
Assaying kinase activity of the TPL-2/NF-κB1 p105/ABIN-2 ... · encouraged to use the Version of...
40
Assaying kinase activity of the TPL-2/NF-κB1 p105/ABIN-2 complex using an optimal peptide substrate Sandra Kümper 1 *, Thorsten Gantke 3 *, Chao-Sheng Chen 3 *, Yasmina Soneji 3 , Michael J. Pattison, Probir Chakravarty 4 , Svend Kjær 5 , Daniel Thomas 2 , Carl Haslam 2 , Bill J. Leavens 2 , David House 1 , David J. Powell 1 & Steven C. Ley 3 1 Crick-GSK Biomedical LinkLabs & 2 RD Platform Technology & Science GlaxoSmithKline Stevenage, U.K. SG1 2NY & 3 Immune Cell Signalling Laboratory The Francis Crick Institute London, U.K. NW1 1AT & 4 Bioinformatics & Biostatistics and 5 Structural Biology Science Technology Platforms The Francis Crick Institute London, U.K. NW1 1AT * These authors contributed equally to this work Corresponding author: [email protected] Short title: Kinase assay of TPL-2/ NF-B1 p105/ABIN-2 complex Key words: ABIN-2; high-throughput assay; inflammation; NF-B1; TPL-2; inhibitor ACCEPTED MANUSCRIPT 10.1042/BCJ20170579 . Please cite using the DOI 10.1042/BCJ20170579 http://dx.doi.org/ up-to-date version is available at encouraged to use the Version of Record that, when published, will replace this version. The most this is an Accepted Manuscript, not the final Version of Record. You are : Biochemical Journal ). http://www.portlandpresspublishing.com/content/open-access-policy#Archiving Archiving Policy of Portland Press ( which the article is published. Archiving of non-open access articles is permitted in accordance with the Use of open access articles is permitted based on the terms of the specific Creative Commons Licence under
Transcript of Assaying kinase activity of the TPL-2/NF-κB1 p105/ABIN-2 ... · encouraged to use the Version of...

Assaying kinase activity of the TPL-2/NF-κB1 p105/ABIN-2 complex
using an optimal peptide substrate
Sandra Kümper1*, Thorsten Gantke3*, Chao-Sheng Chen3*, Yasmina Soneji3,
Michael J. Pattison, Probir Chakravarty4, Svend Kjær5, Daniel Thomas2, Carl
Haslam2, Bill J. Leavens2, David House1, David J. Powell1 & Steven C. Ley3
1Crick-GSK Biomedical LinkLabs & 2RD Platform Technology & Science
GlaxoSmithKline
Stevenage, U.K.
SG1 2NY
&
3Immune Cell Signalling Laboratory
The Francis Crick Institute
London, U.K.
NW1 1AT
&
4Bioinformatics & Biostatistics and 5Structural Biology
Science Technology Platforms
The Francis Crick Institute
London, U.K.
NW1 1AT
* These authors contributed equally to this work
Corresponding author: [email protected]
Short title:
Kinase assay of TPL-2/ NF-B1 p105/ABIN-2 complex
Key words:
ABIN-2; high-throughput assay; inflammation; NF-B1; TPL-2; inhibitor
AC
CE
PT
ED
MA
NU
SC
RIP
T
10.1042/BCJ20170579. Please cite using the DOI 10.1042/BCJ20170579http://dx.doi.org/up-to-date version is available at
encouraged to use the Version of Record that, when published, will replace this version. The most this is an Accepted Manuscript, not the final Version of Record. You are:Biochemical Journal
). http://www.portlandpresspublishing.com/content/open-access-policy#ArchivingArchiving Policy of Portland Press (which the article is published. Archiving of non-open access articles is permitted in accordance with the Use of open access articles is permitted based on the terms of the specific Creative Commons Licence under

TPL-2 complex assay
2
ABSTRACT
The MKK1/2 kinase TPL-2 is critical for the production of Tumour Necrosis Factor
α (TNF) in innate immune responses and a potential anti-inflammatory drug
target. Several earlier pharmaceutical company screens with the isolated TPL-2
kinase domain have identified small molecule inhibitors that specifically block
TPL-2 signalling in cells but none of these have progressed to clinical
development. We have previously shown that TPL-2 catalytic activity regulates
TNF production by macrophages while associated with NF-κB1 p105 and ABIN-2,
independently of MKK1/2 phosphorylation via an unknown downstream substrate.
In this study, we used a positional scanning peptide library to determine the
optimal substrate specificity of a complex of TPL-2, NF-B1 p105 and ABIN-2.
Using an optimal peptide substrate based on this screen and a high-throughput
mass spectrometry assay to monitor kinase activity, we found that the TPL-2
complex has significantly altered sensitivities versus existing ATP competitive
TPL-2 inhibitors than the isolated TPL-2 kinase domain. These results imply that
screens with the more physiologically relevant TPL-2/NF-B1 p105/ABIN-2
complex have the potential to deliver novel TPL-2 chemical series; both ATP
competitive and allosteric inhibitors could emerge with significantly improved
prospects for development as anti-inflammatory drugs.

TPL-2 complex assay
3
INTRODUCTION
The pro-inflammatory cytokine tumour necrosis factor α (TNF) plays an important
role in the pathogenesis of multiple inflammatory diseases and biological agents
that block TNF activity have been successfully used to treat rheumatoid arthritis
(RA), Crohn’s disease, psoriatic arthritis and ankylosing spondylitis [1]. However,
such antibody drugs are expensive, require repeated injection and are only
effective in a fraction of patients. There is therefore a need for more effective,
less expensive and, ideally, orally active drugs to suppress the production of TNF
by the innate immune system [2].
In inflammatory responses to pathogen infection, macrophages are a major
source of TNF, triggered by stimulation of pattern recognition receptors. These
include toll-like receptors (TLRs) such as TLR4, which responds to
lipopolysaccharide (LPS), a surface component of gram-negative bacteria [3].
Tumour Progression Locus 2 (TPL-2) kinase is a critical regulator of TNF
production by TLR4-stimulated macrophages. Production of TNF after LPS
injection is substantially reduced in TPL-2-deficient mice, which are resistant to
septic shock induction [4]. Analyses of Tpl2-/- mice have also indicated key pro-
inflammatory roles for TPL-2 in a number of other disease models, including
inflammatory bowel disease, pancreatitis, liver fibrosis and experimental
autoimmune encephalomyelitis [5-7]. The importance of TPL-2 in inflammation
and its critical role in regulating production of the key pro-inflammatory cytokine
TNF have made TPL-2 an attractive target for development of novel anti-
inflammatory drugs [8].
TPL-2 is a MKK 1/2 kinase, which mediates activation of ERK1/2 MAP kinases by
TLR4 (and all other TLRs), as well as the receptors for TNF and IL-1, in
macrophages [5]. Several pharmaceutical companies have carried out screens
using the isolated TPL-2 kinase domain and MKK1 substrate to identify inhibitors
that block TPL-2 activation of ERK1/2 in cultured macrophages [8, 9]. However,
none of these have progressed to clinical trial, likely reflecting the challenge of
producing ATP competitive inhibitors with the appropriate physicochemical
characteristics required for drug development. An alternative screening paradigm
utilizing TPL-2 in it native context could provide a tractable route to clinical
development.

TPL-2 complex assay
4
In unstimulated cells, TPL-2 forms a stoichiometric complex with NF-B1 p105
and the ubiquitin-binding protein ABIN-2, which are both required for
maintenance of TPL-2 steady-state levels. Binding to p105 also prevents TPL-2
phosphorylation of MKK1/2 [10]. Consequently, TPL-2 activation of the ERK1/2
MAP kinase pathway requires TPL-2 release from p105, which is triggered by IB
kinase (IKK)-induced proteolysis of p105 by the proteasome [11-13].
Surprisingly, we have recently found that TPL-2 catalytic activity regulates TNF
production by macrophages independently of IKK-induced p105 proteolysis and
ERK1/2 activation, while still associated with p105 and ABIN-2 [13]. While the
target substrate that controls TNF production has not been identified, these
results indicate that the TPL-2/NF-B1 p105/ABIN-2 complex actively signals and
that small molecule inhibitors targeting this complex should block TNF production
and possibly other pro-inflammatory functions of TPL-2.
In the present study, we used a positional scanning peptide library to identify the
optimal peptide phosphorylation motif of recombinant TPL-2/NF-B1 p105/ABIN-2
complex [14]. Using a synthetic peptide based on this optimal motif, we
developed a high-throughput screening assay and used this to demonstrate that
the TPL-2 complex has a substantially altered sensitivity to existing TPL-2
inhibitors compared with the isolated TPL-2 kinase domain. These results suggest
that inhibitor screens using TPL-2/NF-B1 p105/ABIN-2 complex instead of the
isolated kinase domain may yield novel series of TPL-2 inhibitors.

TPL-2 complex assay
5
EXPERIMENTAL
Plasmids, peptides and inhibitors
Expression plasmids (pCDNA3 vector; Life Technologies Inc.) encoding
hexahistidine-tagged (6xHis) TPL-2 and TPL-2D270A (NCBI RefSeq:
NP_001231063.1), haemagglutinin (HA)-tagged NF-κB1 p105 and C-terminally
StrepII-tagged ABIN-2 (NCBI RefSeq NP_077285.3) have been described
previously [14, 15]. 6xHis TPL-230-404 was produced by PCR amplification and
subcloned into the pFastBac vector (Invitrogen). The pDNA3 plasmid encoding
HA-VP35 of the Ebola virus species Zaire ebolavirus (Mayinga isolate) has been
described [16].
MKK1 and TPL-2tide peptides (biotinylated at their C-termini) were synthesised
and HPLC-purified (95% purity) by GL Biochem (Shanghai, China). The MKK1
peptide, which corresponded to the activation loop of MKK1
(YAGQLIDSMANSFVGTAGKK), has been previously described [17]. TPL-2tide and
pTPL2tide used in the mass spectrometry assay were synthesised by Cambridge
Research Biochemicals. TPL-2tide (YADDDDDSFLWNAGKK) was an optimised TPL-
2 peptide substrate predicted by the optimal sequence motif. The S5 peptide
(GAFRSAIRRLAARRR-acid) was optimised from an IMAP peptide library screen
(Molecular Devices), which identified the parent peptide, FAM-
GTFRSSIRRLSTRRR-acid, as the most efficient substrate for the isolated TPL-2
kinase domain. The sequence was mutated at four of the five Ser/Thr residue
positions to generate daughter peptides with only a single phosphorylation site.
Subsequent testing identified the S5 version to be the most efficient TPL-2 kinase
domain substrate.
Abbott C41, Wyeth C1, Wyeth C34 and Wyeth C2p TPL-2 inhibitors, as described
[18-21], were synthesised according to published procedures. Proton NMR and
LC-MS spectra were in accord with published data.
Expression of recombinant TPL-2
HEK293 cells (QBI293A cells, Quantum Biotechnologies) were grown in
suspension cultures as described previously [14]. For expression of recombinant
TPL-2/NF-B1 p105/ABIN-2 complex, cells were pelleted by centrifugation and
resuspended at a density of 4.0x106 cells/mL in standard culture medium
(Pro293s-CDM medium (Lonza), supplemented with 1.5 % fetal bovine serum, 2
mM L-glutamine, 50 U/mL penicillin, and 50 U/mL streptomycin). DNA complexed

TPL-2 complex assay
6
with linear polyethyleneimine (25 kDa) at a ratio of 3:1 (w/w) was then added to
a final concentration of 2 mg DNA/mL. After 5 h, cell density was adjusted to
2.0x106 cells/mL and cells cultured for a total of 72 h. Cells were lysed in buffer A
[50 mM Tris-HCl (pH7.5), 0.5 % IGEPAL CA-630, 150 mM NaCl, 10 mM
imidazole, 10 mM Na-Fluoride, 1 mM Na-pyrophosphate, 10 mM β-
glycerophosphate, 0.5 mM tris(2-carboxyethyl)phosphine (TCEP), 10 % glycerol
plus CompleteTM Protease Inhibitor Cocktail (Roche)].
TPL-230-404 protein was produced following a published methodology [9]. Sf9 cells
were co-transfected with pFastbac virus DNA for baculovirus generation. Protein
expression was carried out in 5-liter cell cultures of Sf9 cells with plaque-purified
viruses. Compound 1 (10 μM; [9]) was added to the cell cultures at 24h to
improve recombinant TPL-230-404 protein yields. Cells were harvested 72h after
infection, pellets snap frozen and stored at -80 oC.
Protein purification
For peptide library screening, recombinant 6xHis-TPL-2/ABIN-2-StrepII/HA-p105
complex was purified by sequential affinity purification. Centrifuged lysates were
incubated with Ni-NTA (Ni2+-nitrilotriacetate)-agarose (Qiagen) for 60 min,
washed in DM buffer [50 mM Tris-HCl (pH7.5), 1.8 mM decyl β-D-
maltopyranoside (DM), 150 mM NaCl, 10 mM imidazole, 10mM Na-Fluoride, 1 mM
Na-pyrophosphate, 10 mM β-glycerophosphate, 0.5 mM tris(2-
carboxyethyl)phosphine (TCEP), and 10 % glycerol supplemented with protease
inhibitors]. Bound protein was eluted with DM buffer supplemented with 200 mM
imidazole. After adding EDTA to a final concentration of 1 mM, eluates were
subsequently incubated with StrepTactin Sepharose (GE Healthcare) to
specifically purify StrepII-ABIN-2 containing TPL-2 complexes, washed
extensively in DM buffer and bound protein eluted with 2.5 mM desthiobiotin. To
remove desthiobiotin, eluted His6-TPL-2 was then loaded on to an Ni2+-charged
His-Trap HP column (GE Healthcare), followed by extensive washing and elution
with 200 mM imidazole. Sample purity of the isolated 6xHis-TPL-2/ABIN-2-
StrepII/HA-p105 complex was quantified by measuring the infrared fluorescence
of Coomassie-stained protein SDS-PAGE gels (Odyssey infrared imaging system,
LI-COR Biosciences) and shown to be > 90%.
A modified three-step purification method for recombinant His6–TPL-2/HA–
p105/ABIN-2-StrepII complex was used for mass spectrometric assays of TPL-
2tide phosphorylation. Centrifuged lysates were applied to a StrepTRAP HP

TPL-2 complex assay
7
column (GE healthcare), pre-equilibrated with Buffer A, at 1 mL / min. Bound
protein was eluted with DM buffer containing 2.5 mM desthiobiotin and loaded on
to a Superdex 200 size-exclusion column (GE healthcare) pre-equilibrated with
Buffer B [50 mM Tris-HCl (pH 7.5), 1.8 mM DM, 150 mM NaCl, 0.5 mM TCEP and
10% glycerol]. High molecular weight fractions containing TPL-2 complex were
pooled and applied to a HiTrap Q ion-exchange column (GE healthcare). The
sample was eluted with a 1 M NaCl 10 column volume gradient in Buffer B.
Sample purity (> 90%) was assessed by Coomassie Brilliant Blue staining of
SDS-PAGE gels.
For purification of His6-TPL-230-404, Sf9 cell pellets were resuspendend in lysis
buffer [50 mM Tris-HCl (pH8), 3 mM TCEP, 400 mM NaCl, 10 % glycerol, 20 mM
imidazole, 0.05% Triton X-100, supplemented with protease inhibitors].
Compound 1 [9] (dissolved in DMSO) was added to give a final concentration of
20 μM, followed by homogenization of the cell lysate. Cleared lysates were loaded
on to a HisTrap HP column (GE Healthcare) pre-equilibrated with lysis buffer.
After extensive washing with lysis buffer supplemented with 200 mM NaCl, His6-
TPL-230-404 protein was eluted with a 20 to 300 mM imidazole gradient. Fractions
containing His6-TPL-230-404 were pooled and loaded onto a Superdex 75 size
exclusion column (GE Healthcare), pre-equilibrated with a buffer containing 50
mM Tris-HCl (pH8), 150 mM NaCl, 0.5 mM TCEP and 10% glycerol. Fractions
containing His6-TPL-230-404 were pooled and concentrated by ultrafiltration to 5
mg / ml. Sample purity, assessed by Coomassie Brilliant Blue staining of SDS-
PAGE gels, was approximately 90 %.
Peptide library screening
The positional scanning synthetic peptide library described by Hutti et al. [22]
was purchased from AnaSpec, USA (ANA62017-1 and ANA62335). Peptides were
dissolved in de-gassed DMSO and the concentration of each peptide mix adjusted
to 0.6 mM by addition of Tris-HCl (pH7.5). Peptide library screening was
performed by incubation of 30 nM recombinant TPL-2 complex with 50 μM peptide
substrate in kinase buffer (50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.01 % Brij-
35, 5 mM MnCl2, 2mM DTT, 10 μM ATP and 3 nCi / μL [γ-32P]ATP) at 30°C for 1
h. Control labeling experiments were performed in the presence of 10 μM C34
TPL-2 inhibitor [21]. After incubation, reactions were transferred onto
streptavidin-coated membranes (SAM2, Promega, USA) using a 384-well 2μl slot
pin replicator (V&P Scientific, USA). Membranes were washed two times 15min in
TBS/1% SDS, and then overnight in the same buffer. Membranes were washed a

TPL-2 complex assay
8
further three times 15 min with 2 M NaCl / 1% H3PO4 before washing briefly with
water and 96 % EtOH. Membranes were air dried and peptide labeling quantified
by phosphorimaging (Storm PhosphorImager, GE Healthcare UK) using
ImageQuant TL data analysis software..
Spot intensities from three replicate experiments were quantified using
ImageQuant TL Data Analysis software (Bio-Rad, USA) and mean values
calculated. The intensity of each spot was normalized by the mean intensity of all
spots in a single position relative to the central phosphoacceptor site to calculate
selectivity values (χi) for each amino acid in every position. Selectivity values
reflected positive or negative selection of an amino acid at one position. These
ranged from 0 to 20, indicating either full negative or positive selection,
respectively. Selectivity values of 1.0 indicated neutral selectivity. Position-
specific scoring matrices (PSSM) describing amino acid selectivity at all positions
from P-5 to P+4 were assembled and used to analyse linear sequences with
Scansite 2.0 (http:/www.scansite.mit.edu) [23]. Because of possible
overestimation of serine or threonine preference due to the presence of additional
photoacceptor residues in peptides with serine or threonine at fixed positions, a
second PSSM was assembled which disregards all positive preferences for either
serine or threonine (termed ‘No Ser/Thr’ motif). Scansite analyses linear
sequences by deriving bit scores (s) for each of 7 positions (i) on either sides of
serines and threonines present in a sequence of interest:
𝑆𝑖 =ln (𝜒𝑖)
ln 2
A raw sequence score (𝑆𝑟𝑎𝑤) is then given by
𝑆𝑟𝑎𝑤 = ∑𝑠𝑖
𝑖 (𝑖 = 1 , ≤ 15)
Which is compared to the best possible score (𝑆𝑜𝑝𝑡) to calculate a final sequence
score
𝑆𝑓 =(𝑆𝑜𝑝𝑡 − 𝑆𝑟𝑎𝑤)
𝑆𝑜𝑝𝑡
Final sequence scores describe how closely a linear sequence matches the optimal
selectivity motif described by the PSSM. Accordingly, good sequence matches will
yield scores approximating 𝑆𝑓 = 0, while neither positive nor negative selection for
a given sequence (𝑆𝑟𝑎𝑤 = 0) will result in 𝑆𝑓 = 1.0.

TPL-2 complex assay
9
Kinase assays
Phosphorylation of specific peptides by TPL-2/p105/ABIN-2 complex was
measured using either a radioactive filter binding assay or by mass spectrometry.
Filter binding: Biotinylated peptides (50 μM) were phosphorylated with 30nM
recombinant TPL-2 complex in kinase buffer [50 mM Tris-HCl (pH7.0), 0.03 %
Brij-35, 2 mM DTT, 5 mM β-glycerophosphate, 5 mM MnCl2, 5 % DMSO],
supplemented with 0.1 mM ATP and 0.02 μCi / μL [γ-32P]ATP at 30 C for the
indicated times. Reactions were stopped by addition of EDTA. Reaction mixes
were subjected to ultrafiltration (10kDa cutoff; AcroPrep 384 Filter Plate, PALL
Lifer Sciences, USA) to separate substrates and autophosphorylated kinase,
before spotting onto streptavidin-coated membranes (SAM2, Promega).
Membranes were extensively washed in TBS / 1% SDS, 2 M NaCl, and 2 M NaCl /
1% H3PO4, and peptide phosphorylation quantified by phosphorimaging (Storm
PhosphorImager, GE Healthcare UK) using ImageQuant TL data analysis
software.
RapidFireTM Mass Spectrometry: A 10 mM stock solution of TPL-2tide peptide
containing 30 % (v/v) DMSO (aq) was prepared and used as substrate. Assays
were performed in clear 384-well V-bottom polypropylene plates (Greiner
#781280). The final assay volume was 10 μL in assay buffer (10 mM Tris-HCl
pH7.2, 10 mM MgCl2, 0.05 % NaN3, 0.01 % Tween-20, 1 mM DTT; Molecular
Devices #R8155). Enzyme and peptide concentrations are given in the figure
legends. Enzymes were serially diluted and a substrate solution containing final
concentrations of 100 μM peptide and 400 μM ATP (TPL-2tide) and 10 μM peptide
and 300 μM ATP (S5 peptide) were added.
To compare compound potencies towards TPL-230-404 and TPL-2/p105/ABIN-2
using TPL-2tide, enzyme solutions were added to each well of a 384-well plate
and compounds dispensed using a HP digital dispenser D-300 (Software version
3.0.1). After a 10 min pre-incubation with compound, a substrate solution
containing final concentrations of 100 μM peptide and 400 μM ATP were added.
All assays were run for 30min at room temperature. Reactions were quenched by
addition of 50 μl of 1 % (v/v) Formic Acid (aq).
Quenched plates were sealed and centrifuged at 1550 x g for 10 min and before
analysis on a RapidFireTM high-throughput solid phase extraction (SPE) system

TPL-2 complex assay
10
(Agilent Technologies, MA, USA). The sample was aspirated directly from each
well of quenched assay plates for 600 ms, and loaded onto the RapidFire micro-
scale solid-phase C4 (Agilent type A) extraction cartridge to remove buffer salts
with RapidFire buffer A (0.1% (v/v) formic acid (aq)), for 3 s at a flow rate of 1.5
mL/min. Analytes were then co-eluted directly into the mass spectrometer source
for a further 3 s using RapidFire buffer B (0.1% (v/v) formic acid in 80 %
acetonitrile (aq)) at a flow rate of 1.25 mL/min. This was followed by a 500 ms
re-equilibration step giving a total elution cycle of 7.1 s.
TPL-2tide and the phosphorylated TPL-2tide were monitored on a Sciex API 6500
triple quadrupole mass spectrometer (Applied Biosystems, Concord, Ontario,
Canada) in positive electrospray ionisation (ESI) mode following multiple reaction
monitoring (MRM) transitions initially at (Q1 mass/Q3 mass) 913.7/91.0 Da,
953.6/904.3 Da with a scan rate of 50 ms / transition, respectively. These
masses relate to the doubly charged peptide species. S5 peptide and the
phosphorylated S5 peptide were monitored on a Sciex API 4000 triple quadrupole
mass spectrometer (Applied Biosystems, Concord, Ontario, Canada) in positive
electrospray ionisation (ESI) mode following multiple reaction monitoring (MRM)
transitions initially at (Q1 mass/Q3 mass) 586.2/120 Da, 612.9/120 Da with a
scan rate of 50 ms / transition.
Data Analysis
Peaks were integrated using the RapidFire integrator software to give peak area
counts. Linear or non-linear regression fits were carried using GraFit version 7.0.3
(Erithacus Software Ltd, Surrey, UK), using peak area counts of phosphorylated
TPL-2tide and S5 peptide for data analysis. To compare the different enzymes,
the following formula was used:
𝑇𝑃𝐿 − 2 𝑎𝑐𝑡𝑖𝑣𝑖𝑡𝑦 % = (𝑎
𝑑) × 100
In this equation, a is the product count at each individual inhibitor concentration
and d is the uninhibited product count value. IC50 data were calculated using the
following equation:
Range is the fitted uninhibited value minus the Background, and s is the Hill
coefficient slope factor. The equation assumes that the y value falls with
increasing x. For each compound multiple replicates were run for each
y =Range
1+x
IC50
æ
èç
ö
ø÷
s+ Background

TPL-2 complex assay
11
experiment and the experiment repeated 4-5 times. The IC50 values and standard
errors were generated by combining the data of all experiments.

TPL-2 complex assay
12
RESULTS AND DISCUSSION
Primary phosphorylation sequence preference of the TPL-2/NF-κB
p105/ABIN-2 complex
TPL-2 induces TNF production in LPS-stimulated macrophages independently of
IKK-induced p105 phosphorylation [13]. Consequently, this does not involve TPL-
2 phosphorylations of the MAP 2-kinases MKK1, MKK2, MKK3 or MKK6, which are
blocked by Nfkb1[SSAA] mutation [24, 25]. Furthermore, these results imply that
the key substrate regulating the production of TNF is phosphorylated by TPL-
2/ABIN-2/NF-B1 p105 complex since Nfkb1[SSAA] mutation prevents the release
of TPL-2 from p105 and ABIN-2. In an attempt to develop an assay for the
catalytic activity of complexed TPL-2, we used a positional scanning synthetic
peptide library to determine the primary sequence preference for phosphorylation
by TPL-2 associated with p105 and ABIN-2.
The peptide library consisted of 198 distinct peptide mixtures [23]
(Supplementary Figure 1). Each mixture contained an equimolar mixture of
peptides with Ser or Thr in a fixed central position. In addition, one of the 20
amino acids, or phospho-Thr or phospho-Tyr, was fixed at one position relative to
the central Ser/Thr, while all other positions contained an equimolar mixture of all
other amino acids excluding Ser, Thr and Cys. Ser or Thr residues in the fixed
positions in addition to the central phosphoacceptor site would obscure the kinase
preference for these residues, as it could not be determined which residue was
phosphorylated by autoradiography readout. Within the peptide library, all amino
acids were tested at nine positions relative to a central phosphoacceptor. Peptide
sequences were flanked by additional amino acids on either side to enhance
peptide solubility and facilitate peptide handling during library preparation. A C-
terminal biotin tag allowed substrate capture on streptavidin-coated membranes
following the reaction.
Purified TPL-2/NF-B1 p105/ABIN-2 complex was incubated for 1h with the
peptide mixtures of the oriented degenerate peptide library in kinase buffer
containing [32P]ATP. Reaction mixtures were then transferred to streptavidin
capture membranes, washed and phosphorylation visualized by autoradiography.
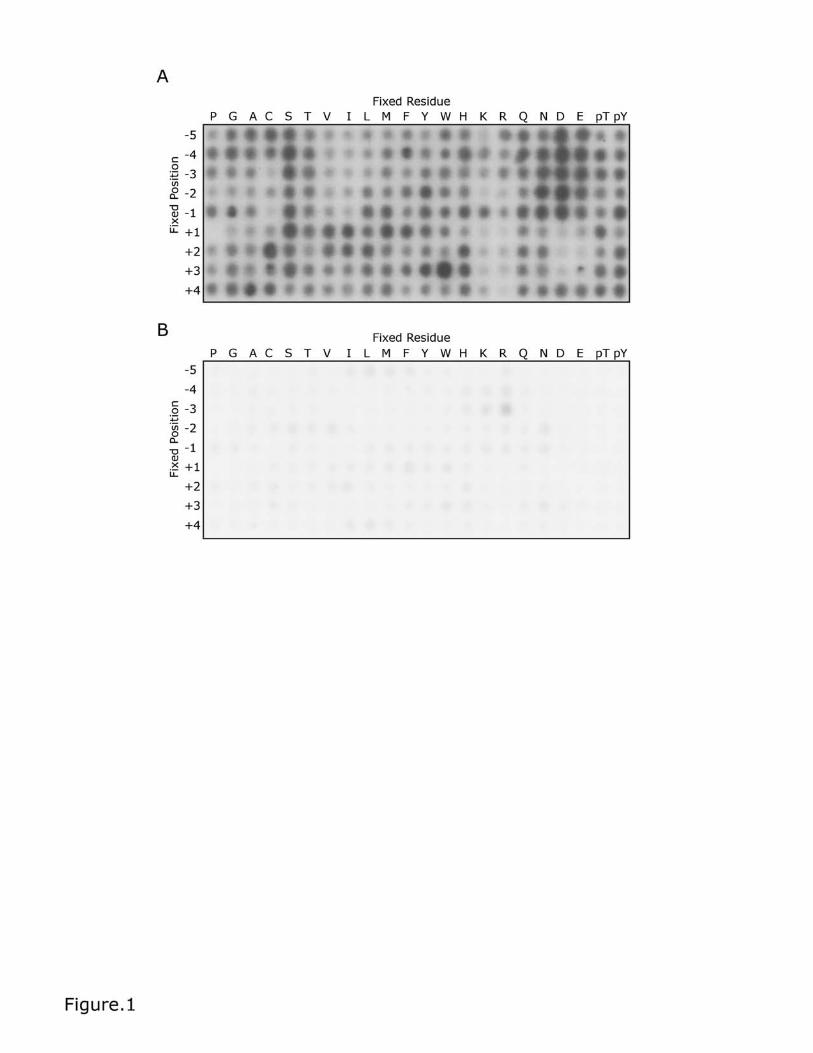
Radioactivity of varying intensity was detected in spots of most reactions (Figure
1A). The addition of 10 μM C34 TPL-2 inhibitor [21] to the reactions significantly
reduced signal intensity, confirming that the kinase assays were detecting TPL-2
activity (Figure 1B). Addition of 10 μM staurosporine, a potent protein kinase

TPL-2 complex assay
13
inhibitor that does not affect TPL-2 catalytic activity [26], did not affect peptide
phosphorylation (data not shown).
Analysis of spot intensities revealed that TPL-2 positively selected for acidic
residues in all ‘minus’ positions from P-5 to P-1, with a preference for Asp over
Glu. TPL-2 had little activity towards peptides containing fixed hydrophobic or
positively charged amino acids at these positions. Peptides containing negatively
or positively charged amino acids at P+1 to P+3 were minimally labeled by TPL-2,
and peptides with a Pro at P+1 were not labeled. Hydrophobic amino acids at P+1
to P+3 were preferentially phosphorylated, with the strongest positive selection
for Trp at P+3. Strong phosphorylation was observed for peptides with fixed Ser
residues at most positions surrounding the central Ser/Thr phosphorylation site.
However, as these peptides contain two phosphorylation sites, these strong
signals may not reflect true selectivity. No strong positive or negative selections
for phosphoamino acids (pThr or pTyr) were observed at any position.
The peptide library screen was repeated three times and selectivity values
calculated from spot intensities (Supplementary Table 1). Selectivity values
quantitatively describe a kinase’s preference for each amino acid at any one of
nine positions surrounding the central phosphorylation site. These values can
range from 0 (strongest negative selection) to 20 (strongest positive selection),
where a value of 1.0 indicates no preference (ie. neutral selectivity). Because of
possible misinterpretation of preference for Ser/Thr, selectivity values were
calculated to disregard positive selections of Ser or Thr, and then combined into a
position-specific scoring matrix (PSSM). This analysis showed that TPL-2
exhibited comparatively weak positive selections, with Trp at P+3 being the
strongest preference with a selectivity value of 3.26. At other positions, TPL-2
generally tolerated several amino acids with similar physicochemical properties
(Figure 2A). These data suggested that TPL-2 selects target residues to
phosphorylate based on primary sequence properties, such as charge and
hydrophobicity, rather than specific amino acid identity at positions relative to the
phosphorylation site.
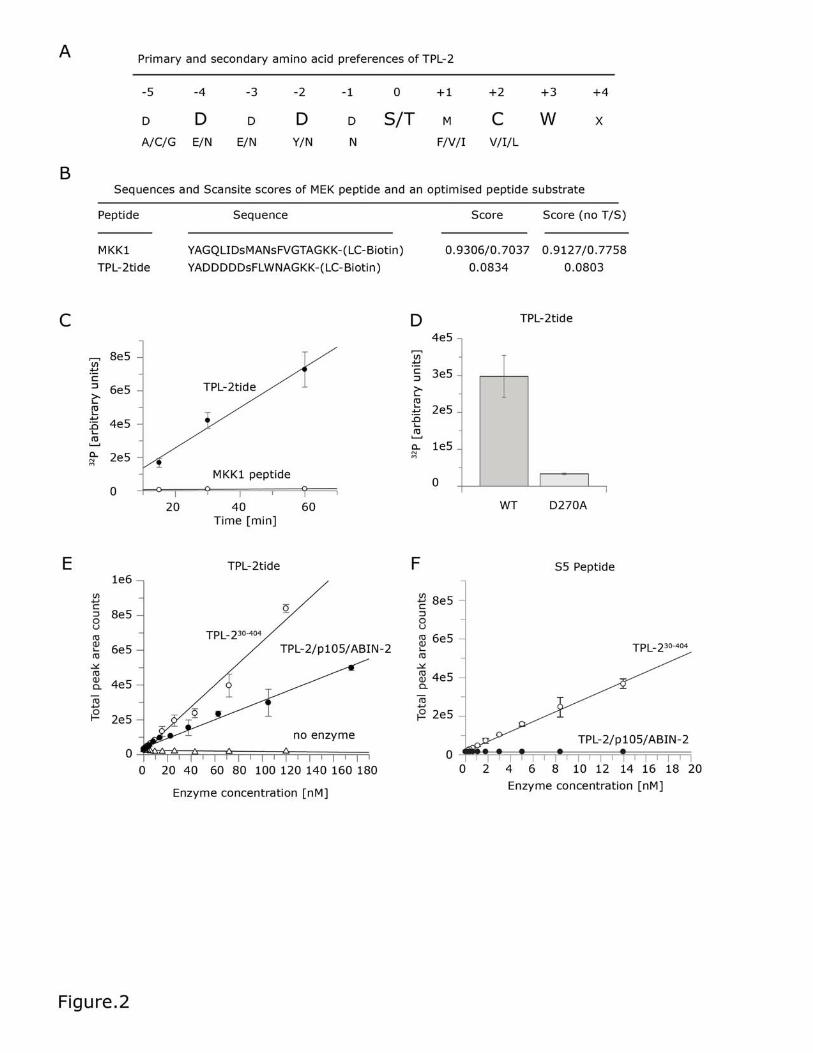
To test whether the peptide library screen could correctly predict phosphorylation
efficiency of linear sequences by TPL-2, a synthetic peptide substrate was
generated corresponding to an optimised sequence predicted by the TPL-2
consensus motif (Figure 2A). The sequence of this optimal peptide was modified
so that Phe and Leu residues replaced the reactive Met and Cys residues at P+1

TPL-2 complex assay
14
and P+2, respectively. Because the selectivity values of these alternative amino
acids were similar, these modifications were expected to have minimal effect on
TPL-2 preference for this peptide. TPL-2 phosphorylation of optimised peptide
substrate (termed TPL-2tide) was then compared to a MKK1 peptide control,
using recombinant purified TPL-2 complex (Figure 2B). TPL-2 complex
phosphorylated TPL-2tide approximately 60-fold more efficiently than MKK1
peptide at all time points (Figure 2C). TPL-2tide was minimally phosphorylated by
kinase-inactive TPL-2[D270A] complex (Figure 2D) demonstrating that the
measured kinase activity was indeed due to TPL-2. These results confirmed the
TPL-2 specificity of the optimal phosphorylation site motif determined by peptide
library screening.
Development of a high throughput kinase assay for the TPL-2 complex
Multiple screens by large pharmaceutical companies using the isolated TPL-2
kinase domain have identified small molecule ATP competitive inhibitors that
specifically block TPL-2 signalling in intact cells, some of which also have activity
in disease models in mice and/or rats [8, 9]. However, none of these inhibitors
has progressed to clinical trials, perhaps as a consequence of sub-optimal
molecular properties or inadequate therapeutic index. Given the intensive
resources previously dedicated to screening with the isolated TPL-2 kinase
domain, a new screening approach, with the potential to expose more molecular
interactions, may ultimately prove the best route to development of anti-
inflammatory drugs from this target.
Our earlier study has indicated that TPL-2 promotes the production of TNF in LPS-
stimulated macrophages independently of ERK1/2 activation while still bound to
NF-κB1 p105 and ABIN-2 [13]. The TPL-2/NF-κB1 p105/ABIN-2 complex is
therefore believed to offer a more a physiologically relevant target for
development of drugs to treat TNF-mediated inflammatory diseases. It is likely
that the TPL-2 kinase domain adopts a different conformation when bound to NF-
κB1 p105 and ABIN-2 than in isolation. In addition, the isolated TPL-2 kinase
domain has a tendency to associate with heat shock proteins and is unstable, in
contrast to the TPL-2/NF-κB1 p105/ABIN-2 complex [14, 27]. Consequently,
screens with the TPL-2 complex might identify different types of inhibitors with
better properties for drug development than those identified with the isolated
TPL-2 kinase domain. These could include non-ATP competitive allosteric
inhibitors, which might have better physicochemical properties. With this aim in

TPL-2 complex assay
15
mind, we developed a high-throughput assay to monitor phosphorylation of TPL-
2tide by the TPL-2/NF-κB1 p105/ABIN-2 complex.
We used the RapidFireTM High-Throughput mass spectrometry system (Agilent
Technologies) to analyse TPL-2tide phosphorylation in a 384-well plate format.
This label-free method directly measures phosphorylated product and is not
commonly prone to interference seen in assays using fluorescent and absorbance
readouts. Multiple reaction monitoring (MRM) was used to simultaneously detect
unphosphorylated and phosphorylated peptide. Due to relatively low substrate
conversion, peak area counts for the phosphorylated peptide were used to
measure TPL-2 activity rather than percentage of substrate conversion from
unphosphorylated to phosphorylated peptide. After 30 min incubation of reaction
mixes, TPL-2tide phosphorylation increased with increasing complexed TPL-2
enzyme concentration. The isolated TPL-2 kinase domain, TPL-230-404 [9], was also
able to phosphorylate TPL-2tide, with slightly higher efficiency than complexed
TPL-2. A large signal-to-noise ratio was achieved with complexed both complexed
TPL-2 and TPL-230-404 with enzyme concentrations at 80 nM, which was used for
subsequent inhibitor assays (Figure 2E). A Z-factor of 0.6-0.7 indicated that this
assay was robust and suitable for high-throughput screening.
Comparative analysis of synthetic MEK-1 and phospho-MEK1 peptides
demonstrated that the RapidFireTM assay was capable of detecting MEK1 peptide
phosphorylation (data not shown). However, we were unable to detect
phosphorylation of a MEK1 activation loop peptide with this assay, using either
complexed TPL-2 or TPL-230-404. Using the TPL-2 PSSM, Scansite 2.0 determined
whether MKK1 peptide exhibited similarity to the optimal TPL-2 complex
consensus motif. Phosphorylation sites in the activation loop of MKK1/2
generated only scored just below 1.00 (0.78 and 0.91), with a score of 1.0
indicating no selection. This suggests that TPL-2 does not select MEK1 as a
substrate based on its linear amino acid sequence. Rather, other mechanisms
such as protein-protein interaction are likely involved. Consistent with this
hypothesis, TPL-2 binds to MEK1 protein and this interaction is blocked by p105,
which inhibits TPL-2 MEK1 kinase activity [10].
S5 (GAFRSAIRRLAARRR), an optimized peptide substrate for the isolated TPL-2
kinase domain identified by IMAP peptide library screen, was not phosphorylated
by complexed TPL-2 (Figure 2F). In contrast, TPL-230-404 phosphorylated S5
peptide efficiently, as expected. These data support the hypothesis that

TPL-2 complex assay
16
complexed TPL-2 does not have the same substrate specificity as the isolated
TPL-2 kinase domain. The isoelectric point of the S5 peptide is 12.54, compared
with 3.74 for TPL-2tide suggesting that peptide substrate selection by the isolated
TPL-2 kinase domain and TPL-2 complex is dependent on the charge of the amino
acids surrounding the phosphorylated serine.
Differential sensitivity of complexed TPL-2 and TPL-2 kinase domain to
TPL-2 inhibitors
Wyeth (Pfizer), Abbott and Novartis have each run large inhibitor screening
programmes targeting the kinase domain of TPL-2 [8, 9, 17, 28]. This has
resulted in a number of ATP-competitive small molecule inhibitors, which block
TPL-2 catalytic activity in vitro and in cells. We next tested the sensitivity of the
TPL-2 complex to a small panel of these inhibitors in comparison with TPL-230-404
kinase domain by measuring TPL-2tide phosphorylation over a range of
compound concentrations (Figure 3 B and C).
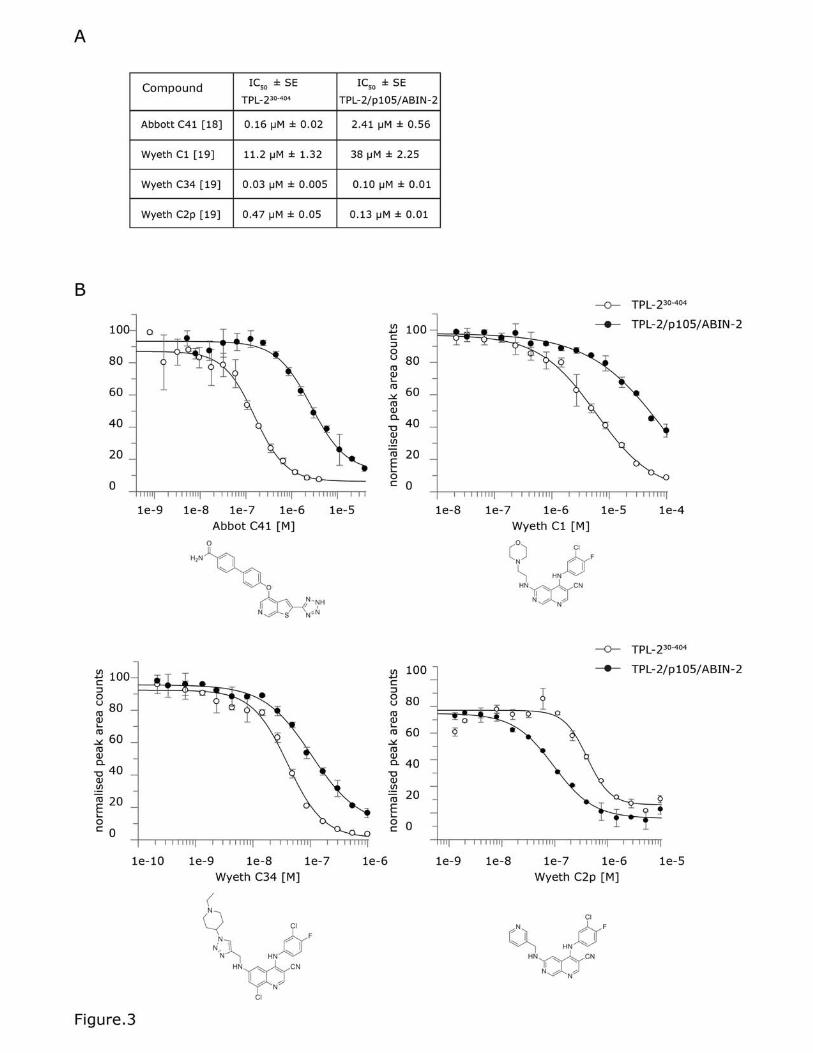
Complexed TPL-2 was substantially less sensitive to inhibition by Wyeth C1 [18,
20] and Abbott C41 [19]. Wyeth C34 [21] also inhibited TPL-2[30-404] slightly
more efficiently than complexed TPL-2. In contrast, Wyeth C2p [18, 20] more
potently inhibited complexed TPL-2 than TPL-230-404. The significant differences in
sensitivities of complexed TPL-2 and TPL-230-404 to ATP competitive inhibitors may
indicate that the ATP binding pocket of TPL-2 associated with NF-κB1 p105 and
ABIN-2 adopts a different conformation to that in TPL-230-404. Consistent with this,
the TPL-2 kinase domain forms interactions both with NF-κB1 p105 and its own
C-terminus [10, 29], which is lacking in TPL-230-397 and TPL-230-404 used in earlier
inhibitor screens.
TPL-2 kinase is a promising drug target for treatment of inflammatory diseases
due to its critical role in regulating the production of TNF and its requirement for
the development of inflammation in multiple mouse disease models [5]. Earlier
screens for TPL-2 inhibitors have used the isolated TPL-2 kinase domain (TPL-230-
397 and TPL-230-404) purified from baculovirus-infected insect cells and MKK1
peptide or protein substrates [8, 9]. Notably, this approach has failed to generate
any inhibitors that have progressed to clinical trials. The present study suggests
that screening with TPL-2/NF-κB1 p105/ABIN-2, using TPL-2tide substrate, has
the potential to identify novel ATP competitive TPL-2 inhibitor series that
preferentially inhibit the kinase domain of complexed TPL-2 in comparison with
the isolated TPL-2 kinase domain. In addition, it is possible that screening with

TPL-2 complex assay
17
the TPL-2 complex will identify allosteric inhibitors (that block TPL-2 signalling
independently of binding to the TPL-2 ATP binding site) that have increased
specificity compared to ATP competitive inhibitors. Furthermore, novel TPL-2
inhibitors identified in screens with the TPL-2/NF-κB1 p105/ABIN-2 complex may
have improved in vivo potency that allows their development through to clinical
trial as anti-inflammatory drugs.

TPL-2 complex assay
18
REFERENCES
[1] C. Monaco, J. Nanchahal, P. Taylor, M. Feldmann, Anti-TNF therapy: past,
present and future., Int. Immunol., 27 (2014) 55 - 62. [2] P. Cohen, D.R. Alessi, Kinase drug discovery - What's next in the field?,
Chem. Biol., 8 (2013) 96 - 104. [3] T. Kawai, S. Akira, The role of pattern-recognition receptors in innate
immunity: update on Toll-like receptors., Nat. Immunol., 11 (2010) 373 - 384. [4] C.D. Dumitru, J.D. Ceci, C. Tsatsanis, D. Kontoyiannis, K. Stamatakis, J.-H.
Lin, C. Patriotis, N.A. Jenkins, N.G. Copeland, G. Kollias, P.N. Tsichlis, TNFa
induction by LPS is regulated post-transcriptionally via a TPL2/ERK-dependent pathway., Cell, 103 (2000) 1071-1083.
[5] T. Gantke, S. Sriskantharajah, M. Sadowski, S.C. Ley, IκB kinase regulation of the TPL-2/ERK MAPK pathway., Immunol. Rev., 246 (2012) 168 - 182.
[6] S. Sriskantharajah, E. Gückel, N. Tsakiri, K. Kierdorf, C. Brender, H. Ben-Addi, M. Veldhoen, P.N. Tsichlis, B. Stockinger, M. Prinz, G. Kollias, S.C. Ley,
Regulation of experimental autoimmune encephalomyelitis by TPL-2., J. Immunol., 192 (2014) 3515 - 3529.
[7] Y. Xiao, J. Jin, M. Chang, M. Nakaya, H. Hu, Q. Zou, X. Zhou, G.C. Brittain, X.
Cheng, S.C. Sun, TPL2 mediates autoimmune inflammation through activation of the TAK1 axis of IL-17 signaling., J. Exp. Med., jem.20132640 (2014).
[8] D. George, A. Salmeron, Cot / TPL-2 protein kinase as a target for the treatment of inflammatory disease., Curr. Topics. Med. Chem., 9 (2009) 611 -
622. [9] S. Gutmann, A. Hinninger, G. Fendrich, P. Drückes, S. Antz, H. Mattes, H.
Möbitz, S. Ofner, N. Schmiedeberg, A. Stojanovic, S. Rieffel, A. Strauss, T. Troxler, R. Glatthar, H. Sparrer, The crystal structure of Cancer Osaka Thyroid
kinase reveals an unexpected kinase domain fold., J. Biol. Chem., 290 (2015)
15210 - 15218. [10] S. Beinke, J. Deka, V. Lang, M.P. Belich, P.A. Walker, S. Howell, S.J.
Smerdon, S.J. Gamblin, S.C. Ley, NF-kB p105 negatively regulates TPL-2 MEK kinase activity., Mol. Cell. Biol., 23 (2003) 4739-4752.
[11] S. Beinke, M.J. Robinson, A. Salmeron, M. Hugunin, H. Allen, S.C. Ley, Lipopolysaccharide activation of the TPL-2/MEK/Extracellular signal-regulated
kinase mitogen-activated protein kinase cascade is regulated by IκB kinase-induced proteolysis of NF-κB1 p105., Mol. Cell. Biol., 24 (2004) 9658-9667.
[12] M. Waterfield, W. Jin, W. Reiley, M.Y. Zhang, S.-C. Sun, IKKb is an essential
component of the TPL-2 signaling pathway., Mol. Cell Biol., 24 (2004) 6040-6048. [13] H.T. Yang, M. Papoutsopoulou, M. Belich, C. Brender, J. Janzen, T. Gantke,
M. Handley, S.C. Ley, Coordinate regulation of TPL-2 and NF-κB signaling in macrophages by NF-κB1 p105., Mol. Cell. Biol., 32 (2012) 3438 - 3451.
[14] T. Gantke, S. Boussouf, J. Janzen, N. Morrice, S. Howell, E. Muhlberger, S.C. Ley, Ebola virus VP35 induces high-level production of recombinant TPL-2-ABIN-
2-NF-κB1 p105 complex in co-transfected HEK-293 cells., Biochem. J., 452 (2013) 359 - 365.
[15] A. Salmeron, J. Janzen, Y. Soneji, N. Bump, J. Kamens, H. Allen, S.C. Ley,
Direct phosphorylation of NF-kB p105 by the IkB kinase complex on serine 927 is essential for signal-induced p105 proteolysis., J. Biol. Chem., 276 (2001) 22215-
22222. [16] M. Schumann, T. Gantke, E. Muhlberger, Ebola virus VP35 antagonizes PKR
activity through its C-terminal interferon inhibitory domain., J. Virol., 83 (2009) 8993 - 8997.
[17] Y. Jia, C.M. Quinn, A. Clabbers, R. Talanian, Y. Xu, N. Wishart, H. Allen, Comparative analysis of various in vitro COT kinase assay formats and their
applications in inhibitor identification and characterization., Anal Biochem., 350
(2006) 268-276.

TPL-2 complex assay
19
[18] L.K. Gavrin, N. Green, Y. Hu, K. Janz, N. Kaila, H.Q. Li, S.Y. Tam, J.R.
Thomason, A. Gopalsamy, G. Ciszewski, J.W. Cuozzo, J.P. Hall, S. Hsu, J.B. Telliez, L.L. Lin, Inhibition of Tpl2 kinase and TNF-alpha production with 1,7-
naphthyridine-3-carbonitriles: Synthesis and structure-activity relationships., Bioorg Med Chem Lett., 15 (2005) 5288-5292.
[19] D. George, M. Friedman, H. Allen, M. Argiriadi, C. Barberis, A. Bischoff, A. Clabbers, K. Cusack, R. Dixon, S. Fix-Stenzel, T. Gordon, B. Janssen, Y. Jia, M.
Moskey, C.M. Quinn, J.A. Salmeron, N. Wishart, K. Woller, Z. Yu, Discovery of thieno [2,3-c]pyridines as potent COT inhibitors., Bioorg. Med. Chem. Lett., 18
(2008) 4952 - 4955.
[20] N. Kaila, N. Green, H.-Q. Li, Y. Hu, K. Janz, L.K. Gavrin, J. Thomason, S. Tam, D.W. Powell, J. Cuozzo, J.P. Hall, J.B. Telliez, S. Hsu, C.L. NIckerson-Nutter,
Q. Wang, L.-L. Lin, Identification of a novel class of selective Tpl2 kinase inhibitors: 4-Alkylamino-[1,7]naphthyridine-3-carbonitriles., bioorg. Med. Chem.,
15 (2007) 6425 - 6442. [21] J. Wu, N. Green, R. Hotchandani, Y. Hu, J. Condon, A. Huang, N. Kaila, H.Q.
Li, S. Guler, W. Li, S.Y. Tam, Q. Wang, J. Pelker, S. Marusic, S. Hsu, P.J. Hall, J.B. Telliez, J. Cui, L.L. Lin, Selective inhibitors of tumor progression loci-2 (Tpl2)
kinase with potent inhibition of TNF-alpha production in human whole blood.,
Bioorg. Med. Chem. Lett., 19 (2009) 3485 - 3488. [22] J.E. Hutti, E.T. Jarrell, J.D. Chang, D.W. Abbott, P. Storz, A. Toker, L.C.
Cantley, B.E. Turk, A rapid method for determining protein kinase phosphorylation specificity., Nat. Methods, 1 (2004) 27 - 29.
[23] M.B. Yaffe, G.G. Leparc, J. Lai, T. Obata, S. Volinia, L.C. Cantley, A motif-based profile scanning approach for genome-wide prediction of signaling
pathways., Nat. Biotech., 19 (2001) 348 - 353. [24] M.J. Pattison, O. Mitchell, H.R. Flynn, C.-S. Chen, H.-T. Yang, A. Ben-Addi,
S. Boeing, B. Snijders, S.C. Ley, TLR and TNFR1 activation of the MKK3/MKK6-
p38alpha axis in macrophages is mediated by TPL-2 kinase., Biochem. J., 473 (2016) 2845 - 2861.
[25] K. Roget, A. Ben-Addi, A. Mambole-Dema, T. Gantke, J. Janzen, H.-T. Yang, N. Shpiro, N. Morrice, D. Abbott, S.C. Ley, IKK2 regulates TPL-2 activation of
ERK-1/2 MAP kinases by direct phosphorylation of TPL-2 serine 400., Mol. Cell. Biol., 32 (2012) 4684 - 4690.
[26] B.S. Luciano, S. Hsu, P.L. Channavajhala, L.-L. Lin, J.W. Cuozzo, Phoshorylation of threonine 290 in the activation loop of Tpl2/Cot is necessary
but not sufficient for kinase activity., J. Biol. Chem., 279 (2004) 52117-52123.
[27] Y. Jia, C.M. Quinn, N.J. Bump, K.M. Clark, A. Clabbers, J. Hardman, A. Gagnon, J. Kamens, M.J. Tomlinson, N. Wishart, H. Allen, Purification and kinetic
characterization of recombinant human mitogen-activated protein kinase kinase kinase COT and the complexes with its cellular partner NF-kB1 p105., Arch.
Biochem. Biophys., 441 (2005) 64 - 74. [28] J.P. Hall, Y. Kurdi, S. Hsu, J. Cuozzo, J. Liu, J.-B. Telliez, K.J. Seidl, A.
Winkler, Y. Hu, N. Green, G.R. Askew, S. Tam, J.D. Clark, L.-L. Lin, Pharmacologic inhibition of TPL-2 blocks inflammatory responses in primary
human monocytes, synoviocytes, and blood., J. Biol. Chem., 282 (2007) 33295-
33304. [29] J.D. Ceci, C.P. Patriotis, C. Tsatsanis, A.M. Makris, R. Kovatch, D.A. Swing,
N.A. Jenkins, P.N. Tsichlis, N.G. Copeland, TPL-2 is an oncogenic kinase that is activated by carboxy-terminal truncation., Gene. Devel., 11 (1997) 688-700.

TPL-2 complex assay
20
FIGURE LEGENDS
Figure 1. The primary amino acid sequence specificity of TPL-2. The
peptide library comprised 198 individual biotinylated peptide mixtures. Each
peptide contained a central phosphoracceptor Ser or Thr, flanked by degenerate
positions, comprising an equimolar mixture of the 17 amino acids, excluding Cys,
Ser and Thr. In each peptide, one position was fixed (fixed residue) with one of
the 20 naturally occurring unmodified amino acids, phosphor-Thr (pT) or
phosphor-Tyr (pY). Peptides were incubated with 30nM recombinant TPL-2/NF-
κB1 p105/ABIN-2 complex at a final substrate concentration of 50 μM at 30 °C
for 1 hour. Assays were performed in TPL-2 kinase buffer plus 10μM ATP and 3
nCi/μL [γ-32P] ATP in the absence (A) or presence (B) of C34 TPL-2 inhibitor (10
μM). Following incubation, reactions were transferred onto SAM2 membranes,
which were then washed extensively. Incorporation of 32P into peptides was
quantified by phosphorimaging.
Figure 2. Testing an optimised peptide substrate for TPL-2/NF-κB1
p105/ABIN-2. (A) The primary and secondary amino acid preferences for
phosphorylation by recombinant TPL-2/NF-κB1 p105/ABIN-2 complex are shown.
Panel (B) shows the sequences and scansite scores for the MKK1 activation loop
and optimised TPL-2tide peptide substrates. (C) Time-course experiment
comparing TPL-2/NF-κB1 p105/ABIN-2 complex phosphorylation of MKK1 and
TPL-2tide peptides (50 μM final concentration). Assays were performed with 30nM
recombinant TPL-2/NF-κB1 p105/ABIN-2 complex in TPL-2 kinase buffer plus
1mM ATP and 0.02 μCi/μL [γ-32P] ATP. Peptides were transferred onto
streptavidin-coated membranes, which were then extensively washed.
Incorporation of 32P into peptides was quantified by phosphorimaging. Linear
regression was fitted with GraFit version 7.0.3. Values are means ± S.D. for three
replicate reactions. (D) Kinase assays as in (C) comparing TPL-2tide peptide
phosphorylation by wild-type (WT) or kinase-inactive (D270A) TPL-2/NF-κB1
p105/ABIN-2 complex (15 min). (E) and (F) TPL-230-404 and TPL-2/NF-κB1
p105/ABIN-2 were titrated (1:1.66) in 384-well plates. A substrate solution
containing ATP and TPL-2tide (E) or S5 peptide (F) was then added. The plates
were analysed using a Sciex API6500 (E) or API4000 (F) Triple Quad with
RapidFireTM Technology. The graph shows total peak area counts for the MRM
transition 953.6/904.3 Da (E; phosphorylated TPL-2tide peptide) and 612.9/120
Da (F; phosphorylated S5 peptide) for the indicated enzyme at increasing
concentrations. Linear regression was fitted using GraFit software.

TPL-2 complex assay
21
Figure 3. High-throughput Rapid FireTM mass spectrometry assay of TPL-
2/NF-κB1 p105/ABIN-2 complex kinase activity.
(A) The efficacy of known TPL-2 inhibitors was tested for TPL-230-404 and the TPL-
2 complex. 5 μL of the enzyme solutions were added to each well (two replicates
for each using final concentrations of 80 nM TPL2 30-404 and 80 nM TPL-2/NF-
κB1 p105/ABIN-2). The compounds were then dispensed using a HP digital
dispenser. The enzyme/compound mix was incubated for 10 min before the
addition of 5 μL of substrate buffer containing final concentrations of 100 μM TPL-
2tide and 400 μM ATP to initiate the reaction. The reactions were incubated for
30 min quenched with 50 μL 1 % formic acid. The plates were analysed using a
Sciex API6500 Triple Quad with RapidFireTM Technology. The data was analysed
using product counts for the MRM transition 953.6/904.3 Da (phosphorylated
peptide) using a 4-parameter logistical equation fitted using GraFit software to
determine IC50 values. For each compound multiple replicates were run for each
experiment and the experiment repeated 4-5 times. The IC50 values shown are
generated from all datasets analysed and include standard errors (SE). (B)
Representative graphs of one experiment for each compound are shown,
including standard deviation from replicates.
Supplementary Figure 1. Oriented degenerate peptide library.
Supplementary Table 1. TPL-2 position-specific scoring matrices
generated by peptide library experiments.
To quantify differences in peptide phosphorylation by the TPL-2 complex, the
peptide library screen was repeated three times and selectivity values calculated
as describe in the Methods section. Selectivity values quantitatively describe TPL-
2’s preference for each amino acid at any one of nine positions surrounding the
central phosphorylation site. These values can range from 0 (strongest negative
selection) to 20 (strongest positive selection), where a value of 1.0 indicates no
preference (ie. neutral selectivity). Because of possible misinterpretation of
preferences for Ser / Thr, selectivity values were calculated to include or
disregard positive selections for Ser or Thr. These values were combined into
position-specific scoring matrices, termed ‘Full Motif’ and ‘No Ser / Thr’.
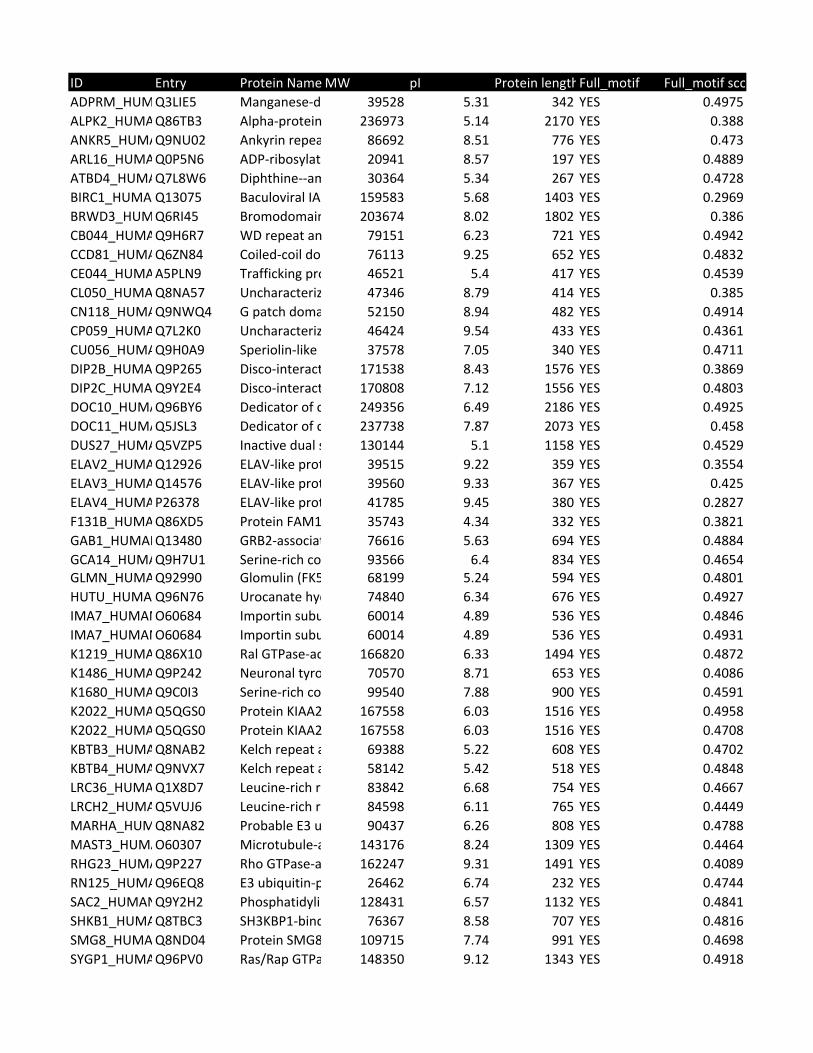
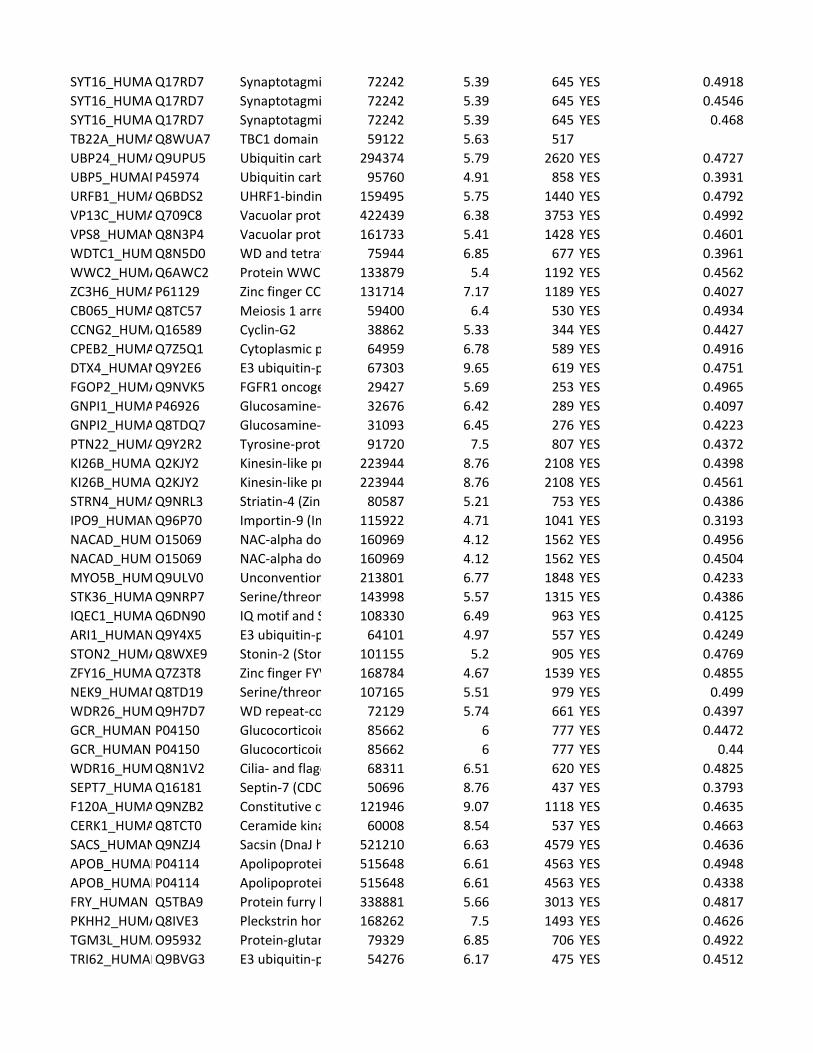
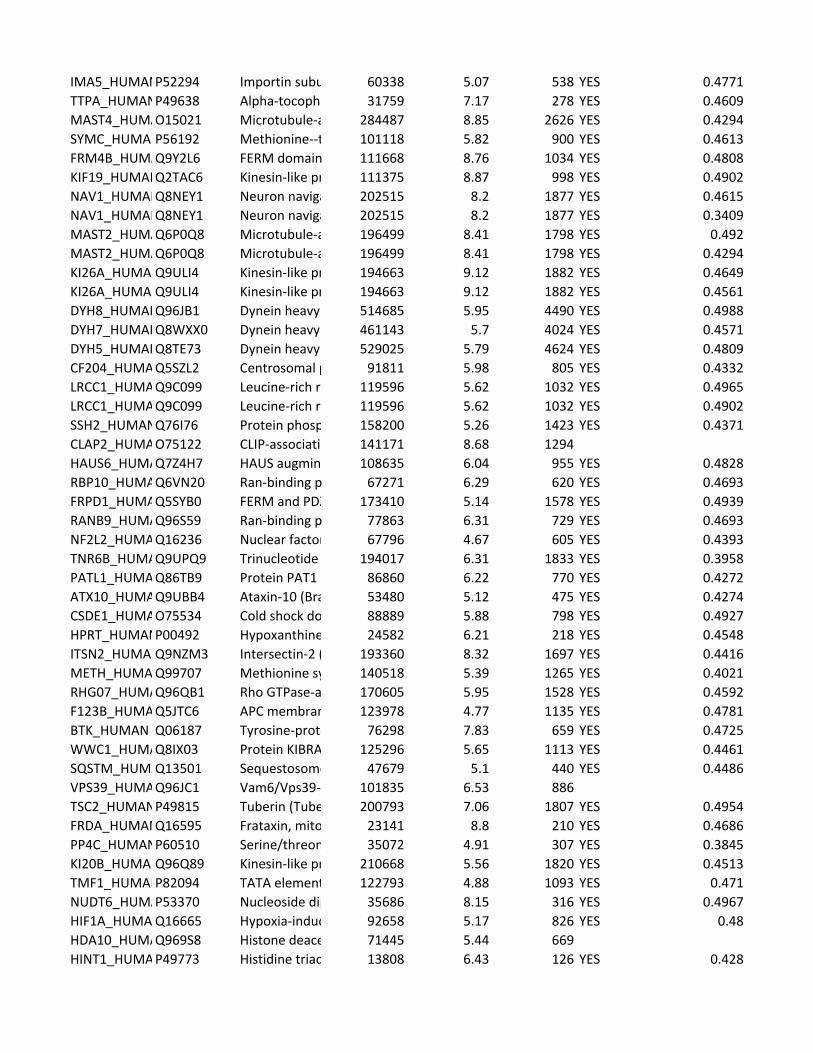
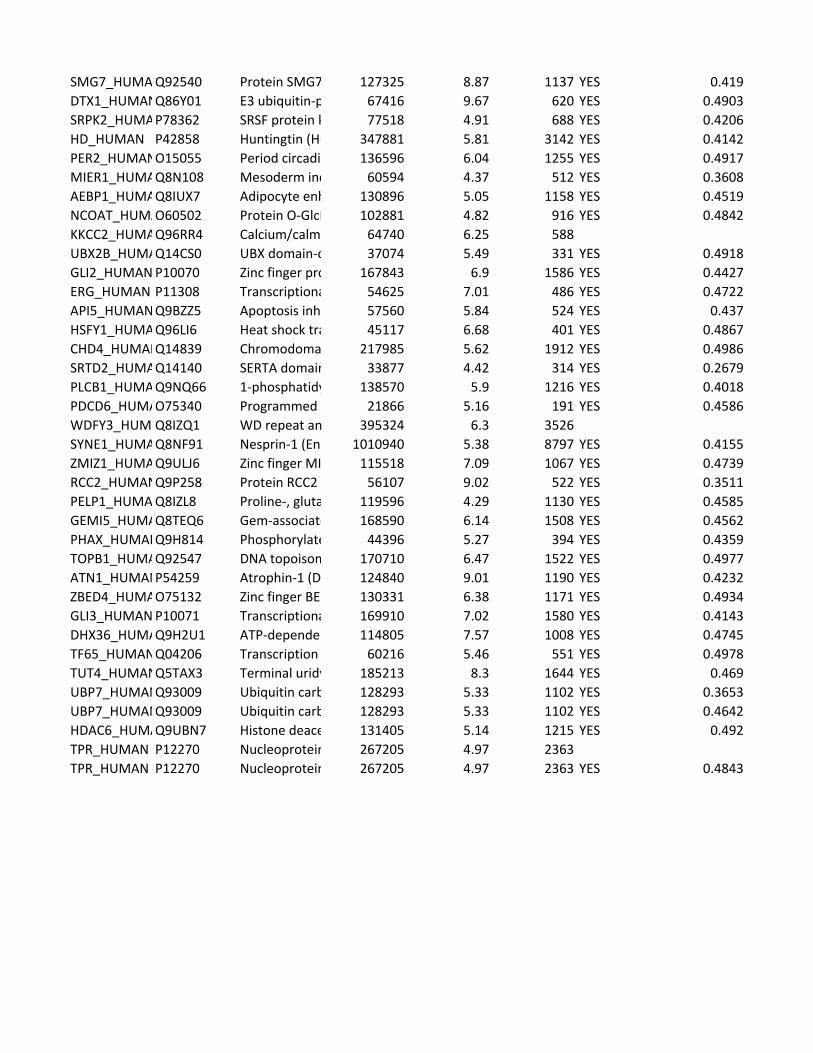
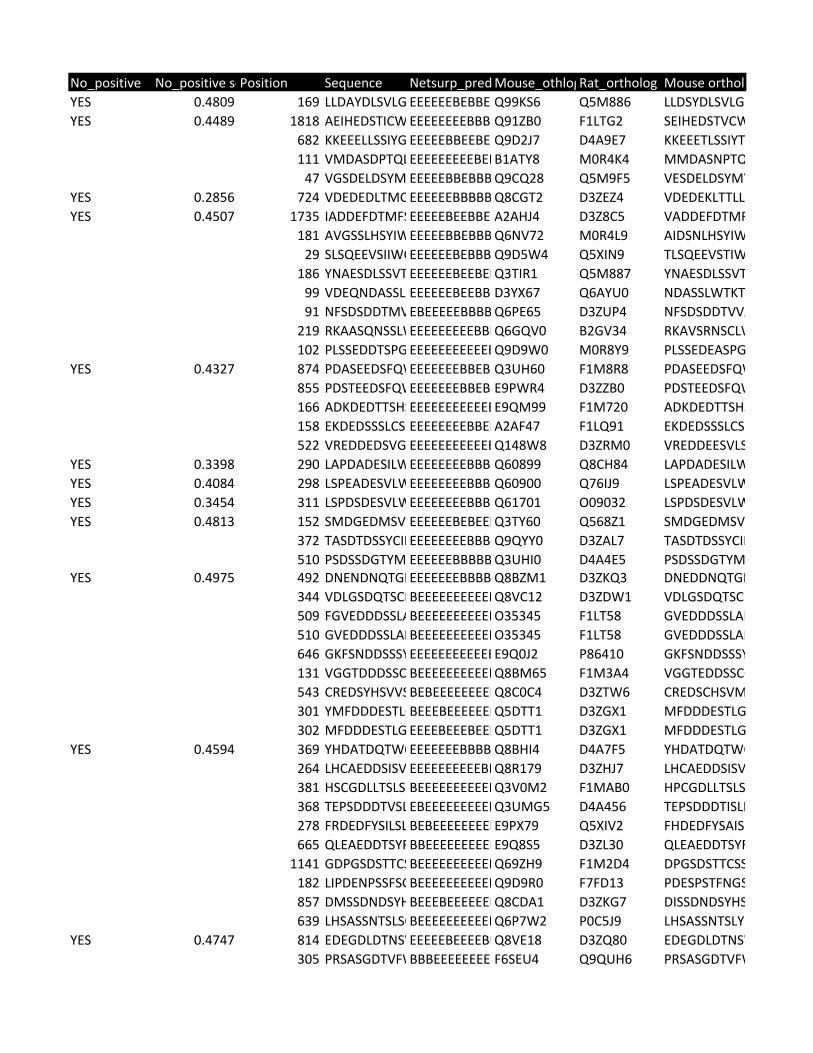
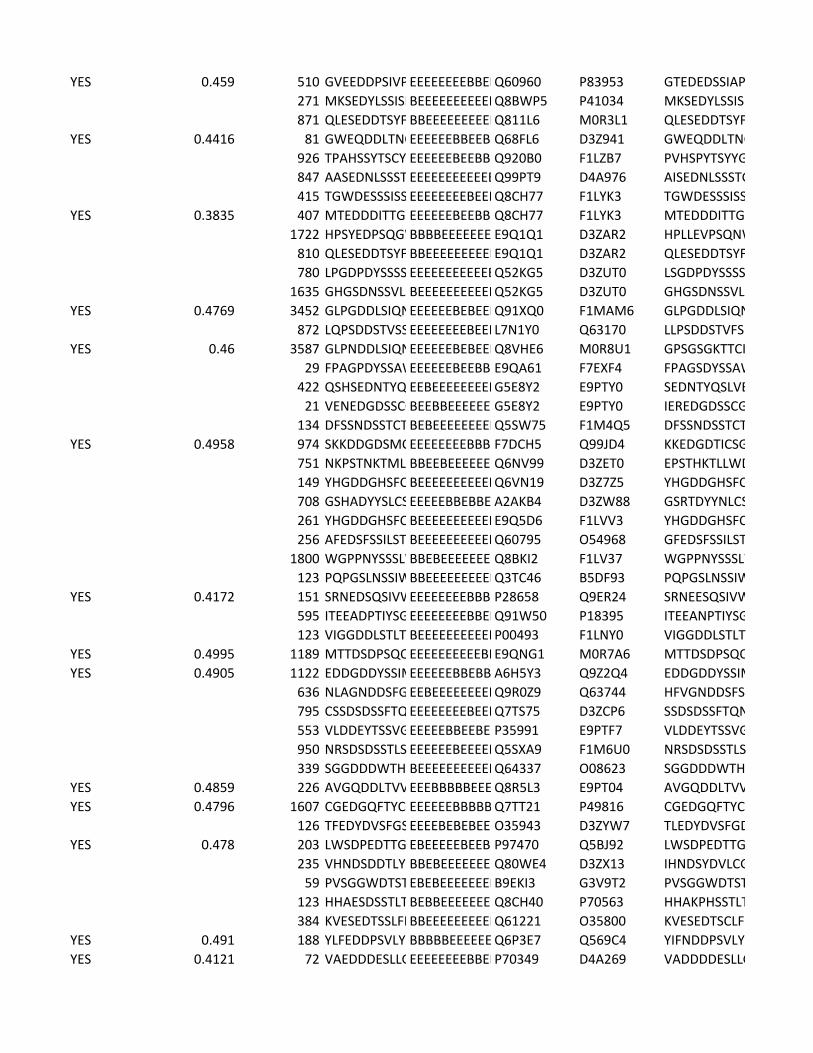
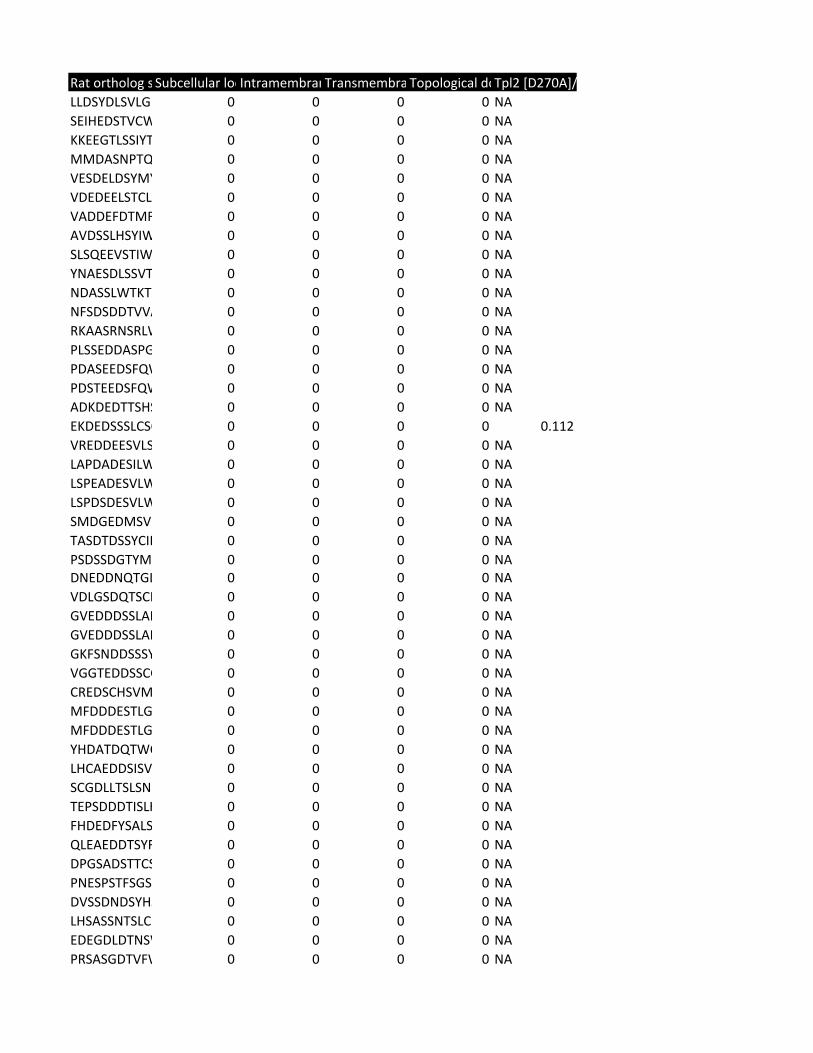
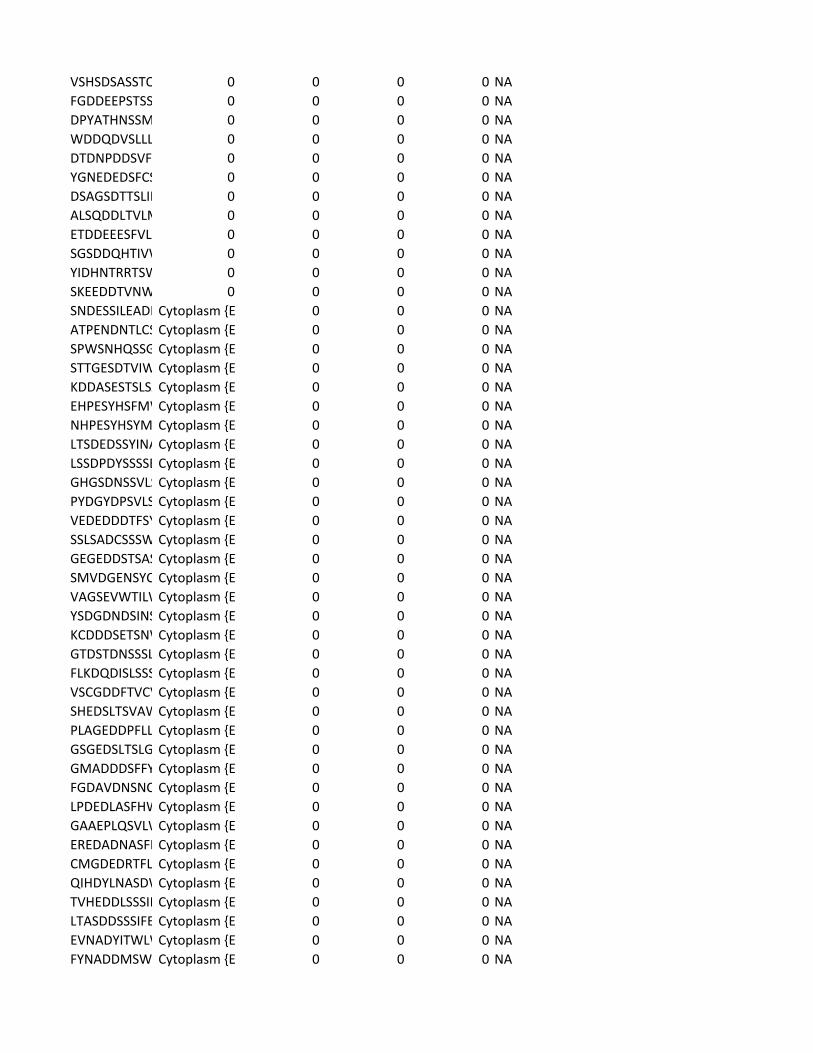
Supplementary Table 2. Predicted substrates for the TPL-2/NF-κB1
p105/ABIN-2 complex. Human proteins predicted to contain a TPL-2 complex
phosphorylation motif were identified using scansite [http://scansite.mit.edu/]

TPL-2 complex assay
22
and the TPL-2 PSSM. Only human peptides with a mouse and rat ortholog were
included. Proteins were excluded as potential substrates if the target amino acid
was not predicted to be surface exposed, as determined by NurSurfP (version 1.1
http://www.cbs.dtu.dk/services/NetSurfP/), or in a transmembrane region, using
TMHMM version 2 (http://www.cbs.dtu.dk/services/TMHMM/). Only cytoplasmic
proteins were included as possible substrates, as determined by Gene Ontology
analysis [http://www.geneontology.org/].

TPL-2 complex assay
23

P G A C S T V I L M F Y W H K R Q N D E
1.15 1.19 1.46 1.26 1 0.79 0.75 0.44 0.57 0.67 0.83 0.89 1.25 1.05 0.65 0.78 1.03 1.28 1.61 1.371.07 1.34 1.32 0.89 1 0.88 0.48 0.49 0.58 0.77 0.55 0.92 1.11 1.21 0.52 0.46 1.11 1.49 2.2 1.611.09 1.05 0.92 0.46 1 1 0.69 0.55 0.68 0.92 0.49 1.32 1.54 0.84 0.46 0.4 1.14 1.51 2.04 1.90.57 0.85 0.83 0.48 1 0.94 0.51 0.67 1.21 0.99 1.16 1.68 1.35 0.78 0.31 0.24 0.9 1.45 2.72 1.371.07 0.55 0.76 0.27 1 0.95 0.72 0.72 1.25 1.24 0.79 1.31 1.1 1.19 0.94 0.66 1.31 1.57 1.6 1.01
0.25 0.64 0.68 0.62 1 1 1.59 1.79 1.19 1.9 1.6 1.48 1.18 0.86 0.4 0.33 1.21 1.1 0.6 0.590.72 1.12 0.93 1.98 1 0.85 1.15 1.26 1.42 1.06 0.96 1.15 1.22 1.12 0.5 0.48 1 1.18 0.48 0.420.93 1.15 0.93 0.66 1 1 0.89 0.82 1.09 1.18 0.99 1.45 3.26 1.19 0.43 0.43 0.91 1.01 0.42 0.251.04 1.38 0.98 1.16 1 1 1.05 0.74 0.88 1.11 0.94 0.88 1.07 1.22 0.63 0.38 1.19 1.38 1.09 0.89
-5-4-3-2-1 0+1+2+3+4
No Ser/Thr
Full motif
P G A C S T V I L M F Y W H K R Q N D E
1.12 1.16 1.43 1.24 1.35 0.77 0.74 0.43 0.56 0.66 0.81 0.87 1.23 1.03 0.63 0.76 1.01 1.25 1.59 1.341.04 1.31 1.29 0.87 1.43 0.86 0.47 0.48 0.56 0.76 0.54 0.9 1.08 1.18 0.51 0.45 1.08 1.46 2.15 1.581.06 1.02 0.89 0.44 1.55 1 0.67 0.53 0.66 0.9 0.48 1.28 1.5 0.82 0.44 0.39 1.11 1.47 1.97 1.840.57 0.85 0.82 0.48 1.04 0.93 0.51 0.66 1.21 0.98 1.16 1.67 1.35 0.78 0.31 0.24 0.9 1.45 2.71 1.361.05 0.54 0.75 0.26 1.28 0.94 0.71 0.7 1.23 1.22 0.78 1.29 1.09 1.17 0.93 0.65 1.29 1.55 1.58 0.99
0.23 0.6 0.64 0.58 1.78 1.34 1.49 1.67 1.12 1.78 1.5 1.39 1.1 0.8 0.38 0.31 1.14 1.03 0.56 0.560.72 1.11 0.93 1.97 1.14 0.85 1.14 1.25 1.41 1.05 0.95 1.14 1.21 1.11 0.5 0.48 0.99 1.17 0.47 0.410.9 1.11 0.9 0.64 1.57 1.07 0.86 0.79 1.05 1.14 0.96 1.4 3.14 1.15 0.41 0.42 0.88 0.97 0.4 0.241.02 1.35 0.97 1.14 1.1 1.22 1.03 0.72 0.86 1.09 0.92 0.87 1.06 1.2 0.62 0.38 1.17 1.35 1.08 0.87
-5-4-3-2-1 0+1+2+3+4
Supplementary Table 1. TPL-2 position-specific scoring matrices generated by peptide library
ID Entry Protein Name MW pI Protein lengthFull_motif Full_motif scoADPRM_HUMQ3LIE5 Manganese-d 39528 5.31 342 YES 0.4975ALPK2_HUMAQ86TB3 Alpha-protein 236973 5.14 2170 YES 0.388ANKR5_HUMAQ9NU02 Ankyrin repea 86692 8.51 776 YES 0.473ARL16_HUMAQ0P5N6 ADP-ribosylat 20941 8.57 197 YES 0.4889ATBD4_HUMAQ7L8W6 Diphthine--am 30364 5.34 267 YES 0.4728BIRC1_HUMA Q13075 Baculoviral IA 159583 5.68 1403 YES 0.2969BRWD3_HUMQ6RI45 Bromodomain 203674 8.02 1802 YES 0.386CB044_HUMAQ9H6R7 WD repeat an 79151 6.23 721 YES 0.4942CCD81_HUMAQ6ZN84 Coiled-coil do 76113 9.25 652 YES 0.4832CE044_HUMAA5PLN9 Trafficking pro 46521 5.4 417 YES 0.4539CL050_HUMA Q8NA57 Uncharacteriz 47346 8.79 414 YES 0.385CN118_HUMAQ9NWQ4 G patch doma 52150 8.94 482 YES 0.4914CP059_HUMAQ7L2K0 Uncharacteriz 46424 9.54 433 YES 0.4361CU056_HUMAQ9H0A9 Speriolin-like 37578 7.05 340 YES 0.4711DIP2B_HUMA Q9P265 Disco-interact 171538 8.43 1576 YES 0.3869DIP2C_HUMA Q9Y2E4 Disco-interact 170808 7.12 1556 YES 0.4803DOC10_HUMAQ96BY6 Dedicator of c 249356 6.49 2186 YES 0.4925DOC11_HUMAQ5JSL3 Dedicator of c 237738 7.87 2073 YES 0.458DUS27_HUMAQ5VZP5 Inactive dual s 130144 5.1 1158 YES 0.4529ELAV2_HUMAQ12926 ELAV-like prot 39515 9.22 359 YES 0.3554ELAV3_HUMAQ14576 ELAV-like prot 39560 9.33 367 YES 0.425ELAV4_HUMAP26378 ELAV-like prot 41785 9.45 380 YES 0.2827F131B_HUMAQ86XD5 Protein FAM1 35743 4.34 332 YES 0.3821GAB1_HUMANQ13480 GRB2-associat 76616 5.63 694 YES 0.4884GCA14_HUMAQ9H7U1 Serine-rich co 93566 6.4 834 YES 0.4654GLMN_HUMAQ92990 Glomulin (FK5 68199 5.24 594 YES 0.4801HUTU_HUMA Q96N76 Urocanate hyd 74840 6.34 676 YES 0.4927IMA7_HUMANO60684 Importin subu 60014 4.89 536 YES 0.4846IMA7_HUMANO60684 Importin subu 60014 4.89 536 YES 0.4931K1219_HUMAQ86X10 Ral GTPase-ac 166820 6.33 1494 YES 0.4872K1486_HUMAQ9P242 Neuronal tyro 70570 8.71 653 YES 0.4086K1680_HUMAQ9C0I3 Serine-rich co 99540 7.88 900 YES 0.4591K2022_HUMAQ5QGS0 Protein KIAA2 167558 6.03 1516 YES 0.4958K2022_HUMAQ5QGS0 Protein KIAA2 167558 6.03 1516 YES 0.4708KBTB3_HUMAQ8NAB2 Kelch repeat a 69388 5.22 608 YES 0.4702KBTB4_HUMAQ9NVX7 Kelch repeat a 58142 5.42 518 YES 0.4848LRC36_HUMAQ1X8D7 Leucine-rich re 83842 6.68 754 YES 0.4667LRCH2_HUMAQ5VUJ6 Leucine-rich re 84598 6.11 765 YES 0.4449MARHA_HUMQ8NA82 Probable E3 u 90437 6.26 808 YES 0.4788MAST3_HUMAO60307 Microtubule-a 143176 8.24 1309 YES 0.4464RHG23_HUMAQ9P227 Rho GTPase-a 162247 9.31 1491 YES 0.4089RN125_HUMAQ96EQ8 E3 ubiquitin-p 26462 6.74 232 YES 0.4744SAC2_HUMANQ9Y2H2 Phosphatidylin 128431 6.57 1132 YES 0.4841SHKB1_HUMAQ8TBC3 SH3KBP1-bind 76367 8.58 707 YES 0.4816SMG8_HUMA Q8ND04 Protein SMG8 109715 7.74 991 YES 0.4698SYGP1_HUMAQ96PV0 Ras/Rap GTPa 148350 9.12 1343 YES 0.4918
SYT16_HUMA Q17RD7 Synaptotagmi 72242 5.39 645 YES 0.4918SYT16_HUMA Q17RD7 Synaptotagmi 72242 5.39 645 YES 0.4546SYT16_HUMA Q17RD7 Synaptotagmi 72242 5.39 645 YES 0.468TB22A_HUMAQ8WUA7 TBC1 domain 59122 5.63 517UBP24_HUMAQ9UPU5 Ubiquitin carb 294374 5.79 2620 YES 0.4727UBP5_HUMANP45974 Ubiquitin carb 95760 4.91 858 YES 0.3931URFB1_HUMAQ6BDS2 UHRF1-bindin 159495 5.75 1440 YES 0.4792VP13C_HUMAQ709C8 Vacuolar prote 422439 6.38 3753 YES 0.4992VPS8_HUMANQ8N3P4 Vacuolar prote 161733 5.41 1428 YES 0.4601WDTC1_HUM Q8N5D0 WD and tetrat 75944 6.85 677 YES 0.3961WWC2_HUMAQ6AWC2 Protein WWC2 133879 5.4 1192 YES 0.4562ZC3H6_HUMAP61129 Zinc finger CCC 131714 7.17 1189 YES 0.4027CB065_HUMAQ8TC57 Meiosis 1 arre 59400 6.4 530 YES 0.4934CCNG2_HUMAQ16589 Cyclin-G2 38862 5.33 344 YES 0.4427CPEB2_HUMAQ7Z5Q1 Cytoplasmic p 64959 6.78 589 YES 0.4916DTX4_HUMANQ9Y2E6 E3 ubiquitin-p 67303 9.65 619 YES 0.4751FGOP2_HUMAQ9NVK5 FGFR1 oncoge 29427 5.69 253 YES 0.4965GNPI1_HUMAP46926 Glucosamine- 32676 6.42 289 YES 0.4097GNPI2_HUMAQ8TDQ7 Glucosamine- 31093 6.45 276 YES 0.4223PTN22_HUMAQ9Y2R2 Tyrosine-prote 91720 7.5 807 YES 0.4372KI26B_HUMA Q2KJY2 Kinesin-like pr 223944 8.76 2108 YES 0.4398KI26B_HUMA Q2KJY2 Kinesin-like pr 223944 8.76 2108 YES 0.4561STRN4_HUMAQ9NRL3 Striatin-4 (Zine 80587 5.21 753 YES 0.4386IPO9_HUMANQ96P70 Importin-9 (Im 115922 4.71 1041 YES 0.3193NACAD_HUMAO15069 NAC-alpha do 160969 4.12 1562 YES 0.4956NACAD_HUMAO15069 NAC-alpha do 160969 4.12 1562 YES 0.4504MYO5B_HUM Q9ULV0 Unconvention 213801 6.77 1848 YES 0.4233STK36_HUMA Q9NRP7 Serine/threon 143998 5.57 1315 YES 0.4386IQEC1_HUMA Q6DN90 IQ motif and S 108330 6.49 963 YES 0.4125ARI1_HUMANQ9Y4X5 E3 ubiquitin-p 64101 4.97 557 YES 0.4249STON2_HUMAQ8WXE9 Stonin-2 (Ston 101155 5.2 905 YES 0.4769ZFY16_HUMA Q7Z3T8 Zinc finger FYV 168784 4.67 1539 YES 0.4855NEK9_HUMANQ8TD19 Serine/threon 107165 5.51 979 YES 0.499WDR26_HUM Q9H7D7 WD repeat-co 72129 5.74 661 YES 0.4397GCR_HUMAN P04150 Glucocorticoid 85662 6 777 YES 0.4472GCR_HUMAN P04150 Glucocorticoid 85662 6 777 YES 0.44WDR16_HUM Q8N1V2 Cilia- and flage 68311 6.51 620 YES 0.4825SEPT7_HUMA Q16181 Septin-7 (CDC 50696 8.76 437 YES 0.3793F120A_HUMAQ9NZB2 Constitutive c 121946 9.07 1118 YES 0.4635CERK1_HUMAQ8TCT0 Ceramide kina 60008 8.54 537 YES 0.4663SACS_HUMANQ9NZJ4 Sacsin (DnaJ h 521210 6.63 4579 YES 0.4636APOB_HUMANP04114 Apolipoprotei 515648 6.61 4563 YES 0.4948APOB_HUMANP04114 Apolipoprotei 515648 6.61 4563 YES 0.4338FRY_HUMAN Q5TBA9 Protein furry h 338881 5.66 3013 YES 0.4817PKHH2_HUMAQ8IVE3 Pleckstrin hom 168262 7.5 1493 YES 0.4626TGM3L_HUMAO95932 Protein-glutam 79329 6.85 706 YES 0.4922TRI62_HUMANQ9BVG3 E3 ubiquitin-p 54276 6.17 475 YES 0.4512
IMA5_HUMANP52294 Importin subu 60338 5.07 538 YES 0.4771TTPA_HUMANP49638 Alpha-tocophe 31759 7.17 278 YES 0.4609MAST4_HUMAO15021 Microtubule-a 284487 8.85 2626 YES 0.4294SYMC_HUMA P56192 Methionine--t 101118 5.82 900 YES 0.4613FRM4B_HUMAQ9Y2L6 FERM domain 111668 8.76 1034 YES 0.4808KIF19_HUMANQ2TAC6 Kinesin-like pr 111375 8.87 998 YES 0.4902NAV1_HUMANQ8NEY1 Neuron naviga 202515 8.2 1877 YES 0.4615NAV1_HUMANQ8NEY1 Neuron naviga 202515 8.2 1877 YES 0.3409MAST2_HUMAQ6P0Q8 Microtubule-a 196499 8.41 1798 YES 0.492MAST2_HUMAQ6P0Q8 Microtubule-a 196499 8.41 1798 YES 0.4294KI26A_HUMA Q9ULI4 Kinesin-like pr 194663 9.12 1882 YES 0.4649KI26A_HUMA Q9ULI4 Kinesin-like pr 194663 9.12 1882 YES 0.4561DYH8_HUMANQ96JB1 Dynein heavy 514685 5.95 4490 YES 0.4988DYH7_HUMANQ8WXX0 Dynein heavy 461143 5.7 4024 YES 0.4571DYH5_HUMANQ8TE73 Dynein heavy 529025 5.79 4624 YES 0.4809CF204_HUMAQ5SZL2 Centrosomal p 91811 5.98 805 YES 0.4332LRCC1_HUMAQ9C099 Leucine-rich re 119596 5.62 1032 YES 0.4965LRCC1_HUMAQ9C099 Leucine-rich re 119596 5.62 1032 YES 0.4902SSH2_HUMANQ76I76 Protein phosp 158200 5.26 1423 YES 0.4371CLAP2_HUMAO75122 CLIP-associatin 141171 8.68 1294HAUS6_HUMAQ7Z4H7 HAUS augmin 108635 6.04 955 YES 0.4828RBP10_HUMAQ6VN20 Ran-binding p 67271 6.29 620 YES 0.4693FRPD1_HUMAQ5SYB0 FERM and PDZ 173410 5.14 1578 YES 0.4939RANB9_HUMAQ96S59 Ran-binding p 77863 6.31 729 YES 0.4693NF2L2_HUMAQ16236 Nuclear factor 67796 4.67 605 YES 0.4393TNR6B_HUMAQ9UPQ9 Trinucleotide 194017 6.31 1833 YES 0.3958PATL1_HUMAQ86TB9 Protein PAT1 86860 6.22 770 YES 0.4272ATX10_HUMAQ9UBB4 Ataxin-10 (Bra 53480 5.12 475 YES 0.4274CSDE1_HUMAO75534 Cold shock do 88889 5.88 798 YES 0.4927HPRT_HUMANP00492 Hypoxanthine 24582 6.21 218 YES 0.4548ITSN2_HUMA Q9NZM3 Intersectin-2 ( 193360 8.32 1697 YES 0.4416METH_HUMA Q99707 Methionine sy 140518 5.39 1265 YES 0.4021RHG07_HUMAQ96QB1 Rho GTPase-a 170605 5.95 1528 YES 0.4592F123B_HUMAQ5JTC6 APC membran 123978 4.77 1135 YES 0.4781BTK_HUMAN Q06187 Tyrosine-prote 76298 7.83 659 YES 0.4725WWC1_HUMAQ8IX03 Protein KIBRA 125296 5.65 1113 YES 0.4461SQSTM_HUMAQ13501 Sequestosome 47679 5.1 440 YES 0.4486VPS39_HUMAQ96JC1 Vam6/Vps39- 101835 6.53 886TSC2_HUMANP49815 Tuberin (Tube 200793 7.06 1807 YES 0.4954FRDA_HUMANQ16595 Frataxin, mito 23141 8.8 210 YES 0.4686PP4C_HUMANP60510 Serine/threon 35072 4.91 307 YES 0.3845KI20B_HUMA Q96Q89 Kinesin-like pr 210668 5.56 1820 YES 0.4513TMF1_HUMANP82094 TATA element 122793 4.88 1093 YES 0.471NUDT6_HUMAP53370 Nucleoside dip 35686 8.15 316 YES 0.4967HIF1A_HUMA Q16665 Hypoxia-induc 92658 5.17 826 YES 0.48HDA10_HUMAQ969S8 Histone deace 71445 5.44 669HINT1_HUMAP49773 Histidine triad 13808 6.43 126 YES 0.428
SMG7_HUMA Q92540 Protein SMG7 127325 8.87 1137 YES 0.419DTX1_HUMANQ86Y01 E3 ubiquitin-p 67416 9.67 620 YES 0.4903SRPK2_HUMAP78362 SRSF protein k 77518 4.91 688 YES 0.4206HD_HUMAN P42858 Huntingtin (Hu 347881 5.81 3142 YES 0.4142PER2_HUMANO15055 Period circadia 136596 6.04 1255 YES 0.4917MIER1_HUMAQ8N108 Mesoderm ind 60594 4.37 512 YES 0.3608AEBP1_HUMAQ8IUX7 Adipocyte enh 130896 5.05 1158 YES 0.4519NCOAT_HUMAO60502 Protein O-GlcN 102881 4.82 916 YES 0.4842KKCC2_HUMAQ96RR4 Calcium/calmo 64740 6.25 588UBX2B_HUMAQ14CS0 UBX domain-c 37074 5.49 331 YES 0.4918GLI2_HUMAN P10070 Zinc finger pro 167843 6.9 1586 YES 0.4427ERG_HUMAN P11308 Transcriptiona 54625 7.01 486 YES 0.4722API5_HUMANQ9BZZ5 Apoptosis inh 57560 5.84 524 YES 0.437HSFY1_HUMAQ96LI6 Heat shock tra 45117 6.68 401 YES 0.4867CHD4_HUMANQ14839 Chromodoma 217985 5.62 1912 YES 0.4986SRTD2_HUMAQ14140 SERTA domain 33877 4.42 314 YES 0.2679PLCB1_HUMAQ9NQ66 1-phosphatidy 138570 5.9 1216 YES 0.4018PDCD6_HUMAO75340 Programmed c 21866 5.16 191 YES 0.4586WDFY3_HUM Q8IZQ1 WD repeat an 395324 6.3 3526SYNE1_HUMAQ8NF91 Nesprin-1 (Ena 1010940 5.38 8797 YES 0.4155ZMIZ1_HUMAQ9ULJ6 Zinc finger MI 115518 7.09 1067 YES 0.4739RCC2_HUMANQ9P258 Protein RCC2 56107 9.02 522 YES 0.3511PELP1_HUMA Q8IZL8 Proline-, gluta 119596 4.29 1130 YES 0.4585GEMI5_HUMAQ8TEQ6 Gem-associate 168590 6.14 1508 YES 0.4562PHAX_HUMANQ9H814 Phosphorylate 44396 5.27 394 YES 0.4359TOPB1_HUMAQ92547 DNA topoisom 170710 6.47 1522 YES 0.4977ATN1_HUMANP54259 Atrophin-1 (D 124840 9.01 1190 YES 0.4232ZBED4_HUMAO75132 Zinc finger BE 130331 6.38 1171 YES 0.4934GLI3_HUMAN P10071 Transcriptiona 169910 7.02 1580 YES 0.4143DHX36_HUMAQ9H2U1 ATP-dependen 114805 7.57 1008 YES 0.4745TF65_HUMANQ04206 Transcription 60216 5.46 551 YES 0.4978TUT4_HUMANQ5TAX3 Terminal uridy 185213 8.3 1644 YES 0.469UBP7_HUMANQ93009 Ubiquitin carb 128293 5.33 1102 YES 0.3653UBP7_HUMANQ93009 Ubiquitin carb 128293 5.33 1102 YES 0.4642HDAC6_HUMAQ9UBN7 Histone deace 131405 5.14 1215 YES 0.492TPR_HUMAN P12270 Nucleoprotein 267205 4.97 2363TPR_HUMAN P12270 Nucleoprotein 267205 4.97 2363 YES 0.4843
No_positive No_positive scPosition Sequence Netsurp_pred Mouse_othlogRat_ortholog Mouse ortholoYES 0.4809 169 LLDAYDLSVLG EEEEEEBEBBE Q99KS6 Q5M886 LLDSYDLSVLGYES 0.4489 1818 AEIHEDSTICW EEEEEEEEBBB Q91ZB0 F1LTG2 SEIHEDSTVCW
682 KKEEELLSSIYG EEEEEBBEEBE Q9D2J7 D4A9E7 KKEEETLSSIYTV111 VMDASDPTQLEEEEEEEEEBEEB1ATY8 M0R4K4 MMDASNPTQ
47 VGSDELDSYM EEEEEBBEBBB Q9CQ28 Q5M9F5 VESDELDSYMYYES 0.2856 724 VDEDEDLTMC EEEEEEBBBBB Q8CGT2 D3ZEZ4 VDEDEKLTTLLYES 0.4507 1735 IADDEFDTMFSEEEEEBEEBBE A2AHJ4 D3Z8C5 VADDEFDTMF
181 AVGSSLHSYIWEEEEEBBEBBB Q6NV72 M0R4L9 AIDSNLHSYIW29 SLSQEEVSIIWGEEEEEEBEBBB Q9D5W4 Q5XIN9 TLSQEEVSTIW
186 YNAESDLSSVT EEEEEEBEEBEEQ3TIR1 Q5M887 YNAESDLSSVT99 VDEQNDASSLWEEEEEEBEEBB D3YX67 Q6AYU0 NDASSLWTKT91 NFSDSDDTMVEBEEEEEBBBB Q6PE65 D3ZUP4 NFSDSDDTVVA
219 RKAASQNSSLWEEEEEEEEEBBEQ6GQV0 B2GV34 RKAVSRNSCLW102 PLSSEDDTSPG EEEEEEEEEEEBQ9D9W0 M0R8Y9 PLSSEDEASPG
YES 0.4327 874 PDASEEDSFQWEEEEEEEBBEB Q3UH60 F1M8R8 PDASEEDSFQW855 PDSTEEDSFQWEEEEEEEBBEB E9PWR4 D3ZZB0 PDSTEEDSFQW166 ADKDEDTTSHSEEEEEEEEEEEEE9QM99 F1M720 ADKDEDTTSHS158 EKDEDSSSLCSQEEEEEEEEBBEEA2AF47 F1LQ91 EKDEDSSSLCSQ522 VREDDEDSVG EEEEEEEEEEEEQ148W8 D3ZRM0 VREDDEESVLS
YES 0.3398 290 LAPDADESILWEEEEEEEEBBB Q60899 Q8CH84 LAPDADESILWYES 0.4084 298 LSPEADESVLWEEEEEEEEBBB Q60900 Q76IJ9 LSPEADESVLWYES 0.3454 311 LSPDSDESVLWEEEEEEEEBBB Q61701 O09032 LSPDSDESVLWYES 0.4813 152 SMDGEDMSV EEEEEEBEBEEEQ3TY60 Q568Z1 SMDGEDMSV
372 TASDTDSSYCIPEEEEEEEEBBB Q9QYY0 D3ZAL7 TASDTDSSYCIP510 PSDSSDGTYM EEEEEEBBBBB Q3UHI0 D4A4E5 PSDSSDGTYM
YES 0.4975 492 DNENDNQTGLEEEEEEEBBBB Q8BZM1 D3ZKQ3 DNEDDNQTGL344 VDLGSDQTSCHBEEEEEEEEEEEQ8VC12 D3ZDW1 VDLGSDQTSCH509 FGVEDDDSSLABEEEEEEEEEEEO35345 F1LT58 GVEDDDSSLAP510 GVEDDDSSLAPBEEEEEEEEEEEO35345 F1LT58 GVEDDDSSLAP646 GKFSNDDSSSYEEEEEEEEEEEEE9Q0J2 P86410 GKFSNDDSSSY131 VGGTDDDSSC BEEEEEEEEEEEQ8BM65 F1M3A4 VGGTEDDSSCG543 CREDSYHSVVSBEBEEEEEEEEEQ8C0C4 D3ZTW6 CREDSCHSVM301 YMFDDDESTLGBEEEBEEEEEEEQ5DTT1 D3ZGX1 MFDDDESTLG302 MFDDDESTLG EEEEBEEEBEEEQ5DTT1 D3ZGX1 MFDDDESTLG
YES 0.4594 369 YHDATDQTWCEEEEEEEBBBB Q8BHI4 D4A7F5 YHDATDQTWC264 LHCAEDDSISV EEEEEEEEEEBEQ8R179 D3ZHJ7 LHCAEDDSISV381 HSCGDLLTSLS BEEEEEEEEEEEQ3V0M2 F1MAB0 HPCGDLLTSLS368 TEPSDDDTVSLEBEEEEEEEEEEQ3UMG5 D4A456 TEPSDDDTISLH278 FRDEDFYSILSL BEBEEEEEEEEEE9PX79 Q5XIV2 FHDEDFYSAISL665 QLEAEDDTSYFBBEEEEEEEEEEE9Q8S5 D3ZL30 QLEAEDDTSYF
1141 GDPGSDSTTCSBEEEEEEEEEEEQ69ZH9 F1M2D4 DPGSDSTTCSS182 LIPDENPSSFSGBEEEEEEEEEEEQ9D9R0 F7FD13 PDESPSTFNGS857 DMSSDNDSYHBEEEBEEEEEEEQ8CDA1 D3ZKG7 DISSDNDSYHS639 LHSASSNTSLSGBEEEEEEEEEEEQ6P7W2 P0C5J9 LHSASSNTSLYG
YES 0.4747 814 EDEGDLDTNSWEEEEEBEEEEBEQ8VE18 D3ZQ80 EDEGDLDTNSW305 PRSASGDTVFWBBBEEEEEEEE F6SEU4 Q9QUH6 PRSASGDTVFW

495 AVSHSDSTSST EEEEEEEEEEEEQ7TN83 D3ZB68 VSHSDSASSTQ185 FGDDEELSTSS EEEEEEEEEEEEQ7TN83 D3ZB68 FGDDEEPSTSS325 DSYATDSSSMWEBEEEEEEEEEEQ7TN83 D3ZB68 DSYATHSSSLW
YES 0.4951 495 HWDDEDISLLLBBEEEEEEEEEEQ8R5A6 D3Z994 WDDQDVSLLLYES 0.4583 1730 DTDNPDDSVF EEEEEEEBBBB B1AY13 F1LSM0 DTDNPDDSVFYES 0.4562 641 YGNEDEDSFCSEEEEEEEEBBEEP56399 D3ZVQ0 YGNEDEDSFCS
901 DSAGSDSTSLVBEEEEEEEEEEEB2KF50 D3ZMR2 DSAGSDTTSLID1876 ALSEDDLTVLMEEEBEBBBEEE Q8BX70 D4A4K4 ALSQDDLTVLM
YES 0.443 81 ETDDEDESFVLEEEEEEEEBBB Q0P5W1 D3ZH26 ETDDEDESFVLYES 0.4329 71 SGSDDQHTIVWEEEEEEBBBBB Q80ZK9 D4A7Z0 SGSDDQHTIVW
927 SSCTEDLSSCTSEEEBEEBEEEEEQ6NXJ0 D4AEJ5 YIDHNTRRTSWYES 0.3881 698 PSKEEDDTVNWEEEEEEEEBEBEQ8BYK8 F1LTU5 SKEEDDTANW
201 DTSNDESSILGTEEEEEEEEBBEEQ9Z0E1 D3ZVK0 SNDESSILEADIYES 0.4287 50 ATPENDNTLCPEEEEEEEEBBEEO08918 B5DFJ6 ATPENDNTLCS
130 SPWSNHQSSGEEBEEEEEEEBEQ812E0 D3ZHK8 SPWSNHQNSGYES 0.4635 570 STTGESDTVIWEEEEEEEEBBB Q6PDK8 Q6TXG2 STTGESDTVIW
237 KDDASESTSLSAEEEEEEEEEBEEQ9CRA9 Q6TA25 KDDASESTSLSAYES 0.4678 87 DHPESYHSFM EEEEEBBBBBBO88958 M0RCH5 DHPESYHSFMYES 0.4802 87 NHPESYHSYM EEEEEBBBBBBQ9CRC9 B5DEZ6 NHPESYHSYM
83 ITSDEDSSYINABEEEEEEEEEEEP29352 D4A2D5 LTSDEDSSYINA855 LSSDPDYSSSSEEEEEEEEEEEEEQ7TNC6 D4ADC4 LSSDPDYSSSSE
1873 GHGSDNSSVLSBEEEEEEEEEEEQ7TNC6 D4ADC4 GHGSDNSSVLS534 PYDGYDPSVLSBEEEEEEEEEEEP58404 F1M6V8 PYDGYDPSVLS
YES 0.3663 393 VEDEDDDTFSYEEEEEEEEBEBEE9QKZ2 D4A857 VEDEDDDTFSY285 SSLSADSSSSWEEEEEEEEEEEEQ5SWP3 F1M6E5 SSLSADSSSSW357 GEGEEDSTSASEEEEEEEEEEEEQ5SWP3 F1M6E5 GEGEDDSTSAS
YES 0.4086 1652 SMADGDNSYCEEEEEEEEBBB G3X9Y9 P70569 SMVDGENSYC879 MSSSEMWTV BBEEBEEEEEE Q69ZM6 D3ZA65 SEVWTILWHR472 YSDGDNDSINSEEEEEEEEEEEEQ8R0S2 M0R5C6 YSDGDNDSINS333 KCDDDSETSNWEEEEEEEEEEBBQ9Z1K5 D3ZXL1 KCDDDSETSNW193 GADSTDNSSSLEBEEEEEEEEEEQ8BZ60 D4AB66 GTDSTDNSSSL
1442 FLKDQDLSILSTEEEBEBBEEEE Q80U44 D4ADF6 FLKDQDISILSS438 VSCGDDFTVCVBBEEEEEEEEEBQ8K1R7 D3ZJT2 VSCGDDFTVCV451 SHEDSLTSVAWEEBEEBBBEEE Q8C6G8 F1LTR1 SHEDSLTSVAW
YES 0.4306 234 PLAGEDDSFLL EEEEEEEEBBB E9PYV1 P06536 PLAGEDDPFLL376 GSGDDNLTSLGEEEEEEBEEBEEE9PYV1 P06536 GSGEDNLTSLG
YES 0.4652 218 IGVDDDDSFFYEEBEEEEBBBB Q5F201 B1WBL5 GMADDDSFFYYES 0.3678 124 FGDAVDNSNCEEEEEEEEEBBEE9Q1G8 Q9WVC0 FGDAVDNSNCYES 0.4668 252 LPDEDLASFHWEEEEEBEEBBB Q6A0A9 D4AB03 LPDEDLASFHWYES 0.451 12 GAAEPLQSVLWEEEEEBEEBBE Q8K4Q7 D3Z9Y3 GAAEPLHSVLW
4218 EREDADNSSFLBBEEEEEEEEEEQ9JLC8 D4A1D3 EREDADNTSFLYES 0.4757 485 CTGDEDYTYLILEEEEEEBBBBB E9Q414 Q7TMA5 CTGNEDHTFLIYES 0.4177 2128 QANDYLNSFN EEEEBBEEBEB E9Q414 Q7TMA5 QIHHYLNASDW
2611 TVHEDDLSSSINEEEEEEBEEEBEE9Q8I9 E9PTY6 TVHEDDLSSSIN333 LTASDDSSSIFEBBEEEEEEEEEBQ8C115 D3ZNS6 LTASDDSSSIFE
YES 0.4704 401 EVNADYITWLWEEEBEBBBBBBQ8BM11 F1M4T7 EVNADYITWLWYES 0.4678 436 FYNADDMSW EEEBEEBBBBBQ80V85 M0R947 FYNADDMSW
YES 0.459 510 GVEEDDPSIVPEEEEEEEEBBEEQ60960 P83953 GTEDEDSSIAP271 MKSEDYLSSISEBEEEEEEEEEEEQ8BWP5 P41034 MKSEDYLSSISE871 QLESEDDTSYF BBEEEEEEEEEEQ811L6 M0R3L1 QLESEDDTSYF
YES 0.4416 81 GWEQDDLTNQEEEEEEBBEEB Q68FL6 D3Z941 GWEQDDLTNQ926 TPAHSSYTSCY EEEEEEBEEBB Q920B0 F1LZB7 PVHSPYTSYYG847 AASEDNLSSST EEEEEEEEEEEEQ99PT9 D4A976 AISEDNLSSSTG415 TGWDESSSISS EEEEEEEEBEEEQ8CH77 F1LYK3 TGWDESSSISS
YES 0.3835 407 MTEDDDITTGWEEEEEEBEEBB Q8CH77 F1LYK3 MTEDDDITTGW1722 HPSYEDPSQGWBBBBEEEEEEE E9Q1Q1 D3ZAR2 HPLLEVPSQNW
810 QLESEDDTSYF BBEEEEEEEEEEE9Q1Q1 D3ZAR2 QLESEDDTSYF780 LPGDPDYSSSS EEEEEEEEEEEEQ52KG5 D3ZUT0 LSGDPDYSSSS
1635 GHGSDNSSVLSBEEEEEEEEEEEQ52KG5 D3ZUT0 GHGSDNSSVLSYES 0.4769 3452 GLPGDDLSIQNEEEEEEBEBEEBQ91XQ0 F1MAM6 GLPGDDLSIQN
872 LQPSDDSTVSSEEEEEEEEBEEBL7N1Y0 Q63170 LLPSDDSTVFSFYES 0.46 3587 GLPNDDLSIQNEEEEEEBEBEEBQ8VHE6 M0R8U1 GPSGSGKTTCI
29 FPAGPDYSSAWEEEEEEBEEBB E9QA61 F7EXF4 FPAGSDYSSAW422 QSHSEDNTYQ EEBEEEEEEEEEG5E8Y2 E9PTY0 SEDNTYQSLVE
21 VENEDGDSSCGBEEBBEEEEEE G5E8Y2 E9PTY0 IEREDGDSSCG134 DFSSNDSSTCT BEBEEEEEEEEEQ5SW75 F1M4Q5 DFSSNDSSTCT
YES 0.4958 974 SKKDDGDSMCEEEEEEEEBBB F7DCH5 Q99JD4 KKEDGDTICSG751 NKPSTNKTMLWBBEEBEEEEEE Q6NV99 D3ZET0 EPSTHKTLLWD149 YHGDDGHSFC BEEEEEEEEEEEQ6VN19 D3Z7Z5 YHGDDGHSFC708 GSHADYYSLCSEEEEEBBEBBE A2AKB4 D3ZW88 GSRTDYYNLCS261 YHGDDGHSFC BEEEEEEEEEEEE9Q5D6 F1LVV3 YHGDDGHSFC256 AFEDSFSSILST BEEEEEEEEEEEQ60795 O54968 GFEDSFSSILST
1800 WGPPNYSSSLWBBEBEEEEEEE Q8BKI2 F1LV37 WGPPNYSSSLW123 PQPGSLNSSIWBBEEEEEEEEEEQ3TC46 B5DF93 PQPGSLNSSIW
YES 0.4172 151 SRNEDSQSIVWEEEEEEEEBBB P28658 Q9ER24 SRNEESQSIVW595 ITEEADPTIYSGEEEEEEEEBBEEQ91W50 P18395 ITEEANPTIYSG123 VIGGDDLSTLT BEEEEEEEEEEEP00493 F1LNY0 VIGGDDLSTLT
YES 0.4995 1189 MTTDSDPSQQEEEEEEEEEEBBE9QNG1 M0R7A6 MTTDSDPSQQYES 0.4905 1122 EDDGDDYSSIMEEEEEEBBEBB A6H5Y3 Q9Z2Q4 EDDGDDYSSIM
636 NLAGNDDSFGEEBEEEEEEEEEQ9R0Z9 Q63744 HFVGNDDSFSS795 CSSDSDSSFTQ EEEEEEEEBEEEQ7TS75 D3ZCP6 SSDSDSSFTQN553 VLDDEYTSSVGEEEEEBBEEBE P35991 E9PTF7 VLDDEYTSSVG950 NRSDSDSSTLS EEEEEEBEEEEEQ5SXA9 F1M6U0 NRSDSDSSTLS339 SGGDDDWTH BEEEEEEEEEEEQ64337 O08623 SGGDDDWTH
YES 0.4859 226 AVGQDDLTVVEEEBBBBBEEE Q8R5L3 E9PT04 AVGQDDLTVVYES 0.4796 1607 CGEDGQFTYCWEEEEEEBBBBB Q7TT21 P49816 CGEDGQFTYCW
126 TFEDYDVSFGSEEEEBEBEBEE O35943 D3ZYW7 TLEDYDVSFGDYES 0.478 203 LWSDPEDTTG EBEEEEEBEEB P97470 Q5BJ92 LWSDPEDTTG
235 VHNDSDDTLYGBBEBEEEEEEE Q80WE4 D3ZX13 IHNDSYDVLCG59 PVSGGWDTSTEBEBEEEEEEEBB9EKI3 G3V9T2 PVSGGWDTST
123 HHAESDSSTLTBEBBEEEEEEE Q8CH40 P70563 HHAKPHSSTLT384 KVESEDTSSLFDBBEEEEEEEEEEQ61221 O35800 KVESEDTSCLFD
YES 0.491 188 YLFEDDPSVLY BBBBBEEEEEE Q6P3E7 Q569C4 YIFNDDPSVLYYES 0.4121 72 VAEDDDESLLGEEEEEEEEBBEEP70349 D4A269 VADDDDESLLG

408 HPHEEDLSSISABEEEEEEEEEEEQ5RJH6 D4ACJ3 HPREDDLSNTNYES 0.4781 574 NTTGESDTVVWEEEEEEEEBBB Q61010 M0RA58 NTTGESDTVVW
426 PNAESDYTYSS EEEEEEEEEEEEO54781 B1WBT4 PNAESDYTYSS1389 QAEQENDTSGEBBEEEEEBEE G3X9H5 P51111 QAEQERDASG
35 LQEDVDMSSGEEEEEEEEEEEEO54943 Q9Z301 LQEDVDMSSGYES 0.4267 131 DADNDDNSGCEEEEEEEEEBEEQ5UAK0 E9PT68 DADNDDNSGCYES 0.4338 707 EDFPDLNSVLWEEBEEBEEBBB Q640N1 A2RUV9 EDFPDLNSVLW
352 MTDSEDSTVSIEBEBEEEEEEEEQ9EQQ9 Q8VIJ5 MTDSEDSTVSIYES 0.4981 189 AYNENDNTYYABBBBBEEEEEE Q8C078 O88831 AYNENDNTYYAYES 0.4752 242 SPEEEDKSILNAEEEEEEEEBBEEQ0KL01 P0C627 SPEEEDKSILNA
1522 IMDDGDHSSL EEEEEEBEEBB Q0VGT2 F1M2B7 IMDDGDHSSL334 LLSDSSNSSCIT EEEEEEEEEBBEP81270 D4A3D6 LLSDSSNSNCIT461 KRASEDTTSGSEEEEEEEEEEEEO35841 B1WC49 KRTSEDTSSGS
95 VESDQFKSISWEBEEEEEEEEEEQ80Y37 Q5XIQ7 VGSDKFKSIWW1245 NKEGEDSSVIHEEEEEEEEBBB Q6PDQ2 E9PU01 CPSSYHIHCLN
YES 0.2927 142 LLEDDDDTFCTEEEEEEEEBBEEQ9JJG5 F7F8X3 LLEDDNDDTFCYES 0.4446 36 VKWDDDSTIV EEBEEEEEBBE Q9Z1B3 P10687 VKWDDDSTIVYES 0.44 26 GAALPDQSFLWEEEBEEEEBBB P12815 G3V7W1 GAALPDQSFLWYES 0.4843 918 ALADEDHSLHPEEEEEBEEEEEEQ6VNB8 D4A7D8 ALADEDHSLHP
8680 SSADELDTSGS EEEEEEEEEEBBQ6ZWR6 D3ZEA8 SSADELDTSGS887 GTNSNDYSSQ EEEBEEEEEEEEQ6P1E1 D4AE97 TSSNDYSSQGN
YES 0.3994 450 IVAADESTISW EEEEEEEEBEBEQ8BK67 F1LVV4 IVAADESTISW787 LESDSDDSVVIVBBBEEEEEEEB Q9DBD5 Q56B11 LESDSDDSVVIV
YES 0.4393 200 GHDDEIHSIAWEEEEEBBEBBB Q8BX17 D3ZGD0 GHDDEIHSIAWYES 0.4819 77 FSDSDDDSCLWEEEEEEEEBBB Q9JJT9 Q63068 DSDDDCSLWK
535 SLQEENQSSVSEEEEEEEEEEBEQ6ZQF0 D3ZRG0 SVQEENKSSVS148 VENDSDSSSGLEEEEEEEEEEEEO35126 P54258 VENDSDSSSGL201 ADAGDLSTILS EEBEEBBEEEE Q80WQ9 D3ZUV0 ADVGDLSTVLS
1516 IIDDGDHSSLMEEEEEEBEEBEEQ61602 F1M9H1 IIDDGDHTSLM326 LQSDPYLSSVS EEEEEEBEEBEEQ8VHK9 D4A2Z8 LQSDSRLSSVS535 LSGDEDFSSIADBEEEEEEEEEEEQ04207 Q7TQN4 LSGDEDFSSIAD798 ADHGQDSSSLSBEEEEEEEBEEBB2RX14 F1M466 ANHGQDSSSL128 CNAESDSTSW BEBEEEEEEEEEE9PXY8 Q4VSI4 CNAESDSTSW127 QCNAESDSTSWEBEBEEEEEEEEE9PXY8 Q4VSI4 CNAESDSTSW
YES 0.4733 665 HMFEDDPSVL EEBEEEBEBBB Q9Z2V5 D3ZVD8 HIFEDDPSVLYVYES 0.4878 1979 DDDDEDDTGMEEEEEEBBBBB F6ZDS4 F1MA98 DDDEEDDTGM
2133 FFDDEDRTVPSEEEEEEEEBEEEF6ZDS4 F1MA98 FFDDEDRTVPS
Rat ortholog s Subcellular locIntramembranTransmembra Topological doTpl2 [D270A]/WT (log 2 ratio)LLDSYDLSVLG 0 0 0 0 NASEIHEDSTVCW 0 0 0 0 NAKKEEGTLSSIYT 0 0 0 0 NAMMDASNPTQ 0 0 0 0 NAVESDELDSYMY 0 0 0 0 NAVDEDEELSTCL 0 0 0 0 NAVADDEFDTMF 0 0 0 0 NAAVDSSLHSYIW 0 0 0 0 NASLSQEEVSTIW 0 0 0 0 NAYNAESDLSSVT 0 0 0 0 NANDASSLWTKT 0 0 0 0 NANFSDSDDTVVA 0 0 0 0 NARKAASRNSRLW 0 0 0 0 NAPLSSEDDASPG 0 0 0 0 NAPDASEEDSFQW 0 0 0 0 NAPDSTEEDSFQW 0 0 0 0 NAADKDEDTTSHS 0 0 0 0 NAEKDEDSSSLCSQ 0 0 0 0 0.112VREDDEESVLS 0 0 0 0 NALAPDADESILW 0 0 0 0 NALSPEADESVLW 0 0 0 0 NALSPDSDESVLW 0 0 0 0 NASMDGEDMSV 0 0 0 0 NATASDTDSSYCIP 0 0 0 0 NAPSDSSDGTYM 0 0 0 0 NADNEDDNQTGL 0 0 0 0 NAVDLGSDQTSCH 0 0 0 0 NAGVEDDDSSLAP 0 0 0 0 NAGVEDDDSSLAP 0 0 0 0 NAGKFSNDDSSSY 0 0 0 0 NAVGGTEDDSSCG 0 0 0 0 NACREDSCHSVM 0 0 0 0 NAMFDDDESTLG 0 0 0 0 NAMFDDDESTLG 0 0 0 0 NAYHDATDQTWC 0 0 0 0 NALHCAEDDSISV 0 0 0 0 NASCGDLLTSLSN 0 0 0 0 NATEPSDDDTISLH 0 0 0 0 NAFHDEDFYSALS 0 0 0 0 NAQLEAEDDTSYF 0 0 0 0 NADPGSADSTTCS 0 0 0 0 NAPNESPSTFSGS 0 0 0 0 NADVSSDNDSYHS 0 0 0 0 NALHSASSNTSLC 0 0 0 0 NAEDEGDLDTNSW 0 0 0 0 NAPRSASGDTVFW 0 0 0 0 NA
VSHSDSASSTQ 0 0 0 0 NAFGDDEEPSTSS 0 0 0 0 NADPYATHNSSM 0 0 0 0 NAWDDQDVSLLL 0 0 0 0 NADTDNPDDSVF 0 0 0 0 NAYGNEDEDSFCS 0 0 0 0 NADSAGSDTTSLID 0 0 0 0 NAALSQDDLTVLM 0 0 0 0 NAETDDEEESFVL 0 0 0 0 NASGSDDQHTIVW 0 0 0 0 NAYIDHNTRRTSW 0 0 0 0 NASKEEDDTVNW 0 0 0 0 NASNDESSILEADI Cytoplasm {EC 0 0 0 NAATPENDNTLCS Cytoplasm {EC 0 0 0 NASPWSNHQSSG Cytoplasm {EC 0 0 0 NASTTGESDTVIW Cytoplasm {EC 0 0 0 NAKDDASESTSLSA Cytoplasm {EC 0 0 0 NAEHPESYHSFMW Cytoplasm {EC 0 0 0 NANHPESYHSYM Cytoplasm {EC 0 0 0 NALTSDEDSSYINA Cytoplasm {EC 0 0 0 NALSSDPDYSSSSE Cytoplasm {EC 0 0 0 NAGHGSDNSSVLS Cytoplasm {EC 0 0 0 NAPYDGYDPSVLS Cytoplasm {EC 0 0 0 NAVEDEDDDTFSY Cytoplasm {EC 0 0 0 NASSLSADCSSSW Cytoplasm {EC 0 0 0 NAGEGEDDSTSAS Cytoplasm {EC 0 0 0 NASMVDGENSYC Cytoplasm {EC 0 0 0 NAVAGSEVWTILW Cytoplasm {EC 0 0 0 NAYSDGDNDSINS Cytoplasm {EC 0 0 0 NAKCDDDSETSNW Cytoplasm {EC 0 0 0 NAGTDSTDNSSSL Cytoplasm {EC 0 0 0 NAFLKDQDISLSSS Cytoplasm {EC 0 0 0 NAVSCGDDFTVCV Cytoplasm {EC 0 0 0 NASHEDSLTSVAW Cytoplasm {EC 0 0 0 NAPLAGEDDPFLL Cytoplasm {EC 0 0 0 NAGSGEDSLTSLG Cytoplasm {EC 0 0 0 NAGMADDDSFFY Cytoplasm {EC 0 0 0 NAFGDAVDNSNC Cytoplasm {EC 0 0 0 NALPDEDLASFHW Cytoplasm {EC 0 0 0 NAGAAEPLQSVLW Cytoplasm {EC 0 0 0 NAEREDADNASFL Cytoplasm {EC 0 0 0 NACMGDEDRTFL Cytoplasm {EC 0 0 0 NAQIHDYLNASDW Cytoplasm {EC 0 0 0 NATVHEDDLSSSIN Cytoplasm {EC 0 0 0 NALTASDDSSSIFE Cytoplasm {EC 0 0 0 NAEVNADYITWLW Cytoplasm {EC 0 0 0 NAFYNADDMSW Cytoplasm {EC 0 0 0 NA
GTEDEDSSIAP Cytoplasm {EC 0 0 0 NAMKSEDYLSSISE Cytoplasm {EC 0 0 0 NAQLESEDDTSYF Cytoplasm {EC 0 0 0 NAGWEQDDLTNQ Cytoplasm {EC 0 0 0 NAPVHSPYTSYYG Cytoplasm, cy 0 0 0 NAAISEDNLSSSTG Cytoplasm, cy 0 0 0 NATGWDESSSISS Cytoplasm, cy 0 0 0 NAMTEDDDITTGW Cytoplasm, cy 0 0 0 NAHPSYEDPSQGW Cytoplasm, cy 0 0 0 NAHPLLEVPSQNW Cytoplasm, cy 0 0 0 NALSGDPDYSSSS Cytoplasm, cy 0 0 0 NAGHGSDNSSVLS Cytoplasm, cy 0 0 0 NAGLPGDDLSIQN Cytoplasm, cy 0 0 0 NALQPSDDSTVFS Cytoplasm, cy 0 0 0 NAGPSGSGKTSCI Cytoplasm, cy 0 0 0 NAFPAGSDYSSAW Cytoplasm, cy 0 0 0 NASEDNTYQSLVE Cytoplasm, cy 0 0 0 NAIEPEDGDSSCK Cytoplasm, cy 0 0 0 NADFSSNDSSTCT Cytoplasm, cy 0 0 0 NAKKEDGDTVCSG Cytoplasm, cy 0 0 0 NASEPSTHKTLLW Cytoplasm, cy 0 0 0 NAYHGDDGHSFC Cytoplasm, cy 0 0 0 NAGSRTDYYSLCS Cytoplasm, cy 0 0 0 NAYHGDDGHSFC Cytoplasm, cy 0 0 0 NAGFEDSFSSILST Cytoplasm, cy 0 0 0 NAWGPPNYSSSLW Cytoplasm, P- 0 0 0 NAPQPGSLNSSIW Cytoplasm, P- 0 0 0 NASRNEDSQSIVW Cytoplasm, pe 0 0 0 NAITEEANPTIYSG Cytoplasm. 0 0 0 NAVIGGDDLSTLT Cytoplasm. 0 0 0 NAMTTDSDPSQQ Cytoplasm. 0 0 0 NAEDDGDDYSSIM Cytoplasm. 0 0 0 NAHFVGNEDSFSS Cytoplasm. Ce 0 0 0 NASSDSDSSFTQN Cytoplasm. Ce 0 0 0 NAVLDDEYTSSVG Cytoplasm. Ce 0 0 0 NAYIDHRNRTTSW Cytoplasm. Cy 0 0 0 NASGGDDDWTH Cytoplasm. La 0 0 0 NaNAVGQDDLTVV Cytoplasm. Ly 0 0 0 NACGEDGQFTYCW Cytoplasm. M 0 0 0 NATLKDYDVSFGD Cytoplasm. M 0 0 0 NALWSDPEDTTG Cytoplasm. N 0 0 0 NAIHNDSYDILCG Cytoplasm. N 0 0 0 NAVSGGWDTSTW Cytoplasm. N 0 0 0 NAHHAEPHLSTLT Cytoplasm. N 0 0 0 NAKVESEDTSCLFD Cytoplasm. N 0 0 0 NAYIFEDDPSVLYF Cytoplasm. N 0 0 0 NAVADDDDESLLG Cytoplasm. N 0 0 0 NA
HPREDDLSSTN Cytoplasm. N 0 0 0 NANTTGESDTVVW Cytoplasm. N 0 0 0 NAPNAESDYTYSS Cytoplasm. N 0 0 0 NAQADQEHDASG Cytoplasm. N 0 0 0 NALQEDVDMSSG Isoform 1: Nu 0 0 0 NADADNDDNSGC Isoform 1: Nu 0 0 0 NAEDFPDLNSVLW Isoform 1: Se 0 0 0 NAMTDSEDSTVSI Isoform 3: Nu 0 0 0 NAAYNENDNTYYA Nucleus {ECO 0 0 0 NASPEEEDKSILNA Nucleus {ECO 0 0 0 NAIMDDGDHSSL Nucleus {ECO 0 0 0 NALLSDSSNSNCIT Nucleus {ECO 0 0 0 NAKRTSEDTSSGS Nucleus {ECO 0 0 0 0.4VGSDKFKSIWW Nucleus {ECO 0 0 0 NACPSSYHIHCLN Nucleus {ECO 0 0 0 NALLEGDDDTFCT Nucleus {ECO 0 0 0 NAVKWDDDSTIV Nucleus mem 0 0 0 NAGAALPDQSFLW Nucleus mem 0 0 0 NAALADEDHSLHP Nucleus mem 0 0 0 NASSADEVDTSGS Nucleus oute 0 TRANSMEM 8 TOPO_DOM 1 NATSSNDYSSQGS Nucleus spec 0 0 0 NAIVAADESTISW Nucleus, nucl 0 0 0 NALESDSDDSVVIV Nucleus, nucl 0 0 0 NAGHDDEIHSIAW Nucleus, nucl 0 0 0 NADSDDDCSLWK Nucleus, nucl 0 0 0 NASVQEENKSSVS Nucleus. Cyto 0 0 0 NAVENDSDSSSGL Nucleus. Cyto 0 0 0 NAADVGDLSTML Nucleus. Cyto 0 0 0 NAIIDDGDHTSLM Nucleus. Cyto 0 0 0 NALQSDSRLSSVS Nucleus. Cyto 0 0 0 NALSGDEDFSSIAD Nucleus. Cyto 0 0 0 NAADHGQDSSSLS Nucleus. Cyto 0 0 0 NACNAESDSTSW Nucleus. Cyto 0 0 0 NACNAESDSTSW Nucleus. Cyto 0 0 0 NAFIFDQDPSVLY Nucleus. Cyto 0 0 0 NADDDEEDDTGM Nucleus. Nuc 0 0 0 NAFFDDEDRTVPS Nucleus. Nuc 0 0 0 NA


















