

AEM Accepts, published online ahead of print on 20 June...
44
1 Biochemical and structural characterization of a thermostable family GH62 α-L- 1 arabinofuranosidase from Streptomyces thermoviolaceus to elucidate the molecular basis for 2 activity towards arabinoxylan 3 4 5 Running title: Structure functional analysis of SthAbf62A 6 7 Weijun Wang 1 , Galina Mai-Gisondi 2 , Peter J. Stogios 1 , Amrit Kaur 1 , Xiaohui Xu 1 , Hong Cui 1 , Ossi 8 Turunen 2 , Alexei Savchenko 1 , Emma R. Master 1, 2# 9 10 11 1 Department of Chemical Engineering and Applied Chemistry, University of Toronto, 200 College 12 Street, Toronto, Ontario, M5S 3E5, Canada. 13 2 Department of Biotechnology and Chemical Technology, Aalto University, Kemistintie 1 D1, Espoo 14 02150, Finland. 15 16 #Corresponding Author 17 Emma R. Master 18 Telephone: 416-946-7861 19 Fax: 416-978-8605 20 Email: [email protected] 21 22 23 AEM Accepts, published online ahead of print on 20 June 2014 Appl. Environ. Microbiol. doi:10.1128/AEM.00685-14 Copyright © 2014, American Society for Microbiology. All Rights Reserved. on August 12, 2019 by guest http://aem.asm.org/ Downloaded from
Transcript of AEM Accepts, published online ahead of print on 20 June...

1
Biochemical and structural characterization of a thermostable family GH62 α-L-1
arabinofuranosidase from Streptomyces thermoviolaceus to elucidate the molecular basis for 2
activity towards arabinoxylan 3
4
5
Running title: Structure functional analysis of SthAbf62A 6
7
Weijun Wang1, Galina Mai-Gisondi2, Peter J. Stogios1, Amrit Kaur1, Xiaohui Xu1, Hong Cui1, Ossi 8
Turunen2, Alexei Savchenko1, Emma R. Master 1, 2# 9
10
11
1Department of Chemical Engineering and Applied Chemistry, University of Toronto, 200 College 12
Street, Toronto, Ontario, M5S 3E5, Canada. 13
2Department of Biotechnology and Chemical Technology, Aalto University, Kemistintie 1 D1, Espoo 14
02150, Finland. 15
16
#Corresponding Author 17
Emma R. Master 18
Telephone: 416-946-7861 19
Fax: 416-978-8605 20
Email: [email protected] 21
22
23
AEM Accepts, published online ahead of print on 20 June 2014Appl. Environ. Microbiol. doi:10.1128/AEM.00685-14Copyright © 2014, American Society for Microbiology. All Rights Reserved.
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

2
ABSTRACT 24
Xylan debranching enzymes facilitate the complete hydrolysis of xylan and can be used to alter 25
xylan chemistry. Herein, the GH62 family α-L-arabinofuranosidase from Streptomyces thermoviolaceus 26
(SthAbf62A) was shown to have a half-life of 60 min at 60 °C, and ability to cleave α-1,3 L-27
arabinofuranose (L-Araf ) from singly-substituted xylopyranosyl (Xylp) backbone residues in wheat 28
arabinoxylan; low activity towards arabinan as well as 4-nitrophenyl α-L-arabinofuranoside was also 29
detected. After selectively removing α-1,3 L-Araf substituents from di-substituted Xylp residues present 30
in wheat arabinoxylan, SthAbf62A could also cleave the remaining α-1,2 L-Araf substituents, 31
confirming the ability of SthAbf62A to remove α-L-Araf residues that are (1→2) and (1→3) linked to 32
mono-substituted β-D-Xylp sugars. Three-dimensional structures of SthAbf62A and its complex with 33
xylotetraose and L-arabinose confirmed a five-bladed β-propeller fold and revealed a molecular Velcro 34
in blade V between the β1 and β21 strands, a disulfide bond between Cys 27 and Cys 297, and a 35
calcium ion coordinated in the central channel of the fold. The enzyme-arabinose complex structure 36
further revealed a narrow and seemingly rigid L-arabinose binding pocket situated at the center of one 37
side of the β propeller, which stabilized the arabinofuranosyl substituent through several hydrogen-38
bonding and hydrophobic interactions. The predicted catalytic amino acids were oriented towards this 39
binding pocket and the catalytic essentiality of Asp53 and Glu213 was confirmed by site-specific 40
mutagenesis. Complex structures with xylotetraose revealed a shallow cleft for xylan backbone binding 41
which is open at both ends and comprises multiple binding subsites above and flanking the L-arabinose 42
binding pocket. 43
44
45
46
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

3
INTRODUCTION 47
Xylan is a major plant cell wall polysaccharide that has been harnessed to produce different 48
chemicals, fuels and materials; the bioactive properties of xylans are also valued in food applications 49
(1-3). Xylan consists of a linear β-1,4-linked xylopyranosyl (β-D-Xylp) backbone, which depending on 50
the source, can be partially substituted at O-2 positions and/or O-3 positions with α-L-arabinofuranosyl 51
(α-L-Araf) substituents and acetyl groups, and at O-2 positions with α-D-glucuronic acid (GlcA) or 4-52
O-methyl GlcA (MeGlcA) (4-6). The O-5 position of some arabinose residues can be further esterified 53
with p-coumaric or ferulic acid (7, 8). Whereas glucuronoxylan is the predominant form of xylan in 54
hardwood trees, xylan from coniferous softwoods contains both arabinofuranosyl and MeGlcA 55
substituents (9); xylans from cereals and grasses resemble softwood xylan although have varying 56
arabinose and MeGlcA contents. Consistent with the molecular diversity of xylans, its complete 57
enzymatic hydrolysis typically requires the concerted action of several enzymes, including endo-58
xylanases and β-xylosidases that target glycosidic linkages within the xylan backbone, along with α-L-59
arabinofuranosidases, α-glucuronidases, acetyl xylan esterases, feruloyl esterases, as well as 60
glucuronoyl esterases, which target branching substituents in different xylan types (10-12). Of these 61
activities, arabinofuransidases are represented in a particularly high number of carbohydrate-active 62
enzyme (CAZy) families (www.cazy.org). 63
Based on amino acid sequence, α-L-arabinofuranosidases (EC 3.2.1.55) have been classified into 64
glycoside hydrolase (GH) families: GH3, GH43, GH51, GH54, and GH62. Moreover, family GH1 65
includes exo-acting arabinofuranosidases acting on p-nitrophenyl-α-arabinofuranoside (pNP-AraF) and 66
α-1,5 arabino-oligosaccharides (13); family GH30 includes activities towards 1,5- α- L-arabinobiose 67
(14), and family GH93 includes exo-acting enzymes that release arabinobiose from the non-reducing 68
end of α-1,5- L-arabinan (15). Several of these enzyme families are further grouped into glycoside 69
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

4
hydrolase clans: GH-A (GH1, GH30 and GH51), GH-E (GH93) and GH-F (GH43 and GH62), 70
reflecting the structural diversity of α- L-arabinofuranosidases. For example, the catalytic domain of 71
enzymes belonging to Clan GH-A adopt a (β/α)8 TIM barrel fold as exemplified by the structure of the 72
GH51 α- L-arabinofuranosidase from Geobacillus stearothermophilus T-6 (16). By contrast, Clan GH-73
E enzymes adopt a six-bladed β-propeller architecture (15), GH43 enzymes in Clan-GH-F display a 74
'non-Velcroed' five-bladed-β-propeller arrangement (17, 18), and the catalytic domain of the GH54 α- 75
L-arabinofuranosidase from Aspergillus kawachiia adopts a β-sandwich fold (19). 76
α- L-Arabinofuranosidases also vary in terms of substrate preference and the linkages they 77
hydrolyze. For example, GH51 as well as GH54 enzymes include α- L-arabinofuranosidases able to 78
remove branching α-L-Araf moieties from arabinan and xylans, although in most cases so far, enzymes 79
from these families tend to prefer p-nitrophenyl-α-arabinofuranoside (pNP-AraF) over polymeric 80
substrates (20-23). GH3 arabinofuranosidases include bifunctional enzymes displaying both α-L-81
arabinofuranosidase and β-D-xylopyranosidase activity when using pNP substrates (24). By comparison, 82
family GH43 includes a broad range of activities, including hydrolysis of pNP-AraF (25), endo-α-L-83
arabinanases (26, 27), exo-α-1,5-L-arabinanases (28), and bifunctional α-L-arabinofuranosidase/β-D-84
xylosidases (29). Family GH43 also includes arabinoxylan α-L-arabinofuranohydrolases (AXH) that 85
remove α-L-Araf residues that are (1→2) and (1→3) linked to mono-substituted β-D-Xylp (AXH-m 2,3) 86
(30), and that cleave α-L-Araf residues that are (1→3) linked to di-substituted β-D-Xylp (AXH-d3) (31). 87
Recently, the first 3-D structures of GH62 enzymes were published, one from Ustilago maydis 88
(UmAbf62A), one from Podospora anserina (PaAbf62A), and one from Streptomyces coelicolor 89
(ScAraf62A) (32, 33). These studies confirmed that GH62 enzymes adopt a five-bladed β-propeller 90
fold common to GH43 enzymes (32). While these studies generated important insight into structure 91
and functional characteristics of GH62 enzymes, there are comparatively few examples of detailed 92
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

5
biochemical and structural analyses of this GH family. Given the high activity of GH62 enzymes on 93
polymeric arabinoxylan over low molecular substrates including pNP-AraF (32-38), GH62 α-L-94
arabinofuranosidases are particularly relevant candidates for fine tuning xylan chemistry as well as 95
enabling the complete saccharification of xylan to sugars. 96
Accordingly, to deepen our understanding of GH62 enzymes, we report the biochemical and 97
structural characterization of a thermostable, bacterial GH62 α-L-arabinofuranosidase (SthAbf62A) 98
from Streptomyces thermoviolaceus subsp. pingens Henssen, which has 67%, 58% and 31% amino acid 99
sequence identity to ScAraf62A, UmAbf62A and PaAbf62A, respectively. Consistent with the first 100
report of SthAbf62A by Tsujibo et al (37), SthAbf62A characterized herein released reducing sugars 101
from polymeric arabinoxylan. The current analysis further demonstrated that SthAbf62A can cleave α-102
L-Araf substituents that are (1→2) and (1→3) linked to mono-substituted β-D-Xylp in arabinoxylan and 103
can also hydrolyze non-reducing terminal α-1,5 linkages in debranched arabinan as well as pNP-104
arabinofuranoside, albeit at much lower specific activities. Structural analysis of SthAbf62A 105
complexes with L-arabinose and xylotetraose revealed a “Velcroed” five-bladed-β-propeller, and 106
provided insight into the molecular basis of substrate selectivity for this enzyme. 107
108
MATERIALS AND METHODS 109
Chemicals. Pfx polymerase was purchased from Invitrogen. The Qiagen stool kit, QIAEX II 110
Gel Extraction Kit, and QIAquick PCR purification kit, were purchased from QIAGEN. Wheat 111
arabinoxylan [high viscosity (WAX-HV) and low viscosity (WAX-LV)], rye flour arabinoxylan (RAX), 112
arabinan from sugar beet, debranched arabinan, rhamnogalacturonan (soy bean), larch arabinogalactan 113
and α-L-arabinofuranosidase from Bifidobacterium sp (E-AFAM2, AXH-d3) were purchased from 114
Megazyme, whereas oat spelt xylan (OSX), birchwood xylan, beechwood xylan and p-nitrophenyl-115
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

6
glycosides were from Sigma-Aldrich. Soluble and insoluble fractions of OSX were prepared as 116
previously reported (39). Streptomyces thermoviolaceus subsp. pingens Henssen (ATCC#19283) was 117
purchased from the American Type Culture Collection (ATCC). All other chemicals were analytical 118
grade and were obtained from Sigma-Aldrich or Fisher Scientific. 119
DNA Manipulations. Streptomyces thermoviolaceus subsp. pingens Henssen was grown in 5 120
mL Luria-Bertani medium at 37°C for 4 days. Genomic DNA was extracted using the Qiagen stool kit 121
according to the manufacturer’s instructions and including a 5 min incubation at 95°C followed by an 122
additional bead beating step using Garnet bead tubes (Mo Bio, 0.70 mm). The gene encoding the mature 123
form of SthAbf62A (Genbank ID: KF958299) without a signal peptide (aa residues 1-35) was 124
amplified using the forward and reverse primers shown in Table S1 and the Pfx DNA polymerase 125
(Invitrogen). The infusion cloning kit from Clontech (California, USA) was used to transfer purified 126
PCR products to the p15Tv-L expression plasmid (GenBank accession EF456736), generating p15Tv-127
L_SthAbf62A. Site-specific mutagenesis was carried out according to a modified QuikChangeTM 128
(Stratagene) method that uses partially overlapping primers (Table S1) (40). All constructs were 129
verified by DNA sequencing at the Center of Applied Genomics at the SickKids Hospital in Toronto. 130
Purification of wild-type and mutant α-L-arabinofuranosidase. Recombinant Escherichia 131
coli BL21(λDE3) codon plus harboring p15Tv-L_SthAbf62A was propagated at 37 °C in 1 L of Luria-132
Bertani medium supplemented with 0.5 M d-sorbitol, 2.5 mM glycine betaine, 34 μg/mL 133
chloramphenicol and 100 μg/mL ampicillin. At OD600nm of 0.6, the cultivation temperature was 134
reduced to 15 °C and recombinant protein expression was induced with 0.5 mM isopropyl β-d-135
thiogalactopyranoside. 136
The next day, induced cultures were harvested by centrifugation at 6000 × g for 10 min. Cell pellets 137
(approx. 4.5 g fresh weight) were suspended in binding buffer [300 mM NaCl, 50 mM 4-(2-138
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

7
hydroxyethyl)-1-piperazinepropane sulfonic acid (HEPES), pH 7.0, 5% glycerol, 5 mM imidazole] and 139
disrupted by sonication. Cell debris was removed by centrifugation (17,500 × g, 20 min) and 140
supernatants were passed through a 0.45-μm filter before being incubated with 5.0 mL bed volume of 141
Ni-NTA resin (Ni2+-nitrilotriacetate, Qiagen) for 45 min at 4°C. Resin samples were then washed with 142
200 mL of washing buffer (300 mM NaCl, 50 mM HEPES, pH 7.0, 5% (v/v) glycerol, 50 mM 143
imidazole) and bound protein was eluted with approximately 30 mL of elution buffer (300 mM NaCl, 144
50 mM HEPES, pH 7.0, 5% v/v glycerol, 250 mM imidazole). Active fractions were pooled, 145
exchanged to 20 mM HEPES buffer (pH 7.0) using a Bio-Gel® P10 column, and then further purified 146
by anion chromatography using the Uno-Q column (5.0 mL) from BioRad. A BIOSHOP Duoflow 147
(Bio-Rad, Canada) system was used to pass 20 mM HEPES buffer (pH 7.0) with a NaCl gradient from 148
0 to 0.5 M through the Uno-Q column over 20 column volumes, and SthAbf62A wild-type as well as 149
mutant enzymes were eluted at 0.15 M NaCl. A Bio-Gel P10 column was then used to transfer the 150
purified protein to 20 mM HEPES buffer (pH 7.0) containing 300 mM NaCl, and centrifugal filter units 151
(10K; Millipore) were used for concentrating protein. Protein aliquots were flash frozen in liquid 152
nitrogen and then stored at -80°C. 153
Protein concentrations were determined using the Bradford assay (41) and bovine serum albumin as 154
a standard. SDS-PAGE was performed and stained with Coomassie Blue according to established 155
procedures. 156
Enzyme activity assays. Hydrolysis of arabinoxylans was monitored by measuring released 157
reducing sugars using the Nelson- Somogyi procedure (42). The standard assay solution contained 0.5% 158
(w/v) (5.0 mg/mL) polysaccharide in 0.5 mL of 100 mM HEPES buffer (pH 7.0). The reaction was 159
initiated by adding an amount of enzyme determined to release products in a linear relation to time 160
when incubated at 45°C for up to 30 min, which were also the temperature and incubation time used to 161
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

8
measure enzyme activities. One unit of the activity was defined as the amount of enzyme releasing 1 162
μmoL of L-(+)-arabinose equivalent per minute. L-(+)-Arabinose was used to generate a standard curve 163
(0.05-0.6 mg/mL). 164
SthAbf62A activity on 4-nitrophenyl (pNP) glycosides was determined by measuring pNP release. 165
The typical reaction mixture contained 2.0 mM substrate in 0.30 mL of 100 mM HEPES buffer (pH 166
7.0). After initiating the reaction by adding enzyme, the reaction mixture was incubated for 20 min at 167
45°C. The reaction was terminated by adding 0.30 mL of 4% (w/v) Na2CO3. The amount of pNP 168
formed was measured at A405nm (extinction coefficient ε 405 = 17,600 M−1 cm−1) and pNP was used to 169
generate a standard curve. As also indicated above, the enzyme added to each assay was optimized to 170
measure initial reaction rates. All enzyme assays were carried out in triplicate. 171
Kinetic parameters were determined using 0.1 mM to 25 mM pNP-AraF; apparent kinetic 172
parameters were also obtained with 0.05% to 1.5% (0.5 to 15 mg/mL) WAX-HV, and 0.1% to 15% (1.0 173
to 150 mg/mL) sugar beet arabinan. Initial rates were obtained by measuring reaction products formed 174
after 30 min at 45°C and pH 7.0 as described in the standard assay. Kinetic parameters were then 175
calculated using the Michaelis-Menten equation or conventional substrate inhibition equation (Eq.1) 176
using GraphPad Prism 5.0 (GraphPad Software, USA): 177
178
(Eq. 1) 179
180
Where v is the initial velocity at various substrate concentrations and Km(app) is the apparent Michaelis 181
constant for substrate. Ksi (app) represents the apparent dissociation constant for the inhibitory SES 182
ternary complex (43). 183
Optimum reaction conditions, enzyme stability, and effects of divalent ions. All enzyme 184
)][
1(][
][
)()(
max
appsiappm K
SSK
SVv
+⋅+
⋅=
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

9
assays were performed using the standard activity assay with 0.5% (w/v) WAX-HV and incubation at 185
45°C for 30 min; reaction products were then detected using the Nelson-Somogyi method. The effect 186
of pH on SthAbf62A activity was determined by performing the activity assay at pH 3.5 to 10.5 with 187
increments of 0.5 pH units; a universal buffer (100 mM acetic acid, 100 mM boric acid and 100 mM 188
phosphoric acid, adjusted to the target pH using sodium hydroxide solution) was used for this analysis. 189
The effect of reaction temperature on enzyme activity was examined by performing the standard assay 190
at temperatures ranging from 5°C to 85°C. To determine pH and temperature stability, the enzyme was 191
pre-incubated in the universal buffer (pH 4 to 10.5) at 4°C for 24 h, or else at 25°C to 80°C for 60 min 192
in 100 mM HEPES buffer (pH 7.0), before being adjusted to standard assay conditions to measure 193
residual SthAbf62A activity. The effect of metal ions was determined by adding 2.0 mM of different 194
metal chloride salts to the standard activity assay. 195
High pressure anion exchange chromatography (HPAEC) detection of reaction products. 196
To confirm the selective hydrolysis of arabinose substituents, products of WAX-HV and sugar beet 197
arabinan from treatment with SthAraf62A were analyzed by HPAEC (Dionex, ICS-5000+ DC) 198
equipped with a pulsed amperometric detector (PAD), Carbopac™ PA1 column (250×2 mm, Dionex) 199
and CarboPac™ PA20 guard column (30×3 mm, Dionex) (44). Reactions (0.5 mL) contained 0.5% 200
(w/v) of WAX-HV or arabinan and SthAbf62A (2.5 μg for WAX-HV and 12.0 μg for arabinan), as 201
described in the standard activity assay. After 15 min, 30 min and 45 min of incubation, reactions were 202
stopped by boiling for 10 min; 100 µL samples were then mixed with 2 volumes of acetone to 203
precipitate polymeric arabinoxylan, and centrifuged at 13,000 x g for 10 min at 4°C. Supernatants 204
containing the liberated sugars were transferred to a clean tube and the acetone was removed using a 205
Speedvac (Thermo). The remaining supernatant was lyophilized, dissolved in 100 µL ultra-pure water 206
(MilliQ), and then filtered through 0.22 µm centrifuge tube filters (Corning). Samples (10 µL) were 207
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

10
injected on the Carbopac™ PA1 column kept at 30°C, and eluted with 50 mM NaOH at 0.25 mL/ min 208
for over 30 min. L-Arabinose and D-xylose (40 µg/ ml) were used as standards. 209
Preparation of wheat arabinoxylan with singly substituted L-arabinose. Arabinoxylan with 210
singly substituted α-1,2 and 1,3-arabinose was generated by selective removal of α-1,3-arabinose from 211
doubly substituted xylose residues using the arabinofuranosidase from Bifidobacterium sp. (AXH-d3) 212
(36). Briefly, 0.2% (w/v) of WAX-HV was treated with 5 U of AXH-d3 in 50 mL 20 mM potassium-213
phosphate buffer (pH 6.0) at 40°C for 24 h; an additional 2 U of AXH-d3 was then added and the 214
reaction was incubated for another 24 h to ensure near complete removal of the α-1,3-arabinose from 215
doubly substituted positions. After boiling for 10 min, arabinoxylan containing singly substituted α-1,2 216
and 1,3-arabinose was precipitated by adding two volumes of 95% ethanol followed by centrifugation 217
(15,000 × g, 10 min). The pellet was dissolved in 50 mL water and again precipitated with 95% ethanol. 218
The precipitate was then dissolved in 20 mL water and lyophilized. 219
1H NMR analysis for the action mode of SthAbf62A towards arabinoxylan. To 220
qualitatively evaluate the regioselectivity of SthAbf62A on arabinoxylan, one-dimensional 1H NMR 221
spectra were collected for intact and AXH-d3 treated WAX-HV before and after treatment with 222
SthAbf62A. Briefly, 1% (w/v) arabinoxylan (intact or singly substituted) was digested for 2 h or 24 h at 223
45°C using 3.0 µg of SthAbf62A in a 0.6 mL reaction solution containing 100 mM HEPES buffer (pH 224
7.0). Reactions were terminated by boiling for 10 min. The treated arabinoxylan was then precipitated 225
by adding 1.2 mL of 95% ethanol and separated by centrifugation (15,000 × g, 10 min). The pellet was 226
suspended in 0.6 mL water and again precipitated with 95% ethanol. To overcome the precipitation of 227
arabinoxylan following treatment with arabinofuranosidases (45, 46), a GH11 endoxylanase from 228
Thermobifida fusca, TfxA(47), was used to hydrolyze the debranched arabinoxylan into arabino-229
xylooligosaccharides (48). The pellet was suspended in 0.6 mL of 10 mM 3-(N-morpholino) 230
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

11
propanesulfonic acid (MOPS) buffer (pH 7.0) and then treated with 1.2 µg TfxA at 50°C. After 24 h, 231
an additional 1.2 µg TfxA was added and the reaction was incubated for another 24 h. Reaction 232
mixtures were subsequently boiled for 15 min to inactivate the endoxylanse, then lyophilized and 233
dissolved in 0.6 mL D2O two times before NMR analysis. 1H NMR spectra were obtained at 25°C in 5.0 234
mm NMR tubes (Norell) using an Agilent DD2 600 MHz spectrometer, with a scan number of 16, and 235
relaxation delay of 10 s. The data were recorded using VnmrJ 4.0 (Agilent) and processed using 236
MestReNova 8.1.0 software (Mestrelab Research). The methylene protons at position 2 of MOPS 237
(2.035 ppm) were used as the internal reference. The NMR signal assignment was based on previously 238
published reports (36, 49). 239
Protein crystallization, data collection and structure determination. SthAbf62A was 240
concentrated to 18 mg/mL in 10 mM HEPES (pH 7.5) containing 500 mM NaCl. The crystallization 241
screen was performed by mixing 0.5 μL of protein with 0.5 μL of reservoir buffer using a Mosquito 242
crystallization robot. The SthAbf62A crystals were obtained by a sitting-drop vapour diffusion 243
technique at 25°C with a condition containing 0.1 M Bis-tris propane (pH 6.5) and 20% polyethylene 244
glycol monomethyl ether (5 K). For the co-crystallization, SthAbf62A (14 mg/mL) was first incubated 245
overnight at 4°C with xylotetraose and L-arabinose at a final concentrations of 5 mg/mL and 200 mM, 246
respectively. The co-crystallization reservoir buffer for xylotetraose contained 0.2 M sodium formate 247
and 20 % polyethylene glycol 3350, and that for L-arabinose contained 0.1 M HEPES (pH 7.5), 0.2 M 248
ammonium acetate, and 25% polyethylene glycol 3350. The crystals were grown at 25°C using the 249
sitting-drop vapour diffusion technique. The crystals were then flash frozen in a 100 K liquid nitrogen 250
stream using paratone-N oil as a cryoprotectant. Diffraction data for SthAbf62A apoenzyme was 251
collected at 100 K on a Rigaku HF-007 rotating anode with a Rigaku R-AXIS IV++ detector. 252
Diffractor data for SthAbf62A-arabinose and SthAbf62A-xylotetraose complexes were collected at the 253
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

12
Argonne National Laboratory, Structural Biology Center at the Advanced Photon Source, beamline 19-254
ID with an ADSC Quantum 315r detector. All diffraction data was processed in HKL3000 (50). The 255
SthAbf62A apoenzyme structure was solved by molecular replacement using the structure of a GH62 256
family α-L-arabinofuranosidase from Scytalidium thermophilum (unpublished) as the search model and 257
PHENIX.phaser (51). All models were refined using PHENIX and Coot (52) with 5% of observed 258
reflections set aside for Rfree calculations. Ligands were built into residual positive Fo–Fc density after 259
building the protein atoms. All geometries were verified with PHENIX.refine and the RCSB PDB 260
Validation server. Conformation of L-arabinose and nucleophilic water was verified by calculation of a 261
simulated annealing omit map, where these atoms were removed, followed by Cartesian simulating 262
annealing using Phenix.refine (default parameters). Average B-factor and bond angle/bond length rmsd 263
values were calculated using PHENIX. Table 3 lists data collection and final model refinement statistics. 264
Structure figures were prepared using PyMol (The PyMOL Molecular Graphics System. DeLano 265
Scientific; San Carlos, CA, USA: 2002). Structures were deposited to the Protein Databank under 266
accession codes 4O8N (SthAbf62A), 4O8O (SthAbf62A complexed with L-arabinose) and 4O8P 267
(SthAbf62A complexed with xylotetraose). 268
269
RESULTS AND DISCUSSION 270
General enzyme properties of SthAbf62A. The recombinant SthAbf62A was functionally 271
expressed in E. coli with an N-terminal His6-tag and purified to homogeneity with a typical protein 272
yield of ~12 mg protein per liter of bacterial culture. The molecular weight of recombinant SthAbf62A 273
estimated by SDS–PAGE was 36.5 kDa (Fig. S1), which is consistent with the predicted molecular 274
mass (38.3 kDa). The optimal pH of SthAbf62A activity on wheat arabinoxylan was approximately pH 275
7.0 (Fig. 1A), which is 2.0 pH units higher than previously reported for the identical α- L-276
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

13
arabinofuranosidase from Streptomyces thermoviolaceus OPC-520 (AB110643.1) (37). Though 277
initially surprising, this difference in pH optima could be explained by the comparatively long 278
incubation time (2 h) used in earlier reports to measure the pH optimum, together with the pH stability 279
of SthAbf62A (see below). For comparison, reported pH optima of other characterized GH62 α- L-280
arabinofuranosidases are: pH 6.0 for SliAraf62 from Streptomyces lividans (34), pH 5.5 for ScAraf62A 281
from S. coelicolor (33), pH 5.0 for PchAraf62 from Penicillium chrysogenum (36), pH 7.0 for 282
CcAraf62A from Coprinopsis cinerea (38), pH 5.0 for UmAbf62A from Ustilago maydis, and pH 5.0 283
for PaAbf62A from Podospora anserina (32). 284
Stability studies showed that SthAbf62A retained over 70% activity after 24 h at 4°C between pH 285
from 6.0 to 9.5 (Fig. 1B). By comparison, CcAra62A retains 80% activity after 1 h at 20°C between pH 286
7.0 to 8.5 (38), and ScAraf62A is stable for 1 h at 35°C between pH 7.0 and 9.0 (33). Further, 287
SthAbf62A showed highest activity at 55°C in a 30 min enzyme assay (Fig. 1C), and retained over 70% 288
and 45% of activity after 60 min at 55°C and 60°C, respectively (Fig. 1D). By contrast, PchAra62, 289
SliAra62, and ScAraf62A lost all activity after 60 min at 60°C (33, 34, 36), and the optimal reaction 290
temperature for both UmAbf62A and PaAbf62A is 37°C and 55°C respectively (32, 53). 291
Consistent with earlier reports (37), the presence of 2.0 mM Zn2+, Cd2+, Hg2+ and Cu2+ reduced 292
sugar release from WAX-HV by SthAbf62A by more than 70%, while 2.0 mM Ca2+ and Mg2+ activated 293
SthAbf62A by at least 60% (Fig. S2). However, addition of 2.0 mM ethylenediaminetetraacetic acid 294
(EDTA) did not significantly affect SthAbf62A activity, suggesting that the enzyme does not strictly 295
depend on ion supplementation despite the coordinated calcium ion observed in the SthAbf62A 296
structure (see below). 297
Substrate specificity of SthAbf62A. SthAbf62A activity was highest on WAX-HV and RAX, 298
followed by water-soluble OSX and WAX-LV (Table 1). This result is consistent with the considerably 299
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

14
higher arabinose-to-xylose ratio in WAX-HV and RAX (ranging from 0.5 to 0.7) (54, 55), compared to 300
WAX-LV (approx. 0.3) (54), and OSX (approx. 0.12) (56, 57). Notably however, when normalizing 301
wheat, rye and water soluble OSX to arabinose content, approximately 8 % (WAX-HV), 6 % ( WAX-302
LV and RAX) and 17 % (OSX) of the total arabinose present was released after 30 min by 0.4 μg 303
SthAbf62A in standard assay conditions. This suggests that arabinose in water-soluble OSX is 304
comparatively accessible to hydrolysis by SthAbf62A, which likely reflects the particular solubility of 305
this OSX preparation. More interestingly, similar extents of arabinose released from wheat and rye 306
arabinoxylan after 30 min suggests that the differences in occurrence of α-L-Araf (1→3) and α- L-Araf 307
(1→2) substituents in WAX-HV, WAX-LV and RAW (54) does not significantly affect initial rates of 308
SthAbf62A activity. 309
Though not detected during the characterization of α- L-arabinofuranosidase from Streptomyces 310
thermoviolaceus OPC-520 (37), SthAbf62A also released arabinose from sugar beet arabinan, which 311
consists of an α-1,5 linked L-arabinofuranose backbone with ca. 60% α-1,3 linked L-arabinofuranose 312
substitutions. HPAEC analyses further confirmed that L-arabinose was the only monosaccharide 313
released from arabinoxylan and arabinan (Fig. S3). Given the polydispersity of the arabinoxylan and 314
arabinan substrates, molar values of these substrates could not be accurately calculated, and so only 315
apparent kinetic parameters could be obtained to evaluate SthAbf62A activity in greater detail. These 316
analyses revealed that the apparent catalytic efficiency (kcat/KM) of SthAbf62A was nearly 70 times 317
higher on wheat arabinoxylan than sugar beet arabinan (Table 2). At the same time, the kinetic data 318
were best explained by the conventional substrate inhibition model (Eq.1), and apparent substrate 319
inhibition constants (Ksi (app)) were lower for arabinoxylan than arabinan (Table 2; Fig. S4). Since 320
sample viscosities did not perceptibly differ at maximum substrate concentrations, the lower Ksi(app) 321
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

15
value for arabinoxylan suggests preferred binding by SthAbf62A of xylan over arabinan backbone 322
structures. 323
Low but detectable activity was also observed towards rhamnogalacturonan, debranched 324
arabinan, as well as pNP-α- L-arabinofuranoside and pNP-α- L-arabinopyranoside (Table 1). Notably, 325
the catalytic efficiency (kcat/KM) of SthAbf62A on pNP-α- L-arabinofuranoside was ~330 times higher 326
than pNP-α- L-arabinopyranoside, which was mainly explained by differences in kcat and reveals the 327
high preference for furanose forms of α- L-arabinose. HPAEC analyses confirmed that L-arabinose was 328
the only product released from debranched α-1,5 linked arabinan (data not shown), and kinetic analyses 329
determined that the kcat for arabinan and pNP-α- L-arabinofuranoside were 30 and 140 times lower than 330
the apparent kcat for WAX-HV, respectively (Table 2). The clear preference of SthAbf62A for 331
polymeric substrates (i.e. arabinoxylan) is consistent with other GH62 members (32-36, 38), and 332
continues to distinguish GH62 enzymes from arabinofuranosidases identified in other GH families. 333
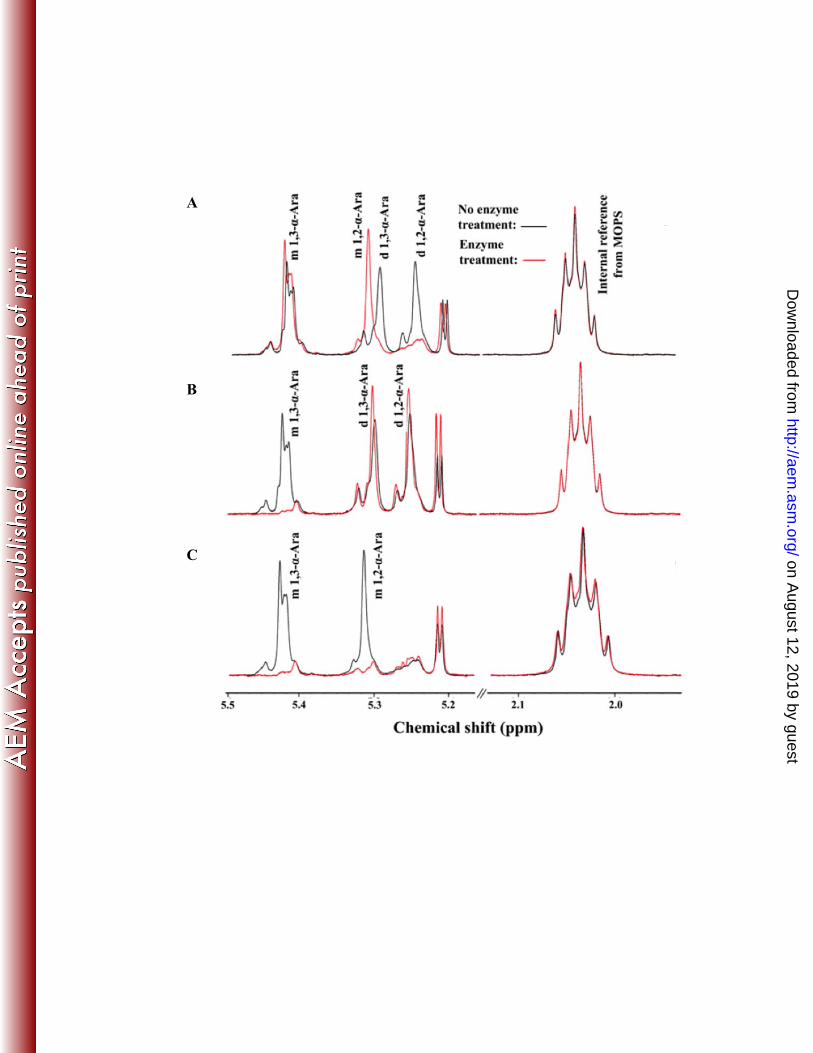
Regio-selectivity of SthAbf62A. 1H NMR analysis was used to then investigate the 334
regiospecificity of SthAbf62A towards α-L-Araf (1→2) and α-L-Araf (1→3) linkages in arabinoxylan. 335
Selective removal of α-L-Araf (1→3) linked to mono-substituted β-D-Xylp by SthAbf62A was shown 336
by the disappearance of the 1H signal at 5.42 ppm (red line; Fig. 2B) (49, 54, 57). To clarify whether 337
SthAbf62A activity was restricted to single α-1,3 linkages or else singly substituted xylose, 338
arabinoxylan with only single α-1,3 and α-1,2 arabinose substitutions was generated using 339
arabinofuranosidase (AXH-d3) from Bifidobacterium sp, which selectively removes α-L-Araf (1→3) 340
linked to di-substituted β-D-Xylp (36, 46). The effect of AXH-d3 treatment was confirmed by the 341
disappearance of signals near 5.28 ppm and 5.22 ppm corresponding the (1→3)-linked and (1→2)-342
linked α-L-Araf substituents of doubly substituted β-D-Xylp, and consequent increase in the signal near 343
5.30 ppm, which corresponds to α-L-Araf (1→2) linked to mono-substituted β-D-Xylp (Fig. 2A)(54). 344
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

16
Subsequent treatment with SthAbf62A significantly reduced signals at both 5.42 ppm and 5.30 ppm 345
(Fig. 2C), consistent with hydrolysis of both of α-1,3 and α-1,2 L-arabinofuranosyl linkages to singly 346
substituted xylose (i.e. AXH-m2,3 activity). This analysis shows that SthAbf62A alone selectively 347
removes singly substituted α-1,3 arabinose from wheat arabinoxylan, but similar to PchAraf62 (36) and 348
a GH43 AXH-m 2,3 from B. subtilis (30), can also target singly substituted α-1,2 linked arabinose after 349
enzymatic removal of α-1,3 L-arabinofuranosyl substituents from di-substituted xylopyranosyl residues. 350
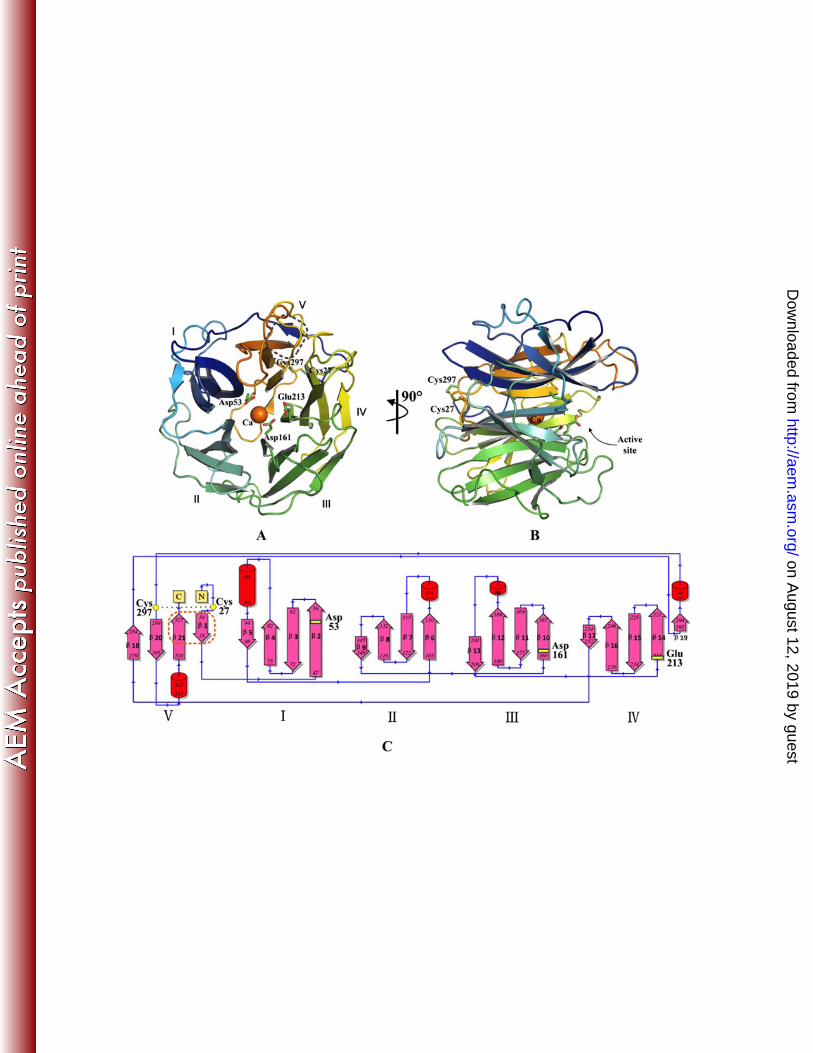
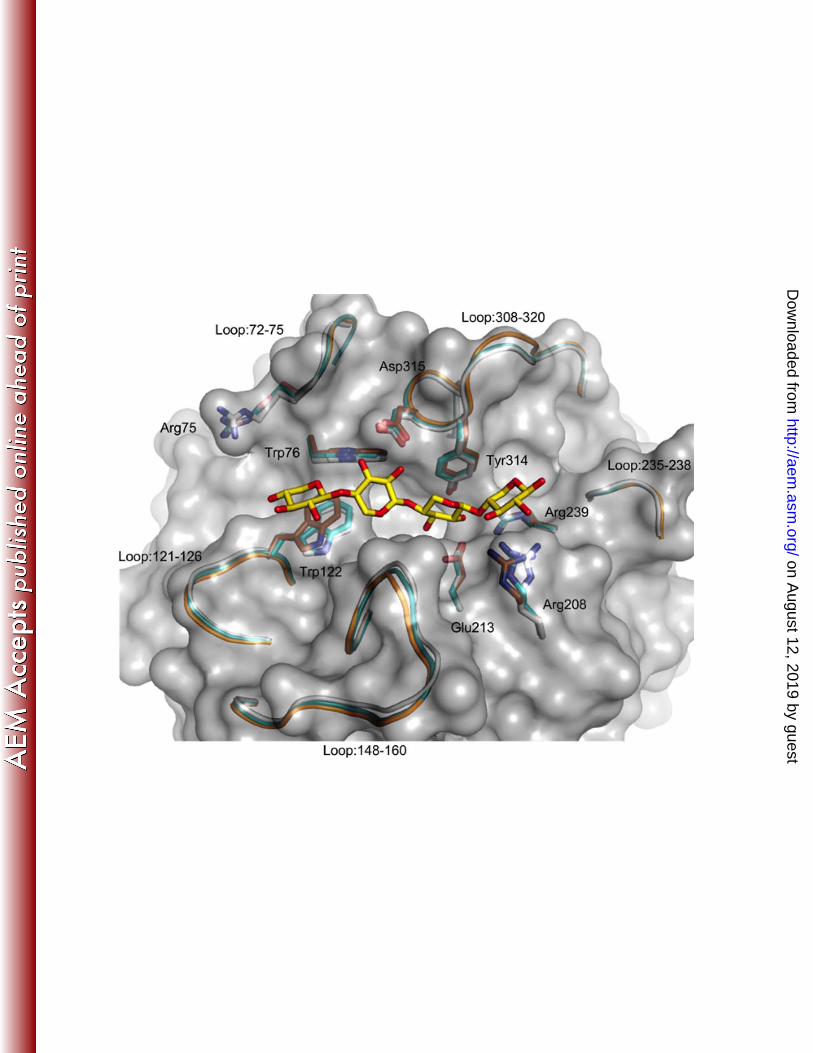
Overall structure. Three structures of SthAbf62A were determined by molecular replacement 351
(Table 3), which included one enzyme structure and two enzyme complex structures co-crystallized 352
with reaction products of xylotetraose and L-arabinose respectively (Protein Data Bank code: 4O8N, 353
4O8O and 4O8P) . The SthAbf62A enzyme adopts a β-propeller fold with five blades (marked I–V) 354
twisted and radially arranged around the pseudo five-fold axis. The blades are linked through loops 355
with variant length, and each blade was made up of four antiparallel β-strands in a typical W-like 356
topology with the first strand being the innermost (Fig. 3A). The only exception is blade IV, which was 357
formed by a typical four antiparallel β-strand arrangement (β14, β15, β16 and β17) plus the small strand 358
of β19 (Fig. 3C). This additional strand closes up one end of the β-propeller together with its flanking 359
loop region. 360
Structure similarity searches of the Protein Data Bank with the Dali Server showed that the 361
overall structure of SthAbf62A resembled that of enzymes in GH43, GH32 and GH68 families, which 362
similarly adopt a five-bladed β-propeller fold (Table S2) (18, 57-62). The overall structure of 363
SthAbf62A superimposed well with the catalytic domain of these enzymes (Table S2), which is 364
consistent with divergent evolution of enzymes from families GH32, 43, 62 and 68 (58). 365
Superimposition of SthAbf62A with enzymes from these families also facilitated the prediction of the 366
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

17
two catalytic residues (Asp53 and Glu213), and of a third residue (Asp161) believed to modulate the 367
pKa of the catalytic general acid (Fig. S5) (18) . 368
While we were completing our analysis of SthAbf62A, the first structures were published for 369
GH62 α-arabinofuranosidases, including UmAbf62A from Ustilago maydis, PaAbf62A from 370
Podospora anserina, and ScAraf62A from Streptomyces coelicolor (32, 33). In line with these first 371
structures of GH62 α-arabinofuranosidases, the five-bladed β-propeller fold of SthAbf62A is stabilized 372
through a molecular Velcro between antiparallel β-strands 1 and 21 in blade V formed through five 373
hydrogen bonds and several hydrophobic interactions (Fig. S6). Belonging to subfamily GH62_2, 374
SthAbf62A shared additional similarities to UmAbf62A and ScAraf62A. For instance, SthAbf62A 375
contains a disulfide bond between Cys27 and Cys297, which constrains the N-terminus of the enzyme 376
in the vicinity of the loop connecting strands 19 and 20 (Fig. 3). Additionally, a calcium ion in the 377
central channel of the fold was buried right under the L-arabinose binding pocket (Fig. 3, 4), displaying 378
a hepta-coordination with His280, Gln216 and five water molecules. 379
Since SthAbf62A shares over 65% sequence identity with ScAraf62A, but demonstrates 380
considerably higher optimal reaction temperature and temperature stability, these enzymes were further 381
compared in terms of 18 sequence characteristics thought to confer thermostability. Relative to 382
ScArab62A, SthAbf62A scored higher in 11 of these 18 parameters, including higher number of 383
predicted hydrogen bonding interactions and salt bridges, and increased core hydrophobicity (Table S3). 384
Though main determinants of thermostability are still difficult to predict, the growing number of GH62 385
enzyme structures, including those from temperature stable members, will undoubtedly facilitate future 386
protein engineering efforts aimed at improving applications of GH62 enzymes. 387
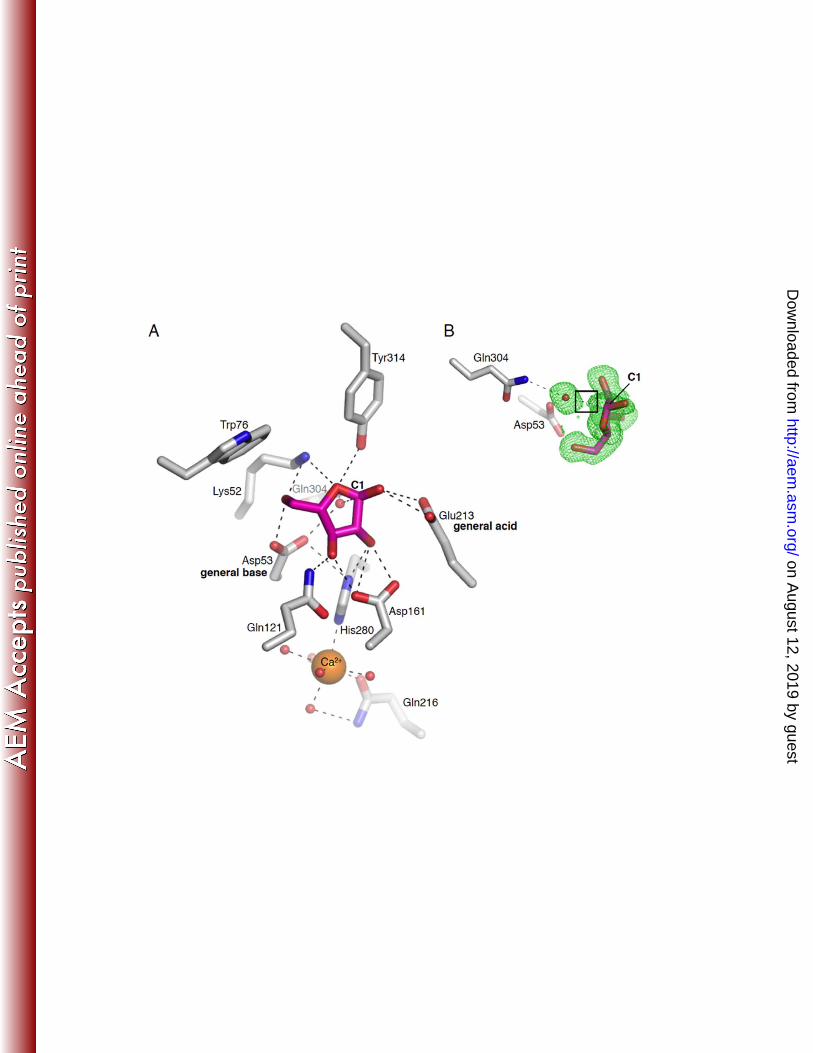
The active site of SthAbf62A. To understand the structural basis for catalysis and substrate 388
binding, the structure of SthAbf62A complexes with its reaction products was obtained by separate co-389
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

18
crystallizations with L-arabinose and xylotetraose. A bound L-arabinose molecule was observed in a 390
narrow and deep pocket (-1 subsite) composed of relatively conserved amino acids, which was open 391
toward the molecular surface (Fig. 3; Fig. S7). It was formed mainly by the five first and innermost β-392
strands of each blade (β2, β6, β10, β14 and β18) and lined by about 12 amino acids (within 4Å distance 393
from L-arabinose), 3 of which were acidic amino acid residues, making this pocket mainly negative 394
charged (Fig. S7). The two predicted catalytic residues (Asp53 and Glu213), along with the putative 395
pKa modulator (Asp161), were pointing into this pocket (Fig. 3A,B), and were donated by the β2 sheet 396
in blade I, β14 in blade IV and β10 in blade III, respectively (Fig. 3C). The Asp53Ala and Glu213Ala 397
substitutions led to complete loss of enzyme activity, whereas the Asp161Ala substitution retained 398
detectable, albeit low activity on WAX-HV (Table S4); the lost activity could not be rescued by NaN3. 399
The site-directed mutagenesis results reported herein support the catalytic importance of Asp53, Glu213 400
and Asp161, and are consistent with mutagenesis studies of ScAraf62A, which similarly implicate 401
corresponding Asp202, Glu361 and Asp309 residues in catalysis (33). 402
The L-arabinofuranose molecule was modeled with an α-configuration (Fig. 4). The ligand was 403
anchored in the L-arabinose binding pocket mainly through several polar interactions (Fig. 4), such as 404
hydrogen bonds between sugar ring O4 and Nζ of Lys52 (3.4 Å) along with Oηof Tyr314, O2 and Nδ1 of 405
His280 (2.9 Å) as well as Oδ1 and Oδ2 of Asp161 (2.7 Å), and O3 with Oδ1 of Asp161 (2.6 Å) and Nζ2 406
of Gln121 (2.8 Å). In particular, the contacts between O5 of L-arabinofuranose and Nζ of Lys52(2.8 Å) 407
as well as Oδ2 of Asp53 (2.6 Å) might be important for discriminating between furanose and pyranose 408
forms of L-arabinose as observed for pNP substrates (Table 2). The catalytic Glu213 side chain was 409
proximal to O1 of L-arabinose in a distance of 2.8 Å. All the residues involved in the above hydrogen 410
bonds are contributed by β sheets and conserved across the sequences in the GH62 family (Fig. S8). In 411
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

19
contrast, residues for hydrophobic interactions (i.e. Trp76, Tyr314, Ile160 and Thr68) were not highly 412
conserved, except Tyr314. Notably, in SthAbf62A, Trp76 along with Tyr314 appeared to contact the L-413
arabinose ring through edge on edge apolar interactions (59) that were further stabilized through 414
interactions with the side chains of Ile160 and Thr68. By comparison, stacking interactions in 415
UmAbf62A were predicted between arabinose and Tyr58 (32). Accordingly, binding of arabinose by 416
SthAbf62A appears more similar to ScAraf62A, which interacts with the flat surface of arabinose 417
through hydrophobic interactions with Ile308 (33). 418
The structure of SthAbf62A in complex with L-arabinofuranose revealed insights into the catalytic 419
mechanism of this enzyme. There was a water molecule closely positioned near the anomeric carbon 420
(2.8 Å) that also formed a hydrogen bond with the expected general base, Asp53 (Fig. 4). We also 421
observed electron density shared between this water molecule and the anomeric carbon (Fig. 4). Finally, 422
the distance between Asp53 and the expected general acid, Glu213, was 8 Å. These observations are all 423
consistent with an inverting mechanism for this enzyme, with the described water molecule acting as 424
the nucleophile for attack on the anomeric carbon. 425
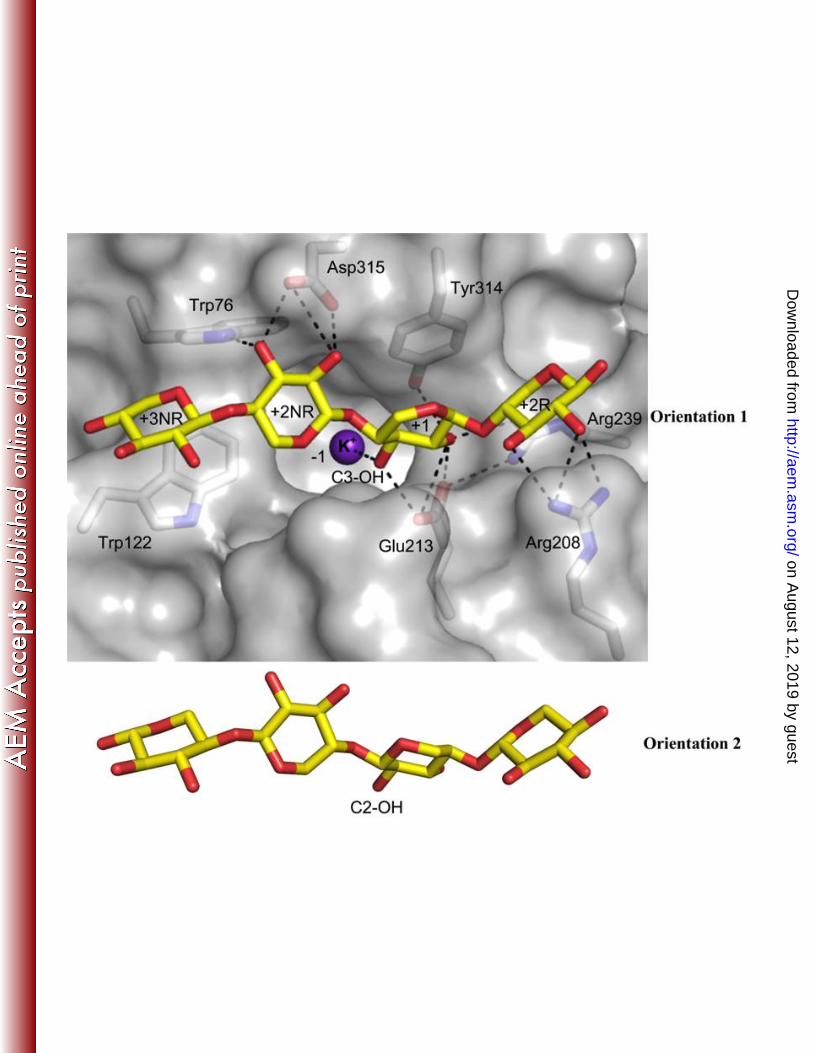
The co-crystallization of SthAbf62A with xylotetraose, a reaction product, revealed the binding 426
mode of the xylan backbone within an open cleft situated above the catalytic pocket. Four xylose 427
binding subsites were observed along the cleft; the xylotetraose chain fit the electron density features 428
equally well in two opposite orientations, which is similar to the model proposed for acetylxylan 429
esterases (60). The nomenclature described in McKee et al (61) was followed herein to name the 430
subsites that bound xylotetraose. In Orientation 1 of xylotetraose, the reducing end xylose at subsite 431
+2R situated above the space between blade IV and V, whereas the non-reducing end xylose at subsite 432
+3NR situated above the space between blade I and II (Fig. 5). Orientation 1 would allow hydrolysis of 433
α-1,3 linked arabinose since O3 of xylose points towards the active site pocket, while the reverse 434
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

20
orientation (Orientation 2) would favor the hydrolysis of α-1,2 linked arabinose. In both orientations, 435
xylose at subsite +1 was stabilized through hydrophobic stacking interactions with Tyr314 and 436
hydrogen bonds with Glu213 (Fig. 5; Table S5). The non-reducing sugar unit at subsite +3NR in 437
Orientation 1, and corresponding reducing end sugar at subsite +3R in Orientation 2, did not form any 438
hydrogen bonds with the enzyme and instead was stabilized via a stacking interaction with Trp122, 439
which is conserved in most sequences with occasional substitution to Gln (Fig. S8). Hydrogen-bonding 440
interactions observed between the xylotetraose backbone and the enzyme are listed in Table S5. 441
Notably, while the +2NR subsite in Orientation 1 and Orientation 2 is characterized by hydrogen bond 442
interactions with Trp76 and Arg208 respectively, Orientation 1 lacks the additional hydrogen bond 443
observed in Orientation 2 between Arg239 and the endocyclic oxygen of the nonreducing end xylose. 444
A cation, modeled as potassium, was bound in the arabinose-binding site (-1 subsite) in the SthAbf62A-445
xylotetraose complex and accordingly this ion could interact with the C3-OH (Orientation 1) or the C2-446
OH (Orientation 2) of xylose at subsite +1(Fig. 5). Taken together, the SthAbf62A substrate binding 447
cleft and active site adopt a “T-like” architecture (Fig. S9), reminiscent of that observed in two GH43 448
side chain-cleaving α-L-arabinofuranosidases (BsAXH-m2,3 (18) and HiAXH-d3 (61)), suggesting the 449
“T-like” architecture is a common feature of debranching glycoside hydrolases. 450
451
Conformation changes following ligand binding. Besides the aromatic residues described 452
above, the xylan binding cleft of SthAbf62A was mainly lined by non-conserved amino acids from loop 453
regions, which was also noted in BsAXH-m2,3, UmAbf62A, PaAbf62A and ScAraf62A (18, 32, 33), 454
suggesting that enzymes from this clan adopt a dynamic substrate binding cleft. Consistent with this 455
prediction, xylotetraose and L-arabinose binding induced small local conformation changes in 456
SthAbf62A (Fig. 6). Superimpositions between the SthAbf62A structure and enzyme complexes 457
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

21
showed pairwise root-mean-square deviation (RMSD) values of 0.362 Å (303 Cα atoms) and 0.173 Å 458
(302 Cα atoms) for xylotetraose and L-arabinose complex structures, respectively. 459
In the SthAbf62A-xylotetraose complex structure, the difference was attributed largely to 460
conformational differences between the five loop regions (72-75,121-126, 148-160, 235-238 and 308-461
320), which were all involved in forming the xylan backbone binding cleft. Indeed, the pairwise RMSD 462
value of Cα atoms computed using only these residues increased to 0.682 Å and 0.411 Å for 463
xylotetraose and L-arabinose complex structures, respectively, also indicating larger conformational 464
differences induced by xylotetraose binding. Movement within these loop regions shifted the side 465
chains of several amino acid residues, including Trp122, Asp315 and Tyr314 along with Trp76 and 466
Arg208 from β-sheets, all of which were involved in xylotetraose binding. Furthermore, the side chain 467
of Arg75 from loop 72-75 shifted towards the ligand by 2.3 Å upon xylotetraose binding (Fig. 6). Such 468
movement within the substrate binding cleft could explain the accommodation of arabinan substrates, 469
and non-catalytic interactions between GH62 arabinofuranosidases and various oligosaccharides, 470
including cello-oligosaccharides that can inhibit enzyme activity (32). 471
Binding of L-arabinose caused similar, albeit smaller, shifts within these mentioned regions. 472
However, the conformation of the L-arabinose binding pocket barely changed (pairwise Cα atoms 473
RMSD values of 0.277 and 0.198 Å for xylotetraose and L-arabinose complex structures, respectively). 474
475
CONCLUSIONS 476
In this study, we conducted a function/structure characterization of a thermostable, bacterial α-477
arabinofuranosidase from family GH62, namely SthAbf62A from Streptomyces thermoviolaceus. In 478
particular, new insights regarding the molecular basis for substrate selectivity among GH62 enzymes 479
were gained by studying complexes of SthAbf62A with L-arabinose and xylotetraose. All structural 480
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

22
features of SthAbf62A underpin the strict specificity for arabinofuranosyl groups bound to backbone 481
moieties through an alpha glycosidic configuration. Additionally, similar to BsAXH-m2,3 (18), the 482
narrowness of the arabinose-binding pocket likely prevents acceptance of α-L-Araf residues that are 483
(1→2) and (1→3) linked to di-substituted β-D-Xylp, which is also consistent with the 1H NMR 484
analyses of SthAbf62A reaction products (Fig. 2B). The complex structure with xylotetraose predicts at 485
least four subsites that bind the xylan backbone, which likely contributes to the clear preference of 486
SthAbf62A and other GH62 arabinofuranosidases for hydrolysis of polymeric arabinoxylan. Although 487
it is unclear how neighboring arabinose substituents may influence the orientation of arabinoxylan 488
substrates, the potential to bind xylo-oligosaccharides in opposite orientations explains the activity of 489
SthAbf62A on both L-Araf substituents that are (1→2) and (1→3) linked to mono-substituted β-D-Xylp 490
in arabinoxylan. 491
In summary, the recent increase in structure and functional characterization of GH62 492
arabinofuranosidases reflects the particular relevance of this enzyme family for modification and 493
characterization of polymeric and oligomeric arabinoxylans. In addition to predicting amino acid 494
residues important to overall enzyme stability, a compelling prospect of these recent insights is to fine-495
tune GH62 activities through better predictions of amino acids that govern regio-selectivity and 496
accommodation of singly versus doubly substituted substrates. 497
498
Acknowledgements 499
We thank Prof. Alexander Yakunin, Greg Brown and Rosa Di Leo from University of Toronto 500
for technical support; Prof. Wilson at Cornell University and Dr. T. Vuong at University of Toronto for 501
providing endoxylanase TfxA from Thermomonospora fusca; Dr. J. Tang and Dr. M. Nejad for NMR 502
spectra acquisition; M. Gudmundsson and Dr. M. Couturier for insightful discussions. We thank B. 503
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

23
Nocek at Argonne National Laboratory, Structural Biology Center at the Advanced Photon Source for 504
x-ray diffraction data collection. Funding for this research was provided by the Government of Ontario 505
for the project "Forest FAB: Applied Genomics for Functionalized Fibre and Biochemicals" (ORF-RE-506
05-005), the Natural Sciences and Engineering Research Council of Canada, and the Finnish Funding 507
Agency for Technology and Innovation (TEKES). 508
509
510
511
REFERENCES 512
1. Ebringerová A. 2005. Structural diversity and application potential of hemicelluloses. Macromol. 513
Sym. 232:1-12. 514
2. Hansen NML, Plackett D. 2008. Sustainable films and coatings from hemicelluloses: a review. 515
Biomacromolecules 9:1493-505. 516
3. Neyrinck AM, Possemiers S, Druart C, Van de Wiele T, De Backer F, Cani PD, Larondelle Y, 517
Delzenne NM. 2011. Prebiotic effects of wheat arabinoxylan related to the increase in 518
bifidobacteria, Roseburia and Bacteroides/Prevotella in diet-induced obese mice. PLoS One 519
6:e20944. 520
4. Aspinall GO. 1980. The biochemistry of plants. J. Preiss, ed. Academic Press, New York. 477-500. 521
5. Aspinall GO. 1959. Structural chemistry of the hemicelluloses. Adv. Carbohydr. Chem. 14:429-468. 522
6. Teleman A, Lundqvist J, Tjerneld F, Stalbrand H, Dahlman O. 2000. Characterization of 523
acetylated 4-O-methylglucuronoxylan isolated from aspen employing 1H and 13C NMR 524
spectroscopy. Carbohydr. Res. 329: 807-815. 525
7. Scheller HV, Ulvskov P. 2010. Hemicelluloses. Annu. Rev. Plant Biol. 61:263-289. 526
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

24
8. Saulnier L, Vigouroux J, Thibault JF. 1995. Isolation and partial characterization of feruloylated 527
oligosaccharides from maize bran. Carbohydr. Res. 272:241-53. 528
9. Timell TE SN. 1967. Recent progress in the chemistry of wood hemicelluloses. Wood Sci. Technol. 529
1:45-70. 530
10. Coughlan MP, Hazlewood GP. 1993. Beta-1,4-D-xylan-degrading enzyme systems: biochemistry, 531
molecular biology and applications. Biotechnol. Appl. Biochem. 17 :259-289. 532
11. Shallom D, Shoham Y. 2003. Microbial hemicellulases. Curr. Opin. Microbiol. 6:219-228. 533
12. de Vries RP, Kester HC, Poulsen CH, Benen JA, Visser J. 2000. Synergy between enzymes 534
from Aspergillus involved in the degradation of plant cell wall polysaccharides. Carbohydr. Res. 535
327:401-410. 536
13. Suzuki H, Murakami A, Yoshida K. 2013. Motif-guided identification of a glycoside hydrolase 537
family 1 alpha-L-arabinofuranosidase in Bifidobacterium adolescentis. Biosci. Biotechnol. 538
Biochem. 77:1709-1714. 539
14. Zhou J, Bao L, Chang L, Zhou Y, Lu H. 2012. Biochemical and kinetic characterization of GH43 540
beta-D-xylosidase/alpha-L-arabinofuranosidase and GH30 alpha-L-arabinofuranosidase/beta-D 541
-xylosidase from rumen metagenome. J. Ind. Microbiol. Biotechnol. 39:143-152. 542
15. Carapito R, Imberty A, Jeltsch JM, Byrns SC, Tam PH, Lowary TL, Varrot A, Phalip V. 543
2009. Molecular basis of arabinobio-hydrolase activity in phytopathogenic fungi: crystal 544
structure and catalytic mechanism of Fusarium graminearum GH93 exo-alpha-L-arabinanase. J. 545
Biol. Chem. 284:12285-12296. 546
16. Hovel K, Shallom D, Niefind K, Belakhov V, Shoham G, Baasov T, Shoham Y, Schomburg D. 547
2003. Crystal structure and snapshots along the reaction pathway of a family 51 alpha-L-548
arabinofuranosidase. Embo. J. 22:4922-4932. 549
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

25
17. Nurizzo D, Turkenburg JP, Charnock SJ, Roberts SM, Dodson EJ, McKie VA, Taylor EJ, 550
Gilbert HJ, Davies GJ. 2002. Cellvibrio japonicus alpha-L-arabinanase 43A has a novel five-551
blade beta-propeller fold. Nat. Struct. Biol. 9:665-668. 552
18. Vandermarliere E, Bourgois TM, Winn MD, van Campenhout S, Volckaert G, Delcour JA, 553
Strelkov SV, Rabijns A, Courtin CM. 2009. Structural analysis of a glycoside hydrolase 554
family 43 arabinoxylan arabinofuranohydrolase in complex with xylotetraose reveals a different 555
binding mechanism compared with other members of the same family. Biochem. J. 418:39-47. 556
19. Miyanaga A, Koseki T, Matsuzawa H, Wakagi T, Shoun H, Fushinobu S. 2004. Crystal 557
structure of a family 54 alpha-L-arabinofuranosidase reveals a novel carbohydrate-binding 558
module that can bind arabinose. J. Biol. Chem. 279:44907-44914. 559
20. Taylor EJ, Smith NL, Turkenburg JP, D'Souza S, Gilbert HJ, Davies GJ. 2006. Structural 560
insight into the ligand specificity of a thermostable family 51 arabinofuranosidase, Araf51, from 561
Clostridium thermocellum. Biochem. J. 395:31-37. 562
21. Beylot MH, McKie VA, Voragen AG, Doeswijk-Voragen CH, Gilbert HJ. 2001. The 563
Pseudomonas cellulosa glycoside hydrolase family 51 arabinofuranosidase exhibits wide 564
substrate specificity. Biochem. J. 358:607-614. 565
22. Birgisson H, Fridjonsson O, Bahrani-Mougeot FK, Hreggvidsson GO, Kristjansson JK, 566
Mattiasson B. 2004. A new thermostable alpha-L-arabinofuranosidase from a novel 567
thermophilic bacterium. Biotechnol. Lett. 26:1347-1351. 568
23. de Wet BJ, Matthew MK, Storbeck KH, van Zyl WH, Prior BA. 2008. Characterization of a 569
family 54 alpha-L-arabinofuranosidase from Aureobasidium pullulans. Appl. Microbiol. 570
Biotechnol. 77:975-983. 571
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

26
24. Lee RC, Hrmova M, Burton RA, Lahnstein J, Fincher GB. 2003. Bifunctional family 3 572
glycoside hydrolases from barley with alpha -L-arabinofuranosidase and beta -D-xylosidase 573
activity. Characterization, primary structures, and COOH-terminal processing. J. Biol. Chem. 574
278:5377-5387. 575
25. Flipphi MJ, Visser J, van der Veen P, de Graaff LH. 1993. Cloning of the Aspergillus niger 576
gene encoding alpha-L-arabinofuranosidase A. Appl. Microbiol. Biotechnol. 39:335-340. 577
26. Flipphi MJ, Panneman H, van der Veen P, Visser J, de Graaff LH. 1993. Molecular cloning, 578
expression and structure of the endo-1,5-alpha-L-arabinase gene of Aspergillus niger. Appl. 579
Microbiol. Biotechnol. 40:318-326. 580
27. McKie VA, Black GW, Millward-Sadler SJ, Hazlewood GP, Laurie JI, Gilbert HJ. 1997. 581
Arabinanase A from Pseudomonas fluorescens subsp. cellulosa exhibits both an endo- and an 582
exo- mode of action. Biochem. J. 323 :547-555. 583
28. Matsuo N, Kaneko S, Kuno A, Kobayashi H, Kusakabe I. 2000. Purification, characterization 584
and gene cloning of two alpha-L-arabinofuranosidases from Streptomyces chartreusis GS901. 585
Biochem. J. 346 :9-15. 586
29. Viborg AH, Sorensen KI, Gilad O, Steen-Jensen DB, Dilokpimol A, Jacobsen S, Svensson B. 587
2013. Biochemical and kinetic characterisation of a novel xylooligosaccharide-upregulated 588
GH43 beta-d-xylosidase/alpha-l-arabinofuranosidase (BXA43) from the probiotic 589
Bifidobacterium animalis subsp. lactis BB-12. AMB Express 3:56. 590
30. Bourgois TM, Van Craeyveld V, Van Campenhout S, Courtin CM, Delcour JA, Robben J, 591
Volckaert G. 2007. Recombinant expression and characterization of XynD from Bacillus 592
subtilis subsp. subtilis ATCC 6051: a GH 43 arabinoxylan arabinofuranohydrolase. Appl. 593
Microbiol. Biotechnol. 75:1309-1317. 594
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

27
31. van den Broek LA, Lloyd RM, Beldman G, Verdoes JC, McCleary BV, Voragen AG. 2005. 595
Cloning and characterization of arabinoxylan arabinofuranohydrolase-D3 (AXHd3) from 596
Bifidobacterium adolescentis DSM20083. Appl. Microbiol. Biotechnol. 67:641-647. 597
32. Siguier B, Haon M, Nahoum V, Marcellin M, Burlet-Schiltz O, Coutinho PM, Henrissat B, 598
Mourey L, MJ OD, Berrin JG, Tranier S, Dumon C. 2014. First structural insights into 599
alpha-L-arabinofuranosidases from the two GH62 glycoside hydrolase subfamilies. J. Biol. 600
Chem. 289:5261-5273. 601
33. Maehara T, Fujimoto Z, Ichinose H, Michikawa M, Harazono K, Kaneko S. 2014. Crystal 602
structure and characterization of the glycoside hydrolase family 62 alpha-L-arabinofuranosidase 603
from Streptomyces coelicolor. J. Biol. Chem. 289:7962-7972 604
34. Vincent P, Shareck F, Dupont C, Morosoli R, Kluepfel D. 1997. New alpha-L-605
arabinofuranosidase produced by Streptomyces lividans: cloning and DNA sequence of the abfB 606
gene and characterization of the enzyme. Biochem. J. 322 :845-852. 607
35. Kimura I, Yoshioka N, Kimura Y, Tajima S. 2000. Cloning, sequencing and expression of an 608
alpha-L-arabinofuranosidase from Aspergillus sojae. J. Biosci. Bioeng. 89:262-266. 609
36. Sakamoto T, Ogura A, Inui M, Tokuda S, Hosokawa S, Ihara H, Kasai N. 2011. Identification 610
of a GH62 alpha-L-arabinofuranosidase specific for arabinoxylan produced by Penicillium 611
chrysogenum. Appl. Microbiol. Biotechnol. 90:137-146. 612
37. Tsujibo H, Takada C, Wakamatsu Y, Kosaka M, Tsuji A, Miyamoto K, Inamori Y. 2002. 613
Cloning and expression of an alpha-L-arabinofuranosidase gene (stxIV) from Streptomyces 614
thermoviolaceus OPC-520, and characterization of the enzyme. Biosci. Biotechnol. Biochem. 615
66:434-438. 616
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

28
38. Hashimoto K, Yoshida M, Hasumi K. 2011. Isolation and characterization of CcAbf62A, a GH62 617
alpha-L-arabinofuranosidase, from the basidiomycete Coprinopsis cinerea. Biosci. Biotechnol. 618
Biochem. 75:342-345. 619
39. Martens EC, Lowe EC, Chiang H, Pudlo NA, Wu M, McNulty NP, Abbott DW, Henrissat B, 620
Gilbert HJ, Bolam DN, Gordon JI. 2011. Recognition and degradation of plant cell wall 621
polysaccharides by two human gut symbionts. PLoS Biol 9:e1001221. 622
40. Liu H, Naismith JH. 2008. An efficient one-step site-directed deletion, insertion, single and 623
multiple-site plasmid mutagenesis protocol. BMC Biotechnol. 8:91. 624
41. Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of 625
protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254. 626
42. Smogyi M. 1952. Notes on sugar determination. J. Biol. Chem. 195:19-23. 627
43. Copeland RA. 2000. Enzymes: A practical introduction to structure, mechanism, and data analysis. 628
7:109-145. 629
44. Ahmed S, Luis AS, Bras JL, Ghosh A, Gautam S, Gupta MN, Fontes CM, Goyal A. 2013. A 630
novel alpha-L-arabinofuranosidase of family 43 glycoside hydrolase (Ct43Araf) from 631
Clostridium thermocellum. PLoS One 8:e73575. 632
45. Sternemalm E, Hoije A, Gatenholm P. 2008. Effect of arabinose substitution on the material 633
properties of arabinoxylan films. Carbohydr. Res. 343:753-7. 634
46. Pitkanen L, Tuomainen P, Virkki L, Tenkanen M. 2011. Molecular characterization and 635
solution properties of enzymatically tailored arabinoxylans. Int. J. Biol. Macromol. 49:963-969. 636
47. Irwin D, Jung ED, Wilson DB. 1994. Characterization and sequence of a Thermomonospora fusca 637
xylanase. Appl. Environ. Microbiol. 60:763-70. 638
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

29
48. Pollet A, Delcour JA, Courtin CM. 2010. Structural determinants of the substrate specificities of 639
xylanases from different glycoside hydrolase families. Crit. Rev. Biotechnol. 30:176-191. 640
49. Swennen K, Courtin CM, Lindemans GCJE, Delcour JA. 2006. Large-scale production and 641
characterisation of wheat bran arabinoxylooligosaccharides. J. Sci. Food Agric. 86:1722-1731. 642
50. Minor W, Cymborowski M, Otwinowski Z, Chruszcz M. 2006. HKL-3000: the integration of 643
data reduction and structure solution--from diffraction images to an initial model in minutes. 644
Acta. Crystallogr. D Biol. Crystallogr. 62:859-866. 645
51. Adams PD, Afonine PV, Bunkoczi G, Chen VB, Davis IW, Echols N, Headd JJ, Hung LW, 646
Kapral GJ, Grosse-Kunstleve RW, McCoy AJ, Moriarty NW, Oeffner R, Read RJ, 647
Richardson DC, Richardson JS, Terwilliger TC, Zwart PH. 2010. PHENIX: a 648
comprehensive Python-based system for macromolecular structure solution. Acta. Crystallogr. 649
D Biol. Crystallogr. 66:213-221. 650
52. Emsley P, Cowtan K. 2004. Coot: model-building tools for molecular graphics. Acta. Crystallogr. 651
D Biol. Crystallogr. 60:2126-2132. 652
53. Couturier M, Haon M, Coutinho PM, Henrissat B, Lesage-Meessen L, Berrin JG. 2011. 653
Podospora anserina hemicellulases potentiate the Trichoderma reesei secretome for 654
saccharification of lignocellulosic biomass. Appl. Environ. Microbiol. 77:237-46. 655
54. Pitkanen L, Virkki L, Tenkanen M, Tuomainen P. 2009. Comprehensive multidetector HPSEC 656
study on solution properties of cereal arabinoxylans in aqueous and DMSO solutions. 657
Biomacromolecules 10:1962-9. 658
55. Izydorczyk MS, Biliaderis CG. 1995. Cereal arabinoxylans: advances in structure and 659
physicochemical properties. Carbohydr. Polym. 28:33-48. 660
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

30
56. Kormelink FJM, Voragen AGJ. 1993. Degradation of different [(glucurono)arabino]xylans by a 661
combination of purified xylan-degrading enzymes. Appl. Microbiol. Biotechnol. 38:688-695. 662
57. Rantanen H, Virkki L, Tuomainen Pi, Kabel M, Schols H, Tenkanen M. 2007. Preparation of 663
arabinoxylobiose from rye xylan using family 10 Aspergillus aculeatus endo-1,4-β-D-xylanase. 664
Carbohydr. Polym. 68:350-359. 665
58. Gerlt JA, Babbitt PC. 2001. Divergent evolution of enzymatic function: mechanistically diverse 666
superfamilies and functionally distinct suprafamilies. Annu. Rev. Biochem. 70:209-46. 667
59. Rogowski A, Basle A, Farinas CS, Solovyova A, Mortimer JC, Dupree P, Gilbert HJ, Bolam 668
DN. 2014. Evidence that GH115 alpha-glucuronidase activity, which is required to degrade 669
plant biomass, is dependent on conformational flexibility. J. Biol. Chem. 289:53-64. 670
60. Biely P. 2012. Microbial carbohydrate esterases deacetylating plant polysaccharides. Biotechnol. 671
Adv. 30:1575-1588. 672
61. McKee LS, Peña MJ, Rogowski A, Jackson A, Lewis RJ, York WS, Krogh KBRM, Viksø-673
Nielsen A, Skjøt M, Gilbert HJ, Marles-Wright J. 2012. Introducing endo-xylanase activity 674
into an exo-acting arabinofuranosidase that targets side chains. Proc. Natl. Acad. Sci. U. S. A. 675
109:6537-42. 676
677 678 679 680
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

FIGURE LEGENDS 1
Figure 1. Effect of pH and temperature on SthAbf62A activity and stability. (A) The effect 2
of pH on enzyme activity; (B) pH stability of purified SthAbf62A; (C) The effect of temperature 3
on SthAbf62A activity; (D) Thermostability of purified SthAbf62A. Activity assays were 4
performed using 0.5% wheat arabinoxylan (high viscosity) and 0.4 μg SthAbf62A. 5
6
Figure 2. 1H NMR analysis of enzyme treated wheat arabinoxylan. m-1,2 and m-1,3-α-L-Ara 7
refer to mono-substituted β-D-Xylp at positions C2 and C3, respectively; d-1,2 and d-1,3-α-L-8
Ara refer to arabinose attached to double-substituted β-D-Xylp. (A) Wheat arabinoxylan (high 9
viscosity) before (black line) and after (red line) treatment with AXH-d3 from Bifidobacterium 10
sp. for 48 h; the effect of AXH-d3 treatment was confirmed by the disappearance of the signals 11
near 5.28 ppm and 5.22 ppm and concomitant increase in the signal near 5.30 ppm (54). (B) 12
Wheat arabinoxylan (high viscosity) before (black line) and after (red line) treatment with 13
SthAbf62A (3.0 μg) in 100 mM HEPES buffer (pH 7.0) at 45°C for 2 h; removal of α-L-Araf 14
(1→3) linked to mono-substituted β-D-Xylp was indicated by the disappearance of the H1 signal 15
at 5.42 ppm (red line); signals at 5.22 ppm and 5.28 ppm for (1→3)-linked and (1→2)-linked α-16
L-Araf substituents of doubly substituted β-D-Xylp remained after treatment for up to 24 h (data 17
not shown). (C) AXH-d3 treated arabinoxylan before (black line) and after (red line) treatment 18
with SthAbf62A (3.0 μg) in 100 mM HEPES buffer (pH 7.0) at 45°C for 2 h; removal of (1→3)-19
linked and (1→2)-linked α-L-Araf substituents of singly substituted β-D-Xylp was indicated by 20
the disappearance of the H1 signal at both of 5.42 ppm and 5.28 ppm (red line). The methylene 21
protons at position 2 of MOPS (2.035 ppm) were used as the internal reference. 22
23
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Figure 3. The overall structure of SthAbf62A revealing a five-bladed β-propeller fold. 24
(A) Top view showing the numbering of 5 blades (I–V), three predicted catalytic residues, a 25
disulfide bond between Cys297 and Cys27, a molecular Velcro boxed in grey dash line and a 26
calcium ion coordinated in the central channel of the fold. (B) Side view showing the active site 27
surrounded by loops. (C) Overall topology of SthAbf62A highlighting the molecular Velcro 28
between β1 and β21 by an orange dashed line, and the position of three predicted catalytic 29
residues. β-strands are indicated by arrows and helices are indicated by cylinders. 30
31
Figure 4. Interactions between SthAbf62A and L-arabinofuranose bound in the active site 32
pocket. 33
(A) L-Arabinofuranose is shown in pink and amino acid residues in grey; polar interactions are 34
indicated by black dotted lines. A calcium ion shown as an orange sphere was buried under the 35
L-arabinose binding pocket, displaying a hepta-coordination with His280, Gln216 and five water 36
molecules indicated as red spheres. (B) Focus on a nucleophilic water molecule, which shows 37
shared electron density with C1 of L-arabinose, as indicated by the black box. The electron 38
density shown is simulated annealing Fo-Fc omit density contoured at 2.0 σ. 39
40
Figure 5. Interactions between SthAbf62A and xylotetraose bound in the substrate binding 41
cleft. Xylotetraose is shown in yellow, amino acids predicted to interact with the bound 42
oligosaccharide are shown in grey, and a potassium ion bound in the arabinose-binding site (-1 43
subsite) is shown as a purple sphere. Black dotted lines indicate polar interactions; all contacts 44
are listed in Table S4. The xylotetraose is shown in Orientation 1. All contacts remained in 45
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Orientation 2, with an extra hydrogen bond between Arg239 and the endocyclic oxygen of 46
xylose unit at +2NR subsite. 47
48
Figure 6. Main conformational changes in SthAbf62A upon ligand binding. 49
Structural elements of SthAbf62A are shown in orange; corresponding regions of the enzyme-50
xylotetraose complex are shown in grey and enzyme-L-arabinose complex are shown in green. 51
Xylotetraose is shown in yellow and the enzyme surface shown in grey corresponds to the 52
enzyme-xylotetraose complex. Superimposition of the three structures was conducted using 53
DaliLite. 54
55
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from
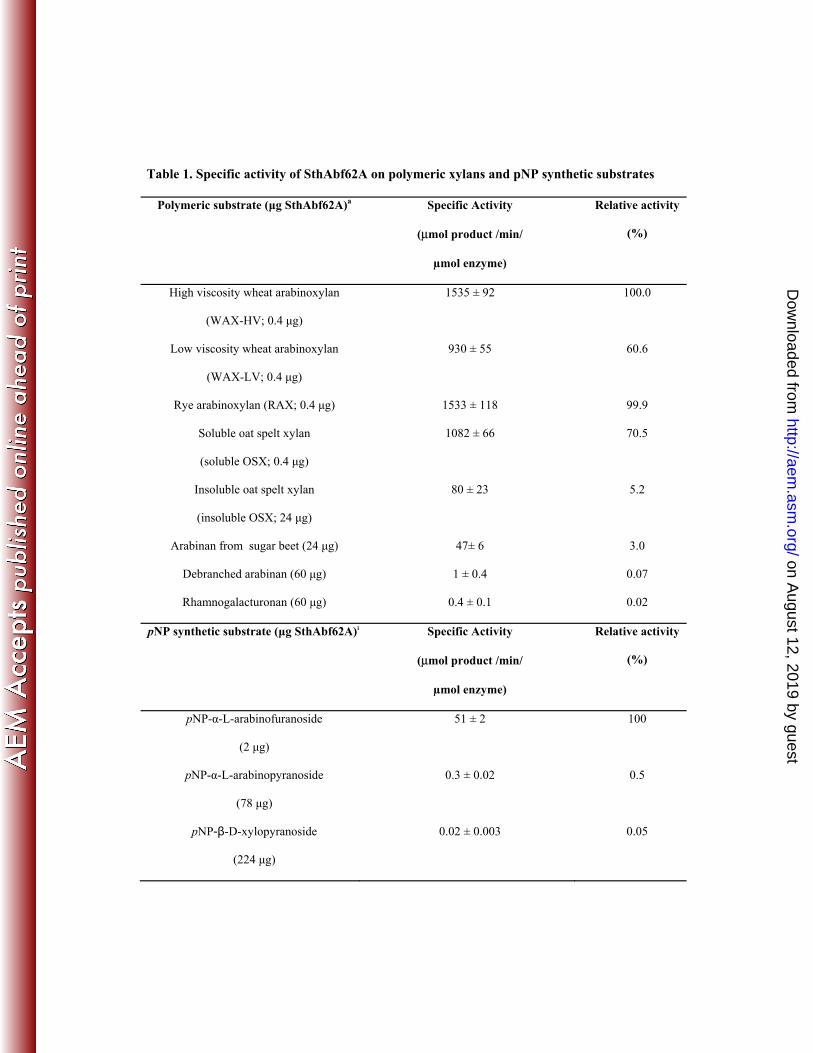
Table 1. Specific activity of SthAbf62A on polymeric xylans and pNP synthetic substrates
Polymeric substrate (μg SthAbf62A)a Specific Activity
(μmol product /min/
µmol enzyme)
Relative activity
(%)
High viscosity wheat arabinoxylan
(WAX-HV; 0.4 μg)
1535 ± 92 100.0
Low viscosity wheat arabinoxylan
(WAX-LV; 0.4 μg)
930 ± 55 60.6
Rye arabinoxylan (RAX; 0.4 μg) 1533 ± 118 99.9
Soluble oat spelt xylan
(soluble OSX; 0.4 μg)
1082 ± 66 70.5
Insoluble oat spelt xylan
(insoluble OSX; 24 μg)
80 ± 23 5.2
Arabinan from sugar beet (24 μg) 47± 6 3.0
Debranched arabinan (60 μg) 1 ± 0.4 0.07
Rhamnogalacturonan (60 μg) 0.4 ± 0.1 0.02
pNP synthetic substrate (μg SthAbf62A)a Specific Activity
(μmol product /min/
µmol enzyme)
Relative activity
(%)
pNP-α-L-arabinofuranoside
(2 μg)
51 ± 2 100
pNP-α-L-arabinopyranoside
(78 μg)
0.3 ± 0.02 0.5
pNP-β-D-xylopyranoside
(224 μg)
0.02 ± 0.003 0.05
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

aAmounts of purified SthAbf62A added to each reaction to measure initial reaction velocities.
Activities obtained with WAX-HV and pNP-α-L-arabinofuranoside were taken as 100% in
respective comparisons. No activity was detected towards birchwood xylan, beechwood xylan,
apple pectin, larch arabinogalactan, pNP-β-L-arabinopyranoside, pNP-α-D-mannopyranoside,
pNP-α-D-galactopyranoside, pNP-α-L-fucopyranoside and pNP-α-D-glucopyranoside.
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from
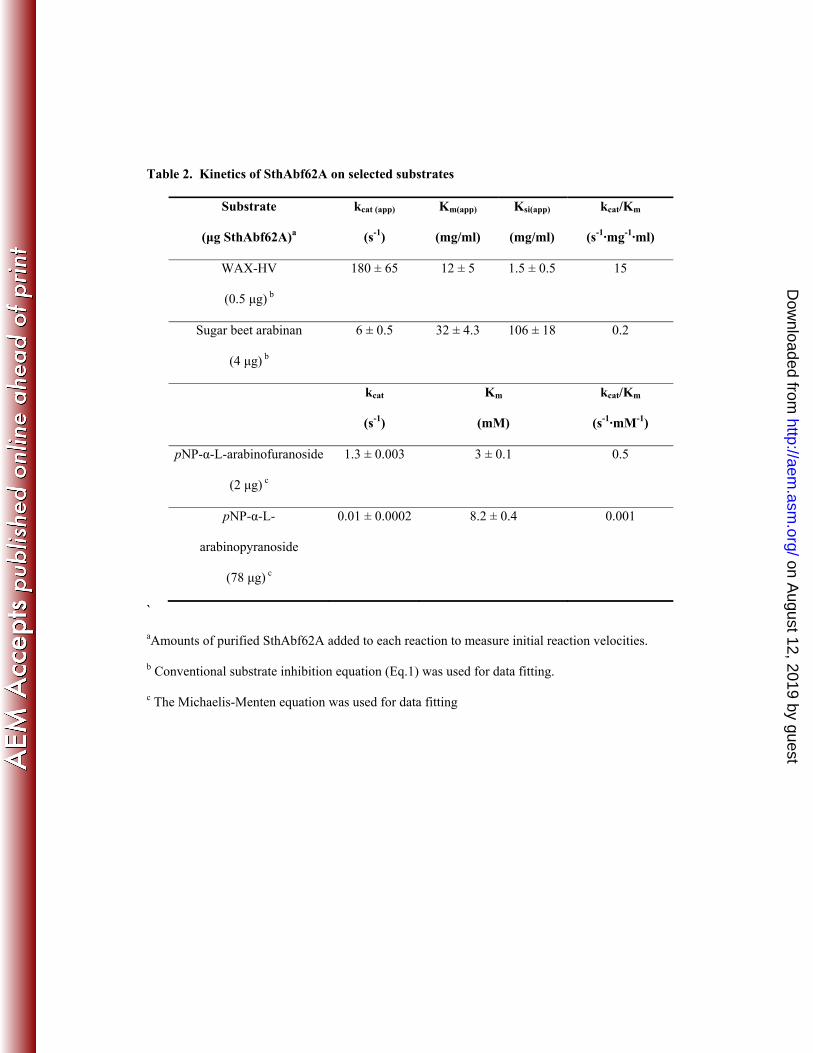
Table 2. Kinetics of SthAbf62A on selected substrates
Substrate
(μg SthAbf62A)a
kcat (app)
(s-1)
Km(app)
(mg/ml)
Ksi(app)
(mg/ml)
kcat/Km
(s-1·mg-1·ml)
WAX-HV
(0.5 μg) b
180 ± 65 12 ± 5 1.5 ± 0.5 15
Sugar beet arabinan
(4 μg) b
6 ± 0.5 32 ± 4.3 106 ± 18 0.2
kcat
(s-1)
Km
(mM)
kcat/Km
(s-1·mM-1)
pNP-α-L-arabinofuranoside
(2 μg) c
1.3 ± 0.003 3 ± 0.1
0.5
pNP-α-L-
arabinopyranoside
(78 μg) c
0.01 ± 0.0002 8.2 ± 0.4 0.001
`
aAmounts of purified SthAbf62A added to each reaction to measure initial reaction velocities.
b Conventional substrate inhibition equation (Eq.1) was used for data fitting.
c The Michaelis-Menten equation was used for data fitting
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from

Table 3. X-ray diffraction data collection and refinement statistics
Ligand
PDB Code
apoenzyme
4O8N
Xylotetraose
4O8P
L-α-arabinose
4O8O
Data collection
Space group P212121 P212121 P212121
Cell dimensions
a, b, c, Å 59.81, 65.68, 84.95 42.49, 64.64, 121.30 60.05, 65.74, 85.20
α, β, γ, ° 90, 90, 90 90, 90, 90 90, 90, 90
Resolution, Å 25.00 – 1.65 25.00 – 1.56 18.00 – 1.18
Rmergea 0.081 (0.587)a 0.086 (0.517)b 0.050 (0.508)c
I / σ(I) 29.96 (4.0) 21.77 (3.4) 31.45 (3.2)
Completeness, % 96.2 (92.7) 99.6 (99.5) 98.1 (96.1)
Redundancy 5.9 (6.0) 4.6 (4.7) 4.9 (4.8)
Refinement
Resolution, Å 24.45 – 1.65 23.68 – 1.56 17.47 – 1.21
No. of reflections 38657 48371 101459
R-factor/free R-factord 12.7/16.3 (18.0/24.0)d 13.0/15.3 (17.2/19.9)e 13.9/15.6 (19.6/21.67)f
No. of refined atoms
Protein
Ca2+
Ligand
Solvent
Water
2391
1
N/A
39
540
2444
1
35
39
474
2405
1
10
0
516
B-factors
Protein
Ca2+
Ligand
15.3
10.8
N/A
13.0
6.1
19.8
14.3
7.8
15.0
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from
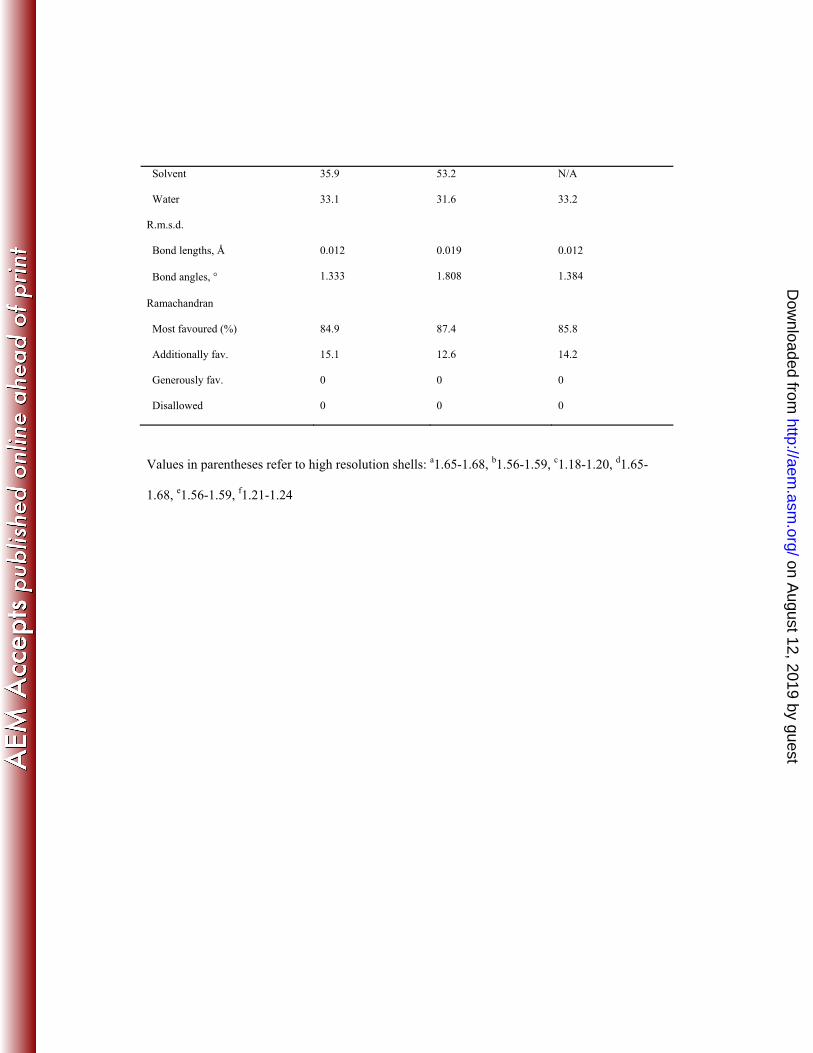
Solvent
Water
35.9
33.1
53.2
31.6
N/A
33.2
R.m.s.d.
Bond lengths, Å
Bond angles, °
0.012
1.333
0.019
1.808
0.012
1.384
Ramachandran
Most favoured (%)
Additionally fav.
Generously fav.
Disallowed
84.9
15.1
0
0
87.4
12.6
0
0
85.8
14.2
0
0
Values in parentheses refer to high resolution shells: a1.65-1.68, b1.56-1.59, c1.18-1.20, d1.65-
1.68, e1.56-1.59, f1.21-1.24
on August 12, 2019 by guest
http://aem.asm
.org/D
ownloaded from



![CdS Nanoparticle-Modified α-Fe2O3/TiO2 Nanorod Array ......3/TiO 2 photoanodes, some narrow band gap semiconductors, like CdS [14, 15] and PbS [16], can be coupled to facilitate the](https://static.fdocument.org/doc/165x107/60e51c272904ea539f2bde32/cds-nanoparticle-modified-fe2o3tio2-nanorod-array-3tio-2-photoanodes.jpg)














