Adaptive immunity 2 - Immune regulatory mechanisms through B cell axis

β-Catenin–related protein WRM-1 is a multifunctionalregulatory subunit of the LIT-1 MAPK complexXiao-Dong Yang, Tejas R. Karhadkar, Jessica Medina, Scott M. Robertson, and Rueyling Lin1
Department of Molecular Biology, University of Texas Southwestern Medical Center, Dallas TX 75390
Edited* by Craig C. Mello, University of Massachusetts Medical School, Worcester, MA, and approved December 5, 2014 (received for review August 25, 2014)
Vertebrate β-catenin has two functions, as a structural componentof the adherens junction in cell adhesion and as the T-cell factor(TCF) transcriptional coactivator in canonical Wnt (wingless-relatedintegration site) signaling. These two functions are split betweenthree of the four β-catenin–related proteins present in the roundworm Caenorhabditis elegans. The fourth β-catenin–related pro-tein, WRM-1, exhibits neither of these functions. Instead, WRM-1binds the MAPK loss of intestine 1 (LIT-1), and these two proteinshave been shown to be essential for the transcription of Wnt targetgenes by phosphorylating and regulating the nuclear level of thesole worm TCF protein. We showed previously that WRM-1 binds toworm TCF and functions as the substrate-binding subunit for LIT-1.In this study, we show that phosphorylation of T220 in the activa-tion loop is essential for LIT-1 kinase activity in vivo and in vitro.T220 can be phosphorylated either through LIT-1 autophosphoryla-tion or directly by the upstream MAP3K MOM-4. Our data supporta model in which WRM-1, which can undergo homotypic interac-tion, binds LIT-1 and thereby generates a kinase complex in whichLIT-1 molecules are situated in a conformation enabling autophos-phorylation as well as promoting phosphorylation of the T220 res-idue by MOM-4. In addition, we show that WRM-1 is essential forthe translocation of the LIT-1 kinase complex to the nucleus, the siteof its TCF substrate. To our knowledge, this is the first report of aMAP3K directly activating a MAPK by phosphorylation within theactivation loop. This study should help uncover novel and as yetunderappreciated functions of vertebrate β-catenin.
MAPK activation | C. elegans | LIT-1 | MOM-4 | MAP3K
The catenins function in cell–cell adhesion by linking thetransmembrane protein cadherin, which forms homotypic
interactions through its extracellular domain, to the underlyingactin cytoskeleton (1). The catenin gene family is composedof three subfamilies, namely p120 (seven members), beta (twomembers), and the structurally unrelated alpha (three members)(reviewed in ref. 2). p120 and beta family members all containa central domain composed of 9–12 armadillo (Arm) repeats (3),which fold upon themselves to form a superhelical ARM domainwith a positively charged central groove (4).The beta subfamily consists of β-catenin and its paralog
γ-catenin (also called “plakoglobin”), with diverged sequences andfunctions (2). β-Catenin functions in adherens junctions, whereasγ-catenin functions in desmosomes. In addition, β-catenin func-tions as a coactivator for the T-cell factor (TCF)/LEF transcriptionfactors in the wingless-related integration site (Wnt) pathway.The β-catenin gene also has undergone species-specific and
phylum-specific duplications in nonvertebrates. For example,planaria have two β-catenins, one functioning in adhesion andthe other in Wnt signaling (5). The Caenorhabditis elegans ge-nome encodes four highly diverged β-catenin–related proteins(HMP-2, BAR-1, SYS-1, and WRM-1), each as dissimilar toeach another in amino acid sequence as they are to DrosophilaArmadillo and mammalian β-catenin (6–10). However, structuredetermination and modeling, along with phylogenetic clusteringbased on protein sequence, support these four proteins beingbona fide 12 Arm repeat-containing homologs of vertebrateβ-catenin (11). In addition, genetic analyses showed that three of
these four β-catenin–related proteins exhibit one or the other ofthe two functions of vertebrate β-catenin (6–9, 12). HMP-2 bindscadherin (HMR-1), and α-catenin (HMP-1), localizes to adherensjunctions, and the hmp-2 mutant phenotype is entirely con-sistent with a defect in cell–cell adhesion (6, 12, 13). Therefore,HMP-2 appears to have retained only the cell–cell adhesionfunction of β-catenin. Both BAR-1 and SYS-1 bind to the con-served N-terminal β-catenin–binding domain of the sole C. elegansTCF protein, POP-1, function with POP-1 to activate transcriptionfrom Wnt reporter constructs in tissue-culture cells and in worms,and do not bind to α-catenin or cadherin (8, 12–14). It appearsthat both BAR-1 and SYS-1 retain the function of β-catenin ascoactivators for the TCF/POP-1 in a canonical Wnt pathway,but act at different developmental stages and in different de-velopmental processes (7, 8, 15–17).The fourth C. elegans β-catenin–related protein, WRM-1, does
not exhibit either of these two well-known functions of verte-brate β-catenin. WRM-1 does not bind to α-catenin, cadherin, orthe conserved N-terminal β-catenin–binding domain of TCF/POP-1, nor does it function as a coactivator of POP-1 in standardin vitro Wnt reporter assays (12–14). However, loss-of-functionand reduction-of-function genetic analyses clearly showed thatWRM-1 is required in early embryos in a Wnt signal-dependentprocess—the specification of the E blastomere in the eight-cellembryo as the sole intestinal precursor (10). Proper specificationof E requires a canonical Wnt signaling event that occurs at thefour-cell stage between EMS, the mother cell of E, and the ad-jacent posterior blastomere, P2 (10, 18–20). The β-catenin SYS-1functions as the coactivator for TCF/POP-1 in the transcriptionalactivation of Wnt target genes in E (15, 17). We have shownpreviously that the E blastomere exhibits a higher level ofβ-catenin SYS-1 and a lower nuclear level of TCF POP-1 than
Significance
Vertebrate β-catenin has two functions: as a structural compo-nent in cell adhesion and as the T-cell factor coactivator in ca-nonical Wnt (wingless-related integration site) signaling. Thesetwo functions are split between three β-catenin–related proteinsin Caenorhabditis elegans. A fourth worm β-catenin–relatedprotein, worm arm motif 1 (WRM-1), exhibits neither of thesefunctions but is essential, together with loss of intestine 1 (LIT-1)MAPK, for transcriptional activation of Wnt target genes. Herewe characterize previously unidentified mechanisms by whichthe LIT-1 kinase is activated and the different regulatory rolesthat WRM-1 plays in LIT-1 activation and LIT-1 subcellular local-ization. This study suggests the potential for novel and as yetundiscovered functions for vertebrate β-catenin.
Author contributions: X.-D.Y., S.M.R., and R.L. designed research; X.-D.Y., T.R.K., and J.M.performed research; X.-D.Y., S.M.R., and R.L. analyzed data; and X.-D.Y., S.M.R., and R.L.wrote the paper.
The authors declare no conflict of interest.
*This Direct Submission article had a prearranged editor.1To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1416339112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1416339112 PNAS | Published online December 29, 2014 | E137–E146
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
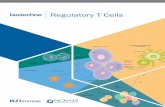
does its anterior sister, MS (15–17). The high SYS-1 and lowPOP-1 levels (a high β-catenin–to–TCF ratio) in E require theP2-to-EMS signal and are crucial for the transcriptional activa-tion of Wnt target genes in E by the SYS-1/POP-1 complex (10,15, 17, 19, 20).The main mechanism by which POP-1 nuclear levels are
lowered in E in response to the P2 signal is through nuclearexport, which requires POP-1 to be phosphorylated at five specificsites (21). We have shown that the MAPK LIT-1 is responsible forPOP-1 phosphorylation at these five sites, both in vivo and in vitro.In lit-1 mutant embryos, POP-1 nuclear levels remain high in boththe MS and E nuclei, and the E intestinal fate is repressed in bothblastomeres (10, 22). WRM-1 is required for LIT-1 phosphory-lation of POP-1, both in C. elegans embryos and in a mammaliantissue-culture cell–based assay (14, 21, 23). Active LIT-1 kinaseactivity has been obtained in vitro only when coexpressed inmammalian cells along with WRM-1 (14, 21, 23). WRM-1 binds toLIT-1 in C. elegans embryos and mammalian tissue-culture cells(14) and in a yeast two-hybrid assay (23). These results all dem-onstrate that LIT-1 activity in C. elegans embryos is regulatedthrough WRM-1 binding. Recent genetic analyses have suggestedthat in larvae CACN-1, a conserved protein of unknown function,also may regulate POP-1 levels partially through LIT-1 (24). Weshowed recently that one essential function of WRM-1 bound tothe LIT-1 kinase is to bind to the kinase substrate, TCF/POP-1(14). WRM-1 binds to a domain of POP-1 distinct from theconserved N-terminal β-catenin–binding domain to which bothSYS-1 and BAR-1 bind. In addition, our analyses showed a secondessential requirement for WRM-1 in LIT-1 phosphorylation ofPOP-1, separate from its substrate-binding function. The secondfunction of WRM-1 in the LIT-1 kinase complex, required forLIT-1 activity, remained unknown.
LIT-1 is an evolutionarily conserved atypical MAPKmost closelyrelated to Drosophila Nemo and mammalian Nemo-like kinase(NLK) (23, 25–27). Typical MAPKs have a conserved motif,threonine-X-tyrosine (TXY, in which X can be any amino acid),in their activation loop (Fig. 1A). Activation of typical MAPKsrequires phosphorylation of both the threonine and tyrosineresidues by an upstream dual-specificity MAP2K (28, 29).However, LIT-1, NEMO, and NLK all have a glutamic acidresidue (E) in the third position of this conserved activationmotif (T220HE, T200QE, and T286QE, respectively). No upstreamMAP2K capable of phosphorylating the threonine in this motifhas yet been identified. Multiple genetic screens for genesfunctioning in the specification of the intestinal precursor did notidentify any MAP2K (10, 20, 30). Unlike C. elegans LIT-1,mammalian NLK, when expressed in tissue-culture cells, doesnot appear to require a binding partner for activation (25, 31,32). Indeed, it was shown that NLK activation can be achievedthrough homodimerization. Upon homodimerization in vitro,NLK can autophosphorylate the T286 residue of its TQE acti-vation motif (33).Homeodomain-interacting protein kinase 2 (HIPK2) and
MAP3K7 (previously TAK1) have been shown to phosphorylateNLK and promote its activation (34–37). However, it is not clearwhether these kinases phosphorylate the activation loop of NLKor promote its autophosphorylation. Among the genes identifiedby genetic screens as functioning in the specification of theC. elegans intestinal precursor was mom-4, the C. elegans homologof MAP3K7 (10, 20, 30). mom-4 was found to function upstreamof lit-1 and wrm-1. However, although depletion of either lit-1 orwrm-1 results in 100% of the embryos lacking intestine (gutlessphenotype), strong loss-of-function mom-4 mutations resultedin no more than 50% of the embryos exhibiting the gutless
A
B C
T220-P
LIT-1
wildtyp
e
lit-1 (
t1512
)
lit-1 (
or131
)
+–CIP
T220-P
LIT-1
D
wrm-1(
-)
T220-P
LIT-1
WT
T220V
T220D
Flag::POP-1 + ++Myc::WRM-1 + ++
GFP::LIT-1 –+
WT
Myc
POP-1
S117-P
T220-P
Myc
GFP IP GFP
Input
E
IP Flag
wildtyp
e
Fig. 1. WRM-1 is important for LIT-1 T220 phosphorylation in worm embryos and mammalian cells. (A) Sequences of the activation loop from mouse (Mm)ERK2, p38α, JNK1, and NLK, Drosophila (Dm) NEMO, and C. elegans (Ce) LIT-1, with identical amino acids highlighted in yellow. Conserved amino acids in theactivation motifs, TXY or TXE, are in red. (B–D) Western blots using anti–T220-P and anti–LIT-1 antibodies on wild-type embryo extracts that had been treated(+) or not treated (−) with calf-intestinal alkaline phosphatase (CIP) (B), embryo extracts derived from wild-type, lit-1(t1512), or lit-1(or131) worms raised at25 °C (C), and embryo extracts derived from wild-type or wrm-1–depleted worms (D). (E) GFP-tagged LIT-1 variants were coexpressed with Myc-tagged WRM-1 and Flag-tagged POP-1 in HeLa cells. Following anti-GFP (LIT-1) or anti-Flag (POP-1) pulldowns, Western blots were performed with the antibodies indicated.Input Myc, anti-Myc blot of extract before pulldowns; IP, immunoprecipitation.
E138 | www.pnas.org/cgi/doi/10.1073/pnas.1416339112 Yang et al.

phenotype (10, 20, 22, 27, 30), indicating that activation of LIT-1 is not entirely dependent upon MOM-4. How MOM-4 func-tioned in the promotion of LIT-1 activity was not known.In this study, we show that phosphorylation of T220 in the
THE motif of the LIT-1 activation loop is essential for LIT-1kinase function in vivo. T220 can be phosphorylated eitherthrough LIT-1 autophosphorylation or directly by the MAP3KMOM-4. WRM-1 is required for LIT-1 T220 autophosphor-ylation and promotes T220 phosphorylation by MOM-4. Weshow that WRM-1 normally forms multimers and by so doingpromotes multimerization of the LIT-1/WRM-1 complex. Thismultimerization results in LIT-1 self-association that favorsautophosphorylation of LIT-1 at T220.
ResultsLIT-1 Is Phosphorylated at T220 in Wild-Type Embryos. LIT-1 T220 islocated in the putative activation loop within kinase domains VIIand VIII and is equivalent to T286 of mouse NLK and T200 ofDrosophila Nemo, corresponding to T180 of human and mousep38 MAPK (Fig. 1A) (23, 30). It has been shown that T220 isrequired for LIT-1 kinase activity in vitro (30). We now showthat T220 is required for LIT-1 activity in C. elegans embryos.The lit-1(t1512) temperature-sensitive (ts) mutation changes leucine177 in the kinase domain of LIT-1, which also is conserved inboth Nemo and NLK, to serine (L177S), resulting in 100%embryonic lethality at the nonpermissive temperature (25 °C),with all embryos lacking intestinal cells (100% gutless, n >300)(10, 22). A transgene that expresses wild-type GFP::LIT-1 spe-cifically in the EMS blastomere fully rescues the intestinal defect(0% gutless, n = 50; Table 1). A transgene that expressed GFP::LIT-1 carrying either a T220-to-valine (T220V) or T220-to-aspartate (T220D) mutation failed to rescue (100% gutless, n =50 and 54, respectively). This result demonstrates that T220 isabsolutely essential for LIT-1 kinase activity in vivo.To elucidate the mechanism by which the LIT-1 kinase is ac-
tivated in vivo, we raised an antibody against a portion of LIT-1that included phosphorylated T220. We will refer to this antibodyas “anti–T220-P.” A control antibody against amino acids 335–454of LIT-1 (anti–LIT-1) was generated also. On Western blots, bothanti–T220-P and anti–LIT-1 detected an expected band of ∼50 kDin wild-type embryo extracts (Fig. 1B). Pretreatment of the embryolysates with phosphatase before electrophoresis abolished theanti–T220-P but not the anti–LIT-1 reactive band, indicatingthat the anti–T220-P antibody recognizes LIT-1 phosphorylated atT220 in vivo. We also performed Western blots using lysates
prepared from lit-1(t1512) embryos, as well as from another tsmutant, lit-1(or131) (22, 27), raised at 25 °C. Although the anti–LIT-1 band was detected in embryo extracts from both mutantstrains at levels comparable to those in wild type, the anti–T220-Pband was not detected (Fig. 1C), showing that both ts lit-1 muta-tions do not affect stability of the LIT-1 protein but do greatlyinterfere with phosphorylation at T220.
WRM-1 Regulates LIT-1 Phosphorylation at T220.Western blots usingthe anti–T220-P antibody and lysates prepared from embryosthat had been depleted of WRM-1 were performed. The deletionmutation wrm-1(tm514) results in embryonic lethality (38) andcannot be maintained as a homozygous stock. We preparedlysates from embryos derived from wrm-1(tm514)/+ mothers thathad been fed with wrm-1(RNAi) bacteria. The anti–T220-P re-activity was greatly reduced in these wrm-1− embryo extracts,compared with wild-type embryo extracts (Fig. 1D), demon-strating that in C. elegans embryos WRM-1 activity is requiredfor LIT-1 phosphorylation at T220.The WRM-1–dependent LIT-1 phosphorylation at T220 ob-
served in C. elegans embryos was recapitulated in mammaliantissue-culture cells (Fig. 1E). LIT-1 is not phosphorylated atT220 when expressed by itself in HeLa cells. However, whencoexpressed with WRM-1, LIT-1 phosphorylation at T220 isdetected by Western blot. We also assayed LIT-1 activity bycoexpressing POP-1 and measuring LIT-1–dependent POP-1phosphorylation using an antibody specific for POP-1 phosphor-ylated at S117 (anti–S117-P). S117 is one of the five residuesphosphorylated by LIT-1 that are required for POP-1 nuclearexport (21). POP-1 S117 phosphorylation was detected only whenPOP-1 was coexpressed with wild-type LIT-1 and WRM-1 but notwhen wild-type LIT-1 was expressed without WRM-1 or when theLIT-1 T220V or T220D variants were coexpressed with WRM-1(Fig. 1E and Table 1). These results demonstrate that T220 isessential for LIT-1 kinase activity and that WRM-1 promotes LIT-1kinase activity by promoting its phosphorylation at T220.
LIT-1 Can Autophosphorylate at T220. We have been unable toobtain active recombinant LIT-1 from bacteria. His-tagged LIT-1purified from bacteria did not undergo autophosphorylationand was unable to phosphorylate POP-1, either by itself or whenbacterially purified maltose-binding protein (MBP)-tagged WRM-1was included. Therefore, to test whether LIT-1 can undergoautophosphorylation, we immunoprecipitated LIT-1 frommammalian cell lysates, incubated it with ATP in an in vitro
Table 1. Summary of transgenic GFP::LIT-1 proteins
TransgenicGFP::LIT-1variant
In vivo rescue of lit-1(t1512) gutless phenotype*, %
gutless (no. ofembryos scored)
In vitro kinase activityusing POP-1 as a substrate
Binding to WRM-1as assayed by co-IP
Subcellular localizationin embryos
None 100 (>300) N.A. N.A. N.A.Wild type 0 (50) Yes Yes N>>CT220V 100 (50) No Yes N<<CT220D 100 (63) No Yes N = CD186N 100 (30) No Yes N ≥ CK89M 100 (32) No Yes N ≥ CK89G 100 (43) No No N<<CL177S 100 (32) No No N<<C (excluded in N)C361Y 0 (21) Yes, reduced Yes N ≥ C
Comparison of GFP::LIT-1 wild type versus seven GFP::LIT-1 variants for (i) the ability to rescue the gutless phenotype of lit-1(t1512) asassayed by presence or absence of gut granules; percent gutless transgenic embryos and number of embryos scored are indicated; (ii)the ability to phosphorylate POP-1 in an in vitro kinase assay, as measured by S117-P by Western blot; (iii) binding to Flag::WRM-1 bycoimmunoprecipitation following coexpression in tissue-culture cells; and (iv) subcellular localization in transgenic embryos. C, cyto-plasm; co-IP, coimmunoprecipitation; N, nucleus; N.A., not applicable.*Scored by the presence or absence of birefringent gut granules.
Yang et al. PNAS | Published online December 29, 2014 | E139
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
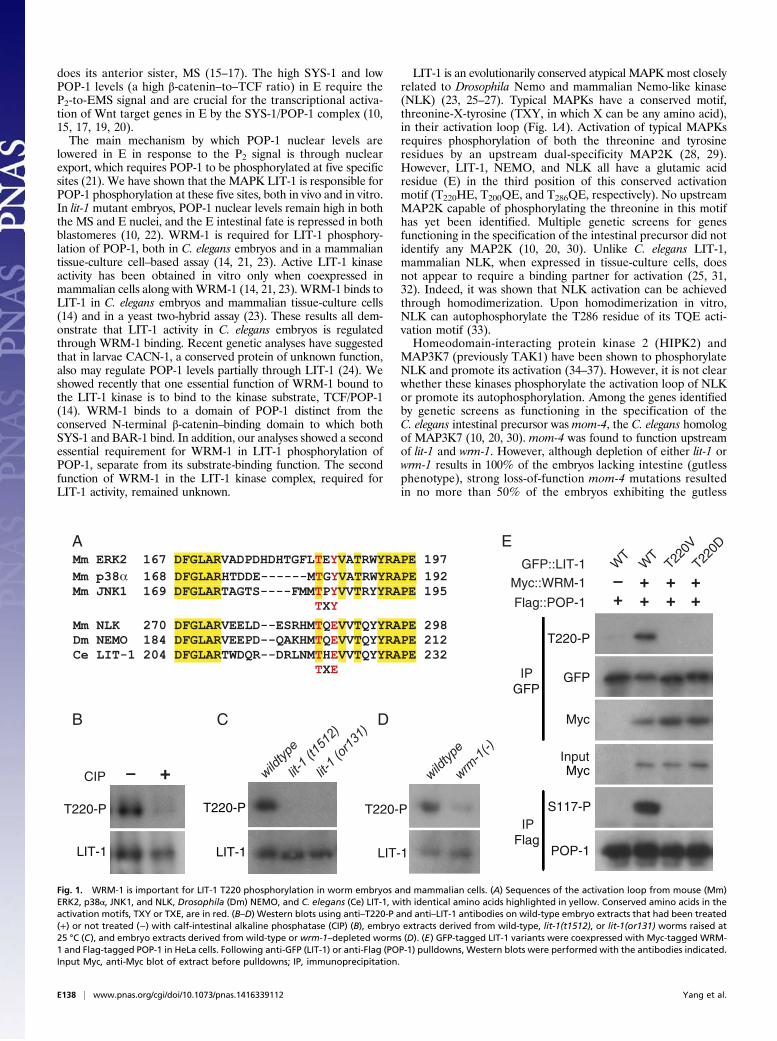
kinase assay, and then evaluated phosphorylation at T220. Whensuch an experiment was performed using lysates from tissue-culture cells expressing LIT-1 by itself, no phosphorylation ofLIT-1 at T220 was observed, even after incubation with ATP inthe kinase assay (Fig. 2A, lanes 1 and 2). However, LIT-1 pulleddown from cell lysates in which WRM-1 had been coexpressedexhibited clear T220 phosphorylation, which was increased fur-ther following incubation with ATP (Fig. 2A, lanes 3 and 4). Noincrease in anti–T220-P level was observed following the addi-tion of ATP to the in vitro kinase assay when a kinase-dead (KD)version of LIT-1, in which the catalytic aspartate was mutated toasparagine (D186N), was coexpressed with WRM-1 (Fig. 2A,lanes 5 and 6). This result demonstrates that the increased T220phosphorylation is caused by LIT-1 autophosphorylation.
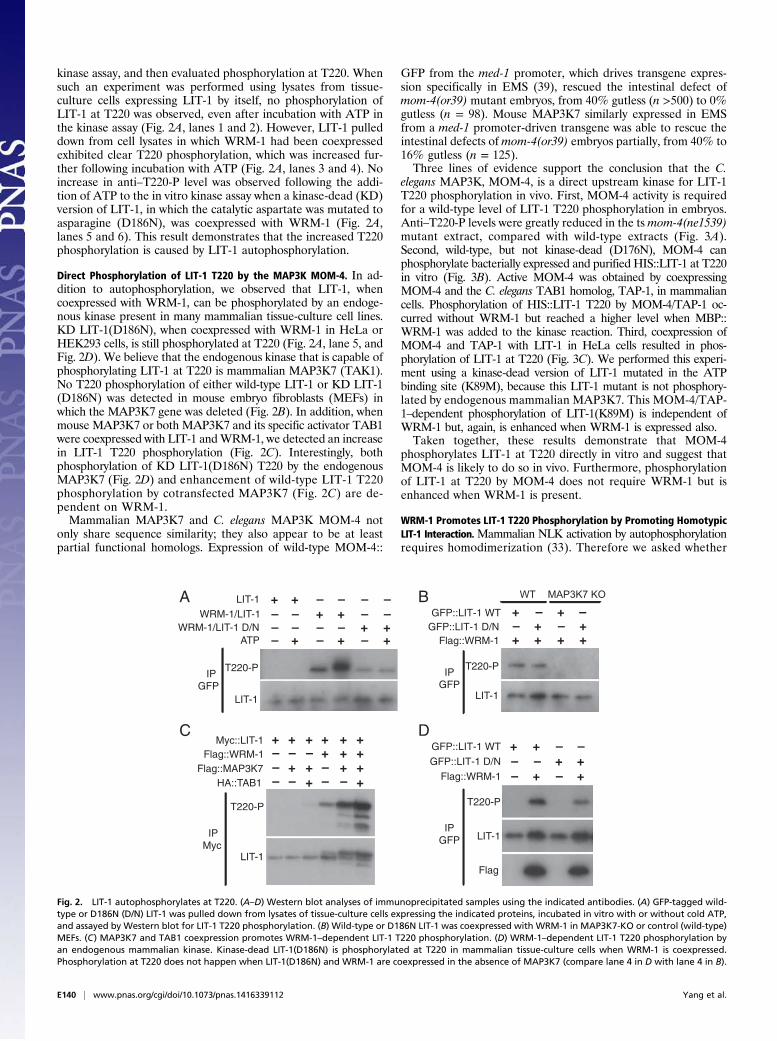
Direct Phosphorylation of LIT-1 T220 by the MAP3K MOM-4. In ad-dition to autophosphorylation, we observed that LIT-1, whencoexpressed with WRM-1, can be phosphorylated by an endoge-nous kinase present in many mammalian tissue-culture cell lines.KD LIT-1(D186N), when coexpressed with WRM-1 in HeLa orHEK293 cells, is still phosphorylated at T220 (Fig. 2A, lane 5, andFig. 2D). We believe that the endogenous kinase that is capable ofphosphorylating LIT-1 at T220 is mammalian MAP3K7 (TAK1).No T220 phosphorylation of either wild-type LIT-1 or KD LIT-1(D186N) was detected in mouse embryo fibroblasts (MEFs) inwhich the MAP3K7 gene was deleted (Fig. 2B). In addition, whenmouse MAP3K7 or both MAP3K7 and its specific activator TAB1were coexpressed with LIT-1 andWRM-1, we detected an increasein LIT-1 T220 phosphorylation (Fig. 2C). Interestingly, bothphosphorylation of KD LIT-1(D186N) T220 by the endogenousMAP3K7 (Fig. 2D) and enhancement of wild-type LIT-1 T220phosphorylation by cotransfected MAP3K7 (Fig. 2C) are de-pendent on WRM-1.Mammalian MAP3K7 and C. elegans MAP3K MOM-4 not
only share sequence similarity; they also appear to be at leastpartial functional homologs. Expression of wild-type MOM-4::
GFP from the med-1 promoter, which drives transgene expres-sion specifically in EMS (39), rescued the intestinal defect ofmom-4(or39) mutant embryos, from 40% gutless (n >500) to 0%gutless (n = 98). Mouse MAP3K7 similarly expressed in EMSfrom a med-1 promoter-driven transgene was able to rescue theintestinal defects ofmom-4(or39) embryos partially, from 40% to16% gutless (n = 125).Three lines of evidence support the conclusion that the C.
elegans MAP3K, MOM-4, is a direct upstream kinase for LIT-1T220 phosphorylation in vivo. First, MOM-4 activity is requiredfor a wild-type level of LIT-1 T220 phosphorylation in embryos.Anti–T220-P levels were greatly reduced in the tsmom-4(ne1539)mutant extract, compared with wild-type extracts (Fig. 3A).Second, wild-type, but not kinase-dead (D176N), MOM-4 canphosphorylate bacterially expressed and purified HIS::LIT-1 at T220in vitro (Fig. 3B). Active MOM-4 was obtained by coexpressingMOM-4 and the C. elegans TAB1 homolog, TAP-1, in mammaliancells. Phosphorylation of HIS::LIT-1 T220 by MOM-4/TAP-1 oc-curred without WRM-1 but reached a higher level when MBP::WRM-1 was added to the kinase reaction. Third, coexpression ofMOM-4 and TAP-1 with LIT-1 in HeLa cells resulted in phos-phorylation of LIT-1 at T220 (Fig. 3C). We performed this experi-ment using a kinase-dead version of LIT-1 mutated in the ATPbinding site (K89M), because this LIT-1 mutant is not phosphory-lated by endogenous mammalian MAP3K7. This MOM-4/TAP-1–dependent phosphorylation of LIT-1(K89M) is independent ofWRM-1 but, again, is enhanced when WRM-1 is expressed also.Taken together, these results demonstrate that MOM-4
phosphorylates LIT-1 at T220 directly in vitro and suggest thatMOM-4 is likely to do so in vivo. Furthermore, phosphorylationof LIT-1 at T220 by MOM-4 does not require WRM-1 but isenhanced when WRM-1 is present.
WRM-1 Promotes LIT-1 T220 Phosphorylation by Promoting HomotypicLIT-1 Interaction. Mammalian NLK activation by autophosphorylationrequires homodimerization (33). Therefore we asked whether
BA
C
IP GFP
T220-P
LIT-1
ATP +– +– +–
WRM-1/LIT-1WRM-1/LIT-1 D/N
LIT-1 + –+ – – –+– +– – –
+– +–– –
T220-P
LIT-1
Flag
IP GFP
GFP::LIT-1 D/N
GFP::LIT-1 WT
–+
+–+
+–
–+Flag::WRM-1 + ––
D
GFP::LIT-1 D/NGFP::LIT-1 WT
–+
+–+
+–
–++
WT
Flag::WRM-1 ++
MAP3K7 KO
T220-P
LIT-1
IP GFP
Myc::LIT-1
HA::TAB1
Flag::WRM-1Flag::MAP3K7
+ ++ + ++
– +– – +–+– – ++ +
+ ++–– –
T220-P
LIT-1
IP Myc
Fig. 2. LIT-1 autophosphorylates at T220. (A–D) Western blot analyses of immunoprecipitated samples using the indicated antibodies. (A) GFP-tagged wild-type or D186N (D/N) LIT-1 was pulled down from lysates of tissue-culture cells expressing the indicated proteins, incubated in vitro with or without cold ATP,and assayed by Western blot for LIT-1 T220 phosphorylation. (B) Wild-type or D186N LIT-1 was coexpressed with WRM-1 in MAP3K7-KO or control (wild-type)MEFs. (C) MAP3K7 and TAB1 coexpression promotes WRM-1–dependent LIT-1 T220 phosphorylation. (D) WRM-1–dependent LIT-1 T220 phosphorylation byan endogenous mammalian kinase. Kinase-dead LIT-1(D186N) is phosphorylated at T220 in mammalian tissue-culture cells when WRM-1 is coexpressed.Phosphorylation at T220 does not happen when LIT-1(D186N) and WRM-1 are coexpressed in the absence of MAP3K7 (compare lane 4 in D with lane 4 in B).
E140 | www.pnas.org/cgi/doi/10.1073/pnas.1416339112 Yang et al.
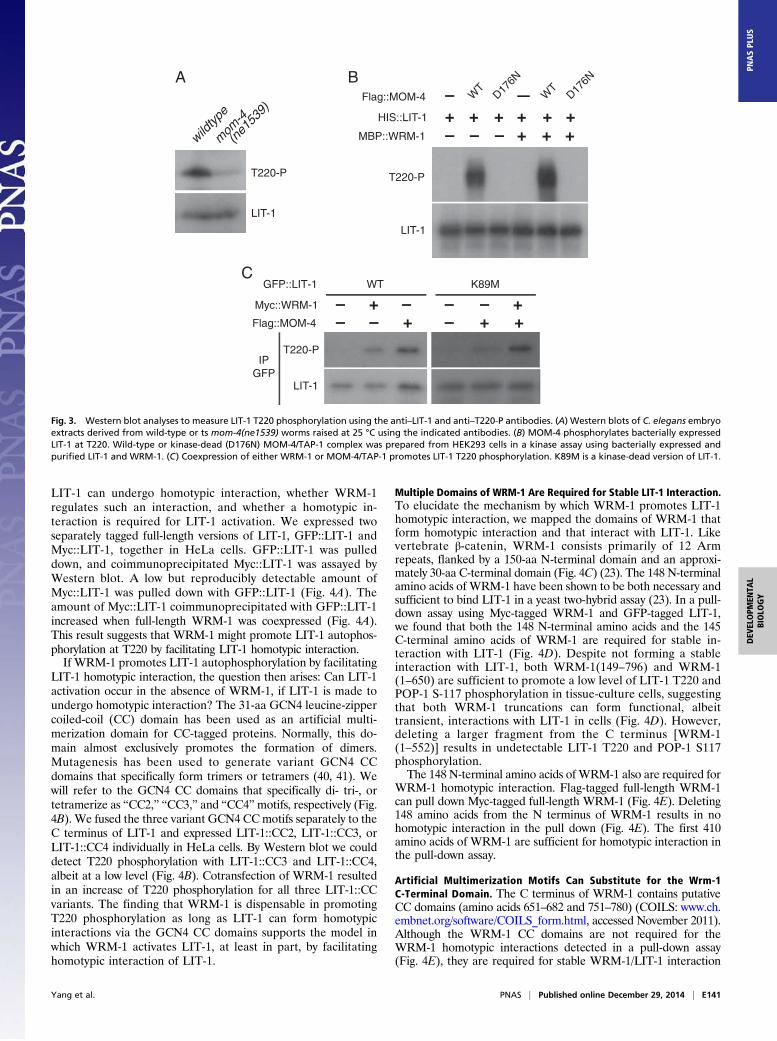
LIT-1 can undergo homotypic interaction, whether WRM-1regulates such an interaction, and whether a homotypic in-teraction is required for LIT-1 activation. We expressed twoseparately tagged full-length versions of LIT-1, GFP::LIT-1 andMyc::LIT-1, together in HeLa cells. GFP::LIT-1 was pulleddown, and coimmunoprecipitated Myc::LIT-1 was assayed byWestern blot. A low but reproducibly detectable amount ofMyc::LIT-1 was pulled down with GFP::LIT-1 (Fig. 4A). Theamount of Myc::LIT-1 coimmunoprecipitated with GFP::LIT-1increased when full-length WRM-1 was coexpressed (Fig. 4A).This result suggests that WRM-1 might promote LIT-1 autophos-phorylation at T220 by facilitating LIT-1 homotypic interaction.If WRM-1 promotes LIT-1 autophosphorylation by facilitating
LIT-1 homotypic interaction, the question then arises: Can LIT-1activation occur in the absence of WRM-1, if LIT-1 is made toundergo homotypic interaction? The 31-aa GCN4 leucine-zippercoiled-coil (CC) domain has been used as an artificial multi-merization domain for CC-tagged proteins. Normally, this do-main almost exclusively promotes the formation of dimers.Mutagenesis has been used to generate variant GCN4 CCdomains that specifically form trimers or tetramers (40, 41). Wewill refer to the GCN4 CC domains that specifically di- tri-, ortetramerize as “CC2,” “CC3,” and “CC4”motifs, respectively (Fig.4B). We fused the three variant GCN4 CCmotifs separately to theC terminus of LIT-1 and expressed LIT-1::CC2, LIT-1::CC3, orLIT-1::CC4 individually in HeLa cells. By Western blot we coulddetect T220 phosphorylation with LIT-1::CC3 and LIT-1::CC4,albeit at a low level (Fig. 4B). Cotransfection of WRM-1 resultedin an increase of T220 phosphorylation for all three LIT-1::CCvariants. The finding that WRM-1 is dispensable in promotingT220 phosphorylation as long as LIT-1 can form homotypicinteractions via the GCN4 CC domains supports the model inwhich WRM-1 activates LIT-1, at least in part, by facilitatinghomotypic interaction of LIT-1.
Multiple Domains of WRM-1 Are Required for Stable LIT-1 Interaction.To elucidate the mechanism by which WRM-1 promotes LIT-1homotypic interaction, we mapped the domains of WRM-1 thatform homotypic interaction and that interact with LIT-1. Likevertebrate β-catenin, WRM-1 consists primarily of 12 Armrepeats, flanked by a 150-aa N-terminal domain and an approxi-mately 30-aa C-terminal domain (Fig. 4C) (23). The 148 N-terminalamino acids of WRM-1 have been shown to be both necessary andsufficient to bind LIT-1 in a yeast two-hybrid assay (23). In a pull-down assay using Myc-tagged WRM-1 and GFP-tagged LIT-1,we found that both the 148 N-terminal amino acids and the 145C-terminal amino acids of WRM-1 are required for stable in-teraction with LIT-1 (Fig. 4D). Despite not forming a stableinteraction with LIT-1, both WRM-1(149–796) and WRM-1(1–650) are sufficient to promote a low level of LIT-1 T220 andPOP-1 S-117 phosphorylation in tissue-culture cells, suggestingthat both WRM-1 truncations can form functional, albeittransient, interactions with LIT-1 in cells (Fig. 4D). However,deleting a larger fragment from the C terminus [WRM-1(1–552)] results in undetectable LIT-1 T220 and POP-1 S117phosphorylation.The 148 N-terminal amino acids of WRM-1 also are required for
WRM-1 homotypic interaction. Flag-tagged full-length WRM-1can pull down Myc-tagged full-length WRM-1 (Fig. 4E). Deleting148 amino acids from the N terminus of WRM-1 results in nohomotypic interaction in the pull down (Fig. 4E). The first 410amino acids of WRM-1 are sufficient for homotypic interaction inthe pull-down assay.
Artificial Multimerization Motifs Can Substitute for the Wrm-1C-Terminal Domain. The C terminus of WRM-1 contains putativeCC domains (amino acids 651–682 and 751–780) (COILS: www.ch.embnet.org/software/COILS_form.html, accessed November 2011).Although the WRM-1 CC domains are not required for theWRM-1 homotypic interactions detected in a pull-down assay(Fig. 4E), they are required for stable WRM-1/LIT-1 interaction
B W
TD17
6N
–Flag::MOM-4 WT
D176N
–HIS::LIT-1
MBP::WRM-1 –+++–– +++
T220-P
LIT-1
+++
A
T220-P
LIT-1
mom
-4
wildtyp
e
(ne1
539)
C
Flag::MOM-4
GFP::LIT-1
Myc::WRM-1
– +––+–
++–+
––
K89MWT
T220-P
LIT-1
IP GFP
Fig. 3. Western blot analyses to measure LIT-1 T220 phosphorylation using the anti–LIT-1 and anti–T220-P antibodies. (A) Western blots of C. elegans embryoextracts derived from wild-type or ts mom-4(ne1539) worms raised at 25 °C using the indicated antibodies. (B) MOM-4 phosphorylates bacterially expressedLIT-1 at T220. Wild-type or kinase-dead (D176N) MOM-4/TAP-1 complex was prepared from HEK293 cells in a kinase assay using bacterially expressed andpurified LIT-1 and WRM-1. (C) Coexpression of either WRM-1 or MOM-4/TAP-1 promotes LIT-1 T220 phosphorylation. K89M is a kinase-dead version of LIT-1.
Yang et al. PNAS | Published online December 29, 2014 | E141
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
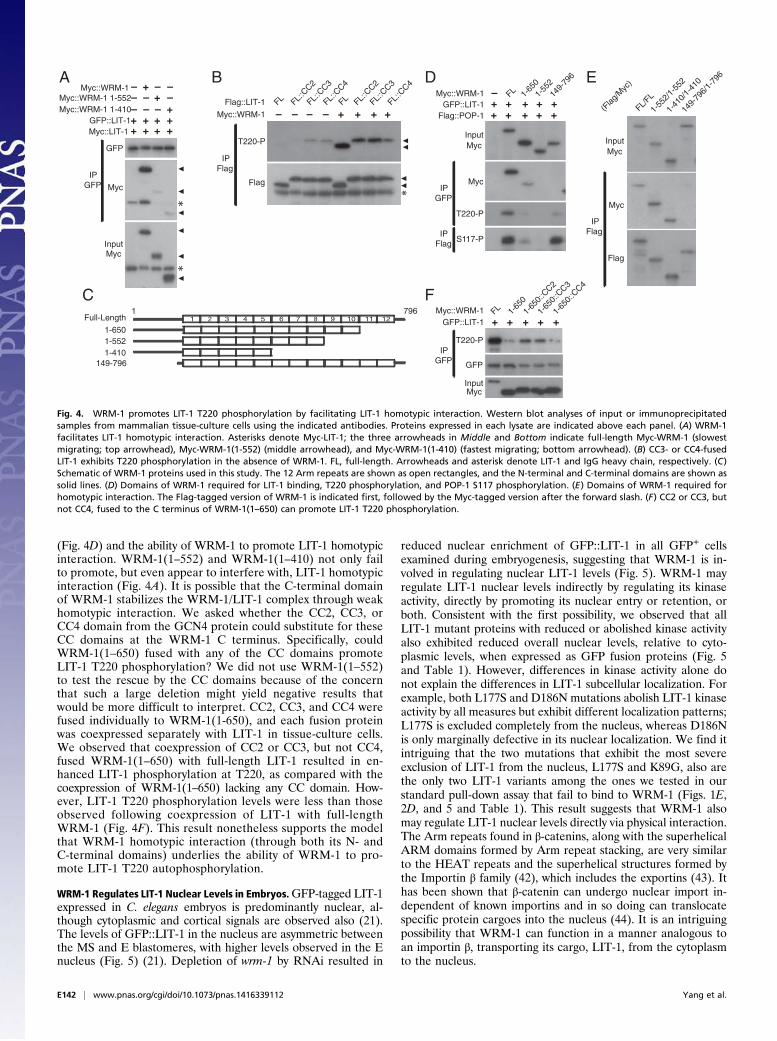
(Fig. 4D) and the ability of WRM-1 to promote LIT-1 homotypicinteraction. WRM-1(1–552) and WRM-1(1–410) not only failto promote, but even appear to interfere with, LIT-1 homotypicinteraction (Fig. 4A). It is possible that the C-terminal domainof WRM-1 stabilizes the WRM-1/LIT-1 complex through weakhomotypic interaction. We asked whether the CC2, CC3, orCC4 domain from the GCN4 protein could substitute for theseCC domains at the WRM-1 C terminus. Specifically, couldWRM-1(1–650) fused with any of the CC domains promoteLIT-1 T220 phosphorylation? We did not use WRM-1(1–552)to test the rescue by the CC domains because of the concernthat such a large deletion might yield negative results thatwould be more difficult to interpret. CC2, CC3, and CC4 werefused individually to WRM-1(1-650), and each fusion proteinwas coexpressed separately with LIT-1 in tissue-culture cells.We observed that coexpression of CC2 or CC3, but not CC4,fused WRM-1(1–650) with full-length LIT-1 resulted in en-hanced LIT-1 phosphorylation at T220, as compared with thecoexpression of WRM-1(1–650) lacking any CC domain. How-ever, LIT-1 T220 phosphorylation levels were less than thoseobserved following coexpression of LIT-1 with full-lengthWRM-1 (Fig. 4F). This result nonetheless supports the modelthat WRM-1 homotypic interaction (through both its N- andC-terminal domains) underlies the ability of WRM-1 to pro-mote LIT-1 T220 autophosphorylation.
WRM-1 Regulates LIT-1 Nuclear Levels in Embryos.GFP-tagged LIT-1expressed in C. elegans embryos is predominantly nuclear, al-though cytoplasmic and cortical signals are observed also (21).The levels of GFP::LIT-1 in the nucleus are asymmetric betweenthe MS and E blastomeres, with higher levels observed in the Enucleus (Fig. 5) (21). Depletion of wrm-1 by RNAi resulted in
reduced nuclear enrichment of GFP::LIT-1 in all GFP+ cellsexamined during embryogenesis, suggesting that WRM-1 is in-volved in regulating nuclear LIT-1 levels (Fig. 5). WRM-1 mayregulate LIT-1 nuclear levels indirectly by regulating its kinaseactivity, directly by promoting its nuclear entry or retention, orboth. Consistent with the first possibility, we observed that allLIT-1 mutant proteins with reduced or abolished kinase activityalso exhibited reduced overall nuclear levels, relative to cyto-plasmic levels, when expressed as GFP fusion proteins (Fig. 5and Table 1). However, differences in kinase activity alone donot explain the differences in LIT-1 subcellular localization. Forexample, both L177S and D186N mutations abolish LIT-1 kinaseactivity by all measures but exhibit different localization patterns;L177S is excluded completely from the nucleus, whereas D186Nis only marginally defective in its nuclear localization. We find itintriguing that the two mutations that exhibit the most severeexclusion of LIT-1 from the nucleus, L177S and K89G, also arethe only two LIT-1 variants among the ones we tested in ourstandard pull-down assay that fail to bind to WRM-1 (Figs. 1E,2D, and 5 and Table 1). This result suggests that WRM-1 alsomay regulate LIT-1 nuclear levels directly via physical interaction.The Arm repeats found in β-catenins, along with the superhelicalARM domains formed by Arm repeat stacking, are very similarto the HEAT repeats and the superhelical structures formed bythe Importin β family (42), which includes the exportins (43). Ithas been shown that β-catenin can undergo nuclear import in-dependent of known importins and in so doing can translocatespecific protein cargoes into the nucleus (44). It is an intriguingpossibility that WRM-1 can function in a manner analogous toan importin β, transporting its cargo, LIT-1, from the cytoplasmto the nucleus.
D
+++GFP::LIT-1 1-
6501-
552
Myc::WRM-1 FL–+
149-
796
+Flag::POP-1 ++++ +
Myc
T220-P
Input
Myc
S117-P IP Flag
IPGFP
7961
1-6501-552
Full-Length
149-796
C
1-410
87 9 10 11 1221 3 4 5 6
MycInput
Myc
IPFlag
Flag
1-55
2/1-
552
149-
796/
1-79
6
(Flag
/Myc
)
1-41
0/1-
410
FL/FL
EB
Flag
Flag::LIT-1 FL::C
C2
FL::C
C3
FL::C
C4
FL FL::C
C2
FL::C
C3
FL::C
C4
FL
*
Myc::WRM-1 – – – – + + + +
T220-P
IPFlag
1-65
0FLMyc::WRM-1 1-
650:
:CC2
1-65
0::C
C3
1-65
0::C
C4
InputMyc
GFP
GFP::LIT-1 + + + ++
T220-P IPGFP
F
A
Myc::LIT-1
–++
GFP::LIT-1+ + ++ + +
Myc::WRM-1Myc::WRM-1 1-552– –
–
Myc
GFP
MycInput
IPGFP
*
*
Myc::WRM-1 1-410– –
–
++
+–
–
Fig. 4. WRM-1 promotes LIT-1 T220 phosphorylation by facilitating LIT-1 homotypic interaction. Western blot analyses of input or immunoprecipitatedsamples from mammalian tissue-culture cells using the indicated antibodies. Proteins expressed in each lysate are indicated above each panel. (A) WRM-1facilitates LIT-1 homotypic interaction. Asterisks denote Myc-LIT-1; the three arrowheads in Middle and Bottom indicate full-length Myc-WRM-1 (slowestmigrating; top arrowhead), Myc-WRM-1(1-552) (middle arrowhead), and Myc-WRM-1(1-410) (fastest migrating; bottom arrowhead). (B) CC3- or CC4-fusedLIT-1 exhibits T220 phosphorylation in the absence of WRM-1. FL, full-length. Arrowheads and asterisk denote LIT-1 and IgG heavy chain, respectively. (C)Schematic of WRM-1 proteins used in this study. The 12 Arm repeats are shown as open rectangles, and the N-terminal and C-terminal domains are shown assolid lines. (D) Domains of WRM-1 required for LIT-1 binding, T220 phosphorylation, and POP-1 S117 phosphorylation. (E) Domains of WRM-1 required forhomotypic interaction. The Flag-tagged version of WRM-1 is indicated first, followed by the Myc-tagged version after the forward slash. (F) CC2 or CC3, butnot CC4, fused to the C terminus of WRM-1(1–650) can promote LIT-1 T220 phosphorylation.
E142 | www.pnas.org/cgi/doi/10.1073/pnas.1416339112 Yang et al.
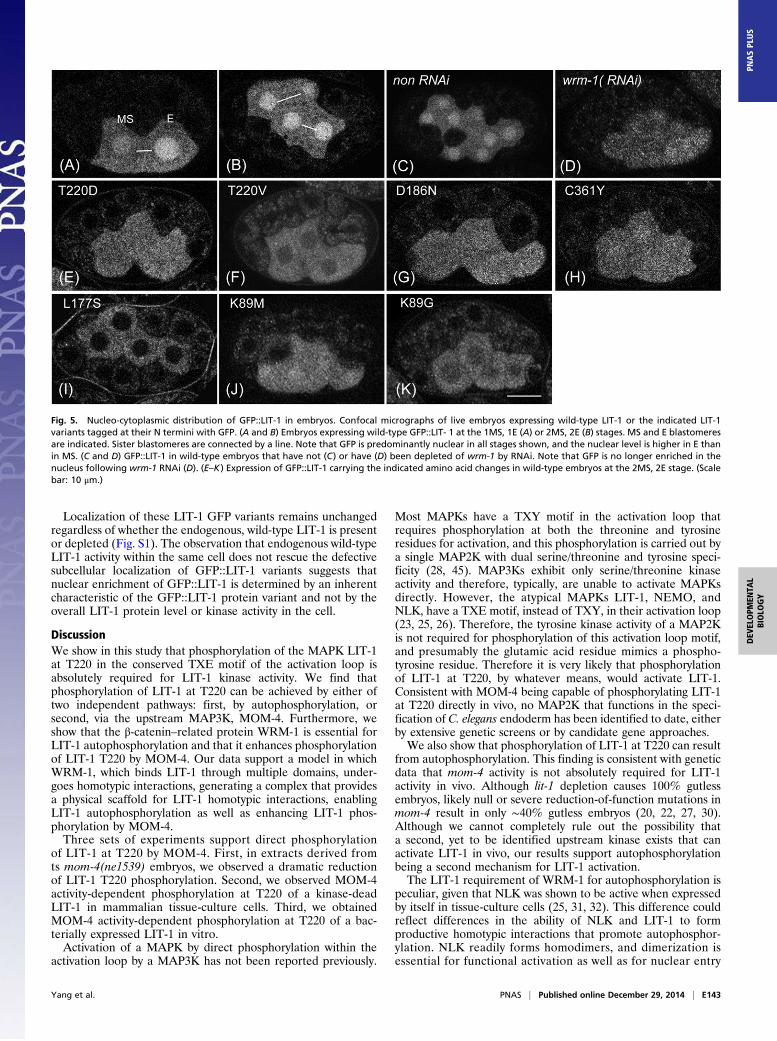
Localization of these LIT-1 GFP variants remains unchangedregardless of whether the endogenous, wild-type LIT-1 is presentor depleted (Fig. S1). The observation that endogenous wild-typeLIT-1 activity within the same cell does not rescue the defectivesubcellular localization of GFP::LIT-1 variants suggests thatnuclear enrichment of GFP::LIT-1 is determined by an inherentcharacteristic of the GFP::LIT-1 protein variant and not by theoverall LIT-1 protein level or kinase activity in the cell.
DiscussionWe show in this study that phosphorylation of the MAPK LIT-1at T220 in the conserved TXE motif of the activation loop isabsolutely required for LIT-1 kinase activity. We find thatphosphorylation of LIT-1 at T220 can be achieved by either oftwo independent pathways: first, by autophosphorylation, orsecond, via the upstream MAP3K, MOM-4. Furthermore, weshow that the β-catenin–related protein WRM-1 is essential forLIT-1 autophosphorylation and that it enhances phosphorylationof LIT-1 T220 by MOM-4. Our data support a model in whichWRM-1, which binds LIT-1 through multiple domains, under-goes homotypic interactions, generating a complex that providesa physical scaffold for LIT-1 homotypic interactions, enablingLIT-1 autophosphorylation as well as enhancing LIT-1 phos-phorylation by MOM-4.Three sets of experiments support direct phosphorylation
of LIT-1 at T220 by MOM-4. First, in extracts derived fromts mom-4(ne1539) embryos, we observed a dramatic reductionof LIT-1 T220 phosphorylation. Second, we observed MOM-4activity-dependent phosphorylation at T220 of a kinase-deadLIT-1 in mammalian tissue-culture cells. Third, we obtainedMOM-4 activity-dependent phosphorylation at T220 of a bac-terially expressed LIT-1 in vitro.Activation of a MAPK by direct phosphorylation within the
activation loop by a MAP3K has not been reported previously.
Most MAPKs have a TXY motif in the activation loop thatrequires phosphorylation at both the threonine and tyrosineresidues for activation, and this phosphorylation is carried out bya single MAP2K with dual serine/threonine and tyrosine speci-ficity (28, 45). MAP3Ks exhibit only serine/threonine kinaseactivity and therefore, typically, are unable to activate MAPKsdirectly. However, the atypical MAPKs LIT-1, NEMO, andNLK, have a TXE motif, instead of TXY, in their activation loop(23, 25, 26). Therefore, the tyrosine kinase activity of a MAP2Kis not required for phosphorylation of this activation loop motif,and presumably the glutamic acid residue mimics a phospho-tyrosine residue. Therefore it is very likely that phosphorylationof LIT-1 at T220, by whatever means, would activate LIT-1.Consistent with MOM-4 being capable of phosphorylating LIT-1at T220 directly in vivo, no MAP2K that functions in the speci-fication of C. elegans endoderm has been identified to date, eitherby extensive genetic screens or by candidate gene approaches.We also show that phosphorylation of LIT-1 at T220 can result
from autophosphorylation. This finding is consistent with geneticdata that mom-4 activity is not absolutely required for LIT-1activity in vivo. Although lit-1 depletion causes 100% gutlessembryos, likely null or severe reduction-of-function mutations inmom-4 result in only ∼40% gutless embryos (20, 22, 27, 30).Although we cannot completely rule out the possibility thata second, yet to be identified upstream kinase exists that canactivate LIT-1 in vivo, our results support autophosphorylationbeing a second mechanism for LIT-1 activation.The LIT-1 requirement of WRM-1 for autophosphorylation is
peculiar, given that NLK was shown to be active when expressedby itself in tissue-culture cells (25, 31, 32). This difference couldreflect differences in the ability of NLK and LIT-1 to formproductive homotypic interactions that promote autophosphor-ylation. NLK readily forms homodimers, and dimerization isessential for functional activation as well as for nuclear entry
Fig. 5. Nucleo-cytoplasmic distribution of GFP::LIT-1 in embryos. Confocal micrographs of live embryos expressing wild-type LIT-1 or the indicated LIT-1variants tagged at their N termini with GFP. (A and B) Embryos expressing wild-type GFP::LIT- 1 at the 1MS, 1E (A) or 2MS, 2E (B) stages. MS and E blastomeresare indicated. Sister blastomeres are connected by a line. Note that GFP is predominantly nuclear in all stages shown, and the nuclear level is higher in E thanin MS. (C and D) GFP::LIT-1 in wild-type embryos that have not (C) or have (D) been depleted of wrm-1 by RNAi. Note that GFP is no longer enriched in thenucleus following wrm-1 RNAi (D). (E–K) Expression of GFP::LIT-1 carrying the indicated amino acid changes in wild-type embryos at the 2MS, 2E stage. (Scalebar: 10 μm.)
Yang et al. PNAS | Published online December 29, 2014 | E143
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US
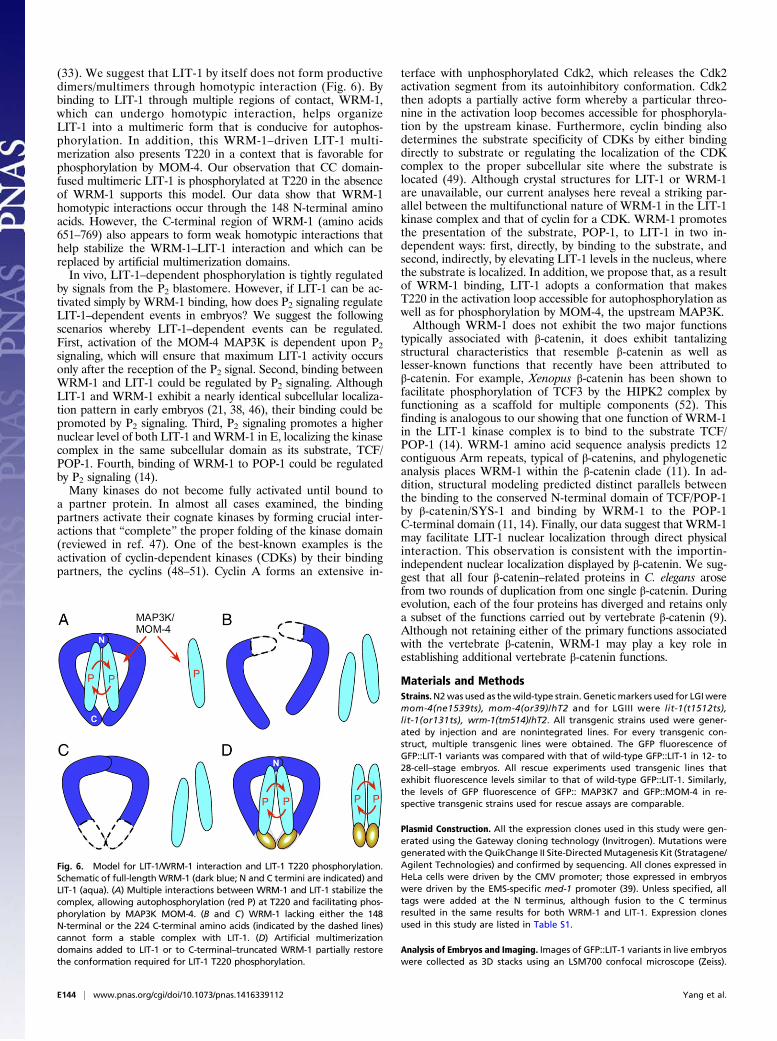
(33). We suggest that LIT-1 by itself does not form productivedimers/multimers through homotypic interaction (Fig. 6). Bybinding to LIT-1 through multiple regions of contact, WRM-1,which can undergo homotypic interaction, helps organizeLIT-1 into a multimeric form that is conducive for autophos-phorylation. In addition, this WRM-1–driven LIT-1 multi-merization also presents T220 in a context that is favorable forphosphorylation by MOM-4. Our observation that CC domain-fused multimeric LIT-1 is phosphorylated at T220 in the absenceof WRM-1 supports this model. Our data show that WRM-1homotypic interactions occur through the 148 N-terminal aminoacids. However, the C-terminal region of WRM-1 (amino acids651–769) also appears to form weak homotypic interactions thathelp stabilize the WRM-1–LIT-1 interaction and which can bereplaced by artificial multimerization domains.In vivo, LIT-1–dependent phosphorylation is tightly regulated
by signals from the P2 blastomere. However, if LIT-1 can be ac-tivated simply by WRM-1 binding, how does P2 signaling regulateLIT-1–dependent events in embryos? We suggest the followingscenarios whereby LIT-1–dependent events can be regulated.First, activation of the MOM-4 MAP3K is dependent upon P2signaling, which will ensure that maximum LIT-1 activity occursonly after the reception of the P2 signal. Second, binding betweenWRM-1 and LIT-1 could be regulated by P2 signaling. AlthoughLIT-1 and WRM-1 exhibit a nearly identical subcellular localiza-tion pattern in early embryos (21, 38, 46), their binding could bepromoted by P2 signaling. Third, P2 signaling promotes a highernuclear level of both LIT-1 andWRM-1 in E, localizing the kinasecomplex in the same subcellular domain as its substrate, TCF/POP-1. Fourth, binding of WRM-1 to POP-1 could be regulatedby P2 signaling (14).Many kinases do not become fully activated until bound to
a partner protein. In almost all cases examined, the bindingpartners activate their cognate kinases by forming crucial inter-actions that “complete” the proper folding of the kinase domain(reviewed in ref. 47). One of the best-known examples is theactivation of cyclin-dependent kinases (CDKs) by their bindingpartners, the cyclins (48–51). Cyclin A forms an extensive in-
terface with unphosphorylated Cdk2, which releases the Cdk2activation segment from its autoinhibitory conformation. Cdk2then adopts a partially active form whereby a particular threo-nine in the activation loop becomes accessible for phosphoryla-tion by the upstream kinase. Furthermore, cyclin binding alsodetermines the substrate specificity of CDKs by either bindingdirectly to substrate or regulating the localization of the CDKcomplex to the proper subcellular site where the substrate islocated (49). Although crystal structures for LIT-1 or WRM-1are unavailable, our current analyses here reveal a striking par-allel between the multifunctional nature of WRM-1 in the LIT-1kinase complex and that of cyclin for a CDK. WRM-1 promotesthe presentation of the substrate, POP-1, to LIT-1 in two in-dependent ways: first, directly, by binding to the substrate, andsecond, indirectly, by elevating LIT-1 levels in the nucleus, wherethe substrate is localized. In addition, we propose that, as a resultof WRM-1 binding, LIT-1 adopts a conformation that makesT220 in the activation loop accessible for autophosphorylation aswell as for phosphorylation by MOM-4, the upstream MAP3K.Although WRM-1 does not exhibit the two major functions
typically associated with β-catenin, it does exhibit tantalizingstructural characteristics that resemble β-catenin as well aslesser-known functions that recently have been attributed toβ-catenin. For example, Xenopus β-catenin has been shown tofacilitate phosphorylation of TCF3 by the HIPK2 complex byfunctioning as a scaffold for multiple components (52). Thisfinding is analogous to our showing that one function of WRM-1in the LIT-1 kinase complex is to bind to the substrate TCF/POP-1 (14). WRM-1 amino acid sequence analysis predicts 12contiguous Arm repeats, typical of β-catenins, and phylogeneticanalysis places WRM-1 within the β-catenin clade (11). In ad-dition, structural modeling predicted distinct parallels betweenthe binding to the conserved N-terminal domain of TCF/POP-1by β-catenin/SYS-1 and binding by WRM-1 to the POP-1C-terminal domain (11, 14). Finally, our data suggest that WRM-1may facilitate LIT-1 nuclear localization through direct physicalinteraction. This observation is consistent with the importin-independent nuclear localization displayed by β-catenin. We sug-gest that all four β-catenin–related proteins in C. elegans arosefrom two rounds of duplication from one single β-catenin. Duringevolution, each of the four proteins has diverged and retains onlya subset of the functions carried out by vertebrate β-catenin (9).Although not retaining either of the primary functions associatedwith the vertebrate β-catenin, WRM-1 may play a key role inestablishing additional vertebrate β-catenin functions.
Materials and MethodsStrains.N2was used as thewild-type strain. Geneticmarkers used for LGIweremom-4(ne1539ts), mom-4(or39)/hT2 and for LGIII were lit-1(t1512ts),lit-1(or131ts), wrm-1(tm514)/hT2. All transgenic strains used were gener-ated by injection and are nonintegrated lines. For every transgenic con-struct, multiple transgenic lines were obtained. The GFP fluorescence ofGFP::LIT-1 variants was compared with that of wild-type GFP::LIT-1 in 12- to28-cell–stage embryos. All rescue experiments used transgenic lines thatexhibit fluorescence levels similar to that of wild-type GFP::LIT-1. Similarly,the levels of GFP fluorescence of GFP:: MAP3K7 and GFP::MOM-4 in re-spective transgenic strains used for rescue assays are comparable.
Plasmid Construction. All the expression clones used in this study were gen-erated using the Gateway cloning technology (Invitrogen). Mutations weregenerated with the QuikChange II Site-DirectedMutagenesis Kit (Stratagene/Agilent Technologies) and confirmed by sequencing. All clones expressed inHeLa cells were driven by the CMV promoter; those expressed in embryoswere driven by the EMS-specific med-1 promoter (39). Unless specified, alltags were added at the N terminus, although fusion to the C terminusresulted in the same results for both WRM-1 and LIT-1. Expression clonesused in this study are listed in Table S1.
Analysis of Embryos and Imaging. Images of GFP::LIT-1 variants in live embryoswere collected as 3D stacks using an LSM700 confocal microscope (Zeiss).
Fig. 6. Model for LIT-1/WRM-1 interaction and LIT-1 T220 phosphorylation.Schematic of full-lengthWRM-1 (dark blue; N and C termini are indicated) andLIT-1 (aqua). (A) Multiple interactions between WRM-1 and LIT-1 stabilize thecomplex, allowing autophosphorylation (red P) at T220 and facilitating phos-phorylation by MAP3K MOM-4. (B and C) WRM-1 lacking either the 148N-terminal or the 224 C-terminal amino acids (indicated by the dashed lines)cannot form a stable complex with LIT-1. (D) Artificial multimerizationdomains added to LIT-1 or to C-terminal–truncated WRM-1 partially restorethe conformation required for LIT-1 T220 phosphorylation.
E144 | www.pnas.org/cgi/doi/10.1073/pnas.1416339112 Yang et al.

Assay for rescue of the E defect in lit-1(t1512) and mom-4(or39) mutantembryos was performed as described (21), with modifications, using anAxioplan microscope (Zeiss) equipped with epifluorescence, polarizing, anddifferential interference contrast (DIC) optics and a MicroMax-512EBFT CCDcamera (Princeton Instruments) controlled by MetaMorph acquisition soft-ware (Molecular Devices) (15, 19). Production of MS-derived pharyngealtissues was scored by DIC optics. Formation of intestine was assayed withboth DIC and polarizing optics.
Antibody Generation. Anti–T220-P, raised against the peptide QRDRLNMT[p]HEVVTQYY, and anti–LIT-1, raised against a bacterially expressed proteincorresponding to amino acids 335–454 of LIT-1e, were generated in rabbitsat Genemed Synthesis, Inc. and Proteintech Group, respectively, and purifiedaccording to the company’s protocol. Anti–T220-P antiserum was subjectedto dual selection at Genemed Synthesis, first for binding to the phospho-peptide and then for lack of binding to the nonphosphorylated peptide.
Lysate Preparation, Immunoprecipitation, and Western Blots. Embryos werecollected from gravid adults, and lysates were prepared as described (14).Transfections were performed into mammalian tissue-culture cells, andlysates were prepared as described (14). All transfections were performedusing HeLa cells, except (i) transfection of MOM-4, TAP-1, MAP3K7, or TAB1into HEK293 cells, and (ii) transfection of wild-type and mutant LIT-1 intowild-type and MAP3K7-KO MEF cells (a gift from Zhijian J. Chen, Universityof Texas Southwestern Medical Center, Dallas).
Antibodies used for immunoprecipitations were camelid anti-GFP anti-body (GFP-Trap; ChromoTek) and anti-FLAG agarose beads (M2; Sigma).Antibodies used in Western blots include anti–S117-P at 1:500 (21), anti–LIT-1(this study) at 1:2,000, anti-T220P (this study) at 1:2,000, anti–c-Myc (9E10;Santa Cruz Biotechnology) at 1:2,000, and anti-FLAG (M2; Sigma) at 1:2,000.Secondary antibodies used were donkey anti-rabbit IgG-HRP (GE Healthcare)and goat anti-mouse IgG1-HRP (Santa Cruz Biotechnology), both at 1:20,000.
Kinase Assays. In vitro kinase assays were performed as described (14). GFP-LIT-1 variants were expressed alone or together with WRM-1 in HeLa cellsand were immunoprecipitated with GFP-Trap beads. Immunoprecipitateswere washed three times with wash buffer (50 mM Hepes, 250 mM NaCl,
0.1% Nonidet P-40), rinsed twice with PBS, rinsed one time with kinasebuffer minus ATP, followed by incubation in 20 μL kinase buffer [20 mMHepes (pH 7.4), 5 mM MgCl2, 1 mM DTT, 2 mM ATP] at 30 °C for 60 min. ForMOM-4 phosphorylation of bacterially purified LIT-1, Flag-tagged wild-typeMOM-4 and kinase-dead mutant MOM-4 D176N were expressed separatelyin HEK293 cells, immunoprecipitated with anti-Flag M2 beads, washed asdescribed above, and incubated with 1 μg bacterially purified LIT-1 in 20 μLkinase buffer [20 mM Tris·HCl (pH 7.5), 5 mM MgCl2, 1 mM DTT, 2 mM ATP]at 30 °C for 30 min. After incubation, reactions were stopped with SDS gel-loading buffer, and phosphorylation levels were assayed by Western blotusing anti–T220-P antibody. Kinase assays were performed initially at 20 °C,25 °C, and 30 °C, with no qualitative differences noted between the threetemperatures. Kinase assays were performed at 30 °C because reactionswere more robust at this temperature.
LIT-1 expressed in E. coli was found to be inactive in all our assays, even ifWRM-1 was coexpressed and copurified with LIT-1. In addition, for reasonsthat remain unclear, phosphatase treatment of the LIT-1/WRM-1 complexpulled down from mammalian cells resulted in dissociation of WRM-1 fromLIT-1 and therefore in the loss of LIT-1 activity. Therefore autophosphor-ylation (Fig. 2A) was assayed using LIT-1 pulled down from tissue-culturelysates for LIT-1 kinase activity-dependent incorporation of ATP at T220.
Recombinant Proteins. His-tagged full-length LIT-1 was expressed alone ortogether with MBP-tagged full-length WRM-1 in E. coli at 16 °C and waspurified using TALON metal affinity resin (BD Biosciences) according to themanufacturer’s protocol. The purity of the eluted proteins was evaluated onSDS/PAGE gels stained with Coomassie Brilliant Blue, and the coimmuno-precipitation of WRM-1 along with LIT-1 was confirmed by Western blot.
ACKNOWLEDGMENTS. We thank members of the R.L. laboratory and Drs.Hongtao Yu and Elizabeth Goldsmith for discussions, Jin Jiang for GCN4 CCmotif plasmids, Zhijian (James) Chen for MAP3K7-KO MEFs and pJC0208,Lijun Sun for technical assistance, and the C. elegans Genome Center(CGC), supported by National Institutes of Health (NIH) Grant P40OD010440, for strains. This work was supported by NIH Grants HD037933and GM084198 (to R.L.).
1. McCrea PD, Gu D (2010) The catenin family at a glance. J Cell Sci 123(Pt 5):637–642.2. Zhao ZM, Reynolds AB, Gaucher EA (2011) The evolutionary history of the catenin
gene family during metazoan evolution. BMC Evol Biol 11:198.3. Shapiro L, Weis WI (2009) Structure and biochemistry of cadherins and catenins. Cold
Spring Harb Perspect Biol 1(3):a003053.4. Xu W, Kimelman D (2007) Mechanistic insights from structural studies of beta-catenin
and its binding partners. J Cell Sci 120(Pt 19):3337–3344.5. Chai G, et al. (2010) Complete functional segregation of planarian beta-catenin-1 and
-2 in mediating Wnt signaling and cell adhesion. J Biol Chem 285(31):24120–24130.6. Costa M, et al. (1998) A putative catenin-cadherin system mediates morphogenesis of
the Caenorhabditis elegans embryo. J Cell Biol 141(1):297–308.7. Eisenmann DM, Maloof JN, Simske JS, Kenyon C, Kim SK (1998) The beta-catenin
homolog BAR-1 and LET-60 Ras coordinately regulate the Hox gene lin-39 duringCaenorhabditis elegans vulval development. Development 125(18):3667–3680.
8. Kidd AR, 3rd, Miskowski JA, Siegfried KR, Sawa H, Kimble J (2005) A beta-cateninidentified by functional rather than sequence criteria and its role in Wnt/MAPK sig-naling. Cell 121(5):761–772.
9. Robertson SM, Lin R (2012) Our evolving view of Wnt signaling in C. elegans: If two’scompany and three’s a crowd, is four really necessary? Worm 1(1):82–89.
10. Rocheleau CE, et al. (1997) Wnt signaling and an APC-related gene specify endodermin early C. elegans embryos. Cell 90(4):707–716.
11. Liu J, Phillips BT, Amaya MF, Kimble J, Xu W (2008) The C. elegans SYS-1 protein isa bona fide beta-catenin. Dev Cell 14(5):751–761.
12. Korswagen HC, Herman MA, Clevers HC (2000) Distinct beta-catenins mediate ad-hesion and signalling functions in C. elegans. Nature 406(6795):527–532.
13. Natarajan L, Witwer NE, Eisenmann DM (2001) The divergent Caenorhabditis elegansbeta-catenin proteins BAR-1, WRM-1 and HMP-2 make distinct protein interactionsbut retain functional redundancy in vivo. Genetics 159(1):159–172.
14. Yang XD, et al. (2011) Distinct and mutually inhibitory binding by two divergentβ-catenins coordinates TCF levels and activity in C. elegans. Development 138(19):4255–4265.
15. Huang S, Shetty P, Robertson SM, Lin R (2007) Binary cell fate specification duringC. elegans embryogenesis driven by reiterated reciprocal asymmetry of TCF POP-1 andits coactivator beta-catenin SYS-1. Development 134(14):2685–2695.
16. Lin R, Hill RJ, Priess JR (1998) POP-1 and anterior-posterior fate decisions in C. elegansembryos. Cell 92(2):229–239.
17. Phillips BT, Kidd AR, 3rd, King R, Hardin J, Kimble J (2007) Reciprocal asymmetry ofSYS-1/beta-catenin and POP-1/TCF controls asymmetric divisions in Caenorhabditiselegans. Proc Natl Acad Sci USA 104(9):3231–3236.
18. Goldstein B (1992) Induction of gut in Caenorhabditis elegans embryos. Nature357(6375):255–257.
19. Shetty P, Lo MC, Robertson SM, Lin R (2005) C. elegans TCF protein, POP-1, convertsfrom repressor to activator as a result of Wnt-induced lowering of nuclear levels. DevBiol 285(2):584–592.
20. Thorpe CJ, Schlesinger A, Carter JC, Bowerman B (1997) Wnt signaling polarizes anearly C. elegans blastomere to distinguish endoderm from mesoderm. Cell 90(4):695–705.
21. Lo MC, Gay F, Odom R, Shi Y, Lin R (2004) Phosphorylation by the beta-catenin/MAPKcomplex promotes 14-3-3-mediated nuclear export of TCF/POP-1 in signal-responsivecells in C. elegans. Cell 117(1):95–106.
22. Kaletta T, Schnabel H, Schnabel R (1997) Binary specification of the embryonic lineagein Caenorhabditis elegans. Nature 390(6657):294–298.
23. Rocheleau CE, et al. (1999) WRM-1 activates the LIT-1 protein kinase to transduceanterior/posterior polarity signals in C. elegans. Cell 97(6):717–726.
24. LaBonty M, et al. (2014) CACN-1/Cactin plays a role in Wnt signaling in C. elegans.PLoS ONE 9(7):e101945.
25. Brott BK, Pinsky BA, Erikson RL (1998) Nlk is a murine protein kinase related to Erk/MAP kinases and localized in the nucleus. Proc Natl Acad Sci USA 95(3):963–968.
26. Choi KW, Benzer S (1994) Rotation of photoreceptor clusters in the developingDrosophila eye requires the nemo gene. Cell 78(1):125–136.
27. Meneghini MD, et al. (1999) MAP kinase and Wnt pathways converge to down-regulate an HMG-domain repressor in Caenorhabditis elegans. Nature 399(6738):793–797.
28. Chen Z, et al. (2001) MAP kinases. Chem Rev 101(8):2449–2476.29. Payne DM, et al. (1991) Identification of the regulatory phosphorylation sites in pp42/
mitogen-activated protein kinase (MAP kinase). EMBO J 10(4):885–892.30. Shin TH, et al. (1999) MOM-4, a MAP kinase kinase kinase-related protein, activates
WRM-1/LIT-1 kinase to transduce anterior/posterior polarity signals in C. elegans. MolCell 4(2):275–280.
31. Ishitani T, et al. (2010) Nemo-like kinase suppresses Notch signalling by interferingwith formation of the Notch active transcriptional complex. Nat Cell Biol 12(3):278–285.
32. Ishitani T, Ishitani S, Matsumoto K, Itoh M (2009) Nemo-like kinase is involved in NGF-induced neurite outgrowth via phosphorylating MAP1B and paxillin. J Neurochem111(5):1104–1118.
33. Ishitani S, Inaba K, Matsumoto K, Ishitani T (2011) Homodimerization of Nemo-likekinase is essential for activation and nuclear localization. Mol Biol Cell 22(2):266–277.
34. Ishitani T, et al. (1999) The TAK1-NLK-MAPK-related pathway antagonizes signallingbetween beta-catenin and transcription factor TCF. Nature 399(6738):798–802.
35. Kanei-Ishii C, et al. (2004) Wnt-1 signal induces phosphorylation and degradation ofc-Myb protein via TAK1, HIPK2, and NLK. Genes Dev 18(7):816–829.
Yang et al. PNAS | Published online December 29, 2014 | E145
DEV
ELOPM
ENTA
LBIOLO
GY
PNASPL
US

36. Ohkawara B, et al. (2004) Role of the TAK1-NLK-STAT3 pathway in TGF-beta-medi-ated mesoderm induction. Genes Dev 18(4):381–386.
37. Smit L, et al. (2004) Wnt activates the Tak1/Nemo-like kinase pathway. J Biol Chem279(17):17232–17240.
38. Nakamura K, et al. (2005) Wnt signaling drives WRM-1/beta-catenin asymmetries inearly C. elegans embryos. Genes Dev 19(15):1749–1754.
39. Maduro MF, Meneghini MD, Bowerman B, Broitman-Maduro G, Rothman JH (2001)Restriction of mesendoderm to a single blastomere by the combined action of SKN-1and a GSK-3beta homolog is mediated by MED-1 and -2 in C. elegans. Mol Cell 7(3):475–485.
40. Harbury PB, Kim PS, Alber T (1994) Crystal structure of an isoleucine-zipper trimer.Nature 371(6492):80–83.
41. Harbury PB, Zhang T, Kim PS, Alber T (1993) A switch between two-, three-, and four-stranded coiled coils in GCN4 leucine zipper mutants. Science 262(5138):1401–1407.
42. Kippert F, Gerloff DL (2009) Highly sensitive detection of individual HEAT and ARMrepeats with HHpred and COACH. PLoS ONE 4(9):e7148.
43. Petosa C, et al. (2004) Architecture of CRM1/Exportin1 suggests how cooperativity isachieved during formation of a nuclear export complex. Mol Cell 16(5):761–775.
44. Asally M, Yoneda Y (2005) Beta-catenin can act as a nuclear import receptor for itspartner transcription factor, lymphocyte enhancer factor-1 (lef-1). Exp Cell Res 308(2):357–363.
45. Krishna M, Narang H (2008) The complexity of mitogen-activated protein kinases(MAPKs) made simple. Cell Mol Life Sci 65(22):3525–3544.
46. Kim S, et al. (2013) Wnt and CDK-1 regulate cortical release of WRM-1/β-catenin tocontrol cell division orientation in early Caenorhabditis elegans embryos. Proc NatlAcad Sci USA 110(10):E918–E927.
47. Bayliss R, Fry A, Haq T, Yeoh S (2012) On the molecular mechanisms of mitotic kinaseactivation. Open Biol 2(11):120136.
48. Morgan DO (1995) Principles of CDK regulation. Nature 374(6518):131–134.49. Morgan DO (1997) Cyclin-dependent kinases: Engines, clocks, and microprocessors.
Annu Rev Cell Dev Biol 13:261–291.50. Morgan DO (2007) The Cell Cycle: Principles of Control (New Science, London), 1st Ed.51. Pavletich NP (1999) Mechanisms of cyclin-dependent kinase regulation: Structures of
Cdks, their cyclin activators, and Cip and INK4 inhibitors. J Mol Biol 287(5):821–828.52. Hikasa H, et al. (2010) Regulation of TCF3 by Wnt-dependent phosphorylation during
vertebrate axis specification. Dev Cell 19(4):521–532.
E146 | www.pnas.org/cgi/doi/10.1073/pnas.1416339112 Yang et al.