
ABCG5/ABCG8: A Structural View on Sterol Transport
30
ABCG5/ABCG8: A Structural View on Sterol Transport Jyh-Yeuan (Eric) Lee University of Texas Southwestern Medical Center at Dallas
-
Upload
jyh-yeuan-eric-lee -
Category
Science
-
view
63 -
download
0
Transcript of ABCG5/ABCG8: A Structural View on Sterol Transport

ABCG5/ABCG8: A Structural View on Sterol Transport
Jyh-Yeuan (Eric) Lee
University of Texas Southwestern Medical Center at Dallas
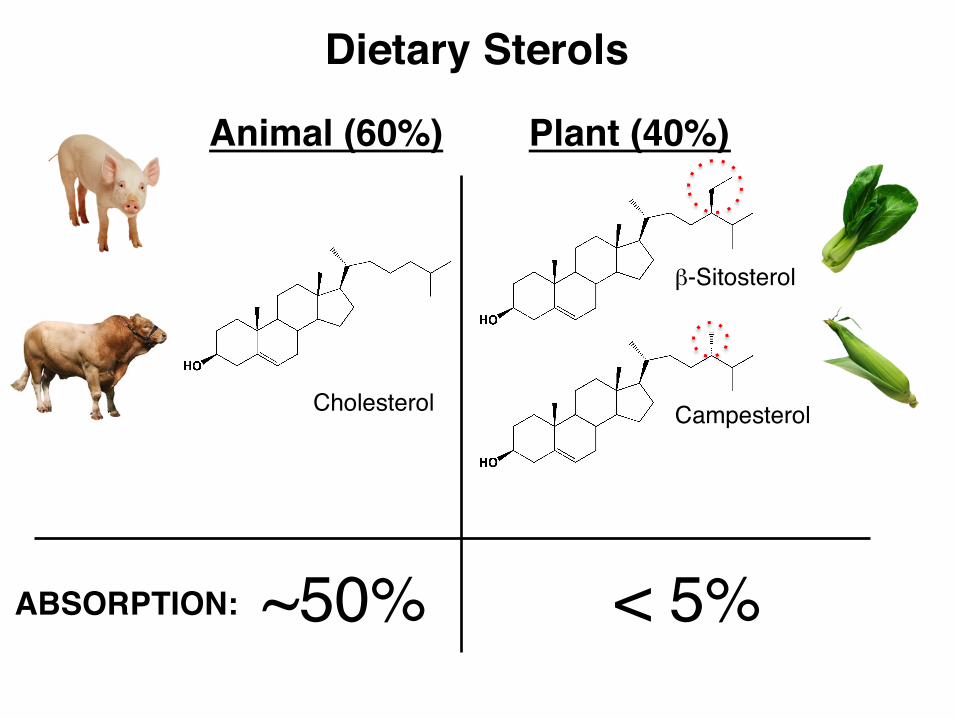
Dietary Sterols
Cholesterol
Animal (60%) Plant (40%)
ABSORPTION: ~50% < 5%
β-Sitosterol
Campesterol
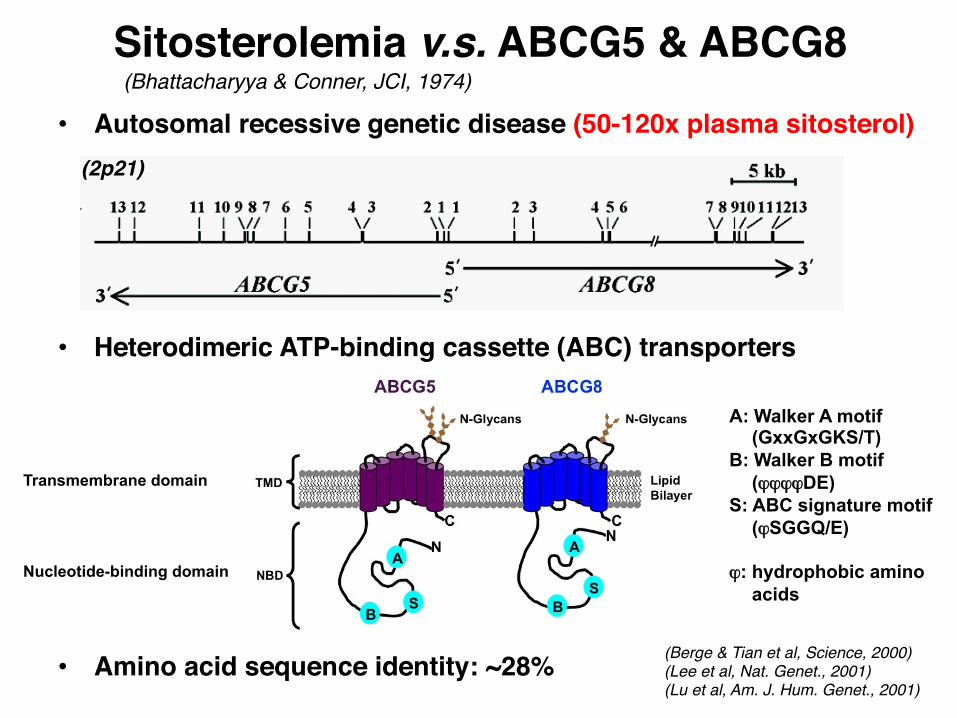
• Autosomal recessive genetic disease (50-120x plasma sitosterol)
• Heterodimeric ATP-binding cassette (ABC) transporters
• Amino acid sequence identity: ~28%
Sitosterolemia v.s. ABCG5 & ABCG8(Bhattacharyya & Conner, JCI, 1974)
(Berge & Tian et al, Science, 2000)(Lee et al, Nat. Genet., 2001)(Lu et al, Am. J. Hum. Genet., 2001)
(2p21)
A: Walker A motif (GxxGxGKS/T)
B: Walker B motif (ϕϕϕϕDE)
S: ABC signature motif (ϕSGGQ/E)
ϕ: hydrophobic amino
acids
Transmembrane domain
Nucleotide-binding domain
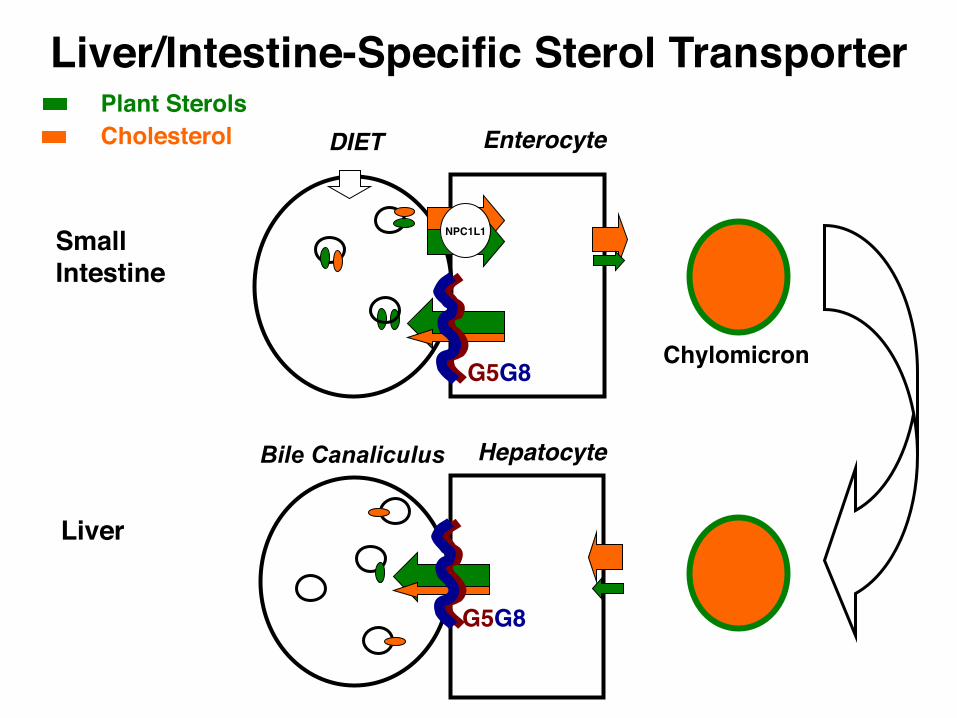
Enterocyte
NPC1L1
Chylomicron
SmallIntestine
CholesterolPlant Sterols
G5G8
DIET
Hepatocyte
Liver
G5G8
Bile Canaliculus
Liver/Intestine-Specific Sterol Transporter
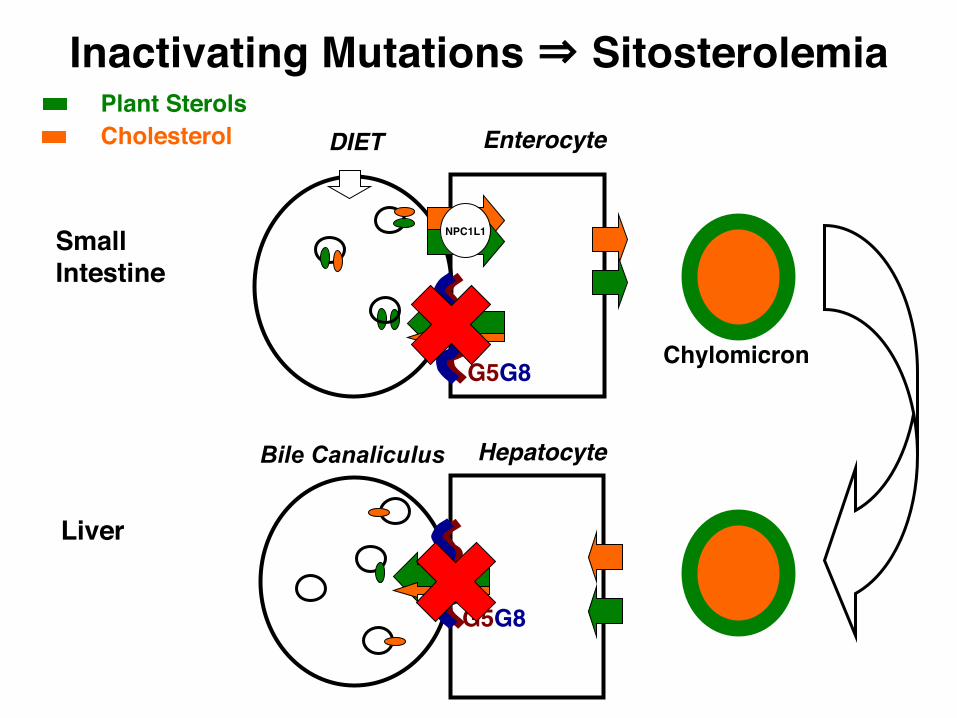
Enterocyte
NPC1L1
Chylomicron
SmallIntestine
CholesterolPlant Sterols
G5G8
DIET
Hepatocyte
Liver
G5G8
Bile Canaliculus
Inactivating Mutations ⇒ Sitosterolemia
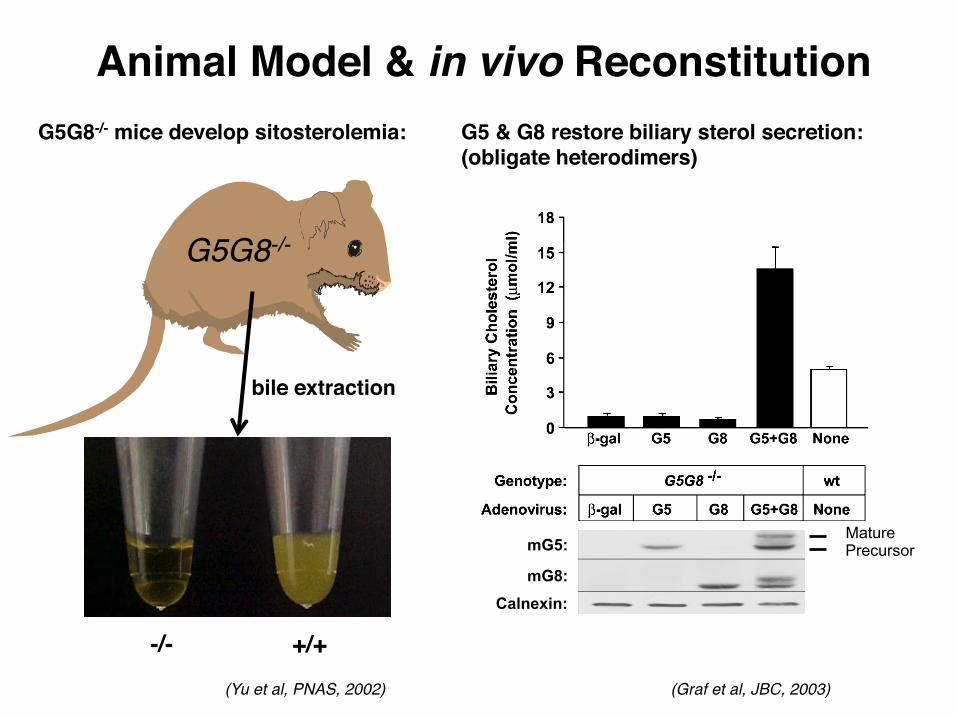
Animal Model & in vivo Reconstitution
(Yu et al, PNAS, 2002)
+/+-/-
G5G8-/-
G5 & G8 restore biliary sterol secretion: (obligate heterodimers)
mG5:
mG8: Calnexin:
Precursor Mature
(Graf et al, JBC, 2003)
bile extraction
G5G8-/- mice develop sitosterolemia:
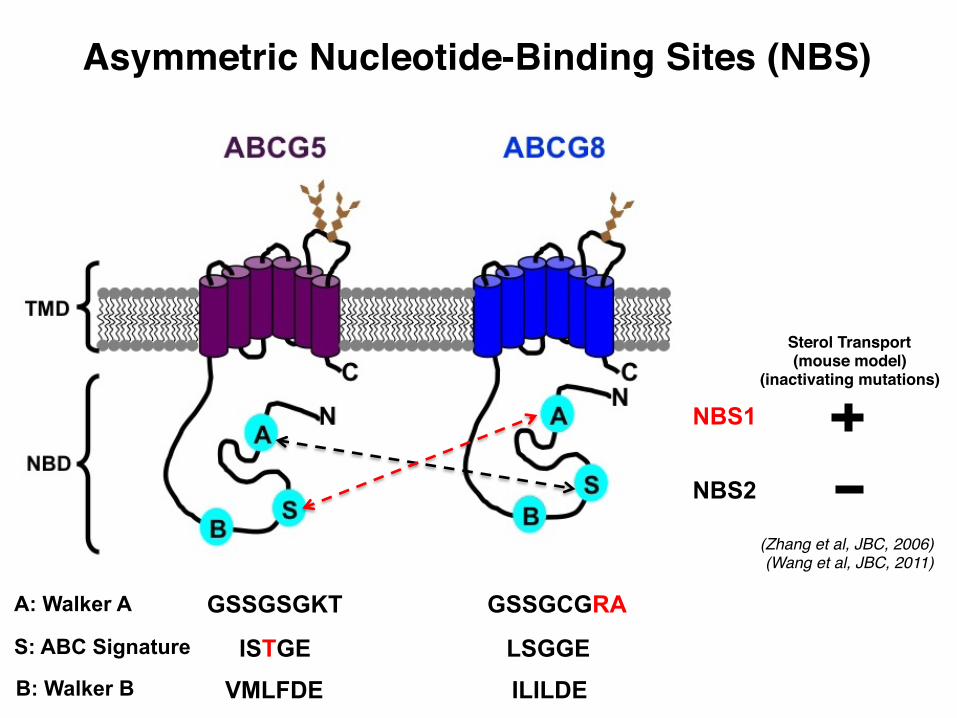
Asymmetric Nucleotide-Binding Sites (NBS)
A: Walker A
S: ABC Signature
GSSGSGKT
LSGGE
GSSGCGRA
ISTGE B: Walker B
NBS1
NBS2
(Zhang et al, JBC, 2006)(Wang et al, JBC, 2011)
Sterol Transport(mouse model)
(inactivating mutations)
VMLFDE ILILDE
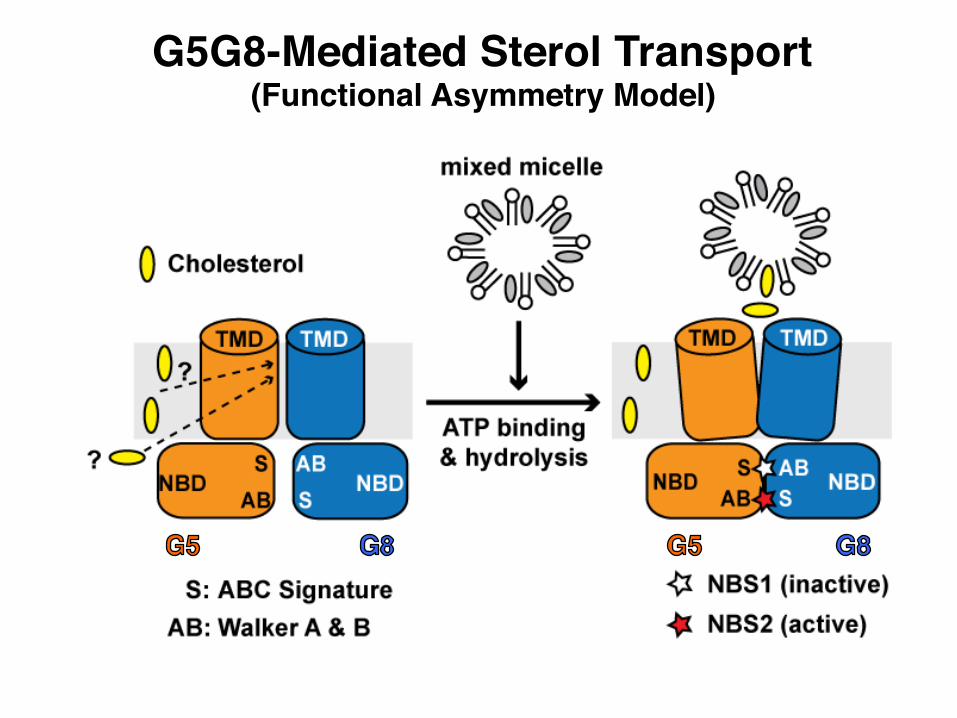
G5G8-Mediated Sterol Transport(Functional Asymmetry Model)
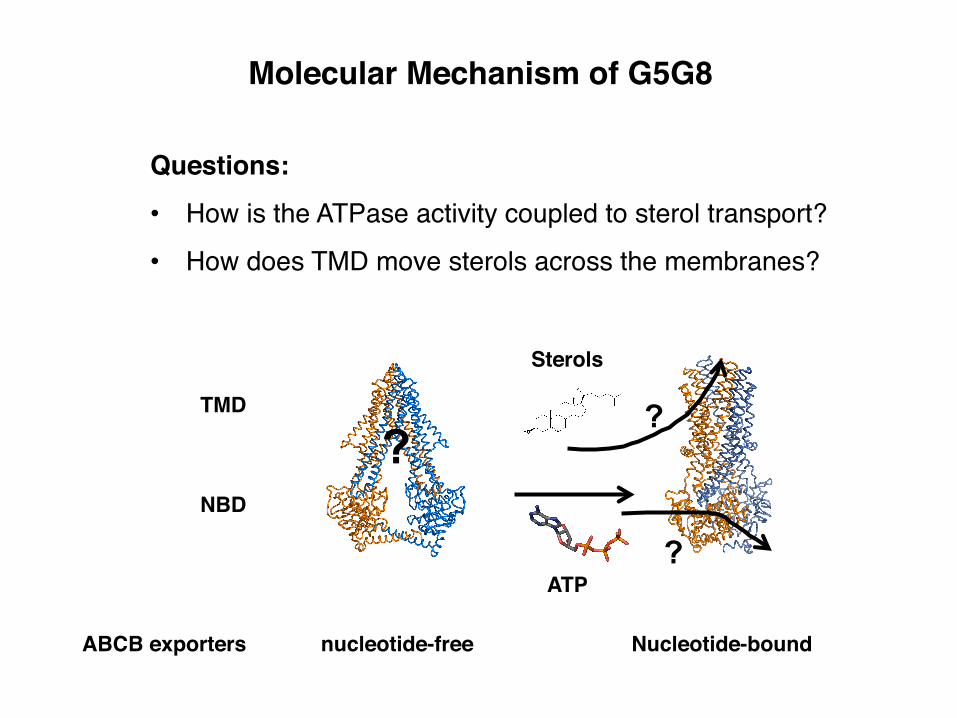
TMD
NBD
Questions:• How is the ATPase activity coupled to sterol transport?
• How does TMD move sterols across the membranes?
Molecular Mechanism of G5G8
?
?
ABCB exporters nucleotide-free Nucleotide-bound
ATP
Sterols
?
0 10 20 30 400
2000
4000
6000
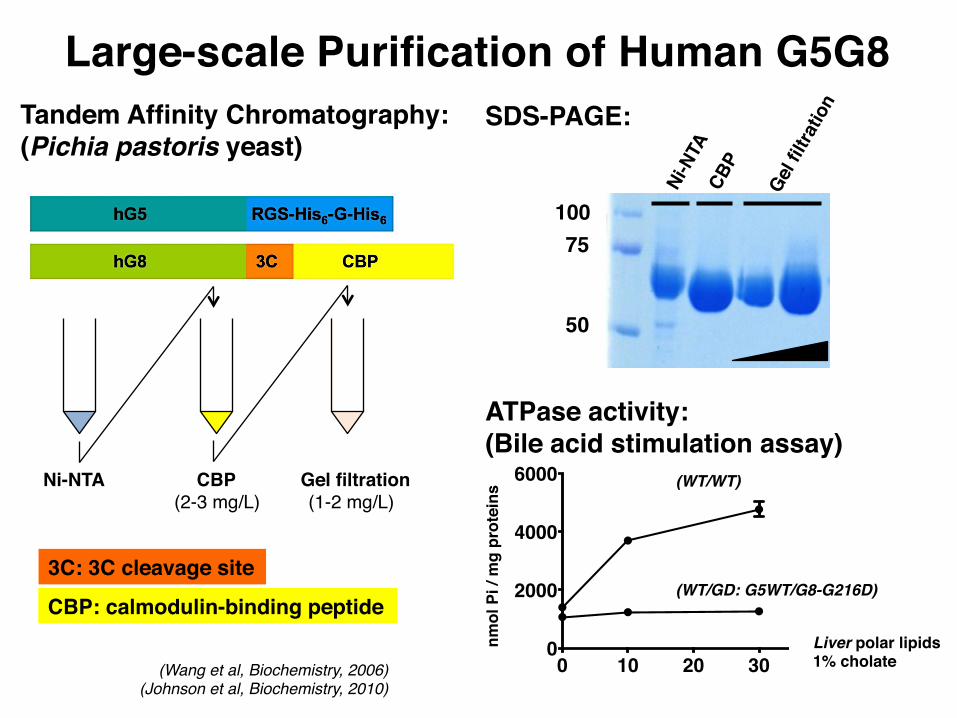
Large-scale Purification of Human G5G8Tandem Affinity Chromatography:(Pichia pastoris yeast)
Ni-NTA CBP Gel filtration
CBP: calmodulin-binding peptide
(2-3 mg/L) (1-2 mg/L)
10075
SDS-PAGE:
3C: 3C cleavage site
ATPase activity:(Bile acid stimulation assay)
Liver polar lipids1% cholate(Wang et al, Biochemistry, 2006)
(Johnson et al, Biochemistry, 2010)
(WT/GD: G5WT/G8-G216D)
50
(WT/WT)nm
ol P
i / m
g pr
otei
ns
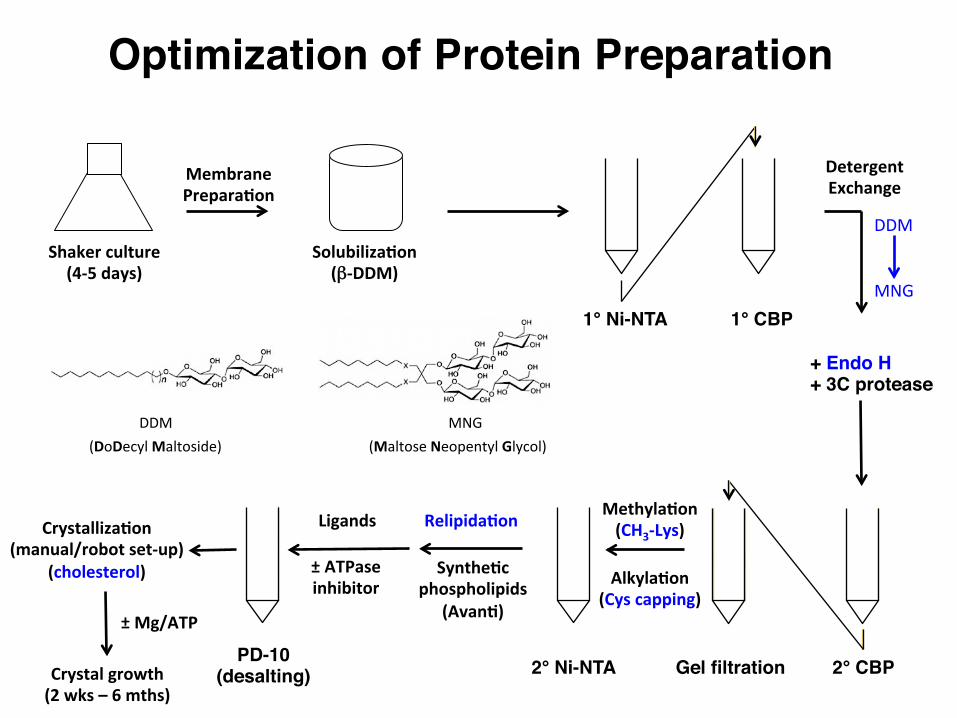
Optimization of Protein Preparation
1° Ni-NTA 1° CBP
Gel filtration
Shakerculture(4-5days)
MembranePrepara:on
Solubiliza:on(β-DDM)
DDM
MNG
+ Endo H+ 3C protease
Methyla:on(CH3-Lys)
Relipida:on
Synthe:cphospholipids
(Avan:)
±ATPaseinhibitor
Crystalliza:on(manual/robotset-up)
(cholesterol)
Crystalgrowth(2wks–6mths)
Ligands
±Mg/ATP
2° CBP2° Ni-NTA
DDM MNG
DetergentExchange
PD-10(desalting)
Alkyla:on(Cyscapping)
(DoDecylMaltoside) (MaltoseNeopentylGlycol)
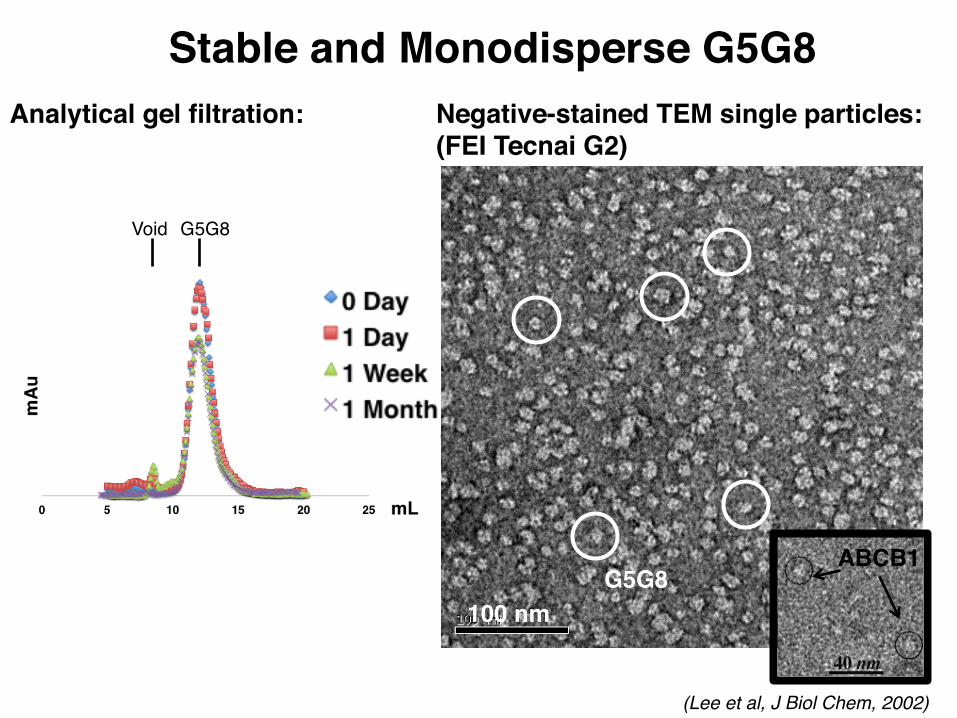
100 nm
Analytical gel filtration: Negative-stained TEM single particles:(FEI Tecnai G2)
ABCB1
Void G5G8
G5G8
(Lee et al, J Biol Chem, 2002)
Stable and Monodisperse G5G8
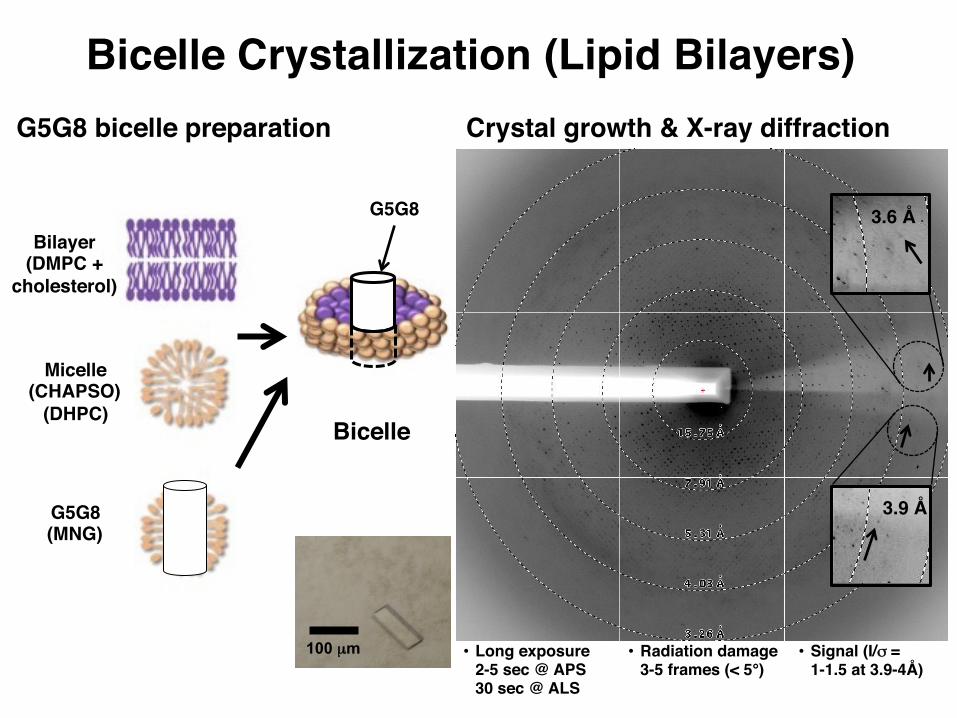
Bicelle Crystallization (Lipid Bilayers)G5G8 bicelle preparation
Bilayer(DMPC +
cholesterol)
Micelle(CHAPSO)
(DHPC)Bicelle
G5G8
G5G8(MNG)
Crystal growth & X-ray diffraction
100 µm • Long exposure2-5 sec @ APS30 sec @ ALS
• Radiation damage3-5 frames (< 5°)
• Signal (I/σ = 1-1.5 at 3.9-4Å)
3.9 Å
3.6 Å
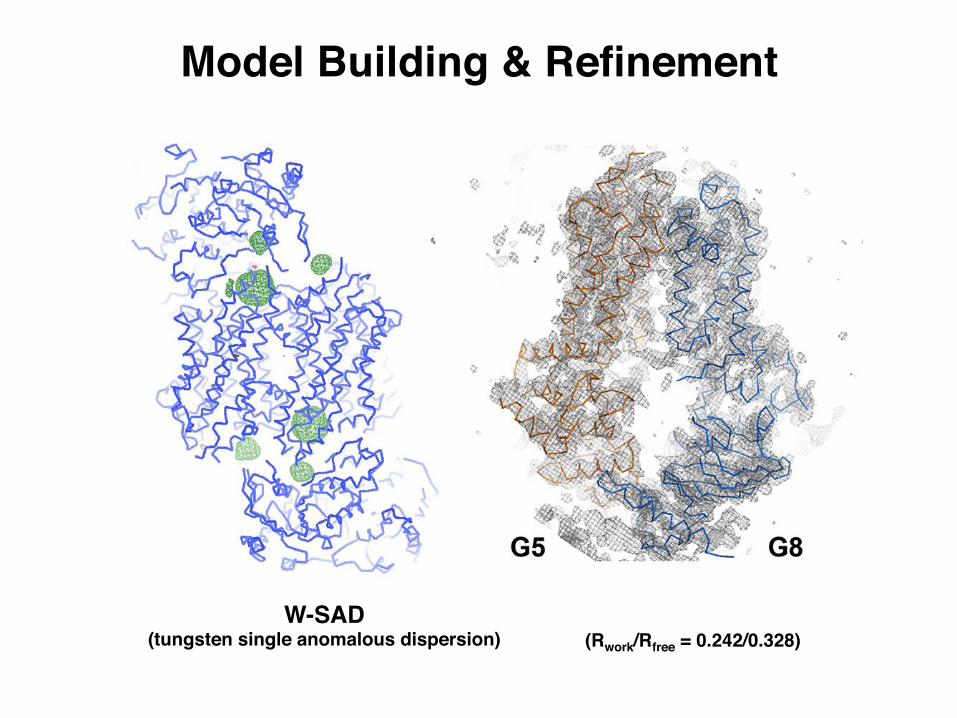
Model Building & Refinement
(Rwork/Rfree = 0.242/0.328)W-SAD
(tungsten single anomalous dispersion)

(Hollenstein et al, Curr Opin Struct Biol, 2007)(Rees et al, Nature Rev Mol Cell Biol, 2009)
New TMD Fold for ABC Transporters
(Lee et al, Nature, 2016)
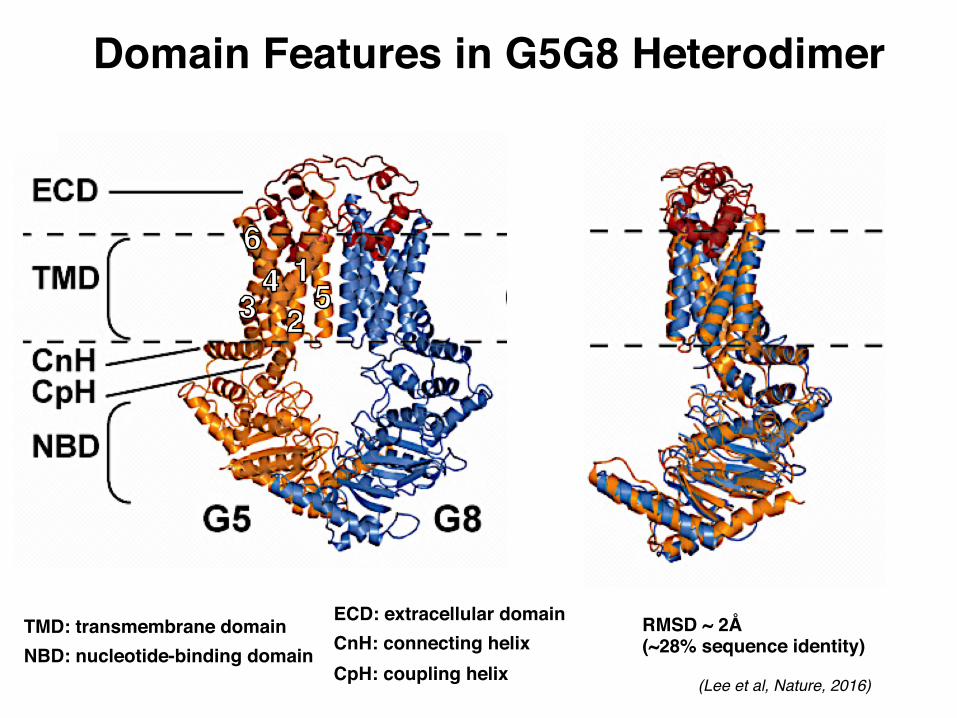
Domain Features in G5G8 Heterodimer
RMSD ~ 2Å(~28% sequence identity)
TMD: transmembrane domainNBD: nucleotide-binding domain
ECD: extracellular domainCnH: connecting helixCpH: coupling helix (Lee et al, Nature, 2016)
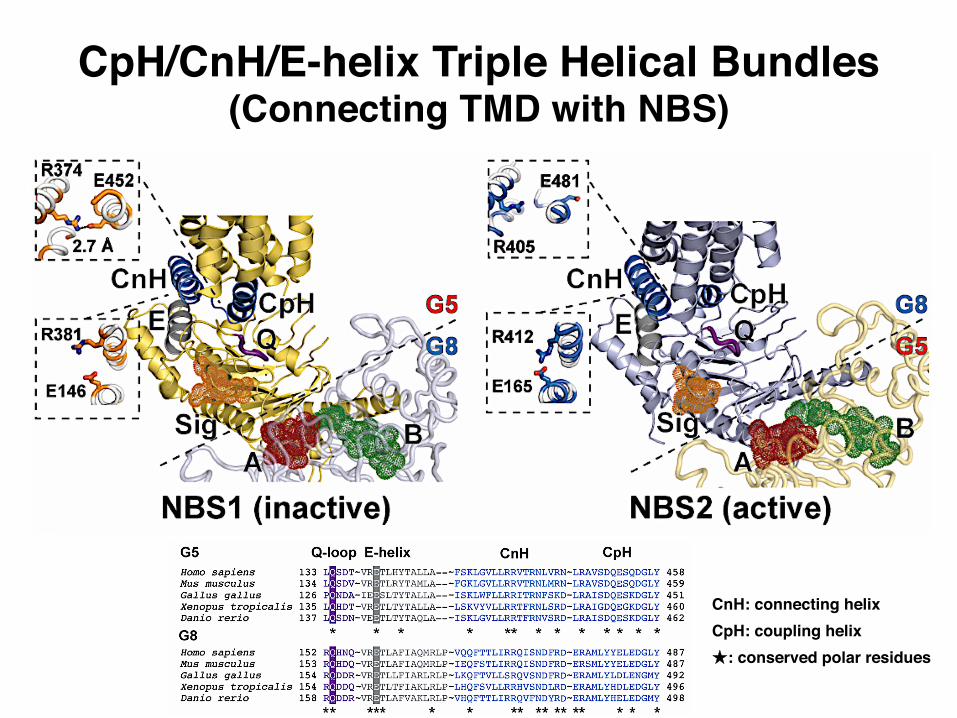
CpH/CnH/E-helix Triple Helical Bundles(Connecting TMD with NBS)
CnH: connecting helixCpH: coupling helix★: conserved polar residues
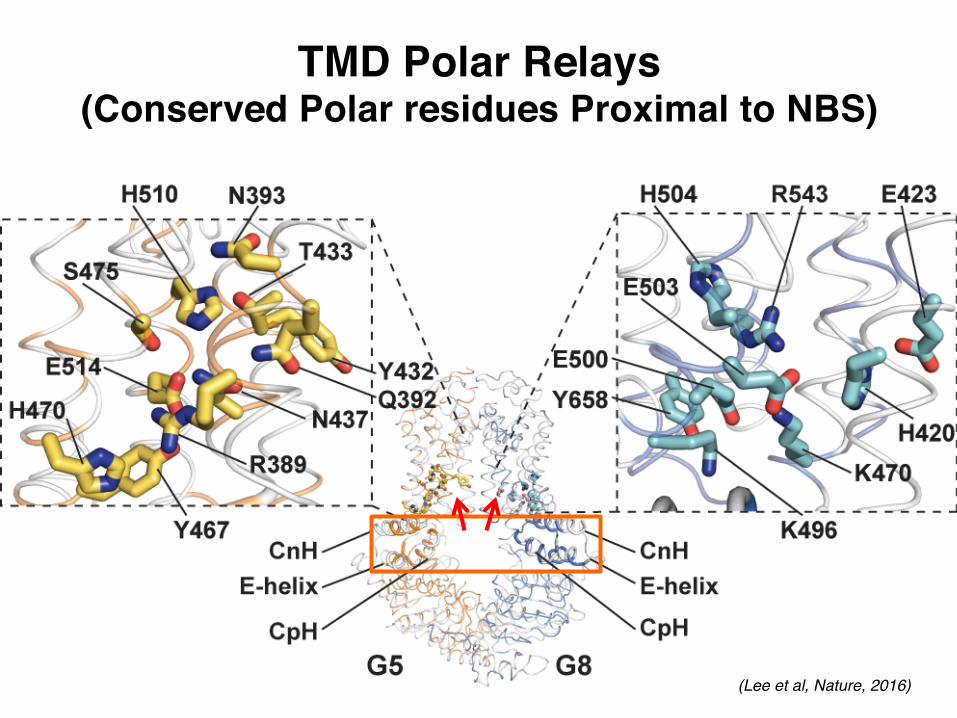
TMD Polar Relays(Conserved Polar residues Proximal to NBS)
(Lee et al, Nature, 2016)
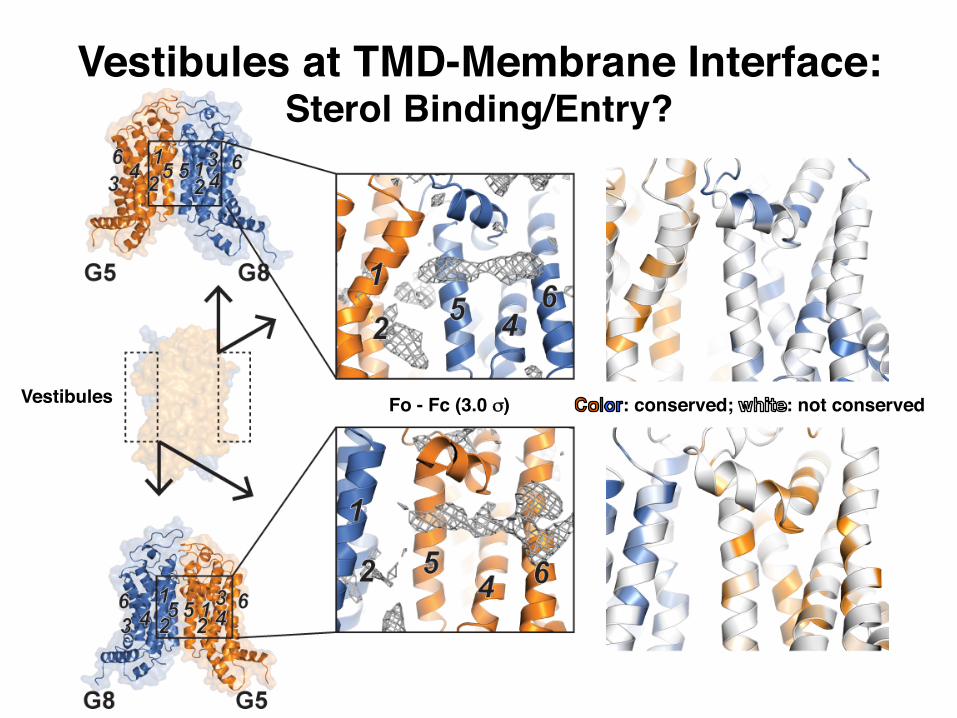
: conserved; : not conservedFo - Fc (3.0 σ)Vestibules
Vestibules at TMD-Membrane Interface:Sterol Binding/Entry?
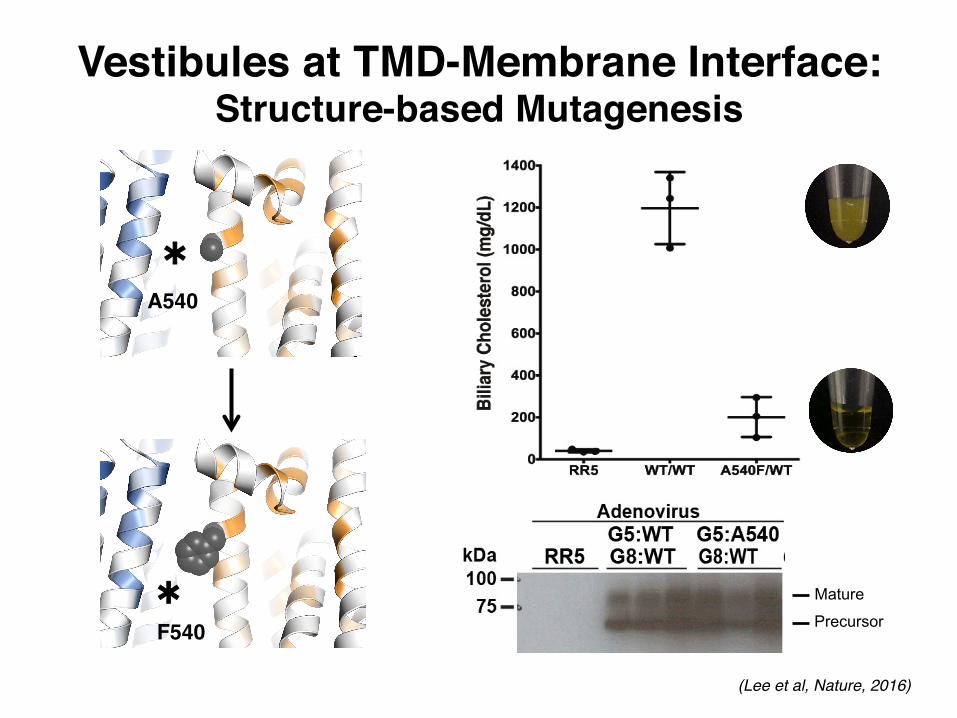
A540
F540*
*
LETTER RESEARCH
Extended Data Figure 5 | Vestibules in the membrane spanning region. a, b, Left: cartoon of G5G8 TMDs in transparent surface (orange and blue). Vestibules on opposing faces of the TMDs are highlighted in boxes. Middle: Fo − Fc difference electron density map contoured at 3.0σ showing extended features at the vestibules. TMHs are numbered. Right: highly conserved residues in G5 and G8 (sequence conservation ≥ 7, see Methods) are shown as orange (G5) and blue (G8) cartoon/sticks. c, Left: in vivo functional reconstitution assay using G5G8 KO mice (G5G8−/− mice)19. Biliary cholesterol levels from mice infected with RR
(empty adenovirus), or with adenoviruses expressing human (h)G5:WT and hG8:WT, or with adenoviruses expressing hG5:A540F and hG8:WT are shown. Pictures of the bile are below the graph. Each experiment represents the mean ± s.d. from three infected mice (n = 3) in each group. Right: expression of hG5 and hG8 detected by immunoblotting using anti-hG5 monoclonal antibodies (see Methods). Connexin was used as the gel-loading control. As a positive control for the mouse and a negative control for the human anti-G5 antibody, we also immunoblotted liver membranes from G5G8+/− mice.
© 2016 Macmillan Publishers Limited. All rights reserved
Precursor
Mature
Vestibules at TMD-Membrane Interface:Structure-based Mutagenesis
(Lee et al, Nature, 2016)
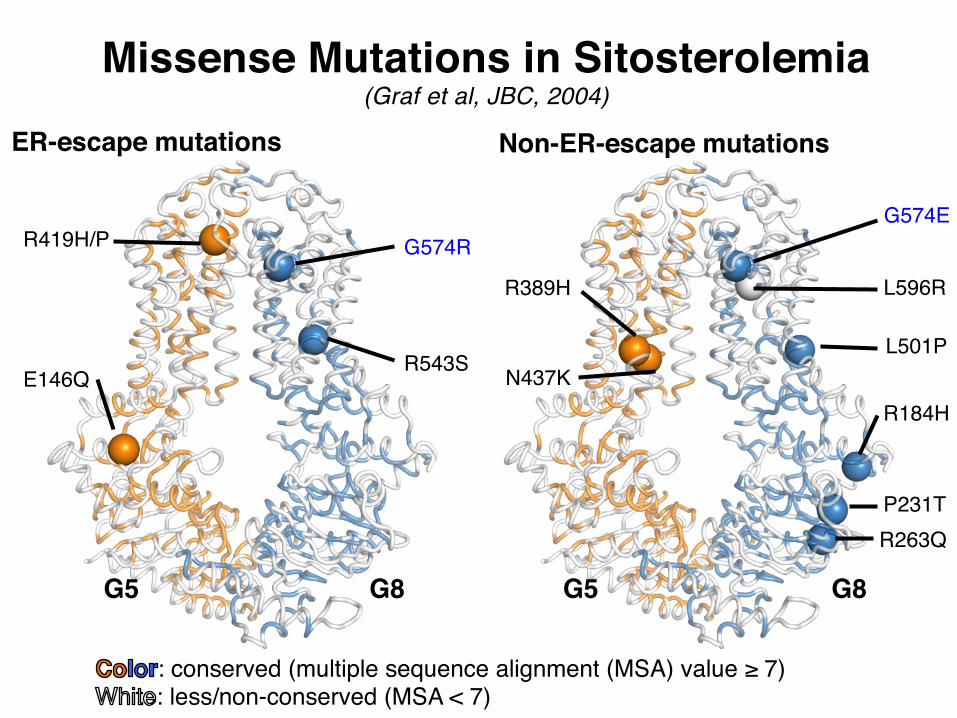
ER-escape mutations Non-ER-escape mutations
E146Q
R419H/P
R543S
G574R
R389H
N437KR184H
P231TR263Q
L501P
G574E
L596R
G5 G5G8 G8
Missense Mutations in Sitosterolemia(Graf et al, JBC, 2004)
: conserved (multiple sequence alignment (MSA) value ≥ 7): less/non-conserved (MSA < 7)
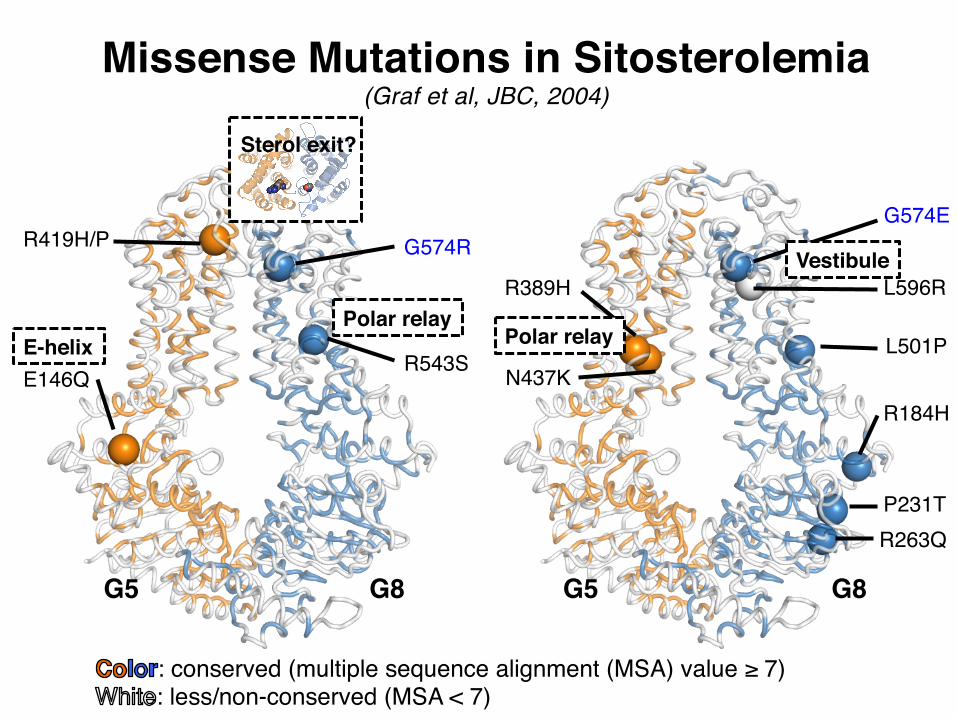
E146Q
R419H/P
R543S
G574R
R389H
N437KR184H
P231TR263Q
L501P
G574E
L596R
G5 G5G8 G8
Missense Mutations in Sitosterolemia(Graf et al, JBC, 2004)
: conserved (multiple sequence alignment (MSA) value ≥ 7): less/non-conserved (MSA < 7)
Polar relayPolar relayE-helix
Sterol exit?
Vestibule
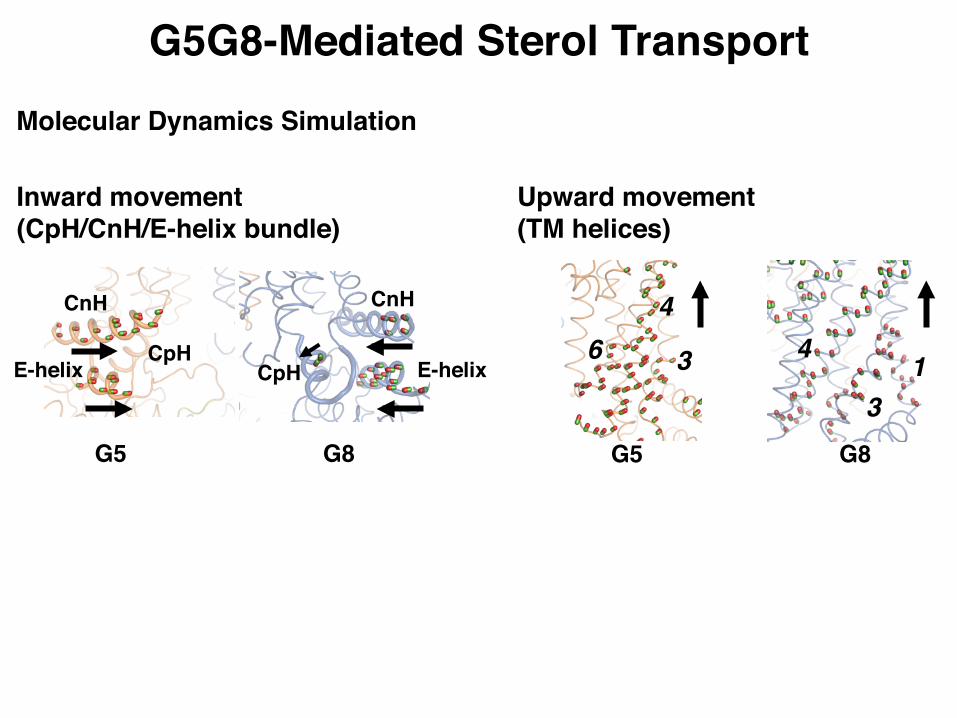
G5G8-Mediated Sterol Transport
Inward movement(CpH/CnH/E-helix bundle)
G5 G8
CnH
CpHE-helix
CnH
CpH E-helix
Upward movement(TM helices)
G5 G8
4
316
4
3
Molecular Dynamics Simulation
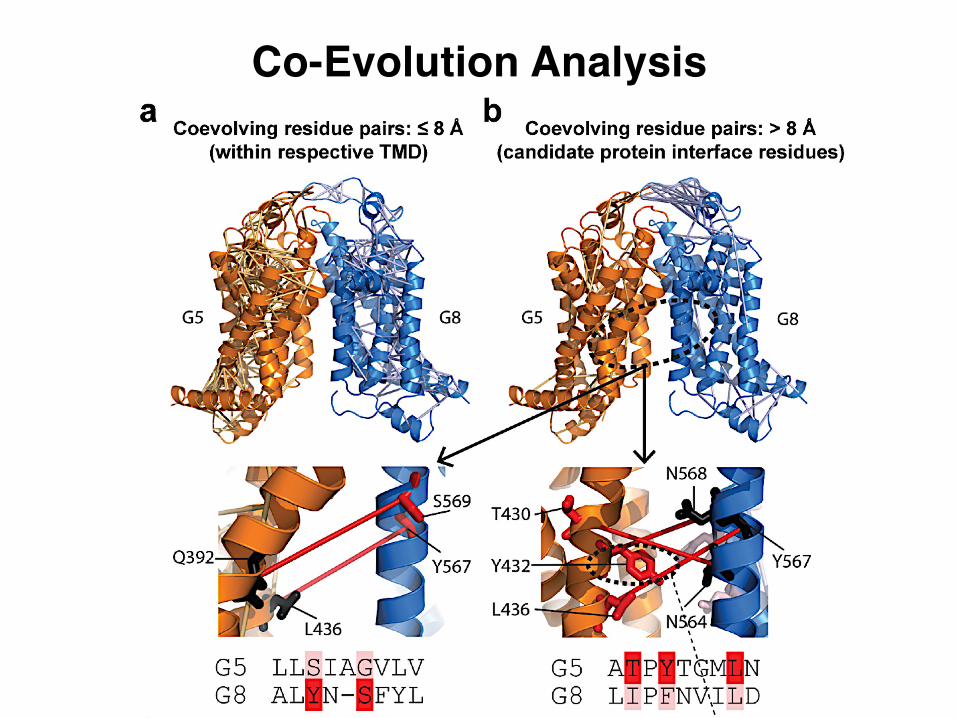
Co-Evolution Analysis
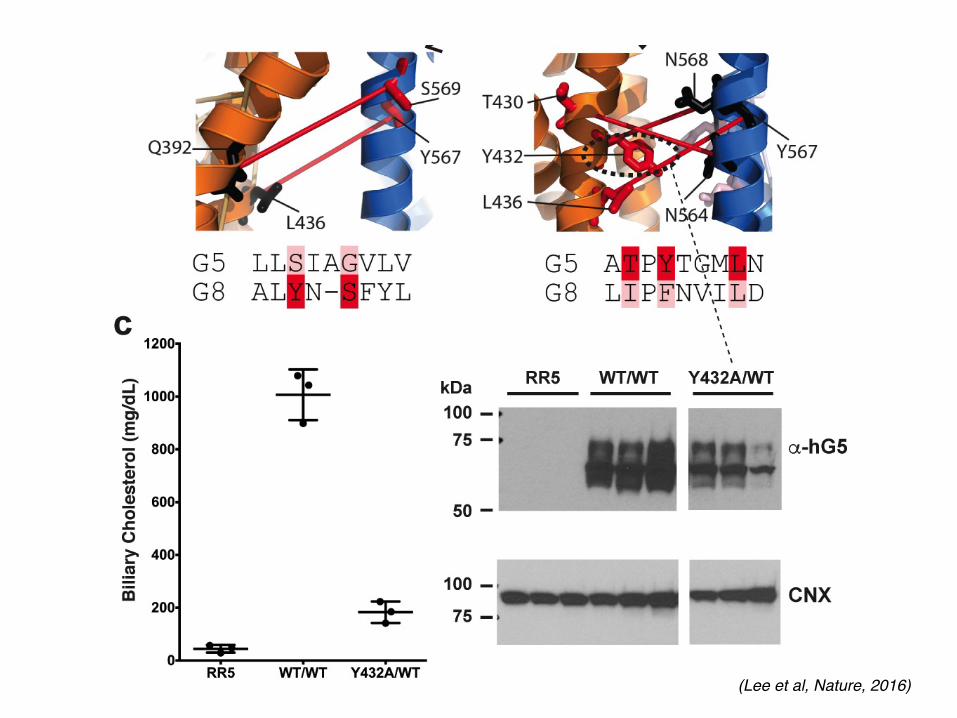
(Lee et al, Nature, 2016)
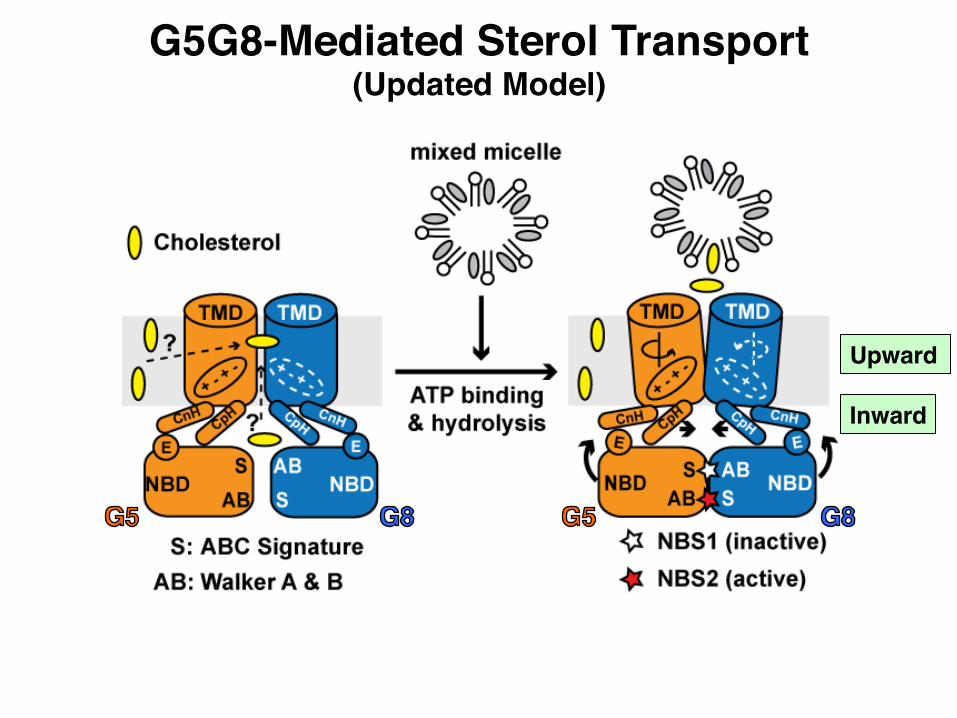
G5G8-Mediated Sterol Transport(Updated Model)
Inward
Upward

Summary
• G5G8 bicelle crystal structure at 3.9 Å(lipid bilayer-bound)
• New TMD fold for ABC transporters
• Cpn/CnH/E-helix bundle connects NBS to TMD• Asymmetric interaction between CnH & CpH• Conserved polar residues proximal to NBS
(TMD polar relays)
• Updated model for G5G8-mediated sterol transport• Structure-based mutagenesis at sterol-interacting site• Structure-based interpretation for disease-causing mutations

Vertebrates
Eukaryotic ABCG Exporters(TMD-based CLANS Network)
(Lee et al, Nature, 2016)

ABC2 Exporter Superfamily CLANS Network
(Lee et al, Nature, 2016)

Acknowledgement• UT Southwestern:
Helen HobbsJonathan Cohen
Daniel Rosenbaum
Zbyszek OtwinowskiDominika Borek
Nick GrishinLisa Kinch
Xiao-Song XieJin Wang
Junmei Wang
Structural Biology Lab (SBL)
• Texas Tech HSC:Ina Urbatsch
• Beamlines:APS 19ID, 23ID (Argonne)ALS 821, 822 (Berkeley)
• Funding:National Institute of HealthAmerican Heart AssociationHoward Hughes Medical Institute


















