
Biological and Functional Analysis of Biosimilar TNF α Drugs
description

Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 83
∆οµή καιλειτουργίατων µεγάλωνβιολογικώνµορίων
Με βάση τη µεγάλη πολυπλοκότητα που χαρα-κτηρίζει τη ζωή στη Γη, θα περίµενε κανείς ναυπάρχει µια εξίσου τεράστια ποικιλία µορίων
στους ζωντανούς οργανισµούς. Ωστόσο, είναι αξιοσηµεί-ωτο ότι τα σηµαντικότερα µεγάλα µόρια σε όλους τουςζωντανούς οργανισµούς –από τα βακτήρια µέχρι τουςελέφαντες– εντάσσονται σε τέσσερεις µόνο κύριες κατη-γορίες: στους υδατάνθρακες, στα λιπίδια, στις πρωτεΐνεςκαι στα νουκλεϊκά οξέα. Οι τρεις από τις τέσσερεις κατη-γορίες (υδατάνθρακες, πρωτεΐνες, νουκλεϊκά οξέα) έχουνµόρια µε πολύ µεγάλο µέγεθος και γι’ αυτό ονοµάζονταιµακροµόρια. Επί παραδείγµατι, µια πρωτεΐνη µπορεί νααποτελείται από χιλιάδες άτοµα τα οποία σχηµατίζουνέναν µοριακό κολοσσό, µε µάζα µεγαλύτερη των 100.000dalton. Αν αναλογιστεί κανείς το µέγεθος και την πολυ-πλοκότητα των µακροµορίων, είναι πράγµατι αξιοθαύµα-
στο το ότι οι βιοχηµικοί έχουν καταφέρει να προσδιορί-σουν µε κάθε λεπτοµέρεια τη δοµή τόσων πολλών πρω-τεϊνών (Εικόνα 5.1).
Η αρχιτεκτονική ενός µεγάλου µορίου µάς βοηθά ναεξηγήσουµε τον τρόπο λειτουργίας του. Όπως συµβαίνειµε το νερό και µε τα άλλα απλά οργανικά µόρια, έτσι καιτα µεγάλα βιολογικά µόρια εµφανίζουν µοναδικές ιδιότη-τες οι οποίες προκύπτουν από τη διευθέτηση των ατόµωντους. Σε αυτό το κεφάλαιο θα εξετάσουµε αρχικά πώς εί-ναι κατασκευασµένες οι τέσσερεις κατηγορίες µεγάλωνβιολογικών µορίων και κατόπιν θα µελετήσουµε τη δοµήκαι τη λειτουργία αυτών των µορίων, δηλαδή των υδα-τανθράκων, των λιπιδίων, των πρωτεϊνών και των νου-κλεϊκών οξέων.
Τ Μ Η Μ Α 5.1
Τα µακροµόρια είναι πολυµερή πουσυντίθενται από µονοµερή
Τα µακροµόρια των τριών από τις τέσσερεις κατηγορίεςοργανικών ενώσεων που συναντάµε στους οργανισµούς –των υδατανθράκων, των πρωτεϊνών και των νουκλεϊκώνοξέων– είναι στην ουσία µόρια-αλυσίδες που ονοµάζονταιπολυµερή. Πολυµερές είναι ένα επίµηκες µόριο αποτε-λούµενο από πολλούς παραπλήσιους ή πανοµοιότυπουςδοµικούς λίθους οι οποίοι συνδέονται µεταξύ τους µε οµοι-οπολικούς δεσµούς, σε µεγάλο βαθµό όπως ένα τρένο απο-τελείται από µια αλυσίδα βαγονιών. Οι επαναλαµβανόµε-νες µονάδες που συνιστούν τους δοµικούς λίθους των πο-λυµερών είναι µικρότερα µόρια τα οποία ονοµάζονται µο-νοµερή. Ορισµένα από τα µόρια που χρησιµεύουν ως µο-νοµερή µπορεί, πέραν του ότι αποτελούν δοµικούς λίθουςτων πολυµερών, να επιτελούν και άλλες λειτουργίες.
Β Α Σ Ι Κ Ε Σ Ε Ν Ν Ο Ι Ε Σ
5.1 Τα µακροµόρια είναι πολυµερή που συντίθενταιαπό µονοµερή
5.2 Οι υδατάνθρακες χρησιµεύουν ως καύσιµα καιδοµικά υλικά
5.3 Τα λιπίδια είναι µια ετερογενής οµάδαυδρόφοβων µορίων
5.4 Οι πρωτεΐνες έχουν διάφορες δοµές, οπότεεµφανίζουν µεγάλο εύρος δράσεων
5.5 Τα νουκλεϊκά οξέα αποθηκεύουν καιµεταβιβάζουν κληρονοµικές πληροφορίες
Ε Π Ι Σ Κ Ο Π Η Σ Η
Tα µόρια της ζωής
Εικόνα 5.1 Γιατί μελετούν οι επιστήμονες τη δομή τωνμακρομορίων;
5

84 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
δύο αδέλφια δεν είναι τίποτε άλλο παρά διαφορές στα πο-λυµερή τους, ιδιαίτερα στο DNA και στις πρωτεΐνες τους.Οι µοριακές διαφορές που παρατηρούνται ανάµεσα σε δύοµη συγγενικά άτοµα είναι µεγαλύτερες, ενώ ανάµεσα σεάτοµα διαφορετικών ειδών είναι ακόµη πιο εκτεταµένες.Με λίγα λόγια, η ποικιλότητα των µακροµορίων στον έµ-βιο κόσµο είναι τεράστια και οι πιθανές παραλλαγές τουςπρακτικά άπειρες.
Ωστόσο, ποια είναι η βάση αυτής της ποικιλότητας τωνπολυµερών που απαντούν στους ζωντανούς οργανισµούς;Για την κατασκευή των συγκεκριµένων µορίων απαιτούν-ται µόλις 40-50 συνήθη µονοµερή και ένας µικρός αριθµόςαπό ορισµένα άλλα, σχετικά σπάνια µονοµερή. Η κατα-σκευή µιας τόσο µεγάλης ποικιλίας πολυµερών από τόσοπεριορισµένο αριθµό µονοµερών είναι κάτι ανάλογο µετην κατασκευή εκατοντάδων χιλιάδων λέξεων από τα 24µόλις γράµµατα του ελληνικού αλφαβήτου. Το κλειδί είναιη διάταξη, δηλαδή η συγκεκριµένη γραµµική σειρά µε τηνοποία διατάσσονται τα γράµµατα. Ωστόσο, ακόµη κι αυ-τή η αναλογία κάθε άλλο παρά περιγράφει επαρκώς τη µε-γάλη ποικιλία των µακροµορίων που συναντάµε στον έµ-βιο κόσµο, διότι τα περισσότερα βιολογικά πολυµερήέχουν πολύ µεγαλύτερο µήκος κι από την πιο µεγάλη λέ-ξη. Οι πρωτεΐνες, παραδείγµατος χάριν, σχηµατίζονται από20 είδη αµινοξέων που διατάσσονται συνήθως σε αλυσίδεςµήκους µερικών εκατοντάδων αµινοξέων. Η µοριακή λο-γική της ζωής είναι απλή και κοµψή. Τα µικρά µόρια που
Σύνθεση και διάσπαση των πολυμερών
Οι ποικίλες κατηγορίες των πολυµερών διαφέρουν ωςπρος τη φύση των µονοµερών τους. Ωστόσο, οι χηµικοίµηχανισµοί µε τους οποίους συντίθενται και διασπώνταιτα διάφορα πολυµερή µέσα στα κύτταρα είναι, κατά κα-νόνα, οι ίδιοι σε όλες τις περιπτώσεις (Εικόνα 5.2). Τα µο-νοµερή συνδέονται µεταξύ τους µέσω µιας αντίδρασηςστην οποία τα δύο µόρια συνδέονται οµοιοπολικά το έναµε το άλλο, µε την ταυτόχρονη απώλεια ενός µορίου νε-ρού. Αυτή η αντίδραση είναι ευρύτερα γνωστή ως αντί-δραση συµπύκνωσης, και πιο συγκεκριµένα ως αντί-δραση αφυδάτωσης, επειδή τα µονοµερή χάνουν κατάτην ένωσή τους ένα µόριο νερού (Εικόνα 5.2α). Κατά τονσχηµατισµό του δεσµού ανάµεσα σε δύο µονοµερή, κάθεµονοµερές προσφέρει ένα τµήµα από το µόριο νερού πουχάνεται. Το ένα µονοµερές προµηθεύει µια υδροξυλοµά-δα (—ΟΗ) και το άλλο µονοµερές προµηθεύει ένα υδρο-γόνο (—Η). Η αντίδραση επαναλαµβάνεται κάθε φοράπου προστίθεται το ένα µονοµερές µετά το άλλο στη µα-κρά αλυσίδα που θα αποτελέσει τελικά το πολυµερές. Ηαντίδραση αφυδάτωσης διευκολύνεται από ειδικά ένζυ-µα, δηλαδή από εξειδικευµένα µακροµόρια των κυττάρωνπου επιταχύνουν τις χηµικές αντιδράσεις.
Τα πολυµερή αποσυναρµολογούνται ξανά σε µονοµε-ρή µέσω της υδρόλυσης, δηλαδή µε µια διαδικασία πουουσιαστικά αποτελεί το αντίστροφο της αντίδρασης αφυ-δάτωσης (Εικόνα 5.2β). Υδρόλυση, όπως υποδηλώνει καιη ίδια η λέξη, σηµαίνει «διάσπαση µε τη χρήση του νε-ρού». Ο δεσµός ανάµεσα σε δύο µονοµερή διασπάται µετην προσθήκη ενός µορίου νερού, κατά την οποία έναυδρογόνο από το µόριο του νερού προστίθεται στο ένα µο-νοµερές και η υδροξυλοµάδα προστίθεται στο άλλο.Υδρόλυση συµβαίνει, παραδείγµατος χάριν, µέσα στο σώ-µα µας κατά τη διαδικασία της πέψης. Το µεγαλύτερο τµή-µα των οργανικών υλικών που εισέρχονται στο σώµα µαςέχει τη µορφή πολυµερών, πράγµα που σηµαίνει ότι ταυλικά αυτά είναι εξαιρετικά µεγάλα και ογκώδη για να ει-σέλθουν στα κύτταρά µας. Για τον λόγο αυτό, στο εσωτε-ρικό του πεπτικού σωλήνα υπάρχουν διάφορα ένζυµα που«επιτίθενται» στα πολυµερή, επιταχύνοντας την υδρόλυ-σή τους. Τα µονοµερή που προκύπτουν από τη συγκεκρι-µένη διεργασία µπορούν κατόπιν να απορροφηθούν καινα διανεµηθούν, µέσω της κυκλοφορίας, σε όλα τα κύτ-ταρα. Στη συνέχεια, τα κύτταρα µπορούν να χρησιµοποι-ήσουν τα εν λόγω µονοµερή για να συνθέσουν νέα πολυ-µερή, ικανά να επιτελέσουν τις ειδικές λειτουργίες πουχρειάζεται κάθε κύτταρο.
Ποικιλομορφία των πολυμερών
Κάθε κύτταρο διαθέτει χιλιάδες διαφορετικά είδη µακρο-µορίων. Η «συλλογή» αυτή ποικίλλει ανάλογα µε τον τύ-πο κάθε κυττάρου, ακόµη και µέσα στον ίδιο οργανισµό.Οι κληρονοµικές διαφορές που εµφανίζονται ανάµεσα σε
H
HO H
Βραχύ πολυμερές
Με την αντίδραση αφυδάτωσηςαπομακρύνεται ένα μόριο νερούκαι σχηματίζεται ένας νέος δεσμός
Επίμηκες πολυμερές
Μη συνδεδεμένο μονομερές
Με την υδρόλυσηπροστίθεται ένα μόριο νερούκαι διασπάται ένας δεσμός
HO
H2O
H2O
1 2 3 4
HO H1 2 3 4
HO HHOH1 2 3
(α) Αντίδραση αφυδάτωσης κατά τη σύνθεση πολυμερούς
(β) Υδρόλυση πολυμερούς
HO H1 2 3
Εικόνα 5.2 Σύνθεση και διάσπαση των πολυμερών.
συνθέτουν τα µακροµόρια είναι κοινά για όλους τους ορ-γανισµούς, αλλά σε κάθε οργανισµό τα µόρια αυτά δια-τάσσονται µε µοναδικό τρόπο.
Παρά την τεράστια αυτή ποικιλότητα, η µοριακή δοµήκαι λειτουργία µπορεί και πάλι να οµαδοποιηθεί χονδρικάσε κατηγορίες. Ας δούµε, φέρ’ ειπείν, τις τέσσερεις µεγά-λες κατηγορίες βιολογικών µορίων. Σε κάθε κατηγορία,τα µεγάλα µόρια που σχηµατίζονται έχουν πρόσθετες ιδιό-τητες οι οποίες δεν υπάρχουν στους µεµονωµένους δοµι-κούς λίθους.
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 85
Τ Μ Η Μ Α 5.2
Οι υδατάνθρακες χρησιµεύουν ωςκαύσιµα και δοµικά υλικά
Οι υδατάνθρακες περιλαµβάνουν τα σάκχαρα και τα πο-λυµερή σακχάρων. Απλούστεροι υδατάνθρακες είναι οιµονοσακχαρίτες, γνωστοι και ως απλά σάκχαρα. Οι δι-σακχαρίτες είναι διπλά σάκχαρα, δηλαδή µόρια αποτε-λούµενα από δύο µονοσακχαρίτες ενωµένους µε µια αντί-δραση αφυδάτωσης. Στους υδατάνθρακες ανήκει και µιαάλλη κατηγορία µακροµορίων, οι πολυσακχαρίτες, δηλαδήπολυµερή αποτελούµενα από πολλά µονοµερή σακχάρων.
Σάκχαρα
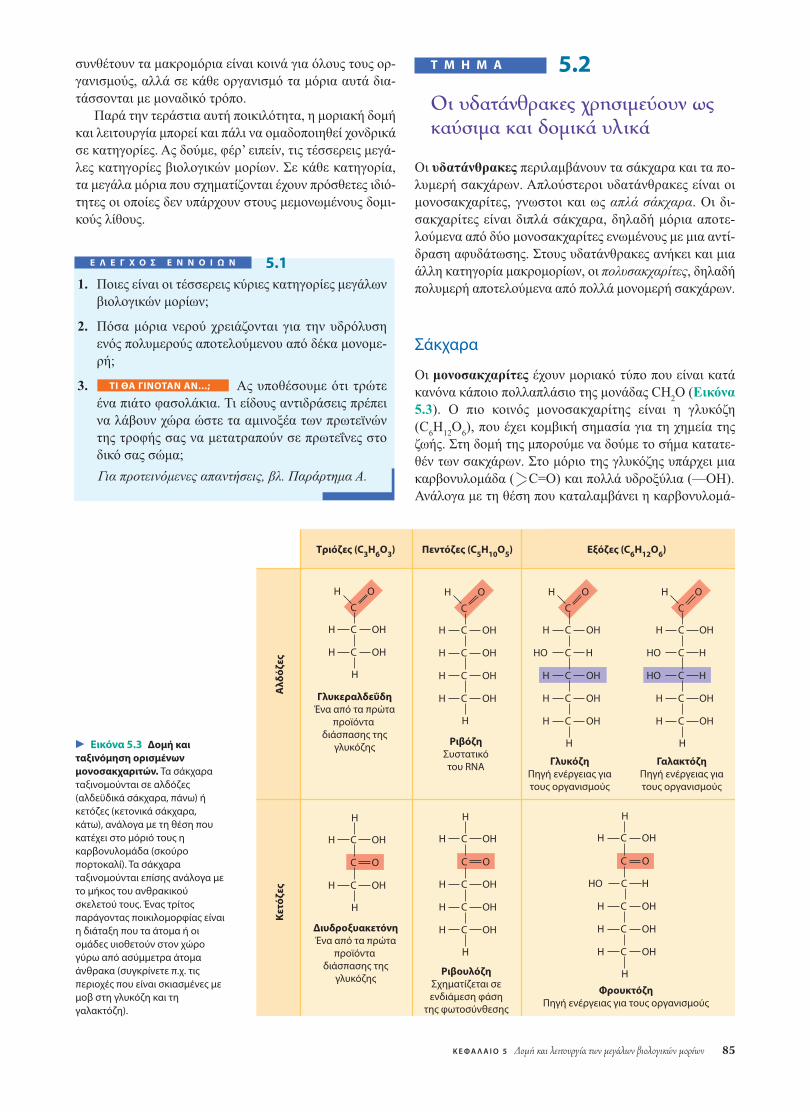
Οι µονοσακχαρίτες έχουν µοριακό τύπο που είναι κατάκανόνα κάποιο πολλαπλάσιο της µονάδας CH2O (Εικόνα5.3). Ο πιο κοινός µονοσακχαρίτης είναι η γλυκόζη(C6H12O6), που έχει κοµβική σηµασία για τη χηµεία τηςζωής. Στη δοµή της µπορούµε να δούµε το σήµα κατατε-θέν των σακχάρων. Στο µόριο της γλυκόζης υπάρχει µιακαρβονυλοµάδα ( C=O) και πολλά υδροξύλια (—OH).Ανάλογα µε τη θέση που καταλαµβάνει η καρβονυλοµά-
Ε Λ Ε Γ Χ Ο Σ Ε Ν Ν Ο Ι Ω Ν 5.11. Ποιες είναι οι τέσσερεις κύριες κατηγορίες µεγάλων
βιολογικών µορίων;
2. Πόσα µόρια νερού χρειάζονται για την υδρόλυσηενός πολυµερούς αποτελούµενου από δέκα µονοµε-ρή;
3. Ας υποθέσουµε ότι τρώτεένα πιάτο φασολάκια. Τι είδους αντιδράσεις πρέπεινα λάβουν χώρα ώστε τα αµινοξέα των πρωτεϊνώντης τροφής σας να µετατραπούν σε πρωτεΐνες στοδικό σας σώµα;Για προτεινόµενες απαντήσεις, βλ. Παράρτηµα Α.
ΤΙ ΘΑ ΓΙΝΟΤΑΝ ΑΝ...;
CH OH
C
OH
H
CH OH
CH OH
H
C O
H
CH OH
CH OH
C
OH
H
CH OH
CH OH
CH OH
CH OH
H
CH OH
CH OH
CH OH
H
C O
CH OH
C
OH
H
CH OH
CH OH
CH OH
H
CH OH
CH OH
CH OH
H
C O
CH OH
C
OH
H
CHO H
CH OH
CH OH
CHO H
CHO H
CHO H
Τριόζες (C3H6O3) Πεντόζες (C5H10O5)
ΓλυκεραλδεΰδηΈνα από τα πρώτα
προϊόνταδιάσπασης της
γλυκόζης
Αλδ
όζες
Κετό
ζες
ΔιυδροξυακετόνηΈνα από τα πρώτα
προϊόνταδιάσπασης της
γλυκόζηςΡιβουλόζη
Σχηματίζεται σεενδιάμεση φάση
της φωτοσύνθεσης
ΡιβόζηΣυστατικότου RNA Γλυκόζη
Πηγή ενέργειας γιατους οργανισμούς
CH OH
ΓαλακτόζηΠηγή ενέργειας γιατους οργανισμούς
ΦρουκτόζηΠηγή ενέργειας για τους οργανισμούς
Εξόζες (C6H12O6)
Εικόνα 5.3 Δομή καιταξινόμηση ορισμένωνμονοσακχαριτών. Τα σάκχαραταξινομούνται σε αλδόζες(αλδεϋδικά σάκχαρα, πάνω) ήκετόζες (κετονικά σάκχαρα,κάτω), ανάλογα με τη θέση πουκατέχει στο μόριό τους ηκαρβονυλομάδα (σκούροπορτοκαλί). Τα σάκχαραταξινομούνται επίσης ανάλογα μετο μήκος του ανθρακικούσκελετού τους. Ένας τρίτοςπαράγοντας ποικιλομορφίας είναιη διάταξη που τα άτομα ή οιομάδες υιοθετούν στον χώρογύρω από ασύμμετρα άτομαάνθρακα (συγκρίνετε π.χ. τιςπεριοχές που είναι σκιασμένες μεμοβ στη γλυκόζη και τηγαλακτόζη).

δα, ένα σάκχαρο µπορεί να είναι είτε αλδόζη (αλδεϋδικόσάκχαρο) είτε κετόζη (κετονικό σάκχαρο). Η γλυκόζη, λό-γου χάρη, είναι µια αλδόζη. Η φρουκτόζη, δοµικό ισοµε-ρές της γλυκόζης, είναι µια κετόζη. (Τα ονόµατα των πε-ρισσότερων σακχάρων καταλήγουν σε -όζη.) Ένα άλλοκριτήριο για την ταξινόµηση των σακχάρων είναι το µέ-γεθος του ανθρακικού σκελετού, το οποίο µπορεί να κυ-µαίνεται από τρία έως επτά άτοµα άνθρακα. Η γλυκόζη, ηφρουκτόζη και όσα άλλα σάκχαρα περιέχουν έξι άτοµαάνθρακα ονοµάζονται εξόζες. Πολύ κοινά µόρια είναι επί-σης οι τριόζες (σάκχαρα µε τρία άτοµα άνθρακα) και οιπεντόζες (σάκχαρα µε πέντε άτοµα άνθρακα).
Μία επιπλέον πηγή ποικιλότητας των απλών σακχά-ρων είναι η διευθέτηση των τµηµάτων τους γύρω απόασύµµετρους άνθρακες. (Θυµίζουµε ότι ασύµµετρος είναιένας άνθρακας που συνδέεται µε τέσσερα διαφορετικάάτοµα ή τέσσερεις διαφορετικές οµάδες ατόµων.) Η γλυ-κόζη και η γαλακτόζη, λόγου χάρη, διαφέρουν µόνο ως
86 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
προς τη διάταξη των τµηµάτων τους γύρω από έναν ασύµ-µετρο άνθρακα (βλ. τις περιοχές της Εικόνας 5.3 που είναισκιασµένες µοβ). Από αυτή την ασήµαντη –εκ πρώτηςόψεως– διαφορά προκύπτουν δύο σάκχαρα µε διαφορετι-κή δοµή και χηµική συµπεριφορά.
Η σχεδίαση του ανθρακικού σκελετού της γλυκόζης µεγραµµικό τρόπο θα ήταν µεν βολική, αλλά όχι εντελώςακριβής. Σε υδατικό διάλυµα, το µόριο της γλυκόζης, όπωςκαι των περισσότερων άλλων σακχάρων, σχηµατίζει δα-κτύλιο (Εικόνα 5.4).
Οι µονοσακχαρίτες, και ειδικά η γλυκόζη, αποτελούνένα από τα κυριότερα θρεπτικά συστατικά των κυττάρων.Τα κύτταρα προσλαµβάνουν χηµική ενέργεια µέσω µιαςσειράς αντιδράσεων (ονοµάζεται κυτταρική αναπνοή) πουξεκινούν µε το µόριο της γλυκόζης. Όµως εκτός από ση-µαντικά καύσιµα µόρια για την ενεργειακή τροφοδότησητων διαφόρων κυτταρικών λειτουργιών, τα απλά σάκχα-ρα χρησιµεύουν και ως πρώτη ύλη για να συντεθούν άλλοι
O
Γλυκόζη Γλυκόζη Μαλτόζη
1
H OOHOH
4
OH
γλυκοζιτικόςδεσμός
1– 4
(α)
H2OH OH
HOH
CH2OH
OH H
H
HO
H OH
HOH
CH2OH
OH H
H
HO
H OH
HOH
CH2OH
OH H
H
H OH
HOH
CH2OH
OH H
H
O
Σακχαρόζη
1 2
γλυκοζιτικόςδεσμός
1– 2
H OH
HOH
CH2OH
OH H
H
HOCH2OH
H
Γλυκόζη Φρουκτόζη
H OOH
H2OH OH
HOH
CH2OH
OH H
H
HO
OH H
OCH2OH
H HOCH2OH
H
OH H
OCH2OH
H HO
Αντίδραση αφυδάτωσης κατά τη σύνθεση της μαλτόζης. Ησύνδεση δύο μονάδων (μονομερών) γλυκόζης σχηματίζει τη μαλτόζη. Ο γλυκοζιτικός δεσμός ενώνει τον άνθρακα 1 της μίας γλυκόζης με τον άνθρακα 4 της άλλης. Αν τα δύο μονομερή ενωθούν διαφορετικά, θα σχηματιστεί άλλος δισακχαρίτης.
(β) Αντίδραση αφυδάτωσης κατά τη σύνθεση της σακχαρόζης. Η σακχαρόζη είναι ένας δισακχαρίτης που σχηματίζεται από γλυκόζη και φρουκτόζη. Παρατηρήστε ότι η φρουκτόζη (αν είναι εξόζη) σχηματίζει δακτύλιο με πέντε έδρες.
Εικόνα 5.5 Παραδείγματα σύνθεσης δισακχαριτών.
H OH
C
OH
H
H OH
H OH
HO H
H OH
H
(α)
(β)
2C
3C
4C
4C
5C
6C
1
H OH
OH H
3 C
C
OH
H
4 C
H
O H
1H
H
O
OH
OH H2C
2C3 COH
H
1H
5C 5C
6 CH2OH 6 CH2OH
C
H
O
Γραμμική μορφή και δακτύλιος. Η χημική ισορροπία ανάμεσα στη γραμμική μορφή της γλυκόζηςκαι σε εκείνη του δακτυλίου ευνοεί σημαντικά τον σχηματισμό δακτυλίων. Οι άνθρακες τωνσακχάρων αριθμούνται από 1 μέχρι 6 με τον τρόπο που βλέπουμε. Για να σχηματιστεί ο δακτύλιοςτης γλυκόζης, ο άνθρακας 1 συνδέεται με το οξυγόνο που είναι συνδεδεμένο στον άνθρακα 5.
Συντετμημένη μορφή του δακτυλίου. Κάθε γωνία αναπαριστά ένα άτομο άνθρακα. Η παχύτερη πλευράτου δακτυλίου σημαίνει ότι ο δακτύλιος αυτός προεξέχει προς εμάς, κάθετα προς το επίπεδο της σελίδας. Τα διάφορα συστατικά που συνδέονται με τις γωνίες (δηλ. τους άνθρακες) του δακτυλίου, προεξέχουν προς τα πάνω ή προς τα κάτω από το επίπεδο που σχηματίζει ο δακτύλιος.
H OH
HOH
CH2OH
OH H
H
HO OH3 2
1
5
6
4
O
Εικόνα 5.4 Η γλυκόζη σε γραμμική μορφή και σε μορφή δακτυλίου.
Ξεκινήστε με τη γραμμική μορφή της φρουκτόζης (βλ. Εικόνα 5.3) και σχεδιάστε τα δύο βήματα που χρειάζονται για να σχηματιστεί οδακτύλιος της γλυκόζης. Αριθμήστε τους άνθρακες. Συνδέστε τον άνθρακα 5, μέσω ενός ατόμου οξυγόνου, με τον άνθρακα 2. Συγκρίνετε τον αριθμό τωνατόμων άνθρακα στους δακτυλίους της φρουκτόζης και της γλυκόζης.
ΣΧΕΔΙΑΣΤΕ!

τύποι µικρών οργανικών µορίων, όπως αµινοξέα και λι-παρά οξέα. Όσα µόρια γλυκόζης δεν χρησιµοποιούνταιάµεσα σε κάποια από τις παραπάνω διεργασίες χρησι-µεύουν ως µονοµερή και ενσωµατώνονται σε δισακχαρί-τες ή πολυσακχαρίτες.
Ένας δισακχαρίτης αποτελείται από δύο µονοσακχα-ρίτες που ενώνονται µε γλυκοζιτικό δεσµό, το είδος τουοµοιοπολικού δεσµού που σχηµατίζεται ανάµεσα σε δύοµονοσακχαρίτες µε ανυδριτική αντίδραση. Λόγου χάρη, ηµαλτόζη είναι ένας δισακχαρίτης που σχηµατίζεται απότην ένωση δύο µορίων γλυκόζης (Εικόνα 5.5α). Η µαλτό-ζη είναι συστατικό ζύµωσης της µπίρας. Ωστόσο, ο πιοκοινός δισακχαρίτης είναι η σακχαρόζη (ή σουκρόζη), δη-λαδή η κοινή ζάχαρη. Τα δύο µονοµερή της σακχαρόζηςείναι η γλυκόζη και η φρουκτόζη (Εικόνα 5.5β). Πρόκει-ται για την κύρια µορφή υδατάνθρακα των φυτών, ο οποί-ος µεταφέρεται από τα φύλλα στις ρίζες και στα άλλα µηφωτοσυνθετικά όργανα. Ένας άλλος κοινός δισακχαρίτηςείναι η λακτόζη του γάλακτος, που αποτελείται από έναµόριο γλυκόζης και ένα µόριο γαλακτόζης.
Πολυσακχαρίτες
Οι πολυσακχαρίτες είναι µεγάλα πολυµερή µόρια (µα-κροµόρια) αποτελούµενα από λίγες εκατοντάδες µέχρι λί-γες χιλιάδες µονοσακχαρίτες, οι οποίοι ενώνονται µεταξύτους µε γλυκοζιτικούς δεσµούς. Μερικοί πολυσακχαρίτεςχρησιµεύουν στα κύτταρα ως αποθήκη θρεπτικών υλικών,τα οποία υδρολύουν κάθε φορά που χρειάζονται σάκχα-ρα. Άλλοι πολυσακχαρίτες χρησιµοποιούνται ως δοµικάυλικά για διάφορες δοµές που προστατεύουν το κύτταροκαι τον οργανισµό. Η δοµή και η λειτουργία ενός πολυ-σακχαρίτη καθορίζεται από τα µονοµερή σάκχαρα από ταοποία αποτελείται, καθώς και από την ακριβή θέση τωνγλυκοζιτικών δεσµών.
Αποθηκευτικοί πολυσακχαρίτες
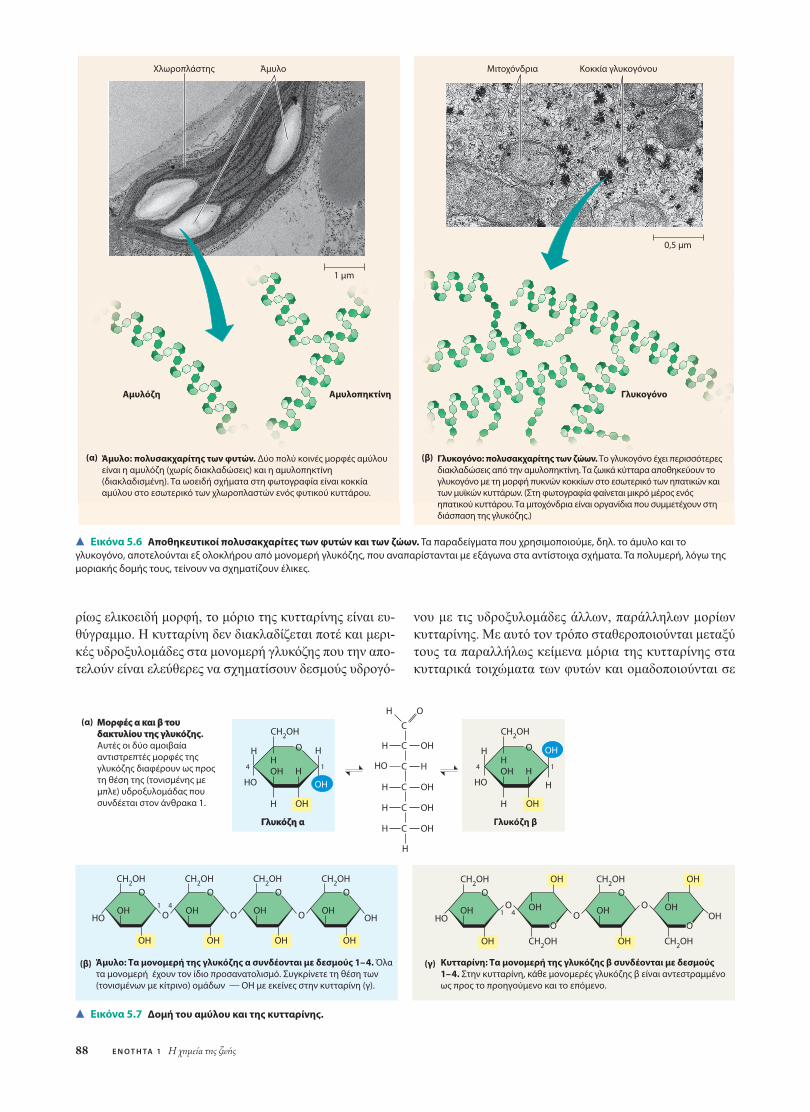
Τόσο τα φυτά όσο και τα ζώα αποθηκεύουν σάκχαρα γιαµεταγενέστερη χρήση, µε τη µορφή των αποθηκευτικώνπολυσακχαριτών. Τα φυτά αποθηκεύουν κοκκία από άµυ-λο (πολυµερές από µονοµερή γλυκόζης) µέσα σε ιδιαίτε-ρα κυτταρικά οργανίδια που ονοµάζονται πλαστίδια (σεαυτά ανήκουν και οι χλωροπλάστες). Η δυνατότητα σύν-θεσης αµύλου επιτρέπει στα φυτά να αποθηκεύουν την πε-ρίσσεια γλυκόζης. Και επειδή η γλυκόζη είναι ένα από τασηµαντικότερα καύσιµα των κυττάρων, το άµυλο λει-τουργεί στην πραγµατικότητα ως αποθηκευµένη ενέργεια.Μονοµερή σάκχαρα µπορούν κατόπιν να ληφθούν από την«τράπεζα» υδατανθράκων µε υδρόλυση, που διασπά τονδεσµό ανάµεσα στα µονοµερή γλυκόζης. Τα περισσότεραζώα, όπως και ο άνθρωπος, διαθέτουν ένζυµα για να υδρο-λύουν το φυτικό άµυλο και, έτσι, να χρησιµοποιούν τηναποθηκευµένη γλυκόζη των φυτών ως θρεπτικό υλικό γιατα δικά τους κύτταρα. Οι βολβοί της πατάτας και οι σπό-
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 87
ροι των δηµητριακών –δηλαδή οι καρποί του σιταριού,του καλαµποκιού, του ρυζιού και άλλων φυτών– είναι οικύριες πηγές αµύλου στη διατροφή του ανθρώπου.
Τα περισσότερα µονοµερή γλυκόζης του αµύλου συν-δέονται µεταξύ τους µε δεσµούς 1-4 (δηλαδή µε δεσµόανάµεσα στον άνθρακα 1 της µίας γλυκόζης και στον άν-θρακα 4 της άλλης). Με τους ίδιους δεσµούς συνδέονταικαι οι δύο γλυκόζες στον δισακχαρίτη µαλτόζη (βλ. Εικό-να 5.5α). Λόγω της γωνίας του συγκεκριµένου δεσµού, τοπολυµερές αποκτά ελικοειδή µορφή. Απλούστερη µορφήαµύλου είναι η αµυλόζη, πολυµερές χωρίς διακλαδώσεις(Εικόνα 5.6α). Η αµυλοπηκτίνη, µια πιο περίπλοκη µορ-φή αµύλου, είναι ένα διακλαδισµένο πολυµερές που στασηµεία διακλάδωσής του σχηµατίζονται δεσµοί 1-6.
Τα ζώα αποθηκεύουν γλυκόζη σε έναν πολυσακχαρίτηπου ονοµάζεται γλυκογόνο και µοιάζει µε την αµυλοπη-κτίνη αλλά διαθέτει ακόµη πιο εκτεταµένες διακλαδώσεις(Εικόνα 5.6β). Ο άνθρωπος και τα άλλα θηλαστικά απο-θηκεύουν το γλυκογόνο κυρίως στα κύτταρα των µυών καιτου ήπατος. Όταν οι απαιτήσεις σε γλυκόζη αυξηθούν, τό-τε τα κύτταρα των µυών και του ήπατος απελευθερώνουνγλυκόζη υδρολύοντας το γλυκογόνο. Αυτή η αποθήκηκαύσιµης ύλης, ωστόσο, δεν επαρκεί και δεν µπορεί νασυντηρήσει ένα ζώο για µεγάλο χρονικό διάστηµα. Στονάνθρωπο, π.χ., τα αποθέµατα γλυκογόνου εξαντλούνταιµέσα σε µία ηµέρα περίπου, και για να αναπληρωθούνπρέπει οπωσδήποτε να καταναλωθεί τροφή. Όσοι ακο-λουθούν δίαιτα µε λίγους υδατάνθρακες πρέπει να λαµ-βάνουν σοβαρά υπ’ όψιν τους αυτό το ζήτηµα.
Δομικοί πολυσακχαρίτες
Με τους πολυσακχαρίτες οι οργανισµοί χτίζουν πολύ ισχυ-ρά υλικά. Λόγου χάρη, ο πολυσακχαρίτης κυτταρίνη είναιτο κύριο συστατικό των σκληρών τοιχωµάτων που περι-βάλλουν τα φυτικά κύτταρα. Σε πλανητική κλίµακα, τα φυ-τά παράγουν σχεδόν 1014 kg (περίπου 100 δισεκατοµµύριατόνους) κυτταρίνης τον χρόνο. Είναι η πιο κοινή οργανικήουσία στη Γη. Η κυτταρίνη, όπως και το άµυλο, είναι έναπολυµερές της γλυκόζης, αλλά διαφέρει από το άµυλοστους γλυκοζιτικούς δεσµούς. Στην πραγµατικότητα, η δια-φορά τους οφείλεται στο ότι υπάρχουν δύο µορφές δακτυ-λίων της γλυκόζης (Εικόνα 5.7α). Όταν η γλυκόζη σχηµα-τίζει δακτύλιο, η υδροξυλοµάδα που συνδέεται στον άν-θρακα 1 βρίσκεται είτε κάτω είτε πάνω από το επίπεδο τουδακτυλίου. Αυτές οι δύο διαµορφώσεις του δακτυλίου τηςγλυκόζης ονοµάζονται, αντίστοιχα, διαµόρφωση άλφα (α)και διαµόρφωση βήτα (β). Στο άµυλο (Εικόνες 5.4 και 5.5),όλα τα µονοµερή γλυκόζης έχουν τη διαµόρφωση α (Ει-κόνα 5.7β). Αντίθετα, όλα τα µονοµερή γλυκόζης στην κυτ-ταρίνη έχουν τη διαµόρφωση β, µε αποτέλεσµα κάθε δεύ-τερο µονοµερές γλυκόζης να είναι αναποδογυρισµένο ωςπρος τους άµεσους γείτονές του (Εικόνα 5.7γ).
Οι διαφορετικοί γλυκοζιτικοί δεσµοί στο άµυλο καιστην κυτταρίνη προσδίδουν στα δύο µόρια αρκετά διαφο-ρετική δοµή στον χώρο. Ενώ το µόριο του αµύλου έχει κυ-
ρίως ελικοειδή µορφή, το µόριο της κυτταρίνης είναι ευ-θύγραµµο. Η κυτταρίνη δεν διακλαδίζεται ποτέ και µερι-κές υδροξυλοµάδες στα µονοµερή γλυκόζης που την απο-τελούν είναι ελεύθερες να σχηµατίσουν δεσµούς υδρογό-
88 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
CH OH
C
OH
H
CH OH
CH OH
CHO H
CH OHΓλυκόζη βΓλυκΓλυκόζη αόζη α
(α)
OH
1
H
4
H
OH
CH2OH
OH H
H
HOOH
1
H
4
H
OH
CH2OH
OH H
H
HO
O
(β)
1
OCH2OH CH2OH CH2OH CH2OH
HO OH4
O
O
O
O
O
OH OH OH OH
OH OH
OH
CH2OH
O1
O
HO4 O
OCH2OH
CH2OHCH2OH
OH OH
OHOH
OHO
O O
OH OH OH OHOH OH OH
ΜορφΜορφές α και β τουδακτυλίου της γλυκόζηςδακτυλίου της γλυκόζης.Αυτές οι δύο αμοιβαία αντιστρεπτές μορφές της γλυκόζης διαφέρουν ως προς τη θέση της (τονισμένης μεμπλε) υδροξυλομάδας που συνδέεται στον άνθρακα 1.
Άμυλο: Τα μονομερή της γλυκόζης α συνδέονται με δεσμούς 1–4. Όλα τα μονομερή έχουν τον ίδιο προσανατολισμό. Συγκρίνετε τη θέση των (τονισμένων με κίτρινο) ομάδων ΟΗ με εκείνες στην κυτταρίνη (γ).
(γ) Κυτταρίνη: Τα μονομερή της γλυκόζης β συνδέονται με δεσμούς 1–4. Στην κυτταρίνη, κάθε μονομερές γλυκόζης β είναι αντεστραμμένο ως προς το προηγούμενο και το επόμενο.
Εικόνα 5.7 Δομή του αμύλου και της κυτταρίνης.
νου µε τις υδροξυλοµάδες άλλων, παράλληλων µορίωνκυτταρίνης. Με αυτό τον τρόπο σταθεροποιούνται µεταξύτους τα παραλλήλως κείµενα µόρια της κυτταρίνης στακυτταρικά τοιχώµατα των φυτών και οµαδοποιούνται σε
Κοκκία γλυκογόνουΜιτοχόνδριαΧλωροπλάστης
Αμυλόζη Αμυλοπηκτίνη Γλυκογόνο
Άμυλο
1 μm
0,5 μm
(α) Άμυλο: πολυσακχαρίτης των φυτών. Δύο πολύ κοινές μορφές αμύλου είναι η αμυλόζη (χωρίς διακλαδώσεις) και η αμυλοπηκτίνη (διακλαδισμένη). Τα ωοειδή σχήματα στη φωτογραφία είναι κοκκία αμύλου στο εσωτερικό των χλωροπλαστών ενός φυτικού κυττάρου.
(β) Γλυκογόνο: πολυσακχαρίτης των ζώων. Το γλυκογόνο έχει περισσότερες διακλαδώσεις από την αμυλοπηκτίνη. Τα ζωικά κύτταρα αποθηκεύουν το γλυκογόνο με τη μορφή πυκνών κοκκίων στο εσωτερικό των ηπατικών και των μυϊκών κυττάρων. (Στη φωτογραφία φαίνεται μικρό μέρος ενός ηπατικού κυττάρου. Τα μιτοχόνδρια είναι οργανίδια που συμμετέχουν στη διάσπαση της γλυκόζης.)
Εικόνα 5.6 Αποθηκευτικοί πολυσακχαρίτες των φυτών και των ζώων. Τα παραδείγματα που χρησιμοποιούμε, δηλ. το άμυλο και τογλυκογόνο, αποτελούνται εξ ολοκλήρου από μονομερή γλυκόζης, που αναπαρίστανται με εξάγωνα στα αντίστοιχα σχήματα. Τα πολυμερή, λόγω τηςμοριακής δομής τους, τείνουν να σχηματίζουν έλικες.

µονάδες που µοιάζουν µε καλώδια και ονοµάζονται µι-κροϊνίδια (Εικόνα 5.8). Τα µικροϊνίδια αποτελούν ισχυρόδοµικό υλικό των φυτών, αλλά έχουν πολύ σηµαντική αξίακαι για τον άνθρωπο, διότι η κυτταρίνη είναι το κύριο συ-στατικό του χαρτιού και, επίσης, το µοναδικό συστατικότου βαµβακιού.
Τα ένζυµα που πέπτουν το άµυλο υδρολύοντας τουςγλυκοζιτικούς δεσµούς α δεν µπορούν να υδρολύσουντους γλυκοζιτικούς δεσµούς β της κυτταρίνης, λόγω τουδιαφορετικού σχήµατος που έχουν στον χώρο τα δύο µό-ρια. Στην πραγµατικότητα, ελάχιστοι οργανισµοί διαθέ-τουν ένζυµα που µπορούν να διασπάσουν την κυτταρίνη.Ο άνθρωπος πάντως δεν µπορεί, µε αποτέλεσµα η κυττα-ρίνη των τροφών µας να παραµένει άπεπτη καθ’ όλο τοµήκος του πεπτικού σωλήνα και τελικά να αποµακρύνε-ται µε τα κόπρανα. Επειδή η κυτταρίνη, κατά τη διαδρο-µή της στην πεπτική οδό, προκαλεί µικροφθορές στα τοι-χώµατα των εντέρων, διεγείρει την έκκριση βλέννας απότα κύτταρα των τοιχωµάτων κι έτσι διευκολύνει τη διέ-λευση των τροφών µέσω του συστήµατος. Έτσι, αν και ηκυτταρίνη δεν έχει καµιά θρεπτική αξία για τον άνθρωπο,παίζει πολύ σηµαντικό ρόλο στη διατροφική υγιεινή του.Τα περισσότερα φρέσκα φρούτα, τα λαχανικά και οι µηαλεσµένοι σπόροι είναι ιδιαιτέρως πλούσια σε κυτταρίνη.
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 89
Η φράση «φυτικές ίνες» στις συσκευασίες των τροφώναναφέρεται κυρίως στην κυτταρίνη.
Υπάρχουν ορισµένοι προκαρυωτικοί οργανισµοί πουπέπτουν την κυτταρίνη διασπώντας την σε µονοµερή γλυ-κόζης. Τέτοιους κυτταρινολυτικούς µικροοργανισµούς φι-λοξενούν, φέρ’ ειπείν, οι αγελάδες στο πρώτο τµήµα τουστοµάχου τους, τη µεγάλη κοιλία (Εικόνα 5.9). Αυτοί οιπροκαρυώτες υδρολύουν την κυτταρίνη των χόρτων καιµετατρέπουν τη γλυκόζη σε άλλες ουσίες τις οποίες µπο-ρεί κατόπιν να χρησιµοποιήσει η αγελάδα για τις δικές της
Μικροϊνίδιακυτταρίνης στοτοίχωμα φυτικούκυττάρου
Κυτταρικότοίχωμα
Μικροϊνίδιο
0,5 μm
10 μm
CH2OH
OH
O
OOOH
CH2OH
OOH
CH2OH
OH
O
O OOOH
OH OH
OH OH
CH2OH
OHO
CH2OH
OH
O
OOOH
CH2OH
OOH
CH2OH
OH
OH
O
O OOOH
CH2OH
OHO
OH OHCH2OH
OH
O
OOOH OH
CH2OH
OOH
CH2OH
Μόριακυτταρίνης
O
O OO
CH2OH
OHO
Τα μικροϊνίδια κυτταρίνηςείναι το κύριο δομικόσυστατικό στο τοίχωμα τωνφυτικών κυττάρων. Για νασχηματιστεί ένα από αυτάσυμμετέχουν γύρω στα 80μόρια κυτταρίνης.
Τα μόρια της κυτταρίνης μπορούν νασυγκρατούνται παράλληλα μεταξύ τους,
μέσω των δεσμών υδρογόνου που σχηματίζονται ανάμεσα στο υδροξύλιο του άνθρακα 3 του ενός μορίου και στο
υδροξύλιο του άνθρακα 6 του παράλληλου μορίου της κυτταρίνης.
Το μόριο της κυτταρίνηςείναι ένα μη διακλαδιζόμενοπολυμερές της γλυκόζης β.
Μονομερέςγλυκόζης β
OH
Εικόνα 5.8 Η διάταξη της κυτταρίνης στο τοίχωμα των φυτικών κυττάρων.
Εικόνα 5.9 Τα μηρυκαστικά, όπως αυτή η αγελάδα, διαθέτουνστο πεπτικό τους σύστημα προκαρυώτες που πέπτουν τηνκυτταρίνη.

διατροφικές ανάγκες. Με παρόµοιο τρόπο εκµεταλλεύον-ται την κυτταρίνη και οι τερµίτες. Αν και οι ίδιοι δεν µπο-ρούν να διασπάσουν την κυτταρίνη, διαθέτουν µικροορ-γανισµούς στον αυλό του εντέρου τους που τρέφονται απότο ξύλο. Την κυτταρίνη πέπτουν επίσης ορισµένοι µύκη-τες, συµβάλλοντας έτσι στην ανακύκλωση των χηµικώνστοιχείων στα χερσαία οικοσυστήµατα.
Ένας άλλος σηµαντικός δοµικός πολυσακχαρίτης εί-ναι η χιτίνη, ο υδατάνθρακας που χρησιµοποιούν τα αρ-θρόποδα (έντοµα, αράχνες, καρκινοειδή και άλλα συγγε-νικά ζώα) για την κατασκευή του εξωσκελετού τους (Ει-κόνα 5.10). Ο εξωσκελετός είναι µια σκληρή επένδυσηπου περιβάλλει τα µαλακά µέρη του ζώου. Η καθαρή χι-τίνη είναι εύκαµπτη, και στην υφή της µοιάζει µε δέρµα,αλλά σκληραίνει πολύ όταν αλληλεπιδράσει µε ανθρακι-κό ασβέστιο. Χιτίνη έχουν επίσης πολλοί µύκητες, οι οποί-οι χρησιµοποιούν αυτό τον πολυσακχαρίτη ως δοµικό υλι-κό (αντί της κυτταρίνης) για την κατασκευή του κυτταρι-κού τους τοιχώµατος. Η χιτίνη µοιάζει µε την κυτταρίνη,αλλά διαφέρει σε µια από τις συνδεδεµένες οµάδες τουανθρακικού δακτυλίου, όπου αντί για υδροξύλιο έχει µιααζωτούχο οµάδα (βλ. Εικόνα 5.10α).
90 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
Τ Μ Η Μ Α 5.3
Τα λιπίδια είναι µια ετερογενήςοµάδα υδρόφοβων µορίων
Τα λιπίδια είναι µια οµάδα που περιλαµβάνει µεγάλα βιο-λογικά µόρια που δεν είναι αληθινά πολυµερή, αλλά ούτεκαι τόσο µεγάλα ώστε να θεωρούνται µακροµόρια. Οι ενώ-σεις που ονοµάζουµε λιπίδια ταξινοµούνται στην ίδια οµά-δα µόνο και µόνο επειδή µοιράζονται ένα κοινό χαρακτη-ριστικό, το ότι δηλαδή αναµειγνύονται ελάχιστα (ή και κα-θόλου) µε το νερό. Η υδρόφοβη συµπεριφορά των λιπιδίωνοφείλεται στη µοριακή τους δοµή. Τα λιπίδια, παρ’ ότι δια-θέτουν ορισµένους πολικούς δεσµούς λόγω της παρουσίαςοξυγόνου στο µόριό τους, αποτελούνται κατά κύριο λόγοαπό περιοχές όπου κυριαρχούν οι υδρογονάνθρακες. Τα λι-πίδια έχουν µεγάλη ποικιλία µορφών και λειτουργιών. Σταλιπίδια ανήκουν οι κηροί και ορισµένες χρωστικές, αλλάεδώ θα εστιάσουµε το ενδιαφέρον µας στα σπουδαιότερα(από βιολογική άποψη) λιπίδια, δηλαδή στα λίπη, τα φω-σφολιπίδια και τα στεροειδή.
Λίπη
Τα λίπη δεν είναι πολυµερή, ωστόσο είναι µεγάλα µόριαπου συντίθενται µε αντιδράσεις αφυδάτωσης από λίγα µι-κρότερα µόρια. Ένα µόριο λίπους κατασκευάζεται απόδύο είδη µικρών µορίων, τη γλυκερόλη και τα λιπαρά οξέα(Εικόνα 5.11α). Η γλυκερόλη είναι µια αλκοόλη µε τρίαάτοµα άνθρακα, καθένα από τα οποία φέρει µια υδροξυ-λοµάδα. Τα λιπαρά οξέα αποτελούνται από έναν επιµήκηανθρακικό σκελετό που στα συνηθέστερα λιπαρά οξέα έχειµήκος 16-18 άτοµα άνθρακα. Ο άνθρακας στο ένα άκροτου λιπαρού οξέος αποτελεί τµήµα µιας καρβοξυλοµάδας,της λειτουργικής οµάδας στην οποία οφείλουν την ονο-µασία τους τα λιπαρά οξέα. Στην καρβοξυλοµάδα συνδέ-
(α) Η δομή του μονομερούς χιτίνης. (β) Η χιτίνη σχηματίζει τον εξωσκελετό των αρθροπόδων. Το τζιτζίκι της φωτογραφίαςεκδύεται, δηλαδή βγαίνει από τον παλιό εξωσκελετό και παίρνει την ενήλικη μορφή του.
(γ) Η χιτίνη χρησιμοποιείται για την κατασκευή ισχυρών και εύκαμπτων χειρουργικών νημάτωνπου αποσυντίθενται μετά το κλείσιμο της πληγήςή της τομής.
C O
CH3
H NH
OHOH
CH2OH
OH H
H
OH H
Εικόνα 5.10 Χιτίνη, ένας δομικός πολυσακχαρίτης.
Ε Λ Ε Γ Χ Ο Σ Ε Ν Ν Ο Ι Ω Ν 5.21. Γράψτε τον τύπο ενός µονοσακχαρίτη που έχει τρία
µόνο άτοµα άνθρακα.
2. Η µαλτόζη σχηµατίζεται από την ένωση δύο µορίωνγλυκόζης µε αντίδραση αφυδάτωσης. Ο µοριακός τύ-πος της γλυκόζης είναι C6H12O6. Ποιος είναι ο µο-ριακός τύπος της µαλτόζης;
3. Τι θα συµβεί αν σε µια αγε-λάδα δοθεί αντιβιοτικό που σκοτώνει όλους τουςπροκαρυωτικούς οργανισµούς του στοµάχου της;Για προτεινόµενες απαντήσεις, βλ. Παράρτηµα Α.
ΤΙ ΘΑ ΓΙΝΟΤΑΝ ΑΝ...;

εται µια επιµήκης αλυσίδα υδρογονάνθρακα. Το γεγονόςότι τα λίπη είναι υδρόφοβα οφείλεται στους σχετικά µηπολικούς δεσµούς C—H της υδρογονανθρακικής αλυσί-δας των λιπαρών οξέων. Τα λίπη δεν αναµειγνύονται µετο νερό, επειδή τα µόρια του νερού σχηµατίζουν µεν δε-σµούς υδρογόνου µεταξύ τους αλλά όχι και µε τα λίπη. Γι’αυτό τον λόγο, αν βάλουµε στο ίδιο µπουκάλι ξίδι (δηλα-δή υδατικό διάλυµα οξικού οξέος) και ένα φυτικό έλαιο(δηλαδή λίπος σε υγρή µορφή, π.χ. ελαιόλαδο), τα δύουγρά δεν θα αναµειχθούν.
Για να συντεθεί ένα µόριο λίπους πρέπει να ενωθούντρία λιπαρά οξέα µε ένα µόριο γλυκερόλης µέσω εστερι-κών δεσµών. Εστερικός δεσµός είναι ο δεσµός που σχη-µατίζεται ανάµεσα σε µια καρβοξυλοµάδα και µια υδρο-ξυλοµάδα. Το λίπος που προκύπτει ονοµάζεται επίσηςτριακυλογλυκερόλη και αποτελείται από τρία λιπαράοξέα που συνδέονται µε ένα µόριο γλυκερόλης. (Από τογεγονός αυτό προκύπτει και η πολύ κοινή ονοµασία «τρι-γλυκερίδια», που αναγράφεται συχνά στη λίστα µε τα συ-στατικά των συσκευασµένων τροφίµων.) Τα τρία λιπαράοξέα στο µορίου του λίπους µπορεί να είναι όλα ίδια, όπωςστην Εικόνα 5.11β, ή να είναι διαφορετικά.
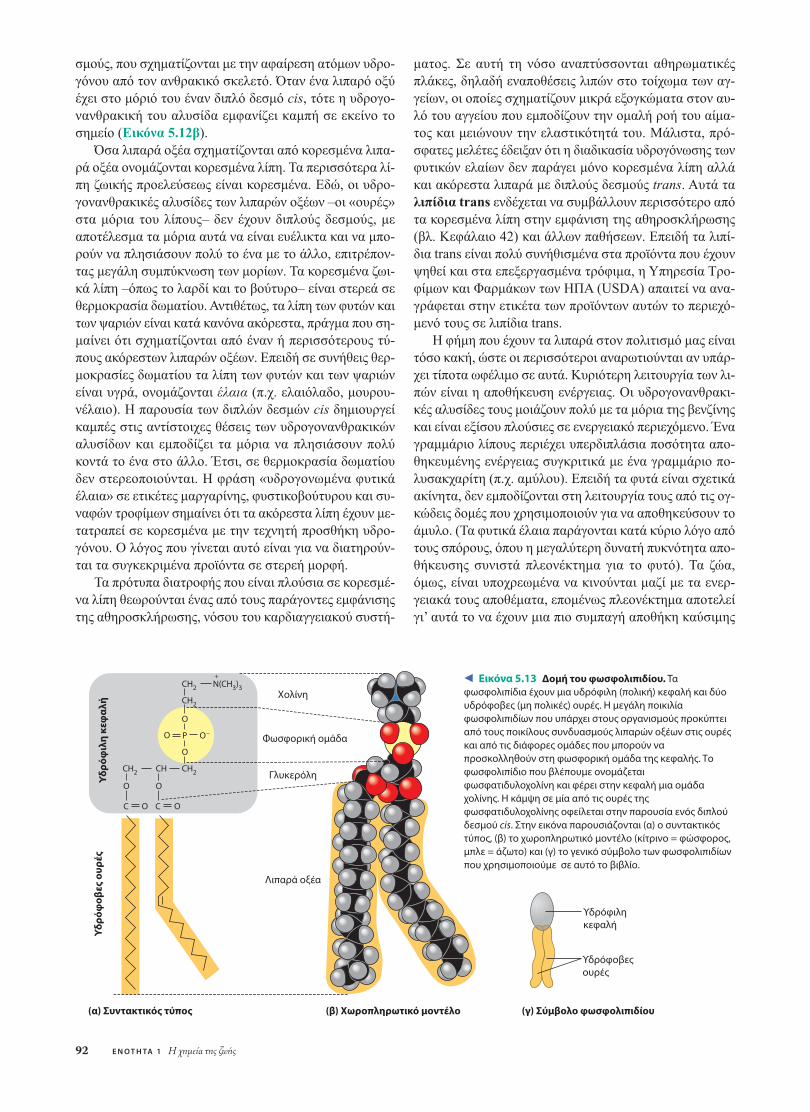
Τα λιπαρά οξέα ποικίλλουν ως προς το µήκος και ωςπρος τη θέση στην οποία µπορεί να φέρουν διπλούς δε-
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 91
σµούς. Ειδικά σε ό,τι αφορά τη διατροφή, χρησιµοποι-ούνται πολύ συχνά οι όροι κορεσµένα και ακόρεστα λιπα-ρά οξέα (Εικόνα 5.12). Αυτοί οι όροι αναφέρονται στη δο-µή της υδρογονανθρακικής αλυσίδας τους. Αν δεν υπάρχειδιπλός δεσµός µεταξύ των ατόµων άνθρακα που συνθέ-τουν την αλυσίδα, τότε ο ανθρακικός σκελετός φέρει τονµεγαλύτερο δυνατό αριθµό συνδεδεµένων ατόµων υδρο-γόνου. Μια τέτοια δοµή λέµε ότι είναι κορεσµένη µε υδρο-γόνο και το αντίστοιχο λιπαρό οξύ ονοµάζεται κορεσµέ-νο λιπαρό οξύ (Εικόνα 5.12α). Τα ακόρεστα λιπαρά οξέαέχουν στο µόριό τους έναν ή περισσότερους διπλούς δε-
Ο διπλός δεσμός cis προκαλεί κάμψη του ανθρακικού σκελετού
(α)
C
H
H
C OH
C
H
OH
O
C
C
C
O
O
O
Κορεσμένα λίπη. Σε θερμοκρασία δωματίου, τα μόρια των κορεσμένων λιπαρών οξέων (όπως το βούτυρο) συσσωρεύονται σε πολύ μικρήαπόσταση μεταξύ τους, με αποτέλεσμα να στερεοποιούνται.
(β) Ακόρεστο λίπος. Σε θερμοκρασία δωματίου, τα μόρια των ακόρεστωνλιπαρών οξέων (όπως το ελαιόλαδο) δεν μπορούν να συσσωρεύονταιπολύ κοντά μεταξύ τους, οπότε παραμένουν σε υγρή μορφή.
Συντακτικός τύπος κορεσμένου μορίου λίπους
Συντακτικός τύπος ακόρεστου μορίου λίπους
C
H
H
C OH
C
H
OH
O
C
C
C
O
O
O
Στεαρικό οξύ, ένα κορεσμένο λιπαρό οξύ
Ελαϊκό οξύ, ένα ακόρεστο λιπαρό οξύ
Εικόνα 5.12 Ακόρεστα και κορεσμένα λιπαρά οξέα και λιπίδια.Σύμφωνα με τη διεθνή χημική σύμβαση, στον δομικό τύπο των λιπώνπαραλείπονται οι άνθρακες και τα συνδεδεμένα σε αυτούς υδρογόνατων υδρογονανθρακικών περιοχών. Στα χωροπληρωτικά μοντέλα τωνλιπαρών οξέων, με μαύρο παριστάνονται τα άτομα του άνθρακα, μεγκρίζο τα άτομα του υδρογόνου και με κόκκινο τα άτομα του οξυγόνου.
(α) Αντίδραση αφυδάτωσης κατά τη σύνθεση λιπιδίου
C
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
HCO
HOC
H
H
C OHH
C
H
OHH
C
H
OH
CH
C
H
H
H2O Λιπαρό οξύ(παλμιτικό οξύ)
Γλυκερόλη
C
O
C
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
H
O C
O
C
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
H
O C
O
C
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
C
H
HC
H
H
H
(β) Λιπίδιο (τριακυλογλυκερόλη)
Εστερικός δεσμός
O H
Εικόνα 5.11 Σύνθεση και δομή λιπιδίου (τριακυλογλυκερόλης).Τα δομικά συστατικά από τα οποία παράγεται ένα λιπίδιο είναι ένα μόριογλυκερόλης και τρία μόρια λιπαρών οξέων. (α) Κάθε φορά πουσυνδέεται ένα μόριο λιπαρού οξέος στη γλυκερόλη αφαιρείται έναμόριο νερού. (β) Μόριο λίπους στο οποίο τα τρία λιπαρά οξέα πουσυνδέονται στη γλυκερόλη είναι ίδια. Οι άνθρακες των λιπαρών οξέωνέχουν διαταχθεί σε πριονωτή διάταξη για να υπενθυμίζουν τονπραγματικό προσανατολισμό που έχουν στον χώρο οι τέσσερεις απλοίδεσμοί κάθε άνθρακα (βλ. Εικόνα 4.3α).
σµούς, που σχηµατίζονται µε την αφαίρεση ατόµων υδρο-γόνου από τον ανθρακικό σκελετό. Όταν ένα λιπαρό οξύέχει στο µόριό του έναν διπλό δεσµό cis, τότε η υδρογο-νανθρακική του αλυσίδα εµφανίζει καµπή σε εκείνο τοσηµείο (Εικόνα 5.12β).
Όσα λιπαρά οξέα σχηµατίζονται από κορεσµένα λιπα-ρά οξέα ονοµάζονται κορεσµένα λίπη. Τα περισσότερα λί-πη ζωικής προελεύσεως είναι κορεσµένα. Εδώ, οι υδρο-γονανθρακικές αλυσίδες των λιπαρών οξέων –οι «ουρές»στα µόρια του λίπους– δεν έχουν διπλούς δεσµούς, µεαποτέλεσµα τα µόρια αυτά να είναι ευέλικτα και να µπο-ρούν να πλησιάσουν πολύ το ένα µε το άλλο, επιτρέπον-τας µεγάλη συµπύκνωση των µορίων. Τα κορεσµένα ζωι-κά λίπη –όπως το λαρδί και το βούτυρο– είναι στερεά σεθερµοκρασία δωµατίου. Αντιθέτως, τα λίπη των φυτών καιτων ψαριών είναι κατά κανόνα ακόρεστα, πράγµα που ση-µαίνει ότι σχηµατίζονται από έναν ή περισσότερους τύ-πους ακόρεστων λιπαρών οξέων. Επειδή σε συνήθεις θερ-µοκρασίες δωµατίου τα λίπη των φυτών και των ψαριώνείναι υγρά, ονοµάζονται έλαια (π.χ. ελαιόλαδο, µουρου-νέλαιο). Η παρουσία των διπλών δεσµών cis δηµιουργείκαµπές στις αντίστοιχες θέσεις των υδρογονανθρακικώναλυσίδων και εµποδίζει τα µόρια να πλησιάσουν πολύκοντά το ένα στο άλλο. Έτσι, σε θερµοκρασία δωµατίουδεν στερεοποιούνται. Η φράση «υδρογονωµένα φυτικάέλαια» σε ετικέτες µαργαρίνης, φυστικοβούτυρου και συ-ναφών τροφίµων σηµαίνει ότι τα ακόρεστα λίπη έχουν µε-τατραπεί σε κορεσµένα µε την τεχνητή προσθήκη υδρο-γόνου. Ο λόγος που γίνεται αυτό είναι για να διατηρούν-ται τα συγκεκριµένα προϊόντα σε στερεή µορφή.
Τα πρότυπα διατροφής που είναι πλούσια σε κορεσµέ-να λίπη θεωρούνται ένας από τους παράγοντες εµφάνισηςτης αθηροσκλήρωσης, νόσου του καρδιαγγειακού συστή-
92 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
µατος. Σε αυτή τη νόσο αναπτύσσονται αθηρωµατικέςπλάκες, δηλαδή εναποθέσεις λιπών στο τοίχωµα των αγ-γείων, οι οποίες σχηµατίζουν µικρά εξογκώµατα στον αυ-λό του αγγείου που εµποδίζουν την οµαλή ροή του αίµα-τος και µειώνουν την ελαστικότητά του. Μάλιστα, πρό-σφατες µελέτες έδειξαν ότι η διαδικασία υδρογόνωσης τωνφυτικών ελαίων δεν παράγει µόνο κορεσµένα λίπη αλλάκαι ακόρεστα λιπαρά µε διπλούς δεσµούς trans. Αυτά ταλιπίδια trans ενδέχεται να συµβάλλουν περισσότερο απότα κορεσµένα λίπη στην εµφάνιση της αθηροσκλήρωσης(βλ. Κεφάλαιο 42) και άλλων παθήσεων. Επειδή τα λιπί-δια trans είναι πολύ συνήθισµένα στα προϊόντα που έχουνψηθεί και στα επεξεργασµένα τρόφιµα, η Υπηρεσία Τρο-φίµων και Φαρµάκων των ΗΠΑ (USDA) απαιτεί να ανα-γράφεται στην ετικέτα των προϊόντων αυτών το περιεχό-µενό τους σε λιπίδια trans.
Η φήµη που έχουν τα λιπαρά στον πολιτισµό µας είναιτόσο κακή, ώστε οι περισσότεροι αναρωτιούνται αν υπάρ-χει τίποτα ωφέλιµο σε αυτά. Κυριότερη λειτουργία των λι-πών είναι η αποθήκευση ενέργειας. Οι υδρογονανθρακι-κές αλυσίδες τους µοιάζουν πολύ µε τα µόρια της βενζίνηςκαι είναι εξίσου πλούσιες σε ενεργειακό περιεχόµενο. Έναγραµµάριο λίπους περιέχει υπερδιπλάσια ποσότητα απο-θηκευµένης ενέργειας συγκριτικά µε ένα γραµµάριο πο-λυσακχαρίτη (π.χ. αµύλου). Επειδή τα φυτά είναι σχετικάακίνητα, δεν εµποδίζονται στη λειτουργία τους από τις ογ-κώδεις δοµές που χρησιµοποιούν για να αποθηκεύσουν τοάµυλο. (Τα φυτικά έλαια παράγονται κατά κύριο λόγο απότους σπόρους, όπου η µεγαλύτερη δυνατή πυκνότητα απο-θήκευσης συνιστά πλεονέκτηµα για το φυτό). Τα ζώα,όµως, είναι υποχρεωµένα να κινούνται µαζί µε τα ενερ-γειακά τους αποθέµατα, εποµένως πλεονέκτηµα αποτελείγι’ αυτά το να έχουν µια πιο συµπαγή αποθήκη καύσιµης
CH2 N(CH3)3
CH2
O
P
O
CH2
O
CH
O
CH2
O O–
C O C O
+
Χολίνη
Φωσφορική ομάδα
Γλυκερόλη
Λιπαρά οξέα
Υδρόφιληκεφαλή
Υδρόφοβεςουρές
(α) Συντακτικός τύπος (β) Χωροπληρωτικό μοντέλο (γ) Σύμβολο φωσφολιπιδίου
Υδρό
φιλ
ηκε
φαλ
ήΥδ
ρόφ
οβες
ουρέ
ς
Εικόνα 5.13 Δομή του φωσφολιπιδίου. Ταφωσφολιπίδια έχουν μια υδρόφιλη (πολική) κεφαλή και δύουδρόφοβες (μη πολικές) ουρές. Η μεγάλη ποικιλίαφωσφολιπιδίων που υπάρχει στους οργανισμούς προκύπτειαπό τους ποικίλους συνδυασμούς λιπαρών οξέων στις ουρέςκαι από τις διάφορες ομάδες που μπορούν ναπροσκολληθούν στη φωσφορική ομάδα της κεφαλής. Τoφωσφολιπίδιο που βλέπουμε ονομάζεταιφωσφατιδυλοχολίνη και φέρει στην κεφαλή μια ομάδαχολίνης. Η κάμψη σε μία από τις ουρές τηςφωσφατιδυλοχολίνης οφείλεται στην παρουσία ενός διπλούδεσμού cis. Στην εικόνα παρουσιάζονται (α) ο συντακτικόςτύπος, (β) το χωροπληρωτικό μοντέλο (κίτρινο = φώσφορος,μπλε = άζωτο) και (γ) το γενικό σύμβολο των φωσφολιπιδίωνπου χρησιμοποιούμε σε αυτό το βιβλίο.

ύλης, που είναι το λίπος. Ο άνθρωπος και πολλά άλλα θη-λαστικά αποθηκεύουν το λίπος για τις µακροπρόθεσµεςενεργειακές τους ανάγκες σε ειδικά αποθηκευτικά κύττα-ρα που ονοµάζονται λιποκύτταρα (βλ. Εικόνα 4.6α), ταοποία διογκώνονται όταν αποθηκεύεται λίπος στο εσωτε-ρικό τους και συρρικνώνονται όταν οι ανάγκες απαιτούντην κατανάλωσή του. Εκτός από αποθήκες ενέργειας, ταλιποκύτταρα επενδύουν διάφορα ζωτικά όργανα, όπωςτους νεφρούς, ενώ υπάρχει και ένα στρώµα λίπους κάτωαπό το δέρµα, που λειτουργεί µονωτικά για το σώµα. Τοστρώµα του υποδόριου λίπους είναι ιδιαίτερα παχύ στιςφάλαινες, τις φώκιες και στα περισσότερα θαλάσσια θη-λαστικά, παρέχοντας προστασία στα ζώα αυτά από το ψυ-χρό νερό των ωκεανών.
Φωσφολιπίδια
Τα κύτταρα δεν θα µπορούσαν να υπάρχουν χωρίς ένανάλλο τύπο λιπιδίων, τα φωσφολιπίδια (Εικόνα 5.13). Ταφωσφολιπίδια έχουν ζωτική σηµασία για το κύτταρο, διό-τι από αυτά συντίθενται οι κυτταρικές µεµβράνες. Η δοµήτους αποτελεί κλασικό παράδειγµα για το πώς η δοµήεναρµονίζεται µε τη λειτουργία, σε µοριακό επίπεδο.Όπως φαίνεται στην Εικόνα 5.13, το µόριο των φωσφολι-πιδίων µοιάζει µε εκείνο των λιπών, αλλά έχει δύο µόνολιπαρά οξέα συνδεδεµένα µε τη γλυκερόλη, αντί για τρία.Το τρίτο υδροξύλιο της γλυκερόλης ενώνεται µε µια φω-σφορική οµάδα που φέρει αρνητικό ηλεκτρικό φορτίο.Κατά συνέπεια, διάφορα µικρά µόρια που φέρουν ηλε-κτρικό φορτίο, ή είναι πολικά, µπορούν να συνδεθούν µετη φωσφορική οµάδα και να σχηµατίσουν µεγάλη ποικιλίαφωσφολιπιδίων.
Τα δύο άκρα των φωσφολιπιδίων εκδηλώνουν διαφο-ρετική συµπεριφορά έναντι του νερού. Οι υδρογονανθρα-κικές ουρές είναι υδρόφοβες και απωθούνται από αυτό.
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 93
Ωστόσο, η φωσφορική οµάδα και όσα φορτισµένα ή πο-λικά µόρια είναι συνδεδεµένα µαζί της σχηµατίζουν µιαυδρόφιλη κεφαλή που εµφανίζει χηµική συγγένεια µε τονερό. Αν προσθέσουµε φωσφολιπίδια στο νερό, συσσω-µατώνονται από µόνα τους σε ένα διπλό στρώµα (µια «δι-πλοστιβάδα») το οποίο σχηµατίζει ένα προστατευτικόστρώµα γύρω από τα υδρόφοβα τµήµατα, εµποδίζονταςτην επαφή τους µε το νερό (Εικόνα 5.14).
Τα φωσφολιπίδια διατάσσονται µε τον ίδιο τρόπο, δη-λαδή σε διπλοστιβάδες, στην επιφάνεια των κυττάρων. Οιυδρόφιλες κεφαλές των µορίων διατάσσονται στις δύοεξωτερικές επιφάνειες της διπλοστιβάδας µε αποτέλεσµανα βρίσκονται σε άµεση επαφή µε το υδατικό διάλυµα, τό-σο στο εξωτερικό όσο και στο εσωτερικό του κυττάρου.Οι υδρόφοβες ουρές διατάσσονται στο εσωτερικό της δι-πλοστιβάδας χωρίς να έρχονται σε επαφή µε το νερό. Οιδιπλοστιβάδες των φωσφολιπιδίων σχηµατίζουν ένα είδοςφραγµού µεταξύ του κυττάρου και του εξωτερικού του πε-ριβάλλοντος. Για να είµαστε ακριβείς, τα κύτταρα δεν θαµπορούσαν να υπάρξουν χωρίς φωσφολιπίδια.
Στεροειδή
Στην κατηγορία των στεροειδών ανήκουν πολλές ορµό-νες, αλλά και η χοληστερόλη. Τα στεροειδή είναι λιπίδιαστα οποία ο ανθρακικός σκελετός αποτελείται από τέσσε-ρεις συγχωνευµένους δακτυλίους (Εικόνα 5.15). Τα διά-φορα στεροειδή λιπίδια διαφέρουν µόνο ως προς τις χηµι-κές οµάδες που είναι συνδεδεµένες σε αυτό το σύνολο τωντεσσάρων δακτυλίων. Η χοληστερόλη είναι ένα πολύ δια-δεδοµένο συστατικό των κυτταρικών µεµβρανών των ζωι-κών κυττάρων, αλλά και ένα πρόδροµο µόριο από τοοποίο µπορούν να συντεθούν άλλα στεροειδή. Στα σπον-δυλωτά, η χοληστερόλη συντίθεται στο ήπαρ. Πολλές ορ-µόνες, µεταξύ των οποίων και οι αναπαραγωγικές ορµόνεςτων θηλαστικών, είναι στεροειδή που παράγονται από τηχοληστερόλη (βλ. Εικόνα 4.9). Η χοληστερόλη, λοιπόν, εί-
CH3
HO
CH3
H3C CH3
CH3
Εικόνα 5.15 Χοληστερόλη, ένα στεροειδές. Η χοληστερόλη είναιένα μόριο από το οποίο συντίθενται πολλά άλλα στεροειδή, μεταξύ τωνοποίων και οι ορμόνες του φύλου. Η ποικιλία των στεροειδών προκύπτειαπό τις διάφορες ομάδες που συνδέονται περιφερειακά στην τετράδατων συζευγμένων δακτυλίων (περιοχή με κίτρινη σήμανση).
ΝΕΡΟ
ΝΕΡΟ
Υδρόφιληκεφαλή
Υδρόφοβηουρά
Εικόνα 5.14 Η διπλοστιβάδα που σχηματίζουν αυθόρμητα ταφωσφολιπίδια όταν βρεθούν σε υδατικό διάλυμα. Η διπλοστιβάδατων φωσφολιπιδίων είναι ο τρόπος με τον οποίο δομούνται όλες οιβιολογικές μεμβράνες. Παρατηρήστε ότι, λόγω της διάταξης τωνφωσφολιπιδίων στη διπλοστιβάδα, οι υδρόφιλες κεφαλές έρχονται σεεπαφή με το νερό, ενώ οι υδρόφοβες ουρές βρίσκονται σε επαφήμεταξύ τους και μακριά από το νερό.

ναι ένα κοµβικό µόριο για τα ζώα, αν και η παρουσία υψη-λών συγκεντρώσεων του µορίου της στο αίµα ενδεχοµέ-νως συµβάλλει στην ανάπτυξη αθηροσκλήρωσης. Τόσοτα κορεσµένα λιπαρά όσο και τα λιπίδια trans φαίνεται ότιασκούν αρνητική επίδραση στην υγεία αυξάνοντας τα επί-πεδα της χοληστερόλης στο αίµα.
94 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
συνάγεται και από το µεγάλο εύρος λειτουργιών που επι-τελούν, οι πρωτεΐνες εµφανίζουν τεράστια ποικιλία στηδοµή τους, ενώ το σχήµα κάθε πρωτεΐνης στον χώρο είναιµοναδικό.
Πολυπεπτίδια
Παρά την ποικιλοµορφία τους, οι πρωτεΐνες δεν παύουννα είναι πολυµερή που όλα συντίθενται από το ίδιο σύνο-λο 20 αµινοξέων. Τα πολυµερή των αµινοξέων ονοµάζον-ται πολυπεπτίδια. Μια πρωτεΐνη αποτελείται από ένα ήπερισσότερα πολυπεπτίδια, καθένα από τα οποία αναδι-πλώνεται και συστρέφεται στον χώρο αποκτώντας τελικάµια πολύ εξειδικευµένη τριδιάστατη δοµή.
Αμινοξέα: τα μονομερή των πρωτεϊνών
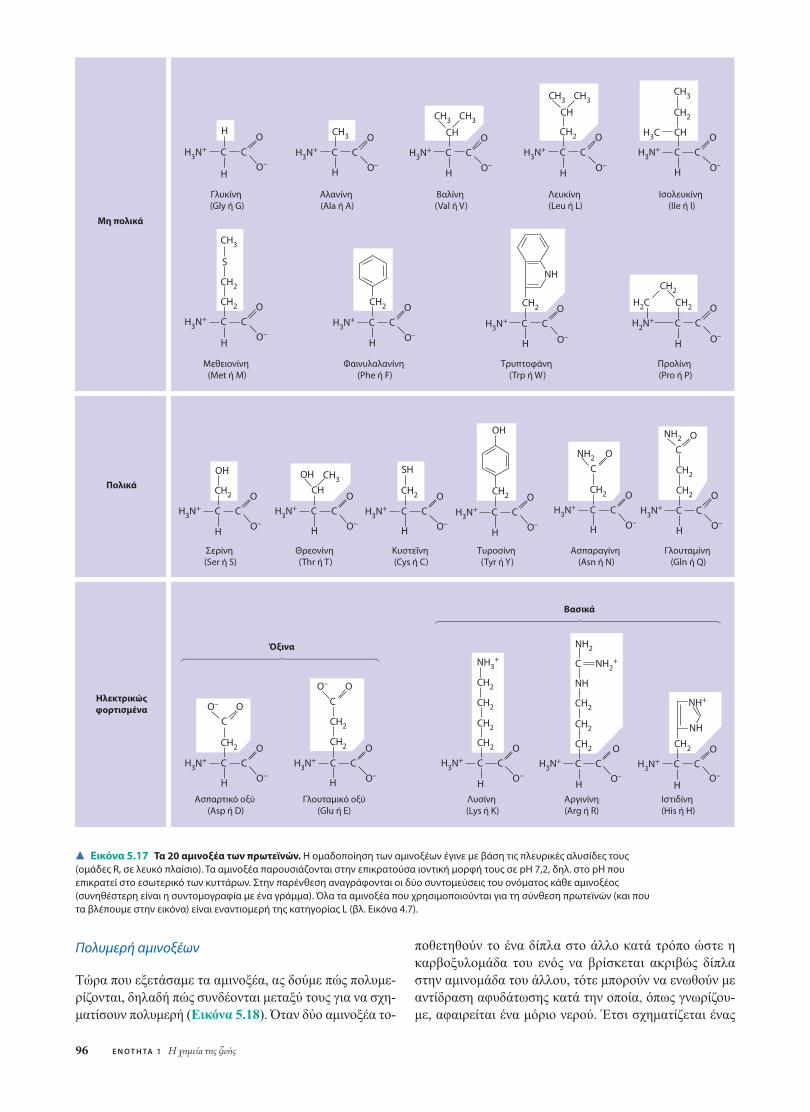
Όλα τα αµινοξέα έχουν την ίδιαδοµή. Τα αµινοξέα είναι οργα-νικά µόρια που φέρουν στο µό-ριό τους ένα καρβοξύλιο και µίααµινοµάδα (βλ. Κεφάλαιο 4). Τοσχεδιάγραµµα στα δεξιά της πα-ραγράφου δείχνει τον γενικό τύ-πο ενός αµινοξέος. Στο κέντρο του υπάρχει ένα ασύµµετροάτοµο άνθρακα, που ονοµάζεται άνθρακας άλφα (α). Οιτέσσερεις χηµικοί συνεταίροι του είναι µία αµινοµάδα, µίακαρβοξυλοµάδα, ένα άτοµο υδρογόνου και µία οµάδα πουποικίλλει από αµινοξύ σε αµινοξύ και συµβολίζεται µε τογράµµα R. Η οµάδα R ονοµάζεται επίσης πλευρική αλυ-σίδα.
Ε Λ Ε Γ Χ Ο Σ Ε Ν Ν Ο Ι Ω Ν 5.31. Συγκρίνετε τη δοµή ενός µορίου λίπους (τριγλυκερι-
δίου) µε τη δοµή ενός φωσφολιπιδίου.
2. Γιατί κατατάσσονται στα λιπίδια οι αναπαραγωγικέςορµόνες του ανθρώπου;
3. Έστω ότι ένα σταγονίδιοελαίου, π.χ. σε κύτταρο φυτικού σπόρου, περιβάλ-λεται από κυτταρική µεµβράνη. Περιγράψτε και εξη-γήστε το σχήµα που θα µπορούσε αυτό να πάρει.Για προτεινόµενες απαντήσεις, βλ. Παράρτηµα Α.
ΤΙ ΘΑ ΓΙΝΟΤΑΝ ΑΝ...;
Τ Μ Η Μ Α 5.4
Οι πρωτεΐνες έχουν διάφορες δοµές,οπότε εµφανίζουν µεγάλο εύροςδράσεων
Όλες σχεδόν οι λειτουργίες ενός ζωντανού οργανισµούεξαρτώνται από τις πρωτεΐνες. Η σπουδαιότητά τους, άλ-λωστε, υπονοείται και από το όνοµά τους. Στα πιο πολλάείδη κυττάρων, οι πρωτεΐνες συνιστούν πάνω από το 50%της ξηρής τους µάζας, ο δε ρόλος τους είναι καθοριστικόςσε οτιδήποτε κάνει ένας οργανισµός. Μερικές πρωτεΐνεςεπιταχύνουν τις χηµικές αντιδράσεις που λαµβάνουν χώ-ρα στους οργανισµούς, ενώ άλλες έχουν δοµική ή υπο-στηρικτική λειτουργία, όπως στην αποθήκευση, τη µετα-φορά, την επικοινωνία µεταξύ κυττάρων, την κίνηση ή τηνάµυνα έναντι ξένων ουσιών (Πίνακας 5.1).
Είναι αδύνατο να υπάρξει ζωή χωρίς ένζυµα, τα πε-ρισσότερα από τα οποία είναι πρωτεΐνες. Οι πρωτεΐνεςπου λειτουργούν ως ένζυµα ρυθµίζουν τον µεταβολισµότου οργανισµού καθώς δρουν ως καταλύτες, δηλαδή ωςχηµικοί παράγοντες που επιταχύνουν επιλεκτικά τις πε-ρισσότερες χηµικές αντιδράσεις, χωρίς οι ίδιες να κατα-ναλώνονται κατά τις αντιδράσεις αυτές (Εικόνα 5.16).Επειδή τα ένζυµα µπορούν να επιτελούν τη λειτουργίατους ξανά και ξανά, θα µπορούσαµε να τα θεωρήσουµεως τους κινητήρες που κρατούν τα κύτταρα σε λειτουρ-γία και διεκπεραιώνουν όλες εκείνες τις διαδικασίες τιςοποίες θα χαρακτηρίζαµε ως έµβιες.
Ένας άνθρωπος διαθέτει δεκάδες χιλιάδες διαφορετικάείδη πρωτεϊνών, το καθένα µε ειδική δράση και λειτουρ-γία. Για την ακρίβεια, οι πρωτεΐνες ανήκουν στα πιο περί-πλοκα από δοµική άποψη µόρια που γνωρίζουµε. Όπως
Ένζυμο(σακχαράση)
Υπόστρωμα(σακχαρόζη)
Γλυκόζη
ΦρουκτόζηH2O
OH
HO
Η ενεργός θέση του ενζύμου είναι διαθέσιμη για να δεχθεί ένα μόριο υποστρώματος, δηλαδή ένα μόριο του αντιδρώντος σώματος πάνω στο οποίο θα ασκήσει τη δράση του το ένζυμο.
1 Το υπόστρωμα δεσμεύεται στο ένζυμο.2
Τα προϊόντα αποδεσμεύονταιαπό το ένζυμο.4 Το υπόστρωμα μετατρέπεται
στα προϊόντα.3
Εικόνα 5.16 Ο καταλυτικός κύκλος ενός ενζύμου. Το ένζυμοσακχαράση (σουκράση) καταλύει την υδρόλυση της σακχαρόζης σεγλυκόζη και φρουκτόζη. Επειδή δρα ως καταλύτης, η σακχαράση δενκαταναλώνεται κατά τη διάρκεια του κύκλου, αλλά παραμένει διαθέσιμηγια περαιτέρω καταλυτική δράση.
OH
O
CCC
R
H
N
άνθρακας a
H
H
Αμινομάδα Καρβο-ξυλομάδα

Στην Εικόνα 5.17 παρουσιάζονται τα 20 αµινοξέα πουχρησιµοποιούν τα κύτταρα για να συνθέσουν τις χιλιάδεςπρωτεΐνες τους. Οι καρβοξυλοµάδες και οι αµινοµάδες πα-ρουσιάζονται στην ιοντισµένη µορφή τους, όπως δηλαδήαπαντούν στις συνθήκες pH που συνήθως επικρατούν στοεσωτερικό των κυττάρων. Η οµάδα R µπορεί να είναι εί-τε απλώς ένα άτοµο υδρογόνου, όπως συµβαίνει στο αµι-νοξύ γλυκίνη (το µόνο χωρίς ασύµµετρο άτοµο άνθρακα,αφού δύο από τους συνεταίρους του, τα άτοµα του υδρο-γόνου, είναι ίδια), είτε ένας ανθρακικός σκελετός στονοποίο συνδέονται διάφορες άλλες λειτουργικές οµάδες,όπως επί παραδείγµατι συµβαίνει στη γλουταµίνη. (Στηνπραγµατικότητα, οι οργανισµοί έχουν και µερικά ακόµηαµινοξέα· ορισµένα από αυτά ανιχνεύονται περιστασιακάστις πρωτεΐνες. Επειδή όµως είναι σχετικά σπάνια, δεν πε-ριλαµβάνονται στην Εικόνα 5.17.)
Οι φυσικές και χηµικές ιδιότητες της πλευρικής αλυσί-δας καθορίζουν τα ιδιαίτερα χαρακτηριστικά που έχει κά-θε αµινοξύ και επηρεάζουν τον λειτουργικό ρόλο που αυ-
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 95
τό παίζει σε ένα πολυπεπτίδιο. Στην Εικόνα 5.17, τα αµι-νοξέα έχουν οµαδοποιηθεί ανάλογα µε τις ιδιότητες τωνπλευρικών αλυσίδων τους. Η µία οµάδα περιλαµβάνει τααµινοξέα που έχουν µη πολικές πλευρικές αλυσίδες, ταοποία είναι υδρόφοβα. Μια άλλη οµάδα αποτελείται απόαµινοξέα µε πολικές πλευρικές αλυσίδες, τα οποία είναιεποµένως υδρόφιλα. Στην οµάδα των όξινων αµινοξέωνανήκουν εκείνα στα οποία η πλευρική αλυσίδα είναι εν γέ-νει αρνητικά φορτισµένη επειδή φέρει µια καρβοξυλοµά-δα που συνήθως διίσταται (ιοντίζεται) στο pH του ενδο-κυττάριου περιβάλλοντος. Τέλος, στην οµάδα των βασι-κών αµινοξέων ανήκουν εκείνα που στην πλευρική αλυσί-δα τους υπάρχει κάποια αµινοµάδα και που, εποµένως, εί-ναι εν γένει θετικά φορτισµένα. (Προφανώς, όλα τα αµι-νοξέα έχουν αµινοµάδα και καρβοξυλοµάδα. Ο όρος όξιναή βασικά αµινοξέα αφορά µόνο τις ιδιότητες που προκύ-πτουν από τις οµάδες των πλευρικών αλυσίδων.) Επειδήτόσο οι όξινες όσο και οι βασικές πλευρικές αλυσίδες είναιφορτισµένες, όλα τα αντίστοιχα αµινοξέα είναι υδρόφιλα.
Πίνακας 5.1 Σύνοψη πρωτεϊνικών λειτουργιών
Είδος πρωτεΐνης Λειτουργία Παραδείγματα
Ένζυµα Επιλεκτική επιτάχυνση Τα πεπτικά ένζυµα καταλύουν την υδρόλυση των πολυµερώνχηµικών αντιδράσεων στα τρόφιµα.
∆οµικές πρωτεΐνες Υποστήριξη Τα έντοµα και οι αράχνες χρησιµοποιούν ίνες από µετάξιγια να κατασκευάσουν (αντίστοιχα) τα κουκούλια καιτους ιστούς τους. Το κολλαγόνο και η ελαστίνη παρέχουντο ινώδες υπόβαθρο για τον συνδετικό ιστό των ζώων.Η κερατίνη είναι η πρωτεΐνη από την οποία φτιάχνονταιοι τρίχες, τα κέρατα, τα πούπουλα και διάφορα άλλα εξαρτήµατατου δέρµατος.
Αποθηκευτικές Αποθήκευση αµινοξέων Η οβαλβουµίνη είναι η πρωτεΐνη στο ασπράδι του αυγούπρωτεΐνες την οποία χρησιµοποιεί το αναπτυσσόµενο έµβρυο ως πηγή
αµινοξέων. Η καζεΐνη, η πρωτεΐνη του γάλακτος, είναι η κύριαπηγή αµινοξέων για τα νεογνά των θηλαστικών. Τα φυτά έχουναποθηκευτικές πρωτεΐνες στους σπόρους τους.
Μεταφορικές Μεταφορά άλλων Η αιµοσφαιρίνη, η σιδηρούχος πρωτεΐνη στο αίµα τωνπρωτεΐνες ουσιών σπονδυλωτών, µεταφέρει οξυγόνο από τους πνεύµονες σε άλλα
τµήµατα του σώµατος. Άλλες πρωτεΐνες µεταφέρουν ουσίες απότη µία πλευρά των κυτταρικών µεµβρανών στην άλλη.
Ορµονικές Συντονισµός Η ινσουλίνη, ορµόνη που εκκρίνεται από το πάγκρεας,πρωτεΐνες δραστηριοτήτων ρυθµίζει τη συγκέντρωση της γλυκόζης στο αίµα
των οργανισµών των σπονδυλωτών.Πρωτεϊνικοί Απόκριση των κυττάρων Οι υποδοχείς που είναι ενσωµατωµένοι στη µεµβράνη
υποδοχείς σε διάφορα χηµικά των νευρικών κυττάρων ανιχνεύουν χηµικά σήµαταερεθίσµατα που απελευθερώνονται από άλλα νευρικά κύτταρα.
Πρωτεΐνες Κίνηση Η ακτίνη και η µυοσίνη είναι υπεύθυνες για τη µυικήσυστολής και συστολή. Άλλες πρωτεΐνες ευθύνονται για τις κινήσειςκίνησης ορισµένων ειδικών κυτταρικών οργανιδίων που ονοµάζονται
βλεφαρίδες και µαστίγια.Αµυντικές Προστασία κατά των Τα αντισώµατα καταπολεµούν βακτήρια και ιούς.
πρωτεΐνες ασθενειών
96 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
CH2
CH2
S
CH3
Φαινυλαλανίνη (Phe ή F)
Μεθειονίνη (Met ή M)
Προλίνη (Pro ή P)
O–
OCC
H
H
H3N+O
CC
H
H3N+
CH2
O–
OCC
H
H3N+
O–
OCH2
CC
H
H3N+
Τρυπτοφάνη (Trp ή W)
O–
OCH2
C
O–
OCC
H
H3N+
NHCH2
CH2
C
H
H2N+
H2C
Μη πολικά
Λευκίνη (Leu ή L)
Βαλίνη (Val ή V)
Αλανίνη (Ala ή A)
Γλυκίνη (Gly ή G)
Ισολευκίνη (Ile ή I)
O–
OCH3
CC
H
H3N+
CH3 CH3
O–
OCH
CH3 CH3
CH
CC
H
H3N+
H3C
O– O–
OCC
H
H3N+
CH
CH3
CH2
O–
OCC
H
H3N+
OH
CH2
OH CH3
O–
OCH
CC
H
H3N+
O–
OCC
H
H3N+
SH
CH2
O–
OCH2
CC
H
H3N+
O–
OCC
H
H3N+
NH2 OC
CH2Πολικά
Τυροσίνη (Tyr ή Y)
Θρεονίνη (Thr ή T)
Σερίνη (Ser ή S)
Ασπαραγίνη (Asn ή N)
Γλουταμίνη (Gln ή Q)
Κυστεΐνη (Cys ή C)
OH
O–
O–
OCC
H
H3N+
O– OC
CH2
H
CH2
CH2
Όξινα
OC
CH2
O–
OCC
H
H3N+
CH2
CH2
CH2
NH3+
CH2
O
O–CC
H
H3N+
O–
OCCH3N+
CH2
NH
CH2
NH2+C
NH2
O
O–
CH2
CC
H
H3N+
NH+
NH
O
CH2
C
CH2
C
H
NH2
Βασικά
Λυσίνη (Lys ή K)
Γλουταμικό οξύ(Glu ή E)
Ασπαρτικό οξύ (Asp ή D)
Αργινίνη(Arg ή R)
Ιστιδίνη (His ή H)
Ηλεκτρικώςφορτισμένα
O–
OCH3N+
Εικόνα 5.17 Τα 20 αμινοξέα των πρωτεϊνών. Η ομαδοποίηση των αμινοξέων έγινε με βάση τις πλευρικές αλυσίδες τους(ομάδες R, σε λευκό πλαίσιο). Τα αμινοξέα παρουσιάζονται στην επικρατούσα ιοντική μορφή τους σε pH 7,2, δηλ. στο pH πουεπικρατεί στο εσωτερικό των κυττάρων. Στην παρένθεση αναγράφονται οι δύο συντομεύσεις του ονόματος κάθε αμινοξέος(συνηθέστερη είναι η συντομογραφία με ένα γράμμα). Όλα τα αμινοξέα που χρησιμοποιούνται για τη σύνθεση πρωτεϊνών (και πουτα βλέπουμε στην εικόνα) είναι εναντιομερή της κατηγορίας L (βλ. Εικόνα 4.7).
Πολυμερή αμινοξέων
Τώρα που εξετάσαµε τα αµινοξέα, ας δούµε πώς πολυµε-ρίζονται, δηλαδή πώς συνδέονται µεταξύ τους για να σχη-µατίσουν πολυµερή (Εικόνα 5.18). Όταν δύο αµινοξέα το-
ποθετηθούν το ένα δίπλα στο άλλο κατά τρόπο ώστε ηκαρβοξυλοµάδα του ενός να βρίσκεται ακριβώς δίπλαστην αµινοµάδα του άλλου, τότε µπορούν να ενωθούν µεαντίδραση αφυδάτωσης κατά την οποία, όπως γνωρίζου-µε, αφαιρείται ένα µόριο νερού. Έτσι σχηµατίζεται ένας

οµοιοπολικός δεσµός, που ονοµάζεται πεπτιδικός δε-σµός. Αν αυτή η διαδικασία επαναληφθεί πολλές φορές,τότε προκύπτει ένα πολυπεπτίδιο, ένα πολυµερές δηλαδήαπό πολλά αµινοξέα που συνδέονται µε πεπτιδικούς δε-σµούς. Στο ένα άκρο της πολυπεπτιδικής αλυσίδας υπάρ-χει µια ελεύθερη αµινοµάδα, ενώ στο αντίθετο άκρο υπάρ-χει µια ελεύθερη καρβοξυλοµάδα. Έτσι, η αλυσίδα έχειένα αµινοτελικό άκρο (άκρο Ν) και ένα καρβοξυτελικόάκρο (άκρο C). Η επαναλαµβανόµενη αλληλουχία τωνατόµων, που έχει τονιστεί µε µοβ στην Εικόνα 5.18β, ονο-µάζεται πολυπεπτιδικός σκελετός. Τα διάφορα άτοµα ήοµάδες που προεξέχουν από τον σκελετό αυτό δεν είναιτίποτε άλλο παρά οι πλευρικές οµάδες των επιµέρους αµι-νοξέων. Το µήκος ενός πολυπεπτιδίου µπορεί να ποικίλλειαπό λίγα µονοµερή έως χίλια, ή και πάνω από χίλια. Κά-θε συγκεκριµένο πολυπεπτίδιο έχει µια µοναδική γραµµι-κή ακολουθία αµινοξέων. Η τεράστια ποικιλία πολυπε-πτιδίων που συναντάµε στη φύση αποτελεί την έµπρακτηέκφραση µιας σηµαντικής έννοιας που έχουµε ήδη ανα-φέρει, δηλαδή του ότι τα κύτταρα µπορούν να κατα-σκευάζουν µια τεράστια ποικιλία πολυµερών από λίγα µό-νο µονοµερή, λόγω της ποικιλόµορφης διάταξης των µο-νοµερών αυτών.
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 97
Δομή και λειτουργία των πρωτεϊνών
Η εξειδικευµένη δράση των πρωτεϊνών προκύπτει από τιςλεπτοµέρειες της δοµής τους στον χώρο. Η δοµή των πρω-τεϊνών έχει διάφορα επίπεδα, αλλά το απλούστερο (καιαυτό απ’ όπου ξεκινά η δοµή µιας πρωτεΐνης) είναι η σει-ρά των αµινοξέων της. Πρωτοπόρος στον προσδιορισµότης ακολουθίας των αµινοξέων µιας πρωτεΐνης ήταν ο Fre-derick Sanger, που µαζί µε τους συναδέλφους του στο Πα-νεπιστήµιο του Cambridge, στην Αγγλία, εργάστηκε γιανα προσδιορίσει τη δοµή της πρωτεΐνης ινσουλίνη στα τέ-λη της δεκαετίας του 1940 και στις αρχές της δεκαετίαςτου 1950. Ο Sanger χρησιµοποίησε ειδικούς παράγοντεςπου διασπούν τα πολυπεπτίδια, για να τεµαχίσει την ιν-σουλίνη σε διάφορα τµήµατα, και κατόπιν προσδιόρισε µεποικίλες χηµικές µεθόδους την ακολουθία των αµινοξέωνσ’ αυτά τα µικρά τµήµατα. Ύστερα από προσπάθειες ετών,ο Sanger και η οµάδα του κατάφεραν να ανασυνθέσουνπλήρως την ακολουθία των αµινοξέων της ινσουλίνης.Έκτοτε, τα περισσότερα βήµατα για τον προσδιορισµό τηςαµινοξικής ακολουθίας ενός πολυπεπτιδίου έχουν αυτο-µατοποιηθεί.
Αν γνωρίζουµε, άραγε, την ακριβή ακολουθία των αµι-νοξέων ενός πολυπεπτιδίου µπορούµε να βγάλουµε κά-ποιο συµπέρασµα για την τριδιάστατη δοµή (συνήθωςονοµάζεται απλώς «δοµή») ή τη λειτουργία µιας πρωτεΐ-νης; ∆εν πρέπει να ξεχνούµε ότι ο όρος πολυπεπτίδιο δενείναι συνώνυµος µε τον όρο πρωτεΐνη. Ακόµα και στην πε-ρίπτωση που µια πρωτεΐνη αποτελείται από ένα µόνο πο-λυπεπτίδιο, η σχέση ανάµεσα στην ακολουθία των αµινο-ξέων και στη δοµή της (κατ’ επέκταση, και στη λειτουργίατης) µοιάζει εκ πρώτης όψεως µε τη σχέση ανάµεσα σεένα καρούλι µε βαµβακερή κλωστή και σε µια µπλούζα µεσυγκεκριµένο µέγεθος και σχέδιο που µπορεί να φτιαχτείαπό τη συγκεκριµένη κλωστή. Μια λειτουργική πρωτεΐνησυνήθως δεν είναι απλώς µια πολυπεπτιδική αλυσίδα, αλ-λά ένα ή περισσότερα πολυπεπτίδια που µε θαυµαστήακρίβεια αναδιπλώνονται, πτυχώνονται και συστρέφονταιγια να σχηµατίσουν τελικά ένα µόριο µε µοναδική διάτα-ξη στον χώρο (Εικόνα 5.19). Εκείνο, όµως, που σε τελευ-ταία ανάλυση θα καθορίσει την ακριβή τριδιάστατη δοµήµιας πρωτεΐνης είναι η ακολουθία των αµινοξέων σε κάθεπολυπεπτιδική αλυσίδα.
Όταν ένα κύτταρο συνθέτει ένα πολυπεπτίδιο, η αλυ-σίδα των αµινοξέων κατά κανόνα αναδιπλώνεται αυθόρ-µητα και παίρνει τη λειτουργική δοµή που αντιστοιχεί στησυγκεκριµένη πρωτεΐνη. Αυτή η αναδίπλωση καθοδηγείταικαι ενισχύεται από τη δηµιουργία πολλών δεσµών υδρο-γόνου ανάµεσα σε διάφορα τµήµατα της αλυσίδας, κάτιπου µε τη σειρά του εξαρτάται από την ίδια την ακολουθίατων αµινοξέων. Πολλές πρωτεΐνες είναι περίπου σφαιρι-κές (σφαιρίνες), ενώ άλλες έχουν επιµήκη δοµή (ινώδειςπρωτεΐνες). Ωστόσο, ακόµη και στις δύο αυτές χονδρικέςκατηγορίες οι παραλλαγές που υπάρχουν είναι άπειρες.
Η συγκεκριµένη δοµή µιας πρωτεΐνης καθορίζει τοντρόπο λειτουργίας της. Η λειτουργία µιας πρωτεΐνης εξαρ-
(α)
H N
H
C
H
C
O
OH
CH2
N C
H
C
O
CH2
OH
N
HH
C
H
C
O
OH
SH
Πεπτιδικόςδεσμός
CH2
(β)
H N
H
C
H
C
O
OH
CH2
N
H
C
H
C
O
CH2
OH
Αμινοτελικό άκρο(άκρο Ν)
Πεπτιδικόςδεσμός
N
H
C
H
C
O
OH
SH
CH2
Καρβοξυτελικό άκρο(άκρο C)
Πεπτιδικόςσκελετός
HOH
Πλευρικέςαλυσίδες
H2O
Εικόνα 5.18 Σύνθεση πολυπεπτιδικής αλυσίδας. (α) Οιπεπτιδικοί δεσμοί που σχηματίζονται από τις αντιδράσεις αφυδάτωσηςσυνδέουν την καρβοξυλομάδα ενός αμινοξέος με την αμινομάδα τουεπόμενου. (β) Οι πεπτιδικοί δεσμοί σχηματίζονται ένας-ένας, ξεκινώνταςκάθε φορά από το αμινοξύ του αμινοτελικού άκρου (άκρο Ν). Κάθεπολυπεπτίδιο αποτελείται από έναν επαναλαμβανόμενο σκελετό (μοβ)απ’ όπου εξέχουν οι πλευρικές αλυσίδες των αμινοξέων του.
Βάλτε μέσα σε κύκλο και σημειώστε ποιες είναι οικαρβοξυλομάδες και οι αμινομάδες του (α) που σχηματίζουν τουςπεπτιδικούς δεσμούς στο (β).
ΣΧΕΔΙΑΣΤΕ!
τάται, στο σύνολο σχεδόν των περιπτώσεων, από την ικα-νότητά της να αναγνωρίζει και να δεσµεύει κάποιο άλλοµόριο. Ένα ιδιαίτερα εντυπωσιακό παράδειγµα του «γά-µου» ανάµεσα στη δοµή και στη λειτουργία µιας πρωτεΐ-νης παρουσιάζεται στην Εικόνα 5.20, όπου φαίνεται µεπόση ακρίβεια αλληλοσυµπληρώνονται συγκεκριµένατµήµατα της δοµής ενός αντισώµατος (µιας πρωτεΐνης τουοργανισµού) και µιας επίσης συγκεκριµένης πρωτεΐνηςαπό έναν ξένο οργανισµό, τον ιό της γρίπης. Η δέσµευσητου αντισώµατος πάνω στην πρωτεΐνη του ιού είναι το
98 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
πρώτο βήµα για την καταστροφή της. Ένα δεύτερο παρά-δειγµα είναι τα ένζυµα. Ένα ένζυµο πρέπει να αναγνωρίσεικαι να δεσµευθεί στενά µε το υπόστρωµά του, δηλαδή µετην ουσία πάνω στην οποία επενεργεί (βλ. Εικόνα 5.16).Επίσης, στο Κεφάλαιο 2 γνωρίσαµε ότι τα σηµατοδοτικάµόρια που φτιάχνει ο ίδιος ο οργανισµός, και τα οποία ονο-µάζονται ενδορφίνες, δεσµεύονται πάνω σε ειδικούς πρω-τεϊνικούς υποδοχείς στην επιφάνεια των εγκεφαλικών κυτ-τάρων του ανθρώπου, προκαλώντας ευφορία και καταστο-λή του πόνου. Η ικανότητα της µορφίνης, της ηρωίνης καιάλλων οπιούχων φαρµάκων να µιµούνται τις ενδορφίνεςοφείλεται στην οµοιότητα που έχουν όλες αυτές οι ενώσειςµεταξύ τους, κάτι που τους δίνει τη δυνατότητα να ταιριά-ξουν και να δεσµευθούν στους υποδοχείς των ενδορφινώνστον εγκέφαλο. Αυτή η συµπληρωµατικότητα είναι πολύεξειδικευµένη και µοιάζει µε τη συµπληρωµατικότητα µιαςκλειδαριάς µε το κλειδί της (βλ. Εικόνα 2.18). Εποµένως,η λειτουργία µιας πρωτεΐνης –όπως λόγου χάρη η ικανότη-τα ενός πρωτεϊνικού υποδοχέα να αναγνωρίζει και να δε-σµεύει ένα συγκεκριµένο αναλγητικό-σηµατοδοτικό µό-ριο– είναι µια νέα ιδιότητα που οφείλεται στη λεπτοµερήδιάταξη των ατόµων και των µορίων στο µοριακό επίπεδο.
Υπάρχουν τέσσερα επίπεδα στη δομή μιας πρωτεΐνης
Όσο περισσότερο γνωρίζουµε τη δοµή µιας πρωτεΐνης τό-σο περισσότερο κατανοούµε τη λειτουργία της. Παρά τηνπολύ µεγάλη ποικιλοµορφία τους, όλες ανεξαιρέτως οιπρωτεΐνες διαθέτουν τρία διαδοχικά επίπεδα οργάνωσηςστη δοµή τους, τα οποία ονοµάζονται πρωτοταγής δοµή,δευτεροταγής δοµή και τριτοταγής δοµή. Υπάρχει και ένατέταρτο επίπεδο, η τεταρτοταγής δοµή, το οποίο όµως εµ-φανίζεται µόνο σε όσες πρωτεΐνες αποτελούνται από δύο
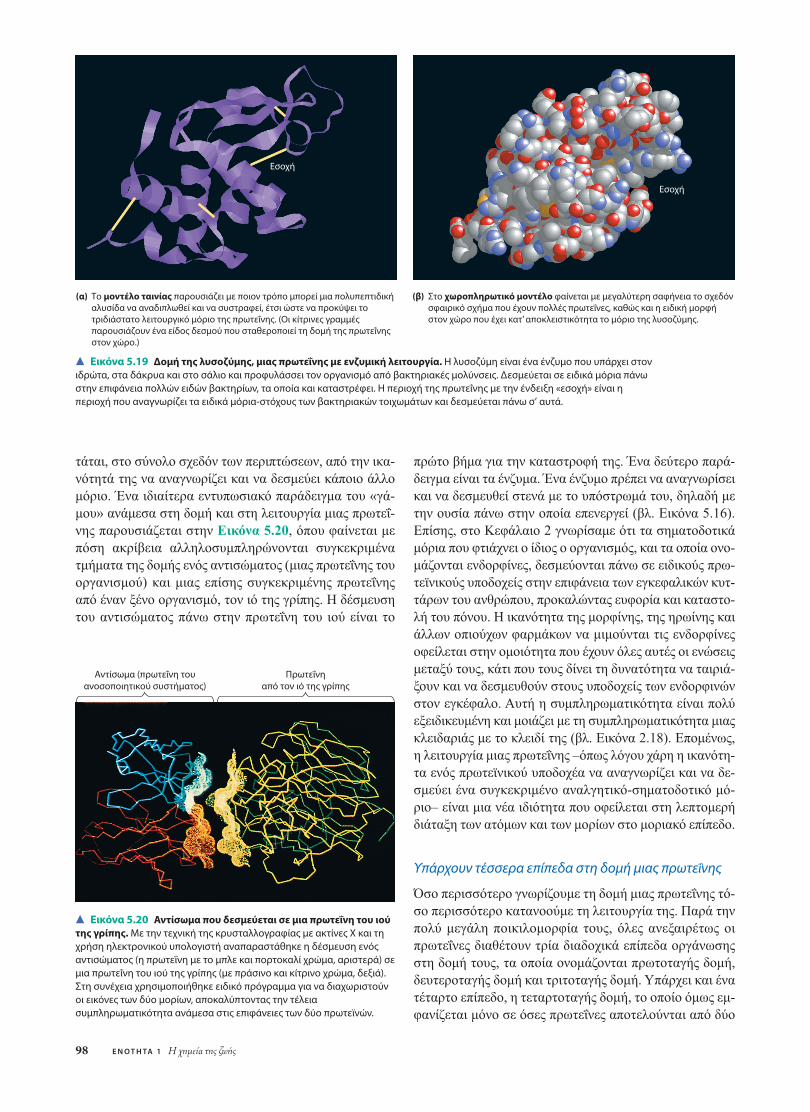
(α) Το μοντέλο ταινίας παρουσιάζει με ποιον τρόπο μπορεί μια πολυπεπτιδικήαλυσίδα να αναδιπλωθεί και να συστραφεί, έτσι ώστε να προκύψει τοτριδιάστατο λειτουργικό μόριο της πρωτεΐνης. (Οι κίτρινες γραμμές παρουσιάζουν ένα είδος δεσμού που σταθεροποιεί τη δομή της πρωτεΐνηςστον χώρο.)
Εσοχή
(β) Στο χωροπληρωτικό μοντέλο φαίνεται με μεγαλύτερη σαφήνεια το σχεδόνσφαιρικό σχήμα που έχουν πολλές πρωτεΐνες, καθώς και η ειδική μορφή στον χώρο που έχει κατ’ αποκλειστικότητα το μόριο της λυσοζύμης.
Εσοχή
Εικόνα 5.19 Δομή της λυσοζύμης, μιας πρωτεΐνης με ενζυμική λειτουργία. Η λυσοζύμη είναι ένα ένζυμο που υπάρχει στονιδρώτα, στα δάκρυα και στο σάλιο και προφυλάσσει τον οργανισμό από βακτηριακές μολύνσεις. Δεσμεύεται σε ειδικά μόρια πάνωστην επιφάνεια πολλών ειδών βακτηρίων, τα οποία και καταστρέφει. Η περιοχή της πρωτεΐνης με την ένδειξη «εσοχή» είναι ηπεριοχή που αναγνωρίζει τα ειδικά μόρια-στόχους των βακτηριακών τοιχωμάτων και δεσμεύεται πάνω σ’ αυτά.
Αντίσωμα (πρωτεΐνη τουανοσοποιητικού συστήματος)
Πρωτεΐνηαπό τον ιό της γρίπης
Εικόνα 5.20 Αντίσωμα που δεσμεύεται σε μια πρωτεΐνη του ιούτης γρίπης. Με την τεχνική της κρυσταλλογραφίας με ακτίνες Χ και τηχρήση ηλεκτρονικού υπολογιστή αναπαραστάθηκε η δέσμευση ενόςαντισώματος (η πρωτεΐνη με το μπλε και πορτοκαλί χρώμα, αριστερά) σεμια πρωτεΐνη του ιού της γρίπης (με πράσινο και κίτρινο χρώμα, δεξιά).Στη συνέχεια χρησιμοποιήθηκε ειδικό πρόγραμμα για να διαχωριστούνοι εικόνες των δύο μορίων, αποκαλύπτοντας την τέλειασυμπληρωματικότητα ανάμεσα στις επιφάνειες των δύο πρωτεϊνών.

ή περισσότερες πολυπεπτιδικές αλυσίδες. Τα τέσσερα επί-πεδα οργάνωσης της δοµής µιας πρωτεΐνης περιγράφονταιστην Εικόνα 5.21, στις δύο επόµενες σελίδες. Πριν συνε-χίσουµε είναι απαραίτητο να τη µελετήσετε προσεκτικά.
Δρεπανοκυτταρική αναιμία: Ένα λάθος στην πρωτοταγήδομή
Ακόµα και µια µικρή αλλαγή στην πρωτοταγή δοµή µιαςπρωτεΐνης µπορεί να επηρεάσει το σχήµα και τη λειτουρ-γικότητά της. Λόγου χάρη, η δρεπανοκυτταρική αναιµία,µια κληρονοµική νόσος του αίµατος, οφείλεται στηνεσφαλµένη αντικατάσταση ενός αµινοξέος στην αµινοξι-κή ακολουθία της αιµοσφαιρίνης, της πρωτεΐνης των ερυ-θρών αιµοσφαιρίων που µεταφέρει το οξυγόνο. Έτσι, αν-τί του σωστού αµινοξέος (γλουταµικό οξύ) σε µια συγκε-κριµένη θέση της αµινοξικής ακολουθίας υπάρχει λάθοςαµινοξύ (βαλίνη). Τα φυσιολογικά ερυθρά αιµοσφαίριαέχουν σχήµα δίσκου, αλλά στη δρεπανοκυτταρική αναι-µία τα µη φυσιολογικά µόρια της αιµοσφαιρίνης έχουν τηντάση να κρυσταλλώνονται, προκαλώντας την παραµόρ-
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 99
φωση µερικών ερυθροκυττάρων, που αποκτούν σχήµαδρεπάνου (Εικόνα 5.22). Η ζωή ενός ανθρώπου µε δρε-πανοκυτταρική αναιµία καθορίζεται από «δρεπανοκυττα-ρικές κρίσεις», που εκδηλώνονται όταν αυτά τα δρεπανο-κύτταρα φράσσουν µικρά αγγεία του αίµατος, εµποδίζον-τας τη ροή του. Το τίµηµα που πληρώνουν οι ασθενείς εί-ναι ένα δραµατικό παράδειγµα του πώς µια µικρή αλλαγήστη δοµή µιας πρωτεΐνης µπορεί να έχει βαρύτατες συνέ-πειες στη λειτουργία της και, κατ’ επέκταση, στη λειτουρ-γία ολόκληρου του οργανισµού.
Tι καθορίζει τη δομή μιας πρωτεΐνης;
Μάθαµε ότι η µοναδικότητα του τριδιάστατου σχήµατοςπροσδίδει σε κάθε πρωτεΐνη συγκεκριµένη λειτουργία.Ποιοι είναι όµως οι κοµβικοί εκείνοι παράγοντες που κα-θορίζουν τη δοµή µιας πρωτεΐνης; Ήδη γνωρίζουµε το µε-γαλύτερο τµήµα της απάντησης. Η πολυπεπτιδική αλυσί-δα δεδοµένης ακολουθίας αµινοξέων διατάσσεται αυθόρ-µητα στον χώρο. Το τριδιάστατο σχήµα ορίζεται και δια-τηρείται από τις ίδιες αλληλεπιδράσεις που ευθύνονται για
Val1
His2
Leu3
Thr4
Pro5
Glu6
Glu7
Val1
His2
Leu3
Thr4
Pro5
Val66
Glu7
Πρωτοταγήςδομή
Δευτεροταγήςκαι τριτοταγήςδομή
Τεταρτοταγήςδομή
Λειτουργία
Φυσιολογικήαιμοσφαιρίνη(κάτοψη)
Τα μόρια δεν αλληλεπιδρούνμεταξύ τους. το καθέναμεταφέρει οξυγόνοανεξάρτητα απότα άλλα.
Τα φυσιολογικά κύτταραείναι γεμάτα απόανεξάρτητα μόριααιμοσφαιρίνης, που τοκαθένα μεταφέρει οξυγόνο.
υπομονάδα β υπομονάδα β
Δρεπανοκυτταρικήαιμοσφαιρίνη
Εκτεθειμένεςυδρόφοβεςπεριοχές
Τα μόρια αλληλεπιδρούνμεταξύ τους και συσσωματώνονται σχηματίζονταςνημάτια, οπότε μειώνεται σημαντικά η ικανότητα μεταφοράςοξυγόνου.
Τα νημάτια της παθολογικήςαιμοσφαιρίνης παραμορφώνουν το ερυθρόαιμοσφαίριο, προσδίδοντάςτου δρεπανοειδές σχήμα.
αa
α
aβ
β
ββ
Σχήμαερυθρούαιμοσφαιρίου
Πρωτοταγήςδομή
Φυσιολογική αιμοσφαιρίνη Δρεπανοκυτταρική αιμοσφαιρίνη
Δευτεροταγήςκαι τριτοταγήςδομή
Τεταρτοταγήςδομή
Λειτουργία
Σχήμαερυθρούαιμοσφαιρίου
10 μm 10 μm
Εικόνα 5.22 Η δρεπανοκυτταρική αναιμία προκαλείται από την αντικατάσταση ενός μόνον αμινοξέος της αιμοσφαιρίνης. Εδώ, οπροσανατολισμός του μορίου της αιμοσφαιρίνης είναι διαφορετικός από εκείνον της Εικόνας 5.21, προκειμένου να φανεί καθαρά ο σχηματισμός τωνπαθολογικών ινιδίων.

C
Καρβοξυτελικό άκροO–
O
Ser
Ser
SerGly Pro
ArgArg
Tyr
TyrTyrSerThr
Thr
Ala
ValVal
Thr Asn ProLys Glu
Ser
ThrIIe
Pro Phe HisHis
Ala
GluVal
ValPheThrAla
Ala
Ala
Leu
LeuSer
Pro
Asn
Asp
Glu
LysAlaLeuGly
IIe
TyrTrp
80
75
85
100105
110
115
120
125
90
95
LysThr
AspIIeGlu
Gly
Gly
GlySer
GluSerLys
Lys
CysPro
Pro
Leu
LeuAsp
Ala
Ala
MetVal
Val Arg
Val
Pro ThrGly
Thr
1
5
10
20
25
15
NN
O
H
CC C C
C NH
NO
HC
C
OC C N
O
HC
CC C N
O
HC
CC C N
O
HC
CC
CHHN
O
HN
O
H
H
N
O
C
ON
O
H
CCNC
H
C
O
N
O
H
CCNC
HR
C
O
N
O
H
CCNC
H
C
O
H HH
+H3NΑμινοτελικό άκρο
Παραδείγματαπεπτιδικών
υπομονάδων
πτυχωτή επιφάνεια β
έλικα αN
C
CC
C
C
C
C
O
H
C C
OO
O
N H
N HH
NC
C R RC
C
C
C
C
OH
OO
O
N H
N H N H
N HHH R H R
HR
HR
HR HR
RRRH
HR
H
R
H
R
R
R
H
R
H
R
H
R
H
R
H
R
H
Εικόνα 5.21
Εξερευνώντας τα επίπεδα της πρωτεϊνικής δομής
Οι σπειροειδείς συλληπτήριες ίνες είναι ελαστικές και υφίστανται
διάταση με το φύσημα του ανέμου,τη βροχή ή την επαφή με έντομοπου παγιδεύτηκε στο εσωτερικό
του ιστού.
Ο ιστός της αράχνης εκκρίνεταιαπό ειδικούς αδένες. Οι μεταξωτής
υφής ίνες του αποτελούνται από μια δομική πρωτεΐνη την οποία
δομούν πτυχωτά φύλλα β.
Οι ακτινωτές ίνες του ιστού είναιξηρές, μεταξωτές και ευθύνονται
για τη διατήρηση του σχήματός του.
Πρωτοταγής δομή Δευτεροταγής δομή
100 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
Πρωτοταγής δοµή µιας πρωτεΐνης είναι η ακολουθία των αµινο-ξέων που την αποτελούν, η οποία είναι µοναδική για κάθε πρωτεΐ-νη. Ας εξετάσουµε επί παραδείγµατι την πρωτεΐνη τρανσθυρετίνη,µια σφαιρίνη που υπάρχει στο αίµα και µεταφέρει τη βιταµίνη Α,αλλά και µία από τις θυρεοειδείς ορµόνες. Η τρανσθυρετίνη αποτε-λείται από τέσσερεις πολυπεπτιδικές αλυσίδες, που καθεµιά συνί-σταται από 127 αµινοξέα. Εδώ παρουσιάζεται ξεδιπλωµένη µία απότις τέσσερεις αλυσίδες, για να φανεί καλύτερα η πρωτοταγής δοµήτης. Κάθε θέση της αλυσίδας των 127 αµινοξέων καταλαµβάνεταιαπό συγκεκριµένο αµινοξύ (ένα από τα 20 συνολικά) και συµβολί-ζεται συντετµηµένα µε τρία γράµµατα. Η πρωτοταγής δοµή είναιόπως η σειρά των γραµµάτων σε µια πολύ µακριά λέξη. Αν η σειράτων αµινοξέων ήταν τυχαία, τότε θα υπήρχαν 20127 διαφορετικοίτρόποι για να φτιαχτεί µια πεπτιδική αλυσίδα µήκους 127 αµινοξέ-
Ορισµένα τµήµατα στην πολυπεπτιδική αλυσίδα των περισσότερωνπρωτεϊνών σχηµατίζουν επαναλαµβανόµενες πτυχές ή έλικες, οι οποί-ες διαµορφώνουν σε µεγάλο βαθµό την τελική µορφή αυτών των πρω-τεϊνών στον χώρο. Όλες οι πτυχώσεις και οι ελικοειδείς δοµές ονοµά-ζονται συνολικά δευτεροταγής δοµή και οφείλονται στους δεσµούςυδρογόνου που αναπτύσσονται ανάµεσα στα επαναλαµβανόµενα συ-στατικά του πολυπεπτιδικού σκελετού (όχι ανάµεσα στις πλευρικέςαλυσίδες των αµινοξέων). Τα άτοµα του οξυγόνου και του αζώτου στονπολυπεπτιδικό σκελετό είναι ηλεκτραρνητικά, εποµένως φέρουν µερι-κό αρνητικό φορτίο (βλ. Εικόνα 2.16). Τα ελαφρώς θετικά φορτισµέναάτοµα του υδρογόνου που συνδέονται στο άτοµο του αζώτου έλκονταιαπό το άτοµο του οξυγόνου του γειτονικού πεπτιδικού δεσµού και σχη-µατίζουν δεσµό υδρογόνου µε αυτό. Κάθε µεµονωµένος δεσµός υδρο-γόνου είναι πολύ ασθενής, αλλά επειδή σχηµατίζονται πολλοί τέτοιοιδεσµοί σε σχετικά µεγάλο τµήµα της πολυπεπτιδικής αλυσίδας, η συ-νολική τους ισχύς είναι αρκετή για να υποστηρίξει την όποια µορφήαποκτά το συγκεκριµένο τµήµα της πρωτεΐνης.
Παράδειγµα δευτεροταγούς δοµής είναι η έλικα α, µια λεπτή σπει-ροειδής δοµή που συγκρατείται από δεσµούς υδρογόνου ανάµεσα σεκάθε τέταρτο αµινοξύ (βλ. αντίστοιχο σχήµα, πάνω). Αν και κάθε υπο-µονάδα της τρανσθυρετίνης διαθέτει µία µόνο έλικα (βλ. εικόνα τριτο-ταγούς δοµής), υπάρχουν πολλές άλλες σφαιρίνες που διαθέτουν αρ-κετά τµήµατα µε διαµόρφωση έλικας α, ανάµεσα στα οποία παρεµ-βάλλονται µη ελικοειδείς περιοχές. Ορισµένες ινώδεις πρωτεΐνες, όπωςη κερατίνη α, από την οποία αποτελούνται οι τρίχες, εµφανίζουν τη δια-µόρφωση της έλικας α στο µεγαλύτερο τµήµα του µορίου τους.
Ο άλλος κύριος τύπος δευτεροταγούς δοµής είναι η πτυχωτή επι-φάνεια β ή πτυχή β. Όπως φαίνεται στο αντίστοιχο σχήµα, στην κο-ρυφή της στήλης αυτής, δύο ή περισσότερες γειτονικές περιοχές τηςπολυπεπτιδικής αλυσίδας συνδέονται µέσω δεσµών υδρογόνου µε πα-ράλληλα τµήµατα του ίδιου πολυπεπτιδικού σκελετού. Τα πτυχωτάφύλλα συνιστούν τον πυρήνα πολλών σφαιρινών, όπως π.χ. της τραν-σθυρετίνης, ενώ είναι και η κυρίαρχη διαµόρφωση σε ορισµένες ινώ-δεις πρωτεΐνες, όπως στις µεταξωτές ίνες που αποτελούν τον ιστό τηςαράχνης. Η συλλογική δύναµη τόσο πολλών δεσµών υδρογόνου καθι-στά κάθε µεταξωτή ίνα στον ιστό της αράχνης ισχυρότερη και από µιαίνα από σίδηρο του ίδιου βάρους.
ων. Η ακριβής πρωτοταγής δοµή µιας πρωτεΐνης, ωστόσο, δεν κα-θορίζεται από την τυχαία σύνδεση των αµινοξέων µεταξύ τους, αλ-λά από τη γενετική πληροφορία που κληρονοµεί κάθε οργανισµόςαπό τους γονείς του.
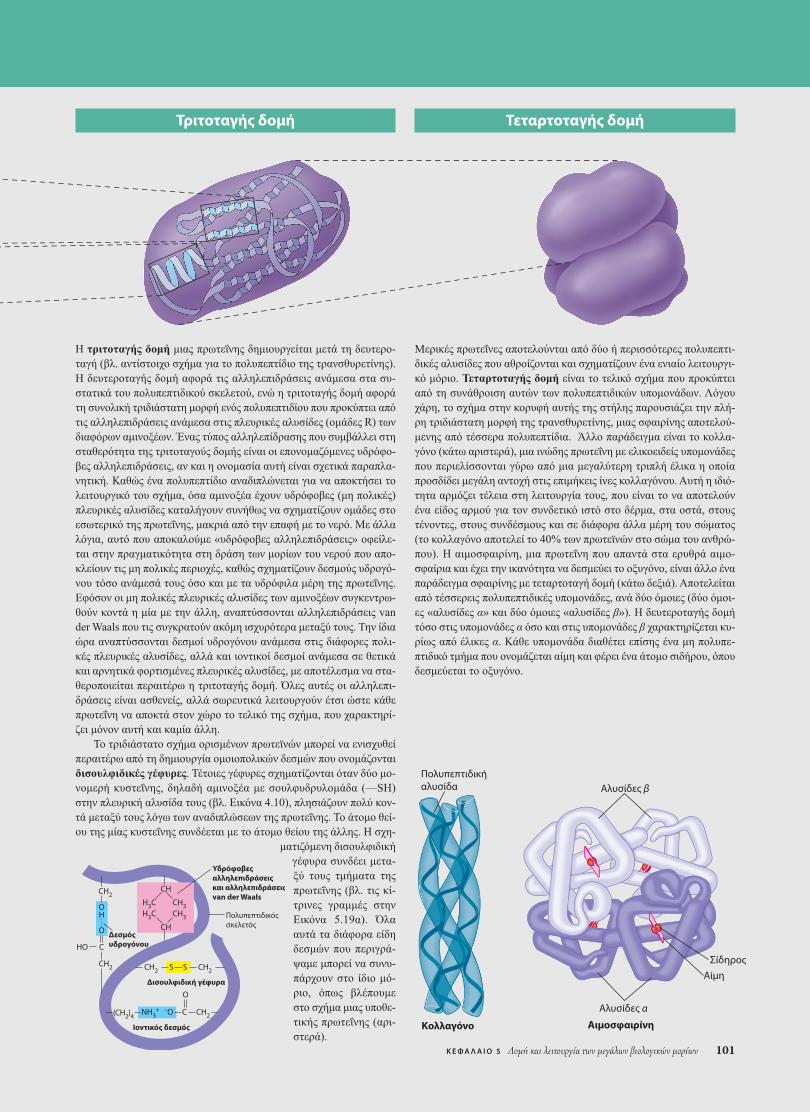
Πολυπεπτιδικήαλυσίδα
Κολλαγόνο
Τριτοταγής δομή Τεταρτοταγής δομή
Αιμοσφαιρίνη
Αλυσίδες β
Αλυσίδες α
ΑίμηΣίδηρος
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 101
Η τριτοταγής δοµή µιας πρωτεΐνης δηµιουργείται µετά τη δευτερο-ταγή (βλ. αντίστοιχο σχήµα για το πολυπεπτίδιο της τρανσθυρετίνης).Η δευτεροταγής δοµή αφορά τις αλληλεπιδράσεις ανάµεσα στα συ-στατικά του πολυπεπτιδικού σκελετού, ενώ η τριτοταγής δοµή αφοράτη συνολική τριδιάστατη µορφή ενός πολυπεπτιδίου που προκύπτει απότις αλληλεπιδράσεις ανάµεσα στις πλευρικές αλυσίδες (οµάδες R) τωνδιαφόρων αµινοξέων. Ένας τύπος αλληλεπίδρασης που συµβάλλει στησταθερότητα της τριτοταγούς δοµής είναι οι επονοµαζόµενες υδρόφο-βες αλληλεπιδράσεις, αν και η ονοµασία αυτή είναι σχετικά παραπλα-νητική. Καθώς ένα πολυπεπτίδιο αναδιπλώνεται για να αποκτήσει τολειτουργικό του σχήµα, όσα αµινοξέα έχουν υδρόφοβες (µη πολικές)πλευρικές αλυσίδες καταλήγουν συνήθως να σχηµατίζουν οµάδες στοεσωτερικό της πρωτεΐνης, µακριά από την επαφή µε το νερό. Με άλλαλόγια, αυτό που αποκαλούµε «υδρόφοβες αλληλεπιδράσεις» οφείλε-ται στην πραγµατικότητα στη δράση των µορίων του νερού που απο-κλείουν τις µη πολικές περιοχές, καθώς σχηµατίζουν δεσµούς υδρογό-νου τόσο ανάµεσά τους όσο και µε τα υδρόφιλα µέρη της πρωτεΐνης.Εφόσον οι µη πολικές πλευρικές αλυσίδες των αµινοξέων συγκεντρω-θούν κοντά η µία µε την άλλη, αναπτύσσονται αλληλεπιδράσεις vander Waals που τις συγκρατούν ακόµη ισχυρότερα µεταξύ τους. Την ίδιαώρα αναπτύσσονται δεσµοί υδρογόνου ανάµεσα στις διάφορες πολι-κές πλευρικές αλυσίδες, αλλά και ιοντικοί δεσµοί ανάµεσα σε θετικάκαι αρνητικά φορτισµένες πλευρικές αλυσίδες, µε αποτέλεσµα να στα-θεροποιείται περαιτέρω η τριτοταγής δοµή. Όλες αυτές οι αλληλεπι-δράσεις είναι ασθενείς, αλλά σωρευτικά λειτουργούν έτσι ώστε κάθεπρωτεΐνη να αποκτά στον χώρο το τελικό της σχήµα, που χαρακτηρί-ζει µόνον αυτή και καµία άλλη.
Το τριδιάστατο σχήµα ορισµένων πρωτεϊνών µπορεί να ενισχυθείπεραιτέρω από τη δηµιουργία οµοιοπολικών δεσµών που ονοµάζονταιδισουλφιδικές γέφυρες. Τέτοιες γέφυρες σχηµατίζονται όταν δύο µο-νοµερή κυστεΐνης, δηλαδή αµινοξέα µε σουλφυδρυλοµάδα (—SH)στην πλευρική αλυσίδα τους (βλ. Εικόνα 4.10), πλησιάζουν πολύ κον-τά µεταξύ τους λόγω των αναδιπλώσεων της πρωτεΐνης. Το άτοµο θεί-ου της µίας κυστεΐνης συνδέεται µε το άτοµο θείου της άλλης. Η σχη-
µατιζόµενη δισουλφιδικήγέφυρα συνδέει µετα-ξύ τους τµήµατα τηςπρωτεΐνης (βλ. τις κί-τρινες γραµµές στηνΕικόνα 5.19α). Όλααυτά τα διάφορα είδηδεσµών που περιγρά-ψαµε µπορεί να συνυ-πάρχουν στο ίδιο µό-ριο, όπως βλέπουµεστο σχήµα µιας υποθε-τικής πρωτεΐνης (αρι-στερά).
Μερικές πρωτεΐνες αποτελούνται από δύο ή περισσότερες πολυπεπτι-δικές αλυσίδες που αθροίζονται και σχηµατίζουν ένα ενιαίο λειτουργι-κό µόριο. Τεταρτοταγής δοµή είναι το τελικό σχήµα που προκύπτειαπό τη συνάθροιση αυτών των πολυπεπτιδικών υποµονάδων. Λόγουχάρη, το σχήµα στην κορυφή αυτής της στήλης παρουσιάζει την πλή-ρη τριδιάστατη µορφή της τρανσθυρετίνης, µιας σφαιρίνης αποτελού-µενης από τέσσερα πολυπεπτίδια. Άλλο παράδειγµα είναι το κολλα-γόνο (κάτω αριστερά), µια ινώδης πρωτεΐνη µε ελικοειδείς υποµονάδεςπου περιελίσσονται γύρω από µια µεγαλύτερη τριπλή έλικα η οποίαπροσδίδει µεγάλη αντοχή στις επιµήκεις ίνες κολλαγόνου. Αυτή η ιδιό-τητα αρµόζει τέλεια στη λειτουργία τους, που είναι το να αποτελούνένα είδος αρµού για τον συνδετικό ιστό στο δέρµα, στα οστά, στουςτένοντες, στους συνδέσµους και σε διάφορα άλλα µέρη του σώµατος(το κολλαγόνο αποτελεί το 40% των πρωτεϊνών στο σώµα του ανθρώ-που). Η αιµοσφαιρίνη, µια πρωτεΐνη που απαντά στα ερυθρά αιµο-σφαίρια και έχει την ικανότητα να δεσµεύει το οξυγόνο, είναι άλλο έναπαράδειγµα σφαιρίνης µε τεταρτοταγή δοµή (κάτω δεξιά). Αποτελείταιαπό τέσσερεις πολυπεπτιδικές υποµονάδες, ανά δύο όµοιες (δύο όµοι-ες «αλυσίδες α» και δύο όµοιες «αλυσίδες β»). Η δευτεροταγής δοµήτόσο στις υποµονάδες α όσο και στις υποµονάδες β χαρακτηρίζεται κυ-ρίως από έλικες α. Κάθε υποµονάδα διαθέτει επίσης ένα µη πολυπε-πτιδικό τµήµα που ονοµάζεται αίµη και φέρει ένα άτοµο σιδήρου, όπουδεσµεύεται το οξυγόνο.
(CH2)4 NH3+ –O C CH2
H3C CH3
CH
H3C CH3
CHH
O
CH2
O
O
CH2 S S CH2
C
CH2
HOΔεσμόςυδρογόνου
Δισουλφιδική γέφυρα
Υδρόφοβεςαλληλεπιδράσειςκαι αλληλεπιδράσειςvan der Waals
Πολυπεπτιδικόςσκελετός
Ιοντικός δεσμός
τη δευτεροταγή και τριτοταγή δοµή. Αυτές οι αναδιπλώ-σεις συµβαίνουν ήδη από την πρώτη στιγµή που αρχίζεινα συντίθεται η ακολουθία των αµινοξέων µέσα στο κύτ-ταρο. Η δοµή µιας πρωτεΐνης, ωστόσο, εξαρτάται και απότις φυσικές και χηµικές συνθήκες που επικρατούν στοκυτταρικό περιβάλλον. Αν µεταβληθούν το pH, η συγκέν-τρωση των αλάτων, η θερµοκρασία ή διάφορες άλλες πα-ράµετροι του άµεσου περιβάλλοντος, η πρωτεΐνη µπορείνα αποδιαταχθεί και να χάσει το φυσιολογικό σχήµα της.Μια τέτοια αλλαγή ονοµάζεται αποδιάταξη ή µετουσίω-ση (Εικόνα 5.23). Επειδή οι αποδιατεταγµένες πρωτεΐνεςχάνουν το φυσιολογικό σχήµα τους, χάνουν και τη βιολο-γική τους δραστικότητα.
Οι περισσότερες πρωτεΐνες αποδιατάσσονται όταν µε-ταφερθούν από ένα υδατικό διάλυµα σε κάποιο άλλο όπουδιαλύτης είναι µια οργανική ουσία, π.χ. ο αιθέρας ή τοχλωροφόρµιο. Σε µια τέτοια περίπτωση, η πολυπεπτιδικήαλυσίδα διατάσσει εκ νέου τις αναδιπλώσεις της, έτσι
102 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
ώστε οι υδρόφοβες περιοχές της να στραφούν προς το εξω-τερικό, δηλαδή προς την πλευρά του διαλύτη. ∆ιάφοροιάλλοι χηµικοί παράγοντες αποδιάταξης εµποδίζουν τονσχηµατισµό των δεσµών υδρογόνου, των ιοντικών δεσµώνκαι των δισουλφιδικών γεφυρών, µε άλλα λόγια εκείνωνακριβώς των δεσµών που διατηρούν σταθερό το σχήµαµιας πρωτεΐνης. Αποδιάταξη µπορεί επίσης να συµβεί καιαπό υπερβολική θέρµανση, όταν οι αναταράξεις που προ-καλούνται στην πολυπεπτιδική αλυσίδα υπερβούν την ισχύτων ασθενών αλληλεπιδράσεων οι οποίες σταθεροποιούντη φυσιολογική δοµή της. Το ασπράδι του αυγού γίνεταιαδιαφανές όταν µαγειρευτεί, επειδή οι αποδιατεταγµένεςπρωτεΐνες είναι αδιάλυτες και στερεοποιούνται. Αυτό εξη-γεί επίσης γιατί µπορεί να αποβεί µοιραίος ένας υπερβο-λικά υψηλός πυρετός, αφού ενδέχεται να προκαλέσει απο-διάταξη των πρωτεϊνών του αίµατος.
Ορισµένες φορές, η αποδιάταξη µιας πρωτεΐνης δεν εί-ναι µόνιµη. Αν προκλήθηκε υπό την επίδραση της θερµό-τητας ή κάποιου χηµικού παράγοντα, τότε είναι δυνατόν ηπρωτεΐνη να ανακτήσει το λειτουργικό σχήµα της. Από τογεγονός αυτό µπορούµε να συµπεράνουµε ότι η πληροφο-ρία για τη συγκεκριµένη και ιδιαίτερη δοµή µιας πρωτεΐ-νης στον χώρο βρίσκεται ενσωµατωµένη στην πρωτοτα-γή της δοµή. Η ακολουθία των αµινοξέων της πρωτεΐνηςκαθορίζει το σχήµα της, δηλαδή το πού ακριβώς θα σχη-µατιστεί µια έλικα α και πού οι πτυχωτές επιφάνειες β, σεποιο σηµείο θα δηµιουργηθούν δισουλφιδικές γέφυρες,πού µπορούν να σχηµατιστούν ιοντικοί δεσµοί κ.ο.κ. Μέ-σα στην πληθώρα µορίων του ενδοκυττάριου περιβάλ-λοντος υπάρχουν και ορισµένες πρωτεΐνες που βοηθούνστην αναδίπλωση των άλλων πρωτεϊνών.
Αναδίπλωση πρωτεϊνών μέσα στο κύτταρο
Σήµερα γνωρίζουµε την ακολουθία αµινοξέων σε τουλάχι-στον 1,2 εκατοµµύρια πρωτεϊνικά µόρια, αλλά και την πλή-
Φυσιολογικήπρωτεΐνη
Αποδιατεταγμένη(μετουσιωμένη) πρωτεΐνη
Αποδιάταξη
Επαναδιάταξη
Εικόνα 5.23 Αποδιάταξη και επαναδιάταξη πρωτεΐνης. Οιυψηλές θερμοκρασίες ή διάφοροι χημικοί παράγοντες προκαλούναποδιάταξη μιας πρωτεΐνης, δηλαδή μεταβάλλουν τη διαμόρφωσή τηςστον χώρο, με αποτέλεσμα την απώλεια της λειτουργίας της. Αν ηαποδιατεταγμένη πρωτεΐνη παραμείνει σε διαλυμένη μορφή, τότε ηεπαναφορά των χημικών και φυσικών παραμέτρων σε φυσιολογικέςτιμές μπορεί συχνά να προκαλέσει επαναδιάταξη της πρωτεΐνης στηναρχική λειτουργική διαμόρφωσή της.
Πολυπεπτίδιο
Στάδια λειτουργίας μιαςσαπερονίνης:
Πρωτεΐνημε ορθήαναδίπλωση
Κούφιοςκύλινδρος
Σαπερονίνη(πλήρως
συναρμολογημένη)
Κάλυμμα
Ένα μη αναδιπλωμένο πολυπεπτίδιο εισέρχεται στον κύλινδρο από το ένα άκρο.
1
Το κάλυμμα προσδένεται στον κύλινδρο και μεταβάλλειέτσι το σχήμα του ώστε να δημιουργείται υδρόφοβο περιβάλλον στο εσωτερικό του, όπου θα συμβεί η αναδίπλωσητου πολυπεπτιδίου.
2 Το κάλυμμα απομακρύνεταικαι η πρωτεΐνη απελευθερώνεταιέχοντας αποκτήσει τη σωστή στερεοδιαμόρφωση.
3
Εικόνα 5.24 Μια σαπερονίνη εν δράσει. Το διάγραμμα αριστερά έχει προκύψει από ηλεκτρονικό υπολογιστή και αναπαριστάένα μεγάλο σύμπλοκο σαπερονίνης. Το σύμπλοκο περιέχει στο εσωτερικό του έναν κενό χώρο που προστατεύει τιςνεοσχηματισμένες πρωτεΐνες, επιτρέποντάς τους να αναδιπλωθούν απρόσκοπτα ώστε να αποκτήσουν την ορθήστερεοδιαμόρφωση. Ένα σύμπλοκο σαπερονίνης αποτελείται από δύο πρωτεΐνες. Η μία σχηματίζει έναν κύλινδρο κενό στοεσωτερικό του, ενώ η άλλη σχηματίζει ένα κάλυμμα που μπορεί να εφαρμόζει σε οποιοδήποτε άκρο του κυλίνδρου.

ρη τριδιάστατη δοµή 8.500 περίπου πρωτεϊνών. Θα έλεγεκανείς ότι αν συσχετίσουµε την πρωτοταγή δοµή µιας πρω-τεΐνης µε το σχήµα της στον χώρο, θα µπορούσαµε να οδη-γηθούµε σχετικά εύκολα στους κανόνες µε τους οποίουςαναδιπλώνεται. ∆υστυχώς, η διαδικασία αναδίπλωσης µιαςπρωτεΐνης δεν είναι τόσο απλή. Οι περισσότερες πρωτεΐνεςπερνούν από διάφορα ενδιάµεσα στάδια πριν αποκτήσουντο τελικό σχήµα τους. Αν γνωρίζουµε µόνο την τελική (στα-θερή) µορφή τους, δεν µπορούµε να συναγάγουµε όλα ταενδιάµεσα στάδια που είναι απαραίτητα για να αποκτηθεί ησυγκεκριµένη µορφή. Οι βιοχηµικοί κατάφεραν ωστόσο ναβρουν µεθόδους για να παρακολουθούν την πορεία µιαςπρωτεΐνης µέσα στα διάφορα στάδια ωρίµανσής της, µέχριδηλαδή να αποκτήσει το τελικό τριδιάστατο σχήµα της στονχώρο. Επίσης, ανακαλύφθηκαν οι σαπερονίνες (ή συνοδοίπρωτεΐνες), ειδικές πρωτεΐνες που βοηθούν στην ορθή ανα-δίπλωση όσων πρωτεϊνών βρίσκονται στο στάδιο της ωρί-µανσης (Εικόνα 5.24). Οι σαπερονίνες δεν καθορίζουν ποιοθα είναι το τελικό σχήµα µιας πρωτεΐνης. Αυτό που κάνουνείναι να προφυλάσσουν το νεοσχηµατισµένα µόρια από τις«κακές επιρροές» την ώρα που αναδιπλώνονται αυθόρµη-τα στο κυτταρόπλασµα. Η σαπερονίνη της Εικόνας 5.24βρέθηκε στο βακτήριο E. coli· είναι ένα γιγάντιο σύµπλοκοπολλών πρωτεΐνών που µοιάζει µε κούφιο κύλινδρο. Η κοι-λότητα στο εσωτερικό του κυλίνδρου λειτουργεί ως «κα-ταφύγιο», όπου µπορούν τα νεοσχηµατιζόµενα πολυπεπτί-δια να ολοκληρώσουν την αναδίπλωσή τους.
Τα σφάλµατα κατά τις πρωτεϊνικές αναδιπλώσεις συ-νιστούν σοβαρό πρόβληµα για τα κύτταρα. Πολλές ασθέ-νειες, όπως η νόσος Alzheimer και η νόσος Parkinson,σχετίζονται µε τη συσσώρευση πρωτεϊνών µε σφάλµαταστις αναδιπλώσεις τους. Πρόσφατα ανακαλύφθηκαν µο-ριακά συστήµατα στο εσωτερικό του κυττάρου που αλλη-λεπιδρούν µε τις σαπερονίνες και ελέγχουν την ορθότητατων αναδιπλώσεων της πρωτεΐνης. Αυτά τα συστήµατα εί-τε αναδιπλώνουν εκ νέου όσες πρωτεΐνες έχουν σφάλµα-τα είτε τις µαρκάρουν για να καταστραφούν.
Ακόµη κι όταν οι επιστήµονες διαθέτουν µια ορθάαναδιπλωµένη πρωτεΐνη, ο προσδιορισµός της τριδιάστα-της δοµής της στον χώρο δεν είναι εύκολη υπόθεση, αφούκαι ένα µόνο πρωτεϊνικό µόριο αποτελείται από χιλιάδεςάτοµα. Ο πρώτος προσδιορισµός τριδιάστατης δοµής έγι-νε το 1959· αφορούσε την αιµοσφαιρίνη και µια συγγενι-κή της πρωτεΐνη. Η µέθοδος που επέτρεψε την επίτευξηαυτού του άθλου ήταν η κρυσταλλογραφία µε ακτίνεςΧ, που έκτοτε έχει χρησιµοποιηθεί για τον προσδιορισµότης τριδιάστατης δοµής πολλών άλλων πρωτεϊνών. Σε έναπρόσφατο παράδειγµα, ο Roger Kornberg και οι συνερ-γάτες του στο Πανεπιστήµιο Stanford χρησιµοποίησαν αυ-τή τη µέθοδο για να αποκρυπτογραφήσουν τη δοµή τηςπολυµεράσης του RNA, ενζύµου που παίζει κοµβικό ρό-λο στην έκφραση των γονιδίων (Εικόνα 5.25). Μια άλληµέθοδος που χρησιµοποιείται σήµερα για τον ίδιο σκοπόείναι η φασµατοσκοπία πυρηνικού µαγνητικού συντονι-σµού (NMR), στην οποία δεν απαιτείται η κρυστάλλωσητων υπό µελέτη πρωτεϊνών.
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 103
Περίθλαση τωνακτίνων Χ
Ψηφιακός ανιχνευτής του προτύπουπερίθλασης ακτίνων Χ
Κρύσταλλος
Δέσμηακτίνων Χ
Πηγήακτίνων X
Εικόνα 5.25 Διερεύνηση
Τι μπορεί να μας πει το τριδιάστατο σχήματου ενζύμου πολυμεράση ΙΙ του RNA όσοναφορά τη λειτουργία του;
Το 2006 απονεμήθηκε το βραβείο NobelΧημείας στον Roger Kornberg για τη χρήση τηςκρυσταλλογραφίας με ακτίνες Χ στον προσδιορισμό τουτριδιάστατου σχήματος της πολυμεράσης ΙΙ του RNA, τουενζύμου που συνδέεται στη διπλή έλικα του DNA και συνθέτειαλυσίδες RNA. Ο Kornberg και οι συνεργάτες του δημιούργησανέναν κρύσταλλο του συμπλόκου με τα τρία αυτά συστατικά(DNA, πολυμεράση του RNA και RNA) και κατόπιν στόχευσαν μιαδέσμη ακτίνων Χ πάνω σ’ αυτόν. Τα άτομα του κρυστάλλουπροκάλεσαν περίθλαση (κάμψη) των ακτίνων Χ με διατεταγμένοτρόπο, ο οποίος καταγράφηκε από ψηφιακό ανιχνευτή ωςδιάταξη σημειακών ιχνών που ονομάζεται πρότυπο περίθλασηςακτίνων Χ.
Χρησιμοποιώντας δεδομένα από τα πρότυπαπερίθλασης ακτίνων Χ και την ακολουθία των αμινοξέων τηςπολυμεράσης ΙΙ του RNA (που προσδιορίστηκαν με χημικέςμεθόδους), ο Kornberg και οι συνεργάτες του κατασκεύασαν ένατριδιάστατο μοντέλο του συμπλόκου με τη βοήθεια ηλεκτρονικούυπολογιστή και ειδικού λογισμικού.
Αναλύοντας το μοντέλο τους, οι ερευνητέςδιατύπωσαν υποθέσεις σχετικά με τη λειτουργία των διαφόρωνπεριοχών της πολυμεράσης ΙΙ του RNA. Λόγου χάρη, υπέθεσαν ότιη περιοχή πάνω από το DNA λειτουργεί ως λαβίδα που συγκρατείτα νουκλεϊκά οξέα στη θέση τους. (Θα μάθουμε περισσότερα γιατο ένζυμο αυτό στο Κεφάλαιο 17.)
A. L. Gnatt et al., Structural basis of transcri-ption: an RNA polymerase II elongation complex at 3.3Å, Science292:1876–1882 (2001).
Αν ήσουν ένας από τους συγγραφείςτης μελέτης αυτής και ανέλυες το μοντέλο, πώς θα περιέγραφες τηδομή του σπειροειδούς πρωτεϊνικού τμήματος που έχει σημανθείμε πράσινο χρώμα, στο κέντρο του μοντέλου;
ΤΙ ΘΑ ΓΙΝΟΤΑΝ ΑΝ...;
ΠΗΓΗ
ΣΥΜΠΕΡΑΣΜΑ
ΑΠΟΤΕΛΕΣΜΑΤΑ
ΠΕΙΡΑΜΑ
DNA
Πολυμεράση II του RNA
RNA

Σε µια ακόµη πιο σύγχρονη µέθοδο γίνεται χρήση τηςβιοπληροφορικής (βλ. Κεφάλαιο 1) για να αναπαρασταθείη τριδιάστατη δοµή των πολυπεπτιδίων µε βάση την ακο-λουθία των αµινοξέων τους. Το 2005, µια ερευνητική οµά-δα στην Αυστρία χρησιµοποίησε υπολογιστικά εργαλείαγια να αναλύσει την ακολουθία και τη δοµή 129 αλλερ-γιογόνων πρωτεϊνών φυτικής προελεύσεως. Οι επιστήµο-νες αυτοί µπόρεσαν να κατατάξουν όλα αυτά τα αλλερ-γιογόνα σε 20 οικογένειες πρωτεϊνών (από ένα σύνολο3.849 πιθανών οικογενειών), ενώ το 65% κατατάχθηκε σε4 µόνο πρωτεϊνικές οικογένειες. Μια πιθανή ερµηνεία αυ-τών των αποτελεσµάτων είναι ότι η πρόκληση των αλ-λεργικών αντιδράσεων οφείλεται στα κοινά πεπτιδικάτµήµατα αυτών των πρωτεϊνικών οµάδων, κάτι που –ανισχύει– θα αποτελέσει στόχο µιας νέας γενεάς αντιαλλερ-γικών φαρµάκων.
Η κρυσταλλογραφία µε ακτίνες Χ, η φασµατοσκοπίαNMR και η βιοπληροφορική είναι συµπληρωµατικές µέ-θοδοι που βοηθούν στο να κατανοήσουµε τη δοµή τωνπρωτεϊνών, ενώ η συνδυασµένη χρήση τους µας έχει δώ-σει πολύτιµες πληροφορίες για τη λειτουργία των πρω-τεϊνών.
104 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
Οι λειτουργίες των νουκλεϊκών οξέων
Οι δύο τύποι νουκλεϊκών οξέων, το δεοξυριβονουκλεϊκόοξύ (DNA) και το ριβονουκλεϊκό οξύ (RNA), επιτρέπουνστους οργανισµούς να αναπαράγουν τα πολύπλοκα συ-στατικά τους από γενεά σε γενεά. Το DNA είναι το µόνοαπ’ όλα τα βιολογικά είδη µορίων που περιέχει οδηγίες γιατην αντιγραφή του εαυτού του. Επιπλέον, το DNA κατευ-θύνει τη σύνθεση του RNA και, µέσω του RNA, ελέγχει τησύνθεση των πρωτεϊνών (Εικόνα 5.26).
Το DNA είναι το γενετικό υλικό που κληρονοµούν οιοργανισµοί από τους γονείς τους. Κάθε χρωµόσωµα πε-ριέχει ένα µακρύ µόριο DNA που µεταφέρει συνήθως µε-ρικές εκατοντάδες γονίδια, ή και περισσότερα. Όταν ένακύτταρο διαιρείται, τα µόρια του DNA του αντιγράφονταικαι µεταφέρονται από τη µία γενεά κυττάρων στην επόµε-νη. Στο DNA περιέχονται οι κωδικοποιηµένες πληροφο-ρίες που προγραµµατίζουν το σύνολο των δραστηριοτήτωνενός κυττάρου. Το DNA, όµως, δεν εµπλέκεται άµεσα στηδιεκπεραίωση των κυτταρικών λειτουργιών. Η σχέση τουDNA µε τις κυτταρικές λειτουργίες είναι ανάλογη της σχέ-σης που έχει το πρόγραµµα ενός υπολογιστή µε την εκτύ-πωση µιας επιταγής ή την αναγνώριση του κωδικού ενόςπροϊόντος. Όπως είναι απαραίτητο να υπάρχει και έναςεκτυπωτής για την εκτύπωση της επιταγής ή ένας σαρω-τής για την ανάγνωση του κωδικού, έτσι απαιτούνται και οιαντίστοιχες πρωτεΐνες στους οργανισµούς για να πραγµα-τοποιηθεί το γενετικό πρόγραµµα. Τα εκτελεστικά τµήµα-τα, δηλαδή τα µοριακά εργαλεία, ενός κυττάρου είναι κα-τά κύριο λόγο οι πρωτεΐνες. Φέρ’ ειπείν, ο µεταφορέας τουοξυγόνου στα ερυθρά αιµοσφαίρια είναι η πρωτεΐνη αιµο-σφαιρίνη, όχι το DNA που καθορίζει τη δοµή της.
Πώς όµως συµµετέχει το RNA, ο δεύτερος τύπος νου-κλεϊκού οξέος, στη ροή της γενετικής πληροφορίας από τοDNA στις πρωτεΐνες; Κάθε γονίδιο µέσα στο µόριο τουDNA κατευθύνει τη σύνθεση µιας κατηγορίας RNA πουονοµάζεται αγγελιαφόρο RNA (mRNA). Το µόριο τουmRNA αλληλεπιδρά µε τις κυτταρικές «µηχανές» πουσυνθέτουν τις πρωτεΐνες και κατευθύνει την παραγωγήενός πολυπεπτιδίου που διαµορφώνεται τελικά σε πρωτεΐ-νη ή σε κάποιο τµήµα µιας πρωτεΐνης. Θα µπορούσαµε νασυνοψίσουµε τη ροή της γενετικής πληροφορίας στο τρί-πτυχο DNA → RNA → πρωτεΐνη (βλ. Εικόνα 5.26). Τοµέρος του κυττάρου όπου συντίθενται οι πρωτεΐνες είναιορισµένες πολύ µικρές δοµές που ονοµάζονται ριβοσώµα-τα. Σε ένα ευκαρυωτικό κύτταρο, τα ριβοσώµατα βρί-σκονται στο κυτταρόπλασµα, ενώ το DNA βρίσκεται στονπυρήνα. Έτσι, το mRNA είναι το µόριο που αναλαµβάνειτη µεταφορά, από τον πυρήνα στο κυτταρόπλασµα, τωνγενετικών οδηγιών για την κατασκευή των πρωτεϊνών. Ταπροκαρυωτικά κύτταρα στερούνται πυρήνα, αλλά και αυ-τά χρησιµοποιούν RNA για να µεταφέρουν το µήνυµα απότο DNA στα ριβοσώµατα και στα άλλα κυτταρικά εργα-λεία που µεταφράζουν την κωδικοποιηµένη πληροφορίασε ακολουθία αµινοξέων. Μόρια RNA, πάντως, επιτελούνκαι άλλες λειτουργίες στο κύτταρο.
Ε Λ Ε Γ Χ Ο Σ Ε Ν Ν Ο Ι Ω Ν 5.41. Γιατί σταµατά να λειτουργεί φυσιολογικά µια απο-
διατεταγµένη πρωτεΐνη;
2. Ποια τµήµατα µιας πολυπεπτιδικής αλυσίδας συµ-µετέχουν στους δεσµούς που συνθέτουν τη δευτερο-ταγή δοµή της; Ποια τµήµατα συµµετέχουν στην τρι-τοταγή δοµή;
3. Αν µια γενετική µετάλλαξηµεταβάλει την πρωτοταγή δοµή µιας πρωτεΐνης, µεποιον τρόπο θα µπορούσε να διαταράξει τη λειτουρ-γία της;Για προτεινόµενες απαντήσεις, βλ. Παράρτηµα Α.
ΤΙ ΘΑ ΓΙΝΟΤΑΝ ΑΝ...;
Τ Μ Η Μ Α 5.5
Τα νουκλεϊκά οξέα αποθηκεύουνκαι µεταβιβάζουν κληρονοµικέςπληροφορίες
Όπως είδαµε, το σχήµα των πρωτεϊνών καθορίζεται απότην πρωτοταγή δοµή των πολυπεπτιδίων. Ωστόσο, τι καιπώς ορίζει αυτή την πρωτοταγή δοµή; Η ακολουθία τωναµινοξέων σε κάθε πολυπεπτίδιο προγραµµατίζεται απόµια αντίστοιχη µονάδα κληρονοµικότητας, που ονοµάζε-ται γονίδιο. Τα γονίδια αποτελούνται από DNA, δηλαδήένα πολυµερές της κατηγορίας ενώσεων που ονοµάζουµενουκλεϊκά οξέα.
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 105
Δομή των νουκλεϊκών οξέων
Τα νουκλεϊκά οξέα είναι πολυνουκλεοτίδια, δηλαδή µα-κροµόρια που υπάρχουν σε πολυµερισµένη µορφή (Εικό-να 5.27α). Όπως υπονοεί και το όνοµά τους, κάθε πολυ-νουκλεοτίδιο αποτελείται από µονοµερή που ονοµάζονταινουκλεοτίδια. Κάθε νουκλεοτίδιο, τώρα, αποτελείται απότρία µέρη: µια αζωτούχο βάση, ένα σάκχαρο µε πέντε άτο-µα άνθρακα (δηλαδή, µια πεντόζη) και µια φωσφορικήοµάδα (Εικόνα 5.27β). Ένα νουκλεοτίδιο χωρίς φωσφο-ρική οµάδα ονοµάζεται νουκλεοζίτης.
Νουκλεοτιδικά μονομερή
Θα ξεκινήσουµε την ανάλυση ενός νουκλεοτιδίου µε δύοαπό τα τρία συστατικά του, την αζωτούχο βάση και τοσάκχαρο (Εικόνα 5.27γ). Υπάρχουν δύο οικογένειες αζω-τούχων βάσεων, οι πυριµιδίνες και οι πουρίνες. Μια πυ-ριµιδίνη έχει έναν εξαµερή δακτύλιο αποτελούµενο απόάτοµα άνθρακα και αζώτου. (Το άτοµο του αζώτου έχειτην τάση να δεσµεύει ιόντα Η+, γι’ αυτό η συγκεκριµένηένωση ονοµάζεται αζωτούχος βάση). Στην οικογένεια τωνπυριµιδινών ανήκουν η κυτοσίνη (C), η θυµίνη (Τ) και ηουρακίλη (U). Η άλλη οικογένεια αζωτούχων βάσεων εί-ναι οι πουρίνες, που έχουν µεγαλύτερο µέγεθος, καθώςαποτελούνται από δύο συγχωνευµένους δακτυλίους, ένανεξαµερή και έναν πενταµερή. Στην οικογένεια των πουρι-
1
mRNA
Ριβόσωμα
ΑμινοξέαΠολυπεπτίδιο
ΚΥΤΤΑΡΟΠΛΑΣΜΑ
DNA
mRNA
ΠΥΡΗΝΑΣ
Σύνθεση τουmRNA στονπυρήνα
1
Μετακίνηση τουmRNA στο κυτταρόπλασμαμέσω των πυρηνικώνπόρων
2
Σύνθεσηπρωτεΐνης
3
Εικόνα 5.26 DNA→ RNA→ πρωτεΐνη. Στα ευκαρυωτικά κύτταρα,το DNA του πυρήνα προγραμματίζει την παραγωγή των πρωτεϊνών στοκυτταρόπλασμα, υπαγορεύοντας τη σύνθεση του αγγελιαφόρου RNA(mRNA). (Στην πραγματικότητα, ο πυρήνας του κυττάρου είναι πολύμεγαλύτερος από τα άλλα οργανίδια της εικόνας.)
(α)
(β) Νουκλεοτίδιο
(γ) Συστατικά των νουκλεοζιτών
NH2
NH2
NH2
C C
NH
O
CH
CH
N
CNH
O
C
CH
HN
C
CH3
O
C
NH
O
CH
CH
HN
C
O
O
Κυτοσίνη (C)
Αζωτούχες βάσεις
Πυριμιδίνες
Σάκχαρα
Πουρίνες
Θυμίνη (T, στο DNA)
C
N
C
C
N
CH
C
Αδενίνη (A)
Δεοξυριβόζη (στο DNA) Ριβόζη (στο RNA)
N
NH
C
NC
NH
CHC HC
N
NH
Γουανίνη (G)
Αζωτούχοςβάση
Νουκλεοζίτης
άκρο 5'
άκρο 3‘
5‘C
Σάκχαρο(πεντόζη)
Φωσφορικήομάδα
O
O–
P–O O
OH
CH2 O
O
O
O
O
4‘ 1‘
5‘
3‘ 2‘
HOCH2 HOCH2OH
H H
H
OH
H
H
O
4‘ 1‘
5‘
3‘ 2‘
OH
H H
H
OH
H
OH
O
Ουρακίλη(U, στο RNA)
Πολυνουκλεοτίδιο ή νουκλεϊκό οξύ
3‘C
3‘C
5‘C
5‘C
3‘C
Εικόνα 5.27 Συστατικά των νουκλεϊκών οξέων. (α) Ένα πολυνουκλεοτίδιοσυνίσταται από έναν σκελετό αποτελούμενο από σάκχαρα και φωσφορικές ομάδες,πάνω στον οποίο συνδέονται και προεξέχουν αζωτούχες βάσεις. (β) Ένα νουκλεοτίδιοαποτελείται από μια αζωτούχο βάση, ένα σάκχαρο και μια φωσφορική ομάδα. Χωρίς τηφωσφορική ομάδα, το μόριο ονομάζεται νουκλεοζίτης. (γ) Ένας νουκλεοζίτηςαποτελείται από μια αζωτούχο βάση (πουρίνη ή πυριμιδίνη) και ένα σάκχαρο με πέντεάτομα άνθρακα (δεοξυριβόζη ή ριβόζη).

νών ανήκουν η αδενίνη (Α) και η γουανίνη (G). Κάθε συγ-κεκριµένη πυριµιδίνη ή πουρίνη διαφέρει από τις υπόλοι-πες της ίδιας οικογένειας ως προς τις χηµικές οµάδες πουείναι συνδεδεµένες στους δακτυλίους. Η αδενίνη, η γουα-νίνη και η κυτοσίνη απαντούν και στα δύο είδη νουκλεϊ-κών οξέων. Η θυµίνη απαντά µόνο στο DNA και η ουρα-κίλη µόνο στο RNA.
Το σάκχαρο που συνδέεται µε την αζωτούχο βάση στανουκλεοτίδια του RNA είναι η ριβόζη, ενώ στα νουκλεο-τίδια του DNA είναι η δεοξυριβόζη (βλ. Εικόνα 5.27γ).Η µόνη διαφορά ανάµεσα σ’ αυτά τα δύο σάκχαρα είναι τοότι η δεοξυριβόζη έχει ένα άτοµο οξυγόνου λιγότερο στονδεύτερο άνθρακα του δακτυλίου (γι’ αυτό άλλωστε ονο-µάζεται δεοξυριβόζη, που σε ελεύθερη απόδοση σηµαίνει«αποξυγονωµένη» ριβόζη). Για να µπορούµε να ξεχωρί-ζουµε τα άτοµα της αζωτούχου βάσης και του σακχάρου,τους δίνουµε έναν αριθµό, προσθέτοντας έναν τόνο µετάτον αριθµό των ατόµων του σακχάρου. Έτσι, ο δεύτεροςάνθρακας στο δακτύλιο του σακχάρου αναφέρεται ως άν-θρακας 2´ και ο άνθρακας που εξέχει από τον δακτύλιοαναφέρεται ως άνθρακας 5´.
Μέχρι στιγµής έχουµε εξετάσει την σύσταση ενός νου-κλεοζίτη. Για να ολοκληρώσουµε την κατασκευή του νου-κλεοτιδίου θα προσθέσουµε µια φωσφορική οµάδα στονάνθρακα 5´ του σακχάρου (βλ. Εικόνα 5.27β). Το µόριοπου προκύπτει µε την προθήκη αυτή είναι ένας µονοφω-σφορικός νουκλεοζίτης, ή νουκλεοτίδιο.
Νουκλεοτιδικά πολυμερή
Τώρα µπορούµε να δούµε µε ποιον τρόπο συνδέονται µε-ταξύ τους τα νουκλεοτίδια για να συνθέσουν ένα πολυ-νουκλεοτίδιο. ∆ύο γειτονικά νουκλεοτίδια ενώνονται µεφωσφοδιεστερικό δεσµό, δηλαδή µε δεσµό στον οποίο τασάκχαρα δύο γειτονικών νουκλεοτιδίων συνδέονται µέσωφωσφορικής οµάδας. Το χαρακτηριστικό αυτού του δε-σµού είναι ότι µε τη διαδοχική σύνδεση πολλών νουκλε-οτιδίων σχηµατίζεται ένας σκελετός από επαναλαµβανό-µενα τµήµατα σακχάρων και φωσφορικών οµάδων (βλ.Εικόνα 5.27α). Αν κοιτάξουµε προσεκτικά, θα δούµε ότιτα δύο ελεύθερα άκρα του σχηµατιζόµενου πολυµερούςδιαφέρουν µεταξύ τους. Στο ένα άκρο υπάρχει µια φω-σφορική οµάδα που συνδέεται µε τον άνθρακα 5´, ενώ στοάλλο άκρο υπάρχει µια υδροξυλοµάδα στον άνθρακα 3´.Από εδώ και στο εξής, θα αναφερόµαστε σε αυτά τα δύοάκρα ως άκρο 5´ και άκρο 3´, αντίστοιχα. Μπορούµε επο-µένως να πούµε ότι ο σακχαροφωσφορικός σκελετός σεµια αλυσίδα DNA έχει, ήδη από τον τρόπο κατασκευήςτου, συγκεκριµένη φορά από το άκρο 5´ προς το άκρο 3´(όπως ένας δρόµος µονής κατεύθυνσης). Καθ’ όλη τη δια-δροµή αυτού του σκελετού προεξέχουν οι αζωτούχες βά-σεις των νουκλεοτιδίων.
Η αλληλουχία των βάσεων κατά µήκος ενός πολυµε-ρούς DNA (ή mRNA) είναι µοναδική για κάθε γονίδιο καιεµπεριέχει µια πολύ συγκεκριµένη πληροφορία για το κύτ-ταρο. Επειδή τα γονίδια έχουν µήκος µερικές εκατοντά-
106 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
δες ή και χιλιάδες νουκλεοτίδια, οι πιθανές αλληλουχίεςνουκλεοτιδικών βάσεων είναι πρακτικά άπειρες. Η συµ-βολή στο κύτταρο ενός γονιδίου είναι κωδικοποιηµένηστην ακριβή αλληλουχία µε την οποία διατάσσονται οιτέσσερεις νουκλεοτιδικές βάσεις του DNA. Λόγου χάρη, ηαλληλουχία AGGTAACTT σηµαίνει ένα πράγµα, ενώ ηαλληλουχία CGCTTTAAC σηµαίνει κάτι διαφορετικό.(Προφανώς, ένα πλήρες γονίδιο έχει πολύ µεγαλύτερο µή-κος.) Η γραµµική σειρά των βάσεων σε ένα γονίδιο καθο-ρίζει την ακολουθία των αµινοξέων –µε άλλα λόγια, τηνπρωτοταγή δοµή– µιας πρωτεΐνης, που µε τη σειρά της κα-θορίζει την τριδιάστατη διαµόρφωση αυτής της πρωτεΐνηςστον χώρο, και εποµένως τη λειτουργία της στο κύτταρο ήστον οργανισµό.
Η διπλή έλικα του DNA
Τα µόρια του κυτταρικού RNA αποτελούνται από µια µο-νή πολυνουκλεοτιδική αλυσίδα, όπως αυτή στην Εικόνα5.27. Αντίθετα, τα µόρια του κυτταρικού DNA αποτελούν-ται από δύο πολυνουκλεοτίδια που περιελίσσονται γύρωαπό έναν φανταστικό άξονα, σχηµατίζοντας διπλή έλικα(Εικόνα 5.28). Το 1953, οι James Watson και FrancisCrick, εργαζόµενοι στο Πανεπιστήµιο του Cambridge, εξέ-φρασαν για πρώτη φορά την άποψη ότι η δοµή του DNAστον χώρο έχει τη µορφή διπλής έλικας. Και οι δύο αυτοίσακχαροφωσφορικοί σκελετοί έχουν κατεύθυνση 5´→ 3´,αλλά η φορά τους είναι αντίθετη, διάταξη που περιγράφε-ται ως αντιπαράλληλη (κάτι σαν τα δύο αντίθετα ρεύµα-τα ενός δρόµου διπλής κατεύθυνσης). Οι σακχαροφωσφο-ρικοί σκελετοί βρίσκονται στην εξωτερική πλευρά της έλι-κας, ενώ οι αζωτούχες βάσεις είναι στραµµένες προς τοεσωτερικό της, έτσι ώστε κάθε βάση της µίας αλυσίδας ναδηµιουργεί ζεύγος µε µια βάση της αντιπαράλληλης αλυ-σίδας. Τα δύο πολυνουκλεοτιδικά µόρια, ή αλυσίδες, όπωςαλλιώς ονοµάζονται, συγκρατούνται µαζί τους χάρη στουςδεσµούς υδρογόνου που αναπτύσσονται ανάµεσα στα ζεύ-γη των βάσεων και στους δεσµούς van der Waals που ανα-πτύσσονται µεταξύ γειτονικών βάσεων κάθε αλυσίδας. Ταπερισσότερα µόρια DNA έχουν πολύ µεγάλο µήκος και οιδύο αλυσίδες του συνδέονται µε χιλιάδες ή και εκατοµµύ-ρια ζεύγη βάσεων. Κάθε δίκλωνο µόριο DNA περιέχει πά-ρα πολλά γονίδια, καθένα από τα οποία καταλαµβάνει συγ-κεκριµένο τµήµα του µορίου του DNA.
Τα ζεύγη των βάσεων που δηµιουργούνται ανάµεσα στιςδύο αντιπαράλληλες αλυσίδες του DNA δεν είναι τυχαία.Για κάθε βάση υπάρχει µία µόνο µε την οποία µπορεί νασχηµατιστεί ζεύγος. Έτσι, η αδενίνη (Α) ζευγαρώνει πάν-τοτε µε θυµίνη (Τ) και η γουανίνη (G) ζευγαρώνει πάντοτεµε κυτοσίνη (C). Αν υποθέσουµε ότι βρισκόµαστε σε έναµοριακό όχηµα που κινείται δίπλα από τη διπλή έλικα ενόςµορίου DNA τότε, καταγράφοντας την αλληλουχία των βά-σεων στη µια αλυσίδα, θα γνωρίζαµε την αλληλουχία βά-σεων και της άλλης. Αν, π.χ., σε ένα τµήµα του δίκλωνουDNA συναντούσαµε στη µια αλυσίδα την αλληλουχία 5´-

AGGTCCG-3´ τότε, σύµφωνα µε τον κανόνα ζευγαρώµα-τος των βάσεων, η άλλη αλυσίδα (για το ίδιο τµήµα τουDNA) θα είχε την αλληλουχία 3´-TCCAGGC-5´. Με άλλαλόγια, οι δύο αλυσίδες στη διπλή έλικα του DNA είναι συµ-πληρωµατικές και καθεµιά τους είναι το προβλέψιµο είδω-λο της άλλης. Αυτή ακριβώς η ιδιότητα του DNA ευθύνε-ται για την κληρονοµικότητα, αφού επιτρέπει στα γονίδιανα αντιγράφονται µε ακρίβεια (βλ. Εικόνα 5.28). Όταν ένακύτταρο ετοιµάζεται να διαιρεθεί, κάθε αλυσίδα του µορί-ου του DNA λειτουργεί ως εκµαγείο, µε βάση το οποίο δια-τάσσονται στη σειρά τα κατάλληλα νουκλεοτίδια για νασυντεθεί η συµπληρωµατική αλυσίδα. Αποτέλεσµα είναινα δηµιουργηθούν δύο πανοµοιότυπα αντίγραφα του αρ-χικού δίκλωνου µορίου του DNA, που κατόπιν διαµοιρά-ζονται στα δύο θυγατρικά κύτταρα. Βλέπουµε, λοιπόν, ότικάθε φορά που αναπαράγεται ένα κύτταρο και τίθεται σελειτουργία η διαδικασία µεταφοράς της γενετικής πληρο-φορίας, η ίδια η δοµή του µορίου του DNA καθορίζει καιτη λειτουργία του.
DNA και πρωτεΐνες ως μέτρο της εξέλιξης
Συνηθίζουµε να θεωρούµε τους κοινούς χαρακτήρες (π.χ.το τρίχωµα ή την παραγωγή γάλακτος, στα θηλαστικά) ως
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 107
απόδειξη κοινής εξελικτικής καταγωγής. Τώρα που γνω-ρίζουµε ότι το DNA µεταφέρει τις κληρονοµικές πληρο-φορίες µέσω των γονιδίων, καταλαβαίνουµε ότι τα γονίδιακαι τα προϊόντα τους (δηλαδή οι πρωτεΐνες) αποτελούν τοκληρονοµικό υπόβαθρο κάθε οργανισµού. Η γραµµική αλ-ληλουχία των νουκλεοτιδίων στο µόριο του DNA µετα-φέρεται από τους γονείς στους απογόνους τους και είναιεκείνη που καθορίζει την ακολουθία των αµινοξέων στιςπρωτεΐνες. Τα αδέλφια εµφανίζουν µεγαλύτερες οµοιότη-τες µεταξύ τους, όσον αφορά το DNA και τις πρωτεΐνες,σε σχέση µε άλλα, µη συγγενικά άτοµα του ίδιου είδους.Εφόσον ισχύει η εξελικτική θεώρηση της ζωής, τότε θαπρέπει να έχουµε τη δυνατότητα να επεκτείνουµε την αν-τίληψη αυτής της «µοριακής γενεαλογίας» και σε ό,τι αφο-ρά τις σχέσεις µεταξύ των ειδών. Θα πρέπει, µε άλλα λό-για, δύο είδη που φαίνεται να είναι στενοί συγγενείς (βά-σει των απολιθωµάτων και της ανατοµικής τους) να έχουνµεγαλύτερο ποσοστό οµοιοτήτων στο DNA και στις πρω-τεΐνες τους απ’ ό,τι δύο πιο αποµακρυσµένα, δηλαδή λι-γότερο συγγενικά, είδη. Πράγµατι, αυτό ισχύει. Ας συγ-κρίνουµε π.χ. την πολυπεπτιδική αλυσίδα της αιµοσφαι-ρίνης του ανθρώπου µε το αντίστοιχο πολυπεπτίδιο της αι-µοσφαιρίνης σε πέντε άλλα σπονδυλωτά. Σε τούτη τηναλυσίδα των 146 αµινοξέων, η αιµοσφαιρίνη του ανθρώ-που διαφέρει από εκείνη του γορίλλα µόνο σε ένα αµινο-
T
T
T
Σάκχαροφωσφορικοίσκελετοί
Ζεύγη βάσεων (συνδέονται μεδεσμούς υδρογόνου)
Νουκλεοτίδιο πουθα προστεθεί στηνέα αλυσίδα
Νέεςαλυσίδες
Προγονικές αλυσίδες
A
G
AG
C
GT
T
A
T
A
A
A
G
C
A
C
C
A
T
G
T
A
G
C
G
G
T
T
T
A T
A
T
A
G
C
C
C
A
A G
C
C
A
TGT
A
G
C G
T
T
TA
TA
GC
C
A
A
GC
C
A
T
άκρο 3‘άκρο 5‘
άκρο 3‘
άκρο 5‘
άκρο 5‘
άκρο 3‘
άκρο 3‘
άκρο 5‘
T
A
G
C
Εικόνα 5.28 Η διπλή έλικα του DNA και ηαντιγραφή της. Το μόριο του DNA αποτελείταισυνήθως από δύο αλυσίδες που περιελίσσονται ημία γύρω από την άλλη. Οι σακχαροφωσφορικοίσκελετοί των δύο αντιπαράλληλων κλώνων (οι δύοταινίες σε αποχρώσεις του μπλε) βρίσκονται στηνεξωτερική πλευρά της έλικας. Οι δύο αλυσίδεςσυγκρατούνται μαζί από ζεύγη αζωτούχων βάσεωνπου συνδέονται μεταξύ τους με δεσμούςυδρογόνου. Κάθε βάση συμβολίζεται με ξεχωριστόσχήμα. Όπως βλέπουμε, η αδενίνη (Α) μπορεί νασχηματίσει ζεύγος μόνο με τη θυμίνη (Τ) και ηγουανίνη (G) μόνο με την κυτοσίνη (C). Όταν ένακύτταρο ετοιμάζεται να διαιρεθεί, οι δύο αλυσίδεςτης διπλής έλικας αποχωρίζονται και καθεμιάλειτουργεί ως εκμαγείο για την ορθή διάταξη τωννουκλεοτιδίων σε μια νέα συμπληρωματικήαλυσίδα (πορτοκαλί χρώμα). Εδώ, κάθε αλυσίδαDNA είναι το δομικό ισοδύναμο ενόςπολυνουκλεοτιδίου, όπως εκείνουστην Εικόνα 5.27α.

ΣΥΝΟΨΗ ΒΑΣΙΚΩΝ ΕΝΝΟΙΩΝ
Τ Μ Η Μ Α 5.1Τα μακρομόρια είναι πολυμερή που συντίθενταιαπό μονομερή (σ. 83-85) Σύνθεση και διάσπαση των πολυμερών Οι υδατάνθρα-
κες, οι πρωτεΐνες και τα νουκλεϊκά οξέα είναι πολυµερή,
δηλαδή αλυσίδες µονοµερών. Τα συστατικά των λιπιδίωνποικίλλουν. Τα µονοµερή σχηµατίζουν µεγαλύτερα µόριαµε το να ενώνονται µεταξύ τους µε αντιδράσεις αφυδάτω-σης, δηλαδή αντιδράσεις στις οποίες απελευθερώνονται µό-ρια νερού. Τα πολυµερή αποδοµούνται µε την αντίστροφηδιαδικασία που ονοµάζεται υδρόλυση.
Ποικιλομορφία των πολυμερών Από έναν µικρό αριθµόµονοµερών µπορεί να προκύψει τεράστια ποικιλία πολυµε-ρών.
Κεφάλαιο5Επανάληψη
108 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
ξύ, ενώ από εκείνη του βατράχου σε 67 αµινοξέα. Πάν-τως, και στις δύο περιπτώσεις πρόκειται για διαφορές πουδεν αλλοιώνουν τη λειτουργικότητα της συγκεκριµένης
πρωτεΐνης. Η µοριακή βιολογία έχει προσθέσει ένα νέο«µέτρο» στην εργαλειοθήκη που χρησιµοποιούν οι βιολό-γοι για να µετρούν τις εξελικτικές συγγένειες.
Ε Λ Ε Γ Χ Ο Σ Ε Ν Ν Ο Ι Ω Ν 5.5
1. Πηγαίνετε στην κορυφή της Εικόνας 5.27α και αριθµήστε όλα τα άτοµα άνθρακα στα σάκχαρα των τριών νου-κλεοτιδίων. Σχεδιάστε έναν κύκλο γύρω από τις αζωτούχες βάσεις και ένα αστέρι πάνω από τις φωσφορικές οµά-δες.
2. Σε µια διπλή έλικα DNA βρίσκουµε, σε µικρό τµήµα της µίας αλυσίδας, την ακόλουθη αλληλουχία αζωτούχωνβάσεων: 5´-TAGGCCT-3´. Αντιγράψτε σε ένα τετράδιο αυτή την αλυσίδα, και τη συµπληρωµατική της, σηµει-ώνοντας µε σαφήνεια στη συµπληρωµατική αλυσίδα τα άκρα 5´ και 3´.
3. (α) Έστω ότι αντικαταστάθηκε κάποια βάση στη µία από τις δύο αλυσίδες της διπλής έλι-κας DNA της ερώτησης 2, και προέκυψε το ακόλουθο ζεύγος συµπληρωµατικών αλληλουχιών:
5´-TAAGCCT-3´
3´-ATCCGGA-5´
Σχεδιάστε τις δύο αλυσίδες και σηµειώστε µε κύκλο τις δύο αταίριαστες βάσεις. (β) Αν αντιγραφεί η πρώτη τρο-ποποιηµένη αλυσίδα (το 5´-TAAGCCT-3´), ποια θα είναι η αλληλουχία βάσεων στη νέα συµπληρωµατική τηςαλυσίδα;
Για προτεινόµενες απαντήσεις, βλ. Παράρτηµα Α.
ΤΙ ΘΑ ΓΙΝΟΤΑΝ ΑΝ...;
Ο κανόνας των αναδυόμενων ιδιοτήτωνστη χημεία της ζωής: Επανάληψη
Υπάρχει µια ιεραρχία στα επίπεδα οργάνωσης της ζωής(βλ. Εικόνα 1.4). Σε κάθε ανώτερο επίπεδο οργάνωσηςπροκύπτουν νέες ιδιότητες της έµβιας ύλης. Στα Κεφάλαια2-5, εξετάσαµε τη χηµεία των ζωντανών οργανισµών και,µελετώντας τον τρόπο µε τον οποίο προκύπτουν νέες ιδιό-τητες σε κάθε ανώτερο επίπεδο οργάνωσης, αποκτήσαµεµια πιο ολοκληρωµένη θεώρηση της ζωής.
Είδαµε ότι οι ιδιότητες του νερού προκύπτουν από τιςαλληλεπιδράσεις των µορίων του και της συγκεκριµένηςδιάταξης των ατόµων υδρογόνου και οξυγόνου από ταοποία αυτό αποτελείται. Απλοποιήσαµε την τεράστια πο-
λυπλοκότητα και ποικιλία των οργανικών ενώσεων, εστιά-ζοντας το ενδιαφέρον µας κυρίως στον ανθρακικό σκελε-τό τους και στις πρόσθετες χηµικές οµάδες µε τις οποίεςσυνδέεται αυτός ο σκελετός. Είδαµε, τέλος, ότι τα βιολο-γικά µακροµόρια συναρµολογούνται από µικρά οργανικάµόρια και γνωρίσαµε τις νέες ιδιότητές τους. Οι γνώσειςπου αποκτήσαµε για τα µακροµόρια µας επιτρέπουν πλέ-ον να προχωρήσουµε στην Ενότητα 2, όπου θα µελετή-σουµε τη δοµή και τη λειτουργία των κυττάρων. Θα προ-σπαθήσουµε να κρατήσουµε µια ισορροπία ανάµεσα στηνανάγκη να επιµερίσουµε τις λειτουργίες της ζωής σεαπλές, θεµελιώδεις διαδικασίες και στην απώτερη ικανο-ποίηση του να κατανοήσουµε ολοκληρωµένα τις διαδικα-σίες αυτές στο ευρύτερο πλαίσιο λειτουργίας τους.
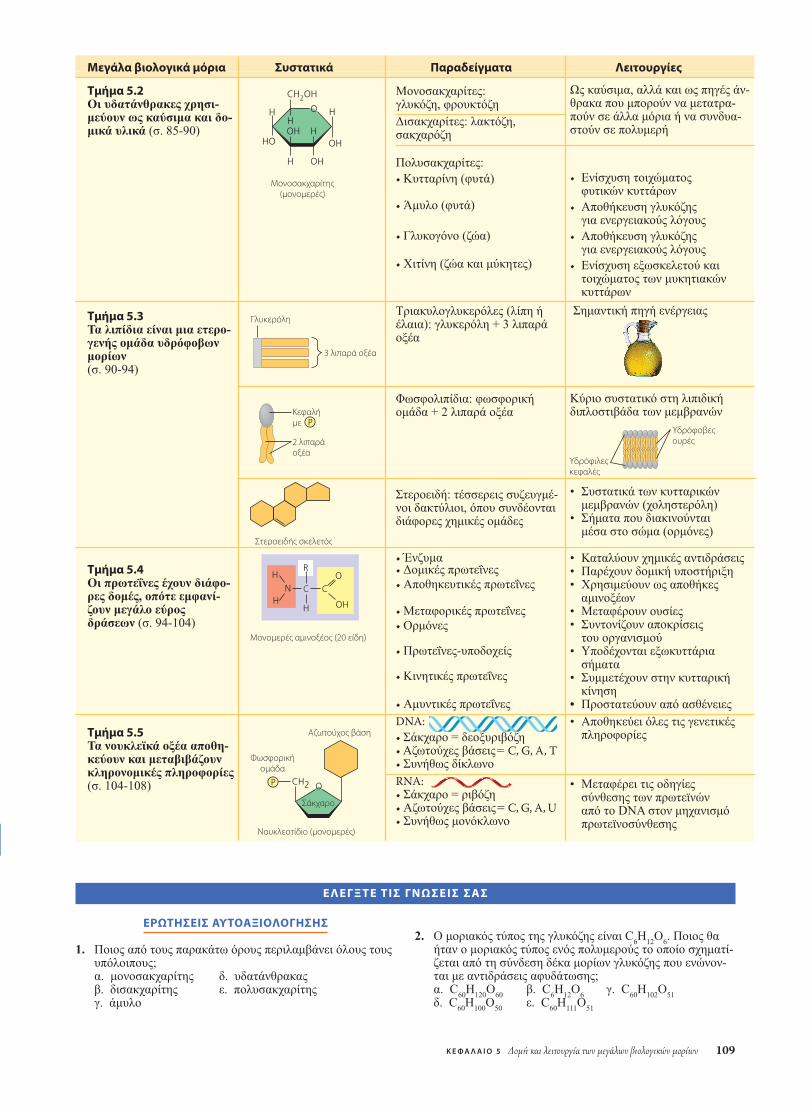
Κ Ε Φ Α Λ Α Ι Ο 5 ∆οµή και λειτουργία των µεγάλων βιολογικών µορίων 109
Μεγάλα βιολογικά μόρια Συστατικά Παραδείγματα Λειτουργίες
Γλυκερόλη
3 λιπαρά οξέα
Υδρόφοβεςουρές
Υδρόφιλεςκεφαλές
OH
O
CCC
R
H
N
H
H
Μονομερές αμινοξέος (20 είδη)
Στεροειδής σκελετός
Αζωτούχος βάση
Σάκχαρο
Νουκλεοτίδιο (μονομερές)
Φωσφορικήομάδα
CH2 OP
Μονοσακχαρίτης(μονομερές)
OH
H
H OH
OH
CH2OH
OH H
H
HO
Τμήμα 5.2Οι υδατάνθρακες χρησι-µεύουν ως καύσιµα και δο-µικά υλικά (σ. 85-90)
Τμήμα 5.3Τα λιπίδια είναι µια ετερο-γενής οµάδα υδρόφοβωνµορίων(σ. 90-94)
Τμήμα 5.4Οι πρωτεΐνες έχουν διάφο-ρες δοµές, οπότε εµφανί-ζουν µεγάλο εύροςδράσεων (σ. 94-104)
Τμήμα 5.5Τα νουκλεϊκά οξέα αποθη-κεύουν και µεταβιβάζουνκληρονοµικές πληροφορίες(σ. 104-108)
Μονοσακχαρίτες:γλυκόζη, φρουκτόζη∆ισακχαρίτες: λακτόζη,σακχαρόζη
Πολυσακχαρίτες:• Κυτταρίνη (φυτά)
• Άµυλο (φυτά)
• Γλυκογόνο (ζώα)
• Χιτίνη (ζώα και µύκητες)
Τριακυλογλυκερόλες (λίπη ήέλαια): γλυκερόλη + 3 λιπαράοξέα
Φωσφολιπίδια: φωσφορικήοµάδα + 2 λιπαρά οξέα
Στεροειδή: τέσσερεις συζευγµέ-νοι δακτύλιοι, όπου συνδέονταιδιάφορες χηµικές οµάδες
• Ένζυµα• ∆οµικές πρωτεΐνες• Αποθηκευτικές πρωτεΐνες
• Μεταφορικές πρωτεΐνες• Ορµόνες
• Πρωτεΐνες-υποδοχείς
• Κινητικές πρωτεΐνες
• Αµυντικές πρωτεΐνεςDNA:• Σάκχαρο = δεοξυριβόζη• Αζωτούχες βάσεις C, G, A, T• Συνήθως δίκλωνοRNA:• Σάκχαρο = ριβόζη• Αζωτούχες βάσεις C, G, A, U• Συνήθως µονόκλωνο
2 λιπαράοξέα
Κεφαλή με P
Ως καύσιµα, αλλά και ως πηγές άν-θρακα που µπορούν να µετατρα-πούν σε άλλα µόρια ή να συνδυα-στούν σε πολυµερή
• Ενίσχυση τοιχώµατοςφυτικών κυττάρων
• Αποθήκευση γλυκόζηςγια ενεργειακούς λόγους
• Αποθήκευση γλυκόζηςγια ενεργειακούς λόγους
• Ενίσχυση εξωσκελετού καιτοιχώµατος των µυκητιακώνκυττάρων
Σηµαντική πηγή ενέργειας
Κύριο συστατικό στη λιπιδικήδιπλοστιβάδα των µεµβρανών
• Συστατικά των κυτταρικώνµεµβρανών (χοληστερόλη)
• Σήµατα που διακινούνταιµέσα στο σώµα (ορµόνες)
• Καταλύουν χηµικές αντιδράσεις• Παρέχουν δοµική υποστήριξη• Χρησιµεύουν ως αποθήκες
αµινοξέων• Μεταφέρουν ουσίες• Συντονίζουν αποκρίσεις
του οργανισµού• Υποδέχονται εξωκυττάρια
σήµατα• Συµµετέχουν στην κυτταρική
κίνηση• Προστατεύουν από ασθένειες• Αποθηκεύει όλες τις γενετικές
πληροφορίες
• Μεταφέρει τις οδηγίεςσύνθεσης των πρωτεϊνώναπό το DNA στον µηχανισµόπρωτεϊνοσύνθεσης
ΕΛΕΓξΤΕ ΤΙΣ ΓΝΩΣΕΙΣ ΣΑΣ
ΕΡΩΤΗΣΕΙΣ ΑΥΤΟΑξΙΟΛΟΓΗΣΗΣ
1. Ποιος από τους παρακάτω όρους περιλαµβάνει όλους τουςυπόλοιπους;α. µονοσακχαρίτης δ. υδατάνθρακαςβ. δισακχαρίτης ε. πολυσακχαρίτηςγ. άµυλο
2. Ο µοριακός τύπος της γλυκόζης είναι C6H12O6. Ποιος θαήταν ο µοριακός τύπος ενός πολυµερούς το οποίο σχηµατί-ζεται από τη σύνδεση δέκα µορίων γλυκόζης που ενώνον-ται µε αντιδράσεις αφυδάτωσης;α. C60H120O60 β. C6H12O6 γ. C60H102O51δ. C60H100O50 ε. C60H111O51

110 Ε Ν Ο Τ Η ΤΑ 1 Η χηµεία της ζωής
3. Το ένζυµο αµυλάση µπορεί να διασπάσει τους γλυκοζιτι-κούς δεσµούς ανάµεσα σε µονοµερή γλυκόζης µόνο εάναυτά τα µονοµερή είναι της µορφής α. Ποια από τα παρα-κάτω µπορεί να διασπάσει η αµυλάση;α. γλυκογόνο, άµυλο και αµυλοπηκτίνηβ. γλυκογόνο και κυτταρίνηγ. κυτταρίνη και χιτίνηδ. άµυλο και χιτίνηε. άµυλο, αµυλοπηκτίνη και κυτταρίνη
4. Σχετικά µε τα ακόρεστα λίπη, ποιες από τις παρακάτω προ-τάσεις είναι αληθείς;α. Είναι πιο συχνά στα ζώα απ’ ό,τι στα φυτά.β. Ο ανθρακικός σκελετός των λιπαρών οξέων τους περιέ-
χει διπλούς δεσµούς.γ. Κατά κανόνα στερεοποιούνται σε θερµοκρασία δωµα-
τίου.δ. Περιέχουν περισσότερα υδρογόνα απ’ ό,τι τα κορεσµέ-
να λίπη µε ίδιο αριθµό ατόµων άνθρακα.ε. Έχουν πολύ λιγότερα µόρια λιπαρών οξέων ανά µόριο
λίπους.
5. Σε µια πρωτεΐνη, ποια δοµή επηρεάζεται λιγότερο από τηδιάσπαση των δεσµών υδρογόνου;α. Η πρωτοταγής δοµή.β. Η δευτεροταγής δοµή.γ. Η τριτοταγής δοµή.δ. Η τεταρτοταγής δοµή.ε. Όλες οι δοµές επηρεάζονται εξίσου.
6. Ποια από τα επόµενα ζεύγη αλληλουχιών βάσεων θα µπο-ρούσαν να σχηµατίσουν µικρό τµήµα µιας φυσιολογικήςδιπλής έλικας DNA;α. 5´-πουρίνη-πυριµιδίνη-πουρίνη-πυριµιδίνη-3´ µε
3´-πουρίνη-πυριµιδίνη-πουρίνη-πυριµιδίνη-5´β. 5´-AGCT-3´ µε 5´-TCGA-3´γ. 5´-GCGC-3´ µε 5´-TATA-3´δ. 5´-ATGC-3´ µε 5´-GCAT-3´ε. Όλα τα ζεύγη είναι σωστά.
7. Τα ένζυµα που διασπούν το DNA καταλύουν την υδρόλυ-ση του οµοιοπολικού δεσµού που συνδέει τα νουκλεοτίδιαµεταξύ τους. Τι θα συνέβαινε στα µόρια του DNA αν επι-δρούσαν πάνω τους αυτά τα ένζυµα;α. Θα αποχωρίζονταν οι δύο αλυσίδες της διπλής έλικας.β. Θα λύονταν οι φωσφοδιεστερικοί δεσµοί ανάµεσα στα
δεοξυριβοζικά σάκχαρα.γ. Οι πουρίνες θα αποχωρίζονταν από τα δεοξυριβοζικά
σάκχαρα.δ. Οι πυριµιδίνες θα αποχωρίζονταν από τα δεοξυριβοζικά
σάκχαρα.ε. Όλες οι βάσεις θα αποχωρίζονταν από τα δεοξυριβοζι-
κά σάκχαρα.
8. Φτιάξτε έναν πίνακα όπου θα ταξινοµούνται λογικά οιακόλουθοι όροι και βάλτε τίτλους στις στήλες και στιςγραµµές.
φωσφοδιεστερικοίδεσµοί πολυπεπτίδια µονοσακχαρίτες
πεπτιδικοί δεσµοί τριακυλογλυκερόλες νουκλεοτίδιαγλυκοζιτικοί δεσµοί πολυνουκλεοτίδια αµινοξέαεστερικοί δεσµοί πολυσακχαρίτες λιπαρά οξέα
9. Σχεδιάστε την πολυνουκλεοτιδική αλυσίδα της Εικόνας5.27 και σηµειώστε τις βάσεις G, T, C και T, ξεκινώνταςαπό το άκρο 5´. Στη συνέχεια, σχεδιάστε τη συµπληρωµα-
τική αλυσίδα της διπλής έλικας, χρησιµοποιώντας τα ίδιασύµβολα για τις φωσφορικές οµάδες (κύκλους), τα σάκχα-ρα (πεντάγωνα) και τις βάσεις. Ονοµατίστε τις βάσεις. Σεκάθε αλυσίδα βάλτε βέλη για την κατεύθυνση 5´→3´. Βε-βαιωθείτε, µε βάση τα βέλη, ότι η δεύτερη αλυσίδα είναιαντιπαράλληλη προς την πρώτη. Συµβουλή: Αφού σχεδιά-σετε την πρώτη αλυσίδα κάθετα, γυρίστε το χαρτί (ή το τε-τράδιό) σας ανάποδα. Έτσι θα είναι ευκολότερο να σχεδιά-σετε τη δεύτερη αλυσίδα από το άκρο 5´ προς το άκρο 3´,καθώς θα προχωράτε από πάνω προς τα κάτω.
Για τις απαντήσεις των ερωτήσεων αυτοαξιολόγησης, βλ. Πα-ράρτηµα Α.
ΣΥΣΧΕΤΙΣΗ ΜΕ ΤΗΝ ΕξΕΛΙξΗ
10. Οι σχέσεις ανάµεσα στις ακολουθίες των αµινοξέων µπο-ρούν να ρίξουν φως στην εξελικτική απόκλιση ανάµεσα σεδιάφορα συγγενικά είδη. Σε δεδοµένο σύνολο ζωντανών ει-δών, θα αναµένατε όλες οι πρωτεΐνες να εµφανίζουν τονίδιο βαθµό απόκλισης; Ναι ή όχι, και γιατί;
ΕΠΙΣΤΗΜΟΝΙΚΗ ΑΝΑΖΗΤΗΣΗ
11. Κατά τους ΝαπολεόντειουςΠολέµους, στις αρχές του19ου αιώνα, σηµειώθηκε έλ-λειψη ζάχαρης στην Ευρώ-πη, επειδή τα εµπορικάπλοία δεν µπορούσαν να κα-ταπλεύσουν στα αποκλει-σµένα λιµάνια. Για να δηµι-ουργήσουν τεχνητές γλυκαν-τικές ουσίες, Γερµανοί επι-στήµονες υδρόλυσαν άµυλο από σιτάρι. Το πέτυχαν, θερ-µαίνοντας διαλύµατα του αµύλου και προσθέτοντας υδρο-χλωρικό οξύ. Με τη µέθοδο αυτή κατάφεραν να διασπά-σουν ορισµένους από τους γλυκοζιτικούς δεσµούς ανάµεσαστα µονοµερή της γλυκόζης. Το σχετικό διάγραµµα δείχνειτο ποσοστό διάσπασης των γλυκοζιτικών δεσµών µε τηνπάροδο του χρόνου. Για ποιον λόγο πιστεύετε ότι οι κατα-ναλωτές έβρισκαν αυτή τη γλυκαντική ουσία λιγότερο γλυ-κιά από τη ζάχαρη; Σχεδιάστε έναν γλυκοζιτικό δεσµό στοάµυλο µε βάση τις Εικόνες 5.5α και 5.7β. ∆είξτε πώς µπο-ρεί το οξύ να διασπά αυτόν το δεσµό. Γιατί πιστεύετε ότι τοοξύ διασπά µόνο το 50% των δεσµών στο άµυλο των σιτη-ρών;
ΕΠΙΣΤΗΜΗ, ΤΕΧΝΟΛΟΓΙΑ ΚΑΙ ΚΟΙΝΩΝΙΑ
12. Ορισµένοι ερασιτέχνες και επαγγελµατίες αθλητέςπαίρνουν αναβολικά στεροειδή για να αυξήσουν τον όγκοή τη δύναµή τους. Οι κίνδυνοι για την υγεία από µια τέτοιαπρακτική έχουν µελετηθεί εκτεταµένα και οι µελέτες έχουντεκµηριωθεί πολύ καλά. Εκτός όµως από τους κινδύνουςγια την υγεία, τι σκέφτεστε εσείς σχετικά µε τη χρήσηχηµικών ουσιών για την ενίσχυση της αθλητικήςαπόδοσης; Είναι εξαπάτηση η χρήση στεροειδώναναβολικών από έναν αθλητή ή µήπως είναι τµήµα τηςαπαραίτητης προετοιµασίας για τους αγώνες; Αναλύστετην άποψή σας.
100
Χρόνος
50
Ποσ
οστό
%τω
νγλ
υκοζ
ιτικ
ών
δεσμ
ών
που
λύον
ται
0


















