
Alstroemeria: A Charming Genus - ΤΕΙ ΗΠΕΙΡΟΥ 683/683_4.pdf · Alstroemeria: A Charming...
15
63 Alstroemeria: A Charming Genus Andrea Mariel Sanso 1 , Marta Camargo de Assis 2 and Cecilia Carmen Xifreda 3 1 CONICET. Instituto de Botánica Darwinion. Labarden 200. C.C. 22. B1642HYD San Isidro. Buenos Aires – Dpto. de Ecología y Genética Evolutiva. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. Argentina 2 EMBRAPA/CNPM (Centro Nacional de Pesquisa em Monitoramento por Satélite) Av. Dr. Júlio Soares de Arruda 803,13088-300 Campinas, SP, Brazil 3 CIC. Laboratorio de Etnobotánica y Botánica Aplicada. Facultad de Ciencias Naturales y Museo. Universidad de La Plata. Paseo del Bosque s/n. La Plata. Argentina Keywords: cut flowers, pot flowers, landscape use, biodiversity, phylogeny. Abstract Alstroemeria (Alstroemeriaceae), the Incas lily, an exclusively South American genus comprises near 90 species ranging from Venezuela (3° North) to Tierra del Fuego, Argentina (53° South). Alstroemeria species and hybrids have gained world-wide importance as cut flower crop due to its attractive flowers and the excellent keeping quality of them. Nevertheless, they are also suitable for growing in pots and for garden and landscape uses. The recent taxonomic revisions of the group for Brazil (Assis, 2001), Argentina (Sanso, 1996), and Chile (Bayer, 1987), the biosystematic research (Aagesen and Sanso, 2003; Sanso and Xifreda, 1998, 1999, 2001, 2003) and the chromosome studies (Sanso and Hunziker, 1998; Sanso, 2002) have facilitated the identification of the taxa and improved the basic knowledge of the genus. The nomenclature of the species, their geographical distribution, ecological characteristics and the interspecific relationships are now better understood. Many wild taxa, that are scarcely known so far, carry genetic traits that open possibilities for improvement and development of more attractive new varieties. For example, some of the Brazilian taxa have an important potential value in breeding programmes because of their showy flowers and the rigid leaves, instead of the very tender foliage that possess most of the species; some Patagonian ones have rather short plant height, which is important for potted plants. Several species of the wild populations are nowadays threatened and with risk of extinction. Most of the species occupy reduced geographical areas. Species that inhabit plain regions are vulnerable because those fields are burned and cultivations are introduced. Patagonian species especially need to be protected, against the grazing of the animals. The cultivation of the native plants would help to the ex situ conservation. Reintroduction in some natural areas might also play an important role in order to maintain the landscape value. Resumen Alstroemeria (Alstroemeriaceae), el lirio de los Incas, es exclusivamente sudamericano y comprende cerca de 90 especies distribuidas desde Venezuela (3° LN) hasta Tierra del Fuego, Argentina (53° LS). Las especies e híbridos han ganado importancia mundial como flor de corte debido a sus atractivas flores y a la excelente calidad de las mismas. Sin embargo, ellas son también apropiadas para ser utilizadas en cultivos en macetas y en jardinería y paisajismo. Las recientes revisiones taxonómicas del grupo para Brasil (Assis, 2001), Argentina (Sanso, 1996) y Chile (Bayer, 1987), los estudios biosistemáticos (Aagesen y Sanso, 2003; Sanso y Xifreda, 1998, 1999, 2001, 2003) y cromosómicos (Sanso y Hunziker, 1998; Sanso, 2002) han facilitado la identificación de los taxa y contribuido al conocimiento básico del género. La nomenclatura de las especies, sus distribuciones geográficas y características ecológicas, así como las relaciones interespecíficas son ahora mejor comprendidas. Muchos taxa silvestres, hasta ahora poco conocidos, portan rasgos genéticos que amplían las posibilidades de mejoramiento y desarrollo de nuevas y Proc. V th IS on New Flor. Crops Eds.: A.F.C. Tombolato and G.M. Dias-Tagliacozzo Acta Hort. 683, ISHS 2005
Transcript of Alstroemeria: A Charming Genus - ΤΕΙ ΗΠΕΙΡΟΥ 683/683_4.pdf · Alstroemeria: A Charming...

63
Alstroemeria: A Charming Genus
Andrea Mariel Sanso1, Marta Camargo de Assis2 and Cecilia Carmen Xifreda31CONICET. Instituto de Botánica Darwinion. Labarden 200. C.C. 22. B1642HYD San Isidro. Buenos Aires – Dpto. de Ecología y Genética Evolutiva. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires. Argentina
2EMBRAPA/CNPM (Centro Nacional de Pesquisa em Monitoramento por Satélite) Av. Dr. Júlio Soares de Arruda 803,13088-300 Campinas, SP, Brazil
3CIC. Laboratorio de Etnobotánica y Botánica Aplicada. Facultad de Ciencias Naturales y Museo. Universidad de La Plata. Paseo del Bosque s/n. La Plata. Argentina
Keywords: cut flowers, pot flowers, landscape use, biodiversity, phylogeny.
AbstractAlstroemeria (Alstroemeriaceae), the Incas lily, an exclusively South
American genus comprises near 90 species ranging from Venezuela (3° North) to Tierra del Fuego, Argentina (53° South). Alstroemeria species and hybrids have gained world-wide importance as cut flower crop due to its attractive flowers and the excellent keeping quality of them. Nevertheless, they are also suitable for growing in pots and for garden and landscape uses. The recent taxonomic revisions of the group for Brazil (Assis, 2001), Argentina (Sanso, 1996), and Chile (Bayer, 1987), the biosystematic research (Aagesen and Sanso, 2003; Sanso and Xifreda, 1998, 1999, 2001, 2003) and the chromosome studies (Sanso and Hunziker, 1998; Sanso, 2002) have facilitated the identification of the taxa and improved the basic knowledge of the genus. The nomenclature of the species, their geographical distribution, ecological characteristics and the interspecific relationships are now better understood. Many wild taxa, that are scarcely known so far, carry genetic traits that open possibilities for improvement and development of more attractive new varieties. For example, some of the Brazilian taxa have an important potential value in breeding programmes because of their showy flowers and the rigid leaves, instead of the very tender foliage that possess most of the species; some Patagonian ones have rather short plant height, which is important for potted plants. Several species of the wild populations are nowadays threatened and with risk of extinction. Most of the species occupy reduced geographical areas. Species that inhabit plain regions are vulnerable because those fields are burned and cultivations are introduced. Patagonian species especially need to be protected, against the grazing of the animals. The cultivation of the native plants would help to the ex situ conservation. Reintroduction in some natural areas might also play an important role in order to maintain the landscape value.
ResumenAlstroemeria (Alstroemeriaceae), el lirio de los Incas, es exclusivamente
sudamericano y comprende cerca de 90 especies distribuidas desde Venezuela (3° LN) hasta Tierra del Fuego, Argentina (53° LS). Las especies e híbridos han ganado importancia mundial como flor de corte debido a sus atractivas flores y a la excelente calidad de las mismas. Sin embargo, ellas son también apropiadas para ser utilizadas en cultivos en macetas y en jardinería y paisajismo. Las recientes revisiones taxonómicas del grupo para Brasil (Assis, 2001), Argentina (Sanso, 1996) y Chile (Bayer, 1987), los estudios biosistemáticos (Aagesen y Sanso, 2003; Sanso y Xifreda, 1998, 1999, 2001, 2003) y cromosómicos (Sanso y Hunziker, 1998; Sanso, 2002) han facilitado la identificación de los taxa y contribuido al conocimiento básico del género. La nomenclatura de las especies, sus distribuciones geográficas y características ecológicas, así como las relaciones interespecíficas son ahora mejor comprendidas. Muchos taxa silvestres, hasta ahora poco conocidos, portan rasgos genéticos que amplían las posibilidades de mejoramiento y desarrollo de nuevas y
Proc. Vth IS on New Flor. Crops Eds.: A.F.C. Tombolato and G.M. Dias-Tagliacozzo Acta Hort. 683, ISHS 2005

64
más atractivas variedades. Por ejemplo, algunos de los taxa brasileros tienen un importante valor potencial en programas de mejoramiento debido a sus vistosas flores y a sus hojas rígidas, en lugar del follaje muy fino que posee la mayoría de las especies; algunas especies patagónicas tienen baja altura, carácter importante para su cultivo en macetas. Diversas poblaciones silvestres están actualmente amenazadas y en riesgo de extinción. La mayor parte de las especies ocupa áreas geográficas reducidas. Aquellas especies que habitan llanuras son vulnerables debido a las quemazones de esos campos y la introducción de cultivos. Las especies patagónicas necesitan ser protegidas, especialmente del pastoreo de animales. El cultivo de las plantas nativas ayudaría a la conservación ex situ. La reintroducción en algunas áreas naturales podría jugar un rol importante para el mantenimiento del valor paisajístico.
INTRODUCTIONAlstroemeria (Alstroemeriaceae), the Incas lily, is an exclusively South American
genus, represented by around 90 species that occupy a diverse range of habitats, from sea level to 4500 m altitude. They are herbaceous perennnials, with fleshy-root tubers. The leaves are frequently resupinate, the inflorescences terminal and umbeliform and the flowers usually zygomorphic with patterned tepals.
The recent taxonomic revisions of the group for Brazil, Argentina and Chile, the biosystematic research (Aagesen and Sanso, 2003; Sanso and Xifreda, 1998, 1999, 2001, 2003) and the chromosome studies (Sanso and Hunziker, 1998; Sanso, 2002) have facilitated the identification of the taxa and improved the basic knowledge of the genus. The nomenclature of the species, their geographical distributions and ecological characteristics and the interspecific relationships are now better understood. The Chilean species monographed by Bayer (1987) are 31. Sanso (1996) reviewed the Argentinean Alstroemeria species and recognized 10 taxa with one endemic species. Recently, Assis (2001) distinguished for Brazil, 38 species distributed in the eastern regions of the country; 35 of them occur exclusively in Brazil and ten of them, were described as new species (Assis and Mello-Silva, 2003). The species from Paraguay occur also in Argentina and Brazil (Sanso and Xifreda, in prep.).
Because of their showy flowers and the excellent keeping quality of them, these plants have been successfully introduced into cultivation and are used as vase flowers. In Europe, were introduced as early as in the century XVI. Since those days, several works of improvement have been carried out which has become the Inca lily in a very popular cut flower. These cut flowers crops has been developed through hybridization, primarily among the Chilean group, particularly A. aurea Graham, A. ligtu L. and A. pelegrina L. and one Brazilian species, A. psittacina Lehm. In Holland is one of the most marketable flowers (Griffin, 1995; Buitendijk, 1998), just as that in Japan (Miyake et al., 1989). Recently an interest in growing Alstroemeria as a pot plant has appeared in U.S.A., Holland, Denmark. It should be possible to increase the variation and enhance the quality of the commercial cultivars by including a larger part of the genetic variation found among wild species. Many other wild taxa, that are scarcely known so far, are also of potential use in breeding programmes. Emphasis has been taken to take species of Brazilian origin with potential ornamental value and plants of low height for use as new pot plants.
RESULTS AND DISCUSSION
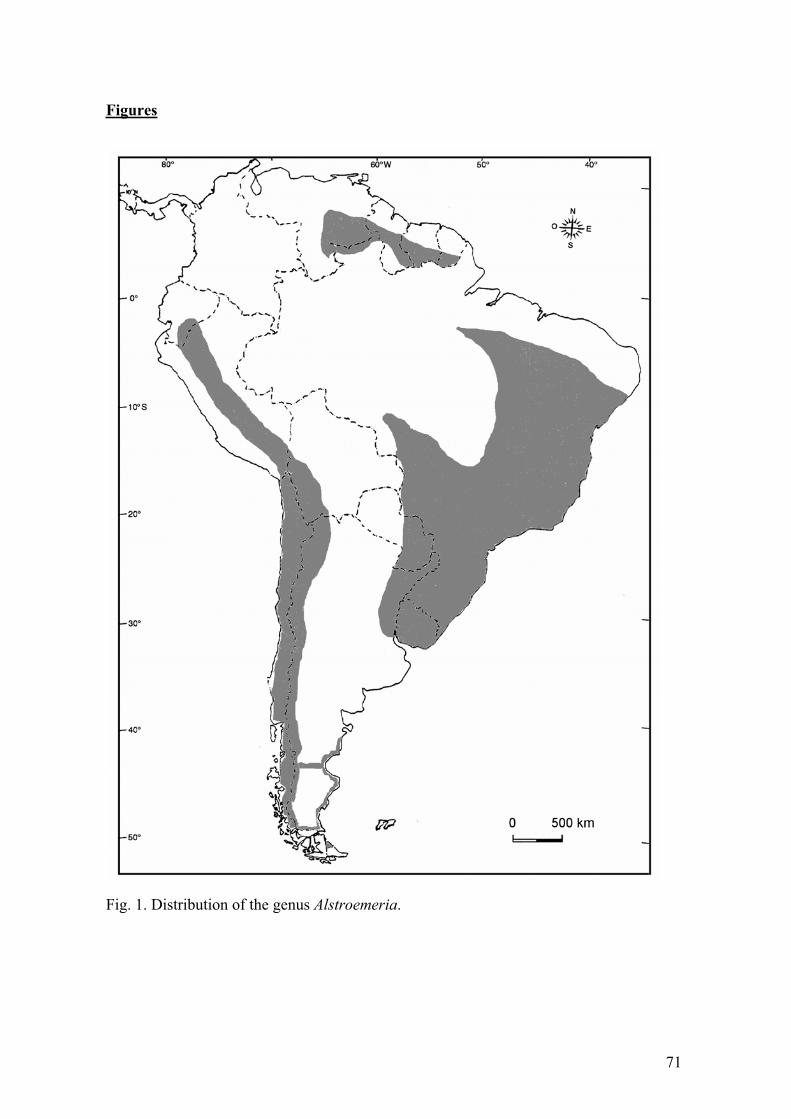
DistributionIts northern limits are southern Venezuela and north-west Brazil (3-4º N) while it
extends as far south as Tierra del Fuego in Argentina (52-53º S), with two main centers of distribution, one in Chile (extending into Peru, Bolivia and Argentina) and the second throughout Brazil, Paraguay and Argentina (Fig. 1). Native species occupy a diverse range of habits from high Andes to marshy lands. In Chile, Alstroemeria is geographically

65
distributed from a latitude of 22º S, near Tocopilla (Region II) in the north (A. pauperculaPhil.), where the vegetation is sparse, with various species of cacti and other xerophytic plants, to 51º S in the south (A. patagonica Phil.). The central region of Chile has the greatest number of species. South of the 36º latitude the number of species is limited to A.aurea, the most widespread species in Chile, and A. presliana Herb.(Ørgaard and Kristiansen, 1998). The Brazilian species occur at different habitats: forest, savanna, high fields, marsh, “campos rupestres” and “caatinga”, from a height of 300 m in the Amazon up to 2,300 m at Serra do Itatiaia. With exception of A. isabellana Herb., A. gardneri Baker, A. apertiflora Baker, A. cunha Vell., A. stenopetala Schenk, A. psittacina and A. longistyla Schenk, most species have a restricted distribution (Assis, 2001).
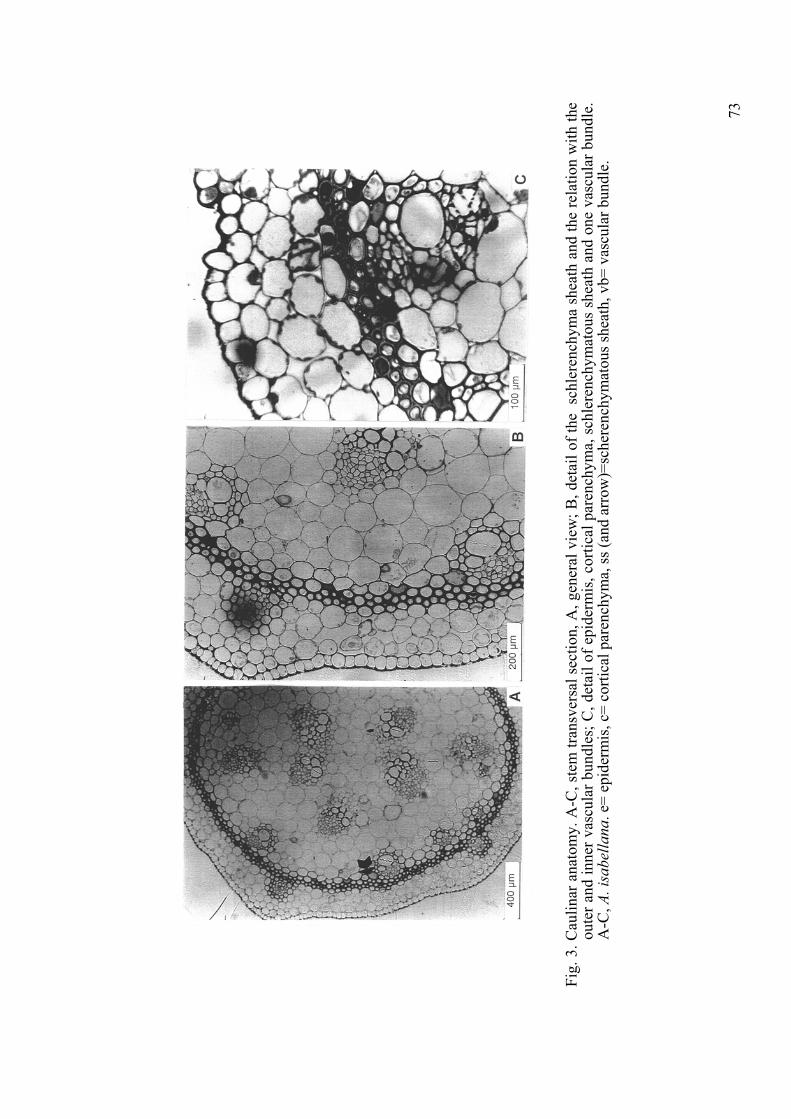
Characters1. Subterranean Organs. In Alstroemeria, the species have either a short, nodose rhizome, as for example in A. pygmaea Herb. or, more frequently, the rhizome is long (Fig. 2A). In some species the rhizomes run horizontally, and in each growing season produce aerial vegetative leafy shoots and, usually, one flowering stem. These stolons or runners allow rapid shoot growth by means of vegetative multiplication. Traditionally propagation of Alstroemeria for horticulture is done by division of the rhizomes. Young roots are thin and fibrous but some of these thicken apically to become cylindrical or fusiform, white or cream-coloured, starch containing storage organs. The fleshy roots can be enlarged as in A. psittacina, + enlarged as in A. aurea and A. patagonica (Fig. 2B) or slender as in A. presliana. They may be directly grouped next to the rhizome as in A.apertiflora (Sanso and Xifreda 2001: Fig. 2A) and A. pygmaea (op. cit.: Fig. 1D) or distinctly separate from the rhizome as in A. patagonica Phil. (Fig. 2B) (Sanso and Xifreda, 2001). According to Bayer (1998), A. graminea Phil., an annual plant, does not develop storage roots. Meerow and al. (1999) described a new species from Goiás and Distrito Federal in Brazil, A. hygrophila Meerow, Tombol. & F.W. Mey, which apparently did not form thickened roots either but later it was synomyzed with A.longistyla and observed that the roots are also enlarged (Assis, 2001). 2. Foliar and Caulinar Anatomy. The stem presents a single-layered epidermis with thickenings of outer and periclinal walls, thinner anticlinal ones and sunken stomata with thick cuticular ridges. Closed and collateral vascular bundles are arranged in an atactostele, with 1-3 large metaxylem vessels surrounded by parenchyma. The central bundles are larger than the outer ones (Fig. 3A). Mechanical tissue as a continuous schlerenchymatous sheath bounds to the cortex, presenting generally 3-6 layers of characteristic short cells (Figs. 3B, C).
Resupinate leaves are characteristic for nearly all members of the genus. The leaves are twisted near the base or along the lamina, hence the abaxial leaf surface becomes the functionally adaxial surface. A comparative survey of leaf anatomy has been performed by Sanso and Xifreda (1998; 2003). The major characteristics are: lamina thickness between 4 to 10 mesophyll cell layers, two types of chlorenchyma distribution, dorsiventral in most of the Alstroemeria species and isolateral in selected ones as some Brazilian species with rigid leaves. Few species exhibit irregularly shaped, lobed chlorenchyma cells and many intercellular spaces as in A. isabellana (Figs. 4A-C), possibly an adaptation to hydrophily. Closed vascular bundles organized in parallel disposition are usually surrounded by a distinct parenchymatic sheath or with schlerenchymatic cells extending as girders towards both epidermises. Anomocytic stomata, usually sunken, occur on one or both surfaces. Idioblasts with oxalate raphides in the mesophyll are widespread in the genus. 3. Inflorescence and Flowers. In few taxa, there is an unique flower, e.g. A. patagonica,A. pygmaea or, two ones, e. g. A. andina Phil. subsp. venustula Ehr. Bayer (Fig. 2E). The flowers are epigynous and are nearly always zygomorphic due to the position of the inner tepals. The zygomorphy is reinforced by the position of the filaments, which are located in the lower part of the flower during anthesis (Fig. 2F). In A. pygmaea and A. patagonica (fig. 2D) the flowers are actinomorphic but with some ones showing some degree of

66
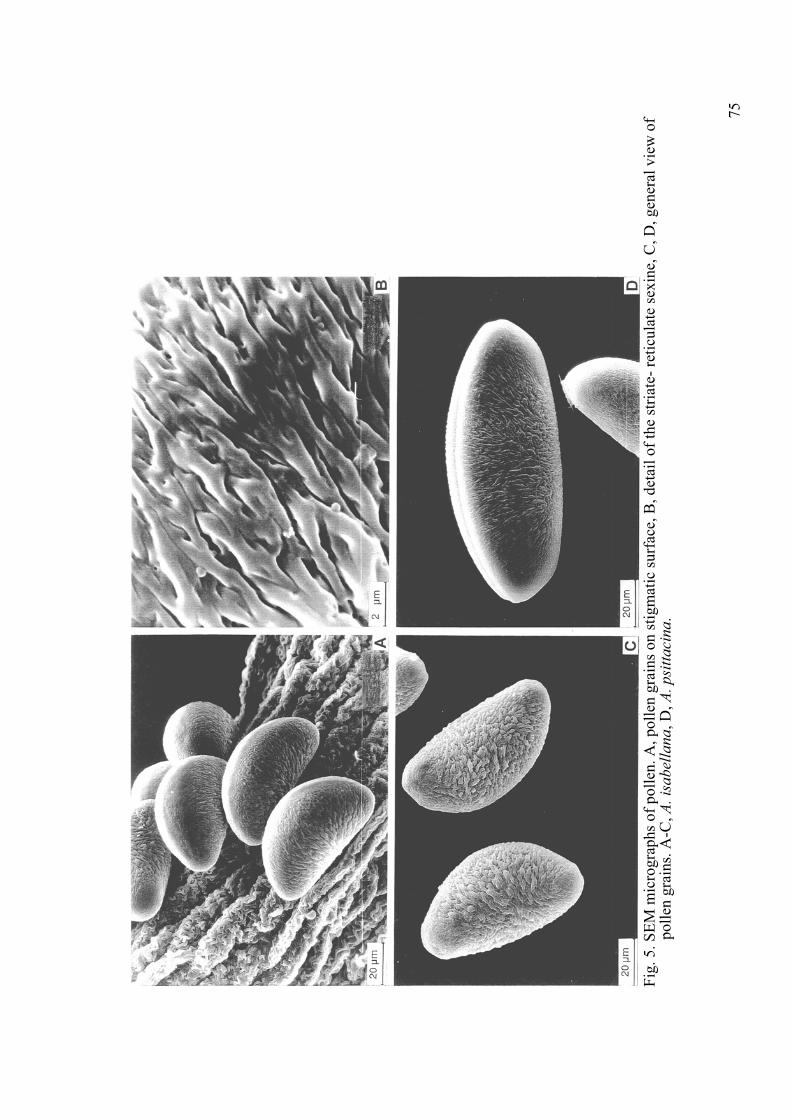
zygomorphy. Most of the floral characters have taxonomic importance. Several of them are also of ornamental weight as the basic colour, which varies from yellow, orange through red-purple and violet to pink and white, the disposition of it, the form of the tepals and the presence or not of spots or streaks on them. A large variation regarding flower colours and patterns is represented. Some species have tiny flowers with a diameter of about 1cm, e.g. A. revoluta Ruiz et. Pav., while other such as A. magnifica Herb. and A. pelegrina L., the type species of the genus, have large showy flowers with a diameter of up 7 cm. Some Brazilian species, as A. isabellana (Fig. 2C), have more tubular flowers with narrower perianth segments. 4. Pollen. The pollen of all species investigated is ellipsoid (boat-shaped), heteropolar and monosulcate (Figs. 5A, C, D). Symmetry is bilateral. Size ranges from 50-120 μm, long axis, x 25-62 μm, short axis. Since pollen size in general ranges from c. 20- 150 μm, these pollen grains would be considered to be large to very large. The exine is brevi-striato-reticulate (Fig. 5B). The striate muri are arranged in a parallel to sub-parallel pattern. In some species small, interconnecting muri are apparent between striae. This structure results in the roundish to elongate perforations and gives a semitectate exine structure. The perforations are fine, for example in A. graminea, A. angustifolia subsp. angustifoliaand A. werdermanii Ehr. Bayer, in A. isabellana, A. pseudospathulata Ehr. Bayer and A. versicolor Ruiz et Pav. the perforations are much coarser. The ultrastructure of the sporoderm is detailed in Sanso and Xifreda (2001). 5. Fruit Morphology and Dehiscence. The fruit of Alstroemeria is a 3-valved
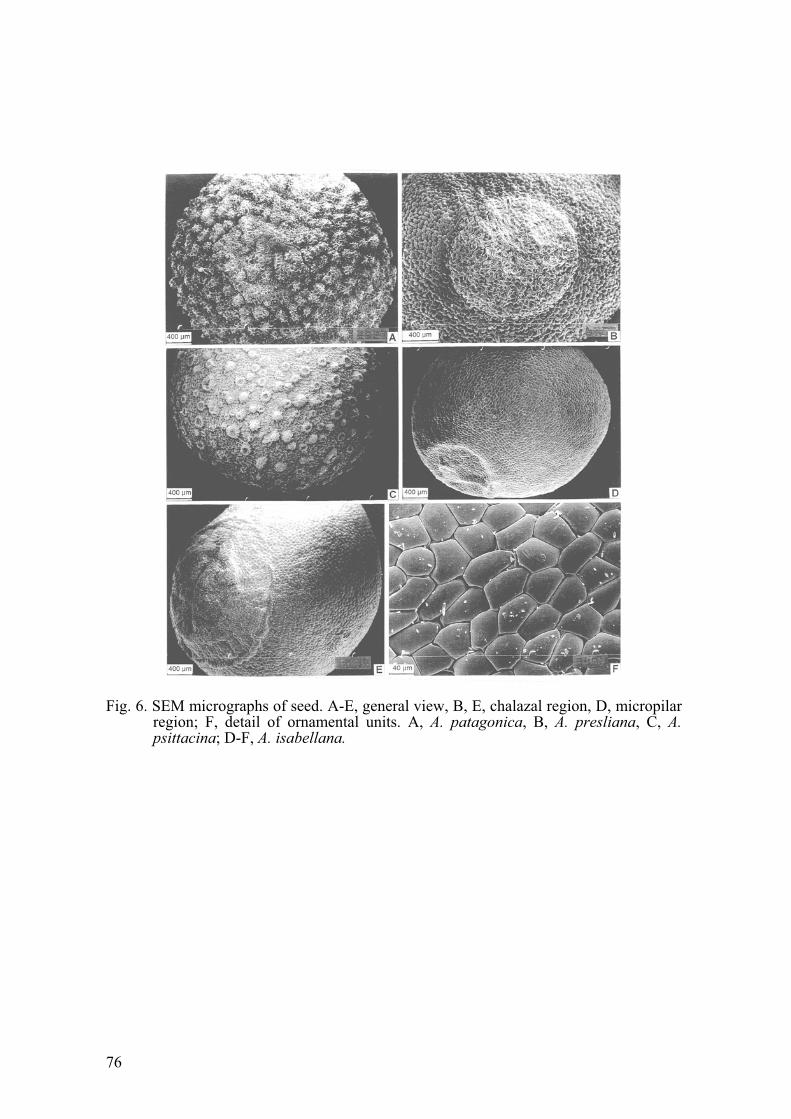
loculicidal capsule. In superior view it is rounded, while in lateral view it is ovoid, 6-ribbed, slightly or notably umbonate towards the apex (Figs. 2G, H), coriaceous and bright light brown when mature. When unripe the fruits are greenish, with six longitudinal ribs, which are joined by a transverse rib below the high, widely conical apex, which is notably marked and more angular than in the mature fruit. Sometimes the ribs are purple (A. presliana). At maturity, the fruits are rounded, the colour turns golden or clear brown and the texture is papiraceous-coriaceous (Sanso and Xifreda, 2001: Fig. 7A). In A. pygmaea and A. patagonica, the inmature fruits are covered by leaves. In some species, the umbo (apex) is short, 0.5-3 mm in length, as in A. aurea, A. andina, A. isabellana (fig. 2H) while in other species it is more pronounced, it may be as much as 20 mm in length, as in A. pseudospathulata (Fig. 2G), A. pelegrina, A. presliana, A. pulchra Sims. The dehiscence is explosive, the capsule splits from the base into three parts. The three valves, together with the seeds, are thrown a considerable distance. Dehiscence takes place when the fruit is dry. The stem remains with three (to six) of the longitudinal ribs more or less remaining in the shape of a claw. Rarely, when the tension is not too great, the capsule is not totally destroyed, and the 3 valves do not completely separate from the stem (Sanso and Xifreda, 2001).6. Seed. In Alstroemeria the seeds per capsule are numerous. The size ranges from 2-4 mm in diameter. They are yellowish-brown, tan or brown, globose or subglobose (Figs. 2I-J, 6A- E). The seed coat is dry and hard. The primary sculpturing consists of conspicuous protuberances, which gives the seeds a peltate (Fig. 6F), papillose or verrucose appearance. In several species the testa is colliculate, for example in A. aurea and A. presliana. High magnification with SEM shows the individual units of the seed surface sculpturing, although basically convex, to have considerable morphological variation between species, some are conical with a wide base, for example in A. andinasubsp. venustula, while in other species very specialized structures occur. In A. psittacina,a central cell is surrounded by 9-10 peripheral cells (Fig. 6C). The cells that comprise these ornamental units, and give roughness of the surface of the seed coat are thick-walled and pitted. The surface of the outer periclinal cell wall may also exhibit micro-ornamentation and secondary sculpturing, for example, irregular hollows or wrinkles (A.andina subsp. venustula, Sanso and Xifreda, 2001: Figs. 8E, H), micro-papillate (verrucate) (A. graminea, op. cit.: figs. 9B-C) or smooth (A. aurea, A. presliana). A dark brownish-black discoid appendage, arising from the chalazal region occurs in the seeds of Alstroemeria (Figs. 6B, E) and the raphe is a shallow longitudinal rib. The thick seed coat

67
consists of just a few cell layers. The seeds are dispersed by a ballistic mechanism. The almost perfect spherical shape guarantees efficient propagation by the centrifugal force exerted on the seed as it is expelled from the capsule. In some cases the seed coat protuberances with large surface cells, may have an adhesive property. A straight, linear and short embryo is enclosed in the endosperm near the micropile (o.c.).7. Chromosomes. Alstroemeria has special interest because of its large chromosomes and its asymmetric karyotypes. It may be considered chromosomally stable with a constancy of the chromosome number, 2n = 16, an unique level of ploidy, 2x, and an asymmetrical complement (Sanso and Hunziker, 1998; Sanso, 2002). Although chromosome sizes vary from species to species, they are karyotypically very similar in terms of relative size relationships between the chromosomes. The proportion of the total complement occupied by the largest chromosome pair varies between 21.4 % and 45 %, with the exception of A.ligtu L: subsp. ligtu (Buitendijk and Ramanna, 1996). Between species, there is a considerable variation in nuclear DNA content, the amount of C-banded heterochromatin (Buitendijk and Ramanna, 1996; Buitendijk and al., 1997) and the presence or lack of satellites on several pairs of chromosomes. Two species differed by about a factor of two in the total chromosome length (A. magnifica = 116 μm; A. pygmaea = 224 μm). However, in Alstroemeria, bimodal karyotypes occur despite these differences in the total chromosome length. It seems that in this group DNA has been added to the complements without altering the karyotype morphology too much (Sanso, 2002).8. Phylogeny. Combining the morphology, rps16 and rbcL data sets and analyzing under equal weights, the strict consensus tree shows that Alstroemeria is monophyletic; within Alstroemeria, the Brazilian species form a clade, which is sister group to the rest of the genus. Among the Chilean/Argentinean species A. patagonica and A. pygmaea is the sister group to the remaining species. All other minor species (A. pseudospathulata,A. graminea, A. andina, and A. werdermannii) are gathered in a clade which is the sister group of a clade consisting of purely Chilean species (Aagesen and Sanso, 2003). When analyzed under implied weights the results differ from those obtained when using equal weights mainly by placing A. andina, A. graminea and A. werdermannii as sister group to the rest of the genus while the Brazilian species are embedded within the Argentinean/Chilean clade, still forming a monophyletic group. Only when the rps16 data are analyzed alone is monophyly of Alstroemeria not maintained. Both morphology and rbcL data support monophyly of the genus. Neither Alstroemeria graminea nor A.pygmaea ever occur in a position that makes their segregation from Alstroemeriapossible. Monophyly of the Brazilian species is supported in all analyses where the three data sets are combined. When analyzed under equal weights, the considered Brazilian species (A. apertiflora, A. isabellana, A. caryophyllaea Jacq. and A. psittacina) are placed near the root of the genus. The flowers of some Brazilian Alstroemeria species superficially resemble Bomarea flowers more than the flowers of some Chilean species do (i.e. smaller and more tubular flowers with the same color pattern on all inner tepals) and there is some support for the Brazilian species being placed near the root of the genus when all data sets are combined as a result of the influence of the morphological characters. However, when implied weights are used the Brazilian species are placed among the Argentinean/Chilean species, which is the position favored by rbcL. At present, monophyly of the Brazilian species seems supported, but morphology and DNA sequence data disagree about the position of the clade (o.c.).
Evaluation of Species for Ornamental Purpose Alstroemeria psittacina, native from SE Brazil, NE of Paraguay, Uruguay and NE
of Argentina, A. caryophyllaea, from Brazil, A. aurea, from Argentina and Chile, A. ligtu,A. versicolor, A. revoluta, A. pelegrina, A. violacea Phil., A. hookeri Schultes, all from Chile, have been used, among other, in crossings from which several commercial cultivars have been obtained. Nevertheless, many others wild taxa, that are scarcely known so far, are also of potential use in breeding programs. Several of the wild species carries genetic traits that wide-open possibilities for improvement and development of

68
more attractive new varieties. For example, some of the Brazilian taxa have rigid leaves, instead of the very tender foliage that possess most of the species; some Patagonian ones have rather short plant height, which is important for potted plants.
Selected Brazilian Alstroemeria species are briefly summarized, the ones that have more potential use in breeding programs: A. isabellana (Fig. 2A) is found in Argentina, Uruguay, Paraguay and Brazil. In this country, is possibly the most widely distributed species of Alstroemeria, occurring at high elevation and along the marshs from the state of Rio de Janeriro to Rio Grande do Sul and west to Minas Gerais. This plant can be up to about 1,5 m tall, has rigid leaves, and pendulous, tubular flowers (Fig. 2C), colored pink, orange red, or yellow and tipped in green. Inner tepals are stripped and spotted. It flowers and fruits in Jan.-Dec. A. longistaminea Mart. ex Schult. & Schult. f. (Fig. 7A) occurs at semi-shaded sites in forests of Pernambuco, Sergipe and Bahia states, flowers in May-Aug. and fruits in Aug.-Sep. This species of tender foliage can be up to 1,5 m tall. It has pendulous flowers with the outer tepals broader then inner, and its colour varying from yellow greenish to pink greenish.A. sellowiana Seub. ex Schenk (Fig. 7B), an odoriferous species, just as A.caryophyllaea, is shorter than the other Brazilian specie although can be up to 0,5 m tall; the leaves are rigid, narrow and the flowers are yellow to orange with dark spots on inner tepals. It occurs at marshs of high fields from Paraná to Rio Grande do Sul states, flowers in Nov.-Feb. and fruits in Jan.-Feb.A. foliosa Mart. ex Schult. & Schult. f. (Fig. 7C) is perhaps, one of the showiest species from Brazil. It is about 0,4-0,6 m tall, the leaves are chartaceus, the flowers are about 0,4-0,6 m long, and the colour varies from yellow throught deep orange or red. The tepals are broader than typical of most Brazilian Alstroemeria species. It grows in high fields of Minas Gerais, Rio de Janeiro and São Paulo states, and flowers in Nov.-Apr. A. speciosa M.C.Assis (Fig. 7D) is another showy species that grows on the shadow at forest of Serra do Mar in São Paulo states, flowers in Dec.-Feb. and fruits in Mar. It can be up to 1,5 m tall; the leaves are large and tender and the flowers are also large, 4-5 cm long., red or yellow colored with the inner tepals dark spotted. A. variegata M.C.Assis (Fig. 7E) is another attractive species with crowded leaves and flowers ranging from red to yellowish on outside, and yellow and dark striated on inside. This species is endemic of campos rupestres of Caldas in Minas Gerais Satates, flowers in Nov.-Feb. and fruits in Jan. A. plantaginea Mart. ex Schult. & Schult. f. (Fig. 7F) grows in “campos rupestres” of Minas Gerais, São Paulo and Bahia states, flowers in Dec.-Apr. and fruits in Feb.-Apr. This species of rigid or chartaceous leaves can be up to 1,2 m tall. The flowers are about 3,5 cm long, with the outer tepals colour ranging from red to orange and the inner ones with a yellow background and dark spots. A. gardneri (fig. 7G), a plant about 0,3-1,5 m tall, with rigid leaves, is very frequent at the Savannahs of Mato Grosso, Goiás and Distrito Federal, flowers and fruits in Jan.-Dec. The flowers size is about 3,5-4,5 cm long and the colour varies from deep red to yellow red on the outside, yellow and spotted internally. A. malmeana Kraenzl. (Fig. 7H) is scarcely found in marshs of Rio Grande do Sul and Paraná states, flowers in Jan.-Feb. and fruits from Jan. This species can be up to 0,9 m tall, the leaves are rigid, amplexicaulos and the flowers, red, pendulous with the stamens and the yellowish stigma exposed. A. stenophylla M.C.Assis (Fig. 7I) is a charming species because of its vegetative characteristics. Its height runs about 0,2-0,7 m tall. The leaves are narrow linear, rigid or chartaceus. The flowers are very similar to A. gardneri, reddish or orange, 2-3.8 cm long. and the inner tepals are dark spotted. It grows on the rocky outcrops and on riverine forest borders in the states of Goiás and Minas Gerais, flowers in Mar.-May and fruits in May.
These species have an important potential use in breeding programs and deserve much attention. However, there are no available studies about pollination and reproduction, neither are there about in situ and ex situ, conservation or in vitro culture in

69
relation to the Brazilian taxa. Plant height also varies considerably in the genus but the species with lower height
are found among the chilean or argentine-chilean taxa. Some of these species with potential use as pots plants are: A. pygmaea, a small herb, 6-15 cm tall, grows at 3500-4400 m in the Andean mountains of Perú, Bolivia and north-west Argentina. The flower is not usually zygomorphic and bright yellow. The upper leaves grow in the soil. A. patagonica (Fig. 2B), 2-14 cm tall, exceptionally can be up to 25 cm. The leaves are linear and the terminal flower is bright yellow or orange, sometimes with small purple spots on the tepals. A. andina subsp. venustula, also a small herb, 5-16 cm tall, occurs at 2800-3700 m above sea in Argentina and Chile. The flowers are erect, infundibuliform, with all the tepals very reflex (Fig. 2E); the outer ones, whitish to rossish, with lilac and the apex greenish, the inner ones, of several colours, ¼ near apex, violet-lilac, ¼ lower, white with a transversal yellow band and near the basis, white, violet spots on all the surface, diminishing in size towards the basis. The another subspecie is also of low height.A. pseudospathulata, another trans-Andean species of Argentina and Chile, inhabits the Patagonian meseta in Argentina. In some cases is taller than the above described species, between 15 and 30 cm tall. The flowers, erect and big (Fig. 2F), yellow, sometimes have dark spots near the thickened apex.A. spathulata Presl., has spatulate and rather succulent leaves and the flowers, with thickened pink tepals, are only raised 4-15 cm above the gravel. A. garaventae Ehr. Bayer, 5-20 cm tall, a species related with the previous one but with the leaves linear to lanceolate. A. umbellata Meyen, 2-8 cm tall, flowers of medium size, pink, infundibuliform with the outer tepals deep-set in the apex. A. hookeri Loddiges, with four subspecies, between 4-10 and 4-22 cm tall. The colour of the flowers varies among pink, rossish and white with different pattern of stripes and spots on the inner tepals.A. graminea, 2,5- 8 cm tall, leaves graminiform with a solitary flower; the outer and the inferior inner tepals rossish, the two superior inner ones, 1/3 inferior rossish and the 2/3 superior yellow with dark spots. This species with the two ones described below occur at Atacama desert. A werdermannii Ehr. Bayer, 4-8 cm tall, with small and elliptic leaves and very narrow outer tepals. A. polyphylla Phil, 6-20 cm tall, leaves wide and sessile, flowers of medium size, red, lilac or white.
The variation among and within species and the many possibilities for species hybridization opens up the possibility for production of a wide range of new cultivars suited for various purposes, such as cut flowers, pot plants and garden and landscape uses. Besides, several of the wild populations are nowadays threatened and with risk of extinction. Most of the species occupies reduced geographical areas. Species that inhabit plain regions are vulnerable because those fields are burned and cultivations are introduced. Patagonian species like A. pseudospathulata or A. patagonica need to be protected especially, from the grazing of the animals, which causes their declination. The cultivation of the native plants would help to the ex situ conservation. Reintroduction in some natural areas might also play an important role in order to maintain the landscape value.
ACKNOWLEDGEMENTS We thank Lic. Rafael Urréjola (La Plata Museum, Argentina) for technical assistance and Sr. Vladimiro Dudás for valuable help with the figures.

70
Literature Cited Aagesen, L. and Sanso, A.M. 2003. Phylogeny of the Alstroemeriaceae, based on
morphology, rps16 intron, and rbsL sequence data. Syst. Bot. 28 (1): 47-69.Assis, M.C. de. 2001. Alstroemeria L. (Alstroemeriaceae) do Brasil. Ph.D. Thesis.
Universidade de São Paulo. São Paulo. Brasil. Assis, M.C. de. and Mello-Silva, R. de. 2003. Brazilian Alstroemeria L.
(Alstroemeriaceae): synopsis. Poster presented at Monocots III, Third Intl. Conference on the Comparative Biol. of the Monocotyledons. California. U.S.A.
Bayer, E. 1897. Die Gattung Alstroemeria in Chile. Miit. der Botanischen Staatssam. München 24: 1-362.
Bayer, E. 1998. Taltalia- eine neue Gattung in der Familie der Alstroemeriaceae. Sendtnera 5: 5-14.
Buitendijk, J.H. 1998. A cytological characterization of genomes of Alstroemeria, the production of interspecific hybrids, and their performance during micropropagation. Ph.D. Thesis. Wageningen.The Netherlands.
Buitendijk, J.H. and Ramanna, M.S. 1996. Giemsa C-banded karyotypes of eight species of Alstroemeria L. and some of their hybrids. Annals of Botany 78: 449-457.
Griffin, M. 1995. Problemas para as Tulipas. Agroanalysis 15 (9): 12-14. Hunziker, A.T. 1973. Notas sobre Alstroemeriaceae. Kurtziana 7: 134. Hunziker, J.H. 1991. Protandry in Alstroemeria psittacina (Alstroemeriaceae). Polish
Botanical Studies 2: 195-198. Hunziker, J.H. and Xifreda, C.C. 1991. Chromosome studies in Bomarea and
Alstroemeria (Alstroemeriaceae). Darwiniana 30 (1-4): 179-183. Meerow, A.W., Tombolato, A.F.C. and Meyer, F. 1999. Two new species of Alstroemeria
L. (Alstroemeriaceae) from Brazil. Brittonia 51: 429-444. Miyake, I. 1989. Breeding spotless Alstroemeria in Japan. Herbertia 45: 40-44. Øogaard, M. and Kristiansen, K. 1998. Alstroemeria in Chile. Herbertia 53: 29-39. Sanso, A.M. 1996. El género Alstroemeria (Alstroemeriaceae) en Argentina. Darwiniana
34: 349-382. Sanso, A.M. 2002. Chromosome studies in Andean taxa of Alstroemeria
(Alstroemeriaceae). Bot. Journ. of Linn. Soc. 138: 451- 459. Sanso, A.M. and Hunziker, J.H. 1998. Karyological studies in Alstroemeria and Bomarea
(Alstroemeriaceae). Hereditas 129: 67-74. Sanso, A.M. and Xifreda, C.C. 1998. Comparative foliar anatomy in Alstroemeriaceae.
Poster presented at Monocots II, Second Intl. Symposium on the Comparative Biol. of the Monocotyledons. Sydney. Australia.
Sanso, A.M. and Xifreda, C.C. 1999. The synonymy of Schickendantzia with Alstroemeria (Alstroemeriaceae). Systematics and Geography of Plants 68: 315-323.
Sanso, A.M. and Xifreda, C.C. 2001. Generic delimitation between Alstroemeria andBomarea (Alstroemeriaceae). Annals of Botany 38: 1057-1069.
Sanso, A.M. and Xifreda, C.C. 2003. Comparative foliar anatomy in Alstroemeriaceae. Submitted to Bot. Journ. Linn. Soc.
Sanso, A.M. and Xifreda, C.C. Alstroemeriaceae. Flora de Paraguay. Edit. Conservatoire et Jardin Botaniques de la Ville de Génève- Missouri Botanical Gardens. In preparation.
71
Figures
Fig. 1. Distribution of the genus Alstroemeria.

72
Fig. 2. General aspect. A-B, subterranean organs, A, vegetative shoot, B, reproductive and vegetative shoots; C-F, flowers, C, tubular perianth, D, more or less actinomorph; E, reflex perianth, F, zygomorphic,; G-H, fruits, G, with a long umbo, H, with a short umbo; I-J, seeds. A, C, H, I, A. isabellana; B, D, J, A.patagonica; E, A. andina subsp. venustula; F, G, A. pseudospathulata. Drawings from Sanso (1996).
73
Fig.
3. C
aulin
ar a
nato
my.
A-C
, ste
m tr
ansv
ersa
l sec
tion,
A, g
ener
al v
iew
; B, d
etai
l of t
he s
chle
renc
hym
a sh
eath
and
the
rela
tion
with
the
oute
r and
inne
r vas
cula
r bun
dles
; C, d
etai
l of e
pide
rmis
, cor
tical
par
ench
yma,
sch
lere
nchy
mat
ous
shea
th a
nd o
ne v
ascu
lar b
undl
e.A
-C,A
. isa
bella
na. e
= ep
ider
mis
, c=
corti
cal p
aren
chym
a, ss
(and
arr
ow)=
sche
renc
hym
atou
s she
ath,
vb=
vas
cula
r bun
dle.

74Fig.
4. F
olia
r ana
tom
y. A
-C, l
eaf t
rans
vers
al se
ctio
n, g
ener
al v
iew
, A. i
sabe
llana
.
75
Fig.
5. S
EM m
icro
grap
hs o
f pol
len.
A, p
olle
n gr
ains
on
stig
mat
ic su
rfac
e, B
, det
ail o
f the
stria
te- r
etic
ulat
e se
xine
, C, D
, gen
eral
vie
w o
f po
llen
grai
ns. A
-C, A
. isa
bella
na, D
, A. p
sitta
cina
.
76
Fig. 6. SEM micrographs of seed. A-E, general view, B, E, chalazal region, D, micropilar region; F, detail of ornamental units. A, A. patagonica, B, A. presliana, C, A. psittacina; D-F, A. isabellana.

77
Fig. 7. Photographs of some Alstroemeria species from Brazil. A, A. longistaminea; B, A. sellowiana; C, A. foliosa; D, A. speciosa; E, A. variegata; F, A. plantaginea; G, A.gardneri; H, A. malmeana; I, A. stenophylla.




![ΓΕΝΙΚΗ ΦΥΤΟΠΑΘΟΛΟΓΙΑ ... · Genus species Albugo candida –(Λευκή ... Microsoft PowerPoint - ERGASTIRIO.5_OOMYCETES.ppt [Λειτουργία συμβατότητας]](https://static.fdocument.org/doc/165x107/5ac2987b7f8b9a1c768e30a9/-species-albugo-candida-.jpg)

![SIEGEL MODULAR FORMS OF GENUS AND LEVEL - arXiv · valued modular forms on the full congruence subgroup Γ[2] of Sp(2,Z) of level 2 together with the action of S6 ∼= Sp(2,Z/2Z)](https://static.fdocument.org/doc/165x107/5f5af4e3fefc036b8e64815e/siegel-modular-forms-of-genus-and-level-arxiv-valued-modular-forms-on-the-full.jpg)



![Total perfect codes in Cayley graphs - arXivin the community of graph theory; see [21, 26, 27, 30, 35, 41] for example. Perfect codes in Cayley graphs are especially charming objects](https://static.fdocument.org/doc/165x107/603fafecff4ef36b9b49103d/total-perfect-codes-in-cayley-graphs-arxiv-in-the-community-of-graph-theory-see.jpg)







