
Autophagy: molecular mechanisms and disease outcomes · 2015-11-05 · Daniel J. Klionsky and Vojo...
2
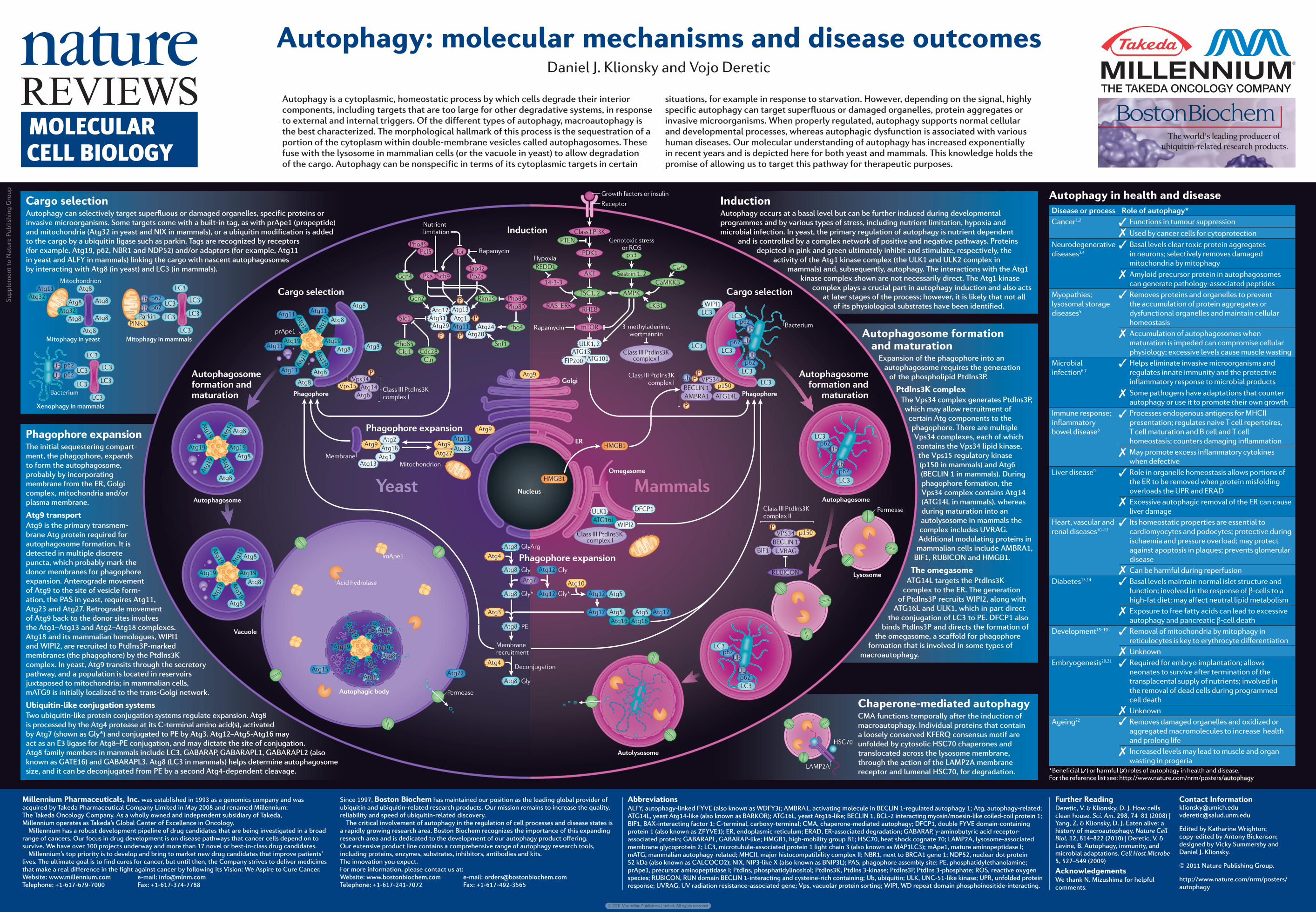
Supplement to Nature Publishing Group #VI #VI #VI #VI #VI #VI #VI #VI #VI #WVQRJCIQUQOG #WVQRJCIQUQOG 2JCIQRJQTG 1OGICUQOG #WVQN[UQUQOG .[UQUQOG 2GTOGCUG 8RU #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI 8RU 2 #VI #VI %NCUU +++ 2VF+PU- EQORNGZ + 2JCIQRJQTG RT#RG #VI 2 2 2 #VI #VI #VI #VI 0WVTKGPV NKOKVCVKQP 2JQ 2EN 6QT 2MC 5KE 5EJ 6CR 2RC 2JQ 2JQ )EP 2JQ %NI %FE %NP 5PH 2JQ #VI 4KO )EP #VI #VI 2 #VI #VI #VI #VI #VI /KVQEJQPFTKQP /GODTCPG #VI #VI #VI #VI #VI +PFWEVKQP %CTIQ UGNGEVKQP %CTIQ UGNGEVKQP 2JCIQRJQTG GZRCPUKQP 2JCIQRJQTG GZRCPUKQP #WVQRJCIQUQOG HQTOCVKQP CPF OCVWTCVKQP #WVQRJCIQUQOG HQTOCVKQP CPF OCVWTCVKQP ;GCUV 8CEWQNG 2GTOGCUG 7.- %NCUU + 2+- 2&- #-6 4*'$ O614 65% #/2- %C/--β .-$ %C 4#5s'4- 26'0 R 5GUVTKP )GPQVQZKE UVTGUU QT 415 OGVJ[NCFGPKPG YQTVOCPPKP *[RQZKC #6) #6) (+2 4CRCO[EKP 4'&& %NCUU +++ 2VF+PU- EQORNGZ + )TQYVJ HCEVQTU QT KPUWNKP 4GEGRVQT /COOCNU )QNIK '4 .% .% R R 7D .% R 7D 7D .% R 7D 2 2 2 7D 825 R $'%.+0 #6). #/$4# %NCUU +++ 2VF+PU- EQORNGZ + .% .% .% .% .% 9+2+ $CEVGTKWO R 7D R 7D .% R 7D #VI #VI #VI #VI #VI #VI #VI #VI #VI #WVQRJCIKE DQF[ #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #EKF J[FTQNCUG O#RG 0WENGWU */)$ */)$ 4CRCO[EKP R $'%.+0 784#) $+( %NCUU +++ 2VF+PU- EQORNGZ ++ 47$+%10 2 &(%2 7.- 9+2+ #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI #VI 2' )N[ )N[ )N[ #VI )N[ )N[ )N[#TI /GODTCPG TGETWKVOGPV &GEQPLWICVKQP #VI #VI #VI #VI %NCUU +++ 2VF+PU- EQORNGZ + #VI 825 #6). #VI #VI #VI #VI #VI #VI #VI #VI #VI .% .% .% .% .% .% .% .% .% .% .% .% R R 7D 7D 2CTMKP 2+0- R R 7D 7D /KVQRJCI[ KP [GCUV :GPQRJCI[ KP OCOOCNU /KVQRJCI[ KP OCOOCNU /KVQEJQPFTKQP $CEVGTKWO Autophagy in health and disease Disease or process Role of autophagy* Cancer 1,2 Functions in tumour suppression Used by cancer cells for cytoprotection Neurodegenerative diseases 3,4 Basal levels clear toxic protein aggregates in neurons; selectively removes damaged mitochondria by mitophagy Amyloid precursor protein in autophagosomes can generate pathology-associated peptides Myopathies; lysosomal storage diseases 5 Removes proteins and organelles to prevent the accumulation of protein aggregates or dysfunctional organelles and maintain cellular homeostasis Accumulation of autophagosomes when maturation is impeded can compromise cellular physiology; excessive levels cause muscle wasting Microbial infection 6,7 Helps eliminate invasive microorganisms and regulates innate immunity and the protective inflammatory response to microbial products Some pathogens have adaptations that counter autophagy or use it to promote their own growth Immune response; inflammatory bowel disease 8 Processes endogenous antigens for MHCII presentation; regulates naive T cell repertoires, T cell maturation and B cell and T cell homeostasis; counters damaging inflammation May promote excess inflammatory cytokines when defective Liver disease 9 Role in organelle homeostasis allows portions of the ER to be removed when protein misfolding overloads the UPR and ERAD Excessive autophagic removal of the ER can cause liver damage Heart, vascular and renal diseases 10–12 Its homeostatic properties are essential to cardiomyocytes and podocytes; protective during ischaemia and pressure overload; may protect against apoptosis in plaques; prevents glomerular disease Can be harmful during reperfusion Diabetes 13,14 Basal levels maintain normal islet structure and function; involved in the response of β-cells to a high-fat diet; may affect neutral lipid metabolism Exposure to free fatty acids can lead to excessive autophagy and pancreatic β-cell death Development 15–19 Removal of mitochondria by mitophagy in reticulocytes is key to erythrocyte differentiation Unknown Embryogenesis 20,21 Required for embryo implantation; allows neonates to survive after termination of the transplacental supply of nutrients; involved in the removal of dead cells during programmed cell death Unknown Ageing 22 Removes damaged organelles and oxidized or aggregated macromolecules to increase health and prolong life Increased levels may lead to muscle and organ wasting in progeria *Beneficial ( ) or harmful ( ) roles of autophagy in health and disease. For the reference list see: http://www.nature.com/nrm/poster s/autophagy .#/2# *5% Autophagy: molecular mechanisms and disease outcomes Daniel J. Klionsky and Vojo Deretic Autophagy is a cytoplasmic, homeostatic process by which cells degrade their interior components, including targets that are too large for other degradative systems, in response to external and internal triggers. Of the different types of autophagy, macroautophagy is the best characterized. The morphological hallmark of this process is the sequestration of a portion of the cytoplasm within double-membrane vesicles called autophagosomes. These fuse with the lysosome in mammalian cells (or the vacuole in yeast) to allow degradation of the cargo. Autophagy can be nonspecific in terms of its cytoplasmic targets in certain situations, for example in response to starvation. However, depending on the signal, highly specific autophagy can target superfluous or damaged organelles, protein aggregates or invasive microorganisms. When properly regulated, autophagy supports normal cellular and developmental processes, whereas autophagic dysfunction is associated with various human diseases. Our molecular understanding of autophagy has increased exponentially in recent years and is depicted here for both yeast and mammals. This knowledge holds the promise of allowing us to target this pathway for therapeutic purposes. Millennium Pharmaceuticals, Inc. was established in 1993 as a genomics company and was acquired by Takeda Pharmaceutical Company Limited in May 2008 and renamed Millennium: The Takeda Oncology Company. As a wholly owned and independent subsidiary of Takeda, Millennium operates as Takeda’s Global Center of Excellence in Oncology. Millennium has a robust development pipeline of drug candidates that are being investigated in a broad range of cancers. Our focus in drug development is on disease pathways that cancer cells depend on to survive. We have over 300 projects underway and more than 17 novel or best-in-class drug candidates. Millennium’s top priority is to develop and bring to market new drug candidates that improve patients’ lives. The ultimate goal is to find cures for cancer, but until then, the Company strives to deliver medicines that make a real difference in the fight against cancer by following its Vision: We Aspire to Cure Cancer. Website: www.millennium.com e-mail: [email protected] Telephone: +1-617-679-7000 Fax: +1-617-374-7788 Since 1997, Boston Biochem has maintained our position as the leading global provider of ubiquitin and ubiquitin-related research products. Our mission remains to increase the quality, reliability and speed of ubiquitin-related discovery. The critical involvement of autophagy in the regulation of cell processes and disease states is a rapidly growing research area. Boston Biochem recognizes the importance of this expanding research area and is dedicated to the development of our autophagy product offering. Our extensive product line contains a comprehensive range of autophagy research tools, including proteins, enzymes, substrates, inhibitors, antibodies and kits. The innovation you expect. For more information, please contact us at: Website: www.bostonbiochem.com e-mail: [email protected] Telephone: +1-617-241-7072 Fax: +1-617-492-3565 Abbreviations ALFY, autophagy-linked FYVE (also known as WDFY3); AMBRA1, activating molecule in BECLIN 1-regulated autophagy 1; Atg, autophagy-related; ATG14L, yeast Atg14-like (also known as BARKOR); ATG16L, yeast Atg16-like; BECLIN 1, BCL-2 interacting myosin/moesin-like coiled-coil protein 1; BIF1, BAX-interacting factor 1; C-terminal, carboxy-terminal; CMA, chaperone-mediated autophagy; DFCP1, double FYVE domain-containing protein 1 (also known as ZFYVE1); ER, endoplasmic reticulum; ERAD, ER-associated degradation; GABARAP, γ-aminobutyric acid receptor- associated protein; GABARAPL, GABARAP-like; HMGB1, high-mobility group B1; HSC70, heat shock cognate 70; LAMP2A, lysosome-associated membrane glycoprotein 2; LC3, microtubule-associated protein 1 light chain 3 (also known as MAP1LC3); mApe1, mature aminopeptidase I; mATG, mammalian autophagy-related; MHCII, major histocompatibility complex II; NBR1, next to BRCA1 gene 1; NDP52, nuclear dot protein 52 kDa (also known as CALCOCO2); NIX, NIP3-like X (also known as BNIP3L); PAS, phagophore assembly site; PE, phosphatidylethanolamine; prApe1, precursor aminopeptidase I; PtdIns, phosphatidylinositol; PtdIns3K, PtdIns 3-kinase; PtdIns3P, PtdIns 3-phosphate; ROS, reactive oxygen species; RUBICON, RUN domain BECLIN 1-interacting and cysteine-rich containing; Ub, ubiquitin; ULK, UNC-51-like kinase; UPR, unfolded protein response; UVRAG, UV radiation resistance-associated gene; Vps, vacuolar protein sorting; WIPI, WD repeat domain phosphoinositide-interacting. Further Reading Deretic, V. & Klionsky, D. J. How cells clean house. Sci. Am. 298, 74–81 (2008) | Yang, Z. & Klionsky, D. J. Eaten alive: a history of macroautophagy. Nature Cell Biol. 12, 814–822 (2010) | Deretic, V. & Levine, B. Autophagy, immunity, and microbial adaptations. Cell Host Microbe 5, 527–549 (2009) Acknowledgements We thank N. Mizushima for helpful comments. Contact Information [email protected] [email protected] Edited by Katharine Wrighton; copy-edited by Antony Bickenson; designed by Vicky Summersby and Daniel J. Klionsky. 2011 Nature Publishing Group. http://www.nature.com/nrm/posters/ autophagy Induction Autophagy occurs at a basal level but can be further induced during developmental programmes and by various types of stress, including nutrient limitation, hypoxia and microbial infection. In yeast, the primary regulation of autophagy is nutrient dependent and is controlled by a complex network of positive and negative pathways. Proteins depicted in pink and green ultimately inhibit and stimulate, respectively, the activity of the Atg1 kinase complex (the ULK1 and ULK2 complex in mammals) and, subsequently, autophagy. The interactions with the Atg1 kinase complex shown are not necessarily direct. The Atg1 kinase complex plays a crucial part in autophagy induction and also acts at later stages of the process; however, it is likely that not all of its physiological substrates have been identified. Cargo selection Autophagy can selectively target superfluous or damaged organelles, specific proteins or invasive microorganisms. Some targets come with a built-in tag, as with prApe1 (propeptide) and mitochondria (Atg32 in yeast and NIX in mammals), or a ubiquitin modification is added to the cargo by a ubiquitin ligase such as parkin. Tags are recognized by receptors (for example, Atg19, p62, NBR1 and NDP52) and/or adaptors (for example, Atg11 in yeast and ALFY in mammals) linking the cargo with nascent autophagosomes by interacting with Atg8 (in yeast) and LC3 (in mammals). Phagophore expansion The initial sequestering compart- ment, the phagophore, expands to form the autophagosome, probably by incorporating membrane from the ER, Golgi complex, mitochondria and/or plasma membrane. Atg9 transport Atg9 is the primary transmem- brane Atg protein required for autophagosome formation. It is detected in multiple discrete puncta, which probably mark the donor membranes for phagophore expansion. Anterograde movement of Atg9 to the site of vesicle form- ation, the PAS in yeast, requires Atg11, Atg23 and Atg27. Retrograde movement of Atg9 back to the donor sites involves the Atg1–Atg13 and Atg2–Atg18 complexes. Atg18 and its mammalian homologues, WIPI1 and WIPI2, are recruited to PtdIns3P-marked membranes (the phagophore) by the PtdIns3K complex. In yeast, Atg9 transits through the secretory pathway, and a population is located in reservoirs juxtaposed to mitochondria; in mammalian cells, mATG9 is initially localized to the trans-Golgi network. Ubiquitin-like conjugation systems Two ubiquitin-like protein conjugation systems regulate expansion. Atg8 is processed by the Atg4 protease at its C-terminal amino acid(s), activated by Atg7 (shown as Gly*) and conjugated to PE by Atg3. Atg12–Atg5-Atg16 may act as an E3 ligase for Atg8–PE conjugation, and may dictate the site of conjugation. Atg8 family members in mammals include LC3, GABARAP, GABARAPL1, GABARAPL2 (also known as GATE16) and GABARAPL3. Atg8 (LC3 in mammals) helps determine autophagosome size, and it can be deconjugated from PE by a second Atg4-dependent cleavage. Autophagosome formation and maturation Expansion of the phagophore into an autophagosome requires the generation of the phospholipid PtdIns3P. PtdIns3K complex The Vps34 complex generates PtdIns3P, which may allow recruitment of certain Atg components to the phagophore. There are multiple Vps34 complexes, each of which contains the Vps34 lipid kinase, the Vps15 regulatory kinase (p150 in mammals) and Atg6 (BECLIN 1 in mammals). During phagophore formation, the Vps34 complex contains Atg14 (ATG14L in mammals), whereas during maturation into an autolysosome in mammals the complex includes UVRAG. Additional modulating proteins in mammalian cells include AMBRA1, BIF1, RUBICON and HMGB1. The omegasome ATG14L targets the PtdIns3K complex to the ER. The generation of PtdIns3P recruits WIPI2, along with ATG16L and ULK1, which in part direct the conjugation of LC3 to PE. DFCP1 also binds PtdIns3P and directs the formation of the omegasome, a scaffold for phagophore formation that is involved in some types of macroautophagy. Chaperone-mediated autophagy CMA functions temporally after the induction of macroautophagy. Individual proteins that contain a loosely conserved KFERQ consensus motif are unfolded by cytosolic HSC70 chaperones and translocated across the lysosome membrane, through the action of the LAMP2A membrane receptor and lumenal HSC70, for degradation. MOLECULAR CELL BIOLOGY © 2011 Macmillan Publishers Limited. All rights reserved
Transcript of Autophagy: molecular mechanisms and disease outcomes · 2015-11-05 · Daniel J. Klionsky and Vojo...
Supp
lem
ent
to N
atur
e Pu
blis
hing
Gro
up
β
Autophagy in health and diseaseDisease or process Role of autophagy*Cancer1,2 Functions in tumour suppression
Used by cancer cells for cytoprotectionNeurodegenerative diseases3,4
Basal levels clear toxic protein aggregates in neurons; selectively removes damaged mitochondria by mitophagyAmyloid precursor protein in autophagosomes can generate pathology-associated peptides
Myopathies; lysosomal storage diseases5
Removes proteins and organelles to prevent the accumulation of protein aggregates or dysfunctional organelles and maintain cellular homeostasisAccumulation of autophagosomes when maturation is impeded can compromise cellular physiology; excessive levels cause muscle wasting
Microbial infection6,7
Helps eliminate invasive microorganisms and regulates innate immunity and the protective inflammatory response to microbial products Some pathogens have adaptations that counter autophagy or use it to promote their own growth
Immune response; inflammatory bowel disease8
Processes endogenous antigens for MHCII presentation; regulates naive T cell repertoires, T cell maturation and B cell and T cell homeostasis; counters damaging inflammationMay promote excess inflammatory cytokines when defective
Liver disease9 Role in organelle homeostasis allows portions of the ER to be removed when protein misfolding overloads the UPR and ERADExcessive autophagic removal of the ER can cause liver damage
Heart, vascular and renal diseases10–12
Its homeostatic properties are essential to cardiomyocytes and podocytes; protective during ischaemia and pressure overload; may protect against apoptosis in plaques; prevents glomerular diseaseCan be harmful during reperfusion
Diabetes13,14 Basal levels maintain normal islet structure and function; involved in the response of β-cells to a high-fat diet; may affect neutral lipid metabolismExposure to free fatty acids can lead to excessive autophagy and pancreatic β-cell death
Development15–19 Removal of mitochondria by mitophagy in reticulocytes is key to erythrocyte differentiation Unknown
Embryogenesis20,21 Required for embryo implantation; allows neonates to survive after termination of the transplacental supply of nutrients; involved in the removal of dead cells during programmed cell deathUnknown
Ageing22 Removes damaged organelles and oxidized or aggregated macromolecules to increase health and prolong lifeIncreased levels may lead to muscle and organ wasting in progeria
*Beneficial ( ) or harmful ( ) roles of autophagy in health and disease. For the reference list see: http://www.nature.com/nrm/posters/autophagy
Autophagy: molecular mechanisms and disease outcomesDaniel J. Klionsky and Vojo Deretic
Autophagy is a cytoplasmic, homeostatic process by which cells degrade their interior components, including targets that are too large for other degradative systems, in response to external and internal triggers. Of the different types of autophagy, macroautophagy is the best characterized. The morphological hallmark of this process is the sequestration of a portion of the cytoplasm within double-membrane vesicles called autophagosomes. These fuse with the lysosome in mammalian cells (or the vacuole in yeast) to allow degradation of the cargo. Autophagy can be nonspecific in terms of its cytoplasmic targets in certain
situations, for example in response to starvation. However, depending on the signal, highly specific autophagy can target superfluous or damaged organelles, protein aggregates or invasive microorganisms. When properly regulated, autophagy supports normal cellular and developmental processes, whereas autophagic dysfunction is associated with various human diseases. Our molecular understanding of autophagy has increased exponentially in recent years and is depicted here for both yeast and mammals. This knowledge holds the promise of allowing us to target this pathway for therapeutic purposes.
Millennium Pharmaceuticals, Inc. was established in 1993 as a genomics company and was acquired by Takeda Pharmaceutical Company Limited in May 2008 and renamed Millennium: The Takeda Oncology Company. As a wholly owned and independent subsidiary of Takeda, Millennium operates as Takeda’s Global Center of Excellence in Oncology.
Millennium has a robust development pipeline of drug candidates that are being investigated in a broad range of cancers. Our focus in drug development is on disease pathways that cancer cells depend on to survive. We have over 300 projects underway and more than 17 novel or best-in-class drug candidates.
Millennium’s top priority is to develop and bring to market new drug candidates that improve patients’ lives. The ultimate goal is to find cures for cancer, but until then, the Company strives to deliver medicines that make a real difference in the fight against cancer by following its Vision: We Aspire to Cure Cancer.Website: www.millennium.com e-mail: [email protected]: +1-617-679-7000 Fax: +1-617-374-7788
Since 1997, Boston Biochem has maintained our position as the leading global provider of ubiquitin and ubiquitin-related research products. Our mission remains to increase the quality, reliability and speed of ubiquitin-related discovery.
The critical involvement of autophagy in the regulation of cell processes and disease states is a rapidly growing research area. Boston Biochem recognizes the importance of this expanding research area and is dedicated to the development of our autophagy product offering. Our extensive product line contains a comprehensive range of autophagy research tools, including proteins, enzymes, substrates, inhibitors, antibodies and kits.The innovation you expect.For more information, please contact us at:Website: www.bostonbiochem.com e-mail: [email protected]: +1-617-241-7072 Fax: +1-617-492-3565
Abbreviations ALFY, autophagy-linked FYVE (also known as WDFY3); AMBRA1, activating molecule in BECLIN 1-regulated autophagy 1; Atg, autophagy-related; ATG14L, yeast Atg14-like (also known as BARKOR); ATG16L, yeast Atg16-like; BECLIN 1, BCL-2 interacting myosin/moesin-like coiled-coil protein 1; BIF1, BAX-interacting factor 1; C-terminal, carboxy-terminal; CMA, chaperone-mediated autophagy; DFCP1, double FYVE domain-containing protein 1 (also known as ZFYVE1); ER, endoplasmic reticulum; ERAD, ER-associated degradation; GABARAP, γ-aminobutyric acid receptor-associated protein; GABARAPL, GABARAP-like; HMGB1, high-mobility group B1; HSC70, heat shock cognate 70; LAMP2A, lysosome-associated membrane glycoprotein 2; LC3, microtubule-associated protein 1 light chain 3 (also known as MAP1LC3); mApe1, mature aminopeptidase I; mATG, mammalian autophagy-related; MHCII, major histocompatibility complex II; NBR1, next to BRCA1 gene 1; NDP52, nuclear dot protein 52 kDa (also known as CALCOCO2); NIX, NIP3-like X (also known as BNIP3L); PAS, phagophore assembly site; PE, phosphatidylethanolamine; prApe1, precursor aminopeptidase I; PtdIns, phosphatidylinositol; PtdIns3K, PtdIns 3-kinase; PtdIns3P, PtdIns 3-phosphate; ROS, reactive oxygen species; RUBICON, RUN domain BECLIN 1-interacting and cysteine-rich containing; Ub, ubiquitin; ULK, UNC-51-like kinase; UPR, unfolded protein response; UVRAG, UV radiation resistance-associated gene; Vps, vacuolar protein sorting; WIPI, WD repeat domain phosphoinositide-interacting.
Further ReadingDeretic, V. & Klionsky, D. J. How cells clean house. Sci. Am. 298, 74–81 (2008) | Yang, Z. & Klionsky, D. J. Eaten alive: a history of macroautophagy. Nature Cell Biol. 12, 814–822 (2010) | Deretic, V. & Levine, B. Autophagy, immunity, and microbial adaptations. Cell Host Microbe 5, 527–549 (2009)
AcknowledgementsWe thank N. Mizushima for helpful comments.
Contact [email protected] [email protected]
Edited by Katharine Wrighton; copy-edited by Antony Bickenson; designed by Vicky Summersby and Daniel J. Klionsky.
2011 Nature Publishing Group.
http://www.nature.com/nrm/posters/autophagy
InductionAutophagy occurs at a basal level but can be further induced during developmental programmes and by various types of stress, including nutrient limitation, hypoxia and microbial infection. In yeast, the primary regulation of autophagy is nutrient dependent
and is controlled by a complex network of positive and negative pathways. Proteins depicted in pink and green ultimately inhibit and stimulate, respectively, the
activity of the Atg1 kinase complex (the ULK1 and ULK2 complex in mammals) and, subsequently, autophagy. The interactions with the Atg1
kinase complex shown are not necessarily direct. The Atg1 kinase complex plays a crucial part in autophagy induction and also acts
at later stages of the process; however, it is likely that not all of its physiological substrates have been identified.
Cargo selectionAutophagy can selectively target superfluous or damaged organelles, specific proteins or invasive microorganisms. Some targets come with a built-in tag, as with prApe1 (propeptide) and mitochondria (Atg32 in yeast and NIX in mammals), or a ubiquitin modification is added to the cargo by a ubiquitin ligase such as parkin. Tags are recognized by receptors (for example, Atg19, p62, NBR1 and NDP52) and/or adaptors (for example, Atg11 in yeast and ALFY in mammals) linking the cargo with nascent auto phagosomes by interacting with Atg8 (in yeast) and LC3 (in mammals).
Phagophore expansionThe initial sequestering compart-ment, the phagophore, expands to form the autophagosome, probably by incorporating membran e from the ER, Golgi complex, mitochondria and/or plasm a membrane.
Atg9 transportAtg9 is the primary transmem-brane Atg protein required for autophagosome formation. It is detected in multiple discrete puncta, which probably mark the donor membranes for phagophore expansion. Anterograde movement of Atg9 to the site of vesicle form-atio n, the PAS in yeast, requires Atg11, Atg23 and Atg27. Retrograde move ment of Atg9 back to the donor sites involves the Atg1–Atg13 and Atg2–Atg18 complexes. Atg18 and its mammalian homologues, WIPI1 and WIPI2, are recruited to PtdIns3P-marked membranes (the phagophore) by the PtdIns3K complex. In yeast, Atg9 transits through the secretory pathway, and a population is located in reservoirs juxtaposed to mitochondria; in mammalian cells, mATG9 is initially localized to the trans-Golgi network.
Ubiquitin-like conjugation systemsTwo ubiquitin-like protein conjugation systems regulate expansion. Atg8 is processed by the Atg4 protease at its C-terminal amino acid(s), activated by Atg7 (shown as Gly*) and conjugated to PE by Atg3. Atg12–Atg5-Atg16 may act as an E3 ligase for Atg8–PE conjugation, and may dictate the site of conjugation. Atg8 family members in mammals include LC3, GABARAP, GABARAPL1, GABARAPL2 (also known as GATE16) and GABARAPL3. Atg8 (LC3 in mammals) helps determine autophagosome size, and it can be deconjugated from PE by a second Atg4-dependent cleavage.
Autophagosome formation and maturation
Expansion of the phagophore into an autophagosome requires the generation
of the phospholipid PtdIns3P.
PtdIns3K complexThe Vps34 complex generates PtdIns3P,
which may allow recruitment of certain Atg components to the phagophore. There are multiple Vps34 complexes, each of which contains the Vps34 lipid kinase, the Vps15 regulatory kinase (p150 in mammals) and Atg6 (BECLIN 1 in mammals). During phagophore formation, the Vps34 complex contains Atg14 (ATG14L in mammals), whereas during maturation into an autolysosome in mammals the complex includes UVRAG. Additional modulating proteins in
mammalian cells include AMBRA1, BIF1, RUBICON and HMGB1.
The omegasomeATG14L targets the PtdIns3K
complex to the ER. The generation of PtdIns3P recruits WIPI2, along with
ATG16L and ULK1, which in part direct the conjugation of LC3 to PE. DFCP1 also
binds PtdIns3P and directs the formation of the omegasome, a scaffold for phagophore
formation that is involved in some types of macroautophagy.
Chaperone-mediated autophagy CMA functions temporally after the induction of macroautophagy. Individual proteins that contain a loosely conserved KFERQ consensus motif are unfolded by cytosolic HSC70 chaperones and translocated across the lysosome membrane, through the action of the LAMP2A membrane receptor and lumenal HSC70, for degradation.
MOLECULAR CELL BIOLOGY
© 2011 Macmillan Publishers Limited. All rights reserved

Supplementary information to poster | Autophagy: molecular mechanisms and disease outcomes.Daniel J. Klionsky and Vojo DereticApril 2010 (http://www.nature.com/nrm/posters/autophagy)
References for Table on Autophagy in health and disease1. Dikic, I., Johansen, T. & Kirkin, V. Selective autophagy in
cancer development and therapy. Cancer Res. 70, 3431–3434 (2010).
2. White, E., Karp, C., Strohecker, A. M., Guo, Y. & Mathew, R. Role of autophagy in suppression of inflammation and cancer. Curr. Opin. Cell Biol. 22, 212–217 (2010).
3. Rubinsztein, D. C. et al. Autophagy and its possible roles in nervous system diseases, damage and repair. Autophagy 1, 11–22 (2005).
4. Irrcher, I. & Park, D. S. Parkinson’s disease: to live or die by autophagy. Sci. Signal. 2, pe21 (2009).
5. Malicdan, M. C., Noguchi, S., Nonaka, I., Saftig, P. & Nishino, I. Lysosomal myopathies: an excessive build-up in autophagosomes is too much to handle. Neuromuscul. Disord. 18, 521–529 (2008).
6. Deretic, V. & Levine, B. Autophagy, immunity, and microbial adaptations. Cell Host Microbe 5, 527–549 (2009).
7. Levine, B. & Deretic, V. Unveiling the roles of autophagy in innate and adaptive immunity. Nature Rev. Immunol. 7, 767–777 (2007).
8. Brest, P. et al. Autophagy and Crohn’s disease: at the crossroads of infection, inflammation, immunity, and cancer. Curr. Mol. Med. 10, 486–502 (2010).
9. Greene, C. M. et al. Alpha-1 antitrypsin deficiency: a conformational disease associated with lung and liver manifestations. J. Inherit. Metab. Dis. 31, 21–34 (2008).
10. Nishida, K., Kyoi, S., Yamaguchi, O., Sadoshima, J. & Otsu, K. The role of autophagy in the heart. Cell Death Differ. 16, 31–38 (2009).
11. Gustafsson, A. B. & Gottlieb, R. A. Eat your heart out: role of autophagy in myocardial ischemia/reperfusion. Autophagy 4, 416–421 (2008).
12. Periyasamy-Thandavan, S., Jiang, M., Schoenlein, P. & Dong, Z. Autophagy: molecular machinery, regulation, and implications for renal pathophysiology. Am. J. Physiol. Renal Physiol. 297, F244–F256 (2009).
13. Gonzalez, C. D. et al. The emerging role of autophagy in the pathophysiology of diabetes mellitus. Autophagy 7, 2–11 (2011).
14. Chen, Z. F. et al. The double-edged effect of autophagy in pancreatic beta cells and diabetes. Autophagy 7, 12–16 (2010).
15. Mizushima, N. & Levine, B. Autophagy in mammalian development and differentiation. Nature Cell Biol. 12, 823–830 (2010).
16. Levine, B. & Klionsky, D. J. Development by self-digestion: molecular mechanisms and biological functions of autophagy. Dev. Cell 6, 463–477 (2004).
17. Qu, X. et al. Autophagy gene-dependent clearance of apoptotic cells during embryonic development. Cell 128, 931–946 (2007).
18. Zhang, J. & Ney, P. A. Reticulocyte mitophagy: monitoring mitochondrial clearance in a mammalian model. Autophagy 6, 405–408 (2010).
19. Kundu, M. et al. Ulk1 plays a critical role in the autophagic clearance of mitochondria and ribosomes during reticulocyte maturation. Blood 112, 1493–1502 (2008).
20. Tsukamoto, S., Kuma, A. & Mizushima, N. The role of autophagy during the oocyte-to-embryo transition. Autophagy 4, 1076–1078 (2008).
21. Kuma, A. et al. The role of autophagy during the early neonatal starvation period. Nature 432, 1032–1036 (2004).
22. Cuervo, A. M. Autophagy and aging: keeping that old broom working. Trends Genet. 24, 604–612 (2008).
NATURE REVIEWS | MOLECULAR CELL BIOLOGY www.nature.com/reviews/molcellbio
SUPPLEMENTARY INFORMATION
© 2011 Macmillan Publishers Limited. All rights reserved


















