
A thioredoxin-like/β-propeller protein maintains the ... · and “qI,” are separated based on...
8
A thioredoxin-like/β-propeller protein maintains the efficiency of light harvesting in Arabidopsis Matthew D. Brooks a,b,c , Emily J. Sylak-Glassman c,d , Graham R. Fleming c,d , and Krishna K. Niyogi a,b,c,1 a Howard Hughes Medical Institute, b Department of Plant and Microbial Biology, University of California, Berkeley, CA 94720; c Physical Biosciences Division, Lawrence Berkeley National Laboratory, Berkeley, CA 94720; and d Department of Chemistry, University of California, Berkeley, CA 94720 Edited by Elisabeth Gantt, University of Maryland, College Park, MD, and approved May 30, 2013 (received for review March 21, 2013) The light-harvesting complexes of plants have evolved the ability to switch between efficient light harvesting and quenching forms to optimize photosynthesis in response to the environment. Several distinct mechanisms, collectively termed “nonphotochemical quench- ing” (NPQ), provide flexibility in this response. Here we report the isolation and characterization of a mutant, suppressor of quenching 1 (soq1), that has high NPQ even in the absence of photosystem II subunit S (PsbS), a protein that is necessary for the rapidly reversible component of NPQ. The formation of NPQ in soq1 was light inten- sity-dependent, and it exhibited slow relaxation kinetics and other characteristics that distinguish it from known NPQ components. Treatment with chemical inhibitors or an uncoupler, as well as crosses to mutants known to affect other NPQ components, showed that the NPQ in soq1 does not require a transthylakoid pH gradient, zeaxanthin formation, or the phosphorylation of light-harvesting complexes, and it appears to be unrelated to the photosystem II damage-and-repair cycle. Measurements of pigments and chloro- phyll fluorescence lifetimes indicated that the additional NPQ in soq1 is the result of a decrease in chlorophyll excited-state lifetime and not pigment bleaching. The SOQ1 gene was isolated by map- based cloning, and it encodes a previously uncharacterized thyla- koid membrane protein with thioredoxin-like and β-propeller domains located in the lumen and a haloacid-dehalogenase domain exposed to the chloroplast stroma. We propose that the role of SOQ1 is to prevent formation of a slowly reversible form of antenna quenching, thereby maintaining the efficiency of light harvesting. P hotoautotrophic organisms harvest energy from the sun, sus- taining themselves and nearly all life on Earth. In many hab- itats, fluctuations between limiting and excess light conditions occur on a wide range of timescales, as predictable diurnal and seasonal changes are integrated with more random events such as cloud cover and shading. Plants form large light-harvesting an- tenna complexes with hundreds of pigment molecules that maxi- mize their ability to harvest energy in low light conditions. As light intensities increase, the rate of photosynthesis becomes saturated, and the excess energy that is absorbed can form damaging re- active oxygen species (1, 2). Therefore, plants have evolved many mechanisms, collectively termed “nonphotochemical quenching” (NPQ), to harmlessly dissipate excess energy absorbed in high light (3, 4). The different components of NPQ, termed “qE,”“qZ,”“qT,” and “qI,” are separated based on their induction and relaxation kinetics as well as by their dependence on known factors. Feed- back de-excitation, or qE, is the fastest component, turning on and relaxing within minutes. When the rate of light absorption exceeds that which can be used by the plant, a high pH gradient builds up across the thylakoid membrane, activating qE, which dissipates the excess energy as heat. The low pH in the thylakoid lumen proto- nates the photosystem II (PSII) subunit S (PsbS) protein (5) and activates violaxanthin de-epoxidase (VDE) (6, 7), which converts the pigment violaxanthin to zeaxanthin. In vivo, qE is known to require these three factors (the pH gradient, PsbS, and zeax- anthin), and in high light it can dissipate up to 75% of the energy absorbed by plants (8). Zeaxanthin-dependent quenching, or qZ, turns on and relaxes in tens of minutes (9). As implied by the name, qZ requires the formation of zeaxanthin, but it is independent of lumen pH once zeaxanthin is present, and it likely involves the minor antenna chlorophyll-binding protein (CP) 26 (10). A third component, state transitions, or qT, displays similar kinetics to qZ but functions to balance light absorption between photo- system I (PSI) and PSII (11) and requires the state transition kinase, STN7 (12). qT is more prominent during low light con- ditions, and is thought to be inhibited in high light via stromal redox regulation (13). NPQ that relaxes more slowly than the components described above is referred to as qI, or photoinhibitory quenching, which is the least-understood component of NPQ. qI is often assumed to be the result of oxidative photoinactivation of PSII reaction cen- ters. Damage to PSII increases linearly with light intensity (14), and its recovery involves synthesis and insertion of a new D1 polypeptide (15). However, the amount of NPQ due to qI often cannot be explained by the amount of reaction center damage (16). This indicates that other slowly relaxing and possibly pho- toprotective mechanisms exist that contribute to qI and are not yet fully characterized (3). An understanding of the different NPQ mechanisms is impor- tant in the effort to improve crop growth and productivity. Mod- eling of canopy photosynthesis has predicted that carbon gain of crop species is decreased by between 13% and 32% due to the slow relaxation kinetics of NPQ mechanisms and that speeding up re- covery could improve photosynthesis (17). Unfortunately, a lack of mechanistic details and genetic insights, particularly when it comes to the slowly relaxing components of NPQ, has limited our ability to study and manipulate these processes. Identification of addi- tional mutants that directly affect these NPQ mechanisms or their regulation is therefore critical to the field. Such mutants will allow researchers to dissect the various NPQ pathways, determine Significance Plants need to regulate light harvesting to match incoming light energy to photosynthetic capacity. Excess excitation can gener- ate reactive oxygen species and damage the photosynthetic machinery. To dissipate excess absorbed light energy, plants have evolved several quenching pathways, each of which acts on a different timescale. Here we identify a chloroplast protein involved in a previously uncharacterized, slowly reversible form of energy dissipation in the photosynthetic antenna. This mutant will be critical in characterizing this quenching pathway and will further our understanding of how plants are able to balance light harvesting and photoprotection to maximize photosynthetic efficiency. Author contributions: M.D.B., E.J.S.-G., G.R.F., and K.K.N. designed research; M.D.B. and E.J.S.-G. performed research; M.D.B., E.J.S.-G., and K.K.N. analyzed data; and M.D.B. and K.K.N. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence should be addressed. E-mail: [email protected]. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1305443110/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1305443110 PNAS | Published online July 1, 2013 | E2733–E2740 PLANT BIOLOGY PNAS PLUS
Transcript of A thioredoxin-like/β-propeller protein maintains the ... · and “qI,” are separated based on...

A thioredoxin-like/β-propeller protein maintains theefficiency of light harvesting in ArabidopsisMatthew D. Brooksa,b,c, Emily J. Sylak-Glassmanc,d, Graham R. Flemingc,d, and Krishna K. Niyogia,b,c,1
aHoward Hughes Medical Institute, bDepartment of Plant and Microbial Biology, University of California, Berkeley, CA 94720; cPhysical Biosciences Division,Lawrence Berkeley National Laboratory, Berkeley, CA 94720; and dDepartment of Chemistry, University of California, Berkeley, CA 94720
Edited by Elisabeth Gantt, University of Maryland, College Park, MD, and approved May 30, 2013 (received for review March 21, 2013)
The light-harvesting complexes of plants have evolved the ability toswitch between efficient light harvesting and quenching forms tooptimize photosynthesis in response to the environment. Severaldistinctmechanisms, collectively termed “nonphotochemical quench-ing” (NPQ), provide flexibility in this response. Here we report theisolation and characterization of a mutant, suppressor of quenching1 (soq1), that has high NPQ even in the absence of photosystem IIsubunit S (PsbS), a protein that is necessary for the rapidly reversiblecomponent of NPQ. The formation of NPQ in soq1 was light inten-sity-dependent, and it exhibited slow relaxation kinetics and othercharacteristics that distinguish it from known NPQ components.Treatment with chemical inhibitors or an uncoupler, as well ascrosses tomutants known to affect other NPQ components, showedthat the NPQ in soq1 does not require a transthylakoid pH gradient,zeaxanthin formation, or the phosphorylation of light-harvestingcomplexes, and it appears to be unrelated to the photosystem IIdamage-and-repair cycle. Measurements of pigments and chloro-phyll fluorescence lifetimes indicated that the additional NPQ insoq1 is the result of a decrease in chlorophyll excited-state lifetimeand not pigment bleaching. The SOQ1 gene was isolated by map-based cloning, and it encodes a previously uncharacterized thyla-koid membrane protein with thioredoxin-like and β-propellerdomains located in the lumen and a haloacid-dehalogenase domainexposed to the chloroplast stroma. We propose that the role ofSOQ1 is to prevent formation of a slowly reversible formof antennaquenching, thereby maintaining the efficiency of light harvesting.
Photoautotrophic organisms harvest energy from the sun, sus-taining themselves and nearly all life on Earth. In many hab-
itats, fluctuations between limiting and excess light conditionsoccur on a wide range of timescales, as predictable diurnal andseasonal changes are integrated with more random events such ascloud cover and shading. Plants form large light-harvesting an-tenna complexes with hundreds of pigment molecules that maxi-mize their ability to harvest energy in low light conditions. As lightintensities increase, the rate of photosynthesis becomes saturated,and the excess energy that is absorbed can form damaging re-active oxygen species (1, 2). Therefore, plants have evolved manymechanisms, collectively termed “nonphotochemical quenching”(NPQ), to harmlessly dissipate excess energy absorbed in highlight (3, 4).The different components of NPQ, termed “qE,” “qZ,” “qT,”
and “qI,” are separated based on their induction and relaxationkinetics as well as by their dependence on known factors. Feed-back de-excitation, or qE, is the fastest component, turning on andrelaxing within minutes. When the rate of light absorption exceedsthat which can be used by the plant, a high pH gradient builds upacross the thylakoidmembrane, activating qE, which dissipates theexcess energy as heat. The low pH in the thylakoid lumen proto-nates the photosystem II (PSII) subunit S (PsbS) protein (5) andactivates violaxanthin de-epoxidase (VDE) (6, 7), which convertsthe pigment violaxanthin to zeaxanthin. In vivo, qE is known torequire these three factors (the pH gradient, PsbS, and zeax-anthin), and in high light it can dissipate up to 75% of the energyabsorbed by plants (8). Zeaxanthin-dependent quenching, or qZ,turns on and relaxes in tens of minutes (9). As implied by the name,
qZ requires the formation of zeaxanthin, but it is independent oflumen pH once zeaxanthin is present, and it likely involves theminor antenna chlorophyll-binding protein (CP) 26 (10). A thirdcomponent, state transitions, or qT, displays similar kinetics toqZ but functions to balance light absorption between photo-system I (PSI) and PSII (11) and requires the state transitionkinase, STN7 (12). qT is more prominent during low light con-ditions, and is thought to be inhibited in high light via stromalredox regulation (13).NPQ that relaxes more slowly than the components described
above is referred to as qI, or photoinhibitory quenching, which isthe least-understood component of NPQ. qI is often assumed tobe the result of oxidative photoinactivation of PSII reaction cen-ters. Damage to PSII increases linearly with light intensity (14),and its recovery involves synthesis and insertion of a new D1polypeptide (15). However, the amount of NPQ due to qI oftencannot be explained by the amount of reaction center damage(16). This indicates that other slowly relaxing and possibly pho-toprotective mechanisms exist that contribute to qI and are not yetfully characterized (3).An understanding of the different NPQ mechanisms is impor-
tant in the effort to improve crop growth and productivity. Mod-eling of canopy photosynthesis has predicted that carbon gain ofcrop species is decreased by between 13%and 32%due to the slowrelaxation kinetics of NPQ mechanisms and that speeding up re-covery could improve photosynthesis (17). Unfortunately, a lack ofmechanistic details and genetic insights, particularly when it comesto the slowly relaxing components of NPQ, has limited our abilityto study and manipulate these processes. Identification of addi-tional mutants that directly affect these NPQ mechanisms or theirregulation is therefore critical to the field. Such mutants willallow researchers to dissect the various NPQ pathways, determine
Significance
Plants need to regulate light harvesting to match incoming lightenergy to photosynthetic capacity. Excess excitation can gener-ate reactive oxygen species and damage the photosyntheticmachinery. To dissipate excess absorbed light energy, plantshave evolved several quenching pathways, each of which actson a different timescale. Here we identify a chloroplast proteininvolved in a previously uncharacterized, slowly reversibleform of energy dissipation in the photosynthetic antenna. Thismutant will be critical in characterizing this quenching pathwayand will further our understanding of how plants are able tobalance light harvesting and photoprotection to maximizephotosynthetic efficiency.
Author contributions: M.D.B., E.J.S.-G., G.R.F., and K.K.N. designed research; M.D.B. andE.J.S.-G. performed research; M.D.B., E.J.S.-G., and K.K.N. analyzed data; and M.D.B. andK.K.N. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1305443110/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1305443110 PNAS | Published online July 1, 2013 | E2733–E2740
PLANTBIOLO
GY
PNASPL
US
their effect on growth and fitness, and probe the conformationalchanges that switch complexes between light-harvesting andenergy-dissipating forms.Previous forward genetics screens for npq mutants in Arabi-
dopsis have mostly identified mutants that specifically affect qE(18–20). These mutants, such as the npq4 mutant that lacks PsbS(19), have been critical for increasing our understanding of thisNPQ component and enabling new experimental approaches. Toisolate mutants that affect other components of NPQ, we per-formed a suppressor screen on mutagenized npq4 seedlings, whichlack qE. Here we describe the isolation and characterization of thesuppressor of quenching 1 (soq1) mutant, which can reach a level ofNPQ similar to that of the wild type. This mutant provides geneticinsights into the regulation of light harvesting and NPQ by iden-tifying a previously uncharacterized form of qI-like NPQ.
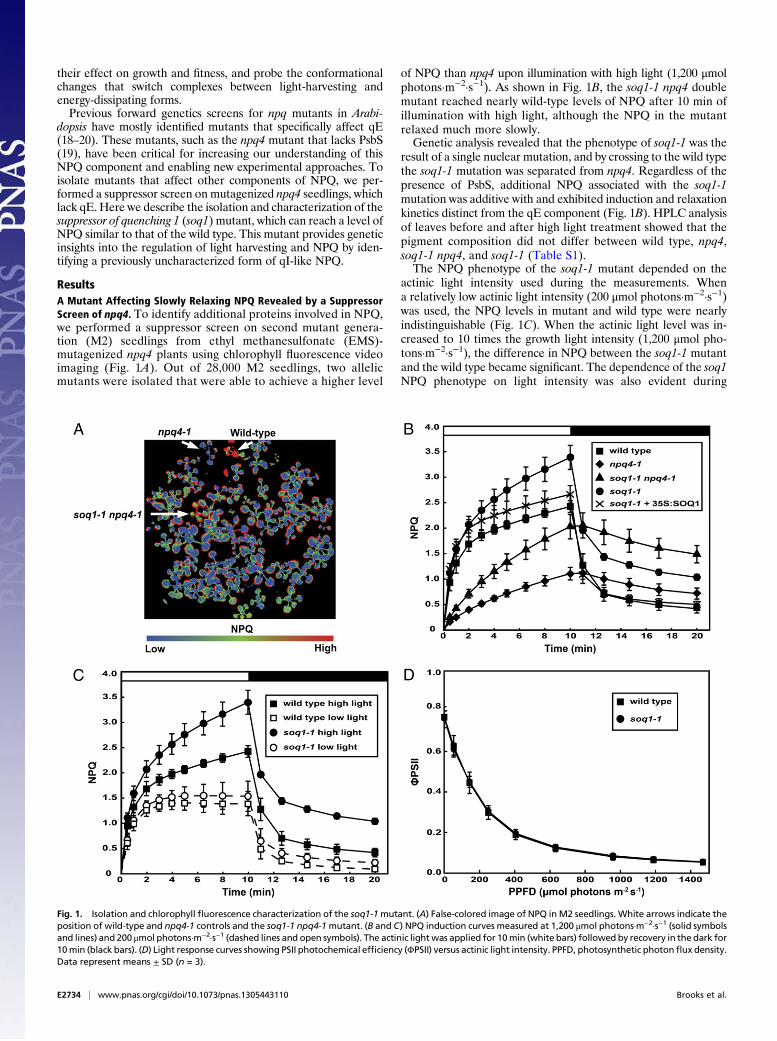
ResultsA Mutant Affecting Slowly Relaxing NPQ Revealed by a SuppressorScreen of npq4. To identify additional proteins involved in NPQ,we performed a suppressor screen on second mutant genera-tion (M2) seedlings from ethyl methanesulfonate (EMS)-mutagenized npq4 plants using chlorophyll fluorescence videoimaging (Fig. 1A). Out of 28,000 M2 seedlings, two allelicmutants were isolated that were able to achieve a higher level
of NPQ than npq4 upon illumination with high light (1,200 μmolphotons·m−2·s−1). As shown in Fig. 1B, the soq1-1 npq4 doublemutant reached nearly wild-type levels of NPQ after 10 min ofillumination with high light, although the NPQ in the mutantrelaxed much more slowly.Genetic analysis revealed that the phenotype of soq1-1 was the
result of a single nuclear mutation, and by crossing to the wild typethe soq1-1 mutation was separated from npq4. Regardless of thepresence of PsbS, additional NPQ associated with the soq1-1mutation was additive with and exhibited induction and relaxationkinetics distinct from the qE component (Fig. 1B). HPLC analysisof leaves before and after high light treatment showed that thepigment composition did not differ between wild type, npq4,soq1-1 npq4, and soq1-1 (Table S1).The NPQ phenotype of the soq1-1 mutant depended on the
actinic light intensity used during the measurements. Whena relatively low actinic light intensity (200 μmol photons·m−2·s−1)was used, the NPQ levels in mutant and wild type were nearlyindistinguishable (Fig. 1C). When the actinic light level was in-creased to 10 times the growth light intensity (1,200 μmol pho-tons·m−2·s−1), the difference in NPQ between the soq1-1 mutantand the wild type became significant. The dependence of the soq1NPQ phenotype on light intensity was also evident during
Fig. 1. Isolation and chlorophyll fluorescence characterization of the soq1-1mutant. (A) False-colored image of NPQ in M2 seedlings. White arrows indicate theposition of wild-type and npq4-1 controls and the soq1-1 npq4-1mutant. (B and C) NPQ induction curves measured at 1,200 μmol photons·m−2·s−1 (solid symbolsand lines) and 200 μmol photons·m−2·s−1 (dashed lines and open symbols). The actinic lightwas applied for 10min (white bars) followed by recovery in the dark for10min (black bars). (D) Light response curves showing PSII photochemical efficiency (ΦPSII) versus actinic light intensity. PPFD, photosynthetic photonflux density.Data represent means ± SD (n = 3).
E2734 | www.pnas.org/cgi/doi/10.1073/pnas.1305443110 Brooks et al.
experiments in which leaves were exposed to a series of in-creasing light intensities (Fig. S1A).Both soq1-1 and soq1-1 npq4 had no visible phenotype in either
low or high light growth conditions, and the maximum photochem-ical efficiency of PSII in the soq1-1 mutant was indistinguishablefrom that of the wild type (Fig. S2). Measurements of the PSIIoperating efficiency (ΦPSII) (Fig. 1D) and electron transport ratethrough PSII (Fig. S1D) in the wild type and soq1-1 indicated thatthere was no defect in photochemical pathways as the mutantplants were exposed to increasing light intensities.
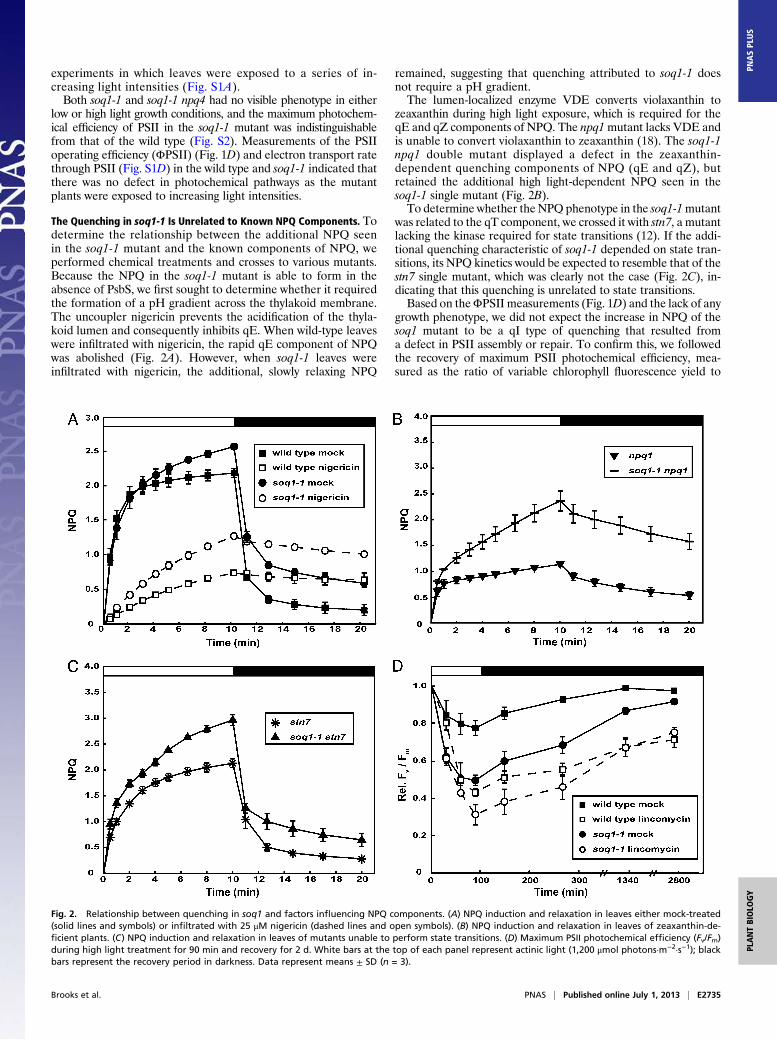
The Quenching in soq1-1 Is Unrelated to Known NPQ Components. Todetermine the relationship between the additional NPQ seenin the soq1-1 mutant and the known components of NPQ, weperformed chemical treatments and crosses to various mutants.Because the NPQ in the soq1-1 mutant is able to form in theabsence of PsbS, we first sought to determine whether it requiredthe formation of a pH gradient across the thylakoid membrane.The uncoupler nigericin prevents the acidification of the thyla-koid lumen and consequently inhibits qE. When wild-type leaveswere infiltrated with nigericin, the rapid qE component of NPQwas abolished (Fig. 2A). However, when soq1-1 leaves wereinfiltrated with nigericin, the additional, slowly relaxing NPQ
remained, suggesting that quenching attributed to soq1-1 doesnot require a pH gradient.The lumen-localized enzyme VDE converts violaxanthin to
zeaxanthin during high light exposure, which is required for theqE and qZ components of NPQ. The npq1mutant lacks VDE andis unable to convert violaxanthin to zeaxanthin (18). The soq1-1npq1 double mutant displayed a defect in the zeaxanthin-dependent quenching components of NPQ (qE and qZ), butretained the additional high light-dependent NPQ seen in thesoq1-1 single mutant (Fig. 2B).To determine whether theNPQ phenotype in the soq1-1mutant
was related to the qT component, we crossed it with stn7, a mutantlacking the kinase required for state transitions (12). If the addi-tional quenching characteristic of soq1-1 depended on state tran-sitions, its NPQ kinetics would be expected to resemble that of thestn7 single mutant, which was clearly not the case (Fig. 2C), in-dicating that this quenching is unrelated to state transitions.Based on theΦPSII measurements (Fig. 1D) and the lack of any
growth phenotype, we did not expect the increase in NPQ of thesoq1 mutant to be a qI type of quenching that resulted froma defect in PSII assembly or repair. To confirm this, we followedthe recovery of maximum PSII photochemical efficiency, mea-sured as the ratio of variable chlorophyll fluorescence yield to
Fig. 2. Relationship between quenching in soq1 and factors influencing NPQ components. (A) NPQ induction and relaxation in leaves either mock-treated(solid lines and symbols) or infiltrated with 25 μM nigericin (dashed lines and open symbols). (B) NPQ induction and relaxation in leaves of zeaxanthin-de-ficient plants. (C) NPQ induction and relaxation in leaves of mutants unable to perform state transitions. (D) Maximum PSII photochemical efficiency (Fv/Fm)during high light treatment for 90 min and recovery for 2 d. White bars at the top of each panel represent actinic light (1,200 μmol photons·m−2·s−1); blackbars represent the recovery period in darkness. Data represent means ± SD (n = 3).
Brooks et al. PNAS | Published online July 1, 2013 | E2735
PLANTBIOLO
GY
PNASPL
US
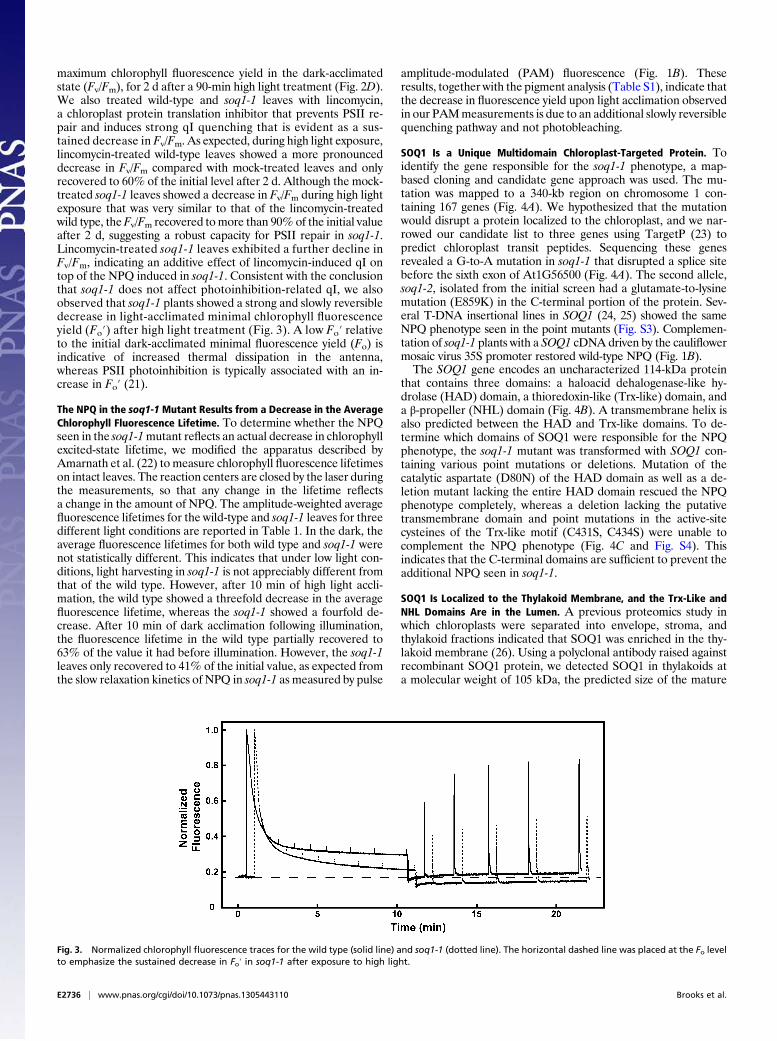
maximum chlorophyll fluorescence yield in the dark-acclimatedstate (Fv/Fm), for 2 d after a 90-min high light treatment (Fig. 2D).We also treated wild-type and soq1-1 leaves with lincomycin,a chloroplast protein translation inhibitor that prevents PSII re-pair and induces strong qI quenching that is evident as a sus-tained decrease in Fv/Fm. As expected, during high light exposure,lincomycin-treated wild-type leaves showed a more pronounceddecrease in Fv/Fm compared with mock-treated leaves and onlyrecovered to 60% of the initial level after 2 d. Although the mock-treated soq1-1 leaves showed a decrease in Fv/Fm during high lightexposure that was very similar to that of the lincomycin-treatedwild type, theFv/Fm recovered tomore than 90% of the initial valueafter 2 d, suggesting a robust capacity for PSII repair in soq1-1.Lincomycin-treated soq1-1 leaves exhibited a further decline inFv/Fm, indicating an additive effect of lincomycin-induced qI ontop of the NPQ induced in soq1-1. Consistent with the conclusionthat soq1-1 does not affect photoinhibition-related qI, we alsoobserved that soq1-1 plants showed a strong and slowly reversibledecrease in light-acclimated minimal chlorophyll fluorescenceyield (Fo′) after high light treatment (Fig. 3). A low Fo′ relativeto the initial dark-acclimated minimal fluorescence yield (Fo) isindicative of increased thermal dissipation in the antenna,whereas PSII photoinhibition is typically associated with an in-crease in Fo′ (21).
The NPQ in the soq1-1Mutant Results from a Decrease in the AverageChlorophyll Fluorescence Lifetime. To determine whether the NPQseen in the soq1-1mutant reflects an actual decrease in chlorophyllexcited-state lifetime, we modified the apparatus described byAmarnath et al. (22) to measure chlorophyll fluorescence lifetimeson intact leaves. The reaction centers are closed by the laser duringthe measurements, so that any change in the lifetime reflectsa change in the amount of NPQ. The amplitude-weighted averagefluorescence lifetimes for the wild-type and soq1-1 leaves for threedifferent light conditions are reported in Table 1. In the dark, theaverage fluorescence lifetimes for both wild type and soq1-1 werenot statistically different. This indicates that under low light con-ditions, light harvesting in soq1-1 is not appreciably different fromthat of the wild type. However, after 10 min of high light accli-mation, the wild type showed a threefold decrease in the averagefluorescence lifetime, whereas the soq1-1 showed a fourfold de-crease. After 10 min of dark acclimation following illumination,the fluorescence lifetime in the wild type partially recovered to63% of the value it had before illumination. However, the soq1-1leaves only recovered to 41% of the initial value, as expected fromthe slow relaxation kinetics of NPQ in soq1-1 as measured by pulse
amplitude-modulated (PAM) fluorescence (Fig. 1B). Theseresults, together with the pigment analysis (Table S1), indicate thatthe decrease in fluorescence yield upon light acclimation observedin our PAMmeasurements is due to an additional slowly reversiblequenching pathway and not photobleaching.
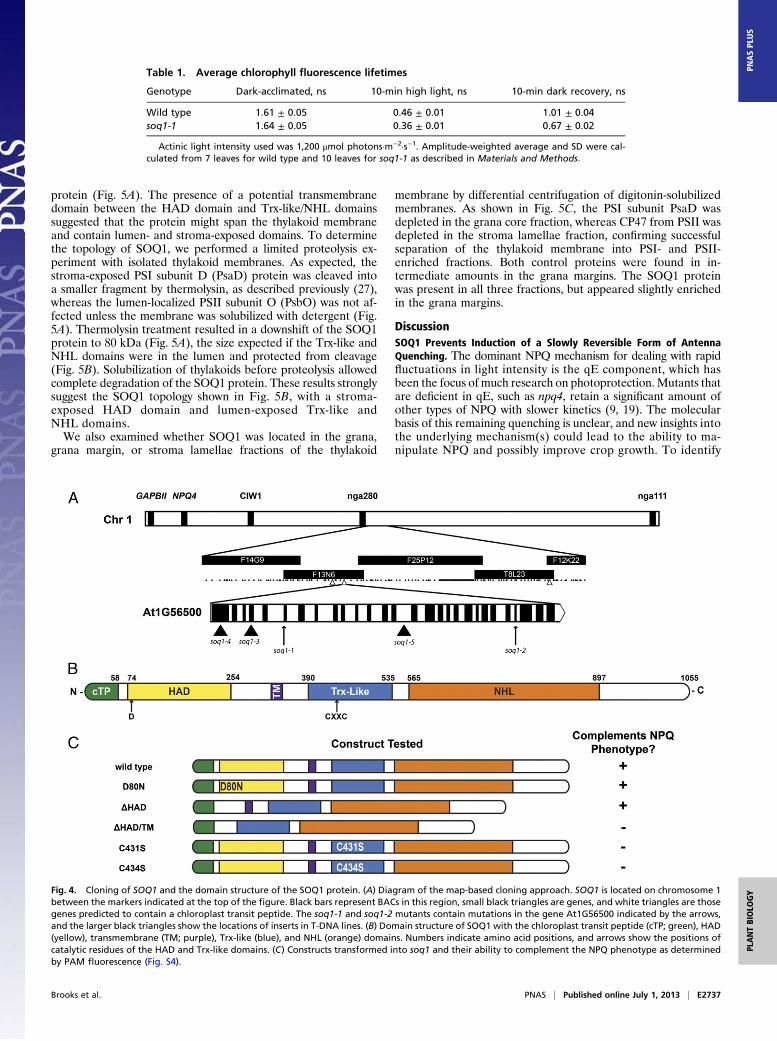
SOQ1 Is a Unique Multidomain Chloroplast-Targeted Protein. Toidentify the gene responsible for the soq1-1 phenotype, a map-based cloning and candidate gene approach was used. The mu-tation was mapped to a 340-kb region on chromosome 1 con-taining 167 genes (Fig. 4A). We hypothesized that the mutationwould disrupt a protein localized to the chloroplast, and we nar-rowed our candidate list to three genes using TargetP (23) topredict chloroplast transit peptides. Sequencing these genesrevealed a G-to-A mutation in soq1-1 that disrupted a splice sitebefore the sixth exon of At1G56500 (Fig. 4A). The second allele,soq1-2, isolated from the initial screen had a glutamate-to-lysinemutation (E859K) in the C-terminal portion of the protein. Sev-eral T-DNA insertional lines in SOQ1 (24, 25) showed the sameNPQ phenotype seen in the point mutants (Fig. S3). Complemen-tation of soq1-1 plants with a SOQ1 cDNA driven by the cauliflowermosaic virus 35S promoter restored wild-type NPQ (Fig. 1B).The SOQ1 gene encodes an uncharacterized 114-kDa protein
that contains three domains: a haloacid dehalogenase-like hy-drolase (HAD) domain, a thioredoxin-like (Trx-like) domain, anda β-propeller (NHL) domain (Fig. 4B). A transmembrane helix isalso predicted between the HAD and Trx-like domains. To de-termine which domains of SOQ1 were responsible for the NPQphenotype, the soq1-1 mutant was transformed with SOQ1 con-taining various point mutations or deletions. Mutation of thecatalytic aspartate (D80N) of the HAD domain as well as a de-letion mutant lacking the entire HAD domain rescued the NPQphenotype completely, whereas a deletion lacking the putativetransmembrane domain and point mutations in the active-sitecysteines of the Trx-like motif (C431S, C434S) were unable tocomplement the NPQ phenotype (Fig. 4C and Fig. S4). Thisindicates that the C-terminal domains are sufficient to prevent theadditional NPQ seen in soq1-1.
SOQ1 Is Localized to the Thylakoid Membrane, and the Trx-Like andNHL Domains Are in the Lumen. A previous proteomics study inwhich chloroplasts were separated into envelope, stroma, andthylakoid fractions indicated that SOQ1 was enriched in the thy-lakoid membrane (26). Using a polyclonal antibody raised againstrecombinant SOQ1 protein, we detected SOQ1 in thylakoids ata molecular weight of 105 kDa, the predicted size of the mature
Fig. 3. Normalized chlorophyll fluorescence traces for the wild type (solid line) and soq1-1 (dotted line). The horizontal dashed line was placed at the Fo levelto emphasize the sustained decrease in Fo′ in soq1-1 after exposure to high light.
E2736 | www.pnas.org/cgi/doi/10.1073/pnas.1305443110 Brooks et al.
protein (Fig. 5A). The presence of a potential transmembranedomain between the HAD domain and Trx-like/NHL domainssuggested that the protein might span the thylakoid membraneand contain lumen- and stroma-exposed domains. To determinethe topology of SOQ1, we performed a limited proteolysis ex-periment with isolated thylakoid membranes. As expected, thestroma-exposed PSI subunit D (PsaD) protein was cleaved intoa smaller fragment by thermolysin, as described previously (27),whereas the lumen-localized PSII subunit O (PsbO) was not af-fected unless the membrane was solubilized with detergent (Fig.5A). Thermolysin treatment resulted in a downshift of the SOQ1protein to 80 kDa (Fig. 5A), the size expected if the Trx-like andNHL domains were in the lumen and protected from cleavage(Fig. 5B). Solubilization of thylakoids before proteolysis allowedcomplete degradation of the SOQ1 protein. These results stronglysuggest the SOQ1 topology shown in Fig. 5B, with a stroma-exposed HAD domain and lumen-exposed Trx-like andNHL domains.We also examined whether SOQ1 was located in the grana,
grana margin, or stroma lamellae fractions of the thylakoid
membrane by differential centrifugation of digitonin-solubilizedmembranes. As shown in Fig. 5C, the PSI subunit PsaD wasdepleted in the grana core fraction, whereas CP47 from PSII wasdepleted in the stroma lamellae fraction, confirming successfulseparation of the thylakoid membrane into PSI- and PSII-enriched fractions. Both control proteins were found in in-termediate amounts in the grana margins. The SOQ1 proteinwas present in all three fractions, but appeared slightly enrichedin the grana margins.
DiscussionSOQ1 Prevents Induction of a Slowly Reversible Form of AntennaQuenching. The dominant NPQ mechanism for dealing with rapidfluctuations in light intensity is the qE component, which hasbeen the focus of much research on photoprotection.Mutants thatare deficient in qE, such as npq4, retain a significant amount ofother types of NPQ with slower kinetics (9, 19). The molecularbasis of this remaining quenching is unclear, and new insights intothe underlying mechanism(s) could lead to the ability to ma-nipulate NPQ and possibly improve crop growth. To identify
Fig. 4. Cloning of SOQ1 and the domain structure of the SOQ1 protein. (A) Diagram of the map-based cloning approach. SOQ1 is located on chromosome 1between the markers indicated at the top of the figure. Black bars represent BACs in this region, small black triangles are genes, and white triangles are thosegenes predicted to contain a chloroplast transit peptide. The soq1-1 and soq1-2 mutants contain mutations in the gene At1G56500 indicated by the arrows,and the larger black triangles show the locations of inserts in T-DNA lines. (B) Domain structure of SOQ1 with the chloroplast transit peptide (cTP; green), HAD(yellow), transmembrane (TM; purple), Trx-like (blue), and NHL (orange) domains. Numbers indicate amino acid positions, and arrows show the positions ofcatalytic residues of the HAD and Trx-like domains. (C) Constructs transformed into soq1 and their ability to complement the NPQ phenotype as determinedby PAM fluorescence (Fig. S4).
Table 1. Average chlorophyll fluorescence lifetimes
Genotype Dark-acclimated, ns 10-min high light, ns 10-min dark recovery, ns
Wild type 1.61 ± 0.05 0.46 ± 0.01 1.01 ± 0.04soq1-1 1.64 ± 0.05 0.36 ± 0.01 0.67 ± 0.02
Actinic light intensity used was 1,200 μmol photons·m−2·s−1. Amplitude-weighted average and SD were cal-culated from 7 leaves for wild type and 10 leaves for soq1-1 as described in Materials and Methods.
Brooks et al. PNAS | Published online July 1, 2013 | E2737
PLANTBIOLO
GY
PNASPL
US

proteins affecting slowly relaxing NPQ components, we screenedfor mutants in the npq4 background and isolated a mutant, soq1,that has higher NPQ (Fig. 1) and a corresponding decrease in theaverage chlorophyll fluorescence lifetime (Table 1). Multiple loss-of-function alleles suggest that the function of the wild-type SOQ1protein is to prevent formation of a slowly relaxing type of NPQwhen plants are exposed to high light.Previous screens have identified mutants with higher qE re-
sulting from additional cyclic electron flow (28), and recent studieson the npq4 mutant have shown that qE-like quenching can formin the absence of PsbS with the aid of chemicals that lower thelumen pH below levels normally attained in vivo (29). This led usto ask whether the quenching seen in soq1 is related to qE or otherknown components of NPQ. The NPQ relaxation kinetics in soq1suggested that the quenching was not the result of a pH gradientthat is sustained after turning off the actinic light (30), because theqE component was able to completely relax whereas the additionalquenching remained (Fig. 1B). Formation of the pH gradient wasalso unnecessary to produce the quenching in soq1, as shown bytreatment with the uncoupler nigericin (Fig. 2A). The xanthophyllcycle, which is necessary for maximal qE in vivo, is also not re-quired for the quenching in soq1, as demonstrated by crossing tothe npq1 mutant (Fig. 2B). Thus, the independence of soq1quenching from all of the three known qE requirements clearlydistinguishes this additional NPQ from qE. Although much less isknown about qZ, these results would also suggest that soq1 is notinvolved in this type of NPQ, because qZ also requires zeaxanthinformation. By showing that soq1-1 stn7 double mutants retain in-creased NPQ (Fig. 2C), we were also able to determine that statetransitions are not involved in the additional quenching seen insoq1 plants.Having ruled out the other known components of NPQ (qE, qZ,
and qT), it seemedmost likely that soq1 affects qI, a slowly relaxingNPQ component often associated with photoinhibition. However,soq1 does not appear to affect the PSII repair cycle that involvesdegradation and replacement of damaged D1 protein. Unlikeother repair mutants (31, 32), soq1 was able to grow in high light,
and the PSII quantum efficiency in soq1 plants did not deviatefrom the wild type during exposure to increasing light intensities(Fig. 1D). In addition, the maximum PSII efficiency in soq1 re-covered to a wild-type level, albeit slowly, after high light treat-ment (Fig. 2D). Finally, the fact that the Fo′ in soq1 remainedlower than the Fo (Fig. 3) indicates that enhanced, slowly re-versible energy dissipation in the antenna is competing with pho-tochemistry (21, 33).Together, these data support the conclusion that the addi-
tional quenching in soq1 is a form of antenna qI that is unrelatedto photoinhibitory qI. It has been observed that the amount ofdamaged PSII measured in plants treated with high light oftencannot account for the amount of qI-type NPQ (16), suggestingthat qI is heterogeneous and consists of multiple mechanisms.Photoprotective qI has been observed in many species, includingoverwintering evergreens and shade plants transferred to highlight (3). This quenching is correlated with zeaxanthin formation(34), and in some cases also requires a pH gradient (30). It isinteresting to speculate that the NPQ seen in soq1 mutants maybe a form of sustained quenching that is activated during pro-longed stress by down-regulation or inhibition of the SOQ1protein. Under normal growth conditions, this type of quenchingis likely to be detrimental, as its slow relaxation kinetics wouldcause it to compete with photochemistry during rapid changes inlight intensities. However, the slow relaxation kinetics could bea form of molecular memory preventing damage during chronichigh light exposure, similar to the role proposed for zeaxanthinin qE and qZ (10, 35) but with a more prolonged effect. Becausethe antenna qI in soq1 does not depend on zeaxanthin formation,it appears to be a previously undescribed form of sustained NPQ.Further experiments will be required to determine whether andunder what conditions this quenching occurs in wild-type plants.
The Trx-Like Domain in the Thylakoid Lumen Is Necessary for SOQ1Function. We mapped the soq1 mutations to a previously un-characterized gene encoding a chloroplast-targeted protein. TheSOQ1 protein contains three domains, with a transmembranehelix separating the HAD domain at the N terminus from the Trx-like and NHL domains in the C-terminal part of the protein. TheHAD superfamily is a large and diverse group of enzymes that usean aspartate as a nucleophile and are most often phosphatasesbut can perform several similar reactions (36). Thioredoxins areenzymes that contain conserved cysteines in a CXXC motif andcatalyze the oxidation or reduction of target proteins. The Trx-like
Fig. 5. Thylakoid localization and topology of SOQ1. (A) Protease pro-tection assay. Isolated thylakoids were treated with thermolysin (Th) in thepresence or absence of Triton X-100 (Tr). Aliquots were removed at thespecified times, samples were separated by SDS/PAGE, and immunode-tection was performed with the specified antibodies. (B) Schematic of thetopology suggested by A. (C) Thylakoids (Thy) were separated into grana(Gra), grana margins (Mar), and stroma lamallae (Str) fractions and sepa-rated by SDS/PAGE before immunodetection with the specified antibodies.
Fig. 6. Model for the role of SOQ1 in regulating light harvesting. Reducingpower generated by linear electron transport through PSI and ferredoxin (FD)is transferred from the stroma, possibly via ferredoxin-thioredoxin reductase(FTR) and Trx, either directly to SOQ1 (path 1) or through CCDA, a plant ho-molog of the bacterial control of cell death protein (path 2). We propose thatSOQ1 is involved in reducing luminal or lumen-exposed target proteins toprevent formation of slowly reversible antenna quenching, either directly orvia another protein (X). When SOQ1 is absent, one or more quenching site(s) (indicated by red stars) forms in the proximal and/or peripheral PSII an-tenna, which decreases the chlorophyll excited-state lifetime.
E2738 | www.pnas.org/cgi/doi/10.1073/pnas.1305443110 Brooks et al.

domain of SOQ1 falls into the thioredoxin-like protein (TlpA)-likefamily, which contains an ∼25-residue insert forming an extraα-helix and β-strand compared with the canonical thioredoxinmotif (37). Furthermore, all eukaryotic members of the TlpAfamily are in the disulfide isomerase-like protein (DipZ)-likesubfamily, some of which have a domain architecture similar to thedisulfide bond formation protein (DsbD) from Escherichia coli.The NHL family is found in prokaryotes and eukaryotes and isa member of the β-propeller clan, a group of structures that con-sists of multiple β-sheets forming blades that surround a centralchannel (38). The functions of β-propeller domains are diverse,with evidence of involvement in catalysis, signaling, ligand binding,protein structure, and protein–protein interactions.Our immunoblot analysis confirmed proteomics data showing
that SOQ1 is associated with thylakoids (Fig. 5A). Furthermore,we demonstrated, using a protease protection assay, that SOQ1 is athylakoid membrane protein with a stroma-exposed HAD domainand Trx-like and NHL domains located in the lumen (Fig. 5).Successful complementation of the NPQ phenotype in soq1 plantswith SOQ1 protein containing a mutation in the catalytic aspartateresidue or lacking the entire HAD domain (Fig. 4C) indicated thatthis domain is not necessary for the function of SOQ1 as a sup-pressor of antenna qI. It is possible that the HAD domain acts asa negative regulator of the luminal domains, or it might havea function in the stroma that is unrelated to NPQ. In contrast,directed mutagenesis showed that the CXXC motif of the Trx-likedomain is necessary for SOQ1 function (Fig. 4C), and the locationof the soq1-2 mutation suggests an important role also for theNHL domain (Fig. 4A).The lumen localization of the Trx-like domain is especially in-
triguing given that the characterized prokaryotic members of theTlpA-like family are found in the oxidizing periplasmic space,where they function indirectly in the reduction of oxidized me-thionine residues (39) or directly in the assembly of cytochromes(40–42) or maintenance of a metallochaperone in a reduced form(43). The reductant for these reactions likely comes from controlof cell death (CcdA) homologs either fused to the Trx-like domainor as a separate protein (39, 41). In bacteria, CcdA is a membraneprotein that receives reducing equivalents from thioredoxin in thecytoplasm and transfers them to periplasmic proteins involved incytochrome biogenesis, protein folding, or reduction of oxidizedcysteines (44). The plant homolog, CCDA, transfers reducingpower to the thylakoid lumen, a space analogous to the periplasm,and along with high chlorophyll fluorescence 164 (HCF164) isinvolved in the maturation of cytochrome f (45, 46). Mutantslacking either CCDA or HCF164 display a high fluorescencephenotype and are unable to grow photoautotrophically. Thesephenotypes are completely different from those described here forsoq1, indicating that SOQ1 is not involved in the cytochromepathway. However, it remains possible that SOQ1 receives re-ducing power from the stroma via CCDA and acts in anotherpathway to reduce luminal proteins (Fig. 6). We hypothesize thatSOQ1 is involved in maintaining target proteins in a reduced statein the oxidizing conditions of the thylakoid lumen, thereby pre-venting, directly or indirectly, the formationof a type of qI quenchingof excitation energy in the light-harvesting antenna (Fig. 6).The isolation and characterization of the soq1mutant presented
in this work open an area of research on the regulation of photo-synthesis. We anticipate that this mutant that affects photoprotec-tive qI will be extremely useful in characterizing both thebiophysical mechanism and physiological role of this type ofquenching. A better understanding of NPQ kinetics, and in par-ticular increasing the rate of relaxation of slower components suchas qI, could lead to greater photosynthetic output and plant growth.
Materials and MethodsStrains and Growth Conditions. The npq4-1 glabrous (gl)1-1 (19) and npq4-5(5) mutants (in the Col-0 wild-type background) were mutagenized with
EMS and grown at 150–200 μmol photons·m−2·s−1 as described (47). T-DNAinsertion alleles SALK_139953, SALK_099617, and SAIL_7_D05 (soq1-3, soq1-4,and soq1-5, respectively) were obtained from the Arabidopsis Biological Re-source Center (24, 25).
Chlorophyll Fluorescence Video Imaging Screen and PAM Fluorescence. Muta-genized seedlings were screened by chlorophyll fluorescence video imagingas described (48) using an actinic light intensity of 1,200 μmol photons·m−2·s−1
for 10 min. Video imaging screening for the complementation experimentsand crosses was done with the IMAGING-PAM M-Series (Heinz Walz). PAMfluorescence measurements were done using an FMS2 (Hansatech Instru-ments) or the Dual-PAM-100 (Heinz Walz) on 4- to 7-wk-old plants as de-scribed (49) under conditions indicated in the figure legends.
Mapping, Crosses, and Complementation. The soq1-1 npq4-1 mutant wascrossed to the Landsberg erecta (Ler) strain, and first filial generation (F1)plants were self-fertilized to generate a mapping population. Rough map-ping using the MapPairs primer set (Research Genetics) was done using aninitial set of 92 plants. Fine mapping was done using an additional 125second filial generation (F2) mutants and additional polymorphisms identi-fied between Ler and Col-0 (wild-type) ecotypes (50). Chloroplast-targetedgenes were identified using TargetP (23) and sequenced for mutations.
The soq1-1 npq4 mutant was crossed to Col-0 to obtain the soq1 mutationin a wild-type background. The single soq1-1 mutant was then used forcrosses to npq1 and stn7 and for complementation and subsequent experi-ments. Complementation was done by cloning a SOQ1 cDNA into pEarley-Gate100 (51), transforming soq1-1 plants with this construct using the floraldip method (52), and selecting for transformants on plates containing 10 μg/mL glufosinate ammonium. Inverse PCR was used to modify the binary vectorand generate the point mutations and domain deletions used in this work.
Chlorophyll Fluorescence Lifetime Measurements. Measurements were doneon medium-sized leaves from 5- to 6-wk-old plants using time-correlatedsingle-photon counting. The fluorescence lifetimes were measured at threetime points: after 30min of dark acclimation, after 10min of illumination withactinic light following the dark acclimation, and after 10 min of dark accli-mation following the illumination. This method is similar to that described byAmarnath et al. (22) with the following modifications. An actinic light in-tensity of 1,200 μmol photons·m2·s−1 was used to illuminate the leaves. At thesample position, the wavelength of the laser was 420 nm. The average powerof the laser at the sample was 5 mW with a pulse energy of 66 pJ. The laserimpinged at a 70° angle to the adaxial side of the leaf. Fluorescence countswere collected for 0.2 s following a period of preillumination by the laser. Thedetection wavelength was set to 684 nm with a monochromator (HORIBAJobin-Yvon; H-20). The detector (Hamamatsu; R3809-U) was computer-con-trolled (Becker & Hickl; DCC-100) with a gain set to 94%. At this gain, theinstrument response function was 35 ps. Single-photon timing was performedwith a specialized card (Becker & Hickl; SPC-630).
The experiment was repeated on 7 wild-type and 10 soq1-1 leaves, and thedata from each genotype’s leaf at a given light condition were summed. Thecurves were fit to a sum of three exponentials.
FðtÞ ¼ ∑iAie
−tτi ;
where the ith component has a fluorescence lifetime τi and an amplitude Ai
such that the sum of the amplitudes equals 1. Using the fit, we calculatedthe amplitude-weighted average lifetimes of wild-type and soq1-1 leaves.
τavg ¼ ∑iAiτi
The SDs of the values of τavg were calculated by performing bootstrappingon the fits.
A description of the sample holder and detection geometry and a fulljustification of the laser intensity and illumination time used will be detailedin a separate publication.
Protease Protection Assay, Thylakoid Subfractionation, and Immunoblots. Forprotease protection assays, thylakoids were isolated as described (53), ex-cept that homogenization of leaves was accomplished by eight 0.5-s pulsesin a precooled blender. Thylakoids were resuspended at 0.5 mg chlorophyllper mL in 0.3 M sorbitol, 2.5 mM EDTA, 5 mM MgCl2, 0.5% (wt/vol) BSA, 20mM Hepes (pH 7.6). Reactions were started by the addition of thermolysin(EMD Millipore) at 10 μg/mL to a final volume of 300 μL. At the indicatedtimes, the reaction was stopped by transferring 50 μL to a tube containingEDTA so that the final concentration of EDTA was 50 mM. The tubes were
Brooks et al. PNAS | Published online July 1, 2013 | E2739
PLANTBIOLO
GY
PNASPL
US

vortexed immediately and sample buffer was added. Separation of grana,grana margins, and stroma lamellae fractions was done as described pre-viously (54).
For immunoblots, samples were loaded by chlorophyll content (2 μg perlane) on a 10–20% Tris·glycine gel (Invitrogen), separated by SDS/PAGE,transferred to a nitrocellulose membrane, blocked with 5% (wt/vol) nonfatdry milk, and blotted with the antibodies indicated in the figure legends andtext. After incubation with HRP-conjugated secondary antibody, bands weredetected with ECL substrate (GE Healthcare).
ACKNOWLEDGMENTS. We thank Wallace Chan and Kevin Li for technicalassistance; the Arabidopsis Biological Resource Center for T-DNA insertionlines; and Kapil Amarnath, Alex Hertle, Lauriebeth Leonelli, Alizée Malnoë,and Julia Zaks for critical reading of the manuscript. This research was sup-ported by the Division of Chemical Sciences, Geosciences, and Biosciences,Office of Basic Energy Sciences, Office of Science, US Department of Energy(Field Work Proposal 449B). E.J.S.-G. was supported by a National ScienceFoundation Graduate Research Fellowship. K.K.N. is an Investigator of TheHoward Hughes Medical Institute and The Gordon and Betty Moore Foun-dation (through Grant GBMF3070).
1. Melis A (1999) Photosystem-II damage and repair cycle in chloroplasts: Whatmodulates the rate of photodamage? Trends Plant Sci 4(4):130–135.
2. Li Z, Wakao S, Fischer BB, Niyogi KK (2009) Sensing and responding to excess light.Annu Rev Plant Biol 60:239–260.
3. AdamsW, III, Zarter CR,Mueh K, Amiard V, Demmig-Adams B (2006), in Photoprotection,Photoinhibition, Gene Regulation, and Environment, eds Demmig-Adams B, Adams W,Mattoo A (Springer, Dordrecht, The Netherlands), pp 49–64.
4. Niyogi KK (1999) Photoprotection revisited: Genetic and molecular approaches. AnnuRev Plant Physiol Plant Mol Biol 50:333–359.
5. Li X-P, Phippard A, Pasari J, Niyogi KK (2002) Structure–function analysis ofphotosystem II subunit S (PsbS) in vivo. Funct Plant Biol 29(10):1131–1139.
6. Yamamoto HY, Bugos RC, Hieber AD (2004) The Photochemistry of Carotenoids, edsFrank HA, Young AJ, Britton G, Cogdell RJ (Springer, Dordrecht, The Netherlands), pp293–303.
7. Jahns P, Holzwarth AR (2012) The role of the xanthophyll cycle and of lutein inphotoprotection of photosystem II. Biochim Biophys Acta 1817(1):182–193.
8. Demmig-Adams B, et al. (1996) Using chlorophyll fluorescence to assess the fraction ofabsorbed light allocated to thermal dissipation of excess excitation. Physiol Plant98(2):253–264.
9. Nilkens M, et al. (2010) Identification of a slowly inducible zeaxanthin-dependentcomponent of non-photochemical quenching of chlorophyll fluorescence generatedunder steady-state conditions in Arabidopsis. Biochim Biophys Acta 1797(4):466–475.
10. Dall’Osto L, Caffarri S, Bassi R (2005) A mechanism of nonphotochemical energydissipation, independent from PsbS, revealed by a conformational change in theantenna protein CP26. Plant Cell 17(4):1217–1232.
11. Murata N (1969) Control of excitation transfer in photosynthesis. I. Light-inducedchange of chlorophyll a fluorescence in Porphyridium cruentum. Biochim BiophysActa 172(2):242–251.
12. Bellafiore S, Barneche F, Peltier G, Rochaix J-D (2005) State transitions and light adap-tation require chloroplast thylakoid protein kinase STN7. Nature 433(7028):892–895.
13. Rintamäki E, Martinsuo P, Pursiheimo S, Aro E-M (2000) Cooperative regulation oflight-harvesting complex II phosphorylation via the plastoquinol and ferredoxin-thioredoxin system in chloroplasts. Proc Natl Acad Sci USA 97(21):11644–11649.
14. Tyystjärvi E, Aro EM (1996) The rate constant of photoinhibition, measured inlincomycin-treated leaves, is directly proportional to light intensity. Proc Natl Acad SciUSA 93(5):2213–2218.
15. Yokthongwattana K, Melis A (2006) Photoprotection, Photoinhibition, GeneRegulation, and Environment, eds Demmig-Adams B, Adams W, Mattoo A (Springer,Dordrecht, The Netherlands), pp 175–191.
16. Walters R, Horton P (1993) Theoretical assessment of alternative mechanisms for non-photochemical quenching of PS II fluorescence in barley leaves. Photosynth Res 36(2):119–139.
17. Zhu XG, Ort DR, Whitmarsh J, Long SP (2004) The slow reversibility of photosystem IIthermal energy dissipation on transfer from high to low light may cause large lossesin carbon gain by crop canopies: A theoretical analysis. J Exp Bot 55(400):1167–1175.
18. Niyogi KK, Grossman AR, Björkman O (1998) Arabidopsis mutants define a centralrole for the xanthophyll cycle in the regulation of photosynthetic energy conversion.Plant Cell 10(7):1121–1134.
19. Li X-P, et al. (2000) A pigment-binding protein essential for regulation ofphotosynthetic light harvesting. Nature 403(6768):391–395.
20. Jung H-S, Niyogi KK (2010) Mutations in Arabidopsis YCF20-like genes affect thermaldissipation of excess absorbed light energy. Planta 231(4):923–937.
21. Demmig B, Björkman O (1987) Comparison of the effect of excessive light onchlorophyll fluorescence (77K) and photon yield of O2 evolution in leaves of higherplants. Planta 171(2):171–184.
22. Amarnath K, Zaks J, Park SD, Niyogi KK, Fleming GR (2012) Fluorescence lifetimesnapshots reveal two rapidly reversible mechanisms of photoprotection in live cells ofChlamydomonas reinhardtii. Proc Natl Acad Sci USA 109(22):8405–8410.
23. Emanuelsson O, Nielsen H, Brunak S, von Heijne G (2000) Predicting subcellularlocalization of proteins based on their N-terminal amino acid sequence. J Mol Biol300(4):1005–1016.
24. Sessions A, et al. (2002) A high-throughput Arabidopsis reverse genetics system. PlantCell 14(12):2985–2994.
25. Alonso JM, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsisthaliana. Science 301(5633):653–657.
26. Ferro M, et al. (2010) AT_CHLORO, a comprehensive chloroplast proteome databasewith subplastidial localization and curated information on envelope proteins. MolCell Proteomics 9(6):1063–1084.
27. Minai L, Cohen Y, Chitnis PR, Nechushtai R (1996) The precursor of PsaD assemblesinto the photosystem I complex in two steps. Proc Natl Acad Sci USA 93(13):6338–6342.
28. Livingston AK, Cruz JA, Kohzuma K, Dhingra A, Kramer DM (2010) An Arabidopsismutant with high cyclic electron flow around photosystem I (hcef) involving theNADPH dehydrogenase complex. Plant Cell 22(1):221–233.
29. Johnson MP, Ruban AV (2011) Restoration of rapidly reversible photoprotectiveenergy dissipation in the absence of PsbS protein by enhanced DeltapH. J Biol Chem286(22):19973–19981.
30. Gilmore A, Björkman O (1995) Temperature-sensitive coupling and uncoupling ofATPase-mediated, nonradiative energy dissipation: Similarities between chloroplastsand leaves. Planta 197(4):646–654.
31. Armbruster U, et al. (2010) The Arabidopsis thylakoid protein PAM68 is required forefficient D1 biogenesis and photosystem II assembly. Plant Cell 22(10):3439–3460.
32. Sun X, et al. (2010) The stromal chloroplast Deg7 protease participates in the repair ofphotosystem II after photoinhibition in Arabidopsis. Plant Physiol 152(3):1263–1273.
33. Krause GH (1988) Photoinhibition of photosynthesis. An evaluation of damaging andprotective mechanisms. Physiol Plant 74(3):566–574.
34. Verhoeven AS, Adams WW, Demmig-Adams B (1998) Two forms of sustainedxanthophyll cycle-dependent energy dissipation in overwintering Euonymuskiautschovicus. Plant Cell Environ 21(9):893–903.
35. Demmig-Adams B, Adams WW (1996) The role of xanthophyll cycle carotenoids in theprotection of photosynthesis. Trends Plant Sci 1(1):21–26.
36. Koonin EV, Tatusov RL (1994) Computer analysis of bacterial haloacid dehalogenasesdefines a large superfamily of hydrolases with diverse specificity. Application of aniterative approach to database search. J Mol Biol 244(1):125–132.
37. Capitani G, et al. (2001) Structure of the soluble domain of a membrane-anchoredthioredoxin-like protein from Bradyrhizobium japonicum reveals unusual properties.J Mol Biol 311(5):1037–1048.
38. Slack FJ, Ruvkun G (1998) A novel repeat domain that is often associated with RINGfinger and B-box motifs. Trends Biochem Sci 23(12):474–475.
39. Wu J, Neiers F, Boschi-Muller S, Branlant G (2005) The N-terminal domain of PILB fromNeisseria meningitidis is a disulfide reductase that can recycle methionine sulfoxidereductases. J Biol Chem 280(13):12344–12350.
40. Loferer H, Bott M, Hennecke H (1993) Bradyrhizobium japonicum TlpA, a novelmembrane-anchored thioredoxin-like protein involved in the biogenesis of cytochromeaa3 and development of symbiosis. EMBO J 12(9):3373–3383.
41. Edeling MA, Guddat LW, Fabianek RA, Thöny-Meyer L, Martin JL (2002) Structure ofCcmG/DsbE at 1.14 Å resolution: High-fidelity reducing activity in an indiscriminatelyoxidizing environment. Structure 10(7):973–979.
42. Crow A, Le Brun NE, Oubrie A (2005) The role of ResA in type II cytochrome cmaturation. Biochem Soc Trans 33(Pt 1):149–151.
43. Mohorko E, et al. (2012) Thioredoxin-like protein TlpA from Bradyrhizobiumjaponicum is a reductant for the copper metallochaperone ScoI. FEBS Lett 586(23):4094–4099.
44. Ito K, Inaba K (2008) The disulfide bond formation (Dsb) system. Curr Opin Struct Biol18(4):450–458.
45. Page MLD, et al. (2004) A homolog of prokaryotic thiol disulfide transporter CcdA isrequired for the assembly of the cytochrome b6f complex in Arabidopsis chloroplasts.J Biol Chem 279(31):32474–32482.
46. Lennartz K, et al. (2001) HCF164 encodes a thioredoxin-like protein involved in thebiogenesis of the cytochrome b(6)f complex in Arabidopsis. Plant Cell 13(11):2539–2551.
47. Li Z, et al. (2009) Lutein accumulation in the absence of zeaxanthin restoresnonphotochemical quenching in the Arabidopsis thaliana npq1 mutant. Plant Cell21(6):1798–1812.
48. Niyogi KK, Björkman O, Grossman AR (1997) Chlamydomonas xanthophyll cyclemutants identified by video imaging of chlorophyll fluorescence quenching. Plant Cell9(8):1369–1380.
49. Brooks MD, Niyogi KK (2011) Use of a pulse-amplitude modulated chlorophyllfluorometer to study the efficiency of photosynthesis in Arabidopsis plants. MethodsMol Biol 775:299–310.
50. Jander G, et al. (2002) Arabidopsis map-based cloning in the post-genome era. PlantPhysiol 129(2):440–450.
51. Earley KW, et al. (2006) Gateway-compatible vectors for plant functional genomicsand proteomics. Plant J 45(4):616–629.
52. Zhang X, Henriques R, Lin S-S, Niu Q-W, Chua N-H (2006) Agrobacterium-mediatedtransformation of Arabidopsis thaliana using the floral dip method. Nat Protoc 1(2):641–646.
53. Casazza AP, Tarantino D, Soave C (2001) Preparation and functional characterizationof thylakoids from Arabidopsis thaliana. Photosynth Res 68(2):175–180.
54. Kyle DJ, Staehelin LA, Arntzen CJ (1983) Lateral mobility of the light-harvestingcomplex in chloroplast membranes controls excitation energy distribution in higherplants. Arch Biochem Biophys 222(2):527–541.
E2740 | www.pnas.org/cgi/doi/10.1073/pnas.1305443110 Brooks et al.


















