
New J/ suppression from 2008 d+Au data compared to theory (Colorado, LANL, FSU)

of April 12, 2018.This information is current as AP-1
Superinduction of ATF-3 and Suppression of viaγMonocytes Is Suppressed by IFN-
Matrix Metalloproteinases in Human Lipopolysaccharide-Induced Expression of
IvashkivHao H. Ho, Taras T. Antoniv, Jong-Dae Ji and Lionel B.
http://www.jimmunol.org/content/181/7/5089doi: 10.4049/jimmunol.181.7.5089
2008; 181:5089-5097; ;J Immunol
Referenceshttp://www.jimmunol.org/content/181/7/5089.full#ref-list-1
, 27 of which you can access for free at: cites 72 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2008 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Lipopolysaccharide-Induced Expression of MatrixMetalloproteinases in Human Monocytes Is Suppressedby IFN-� via Superinduction of ATF-3 and Suppressionof AP-11
Hao H. Ho,*† Taras T. Antoniv,* Jong-Dae Ji,*‡ and Lionel B. Ivashkiv2*§
Matrix metalloproteinases (MMPs) are induced during inflammatory responses and are important for immune regulation, an-giogenesis, wound healing, and tissue remodeling. Expression of MMPs needs to be tightly controlled to avoid excessive tissuedamage. In this study, we investigated the regulation of MMP expression by inflammatory factors in primary human monocytesand macrophages. IFN-�, which augments inflammatory cytokine production in response to macrophage-activating factors suchas TLR ligands, instead broadly suppressed TLR-induced MMP expression. Inhibition of MMP expression was dependent onSTAT1 and required de novo protein synthesis. IFN-� strongly enhanced TLR-induced expression of the transcriptional repressoractivating transcription factor (ATF-3) in a STAT1-dependent manner, which correlated with recruitment of ATF-3 to theendogenous MMP-1 promoter as detected by chromatin immunoprecipitation assays. RNA interference experiments furthersupported a role for ATF-3 in suppression of MMP-1 expression. In addition, IFN-� suppressed DNA binding by AP-1 tran-scription factors that are known to promote MMP expression and a combination of supershift, RNA interference and overex-pression experiments implicated AP-1 family member Fra-1 in the regulation of MMP-1 expression. These results define anIFN-�-mediated homeostatic loop that limits the potential for tissue damage associated with inflammation, and identify tran-scriptional factors that regulate MMP expression in myeloid cells in inflammatory settings. The Journal of Immunology, 2008,181: 5089–5097.
S ince the discovery of the first matrix metalloproteinase(MMP)3 in the regressing tadpole tail by Gross andLapiere in 1962 (1), the list of functions assigned to var-
ious MMPs has been expanding rapidly. MMPs were initiallythought to be a group of enzymes with the sole function of regu-lating extracellular matrix (ECM) composition. However, intenseresearch in recent years has shown that ECM degradation is not thesole and possibly not the main function of these proteinases. In-deed, plenty of evidence supports roles of MMPs in physiologicalprocesses such as embryogenesis and wound healing/tissue remod-eling and also in pathological processes that include arthritis, pul-monary diseases, cardiovascular ailments, and cancer (2–7). In ad-dition to ECM remodeling MMPs regulate signal transduction andgrowth factor function, and recent findings also indicate that
MMPs are immunoregulators by acting on inflammatory cyto-kines, chemokines, and other immune proteins (8–18).
Not surprisingly, the activity of MMPs is tightly regulated atmultiple levels that include gene expression, compartmentaliza-tion, proenzyme activation, and enzyme inactivation, catabolism,and clearance (3, 18). Expression of many MMPs is normally lowor absent in healthy tissues, but they are induced when tissue re-modeling is required, during inflammation, and in disease statessuch as rheumatoid arthritis and cancer in which high levels ofMMPs may signify a poor prognosis in human patients (19). MMPgene expression is mainly regulated at the transcriptional level inresponse to hormones, growth factors, cytokines, and cell-cell/cell-matrix interactions (3, 5, 20). Though the expression of MMPs indiseased states is intended to restore normal homeostasis, in a set-ting such as inflammation the up-regulation of various MMPs maylead to more damage than repair (5).
Rheumatoid arthritis and atherosclerosis are disorders of chronicinflammation, and macrophages are central players in both of thesepathological conditions (21–23). Though the importance of mac-rophage-derived MMPs in cartilage and/or bone destruction inrheumatoid arthritis has not been fully elucidated, their role inatherosclerosis progression is well documented (21, 24). In ath-erosclerosis, macrophages accumulate lipid to form foam cells.The accumulation of foam cells in an atheroma is reversible anddoes not in itself cause clinical consequences. However, the se-cretion of MMPs by the atheromatous plaque-associated macro-phages can cause plaque destabilization, leading to acute coronaryevents with high morbidity and mortality (22, 23, 25). Among theMMPs produced by macrophages, MMP-1 (collagenase-1),MMP-2 (gelatinase A), MMP-3 (stromelysin-1), MMP-7 (matri-lysin-1), MMP-9 (gelatinase B), MMP-12 (metalloelastase), and
*Arthritis and Tissue Degeneration Program, Department of Medicine, Hospital forSpecial Surgery, †Department of Pathology, New York University School of Medi-cine, New York, NY 10016; ‡Division of Rheumatology, College of Medicine, KoreaUniversity, Seoul, Korea; and §Graduate Program in Immunology, Weill GraduateSchool of Medical Sciences of Cornell University, New York, NY 10021
Received for publication September 11, 2007. Accepted for publication July 23, 2008.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by grants from the National Institutes of Health (to L.B.I.).2 Address correspondence and reprint requests to Dr. Lionel B. Ivashkiv, Departmentof Medicine, Hospital for Special Surgery, 535 East 70th Street, New York, NY10021. E-mail address: [email protected] Abbreviations used in this paper: MMP, matrix metalloproteinase; ECM, extracel-lular matrix; ATF, activating transcription factor; PGE2, prostaglandin E2; RNAi,RNA interference; eGFP, enhanced GFP; shRNA, short hairpin RNA; siRNA, shortinterfering RNA duplex; ChIP, chromatin immunoprecipitation; GSK-3, glycogensynthase kinase 3.
Copyright © 2008 by The American Association of Immunologists, Inc. 0022-1767/08/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
MMP-13 (metalloproteinase-13) have been associated with athero-sclerotic plaque instability and rupture (22, 23, 25–27). Therefore,understanding the regulation of MMPs in inflammatory conditionsis crucial in formulating a therapeutic intervention to prevent dis-ease progression.
Most studies on regulation of MMP expression have been per-formed using fibroblasts, chondrocytes, osteoblasts, and cell linessuch as HEK293, HeLa and NIH-3T3 cells (7, 28–45), with veryfew studies using monocytes/macrophages (46–48). BecauseMMP expression is regulated differently in the various cell typesand cell lines studied to date, we were interested in understandingMMP regulation in primary human monocytes and macrophagesthat are relevant for inflammatory diseases, and defining mecha-nisms that inhibit MMP expression. In this study, we found that inprimary human macrophages IFN-� globally inhibits induction ofMMPs by Toll-like receptors, innate immune receptors that play akey role in activation of macrophages by microbial products andendogenous ligands that include ECM degradation products. Fur-ther analysis of the regulation of MMP-1 showed that the mech-anism of IFN-�-mediated inhibition involves induction of the tran-scriptional repressor activating transcription factor (ATF)-3 andsuppression of DNA binding by AP-1 proteins. Prostaglandin E2
(PGE2) also down-regulated LPS-induced MMP-1 expression, andsuppression by PGE2 and IFN-� converged on inhibition of AP-1proteins and regulation of AP-1 family member Fra-1. These re-sults define mechanisms that inhibit MMP expression in primaryhuman macrophages.
Materials and MethodsBiological reagents and cell culture
PBMC were obtained from whole blood from disease-free volunteers bydensity gradient centrifugation with Ficoll (Invitrogen). CD14� monocyteswere purified from fresh PBMCs with anti-CD14 magnetic beads (MiltenyiBiotec), as recommended by the manufacturer. Purity of monocytes was�97% as verified by FACS, and freshly isolated monocytes were used inmany experiments, as noted in text and figure legends. Some experimentswere performed using macrophages, which were derived by culturingmonocytes in RPMI 1640 medium (Invitrogen) supplemented with 10%FBS (HyClone) in the presence of 10 ng/ml of human M-CSF (PeproTech).THP-1 human monocytic cells were obtained from ATCC and cultured inRPMI 1640 medium (Invitrogen) supplemented with 10% FBS (HyClone).LPS (100 ng/ml), PMA (100 ng/ml), LiCl (20 mM), cycloheximide (20�g/ml), and SB216763 (10 �M) were purchased from Sigma-Aldrich.PGE2 was obtained from Sigma-Aldrich and Calbiochem.
Lentiviral gene transduction and RNA interference (RNAi)
A lentivirus-based vector expressing the human STAT1 or Fra-1 cDNAdriven by a human phospho-glycerol kinase promoter was used to generaterecombinant lentiviral particles as described (49). A construct that con-tained a transcription cassette encoding enhanced GFP (eGFP) driven bythe human phospho-glycerol kinase promoter was used to generate controlviral particles for STAT1 and Fra-1 expression experiments. RNAi in theTHP-1 monocytic cell line was performed using lentiviral transduction tostably express shRNAs that targeted STAT1 or ATF-3, or DSRed2 as acontrol. For STAT1 RNA interference, oligonucleotides encoding severaldifferent short hairpin RNAs (shRNAs) that target human STAT1 werecloned into the lentivirus-based RNAi vector pLL3.7 that also contains atranscription cassette encoding eGFP driven by a CMV promoter (49).Lentivirus-based RNAi vectors against ATF3 were purchased from Sigma-Aldrich. Constructs that were effective in suppressing STAT1 or ATF-3expression were identified using transient cotransfection of HEK 293Tcells with expression plasmids encoding the respective genes. The con-structs that were most effective in HEK 293T cells (containing the shRNAsequences 5�-GCGTAATCTTCAGGATAAT-3� or 5�-ACCTTGCAGAACAGAGAAC-3� for human STAT1 and Sigma-Aldrich’s TRC Numbersof 0000013570 and 0000013572 for human ATF-3) were used to generaterecombinant lentiviral particles. Lentiviral particles encoding interferingshRNA against red fluorescence protein DSRed2 (5�-GTGGGAGCGCGTGATGAAC-3�) were used as a control for STAT1/ATF3 RNAi experi-ments. THP-1 cells were incubated overnight with recombinant lentiviral
particles at a ratio of 1:50 in the presence of 4 �g/ml polybrene. Theefficiency of transduction was evaluated using flow cytometry and fluores-cence microscopy to monitor eGFP expression and was typically �90%.For RNAi experiments with primary human monocytes, we used nucleo-fection of short interfering RNA duplexes (siRNA) that targeted GSK3� orFra-1, or control siRNAs. GSK3�- and Fra-1-specific prevalidated siRNAsand nontargeting control siRNAs were purchased from Dharmacon.siRNAs were transfected into primary human macrophages using theAmaxa Nucleofector device set to program Y-001 with the Human Mono-cyte Nucleofector Kit (Amaxa).
Real-time quantitative RT-PCR
For real-time PCR, total RNA was extracted using an RNeasy mini kit and1 �g of total RNA was reverse transcribed using a First Strand cDNA
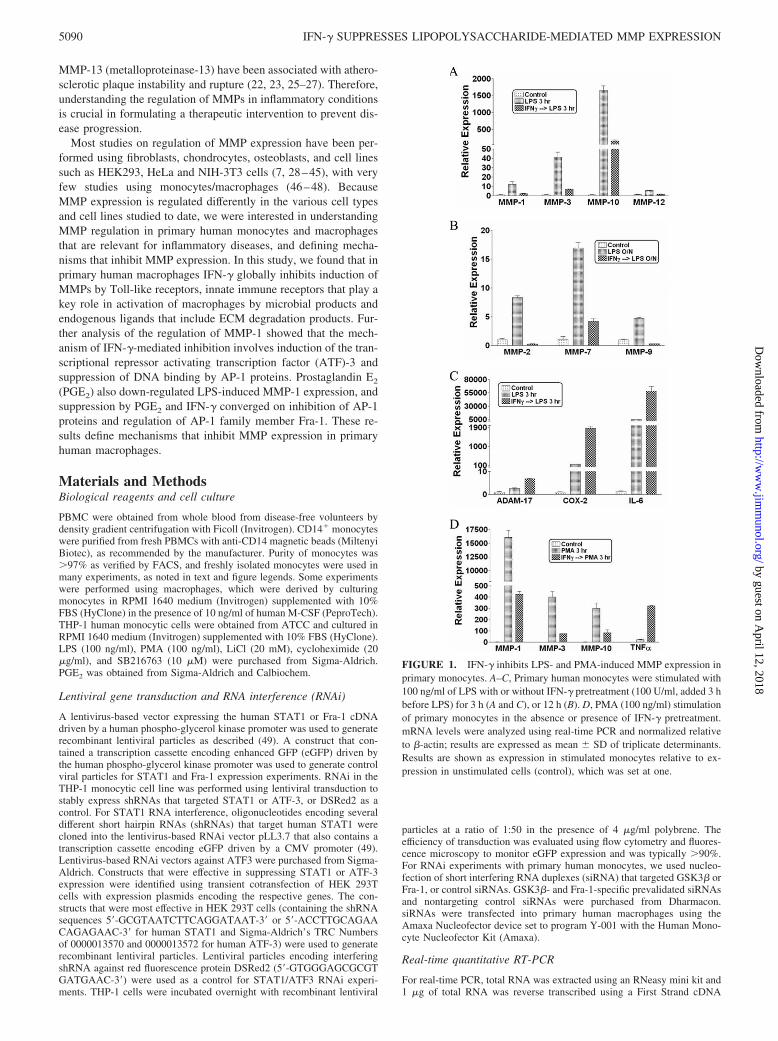
FIGURE 1. IFN-� inhibits LPS- and PMA-induced MMP expression inprimary monocytes. A–C, Primary human monocytes were stimulated with100 ng/ml of LPS with or without IFN-� pretreatment (100 U/ml, added 3 hbefore LPS) for 3 h (A and C), or 12 h (B). D, PMA (100 ng/ml) stimulationof primary monocytes in the absence or presence of IFN-� pretreatment.mRNA levels were analyzed using real-time PCR and normalized relativeto �-actin; results are expressed as mean � SD of triplicate determinants.Results are shown as expression in stimulated monocytes relative to ex-pression in unstimulated cells (control), which was set at one.
5090 IFN-� SUPPRESSES LIPOPOLYSACCHARIDE-MEDIATED MMP EXPRESSION
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Synthesis kit (Fermentas). Real-time, quantitative PCR was performed us-ing iQ SYBR-Green Supermix and iCycler iQ thermal cycler (Bio-Rad)following the manufacturer’s protocols. Triplicate reactions were run foreach sample and mRNA levels were normalized relative to �-actin. Thegeneration of only the correct size amplification products was confirmedusing agarose gel electrophoresis.
Chromatin immunoprecipitation (ChIP) assays
ChIP assays were performed using the ChIP-IT Express Kit (Active Motif)according to the manufacturer’s instructions. Chromatin was prepared from2 � 107 primary human monocytes per condition and was sheared to anaverage size of 500 bp using a Missonics Sonicator 3000. Rabbit poly-clonal Abs against ATF-3 (sc-188) or control rabbit IgG (sc-2027) (SantaCruz Biotechnology) were used for immunoprecipitation; each immuno-precipitation contained chromatin derived from 2 � 106 monocytes. Im-munoprecipitates were subjected to PCR with primers that span the ATFsite located 2017 bp upstream of the transcription start site of the MMP1gene. Primer sequences are: upstream, 5�-AGGGATTTTGTTTAAGTAAAGG-3�; downstream, 5�-GCATGTGACCATCTGTGATT-3�.
Immunoblotting and EMSA
Whole cell and nuclear extracts were obtained, and protein levels quanti-tated using the Bradford assay (Bio-Rad), as previously described (50). Forimmunoblotting, 30 �g of THP-1 whole cell lysates were fractionated on7.5% polyacrylamide gels using SDS-PAGE, transferred to polyvinylidenefluoride membranes (Millipore), and ECL was used for detection. ForEMSA, 5 �g of nuclear extracts from primary monocytes were incubatedfor 15 min at room temperature with 0.5 ng of 32P-labeled double strandedAP-1 (5�- AAA GCA TGA GTC AGA CAC - 3� and 5�- GTG TCT GACTCA TGC TTT �3�) or CRE (5�- TTC CAG GGT GAC GTC TTA GGC-3� and 5�-GCC TAA GAC GTC ACC CTG GAA-3�) oligonucleotidesderived from the human MMP-1 promoter in a 15 �l binding reactioncontaining 40 mM NaCl and 2 �g of poly-dI-dC (Pharmacia), and com-plexes were resolved on nondenaturing 4.5% polyacrylamide gels. For su-pershift assays, nuclear extracts were incubated with Abs on ice for 60 min
before the addition of radiolabeled oligonucleotides as previously de-scribed (51). All supershift Abs used were purchased from Santa CruzBiotechnology.
ResultsIFN-� suppresses TLR- and PMA-induced MMP expression inprimary human monocytes
We wished to identify physiological mechanisms that suppressMMP expression in primary human myeloid cells. We first deter-mined which MMPs are induced in monocytes and macrophagesby inflammatory stimuli. Primary monocytes were treated with in-flammatory factors for 3 h or overnight (12 h) and MMP mRNAlevels were measured using real-time PCR. The TLR4 ligand LPSwas the most potent inducer of MMP expression (data not shown),and activated expression of at least seven MMP genes more than5-fold at either an early time point (3 h) (MMP-1, MMP-3, MMP-10, and MMP-12; Fig. 1A), or at a later time point after LPS stim-ulation (12 h) (MMP-2, MMP-7, and MMP-9; Fig. 1B). Expres-sion of many MMPs, with certain exceptions such as MMP-2, isregulated by AP-1 transcription factors and we and others hadpreviously shown that IFN-� can suppress AP-1-mediated geneexpression (52). Therefore, we tested whether IFN-� could sup-press LPS-induced MMP expression. Pretreatment of monocyteswith IFN-� for 3 h strikingly decreased LPS-induced expression ofthe MMPs that were studied (Fig. 1, A and B); similar results wereobtained with primary M-CSF-differentiated macrophages (datanot shown). As IFN-� typically synergizes with LPS to superin-duce expression of inflammatory genes such as COX-2 and IL-6,we measured COX-2 and IL-6 mRNA to rule out nonspecific sup-pressive effects of IFN-� on gene expression. As expected, IFN-�
FIGURE 2. IFN-� inhibition of LPS-induced MMP expression is not dependent on GSK-3. A, IFN-� pretreated primary monocytes were stimulated with LPSfor 3 h in the absence or presence of GSK-3 inhibitors LiCl or SB216763 (added 20 min before IFN-�). B and C, Primary monocytes transfected with controlor GSK-3� siRNAs were stimulated with LPS (B) or PMA (C) with or without IFN-� pretreatment. D, Primary monocytes were transfected with controlor GSK-3� siRNAs, and GSK-3� mRNA and protein levels were measured. mRNA levels were measured using real-time PCR and normalized relativeto �-actin. Results are expressed as mean � SD of triplicate determinants. Results are shown as expression in stimulated monocytes relative to expressionin unstimulated cells (ctrl), which was set at one. GSK-3� protein levels were measured using immunoblotting.
5091The Journal of Immunology
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
increased LPS-induced accumulation of COX-2 and IL-6 mRNA(Fig. 1C). In addition, IFN-� augmented LPS-induced expressionof ADAM-17, a cell surface MMP (Fig. 1C). These results showthat IFN-� specifically and strongly suppressed LPS-induced ex-pression of multiple secreted MMPs that have AP-1 binding se-quences (also known as phorbol ester-responsive elements orTPA-responsive elements) in their promoters and are known to bepositively regulated by AP-1 proteins (3, 53–55). The phorbol es-ter PMA is a strong activator of AP-1-mediated transcription, andIFN-� also suppressed PMA-induced expression of MMPs, but notof TNF-� (Fig. 1D). PMA was a stronger inducer of MMP ex-pression than TLR ligands, which correlated with stronger induc-tion of AP-1 proteins by PMA (data not shown). These resultsshow that IFN-� strongly suppresses MMP expression in primaryhuman monocytes. The dependence of expression of many MMPgenes on AP-1, together with the previously reported inhibition ofAP-1 proteins by IFN-�, suggested that IFN-� may suppress MMPexpression at least in part by inhibiting AP-1. However, IFN-�-mediated inhibition of MMP-2 (Fig. 1B), which is not regulated byAP-1, suggests IFN-� inhibits MMP expression by severalmechanisms.
Previously we (52) and others (56) have shown that IFN-� in-hibits TLR-induced IL-10 production by enhancing glycogen syn-thase kinase 3 (GSK-3) activity and that inhibition of GSK-3 re-verses IFN-� suppression of IL-10 production. This prompted us toinvestigate whether the same mechanism was also applicable toIFN-� regulation of MMPs. However, GSK inhibitors did not haveany effect on IFN-�-mediated suppression of MMP expression(Fig. 2A); reversal of IFN-�-mediated suppression of IL-10 ex-pression confirmed the efficacy of GSK-3 inhibition (data notshown). In addition, we used siRNA to knockdown GSK-3� ex-pression in primary monocytes to �30% of control at both mRNAand protein levels (Fig. 2, B and D), a level sufficiently low toattenuate IFN-�-mediated inhibition of IL-10 expression (Ref. 52and data not shown). However, reduced GSK-3� expression hadno discernable effect on IFN-� suppression of TLR4-inducedMMP expression (Fig. 2B), and similar results were obtained when
PMA was used to induce MMP expression (Fig. 2C). Our resultssuggest that the mechanisms by which IFN-� suppresses LPS in-duction of MMP genes differ from mechanisms used to suppressinduction of the potent anti-inflammatory factor IL-10.
Suppression of MMP expression by IFN-� is dependent onSTAT1 and de novo protein synthesis
Activation of gene expression by IFN-� is mediated by STAT1,but mechanisms by which IFN-� inhibits gene expression are notwell understood (57). Therefore, we investigated whether IFN-�-mediated suppression of MMP expression was dependent on
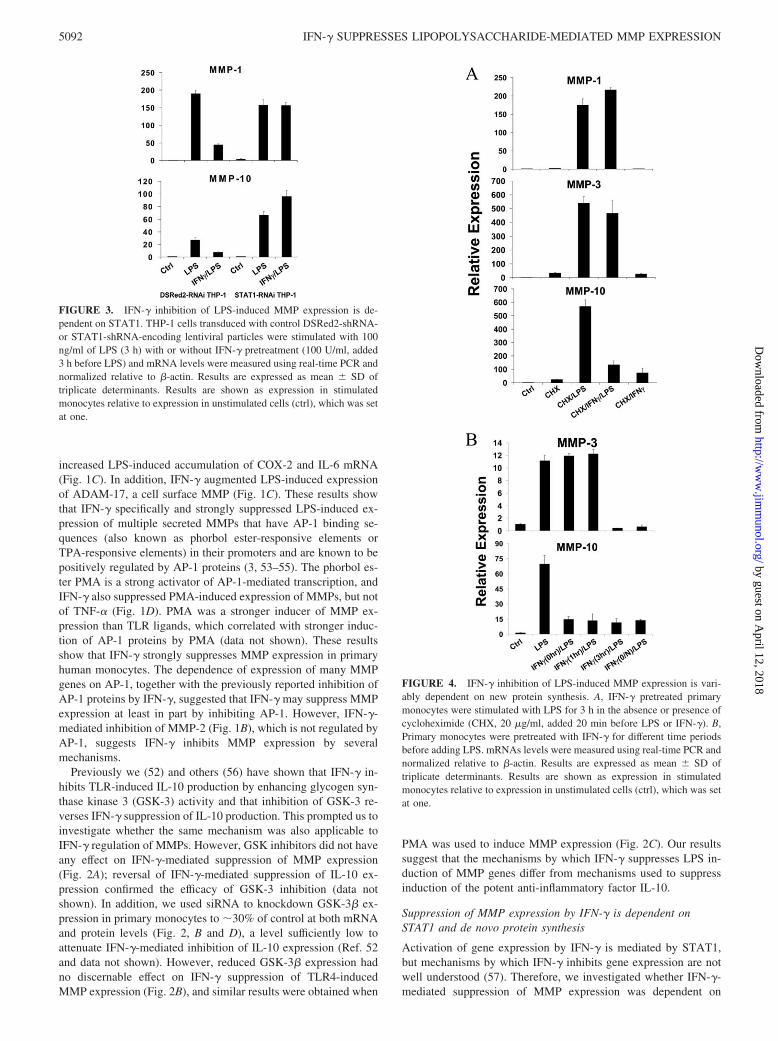
FIGURE 3. IFN-� inhibition of LPS-induced MMP expression is de-pendent on STAT1. THP-1 cells transduced with control DSRed2-shRNA-or STAT1-shRNA-encoding lentiviral particles were stimulated with 100ng/ml of LPS (3 h) with or without IFN-� pretreatment (100 U/ml, added3 h before LPS) and mRNA levels were measured using real-time PCR andnormalized relative to �-actin. Results are expressed as mean � SD oftriplicate determinants. Results are shown as expression in stimulatedmonocytes relative to expression in unstimulated cells (ctrl), which was setat one.
FIGURE 4. IFN-� inhibition of LPS-induced MMP expression is vari-ably dependent on new protein synthesis. A, IFN-� pretreated primarymonocytes were stimulated with LPS for 3 h in the absence or presence ofcycloheximide (CHX, 20 �g/ml, added 20 min before LPS or IFN-�). B,Primary monocytes were pretreated with IFN-� for different time periodsbefore adding LPS. mRNAs levels were measured using real-time PCR andnormalized relative to �-actin. Results are expressed as mean � SD oftriplicate determinants. Results are shown as expression in stimulatedmonocytes relative to expression in unstimulated cells (ctrl), which was setat one.
5092 IFN-� SUPPRESSES LIPOPOLYSACCHARIDE-MEDIATED MMP EXPRESSION
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
STAT1. STAT1 protein expression was too stable to achieve suc-cessful knockdown of expression in primary monocytes, so insteadwe used THP-1 monocytic cells in which STAT1 expression wasstably diminished secondary to transduction with lentiviral vectorsthat express shRNAs targeting STAT1 mRNA (58). IFN-� effec-tively suppressed TLR4-induced MMP-1 and MMP-10 expressionin control THP-1 cells that expressed shRNA that targets the ir-relevant DSRed2 mRNA, but IFN-�-mediated suppression ofMMP induction was completely abrogated in THP-1 cells express-ing low STAT1 levels (Fig. 3). Effects of IFN-� alone on MMPexpression were not consistently observed (data not shown). Thus,suppression of MMP expression by IFN-� is dependent on STAT1.
STAT1 could suppress MMP expression directly by binding toMMP promoters and recruiting transcriptional corepressors, al-though there is minimal precedent for such a direct repressivemechanism. Alternatively, STAT1 could activate expression ofgenes that encode transcriptional repressors that target MMPgenes. To differentiate between these two possibilities, we testedwhether the inhibitory effect of IFN-� on MMP expression wasdependent on new protein synthesis. Primary monocytes werestimulated with LPS and IFN-� in the presence of cycloheximide(added 15 min before IFN-� or LPS) to block protein synthesis. Asshown in Fig. 4A, LPS induced MMP-1, MMP-3 and MMP-10mRNAs in the absence of protein synthesis, indicating that LPSincreases their expression at least in part by inducing posttransla-tional modification of pre-existing proteins. However, cyclohexi-mide abolished the inhibition of MMP-1 and MMP-3 expressionby IFN-�, whereas suppression of MMP-10 expression remainedintact (Fig. 4A). These results suggest that IFN-� induction of arepressor is required for suppression of MMP-1 and MMP-3 ex-
pression, whereas inhibition of MMP-10 expression is more direct.This notion was further supported by a kinetic analysis of IFN-�-mediated inhibition. Down-regulation of LPS-induced MMP-3 ex-pression required greater than 1 h of pretreatment with IFN-�,consistent with the requirement for synthesis of an inhibitor,whereas MMP-10 expression was suppressed even when IFN-�was added simultaneously with LPS (Fig. 4B).
Inhibition of MMP-1 expression is mediated byIFN-�-induced ATF-3
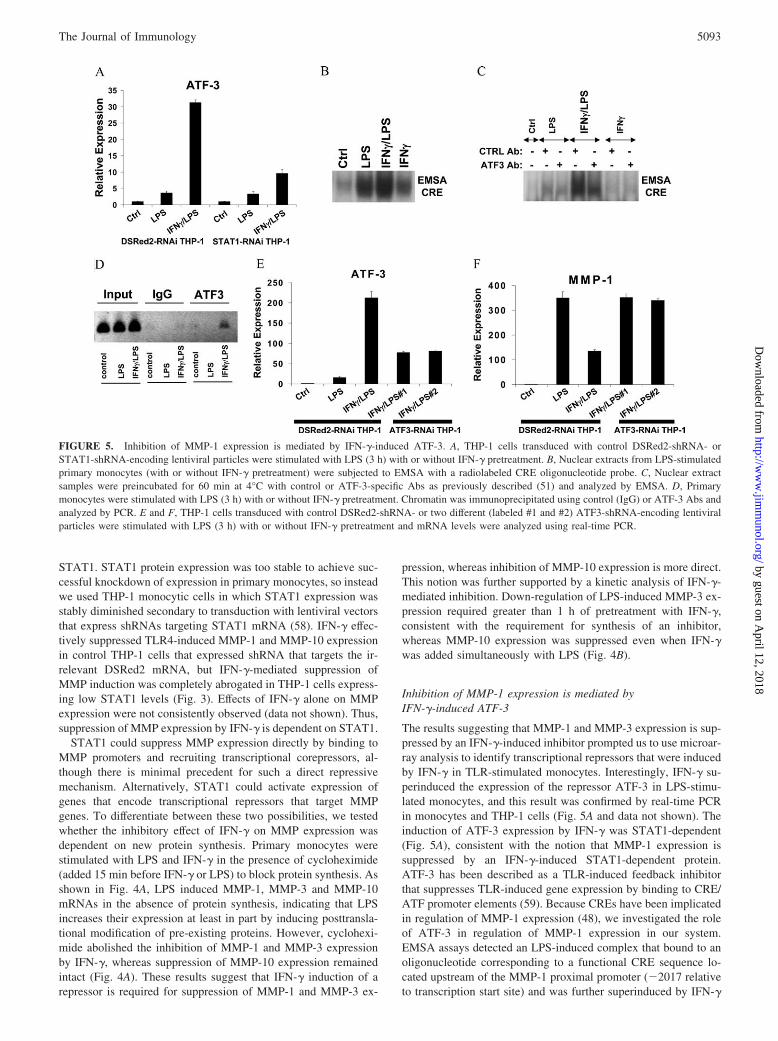
The results suggesting that MMP-1 and MMP-3 expression is sup-pressed by an IFN-�-induced inhibitor prompted us to use microar-ray analysis to identify transcriptional repressors that were inducedby IFN-� in TLR-stimulated monocytes. Interestingly, IFN-� su-perinduced the expression of the repressor ATF-3 in LPS-stimu-lated monocytes, and this result was confirmed by real-time PCRin monocytes and THP-1 cells (Fig. 5A and data not shown). Theinduction of ATF-3 expression by IFN-� was STAT1-dependent(Fig. 5A), consistent with the notion that MMP-1 expression issuppressed by an IFN-�-induced STAT1-dependent protein.ATF-3 has been described as a TLR-induced feedback inhibitorthat suppresses TLR-induced gene expression by binding to CRE/ATF promoter elements (59). Because CREs have been implicatedin regulation of MMP-1 expression (48), we investigated the roleof ATF-3 in regulation of MMP-1 expression in our system.EMSA assays detected an LPS-induced complex that bound to anoligonucleotide corresponding to a functional CRE sequence lo-cated upstream of the MMP-1 proximal promoter (�2017 relativeto transcription start site) and was further superinduced by IFN-�
FIGURE 5. Inhibition of MMP-1 expression is mediated by IFN-�-induced ATF-3. A, THP-1 cells transduced with control DSRed2-shRNA- orSTAT1-shRNA-encoding lentiviral particles were stimulated with LPS (3 h) with or without IFN-� pretreatment. B, Nuclear extracts from LPS-stimulatedprimary monocytes (with or without IFN-� pretreatment) were subjected to EMSA with a radiolabeled CRE oligonucleotide probe. C, Nuclear extractsamples were preincubated for 60 min at 4°C with control or ATF-3-specific Abs as previously described (51) and analyzed by EMSA. D, Primarymonocytes were stimulated with LPS (3 h) with or without IFN-� pretreatment. Chromatin was immunoprecipitated using control (IgG) or ATF-3 Abs andanalyzed by PCR. E and F, THP-1 cells transduced with control DSRed2-shRNA- or two different (labeled #1 and #2) ATF3-shRNA-encoding lentiviralparticles were stimulated with LPS (3 h) with or without IFN-� pretreatment and mRNA levels were analyzed using real-time PCR.
5093The Journal of Immunology
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

(Fig. 5, B and C). DNA binding by this IFN-�-superinduced com-plex was disrupted by ATF-3 Abs but not by control Abs (Fig. 5C).Furthermore, treatment of monocytes with LPS plus IFN-� in-duced recruitment of ATF-3 to the endogenous MMP-1 promoterin the vicinity of the �2017 ATF site (Fig. 5D). Taken togetherwith the real-time PCR results (Fig. 5A), these results suggest thatIFN-� induces DNA binding of the ATF-3 transcriptional repres-sor that can potentially suppress MMP-1 expression. The role ofATF-3 in regulating MMP-1 expression was investigated by usinglentivirus-mediated transduction of THP-1 monocytes to createtwo stable cell lines that express different shRNAs targetingATF-3. In those ATF3-RNAi THP-1 cells, the synergistic LPS/IFN-� induction of ATF-3 was reduced to �40% that of control(Fig. 5E), and decreased ATF-3 expression resulted in reversal ofIFN-�-mediated suppression of LPS-induced MMP-1 gene expres-sion (Fig. 5F). Attempts to test the effects of ATF-3 overexpres-sion were not successful as increased expression of ATF-3 inducedcell death (data not shown). These results implicate IFN-�-induced
ATF-3 in mediating the suppressive effects of IFN-� on MMPexpression.
IFN-� regulates AP-1 and Fra-1 DNA binding
The AP-1 element located between base pairs �72 and �66 of theMMP-1 promoter is crucial for MMP-1 gene expression (41, 55,60–62) and we and others have shown that IFN-� suppresses AP-1-dependent gene expression. Thus, despite the negative data onthe role of GSK-3 (Fig. 2), we further investigated the effects ofIFN-� on LPS-induced AP-1 proteins. EMSA experiments usingthe MMP-1 promoter AP-1 site showed an LPS-induced DNA-binding complex (Fig. 6A). Interestingly, LPS-induced binding tothe AP-1 oligonucleotide was suppressed by IFN-� (Fig. 6A), sug-gesting suppression of AP-1 proteins as a complementary mech-anism by which IFN-� suppresses MMP-1 expression. Supershiftexperiments showed that the LPS-induced complex was reactivewith Fra-1 Abs, but not with Fos or Jun Abs (Fig. 6B). Because theMMP-1 promoter-derived AP-1 oligonucleotide contains an over-lapping Ets site, we also investigated whether the LPS-inducedcomplexes contained Ets proteins, but supershift experiments withPEA-3, PU.1, ETS-1, ETS-2, and ERG 1/2/3 showed no reactivity(Fig. 6B and data not shown). These results suggested a role forFra-1 in LPS and IFN-� regulation of MMP-1 expression, and thiswas further investigated using RNA interference and overexpres-sion of Fra-1. Indeed, knockdown of Fra-1 expression resulted indiminished LPS-induced MMP-1 expression (Fig. 6C), and con-versely overexpression of Fra-1 increased MMP-1 expression (Fig.6D), thus implicating Fra-1 in induction of MMP-1 expression by
FIGURE 6. IFN-� inhibits LPS-induced AP-1 DNA-binding complex.A, Nuclear extracts from LPS stimulated primary monocytes (with or with-out IFN-� pretreatment) were subjected to EMSA with a radiolabeledAP-1 oligonucleotide probe derived from the MMP-1 promoter. B, Nu-clear extracts were preincubated for 60 min at 4°C with control or Absagainst various ETS and AP-1 proteins before EMSA. C, Primarymonocytes transfected with control or Fra-1 siRNAs were stimulatedwith LPS. D, THP-1 cells transduced with eGFP- or Fra-1-encodinglentiviral particles were stimulated with LPS and RNA levels were mea-sured using real-time PCR.
FIGURE 7. PGE2 inhibits LPS-induced MMP-1 expression and sup-presses Fra-1 expression and DNA binding. A and B, Primary monocyteswere stimulated with LPS in the absence or presence of PGE2 (1 �M added20 min before LPS) and RNA was measured using real-time PCR. C,Nuclear extracts from primary monocytes stimulated with LPS in the ab-sence or presence of PGE2 were incubated with control or Fra-1 Abs andsubjected to EMSA with a radiolabeled AP-1 oligonucleotide probe.
5094 IFN-� SUPPRESSES LIPOPOLYSACCHARIDE-MEDIATED MMP EXPRESSION
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

LPS. We did not detect regulation of Fra-1 expression by IFN-�(data not shown), but, interestingly, PGE2 which suppressedMMP-1 expression in monocytes (Fig. 7A) also suppressed Fra-1expression (Fig. 7B). EMSA using the MMP-1 promoter AP-1 sitedemonstrated that in the presence of PGE2, the composition ofthe LPS-induced DNA binding complex was altered such that thecomplex was no longer shifted by Fra-1 Abs (Fig. 7C). Theseresults support the notion that PGE2 inhibits LPS-induced MMP-1gene expression by suppressing expression of Fra-1 that contrib-utes to induction of MMP-1 expression through the proximal pro-moter AP-1 site. Overall, our findings support a role for Fra-1 inthe positive and negative regulation of MMP-1 expression, andsuggest that IFN-� inhibits MMP-1 transcription in part by sup-pressing DNA binding by Fra-1. Although IFN-� also suppressesother AP-1 proteins such as Fos and Jun (52), we were not able tolink regulation of Fos and Jun with regulation of MMP-1 expres-sion in our system.
DiscussionMMP expression needs to be tightly regulated during inflammationto allow necessary angiogenesis and tissue remodeling but to avoidexcessive tissue damage. In this study, we found that IFN-�, apotent macrophage activating cytokine, globally inhibits MMP ex-pression at the same time that it synergistically enhances the pro-duction of proinflammatory cytokines. Thus, as the intensity ofinflammation increases in response to IFN-� there is a built-inmechanism to restrain associated tissue damage. We have alsoidentified two mechanisms by which IFN-� inhibits MMP expres-sion, by inducing expression of the transcriptional repressor ATF-3and simultaneously suppressing the function of AP-1 transcrip-tional activators.
The regulation of MMP gene expression has been extensivelystudied, although much of this work has utilized reporter geneapproaches in cell lines and nonphysiological activators such asPMA (7, 28–45). This previous work has revealed the importanceof proximal promoter sequences, especially a composite AP-1/Etssite, in basal and inducible MMP-1 and MMP-3 expression. Morerecently, the importance of promoter and enhancer sequences out-side MMP proximal promoters is becoming increasingly apparent,including upstream CRE sites (48, 63). Surprisingly little is knownabout MMP regulation in primary cells, especially hematopoieticcells such as monocytes and macrophages. In addition, althoughimportant MMP promoter elements have been extensively ana-lyzed, less is known about transcription factors that regulate MMPexpression. Our work implicates ATF-3 in the regulation ofMMP-1 expression. Because ATF-3 is induced by TLR stimula-tion alone, it participates in feedback inhibition of TLR-inducedgene expression, as has been previously reported in other systems(59). In addition, superinduction of ATF-3 by IFN-� contributes toIFN-�-mediated suppression of MMP-1 gene expression. Trans-fection of primary monocytes or THP-1 cells with MMP promoter-driven reporter plasmids resulted in cell activation, presumablysecondary to an innate immune response to exogenous DNA, andrapid extinction of reporter gene expression (H. Ho, unpublisheddata). Thus, the reporter gene approach was not informative instudying LPS and IFN-� regulation of MMP expression.
A consistent finding in multiple systems and cell types has beenthe important role of the proximal promoter AP-1 site in the ex-pression of MMP-1 and MMP-3 (3, 41, 55, 60–62). Therefore,given our previous findings that IFN-� can suppress AP-1-medi-ated IL-10 transcription by a GSK-3-dependent mechanism, weinvestigated a potential role for regulation of AP-1 proteins byIFN-� in suppression of MMP expression. LPS induced a Fra-1-containing complex that bound to the MMP-1 promoter AP-1 site,
and the binding of this complex was suppressed by IFN-�. Fra-1has been previously implicated in promoting MMP expression (64,65), although its regulation by LPS and IFN-� has not been pre-viously appreciated. We confirmed the importance of Fra-1 inMMP-1 gene expression using both knockdown and overexpres-sion approaches and implicated Fra-1 in mediating PGE2 regula-tion of MMP-1 expression. In contrast to PGE2, IFN-� did notinhibit Fra-1 expression, suggesting that IFN-� may suppress DNAbinding of Fra-1 by posttranslational modifications or indirectly byregulating other members of the AP-1 family. Overall, the datashow that IFN-� regulated MMP-1 expression by at least twomechanisms, induction of ATF-3 and suppression of DNA bindingby a Fra-1-containing AP-1 complex. Of the MMPs studied whoseexpression was inhibited by IFN-� (Fig. 1), all contained AP-1sites with the exception of MMP-2 (66). Thus, although suppres-sion of AP-1 may contribute to diminished expression of severalMMPs, it cannot explain inhibition of MMP-2 by IFN-�. Interest-ingly, ATF-3 has been implicated in negative regulation of MMP-2(59), suggesting a role for ATF-3 in regulation of MMP gene ex-pression beyond the regulation of MMP-1 expression that was in-vestigated in this study. Collectively, our data and previous reportssuggest that IFN-� can suppress MMP expression by differentmechanisms. This notion is further supported by our results sug-gesting that MMP-10 expression was suppressed by IFN-� by amechanism that differed from suppression of MMP-1 because in-hibition of MMP-10 did not require new protein synthesis, andIFN-� inhibits MMP-9 expression by modifying chromatin andtranscriptional coactivator recruitment (28, 30, 35, 67, 68). Thesefindings highlight that IFN-� suppresses MMP expression by sev-eral mechanisms and provide insights into how IFN-� down-reg-ulates gene expression.
Prior studies from several laboratories including ours have de-scribed antagonistic functions for STAT1 and STAT3 in the reg-ulation of cell growth/survival and inflammatory gene expression(58, 69). It appears that this opposition of STAT1 and STAT3 alsoextends to the regulation of MMP expression. STAT3 has beenshown to be an important and strong inducer of MMPs such asMMP-1 and MMP-10 (70, 71), and was found to bind directly toa STAT-binding element located between base pairs �53 and �45upstream in the MMP-1 proximal promoter after stimulation withEGF (71). We have shown that inhibition of MMP-1 expression byIFN-� is dependent on STAT1, but likely occurs indirectly viaregulation of ATF-3 and AP-1 proteins. Consistent with these re-sults, we were not able to detect binding of STAT1 to the MMP-1promoter AP-1 site or the previously described STAT-biding ele-ment that binds STAT3 (H. Ho, unpublished observations), sug-gesting that STAT1 does not directly compete with STAT3 forbinding to the MMP-1 promoter. On the other hand, it is possiblethat STAT-1 inhibits MMP-10 expression by a direct mechanism,as has been shown for IFN-� suppression of the Perlecan gene(72), because inhibition of MMP-10 expression by IFN-� wasSTAT1-dependent and rapid and independent of new proteinsynthesis.
In summary, our results demonstrate that IFN-� differentiallyregulates gene expression in primary human monocytes and mac-rophages by augmenting the production of inflammatory cytokinessuch as IL-6 and TNF-�, but at the same time globally suppressingMMP production. The suppression of MMPs by IFN-� may rep-resent a self-protective mechanism to limit undesirable damage totissues during intense immune and inflammatory reactions that arerequired to eradicate infectious pathogens. The results also revealthat IFN-� modulates MMP expression by regulating the expres-sion and function of ATF-3, Fra-1 and potentially other AP-1 pro-teins. Understanding the mechanisms by which IFN-� regulates
5095The Journal of Immunology
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

these genes and transcription factors may yield useful insights intoformulating future approaches to enhancing immune clearance ofinvading pathogens and yet limiting damage to tissues and organsystems.
AcknowledgmentsWe thank Janice Chen for helpful discussions, Kyung-Kyun Park-Min forreview of the manuscript, and Inez Rogatsky and Barbara Nikolajczyk foradvice concerning ChIP assays.
DisclosuresThe authors have no financial conflict of interest.
References1. Gross, J., and C. M. Lapiere. 1962. Collagenolytic activity in amphibian tissues:
a tissue culture assay. Proc. Natl. Acad. Sci. USA 48: 1014–1022.2. Woessner, J. F., Jr. 1991. Matrix metalloproteinases and their inhibitors in con-
nective tissue remodeling. FASEB J. 5: 2145–2154.3. Chakraborti, S., M. Mandal, S. Das, A. Mandal, and T. Chakraborti. 2003. Reg-
ulation of matrix metalloproteinases: an overview. Mol. Cell. Biochem. 253:269–285.
4. Sounni, N. E., and A. Noel. 2005. Membrane type-matrix metalloproteinases andtumor progression. Biochimie 87: 329–342.
5. Lemaitre, V., and J. D’Armiento. 2006. Matrix metalloproteinases in develop-ment and disease. Birth Defects Res. C Embryo Today 78: 1–10.
6. Vincenti, M. P., and C. E. Brinckerhoff. 2002. Transcriptional regulation of col-lagenase (MMP-1, MMP-13) genes in arthritis: integration of complex signalingpathways for the recruitment of gene-specific transcription factors. Arthritis Res.4: 157–164.
7. Huntington, J. T., J. M. Shields, C. J. Der, C. A. Wyatt, U. Benbow,C. L. Slingluff, Jr., and C. E. Brinckerhoff. 2004. Overexpression of collagenase1 (MMP-1) is mediated by the ERK pathway in invasive melanoma cells: role ofBRAF mutation and fibroblast growth factor signaling. J. Biol. Chem. 279:33168–33176.
8. McQuibban, G. A., J. H. Gong, E. M. Tam, C. A. McCulloch, I. Clark-Lewis, andC. M. Overall. 2000. Inflammation dampened by gelatinase A cleavage of mono-cyte chemoattractant protein-3. Science 289: 1202–1206.
9. Wilson, C. L., A. J. Ouellette, D. P. Satchell, T. Ayabe, Y. S. Lopez-Boado,J. L. Stratman, S. J. Hultgren, L. M. Matrisian, and W. C. Parks. 1999. Regulationof intestinal �-defensin activation by the metalloproteinase matrilysin in innatehost defense. Science 286: 113–117.
10. Powell, W. C., B. Fingleton, C. L. Wilson, M. Boothby, and L. M. Matrisian.1999. The metalloproteinase matrilysin proteolytically generates active solubleFas ligand and potentiates epithelial cell apoptosis. Curr. Biol. 9: 1441–1447.
11. Hartzell, W., and S. D. Shapiro. 1999. Macrophage elastase prevents Gemellamorbillorum infection and improves outcome following murine bone marrowtransplantation. Chest 116: 31S–32S.
12. Lopez-Boado, Y. S., C. L. Wilson, L. V. Hooper, J. I. Gordon, S. J. Hultgren, andW. C. Parks. 2000. Bacterial exposure induces and activates matrilysin in mu-cosal epithelial cells. J. Cell Biol. 148: 1305–1315.
13. Corry, D. B., K. Rishi, J. Kanellis, A. Kiss, L. Z. Song Lz, J. Xu, L. Feng,Z. Werb, and F. Kheradmand. 2002. Decreased allergic lung inflammatory cellegression and increased susceptibility to asphyxiation in MMP2-deficiency. Nat.Immunol. 3: 347–353.
14. Li, Q., P. W. Park, C. L. Wilson, and W. C. Parks. 2002. Matrilysin shedding ofsyndecan-1 regulates chemokine mobilization and transepithelial efflux of neu-trophils in acute lung injury. Cell 111: 635–646.
15. Haro, H., H. C. Crawford, B. Fingleton, J. R. MacDougall, K. Shinomiya,D. M. Spengler, and L. M. Matrisian. 2000. Matrix metalloproteinase-3-depen-dent generation of a macrophage chemoattractant in a model of herniated discresorption. J. Clin. Invest. 105: 133–141.
16. Liu, Z., X. Zhou, S. D. Shapiro, J. M. Shipley, S. S. Twining, L. A. Diaz,R. M. Senior, and Z. Werb. 2000. The serpin alpha1-proteinase inhibitor is acritical substrate for gelatinase B/MMP-9 in vivo. Cell 102: 647–655.
17. Haro, H., H. C. Crawford, B. Fingleton, K. Shinomiya, D. M. Spengler, andL. M. Matrisian. 2000. Matrix metalloproteinase-7-dependent release of tumornecrosis factor-� in a model of herniated disc resorption. J. Clin. Invest. 105:143–150.
18. Parks, W. C., C. L. Wilson, and Y. S. Lopez-Boado. 2004. Matrix metallopro-teinases as modulators of inflammation and innate immunity. Nat. Rev. Immunol.4: 617–629.
19. Egeblad, M., and Z. Werb. 2002. New functions for the matrix metalloproteinasesin cancer progression. Nat. Rev. Cancer 2: 161–174.
20. Schroen, D. J., and C. E. Brinckerhoff. 1996. Nuclear hormone receptors inhibitmatrix metalloproteinase (MMP) gene expression through diverse mechanisms.Gene Expression 6: 197–207.
21. Szekanecz, Z., and A. E. Koch. 2007. Macrophages and their products in rheu-matoid arthritis. Curr. Opin. Rheumatol. 19: 289–295.
22. Stoll, G., and M. Bendszus. 2006. Inflammation and atherosclerosis: novel in-sights into plaque formation and destabilization. Stroke 37: 1923–1932.
23. Takahashi, K., M. Takeya, and N. Sakashita. 2002. Multifunctional roles of mac-rophages in the development and progression of atherosclerosis in humans andexperimental animals. Med. Electron Microsc. 35: 179–203.
24. Boyle, J. J. 2005. Macrophage activation in atherosclerosis: pathogenesis andpharmacology of plaque rupture. Curr. Vasc. Pharmacol. 3: 63–68.
25. Libby, P. 2000. Changing concepts of atherogenesis. J. Intern. Med. 247:349–358.
26. Shah, P. K., E. Falk, J. J. Badimon, A. Fernandez-Ortiz, A. Mailhac,G. Villareal-Levy, J. T. Fallon, J. Regnstrom, and V. Fuster. 1995. Human mono-cyte-derived macrophages induce collagen breakdown in fibrous caps of athero-sclerotic plaques: potential role of matrix-degrading metalloproteinases and im-plications for plaque rupture. Circulation 92: 1565–1569.
27. Sukhova, G. K., U. Schonbeck, E. Rabkin, F. J. Schoen, A. R. Poole,R. C. Billinghurst, and P. Libby. 1999. Evidence for increased collagenolysis byinterstitial collagenases-1 and -3 in vulnerable human atheromatous plaques. Cir-culation 99: 2503–2509.
28. Xu, Y., L. Wang, G. Buttice, P. K. Sengupta, and B. D. Smith. 2004. Majorhistocompatibility class II transactivator (CIITA) mediates repression of collagen(COL1A2) transcription by interferon � (IFN-�). J. Biol. Chem. 279:41319–41332.
29. Lewis, M., E. P. Amento, and E. N. Unemori. 1999. Transcriptional inhibition ofstromelysin by interferon-� in normal human fibroblasts is mediated by the AP-1domain. J. Cell. Biochem. 72: 373–386.
30. Ma, Z., M. J. Chang, R. C. Shah, and E. N. Benveniste. 2005. Interferon-�-activated STAT-1� suppresses MMP-9 gene transcription by sequestration of thecoactivators CBP/p300. J. Leukocyte Biol. 78: 515–523.
31. Yokota, H., M. B. Goldring, and H. B. Sun. 2003. CITED2-mediated regulationof MMP-1 and MMP-13 in human chondrocytes under flow shear. J. Biol. Chem.278: 47275–47280.
32. Andrews, H. J., R. A. Bunning, C. A. Dinarello, and R. G. Russell. 1989. Mod-ulation of human chondrocyte metabolism by recombinant human interferon �:in-vitro effects on basal and IL-1-stimulated proteinase production, cartilage deg-radation and DNA synthesis. Biochim. Biophys. Acta 1012: 128–134.
33. Unemori, E. N., M. J. Bair, E. A. Bauer, and E. P. Amento. 1991. Stromelysinexpression regulates collagenase activation in human fibroblasts: dissociable con-trol of two metalloproteinases by interferon-�. J. Biol. Chem. 266: 23477–23482.
34. Varga, J., T. Yufit, and R. R. Brown. 1995. Inhibition of collagenase and strome-lysin gene expression by interferon-� in human dermal fibroblasts is mediated inpart via induction of tryptophan degradation. J. Clin. Invest. 96: 475–481.
35. Nozell, S., Z. Ma, C. Wilson, R. Shah, and E. N. Benveniste. 2004. Class II majorhistocompatibility complex transactivator (CIITA) inhibits matrix metallopro-teinase-9 gene expression. J. Biol. Chem. 279: 38577–38589.
36. Fenrick, R., L. Wang, J. Nip, J. M. Amann, R. J. Rooney, J. Walker-Daniels,H. C. Crawford, D. L. Hulboy, M. S. Kinch, L. M. Matrisian, and S. W. Hiebert.2000. TEL, a putative tumor suppressor, modulates cell growth and cell mor-phology of ras-transformed cells while repressing the transcription of stromely-sin-1. Mol. Cell. Biol. 20: 5828–5839.
37. Barchowsky, A., D. Frleta, and M. P. Vincenti. 2000. Integration of the NF-�Band mitogen-activated protein kinase/AP-1 pathways at the collagenase-1 pro-moter: divergence of IL-1 and TNF-dependent signal transduction in rabbit pri-mary synovial fibroblasts. Cytokine 12: 1469–1479.
38. Quinones, S., J. Saus, Y. Otani, E. D. Harris, Jr., and M. Kurkinen. 1989. Tran-scriptional regulation of human stromelysin. J. Biol. Chem. 264: 8339–8344.
39. Buttice, G., and M. Kurkinen. 1993. A polyomavirus enhancer A-binding pro-tein-3 site and Ets-2 protein have a major role in the 12-O-tetradecanoylphorbol-13-acetate response of the human stromelysin gene. J. Biol. Chem. 268:7196–7204.
40. Buttice, G., S. Quinones, and M. Kurkinen. 1991. The AP-1 site is required forbasal expression but is not necessary for TPA-response of the human stromelysingene. Nucleic Acids Res. 19: 3723–3731.
41. Bidder, M., A. P. Loewy, T. Latifi, E. P. Newberry, G. Ferguson, D. M. Willis,and D. A. Towler. 2000. Ets domain transcription factor PE1 suppresses humaninterstitial collagenase promoter activity by antagonizing protein-DNA interac-tions at a critical AP1 element. Biochemistry 39: 8917–8928.
42. Jinnin, M., H. Ihn, Y. Mimura, Y. Asano, K. Yamane, and K. Tamaki. 2005.Matrix metalloproteinase-1 up-regulation by hepatocyte growth factor in humandermal fibroblasts via ERK signaling pathway involves Ets1 and Fli1. NucleicAcids Res. 33: 3540–3549.
43. Jayaraman, G., R. Srinivas, C. Duggan, E. Ferreira, S. Swaminathan,K. Somasundaram, J. Williams, C. Hauser, M. Kurkinen, R. Dhar, et al. 1999.p300/cAMP-responsive element-binding protein interactions with ets-1 and ets-2in the transcriptional activation of the human stromelysin promoter. J. Biol.Chem. 274: 17342–17352.
44. Nakajima, H., Y. Hiyama, W. Tsukada, H. Warabi, S. Uchida, and S. Hirose.1990. Effects of interferon � on cultured synovial cells from patients with rheu-matoid arthritis: inhibition of cell growth, prostaglandin E2, and collagenase re-lease. Ann. Rheum. Dis. 49: 512–516.
45. Schroen, D. J., J. D. Chen, M. P. Vincenti, and C. E. Brinckerhoff. 1997. Thenuclear receptor corepressor SMRT inhibits interstitial collagenase (MMP-1)transcription through an HRE-independent mechanism. Biochem. Biophys. Res.Commun. 237: 52–58.
46. Khan, K. M., L. R. Howe, and D. J. Falcone. 2004. Extracellular matrix-inducedcyclooxygenase-2 regulates macrophage proteinase expression. J. Biol. Chem.279: 22039–22046.
47. Zhang, Y., K. McCluskey, K. Fujii, and L. M. Wahl. 1998. Differential regulationof monocyte matrix metalloproteinase and TIMP-1 production by TNF-�, gran-ulocyte-macrophage CSF, and IL-1 � through prostaglandin-dependent and -in-dependent mechanisms. J. Immunol. 161: 3071–3076.
5096 IFN-� SUPPRESSES LIPOPOLYSACCHARIDE-MEDIATED MMP EXPRESSION
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

48. Lai, W. C., M. Zhou, U. Shankavaram, G. Peng, and L. M. Wahl. 2003. Differ-ential regulation of lipopolysaccharide-induced monocyte matrix metalloprotein-ase (MMP)-1 and MMP-9 by p38 and extracellular signal-regulated kinase 1/2mitogen-activated protein kinases. J. Immunol. 170: 6244–6249.
49. Rubinson, D. A., C. P. Dillon, A. V. Kwiatkowski, C. Sievers, L. Yang,J. Kopinja, M. Zhang, M. T. McManus, F. B. Gertler, M. L. Scott, andL. Van Parijs. 2003. A lentivirus-based system to functionally silence genes inprimary mammalian cells, stem cells and transgenic mice by RNA interference.Nat. Genet. 33: 401–406.
50. Tassiulas, I., X. Hu, H. Ho, Y. Kashyap, P. Paik, Y. Hu, C. A. Lowell, andL. B. Ivashkiv. 2004. Amplification of IFN-�-induced STAT1 activation andinflammatory function by Syk and ITAM-containing adaptors. Nat. Immunol. 5:1181–1189.
51. Sengupta, T. K., A. Chen, Z. Zhong, J. E. Darnell, Jr., and L. B. Ivashkiv. 1995.Activation of monocyte effector genes and STAT family transcription factors byinflammatory synovial fluid is independent of interferon �. J. Exp. Med. 181:1015–1025.
52. Hu, X., P. K. Paik, J. Chen, A. Yarilina, L. Kockeritz, T. T. Lu, J. R. Woodgett,and L. B. Ivashkiv. 2006. IFN-� suppresses IL-10 production and synergizes withTLR2 by regulating GSK3 and CREB/AP-1 proteins. Immunity 24: 563–574.
53. Angel, P., I. Baumann, B. Stein, H. Delius, H. J. Rahmsdorf, and P. Herrlich.1987. 12-O-tetradecanoyl-phorbol-13-acetate induction of the human collagenasegene is mediated by an inducible enhancer element located in the 5�-flankingregion. Mol. Cell. Biol. 7: 2256–2266.
54. Westermarck, J., and V. M. Kahari. 1999. Regulation of matrix metalloproteinaseexpression in tumor invasion. FASEB J. 13: 781–792.
55. Benbow, U., and C. E. Brinckerhoff. 1997. The AP-1 site and MMP gene regu-lation: what is all the fuss about? Matrix Biol. 15: 519–526.
56. Martin, M., K. Rehani, R. S. Jope, and S. M. Michalek. 2005. Toll-like receptor-mediated cytokine production is differentially regulated by glycogen synthasekinase 3. Nat. Immunol. 6: 777–784.
57. Ramana, C. V., M. P. Gil, R. D. Schreiber, and G. R. Stark. 2002. Stat1-depen-dent and -independent pathways in IFN-�-dependent signaling. Trends Immunol.23: 96–101.
58. Ho, H. H., and L. B. Ivashkiv. 2006. Role of STAT3 in type I interferon re-sponses: negative regulation of STAT1-dependent inflammatory gene activation.J. Biol. Chem. 281: 14111–14118.
59. Gilchrist, M., V. Thorsson, B. Li, A. G. Rust, M. Korb, K. Kennedy, T. Hai,H. Bolouri, and A. Aderem. 2006. Systems biology approaches identify ATF3 asa negative regulator of Toll-like receptor 4. Nature 441: 173–178.
60. Westermarck, J., A. Seth, and V. M. Kahari. 1997. Differential regulation ofinterstitial collagenase (MMP-1) gene expression by ETS transcription factors.Oncogene 14: 2651–2660.
61. Oikawa, T. 2004. ETS transcription factors: possible targets for cancer therapy.Cancer Sci. 95: 626–633.
62. Schroen, D. J., and C. E. Brinckerhoff. 1996. Inhibition of rabbit collagenase(matrix metalloproteinase-1; MMP-1) transcription by retinoid receptors: evi-dence for binding of RARs/RXRs to the -77 AP-1 site through interactions withc-Jun. J. Cell. Physiol. 169: 320–332.
63. Benbow, U., J. L. Rutter, C. H. Lowrey, and C. E. Brinckerhoff. 1999. Tran-scriptional repression of the human collagenase-1 (MMP-1) gene in MDA231breast cancer cells by all-trans-retinoic acid requires distal regions of the pro-moter. Br. J. Cancer 79: 221–228.
64. Tower, G. B., C. I. Coon, K. Belguise, D. Chalbos, and C. E. Brinckerhoff. 2003.Fra-1 targets the AP-1 site/2G single nucleotide polymorphism (ETS site) in theMMP-1 promoter. Eur. J. Biochem. 270: 4216–4225.
65. Newberry, E. P., D. Willis, T. Latifi, J. M. Boudreaux, and D. A. Towler. 1997.Fibroblast growth factor receptor signaling activates the human interstitial col-lagenase promoter via the bipartite Ets-AP1 element. Mol. Endocrinol. 11:1129–1144.
66. Yan, C., and D. D. Boyd. 2007. Regulation of matrix metalloproteinase geneexpression. J. Cell. Physiol. 211: 19–26.
67. Shapiro, S. D., E. J. Campbell, D. K. Kobayashi, and H. G. Welgus. 1990. Im-mune modulation of metalloproteinase production in human macrophages: selec-tive pretranslational suppression of interstitial collagenase and stromelysin bio-synthesis by interferon-�. J. Clin. Invest. 86: 1204–1210.
68. Nguyen, J., P. Knapnougel, P. Lesavre, and B. Bauvois. 2005. Inhibition of ma-trix metalloproteinase-9 by interferons and TGF-�1 through distinct signalingsaccounts for reduced monocyte invasiveness. FEBS Lett. 579: 5487–5493.
69. Hu, X., K. H. Park-Min, H. H. Ho, and L. B. Ivashkiv. 2005. IFN-�-primedmacrophages exhibit increased CCR2-dependent migration and altered IFN-�responses mediated by Stat1. J. Immunol. 175: 3637–3647.
70. Tsareva, S. A., R. Moriggl, F. M. Corvinus, B. Wiederanders, A. Schutz,B. Kovacic, and K. Friedrich. 2007. Signal transducer and activator of transcrip-tion 3 activation promotes invasive growth of colon carcinomas through matrixmetalloproteinase induction. Neoplasia 9: 279–291.
71. Itoh, M., T. Murata, T. Suzuki, M. Shindoh, K. Nakajima, K. Imai, andK. Yoshida. 2006. Requirement of STAT3 activation for maximal collagenase-1(MMP-1) induction by epidermal growth factor and malignant characteristics inT24 bladder cancer cells. Oncogene 25: 1195–1204.
72. Sharma, B., and R. V. Iozzo. 1998. Transcriptional silencing of perlecan geneexpression by interferon-�. J. Biol. Chem. 273: 4642–4646.
5097The Journal of Immunology
by guest on April 12, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from


















