. 275-294.
Bowers, L., Carr, P., 1980. Immobilized enzymes in analytical chemistry. In, Advances in Biochemical Engineering, Volume 15. Springer Berlin Heidelberg, pp. 89-129.
Cao, L., 2006. Covalent Enzyme Immobilization. In, Carrier-bound Immobilized Enzymes. Wiley-VCH Verlag GmbH & Co. KGaA, pp. 169-316.
Champ, M., Langkilde, A.-M., Brouns, F., Kettlitz, B., Collet, Y.L.B., 2003. Advances in dietary fibre characterisation. 1. Definition of dietary fibre, physiological relevance, health benefits and analytical aspects. Nutrition Research Reviews 16 (01), 71-82.
Choi, H.-J., Kim, C.S., Kim, P., Jung, H.-C., Oh, D.-K., 2004. Lactosucrose Bioconversion from Lactose and Sucrose by Whole Cells of Paenibacillus polymyxa Harboring Levansucrase Activity. Biotechnology Progress 20 (6), 1876-1879.

146
Chouyyok, W., Panpranot, J., Thanachayanant, C., Prichanont, S., 2009. Effects of pH and pore characters of mesoporous silicas on horseradish peroxidase immobilization. Journal of Molecular Catalysis B: Enzymatic 56 (4), 246-252.
Crittenden, R.G., Playne, M.J., 1996. Production, properties and applications of food-grade oligosaccharides. Trends in Food Science & Technology 7 (11), 353-361.
Dashevsky, A., 1998. Protein loss by the microencapsulation of an enzyme (lactase) in alginate beads. International Journal of Pharmaceutics 161 (1), 1-5.
Daude, D., Remaud-Simeon, M., Andre, I., 2012. Sucrose analogs: an attractive (bio)source for glycodiversification. Natural Product Reports 29 (9), 945-960.
Desch, R.J., Kim, J., Thiel, S.W., 2014. Interactions between biomolecules and an iron-silica surface. Microporous and Mesoporous Materials 187, 29-39.
Domínguez, E., Nilsson, M., Hahn-Hägerdal, B., 1988. Carbodiimide coupling of β-galactosidase from Aspergillus oryzae to alginate. Enzyme and Microbial Technology 10 (10), 606-610.
Emaga, T.H., Rabetafka, N., Blecker, C.S., Paquot, M., 2012. Kinetics of the hydrolysis of polysaccharide galacturonic acid and neutral sugars chains from flaxseed mucilage. Biotechnology, Agronomy and Society and Environment 16 (2), 139-147.
End, N., Schöning, K.-U., 2004. Immobilized Biocatalysts in Industrial Research and Production. In: Kirschning, A. (Ed.), Immobilized Catalysts. Springer Berlin Heidelberg, pp. 273-317.
End, N., Schöning, K.-U., 2004. Immobilized Catalysts in Industrial Research and Application. In: Kirschning, A. (Ed.), Immobilized Catalysts. Springer Berlin Heidelberg, pp. 241-271.
Falahati, M., Saboury, A., Shafiee, A., Ma’mani, L., Moosavi-Movahedi, A., 2012. Immobilization of superoxide dismutase onto ordered mesoporous silica nanoparticles and improvement of its stability. Journal of the Iranian Chemical Society 9 (2), 157-161.
Fan, J., Yu, C., Lei, J., Zhang, Q., Li, T., Tu, B., Zhou, W., Zhao, D., 2005. Low-Temperature Strategy to Synthesize Highly Ordered Mesoporous Silicas with Very Large Pores. Journal of the American Chemical Society 127 (31), 10794-10795.
Feigin, L.A., Svergun, D.I., Taylor, G., 1987. Determination of the Integral Parameters of Particles. In: Taylor, G. (Ed.), Structure Analysis by Small-Angle X-Ray and Neutron Scattering. Springer US, pp. 59-105.
Freitas, F.F., Marquez, L.D.S., Ribeiro, G.P., Brandão, G.C., Cardoso, V.L., Ribeiro, E.J., 2011. A comparison of the kinetic properties of free and immobilized Aspergillus oryzae β-galactosidase. Biochemical Engineering Journal 58–59 (0), 33-38.
Fuchsbauer, H.L., Gerber, U., Engelmann, J., Seeger, T., Sinks, C., Hecht, T., 1996. Influence of gelatin matrices cross-linked with transglutaminase on the properties of an enclosed bioactive material using β-galactosidase as model system. Biomaterials 17 (15), 1481-1488.
Fujita, K., Hara, K., Hashimoto, H., Kitahata, S., 1990. Transfructosylation Catalyzed by β-Fructofurariosidase I from Arthrobacter sp. K-1(Biological Chemistry). Agricultural and biological chemistry 54 (10), 2655-2661.
Fusako, T., KAZUHITO, R., KAWAKAMI., Y., FUJIMURA., Y., UCHIDA., J., OKU., K., OKA., M., YONEYAMA., M., 1996. Effect of 4G-β -D-galactosylsucrose (lactosucrose) on fecal microflora in patients with chronic inflammatory bowel disease. Journal of Gastroenterol 31, 33-39.

147
Gänzle, M.G., 2012. Enzymatic synthesis of galacto-oligosaccharides and other lactose derivatives (hetero-oligosaccharides) from lactose. International Dairy Journal 22 (2), 116-122.
Gänzle, M.G., 2011. Lactose and Oligosaccharides | Lactose: Derivatives. In: Fuquay, J.W. (Ed.), Encyclopedia of Dairy Sciences (Second Edition). Academic Press, San Diego, pp. 202-208.
Gänzle, M.G., Haase, G., Jelen, P., 2008. Lactose: Crystallization, hydrolysis and value-added derivatives. International Dairy Journal 18 (7), 685-694.
Gaur, R., Pant, H., Jain, R., Khare, S.K., 2006. Galacto-oligosaccharide synthesis by immobilized Aspergillus oryzae β-galactosidase. Food Chemistry 97 (3), 426-430.
Guerrero, C., Vera, C., Plou, F., Illanes, A., 2011. Influence of reaction conditions on the selectivity of the synthesis of lactulose with microbial β-galactosidases. Journal of Molecular Catalysis B: Enzymatic 72 (3-4), 206-212.
Guimarães, P.M.R., Teixeira, J.A., Domingues, L., 2010. Fermentation of lactose to bio-ethanol by yeasts as part of integrated solutions for the valorisation of cheese whey. Biotechnology Advances 28 (3), 375-384.
Güleç, H., Gürdaş, S., Albayrak, N., Mutlu, M., 2010. Immobilization of Aspergillus oryzae β-galactosidase on low-pressure plasma-modified cellulose acetate membrane using polyethyleneimine for production of galactooligosaccharide. Biotechnology and Bioprocess Engineering 15 (6), 1006-1015.
Hamed, R.B., Gomez-Castellanos, J.R., Henry, L., Ducho, C., McDonough, M.A., Schofield, C.J., 2013. The enzymes of [small beta]-lactam biosynthesis. Natural Product Reports 30 (1), 21-107.
Han, W.-C., Sun-Ho Byun, Mi-Hyun Kim, Eun Hwa Sohn, Jung Dae Lim, Byung Hun Um, Chul Ho Kim, Soon Ah Kang, and Ki-Hyo Jang, 2009. Production of Lactosucrose from Sucrose and Lactose by a Levansucrase from Zymomonas mobilis. Journal of Microbiology and Biotechnology 19, 1153–1160.
Honda K1, M.T., Kuroki F, Iida M, Oka M, Sawatani I, 1999. Protective effect of lactosucrose on intracolonic indomethacin-induced small-intestinal ulcers in rats. Scandinavian Journal of Gastroenterology 34 (3), 264-269.
Hu, Z.-C., Korus, R., Stormo, K., 1993. Characterization of immobilized enzymes in polyurethane foams in a dynamic bed reactor. Applied Microbiology and Biotechnology 39 (3), 289-295.
Huang, L., Kruk, M., 2012. Synthesis of ultra-large-pore FDU-12 silica using ethylbenzene as micelle expander. Journal of Colloid and Interface Science 365 (1), 137-142.
Hudson, S., Magner, E., Cooney, J., Hodnett, B.K., 2005. Methodology for the Immobilization of Enzymes onto Mesoporous Materials. The Journal of Physical Chemistry B 109 (41), 19496-19506.
Huerta, L.M., Vera, C., Guerrero, C., Wilson, L., Illanes, A., 2011. Synthesis of galacto-oligosaccharides at very high lactose concentrations with immobilized β-galactosidases from Aspergillus oryzae. Process Biochemistry 46 (1), 245-252.
Humphrey, H.P., Wright, P.A., 2004. Nanoporous materials as supports for enzyme immobilization. In: Lu, G.Q.Z., X.S. (Ed.), Nanoporous Materials - Science and Engineering. World Scientific, pp. 849-872.
Husain, Q., 2010. β Galactosidases and their potential applications: a review. Critical Reviews in Biotechnology 30 (1), 41–62.
Husain, Q., Ansari, S.A., Alam, F., Azam, A., 2011. Immobilization of Aspergillus oryzae β galactosidase on zinc oxide nanoparticles via simple adsorption mechanism. International Journal of Biological Macromolecules 49 (1), 37-43.

148
Hussein, H.S., Flickinger, E.A., Fahey, G.C., 1999. Petfood Applications of Inulin and Oligofructose. The Journal of Nutrition 129 (7), 1454S-1456s.
Irazoqui, G., Giacomini, C., Batista-Viera, F., Brena, B.M., Cardelle-Cobas, A., Corzo, N., Jimeno, M.L., 2009. Characterization of Galactosyl Derivatives Obtained by Transgalactosylation of Lactose and Different Polyols Using Immobilized β-Galactosidase from Aspergillus oryzae. Journal of Agricultural and Food Chemistry 57 (23), 11302-11307.
Ito, M., Kimura, M., 1993. Influence of Lactose on Faecal Microflora in Lactose Maldigestors. Microbial Ecology in Health and Disease 6 (2), 73-76.
Joiner, B.L., Rosenblatt, J.R., 1971. Some Properties of the Range in Samples from Tukey's Symmetric Lambda Distributions. Journal of the American Statistical Association 66 (334), 394-399.
Kannan, K., Jasra, R.V., 2009. Immobilization of alkaline serine endopeptidase from Bacillus licheniformis on SBA-15 and MCF by surface covalent binding. Journal of Molecular Catalysis B: Enzymatic 56 (1), 34-40.
Kawase, M., Pilgrim, A., Araki, T., Hashimoto, K., 2001. Lactosucrose production using a simulated moving bed reactor. Chemical Engineering Science 56 (2), 453-458.
Kim, J., Jia, H., Lee, C.-w., Chung, S.-w., Kwak, J.H., Shin, Y., Dohnalkova, A., Kim, B.-G., Wang, P., Grate, J.W., 2006. Single enzyme nanoparticles in nanoporous silica: A hierarchical approach to enzyme stabilization and immobilization. Enzyme and Microbial Technology 39 (3), 474-480.
Kim, M.I., Kim, J., Lee, J., Jia, H., Na, H.B., Youn, J.K., Kwak, J.H., Dohnalkova, A., Grate, J.W., Wang, P., Hyeon, T., Park, H.G., Chang, H.N., 2007. Crosslinked enzyme aggregates in hierarchically-ordered mesoporous silica: A simple and effective method for enzyme stabilization. Biotechnology and Bioengineering 96 (2), 210-218.
Kim, M.I., Kim, J., Lee, J., Shin, S., Na, H.B., Hyeon, T., Park, H.G., Chang, H.N., 2008. One-dimensional crosslinked enzyme aggregates in SBA-15: Superior catalytic behavior to conventional enzyme immobilization. Microporous and Mesoporous Materials 111 (1–3), 18-23.
Kosikowski, F.V., 1979. Whey Utilization and Whey Products. Journal of Dairy Science 62 (7), 1149-1160.
Kruk, M., Hui, C.M., 2008. Synthesis and characterization of large-pore FDU-12 silica. Microporous and Mesoporous Materials 114 (1-3), 64-73.
Lee, C.H., Lin, T.S., Mou, C.Y., 2009. Mesoporous materials for encapsulating enzymes. Nano Today 4 (2), 165-179.
Lee, J., Kim, J., Kim, J., Jia, H., Kim, M.I., Kwak, J.H., Jin, S., Dohnalkova, A., Park, H.C., Chang, H.N., Wang, P., Grate, J.W., Hyeon, T., 2005. Simple synthesis of hierarchically ordered mesocellular mesoporous silica materials hosting crosslinked enzyme aggregates. Small 1 (7), 744-753.
Li, W., Xiang, X., Tang, S., Hu, B., Tian, L., Sun, Y., Ye, H., Zeng, X., 2009. Effective Enzymatic Synthesis of Lactosucrose and Its Analogues by β-d-Galactosidase from Bacillus circulans. Journal of Agricultural and Food Chemistry 57 (9), 3927-3933.
López-Gallego, F., Betancor, L., Mateo, C., Hidalgo, A., Alonso-Morales, N., Dellamora-Ortiz, G., Guisán, J.M., Fernández-Lafuente, R., 2005. Enzyme stabilization by glutaraldehyde crosslinking of adsorbed proteins on aminated supports. Journal of Biotechnology 119 (1), 70-75.
Luckarift -Heather, R., Spain- Jim, C., 2008. Continuous-Flow Applications of Silica-Encapsulated Enzymes. In, Biomolecular Catalysis. American Chemical Society, pp. 243-253.

149
Lynch, J., 2001. ANALYSE PHYSICO-CHIMIQUE DES CATALYSEURS INDUSTRIELS: MANUEL PRATIQUE DE CARACTÉRISATION. In: Bartling, S., Friesike, S. (Eds.), Opening Science. Springer International Publishing.
Marriott, B., 2000. Functional Foods: an Ecologic Perspective. Am J Clin Nutr 71(6 Suppl), 1728S-1734S.
Martínez-Villaluenga, C., Cardelle-Cobas, A., Corzo, N., Olano, A., Villamiel, M., 2008. Optimization of conditions for galactooligosaccharide synthesis during lactose hydrolysis by β-galactosidase from Kluyveromyces lactis (Lactozym 3000 L HP G). Food Chemistry 107 (1), 258-264.
Mawson, A.J., 1994. Bioconversions for whey utilization and waste abatement. Bioresource Technology 47 (3), 195-203.
McDonald, N., Novembre 2006. Profil sectoriel: le fromage. Agriculture et agroalimentaire canada; Centre canadien d’information laitière (CCIL); Euromonitor, Dairy Products in Canada, février 2006; Faostat; Global Trade Atlas, novembre 2006; Commission canadienne du lait, Ingrédients laitiers; Statistique Canada.
Menrad, K., 2003. Market and marketing of functional food in Europe. Journal of Food Engineering 56 (2–3), 181-188.
Mizote, A., Taniguchi, Y., Takei, Y., Koya-Miyata, S., Kohno, K., Iwaki, K., Kurose, M., Oku, K., Chaen, H., Fukuda, S., 2009. Lactosucrose Inhibits Body Fat Accumulation in Rats by Decreasing Intestinal Lipid Absorption. Bioscience, Biotechnology, and Biochemistry 73 (3), 582-587.
Mohy Eldin, M.S., El-Aassar, M.R., El. Zatahry, A.A., Al-Sabah, M.M.B., 2014. Covalent Immobilization of β-Galactosidase onto Amino-Functionalized Polyvinyl Chloride Microspheres: Enzyme Immobilization and Characterization. Advances in Polymer Technology 33 (1), n/a-n/a.
Mu, W., Chen, Q., Wang, X., Zhang, T., Jiang, B., 2013. Current studies on physiological functions and biological production of lactosucrose. Applied Microbiology and Biotechnology 97 (16), 7073-7080.
Mussatto, S.I., Mancilha, I.M., 2007. Non-digestible oligosaccharides: A review. Carbohydrate Polymers 68 (3), 587-597.
Neri, D.F.M., Balcão, V.M., Cardoso, S.M., Silva, A.M.S., Domingues, M.D.R.M., Torres, D.P.M., Rodrigues, L.R.M., Carvalho, L.B., Teixeira, J.A.C., 2011. Characterization of galactooligosaccharides produced by β-galactosidase immobilized onto magnetized Dacron. International Dairy Journal 21 (3), 172-178.
Ohkusa, T.O., Yoshinori; Sato, Chifui; Mikuni, Katsuhiko; Ikeda, Hiroshi, 1995. Long-term ingestion of lactosucrose increases Bifidobacterium sp in human fecal flora. Digestion 56, 415–420.
Oliveira, C., Guimarães, P.M.R., Domingues, L., 2011. Recombinant microbial systems for improved β-galactosidase production and biotechnological applications. Biotechnology Advances 29 (6), 600-609.
Pandya, P.H., Jasra, R.V., Newalkar, B.L., Bhatt, P.N., 2005. Studies on the activity and stability of immobilized α-amylase in ordered mesoporous silicas. Microporous and Mesoporous Materials 77 (1), 67-77.
Panesar, P.S., Kennedy, J.F., 2012. Biotechnological approaches for the value addition of whey. Critical Reviews in Biotechnology 32 (4), 327-348.
Panesar, P.S., Kumari, S., Panesar, R., 2010. Potential applications of immobilized β-galactosidase in food processing industries. Enzyme Research 2010.

150
Panesar, P.S., Panesar, R., Singh, R.S., Kennedy, J.F., Kumar, H., 2006. Microbial production, immobilization and applications of β-D-galactosidase. Journal of Chemical Technology & Biotechnology 81 (4), 530-543.
Parhi, P., Manivannan, V., Kohli, S., McCurdy, P., 2008. Synthesis and characterization of M3V2O8 (M = Ca, Sr and Ba) by a solid-state metathesis approach. Bulletin of Materials Science 31 (6), 885-890.
Park, Y.K., De Santi, M.S.S., Pastore, G.M., 1979. Production and characteriza of β-galactosidase from aspergillus oryzae. Journal of Food Science 44 (1), 100-103.
Pereira-Rodríguez, Á., Fernández-Leiro, R., González-Siso, M.I., Cerdán, M.E., Becerra, M., Sanz-Aparicio, J., 2012. Structural basis of specificity in tetrameric Kluyveromyces lactis β-galactosidase. Journal of Structural Biology 177 (2), 392-401.
Qiang, X., YongLie, C., QianBing, W., 2009. Health benefit application of functional oligosaccharides. Carbohydrate Polymers 77 (3), 435-441.
Richmond, M.L., Gray, J.I., Stine, C.M., 1981. Beta-Galactosidase: Review of Recent Research Related to Technological Application, Nutritional Concerns, and Immobilization. Journal of Dairy Science 64 (9), 1759-1771.
Sako, T., Matsumoto, K., Tanaka, R., 1999. Recent progress on research and applications of non-digestible galacto-oligosaccharides. International Dairy Journal 9 (1), 69-80.
Salis, A., Meloni, D., Ligas, S., Casula, M.F., Monduzzi, M., Solinas, V., Dumitriu, E., 2005. Physical and Chemical Adsorption of Mucor javanicus Lipase on SBA-15 Mesoporous Silica. Synthesis, Structural Characterization, and Activity Performance. Langmuir 21 (12), 5511-5516.
Schmidt-Winkel, P., Lukens, W.W., Zhao, D., Yang, P., Chmelka, B.F., Stucky, G.D., 1999. Mesocellular Siliceous Foams with Uniformly Sized Cells and Windows. Journal of the American Chemical Society 121 (1), 254-255.
Schmidt-Winkel, P., Lukens, W.W., Zhao, D., Yang, P., Chmelka, B.F., Stucky, G.D., 1998. Mesocellular Siliceous Foams with Uniformly Sized Cells and Windows. Journal of the American Chemical Society 121 (1), 254-255.
Seki, N., Saito, H., 2012. Lactose as a source for lactulose and other functional lactose derivatives. International Dairy Journal 22 (2), 110-115.
Sheik-Asraf, S., Gunasekaran, P., 2010. Current trends of ß-galactosidase research and application. Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology, 2 (Antonio Mendez Vilas (Ed.)), 880-890.
Siso, M.I.G., 1996. The biotechnological utilization of cheese whey: A review. Bioresource Technology 57 (1), 1-11.
Smith, P.K., Krohn, R.I., Hermanson, G.T., Mallia, A.K., Gartner, F.H., Provenzano, M.D., Fujimoto, E.K., Goeke, N.M., Olson, B.J., Klenk, D.C., 1985. Measurement of protein using bicinchoninic acid. Analytical Biochemistry 150 (1), 76-85.
Smithers, G.W., 2008. Whey and whey proteins—From ‘gutter-to-gold’. International Dairy Journal 18 (7), 695-704.
Smithers, G.W., John Ballard, F., Copeland, A.D., de Silva, K.J., Dionysius, D.A., Francis, G.L., Goddard, C., Grieve, P.A., McIntosh, G.H., Mitchell, I.R., Pearce, R.J., Regester, G.O., 1996. New Opportunities from the Isolation and Utilization of Whey Proteins. Journal of Dairy Science 79 (8), 1454-1459.
Takahashi, H., Miyazaki-Imamura, C., 2008. Enzyme Stabilization Involving Molecular Evolution and Immobilization in Mesoporous Materials. In, Biomolecular Catalysis. American Chemical Society, pp. 61-75.

151
Terada, A., Hara, H., Oishi, T., Matsui, S., Mitsuoka, T., Nakajyo, S., Fujimori, I., Hara, K., 1992. Effect of Dietary Lactosucrose on Faecal Flora and Faecal Metabolites of Dogs. Microbial Ecology in Health and Disease 5 (2), 87-92.
Teramoto, F., Rokutan, K., Kawakami, Y., Fujimura, Y., Uchida, J., Oku, K., Oka, M., Yoneyama, M., 1996. Effect of 4G-β-D-galactosylsucrose (lactosucrose) on fecal microflora in patients with chronic inflammatory bowel disease. Journal of Gastroenterology 31 (1), 33-39.
Teramoto, F., Rokutan, K., Sugano, Y., Oku, K., Kishino, E., Fujita, K., Hara, K., Kishi, K., Fukunaga, M., Morita, T., 2006. Long-Term Administration of 4G-β -D-Galactosylsucrose (Lactosucrose) Enhances Intestinal Calcium Absorption in Young Women: A Randomized, Placebo-Controlled 96-wk Study. Journal of Nutritional Science and Vitaminology 52 (5), 337-346.
Torres, D.P.M., Gonçalves, M.d.P.F., Teixeira, J.A., Rodrigues, L.R., 2010. Galacto-Oligosaccharides: Production, Properties, Applications, and Significance as Prebiotics. Comprehensive Reviews in Food Science and Food Safety 9 (5), 438-454.
Trevisan, H.C., Bergamo, E.P., Contiero, J., Hojo, O., Mont, R., 1997. β-galactosidase immobilization on controlled pore silica. Brazilian Journal of Chemical Engineering 14 (4), 315-319.
Urrego, S., Serra, E., Alfredsson, V., Blanco, R.M., Díaz, I., 2010. Bottle-around-the-ship: A method to encapsulate enzymes in ordered mesoporous materials. Microporous and Mesoporous Materials 129 (1–2), 173-178.
Urrutia, P., Rodriguez-Colinas, B., Fernandez-Arrojo, L., Ballesteros, A.O., Wilson, L., Illanes, A., Plou, F.J., 2013. Detailed Analysis of Galactooligosaccharides Synthesis with β-Galactosidase from Aspergillus oryzae. Journal of Agricultural and Food Chemistry 61 (5), 1081-1087.
Vera, C., Guerrero, C., Illanes, A., 2011. Determination of the transgalactosylation activity of Aspergillus oryzae β-galactosidase: effect of pH, temperature, and galactose and glucose concentrations. Carbohydrate Research 346 (6), 745-752.
Vilain, A.-C., 2010. Qu'est-ce que le lait ? = What's milk? Revue française d'allergologie 50 (3), 124-127.
Vinu, A., Murugesan, V., Tangermann, O., Hartmann, M., 2004. Adsorption of Cytochrome c on Mesoporous Molecular Sieves: Influence of pH, Pore Diameter, and Aluminum Incorporation. Chemistry of Materials 16 (16), 3056-3065.
Wu, Z., Wang, Z., Guan, B., Wang, X., Zhang, Y., Xiao, Y., Zhi, B., Liu, Y., Li, Z., Huo, Q., 2013. Improving the properties of [small beta]-galactosidase from Aspergillus oryzae via encapsulation in aggregated silica nanoparticles. New Journal of Chemistry 37 (11), 3793-3797.
Yiu, H.H.P., Wright, P.A., 2005. Enzymes supported on ordered mesoporous solids: a special case of an inorganic-organic hybrid. Journal of Materials Chemistry 15 (35-36), 3690-3700.
Zhang, N., Cao, J., Peng, Z., 2001. Characteristics of β-D-fructofurannosidase in Organic Solvent-water Biphasic System. Food and Fermentation Industries.
Zhang Ning, Cao Jinsong, Zhiying, P., 2001. Characteristics of β-D-fructofurannosidase in Organic Solvent-water Biphasic System. Food and Fermentation Industries.
Zhang Ning, Cao Jinsong, Zhiying, P., 2001. Characteristics of β-D-fructofurannosidase in Organic Solvent-water Biphasic System. Food and Fermentation Industries 49, 122-127.
Zhao, D., Feng, J., Huo, Q., Melosh, N., Fredrickson, G.H., Chmelka, B.F., Stucky, G.D., 1998. Triblock Copolymer Syntheses of Mesoporous Silica with Periodic 50 to 300 Angstrom Pores. Science 279 (5350), 548-552.

152
Zhao, X., 2006. Synthesis and characterization of a novel hyaluronic acid hydrogel. Journal of Biomaterials Science, Polymer Edition 17 (4), 419-433.
Zhou, G., Chen, Y., Yang, J., Yang, S., 2007. From cylindrical-channel mesoporous silica to vesicle-like silica with well-defined multilamella shells and large inter-shell mesopores. Journal of Materials Chemistry 17 (27), 2839-2844.
Zhou, G., Chen, Y., Yang, S., 2009. Comparative studies on catalytic properties of immobilized Candida rugosa lipase in ordered mesoporous rod-like silica and vesicle-like silica. Microporous and Mesoporous Materials 119 (1–3), 223-229.
Zhou, Q.Z.K., Chen, X.D., 2001. Effects of temperature and pH on the catalytic activity of the immobilized β-galactosidase from Kluyveromyces lactis. Biochemical Engineering Journal 9 (1), 33-40.

153
Annexes
Annexe 1 : La présence de glucose et de galactose seulement à la fin de la réaction (l’hydrolyse de tous les
produits)

154
Annexe 2: Les résultats de SAS pour l’effet de pH sur la rétention des matériaux.
Fichier de données phm
Obs mat pH rep ret
1 1 5.0 1 93.7
2 1 5.0 2 94.1
3 1 5.0 3 94.1
4 1 7.0 1 77.0
5 1 7.0 2 58.6
6 1 7.0 3 69.7
7 1 3.5 1 89.2
8 1 3.5 2 90.9
9 1 3.5 3 90.8
10 2 5.0 1 80.5
11 2 5.0 2 89.8
12 2 5.0 3 85.0
13 2 7.0 1 63.7
14 2 7.0 2 67.4
15 2 7.0 3 67.4
16 2 3.5 1 85.2
17 2 3.5 2 85.2
18 2 3.5 3 89.5
19 3 5.0 1 93.7
20 3 5.0 2 95.3
21 3 5.0 3 89.1
22 3 7.0 1 72.1
23 3 7.0 2 78.0
24 3 7.0 3 80.0
25 3 3.5 1 90.2

155
Obs mat pH rep ret
26 3 3.5 2 89.9
27 3 3.5 3 91.3
28 4 5.0 1 96.7
29 4 5.0 2 96.8
30 4 5.0 3 89.9
31 4 7.0 1 73.0
32 4 7.0 2 57.9
33 4 7.0 3 69.8
34 4 3.5 1 95.6
35 4 3.5 2 94.0
36 4 3.5 3 92.8
37 5 5.0 1 98.8
38 5 5.0 2 91.8
39 5 5.0 3 99.4
40 5 7.0 1 78.2
41 5 7.0 2 76.7
42 5 7.0 3 71.9
43 5 3.5 1 88.9
44 5 3.5 2 90.6
45 5 3.5 3 89.8
46 6 5.0 1 90.4
47 6 5.0 2 94.4
48 6 5.0 3 96.2
49 6 7.0 1 77.2
50 6 7.0 2 75.4
51 6 7.0 3 81.0
52 6 3.5 1 91.5
53 6 3.5 2 90.7
54 6 3.5 3 92.9
55 7 5.0 1 87.6
56 7 5.0 2 93.7

156
Obs mat pH rep ret
57 7 5.0 3 96.8
58 7 7.0 1 62.6
59 7 7.0 2 76.5
60 7 7.0 3 78.4
61 7 3.5 1 89.9
62 7 3.5 2 91.2
63 7 3.5 3 90.2
Comparaisaon des materiaux en fonction du pH
The GLM Procedure
Class Level Information
Class Levels Values
rep 3 1 2 3
mat 7 1 2 3 4 5 6 7
pH 3 3.5 5 7
Number of Observations Read 63
Number of Observations Used 63
Comparaisaon des materiaux en fonction du pH
The GLM Procedure
Dependent Variable: ret
Source DF Sum of Squares Mean Square F Value Pr > F
Model 20 6186.002222 309.300111 17.45 <.0001
Error 42 744.560000 17.727619
Corrected Total 62 6930.562222
R-Square Coeff Var Root MSE ret Mean
0.892569 4.948258 4.210418 85.08889

157
Source DF Type I SS Mean Square F Value Pr > F
mat 6 438.793333 73.132222 4.13 0.0024
pH 2 5430.653651 2715.326825 153.17 <.0001
mat*pH 12 316.555238 26.379603 1.49 0.1671
Source DF Type III SS Mean Square F Value Pr > F
mat 6 438.793333 73.132222 4.13 0.0024
pH 2 5430.653651 2715.326825 153.17 <.0001
mat*pH 12 316.555238 26.379603 1.49 0.1671

158
Comparaisaon des materiaux en fonction du pH
The GLM Procedure

159
Comparaisaon des materiaux en fonction du pH
The GLM Procedure
Tukey's Studentized Range (HSD) Test for ret
Note: This test controls the Type I experimentwise error rate, but it generally has a higher Type II error rate than REGWQ.
Alpha 0.05
Error Degrees of Freedom 42
Error Mean Square 17.72762
Critical Value of Studentized Range 4.37776
Minimum Significant Difference 6.1441
Means with the same letter are not significantly different.
Tukey Grouping Mean N mat
A 87.744 9 6
A
A 87.344 9 5
A
A 86.622 9 3
A
B A 85.211 9 7
B A
B A 85.167 9 4
B A
B A 84.233 9 1
B
B 79.300 9 2

160
Comparaisaon des materiaux en fonction du pH
The GLM Procedure
Comparaisaon des materiaux en fonction du pH
The GLM Procedure
Tukey's Studentized Range (HSD) Test for ret
Alpha 0.05
Error Degrees of Freedom 42
Error Mean Square 17.72762
Critical Value of Studentized Range 3.43580
Minimum Significant Difference 3.1568
Means with the same letter are not significantly different.
Tukey Grouping Mean N pH

161
Means with the same letter are not significantly different.
Tukey Grouping Mean N pH
A 92.752 21 5
A
A 90.490 21 3.5
B 72.024 21 7
Vérification de la normalité des résidus
The UNIVARIATE Procedure Variable: rret
Moments
N 63 Sum Weights 63
Mean 0 Sum Observations 0
Std Deviation 1.00803226 Variance 1.01612903
Skewness -0.7307808 Kurtosis 1.56679638
Uncorrected SS 63 Corrected SS 63
Coeff Variation . Std Error Mean 0.12700013
Basic Statistical Measures
Location Variability
Mean 0.00000 Std Deviation 1.00803
Median 0.14544 Variance 1.01613
Mode -1.33807 Range 5.37167
Interquartile Range 1.03749
Note: The mode displayed is the smallest of 4 modes with a count of 2.

162
Tests for Location: Mu0=0
Test Statistic p Value
Student's t t 0 Pr > |t| 1.0000
Sign M 3.5 Pr >= |M| 0.4500
Signed Rank S 103 Pr >= |S| 0.4851
Tests for Normality
Test Statistic p Value
Shapiro-Wilk W 0.944001 Pr < W 0.0064
Kolmogorov-Smirnov D 0.13323 Pr > D <0.0100
Cramer-von Mises W-Sq 0.214887 Pr > W-Sq <0.0050
Anderson-Darling A-Sq 1.226921 Pr > A-Sq <0.0050
Quantiles (Definition 5)
Quantile Estimate
100% Max 2.491910
99% 2.491910
95% 1.367157
90% 0.959919
75% Q3 0.649642
50% Median 0.145442
25% Q1 -0.387846
10% -1.338068
5% -1.483510
1% -2.879756
0% Min -2.879756
Extreme Observations
Lowest Highest
Value Obs Value Obs
-2.87976 58 1.19263 57
-2.86036 5 1.36716 11
-2.61796 32 1.71622 60

163
Extreme Observations
Lowest Highest
Value Obs Value Obs
-1.48351 55 1.77439 31
-1.41564 38 2.49191 4
Stem Leaf # Boxplot
2 5 1 0
2
1 78 2 |
1 0224 4 |
0 6677888889 10 +-----+
0 000122222333344444 18 *--+--*
-0 444333222111100 15 +-----+
-0 77 2 |
-1 4333100 7 |
-1 5 1 |
-2
-2 996 3 0
----+----+----+----+
Normal Probability Plot
2.75+ +
| +++*+
| ++*+*
| +++*+**
| +********
| ********
| *******+
| ***+++
| **+****
| +++++
| +++++
-2.75++ * * *
+----+----+----+----+----+----+----+----+----+----+
-2 -1 0 +1 +2

164
Annexe 3 : Résultats de SAS pour l’effet de Concentration d’enzyme sur la rétention
Fichier de données Ezm
Obs mat Ez rep ret
1 1 25 1 93.2
2 1 25 2 95.2
3 1 25 3 94.1
4 1 60 1 94.0
5 1 60 2 93.7
6 1 60 3 92.8
7 1 75 1 94.4
8 1 75 2 93.6
9 1 75 3 94.2
10 1 100 1 93.6
11 1 100 2 91.8
12 1 100 3 94.1
13 2 25 1 90.5
14 2 25 2 91.8
15 2 25 3 93.2
16 2 60 1 89.8
17 2 60 2 93.8
18 2 60 3 91.5
19 2 75 1 94.6
20 2 75 2 94.0
21 2 75 3 92.8
22 2 100 1 93.6
23 2 100 2 91.5
24 2 100 3 90.4
25 3 25 1 82.6
26 3 25 2 86.1

165
Obs mat Ez rep ret
27 3 25 3 84.6
28 3 60 1 85.3
29 3 60 2 93.6
30 3 60 3 92.6
31 3 75 1 96.1
32 3 75 2 94.2
33 3 75 3 94.3
34 3 100 1 95.6
35 3 100 2 94.8
36 3 100 3 93.9
37 4 25 1 87.5
38 4 25 2 90.2
39 4 25 3 89.0
40 4 60 1 96.7
41 4 60 2 93.9
42 4 60 3 92.8
43 4 75 1 97.2
44 4 75 2 96.3
45 4 75 3 96.9
46 4 100 1 95.9
47 4 100 2 96.5
48 4 100 3 94.8
49 5 25 1 88.2
50 5 25 2 88.7
51 5 25 3 89.3
52 5 60 1 96.7
53 5 60 2 97.3
54 5 60 3 95.6
55 5 75 1 93.7

166
Obs mat Ez rep ret
56 5 75 2 93.2
57 5 75 3 93.3
58 5 100 1 94.7
59 5 100 2 93.9
60 5 100 3 94.8
61 6 25 1 92.1
62 6 25 2 91.4
63 6 25 3 91.7
64 6 60 1 96.2
65 6 60 2 95.1
66 6 60 3 95.4
67 6 75 1 94.2
68 6 75 2 93.9
69 6 75 3 94.5
70 6 100 1 95.0
71 6 100 2 93.9
72 6 100 3 94.5
73 7 25 1 83.1
74 7 25 2 87.9
75 7 25 3 91.2
76 7 60 1 96.8
77 7 60 2 94.5
78 7 60 3 92.5
79 7 75 1 97.7
80 7 75 2 96.1
81 7 75 3 95.6
82 7 100 1 95.8
83 7 100 2 94.0
84 7 100 3 94.1

167
Comparaisaon des materiaux en fonction de Ez
The GLM Procedure
Class Level Information
Class Levels Values
rep 3 1 2 3
mat 7 1 2 3 4 5 6 7
Ez 4 25 60 75 100
Number of Observations Read 84
Number of Observations Used 84
Comparaisaon des materiaux en fonction de Ez
The GLM Procedure
Dependent Variable: ret
Source DF Sum of Squares Mean Square F Value Pr > F
Model 27 684.1995238 25.3407231 10.09 <.0001
Error 56 140.6400000 2.5114286
Corrected Total 83 824.8395238
R-Square Coeff Var Root MSE ret Mean
0.829494 1.702244 1.584749 93.09762
Source DF Type I SS Mean Square F Value Pr > F
mat 6 78.0478571 13.0079762 5.18 0.0003
Ez 3 352.6376190 117.5458730 46.80 <.0001
mat*Ez 18 253.5140476 14.0841138 5.61 <.0001
Source DF Type III SS Mean Square F Value Pr > F
mat 6 78.0478571 13.0079762 5.18 0.0003
Ez 3 352.6376190 117.5458730 46.80 <.0001
mat*Ez 18 253.5140476 14.0841138 5.61 <.0001

168

169
Comparaisaon des materiaux en fonction de Ez
The GLM Procedure
Comparaisaon des materiaux en fonction de Ez
The GLM Procedure
Tukey's Studentized Range (HSD) Test for ret
Alpha 0.05
Error Degrees of Freedom 56
Error Mean Square 2.511429
Critical Value of Studentized Range 4.32467
Minimum Significant Difference 1.9784

170
Means with the same letter
are not significantly different.
Tukey Grouping Mean N mat
A 93.9917 12 6
A
A 93.9750 12 4
A
A 93.7250 12 1
A
A 93.2833 12 5
A
A 93.2750 12 7
A
B A 92.2917 12 2
B
B 91.1417 12 3

171
Comparaisaon des materiaux en fonction de Ez
The GLM Procedure

172
Comparaisaon des materiaux en fonction de Ez
The GLM Procedure
Tukey's Studentized Range (HSD) Test for ret
Note: This test controls the Type I experimentwise error rate, but it generally has a higher Type II error rate than REGWQ.
Alpha 0.05
Error Degrees of Freedom 56
Error Mean Square 2.511429
Critical Value of Studentized Range 3.74468
Minimum Significant Difference 1.295
Means with the same letter
are not significantly different.
Tukey Grouping Mean N Ez
A 94.8000 21 75
A
A 94.1524 21 100
A
A 93.8381 21 60
B 89.6000 21 25

173
Comparaisaon des materiaux en fonction de Ez
The GLM Procedure
Level of
mat
Level of
Ez
N ret
Mean Std Dev
1 25 3 94.1666667 1.00166528
1 60 3 93.5000000 0.62449980
1 75 3 94.0666667 0.41633320
1 100 3 93.1666667 1.20968315
2 25 3 91.8333333 1.35030861
2 60 3 91.7000000 2.00748599
2 75 3 93.8000000 0.91651514
2 100 3 91.8333333 1.62583312
3 25 3 84.4333333 1.75594229

174
Level of
mat
Level of
Ez
N ret
Mean Std Dev
3 60 3 90.5000000 4.53100430
3 75 3 94.8666667 1.06926766
3 100 3 94.7666667 0.85049005
4 25 3 88.9000000 1.35277493
4 60 3 94.4666667 2.01080415
4 75 3 96.8000000 0.45825757
4 100 3 95.7333333 0.86216781
5 25 3 88.7333333 0.55075705
5 60 3 96.5333333 0.86216781
5 75 3 93.4000000 0.26457513
5 100 3 94.4666667 0.49328829
6 25 3 91.7333333 0.35118846
6 60 3 95.5666667 0.56862407
6 75 3 94.2000000 0.30000000
6 100 3 94.4666667 0.55075705
7 25 3 87.4000000 4.07308237
7 60 3 94.6000000 2.15174348
7 75 3 96.4666667 1.09696551
7 100 3 94.6333333 1.01159939

175
Vérification de l'homogénéité des variances
Plot of rret*pret. Legend: A = 1 obs, B = 2 obs, etc.
rret |
|
5.0 +
|
|
| A
2.5 + A
| A A B
| A A B A A
| A A BA AA BA AA A
0.0 + A AA AE D AF DA AA AAB
| A A AB EC AA AAA
| A A AB A A
| A
-2.5 +
| A
| A
|
-5.0 +
|
--+----------+----------+----------+----------+----------+---------
-+--
82.5 85.0 87.5 90.0 92.5 95.0
97.5
pret

176
Vérification de la normalité des résidus
The UNIVARIATE Procedure
Variable: rret
Moments
N 84 Sum Weights 84
Mean 0 Sum Observations 0
Std Deviation 1.00600606 Variance 1.01204819
Skewness -0.6411047 Kurtosis 3.95957293
Uncorrected SS 84 Corrected SS 84
Coeff Variation . Std Error Mean 0.10976426
Basic Statistical Measures
Location Variability
Mean 0.00000 Std Deviation 1.00601
Median 0.01288 Variance 1.01205
Mode -0.43794 Range 6.95549
Interquartile Range 0.86300
Note: The mode displayed is the smallest of 4 modes with a count of 2.
Tests for Location: Mu0=0
Test Statistic p Value
Student's t t 0 Pr > |t| 1.0000
Sign M 0.5 Pr >= |M| 1.0000
Signed Rank S 15 Pr >= |S| 0.9462
Tests for Normality
Test Statistic p Value
Shapiro-Wilk W 0.927041 Pr < W 0.0001
Kolmogorov-Smirnov D 0.105473 Pr > D 0.0212
Cramer-von Mises W-Sq 0.252547 Pr > W-Sq <0.0050
Anderson-Darling A-Sq 1.538443 Pr > A-Sq <0.0050

177
Quantiles (Definition 5)
Quantile Estimate
100% Max 2.9367624
99% 2.9367624
95% 1.6229476
90% 1.0562040
75% Q3 0.4250577
50% Median 0.0128805
25% Q1 -0.4379383
10% -1.0562040
5% -1.4168591
1% -4.0187275
0% Min -4.0187275
Extreme Observations
Lowest Highest
Value Obs Value Obs
-4.01873 28 1.62295 17
-3.32318 73 1.70023 76
-1.62295 78 1.72599 40
-1.46838 16 2.39578 29
-1.41686 25 2.93676 75

178
Stem Leaf # Boxplot
2 9 1 0
2 4 1 0
1 6677 4 0
1 000134 6 |
0 56666789 8 |
0 00011111122222333334444 23 +--+--+
-0 4444444443332221111000 22 +-----+
-0 877777555 9 |
-1 431110 6 |
-1 65 2 |
-2
-2
-3 3 1 *
-3
-4 0 1 *
----+----+----+----+---
Normal Probability Plot
2.75+ *+
| +*+++
| ***+*
| ++***
| +++****
| ********
| *********
-0.75+ ****+++
| ******++
| *++++
| +++++
|+
| *
|
-4.25+ *
+----+----+----+----+----+----+----+----+----+----+
-2 -1 0 +1 +2

179
Annexe 4 : Résultats de SAS pour l’effet de GA sur la rétention des enzymes
Fichier de données GAm
Obs mat GA rep ret
1 1 0.1 1 90.0
2 1 0.1 2 90.8
3 1 0.1 3 92.0
4 1 1.0 1 79.2
5 1 1.0 2 77.6
6 1 1.0 3 75.6
7 1 2.0 1 71.8
8 1 2.0 2 71.6
9 1 2.0 3 77.6
10 2 0.1 1 85.4
11 2 0.1 2 82.0
12 2 0.1 3 86.6
13 2 1.0 1 78.6
14 2 1.0 2 80.0
15 2 1.0 3 79.2
16 2 2.0 1 87.0
17 2 2.0 2 81.6
18 2 2.0 3 83.8
19 3 0.1 1 93.4
20 3 0.1 2 91.0
21 3 0.1 3 91.6
22 3 1.0 1 82.6
23 3 1.0 2 82.6
24 3 1.0 3 78.6
25 3 2.0 1 79.0
26 3 2.0 2 80.4

180
Obs mat GA rep ret
27 3 2.0 3 82.6
28 4 0.1 1 81.4
29 4 0.1 2 81.6
30 4 0.1 3 82.4
31 4 1.0 1 83.0
32 4 1.0 2 80.6
33 4 1.0 3 82.0
34 4 2.0 1 77.0
35 4 2.0 2 78.4
36 4 2.0 3 81.0
37 5 0.1 1 90.0
38 5 0.1 2 91.4
39 5 0.1 3 94.6
40 5 1.0 1 84.2
41 5 1.0 2 85.0
42 5 1.0 3 86.8
43 5 2.0 1 83.8
44 5 2.0 2 84.6
45 5 2.0 3 89.0
46 6 0.1 1 91.0
47 6 0.1 2 91.2
48 6 0.1 3 92.0
49 6 1.0 1 87.8
50 6 1.0 2 86.2
51 6 1.0 3 88.2
52 6 2.0 1 78.8
53 6 2.0 2 85.8
54 6 2.0 3 86.6
55 7 0.1 1 91.4

181
Obs mat GA rep ret
56 7 0.1 2 88.2
57 7 0.1 3 91.8
58 7 1.0 1 86.6
59 7 1.0 2 89.0
60 7 1.0 3 87.8
61 7 2.0 1 83.0
62 7 2.0 2 84.4
63 7 2.0 3 86.0
Comparaisaon des materiaux en fonction du GA
The GLM Procedure
Class Level Information
Class Levels Values
rep 3 1 2 3
mat 7 1 2 3 4 5 6 7
GA 3 0.1 1 2
Number of Observations Read 63
Number of Observations Used 63
Comparaisaon des materiaux en fonction du GA
The GLM Procedure
Dependent Variable: ret
Source DF Sum of Squares Mean Square F Value Pr > F
Model 20 1576.816508 78.840825 18.80 <.0001
Error 42 176.106667 4.193016
Corrected Total 62 1752.923175

182
R-Square Coeff Var Root MSE ret Mean
0.899535 2.422705 2.047685 84.52063
Source DF Type I SS Mean Square F Value Pr > F
mat 6 541.7942857 90.2990476 21.54 <.0001
GA 2 660.7098413 330.3549206 78.79 <.0001
mat*GA 12 374.3123810 31.1926984 7.44 <.0001
Source DF Type III SS Mean Square F Value Pr > F
mat 6 541.7942857 90.2990476 21.54 <.0001
GA 2 660.7098413 330.3549206 78.79 <.0001
mat*GA 12 374.3123810 31.1926984 7.44 <.0001

183
Comparaisaon des materiaux en fonction du GA
The GLM Procedure

184
Comparaisaon des materiaux en fonction du GA
The GLM Procedure
Tukey's Studentized Range (HSD) Test for ret
Alpha 0.05
Error Degrees of Freedom 42
Error Mean Square 4.193016
Critical Value of Studentized Range 4.37776
Minimum Significant Difference 2.9881
Means with the same letter
are not significantly different.
Tukey Grouping Mean N mat
A 87.7111 9 5
A
B A 87.5778 9 7
B A
B A 87.5111 9 6
B
B C 84.6444 9 3
C
D C 82.6889 9 2
D
D 80.8222 9 4
D
D 80.6889 9 1

185
Comparaisaon des materiaux en fonction du GA
The GLM Procedure

186
Comparaisaon des materiaux en fonction du GA
The GLM Procedure
Tukey's Studentized Range (HSD) Test for ret
Note: This test controls the Type I experimentwise error rate, but it generally has a higher Type II error rate than REGWQ.
Alpha 0.05
Error Degrees of Freedom 42
Error Mean Square 4.193016
Critical Value of Studentized Range 3.43580
Minimum Significant Difference 1.5353
Means with the same letter
are not significantly different.
Tukey Grouping Mean N GA
A 89.0381 21 0.1
B 82.9143 21 1
B
B 81.6095 21 2

187
Comparaisaon des materiaux en fonction du GA
The GLM Procedure
Level of
mat
Level of
GA
N ret
Mean Std Dev
1 0.1 3 90.9333333 1.00664459
1 1 3 77.4666667 1.80369990
1 2 3 73.6666667 3.40783411
2 0.1 3 84.6666667 2.38607069
2 1 3 79.2666667 0.70237692
2 2 3 84.1333333 2.71538825
3 0.1 3 92.0000000 1.24899960
3 1 3 81.2666667 2.30940108
3 2 3 80.6666667 1.81475435

188
Level of
mat
Level of
GA
N ret
Mean Std Dev
4 0.1 3 81.8000000 0.52915026
4 1 3 81.8666667 1.20554275
4 2 3 78.8000000 2.02977831
5 0.1 3 92.0000000 2.35796522
5 1 3 85.3333333 1.33166562
5 2 3 85.8000000 2.80000000
6 0.1 3 91.4000000 0.52915026
6 1 3 87.4000000 1.05830052
6 2 3 83.7333333 4.29107601
7 0.1 3 90.4666667 1.97315314
7 1 3 87.8000000 1.20000000
7 2 3 84.4666667 1.50111070
Vérification de l'homogénéité des variances
Vérification de la normalité des résidus
The UNIVARIATE Procedure
Variable: rret
Moments
N 63 Sum Weights 63
Mean 0 Sum Observations 0
Std Deviation 1.00803226 Variance 1.01612903
Skewness -0.096176 Kurtosis 0.16414867
Uncorrected SS 63 Corrected SS 63
Coeff Variation . Std Error Mean 0.12700013

189
Basic Statistical Measures
Location Variability
Mean 0.00000 Std Deviation 1.00803
Median -0.07975 Variance 1.01613
Mode -1.59496 Range 5.30326
Interquartile Range 1.51522
Note: The mode displayed is the smallest of 4 modes with a count of 2.
Tests for Location: Mu0=0
Test Statistic p Value
Student's t t 0 Pr > |t| 1.0000
Sign M -2.5 Pr >= |M| 0.6147
Signed Rank S -4 Pr >= |S| 0.9784
Tests for Normality
Test Statistic p Value
Shapiro-Wilk W 0.989649 Pr < W 0.8767
Kolmogorov-Smirnov D 0.056991 Pr > D >0.1500
Cramer-von Mises W-Sq 0.030067 Pr > W-Sq >0.2500
Anderson-Darling A-Sq 0.201602 Pr > A-Sq >0.2500
Quantiles (Definition 5)
Quantile Estimate
100% Max 2.3525731
99% 2.3525731
95% 1.7145872
90% 1.2360977
75% Q3 0.7974824
50% Median -0.0797482
25% Q1 -0.7177342
10% -1.1962236

190
Quantiles (Definition 5)
Quantile Estimate
5% -1.5152166
1% -2.9506849
0% Min -2.9506849
Extreme Observations
Lowest Highest
Value Obs Value Obs
-2.95068 52 1.55509 39
-1.59496 24 1.71459 54
-1.59496 11 1.71459 16
-1.51522 17 1.91396 45
-1.35572 56 2.35257 9

191
Stem Leaf # Boxplot
2 4 1 |
1 6779 4 |
1 02223 5 |
0 56677888899 11 +-----+
0 01124444 8 | + |
-0 44222222211100 14 *-----*
-0 98777766 8 +-----+
-1 42221110 8 |
-1 665 3 |
-2 |
-2 |
-3 0 1 |
----+----+----+----+
Normal Probability Plot
2.25+ +++*+
| ***+*
| ****+
| ******
| +*****
| *******
| *****
| *+****
| *+**+
| +++++
|+ *
-3.25+
+----+----+----+----+----+----+----+----+----+----+
-2 -1 0 +1 +2