
Amino kiseline i proteini
68
1 2.3. PROTEINI 2D-PAGE proteinâ humanog hepatoblastoma katalaza hemoglobin, β-podjedinica Hsp60 serumalbumin β-aktin ATP-sintaza, δ-podjedinica trioza-fosfat izomeraza Pod proteinima se podrazumevaju peptidi koji sadrže veliki broj aminokiselinskih ostataka. Različiti izvori navode različite granične dužine lanca – 20-30 AK, 50 AK ili 100 AK. U stvarnosti, ne postoji oštra granica između polipeptida i proteina. Razlika je u stvari vezana za 3D strukturu molekula – dok polipeptidi nemaju stabilnu 3D strukturu, proteini nakon sinteze zauzimaju jedinstvenu, dobro definisanu nativnu konformaciju. Poznat je veliki broj proteina – u PDB (Protein Data Bank) je u februaru 2013. bilo evidentirano preko 88000 struktura; sisari sadrže oko 20 000 različitih proteina, a E. coli oko 4000. Ova jedinjenja značajno se razlikuju po broju AK ostataka odn. molekulskoj masi – dok neki imaju mase od svega nekoliko kDa, drugi mogu preći 200 kDa, a najveći poznati oligomerni protein – titin (konektin) iz mišića – sadrži oko 30 000 AK i ima masu skoro 4 MDa. [slika: sa Swiss-2D PAGE sajta, world-2dpage.expasy.org/swiss-2dpage/]
description
Formiranje proteina od amino kiselina, klasifikacija proteina, funkcija proteina, DNK
Transcript of Amino kiseline i proteini

1
2.3. PROTEINI2D-PAGE proteinâ humanog hepatoblastoma
katalaza
hemoglobin, β-podjedinica
Hsp60serumalbumin
β-aktin
ATP-sintaza,δ-podjedinica
trioza-fosfatizomeraza
Pod proteinima se podrazumevaju peptidi koji sadrže veliki broj aminokiselinskih ostataka. Različiti izvori navode različite granične dužine lanca – 20-30 AK, 50 AK ili 100 AK. U stvarnosti, ne postoji oštra granica između polipeptida i proteina. Razlika je u stvari vezana za 3D strukturu molekula – dok polipeptidi nemaju stabilnu 3D strukturu, proteini nakon sinteze zauzimaju jedinstvenu, dobro definisanu nativnu konformaciju. Poznat je veliki broj proteina – u PDB (Protein Data Bank) je u februaru 2013. bilo evidentirano preko 88000 struktura; sisari sadrže oko 20 000 različitih proteina, a E. coli oko 4000. Ova jedinjenja značajno se razlikuju po broju AK ostataka odn. molekulskoj masi – dok neki imaju mase od svega nekoliko kDa, drugi mogu preći 200 kDa, a najveći poznati oligomerni protein – titin (konektin) iz mišića – sadrži oko 30 000 AK i ima masu skoro 4 MDa.
[slika: sa Swiss-2D PAGE sajta, world-2dpage.expasy.org/swiss-2dpage/]

2
2.3.1. Klasifikacija proteina
Prema složenostiA) Prosti (jednostavni) – samo AKB) Složeni (konjugovani, proteidi) – AK + neproteinska komponenta
• H3PO4 – fosfoproteini• lipidi – lipoproteini• ugljeni hidrati
• glikoproteini (protein + oligosaharid), • proteoglikani (protein + polisaharidi)
• hromoproteini – sa obojenim prostetskim grupama: • hemoproteini (hem), • flavoproteini (flavin-nukleotidi), • metaloproteini (Fe2+, Fe3+, Cu2+, Zn2+, Mg2+, Mn2+)
• nukleinske kiseline - nukleoproteini
Prema 3D strukturiA) Fibrozni (fibrilarni, skleroproteini): kolageni, elastini, keratini, fibroini, retikuliniB) Globularni: albumini, globulini, glutelini, protamini, histoni, prolamini
Prema broju pp lanacaA) monomerniB) oligomerni
Proteini se mogu klasifikovati na različite načine.Prema broju lanaca, proteini se dele na monomerne i oligomerne. Monomerni proteini
sastoje se samo od jednog polipeptidnog lanca, dok oligomerni sadrže više nezavisno umotanih lanaca povezanih nekovalentnim interakcijama.
Prema složenosti, razlikujemo proste proteine, sačinjene samo od peptidnih lanaca, i složene, koji sadrže kovalentno ili nekovalentno vezanu nepeptidnu komponentu.
Na osnovu oblika odn. 3D strukture, proteini se mogu podeliti na globularne i fibrilarne. Globularni proteini su grubo sferičnog oblika, kompaktni, tesno umotani, sa hidrofilnom površinom i hidrofobnom unutrašnjošću. Najčešće imaju funkciju enzima, nosača, regulatornu funkciju itd. Fibrilarni proteini su izduženi, formiraju duga vlakna, i obezbeđuju mehaničku potporu ćelijama, tkivima i organizmu.
[detalji o pojedinim klasama: praktikum]

3
2.3.2. Funkcija proteina1. Održavanje i uspostavljanje oblika:
strukturni proteini – proteini vezivnog tkiva (kolagen, elastin), proteini mišića (miozin, aktin, tropomiozin); histoni –organizuju DNK u hromatinu.
2. Transportni proteini: hemoglobin, prealbumin (transport tiroidnih hormona u krvi), membranski transporteri (nosači, kanali, pumpe), LDL
3. Odbrambena i zaštitna funkcija: imunoglobulini
4. Kontrola i regulacija metabolizma: hormoni, receptori, DNA-vezujući proteini (efektori, induseri, represori)
5. Katalitička funkcija: enzimi
6. Motorni proteini: aktin i miozin
7. Skladišna funkcija: ceruloplazmin, feritin
8. Toksini
Proteini zahvaljujući strukturnoj raznovrsnosti imaju brojne f-je, koje se često preklapaju:1. Strukturna f-ja: Strukturni proteini obezbeđuju definisanje i održavanje oblika subcelularnih
komponenti, ćelija, organa i organizma u celini. Primeri su histoni koji učestvuju u izgradnji hromozoma, aktin i tubulini iz citoskeleta, kolagen i elastin iz vezivnog tkiva, miozin iz mišića, keratin iz kože i rožnatih struktura...
2. Transportna f-ja: Ovi proteini omogućavaju transport različitih molekula u organele, ćelije ili kroz organizam. Primer su nosači, jonski kanali i pumpe koje omogućavaju ćeliji i organelama razmenu materije, serumalbumin koji transportuje masne kiseline i trigliceride kroz krvotok, hemoglobin koji transportuje O2...
3. Odbrambena i zaštitna f-ja: Neki proteini (imunoglobulini – antitela) štite organizam od patogenih mikroorganizama, dok neki (fibrinogen, trombin) učestvuju u koagulaciji krvi i time sprečavaju iskrvavljenje prilikom povreda.
4. Regulacija: Neki od proteina imaju ulogu u regulaciji ćelijskih funkcija. Primeri su neki hormoni (adrenokortikotropni hormon – ACTH, reguliše biosintezu kortikoida, insulin – reguliše koncentraciju glukoze u krvi), receptori (omogućavaju ćelijama i tkivima da reaguju na hemijske signale), DNK-vezujući proteini (regulišu proces transkripcije)
5. Katalitička f-ja: Jedna od najznačajnijih grupa proteina su enzimi, koji katalizuju skoro sve reakcije u organizmu.
6. Motorna f-ja: Različiti proteini omogućavaju proces kretanja. Miozin i aktin omogućavaju kontrakcije mišića, dok dinein pokreće cilije i flagele mikroorganizama i omogućava kretanje organela unutar ćelije.
7. Skladišna f-ja: Neki proteini predstavljaju zalihu aminokiselina za potomstvo, npr. glijadin iz pšeničnog zrna ili ovalbumin iz jajeta. Drugi mogu služiti kao depo određenih materija, npr. feritin koji skladišti i po potrebi otpušta gvožđe.
8. Toksini: Neki proteini već pri veoma niskim koncentracijama mogu ispoljavati izuzetno toksično delovanje. Ovde spadaju životinjski toksini (zmijski, pčelinji otrov), biljni (ricin) i mikrobijalni (botulinum, tetanus toksin).

4
2.3.3. Nivoi strukture proteina• peptidi nemaju stabilnu 3D-strukturu
• proteini se umotavaju u specifičnu stabilnu 3D strukturu uslovljenu sekvencom → jedinstvena nativna konformacija
• 3D strukturu u najvećoj meri stabilizuju nekovalentne interakcije
• struktura nije rigidna; narušavanje konformacije → promene u aktivnosti
• nivoi: primarna, sekundarna, tercijerna, kvaternerna
1 2 3 n
sekvenca 1
1 2 3 n
sekvenca 2
Kao što je pomenuto, polipeptidi kraćeg lanca nemaju stabilnu, kompaktnu 3D strukturu. S druge strane, proteini se skoro uvek umotavaju u specifičnu stabilnu 3D strukturu koja je direktno uslovljena AK sekvencom. Posledica je da svaki protein ima jednu jedinstvenu nativnu konformaciju, koja ispoljava određenu aktivnost i funkciju u organizmu.
3D strukturu u najvećoj meri stabilizuju nekovalentne interakcije (kasnije će biti više reči o tome), ali se kod određenih proteina javljaju i disulfidni mostovi i druge kovalentne veze. 3D struktura proteina nije rigidna, i može se delimično ili potpuno narušiti vezivanjem ligandâ, promenom uslova okoline i sl., što dovodi do promena u aktivnosti.

5
H
O
OH
HH
H
δ+δ+
δ−
δ−
.. ..
....
+I
klasični organski molekuli proteini
empirijskaformula CH2O
brutoformula C2H4O2
molekulskaformula
CH3COOH
strukturnaformula CH3 C
OH
O
sekundarnastruktura
primarnastruktura Ala-Met-Ser-Val-Val-Thr-Ile-
tercijernastruktura
kvaternernastruktura
3D struktura proteina veoma je kompleksna, i tipično se posmatra na 4 nivoa (isto kao što se struktura organskih jedinjenja može posmatrati na različitim nivoima).
Za svaki protein možemo definisati do 4 nivoa strukture. Primarna struktura prvenstveno opisuje redosled AK u polipeptidnom lancu, tj. opisuje položaj kovalentnih veza. Sekundarna, tercijerna i kvaternerna struktura opisuju 3D strukturu proteina, koju prvenstveno stabilizuju nekovalentne interakcije. Sekundarna struktura opisuje lokalno uvijanje polipeptidnog lanca u oblike kao što su α-heliks, β-trake i zavijutci, dok tercijerna struktura opisuje kompletno umotan polipeptidni lanac. Kod oligomernih proteina, kvaternerna struktura opisuje prostorni raspored pojedinačnih, prethodno uvijenih podjedinica.

6
2.3.4. Primarna struktura proteina
• 1° struktura: sekvenca, postranslacione modifikacije, umrežavanje (-SS-)• svi proteini izgrađeni od istih 20 AK• moguće bezbroj kombinacija• jedinstveni redosled (sekvenca) AK određuje strukturu i funkciju datog
proteina:• hidrofobne: u unutrašnjosti, hidrofilne: na površini• Pro: cikličan + ne gradi H-veze → neuređene strukture• Gly: mali, omogućava približavanje lanaca (bitno za petlje i zaokrete)• Ser, Thr: grade vodonične veze, nukleofili, fosforilacija• Cys: gradi disulfidne mostove• His: amfoteran, značajan za enzimsku aktivnost, ligand• Asp, Asn, Lys, Arg: nealektrisane → stabilizacija, elektronski efekti
• održavaju je kovalentne veze: peptidne veze i disulfidni mostovi
Phe–Pro–Phe–Cys–Asn–Gln–Tyr–Val–Cys–Pro–Cys–Leu
S S
1 2 3 4 5 6 7 8 9 10 11 12
Kao što je već pomenuto, iako se svi peptidi sastoje od istih 20 osnovnih proteinogenih AK i njihovih derivata, poznat je veoma veliki broj proteina sa različitom 3D strukturom, svojstvima i funkcijom. Uzrok tome je činjenica da kod peptida, kao informacionih biomolekula, ključnu ulogu igra redosled aminokiselinskih ostataka (sekvenca). Svaki protein karakteriše jedinstvena sekvenca, koja je za dati organizam konstantna*, a često je i između različitih organizama identična ili veoma slična.
Primarna struktura proteina predstavlja njegovu konstitutivnu formulu, odn. opisuje kovalentno povezivanje u molekulu. Ona opisuje sekvencu aminokiselina (povezanih peptidnim vezama), kao i eventualne posttranslacione modifikacije i kovalentno umrežavanje (npr. pozicije disulfidnih mostova). Zbog jačine kovalentnih veza, sekvenca je stabilna i pri fiziološkim uslovima tipično se može narušiti samo dejstvom hidrolitičkih enzima, za razliku od 2°, 3° i 4° strukture, koje su stabilizovane slabim nekovalentnim interakcijama i stoga se lako narušavaju promenom uslova okoline.
Različite AK daju različite doprinose svojstvima i funkciji proteina [već obrađeno kod AK].
*uz neke izuzetke – sekvenca određenih delova molekula imunoglobulina je visoko varijabilna

7
*• anemija srpastih ćelija• hemoglobin = 4 hema + 2 α-globina (po 141 AK) + 2 β-globina (po 146 AK)• tačkasta mutacija → u β-globinskom lancu Glu6 (hidrofilna) zamenjena sa Val
(hidrofobna)• favorizovana agregacija Hb → polimerizacija → vlakna → deformacija
eritrocita, gubitak elastičnosti• → skraćen život eritrocita → anemija• → otežan prolazak kroz kapilare → ishemija
normalni Hb polimer HbS
Na značaj primarne strukture najbolje ukazuju obolenja koja nastaju kao posledica pogrešno izgrađene sekvence. Dobar primer je anemija srpastih ćelija, koja je posledica greške u formiranju β-globinskog lanca*. Tačkasta mutacija (zamena jedne azotne baze) u genu koji kodira β-globin dovodi do zamene glutaminske kiseline u poziciji 6 valinom (dakle, zamenjena je samo jedna od 146 AK). Zamena hidofilne AK hidrofobnom dovodi do formiranja hidrofobnog džepa na površini, koji favorizuje agregaciju molekula hemoglobina i polimerizaciju u duga vlakna. Formirane vlaknaste strukture dovode do deformacije eritrocita (dobijaju karakteristični srpasti oblik) što im s jedne strane skraćuje životni vek i vodi anemiji, a s druge otežava prolaz kroz kapilare i dovodi do ishemije (slabe prokrvljenosti).
*hemoglobin se sastoji od proteina – globina – sastavljenog od 2 α i 2 β lanca; za svaki od lanaca vezan je po jedan hem

8
2.3.5. 3D struktura proteina• svi proteini imaju definisanu 3D strukturu• opisana sekundarnom, tercijernom i kvaternernom strukturom• najstabilnija i najaktivnija konformacija – nativna konformacija• 3D struktura fleksibilna → promene u aktivnosti, prilagođavanje supstratu...• aktivnost i f-ja – uslovljene 3D rasporedom funkcionalnih grupa
Skoro svi* proteini imaju dobro definisanu, uređenu, stabilnu 3D strukturu koja se opisuje sekundarnom, tercijernom i (kod oligomernih proteina) kvaternernom strukturom. Od bezbroj mogućih konformacija, protein se u ćeliji uvija u jednu određenu, nativnu konformaciju, koja je pri fiziološkim uslovima nastabilnija i istovremeno najaktivnija. Ova 3D struktura je donekle fleksibilna (moguća je ograničena rotacija oko jednostrukih veza, pomeranje pojedinih domena, bočnih nizova...) čime se omogućava prilagođavanje supstratu (ligandu), promena u aktivnosti, kretanje itd. 3D struktura, odn. 3D raspored funkcionalnih grupa, kritična je za ispoljavanje aktivnosti i funkcije; narušavanje nativne konformacije rezultuje smanjenjem ili gubitkom aktivnosti.
Uobičajene hemijske formule nepraktične su za dočaravanje 3D strukture proteina, te se koriste različiti specijalizovani prikazi. Spacefill/balls prikazuje svaki atom kao sferu. Dobro dočarava oblik proteina, ali ne daje informacije o sekundarnoj strukturi, kao i o unutrašnjosti molekula. Sticks prikazuje samo veze, što pojednostavljuje prikaz i pruža i uvid u unutrašnjost globule. Backbone prikazuje samo polipeptidnu kičmu (ili čak samo αC), bez bočnih nizova, što olakšava razumevanje 3D uvijanja proteina. Elementi sekundarne strukture najbolje se uočavaju u cartoon prikazu, koji α-helikse obeležava spiralama/cilindrima, β-trake – trakastim strelicama, a neuređene regione – tankim cevima. Na kraju, prikaz površine proteina omogućava uočavanje žljebova na površini, koji mogu imati funkciju mesta za vezivanje liganda.
[na slikama je metalošaperon 1cc8; slike su generisane korišćenjem programa YASARA View]*Intrinsically disordered proteins nemaju stabilnu 2° i 3° strukturu

9
2.3.5.1. Faktori stabilizacije 3D strukture
• uvijanje proteina u nativnu konformaciju je spontano • promena Gibsove energije ΔG = ΔH – TΔS ΔG < 0 • kod uvijanja proteina:
• uspostavljene interakcije AK-AK• uspostavljene H-veze voda-voda
• molekuli vode oslobođenipovoljno
• raskinute H-veze AK-voda• kruta struktura → AK fiksiranenepovoljno
entalpija, ΔHentropija, ΔS
• 3D struktura uslovljena i stabilizovana nekovalentnim interakcijama:1. elektrostatičke2. vodonične veze3. van der Waals-ove interakcije4. hidrofobne “interakcije”
• slabe – energija veze 2–30 kJ/mol vs. 200–800 kJ/mol kod kovalentnih• brojne• omogućavaju izvesnu fleksibilnost• dodatno: kovalentne veze – disulfidni mostovi...
Uvijanje proteina (protein folding) u nativnu konformaciju spontano se odvija u ćeliji, što znači da je proces egzergon (ΔG < 0). Energetski bilans uvijanja (ΔG) sastoji se od dve komponente – ΔH i ΔS. Entropijski faktor povezan je sa slobodom kretanja atoma i molekula; prilikom umotavanja proteina nastaje relativno kruta struktura, što je entropijski nepovoljno (ΔSprotein < 0). Međutim, istovremeno se oslobađa veliki broj molekula vode (koji su do tada solvatisali polarne grupe lanca ili gradili klatrate oko nepolarnih), zbog čega je ΔSokoline > 0 i ΔSukupno > 0 tj. proces je entropijski povoljan. Uvijanje je povoljno i entalpijski – iako dolazi do narušavanja privlačnih interakcija između molekula vode i aminokiselinskih ostataka (endoterman proces, ΔH > 0), istovremeno se uspostavlja veliki broj nekovalentnih interakcija između ostataka aminokiselina (o čemu će dalje biti reči) a, dodatno, oslobođeni molekuli vode sada se mogu vodonično vezivati za okolne molekule vode. Zbog svega ovoga, uvijanje je i entalpijski povoljno odn. egzotermno (ΔH < 0).
Interakcije između AK koje stabilizuju 3D strukturu proteina pretežno su nekovalentne prirode, i uključuju elektrostatičke, van der Waals-ove, hidrofobne interakcije i vodonične veze. Ove interakcije su slabe (mnogo slabije od kovalentnih) ali brojne, te je njihov sumarni efekat značajan. Istovremeno, one dopuštaju proteinskom molekulu izvesnu fleksibilnost, koja je neophodna za mnoge funkcije.
Pored nekovalentnih interakcija, proteinsku strukturu mogu dodatno stabilizovati i kovalentne veze kao što su disulfidni mostovi.

10
• Elektrostatičke interakcije• najjače (5,9–12 kJ/mol)• privlačne/odbojne• između naelektrisanih grupa• po Coulomb-ovom zakonu: 1 2
20
q q 1Fr 4πε⋅= ⋅
O
O-
NH3+
• ukoliko se istovremeno uspostavlja i H-veza: soni most• na ove interakcije značajno utiče prisustvo rastvorenih supstanci:
O
O- O
O
-Mg
2+
naelektrisanje katjona zaklanja nael. anjona ⇒ manje odbijanje
NO
O
OH
N
NN
NH2
OH
P
O
O-
P
O
O
O-
O-
O-
O
O
P
Mg2+
Najjače nekovalentne interakcije su elektrostatičke (Coulomb-ovske, jonske) interakcije, koje se javljaju između naelektrisanih grupa (grupa sa permanentnom šaržom). Ove interakcije mogu biti privlačne (ukoliko su grupe jednako naelektrisane) ili odbojne (ukoliko su naelektrisanja suprotnog znaka), i opisuju se Coulomb-ovim zakonom. Iako su snažne, doprinos stabilizaciji proteinske strukture im je mali, pošto se AK sa naelektrisanim bočnim nizom pretežno nalaze na površini proteinske globule i interaguju sa vodom. Međutim, u nekim slučajevima mogu se formirati i jonski parovi između pozitivno i negativno naelektrisanih grupa unutar globule. Kod nekih grupa dodatno se formiraju i vodonične veze, pri čemu nastaju tzv. soni mostovi (salt bridges).
Na ove interakcije značajno utiče prisustvo rastvorenih supstanci (o čemu će biti više reči kod denaturacije). Promena pH dovodi do protonacije/deprotonacije i promene naelektrisanja funkcionalnih grupa, što dovodi do raskidanja/formiranja jonskih parova. Organski rastvarači koji smanjuju dielektričnu konstantu (ε) po Coulomb-ovom zakonu pojačavaju elektrostatičke interakcije. S druge strane, metalni katjoni mogu maskirati naelektrisanje negativno naelektisanih grupa i smanjiti njihovo odbijanje, čime stabilizuju strukturu. Odličan (neproteinski) primer imamo kod ATP, kod koga vezivanje Mg2+ ublažava odbijanje 4 negativne šarže u trifosfatnom ostatku.

11
• Vodonične veze• privlačne• 4–30 kJ/mol – jače od vtW interakcija, slabije od kovalentnih veza• između donora – H-atoma vezanog za elektronegativan atom (O, N, S) – i
akceptora – drugog elektronegativnog atoma:
δ–X–Hδ+ + Yδ– → X–H···Y
• u izgradnji H-veze učestvuju elektrostatičke interakcije, prenos šarže sa donora na akceptor (→ parcijalna kovalentna veza) i disperzione interakcije
• donori: >N–H, –O–H• akceptori: >O, –O–H, –COO–, >N–
interlančaneH-veze
intralančaneH-veze
Za razliku od elektrostatičkih interakcija, vodonične veze* uvek su privlačne. Jače su od van der Waals-ovih interakcija, mada i dalje slabije od kovalentnih i jonskih veza. Uvek se uspostavljaju između donora, odn. atoma vodonika koji je vezan za elektronegativni atom i stoga nosi parcijalnu pozitivnu šaržu, i akceptora – nekog elektronegativnog atoma. Ne predstavljaju jednostavne dipol-dipol interakcije, nego uključuju i delimično uspostavljanje kovalentne veze i disperzione interakcije (indukovani dipol-indukovani dipol).
Kod biomolekula, tipični donori vodonične veze su amino-, amidne i hidroksilne grupe, a akceptori – hidroksilne, amino-grupe, karboksilatni joni, heterociklična jedinjenja sa N itd.
Vodoničnim povezivanjem donora i akceptora formiraju se mostovi koji povezuju različite delove lanca (intralančane H-veze) ili različite lance (interlančane H-veze). Pošto kod proteina svaka peptidna grupa može da gradi po dve H-veze (jednu između kiseonika i nekog donora, drugu između N-vezanog vodonika i donora), vodonična veza značajno doprinosi stabilizaciji strukture i ključna je za prvu fazu umotavanja proteina (formiranje sekundarne strukture).
*IUPAC: The hydrogen bond is an attractive interaction between a hydrogen atom from a molecule or a molecular fragment X–H in which X is more electronegative than H, and an atom or a group of atoms in the same or a different molecule, in which there is evidence of bond formation.
12
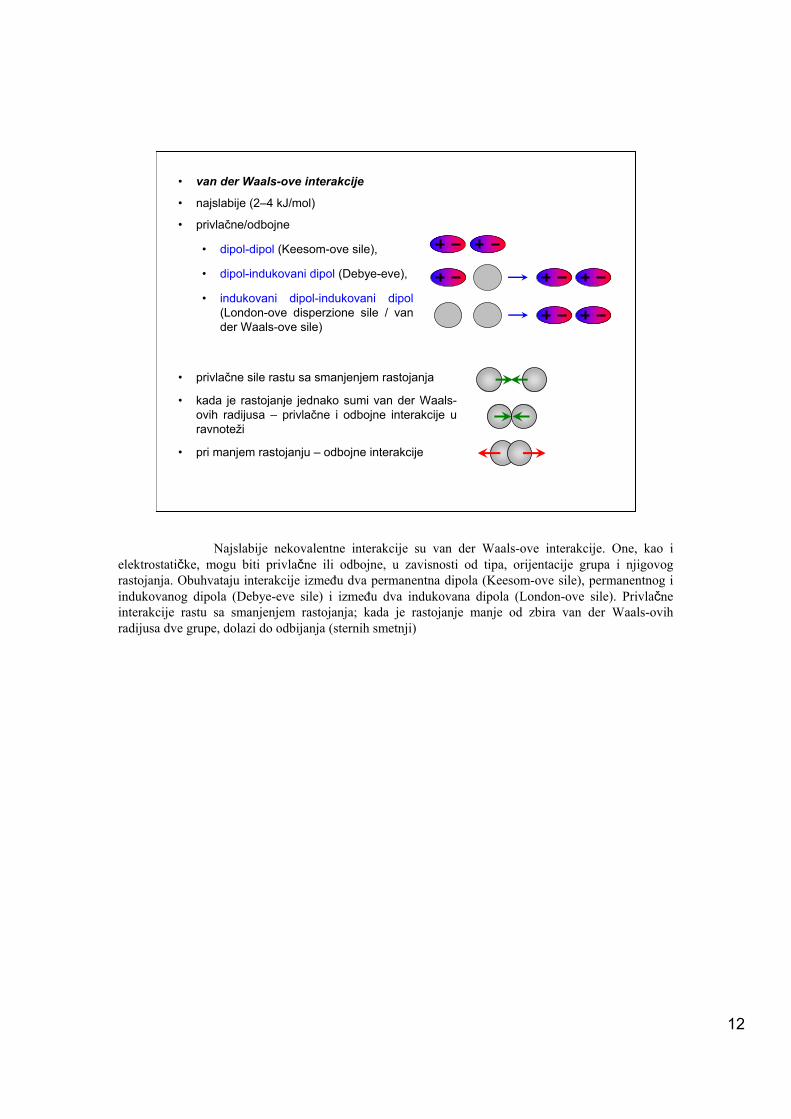
• van der Waals-ove interakcije
• najslabije (2–4 kJ/mol)
• privlačne/odbojne
• dipol-dipol (Keesom-ove sile),
• dipol-indukovani dipol (Debye-eve),
• indukovani dipol-indukovani dipol(London-ove disperzione sile / van der Waals-ove sile)
• privlačne sile rastu sa smanjenjem rastojanja
• kada je rastojanje jednako sumi van der Waals-ovih radijusa – privlačne i odbojne interakcije u ravnoteži
• pri manjem rastojanju – odbojne interakcije
Najslabije nekovalentne interakcije su van der Waals-ove interakcije. One, kao i elektrostatičke, mogu biti privlačne ili odbojne, u zavisnosti od tipa, orijentacije grupa i njigovog rastojanja. Obuhvataju interakcije između dva permanentna dipola (Keesom-ove sile), permanentnog i indukovanog dipola (Debye-eve sile) i između dva indukovana dipola (London-ove sile). Privlačne interakcije rastu sa smanjenjem rastojanja; kada je rastojanje manje od zbira van der Waals-ovih radijusa dve grupe, dolazi do odbijanja (sternih smetnji)

13
• naročito značajne kod interakcija hidrofobnih grupa – alkil, aril
slaganje prstenova
slaganje baza kod NKinterakcije alkil-nizova kod lipida u membrani
Van der Waals-ove interakcije su naročito značajne kod hidrofobnih grupa –ugljovodoničnih nizova i prstenova – koji uglavnom ne mogu interagovati na druge načine, a koji tipično čine 30–50 % bočnih nizova u proteinu. Jedine privlačne sile moguće između alkil-nizova su indukovani dipol-indukovani dipol (Londonove interakcije), i one su naročito značajne kod derivata masnih kiselina (lipida), mada se mogu javiti i proteina, između nepolarnih bočnih nizova unutar globule.
Van der Waals-ove interakcije javljaju se i između π-sistema aromatičnih prstenova, i rezultuju karakterističnim paralelnim slaganjem prstenova (naročito bitno kod nukleinskih kiselina). Energija ovih, tzv. π-interakcija, iznosi oko 10 kJ/mol.
Treba napomenuti da ovde navedene interakcije između hidrofobnih grupa ne spadaju u “hidrofobne interakcije”.

14
• hidrofobne interakcije (hidrofobni efekat)
hidrofobni molekul zamenjuje H2O u mreži H-vezanih molekula → manji broj H-veza → ΔH > 0
1 2
3
54
reorganizacija vode → klatrati→ više H-veza → ΔH < 0→ više fiksiranih molekula H2O → ΔS < 0
agregacija kavezâ → manja površina (manje fiksiranih molekula H2O) → ΔS > 0
Hidrofobne interakcije zapravo ne predstavljaju realne privlačne/odbojne sile per se, te ih je pravilnije zvati hidrofobnim efektom. Ovaj efekat ispoljava se kao težnja hidrofobnih molekula ili grupa da se u vodenoj sredini agregiraju. Međutim, on zapravo predstavlja posledicu težnje vode sa smanji površinu kontakta sa hidrofobnim česticama, a u cilju povećanja broja molekula vode koji su slobodni (imaju visoku entropiju) i vodonično vezani (niska entalpija).
Vodena sredina sastoji se od molekula vode povezanih vodoničnim vezama u mrežu. Kada se hidrofobni molekul nađe u vodenoj sredini, on istiskuje deo molekula vode iz mreže, pri čemu se smanjuje broj vodoničnih veza. Raskidanje H-veza entalpijski je nepovoljan proces (ΔH > 0). U cilju maksimizovanja broja H-veza (smanjenja H), rešetka se reorganizuje i stvaraju se kavezi (klatrati) od molekula vode raspoređenih oko hidrofobnog rastvorka. Klatrati su veoma uređene strukture, koje sadrže mnogo molekula sa ograničenom slobodom kretanja, što je entropijski nepovoljno (ΔS < 0). Kavezi se dalje agregiraju, čime se ukupna površina smanjuje, a samim tim smanjuje se i broj “uređenih”, vezanih molekula vode. Povećan broj slobodnih molekula H2O entropijski je povoljan (ΔS> 0), što čini ceo proces povoljnim i spontanim (ΔG < 0).

15
• posledica kod proteina: hidrofilne grupe spolja, hidrofobne u globuli
nepolarnepolarne nenaelektrisanepolarne naelektrisane
površina
unutrašnjost
Hidrofobni efekat ključan je za formiranje 3D strukture proteina – u vodenoj sredini proteinski lanac kolabira tako da hidrofobni bočni nizovi aminokiselinskih ostataka grade kompaktnu globulu (dok hidrofilnije grupe pretežno ostaju na površini).

16
• disulfidni mostovi
• kovalentne veze nastale oksidacijom Cys• proteini sa brojnim S-S mostovima
pretežno u ekstracelularnom prostoru (koji je oksidujući)
• redukciona sredina u citoplazmi destabilizuje S-S mostove
–S-S––SHHS–
interlančani
interlančaniintralančani
Kao što je već pomenuto kod aminokiselina, oksidacijom bočnih grupa (SH-grupa) dva cisteinska ostatka formira se disulfidni most, koji takođe može učestvovati u stabilizaciji strukture. Disulfidni mostovi nisu uobičajeni kod intracelularnih proteina. Pretežno se javljaju kod proteina u ekstracelularnom prostoru (koji je oksidujući) i sekretornih proteina (koji su izloženi nepovoljnim uslovima u spoljašnjoj sredini).
Disulfidni mostovi nemaju ulogu u usmeravanju uvijanja proteina kao ostale interakcije. Za uspostavljanje mostova na korektnim pozicijama, neophodno je da protein već bude potpuno uvijen. Međutim, oni doprinose stabilizaciji 3D strukture već uvijenog proteina.

17
2.3.5.3. Sekundarna struktura proteina• lokalno uređenje strukture (konformacije) polipeptidne kičme• ne razmatra se konformacija bočnih ostataka
• prilikom umotavanja:→ maksimalni broj H-veza između N–H i C=O peptidnih grupa→ minimalne sterne smetnje
• dolazi do uvijanja kičme, uz formiranje različitih elemenata sekundarne strukture:
• heliks (α, π, 310)• β-niz → β-nabrana struktura• β-zavijutak...
β-traka
petljaα-heliks
• uslovljena 1° strukturom polipeptidnog lanca (prirodom bočnih nizova)
Sekundarna struktura opisuje lokalno 3D uređenje (uvijanje) strukture polipeptidnogniza, odn. konformaciju polipeptidne kičme. Konformacije bočnih nizova svake aminokiseline pritom se ne razmatraju. Sekundarna struktura okarakterisana je kada se za svaki AK ostatak definiše konformacija.
Sekundarna struktura uslovljena je primarnom – od prirode bočnih nizova zavisiće najpovoljnije konformacija koje će date aminokiseline zauzeti pri uvijanju lanca. Prilikom uvijanja, polipeptidni lanac zauzimaće takve konformacije da se obezbedi maksimalan broj vodoničnih veza između polarnih grupa (N–H i C=O peptidnih grupa), uz minimalne sterne smetnje. Najčešće konformacije koje pp lanac poprima uključuju α-heliks (i, ređe, druge tipove heliksa), β-nizove (β-trake) koje se dalje udružuju u β-nabrane strukture, kao i različite neuređene strukture – zavijutke i petlje.

18
N
O
H
N+
O-
H• C–N veza: parcijalni karakter dvostruke veze → otežana rotacija → torzioni ugao ω =180° (trans)
• sp2 → koplanarni atomi: Cα, C=O, N-H, Cα
• peptidna veza ograničava broj mogućih konformacija pp lanca
• svaki AK ostatak ima 2 stepena slobode (2 veze oko kojih je moguća rotacija); konformacija je opisana sa dva torziona ugla:
• αC—C(=O) - ugao ψ
• N–αC - ugao φ
• 3D struktura pp kičme potpuno je opisana ψi φ uglovima svih AK
Zašto pp lanac formira samo nekoliko različitih konformacija? Moglo bi se očekivati da je oko svake od 3 jednostruke veze koje grade peptidnu kičmu (N–αC, αC–CO i CO–N) moguća potpuno slobodna rotacija i neograničen broj konformacija.
Međutim, u stvarnosti postoji niz ograničenja. Kao što je već pomenuto, peptidna C–N veza ima parcijalni karakter dvostruke veze, te je rotacija oko nje otežana, što ograničava broj mogućih konformacija polipeptidnog lanca. Torzioni ugao peptidne (CO–N) veze, ω, najčešće iznosi 180°, odn. ona ima trans konformaciju (sem u slučaju Pro, kod koga cis nije mnogo nepovoljnija).
Iako svi atomi peptidne grupe (αC, C=O, NH i αC) leže u istoj ravni, svaki AK ostatak i dalje ima 2 stepena slobode, odn. rotacija je moguća oko 2 veze: N–αC i αC–C(O). 3D struktura polipeptidne kičme potpuno je opisana sa dva torziona ugla, ψ (psi) i φ (fi). Ugao ψ odnosi se na vezu između αC i karbonilnog C, a ugao φ – na vezu između α-N i α-C.

19
teorijski Ramachandran-ov dijagram
• neki ψ i φ uglovi verovatniji i pogodniji od drugih:
• sterne smetnje (kolizija između grupa)
• manji bočni ostaci (npr. Gly, Ala...) imaju više dozvoljenih konformacija nego voluminozni (npr. Trp)
• kod Pro zbog ciklične strukture φ je fiksiran na –60° do –77° → zavoj u pp kičmi
ψ=0φ=0
• α-heliks: ψ ≈ –47°, φ ≈ –57°• 310-heliks: ψ ≈ –49°, φ ≈ –26°• β-traka: ψ ≈ 135°, φ ≈ –135°
Iako su αC–CO i N–αC čiste jednostruke veze oko kojih je rotacija slobodna, neki ψ i φ uglovi verovatniji su i pogodniji od drugih. Naime, rotacija oko N–αC i αC–C(O) veza ograničena je sternim smetnjama između atoma kičme i atoma iz bočnih nizova (npr. između R i karbonilnog O). Rotaciju naročito ograničavaju voluminozni bočni nizovi (npr. aromatičnih aminokiselina). Prolin je specijalan slučaj – zbog ciklične strukture, rotacija oko N–αC veze je nemoguća, i ugao φ fiksiran je na oko –60° do –77°. S druge strane, glicin ima znatno veću slobodu od ostalih AK zahvaljujući odsustvu β-C.
Sve moguće konformacije polipeptidne kičme prikazuju se Ramachandran-ovim dijagramom. Ose predstavljaju ceo opseg vrednosti φ i ψ, od –180° do +180°. Beli regioni predstavljaju konformacije koje su nepovoljne ili nemoguće zbog sternih smetnji (sem za Gly), svetloplavi regioni –dozvoljene konformacije za aminokiseline sa malim bočnim nizom koji ne ograničava rotaciju (npr. Ala), a tamnoplavi – dozvoljene konformacije za većinu AK. Kao što se može primetiti, samo neke konformacije su povoljne, i one odgovaraju elementima sekundarne strukture detektovanim u proteinima – α-heliksu, β-traci, zavijutcima itd.

20
α-Heliks
• bočni ostaci – na spoljnoj strani heliksa, skoro normalni na osu
• nastaje uvijanjem pp lanca u desnu spiralu
• jedan navoj dug je 0,54 nm i sadrži 3,6 AK
• unutrašnjost heliksa nije prazna
α-heliks nastaje uvijanjem polipeptidnog lanca u desnu spralu [pravilo desne ruke]. Dok je leva spirala takođe moguća, kod L-AK manje je uobičajena zbog sternih smetnji. Kod idealnog α-heliksa, jedan namotaj dug je 0,54 nm i sadrži 3,6 AK ostataka; u praksi, kod proteina, dolazi do izvesnog krivljenja. Unutrašnjost α-heliksa nije prazna (ne formira se kanal), nego kompaktna –polipeptidna kičma tesno je uvijena.
Bočni AK ostaci se nalaze na spoljašnjoj strani heliksa i skoro su normalni na osu. Priroda bočnih ostataka može uticati na stabilnost heliksa, zbog čega se neke AK češće nalaze u sklopu heliksa od drugih. Na primer, Ala (sa malim bočnim nizom) dobro se uklapa u heliks, dok su Tyr i Arg manje uobičajeni. Gly, zbog odsustva β-C, ima veliku fleksibilnost odn. konformacionu slobodu (entropijski je nepovoljno da bude fiksiran), te destabilizuje heliks. Prolin takođe narušava i prekida strukturu α-heliksa – zbog odsustva H-atoma na amidnom N ne gradi vodoničnu vezu, krut je te se loše uklapa u konformaciju ostatka kičme, a njegov bočni niz preklapao bi se sa bočnim nizovima susednih AK. Prolin se zbog toga retko nalazi u α-heliksima, mada može da ulazi u sastav nekih drugih.

21
• sve amidne grupe međusobno povezane vodoničnim vezama: veže se >C=O: grupa n-te AK sa N-H grupom (n+4)-te AK
• vodonične veze paralelne sa prostiranjem lanca
• svaka peptidna grupa učestvuje u H-vezivanju → maksimalna stabilnost
• α-heliks najstabilnija konformacija pp niza → spontano formiranje
Kod α-heliksa, karbonilni kiseonik svake n-te AK povezan je vodoničnom vezom sa amidnim vodonikom (n+4)-te AK, tj. AK iz sledećeg zavoja. Ove vodonične veze skoro su paralelne sa osom prostiranja heliksa. Iako su vodonične veze slabe, njihova brojnost rezultuje stabilizacijom 3D strukture α-heliksa i njegovom rigidnošću. α-heliks je, generalno, najstabilnija konformacija polipeptidnog lanca i spontano se formira.
Pored ovakvog, 3.613 heliksa (3.6 AK po namotaju, H-veza zatvara 13-člani prsten), kod nekih proteina javlja se i 310 heliks, koji je kompaktniji – H-veza uspostavlja se između n-te i (n+3)-e AK – ali i manje stabilan zbog sternih smetnji.

22
• pretežno nepolarne AK + amidne grupe već uključene u građenje H-veza → α-heliks je hidrofoban → često je u sklopu membranskih proteina
nepolarnepolarne nenaelektrisanepolarne naelektrisane
membranskilipidi
hidrofobna strana
hidrofilna strana
Pošto su sve amidne grupe α-heliksa uključene u građenje intramolekulskih vodoničnih veza, njihove C=O i N-H grupe ne mogu graditi vodonične veze sa okolnim rastvaračem, te je α-heliks hidrofoban. Bočni nizovi nepolarnih aminokiselina dodatno mogu povećavati hidrofobnost heliksa. Zahvaljujući ovome, α-heliks se često nalazi u sklopu transmembranskih proteina (proteina koji prodiru kroz lipidni dvosloj bioloških membrana).[na slici: fotosistem II, 3bz2]
Druga mogućnost je da je heliks amfipatičan – na jednoj strani zastupljene su hidrofobne AK, na drugoj hidrofilne. Ova situacija javlja se kod proteina na površini membrane. Heliks je položen na membranu, tako da je hidrofobna strana orijentisana ka hidrofobnoj unutrašnjosti, a hidrofilna ka spoljašnjoj sredini.

23
β-struktura• izduženi polipeptidni lanci• kod globularnih proteina – do 15 AK ostataka (u proseku 6)• 2-15 β-traka/nizova (β-strand) postavljenih paralelno ili antiparalelno
→ β-nabrana struktura/pločica (β-pleated sheet)• H-veze – između dve susedne trake
paralelni antiparalelni
Drugi uobičajeni tip sekundarne strukture je β-struktura, koja uključuje β-trake (β-strands) i njihovim povezivanjem nastale β-nabrane pločice (β-pleated sheet). Za razliku od α-heliksa, koji su uvijeni i kompaktni, polipeptidni lanci su kod β-strukture skoro potpuno istegnuti. Pojedinačne β-trake su retke, jer nisu ništa stabilnije od drugih mogućih konformacija. Međutim, kada se trake postave jedna pored druge, dolazi do uspostavljanja brojnih vodoničnih veza između peptidnih grupa iz dve susedne trake, čime nastaje mnogo stabilnija β-ploča. Trake koje se povezuju mogu biti različiti delovi istog lanca, ili čak različiti polipeptidni lanci.
β-trake koje izgrađuju β-pločice mogu biti postavljene paralelno ili antiparalelno. Kod paralelne β-pločice, svaka AK povezuje se vodonično sa 2 AK iz susednog lanca, dok se kod antiparalelne svaka AK povezuje samo sa jednom AK iz drugog lanca, pri čemu sa njom gradi 2 vodonične veze koje su normalne na osu prostiranja trake (što je verovatno uzrok većoj stabilnosti antiparalelne strukture).

24
• pp niz nije potpuno istegnut – nabrana struktura• bočni ostaci – iznad i ispod ravni• pločice obično nisu planarne, nego izvijene udesno
Polipeptidni niz kod β-strukture nije potpuno istegnut, nego je naboran (kao harmonika), zbog čega se ona često naziva β-nabranom strukturom. Bočni ostaci AK smešteni su naizmenično iznad i ispod ravni pločice, pri čemu jedna površina može biti hidrofilna a druga hidrofobna. Dalje, pločice nisu savršeno planarne, nego su često izvijene (tipično, udesno odn. u smeru kazaljke na satu, ako se posmatra duž jednog lanca).

25
Neuređene strukture
• kraći neuređeni delovi koji omogućavaju promenu pravca pp kičme
petlja• zavijutci (engl. turns) – 1–5 AK ostataka (δ, γ, β, α i π-zavijutak)
• formiraju se 1-2 vodonične veze
• najčešći – β-zavijutak (4 AK), menja smer pp lanca za 180°; povezuje dve antiparalelne β-trake
• ω-petlja – duži ili neuređeni zaokreti bez fiksnih vodoničnih veza
I α-helikse i β-trake čini niz AK koje se nalaze u sličnoj konformaciji, odn. imaju slične φ i ψ uglove, te predstavljaju uređene, ponavljajuće strukture. Pored njih, u proteinima se javljaju i petlje – segmenti sa neponavljajućom 3D-strukturom, koji mogu činiti i do 50 % strukture kod uobičajenih proteina. One povezuju pojedinačne α-helikse i/ili β-trake i omogućavaju da se protein umota u globularnu strukturu. Često sadrže hidrofilne AK i nalaze se na površini proteina, izložene vodi sa kojom grade vodonične veze.
Petlje koje sadrže manji broj AK ostataka (1–5) a koje dovode do promene pravca i/ili smera polipeptidne kičme nazivaju se zavijutcima. Prema broju peptidnih veza, označavaju se sa δ(jedna veza tj. 2 AK), γ (2 veze), β (3 veze), α (4 veze) i π-zavijutak (5 peptidnih veza). Najuobičajeniji su β-zavijutci, koje čine 4 AK, a koji menjaju smer lanca za 180° čime, pored ostalog, omogućavaju povezivanje dve antiparalelne β-trake*. Između 1. i 4. AK u zavijutku uspostavlja se vodonična veza.
Duge, neuređene petlje bez fiksnih vodoničnih veza označavaju se kao ω-petlje.
*zavijutci koji povezuju antiparalelne β-trake često se nazivaju β-ukosnice (bez obzira na konkretan tip zavijutka)

26
2.3.5.4. Tercijarna struktura proteina• kompletna 3D struktura celokupnog polipeptida (svih atoma), uključujući i konformacije bočnih ostataka
• formirane 2° strukture orijentišu se i pakuju u konačni oblik – nativnu konformaciju
• stabilizovana nekovalentnim interakcijama (elektrostatičke, H-veze, van der Waals-ove, hidrofobne interakcije) i disulfidnim mostovima
• ključna za funkciju – formiranje aktivnog centra
• nije kruta, oblik molekula je dinamičan → prilagođavanje enzima supstratu, kretanje...
Tercijerna struktura opisuje kompletnu 3D strukturu celokupnog monomernog proteina, uključujući ne samo φ, ψ i ω torzione uglove polipeptidne kičme nego i konformacije svih bočnih ostataka. Nastaje daljim umotavanjem peptida (sa potpuno ili delimično formiranim elementima sekundarne strukture) u kompaktnu 3D strukturu. Pritom, AK koje su udaljene u sekvenci orijentišu dolaze u bliski kontakt i između njih se mogu uspostaviti nekovalentne interakcije (ili čak i kovalentne veze, kao što su disulfidni mostovi). Prostorno orijentisanje AK ostataka ključno je za ispoljavanje biološke aktivnosti i funkcije, jer omogućava formiranje aktivnih centara (koje tipično čine bočni nizovi AK udaljenih u sekvenci). Treba napomenuti da tercijerna struktura nije kruta – naročito zahvaljujući petljama, oblik molekula je dinamičan i konformacija se može u izvesnoj mer menjati, čime se omogućava prilagođavanje obliku supstrata (kod enzima), modifikacija aktivnosti, mehanički rad (kretanje) itd.
Dok sekundarnu strukturu stabilizuju vodonične veze između peptidnih grupa, tercijernu strukturu prvenstveno stabilizuju ostale nekovalentne interakcije – elektrostatičke, van der Waals-ove i naročito hidrofobne interakcije.
27
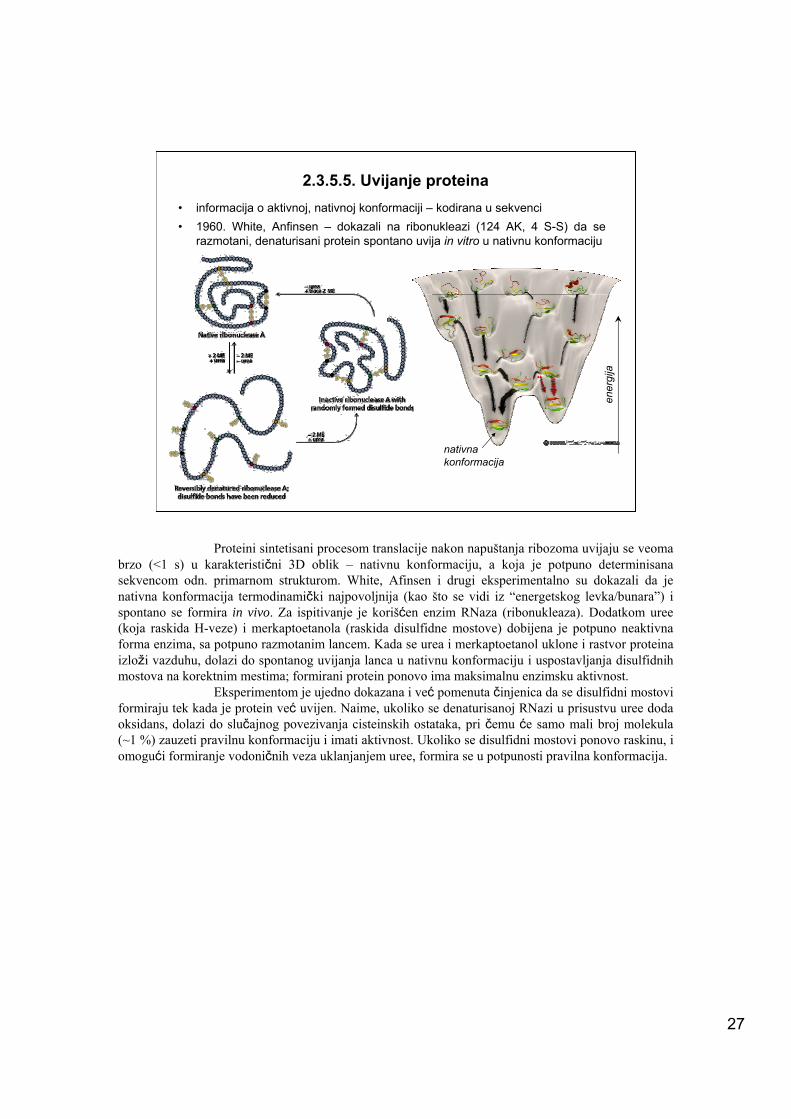
2.3.5.5. Uvijanje proteina• informacija o aktivnoj, nativnoj konformaciji – kodirana u sekvenci• 1960. White, Anfinsen – dokazali na ribonukleazi (124 AK, 4 S-S) da se
razmotani, denaturisani protein spontano uvija in vitro u nativnu konformaciju
ener
gija
nativnakonformacija
Proteini sintetisani procesom translacije nakon napuštanja ribozoma uvijaju se veoma brzo (<1 s) u karakteristični 3D oblik – nativnu konformaciju, a koja je potpuno determinisana sekvencom odn. primarnom strukturom. White, Afinsen i drugi eksperimentalno su dokazali da je nativna konformacija termodinamički najpovoljnija (kao što se vidi iz “energetskog levka/bunara”) i spontano se formira in vivo. Za ispitivanje je korišćen enzim RNaza (ribonukleaza). Dodatkom uree (koja raskida H-veze) i merkaptoetanola (raskida disulfidne mostove) dobijena je potpuno neaktivna forma enzima, sa potpuno razmotanim lancem. Kada se urea i merkaptoetanol uklone i rastvor proteina izloži vazduhu, dolazi do spontanog uvijanja lanca u nativnu konformaciju i uspostavljanja disulfidnih mostova na korektnim mestima; formirani protein ponovo ima maksimalnu enzimsku aktivnost.
Eksperimentom je ujedno dokazana i već pomenuta činjenica da se disulfidni mostovi formiraju tek kada je protein već uvijen. Naime, ukoliko se denaturisanoj RNazi u prisustvu uree doda oksidans, dolazi do slučajnog povezivanja cisteinskih ostataka, pri čemu će samo mali broj molekula (~1 %) zauzeti pravilnu konformaciju i imati aktivnost. Ukoliko se disulfidni mostovi ponovo raskinu, i omogući formiranje vodoničnih veza uklanjanjem uree, formira se u potpunosti pravilna konformacija.

28
• uvijanje teče hijerarhijski:1. formiranje sekundarnih struktura – uslovljeno sekvencom
2. formiranje supersekundarnih struktura – motivâ3. formiranje domena (kompaktnih, stabilnih, nezavisnih delova)4. uspostavljanje tercijarne strukture
domenmotiv
Uvijanje proteina od izduženog niza do konačne, kompaktne 3D strukture teče hijerarhijski. Glavna pokretačka sila za formiranje 2° struktura je građenje H-veza, a za 3° strukturu –hidrofobni efekat.
U prvoj fazi, formiraju se elementi sekundarne strukture (α-heliksi, β-trake itd.) pravilnim orijentisanjem AK ostataka i njihovim povezivanjem vodoničnim vezama. Konačna konformacija polipeptidne kičme strogo je uslovljena sekvencom.
U sledećoj fazi, heliksi, trake i petlje povezuju se u supersekundarne strukture, tzv. motive, kao što su β-ukosnica (2 antiparalelne β-trake povezane zavijutkom), heliks-petlja-heliks, β-meander i grčki ključ (niz antiparalelnih β-traka povezanih zavijutcima), β-α-β (dve paralelne β-trake između kojih je α-heliks, za njih povezan zavijutcima) itd.
Dalje se kod mnogih proteina formiraju domeni – kompaktne jedinice koje se uvijaju nezavisno od ostatka molekula, i koje mogu da sadrže od 25 do preko 300 AK. Pojedinačni domeni često predstavljaju funkcionalne celine, odn. imaju definisanu strukturu u sklopu datog proteina (npr. sadrže aktivni centar, omogućavaju vezivanje alosternih aktivatora ili inhibitora). Kod drugih proteina aktivni centar formira se na graničnoj površini između dva domena.
Finalnim uvijanjem formira se konačna tercijerna struktura – nativna konformacija.
* napomena: po nekim simulacijama, formiranje sekundarnih struktura odvija se istovremeno sa globalnim uvijanjem (hidrofobnim kolapsom)
29
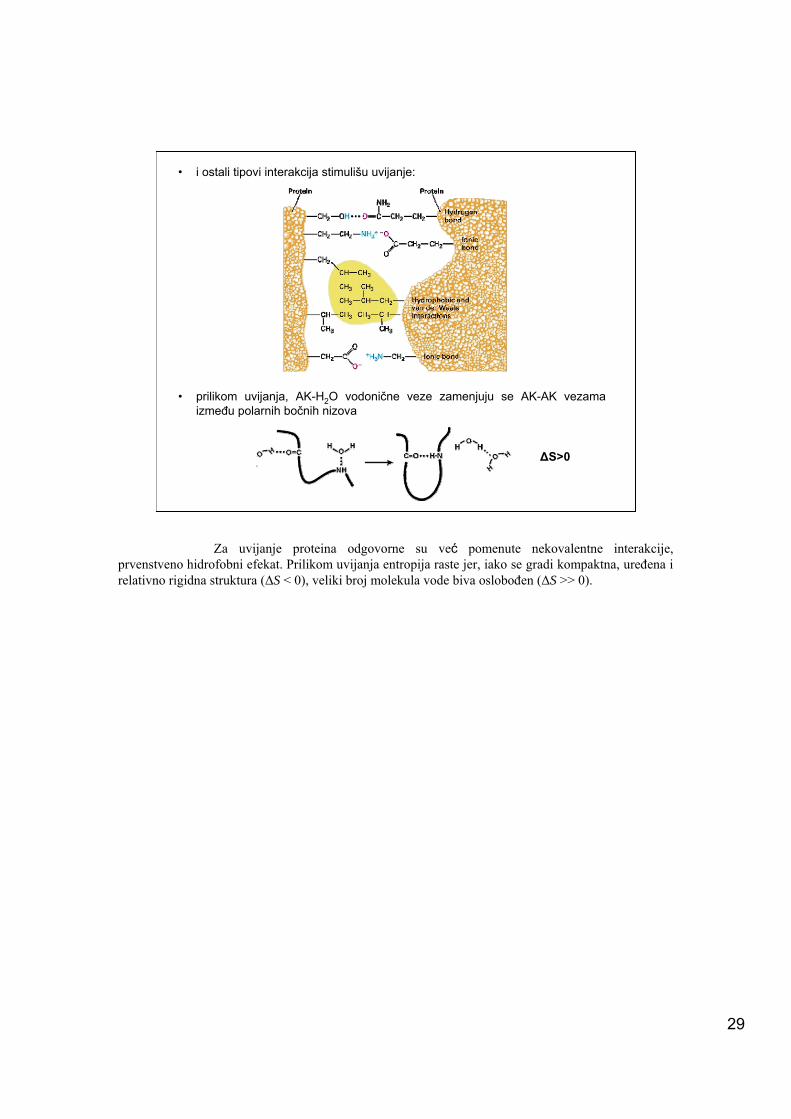
• i ostali tipovi interakcija stimulišu uvijanje:
• prilikom uvijanja, AK-H2O vodonične veze zamenjuju se AK-AK vezama između polarnih bočnih nizova
ΔS>0
Za uvijanje proteina odgovorne su već pomenute nekovalentne interakcije, prvenstveno hidrofobni efekat. Prilikom uvijanja entropija raste jer, iako se gradi kompaktna, uređena i relativno rigidna struktura (ΔS < 0), veliki broj molekula vode biva oslobođen (ΔS >> 0).
30
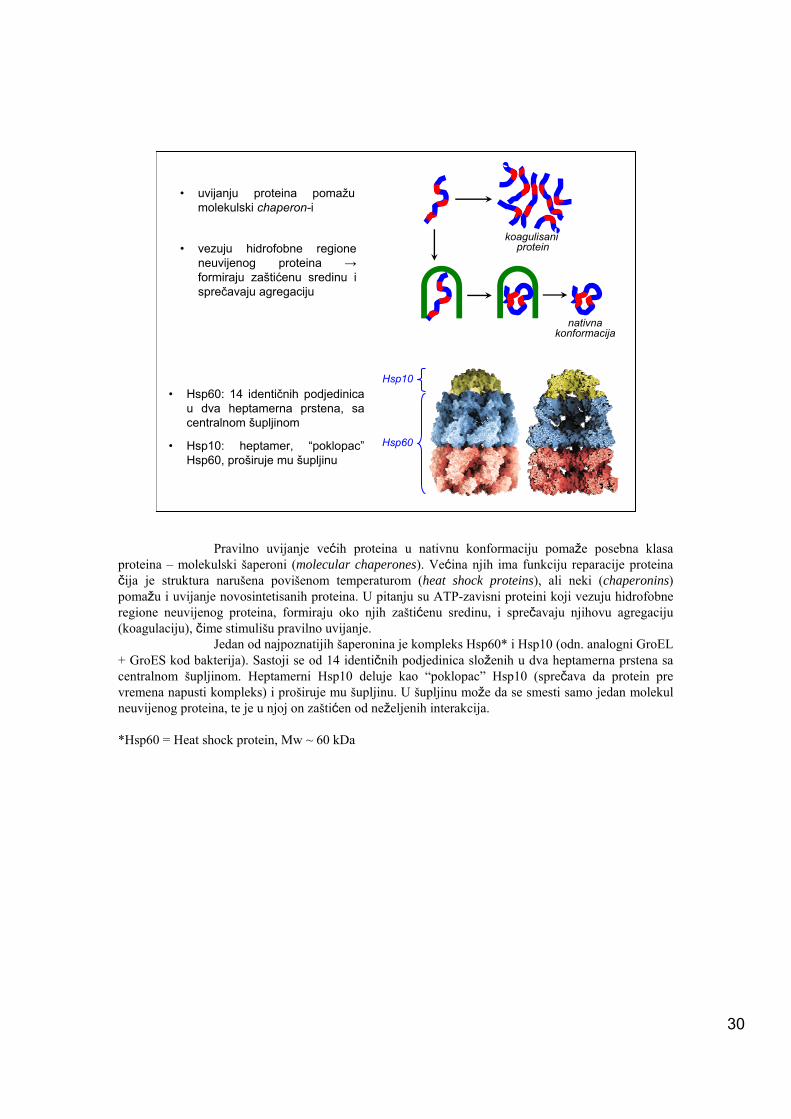
• uvijanju proteina pomažu molekulski chaperon-i
• vezuju hidrofobne regione neuvijenog proteina →formiraju zaštićenu sredinu i sprečavaju agregaciju
Hsp60
Hsp10• Hsp60: 14 identičnih podjedinica
u dva heptamerna prstena, sa centralnom šupljinom
• Hsp10: heptamer, “poklopac”Hsp60, proširuje mu šupljinu
koagulisani protein
nativnakonformacija
Pravilno uvijanje većih proteina u nativnu konformaciju pomaže posebna klasa proteina – molekulski šaperoni (molecular chaperones). Većina njih ima funkciju reparacije proteina čija je struktura narušena povišenom temperaturom (heat shock proteins), ali neki (chaperonins) pomažu i uvijanje novosintetisanih proteina. U pitanju su ATP-zavisni proteini koji vezuju hidrofobne regione neuvijenog proteina, formiraju oko njih zaštićenu sredinu, i sprečavaju njihovu agregaciju (koagulaciju), čime stimulišu pravilno uvijanje.
Jedan od najpoznatijih šaperonina je kompleks Hsp60* i Hsp10 (odn. analogni GroEL + GroES kod bakterija). Sastoji se od 14 identičnih podjedinica složenih u dva heptamerna prstena sa centralnom šupljinom. Heptamerni Hsp10 deluje kao “poklopac” Hsp10 (sprečava da protein pre vremena napusti kompleks) i proširuje mu šupljinu. U šupljinu može da se smesti samo jedan molekul neuvijenog proteina, te je u njoj on zaštićen od neželjenih interakcija.
*Hsp60 = Heat shock protein, Mw ~ 60 kDa
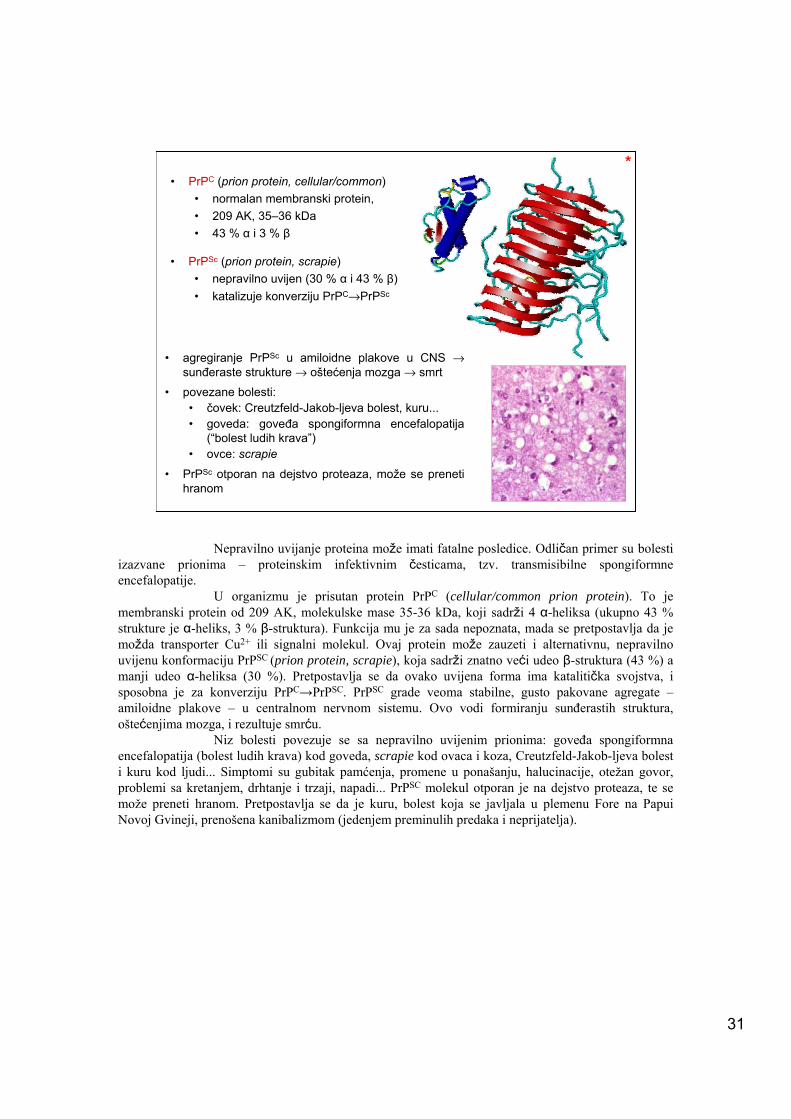
31
• PrPC (prion protein, cellular/common)• normalan membranski protein, • 209 AK, 35–36 kDa• 43 % α i 3 % β
• agregiranje PrPSc u amiloidne plakove u CNS → sunđeraste strukture → oštećenja mozga → smrt
• povezane bolesti: • čovek: Creutzfeld-Jakob-ljeva bolest, kuru...• goveda: goveđa spongiformna encefalopatija
(“bolest ludih krava”)• ovce: scrapie
• PrPSc otporan na dejstvo proteaza, može se preneti hranom
*
• PrPSc (prion protein, scrapie) • nepravilno uvijen (30 % α i 43 % β)• katalizuje konverziju PrPC→PrPSc
Nepravilno uvijanje proteina može imati fatalne posledice. Odličan primer su bolesti izazvane prionima – proteinskim infektivnim česticama, tzv. transmisibilne spongiformne encefalopatije.
U organizmu je prisutan protein PrPC (cellular/common prion protein). To je membranski protein od 209 AK, molekulske mase 35-36 kDa, koji sadrži 4 α-heliksa (ukupno 43 % strukture je α-heliks, 3 % β-struktura). Funkcija mu je za sada nepoznata, mada se pretpostavlja da je možda transporter Cu2+ ili signalni molekul. Ovaj protein može zauzeti i alternativnu, nepravilno uvijenu konformaciju PrPSC (prion protein, scrapie), koja sadrži znatno veći udeo β-struktura (43 %) a manji udeo α-heliksa (30 %). Pretpostavlja se da ovako uvijena forma ima katalitička svojstva, i sposobna je za konverziju PrPC→PrPSC. PrPSC grade veoma stabilne, gusto pakovane agregate –amiloidne plakove – u centralnom nervnom sistemu. Ovo vodi formiranju sunđerastih struktura, oštećenjima mozga, i rezultuje smrću.
Niz bolesti povezuje se sa nepravilno uvijenim prionima: goveđa spongiformna encefalopatija (bolest ludih krava) kod goveda, scrapie kod ovaca i koza, Creutzfeld-Jakob-ljeva bolest i kuru kod ljudi... Simptomi su gubitak pamćenja, promene u ponašanju, halucinacije, otežan govor, problemi sa kretanjem, drhtanje i trzaji, napadi... PrPSC molekul otporan je na dejstvo proteaza, te se može preneti hranom. Pretpostavlja se da je kuru, bolest koja se javljala u plemenu Fore na Papui Novoj Gvineji, prenošena kanibalizmom (jedenjem preminulih predaka i neprijatelja).

32
2.3.5.6. Denaturacija proteina• (intramolekulske interakcije u unutrašnjosti proteina) + (interakcija proteina
sa okolinom) → nativna konformacija proteina
• promene u okolini → promene u konformaciji → delimično / potpuno odvijanje → gubitak funkcije (denaturacija)
• reverzibilna/ireverzibilna
• uzroci:• promena pH• jake kiseline i baze• visoke i niske t• promena jonske jačine
• promena ε (rastvarača)• reagensi koji raskidaju S-S veze• reagensi koji raskidaju H-veze• deterdženti• ...
• → umrežene, nerastvorne strukture → koagulacija proteina
koagulisani protein
nativnakonformacija
denaturisaniprotein
Kao što je pomenuto, interakcije proteina sa okolinom – hidrofobni efekat, vodonične veze, elektrostatičke sile – imaju ključnu ulogu u uvijanju odn. formiranju 3D strukture proteina. Nativna konformacija posledica je ravnoteže između intramolekulskih interakcija u unutrašnjosti proteina i interakcija proteina sa okolnom sredinom. Zbog toga će promene u okolini (rastvarač, joni, drugi proteini i mali molekuli, pH itd.) uticati na strukturu proteina odn. dovoditi do promene u konformaciji. U slučaju značajnijih promena, dolazi do delimičnog ili potpunog uvijanja odn. narušavanja nativne konformacije. Pošto ovo dovodi do delimičnog ili potpunog gubitka funkcije, pojava se naziva denaturacijom. Denaturacija može biti reverzibilna ili ireverzibilna, u zavisnosti od faktora koji su izazvali denaturaciju i prirode samog proteina.
Niz faktora može dovesti do denaturacije proteina, što značajno otežava njihovu izolaciju i analitiku. Najuobičajeniji faktor je topolota – već malo povišenje temperature dovodi do narušavanja slabih nekovalentnih interakcija koje stabilizuju 3D strukturu proteina. Većina proteina pri temperaturama iznad 50–60 °C potpuno gubi funkciju, te velika većina organizama ne može preživeti visoke temperature (izuzetak su termofilne bakterije, koje zahvaljujući stabilnijim proteinima i mehanizmima reparacije preživljavaju i 100–105 °C). Prilikom denaturacije, usled razmotavanja lanaca i izlaganja hidrofobnih regiona može doći do njihove interakcije, umrežavanja i formiranja nerastvornih struktura, odn. do koagulacije.
Pored toplote, denaturaciju mogu izazvati i deterdženti, koji narušavaju hidrofobne interakcije u unutrašnjosti proteinske globule odn. omogućavaju “emulgovanje” hidrofobnih regiona u vodi.

33
• denaturacija kiselinama i bazama
• IET: Protein– + H+ → Protein0 → (Protein)n
• desolvatacija: Protein(aq) + H+ → (Protein)n + H+(aq)
• raskidanje sonih mostova:
O
O-
NH3+
Glu Lys
O
OH
NH3+
H+
• denaturacija reducensima
• raskidanje S-S mostova:
SS[H]
SH SH
Kiseline i baze su takođe uobičajeni faktori denaturacije, po bar 3 mehanizma.Prvo, proteini pri fiziološkim uslovima često nose naelektrisanje (npr. albumin je
negativno naelektrisan), te se u rastvoru odbijaju. Podešavanjem pH može se dostići izoelektrična tačka proteina, pri kojoj su proteinske čestice ukupno elektroneutralne, te se lakše mogu približiti jedna drugoj i povezivati u agregate, što dovodi do koagulacije. Zbog ovoga se izolacija proteina tipično izvodi pri njihovoj pH(I).
Drugo, u unutrašnjosti proteina formiraju se soni mostovi između suprotno naelektrisanih bočnih nizova AK, koji povezuju različite delove istog lanca ili različite lance. Promena pH dovešće do neutralizacije naelektrisanja jedne od grupa (protonovanjem -COO– ili deprotonovanjem –NH3
+), čime se ovi mostovi raskidaju.Na kraju, koncentrovane kiseline (i uopšte koncentrovane veoma polarne supstance)
deluju i putem desolvatacije. Naime, protein je okružen solvatacionom sferom, koja delimično neutrališe naelektrisane grupe i štiti hidrofobne regione. Kada se npr. koncentrovana kiselina rastvori u vodi, njeni joni (koji imaju veliku gustinu naelektrisanja) će oduzimati proteinima molekule vode da bi izgradili sopstvenu solvatacionu sferu, odn. doći će do kompeticije za molekule vode.
Kod nekih proteina, značajnu ulogu u stabilizaciji ima i kovalentno povezivanje disulfidnim mostovima, koji nastaju oksidativnim povezivanjem dva cisteinska ostatka. Neki reagensi sposobni su za redukciju disulfidnih mostova do tiolnih grupa, čime protein čine podložnijim denaturaciji. Primeri ovakvih reagenasa su 2-merkaptoetanol (HSCH2CH2OH) i ditiotreitol (HSCH2-CH(OH)-CH(OH)-CH2SH).

34
• denaturacija alkoholom
• niža ε → jače interakcije između molekula
• izlaganje hidrofobnih regiona
• raskidanje H-veza
1 22
0
q q 1Fr 4πε⋅= ⋅
C2H5OH
O
O-
C2H5OH
OHO
O-
OH
Alkoholi takođe mogu izazvati denaturaciju. Mehanizam nije tačno poznat, ali se pretpostavljaju bar 3 efekta.
Pošto su alkoholi manje polarni od vode, odn. imaju nižu dielektričnu konstantu ε, oni će manje maskirati naelektrisanje jonizovanih funkcionalnih grupa na površini proteina. Interakcije ovih grupa biće jače, što može dovesti do agregacije molekulâ proteina.
Dalje, hidrofobni regioni na površini proteina okruženi su klatratima sačinjenim od molekula vode, a koji otežavaju kontakt sa odgovarajućim regionima drugih molekula i njihovo povezivanje hidrofobnim efektom. Alkoholi, budući da su manje polarni, mogu bliže prići hidrofobnim regionima i solvatisati ih, te ne dolazi do formiranja klatrata.
Na kraju, molekuli alkohola mogu graditi vodonične veze sa peptidnim grupama i bočnim AK nizovima unutar proteinske globule, koji su inače međusobno povezani i stabilizuju 3D strukturu. Raskidanje intramolekulskih vodoničnih veza dovodi do razmotavanja proteina i denaturacije.

35
2.3.6. Kvaternerna struktura proteina• većina proteina su oligomerni – više istih ili različitih polipeptidnih lanaca (homopolimeri / heteropolimeri)• svaki lanac ima 3° strukturu, i sa susednim se povezuje nekov. interakcijama• najjednostavnija ponavljajuća jedinica – protomer• 4° struktura: broj i prostorni raspored pojedinačnih podjedinica
Kvaternerna struktura opisuje organizaciju i uređenje podjedinica unutar oligomernog proteina. Naime, većina proteina su oligomerni – sastoje se od više istih (homopolimeri) ili različitih polipeptidnih lanaca – podjedinica (heteropolimeri). Uobičajeno je da se pojedinačni tipovi podjedinica označavaju grčkim slovima. Tako se struktura hemoglobina može označiti sa α2β2. Najjednostavnija ponavljajuća jedinica oligomernog proteina naziva se protomer (u slučaju pomenutog α2β2, protomer je αβ). Broj podjedinica može biti od 2, pa i do mnogo hiljada (kod strukturnih polimernih proteina). Podjedinice uvek imaju dobro definisanu stehiometriju i prostorno uređenje. Svaka podjedinica (sa prethodno formiranom 2° i 3° strukturom) orijentiše se prema ostalim i sa njima povezuje, najčešće slabim, nekovalentnim interakcijama (hidrofobnim, mada mogu učestvovati i elektrostatičke).
U nekim slučajevima, oligomerni proteini mogu imati veoma kompleksne strukture i sastojati se od više desetina različitih podjedinica sa definisanim funkcijama (npr. ATP-sintaza, bakterijske flagele...).

36
4. genetska ekonomičnost
5. multienzimski kompleksi
• prednosti:
2. formiranje aktivnog/alosternog centra
3. kooperativnost, grupna regulacija
1. stabilnost, mehanička svojstva
T TT T
T TT T
O2
T TT
O2
RR RR R
O2
Zašto se proteini veoma često javljaju u vidu oligomera? Pretpostavlja se nekoliko mogućih razloga.
Prvo, oligomeri su stabilniji od pojedinačnih podjedinica, odn. kvaternerna struktura može da produži vreme života proteina.
Dalje, kod nekih enzima aktivni centar ili mesto za vezivanje alosternog efektora formira se između podjedinica, odn. čine ga AK ostaci iz različitih lanaca.
Kod oligomera se može javiti i pojava kooperativnosti – promena konformacije i aktivnosti usled vezivanja liganda na jednoj podjedinici dovodi do jednakog odgovora i na ostalim podjedinicama, što omogućava grupnu regulaciju aktivnosti.
Formiranje oligomera obezbeđuje i genetsku ekonomičnost – više različitih proteina može koristiti istu podjedinicu sa definisanom funkcijom (npr. kalmodulin omogućava vezivanje Ca2+, koji služi kao glasnik u ćeliji), zbog čega je dovoljno jednom je kodirati u DNK.
Kod multienzimskih kompleksa, povezano je više enzima koji katalizuju niz uzastopnih reakcija; svaki enzim prosleđuje svoj produkt sledećem. Na ovaj način, povećava se brzina reakcije, a smanjuje se i verovatnoća sporednih reakcija.

37
2.3.7. Globularni i fibrilarni proteini
• prema globalnoj 3D strukturi (3° i 4°) i rastvorljivosti proteini se dele na:
1. fibrilarne proteine (skleroproteine) - izdužene, vlaknaste strukture; nerastvorni u vodi; strukturna, motorna, zaštitna f-ja
2. globularne proteine (sferoproteine) – pribl. sferični, rastvorni u vodi ili rastvorima soli; različite f-je
Proteini se prema globalnoj 3D strukturi – tercijernoj i kvaternernoj – mogu podeliti u dve velike grupe.
Fibrilarni (fibrozni) proteini ili skleroproteini imaju izdužene, vlaknaste strukture. Nerastvorni su u vodi.
Globularni proteini (sferoproteini) imaju približno sferne strukture. Manje ili više su rastvorni u vodi i/ili vodenim rastvorima soli. Funkcionalno su mnogo raznovrsniji, i uključuju enzime, signalne molekule, receptore, transportere, depoe AK...

38
2.3.7.1. Fibrilarni proteini
• primeri: kolagen, keratin, elastin, fibroin, aktin, miozin, tubulin
• veoma izduženi molekuli (štapići/vlakna)
• helikoidalne strukture → uvijanje (kolagen...)
• globularne podjedinice → spiralno slaganje (tubulin)
• sekvenca: ponavljajuće podjedinice
• hidrofobni bočni nizovi → agregacija → nerastvorni u vodi
• često: umreženi (S-S mostovi između lanaca...)
• ponekad: neuobičajene 2° strukture (kolagenski heliks...)
• strukturni elementi ćelija i tkiva (citoskelet, koža, kosti, kosa, rožnate strukture), ponekad zaštitna (fibrin), skladišna f-ja
Kao što je pomenuto, fibrilarne proteine karakterišu veoma izduženi – štapićasti ili vlaknasti – molekuli. Struktura se može formirati na dva osnovna načina. Kod jedne grupe (u koju spadaju kolagen, α-keratin, miozin...), dominantne su helikoidalne strukture (α-heliksi i dr.), koje su po prirodi izdužene. Tipično se više pojedinačnih heliksa uvija jedan oko drugog, gradeći superheliks. Proteini druge grupe (u koju spadaju tubulin, aktin...) sastavljeni su od globularnih podjedinica, koje se spiralno slažu.
Fibrilarni proteini tipično imaju jednostavnu sekvencu, koja se sastoji ponavljajućih jedinica – npr. kod kolagena (Gly-Xaa-Pro/Hyp)n. Zahvaljujući visokom udelu hidrofobnih aminokiselina, čiji su bočni nizovi usmereni ka spoljašnjoj sredini, skloni su agregaciji što ih čini nerastvornima u vodi. Dodatno, mnogi od njih grade umrežene strukture nastale povezivanjem različitih lanaca kovalentnim vezama (npr. -S-S-), što im povećava stabilnost (teže se denaturišu od globularnih). Kod ove grupe proteina mogu se javiti i neke neuobičajene sekundarne strukture, kao što je poliprolinski heliks.
Tipično imaju strukturnu funkciju – izgrađuju citoskelet (aktin, tubulin...), kožu, rožnate strukture, kosu (keratin), kosti (kolagen), vezivna tkiva... Dodatno, učestvuju u mišićnim kontrakcijama (aktin, miozin), a mogu imati i zaštitnu funkciju (fibrin, koji učestvuje u koagulaciji krvi) i skladišnu funkciju.

39
• Keratini
• u višim kičmenjacima• u epidermalnom tkivu (gde čine 85 % celularnih proteina), u rožnatim
tvorevinama (kandže, kopita, dlake, perje, rogovi, krljušt gmizavaca, oklop kornjača, kljun...)
• kod sisara – α-keratini, kod ptica i gmizavaca – β-keratini
• α-Keratini
• osnovna komponenta epidermisa i rožnatih struktura sisara• helikoidalna struktura slična α-heliksu → superheliksi• ponavljajuća heptada (a-b-c-d-e-f-g)
• bogati nepolarnim AK: Ala, Val, Leu, Ile, Met , Phe• pozicija a i d – nepolarne AK → nepolarna traka duž jedne ivice
Prva grupa fibrilarnih proteina koja će biti pomenuta su keratini. Ovi proteini zastupljeni su u epidermisu viših kičmenjaka (gde čine 85 % celularnih proteina) i rožnatim tvorevinama (nokti, kandže, kopita, dlake, perje, rogovi, krljušt gmizavaca, oklop kornjača, kljun...). Poznato je preko 30 proteina iz ove grupe. Kod sisara prisutni su α-keratini, a kod ptica i gmizavaca –β-keratini.
α-keratini izgrađuju epidermis i rožnate strukture kod sisara (nokte, kandže, rogove, kopita, kosu i vunu). Molekule α-keratina karakterišu nehelikoidalni N- i C-terminus i helikoidalni centralni deo lanca (~300+ AK), čija se struktura neznatno razlikuje od uobičajenih α-heliksa (sa 3,5 AK po namotaju, umesto 3,6). U sekvenci se javlja ponavljajuća heptapeptidna jedinica, u kojoj pozicije 1 i 4 zauzimaju nepolarne AK. Posledica je da se duž jedne ivice heliksa formira nepolarna traka, koja stimuliše agregaciju lanaca u vrlo stabilne superhelikse.
40
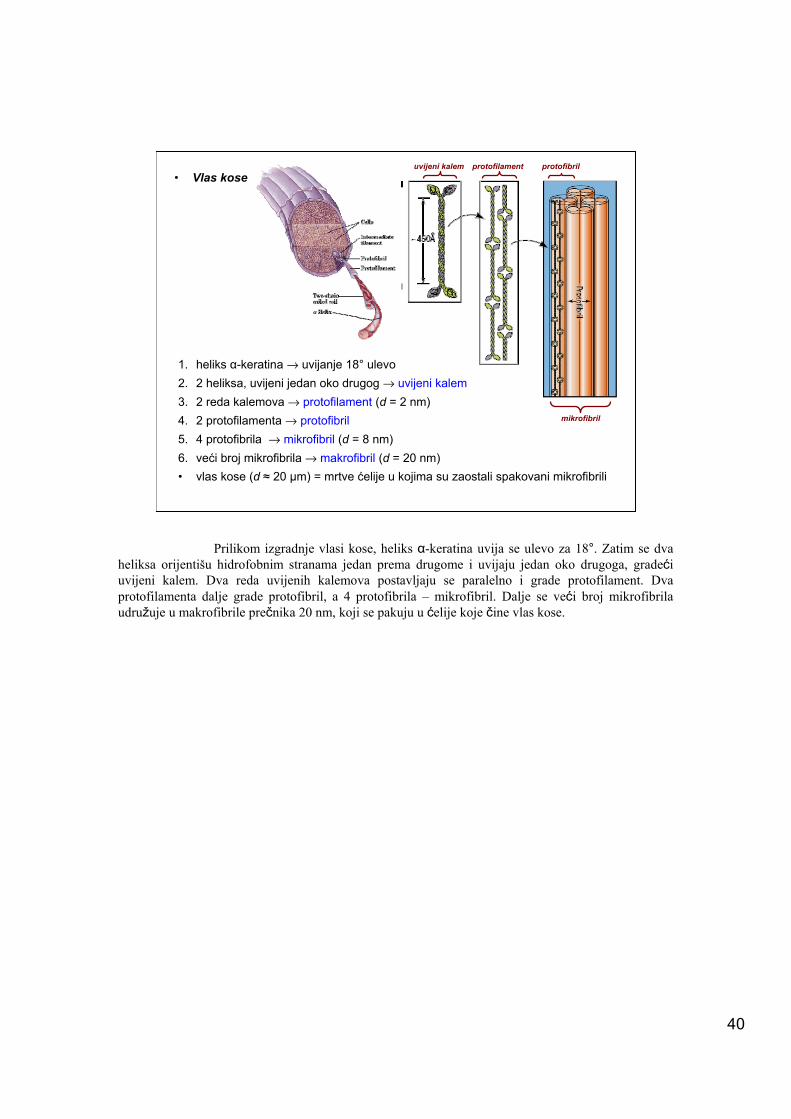
1. heliks α-keratina → uvijanje 18° ulevo2. 2 heliksa, uvijeni jedan oko drugog → uvijeni kalem3. 2 reda kalemova → protofilament (d = 2 nm)4. 2 protofilamenta → protofibril5. 4 protofibrila → mikrofibril (d = 8 nm)6. veći broj mikrofibrila → makrofibril (d = 20 nm)• vlas kose (d ≈ 20 μm) = mrtve ćelije u kojima su zaostali spakovani mikrofibrili
• Vlas koseprotofilament protofibril
mikrofibril
uvijeni kalem
Prilikom izgradnje vlasi kose, heliks α-keratina uvija se ulevo za 18°. Zatim se dva heliksa orijentišu hidrofobnim stranama jedan prema drugome i uvijaju jedan oko drugoga, gradeći uvijeni kalem. Dva reda uvijenih kalemova postavljaju se paralelno i grade protofilament. Dva protofilamenta dalje grade protofibril, a 4 protofibrila – mikrofibril. Dalje se veći broj mikrofibrila udružuje u makrofibrile prečnika 20 nm, koji se pakuju u ćelije koje čine vlas kose.

41
• α-keratin je bogat cistinom → interlančani S-S mostovi → povećana čvrstoća
• prema sadržaju Cys – meki keratini (dlake...) i tvrdi (bodlje bodljikavog praseta, rogovi...)
• S-S mostovi, istegljivi α-heliksi → elastičnost, kovrdžavost kose
trajna ondulacija:1. namotavanje kose oko viklera2. reducens (HSCH2COO–NH4
+) → raskidanje S-S mostova3. zagrevanje → (α-heliksi→β-nabrana) → izduživanje oko viklera4. oksidans (H2O2) → uspostavljanje S-S mostova na novim mestima5. pranje i hlađenje → obnavljanje helikoidalne strukture
Pored navedenih nepolarnih AK, α-keratin je bogat i cisteinom (odn. njegovim dimerom, cistinom). Disulfidni mostovi formirani između različitih lanaca α-keratina formiraju umreženu strukturu, čime se povećava čvrstoća vlakna. Prema sadržaju cisteina, keratini se mogu podeliti na meke, koji ulaze u sastav dlake, i sadrže mali broj S-S mostova, i tvrde, koji grade npr. rogove, koji su više umreženi i stoga čvršći.
Kosa je elastična (može se izdužiti pri primeni sile, i ponovo vratiti u prvobitni oblik kada se sila ukloni) zahvaljujući istegljivosti α-heliksa i prisustvu S-S mostova.
Na činjenici da se disulfidni mostovi reverzibilno mogu raskinuti reducensima zasniva se trajna ondulacija kose (mini-val). Kod ovog postupka, kosa se nakon namotavanja oko viklera tretira reducensom (amonijum-tioglikolat, HSCH2COO–NH4
+), pri čemu dolazi do raskidanja disulfidnih mostova koji povezuju vlakna. Zagrevanjem se vlažna kosa prevodi iz α-heliksa u β-nabranu strukturu, pri čemu dolazi do istezanja oko viklera. Primenom oksidansa (H2O2) ponovo se uspostavljaju disulfidni mostovi, ali koji sada fiksiraju kosu u uvijenoj formi. Na kraju, pranjem i hlađenjem obnavlja se helikoidalna struktura α-keratina.

42
• Svila• insekti (svilena buba), paukovi
• fibroin (strukturni centar) + sericin(lepljiva materija)
• fibroin: ponavljajuća sekvenca (Gly-Ser-Gly-Ala-Gly-Ala)n
• slojevi antiparalelnih β-pločica
• bočni ostaci Gly na jednoj strani pločice, ostaci Ala (CH3) na drugoj → CH3-grupe uklještene →mehanička otpornost, rezistencija na istezanje
Drugi fibrilarni protein koji će biti pomenut je fibroin svile, koji je donekle sličan β-keratinu. Svila je inače prirodni vlaknasti materijal koga luče različiti zglavkari. Insekti svilu uglavnom koriste za formiranje čaure u kojoj larva podleže metamorfozi (svila larve svilene bube, Bombyx mori, koristi se i komercijalno), dok paukovi od nje izgrađuju mreže.
Svila svilene bube sastoji se od dva proteina – fibroina, koji je strukturna osnova, i sericina, koji predstavlja lepljivu komponentu.
Primarnu strukturu fibroina pretežno čine ponavljajuće heksapeptidne jedinice GSGAGA. Sekundarnom strukturom dominiraju antiparalelne β-nabrane pločice, koje se slažu u slojeve, pri čemu se formira mikrokristalna struktura. Bočni ostaci se nalaze naizmenično iznad i ispod ravni pločica. Unutar jedne β-trake, svi bočni ostaci Gly (H) su sa jedne strane, a bočni ostaci Ala i Ser (CH3 i CH2OH) sa druge strane. Relativno mali bočni nizovi omogućavaju tesno pakovanje β-pločica, što zajedno sa uklještenjem CH3-grupa rezultuje velikom mehaničkom otpornošću i rezistencijom na istezanje.

43
• u pojedinim delovima fibroina: Asp, Arg, Tyr, Val → voluminozne bočne grupe → narušavanje 2° strukture → amorfni regioni
• mikrokristalne, β-strukture daju čvrstoću, a amorfne – istegljivost
U pojedinim delovima fibroinskog lanca javljaju se segmenti sa voluminoznim AK: Asp, Arg, Tyr, Val. Ove AK ne uklapaju se u slojevitu β-nabranu strukturu, te dolazi do formiranja amorfnih regiona. Ovi regioni daju svili izvesnu istegljivost.

44
• Kolagen
• glavni sastojak vezivnog tkiva – u kostima, koži, rožnjači, hrskavici, ligamentima, krvnim sudovima
• ponavljajuća sekvenca: (Gly-Xaa-Pro/Hyp)n
• Pro i Hyp → sprečeno formiranje α-heliksa i β-niza
• formira se levo uvijena strma spirala (heliks tipa poliprolina II) – ”α-niz”
• 3 spirale → blago uvijanje (10 AK po navoju) → u desni trostruki heliks (superheliks), stabilizovan interlančanim H-vezama
• Gly (mali bočni niz) omogućava formiranje kompaktne spirale
• ostaci Pro - spoljaGly
Poslednji fibrilarni protein o kome će biti reči je kolagen. Kolagen je glavni sastojak vezivnog tkiva kičmenjaka, i kod sisara čini ~30 od ukupnih proteina. Nalazi se u kostima, zubima, koži, rožnjači oka, hrskavici, ligamentima, krvnim sudovima, crevima itd.
Primarnu strukturu čini ponavljajuća tripeptidna jedinica (Gly-Xaa-Pro)n, u kojoj je Pro često modifikovan (oksidovan do hidroksiprolina). Pro i Hyp zbog svoje krutosti (neodgovarajući φi ψ) sprečavaju formiranje α-heliksa i β-niza. Umesto toga, nastaje levo uvijena strma spirala – heliks tipa poliprolina I, unutar koje peptidne grupe nisu vodonično vezane. 3 ovakva α-niza uvijaju se blago (10 AK po navoju) u desni trostruki heliks – superheliks, koji je stabilizovan interlančanim H-vezama. Ostaci glicina locirani su duž jedne ivice heliksâ (poravnati pošto lanac sadrži 3 AK po navoju), i zahvaljujući malom bočnom nizu omogućavaju tesno pakovanje heliksâ i formiranje kompaktnog superheliksa.

45
• formiranje:
1. na ribozomima: sinteza pro-α-nizova (~1000 AK)
2. posttranslacione modifikacije: Lys→Hyl, Pro→Hyp, glikozilacija → α-nizovi
3. blago uvijanje 3 α-niza u prokolagen
4. otkidanje 180 AK-ostatka sa N-terminusa i 300 AK-ostatka sa C-terminusa → tropokolagen
5. lanci tropokolagena → kovalentno poprečno povezivanje (unutar i između jedinica tropokolagena) → mikrofibrili
NH2 NH2
NH2 O
N
Lizin
Alizin
lizil oksidaza
Inače, proteinski lanci koji izgrađuju kolagen sintetišu se translacijom, pri čemu nastaju pro-α-nizovi od ~1000 AK. U sledećoj fazi, dolazi do posttranslacionih modifikacija –hidroksilacije Pro i Lys do Hyp i Hyl, i glikozilacije Hyl (svrha glikozilacije još uvek je nepoznata), čime se stvaraju α-nizovi. Prolin i hidroksiprolin ključni su za termičku stabilnost kolagena. Za reakciju hidroksilacije Pro i Lys neophodan je vitamin C (askorbinska kiselina). U slučaju nedostatka vitamina C, Hyp i Hyl ne nastaju, kolagen je manje stabilan, i javlja se obolenje skorbut, koje se manifestuje usporenim rastom kostiju, sporim zarastanjem rana, fragilnim kapilarima...
Po 3 α-niza dalje se blago uvijaju u superheliks prokolagena, koji se uekstracelularnom prostoru otkidanjem N- i C-terminalnih sekvenci konvertuje u tropokolagen.
Lanci tropokolagena se zatim asociraju u mikrofibrile uz kovalentno poprečno povezivanje (unutar tropokolagenskih jedinica, i između njih). Kovalentno umrežavanje omogućava enzim lizil oksidaza, koja jedan od lizinskih ostataka oksiduje do alizina. ε-Amino-grupa lizina i aldehidna grupa alizina zatim se povezuju u jedinjenje tipa imina (Šifove baze). Alternativno, dva alizinska ostatka mogu stupiti u reakciju tipa aldolne kondenzacije. Ova reakcija značajno doprinosi čvrstoći kolagena. Jedinjenje β-aminopropionitril (NH2CH2CH2C≡N, BAPN) iz mahunarke Lathyrus sativus inhibira lizil oksidazu, što sprečava umrežavanje kolagena, zbog čega on postaje nestabilan, što rezultuje osteolatirizmom, koji se manifestuje slabošću i krhkošću vezivnih tkiva (u koži, kostima, krvnim sudovima). S druge strane, formiranje prevelikog broja mostova (dešava se u starosti) rezultuje tvrdim, neelastičnim tetivama i neelastičnim, lomljivim kostima.
46
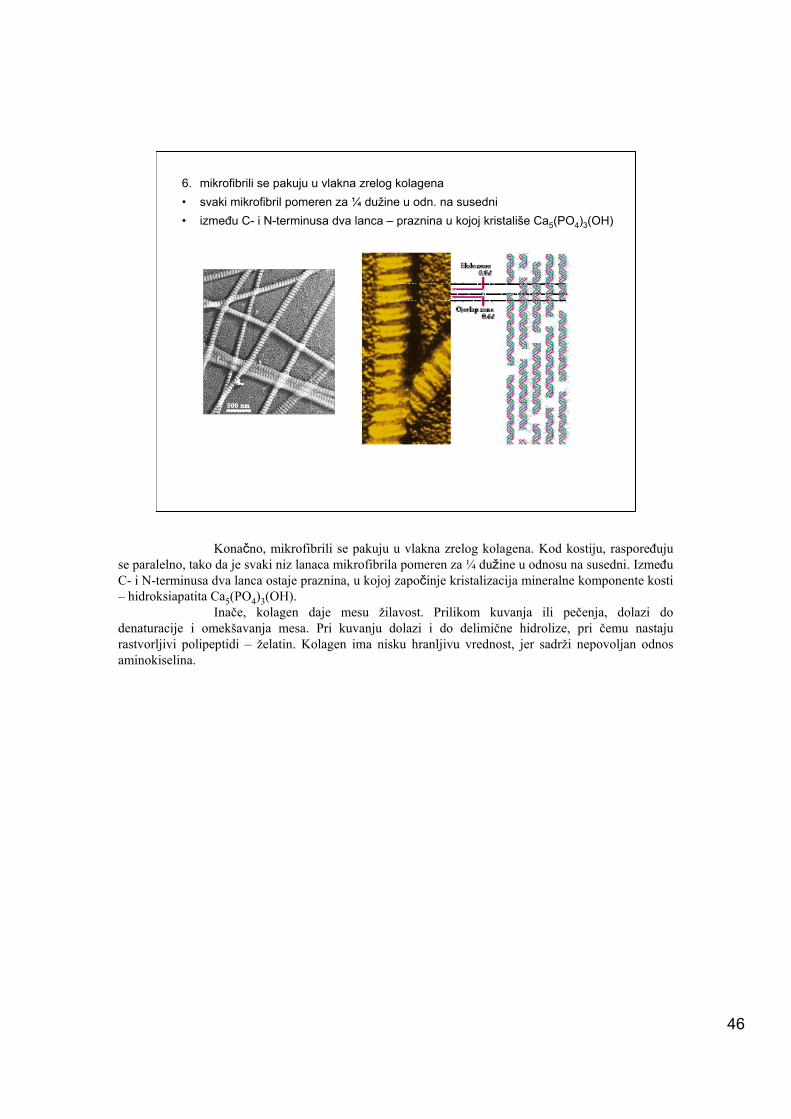
6. mikrofibrili se pakuju u vlakna zrelog kolagena• svaki mikrofibril pomeren za ¼ dužine u odn. na susedni• između C- i N-terminusa dva lanca – praznina u kojoj kristališe Ca5(PO4)3(OH)
Konačno, mikrofibrili se pakuju u vlakna zrelog kolagena. Kod kostiju, raspoređuju se paralelno, tako da je svaki niz lanaca mikrofibrila pomeren za ¼ dužine u odnosu na susedni. Između C- i N-terminusa dva lanca ostaje praznina, u kojoj započinje kristalizacija mineralne komponente kosti – hidroksiapatita Ca5(PO4)3(OH).
Inače, kolagen daje mesu žilavost. Prilikom kuvanja ili pečenja, dolazi do denaturacije i omekšavanja mesa. Pri kuvanju dolazi i do delimične hidrolize, pri čemu nastaju rastvorljivi polipeptidi – želatin. Kolagen ima nisku hranljivu vrednost, jer sadrži nepovoljan odnos aminokiselina.

47
2.3.7.2. Globularni proteini
• Mioglobin (Mb)• prvi protein kome je određena 3° struktura (1950. Kendrew et al.)• 1 pp lanac, 153 AK, Mr = 16,7 kDa• 8 α-heliksa (~75 % pp lanca), 7 neuređenih delova • prostetska grupa – hem – u nepolarnom džepu
Globularni proteini strukturno su i funkcionalno veoma raznovrsni – predstavljaju enzime, transportere, glasnike, receptore, skladišne proteine itd. Ovde će biti opisana struktura i funkcija dva srodna proteina – hemoglobina i mioglobina. Hemoglobin i mioglobin su hemoproteini –pored proteinske komponente (iz grupe globina), oni sadrže i prostetsku grupu – hem. Oba proteina imaju ulogu u transportu O2.
Mioglobin je prvi protein kome je određena tercijerna struktura. To je monomerni protein sačinjen od 153 AK, molekulske mase 16,7 kDa. ~75 % peptidnog lanca je u α-helikoidalnoj konformaciji. Pored 8 α-heliksâ, u sekundarnoj strukturi javlja se i 7 neuređenih segmenata. U unutrašnjosti molekula dominiraju bočni nizovi hidrofobnih AK (Val, Leu, Ile, Phe, Met). U hidrofobnom žljebu koji se formira između heliksâ smešta se prostetska grupa – hem.

48
• hem = protoporfirin IX sa centralno vezanim Fe2+
• O2 ne može da se vezuje za AK; lako se veže za Fe i Cu jone
• Fe2+: 2 koordinativna mesta - jednim se vezuje za globin (His93), a drugo vezuje O2
• pri vezivanju O2 Fe(II) se ne oksiduje (nepolarni džep ga štiti)
• Hb / Mb sa Fe3+ (metMb / metHb) ne može da veže O2; enzim metHb reduktaza mora da redukuje metHb do Hb
• za 6. koordinativno mesto Fe mogu se vezati i CO, NO i H2S, sa većim afinitetom od O2
• oba reverzibilno vezuju O2, ali se afiniteti i f-je razlikuju
FeN N
NN
O O-
O O-
Hem predstavlja kompleks tetrapirolnog organskog jedinjenja – protoporfirina IX –sa Fe2+ jonom. Protoporfirinska komponenta vezana je za globin samo nekovalentnim interakcijama –hidrofobnim, van der Waals-ovim, vodoničnim vezama.
Hem je funkcionalni deo hemoglobina i mioglobina, budući da O2 ne može da se veže ni za jednu AK, ali se lako reverzibilno vezuje za metalne jone kao što su Fe2+ i Cu2+. Fe2+ hema ima slobodna 2 koordinativna mesta. Jednim se vezuje za globin, preko proksimalnog histidina (His F8, His93), a drugo služi za vezivanje O2. Prilikom vezivanja O2, Fe(II) se ne oksiduje jer ga nepolarni džep štiti (slobodni hem, bez proteina, odmah bi se oksidovao). Ukoliko do oksidacije dođe (Fe2+ pređe u Fe3+), nastala forma (methemoglobin/metmioglobin) ne može da veže O2 i mora se enzimatski redukovati da bio ponovo vršila svoju ulogu. Inače, pored O2, za 6. koordinativno mesto Fe2+ mogu se vezati i CO, NO i H2S. Pritom, njihov afinitet prema hemu je veći nego kod O2, te oni blokiraju njegovu funkciju.
49
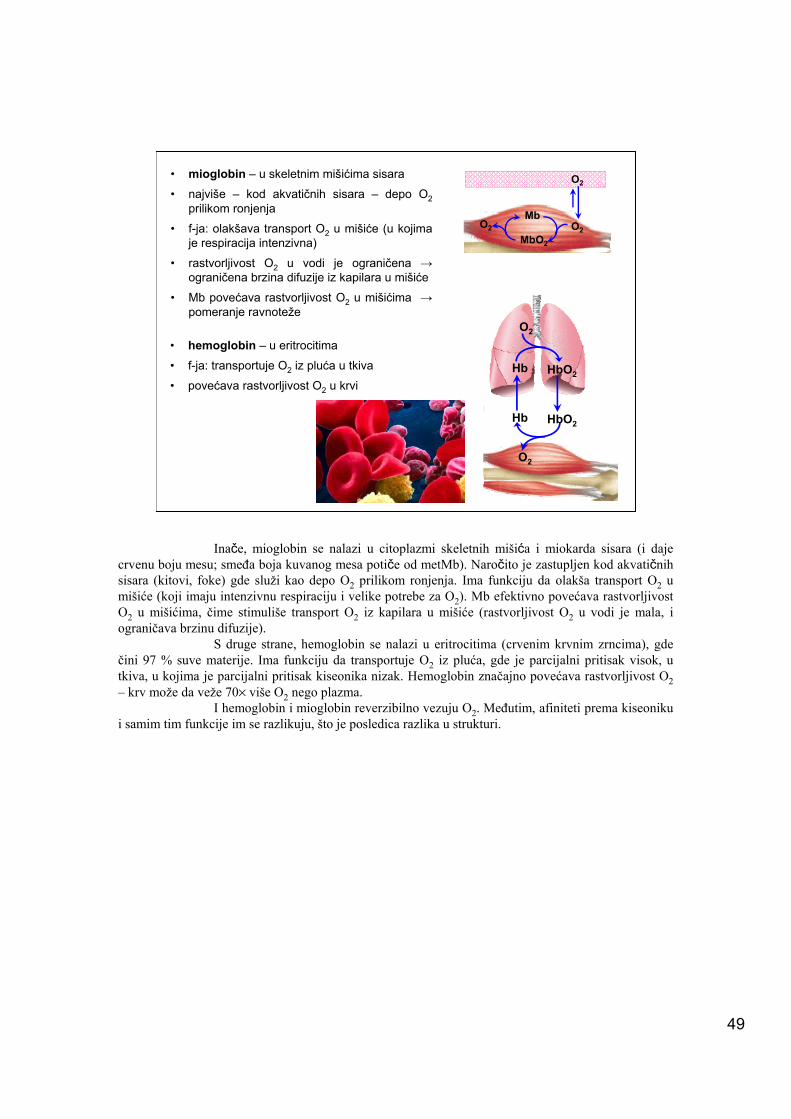
• mioglobin – u skeletnim mišićima sisara
• najviše – kod akvatičnih sisara – depo O2prilikom ronjenja
• f-ja: olakšava transport O2 u mišiće (u kojima je respiracija intenzivna)
• rastvorljivost O2 u vodi je ograničena →ograničena brzina difuzije iz kapilara u mišiće
• Mb povećava rastvorljivost O2 u mišićima →pomeranje ravnoteže
Mb
MbO2
O2 O2
O2
• hemoglobin – u eritrocitima
• f-ja: transportuje O2 iz pluća u tkiva
• povećava rastvorljivost O2 u krviHb HbO2
Hb HbO2
O2
O2
Inače, mioglobin se nalazi u citoplazmi skeletnih mišića i miokarda sisara (i daje crvenu boju mesu; smeđa boja kuvanog mesa potiče od metMb). Naročito je zastupljen kod akvatičnih sisara (kitovi, foke) gde služi kao depo O2 prilikom ronjenja. Ima funkciju da olakša transport O2 u mišiće (koji imaju intenzivnu respiraciju i velike potrebe za O2). Mb efektivno povećava rastvorljivost O2 u mišićima, čime stimuliše transport O2 iz kapilara u mišiće (rastvorljivost O2 u vodi je mala, i ograničava brzinu difuzije).
S druge strane, hemoglobin se nalazi u eritrocitima (crvenim krvnim zrncima), gde čini 97 % suve materije. Ima funkciju da transportuje O2 iz pluća, gde je parcijalni pritisak visok, u tkiva, u kojima je parcijalni pritisak kiseonika nizak. Hemoglobin značajno povećava rastvorljivost O2– krv može da veže 70× više O2 nego plazma.
I hemoglobin i mioglobin reverzibilno vezuju O2. Međutim, afiniteti prema kiseonikui samim tim funkcije im se razlikuju, što je posledica razlika u strukturi.

50
Mioglobin• monomer• 153 AK
Hemoglobin• tetramer• 2×141 + 2×146 AK
Već je pomenuto da je mioglobin monomerni protein.Hemoglobin je kompleksniji od mioglobina, jer predstavlja oligomerni protein.
Sastoji se od 4 globinske podjedinice – 2 α-globina (141 AK) i 2 β-globina (146 AK), tj. α2β2. Obe globinske jedinice slične su po sekvenci i tercijernoj strukturi mioglobinu. Hemoglobin je prvi protein kome je određena kvaternerna struktura (Perutz 1959).
51
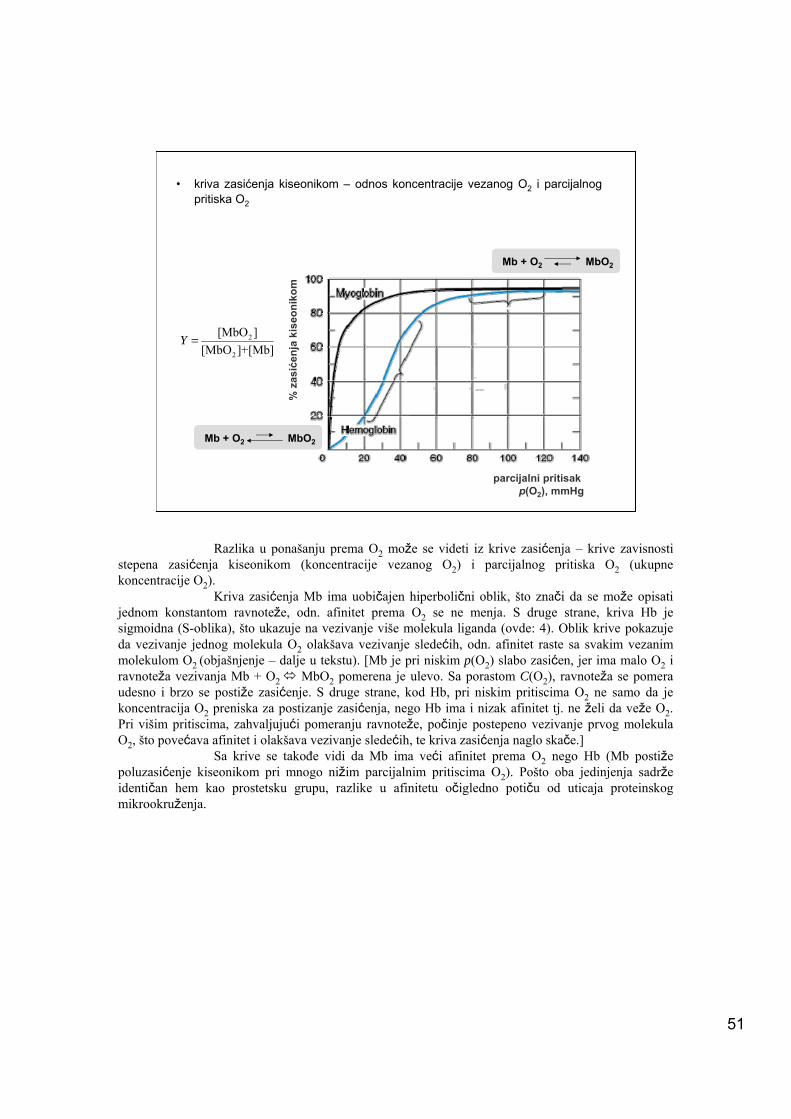
• kriva zasićenja kiseonikom – odnos koncentracije vezanog O2 i parcijalnog pritiska O2
parcijalni pritisak p(O2), mmHg
% z
asić
enja
kis
eoni
kom
2
2
[MbO ][MbO ]+[Mb]
Y =
Mb + O2 MbO2
Mb + O2 MbO2
Razlika u ponašanju prema O2 može se videti iz krive zasićenja – krive zavisnosti stepena zasićenja kiseonikom (koncentracije vezanog O2) i parcijalnog pritiska O2 (ukupne koncentracije O2).
Kriva zasićenja Mb ima uobičajen hiperbolični oblik, što znači da se može opisati jednom konstantom ravnoteže, odn. afinitet prema O2 se ne menja. S druge strane, kriva Hb je sigmoidna (S-oblika), što ukazuje na vezivanje više molekula liganda (ovde: 4). Oblik krive pokazuje da vezivanje jednog molekula O2 olakšava vezivanje sledećih, odn. afinitet raste sa svakim vezanim molekulom O2 (objašnjenje – dalje u tekstu). [Mb je pri niskim p(O2) slabo zasićen, jer ima malo O2 i ravnoteža vezivanja Mb + O2 MbO2 pomerena je ulevo. Sa porastom C(O2), ravnoteža se pomera udesno i brzo se postiže zasićenje. S druge strane, kod Hb, pri niskim pritiscima O2 ne samo da je koncentracija O2 preniska za postizanje zasićenja, nego Hb ima i nizak afinitet tj. ne želi da veže O2. Pri višim pritiscima, zahvaljujući pomeranju ravnoteže, počinje postepeno vezivanje prvog molekula O2, što povećava afinitet i olakšava vezivanje sledećih, te kriva zasićenja naglo skače.]
Sa krive se takođe vidi da Mb ima veći afinitet prema O2 nego Hb (Mb postiže poluzasićenje kiseonikom pri mnogo nižim parcijalnim pritiscima O2). Pošto oba jedinjenja sadrže identičan hem kao prostetsku grupu, razlike u afinitetu očigledno potiču od uticaja proteinskog mikrookruženja.
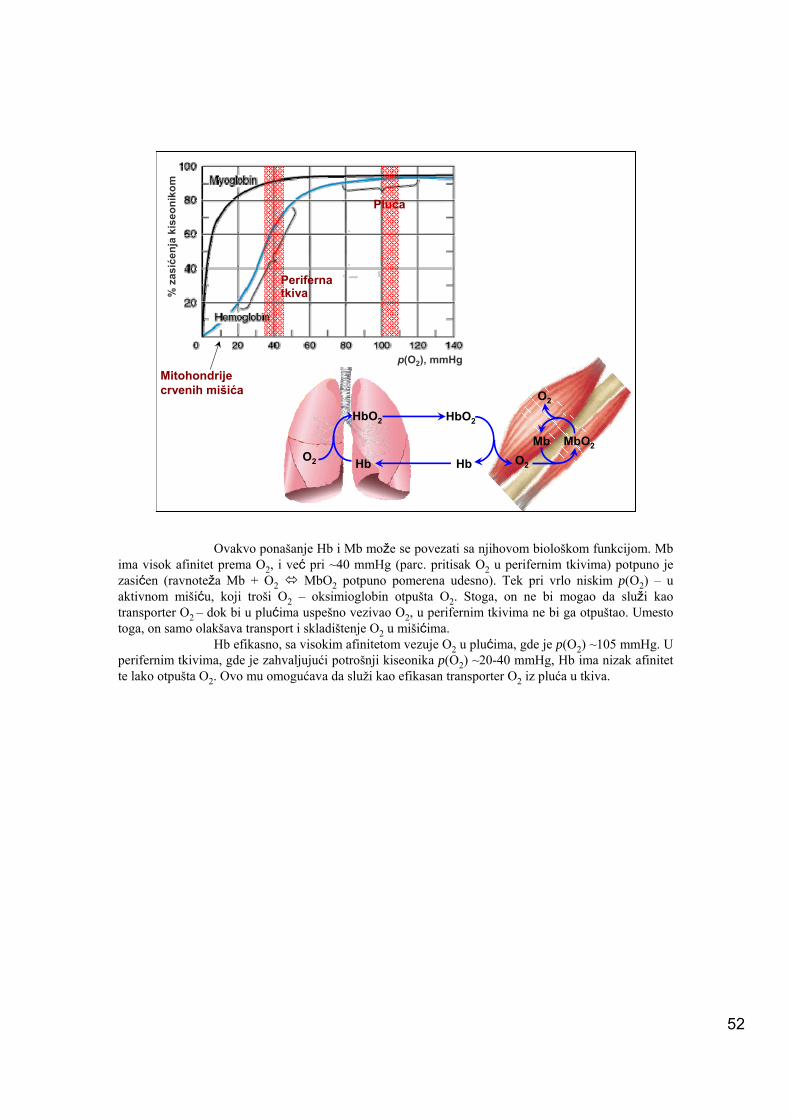
52
Hb
HbO2
Hb
HbO2
O2 O2
MbO2Mb
O2
Mitohondrije crvenih mišića
Perifernatkiva
Pluća
p(O2), mmHg
% z
asić
enja
kis
eoni
kom
Ovakvo ponašanje Hb i Mb može se povezati sa njihovom biološkom funkcijom. Mb ima visok afinitet prema O2, i već pri ~40 mmHg (parc. pritisak O2 u perifernim tkivima) potpuno je zasićen (ravnoteža Mb + O2 MbO2 potpuno pomerena udesno). Tek pri vrlo niskim p(O2) – u aktivnom mišiću, koji troši O2 – oksimioglobin otpušta O2. Stoga, on ne bi mogao da služi kao transporter O2 – dok bi u plućima uspešno vezivao O2, u perifernim tkivima ne bi ga otpuštao. Umesto toga, on samo olakšava transport i skladištenje O2 u mišićima.
Hb efikasno, sa visokim afinitetom vezuje O2 u plućima, gde je p(O2) ~105 mmHg. U perifernim tkivima, gde je zahvaljujući potrošnji kiseonika p(O2) ~20-40 mmHg, Hb ima nizak afinitet te lako otpušta O2. Ovo mu omogućava da služi kao efikasan transporter O2 iz pluća u tkiva.
53
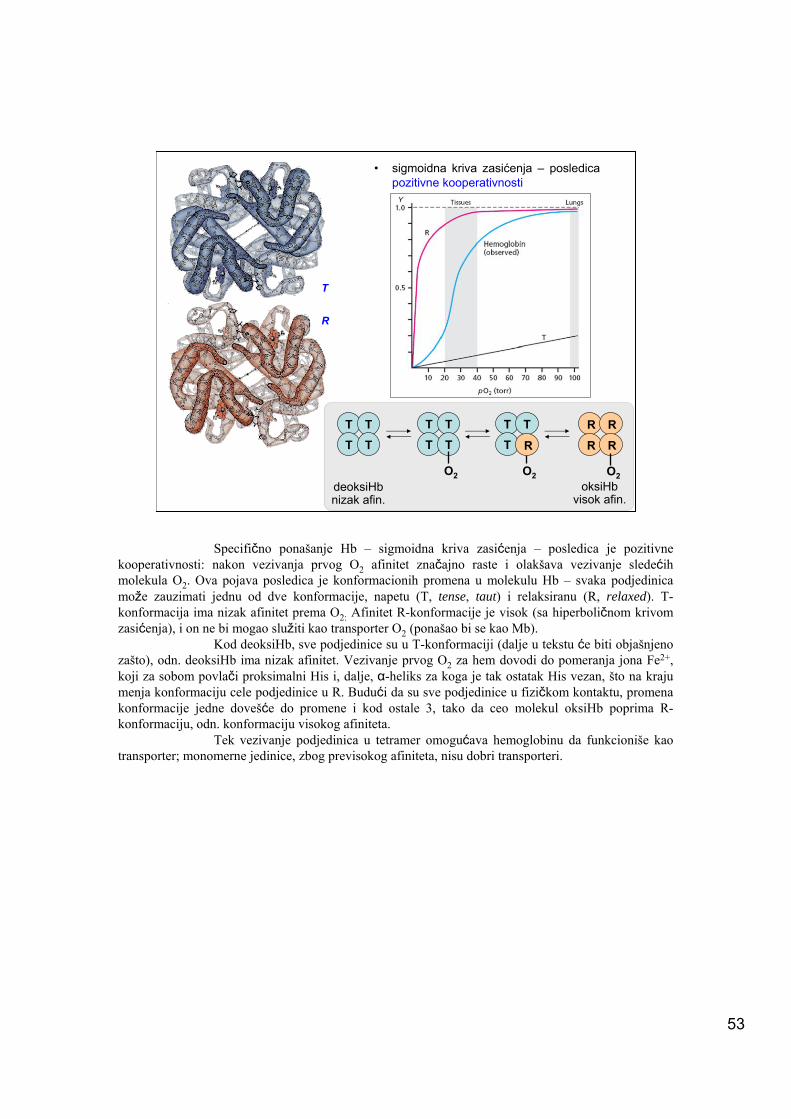
• sigmoidna kriva zasićenja – posledica pozitivne kooperativnosti
T
R
T TT T
deoksiHbnizak afin.
T TT T
O2
T TT
O2
R
R R
R R
O2oksiHb
visok afin.
Specifično ponašanje Hb – sigmoidna kriva zasićenja – posledica je pozitivne kooperativnosti: nakon vezivanja prvog O2 afinitet značajno raste i olakšava vezivanje sledećih molekula O2. Ova pojava posledica je konformacionih promena u molekulu Hb – svaka podjedinica može zauzimati jednu od dve konformacije, napetu (T, tense, taut) i relaksiranu (R, relaxed). T-konformacija ima nizak afinitet prema O2: Afinitet R-konformacije je visok (sa hiperboličnom krivom zasićenja), i on ne bi mogao služiti kao transporter O2 (ponašao bi se kao Mb).
Kod deoksiHb, sve podjedinice su u T-konformaciji (dalje u tekstu će biti objašnjeno zašto), odn. deoksiHb ima nizak afinitet. Vezivanje prvog O2 za hem dovodi do pomeranja jona Fe2+, koji za sobom povlači proksimalni His i, dalje, α-heliks za koga je tak ostatak His vezan, što na kraju menja konformaciju cele podjedinice u R. Budući da su sve podjedinice u fizičkom kontaktu, promena konformacije jedne dovešće do promene i kod ostale 3, tako da ceo molekul oksiHb poprima R-konformaciju, odn. konformaciju visokog afiniteta.
Tek vezivanje podjedinica u tetramer omogućava hemoglobinu da funkcioniše kao transporter; monomerne jedinice, zbog previsokog afiniteta, nisu dobri transporteri.

54
• 2,3-bisfosfoglicerat (2,3-BPG) se vezuje u centralnu šupljinu
• 2,3-BPG stabilizuje T-konformaciju → smanjen afinitet → otpuštanje O2
O
O
O-
PO
O-
O-
O
PO
O-
O-
• na višim nadmorskim visinama, povećan sadržaj 2,3-BPG
(normalno u eritrocitima)
(na višim nadmorskim visinama)
(bez BPG)
• lako se vezuje za deoksiHb, teško za oksiHb (R)
• u plućima (oksiHb, R) ne ometa vezivanje; u tkivima stimuliše otpuštanje O2
T RT
T T
R
RR
T T
T T
Pomenuto je da se deoksiHb nalazi u T-konformaciji. Uzrok tome je vezivanje alosternog efektora – 2,3-bisfosfoglicerata (2,3-BPG). Hb je alosterni protein – regulatorni centar udaljen je od vezivnog (gr. allos – drugi, stereos – telo, objekat). 2,3-BPG vezuje se u centralnu šupljinu Hb, između 4 podjedinice, i gradi sone mostove sa obe β-podjedinice. 2,3-BPG je negativni efektor Hb – on stabilizuje T-konformaciju, čime smanjuje afinitet i stimuliše otpuštanje O2. Lako se vezuje za deoksiHb, a teško za oksiHb jer je šupljina između podjedinica u R konformaciji mnogo manja. Zahvaljujući tome, u plućima, gde je p(O2) visok i favorizovana R-konformacija, 2,3-BPG ne ometa vezivanje; u tkivima, on stimuliše maksimalno otpuštanje O2. Bez 2,3-BPG, Hb bi prejako vezivao O2 jer bi bila favorizovana R-konformacija, te ne bi mogao da vrši svoju funkciju (već na ~20 mmHg, tj. u tkivima, bio bi zasićen).
Koncentracija 2,3-BPG u eritrocitima približno je jednaka koncentraciji Hb. Na višim nadmorskim visinama, u slučaju zagušenja disajnih puteva i sl., koncentracija 2,3-BPG se povećava da bi se stimulisalo otpuštanje dovoljnih količina O2 u tkiva i sprečila hipoksija.
Mb, pošto ima samo jednu podjedinicu, ne pokazuje pojavu kooperativnosti.
* NAPOMENA: U literaturi se ponegde javlja pogrešan naziv „difosfoglicerat”. „Bisfosfo-” označava dve nezavisno vezane fosfatne grupe, a „difosfo-” vezanu difosfatnu (pirofosfatnu) grupu!
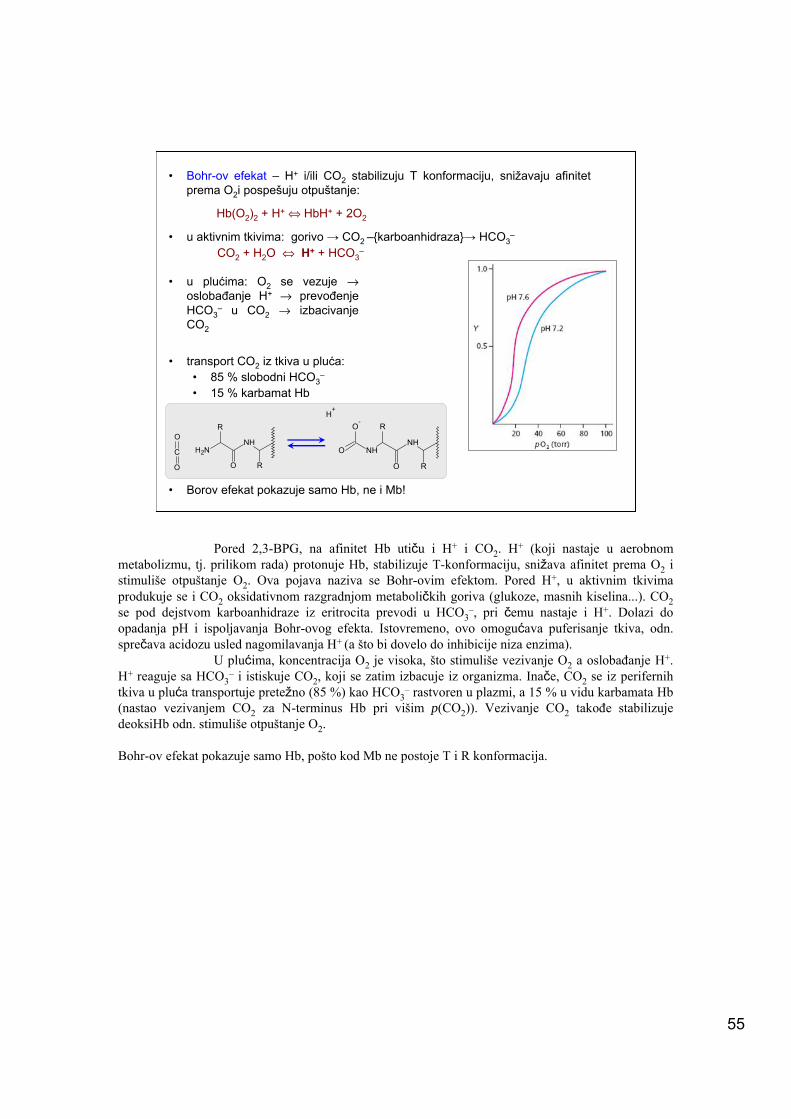
55
• Bohr-ov efekat – H+ i/ili CO2 stabilizuju T konformaciju, snižavaju afinitet prema O2i pospešuju otpuštanje:
Hb(O2)2 + H+ ⇔ HbH+ + 2O2
• u aktivnim tkivima: gorivo → CO2 –{karboanhidraza}→ HCO3–
CO2 + H2O ⇔ H+ + HCO3–
• u plućima: O2 se vezuje → oslobađanje H+ → prevođenje HCO3
– u CO2 → izbacivanje CO2
• Borov efekat pokazuje samo Hb, ne i Mb!
• transport CO2 iz tkiva u pluća:• 85 % slobodni HCO3
–
• 15 % karbamat Hb
NH2NH
R
O R
C
O
O
NHNH
R
O R
O-
O
H+
Pored 2,3-BPG, na afinitet Hb utiču i H+ i CO2. H+ (koji nastaje u aerobnom metabolizmu, tj. prilikom rada) protonuje Hb, stabilizuje T-konformaciju, snižava afinitet prema O2 i stimuliše otpuštanje O2. Ova pojava naziva se Bohr-ovim efektom. Pored H+, u aktivnim tkivima produkuje se i CO2 oksidativnom razgradnjom metaboličkih goriva (glukoze, masnih kiselina...). CO2se pod dejstvom karboanhidraze iz eritrocita prevodi u HCO3
–, pri čemu nastaje i H+. Dolazi do opadanja pH i ispoljavanja Bohr-ovog efekta. Istovremeno, ovo omogućava puferisanje tkiva, odn. sprečava acidozu usled nagomilavanja H+ (a što bi dovelo do inhibicije niza enzima).
U plućima, koncentracija O2 je visoka, što stimuliše vezivanje O2 a oslobađanje H+. H+ reaguje sa HCO3
– i istiskuje CO2, koji se zatim izbacuje iz organizma. Inače, CO2 se iz perifernih tkiva u pluća transportuje pretežno (85 %) kao HCO3
– rastvoren u plazmi, a 15 % u vidu karbamata Hb (nastao vezivanjem CO2 za N-terminus Hb pri višim p(CO2)). Vezivanje CO2 takođe stabilizuje deoksiHb odn. stimuliše otpuštanje O2.
Bohr-ov efekat pokazuje samo Hb, pošto kod Mb ne postoje T i R konformacija.
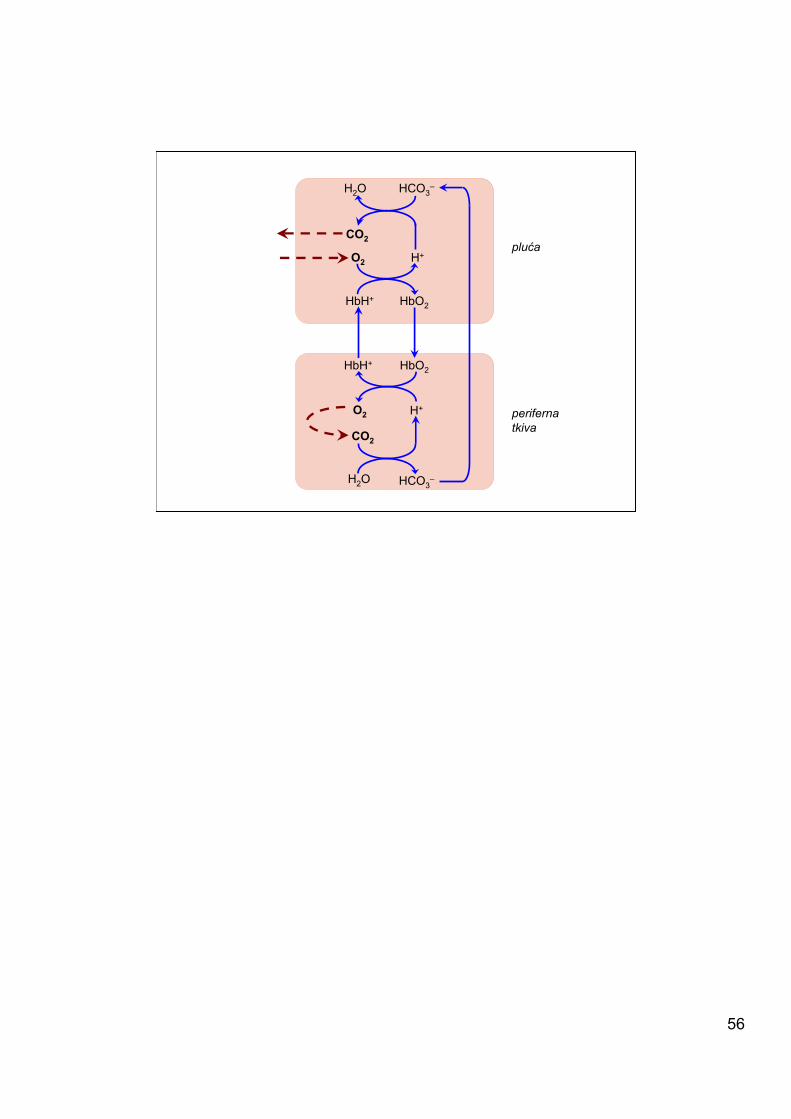
56
pluća
perifernatkiva
HbH+ HbO2
O2 H+
CO2
HCO3–H2O
HbH+ HbO2
O2 H+
CO2
HCO3–H2O

57
2.3.8. Analiza i sinteza proteina2.3.8.1. Razdvajanje proteina
zasnovane na dimenzijama čestica• ekskluziona hromatografija• ultrafiltracija• ultracentrifugiranje
zasnovane na naelektrisanju• jonoizmenjivačka hromatografija• elektroforeza• izoelektrično taloženje
ostale metode• afinitetna hromatografija• RP hromatografija• hromatografija na stubu
gel filtracija afinitetna hr.
ultracentrifugiranje ultrafiltracija
Zbog izuzetnog značaja proteina za živi svet, javila se potreba za njihovom analizom, u smislu određivanja aminokiselinskog sastava, sekvence, 3D strukture i koncentracije. Ćelije i biološki materijali uopšte veoma su kompleksnog sastava, sa hiljadama biomolekula, te prvi korak u analizi nekog proteina mora biti njegovo razdvajanje od matriksa i od ostalih proteina. Metode razdvajanja proteina zasnivaju se uglavnom na razlici u dimenzijama ili razlici u naelektrisanju. Prva grupa uključuje ekskluzionu hromatografiju (gel filtracionu hromatografiju), ultrafiltraciju (kroz polupropustljivu membranu, koja propušta male molekule a ne propušta proteine) i ultracentrifugiranje (do 105 g), a druga – elektroforezu, jonoizmenjivačku hromatografiju (o kojoj je već bilo reči) i izoelektrično taloženje (taloženje pri pH(I) usled smanjenja odbojnih sila). Na kraju, treba pomenuti i tehnike kao što su reverznofazna, afinitetna i drugi tipovi tečne hromatografije.
Jedna od često korišćenih tehnika za razdvajanje proteina po dimenzijama čestica, kao i za određivanje približne molekulske mase, je gel filtracija. Kod ove tehnike, koristi se stacionarna faza na bazi perli poroznog gela. Manji proteini ulaziće dublje u pore gela, duže će se zadržavati u njemu i kasnije će eluirati, dok će veliki molekuli zaobilaziti gel i najranije eluirati sa kolone.
Afinitetna hromatografija je najselektivniji tip hromatografije. Zasniva se na specifičnim interakcijama između analita (ovde: proteina) i nekog molekula vezanog za stacionarnu fazu, koji može biti ligand ili supstrat koga protein inače vezuje in vivo, antitelo koje prepoznaje dati protein, neki drugi protein... Analizirani protein vezuje se za stac. fazu, nakon čega se ostatak uzorka spira sa kolone; eluiranje proteina postiže se narušavanjem interakcija (rastvorom soli) ili istiskivanjem (dodatkom liganda koga protein veže).

58
gel elektroforeza
pufer
pufer
gel nakonbojenja
jamice sauzorcima
gel
izvorstruje
smermigracije
Gel elektroforeza jedna je od najčešće korišćenih tehnika u biohemiji. Zasniva se na migraciji naelektrisanih čestica (npr. proteina u rastvoru puferisanom na određenu pH vrednost) u električnom polju, a kroz poliakrilamidni gel kao medijum. Pritom, brzina migracije zavisiće i od naelektrisanja (više naelektrisane čestice brže se kreću) i od dimenzija čestica (sitnije čestice lakše nalaze slobodan put kroz umreženi gel). Kada se električno polje isključi, molekuli proteina ostaju fiksirani u gelu i mogu se dokazivati (bojenjem) ili izolovati (sečenje i elektroeluiranje mrlja, ili blotting).
Ukoliko je potrebno odrediti veličinu (molekulsku masu) proteina, primenjuje se modifikovana varijanta SDS-PAGE (sodium dodecyl sulfate polyacrylamide gel electrophoresis). Uzorku se dodaje deterdžent Na-dodecil-sulfat, čiji molekuli pokrivaju molekul proteina i negativno ga naelektrišu, pri čemu će naelektrisanje takođe srazmerno Mr, te će put koji protein pređe biti srazmeran samo Mr. Paralelno sa uzorkom analizira se i smeša proteina poznatih Mr, u cilju kalibracije.

59
2.3.8.2. Sekvencionisanje polipeptidnog niza
1. razdvajanje lanaca2. fragmentacija pp lanca3. prečišćavanje fragmenata4. sekvencionisanje AK u fragmentima5. preklapanje fragmenata → određivanje sekvence pp
1. razdvajanje lanaca (raskidanje inter- i intramolekulskih veza)
• vodonične veze: urea, guanidin-hidrohlorid• disulfidni mostovi: merkaptoetanol, permravlja kiselina, ditiotreitol
CO
NH
SH2
SH SH
OH OH
S S
OH OH
2
COOH
I
HICO
NH
S
CO
NH
SCO
NH
S
COOH
Prvo uspešno sekvencionisanje proteina (insulina) izvršio je Frederick Sanger 1953., za šta je dobio Nobelovu nagradu.
Sekvencionisanje polipeptida/proteina obuhvata nekoliko faza...Prva faza obuhvata raskidanje eventualnih poprečnih veza u proteinu, uključujući
inter- i intralančane disulfidne mostove, kao i razmotavanje lanca (raskidanjem vodoničnih veza). Za raskidanje S-S mostova mogu se koristiti različiti reagensi. Permravlja kiselina (HCOOOH) oksiduje -S-S- grupu cistina do –SO3H cisteinske kiseline. S druge strane, reducensi (merkaptoetanol, HSCH2CH2OH, ditiotreitol, HSCH2CH(OH)CH(OH)CH2SH) redukuju cistin do cisteina. U tom slučaju, neophodno je sprečiti ponovno formiranje S-S mostova (na istim ili drugim pozicijama), što se može postići kovalentnim vezivanjem zaštitnih grupa (npr. reakcijom sa jodoacetatom).

60
2. raskidanje lanaca (parcijalno)
• kisela hidroliza (neke od AK se degradiraju!)• hemijski reagensi• proteaze
aromatične (Phe, Tyr, Trp), Leu i kisele AK (Glu, Asp) pepsinAsp, Gluproteaze stafilokokaC-terminalne AK (sem Arg, Lys i Pro)karboksipeptidaza AArghlostripanArgtrombinTyr, Trp, Phe, Leu i MethimotripsinArg, Lystripsin
B. Enzimi
Cys2-nitro-5-tiocijanobenzoatAsn-Gly vezahidroksilaminTrpo-jodozobenzoatMetbromcijan
A. Hemijski reagensiMesto cepanja pp lancaReagens
Pošto je duge peptidne lance veoma teško ili nemoguće analizirati, u praksi se primenjuje njihovo parcijalno raskidanje na kraće lance, koji su dalje pogodniji za analizu. Jedan od najstarijih postupaka je kisela hidroliza, koja se može koristiti i za ispitivanje AK sastava proteina (6 mol/L HCl, t → slobodne AK → aminokiselinski analizator). Međutim, prilikom ove reakcije neke AK se potpuno (Trp) ili delimično (Ser, Thr, Tyr) razlažu, a Asn i Gln prelaze u Asp i Glu. Alternativno, za selektivno, ciljano raskidanje peptidnih veza mogu se koristiti različiti hemijski reagensi (dati na slajdu), kao i enzimi iz grupe peptidaza i proteaza. Ovi reagensi raskidaju samo peptidne veze koje uspostavljaju određene AK, što olakšava kasniju interpretaciju rezultata. Na primer, tripsin raskida samo peptidnu vezu nakon Arg i Lys, himotripsin – peptidne veze nakon aromatičnih aminokiselina, BrCN – nakon Met...

61
• određivanje N-terminusa - automatizovana Edmanova tehnika
CH C ONH
R
O
CH CNH2
R
O
H
n
N C S
fenil-izotiocijanat
CH C ONH
R
O
CH CNH
R
O
HNH
Sn
4. sekvencionisanje fragmenata
CH C ONH
R
O
H H
n
NH
RO
N
S
H+
feniltiohidantoin sekvenator (sekvencer)
U sledećoj fazi, određuje se konačno sekvenca dobijenih fragmenata. 1950. godine Pehr Edman razvio je postupak – Edmanovu degradaciju – za identifikaciju N-terminalnog AK ostatka. Protein se u alkalnoj sredini tretira Edmanovim reagensom (fenil-izotiocijanat, PITC). PITC reaguje sa slobodnom, N-terminalnom amino-grupom gradeći feniltiokarbamoil- (PTC-) derivat. U sledećoj fazi, PTC-peptid tretira se anhidrovanom trifluorosirćetnom kiselinom pri čemu dolazi do selektivnog otkidanja N-terminalnog anilinotiazolina, koji se nakon ekstrakcije organskim rastvaračem dodatkom vodenog rastvora kiseline prevodi u feniltiohidantoinski derivat AK (PTH-aminokiselinu). PTH-AK može se zatim identifikovati HPLC tehnikom, čime se identifikuje i N-terminalna AK.
Ono što je značajno je da nakon ovog postupka ostaje netaknuti peptid kraći za jednu AK jedinicu, koji se može ponovo podvrgnuti degradaciji. Ceo postupak može se automatizovati korišćenjem instrumenta – sekvencera (sekvenatora). U praksi, sa 50 pmol (0,1 μg) uzorka može se izvesti do 50 ciklusa sa peptidom dužine 100–200 AK, čime se determiniše sekvenca prvih 50-ak AK u proteinu. Za veće peptide, reakcija je manje efikasna (zbog akumulacije neizreagovalih peptida). Zbog toga se u praksi i primenjuje hemijsko i/ili enzimsko raskidanje lanca na manje fragmente koji se mogu analizirati.

62
• određivanje C-terminusa
a) specifične karboksipeptidaze
b) hidrazinoliza (N2H4, 100 °C, 20-100 h, katalizator: jonoizmenjivač) –potpuno raskidanje lanca; sve AK sem C-terminalne daju aminoacil-hidrazide
+ CH C OHNH
R
O
CH CNH
R1
O
H
n
n CH CNH2
R
O
OHCH C NHNH2
R1
O
NH2
C-terminalna AK tipično se identifikuje enzimatski. Enzimi iz grupe karboksipeptidaza otkidaju C-terminalnu aminokiselinu, što omogućava njenu identifikaciju. Karboksipeptidaza Y može se primeniti u automatskim sekvenatorima sličnim Edmanovom. Drugi postupak podrazumeva korišćenje hidrazina kao reagensa. Kuvanjem sa hidrazinom na 100 °C tokom 20–100 h uz jonoizmenjivač kao katalizator postiže se potpuno raskidanje lanca i prevođenje svih AK ostataka u aminoacil-hidrazone. Jedino C-terminalna AK ostaje u vidu slobodne kiseline, i može se identifikovati.

63
Trp–Glu–Val
• rekonstrukcija sekvence
tripsin (posle Arg, Lys): himotripsin (posle Phe, Tyr, Trp):
Gly–Arg–Ala–Ser–Phe–Gly–Asn–Lys–Trp–Glu–Val
Gly–Arg
Ala–Ser–Phe–Gly–Asn–Lys Gly–Arg–Ala–Ser–Phe
Gly–Asn–Lys–Trp
Glu–ValN-terminusGly
C-terminusVal
NH2–Gly–Asn–Lys–Trp~~~
NH2–Gly–Arg–Ala–Ser–Phe~~~ ~~~Trp–Glu–Val–COOH
Ala–Ser–Phe–Gly–Asn–LysGly–Asn–Lys–Trp
Paralelnom primenom nekoliko različitih tehnika fragmentacije polipeptidnog lanca (npr. BrCN, tripsin, V8 proteaza, himotripsin) mogu se dobiti peptidni fragmenti različite dužine, koji se sekvencionišu Edmanovom degradacijom. Finalnu fazu sekvencionisanja čini rekonstrukcija sekvence celog lanca na osnovu preklapanja sekvenci fragmenata dobijenih različitim tehnikama.
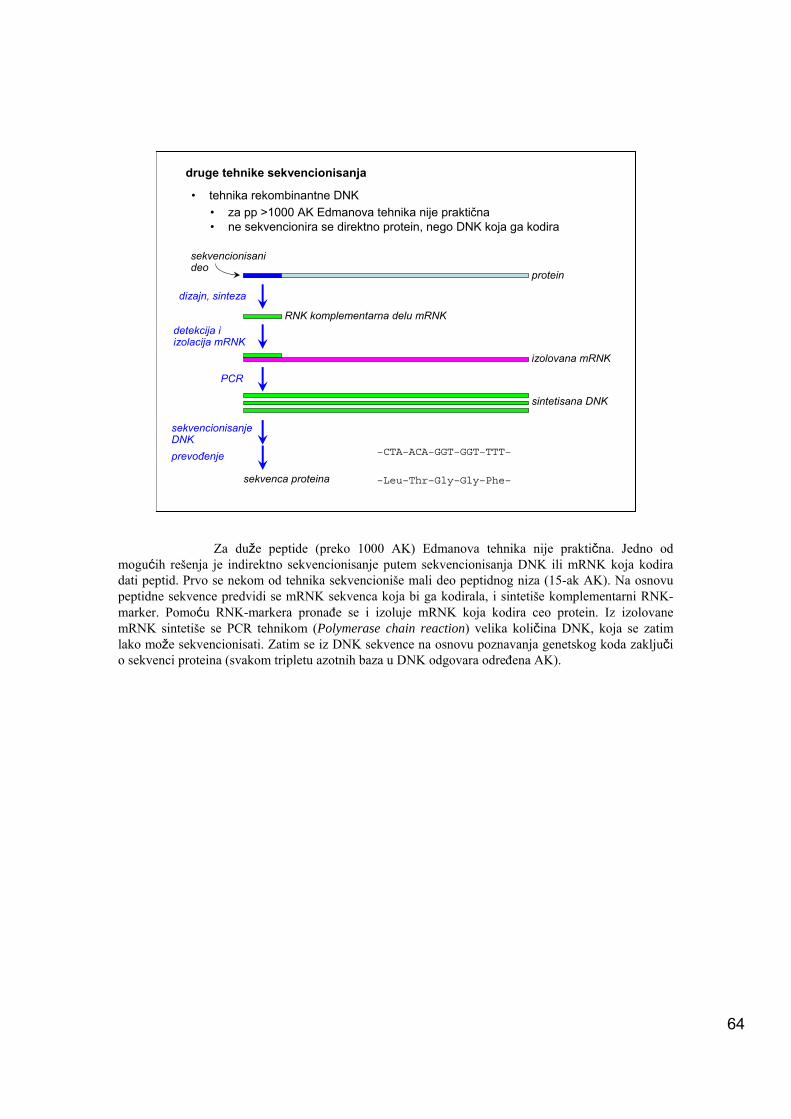
64
• tehnika rekombinantne DNK• za pp >1000 AK Edmanova tehnika nije praktična• ne sekvencionira se direktno protein, nego DNK koja ga kodira
druge tehnike sekvencionisanja
protein
sekvencionisanideo
sintetisana DNK
PCR
izolovana mRNK
detekcija i izolacija mRNK
RNK komplementarna delu mRNK
dizajn, sinteza
sekvenca proteina
–CTA–ACA–GGT–GGT–TTT–
–Leu–Thr–Gly–Gly–Phe–
sekvencionisanje DNKprevođenje
Za duže peptide (preko 1000 AK) Edmanova tehnika nije praktična. Jedno od mogućih rešenja je indirektno sekvencionisanje putem sekvencionisanja DNK ili mRNK koja kodira dati peptid. Prvo se nekom od tehnika sekvencioniše mali deo peptidnog niza (15-ak AK). Na osnovu peptidne sekvence predvidi se mRNK sekvenca koja bi ga kodirala, i sintetiše komplementarni RNK-marker. Pomoću RNK-markera pronađe se i izoluje mRNK koja kodira ceo protein. Iz izolovane mRNK sintetiše se PCR tehnikom (Polymerase chain reaction) velika količina DNK, koja se zatim lako može sekvencionisati. Zatim se iz DNK sekvence na osnovu poznavanja genetskog koda zaključi o sekvenci proteina (svakom tripletu azotnih baza u DNK odgovara određena AK).
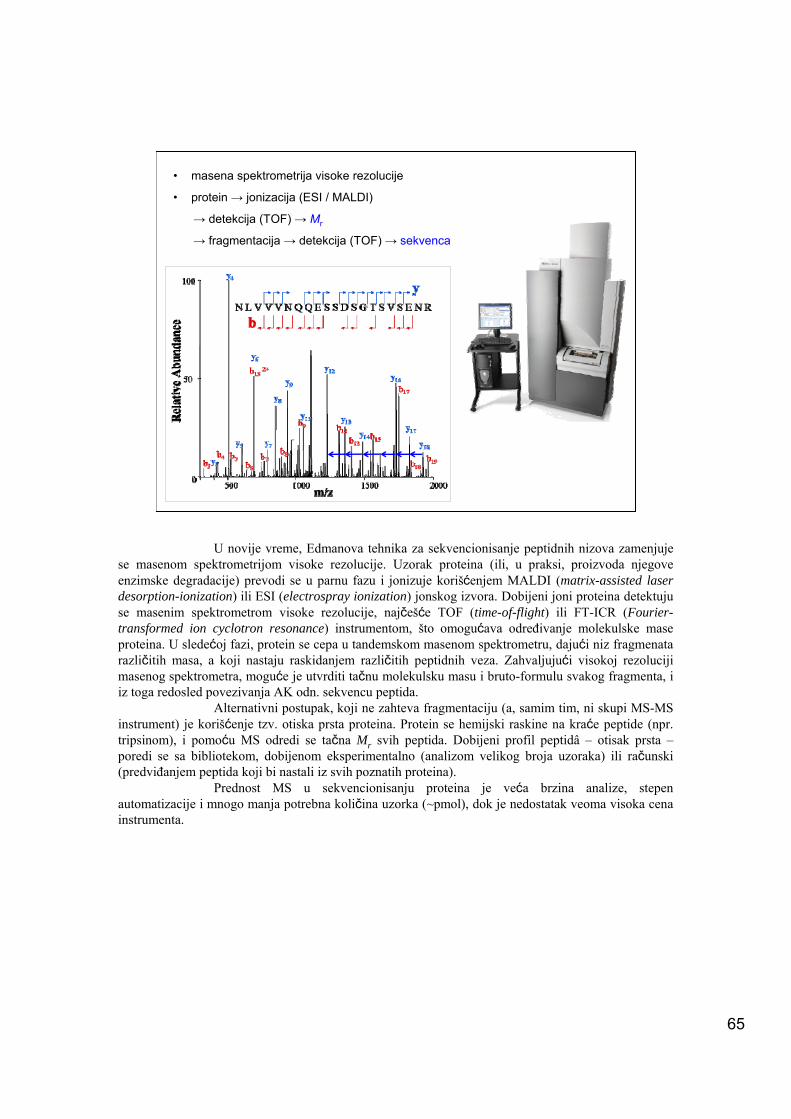
65
• masena spektrometrija visoke rezolucije
• protein → jonizacija (ESI / MALDI)
→ detekcija (TOF) → Mr
→ fragmentacija → detekcija (TOF) → sekvenca
U novije vreme, Edmanova tehnika za sekvencionisanje peptidnih nizova zamenjuje se masenom spektrometrijom visoke rezolucije. Uzorak proteina (ili, u praksi, proizvoda njegove enzimske degradacije) prevodi se u parnu fazu i jonizuje korišćenjem MALDI (matrix-assisted laser desorption-ionization) ili ESI (electrospray ionization) jonskog izvora. Dobijeni joni proteina detektuju se masenim spektrometrom visoke rezolucije, najčešće TOF (time-of-flight) ili FT-ICR (Fourier-transformed ion cyclotron resonance) instrumentom, što omogućava određivanje molekulske mase proteina. U sledećoj fazi, protein se cepa u tandemskom masenom spektrometru, dajući niz fragmenata različitih masa, a koji nastaju raskidanjem različitih peptidnih veza. Zahvaljujući visokoj rezoluciji masenog spektrometra, moguće je utvrditi tačnu molekulsku masu i bruto-formulu svakog fragmenta, i iz toga redosled povezivanja AK odn. sekvencu peptida.
Alternativni postupak, koji ne zahteva fragmentaciju (a, samim tim, ni skupi MS-MS instrument) je korišćenje tzv. otiska prsta proteina. Protein se hemijski raskine na kraće peptide (npr. tripsinom), i pomoću MS odredi se tačna Mr svih peptida. Dobijeni profil peptidâ – otisak prsta –poredi se sa bibliotekom, dobijenom eksperimentalno (analizom velikog broja uzoraka) ili računski (predviđanjem peptida koji bi nastali iz svih poznatih proteina).
Prednost MS u sekvencionisanju proteina je veća brzina analize, stepen automatizacije i mnogo manja potrebna količina uzorka (~pmol), dok je nedostatak veoma visoka cena instrumenta.

66
2.3.8.3. Određivanje 3D-strukture proteina*
• rendgenska kristalografija (X-ray crystallography)
izvor X-zraka kristalproteina
film
Za određivanje 3D strukture proteina najviše se koristi tehnika rendgenske kristalografije. Snop X-zraka propušta se kroz kristal proteina, pri čemu dolazi do njihove difrakcije. Zraci padaju na fotografski film stvarajući složene difrakcione obrasce, čijom se matematičkom (kompjuterskom) obradom dobija 3D raspodela elektronskih oblaka u molekulu, odn. 3D-raspodela atoma. Dalje se korišćenjem principa hemijskog vezivanja zaključuje o položaju pojedinačnih veza i strukturi celog molekula (proces je znatno lakši ukoliko je poznata primarna struktura). U poslednje vreme, određivanje strukture ovom tehnikom znatno je unapređeno razvojem jeftinih brzih kompjutera, boljih izvora X-zraka i automatskih sistema za pripremu kristala proteina. Zahvaljujući tome, okarakterisana je 3D struktura za desetine hiljada proteina (strukture su deponovane u Protein Data Bank, www.rcsb.org).
Druga tehnika koja omogućava određivanje 3D strukture proteina je NMR spektrometrija. Pošto NMR koristi uzorak u rastvoru, nema potrebe za izolacijom čistih kristala. NMR daje informacije o interakcijama između atoma, odn. ukazuje na prostorno bliske atome. Ukoliko je poznata sekvenca proteina, moguće je izračunati koje od mogućih 3D struktura zadovoljavaju uočene interakcije. Tehnika je ograničena na manje molekule – veći proteini daju previše složene spektre.

67
2.3.8.4. Sinteza polipeptidnog niza
• početkom XX veka: prva sinteza peptida (16 AK) – Emil Fischer• 1953: prvi biološki aktivni peptid (oksitocin) – Vincent du Vigneaud
• problem kod klasične, hemijske sinteze: sporedne reakcije
AK1 + AK2 ➜ AK1-AK2 + AK2-AK1 + AK1-AK1 + AK2-AK2
• korišćenjem zaštitnih grupa:
AK1 + z1 ➜ z1–AK1
AK2 + z2 ➜ AK2–z2
z1–AK1 + AK2–z2 ➜ z1–AK1–AK2–z2
z1–AK1–AK2–z2
z1–AK1–AK2–z2 ➜ z1–AK1–AK2 + z2
maskiranje AK1:
maskiranje AK2:
povezivanje:
prečišćavanje:
demaskiranje:
Zbog primene u medicini i nauci, postoji potreba za sintezom polipeptidnih i proteinskih lanaca tačno definisane sekvence. Prvi, kratkolančani peptidi sintetisani su već u prvoj polovini XX veka. Iako je sama reakcija formiranja peptidne veze hemijski jednostavan proces, situaciju komplikuje stvaranje neželjenih proizvoda. Već u sistemu od 2 različite AK mogu nastati najmanje 4 različita dipeptida (kao i veći broj tri-, tetra-... peptida) ukoliko se ne uzimaju u obzir i reakcije bočnih nizova. Zbog toga, da bi reakcija povezivanja AK bila kontrolisana i rezultovala samo jednim, željenim proizvodom, neophodno je sve reaktivne funkcionalne grupe sem jedne COOH i jedne NH2 maskirati uvođenjem zaštitnih grupa. Nakon uspešne reakcije i prečišćavanja proizvoda, jedna od grupa se deprotektuje i proces ponavlja sa sledećom maskiranom AK.
Najveći problem kod ovog postupka je potreba za prečišćavanjem svakog sintetisanog peptida, kao i činjenica da svaka faza zahteva manuelni rad. Ovi nedostaci rešeni su primenom sinteze na čvrstoj fazi (solid-phase peptide synthesis, SPPS).

68
• Merrifield-ova sinteza
1. C-terminalna AK1 veže se za smolu
2. + AK2 sa zaštićenom -NH2 (tBoc-grupa – t-butoksikarbonil)
3. građenje peptidne veze
4. ispiranje
5. deprotekcija AK2
6. → korak 2
7. odvajanje gotovog pp NH2 CH CO
R
tBoc-NH CH COOH
R
tBoc-NH CH CO
R
NH CH CO
R
NH2 CH COOH
R
N
C
N
DCC(dicikloheksilkarbodiimid)
NH
C
NH
O
dicikloheksilurea
N
NH
tBoc-NH CH CO
R
Sinteza na čvrstoj fazi, koju je razvio Merrifield, zasniva se na korišćenju čvrste, porozne smole pokrivene funkcionalnim grupama – linkerima – za koje se kovalentno mogu vezati peptidi.
Ceo proces sinteze sastoji se od ciklusa kuplovanje-ispiranje-deprotekcija-ispiranje. C-terminalna AK vezuje se kovalentno za smolu. Kod sledeće AK, koja treba da se veže za slobodnu NH2 grupu C-terminalne AK, NH2 grupa maskira se vezivanjem t-butoksikarbonil (tBOC) ili neke druge grupe, a COOH grupa se aktivira vezivanjem za dicikloheksilkarbodiimid (DOC). Aktivirana AK reaguje sa vezanom uz formiranje dipeptida. Višak reagenasa uklanja se jednostavnim ispiranjem (budući da željeni peptid ostaje sve vreme imobilisan na čvrstoj fazi), nakon čega se zaštitna tBoc grupa uklanja trifluorsirćetnom kiselinom, višak reagenasa ponovo ispira, i ceo ciklus ponavlja. Na kraju, kada je formiran željeni polipeptid, on se odvaja sa smole pomoću HF.
Proces je u potpunosti moguće automatizovati; automatski sintetizatori proteina komercijalno su dostupni.


















