
Καρακιουλάκη-εργασία πρακτικής- ΤΕΛΙΚΟ
38
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ Μοριακή διάγνωση μεταλλάξεων του γονιδίου CFTR με RT-PCR και ARMS-PCR Καρακιουλάκη Μερόπη 5785 Επόπτρια Καθηγήτρια: Δροσοπούλου Ελένη Μονάδα Εκτέλεσης: LabNET Εκπαιδευτής Μονάδας: Χατζηδημητρίου Δημήτριος Θεσσαλονίκη, 2016
-
Upload
meropi-karakioulaki -
Category
Documents
-
view
95 -
download
3
Transcript of Καρακιουλάκη-εργασία πρακτικής- ΤΕΛΙΚΟ

ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ
Μοριακή διάγνωση µεταλλάξεων του γονιδίου
CFTR µε RT-PCR και ARMS-PCR
Καρακιουλάκη Μερόπη 5785
Επόπτρια Καθηγήτρια: Δροσοπούλου Ελένη Μονάδα Εκτέλεσης: LabNET Εκπαιδευτής Μονάδας: Χατζηδηµητρίου Δηµήτριος
Θεσσαλονίκη, 2016

2
ΑΡΙΣΤΟΤΕΛΕΙΟ ΠΑΝΕΠΙΣΤΗΜΙΟ ΘΕΣΣΑΛΟΝΙΚΗΣ ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ
ARISTOTLE UNIVERSITY OF THESSALONIKI BIOLOGY DEPARTMENT
Μοριακή διάγνωση µεταλλάξεων του γονιδίου
CFTR µε RT-PCR και ARMS-PCR
Molecular diagnosis of mutations in the CFTR gene by RT-PCR and ARMS -PCR
Πρακτική Άσκηση σε Κλινική Βιοδιαγνωστική Practical training in Clinical Diagnostic Center
Καρακιουλάκη Μερόπη
Κarakioulaki Meropi Επόπτρια Καθηγήτρια: Δροσοπούλου Ελένη University Supervisor: Drosopoulou Eleni Μονάδα Εκτέλεσης: LabNET Training Unit: LabNET Εκπαιδευτής Μονάδας: Χατζηδηµητρίου Δηµήτριος Training Unit Supervisor: Chatzidimitriou Dimitrios

3
ΠΕΡΙΕΧΟΜΕΝΑ
1. ΠΡΟΛΟΓΟΣ............................................................................................................4
2. ΘΕΩΡΗΤΙΚΟ ΜΕΡΟΣ...........................................................................................5
I. H ΚΥΣΤΙΚΗ ΙΝΩΣΗ..............................................................................................5II. Η ΜΕΘΟΔΟΣ ΤΗΣ ΑΛΥΣΙΔΩΤΗΣ ΑΝΤΙΔΡΑΣΗΣ ΠΟΛΥΜΕΡΑΣΗΣ (POLYMERASE CHAIN REACTION – PCR)...........................................................14
III. MULTIPLEX PCR............................................................................................16IV. PCR ΠΡΑΓΜΑΤΙΚΟΥ ΧΡΟΝΟΥ (REAL TIME-POLYMERASE CHAIN REACTION – RT-PCR)...............................................................................................17
3. ΠΡΑΚΤΙΚΟ ΜΕΡΟΣ............................................................................................23
I. ΑΠΟΜΟΝΩΣΗ ΓΕΝΩΜΙΚΟΥ DNA ΑΠΟ ΠΕΡΙΦΕΡΙΚΟ ΑΙΜΑ...................23II. ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΜΕΤΑΛΛΑΞΗΣ F508DEL (ΔF508) ME TH ΒΟΗΘΕΙΑ ΤΗΣ RT-PCR................................................................................................................25
III. ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΑΠΟΤΕΛΕΣΜΑΤΩΝ...................................................27IV. ΑΝΙΧΝΕΥΣΗ 29 ΜΕΤΑΛΛΑΞΕΩΝ ΤΗΣ ΚΥΣΤΙΚΗΣ ΙΝΩΣΗΣ ΜΕ ΤΗ ΒΟΗΘΕΙΑ ΤΗΣ PCR...................................................................................................29
V. ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΑΠΟΤΕΛΕΣΜΑΤΩΝ...................................................32
4. ΕΠΙΛΟΓΟΣ...........................................................................................................34
5. ΒΙΒΛΙΟΓΡΑΦΙΑ...................................................................................................35

4
1. ΠΡΟΛΟΓΟΣ Στα πλαίσια της Πρακτικής Άσκησης που διετέλεσα στο 8ο εξάµηνο σπουδών µου στο Τµήµα
Βιολογίας ΑΠΘ είχα την ευκαιρία να εκπαιδευτώ για 2 µήνες στο εργαστήριο LABnet στη
Θεσσαλονίκη, ένα υπερσύγχρονο διαγνωστικό κέντρο που υποστηρίζει τα περισσότερα
βιοπαθολογικά εργαστήρια της περιοχής, µέσω µιας πληθώρας βιοχηµικών, µοριακών και
µικροβιολογικών εξετάσεων που εκτελεί. Η πρακτική άσκηση µου έδωσε την δυνατότητα να
εξοικειωθώ µε τον τρόπο λειτουργίας και οργάνωσης µιας τέτοιας επιχείρησης, η οποία
καθηµερινά καλείται να εκτελέσει αναλύσεις σε πάνω από 1000 δείγµατα ασθενών.
Αναλυτικότερα, έµαθα να χρησιµοποιώ το σύστηµα “slis” (Laboratory and Radiology
Information System), στο οποίο καταγράφονται όλες οι εντολές των γιατρών που
υποδεικνύουν τις διαγνωστικές εξετάσεις που πρέπει να εκτελεστούν σε κάθε δείγµα (τα
δείγµατα, έτσι, αποκτούν συγκεκριµένα barcodes). Ακόµη µου δόθηκε η δυνατότητα να
συµµετάσχω στη διαδικασία προετοιµασίας των δειγµάτων, µε σκοπό αυτά να µπουν στους
αυτόµατους αναλυτές, οι οποίοι εκτελούν αυτοµατοποιηµένα µια πληθώρα βιοχηµικών και
ανοσολογικών εξετάσεων. Επιπλέον, παρακολούθησα πολλές βιοπαθολογικές αναλύσεις,
όπως:
• διάφορες µοριακές αναλύσεις (ανίχνευση του γονιδίου της θροµβοφιλίας, του
αντιγόνου ιστοσυµβατότητας HLA-B27, της µετάλλαξης F508del και 29 άλλων
µεταλλάξεων της κυστικής ίνωσης, των αλληλοµόρφων των TGFb2 και TGFb3, του
ιού της ηπατίτιδας Β)
• τεχνικές elisa (ανίχνευση της 17 υδροξυ-προγεστερόνης και του κυτταροµεγαλοϊού)
• µεθόδους ανοσοφθορισµού (ανίχνευση αυτοάνοσων νοσηµάτων -λύκος- και listeria)
• εξετάσεις γλυκοζυλιωµένης αιµοσφαιρίνης, ηλεκτροφόρησης αιµοσφαιρίνης
(capilarys flex piercing), πήξης αίµατος, γενικές εξετάσεις αίµατος
• αποµόνωση DNA από περιφερικό αίµα
• ταυτοποίηση µικροβίων (χρώση Gram, καλλιέργεια σε εκλεκτικά υποστρώµατα,
µορφολογία, τεστ οξειδάσης, αντίδραση καταλάσης, ταυτοποίηση εντεροβακτηρίων µε
enteropluri, αντιβιόγραµµα)
• αιµατολογικούς ελέγχους (µέτρηση αιµοπεταλίων, µορφολογία ερυθρών, τεστ
δρεπάνωσης, αντιδράσεις Coombs για ανίχνευση αυτοάνοσης αιµολυτικής αναιµίας,
ανίχνευση δικτυοερυθροκυττάρων σε αίµα)
• σπερµοδιαγράµµατα, αξιολόγηση κινητικότητας σπερµατοζωαρίων

5
• παρασιτολογικούς ελέγχους (κόπρανα και νύχια)
Από όλες αυτές τις αναλύσεις που παρακολούθησα, στην παρούσα εργασία εστίασα στην
περιγραφή της µοριακής διάγνωσης 29 µεταλλάξεων της κυστικής ίνωσης, µε τη βοήθεια της
αλυσιδωτής αντίδρασης πολυµεράσης (PCR).
2. ΘΕΩΡΗΤΙΚΟ ΜΕΡΟΣ
I. H ΚΥΣΤΙΚΗ ΙΝΩΣΗ
H κυστική ίνωση (CF- Cystic Fibrosis) είναι η πιο συχνή αυτοσωµική υπολειπόµενη γενετική
ασθένεια που εµφανίζεται στον Καυκάσιο πληθυσµό (συχνότητα ασθενών 1:2500, συχνότητα
φορέων 1:25)1. Οφείλεται σε µεταλλάξεις που συµβαίνουν στο γονίδιο CFΤR (Cystic Fibrosis
Transmembrane Conductance Regulator – ρυθµιστής διαµεµβρανικής διακίνησης ιόντων)
(ΟΜΙΜ 602421; PubMed 2772657), το οποίο βρίσκεται στον µακρύ βραχίονα του
χρωµοσώµατος 7 (περιοχή 7q31-32). Το γονίδιο αυτό έχει πολύ µεγάλο µέγεθος (230kb) και
περιλαµβάνει 27 εξώνια που οδηγούν στην παραγωγή ενός mRNA µε µέγεθος 6.5kb, το οποίο
κωδικοποιεί την CFTR πρωτεΐνη1–8
Η πρωτεΐνη CFTR αποτελείται από 1480 αµινοξέα και σχηµατίζεται από 2 επαναλαµβανόµενα
τµήµατα, το καθένα από τα οποία περιλαµβάνει µια διαµεµβρανική περιοχή µε 6
διαµεµβρανικά τµήµατα (MSD1, MSD2), και µία περιοχή δεύσµευσης ATP (NBD1, NBD2).
Tα δύο αυτά τµήµατα ενώνονται διαµέσου της ρυθµιστικής περιοχής (R), η οποία διαθέτει
πολλές θέσεις φωσφορυλίωσης9,10 [Εικόνα 1]. H πρωτεΐνη CFTR βρίσκεται στη µεµβράνη των
αναπνευστικών και εκκριτικών επιθηλιακών κυττάρων και παρουσιάζει ποικίλες
δραστηριότητες, η κυριότερη από τις οποίες είναι η λειτουργία της ως κανάλι ιόντων χλωρίου
(κίνηση ιόντων χλωρίου µέσα και έξω από τα κύτταρα) που ενεργοποιείται από την παρουσία
cAMP. Η CFTR µε τη σειρά της ρυθµίζει τη λειτουργία άλλων µεµβρανικών διαύλων ιόντων,
όπως οι δίαυλοι που µεταφέρουν ιόντα νατρίου, και οι οποίοι είναι απαραίτητοι για τη σωστή
λειτουργία πολλών οργάνων, όπως οι πνεύµονες και το πάγκρεας11. Το ΑΤP προσδένεται πάνω
στην πρωτεΐνη και υδρολύεται, γεγονός που παράγει την απαραίτητη ενέργεια για το άνοιγµα
και το κλείσιµο του καναλιού χλωρίου. Η ρύθµιση του καναλιού ελέγχεται από τη
φωσφορυλίωση της ρυθµιστικής περιοχής R10.
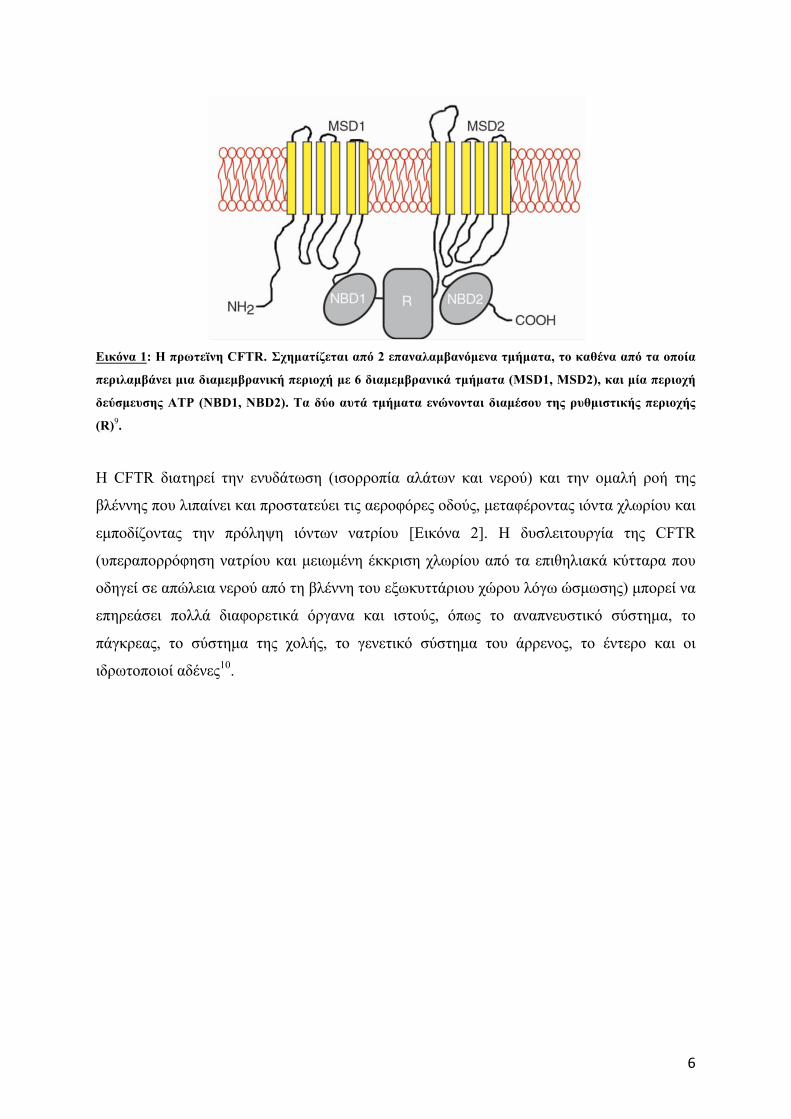
6
Εικόνα 1: Η πρωτεϊνη CFTR. Σχηµατίζεται από 2 επαναλαµβανόµενα τµήµατα, το καθένα από τα οποία
περιλαµβάνει µια διαµεµβρανική περιοχή µε 6 διαµεµβρανικά τµήµατα (MSD1, MSD2), και µία περιοχή
δεύσµευσης ATP (NBD1, NBD2). Tα δύο αυτά τµήµατα ενώνονται διαµέσου της ρυθµιστικής περιοχής
(R)9.
Η CFTR διατηρεί την ενυδάτωση (ισορροπία αλάτων και νερού) και την οµαλή ροή της
βλέννης που λιπαίνει και προστατεύει τις αεροφόρες οδούς, µεταφέροντας ιόντα χλωρίου και
εµποδίζοντας την πρόληψη ιόντων νατρίου [Εικόνα 2]. Η δυσλειτουργία της CFTR
(υπεραπορρόφηση νατρίου και µειωµένη έκκριση χλωρίου από τα επιθηλιακά κύτταρα που
οδηγεί σε απώλεια νερού από τη βλέννη του εξωκυττάριου χώρου λόγω ώσµωσης) µπορεί να
επηρεάσει πολλά διαφορετικά όργανα και ιστούς, όπως το αναπνευστικό σύστηµα, το
πάγκρεας, το σύστηµα της χολής, το γενετικό σύστηµα του άρρενος, το έντερο και οι
ιδρωτοποιοί αδένες10.

7
Εικόνα 2: H λειτουργία της CFTR πρωτεΐνης στους αεραγωγούς (Lung airways). Αριστερά απεικονίζεται
ο αεραγωγός ενός υγιούς ατόµου (Lung airway of unaffected person). Τα κανάλια CFTR λειτουργούν οµαλά
και επιτρέπουν την έξοδο ιόντων χλωρίου από τα επιθηλιακά κύτταρα προς τον αεραγωγό. Η βλέννη του
αεραγωγού, έτσι, παραµένει ενυδατωµένη, καθώς µόρια νερού εισέρχονται στον αεραγωγό λόγω ώσµωσης.
Δεξιά απεικονίζεται ο αεραγωγός ενός ατόµου που πάσχει από κυστική ίνωση (Lung airway of person with
cystic fibrosis). Τα κανάλια CFTR δεν λειτουργούν οµαλά και δεν επιτρέπουν την έξοδο ιόντων χλωρίου
από τα επιθηλιακά κύτταρα προς τον αεραγωγό. Η βλέννη του αεραγωγού, έτσι, δεν ενυδατώνεται (τα µόρια
νερού δεν εισέρχονται στον αεραγωγό), γίνεται πηκτή, κολλώδης, δυσκίνητη και παγιδεύει βακτήρια που
δεν µπορούν να αποµακρυνθούν από το κροσσωτό επιθήλιο. [πηγή:
http://www.nchpeg.org/nutrition/index.php?option=com_content&view=article&id=468&tmpl=compone
nt ]
Οι αφυδατωµένες και κολλώδεις εκκρίσεις στους πνεύµονες των ασθενών µε κυστική ίνωση
έχουν πολλές δυσµενείς επιπτώσεις4,10:
• εµποδίζουν την κάθαρση του κροσσωτού επιθηλίου από τις βλέννες,
• αναστέλλουν τη λειτουργία των φυσιολογικά αναπτυσσόµενων αντιµικροβιακών
πεπτιδίων,
• παρέχουν µέσο για την ανάπτυξη παθογόνων οργανισµών
• αποφράσσουν την κυκλοφορία του αέρα.
Εντός των πρώτων µηνών ζωής, αυτές οι εκκρίσεις και τα βακτήρια που τις εποικίζουν
ξεκινούν µια φλεγµονώδη αντίδραση, η οποία χαρακτηρίζεται από απελευθέρωση
φλεγµονωδών κυτοκινών, αντιβακτηριακών ενζύµων και βακτηριακών ενζύµων που
καταστρέφουν τα βρογχιόλια. Επαναλαµβανόµενοι κύκλοι λοιµώξεων, φλεγµονής και ιστικής
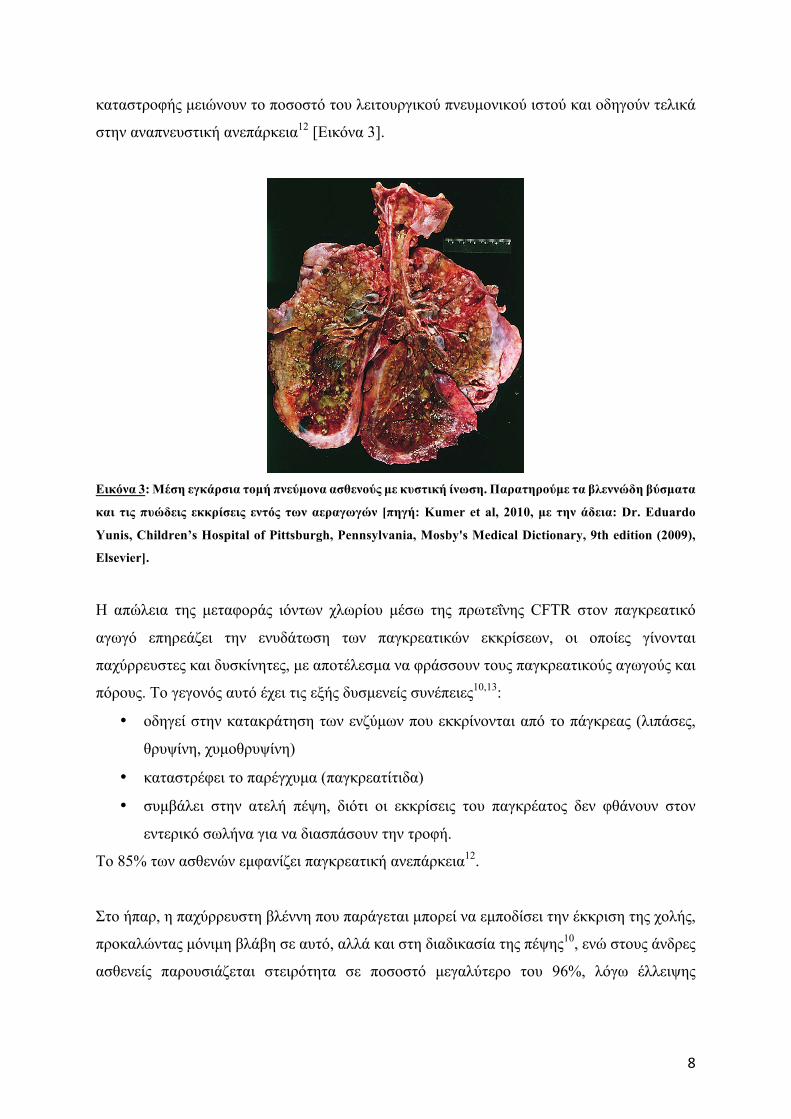
8
καταστροφής µειώνουν το ποσοστό του λειτουργικού πνευµονικού ιστού και οδηγούν τελικά
στην αναπνευστική ανεπάρκεια12 [Εικόνα 3].
Εικόνα 3: Μέση εγκάρσια τοµή πνεύµονα ασθενούς µε κυστική ίνωση. Παρατηρούµε τα βλεννώδη βύσµατα
και τις πυώδεις εκκρίσεις εντός των αεραγωγών [πηγή: Κumer et al, 2010, µε την άδεια: Dr. Eduardo
Yunis, Children’s Hospital of Pittsburgh, Pennsylvania, Mosby's Medical Dictionary, 9th edition (2009),
Elsevier].
Η απώλεια της µεταφοράς ιόντων χλωρίου µέσω της πρωτεΐνης CFTR στον παγκρεατικό
αγωγό επηρεάζει την ενυδάτωση των παγκρεατικών εκκρίσεων, οι οποίες γίνονται
παχύρρευστες και δυσκίνητες, µε αποτέλεσµα να φράσσουν τους παγκρεατικούς αγωγούς και
πόρους. Το γεγονός αυτό έχει τις εξής δυσµενείς συνέπειες10,13:
• οδηγεί στην κατακράτηση των ενζύµων που εκκρίνονται από το πάγκρεας (λιπάσες,
θρυψίνη, χυµοθρυψίνη)
• καταστρέφει το παρέγχυµα (παγκρεατίτιδα)
• συµβάλει στην ατελή πέψη, διότι οι εκκρίσεις του παγκρέατος δεν φθάνουν στον
εντερικό σωλήνα για να διασπάσουν την τροφή.
Το 85% των ασθενών εµφανίζει παγκρεατική ανεπάρκεια12.
Στο ήπαρ, η παχύρρευστη βλέννη που παράγεται µπορεί να εµποδίσει την έκκριση της χολής,
προκαλώντας µόνιµη βλάβη σε αυτό, αλλά και στη διαδικασία της πέψης10, ενώ στους άνδρες
ασθενείς παρουσιάζεται στειρότητα σε ποσοστό µεγαλύτερο του 96%, λόγω έλλειψης

9
(αµφίπλευρη ή µονόπλευρη) του σπερµατικού πόρου (η έλλειψη αυτή δηµιουργεί προβλήµατα
στη µεταφορά σπέρµατος από τους όρχεις)12.
Παραδείγµατα µονοσυµπτωµατικών µορφών της νόσου αποτελούν η ανδρική στειρότητα, η
χρόνια ή οξεία παγκρεατίτιδα, η παραρρινοκολπίτιδα, οι διάχυτες βρογχεκτασίες, η χρόνια
αποφρακτική πνευµονοπάθεια και το άσθµα 14–18 .
Η αυξηµένη συγκέντρωση άλατος (NaCl), µεγαλύτερη από 60mEq/L, στον ιδρώτα των
ασθενών αποτελεί το κυριότερο διαγνωστικό µέσο12. O µέσος χρόνος ηλικίας διάγνωσης είναι
7 µηνών και σε περίπου το 60% των πασχόντων η διάγνωση πραγµατοποιείται µέχρι τον πρώτο
χρόνο ζωής. Χωρίς θεραπευτική αντιµετώπιση, η κυστική ίνωση καταλήγει σε θάνατο κατά τη
νεογνική και παιδική περίοδο12 .
Σήµερα δεν υπάρχει αποτελεσµατική θεραπεία για την κυστική ίνωση, όµως νέα θεραπευτικά
πρωτόκολλα αντιµετώπισης των συµπτωµάτων έχουν αυξήσει το µέσο όρο ζωής στα 30-40
χρόνια. Οι ασθενείς, όµως, συνεχίζουν να αντιµετωπίζουν σοβαρά προβλήµατα υγείας λόγω
της δηµιουργίας παχύρρευστης βλέννας και των συχνών αναπνευστικών λοιµώξεων που
οδηγούν σε αποφρακτική πνευµονοπάθεια5.
Οι στόχοι θεραπευτικής παρέµβασης της κυστικής ίνωσης είναι οι εξής10,12:
• Κάθαρση πνευµονικών εκκρίσεων
• Έλεγχος πνευµονικών λοιµώξεων
• Χορήγηση παγκρεατικών ενζύµων για αποκατάσταση της πέψης και της θρέψης
• Πρόληψη εντερικής απόφραξης
Αν και η ιατρική περίθαλψη επιβραδύνει την πρόοδο της πνευµονικής νόσου, η µόνη
αποτελεσµατική θεραπεία της πνευµονικής ανεπάρκειας είναι η µεταµόσχευση πνευµόνων. Η
υποκατάσταση των παγκρεατικών ενζύµων και η χορήγηση συµπληρωµατικών λιποδιαλυτών
βιταµινών αντιµετωπίζουν αποτελεσµατικά την ατελή πέψη. Λόγω των αυξηµένων θερµιδικών
αναγκών και της ανορεξίας, µερικοί ασθενείς χρειάζονται θερµιδικά συµπληρώµατα. Επίσης,
αναγκαία είναι για τους περισσότερους ασθενείς η εκτεταµένη ψυχολογική υποστήριξη για να
αντιµετωπίσουν τη χρόνια αυτή θανατηφόρα νόσο10.

10
H ασθένεια έχει ιστοειδική έκφραση και σήµερα έχουν βρεθεί παραπάνω από 2000
διαφορετικές µεταλλάξεις στο γονίδιο CFTR, οι περισσότερες από τις οποίες θεωρούνται
παθολογικές και παρουσιάζουν συχνότητα που διαφέρει πληθυσµιακά και γεωγραφικά6,9. Οι
φορείς της νόσου είναι φυσιολογικοί, ενώ στους οµοζυγώτες η νόσος παρουσιάζει µεγάλη
ετερογένεια ως προς τη βαρύτητά της, λόγω του µεγάλου αριθµού µεταλλάξεων που έχουν
ανιχνευθεί στο γονίδιο CFTR. Αυτό συµβαίνει διότι οι µεταλλάξεις έχουν διαφορετικές
επιπτώσεις στη λειτουργία της πρωτεΐνης, οδηγώντας µε διαφορετικούς µηχανισµούς στην
παραγωγή µιας µη λειτουργικής πρωτεΐνης CFTR, ή µιας CFTR µε µειωµένη
λειτουργικότητα19,20. Ορισµένοι ασθενείς, µάλιστα, δεν εµφανίζουν τα συµπτώµατα της νόσου
µέχρι την ενήλικη ζωή12. Ανάλογα µε τις επιπτώσεις τους στη λειτουργία της πρωτεΐνης, οι
µεταλλάξεις στο γονίδιο CFTR κατατάσσονται σε 6 οµάδες10,11,21:
• 1η οµάδα: ανερµηνεύσιµες µεταλλάξεις (µονονουκλεοτιδικές αλλαγές που οδηγούν
στη δηµιουργία πρόωρου κωδικονίου τερµατισµού, µε αποτέλεσµα την παραγωγή µη
λειτουργικής πρωτεΐνης µε µέγεθος µικρότερο του φυσιολογικού) και µεταλλάξεις
αλλαγής πλαισίου ανάγνωσης µε µηδενική παραγωγή πρωτεΐνης (ασταθή µόρια
mRNA).
• 2η οµάδα: δυσερµηνεύσιµες µεταλλάξεις (µονονουκλεοτιδικές αλλαγές σε µια που
οδηγούν στην αλλαγή ενός αµινοξέως- µη συνώνυµη αλλαγή) και ελλείµµατα
αµινοξέων που οδηγούν στην παραγωγή πρωτεΐνης που δεν ωριµάζει φυσιολογικά (δεν
αναδιπλώνεται σωστά και άρα δεν µπορεί να υποστεί γλυκοσυλίωση εντός του
ενδοπλασµατικού δικτύου και του συµπλέγµατος Golgi, έτσι ώστε να εκκριθεί προς
την κυτταρική µεµβράνη) και έτσι καταστρέφεται από τους µηχανισµούς του
κυττάρου.
• 3η οµάδα: δυσερµηνεύσιµες µεταλλάξεις που αφορούν τις περιοχές δέσµευσης ATP
(NΒD1, NBD2) και τη ρυµθιστική περιοχή R, και έτσι επηρεάζουν τη ρύθµιση της
πρωτεΐνης, καταλήγοντας σε µειωµένη δραστικότητα.
• 4η οµάδα: δυσερµηνεύσιµες µεταλλάξεις που εντοπίζονται στις διαµεµβρανικές
περιοχές της πρωτεΐνης (TMD1, TMD2) και προκαλούν ελαττωµατική αγωγιµότητα
χλωρίου.
• 5η οµάδα: µεταλλάξεις στις περιοχές µατίσµατος που οδηγούν σε µειωµένη σύνθεση
της CFTR. Σε αυτή την περίπτωση µπορεί να υπάρχει µερική παραγωγή ενός µικρού
ποσοστού λειτουργικής πρωτεΐνης, οδηγώντας σε περισσότερο ήπιους φαινότυπους.

11
• 6η οµάδα: µεταλλάξεις που επηρεάζουν την ικανότητα ρύθµισης από την CFTR άλλων
διαµεµβρανικών αγωγών ή προκαλούν µείωση της σταθερότητάς της στη µεµβράνη
των επιθηλιακών κυττάρων.
Οι µεταλλάξεις των οµάδων 1-3 χαρακτηρίζονται ως βαριές, και συσχετίζονται µε την
παγκρεατική ανεπάρκεια. Οι µεταλλάξεις των οµάδων 4-6 θεωρούνται ήπιες και επιτρέπουν
την παραγωγή ενός ποσοστού φυσιολογικής πρωτεΐνης. Οι ασθενείς που φέρουν τις τελευταίες
χαρακτηρίζονται από παγκρεατική επάρκεια, έχουν καλύτερη ανάπτυξη και συνολική
πρόγνωση 10.
Μία συγκεκριµένη µετάλλαξη, εµφανίζει πολύ υψηλή συχνότητα εµφάνισης σε σχέση µε τις
υπόλοιπες. Η µετάλλαξη αυτή ονοµάζεται F508del ή ΔF508 και αφορά το έλλειµα 3 βάσεων
(αλληλουχία: CTT) που οδηγεί στην απώλεια ενός αµινοξέως φαινυλαλανίνης στο κωδικόνιο
508 της πρωτεΐνης, δηλαδή την πρώτη περιοχή σύνδεσης της πρωτεΐνης µε το ATP, NBD1
(ανήκει στη 2η οµάδα µεταλλάξεων). Η µετάλλαξη αυτή οφείλεται τουλάχιστον για το 70%
των περιστατικών κυστικής ίνωσης σε Καυκάσιους (στους πληθυσµούς αυτούς µόνο 7 άλλες
µεταλλάξεις έχουν συχνότητα µεγαλύτερη από 0,5%) και θεωρείται από τις βαρύτερες σε
συµπτωµατολογία µεταλλάξεις, διότι οδηγεί στην παραγωγή µιας CFTR πρωτεΐνης που δεν
είναι ικανή να ωριµάσει φυσιολογικά και καταστρέφεται από τους µηχανισµούς του κυττάρου 10,22–25. Στις βορειοευρωπαϊκές χώρες η συχνότητα της µετάλλαξης F508del είναι 70-80%, στις
Νοτιοευρωπαϊκές 30-54%, ενώ στην Ελλάδα 54.5% 25,26. Η µεγάλη επικράτηση της
µετάλλαξης αυτής (σε ετερόζυγη κατάσταση) ίσως να σχετίζεται µε την απόδοση ενός
πλεονεκτήµατος επιβίωσης, όπως για παράδειγµα η αυξηµένη αντίσταση στη µόλυνση από το
βακτήριο Salmonella typhi. Το βακτήριο αυτό χρησιµοποιεί το κανάλι CFTR για να εισέλθει
στα επιθηλιακά κύτταρα, ωστόσο όταν υπάρχει η µετάλλαξη F508del η είσοδός του
παρεµποδίζεται27. Ορισµένες µεταλλάξεις, συµπεριλαµβανοµένου της F508del, οι οποίες
έχουν εντοπιστεί σε όλες τις πληθυσµιακές οµάδες φαίνονται στον Πίνακα 1 12,25.

12
Πίνακας 1: Οι συχνότερες µεταλλάξεις της κυστικής ίνωσης παγκοσµίως και τα ποσοστά εµφάνισής τους25.
Μετάλλαξη Συχνότητα παγκοσµίως
ΔF508 66%-70%
G542X 2.4%
G551D 1.6%
N1303K 1.3%
W1282X 1.2%
Όλες οι άλλες 27.5%
Λόγω του µεγάλου αριθµού µεταλλάξεων, αλλά και της διαφορετικής συχνότητας εµφάνισής
τους στους διάφορους πληθυσµούς, είναι σχεδόν αδύνατο να ταυτοποιηθεί µια συγκεκριµένη
οµάδα µεταλλάξεων που θα ελέγχεται παγκοσµίως. Ο προληπτικός έλεγχος της νόσου,
σύµφωνα µε ευρωπαϊκές κατευθυντήριες οδηγίες πρέπει να καλύπτει το 75-85% των
µεταλλάξεων του πληθυσµού28. Σε ασθενείς ελληνικής καταγωγής έχουν βρεθεί περισσότερες
από 120 διαφορετικές µεταλλάξεις, µερικές εκ των οποίων είναι ιδιαίτερα σπάνιες. Στην
Ελλάδα και στην περιοχή της Μεσογείου παρουσιάζεται παρουσιάζει µεγάλη µοριακή
ετερογένεια (ποικιλοµορφία µεταλλάξεων), κάτι που καθιστά τον µοριακό έλεγχο ιδιαίτερα
ακριβό και δύσκολο12,29–31. Η εφαρµογή, για παράδειγµα, πρωτοκόλλων που έχουν σχεδιαστεί
για οµοιογενείς πληθυσµούς (π.χ. Βόρεια Ευρώπη, Αµερική) δεν καλύπτουν επαρκώς τον
Ελληνικό πληθυσµό 26,29,30. Οι 4 συχνότερες µεταλλάξεις στον ελλαδικό χώρο, που καλύπτουν
ένα ποσοστό ~67% φαίνονται στον Πίνακα 2.
Πίνακας 2: Οι 4 συχνότερες µεταλλάξεις και η συχνότητα εµφάνισής τους στον ελλαδικό χώρο. Οι
µεταλλάξεις αυτές καλύπτουν ένα ποσοστό ~67% των συνολικών µεταλλάξεων που εµφανίζονται στην
Ελλάδα12,31. Μετάλλαξη Συχνότητα στην Ελλάδα
ΔF508 54.5%
621+1G>T 6.3%
G542X 3.9%
N1303K 2.3%
Η κυστική ίνωση κληρονοµείται µε υπολειπόµενο σωµατικό χαρακτήρα και τα πάσχοντα
άτοµα κληρονοµούν ένα παθολογικό αλληλόµορφο από κάθε γονέα. Έτσι, ένα ζευγάρι φορέων

13
έχει πιθανότητα 25% να αποκτήσει ένα πάσχον παιδί σε κάθε εγκυµοσύνη12. O ανιχνευτικός
έλεγχος εφαρµόζεται σε όλες τις οµάδες υψηλού κινδύνου, οι οποίες είναι οι εξής10,12:
• µέλη οικογενειών µε κυστική ίνωση (ανίχνευση συγκεκριµένης µετάλλαξης).
• το άλλο µέλος ενός ζευγαριού, στο οποίο ένα άτοµο είναι φορέας της νόσου.
• άντρες µε στειρότητα λόγω έλλειψης του σπερµατικού πόρου (έλεγχος
υπογονιµότητας).
• άτοµα µε χρόνια αποφρακτική πνευµονοπάθεια και ιδιοπαθή χρόνια παγκρεατίτιδα, η
παθογένεια των οποίων δεν έχει διευκρινιστεί.
• εγκυµοσύνες µε υπερηχογένεια εντέρου στο έµβρυο κατά τον υπερηχογραφικό έλεγχο
στο δεύτερο τρίµηνο της εγκυµοσύνης.
Ο µοριακός έλεγχος πραγµατοποιείται συνήθως σε ζευγάρια πριν την εγκυµοσύνη, ή κατά τα
αρχικά στάδια µιας κύησης. Αν η µητέρα βρεθεί φορέας κάποιας µετάλλαξης, τότε επιβάλλεται
να εξεταστεί και ο πατέρας µε διευρυµένο έλεγχο µεταλλάξεων, αφού ο κίνδυνος να γεννηθεί
παιδί µε τη νόσο αυξάνεται πολύ. Πιο συγκεκριµένα, αν η µητέρα βρεθεί φορέας-
ετεροζυγώτης, τότε η πιθανότητα να γεννηθεί πάσχον παιδί είναι29:
• 1%, αν δεν εξεταστεί καθόλου ο πατέρας
• 0,14% αν ο πατέρας ελεγχθεί και βρεθεί αρνητικός για το 85% των µεταλλάξεων
(διευρυµένος µοριακός έλεγχος).
Αν αποδειχθούν και τα δύο µέλη ενός ζευγαριού ότι είναι φορείς της νόσου πραγµατοποιείται
προγεννητική διάγνωση για την κυστική ίνωση µε λήψη τροφοβλάστης, χοριακών λαχνών ή
αµνιακού υγρού. Μια ακόµη επιλογή που έχουν τα ζευγάρια είναι αυτή της Προεµφυτευτικής
Γενετικής Διάγνωσης (Preimplantation Genetic Diagnosis – PGD), η οποία δίνει τη
δυνατότητα γενετικής διάγνωσης και µεταφοράς στη µήτρα µόνο των υγιών εµβρύων που
προκύπτουν από εξωσωµατική γονιµοποίηση. Κατά τη µέθοδο αυτή πραγµατοποιείται λήψη
και εξέταση ενός κυττάρου από το έµβρυο στο στάδιο των 6-8 κυττάρων (3η µέρα µετά τη
γονιµοποίηση). Για να εφαρµοστεί η Προεµφυτευτική Γενετική Διάγνωση πρέπει να είναι
γνωστές οι µεταλλάξεις που φέρουν οι υποψήφιοι γονείς26,29 .

14
II. Η ΜΕΘΟΔΟΣ ΤΗΣ ΑΛΥΣΙΔΩΤΗΣ ΑΝΤΙΔΡΑΣΗΣ ΠΟΛΥΜΕΡΑΣΗΣ
(POLYMERASE CHAIN REACTION – PCR)
Η αλυσιδωτή αντίδραση πολυµεράσης είναι µία ευρέως χρησιµοποιούµενη τεχνική που
επιτρέπει τον πολλαπλασιασµό προεπιλεγµένης αλληλουχίας DNA σε πολλά αντίγραφα, σε
σύντοµο χρονικό διάστηµα.
Μεγάλη σηµασία για την επιτυχία της µεθόδου έχουν οι εκκινητές (primers) που
χρησιµοποιούνται, οι οποίοι είναι συνθετικά νουκλεοτίδια, µήκους 15-20 βάσεων,
συµπληρωµατικά µε τις περιοχές του DNA που επιθυµούµε να ενισχύσουµε. Οι εκκινητές
υβριδίζονται και στους δύο κλώνους DNA και αποτελούν το σηµείο έναρξης της επιµήκυνσης
του DNA από την DNA πολυµεράση32.
Η DNA πολυµεράση που χρησιµοποιείται ονοµάζεται Taq πολυµεράση και έχει αποµονωθεί
από το θερµόφιλο βακτήριο Thermus aquaticus, παρουσιάζοντας ιδιαίτερα µεγάλη αντοχή σε
υψηλές θερµοκρασίες (72-80οC). Για το λόγο αυτό, η Taq πολυµεράση διατηρεί την ενζυµική
δραστικότητά της στις συνθήκες της αντίδρασης. Σήµερα µε τις µεθόδους της Μοριακής
Βιολογίας και της Βιοτεχνολογίας έχουν παραχθεί διάφορα παράγωγα της Taq πολυµεράσης
µε βελτιωµένα χαρακτηριστικά και µεγαλύτερη εξειδίκευση. Οι βελτιωµένες αυτές
πολυµεράσες διαθέτουν επιπλέον µια 3’ προς 5’ επιδιορθωτική ικανότητα, την οποία η Taq
πολυµεράση φυσιολογικά δεν διαθέτει: αναγνωρίζουν εάν κάποιο νουκλεοτίδιο τοποθετηθεί
λανθασµένα και το αντικαθιστούν το µε το σωστό32.
Μέσα στο διάλυµα της αντίδρασης, εκτός από τους εκκινητές και την πολυµεράση,
τοποθετούνται επίσης τριφωσφορικά δεοξυριβονουκλεοτίδια (dNTPs: ATP, TTP, CTP, GTP),
ιόντα µαγνησίου (απαραίτητα για τη λειτουργία της πολυµεράσης) και ένα ρυθµιστικό διάλυµα
για τη διεξαγωγή της αντίδρασης στις βέλτιστες ιονικές συνθήκες32.
Η PCR στηρίζεται στη συνεχή επανάληψη ενός κύκλου που αποτελείται από τρία διαδοχικά
στάδια33 [Εικόνα 4]:
1. Αποδιάταξη του δίκλωνου DNA (denaturation)
Κατά το στάδιο αυτό το DNA υποβάλλεται σε θερµοκρασία 94oC προκειµένου να
επιτευχθεί ο διαχωρισµός των αλυσίδων του δίκλωνου DNA.
2. Υβριδοποίηση εκκινητών (primer annealing) στις αλληλουχίες του DNA- στόχου

15
Η θερµοκρασία µειώνεται στους 50-60οC και έτσι επιτυγχάνεται η ένωση των εκκινητών
µε τις συµπληρωµατικές αλληλουχίες DNA σε κάθε αλυσίδα.
3. Επιµήκυνση εκκινητών (extension)
Η θερµοκρασία αυξάνεται στους 72οC και η DNA πολυµεράση προσθέτει νουκλεοτίδια
(dNTPs) στο 3’ άκρο των εκκινητών, µε κατεύθυνση 5’ προς 3’. Έτσι επιτυγχάνεται η
σύνθεση των νέων συµπληρωµατικών αλυσίδων DNA.
Εικόνα 4: Τα βασικά στάδια της PCR). Στάδιο 1 (Step 1): Αποδιάταξη της διπλής αλυσίδας του τµήµατος
DNA (denaturation), Στάδιο 2 (Step 2): Ένωση των εκκινητών σε κάθε αλυσίδα (annealing), Στάδιο 3 (Step
3): Σύνθεση των νέων µορίων DNΑ (extension)33 .
H διαδικασία της PCR χωρίζετε σε τρεις φάσεις32:
1. Εκθετική φάση (exponential)
Είναι η φάση κατά την οποία έχει αρχίσει ο πολλαπλασιασµός της προεπιλεγµένης
αλληλουχίας DNA. Σε αυτή τη φάση η αντίδραση είναι πολύ αποτελεσµατική, καθώς σε
κάθε κύκλο διπλασιάζεται η προεπιλεγµένη αλληλουχία DNA.
2. Γραµµική φάση (linear)
Είναι η φάση στην οποία παρατηρείται µειωµένη παραγωγή αντιγράφων DNA, εξαιτίας
της µειωµένης ενεργότητας των αντιδραστηρίων.
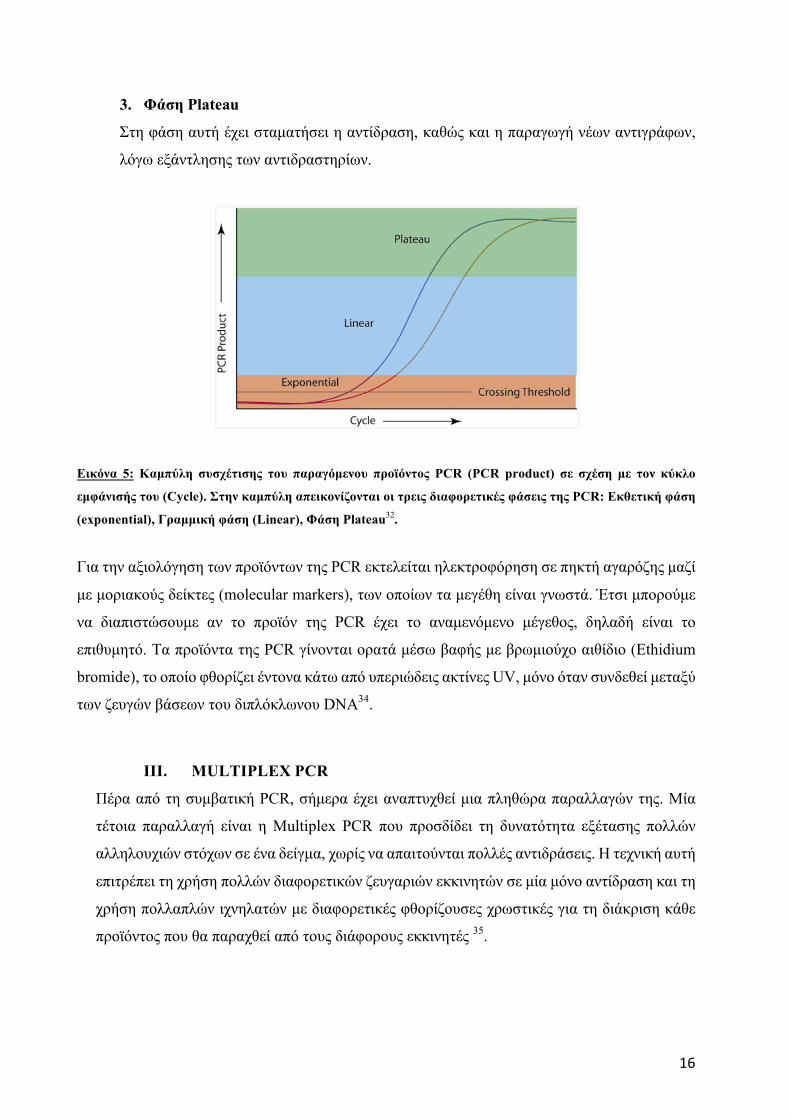
16
3. Φάση Plateau
Στη φάση αυτή έχει σταµατήσει η αντίδραση, καθώς και η παραγωγή νέων αντιγράφων,
λόγω εξάντλησης των αντιδραστηρίων.
Εικόνα 5: Καµπύλη συσχέτισης του παραγόµενου προϊόντος PCR (PCR product) σε σχέση µε τον κύκλο
εµφάνισής του (Cycle). Στην καµπύλη απεικονίζονται οι τρεις διαφορετικές φάσεις της PCR: Εκθετική φάση
(exponential), Γραµµική φάση (Linear), Φάση Plateau32.
Για την αξιολόγηση των προϊόντων της PCR εκτελείται ηλεκτροφόρηση σε πηκτή αγαρόζης µαζί
µε µοριακούς δείκτες (molecular markers), των οποίων τα µεγέθη είναι γνωστά. Έτσι µπορούµε
να διαπιστώσουµε αν το προϊόν της PCR έχει το αναµενόµενο µέγεθος, δηλαδή είναι το
επιθυµητό. Τα προϊόντα της PCR γίνονται ορατά µέσω βαφής µε βρωµιούχο αιθίδιο (Ethidium
bromide), το οποίο φθορίζει έντονα κάτω από υπεριώδεις ακτίνες UV, µόνο όταν συνδεθεί µεταξύ
των ζευγών βάσεων του διπλόκλωνου DNA34.
III. MULTIPLEX PCR
Πέρα από τη συµβατική PCR, σήµερα έχει αναπτυχθεί µια πληθώρα παραλλαγών της. Μία
τέτοια παραλλαγή είναι η Multiplex PCR που προσδίδει τη δυνατότητα εξέτασης πολλών
αλληλουχιών στόχων σε ένα δείγµα, χωρίς να απαιτούνται πολλές αντιδράσεις. Η τεχνική αυτή
επιτρέπει τη χρήση πολλών διαφορετικών ζευγαριών εκκινητών σε µία µόνο αντίδραση και τη
χρήση πολλαπλών ιχνηλατών µε διαφορετικές φθορίζουσες χρωστικές για τη διάκριση κάθε
προϊόντος που θα παραχθεί από τους διάφορους εκκινητές 35.
17
IV. PCR ΠΡΑΓΜΑΤΙΚΟΥ ΧΡΟΝΟΥ (REAL TIME-POLYMERASE
CHAIN REACTION – RT-PCR)
Η αλυσιδωτή αντίδραση πολυµεράσης πραγµατικού χρόνου, RT-PCR (Real Time- Polymerase
Chain Reaction) αποτελεί ακόµα µία παραλλαγή της συµβατικής PCR και επιτρέπει τον
πολλαπλασιασµό, την ανίχνευση και την προσοτικοποίηση ενός συγκεκριµένου τµήµατος
DNA. Αυτό επιτυγχάνεται µε την παρακολούθηση της σύνθεσης νέων προϊόντων
πολλαπλασιασµού (amplicon molecules) της αλυσιδωτής αντίδρασης πολυµεράσης σε κάθε
κύκλο36.
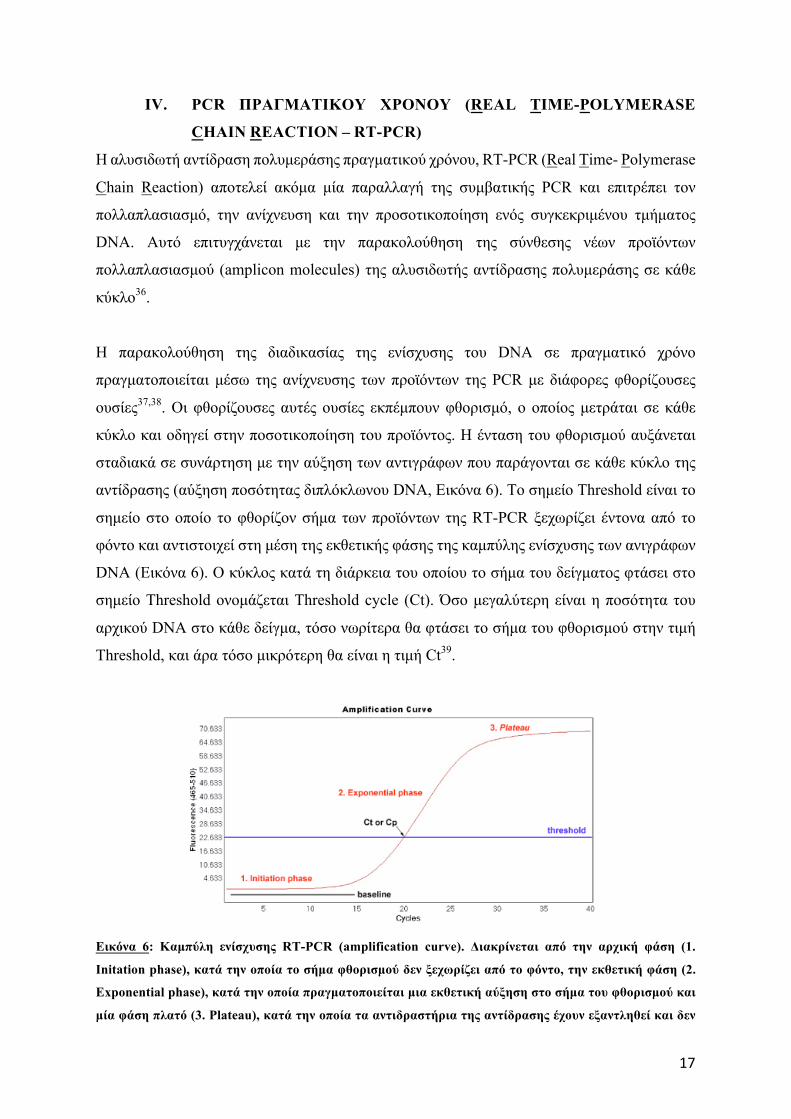
H παρακολούθηση της διαδικασίας της ενίσχυσης του DNA σε πραγµατικό χρόνο
πραγµατοποιείται µέσω της ανίχνευσης των προϊόντων της PCR µε διάφορες φθορίζουσες
ουσίες37,38. Οι φθορίζουσες αυτές ουσίες εκπέµπουν φθορισµό, ο οποίος µετράται σε κάθε
κύκλο και οδηγεί στην ποσοτικοποίηση του προϊόντος. Η ένταση του φθορισµού αυξάνεται
σταδιακά σε συνάρτηση µε την αύξηση των αντιγράφων που παράγονται σε κάθε κύκλο της
αντίδρασης (αύξηση ποσότητας διπλόκλωνου DNA, Eικόνα 6). Το σηµείο Threshold είναι το
σηµείο στο οποίο το φθορίζον σήµα των προϊόντων της RT-PCR ξεχωρίζει έντονα από το
φόντο και αντιστοιχεί στη µέση της εκθετικής φάσης της καµπύλης ενίσχυσης των ανιγράφων
DNA (Εικόνα 6). Ο κύκλος κατά τη διάρκεια του οποίου το σήµα του δείγµατος φτάσει στο
σηµείο Threshold ονοµάζεται Threshold cycle (Ct). Όσο µεγαλύτερη είναι η ποσότητα του
αρχικού DNA στο κάθε δείγµα, τόσο νωρίτερα θα φτάσει το σήµα του φθορισµού στην τιµή
Threshold, και άρα τόσο µικρότερη θα είναι η τιµή Ct39.
Εικόνα 6: Καµπύλη ενίσχυσης RT-PCR (amplification curve). Διακρίνεται από την αρχική φάση (1.
Initation phase), κατά την οποία το σήµα φθορισµού δεν ξεχωρίζει από το φόντο, την εκθετική φάση (2.
Exponential phase), κατά την οποία πραγµατοποιείται µια εκθετική αύξηση στο σήµα του φθορισµού και
µία φάση πλατό (3. Plateau), κατά την οποία τα αντιδραστήρια της αντίδρασης έχουν εξαντληθεί και δεν
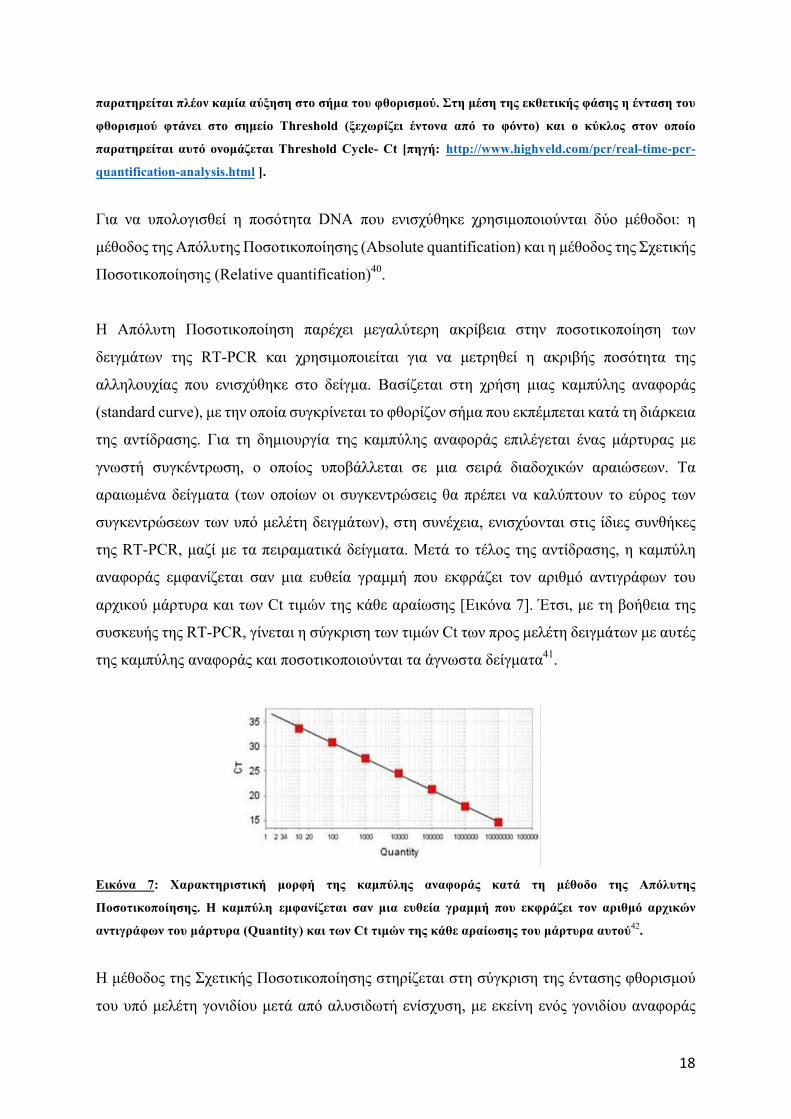
18
παρατηρείται πλέον καµία αύξηση στο σήµα του φθορισµού. Στη µέση της εκθετικής φάσης η ένταση του
φθορισµού φτάνει στο σηµείο Threshold (ξεχωρίζει έντονα από το φόντο) και ο κύκλος στον οποίο
παρατηρείται αυτό ονοµάζεται Threshold Cycle- Ct [πηγή: http://www.highveld.com/pcr/real-time-pcr-
quantification-analysis.html ].
Για να υπολογισθεί η ποσότητα DNA που ενισχύθηκε χρησιµοποιούνται δύο µέθοδοι: η
µέθοδος της Απόλυτης Ποσοτικοποίησης (Absolute quantification) και η µέθοδος της Σχετικής
Ποσοτικοποίησης (Relative quantification)40.
H Aπόλυτη Ποσοτικοποίηση παρέχει µεγαλύτερη ακρίβεια στην ποσοτικοποίηση των
δειγµάτων της RT-PCR και χρησιµοποιείται για να µετρηθεί η ακριβής ποσότητα της
αλληλουχίας που ενισχύθηκε στο δείγµα. Βασίζεται στη χρήση µιας καµπύλης αναφοράς
(standard curve), µε την οποία συγκρίνεται το φθορίζον σήµα που εκπέµπεται κατά τη διάρκεια
της αντίδρασης. Για τη δηµιουργία της καµπύλης αναφοράς επιλέγεται ένας µάρτυρας µε
γνωστή συγκέντρωση, ο οποίος υποβάλλεται σε µια σειρά διαδοχικών αραιώσεων. Τα
αραιωµένα δείγµατα (των οποίων οι συγκεντρώσεις θα πρέπει να καλύπτουν το εύρος των
συγκεντρώσεων των υπό µελέτη δειγµάτων), στη συνέχεια, ενισχύονται στις ίδιες συνθήκες
της RT-PCR, µαζί µε τα πειραµατικά δείγµατα. Μετά το τέλος της αντίδρασης, η καµπύλη
αναφοράς εµφανίζεται σαν µια ευθεία γραµµή που εκφράζει τον αριθµό αντιγράφων του
αρχικού µάρτυρα και των Ct τιµών της κάθε αραίωσης [Εικόνα 7]. Έτσι, µε τη βοήθεια της
συσκευής της RT-PCR, γίνεται η σύγκριση των τιµών Ct των προς µελέτη δειγµάτων µε αυτές
της καµπύλης αναφοράς και ποσοτικοποιούνται τα άγνωστα δείγµατα41.
Εικόνα 7: Χαρακτηριστική µορφή της καµπύλης αναφοράς κατά τη µέθοδο της Απόλυτης
Ποσοτικοποίησης. Η καµπύλη εµφανίζεται σαν µια ευθεία γραµµή που εκφράζει τον αριθµό αρχικών
αντιγράφων του µάρτυρα (Quantity) και των Ct τιµών της κάθε αραίωσης του µάρτυρα αυτού42.
Η µέθοδος της Σχετικής Ποσοτικοποίησης στηρίζεται στη σύγκριση της έντασης φθορισµού
του υπό µελέτη γονιδίου µετά από αλυσιδωτή ενίσχυση, µε εκείνη ενός γονιδίου αναφοράς

19
(reference-control gene). Το γονίδιο αναφοράς προέρχεται από τον ίδιο οργανισµό και
ενισχύεται στην ίδια αντίδραση34.
H µεθοδολογία της αντίδρασης της RT-PCR θα πρέπει να περιλαµβάνει πάντα και αρνητικούς
µάρτυρες µαζί µε τα προς ανάλυση δείγµατα. Οι αρνητικοί µάρτυρες αφορούν την εφαρµογή
κανονικής αντίδρασης µε χρήση όλων των υλικών της PCR, πλην της προσθήκης του DNA.
Συνήθως χρησιµοποιείται νερό αντί του αντίστοιχου όγκου DNA, για να εξεταστεί η
περίπτωση επιµόλυνσης του διαλύµατος της αντίδρασης PCR µε ξένο DNA.
Προκειµένου να ποσοτικοποιηθεί η αλληλουχία του DNA που ενισχύθηκε κατά την αντίδραση
χρησιµοποιούνται ειδικά ολιγονουκλεοτίδια-ιχνηλάτες (probes). Οι ιχνηλάτες αυτοί έχουν
συνδεδεµένη στο ένα τους άκρο µια φθορίζουσα χρωστική (fluorophore) και στο άλλο άκρο
τους έχουν συνδεδεµένο ένα µόριο-αποσβέστη του φθορισµού (quencher). Οι ιχνηλάτες αυτοί
µάλιστα έχουν σχεδιαστεί µε βάση την αλληλουχία του DNA στόχου, έτσι ώστε να
υβριδοποιούνται µαζί της µόλις την εντοπίσουν41 . Όταν υβριδοποιούνται µε το DNA στόχο,
οι ιχνηλάτες φθορίζουν, διότι πραγµατοποιείται διαχωρισµός της φθορίζουσας χρωστικής από
το µόριο αποσβέστη34,41. Όταν ο ιχνηλάτης δεν βρει τη συµπληρωµατική του αλληλουχία στο
δείγµα, δεν υβριδοποιείται µε αυτό, µε αποτέλεσµα η φθορίζουσα χρωστική και ο αποσβέστης
να παραµένουν ενωµένοι και ο αποσβέστης να απορροφά το φθορισµό της χρωστικής41. Οι
ιχνηλάτες είναι δυνατό να σηµανθούν µε διαφορετικές φθορίζουσες χρωστικές και έτσι
καθιστούν δυνατή την ανίχνευση περισσότερων από µία αλληλουχιών σε µία µόνο αντίδραση.
Τα είδη ιχνηλατών που χρησιµοποιούνται είναι τα εξής:
1. Γραµµικοί ιχνηλάτες (TaqMan probes)
Είναι ολιγονουκλεοτίδια 20-30 βάσεων και είναι οι πιο διαδεδοµένοι. Φέρουν στο 5’ άκρο
τους µια φθορίζουσα χρωστική (fluorophore) και στο 3’ άκρο τους ένα µόριο αποσβέστη του
φθορισµού (quencher). Ο ιχνηλάτης συνδέεται στο DNA µετά το σηµείο που έχει τοποθετηθεί
ο εκκινητής. Όταν η Taq πολυµεράση φτάσει στον ιχνηλάτη, εξαιτίας της επιδιορθωτικής της
ικανότητας (3’ προς 5’ εξωνουκλεάση) ελευθερώνει τη φθορίζουσα χρωστική στο διάλυµα.
Έτσι η φθορίζουσα χρωστική αποµακρύνεται από τον αποσβέστη και όταν διεγερθεί από µία
εξωγενή πηγή φωτός αρχίζει να φθορίζει36,41 [Εικόνα 8].

20
Εικόνα 8: Μηχανισµός λειτουργίας των γραµµικών ιχνηλατών (TaqMan probes). Οι δύο κλώνοι του DNA
αποδιατάσσονται (Denaturate) και ο ιχνηλάτης συνδέεται στο DNA µετά τον εκκινητή (Anneal). Κατά την
επιµήκυνση του εκκινητή από την DNA πολυµεράση (Εxtend) αφαιρείται η φθορίζουσα χρωστική του
ιχνηλάτη (R) και αποµακρύνεται από τον αποσβέστη (Q). Αυτό επιτρέπει στη φθορίζουσα χρωστική να
εκπέµψει σήµα φθορισµού, αφού διεγερθεί από µία εξωγενή πηγή φωτός µε ορισµένο µήκος κύµατος (hv).
H ποσότητα του φθορισµού που εκπέµπεται σε κάθε κύκλο PCR είναι ανάλογη µε την ποσότητα του
παραγόµενου προϊόντος43.
2. Ιχνηλάτες µε δευτεροταγή δοµή (Molecular Beacons)
Οι ιχνηλάτες αυτοί έχουν τη µορφή «έµµισχης θηλιάς» που µοιάζει µε φουρκέτα. Η
αλληλουχία της θηλιάς (µονόκλωνη) είναι συµπληρωµατική µε την αλληλουχία του DNA
στόχου, ενώ ο µίσχος (δίκλωνο DNA) έχει µια φθοροχρωστική (fluorophore) στο ένα του άκρο
και έναν αποσβέστη φθορισµού (quencher) στο άλλο. O µηχανισµός λειτουργίας των
Molecular Beacons βασίζεται στις υψηλές θερµοκρασίες που δηµιουργούνται κατά τη
διάρκεια της αντίδρασης PCR41,44. Σε υψηλές θερµοκρασίες ο ιχνηλάτης συνδέεται µε την
αλληλουχία-στόχο και η θηλιά αποδιατάσσεται και ανοίγει. Τα δύο άκρα του µίσχου έτσι
αποµακρύνονται και η φθοροχρωστική είναι ικανή να εκπέµψει σήµα φθορισµού, εάν
διεγερθεί από µια εξωτερική πηγή φωτός [Εικόνα 9].

21
Εικόνα 9: Δοµή και τρόπος λειτουργίας των ιχνηλατών µε δευτεροταγή δοµή (molecular beacons).
Αποτελούνται από µία θηλιά που αναγνωρίζει τη συγκεκριµένη αλληλουχία του DNA στόχου που θέλουµε
να ανιχνεύσουµε και µία συµπληρωµατική αλληλουχία που σχηµατίζει ένα διπλόκλωνο µίσχο (δοµή που
µοιάζει µε φουρκέτα). Στα άκρα του µορίου συνδέονται η φθορίζουσα χρωστική (fluorescent reporter dye)
και ο αποσβέστης (quencher dye). Στη διαλυτή του µορφή, το µόριο διατηρεί τη δοµή φουρκέτας και έτσι
ο αποσβέστης και η φθοροχρωστική βρίσκονται κοντά, κάτι που επιτρέπει στον αποσβέστη να
απορροφήσει το φθορισµό της φθοροχρωστικής. Όταν όµως το µόριο συνδεθεί µε την αλληλουχία στόχο,
αποδιατάσσεται και η φθοροχρωστική αποµακρύνεται από τον αποσβέστη, κάτι που της επιτρέπει να
φθορίζει όταν διεγείρεται από µία εξωγενή πηγή φωτός [πηγή: http://www.biotechspace.site90.com/?p=33].
3. Iχνηλάτες τύπου σκορπιού (Scorpion probes)
Οι ιχνηλάτες αυτοί διαθέτουν µια έµµισχη θηλιά συµπληρωµατική προς την αλληλουχία του
DNA που θέλουµε να εντοπίσουµε, η οποία είναι επιπλέον ενσωµατωµένη στον ένα εκκινητή.
Η φθορίζουσα χρωστική βρίσκεται στο 5’ άκρο του ιχνηλάτη, ενώ στο 3’ άκρο βρίσκεται ο
εκκινητής. Ο αποσβέστης βρίσκεται ανάµεσα στον εκκινητή και στον ιχνηλάτη. Όταν συντεθεί
η αλληλουχία στόχος της θηλιάς κατά την επιµήκυνση του DNA, ο ιχνηλάτης αποδιατάσσεται
και υβριδίζεται στο DNA, κάτι που αποµακρύνει τη φθοροχρωστική µακριά από τον
αποσβέστη45 [Εικόνα 10].

22
Εικόνα 10: Η δοµή και ο τρόπος λειτουργίας ενός ιχνηλάτη τύπου σκορπιού. Η φθοροχρωστική (κόκκινο)
βρίσκεται συνδεδεµένη στο 5’ άκρο, ενώ ο εκκινητής (primer) βρίσκεται στο 3’ άκρο του ιχνηλάτη. Ο
αποσβέστης (πράσινο) βρίσκεται µεταξύ ιχνηλάτη και εκκινητή. Όταν συντεθεί η αλληλουχία στόχος της
θηλιάς του ιχνηλάτη (DNA synthesis) η θηλιά υβριδίζεται µε την αλληλουχία αυτή (annealing) και έτσι η
φθοροχρωστική αποµακρύνεται από τον αποσβέστη και γίνεται δυνατή η εκποµπή σήµατος φθορισµού από
αυτήν [πηγή: http://pubs.acs.org/subscribe/archive/mdd/v05/i02/html/alltoolbox.html ].
4. Iχνηλάτες τύπου FRET (Fluorescence Resonance Energy Transfer)
Για την υβριδοποίηση τύπου FRET απαιτούνται 2 εκκινητές και 2 ιχνηλάτες. Οι ιχνηλάτες
τύπου FRET έχουν σηµανθεί µε διαφορετικές χρωστικές και όταν υβρίζονται στην αλληλουχία
στόχο τους βρίσκονται µεταξύ των εκκινητών. Ο ένας ιχνηλάτης φέρει µια φθορίζουσα
χρωστική δότη στο 3’ άκρο, ενώ ο άλλος φέρει µια φθορίζουσα χρωστική αποδέκτη στο
5΄άκρο. Όταν οι δυο ιχνηλάτες υβριδιστούν ο ένας κοντά στον άλλο, τότε ενέργεια φθορισµού
µεταφέρεται από το δότη στον αποδέκτη και έτσι γίνεται η εκποµπή του φθορίζοντος σήµατος
που καταγράφεται από το µηχάνηµα της RT-PCR36 [Εικόνα 11].
Εικόνα 11: Δοµή και τρόπος λειτουργίας των ιχνηλατών τύπου FRET. Οι δύο ιχνηλάτες έχουν σηµανθεί µε
διαφορετικές χρωστικές, οι οποίες εντοπίζονται στο 5’ άκρο του ενός και στο 3’ άκρο του άλλου (κίτρινο
και πράσινο). Υβριδίζονται σε αλληλουχίες στόχους µεταξύ των εκκινητών (hybridisation) και η
φθορίζουσα χρωστική του ενός ιχνηλάτη- δότη (donor) µεταφέρει ενέργεια φθορισµού στη φθορίζουσα

23
χρωστική του ιχνηλάτη- αποδέκτη (acceptor), µε αποτέλεσµα η τελευταία να εκπέµπει ανιχνεύσιµο σήµα
φθορισµού (detection) [πηγή: https://www.eurofinsgenomics.eu/en/dna-rna-oligonucleotides/optimised-
application-oligos/qpcr-probes/lightcycler-probes.aspx ].
3. ΠΡΑΚΤΙΚΟ ΜΕΡΟΣ Στο πλαίσιο της παρούσας εργασίας εφαρµόστηκαν δυο διαφορετικές µέθοδοι ανίχνευσης
µεταλλάξεων της κυστικής ίνωησης:
• Η ανίχνευση µόνο της µετάλλαξης F508del, µε τη βοήθεια της RT-PCR.
• Η ταυτόχρονη ανίχνευση 29 συχνών µεταλλάξων (συµπεριλαµβανοµένου της
F508del) που οδηγούν στην εµφάνιση της νόσου της κυστικής ίνωσης, µε τη βοήθεια
της µεθόδου PCR και της ηλεκτροφόρησης σε πηκτή αγαρόζης.
I. ΑΠΟΜΟΝΩΣΗ ΓΕΝΩΜΙΚΟΥ DNA ΑΠΟ ΠΕΡΙΦΕΡΙΚΟ ΑΙΜΑ
Αρχικά, πραγµατοποιήθηκε αποµόνωση γενωµικού DNA από το περιφερικό αίµα των προς
εξέταση ατόµων, µε τη βοήθεια ειδικού kit, διαθέσιµου στο εµπόριο. Τοποθετήθηκαν 25µL
Πρωτεϊνάσης Κ σε ειδικό σωλήνα λύσης (Lysis Tube, 1.5mL) του kit. H Πρωτεϊνάση Κ οδηγεί
στη λύση των κυττάρων. Στη συνέχεια προστέθηκαν 200µL από το δείγµα αίµατος τα οποία
αναµίχθηκαν µε την Πρωτεϊνάση Κ και 200µL διαλύµατος λύσης (Buffer B3), το οποίο
περιέχει απορρυπαντικό. Το µίγµα αναδεύτηκε µε στροβιλισµό για 10 δευτερόλεπτα.
Ακολούθησε επώαση του µίγµατος για 5 λεπτά σε θερµοκρασία δωµατίου (18-25οC) και για
10 λεπτά στους 70οC. Μετά την πάροδο του χρόνου το µίγµα αναδεύτηκε µε στροβιλισµό για
5 δευτερόλεπτα. Στη συνέχεια, το µίγµα φυγοκεντρήθηκε για 1 δευτερόλεπτο, µε σκοπό να
αποµακρυνθούν τυχόν σταγόνες από το καπάκι του σωλήνα λύσης, και ακολούθησε προσθήκη
210µL απόλυτης αιθανόλης και ανάδευση µε στροβιλισµό για 5 δευτερόλεπτα. Έπειτα, ο
σωλήνας λύσης φυγοκεντρήθηκε για 1 δευτερόλεπτο, µε σκοπό να αποµακρυνθούν τυχόν
σταγόνες από το καπάκι και στη συνέχεια όλο το περιεχόµενό του µεταφέρθηκε στην ειδική
στήλη του kit, η οποία τοποθετήθηκε µέσα σε έναν σωλήνα συλλογής (Collection Tube). Το
δείγµα φυγοκεντρήθηκε για 1 λεπτό στα 11.000 g, µε αποτέλσµα το DNA να παραµείνει στο
ειδικό φίλτρο που διαθέτει η στήλη και το υπόλοιπο δείγµα να καταλήξει στο σωλήνα
συλλογής. Να σηµειωθεί πως η αιθανόλη αποτελεί αναστολέα των αντιδράσεων της RT-PCR,
ωστόσο µε τις συνεχείς φυγοκεντρήσεις, το DNA στεγνώνει από την αιθανόλη και έτσι δεν
δηµιουργείται κάποιο πρόβληµα στη συνέχεια. Ένας νέος σωλήνας συλλογής αντικαθιστά τον

24
πρώτο και σε αυτόν τοποθετείται η στήλη που στη µεµβράνη της περιέχει το DNA. Έπειτα στη
στήλη προστίθενται 500µL από το Buffer πλύσης (BW) του kit και ακολουθεί φυγοκέντρηση
για 1 λεπτό στα 11.000 g. Το DNA παραµένει στο φίλτρο της στήλης, ενώ ο σωλήνας συλλογής
αντικαθίσταται από νέο σωλήνα. Στη συνέχεια προστίθενται 600µL από το Buffer πλύσης (B5)
του kit και ακολουθεί φυγοκέντρηση για 1 λεπτό στα 11.000 g. Το DNA βρίσκεται ακόµα στο
φίλτρο της στήλης. Η στήλη τοποθετείται σε ένα σωλήνα έκλουσης του kit (Elution Tube,
1.5mL) και σε αυτή προστίθενται 50-200µL από το Buffer έκλουσης του kit (BE). Ακολουθεί
φυγοκέντρηση για 1 λεπτό στα 11.000 g και το DNA εκλούεται από το φίλτρο στο διάλυµα
και συλλέγεται στο σωλήνα έκλουσης [Εικόνα 12].
Εικόνα 12: Περιληπτική απεικόνιση των σταδίων αποµόνωσης γενωµικού DNA από περιφερικό αίµα µε
τη βοήθεια ειδικού kit διαθέσιµου στο εµπόριο [πηγή: http://www.mn-
net.com/Portals/8/attachments/Redakteure_Bio/Protocols/Genomic%20DNA/UM_gDNADxBlood.pdf ].

25
II. ΑΝΙΧΝΕΥΣΗ ΤΗΣ ΜΕΤΑΛΛΑΞΗΣ F508DEL (ΔF508) ME TH
ΒΟΗΘΕΙΑ ΤΗΣ RT-PCR
Για την ανίχνευση της µετάλλαξης F508del στο γνωµικό DNA που αποµονώθηκε από
περιφερικό αίµα χρησιµοποιήθηκε ειδικό kit διαθέσιµο στο εµπόριο. Προκειµένου να γίνει
γενοτύπηση του ατόµου (δηλαδή να χαρακτηριστούν τα αλληλόµορφα του γονιδίου CFTR που
φέρει ως προς τη µετάλλαξη F508del) το kit που χρησιµοποιήθηκε περιέχει δυο σηµασµένους
ιχνηλάτες, οµόλογους στους δύο διαφορετικούς γενότυπους που διερευνώνται. O ιχνηλάτης
που είναι οµόλογος και συνδέεται στου αγρίου τύπου γονίδιο CFTR φέρει τη φθορίζουσα
χρωστική ROX, ενώ ο ιχνηλάτης που συνδέεται στο γονίδιο CFTR που φέρει τη µετάλλαξη
F508del είναι σηµασµένος µε τη φθορίζουσα χρωστική VIC. Κατά την ενίσχυση του γονιδίου
CFTR µε τη βοήθεια των αντιδράσεων της RT-PCR, οι δύο ιχνηλάτες ανταγωνίζονται για
πρόσδεση στην περιοχή του γονιδίου CFTR που εµφανίζεται η µετάλλαξη F508del. O
ιχνηλάτης, όµως, που είναι 100% οµόλογος µε την αλληλουχία του DNA σε αυτή την περιοχή
συνδέεται µε µεγαλύτερη ευκολία και δίνει σήµα φθορισµού (ROX ή VIC) καθώς προχωρούν
οι αντιδράσεις της PCR. Εάν το δείγµα είναι οµόζυγο για το αγρίου τύπου αλληλόµορφο, τότε
θα δηµιουργηθεί ένα πολύ ισχυρό σήµα φθορισµού από τον ιχνηλάτη που είναι οµόλογος µε
την αγρίου τύπου αλληλουχία (σήµα από τη χρωστική ROX). Εάν το δείγµα είναι οµόζυγο για
τη µετάλλαξη F508del, τότε θα παραχθεί έντονο σήµα φθορισµού από τον ιχνηλάτη που
συνδέεται στην µεταλλαγµένη αλληλουχία (σήµα από τη χρωστική VIC). Τέλος, εάν το δείγµα
είναι ετερόζυγο, θα περιέχει τις θέσεις σύνδεσης και των δύο ιχνηλατών και έτσι θα παραχθούν
δύο µέσης έντασης σήµατα φθορισµού, ένα από τον κάθε ιχνηλάτη (σήµα και από τη χρωστική
ROX και από τη VIC). Με άλλα λόγια, οι τρεις διαφορετικοί φαινότυποι µπορούν να
ελεγχθούν (γενοτύπηση- genotyping) συγκρίνοντας την παραγόµενη ένταση φθορισµού στο
τέλος της αντίδρασης.
To kit περιέχει επίσης ένα θετικό µάρτυρα (positive control) για καθένα από τους 2 γενότυπους
(µεταλλαγµένος και άγριου τύπου). Αυτά τα control τοποθετούνται στο µηχάνηµα της RT-
PCR παράλληλα µε το δείγµα (ίδιες συνθήκες) και δίνουν τα σήµατα φθορισµού που πρέπει
να ανιχνευθούν για κάθε γενότυπο. Για να είναι τα σήµατα αυτά σχετικά µε τα σήµατα
φθορισµού που θα παράξουν τα δείγµατα, θα πρέπει το DNA των θετικών µαρτύρων να
βρίσκεται στην ίδια αρχική ποσότητα µε το DNA του δείγµατος. Για να επιτευχθεί αυτό,
πραγµατοποιείται κατάλληλη αραίωση στο θετικό µάρτυρα (η οποία που υποδεικνύεται από
το kit), ανάλογα µε την ποσότητα γενωµικού DNA που περιέχεται στο δείγµα.
26
Για να ελεγχθεί η πιθανότητα επιµόλυνσης µε ξένο DNA στην αντίδραση τοποθετούνται και
αρνητικοί µάρτυρες (negative controls) στους οποίους τοποθετούνται όλα τα υλικά της
αντίδρασης, αλλά αντί για DNA, προστίθεται ίση ποσότητα νερού, επεξεργασµένου
κατάλληλα, ώστε να µην περιέχει RNAάσες και DNAάσες . Εάν σε αυτό τον αρνητικό
µάρτυρα ανιχνευθεί σήµα, τότε το γεγονός αυτό αποτελεί ένδειξη επιµόλυνσης των
αντιδραστηρίων µε ξένο DNA και η αντίδραση της RT-PCR πρέπει να επαναληφθεί εκ νέου
για όλα τα δείγµατα.
To Master Mix που χρησιµοποιήθηκε για τις αντιδράσεις της RT-PCR περιείχε το ένζυµο της
Taq πολυµεράσης, τα νουκλεοτίδια, το ρυθµιστικό διάλυµα που απαιτείται για την εκτέλεση
της αντίδρασης (buffer) και τα απαραίτητα άλατα, όλα στη σωστή συγκέντρωση για να γίνει
µε επιτυχία η ενίσχυση του γονιδίου CFTR.
Για να µειωθεί το ρίσκο επιµόλυνσης των δειγµάτων µε ξένο DNA όλη η διαδικασία
πραγµατοποιείται σε αποστειρωµένο χώρο και στις πιπέτες τοποθετούνται ρύγχη µε φίλτρο
(filtered tips). To θετικό control είναι µία πολύ πιθανή πηγή επιµόλυνσης, οπότε πιπετάρεται
µε προσοχή, µετά το αρνητικό control και τα δείγµατα.
Αρχικά προετοιµάζεται το διάλυµα της αντίδρασης µε ποσότητες ανάλογες µε τον αριθµό των
αντιδράσεων που θα εκτελεστούν για τα δείγµατα, αλλά και για 2 επιπλέον αντιδράσεις που
θα πραγµατοποιηθούν για τους µάρτυρες (αρνητικός και θετικός). Οι ποσότητες
αντιδραστηρίων που αντιστοιχούν σε 1 µόνο αντίδραση φαίνονται στον Πίνακα 3:
Πίνακας 3: Ποσότητες αντιδραστηρίων του kit που απαιτούνται για την εκτέλεση 1 αντίδρασης RT-PCR.
Αντιδραστήριο Ποσότητα
Μaster Μix 10µl
Genotyping primer/probe mix (περιέχει τους
εκκινητές και τους ιχνηλάτες) 1µl
Νερό ελεύθερο από RNAάσες/DNAάσες 4µl
ΤΕΛΙΚΟΣ ΟΓΚΟΣ 15µl
Σε κάθε αντίδραση προστίθενται επιπλέον 5µl από το γενωµικό DNA που αποµονώθηκε (το
οποίο έχει αραιωθεί, έτσι ώστε η τελική του συγκέντρωση να είναι 2ng/µl) και στο αρνητικό
27
control προστίθενται 5µl νερού χωρίς RNAάσες/DNAάσες. Στο θετικό control
πραγµατοποιείται κατάλληλη αραίωση σύµφωνα µε τον Πίνακα 4, έτσι ώστε και αυτό να έχει
συγκέντρωση DNA 2ng/µl.
Πίνακας 4: Κατάλληλες αραιώσεις του θετικού µάρτυρα, έτσι ώστε να έχει την ίδια συγκρέντρωση µε το
DNA των δειγµάτων πριν ξεκινήσουν οι αντιδράσεις της RT-PCR.
Συγκέντρωση DNA δείγµατος Αραίωση Θετικού Μάρτυρα
10ng 1:25
5ng 1:50
1ng 1:250
Στη συνέχεια τα δείγµατα τοποθετούνται στο µηχάνηµα της RT-PCR (7500 RT-PCR System,
Applied Biosystems®) το οποίο προγραµµατίζεται µέσω του 7500 Software v2.0.5 για να
εκτελέσει το πρωτόκολλο ενίσχυσης (amplification protocol) που φαίνεται στον Πίνακα 5.
Πίνακας 5: Πρωτόκολλο ενίσχυσης της RT-PCR που εκτελείται µε σκοπό τη γενοτύπηση ενός ατόµου ως
προς τη µετάλλαξη F508del.
III. ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΑΠΟΤΕΛΕΣΜΑΤΩΝ
O ιχνηλάτης που συνδέεται στην αγρίου τύπου αλληλουχία είναι σηµασµένος µε τη
φθοροχρωστική ROX, ενώ ο ιχνηλάτης που συνδέεται στη µεταλλαγµένη αλληλουχία είναι
σηµασµένος µε τη φθοροχρωστική VIC. Σε περίπτωση που το δείγµα είναι οµόζυγο για του
αγρίου τύπου αλληλόµορφο, η φθοροχρωστική ROX θα δώσει ένα ισχυρό σήµα ενίσχυσης,
ενώ η χρωστική VIC ελάχιστο, εώς καθόλου. Το αντίθετο θα συµβεί σε δείγµατα οµόζυγα για
Βήµα Χρόνος Θερµοκρασία
Ενεργοποίηση
ενζύµου 2 min 95oC
10 ΚΥΚΛΟΙ Αποδιάταξη 10 s 95oC
Επιµήκυνση 60s 60oC
35 ΚΥΚΛΟΙ
Αποδιάταξη 10s 95oC
Επιµήκυνση-
Συλλογή
Δεδοµένων
60s 66oC

28
το µεταλλαγµένο αλληλόµορφο, ενώ στα ετερόζυγα δείγµατα τόσο η VIC όσο και η ROX
φθοροχρωστικές δίνουν ένα ενδιάµεσο σήµα ενίσχυσης [Εικόνα 13].
Εικόνα 13: Αξιολόγηση αποτελεσµάτων της RT-PCR κατά τη γενοτύπηση των ατόµων για τη µετάλλαξη
F508del του γονιδίου CFTR. Εάν το άτοµο είναι οµόζυγο για το άγριου τύπου αλληλόµορφο, τότε η
χρωστική ROX (πράσινη καµπύλη) δίνει πιο έντονο σήµα ενίσχυσης από τη χρωστική VIC (κόκκινη
καµπύλη). Το αντίθετο συµβαίνει στην περίπτωση που το άτοµο είναι οµόζυγο για τη µετάλλαξη F508del,
ενώ στην περίπτωση που το άτοµο είναι ετερόζυγο (φέρει τόσο το µεταλλαγµένο όσο και το αγρίου τύπου
αλληλόµορφο) ανιχνεύεται σήµα ενδιάµεσης έντασης τόσο από τη ROX όσο και από τη VIC.

29
IV. ΑΝΙΧΝΕΥΣΗ 29 ΜΕΤΑΛΛΑΞΕΩΝ ΤΗΣ ΚΥΣΤΙΚΗΣ ΙΝΩΣΗΣ ΜΕ
ΤΗ ΒΟΗΘΕΙΑ ΤΗΣ PCR
Προκειµένου να πραγµατοποιηθεί ταυτόχρονη ανίχνευση 29 διαφορετικών µεταλλάξεων του
γονιδίου CFTR της κυστικής ίνωσης [Πίνακας 6] χρησιµοποιήθηκε ειδικό kit διαθέσιµο στο
εµπόριο.
Πίνακας 6: Οι 29 διαφορετικές µεταλλάξεις του γονιδίου CFTR που ανιχνεύονται µε τη βοήθεια ειδικού
kit διαθέσιµου στο εµπόριο. Στην πρώτη στήλη αναγράφεται το όνοµα της µετάλλαξης, στη δεύτερη το
όνοµα του cDNA που παράγεται και στην τρίτη το όνοµα της µεταλλαγµένης πρωτεΐνης.
H µέθοδος πάνω στην οποία βασίζεται το kit ονοµάζεται ARMS (Amplification Refractory
Mutation System) και είναι µία στρατηγική ενίσχυσης στην οποία οι εκκινητές της PCR έχουν
σχεδιαστεί µε τέτοιο τρόπο, ώστε να επιτρέπουν τη διάκριση µεταξύ δειγµάτων DNA που
µπορεί να διαφέρουν µεταξύ τους εώς και 1 νουκλεοτίδιο. Με άλλα λόγια, οι εκκινητές είναι
σχεδιασµένοι έτσι ώστε να µπορούν να ενισχύουν ένα συγκεκριµένο αλληλόµορφο ενός
γονιδίου, το οποίο γονίδιο µπορεί να έχει εµφανίζει πολλά διαφορετικά αλληλόµορφα που
30
διαφέρουν το ένα από το άλλο εώς και µία βάση46. Αυτή η στρατηγική επιτρέπει τον εντοπισµό
σηµειακών µεταλλάξεων και απωλειών µικρού αριθµού βάσεων στην αλληλουχία κάποιου
γονιδίου.
Αρχικά το µηχάνηµα της PCR (θερµικός κυκλοποιητής- thermal cycler) προγραµµατίζεται
όπως φαίνεται στον Πίνακα 7:
Πίνακας 7: Πρωτόκολλο ενίσχυσης της PCR που εκτελείται µε σκοπό την ανίχνευση 29 διαφορετικών
µεταλλάξεων του γονιδίου CFTR µε τη βοήθεια του Elucigene CF29v2 kit της Elucigene Diagnοstics. Να
σηµειωθεί ότι στον τελευταίο κύκλο, το στάδιο ενίσχυσης στους 72οC πραγµατοποιείται για 20 λεπτά
παραπάνω.
Σε κάθε αντίδραση PCR είναι απαραίτητο να συµπεριληφθεί ένα αρνητικό δείγµα (νερό αντί
για DNA), προκειµένου να ελεγχθεί η πιθανότητα επιµόλυνσης µε ξένο DNA.
Με σκοπό να πραγµατοποιηθούν 10 αντιδράσεις προετοιµάζεται το µίγµα του ενζύµου
(Enzyme Dilution) και τοποθετούνται 85µl αποστειρωµένου και απιονισµένου νερού, 25µl
ρυθµιστικού διαλύµατος “Dilution Buffer”, 125µL χρωστικής φορτώµατος (Loading Dye) και
15µL από την πολυµεράση “AmpliTaq Gold” σε ένα σωλήνα των 1.5mL. Το µίγµα του
ενζύµου αναδεύεται καλά.
Στη συνέχεια προετοιµάζεται το µίγµα της αντίδρασης (Reaction Mix) για κάθε διαφορετικό
µίγµα εκκινητών (Primer Mix). Για 10 αντιδράσεις το µίγµα της αντίδρασης περιέχει: 165µL
από ένα συγκεκριµένο µίγµα εκκινητών (χρησιµοποιούνται 4 διαφορετικά µίγµατα εκκινητών:
TA, TB, TC, TD) σε 55µl µίγµατος ενζύµου. Ακολουθεί καλή ανάδευση και σε κάθε σωλήνα
της PCR τοποθετούνται 20µl από το µίγµα της αντίδρασης.
Βήµα Χρόνος Θερµοκρασία
Ενεργοποίηση ενζύµου
“AmpliTaq Gold” 20min 94oC
35 ΚΥΚΛΟΙ
Αποδιάταξη 30s 94oC
Υβριδοποίηση 2min 58oC
Επιµήκυνση 1min 72oC

31
Έπειτα σε κάθε σωλήνα της αντίδρασης τοποθετούνται 5µl από το γενωµικό DNA που
αποµονώθηκε από το περιφερικό αίµα, και το οποίο έχει αραιωθεί κατάλληλα, ώστε να η
τελική του συγκέντρωση να είναι 2ng/µl. Στο σωλήνα µε το αρνητικό control τοποθετούνται
5µl νερού ελεύθερου από DNAάσες και RNAάσες.
Οι σωλήνες της αντίδρασης τοποθετούνται στο θερµικό κυκλοποιητή και ξεκινάει το
πρωτόκολλο ενίσχυσης, µε το οποίο αυτός έχει προγραµµατιστεί νωρίτερα.
Μετά το τέλος του πρωτοκόλλου ενίσχυσης, τα PCR προϊόντα από κάθε σωλήνα συλλέγονται
και 15µl από το κάθε ένα ηλεκτροφορούνται σε πηκτή αγαρόζης το οποίο βρίσκεται σε
TBE/EtBr (Tris-borate/ Εthidium Βromide) ρυθµιστικό διάλυµα ηλεκτροφόρησης (Running
Buffer).
Το ρυθµιστικό διάλυµα ηλεκτροφόρησης TBE/EtBr περιέχει:
• 134mM (16.2g/L) Tris-base
• 74.9mM (4.63g/L) Βορικό οξύ
• 2.55mM (0.95g/L) EDTA
• 0.1 µg/mL Βρωµιούχο Αιθίδιο.
Στην πηκτή φορτώνονται και 15µL από ένα διάλυµα µάρτυρα (50 Βase-Pair Ladder,
1.5µg/15µL) µε την εξής σύσταση:
• 80µL αποστειρωµένου νερού
• 10µL χρωστικής φορτώµατος (loading dye)
• 10µL από τον µάρτυρα (50 Βase-Pair Ladder).
Τα δείγµατα και ο µάρτυρας ηλεκτροφορούνται στα 5 µε 6 volts/cm (τα cm υπολογίζονται από
την απόσταση µεταξύ των ηλεκτροδίων), µέχρι η χρωστική φορτώµατος να κινηθεί για 5cm
(τα cm υπολογίζονται από τις κυψελίδες που φορτώθηκαν τα δείγµατα προς την άνοδο). Μετά
την ηλεκτροφόρηση, τα δείγµατα τοποθετούνται κάτω από λάµπα UV (260nm) και
φωτογραφίζονται.

32
V. ΑΞΙΟΛΟΓΗΣΗ ΤΩΝ ΑΠΟΤΕΛΕΣΜΑΤΩΝ
Μετά την ηλεκτροφόρηση τα προϊόντα της PCR φαίνονται σαν µπάντες πάνω στο gel
αγαρόζης [Εικόνα 14]. Η πάνω και η κάτω µπάντα σε κάθε στήλη αποτελούν µάρτυρες
επιτυχούς διεξαγωγής της PCR (upper και lower control) και πρέπει να εµφανίζονται σε κάθε
δείγµα (και στο αρνητικό control). Στο αρνητικό control, βέβαια, δεν πρέπει να εµφανίζονται
καθόλου µπάντες µεταξύ της πάνω και κάτω µπάντας-µάρτυρα. Εάν εµφανιστεί κάποια
µπάντα σε αυτό, τότε δεν πρέπει να ληφθεί υπόψη σε περίπτωση που εµφανισθεί και στα προς
εξέταση δείγµατα, καθώς αποτελεί ένδειξη επιµόλυνσης µε ξένο DNA.
Εικόνα 14: Τυπική εικόνα αποτελεσµάτων ανίχνευσης 29 µεταλλάξεων του γονιδίου CFTR µε τη βοήθεια
ειδικού kit διαθέσιµου στο εµπόριο. Μετά την ηλεκτροφόρηση και εµφάνιση στο UV, τα προϊόντα της PCR
φαίνονται σαν µπάντες πάνω στο gel αγαρόζης. Ανάλογα µε το µοριακό βάρος της κάθε µπάντας (σύγκριση
µε Ladder), αλλά και ανάλογα µε το µίγµα εκκινητών το οποίο χρησιµοποιήθηκε για την ενίσχυση του
δείγµατος που τη φέρει, µπορούµε να συµπεράνουµε σε ποια ακριβώς µετάλλαξη αντιστοιχεί [Εικόνα 15].
[πηγή: http://www.elucigene.com/wp-content/uploads/2014/08/CF29v2.pdf ]
Η τεχνική αυτή µας επιτρέπει να ανιχνεύσουµε εάν ένα άτοµο είναι οµόζυγο ή ετερόζυγο µόνο
για τη µετάλλαξη F508del. Για τις υπόλοιπες 28 µεταλλάξεις µπορούµε απλά να ανιχνεύσουµε
εάν υπάρχει ή όχι το µεταλλαγµένο αλληλόµορφο. Εάν, δηλαδή, ανιχνευτεί κάποια άλλη
µετάλλαξη, µπορεί στη συνέχεια να πραγµατοποιηθεί γενοτύπηση µε άλλες µεθόδους (π.χ. RT-
PCR) προκειµένου να δειχθεί εάν το άτοµο είναι οµόζυγο ή ετερόζυγο για αυτήν.
Συγκεκριµένα, στο µίγµα εκκινητών Β, υπάρχουν εκκινητές για το γονίδιο CFTR που φέρει
την κανονική αλληλουχία στην περιοχή της F508del µετάλλαξης (µη µεταλλαγµένο µε την
F508del). Το γονίδιο αυτό µπορεί να εντοπιστεί στην περιοχή που τρέχουν τα δείγµατα στα
οποία τοποθετήθηκε το µίγµα εκκινητών Β, στο ύψος των 160bp. Για το γονίδιο που φέρει τη
µεταλλαγµένη αλληλουχία στην περιοχή της F508del µετάλλαξης υπάρχουν εκκινητές στο
µίγµα εκκινητών Α. Έτσι το γονίδιο που φέρει τη µετάλλαξη αυτή µπορεί να εντοπιστεί στην
περιοχή που τρέχουν τα δείγµατα στα οποία τοποθετήθηκε το µίγµα εκκινητών Α, στο ύψος

33
των 160bp. Εάν ένα δείγµα εµφανίσει µία µπάντα στο ύψος των 160bp, µόνο στην περιοχή
που ενισχύθηκε µε το µίγµα εκκινητών Α, τότε είναι οµόζυγο για τη µετάλλαξη F508del. Εάν
ένα δείγµα εµφανίσει µία µπάντα στο ύψος των 160bp και στην περιοχή που ενισχύθηκε µε το
µίγµα εκκινητών Α αλλά και στην περιοχή που ενισχύθηκε µε το µίγµα εκκινητών Β, τότε είναι
ετερόζυγο για τη µετάλλαξη F508del, ενώ δεν φέρει αυτή τη µετάλλαξη, εάν εµφανίσει µία
µπάντα στο ύψος των 160bp µόνο στην περιοχή που ενισχύθηκε µε το µίγµα εκκινητών Β
[Εικόνα 15].
Αντίστοιχα, εάν ένα δείγµα εµφανίσει µία µπάντα µεταξύ της πάνω και κάτω µπάντας-
µάρτυρα, ανάλογα µε το µοριακό βάρος της µπάντας (σύγκριση µε Ladder), αλλά και ανάλογα
µε το µίγµα εκκινητών στο οποίο ενισχύθηκε το δείγµα που τη φέρει, µπορούµε να
συµπεράνουµε σε ποια ακριβώς µετάλλαξη του γονιδίου CFTR αντιστοιχεί [Εικόνα 15].
Εικόνα 15: Οι 29 διαφορετικές µεταλλάξεις που ανιχνεύονται µε τη βοήθεια ειδικού kit που είναι διαθέσιµο
στο εµπόριο. Στα δεξιά (πορτοκαλί) φαίνονται οι µπάντες που δίνει ο µάρτυρας (50 Base Pair Ladder), ενώ
αριστερά φαίνονται τα σηµεία όπου εντοπίζονται οι µπάντες (µωβ) που αντιστοιχούν στις διάφορες

34
µεταλλάξεις. Οι στήλες Α, Β, C, D αντιστοιχούν στα διαφορετικά µίγµατα εκκινητών στα οποία έχει
ενισχυθεί το δείγµα. Τα µίγµατα αυτά ενισχύουν διαφορετικά µεταλλαγµένα αλληλόµορφα του γονιδίου
CFTR που µπορεί να διαφέρουν ακόµα και σε 1 βάση. Με πράσινο συµβολίζεται η µπάντα που αντιστοιχεί
στη φυσιολογική αλληλουχία στην περιοχή που ανιχνεύεται η µετάλλαξη F508del. Η φυσιολογική αυτή
αλληλουχία ανιχνεύεται όταν προστεθεί το µίγµα εκκινητών Β. Με µπλε συµβολίζονται οι πάνω και κάτω
µπάντες-µάρτυρες (lower, upper control), οι οποίες πρέπει να εµφανίζονται σε κάθε δείγµα, ακόµα και στο
αρνητικό control, µιας και υποδεικνύουν πως η αντίδραση της PCR έχει διεξαχθεί οµαλά. [πηγή:
http://www.elucigene.com/wp-content/uploads/2014/08/CF029BYEN-001.pdf ]
4. ΕΠΙΛΟΓΟΣ Η κυστική ίνωση είναι µια πολύ σοβαρή και επικίνδυνη γενετική νόσος µε ιδιαίτερα υψηλή
συχνότητα εµφάνισης στον Καυκάσιο πληθυσµό. Λόγω της µεγάλης ετερογένειας των
συµπτωµάτων της νόσου, σήµερα δεν έχει βρεθεί αποτελεσµατική θεραπεία. Ωστόσο, η
µοριακή ανίχνευση των φορέων των διαφόρων µεταλλάξεών της, αλλά και των οµοζυγωτών
απογόνων κατά τον προγεννητικό έλεγχο, µπορεί να συµβάλει στη µείωση της συχνότητας
εµφάνισης της νόσου. Η µοριακή διάγνωση µπορεί επίσης να συµβάλει στη γένεση υγιών
απογόνων από ζευγάρια φορέων, µέσω εξωσωµατικής γονιµοποίησης και µε εκτέλεση
Προεµφυτευτικής Γενετικής Διάγνωσης.
Για τους λόγους αυτούς, είναι σηµαντικό να αναπτυχθούν όσο το δυνατόν πιο ακριβείς και
γρήγορες µέθοδοι ανίχνευσης των µεταλλάξεων της νόσου στους διάφορους πληθυσµούς στον
κόσµο. Η βελτίωση των µεθόδων µοριακής ανίχνευσης των µεταλλάξεων του γονιδίου CFTR
θα µπορέσει να βοηθήσει στην πιο ακριβή και έγκαιρη διάγνωση, αποτελεσµατική θεραπεία,
αλλά και στη µείωση της συχνότητας εµφάνισης της νόσου. Να σηµειωθεί, ωστόσο, ότι ο
µόνος τρόπος να πραγµατοποιηθεί αποτελεσµατικός έλεγχος σε ένα άτοµο για όλες τις
µεταλλάξεις που έχουν εντοπιστεί στο γονίδιο CFTR είναι να αναλυθεί η αλληλουχία των
νουκλεοτιδίων του γονιδίου αυτού (sequencing).
Ακόµα, σηµαντικό είναι να επανεξετασθούν τα κριτήρια διάγνωσης της κυστικής ίνωσης.
Υπάρχουν πάρα πολλές διαφορετικοί παράµετροι που πρέπει να ληφθούν υπόψη, όπως για
παράδειγµα η ηλικία στην οποία εµφανίζονται τα συµπτώµατα και η οποία παρουσιάζει
µεγάλη ετερογένεια. Η όλο και αυξανόµενη γνώση µας για την πολυπλοκότητα των
µεταλλάξεων του γονιδίου CFTR, µε παραπάνω από 2000 µεταλλάξεις να έχουν ανιχνευθεί
σήµερα, αλλά και η συστηµατική ανάλυση του γονιδίου CFTR (αλληλούχιση εξωνίων και
ιντρονίων, εύρεση θέσεων µατίσµατος, ανίχνευση ελλειµµάτων και διπλασιασµών) έχει

35
αυξήσει την γνώση για τη πληθώρα των µεταλλαγών που µπορούν να ανιχνευθούν σε
συγκεκριµένες πληθυσµιακές οµάδες, αλλά και για τη σχέση µεταξύ φαινοτύπου και
γενοτύπου. Η σχέση αυτή µπορεί να µας βοηθήσει ιδιαίτερα στο να συσχετίσουµε τις διάφορες
µεταλλάξεις µε συγκεκριµένους τύπους δυσλειτουργιών της πρωτεΐνης CFTR και µε την
εµφάνιση συγκεκριµένων συµπτωµάτων στους ασθενείς. Το γεγονός αυτό θα συµβάλει στην
εγκυρότερη διάγνωση της νόσου και στην έγκαιρη θεραπεία των συµπτωµάτων της, έτσι ώστε
να αυξηθεί το προσδοκώµενο όριο ηλικίας των ατόµων που πάσχουν από τη χρόνια αυτή
θανατογόνο νόσο.
5. ΒΙΒΛΙΟΓΡΑΦΙΑ 1. Collins, F. S. Cystic fibrosis: molecular biology and therapeutic implications. Science
256, 774–779 (1992).
2. Devidas, S. & Guggino, W. B. CFTR: domains, structure, and function. J. Bioenerg.
Biomembr. 29, 443–451 (1997).
3. Sheppard, D. N. & Ostedgaard, L. S. Understanding how cystic fibrosis mutations cause
a loss of Cl- channel function. Mol. Med. Today 2, 290–297 (1996).
4. Kirk, K. L. New paradigms of CFTR chloride channel regulation. Cell. Mol. Life Sci.
CMLS 57, 623–634 (2000).
5. Welsh, M., Ramsey, B., Accurso, F. & Cutting, G. The metabolic and molecular bases of
inherited disease. (McGraw-Hill, 2001).
6. Kerem, B. et al. Identification of the cystic fibrosis gene: genetic analysis. Science 245,
1073–1080 (1989).
7. Riordan, J. R. et al. Identification of the cystic fibrosis gene: cloning and characterization
of complementary DNA. Science 245, 1066–1073 (1989).
8. Rommens, J. M. et al. Identification of the cystic fibrosis gene: chromosome walking and
jumping. Science 245, 1059–1065 (1989).
9. Tsui, L. Cystic Fibrosis Mutation Database. Cystic Fibrosis Genetic Analysis Consortium
(CFGAC)
10. Thompson, Nussbaum, R., McInnes, R. & Willard, H. Ιατρική Γενετική. (Πασχαλίδης).
11. Vankeerberghen, A., Cuppens, H. & Cassiman, J.-J. The cystic fibrosis transmembrane
conductance regulator: an intriguing protein with pleiotropic functions. J. Cyst. Fibros.
Off. J. Eur. Cyst. Fibros. Soc. 1, 13–29 (2002).

36
12. Kanavakis, E. & Tzetis, M. Screening for cystic fibrosis mutations: Methods for
molecular diagnosis, prenatal diagnosis, and carrier identification amongst high-risk
individuals – the Greek experience. Arch. Hell. Med. 23(5):455–472 (2006).
13. Cohn, J. A. Reduced CFTR function and the pathobiology of idiopathic pancreatitis. J.
Clin. Gastroenterol. 39, S70–77 (2005).
14. Kanavakis, E. et al. Cystic fibrosis mutation screening in CBAVD patients and men with
obstructive azoospermia or severe oligozoospermia. Mol. Hum. Reprod. 4, 333–337
(1998).
15. Tzetis, M. et al. CFTR gene mutations--including three novel nucleotide substitutions--
and haplotype background in patients with asthma, disseminated bronchiectasis and
chronic obstructive pulmonary disease. Hum. Genet. 108, 216–221 (2001).
16. Noone, P. G. & Knowles, M. R. ‘CFTR-opathies’: disease phenotypes associated with
cystic fibrosis transmembrane regulator gene mutations. Respir. Res. 2, 328–332 (2001).
17. Casals, T. et al. Bronchiectasis in adult patients: an expression of heterozygosity for
CFTR gene mutations? Clin. Genet. 65, 490–495 (2004).
18. Ahmed, N. et al. Molecular consequences of cystic fibrosis transmembrane regulator
(CFTR) gene mutations in the exocrine pancreas. Gut 52, 1159–1164 (2003).
19.http://www.cytogenomics.gr/index.php?option=com_content&view=article&id=144:cysti
c-fibrosis&catid=48:examvivliography&Itemid=147.
20. Zielenski, J. & Tsui, L. C. Cystic fibrosis: genotypic and phenotypic variations. Annu.
Rev. Genet. 29, 777–807 (1995).
21. Zielenski, J. Genotype and phenotype in cystic fibrosis. Respir. Int. Rev. Thorac. Dis. 67,
117–133 (2000).
22. Chertkoff, L. et al. Spectrum of CFTR mutations in Argentine cystic fibrosis patients.
Clin. Genet. 51, 43–47 (1997).
23. Laufer-Cahana, A. et al. Cystic fibrosis mutations in Israeli Arab patients. Hum. Mutat.
14, 543 (1999).
24. Alonso, M. J. et al. Spectrum of mutations in the CFTR gene in cystic fibrosis patients of
Spanish ancestry. Ann. Hum. Genet. 71, 194–201 (2007).
25. Bobadilla, J. L., Macek, M., Fine, J. P. & Farrell, P. M. Cystic fibrosis: a worldwide
analysis of CFTR mutations--correlation with incidence data and application to
screening. Hum. Mutat. 19, 575–606 (2002).
26. http://www.cysticfibrosis.gr/el/η-νόσος/πρόληψη-διάγνωση.

37
27. Pier, G. B. et al. Salmonella typhi uses CFTR to enter intestinal epithelial cells. Nature
393, 79–82 (1998).
28. Castellani, C. et al. European best practice guidelines for cystic fibrosis neonatal
screening. J. Cyst. Fibros. Off. J. Eur. Cyst. Fibros. Soc. 8, 153–173 (2009).
29. Kanavakis, E. et al. Cystic fibrosis in Greece: molecular diagnosis, haplotypes, prenatal
diagnosis and carrier identification amongst high-risk individuals. Clin. Genet. 63, 400–
409 (2003).
30. Casals, T., Gimenez, J., Ramos, M. D., Nunes, V. & Estivill, X. Prenatal diagnosis of
cystic fibrosis in a highly heterogeneous population. Prenat. Diagn. 16, 215–222 (1996).
31. Tzetis, M. et al. Characterization of more than 85% of cystic fibrosis alleles in the Greek
population, including five novel mutations. Hum. Genet. 99, 121–125 (1997).
32. VanGuilder, H. D., Vrana, K. E. & Freeman, W. M. Twenty-five years of quantitative
PCR for gene expression analysis. BioTechniques 44, 619–626 (2008).
33. Vierstraete, A. Principle of PCR. (1999).
34. Gibson, N. J. The use of real-time PCR methods in DNA sequence variation analysis.
Clin. Chim. Acta Int. J. Clin. Chem. 363, 32–47 (2006).
35. Hayden, M. J., Nguyen, T. M., Waterman, A. & Chalmers, K. J. Multiplex-ready PCR: a
new method for multiplexed SSR and SNP genotyping. BMC Genomics 9, 80 (2008).
36. Houghton, S. G. & Cockerill, F. R. Real-time PCR: overview and applications. Surgery
139, 1–5 (2006).
37. Heid, C. A., Stevens, J., Livak, K. J. & Williams, P. M. Real time quantitative PCR.
Genome Res. 6, 986–994 (1996).
38. Higuchi, R., Fockler, C., Dollinger, G. & Watson, R. Kinetic PCR analysis: real-time
monitoring of DNA amplification reactions. Biotechnol. Nat. Publ. Co. 11, 1026–1030
(1993).
39. Wong, M. L. & Medrano, J. F. Real-time PCR for mRNA quantitation. BioTechniques
39, 75–85 (2005).
40. Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-
time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods San Diego Calif
25, 402–408 (2001).
41. Stratagene. Introduction to Quantitive PCR. (2004).
42. Qiu, F., Cao, J., Su, Q., Yi, Y. & Bi, S. Multiplex hydrolysis probe real-time PCR for
simultaneous detection of hepatitis A virus and hepatitis E virus. Int. J. Mol. Sci. 15,
9780–9788 (2014).

38
43. Koch, W. H. Technology platforms for pharmacogenomic diagnostic assays. Nat. Rev.
Drug Discov. 3, 749–761 (2004).
44. Espy, M. J. et al. Real-time PCR in clinical microbiology: applications for routine
laboratory testing. Clin. Microbiol. Rev. 19, 165–256 (2006).
45. Arya, M. et al. Basic principles of real-time quantitative PCR. Expert Rev. Mol. Diagn. 5,
209–219 (2005).
46. Lo, Y. M. The amplification refractory mutation system. Methods Mol. Med. 16, 61–69
(1998).


















