1 Running head: leaf-water δ18O in atmospheric CAM epiphytes · This shift lead to morphological...
51
1 Running head: leaf-water δ 18 O in atmospheric CAM epiphytes 1 2 Brent R. Helliker 3 Department of Biology 4 University of Pennsylvania 5 433 S. University Ave. 6 Philadelphia, PA 19104 7 Tel: 215-746-6217 8 Fax: 215-898-8780 9 Email: [email protected] 10 11 12 Whole Plant and Ecophysiology 13 14 15 Plant Physiology Preview. Published on February 7, 2011, as DOI:10.1104/pp.111.172494 Copyright 2011 by the American Society of Plant Biologists www.plantphysiol.org on May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
Transcript of 1 Running head: leaf-water δ18O in atmospheric CAM epiphytes · This shift lead to morphological...

1
Running head: leaf-water δ18O in atmospheric CAM epiphytes 1
2
Brent R. Helliker 3
Department of Biology 4
University of Pennsylvania 5
433 S. University Ave. 6
Philadelphia, PA 19104 7
Tel: 215-746-6217 8
Fax: 215-898-8780 9
Email: [email protected] 10
11 12 Whole Plant and Ecophysiology 13 14
15
Plant Physiology Preview. Published on February 7, 2011, as DOI:10.1104/pp.111.172494
Copyright 2011 by the American Society of Plant Biologists
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

2
On the controls of leaf-water δ18O in the atmospheric CAM epiphyte Tillandsia 16
usneoides. 17
18
19
Brent R. Helliker 20
Department of Biology, University of Pennsylvania, 415 S. University Ave, Philadelphia, 21
PA 19104. 22
23
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

3
Financial source: 24
This work was supported by the National Science Foundation under award number IOB-25
0615501. Any opinions, findings and conclusions or recommendations expressed in this 26
material are those of the author(s) and do not necessarily reflect those of the National 27
Science Foundation. 28
29
Corresponding author: Brent R. Helliker, [email protected] 30
31
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

4
32
Abstract 33
Previous theoretical work showed that leaf-water isotope ratio (δ18OL) of CAM 34
epiphytes was controlled by the δ18O of atmospheric water vapor (δ18Oa), and 35
observed δ18OL could be explained by both a non-steady state model and a 36
‘maximum enrichment’ steady-state model (δ18OL-M); the latter requiring only δ18Oa 37
and relative humidity (h) as inputs. δ18OL should therefore contain an extractable 38
record of δ18Oa. Previous empirical work supported this hypothesis, but raised 39
many questions: How does changing δ18Oa and h affect δ18OL? Do hygroscopic 40
trichomes affect observed δ18OL? Are observations of changes in water content (W) 41
required for prediction of δ18OL? Does the leaf need to be at full isotopic steady state 42
for observed δ18OL to equal δ18OL-M? These questions were examined with a climate-43
controlled experimental system capable of holding δ18Oa constant for several weeks. 44
Water adsorbed to trichomes required a correction ranging from 0.5 to 1 ‰. δ18OL 45
could be predicted using constant values of W and even total conductance (gtot). 46
Tissue rehydration caused a transitory change in δ18OL but the consequent increase 47
in gtot led to a tighter coupling with δ18Oa. The non-steady-state leaf water models 48
explained observed δ18OL (y = 0.93*x - 0.07, R2 = 0.98) over a wide range of δ18Oa 49
and h. Predictions of δ18OL-M agreed with observations of δ18OL (y = 0.87*x - 0.99, R2 50
= 0.92), and when h > 0.9, the leaf did not need to be at isotopic steady state for the 51
δ18OL-M model to predict δ18OL in the CAM epiphyte Tillandsia usneoides. 52
53
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

5
Introduction 54
Tropical and subtropical epiphytic higher plants have long been a curiosity to plant 55
physiologists because of the multiple and unique constraints on physiology that the 56
epiphytic lifeform represents (Mez, 1904; Benzing, 1970; Medina and Troughton, 1974). 57
While the competition for light has no doubt lead the plants to the tree tops, the 58
concomitant loss of roots effectively removed the plants from an environment of 59
abundant rainfall to one of arid conditions (Griffiths et al., 1986; Smith et al., 1986; 60
Winter and Smith, 1996). In a sense, the plants shifted biomes by immigrating vertically 61
instead of horizontally. This shift lead to morphological adaptations like the development 62
of modified leaf hairs to absorb water and the formation of tanks via the overlapping of 63
leaf bases as well as physiological adaptations such as maintaining high leaf osmotic 64
potential and, perhaps most notably, the evolution of the CO2 concentrating mechanism 65
known as Crassulacean acid metabolism (CAM) photosynthesis (Medina and Troughton, 66
1974; Benzing et al., 1976; Griffiths and Smith, 1983; Smith et al., 1986; Martin and 67
Schmitt, 1989; Martin et al., 2004; Ohrui et al., 2007). This unique physiology of tropical 68
and subtropical CAM epiphytes also presents an intriguing contrast to the general way we 69
view oxygen isotope ratios (δ18O) in plant water and organic material (Helliker and 70
Griffiths, 2007; Helliker and Noone, 2009). 71
72
The enrichment of 18O in leaf water during plant transpiration leads to a suite of 73
physiology-based tracers that inform us of current and past plant-environment 74
interactions. Leaf water δ18O (δ18OL) labels CO2 and allows for partitioning of the gross 75
components of net CO2 flux from ecosystem to regional scales (Yakir and Wang, 1996; 76
Ciais et al., 1997; Styles et al., 2002; Cuntz et al., 2003; Ogee et al., 2004; Helliker et al., 77
2005). The production of isotopically distinct O2 by photosynthesis allows for global-78
scale estimates of productivity over millennia (Guy et al., 1993; Luz et al., 1999; 79
Hoffmann et al., 2004). The isotopic record of δ18OL in leaf and tree ring cellulose allows 80
for the reconstruction of growth environment and/or physiological responses to that 81
growth environment. (Epstein et al., 1977; Anderson et al., 1998; Switsur and 82
Waterhouse, 1998; Barbour and Farquhar, 2000; Barbour et al., 2000; Roden and 83
Ehleringer, 2000; Ferrio and Voltas, 2005; Poussart and Schrag, 2005; Helliker and 84
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

6
Richter, 2008). Much of the error associated with the above δ18O applications could be 85
decreased with better estimates of the isotope ratio of atmospheric water vapor (δ18Oa), a 86
primary control on leaf-water 18O enrichment (Farquhar and Cernusak, 2005; Helliker 87
and Griffiths, 2007), and the study of δ18OL in CAM epiphytes may offer better estimates 88
of δ18Oa through time and space (Helliker and Griffiths, 2007). 89
90
The epiphyte— and specifically the CAM vascular epiphyte— offers an extreme in the 91
way δ18O in plant water and organic material is viewed. For most vascular plants, δ18OL 92
is controlled by a balance of soil water δ18O and δ18Oa; soil water comes into the leaves 93
via the root system and atmospheric vapor diffuses into to leaf through open stomata. The 94
balance of the two water sources is determined by relative humidity (h); if h is high then 95
δ18OL is controlled proportionally more by δ18Oa, if h is unity then δ18OL is at equilibrium 96
with δ18Oa. If these plants happen to lose water during a humid night, then leaf water 97
exchange can lead to control by— and possibly equilibrium with— δ18Oa (Lai et al., 98
2008). Typically, the dramatic increase of transpiration during the daytime moves the 99
δ18OL away from equilibrium with δ18Oa. Hence, the nocturnal movement of δ18OL 100
towards equilibrium with δ18Oa in most vascular plants is not likely recorded in plant 101
organic material and is only important to the isotopic composition of CO2 during 102
nocturnal respiration (Cernusak et al., 2004). In the CAM epiphyte, rainwater and dewfall 103
rehydrates epiphyte tissue, and due to stomata being open at night when relative humidity 104
is near unity, rehydration water is quickly exchanged with atmospheric water vapor. This 105
leads to the situation, hypothesized by Helliker and Griffiths (2007), where rainfall and 106
dewfall (which are in equilibrium with δ18Oa; (Gat, 1996)) control tissue water status, and 107
the δ18O of this tissue water is continuously controlled by δ18Oa through subsequent 108
nocturnal vapor exchange. Because stomata are typically closed during the day, organic 109
material synthesized by the photosynthetic carbon reduction cycle should obtain the δ18O 110
signature of δ18Oa-controlled leaf water (Helliker and Griffiths, 2007). It is this situation 111
that yields new applications for δ18O that is unique to epiphytes: the reconstruction of 112
physiological responses to the epiphytic growth habit (Reyes-Garcia et al., 2008) and the 113
reconstruction of δ18Oa (Helliker and Griffiths, 2007). It was recently shown through 114
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

7
empirical and theoretical work that lichen thalli obtain equilibrium with δ18Oa even under 115
non-saturating conditions (Hartard et al., 2009). The lichen system of equilibrium with 116
δ18Oa represents an important contrast to that of CAM epiphytes, and this is discussed 117
later. The theoretical work presented by Hartard et al., however, is extremely helpful for 118
the interpretation of isotopic signals in CAM epiphytes. 119
120
Helliker and Griffiths (2007) developed the theoretical underpinnings for δ18Oa as a 121
control on CAM epiphyte δ18OL. They extended the thoughts of Farquhar and Cernusak 122
(2005) to show that at the high nocturnal relative humidity (h) experienced by CAM 123
epiphytes, gross exchange fluxes of water vapor into the leaf from the atmosphere can be 124
several times the transpirational flux out of the leaf. Model simulations showed that 125
constant environmental conditions and continuous water loss lead to an isotopic steady 126
state where both δ18OL and transpired water (δ18OE) converged to a single isotopic value 127
controlled by the exchange of δ18Oa and described by the following ‘maximum 128
enrichment’ equation (Farquhar and Gan, 2003; Helliker and Griffiths, 2007; Hartard et 129
al., 2009): 130
131
RL−M ≈ Rah1
α *− α K + α K h
132
133
Where the steady-state δ18OL (RL) is determined solely by δ18Oa (Ra), h, the temperature 134
dependent equilibrium fractionation factor α*, and the balance of the ratio of diffusivities 135
of light to heavy water molecules through the stomata and through the leaf boundary 136
layer, αK (Flanagan et al., 1991; Farquhar and Lloyd, 1993; Farquhar and Cernusak, 137
2005). The non-steady state analytical solution for δ18OL developed in by Hartard et al. 138
(2009; Equation 6 in Materials and Methods, hereafter referred to as the Hartard-Cuntz 139
solution) clearly demonstrates that through time in the non-steady state δ18OL is 140
continuously moving towards the value of RL-M (δ18OL-M). In a similar manner, the 141
simulations of Helliker and Griffiths suggested that even at values of nocturnal h of 0.8— 142
which is low for a CAM epiphyte (Garth, 1964; Smith et al., 1986)— δ18OL was 143
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

8
controlled almost entirely by the isotope ratio of atmospheric water vapor and that the 144
maximum enrichment equation above would ultimately predict δ18OL. The simulations 145
also showed that the approach to steady was faster and occurred through less water loss 146
as h increased. In general, their simulations showed that δ18OL approached the steady-147
state value, or δ18OL-M, much faster than δ18OE and this yields an important prediction 148
that is tested by current study: the leaf does not need to be at full isotopic steady state for 149
δ18OL to be at or near the steady-state, maximum enrichment value (δ18OL-M). 150
151
There are many unresolved questions as to the agreement between models and 152
observations that underpin the efficacy of using δ18OL to reconstruct δ18Oa. The 153
empirical work of Helliker and Griffiths (2007) did show support for the modeling 154
exercises, but only over a narrow range of conditions. Their experimental setup was 155
limited as it was developed to demonstrate a preliminary proof of concept. Also, the 156
invariably high h and noisy δ18Oa (± 1.2 ‰) of Helliker and Griffiths could lead one to 157
erroneously conclude that δ18OL was in direct equilibrium with δ18Oa even when h was 158
less than unity, which should not be the case. The primary controls of CAM epiphyte 159
δ18OL are h and δ18Oa and the manner in which changes in δ18Oa and h affect δ18OL 160
through time must be assessed. Like many CAM epiphytes, the study species Tillandsia 161
usneoides has a heavy covering of hygroscopic trichomes. While the water adsorbed to 162
these trichomes does not lead to water uptake by living cells at sub-saturating conditions 163
(Martin and Schmitt, 1989), the water is inevitably sampled and extracted for δ18OL 164
determination. Hence, the isotopic offset caused by these trichomes must be determined. 165
Total plant conductance to water loss (gtot; stomatal and boundary layer conductance) 166
controls the rate of exchange of tissue water with water vapor, and the relative water 167
content (RWC) of the plant. Therefore the effect of changes in RWC on δ18OL — through 168
both water loss and rehydration— must be determined. Previous work has shown that 169
changes in water volume are not important to predictions of δ18OL (Cernusak et al., 2002; 170
Cuntz et al., 2007) and a similar finding in CAM epiphytes could greatly simplify our 171
interpretation of observed δ18OL. In summary, the goal of this study was to construct 172
controlled-environment experiments to assess how well observed δ18OL could be 173
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

9
predicted over a range of conditions through both steady state and non-steady state 174
approaches of Helliker and Griffiths (2007) and Hartard et al. (2009). 175
176
Results 177
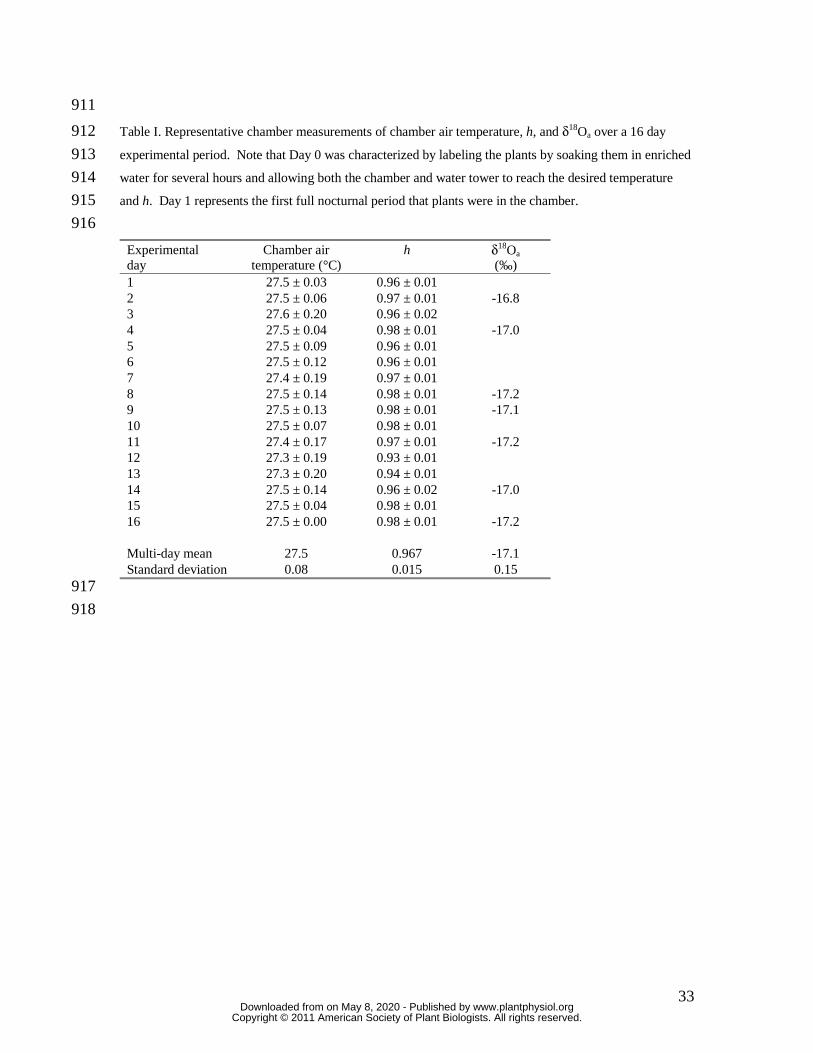
Representative chamber measurements of h, air temperature, and δ18Oa over a 16 day 178
experimental period are presented in Table I. Mean values of δ18Oa, h and the number of 179
days that T. usneoides strands were in the chamber for each experimental run are 180
presented in Table II. An image of the experimental setup and representative diurnal 181
cycles are presented in the Supplementary Material. Data from the 4 initial experimental 182
runs that were used to test the chamber setup were not included. Throughout this paper, h 183
= ea/ei and it is assumed that h equaled relative humidity in all chamber studies because 184
the chamber fans assured little boundary layer development around the T. usneoides 185
ramets. Support for this assumption is provided from several early runs of the chamber 186
setup where T. usneoides leaf temperatures were measured and they were not 187
significantly different from measures of air temperature (data not shown), so ei was equal 188
to esat at air temperature. The chamber held temperature and h relatively constant 189
(typically standard deviation for temperature = 0.08 °C and for h = 0.015; Table 1). 190
Maintaining constant h above 0.9 for long periods of time is difficult and, to my 191
knowledge, has not been attempted in many plant gas exchange studies. Condensation in 192
tubing and within the chamber, and directly on any chamber temperature control device 193
like the peltier unit, can systematically and dynamically alter h within the chamber. There 194
were occasional, unexplained changes in h as seen on days 12 and 13 (Table I). These 195
changes were manifest as a systematic drop in h that was caused by a slight drop in ea and 196
not changes in temperature (thus, not a change in ei). Fortunately such variability in h did 197
not appear to compromise the long-term stability of δ18Oa. The source for δ18Oa was the 198
23 L capacity temperature-controlled water tower, the volume and specific heat of which 199
lead to a low multi-day standard deviation for δ18Oa of ± 0.15 ‰. 200
201
To quantify the amount of water adsorbed to trichomes (Figure 1) and to determine how 202
the δ18O of this trichome water should affect observed δ18OL (Figure 2) the gain in mass 203
from water adsorption to T. usneoides trichomes was determined at 23 separate settings 204
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

10
of h from 0.34 to unity. The data in Figure 1 represent the water adsorbed to trichomes 205
expressed as RWC, therefore this value is comparable to RWC values elsewhere in this 206
study. Below an h of about 0.8, the water absorbed to trichomes was less than 8 % RWC, 207
however, above 0.8 the trichome water increased to more than 40% at h =1. While the 208
latter value may seem high, these results are consistent with water hydration studies of 209
non-lipid plant cuticle components (Dominguez and Heredia, 1999). These data were 210
used to parameterize the Hailwood and Horrobin model which was then used to predict 211
the ‘trichome offset’ for a variety of T. usneoides relative water contents (RWC) and h 212
(Figure 2). For the model values of Figure 2, RWC includes both tissue water and water 213
adsorbed to trichomes. The expected isotopic offset can be as high as 2 ‰ if RWC is low 214
and the h during sampling is high, however, for most of the sampling scenarios 215
encountered in this study, the trichome offset was between 0.5 and 1 ‰, which is still 216
well above the measurement precision of 0.2 ‰. These results show that the trichome 217
offset must be considered for this study because sampling occurred during nocturnal 218
periods for the plants, when h was highest. All steady and non-steady-state predicted 219
values of δ18OL were corrected for trichome water adsorption based on these observations 220
(see Materials and Methods). In field sampling conditions, trichome offsets should be less 221
of a worry because of typically low h during the day. For example, at an h of 0.65 and 222
RWC of 100 %, the trichome offset is near the sampling precision for δ18O (~ 0.2 ‰). If 223
the RWC is decreased to 50 % (thereby increasing the proportional representation of 224
trichome water) the offset increases to 0.36 ‰. 225
226
To examine how δ18OL changed as a consequence of rehydration, T. usneoides strands 227
that were not watered for one-week were placed in enriched water and removed at set 228
times from 10 minutes to 24 hours (Figure 3). A mass balance using the δ18O of wetting 229
water (11.5 ‰), the initial δ18OL (-3.3 ± 0.31 ‰) and the final δ18OL (3.7 ± 0.57 ‰) 230
showed that approximately 47% of the final water volume was the wetting water, or that 231
plant water volume nearly doubled due to the rewetting. After 5 hours of submersion in 232
water, the δ18OL was statistically indistinguishable from δ18OL after 24 hours submersed 233
(Figure 3). These results are similar to previous work showing that T. usneoides attains 234
saturated water content in approximately 4 hours. After only 20 minutes submerged, 235
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

11
δ18OL became significantly enriched by 2.5 ‰ and approximately 70 % of the “new” 236
water was absorbed. Therefore, substantial changes in δ18OL can occur over relatively 237
short time spans due to water absorption. 238
239
Figure 4 shows total plant conductance (gtot; mmol m-2 s-1; stomatal and boundary layer 240
conductance) for water vapor from T. usneoides versus RWC for average chamber h 241
ranging from 0.556 to 0.997. There was a highly significant (R2 = 0.78, p < 0.001) 242
correlation between RWC and gtot. Across treatments there was also a significant effect of 243
vapor pressure deficit (VPD) on gtot, but VPD explained only about 17 percent of the 244
variation in gtot (data not shown). 245
246
A general empirical sense of how the control of δ18OL by δ18Oa is regulated by h, and that 247
as h approaches unity δ18OL approaches equilibrium with δ18Oa can be seen in Figure 5. 248
Helliker and Griffiths (2007) showed that, under reasonably constant conditions, δ18OL 249
tended towards maximum enrichment (Equation 4 in Materials and Methods) after four 250
days. Figure 1 shows the difference between δ18OL and δ18Oa (δ18OL – δ18Oa) plotted 251
against h for every δ18OL sample that had been in the chamber for at least 4 days (n = 252
169). The data showed a tight relationship (y = -44.9x + 52.8, p < 0.0001, R2 = 0.87) 253
where h explained 87 percent of observed δ18OL. However, when the range of h was 254
restricted to 0.8 and above— conditions more in line with natural field conditions— h 255
explained the observed data scatter less well (y = - 59x + 66.0, p < 0.0001, R2 = 0.67). 256
257
Figures 6 and 7 show day to day observations and predictions of δ18OL for representative 258
chamber studies at three high (0.96 and 0.92 in Figure 6, and 0.97 in Figure 7A), one mid 259
(0.84; Figure 7B) and one low (0.56; Figure 7C) values of mean h. The leaf water models 260
(Equations 1, 4 and 6) were parameterized with observed nocturnal h, temperature, δ18Oa, 261
and gtot data for each day and the solutions were corrected for water adsorbed to 262
trichomes (δ18Otrichome) using observed RWC, and the empirically derived offset between 263
δ18Oa and δ18Otrichome (see Materials and Methods). The models were initialized with 264
observed Day 0 data for δ18OL, so that both observed and predicted values were equal at 265
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

12
the start. For the Hartard-Cuntz solution (Equation 6) I used the following schemes: 266
observed gtot and changing W, observed gtot and constant W (9 mol/m-2) and constant gtot 267
(0.002 mmol m-2 s-1) and W. Both Figures show the march of non-steady-state predictions 268
and observed δ18OL towards maximal enrichment (RL-M, Equation 4). Figure 6 further 269
shows that the non-steady-state predictions from the Hartard-Cuntz solution match 270
observations well, whether or not W, or gtot and W, were kept constant. Where h was 271
above 0.9, maximal enrichment was achieved in 4 to 9 days. Within similar levels of h 272
(Figure 6A vs. Figure 7A), the length of time to reach δ18OL-M was determined by the 273
isotopic distance between δ18OL-M and δ18OL at Day 0. Finally, δ18OE (RE; Figure 6 and 274
Figure 7A) slowly approached and occasionally reached the value of δ18OL-M as would be 275
expected at isotopic steady state. In contrast, observed δ18OL quickly matched δ18OL-M 276
and generally followed δ18OL-M even with slight changes in h (e.g. Days 16-18 in Figure 277
6A). 278
279
For all experimental runs, predicted δ18OL using the full non-steady-state leaf-water 280
model, Equation (1), agreed very well with measured δ18OL (Figure 8A). Every sampling 281
period of the chamber studies with the exception of Days 0 are presented, therefore 282
Figure 8 includes all of the within-experiment dynamism shown in Figures 6 and 7, and 283
the broad range of h and δ18Oa shown in Table II. The gray line represents a perfect 284
correlation between predicted and observed values. The solid line represents the 285
relationship between predicted and observed, the slope of which was near 1 with an 286
intercept near zero (y = 0.93*x - 0.07, R2 = 0.975, p < 0.0001). δ18OL-M predictions from 287
the maximum enrichment model, Equation (5), also agreed well with all observed δ18OL 288
that were sampled at h > 0.8 and after four days in the chamber (Figure 8B). That data in 289
Figure 8B include all of the data in Figure 1 in which h > 0.8. 290
291
Rehydration of plants during experimental runs did cause a significant change in δ18OL, 292
but the change was short-term. In 4 out of 12 of the high h experimental runs (h > 0.9), 293
two populations of T. usneoides were maintained within the chamber, one wetted and one 294
non-wetted. The frequency of the wetting varied from 2 to 7 days, and the rehydration 295
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

13
water was enriched to artificially enhance the difference between the wetted and non-296
wetted δ18OL. Representative data from these comparisons are presented in Table III. As 297
in all experiments, all plants were fully hydrated on Day 0. From Day 1 to Day 6 all 298
plants received no rehydration water. On Day 7, δ18OL samples were taken and half of the 299
plants were submerged in the wetting water for an hour. When all plants were sampled 300
for δ18OL on day 10, there was no significant difference between the wetted and non-301
wetted plants. The wetted subset of plants were again rehydrated on Day 10 (and again on 302
Day 14) and in each subsequent sampling period there were no significant differences 303
between the wetted and non-wetted plants. δ18OL for the wetted plants was sampled 304
shortly after rehydration on Day 10 to show that the wetting water did in fact cause a 305
significant enrichment of δ18OL, yet this difference was not apparent on Day 14. With one 306
exception, the results in Table III are comparable to all the wetted versus non-wetted 307
comparisons. In only one part of one trial was there a significant difference between 308
wetted and non-wetted strands. In this trial, the wetted plants were rehydrated every two 309
days and the non-wetted plants were very dry (RWC below 80 %) and exchanging 310
(losing) very little water with chamber water vapor (gtot was very low). 311
312
Discussion 313
It is a long-term goal to use the δ18O of epiphytic CAM plants— both the δ18O of leaf 314
water (δ18OL) and organic material that is labeled by δ18OL— to illuminate plant 315
physiological responses to the unique epiphytic growth habit (Reyes-Garcia et al., 2008) 316
and to reconstruct the δ18O of atmospheric water (δ18Oa; Helliker and Griffiths 2007); the 317
latter goal being much more broadly important to the use of stable isotopes in plant 318
physiological and ecophysiological research. The largely abiotic factors h and δ18Oa are 319
the primary controls of δ18OL (Figure 1) and their control under both constant and 320
dynamical conditions and must be considered. In addition, epiphyte morphology and 321
physiology act as important secondary controls on δ18OL and not accounting for both 322
could potentially obscure any δ18O interpretation. Plant physiology affects δ18OL 323
primarily through total plant conductance (gtot; stomatal and boundary layer conductance) 324
which controls the rate of exchange of tissue water with water vapor, and the relative 325
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

14
water content (RWC) of the plant. Plant morphology should affect epiphytic δ18OL 326
through water trapped in tanks formed at leaf bases (although this is not a concern for T. 327
usneoides), and through water adsorbed to hygroscopic trichomes. Therefore, the goal of 328
this study was to use controlled experiments on the CAM epiphyte T. usneoides to assess 329
how well observed δ18OL could be explained through controlled variation in environment 330
and physiology. 331
332
Water adsorbed to trichomes was shown to have a substantial potential impact (Figures 2 333
and 3) on observed and predicted δ18OL, but the actual impact and the need for 334
accounting in field and lab situations needs clarification. For all predictions shown in 335
Figure 7, the relationship between predicted δ18OL using the trichome offset and 336
predictions not using the offset was y = 0.99x - 0.81 (R2 = 0.997), so the non-corrected 337
predictions of δ18OL were about 0.8 ‰ more enriched than the predictions using the 338
trichome offset. This overall difference of about 1 ‰ is clearly a substantial offset. 339
However, the experimental setup exacerbated this offset because we sampled δ18OL 340
during the nocturnal period when h was highest. This setup was chosen so that δ18Oa 341
could be sampled during the period when most vapor exchange was occurring. Under 342
field conditions, δ18OL would likely be sampled during the low humidity day then the 343
trichome offset would be less of a problem, and perhaps not a problem at all. The 344
exception to this would be sampling leaves for δ18OL near dawn or dusk when h would 345
likely be high. It should be pointed out that while most of the adsorbed water was likely 346
onto the trichomes, adsorption to the cuticle could not be ruled out. This is somewhat 347
inconsequential as discussed below. Also, the apparent isotopic offset between the 348
trichome water and δ18Oa was 7.4 ± 0.1 ‰ while the equilibrium fractionation factor for 349
condensation was around 9.2 ‰. It is unclear at this time how similar adsorption of 350
water to leaf cuticles and trichomes of more common plant types (i.e. non-epiphytic) 351
could bias observations of samples taken near dawn or dusk. Similarly, this is an unlikely 352
problem in field studies with the exception of diurnal sampling schemes. In the latter 353
case, water adsorption to leaf cuticles/trichomes could lead to lower than expected 354
enrichment during high humidity morning and late afternoon sampling. 355
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

15
356
When h is below 1, The direct adsorption of atmospheric water vapor by the dead 357
trichomes of T. usneoides does not lead to any apparent absorption by the living cells of 358
the plants (Martin and Schmitt, 1989). Several studies have reported direct uptake of 359
water by atmospheric epiphyte leaves, but this is under conditions of saturating vapor 360
and/or leaf wetness (Benzing et al., 1976; Martin and Schmitt, 1989; Andrade, 2003). The 361
trichomes of T. usneoides are highly hygroscopic and can, within gas exchange 362
chambers, lead to what appears as a ‘negative’ transpiration while equilibrating to an 363
increase in h. This negative transpiration is the process of water adsorbing onto the 364
trichomes and it stops once equilibrium is reached with h. This process is also what was 365
quantified in isolation here in with dead T. usneoides (Figure 2). The dead trichomes of 366
T. usneoides likely equilibrate with the water potential of ambient air much like what is 367
seen in the lichen thallus (Lange et al., 2001; Hartard et al., 2009) and isotopically the 368
adsorption of water onto the trichomes may be very similar to that seen by the thallus of a 369
lichen. This is, however, where the similarities end. Lichens are poikilohydric and some 370
algal-symbiotic species can become physiologically active solely through this water 371
adsorption (yet only at relatively high h); thus, the adsorption of water vapor by the 372
thallus matrix can lead to a cellular absorption (Lange et al., 2001). Such an absorption of 373
atmospheric water vapor does not occur in homoiohydric T. usneoides and is unlikely to 374
occur in other vascular plant atmospheric-type epiphytes (e.g. Bromeliads, Orchids, 375
Lycophytes, and Ferns; (Martin et al., 2004)). 376
377
T. usneoides, like most epiphytic higher plants that have been observed, maintains very 378
high (near zero) plant water potentials with osmotic potentials maintained consistently in 379
the realm of aquatic plants (Martin et al., 2004). In general, the measured water potential 380
of atmospheric epiphytes, T. usneoides included, has rarely been observed below -1 MPa 381
(Martin, 1994). In one experiment with T. ionantha, total plant water potential did not 382
drop below -0.75 MPa after 60 days of imposed drought (Nowak and Martin, 1997). 383
Considering that the water potential of air at h = 0.99 is -1.4 MPa (and -2.8 MPa at h = 384
0.98, both at 20 °C), it is hard to imagine a scenario in which water adsorbed onto 385
trichomes that are in equilibrium with subsaturated air could move against the water 386
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

16
potential gradient into the living cells of T. usneoides, and the work of Martin and 387
Schmitt (1989) corroborates this. Therefore, there should be no consideration of water 388
vapor uptake by trichomes affecting δ18OL. Once saturation is realized and the plant 389
tissue is covered with liquid water, the trichomes become appressed against the cuticle 390
and channel the absorption of liquid water into the underlying living cells (Benzing et al., 391
1976). This sort of precipitation event is explicitly considered through Equation 1. 392
Furthermore, the maintenance of extremely high water and largely invariant water 393
potential in the face of drought is likely the driving force behind the austere physiology 394
of epiphytes (Martin et al., 2004) in terms of both carbon assimilation and water loss. For 395
T. usneoides, well-watered measurements of CO2 uptake as low as 0.2 μmol m-2 s-1 396
(Martin & Schmitt, 1989) and transpiration of 0.002 mmol m-2 s-1 are common (this study 397
and (Martin, 1994)). The tight coupling of plant water potential and gtot likely led to the 398
rapid decrease in gtot as RWC decreased (Figure 5) and could explain why gtot was more 399
highly correlated with RWC than with VPD. The inherently low levels of water loss 400
meant that estimates of gtot could be determined once per day or even once every other 401
day, thus maintaining constant chamber conditions for longer periods of time (see 402
Supplementary Material). 403
404
Transpiration forced δ18OL towards the value of maximum enrichment, δ18OL-M, and the 405
closer the value of h to unity, the faster was the approach to δ18OL-M. This overall 406
tendency can be seen in Figures 6 and 7 where all values of δ18OL moved towards the 407
predicted δ18OL-M. Yet only when values of h were greater than 0.9 (Figures 6 and 7A), 408
did observed δ18OL actually reach δ18OL-M. At higher h, the gross flux of water into the 409
leaf (gtot • wa) was proportionally much higher than at low h and therefore the 410
replacement of leaf water with atmospheric water was faster. At lower h, a considerably 411
greater volume of leaf water must be lost before δ18OL becomes equal to δ18OL-M 412
(Helliker and Griffiths, 2007) and the response of gtot to RWC tends stop any great 413
reduction in leaf water volume (Figure 5). A quick comparison of Figures 6A and 7A 414
shows two experimental runs at similar h that took drastically different amounts of time 415
for δ18OL to equal δ18OL-M. For Figure 7A, the apparent leaf-water turnover time was 416
about 4 days, similar to the results of Helliker and Griffiths (2007), whereas in Figure 6A 417
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

17
the apparent leaf-water turnover time was 9 days. The reason for this discrepancy was 418
the different starting points of δ18OL. In Figure 6A, the initial δ18OL was approximately 419
15 ‰ more enriched than δ18OL-M and for Figure 7A the starting δ18OL was 420
approximately 7 ‰ more enriched than δ18OL-M. 421
422
In general, as water is lost from CAM epiphytes δ18OL is more and more likely to be 423
described by the maximum enrichment model, δ18OL-M. Yet it should be recognized that 424
as the period between rehydration events increases, RWC decreases and so does gtot, 425
which leads to an inevitable decrease in the water vapor exchange between the leaf and 426
the atmosphere. So, as the plant dries, δ18Oa still controls δ18OL, but the turnover of leaf 427
water and consequently any affect that changes in ambient δ18Oa has on δ18OL are 428
minimized. This can be seen distinctly in Figure 6C where even after several days at low 429
humidity, the enrichment of 18O in leaf water ceased. As T. usneoides dries out, the 430
overall tissue water exchange with atmospheric water vapor slows until δ18OL becomes 431
decoupled from δ18Oa. 432
433
The largest potential hurdle in using δ18OL-M to describe observed δ18OL appears to be the 434
effect that the isotope ratio of rehydration water (simulated precipitation events) has on 435
δ18OL and there are two components of this: the absolute difference in isotopic space 436
between the rehydration water and δ18OL-M and the total amount of rehydration water 437
absorbed by the plant. As a general response, water gain through precipitation has the 438
initial tendency to move δ18OL farther away from δ18OL-M, but then— due to increased 439
gtot—rapidly trends toward δ18OL-M. The more specific response of T. usneoides δ18OL to 440
precipitation events was examined three ways: The first was through the rehydration 441
study (Figure 4) which showed that the absorption of precipitation can lead to a rapid and 442
significant change in δ18OL after only 20 minutes in liquid water and after five hours the 443
change in δ18OL was complete. The second was starting all experimental runs by soaking 444
plants in an enriched water source, thereby ensuring that each experimental run began 445
after a simulated precipitation event. This method showed that, at high h, the 446
precipitation isotope label could exchange quickly with atmospheric water vapor and the 447
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

18
speed of exchange depended on the size of disparity between the initial label leaf water 448
and δ18OL-M. The final assessment of how precipitation inputs affect T. usneoides δ18OL 449
was designed to examine the differential effect of rehydrating one subset of strands while 450
keeping the other dry during the same experimental runs (Table III). This approach 451
explored a potential situation in the field where rainfall or dew formation could vary 452
spatially either across a field site or within a single clump of T. usneoides depending on 453
the intensity of a precipitation event. This method was performed in several of the high h 454
experimental runs, the frequency of the wetting varied from 2 to 7 days, and the 455
rehydration water was enriched. In general, there was no sustained, significant difference 456
between the wetted and non-wetted plants. Even though significant enrichment occurred 457
in as little as 20 minutes (Figure 4), it appears that this water was quickly turned over 458
through nocturnal exchange of water vapor. This rapid turnover is likely due to the fact 459
that a rehydration allows for an uptick in gtot which then allows for increased gross 460
exchange of vapor between the plant and the atmosphere. The end result being that both 461
wetted and non-wetted strands tracked δ18Oa, although it seems likely that strands with 462
higher RWC would respond to changes in δ18Oa more quickly than drier strands, as 463
mentioned above. It appears that a relatively consistent input of dew or precipitation 464
water could lead to a tighter coupling of δ18OL and δ18Oa because of enhanced vapor 465
exchange. Additionally, as discussed below, I hypothesize that under field conditions 466
precipitation itself should lead to a greater coupling between δ18OL-M. 467
468
The use of enriched rehydration water allowed for easier observation of how precipitation 469
affected δ18OL, but also led to a potential misinterpretation of the coupling between δ18OL 470
and δ18Oa. Both the rapid change in δ18OL during rehydration (Figure 4) and the multi-471
day turnover of δ18OL could lead one to conclude that a rehydration event pulls δ18OL too 472
far away from δ18OL-M to use δ18OL as a decent proxy for δ18Oa. However the δ18O of the 473
wetting water was typically 15 ‰ more enriched in 18O than the δ18Oa-equilibrated of 474
δ18OL and 25 ‰ to 30 ‰ more enriched than δ18Oa. While this isotopic span allowed for 475
a clearer picture of rehydration effects and tissue-water turnover times, it is highly 476
unlikely that such a span would occur in the field. Precipitation (dewfall or rainfall) 477
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

19
should also be near equilibrium with δ18Oa (Gat 1996). Therefore, isotopic differences 478
between rain and/or dew δ18O and δ18OL (and δ18OL-M) on the scale of 15 ‰ would not be 479
expected— not even considering large-scale episodic events that can dramatically alter 480
δ18O of precipitation (Miller et al., 2006)— and precipitation should enhance the 481
coupling of δ18OL and δ18Oa. 482
483
When variation in δ18Oa, h, gtot, RWC and trichome water adsorption were considered 484
through the leaf water models (Equations 1, 4 and 6) and through a wide-range of 485
conditions (Table II), observed δ18OL was explained very well (Figures 6, 7 and 8). Both 486
variations of the non-steady state model (Equations 1 and 6) predicted observed δ18OL 487
well and this included runs when gtot and W were held constant (Figure 6). Previous work 488
on non-epiphytic plants has also shown measurements of dW
dt are not crucial for accurate 489
predictions of non-steady-state models (Cernusak et al., 2002; Farquhar and Cernusak, 490
2005; Cuntz et al., 2007; Ogee et al., 2007; Kahmen et al., 2008). The modeled estimates 491
of δ18OE (RE; Figure 6 and Figure 7A) approached and only occasionally matched the 492
value of δ18OL-M as would be expected at isotopic steady state. It can be concluded then 493
that δ18OL was rarely at true isotopic steady state, however observed δ18OL often matched 494
δ18OL-M at values of h > 0.9 and generally followed δ18OL-M even with slight changes in h. 495
This apparent contrast leads to one of the more important conclusions of this study: that, 496
in agreement with the predictions of Helliker and Griffiths (2007), leaf water does not 497
need to be at full isotopic steady state for observed δ18OL to equal δ18OL-M. 498
499
The extreme sensitivity of δ18OL to h > 0.9 means that Equation 7— the full equilibrium 500
equation— is rarely useful. This may seem contradictory to the some of the data 501
presentation of Helliker and Griffiths (2007) which showed a prediction range for 502
Equation 5 on some figures. The experimental setup of Helliker and Griffiths had an 503
invariably high h and noisy δ18Oa (± 1.2 ‰) and we used the range of predictions 504
stemming from the variability in δ18Oa to show that observations were near these values. 505
Unfortunately, this approach could lead one to erroneously conclude that δ18OL was in 506
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

20
direct equilibrium with δ18Oa even when h was less than unity, which is not the case. The 507
value of h should be known to decide which equation to use when extracting δ18OL from 508
measures of δ18Oa using CAM epiphytes or non-epiphytic plants as in the pre-dawn 509
sampling approach developed by Lai et al. (2008). While predictions of δ18OL-M 510
(Equation 4) matched observed δ18OL at h> 0.9, the predictions from this model showed 511
good agreement with observed δ18OL over the broader range of h > 0.8 (y = 0.87x - 0.99, 512
R2 = 0.92; Figure 8B) after 4-5 days. The primary need to exclude data under four days 513
results from the artifact of using an enriched water source for rehydration, as mentioned 514
above. A more complete test of Equation 4, the maximum enrichment model δ18OL-M, has 515
been performed under field conditions and is the subject of subsequent publications. 516
517
518
Conclusions 519
Helliker and Griffiths (2007) proposed that the δ18OL of CAM atmospheric epiphytes 520
could be used to reconstruct δ18Oa. An inherent assumption of this proposal was that the 521
environmental controls on δ18OL— aside from δ18Oa — are known and can be accounted 522
for in the framework of a mechanistic model. The solid agreement between observations 523
of δ18OL and predictions from Equations 1, 4 and 6 appears to satisfy this assumption and 524
shows a fair understanding of the controls on δ18OL in T. usneoides, at least under 525
laboratory conditions. Hygroscopic trichomes can affect interpretations of observed 526
δ18OL but only when leaf water is sample under high h; a condition that is not often 527
satisfied in the field. Observed δ18OL could be predicted well through the non-steady 528
state equations even by using constant values of W and gtot. Therefore, the non-steady-529
state models can potentially be applied more easily. Also, the leaf does not need to be at 530
full isotopic steady state for observed δ18OL to equal δ18OL-M (Equation 4) so only δ18Oa 531
and relative humidity (h) are required as predictive inputs. The poor performance of 532
Equation 4 in the first 4-5 days of experimentation was likely an artifact of using 533
artificially enriched rehydration water to start each experiment. Tissue rehydration using 534
enriched water caused a significant, but transitory change in δ18OL because the 535
consequent increase in gtot ultimately led to a faster coupling with δ18Oa. The inherent 536
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

21
coupling between precipitation water δ18O and δ18Oa in the field should mean that δ18OL 537
is more tightly coupled to δ18Oa after rehydration. This enhanced understanding about the 538
controls of δ18OL in CAM epiphytes under controlled laboratory conditions will improve 539
the interpretation of δ18OL observations in the more varied natural environment. 540
541
542
Materials and Methods 543
Modeling leaf water isotopes 544
The isotope ratio of leaf water (δ18OL) for a CAM epiphyte such as T. usneoides can be 545
explained by the following non-steady state equation (Flanagan et al., 1991; Farquhar and 546
Lloyd, 1993; Farquhar and Cernusak, 2005; see Helliker and Griffiths 2007 for a full 547
derivation): 548
549
RL = α * RpJp − d(WRL )
dt
⎛ ⎝ ⎜
⎞ ⎠ ⎟
α K
gtotwi
+ Ra
wa
wi
⎡
⎣ ⎢
⎤
⎦ ⎥ (1). 550
551
where R is the molar isotope ratio the subscripts L and P represent leaf and precipitation 552
(rainfall or dewfall) water, respectively. JP is the flux of water into the plant, W is leaf 553
water volume (mol m-2), gtot is total conductance (stomatal and boundary layer; mol m-2 s-554 1) and wi and wa represent the mole fraction of water vapor in the substomatal cavity and 555
ambient air, respectively. the mole fraction of water vapor, w, is equal to the partial vapor 556
pressure e divided by total atmospheric pressure. The temperature dependent equilibrium 557
fractionation factor is represented by α*, and αK is the balance of the ratio of diffusivities 558
of light to heavy water molecules through the stomata and through the leaf boundary 559
layer (Merlivat, 1978; Luz et al., 2009). Between rewetting events, when RPJP is zero, RL 560
can be explained by: 561
562
RL = α * − d(WRL )
dt
⎛ ⎝ ⎜
⎞ ⎠ ⎟
α K
gwi
+ Ra
wa
wi
⎡
⎣ ⎢
⎤
⎦ ⎥ (2) 563
564
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

22
In between rain events d(WRL )
dt= −RE E , where E is the transpiration flux from the leaves 565
(E = g (wi –wa)). If we define h = wa/wi, then equation 2 can be rewritten as: 566
567
RL = α * α K RE (1− h) + Rah[ ] (3) 568
569
Throughout this study, h = ambient relative humidity, so in Equation (3) the effect of h 570
on RL can be seen easily where, as h increases towards unity, Ra assumes a greater control 571
over RL. Under constant conditions and water loss, steady state is achieved and RE in 572
Equation 3 can be replaced by RL arriving at maximum enrichment, RL-M: 573
574
RL−M ≈ Rah1
α *− α K + α K h
(4). 575
Equation 4 is similar to the derivation of maximum enrichment of Farqhar and Gan 576
(2003) and Hartard et al. (2009): 577
578
RL−M ≡ α * hRa
1−α Kα * 1− h( ) (5). 579
Hartard et al. (2009) further developed an analytical solution to the non-steady state 580
equation, referred to throughout as the Hartard & Cuntz solution: 581
RL = RL−M + R0 − RL−M[ ]exp− 1
α K α*(1−h )−1
⎡
⎣ ⎢
⎤
⎦ ⎥
E
Wt
⎧ ⎨ ⎩
⎫ ⎬ ⎭ (6) 582
583
Lastly, when h = 1, RL is equal to the isotope composition of atmospheric water vapor, 584
corrected for the equilibrium fractionation factor, 585
586
RL = Raα * (7). 587
588
In this paper, the majority of model solutions were obtained through iterative solution of 589
Equation (2). The Hartard & Cuntz solution was compared to outputs of equation (2) 590
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

23
using changing and constant values of gtot and W. Equation (5) was examined as a 591
simplified method of predicting RL for eventual field applications. 592
593
594
Plant Material 595
The study species for this work was the CAM epiphytic Bromeliad Tillandsia usneoides. 596
T. usneoides is a subtropical-to-tropical plant that ranges from coastal Virginia, USA 597
through the tropics to Argentina. The broad distribution is determined by high nocturnal 598
relative humidity (Garth, 1964) and makes it a model organism for the reconstructing 599
δ18Oa. In October 2006 and 2008, plants were obtained from the northernmost point of 600
distribution for T. usneoides in North America, First Landing State Park in Virginia 601
Beach, VA, USA. The plants were subsequently maintained in a high-humidity 602
greenhouse at the University of Pennsylvania until experimentation. Approximately 5 603
days prior to the start of an experiment, clumps of T. usneoides were separated into 604
strands of 10 to 20 ramets (ramets consist of 2 to 5 leaves that were 3 –6 cm in length) 605
each and placed in the growth chamber under a reversed 12h photoperiod (i.e. forcing the 606
plants nocturnal cycle to coincide with the researchers normal diurnal cycle). Prior to this 607
acclimation period all strands were submersed in enriched water (ca. 11 ‰) for 2-4 hours 608
and plants were watered daily thereafter until the start of an experiment. 609
610
Experimental Conditions 611
For experiments, plants were maintained in a climate-controlled chamber coupled to a 612
large volume dew-point generator (the ‘water tower’) that was capable of maintaining a 613
constant δ18Oa for up to 3 weeks (s.d. ± 0.15 ‰). The chamber was fabricated from a 614
cubic 22.2 L acrylic desiccator cabinet (Fisher Scientific) which was back-fitted with 615
tubing inlets and a 75 W, 24 VDC Peltier type thermoelectric air conditioner (INB260-616
24-AA, Watronix Inc., CA, USA). A temperature controller (model CN77322-C2; 617
Omega Engineering, Stamford, CT) signaled a relay to change the DC polarity to either 618
heat or cool the chamber based on the prescribed set point; separate day/night 619
temperature setpoints were controlled by a Grässlin Digi-20E timer (Intermatic, IL, 620
USA). 20 cm long strands of T. usneoides were placed horizontally in the chamber on top 621
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

24
of a wire mesh shelf. A 400 W metal-halide light source (Sun System 3, Sunlight Supply 622
Inc., Vancouver, WA, USA ) was placed above the chamber; photosynthetically active 623
radiation was 500 μmol m-2 s-1 at plant height. 624
625
The water tower consisted of two 132 cm long, 0.64 cm wall thickness, extruded acrylic 626
cylinders— one inside of the other (outer cylinder ID = 20.3 cm, inner cylinder ID = 15.2 627
cm) — which were sealed off with 1 cm thick Aluminum plates at either end. The effect 628
was to create an outer cavity for a temperature-controlled water jacket that surrounded 629
the inner cylinder. The inner cylinder was filled with 23 L of water which was the source 630
for water vapor in the chamber. The water jacket temperature was controlled by pumping 631
water from a circulating water bath (water circulated between the outer and inner 632
cylinders). Ambient air was pumped (KNF Neuberger Inc., Freiberg, Germany) through 633
the length of the inner cylinder at 2 L min-1 (MKS flow controller, Andover, MA, USA) 634
to create δ18Oa and dewpoint values that were fed to the plant chamber. The temperature 635
of the water tower controlled the ambient vapor pressure (ea) values for the climate 636
controlled chamber and the temperature of the chamber set the saturated vapor pressure 637
(esat) values. Because of the high heat capacity of the water tower and the tight 638
temperature control of the chamber, a fairly constant ea, and therefore h (ea/esat), could be 639
maintained (see supplementary data for picture of chamber/water tower setup and a 640
representative diurnal cycles of h and air temperature). 641
642
Five days prior to each experiment, 30 to 50, 30 cm long strands of T. usneoides were 643
removed from the greenhouse and placed in a growth chamber set on reversed, 12h 644
photoperiod to acclimate the plants. The reversed photoperiod allowed for easy sampling 645
of δ18Oa during the period of T. usneoides water loss (and isotopic exchange). Each 646
experimental run was started (Day 0) by submerging the strands in a tub of enriched 647
water (typically 10 ‰ to 20 ‰) for two hours. This submersion assured hydration of the 648
strands and assured that the initial δ18O of leaf water was considerably different from any 649
chamber-induced δ18OL. The strands were surface-dried and then placed in the 650
experimental chamber. 4 of the strands were sealed in glass vials immediately to sample 651
for initial δ18OL and 4 of the strands were tagged for measurements of relative water 652
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

25
content and water loss (through loss of mass) during every sampling period. Typically 653
every third day, 3 to 5 strands of T. usneoides were removed from the chamber and 654
placed into glass vials for eventual water extraction and determination of δ18O. 655
656
Isotope sampling and analysis 657
Water vapor was sampled from the chamber exhaust tubing for at 2.5 to 3 hours prior to 658
opening the chamber to sample leaf water in a 200 ml internal volume condenser inserted 659
into a dry ice/ethanol slush (-69 °C). The air exiting the chambers was split and 660
monitored with a rotometer to assure that flow rates of air through the condenser never 661
exceeded 0.5 L m-1 (Helliker et al., 2002). Leaf water was extracted cryogenically 662
(Ehleringer et al., 2000). Water samples (0.5 mL) were equilibrated for at least 48 hours 663
in 3 ml Exetainer® vials (Labco Limited, UK) with 10/90 mixture of CO2/He. 664
Approximately 100 μL of the headspace gas were injected into a GC and carried in a 665
helium air stream to a Delta Plus (Thermo-Finnigan, Germany) isotope ratio mass 666
spectrometer. All molar isotope ratios were expressed in the standard ‘delta’ notation on a 667
per mil basis by δ = (Rsample/Rstandard) * 1000, where R = 18O/16O and the standard was 668
Standard Mean Ocean Water (V-SMOW R = 0.002005). 669
670
Estimating relative water content and conductance to water loss 671
After each sampling period for δ18OL, three to four strands were placed in plastic bags to 672
assure no immediate loss of water from the trichomes and were removed from the 673
chamber. One by one the strands were taken out of the bags and the mass measured. 674
Relative water content was determined in the same manner as Martin and Schmitt (1989). 675
676
RWC = [(FW – DW)/DW] * 100 (8) 677
678
Where FW is the measured fresh weight and DW is the measured dry weight after 3 days 679
in a 60 °C drying oven. Plant conductance to water loss (gtot; mmol m-2 s-1) was 680
determined by: 681
682
gtot = E/(wi – wa) (9) 683
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

26
684
Plant transpiration (E) was estimated from the measures of fresh weight above, wi and wa 685
were obtained from measurements of chamber h and temperature, pressure was assumed 686
constant at 100 kPa. Leaf area was determined using the fresh weight to leaf area 687
conversions presented in Martin and Schmitt (1989). Based on direct measurements, leaf 688
temperature and air temperature were not different, therefore wi was equal to wsat at 689
chamber temperature. 690
691
Correcting for rehydration and water adsorbed to trichomes 692
The effect that rehydration had on T. usneoides δ18OL was determined by fully 693
submerging 45, 30 cm long strands for several hours to rehydrate. The strands were then 694
placed in the greenhouse and not watered for one-week. After a week the strands were 695
placed in enriched water (11.5 ‰) and 5 strands were removed from the rehydrating 696
water at intervals of 10 min, 20 min, 40 min, 1 hour, 1.5 hours, 2.5 hours, 5 hours, 7 697
hours and 24 hours. 698
699
In prior work we assumed water was adsorbed to trichomes at the time of sampling and 700
that this water was inevitably extracted with tissue water when bulk leaf water was 701
cryogenically extracted. In our prior attempt to correct for the isotope ratio of this water 702
to obtain tissue water (δtissue) several untested assumptions were made (Helliker and 703
Griffiths 2007): (i) that water adsorption by the trichomes from the atmosphere did not 704
fractionate and (ii) the humidity-driven proportion of adsorbed water on trichomes 705
(ƒtrichome) could be explained by a simple parameterization based on two measured 706
humidities (Martin and Schmitt 1989). The first assumption was examined by placing 707
several clumps of dead T. usneoides (about 2g of material, killed via desiccation at 60 ° 708
C) into the growth chamber for two days and then extracting the water adsorbed onto the 709
dead tissue for isotope analysis. This yielded an apparent fractionation factor (εtrichome) 710
between atmospheric water vapor and the water adsorbed to trichomes of 7.4 ± 0.1 ‰. 711
712
δ18Otrichome =δ18Oa + εtrichome (10) 713
714
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

27
The second assumption was examined by building an empirical relationship between the 715
mass of dead T. usneoides and h ranging from 0.34 to saturation. This was accomplished 716
by placing dead T. usneoides of a variety of masses into nylon pouches (made from 717
pantyhose, L’eggs®, Hanes, Winston-Salem, NC) and comparing oven-dried mass to the 718
mass after placement in a growth chamber at a known humidity for 24 hours. Empty 719
nylon pouches were also put through this exercise to correct for any water adsorption 720
onto the nylon (the maximum for nylon adsorption was less than 6 % of dry nylon mass 721
at saturating humidity). The relationship between trichome water content (ƒtrichome) and h 722
was used to parameterize the Hailwood and Horrobin (Hailwood and Horrobin, 1946) 723
model for predicting water adsorption of solid polymer solutions as a function of h: 724
725
f t = k1
k2h
1+ k2h+ k3h
1− k3h
⎡
⎣ ⎢
⎤
⎦ ⎥ ×100 (11) 726
727
Where k1 is the number of binding sites, k2 is the affinity of the material for adsorption, k3 728
is the water activity. The values for k1, k2 and k3 were changed so that the adsorption 729
curve fit the data over a range of h. These values were k1 = 0.02, k2 = 10 and k3 = 0.95. 730
731
Finally, modeled values of δ18OL from Equations (3) (4) (5) and (6) were corrected for 732
trichome adsorbed water as a proportion of RWC by: 733
734
δ18Opredicted =δ18OEqns3,4,5,6 • 1− f t
RWC
⎛ ⎝ ⎜
⎞ ⎠ ⎟ +δ18Otrichome • f t
RWC
⎛ ⎝ ⎜
⎞ ⎠ ⎟ (12) 735
736
Acknowledgements 737
M. Suplick, F. Letterio and J. Andrews-Labinski offered a great deal of assistance in 738
constructing the chamber and water tower. M. Vergari, C. Hulshof, B. Waring, C. 739
Policastro and A. Dinerstein helped with water extractions. D. Vann was instrumental in 740
performing isotopic analyses. J. Lloyd-Helliker assisted in obtaining sample material. I 741
am grateful to First Landing State Park, Virginia State Parks and in particular Erik 742
Molleen for permission and assistance in obtaining sample material of T. usneoides. Two 743
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

28
anonymous reviewers gave extremely helpful comments to improve this manuscript. 744
745
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

29
References 746
747
Anderson W, Bernasconi S, McKenzie J (1998) Oxygen and carbon isotopic record of 748 climatic variability in tree ring cellulose (Picea abies): An example from central 749 Switzerland (1913-1995). Journal of Geophysical Research 103: 31,625-631,636 750
Andrade JL (2003) Dew deposition on epiphytic bromeliad leaves: an important event in 751 a Mexican tropical dry deciduous forest. Journal of Tropical Ecology 19: 479-488 752
Barbour MM, Farquhar GD (2000) Relative humidity- and ABA-induced variation in 753 carbon and oxygen isotope ratios of cotton leaves. Plant Cell and Environment 754 23: 473-485 755
Barbour MM, Fischer RA, Sayre KD, Farquhar GD (2000) Oxygen isotope ratio of 756 leaf and grain material correlates with stomatal conductance and grain yield in 757 irrigated wheat. Australian Journal of Plant Physiology 27: 625-637 758
Benzing DH (1970) Foliar Permeability and Absorption of Minerals and Organic 759 Nitrogen by Certain Tank Bromeliads. Botanical Gazette 131: 23-& 760
Benzing DH, Henderson K, Kessel B, Sulak J (1976) The absorptive capacities of 761 bromeliad trichomes. American Journal of Botany 63: 1009-1014 762
Cernusak LA, Farquhar GD, Wong SC, Stuart-Williams H (2004) Measurement and 763 interpretation of the oxygen isotope composition of carbon dioxide respired by 764 leaves in the dark. Plant Physiology in press 765
Cernusak LA, Pate JS, Farquhar GD (2002) Diurnal variation in the stable isotope 766 composition of water and dry matter in fruiting Lupinus angusifolius under field 767 conditions. Plant, Cell and Environment 25: 893-907 768
Ciais P, Tans PP, Denning AS, Francey RJ, Trolier M, Meijer HAJ, White JWC, 769 Berry JA, Randall DA, Collatz GJ, Sellers PJ, Monfray P, Heimann M 770 (1997) A three-dimensional synthesis study of δ18O in atmospheric CO2 2. 771 Simulations with the TM2 transport model. 102: 5873-5883 772
Cuntz M, Ciais P, Hoffmann G, Allison CE, Francey RJ, Knorr W, Tans PP, White 773 JWC, Levin I (2003) A comprehensive global three-dimensional model of ∂18O 774 in atmospheric CO2: 2. Mapping the atmospheric signal. Journal of Geophysical 775 Research 108: 4528 776
Cuntz M, Ogee J, Farquhar GD, Peylin P, Cernusak LA (2007) Modelling advection 777 and diffusion of water isotopologues in leaves. Plant, Cell & Environment 30: 778 829-909 779
Dominguez E, Heredia A (1999) Water hydration in cutinized cell walls: a physico-780 chemical analysis. Biochimica Et Biophysica Acta-General Subjects 1426: 168-781 176 782
Ehleringer JR, Roden JR, Dawson TE (2000) Assessing ecosystem-level water 783 relations through stable isotope ratio analyses. In OE Sala, RB Jackson, HA 784 Mooney, RW Howarth, eds, Methods in ecosystem science. Springer, New York, 785 pp 181-198 786
Epstein S, Thompson P, Yapp CJ (1977) Oxygen and hydrogen isotopic ratios in plant 787 cellulose. Science 198: 1209-1215 788
Farquhar GD, Cernusak LA (2005) On the isotopic composition of leaf water in the 789 non-steady state. Functional Plant Biology 32: 293-303 790
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

30
Farquhar GD, Gan KS (2003) On the progressive enrichment of the oxygen isotope 791 composition of water along a leaf. Plant Cell & Environment 26: 1579-1597 792
Farquhar GD, Lloyd J (1993) Carbon and oxygen isotope effects in the exchange of 793 carbon dioxide between terrestrial plants and the atmosphere. In JR Ehleringer, 794 AE Hall, GD Farquhar, eds, Stable Isotopes and Plant Carbon/Water Relations. 795 Academic Press, San Diego, pp 47-70 796
Ferrio JP, Voltas J (2005) Carbon and oxygen isotope ratios in wood constituents of 797 Pinus halepensis as indicators of precipitation, temperature and vapour pressure 798 deficit. Tellus B 57: 164-173 799
Flanagan LB, Comstock JP, Ehleringer JR (1991) Comparison of modeled and 800 observed environmental influences on the stable oxygen and hydrogen isotope 801 composition of leaf water in Phaseolus vulgaris L. Plant Physiol 96: 588-596 802
Garth RE (1964) Ecology of Spanish Moss ( Tillandsia Usneoides ) - Its Growth + 803 Distribution. Ecology 45: 470-& 804
Gat JR (1996) Oxygen and hydrogen isotopes in the hydrologic cycle. Annu Rev Earth 805 Planet Sci 24: 225-262 806
Griffiths H, Luttge U, Stimmel KH, Crook CE, Griffiths NM, Smith JAC (1986) 807 Comparative ecophysiology of CAM and C3 Bromeliads. 3. Environmental 808 influences on CO2 assimilation and transpiration. Plant Cell and Environment 9: 809 385-393 810
Griffiths H, Smith JAC (1983) Photosynthetic pathways in the Bromeliaceae of 811 Trinidad: relations between life-forms, habitat preference and the occurrence of 812 CAM. Oecologia 60: 176-184 813
Guy RD, Fogel ML, Berry JA (1993) Photosynthetic Fractionation of the Stable 814 Isotopes of Oxygen and Carbon. Plant Physiol 101: 37-47 815
Hailwood AJ, Horrobin S (1946) Absorption of water by polymers: analysis in terms of 816 a simple model. Transactions of the Faraday Society 42B: 84-101 817
Hartard B, Cuntz M, Maguas C, Lakatos M (2009) Water isotopes in desiccating 818 lichens. Planta 231: 179-193 819
Helliker BR, Berry J, Bakwin PS, Davis KJ, Denning AS, Ehleringer JR, Miller JB, 820 Butler MP, Ricciuto DM (2005) Regional-scale estimates of forest CO2 and 821 isotope flux based on monthly CO2 budgets of the atmospheric boundary layer. In 822 H Griffiths, PG Jarvis, eds, The Carbon Balance of Forest Biomes Taylor & 823 Francis, New York, pp 77-92 824
Helliker BR, Griffiths H (2007) Toward a plant-based proxy for the isotope ratio of 825 atmospheric water vapor. Global Change Biology 13: 723-733 826
Helliker BR, Noone D (2009) Novel approaches to spatial monitoring of water vapor 827 isotope ratios. . In GJ Bowen, JB West, TE Dawson, eds, Isocapes: Understanding 828 movement, pattern, and process on Earth through isotope mapping, 829
Helliker BR, Richter SL (2008) Subtropical to boreal convergence of tree-leaf 830 temperatures. Nature 454: 511-514 doi: 510.1038/nature07031 831
Hoffmann G, Cuntz M, Weber C, Ciais P, Friedlingstein P, Heimann M, Jouzel J, 832 Kaduk J, Maier-Reimer E, Seibt U, Six K (2004) A model of the earth's dole 833 effect. Global Biochemical Cycles 18 834
Kahmen A, Simonin K, Tu KP, Merchant A, Callister A, Siegwolf R, Dawson TE, 835 Arndt SK (2008) Effects of environmental parameters, leaf physiological 836
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

31
properties and leaf water relations on leaf water delta O-18 enrichment in 837 different Eucalyptus species. Plant Cell and Environment 31: 738-751 838
Lai CT, Ometto J, Berry JA, Martinelli LA, Domingues TF, Ehleringer JR (2008) 839 Life form-specific variations in leaf water oxygen-18 enrichment in Amazonian 840 vegetation. Oecologia 157: 197-210 841
Lange OL, Green TGA, Heber U (2001) Hydration-dependent photosynthetic 842 production of lichens: what do laboratory studies tell us about field performance? 843 Journal of Experimental Botany 52: 363 844
Luz B, Barkan E, Bender ML, Thiemens MH, Boering KA (1999) Triple-isotope 845 composition of atmospheric oxygen as a tracer of biosphere productivity. Nature 846 400: 547-550 847
Luz B, Barkan E, Yam R, Shemesh A (2009) Fractionation of oxygen and hydrogen 848 isotopes in evaporating water. Geochimica Et Cosmochimica Acta 73: 6697-6703 849
Martin CE (1994) Physiological Ecology of the Bromeliaceae. Botanical Review 60: 1-850 82 851
Martin CE, Lin TC, Lin KC, Hsu CC, Chiou W (2004) Causes and consequences of 852 high osmotic potentials in epiphytic higher plants. Journal of Plant Physiology 853 161: 1119-1124 854
Martin CE, Schmitt AK (1989) Unusual water relations in the CAM atmospheric 855 epiphyte Tillandsia usneoides L. (Bromeliaceae). Bot Gaz 150: 1-8 856
Medina E, Troughton JH (1974) Dark CO2 fixation and the carbon isotope ratio in 857 Bromeliaceae. Plant Science Letters 2: 357-362 858
Merlivat L (1978) Molecular diffusivities of H216O, HD16O and H2
18O in gases. J Chem 859 Phys 69: 2864-2871 860
Mez C (1904) Physiologische Bromeliaceen-Studien. I. Die wasser-Oekonomie der 861 extrem atmosphaerischen Tillandsien. Jahrbuecher fuer wissenschaftliched 862 Botanik 40: 157-229 863
Miller DL, Mora CI, Grissino-Mayer HD, Mock CJ, Uhle ME, Sharp Z (2006) Tree-864 ring isotope records of tropical cyclone activity. Proceedings of the National 865 Academy of Sciences of the United States of America 103: 14294-14297 866
Nowak EJ, Martin CE (1997) Physiological and anatomical responses to water deficits 867 in the CAM epiphyte Tillandsia ionantha (Bromeliaceae). International Journal of 868 Plant Sciences 158: 818-826 869
Ogee J, Cuntz M, P P, Bariac T (2007) Non-steady-state, non-uniform transpiration rate 870 and leaf anatomy effects on the progressive stable isotope enrichment of leaf 871 water along monocot leaves. Plant, Cell and Environment 30: 367-387 872
Ogee J, Peylin P, Cuntz M, Bariac T, Brunet Y, Berbigier P, Richard P, Ciais P 873 (2004) Partitioning net ecosystem carbon exchange into net assimilation and 874 respiration with canopy-scale isotopic measurements: An error propagation 875 analysis with (CO2)-C-13 and (COO)-O-18 data. Global Biogeochemical Cycles 876 18 877
Ohrui T, Nobira H, Sakata Y, Taji T, Yamamoto C, Nishida K, Yamakawa T, 878 Sasuga Y, Yaguchi Y, Takenaga H, Tanaka S (2007) Foliar trichome- and 879 aquaporin-aided water uptake in a drought-resistant epiphyte Tillandsia ionantha 880 Planchon. Planta 227: 47-56 881
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

32
Poussart PF, Schrag DP (2005) Seasonally resolved stable isotope chronologies from 882 northern Thailand deciduous trees. Earth and Planetary Science Letters 235: 752-883 765 884
Reyes-Garcia C, Mejia-Chang M, Jones GD, Griffiths H (2008) Water vapour 885 isotopic exchange by epiphytic bromeliads in tropical dry forests reflects niche 886 differentiation and climatic signals Plant, Cell and Environment 31: 828-841 887
Roden JS, Ehleringer JR (2000) Hydrogen and oxygen isotope ratios of tree ring 888 cellulose for field-grown riparian trees. Oecologia 123: 481-489 889
Smith JAC, Griffiths H, Luttge U (1986) Comparative ecophysiology of CAM and C3 890 Bromeliads. 1. The ecology of the Bromeliaceae in Trinidad. Plant Cell and 891 Environment 9: 359-376 892
Smith JAC, Griffiths H, Luttge U, Crook CE, Griffiths NM, Stimmel KH (1986) 893 Comparative ecophysiology of CAM and C3 Bromeliads. 3. Plant water relations. 894 Plant Cell and Environment 9: 395-410 895
Styles JM, Lloyd J, Zolotoukhine D, Lawton KA, Tchebakova N, Francey RJ, 896 Arneth A, Salamakho D, Kolle O, Schulze E-D (2002) Estimates of regional 897 surface carbon dioxide exchange and carbon and oxygen isotope discrimination 898 during photosynthesis from concentration profiles in the atmospheric boundry 899 layer. Tellus 54: 768-783 900
Switsur R, Waterhouse J (1998) Stable isotopes in tree ring cellulose. In H Griffiths, ed, 901 Stable Isotopes: integration of biological, ecological and geochemical processes. 902 BIOS Scientific Publishers Ltd, Oxford, pp 303-321 903
Winter K, Smith JAC, eds (1996) Crassulacean Acid Metabolism: Biochemistry, 904 Ecophysiology, and Evolution. , Vol 114. Springer Verlag, Berlin 905
Yakir D, Wang XF (1996) Fluxes of CO2 and water between terrestrial vegetation and 906 the atmosphere estimated from isotope measurements. Nature 380: 515-517 907
908 909
910
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
33
911
Table I. Representative chamber measurements of chamber air temperature, h, and δ18Oa over a 16 day 912
experimental period. Note that Day 0 was characterized by labeling the plants by soaking them in enriched 913
water for several hours and allowing both the chamber and water tower to reach the desired temperature 914
and h. Day 1 represents the first full nocturnal period that plants were in the chamber. 915
916
Experimental day
Chamber air temperature (°C)
h
δ18Oa
(‰) 1 27.5 ± 0.03 0.96 ± 0.01 2 27.5 ± 0.06 0.97 ± 0.01 -16.8 3 27.6 ± 0.20 0.96 ± 0.02 4 27.5 ± 0.04 0.98 ± 0.01 -17.0 5 27.5 ± 0.09 0.96 ± 0.01 6 27.5 ± 0.12 0.96 ± 0.01 7 27.4 ± 0.19 0.97 ± 0.01 8 27.5 ± 0.14 0.98 ± 0.01 -17.2 9 27.5 ± 0.13 0.98 ± 0.01 -17.1 10 27.5 ± 0.07 0.98 ± 0.01 11 27.4 ± 0.17 0.97 ± 0.01 -17.2 12 27.3 ± 0.19 0.93 ± 0.01 13 27.3 ± 0.20 0.94 ± 0.01 14 27.5 ± 0.14 0.96 ± 0.02 -17.0 15 27.5 ± 0.04 0.98 ± 0.01 16 27.5 ± 0.00 0.98 ± 0.01 -17.2 Multi-day mean 27.5 0.967 -17.1 Standard deviation 0.08 0.015 0.15
917
918
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
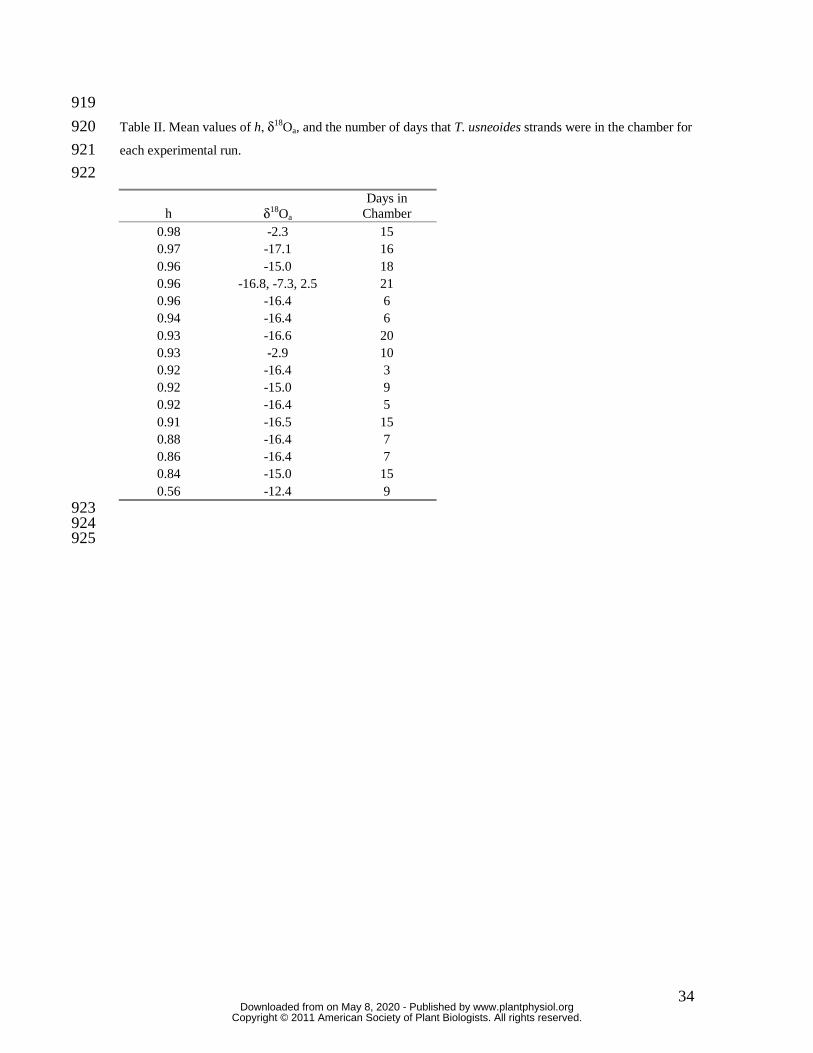
34
919
Table II. Mean values of h, δ18Oa, and the number of days that T. usneoides strands were in the chamber for 920
each experimental run. 921
922
h δ18Oa Days in
Chamber 0.98 -2.3 15 0.97 -17.1 16 0.96 -15.0 18 0.96 -16.8, -7.3, 2.5 21 0.96 -16.4 6 0.94 -16.4 6 0.93 -16.6 20 0.93 -2.9 10 0.92 -16.4 3 0.92 -15.0 9 0.92 -16.4 5 0.91 -16.5 15 0.88 -16.4 7 0.86 -16.4 7 0.84 -15.0 15 0.56 -12.4 9
923 924
925
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

35
926 Table III. The representative effect of rehydration on δ18OL through time during the chamber experiments. 927 On Day 10, one population of T. usneoides strands within the chamber were watered, while one population 928 remained dry. This scheme was repeated in 1/3 of the high h experiments in Table II with similar results. 929
Wetted plants δ18OL (‰)
Non-wetted plants δ18OL (‰)
Day 0 -3.0 ± 0.4 Day 7 -6.8 ± 0.6 Day 10 -7.4 ± 0.2 -6.8 ± 0.1 ns Day 10, 1 hour after rehydration 2.5 ± 0.6 Day 14 -6.1 ± 0.2 -6.3 ± 0.6 ns Day 21 -6.5 ± 0.4 -7.1 ± 0.7 ns
930 931
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
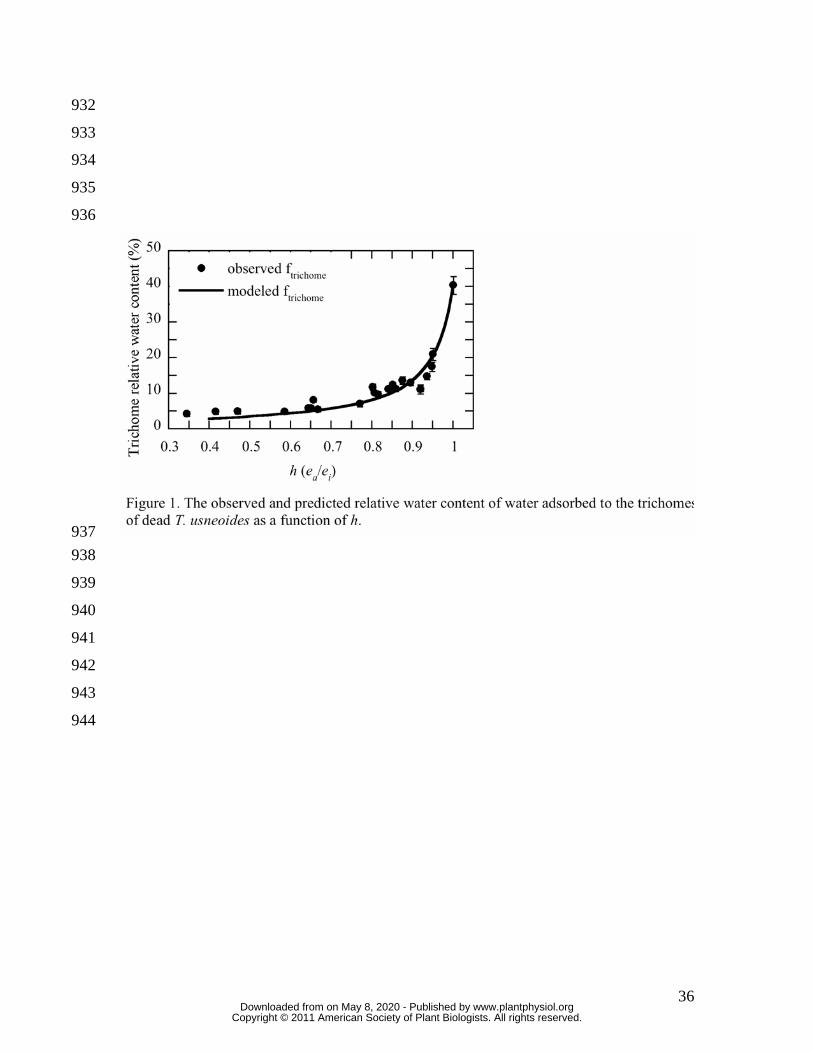
36
932
933
934
935
936
937
938
939
940
941
942
943
944
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
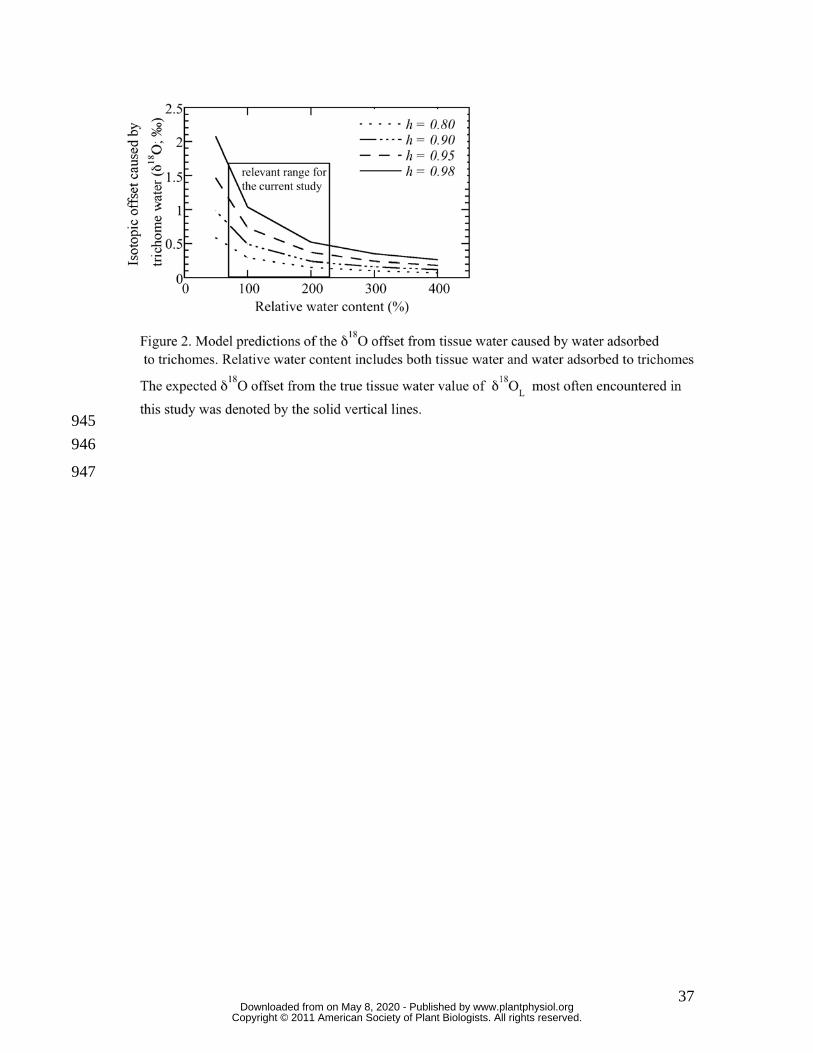
37
945
946
947
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
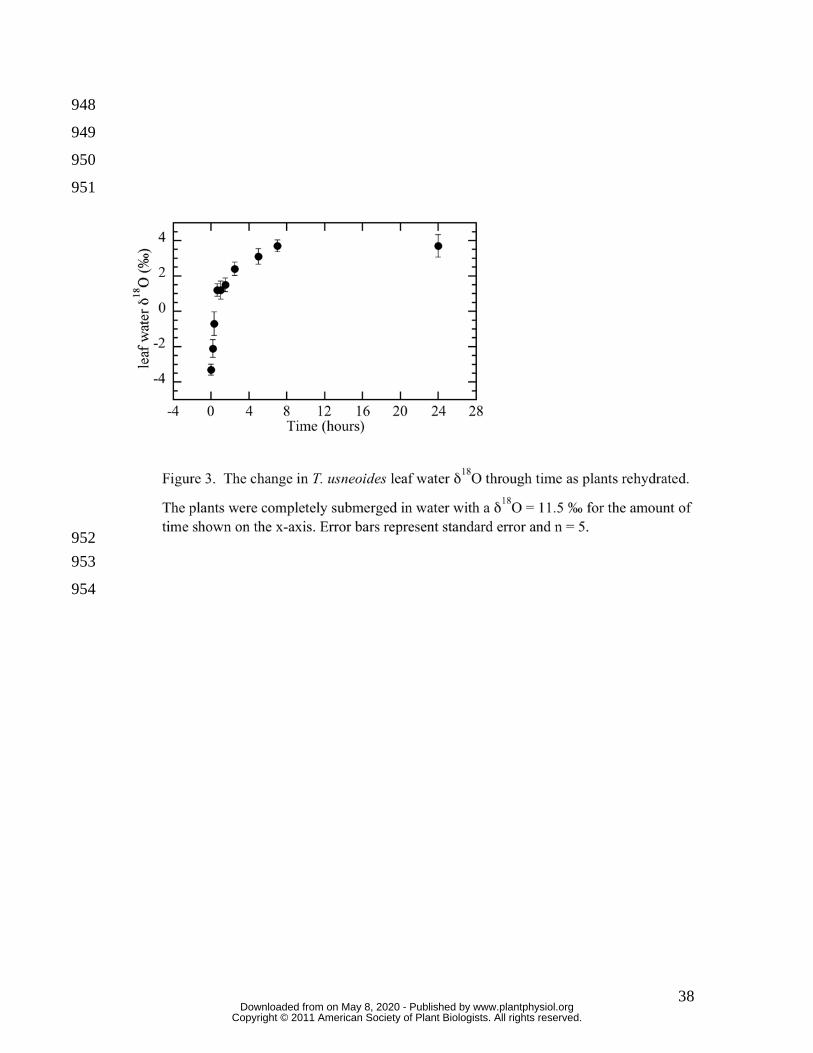
38
948
949
950
951
952
953
954
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
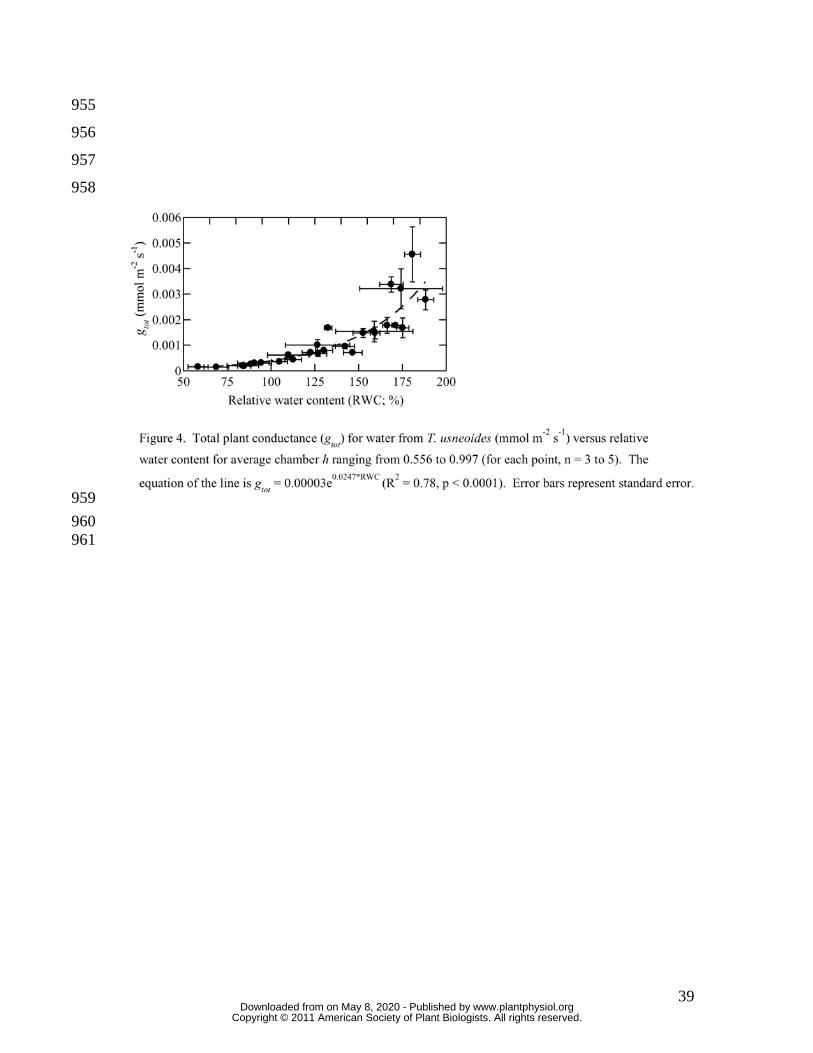
39
955
956
957
958
959
960 961
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
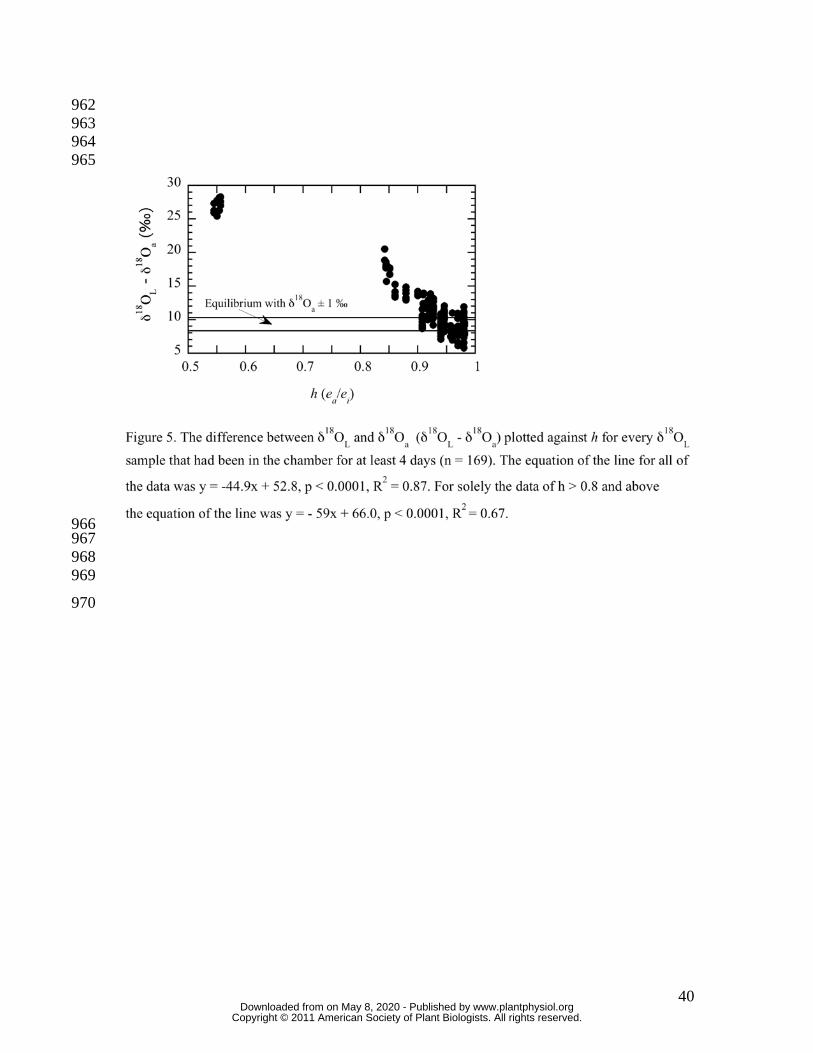
40
962 963 964 965
966 967 968 969
970
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
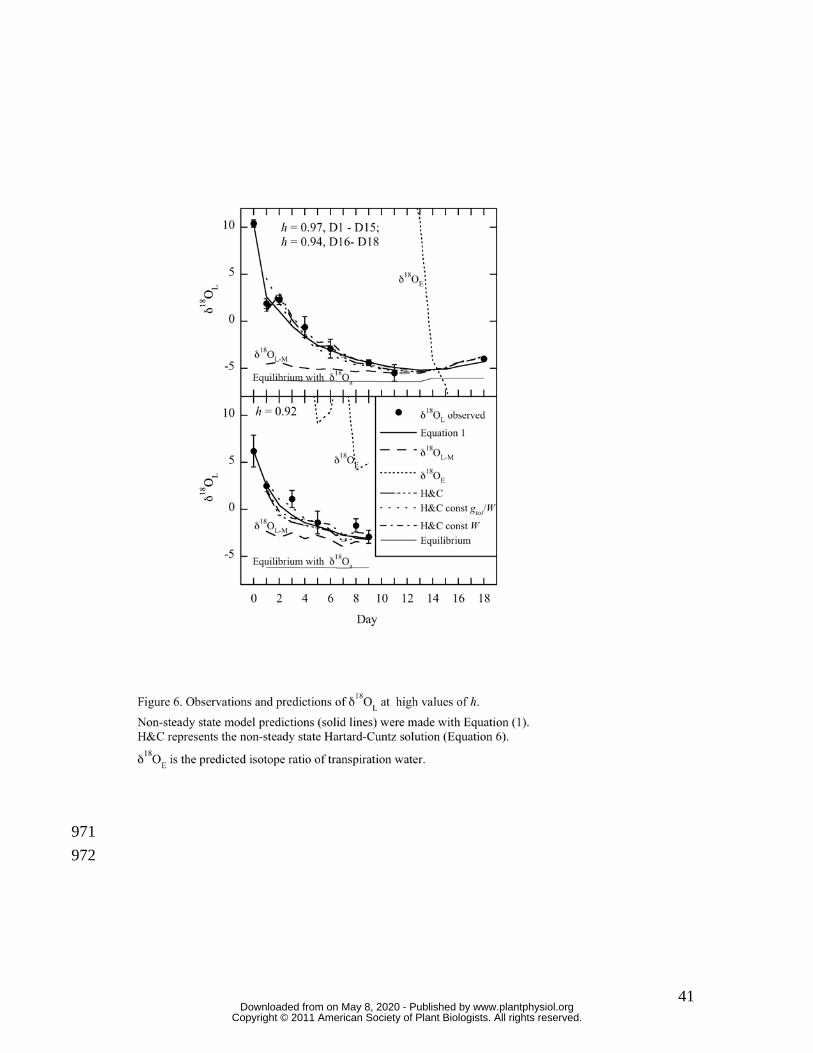
41
971
972
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

42
973
974
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
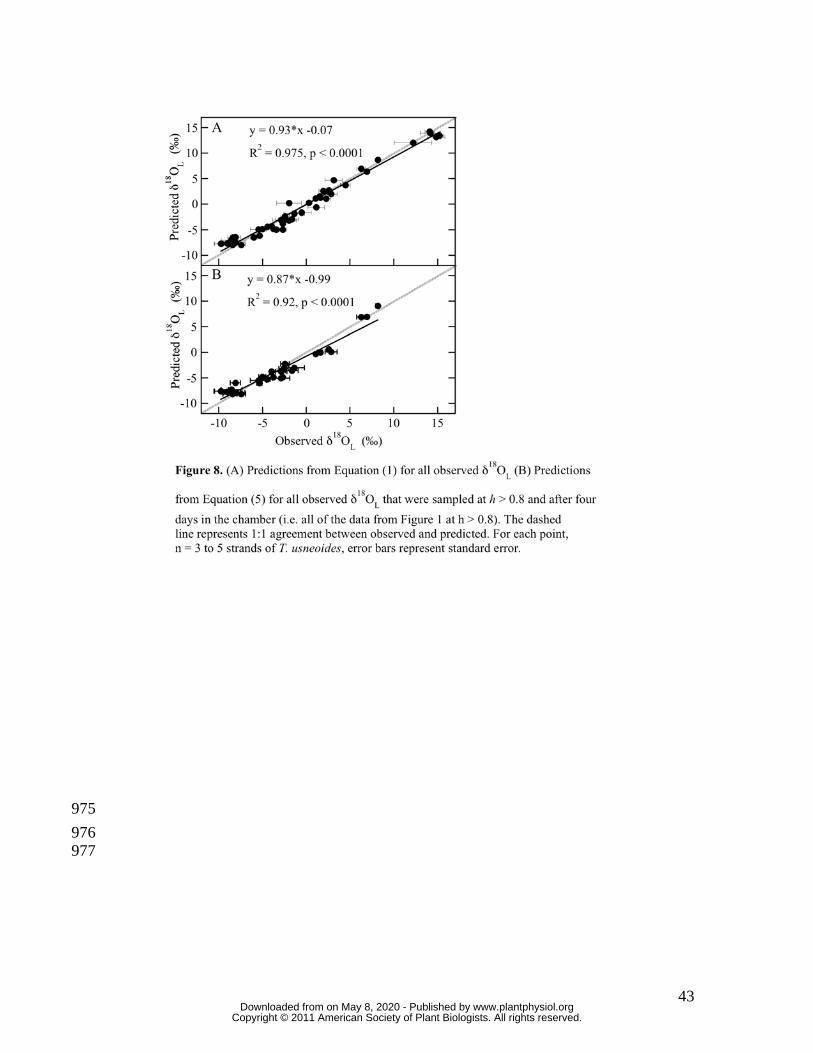
43
975
976 977
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
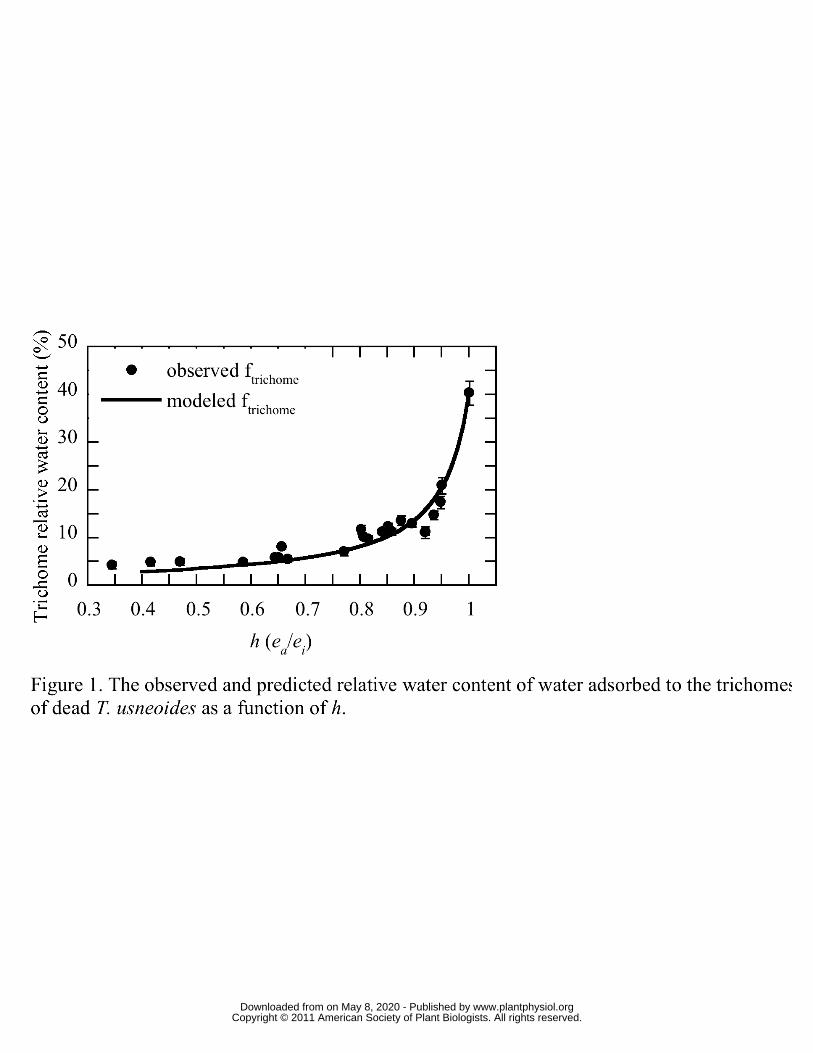
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
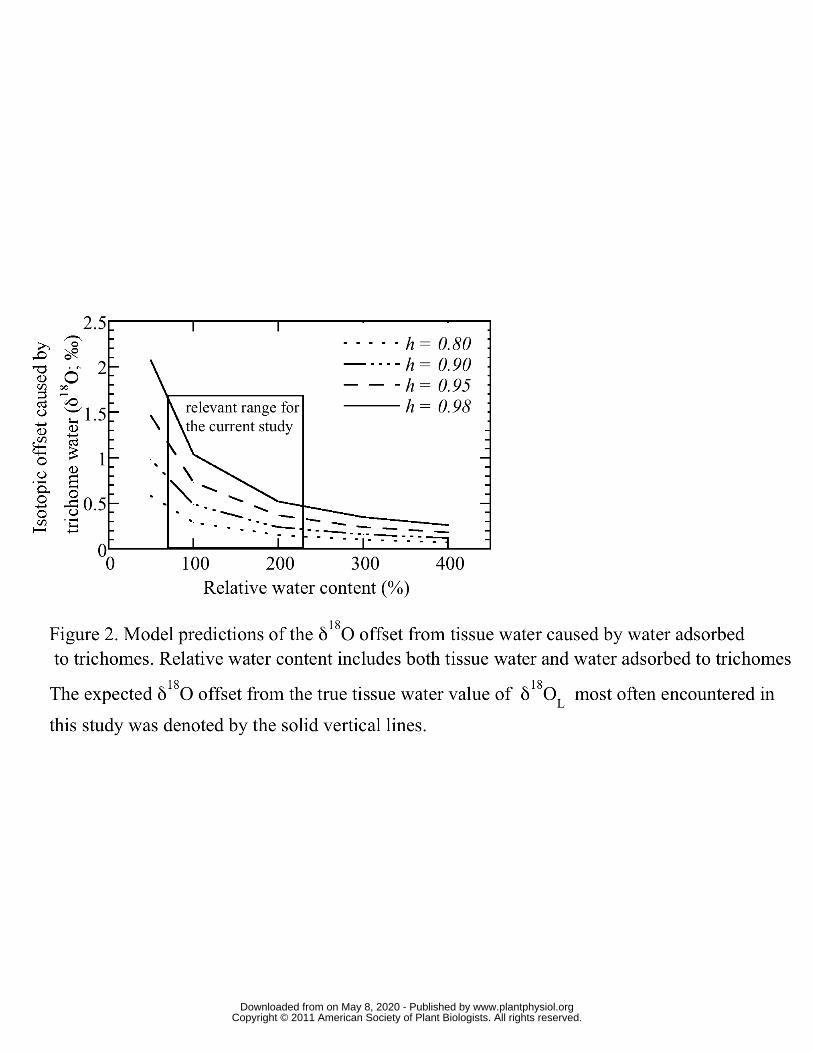
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
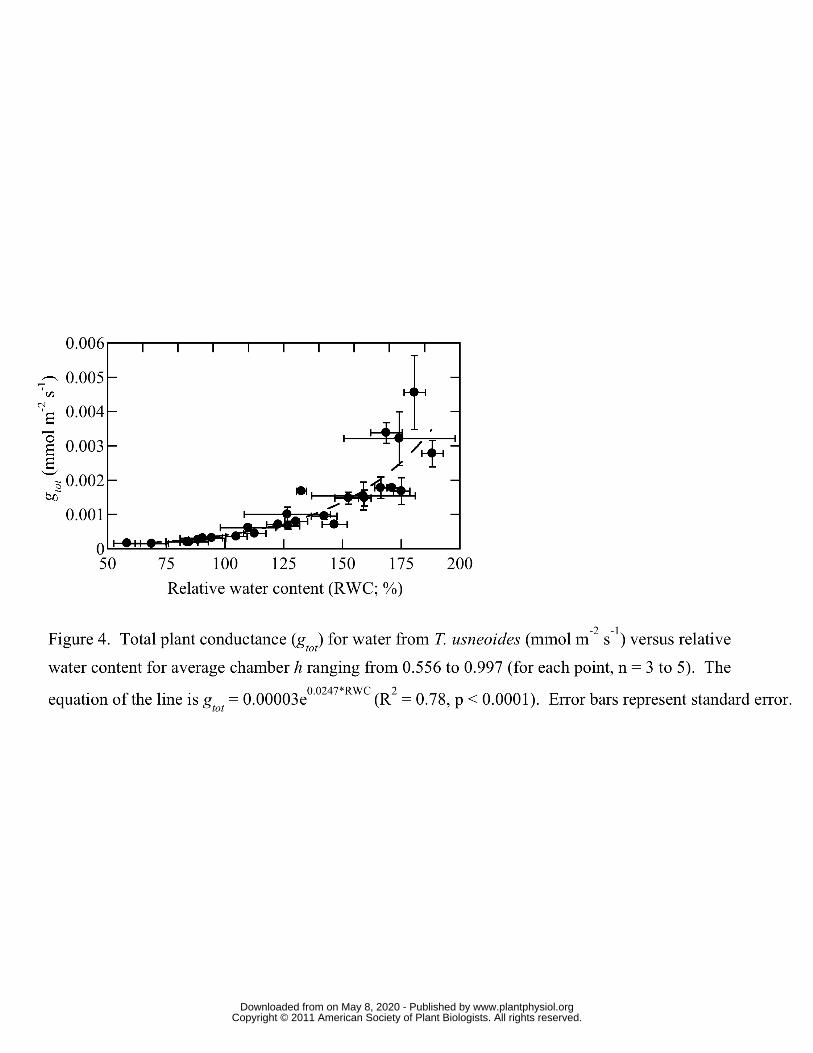
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
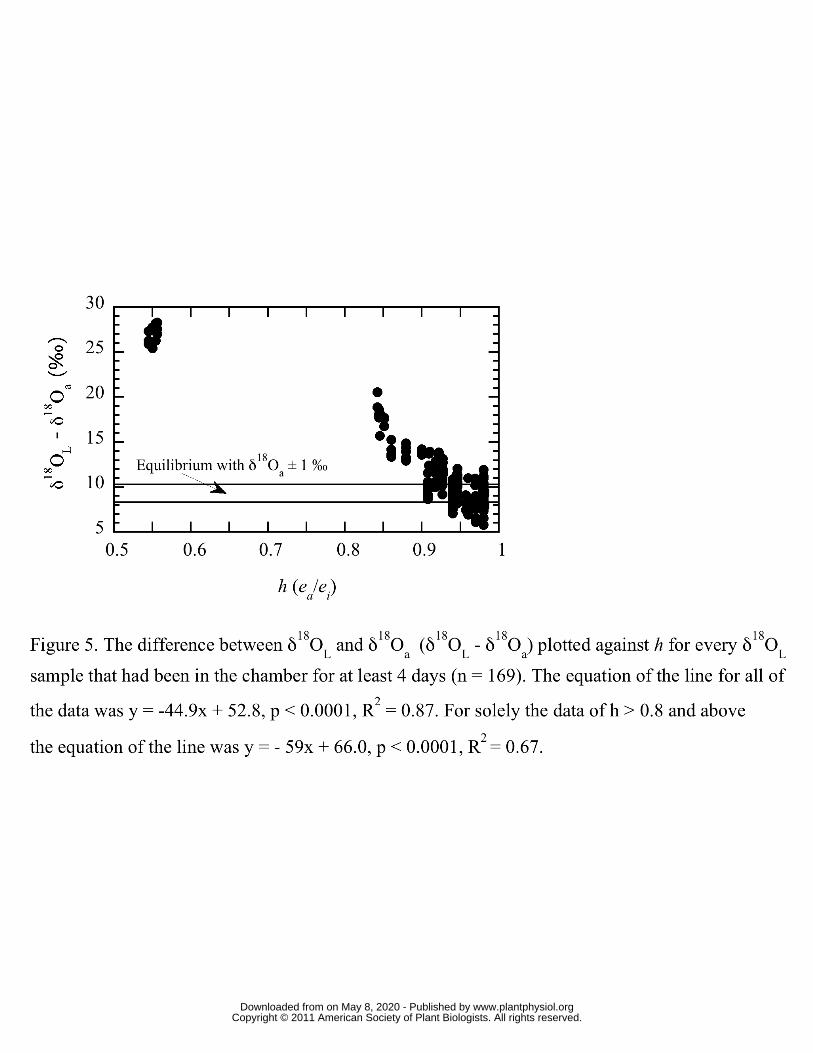
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
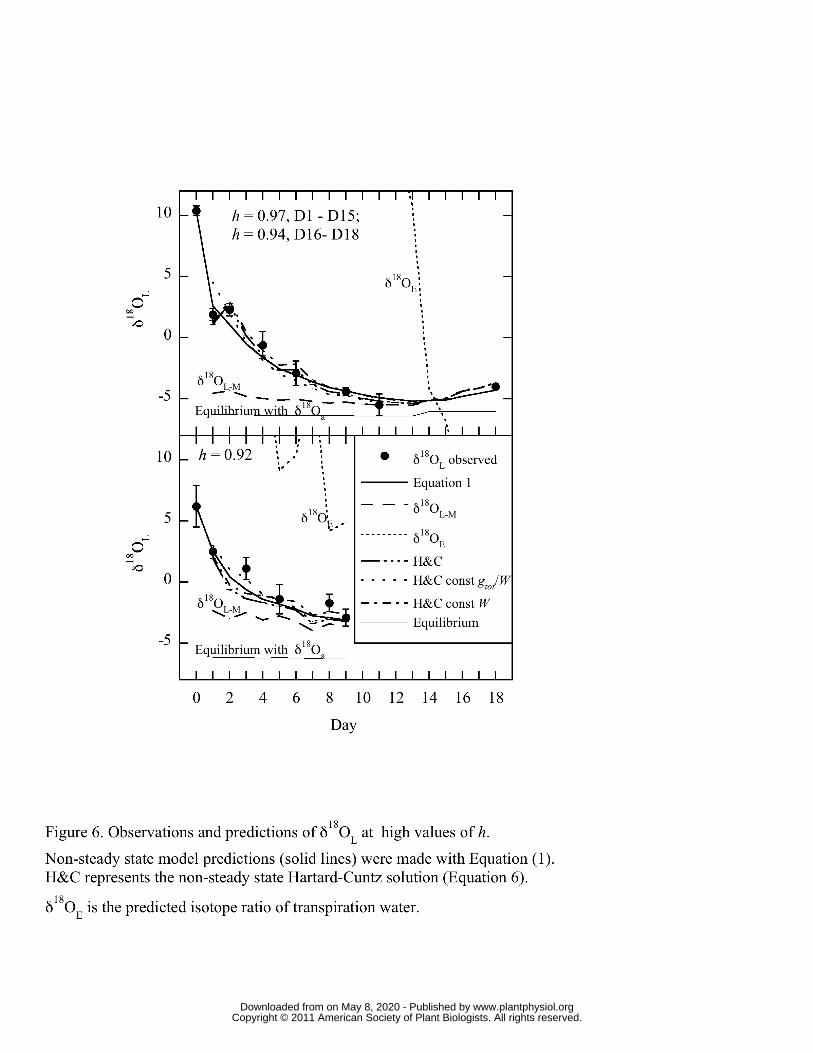
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
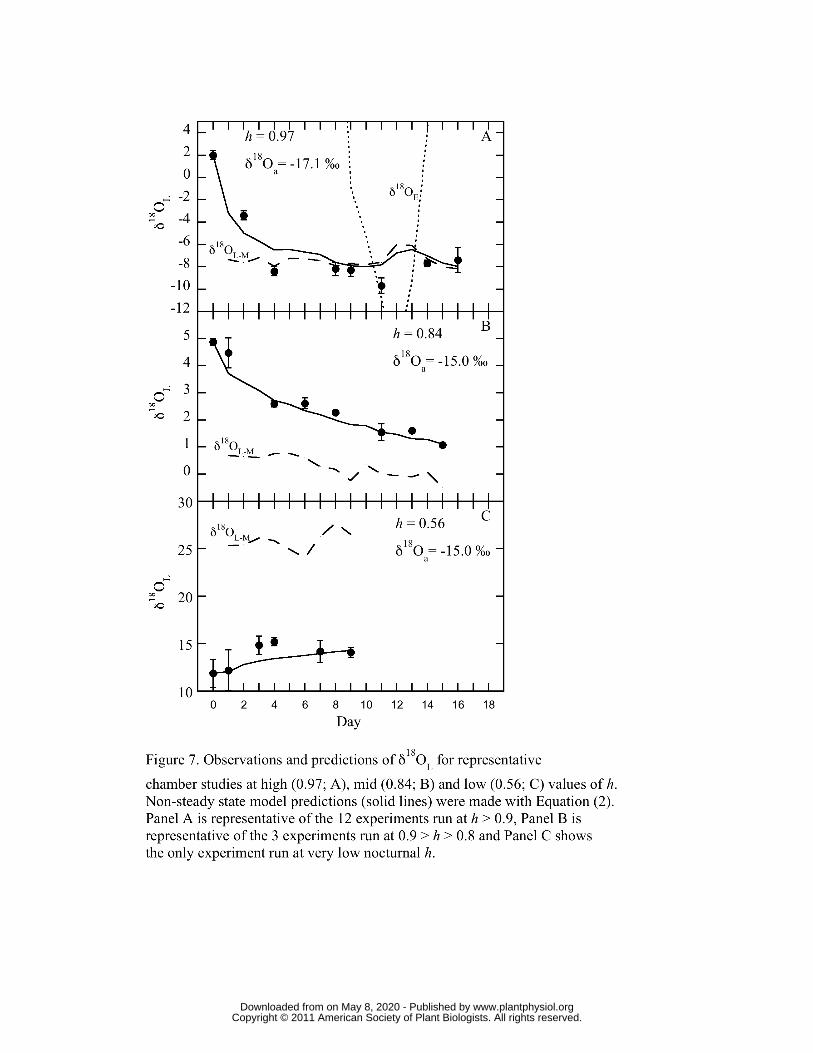
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.
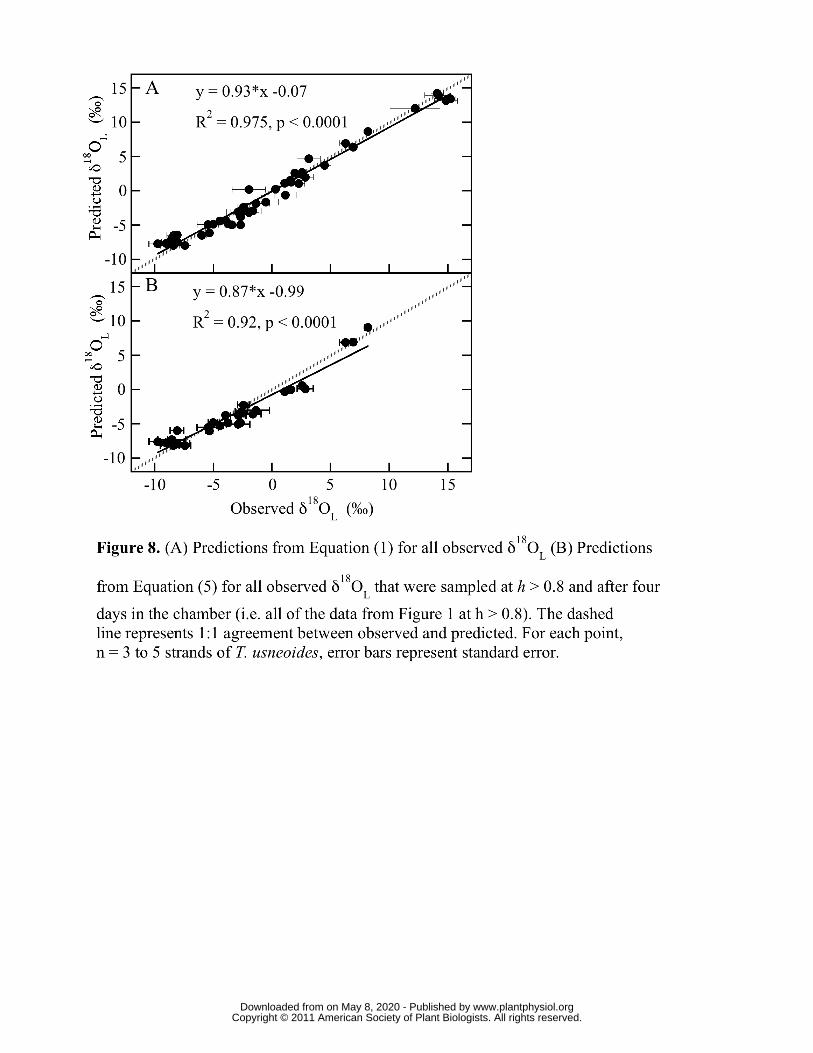
www.plantphysiol.orgon May 8, 2020 - Published by Downloaded from Copyright © 2011 American Society of Plant Biologists. All rights reserved.